thigmomorphogenesis: morphological, biochemical · 1 thigmomorphogenesis: morphological,...
TRANSCRIPT

Draft
Thigmomorphogenesis: morphological, biochemical
changes, and transcriptional level changes in response to
mechanical stress in Acacia koa A. Gray
Journal: Canadian Journal of Forest Research
Manuscript ID cjfr-2016-0356.R2
Manuscript Type: Article
Date Submitted by the Author: 08-Dec-2016
Complete List of Authors: Ishihara, Kazue; University of Hawaii at Manoa, Department of Molecular
Biosciences and Bioengineering Lee, Eric; University of Hawaii at Manoa, Department of Molecular Biosciences and Bioengineering Borthakur, Dulal; University of Hawaii at Manoa
Keyword: Acacia koa, thigmomorphogenesis, mechanical stress, lignin, disease resistance
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
1
Thigmomorphogenesis: morphological, biochemical changes, and transcriptional level 1
changes in response to mechanical stress in Acacia koa A. Gray 2
3
Kazue L. Ishihara, Eric K.W. Lee, Dulal Borthakur 4
University of Hawaii at Manoa 5
Department of Molecular Biosciences and Bioengineering 6
1955 East-West Road, Agricultural Sciences 218 7
Honolulu, Hawaii 96822 8
9
Kazue L. Ishihara 10
Eric K.W. Lee 12
Dulal Borthakur 14
(808) 956-6600 15
17
1 Table and 5 Figures 18
Supplementary Information: 5 Tables and 1 Figure 19
20
Page 1 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
2
ABSTRACT 21
Acacia koa A. Gray, an economically important timber-wood tree growing in the 22
Hawaiian Islands, is affected by many environmental stresses, including drought, strong wind, 23
heavy rain, and infection by fungal pathogens. Previous studies have shown that some 24
morphological and biochemical changes that take place as a result of environmental stresses in 25
plants can be also induced by mechanical stresses, such as touching and bending. We studied 26
morphological and biochemical changes and levels of gene transcription in A. koa plants due to 27
mechanical stress. For a mechanical stress treatment, A. koa seedlings were gently bent in four 28
cardinal directions daily for 2-6 months, after which morphological and biochemical changes 29
were quantified. The stressed A. koa had significantly increased stem diameter, number of xylem 30
cells, anthocyanin and lignin contents, and significantly reduced stem length. The gene 31
expression analyses showed that 53 genes, including the genes for calcium signaling, ethylene 32
biosynthesis, abscisic acid degradation, stress-related transcriptional regulation, and disease 33
resistance, were induced more than twofold within 10-60 min following mechanical stress. The 34
observation that the genes for disease resistance, such as NBS-LRR, can be induced by 35
mechanical stress suggests that strong wind and rain in the natural forest may also induce disease 36
resistance in trees. 37
38
KEY WORDS 39
Acacia koa; thigmomorphogenesis; mechanical stress; lignin; disease resistance 40
41
Page 2 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
3
INTRODUCTION 42
Mechanical perturbations, such as predation, rain, and wind, are common environmental 43
stresses for plants. Because plants are sessile and cannot travel to a new habitat on their own, the 44
ability to sense, respond, and adapt to such environmental stresses is important for their survival. 45
Responses can be rapid and apparent in some plants with specialized sensory cells. For example, 46
Dionaea muscipula closes bi-lobed leaves as soon as an insect lands on the ventral leaf surface, 47
which has ‘trigger hairs’ with sensory cells at the base (Volkov et al. 2008). Another plant 48
species Mimosa pudica has specialized pulvinar motor tissues (Visnovitz et al. 2007), and the 49
leaflets fold up quickly after touch stimulation (Temmei et al. 2005; Volkov et al. 2010); this 50
behavior may protect the leaves from insect predation or damage during rain or windstorms 51
(Koller 2011). 52
Plants without specialized sensory cells also respond to mechanical stimuli; 53
morphological responses in such plants may not be noticeable immediately, but their growth and 54
development are slowly altered over time in response to prolonged mechanical stimulation. Such 55
physiological and developmental responses of plants to mechanical stimuli are called 56
thigmomorphogenesis (Jaffe 1973). For example, plants repeatedly swayed by wind typically 57
respond by growing shorter and stockier as to protect themselves from the stimuli (Jacobs 58
1954). In experimental studies, wind-induced motion of plants has been artificially generated 59
under controlled laboratory conditions by touching or bending stems back and forth (Mitchell et 60
al. 1975, Jaffe 1976, Biro et al. 1980, Biddington and Dearman 1985). Such mechanical 61
perturbations commonly result in decreased elongation and increased thickness, and plants 62
respond to wind in the same way (Telewski and Jaffe 1986a; Braam and Davis 1990; Lee et al. 63
2005). These typical thigmomorphogenetic responses are observed in various plants, including 64
Page 3 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
4
Arabidopsis thaliana, Carcica papaya, and woody plants, such as Populus plants, Abies fraseri, 65
and Pinus taeda (Jacobs 1953; Biro et al. 1980; Telewski and Jaffe 1986a 1986b; Pruyn et al. 66
2000; Lee et al. 2005; Porter et al. 2009). Similar morphological changes have been also 67
observed to other mechanical stimuli, such as rubbing and brushing of the stems, in various 68
plants, including Arabidopsis, Bryonia dioica, Phaseolus vulgaris, and Solanum lycopersicum 69
(Jaffe 1973; Saidi et al. 2009; Paul-Victor and Rowe 2011). 70
Interestingly, some plants do not show typical thigmomorphogenetic responses, and also 71
show other diverse responses. For example, Cucurbita pepo, Pisum sativum, Triticum aestivum 72
display little to no changes in elongation after mechanical perturbations (Jaffe 1973), and the 73
stem diameter, instead of increasing as in several other thigmomorphogenetic plants, decreased 74
in Brassica oleracea (Biddington and Dearman 1985). Different morphological changes have 75
been observed in different plant species, such as a delay in flowering and an increase in number 76
of rosette paraclades in Arabidopsis (Johnson et al. 1998), tissue outgrowths in petioles, and 77
decreased anthocyanin contents in C. papaya (Porter et al. 2009). 78
Although morphological changes occur gradually, genes induced by mechanical stimuli 79
are immediately detectable (Braam and Davis 1990; Lee et al. 2005; Porter et al. 2009). In 80
Arabidopsis, approximately 2.5% of all genes were upregulated at least twofold within 30 min 81
following mechanical stimulation (Lee et al. 2005). These induced genes encode proteins with 82
diverse functions, such as calcium-binding proteins, cell-wall-modifying enzymes, kinases, 83
transcription factors, and disease resistance proteins, indicating complex molecular mechanisms 84
involved in the perception of mechanical stimulation (Lee et al. 2005). Because some 85
thigmomorphogenetic responses appear to be species-specific, different pathways may be 86
Page 4 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
5
involved in the perception of mechanical stimulation in different plants; therefore, it is important 87
to study thigmomorphogenesis in various plants. 88
In this study, we present the occurrence of changes in morphology, biochemistry, and 89
levels of gene transcription in response to mechanical stimulation in Acacia koa (koa), a 90
leguminous timber tree endemic to the Hawaiian Islands. A. koa serves as a vital economical 91
resource for the Hawaiian Islands as its timber is a highly priced commodity due to the beautiful 92
texture, hardiness, and carving quality of the wood. With a current market value of up to $125 93
per board foot, the gross value of A. koa timber and the wood products produced is estimated to 94
be in the range of $20-$30 million annually (Yanagida et al. 2004; Baker et al. 2009). To grow A. 95
koa successfully under various conditions of mechanical stimulation, such as heavy rain, 96
predation, and wind, understanding how mechanical stimulation influences the plant growth and 97
development is crucial. This study of morphological, physiological, and molecular aspects of 98
thigmomorphogenesis may contribute towards A. koa improvement programs and further 99
understanding of this phenomenon. 100
101
MATERIALS AND METHODS 102
Plant growth and treatment 103
Seeds of A. koa were obtained from three different mother trees grown in the Maunawili 104
Research Station of Hawaii Agriculture Research Center (HARC), Kailua, Hawaii. In A. koa 105
improvement program, half-sib progenies grown from the seeds obtained from a single mother 106
tree are considered a family. The seeds were immersed in concentrated sulfuric acid for 10 min 107
for scarification, rinsed with sterilized water six times, and incubated in petri dishes with wet 108
paper towels at 28 ᵒC till they germinated in 3-5 days (Rushanaedy et al. 2012). The resulting 109
Page 5 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
6
germinated seedlings were then planted into 4-inch pots containing a vermiculite-perlite mixture 110
pre-wetted with Hoagland’s solution. The seedlings were grown in a controlled growth room at 111
25 °C ± 2 °C with a 16/8-h light/dark photoperiod with an irradiance of 30 µmol s-1 m-1 for a 112
month. Plants were watered with Hoagland’s solution once in two days. For a mechanical stress, 113
stems were gently bent from their position at 90º with the ground level to a position at 114
approximately 60º with the ground level in each of all the four cardinal directions manually. 115
Plants were mechanically treated daily starting from a week after seedlings were transferred to 116
pots. The seedlings in the control group were maintained similarly, except that they were not 117
given the mechanical stress. 118
119
Phenotypic measurements and microscopy 120
Plant heights, stem diameters, and fresh and dry weights of stems, roots, and leaves of 2- 121
and 6-month-old koa seedlings were quantified. Plant heights were measured from the base of 122
the stem to the shoot tip. Stem diameters were measured at the stem base using a 5-inch digital 123
caliper. Each of the stressed and unstressed groups represented 39 2-month-old individual 124
seedlings and 15 6-month-old individual seedlings. For microscopic analysis, 2-month-old koa 125
seedlings were used; 30-µm cross-sections of stems at 5 mm below the top of the hypocotyls 126
were made using a VT1000S vibratome (Leica Biosystems Inc., IL, USA), and the cross-sections 127
were observed under an inverted microscope DMIL (Leica Biosystems Inc.). The number and 128
area of the xylem cells in three radial files were manually quantified using the images obtained 129
from the microscope. Three plants were randomly selected from each of the stressed and 130
unstressed groups for this analysis. Statistical significance for change in the stressed plants 131
compared to unstressed plants was determined by comparing the average of the two groups 132
Page 6 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
7
through Student’s two-tailed t-test with a cutoff p-value of 0.05. Similarly, the stem cross-133
sections of 3-month old unstressed plants were analyzed and compared with those of the 2-month 134
old stressed plants. 135
136
Chlorophyll, anthocyanin, and lignin quantification 137
Stem tissue of 2-month-old koa seedlings was frozen in liquid nitrogen and ground using 138
a mortar and pestle for tissue homogenization. For every 0.1g of the stem tissue, 1.0 mL of EtOH 139
containing 1% HCl was used to extract anthocyanin. After mixing the tissue and the solution, 140
each sample was incubated at 4 ºC overnight and then centrifuged for 10 min at 20,000 rcf. 141
Absorbance of the supernatant was quantified at 529 nm and 650 nm using a UV-visible 142
spectrophotometer, UV-1600PC (VWR, PA, USA). The equation below was used to calculate 143
the corrected anthocyanin absorbance. 144
Anthocyanin absorbance = A529 – (0.228) (A650) (1) 145
The anthocyanin concentration was calculated using the Beer-Lambert expression, A = εcl, 146
where A is absorption, ε is a molar absorbance coefficient, c is concentration, and l is path length. 147
The molar absorbance coefficient ε of anthocyanin at 529 nm was assumed to be 30,000 L mol-1 148
cm-1 (Sims and Gamon 2002). Similarly, chlorophyll was extracted using 1.0 mL of 80% acetone 149
per 0.1 g of sample. Absorbance of the supernatant was quantified at 647 nm, and 664 nm, and 150
the following equations were used to calculate the concentrations of chlorophyll a, chlorophyll b, 151
and total chlorophyll (Porra 2002). For quantification of anthocyanins and chlorophylls, 15 152
individual plants were analyzed for each of the stressed and unstressed groups. 153
Page 7 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
8
Chlorophyll a (µg/mL) = 12.25 (A664) - 2.55 (A647) (2) 154
Chlorophyll b (µg/mL) = 20.31 (A647) - 4.91 (A664) (3) 155
Total chlorophyll (µg/mL) = 17.76 (A647) + 7.34 (A664) (4) 156
Stem lignin analysis was conducted by the Agricultural Diagnostic Services Center, University 157
of Hawaii at Manoa through an ANKOM 200 Automated Fiber Analyzer (ANKOM Technology, 158
Macedon, NY) as described by the manufacturer’s protocol for lignin quantification in Method 8 159
(https://www.ankom.com/). Three biological replicates were analyzed, and each replicate 160
represented a pool of 10 hypocotyls. For each analysis, statistical significance for change in the 161
stressed plants relative to unstressed plants was determined by comparing the average of the two 162
groups through Student’s two-tailed t-test with a cutoff p-value of 0.05. 163
164
Plant treatment and RNA extraction 165
Stems were harvested via flash-freezing in liquid nitrogen after 10, 30, and 60 min 166
following the mechanical stress treatment. To extract total RNA, the frozen stems were ground to 167
a fine powder in liquid nitrogen with a mortar and a pestle that were baked for 6 h at 300 °C 168
prior to use. Total RNA was extracted with the modified method using Qiagen RNeasy Plant Kit 169
(Valencia, CA, USA) and Fruit-mate (Takara, Japan) as described by Ishihara et al. (2016). RNA 170
was treated with TURBO DNA-free Kit (Ambion, CA, USA) to remove genomic DNA 171
contamination. The quantity and quality of the RNA were assessed at wavelengths of 230, 260, 172
and 280 nm using a NanoDrop ND-1000 (NanoDrop Technologies, DE, USA). The integrity of 173
RNA was confirmed with RNA Integrity Numbers (RIN) of ≥ 9 obtained through an Agilent 174
2100 Bioanalyzer (Agilent Technologies, CA, USA). 175
Page 8 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
9
176
Microarray analysis 177
Relative gene expression levels of the stressed plants relative to the unstressed control 178
were determined through a microarray analysis. An equal amount of extracted RNA of the A. koa 179
seedlings harvested at 10, 30, and 60 min following the mechanical stress treatment was pooled 180
for the analysis. The RNA extracted from the seedlings without the mechanical stress treatment 181
was used as the unstressed control. A 4x72K microarray analysis was performed by the Roy J. 182
Carver Center for Genomics at the University of Iowa to compare gene expression levels of 2 183
biological replicates of 30 stressed plants (10 plants/time point) and 2 biological replicates of 20 184
unstressed control plants. From the previous study of transcriptome analysis (Ishihara et al. 185
2015), 4,000 cDNA sequences longer than 360 bp were selected on the basis of similarities with 186
genes that might be related to plant growth and development. Six 60-bp probes per sequence 187
were designed by the Carver Center for Genomics; each probe had three replicates in a DNA 188
chip. The designs of probes and microarray were deposited into GEO (Record No. GSE73889). 189
The array was scanned using a NimbleGen MS 200 Microarray Scanner (Roche NimbleGen, 190
Inc.). The NimbleScan software v.2.6 (Roche NimbleGen, Inc.) extracted raw intensities from 191
the images generated by the scanner, which were corrected for background noise and normalized 192
between arrays using a Robust Multichip Average (RMA) algorithm included in the NimbleScan 193
software. The normalized probe intensity values were averaged to give a single intensity value 194
per transcript and per sample. Statistical significance for change in the stressed plants relative to 195
unstressed plants was determined by comparing the average of the two groups through Student’s 196
two-tailed t-test with a cutoff p-value of 0.05. 197
198
Page 9 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
10
Confirmation of microarray data by NanoString nCounter and qRT-PCR 199
To verify if the 57 sequences shown to be upregulated in the microarray analysis were 200
truly upregulated, the NanoString nCounter System, a digital multiplexed gene-expression 201
analysis, was used at NanoString Technologies, Seattle WA. Briefly, total RNA extracted (100 202
ng) from seedlings at 10, 30, and 60 min following the stress treatment and from seedlings 203
without the stress treatment was prepared. Three biological replicates of RNA from ten stressed 204
plants for each time point were compared with three biological replicates of RNA from ten 205
unstressed plants. RNA was directly hybridized with gene-specific color-coded probes, and data 206
collection was carried out in the nCounter Digital Analyzer as described by the manufacturer 207
(NanoString Technologies). The NanoString Codeset (Supplementary Table S1) was designed 208
and synthesized by Nanostring Technologies, which included 3 reference genes, eflα, β-actin, 209
and ubiquitin; six positive-control and eight negative-control probes were also added to each 210
reaction to normalize the results. The data analysis was performed through nSolver Analysis 211
Software (Nanostring Technologies) using the default settings. Statistical significance for 212
increase in gene expression levels in the stressed plants compared to unstressed plants was 213
determined using Student’s one-tailed t-test with a cutoff p-value of 0.05. 214
The nCounter results were further confirmed through quantitative real-time PCR (qRT-215
PCR) analysis of five genes. Total RNA extracted prior to the mechanical stress treatment and 216
after 10, 30, and 60 min following the mechanical stress treatment was treated with TURBO 217
DNAfree Kit (Invitrogen, Carlsbad, CA) to remove any genomic DNA contamination. First-218
strand cDNA was synthesized from 2 µg of DNase-treated RNA using M-MLV Reverse 219
Transcriptase (Promega, WI, USA) with random hexamers according to the manufacturer’s 220
instructions. The qRT-PCR was performed using a 10 µL PCR reaction consisting of 0.25 µL 221
Page 10 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
11
forward primer (10 µM), 0.25 µL reverse primer (10 µM), 5 µL iQTM SYBR® Green Supermix 222
(Bio-Rad Laboratories, Hercules, CA), and 1 µL of first strand cDNA. Reaction conditions were 223
95 °C for 3 min, 35-40 cycles of 95 °C for 15 s, annealing for 30 s, and 72 °C for 30 s, followed 224
by melting curve analysis of the amplicon to confirm the specificities of primers. The annealing 225
temperature was at 54 °C for all the tested genes, except that 58 °C was used for the RING gene. 226
Each assay consisted of three biological and three technical replicates and was performed using 227
StepOne Real-Time PCR System (Applied Biosystems, Foster City, CA). The PCR protocol 228
produced a PCR efficiency of 90% to 110% for each primer set. The primer sequences used for 229
this study are listed in Supplementary Table S2. 230
To select internal reference genes for relative quantification of target gene expressions, 231
six candidate reference genes: ef1α, β-actin, 18S rRNA, 5.8S rRNA, ubiquitin-5, and tubulin-1 232
were tested on first strand cDNA samples from the A. koa stem with the primer sequences as 233
described in Negi et al. (2011). The qRT-PCR protocol was performed as described previously, 234
with the annealing temperature at 58 °C. The cycle threshold (Ct) values of the candidate genes 235
were used to evaluate their expression stability by using NormFinder applet for MS Excel 236
(Andersen et al. 2004). NormFinder allows the user to determine intra- and inter-group variances 237
as well as the stability value of each candidate gene. One gene or a combination of two genes 238
with the lowest stability value were selected as a reference control to normalize the expression 239
levels at each time point: a combination of two genes, 18S and ef1α, for the time point at 10 min 240
following the treatment, and 18S rRNA for the time points at 30 and 60 min following the 241
treatment (Supplementary Table S3). The fold changes of target gene expression levels relative 242
to the unstressed control plants were determined using the selected reference genes for 243
normalization through the 2-∆∆Ct method (Livak and Schmittgen 2001). Statistical significance for 244
Page 11 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
12
increase in gene expression levels in the stressed plants compared to unstressed plants was 245
determined using Student’s one-tailed t-test with a cutoff p-value of 0.05. 246
247
Page 12 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
13
RESULTS 248
Mechanical stress alters plant morphology by reducing stem elongation and increasing 249
stem diameter in A. koa 250
Following two months of the daily mechanical perturbation, the stressed A. koa seedlings 251
showed a decrease in stem elongation by 32.2% and an increase in hypocotyl diameter by 17.8% 252
(Fig. 1A, C, D). The mechanical stress also decreased the size of the leaflets by 19.8% and the 253
dry weight of root by 44.6% (Fig. 1E, F). After six months of the treatment, plants showed 254
greater developmental changes compared to the plants that were stressed for two months, except 255
for the root dry weight. The stressed plants were stunted by 60.9% and had a significantly 256
increased hypocotyl diameter by 23.6% (Fig. 1B, D, C). The leaflet size decreased approximately 257
by 32.2% (Fig. 1E). Interestingly, the root dry weight decreased by 27.2% (Fig. 1F), but the 258
percentage in decrease was greater after two months (44.6%) than after 6 months. There was no 259
significant difference in dry weight of hypocotyls in spite of the difference in height (data not 260
shown). 261
262
Mechanical stress increases anthocyanin and lignin but not chlorophyll 263
Mechanical stress increased the anthocyanin content in stems of the stressed seedlings by 264
91.0% following 2 months of the treatment (Fig. 2B). It was visually observable as well (Fig. 265
2A); the stressed seedlings exhibited anthocyanin accumulation in their stems with reddish color. 266
The lignin composition in the stressed stems also increased by 32.3% (Fig. 2C). There was no 267
significant difference in chlorophyll concentration (data not shown). 268
269
270
Page 13 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
14
271
Mechanical stress promotes xylem cell division 272
Microscopic analysis of hypocotyl cross-sections showed an increase in the density of 273
xylem cells (number of cells/10,000 µm2) by 45.7% in the stressed seedlings compared to 274
unstressed of the same age (2-month old; Fig. 3A, B, and E). The increase in cell density was due 275
to the reduction in the cell size, as the average size of a xylem cell was smaller in stressed plants 276
by 32.4% (Fig. 3D). Regardless of the increase in the cell density following the mechanical stress, 277
the ratio between the radii of the stem and the xylem did not change (data not shown). To 278
determine if xylem cell size was correlated with stem diameter, stem cross-sections of 2-month-279
old stressed seedlings were also compared with those of 3-month-old unstressed seedlings with 280
similar stem diameters; the xylem cell density of 2-month-old stressed plants was 66.3% higher 281
than 3-month-old unstressed plants whereas the cell size of the 2-month-old stressed plants was 282
lower by 40.5% than 3-month-old unstressed plants (Supplementary Table S4). The results 283
showed no correlation between the stem diameter and the xylem cell size, and 284
thigmomorphogenetic plants had higher cell density with smaller cells, while unstressed plants 285
had lower cell density with bigger cells. 286
287
Genes induced by mechanical stress 288
The microarray analysis indicated that out of the 4,000 cDNA sequences, 60 sequences 289
were upregulated more than twofold in the mechanically stressed plants within 60 min following 290
the treatment (Fig. 4). We grouped those 60 sequences by their putative functions; 28 were 291
classified in transcriptional regulation, 14 in signal transduction, 5 in carbohydrate metabolism, 4 292
in protein degradation, 3 in disease resistance, and 2 in hormone metabolism (Fig. 5). Some of 293
Page 14 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
15
the most upregulated genes are those that have homology with sequences encoding RING finger 294
proteins (>18-fold), zinc-finger transcription factors (>15-fold), ethylene-responsive-element 295
binding proteins (EREBPs; >9-fold), WRKY transcription factors (>8-fold), protein phosphatase 296
2C (>8-fold), dehydration responsive element binding (DREB) proteins, and disease resistance 297
proteins (>4-fold; Supplementary Table S5). 298
The Nanostring nCounter analysis verified upregulation of 53 sequences out of the 60, 299
which was further confirmed by qRT-PCR (Table 1; Supplementary Fig. S1). The expression 300
levels of most of the sequences were low (~2-fold) at 10 min following the stress treatment but 301
peaked at 30 min. Two sequences that have homology with sequences encoding a RING finger 302
protein or a zinc finger protein (Acc. No. GBYE01027655 and GBYE01019260, respectively) 303
had the highest fold-change of ~11 at 10 min, showing a rapid response of the plant to the 304
mechanical stimulus. The sequence encoding the putative RING finger protein had the highest 305
expression of 38.1-fold at 30 min after the treatment, whereas the sequence encoding the putative 306
zinc finger protein was upregulated 45-fold within 30 min. The most represented putative 307
function among the 53 upregulated sequences was ethylene-related transcription factors (9 308
sequences homologous to AP2/ERF, DREB, and EREBPs), followed by WRKY transcription 309
factors (6 sequences), and calcium-binding proteins (5 sequences). Three sequences homologous 310
to sequences encoding disease-resistance proteins were also confirmed to be induced by 311
mechanical stress, and they were upregulated more than 10 times at 30 min following the stress 312
treatment. 313
314
DISCUSSION 315
Page 15 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
16
Acacia koa inhabits a wide range of areas at elevations from the sea level to 2,000 m, in 316
wet or mesic forests with annual rainfall from 1,850 to 5,000 mm (Harrington et al. 1995; 317
Wilkinson and Elevitch 2005; Baker et al. 2009). A. koa trees show various sizes and shapes in 318
different geological locations, and it has been reported that wood properties of A. koa are also 319
influenced by environmental factors to a large extent (Loudat and Kanter 1996; Dudley and 320
Yamasaki 2000). Mechanical stimuli, such as rain and wind, are common environmental factors 321
that affect growth and development in plants. It is especially important to understand the effects 322
of wind on A. koa because in areas with steady winds, A. koa plants are reported to have 323
relatively slower growth (Elevitch et al. 2006). In this study, physical stress similar to that 324
produced by wind was artificially created in A. koa seedlings through bending stems in four 325
cardinal directions to observe morphological, physiological, and biochemical effects of such 326
mechanical stress. 327
328
Physical adaptation of A. koa to mechanical stress 329
Non-wounding swaying of stems in four cardinal directions had a profound effect on the 330
morphology of A. koa (Fig. 1), making it capable to withstand further mechanical stress. The 331
reduced height and increased stem diameter after the stress treatment resembled the 332
thigmomorphogenetic changes previously observed with other woody plants, such as Pinus taeda 333
(Telewski and Jaffe 1986a) and Ulmus americana (Telewski and Pruyn 1998), and other 334
dicotyledonous plants, including, Solanum lycopersicum (Coutand et al. 2000), and Nicotiana 335
tabacum (Anten et al. 2005). Interestingly, little to no overall changes have been reported in 336
some plants, in which the cambium is limited or absent; for example, there were a little change in 337
inflorescence stem diameter of Arabidopsis and no change in stem diameter of Helianthus annus 338
Page 16 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
17
following mechanical stress (Smith and Ennos 2003; Paul-Victor and Rowe 2011). Therefore, 339
elevated cambial activity may be a major factor that increases the overall stem diameter (Biro et 340
al. 1980; Jacobs 1953; Pruyn et al. 2000; Telewski and Jaffe 1986a, 1986b); as such, increase in 341
xylem cells, which are derived from the cambium (Lev-Yadun and Flaishman 2001), was 342
observed in several mechanically perturbed plants, such as Phaseolus vulgaris (Biro et al. 1980) 343
and N. tabacum (Hepworth et al. 1999). Similarly, in the stressed A. koa, there was an increase in 344
number of xylem cells (Fig. 3A, B); however, the ratio of diameters between xylem and stem and 345
between pith and stem showed no significant difference between the stressed and unstressed A. 346
koa (data not shown). Therefore, expansion of xylem and also other tissues appeared to 347
contribute to the increase in overall stem thickness. In addition, the stressed A. koa had denser 348
xylems with smaller cells (Fig. 3D), and similar xylem characteristics were observed in 349
mechanically stressed Abies fraseri (Telewski 1989). Therefore, thigmomorphogenetic responses 350
in A. koa may include the enhancement of secondary growth and modification of wood 351
properties; further analysis of responses to mechanical stimuli in A. koa may reveal unique 352
genetic and physiological responses that will help study wood formation and development. 353
354
Increase in phenylpropanoid biosynthesis in stems of thigmomorphogenetic A. koa 355
The significant elevation of lignin and anthocyanin contents in the stressed A. koa 356
indicated that thigmomorphogenesis affected the phenylpropanoid biosynthesis pathway, which 357
is responsible for synthesis of a wide variety of secondary metabolites, including monolignols 358
and flavonoids (Mayer et al. 2001; Fig. 2). Monolignols are monomers of lignin, which is an 359
important component of wood, constituting 25-35% of the secondary cell wall (Plomion et al. 360
2001); the stressed A. koa should have higher deposit of lignin and therefore may have thicker 361
Page 17 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
18
cell walls and stronger structural support (for review, see Vanholme et al. 2010) so that they 362
would have higher resistance against the swaying movement. In Bryonia dioica and S. 363
lycoperisicum, increased lignin and elevated activities of enzymes involved in lignification, such 364
as phenylalanine ammonia-lyase (PAL) and cinnamyl alcohol dehydrogenase (CAD), have been 365
observed 6 h following the mechanical stimulus (Saidi et al. 2009; de Jaegher et al. 1985). 366
However, the genes for enzymes involved in the monolignol biosynthesis, including PAL and 367
CAD, were not induced in A. koa within 60 min following the mechanical stress. Since we did 368
not determine the expression levels of the genes beyond 60 min, further studies in future will be 369
needed to elucidate effects of mechanical stress treatment on the transcription of the 370
phenylpropanoid pathway genes in A. koa. 371
We observed elevated concentration of anthocyanins, a type of flavonoids, in the stressed 372
A. koa; such increases in anthocyanin levels due to the mechanical stress have not been reported 373
in other studies of thigmomorphogenesis. On the contrary, decrease in anthocyanin content has 374
been observed in thigmomorphogenetic C. papaya (Porter et al. 2009). It shows that molecular 375
responses to mechanical stress vary among species as anthocyanin production was repressed in 376
the stressed C. papaya, while it was enhanced in the stressed A. koa. Anthocyanins and other 377
flavonoids are derived from p-coumaroyl CoA, a major pivotal intermediate of the 378
phenylpropanoid pathway, which is also an intermediate product for biosynthesis of monolignols 379
and other phenylpropanoids. It is the substrate for the monolignol biosynthesis enzymes, 380
including hydroxycinnamoyl-CoA shikimate/quinate hydroxycinnamoyl transferase (HCT) and 381
caffeoyl-CoA O-methyltransferase (CCoAOMT), as well as for the flavonoid biosynthesis 382
enzyme chalcone synthase (CHS). Therefore, it is likely that p-coumaroyl CoA accumulated in 383
response to the mechanical stress, feeding both pathways of monolignol and flavonoid 384
Page 18 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
19
biosynthesis. Lignin production is shown to be induced by mechanical perturbations in several 385
other plants (de Jaegher et al. 1985; Bourgeade et al. 1989; Porter et al. 2009) while anthocyanin 386
production is not; therefore, the anthocyanins in the stressed A. koa may be inadvertently 387
produced due to enhancement of the lignin biosynthesis pathway. It is thus possible that 388
thigmomorphogenetic A. koa produced other phenylpropanoids, such as coumarins, tannins, and 389
other aromatic compounds, from p-coumaryol CoA, (VanEtten et al. 1989; Vogt 2010). 390
The thigmomorphogenetic plants produced in this research can be considered as 391
phenotypic mutants for biosynthesis of lignin, its precursor phenolic compounds, and 392
anthocyanins. Many of the phenolic compounds, including lignin, are known to be involved in 393
plant defense mechanisms in various plants (Uppalapati et al. 2009; Kamphuis 2012; Purwar et 394
al. 2012; Boubakri et al. 2013). Therefore, besides being more tolerant to mechanical stress, the 395
thigmomorphogenetic A. koa plants may also grow more tolerant to other stresses, including 396
resistance to pathogens and insects, compared to the unstressed A. koa plants; further analysis of 397
the mechanically stressed plants by exposing them to pathogens will verify if mechanical stress 398
induces disease resistance in plants. We may be able to relate this mutant phenotype with aspects 399
of wood quality in future studies. Lignin is a major component of the cell wall, and its 400
composition is one of the major determinants of physical characteristics of wood (Novaes et al., 401
2010). Wood stiffness and strength have been shown to be related to the lignin composition 402
(Voelker et al. 2011). Woods of transgenic poplars containing a higher syringyl lignin content 403
have been shown to be resistant to decay by wood-decaying fungi (Skyba et al. 2013); therefore, 404
we can expect that A. koa wood having higher lignin content may also be resistant to decay by 405
such fungi. In addition to lignin, anthocyanins may be a determinant of wood characteristics. 406
Anthocyanins are known to accumulate in the vacuole (Evert 2006; Zhao et al. 2006). It is likely 407
Page 19 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
20
that they are converted into anthocyanidins through removal of the sugar group by vacuole 408
glycosidase under certain physiological conditions. Anthocyanidins, which are also stored in the 409
vacuole, are important because they are oligomers of proanthocyanidins (condensed tannins; 410
Zhao et al. 2006); proanthocyanidins are known to be involved in wood color and defense 411
mechanisms in plants (Hon and Minemura 2000; War et al. 2012; Constabel et al. 2014). Thus, 412
lignin and anthocyanins may be used as biochemical markers for seedling selection in both 413
disease resistance and wood quality improvement programs for A. koa as well as for other timer-414
wood tree species. 415
Regulation of chlorophyll biosynthesis in leaves or total tissues in thigmomorphogenetic 416
plants has been reported. For instance, chlorophyll content went up in Medicago truncatula, 417
Apium graveolens, Lactuca sativa, and S. lycopersicum (Biddington and Dearman 1985; Mitchell 418
et al. 1975; Tretner et al. 2008) whereas it decreased in B. oleracea and C. papaya (Biddington 419
and Dearman 1985; Porter et al. 2009). On the contrary, in the present study, chlorophyll content 420
in the stem, including petioles, did not show any significant difference between the mechanically 421
stressed and unstressed A. koa. Chlorophyll content of the leaves was not determined in this 422
study. 423
424
Calcium signaling in thigmomorphogenesis of A. koa 425
Calcium (Ca2+) is one of the major signaling molecules released from the endoplasmic 426
reticulum to the cytosol in response to mechanical perturbations (Braam 2005; Telewski 2006; 427
Chehab et al. 2009). Studies have shown upregulation of genes for calmodulins and calmodulin-428
related proteins in various plants, including Arabidopsis, C. papaya, Brassica napus, and Vigna 429
radiata (Braam and Davis 1990; Botella and Arteca 1994; Botella et al. 1996; Oh et al. 1996; 430
Page 20 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
21
Lee et al. 2005; Porter et al. 2009). The gene for calmodulin was one of the first identified touch-431
inducible (TCH) genes (TCH1; Acc. No. AAA32762.1; Braam and Davis 1990) in Arabidopsis, 432
and its homolog in A. koa (Acc. No. GBYE01000487; 97.6% sequence identity) was among the 433
5 sequences coding for putative calcium-binding proteins that are upregulated in response to 434
mechanical stress (Table 1). Moreover, in stressed A. koa, two putative mitogen-activated protein 435
kinase (MAPK) sequences were upregulated more than 20-fold within 30 min following the 436
stress treatment. MAPK sequences are known to be indirectly or directly regulated by Ca2+ in 437
plants (Wurzinger et al. 2011). The increased expression levels of genes encoding MAPK and 438
other protein kinases, possibly calcium-dependent, following mechanical stimulation have been 439
observed in other plants (Seo et al. 1995; Mizoguchi et al. 1996; Bögre et al. 1997). Therefore, it 440
is possible that calcium signaling may induce MAPK cascades in the stressed A. koa as well. 441
442
Regulation of phytohormones in thigmomorphogenesis in mechanically stressed A. koa 443
Ethylene, a plant growth hormone, appears to be involved in early thigmomorphogenic 444
responses in A. koa. A sequence for putative 1-aminocyclopropane 1-carboxylate synthase 445
(ACS), a key enzyme for ethylene production, was rapidly induced within 10 min following the 446
stress (Table 1). In addition, the expression of nice sequences for putative ethylene-responsive 447
transcription factors was also enhanced. In other plants, ethylene production is shown to be one 448
of the early thigmomorphogenetic responses (de Jaegher et al. 1987; Botella et al. 1995; Arteca 449
and Arteca 1999; Tatsuki and Mori 1999). Increase in the activity of ACS and upregulation of an 450
ethylene-responsive transcription factor were also observed rapidly following mechanical 451
stimulation in Phaseolus vulgaris and Arabidopsis (Biro and Jaffe 1984; Lee et al. 2005). 452
Subsequent to exogenous application of ethylene, stunted growth was observed in various plants, 453
Page 21 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
22
resembling thigmomorphogenic responses (Jaffe and Biro 1979; de Jaegher et al. 1987; Erner 454
and Jaffe 1982). However, ethylene-insensitive Arabidopsis mutants still showed 455
thigmomorphogenetic changes following mechanical stimulation, so ethylene is unlikely to be 456
the ‘primary’ signaling molecule (Johnson et al. 1998), at least in Arabidopsis. 457
Another growth phytohormone, abscisic acid (ABA) may be involved in 458
thigmomorphogenesis in A. koa. Many plants, such as H. annuus and Zea mays, had increased 459
ABA levels following the mechanical stimulation (Whitehead and Luti 1962; Beyl and Mitchell 460
1983), and exogenous application of ABA also led to morphological changes similar to 461
thigmomorphogenetic responses (Erner and Jaffe 1982). On the contrary, a sequence for a key 462
enzyme for ABA degradation, ABA 8’-hydroxylase, was upregulated in the stressed A. koa (> 463
35.8-fold within 60 min following the stress); thus, the induction of the gene would result in 464
reduction of ABA levels in thigmomorphogenetic A. koa. It may be possible that ABA synthesis 465
was downregulated due to feedback inhibition. Different plants respond to mechanical stress 466
differently; further analyses on phytohormones and their quantification in the stressed and 467
unstressed A. koa may elucidate their roles in thigmomorphogenesis. 468
469
Abiotic- and biotic-stress-related genes induced by mechanical stress in A. koa 470
The gene expression analyses suggested that thigmomorphogenetic responses in A. koa 471
might influence a wide range of cellular mechanisms, such as transcriptional regulation, signal 472
transduction, carbohydrate metabolism, and hormone metabolism. Interestingly, many of them 473
are related to abiotic and biotic stresses as well as plant growth and development. Among the 474
upregulated 23 sequences coding for transcription factors, 15 sequences encode putative WRKYs, 475
MYBs, NACs, DREBs, and zinc finger proteins, many of which are known to be involved in 476
Page 22 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
23
disease resistance, defense, and abiotic stress tolerance, such as drought and high-salinity 477
stresses (Lata and Prasad 2011; Nuruzzaman et al. 2013; Ambawat et al. 2013; Bakshi and 478
Oelmüller 2014). Among the transcription factors, two sequences for zinc finger proteins were 479
highly upregulated within 10 min after the treatment, and they were found to be homologous to a 480
ZAT-10 protein, which is expressed in response to drought, cold, salt, and photo-oxidative 481
stresses in Arabidopsis (Mittler et al. 2006; Rossel et al. 2007). Among the 57 tested sequences 482
in the nCounter analysis, it is one of the most rapidly upregulated genes in response to the 483
mechanical stress. The upregulation of these abiotic stress-related transcription factors suggests 484
that the mechanically stressed A. koa may also develop higher tolerance to other environmental 485
stresses. 486
It is notable that sequences for disease resistance (R) proteins, or nucleotide-binding site 487
leucine-rich repeat (NBS-LRR) proteins, were induced by mechanical perturbations in A. koa. R 488
proteins are involved in disease resistance as they recognize invading pathogens by directly 489
binding to or indirectly monitoring effector molecules secreted by pathogens (McHale et al. 490
2006). Although the mechanically stressed A. koa were not exposed to pathogens, three 491
sequences encoding R proteins were upregulated over 10-fold within 30 min after the stress 492
treatment (Table 1). Similarly, the microarray analysis of the mechanically inducible genes in 493
Arabidopsis showed upregulation of 14 R proteins (> 2-fold; Lee et al. 2005). In addition to R 494
proteins, ATL family RING finger ubiquitin ligase is shown to regulate defense mechanisms and 495
cell death regulations in response to pathotoxins (Guzmán 2012). One homolog of ATL family 496
RING finger ubiquitin ligases was highly upregulated in response to the mechanical stress in A. 497
koa, increasing by over 10-fold within 10 min and over 38-fold within 30 min following the 498
mechanical stress (Table 1). Higher expression levels of the R proteins and ATL family RING 499
Page 23 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
24
finger proteins in the stressed A. koa indicates that mechanical stimulation may also induce 500
tolerance to pathogens. 501
502
Conclusion 503
In this study, morphological, biochemical changes and levels of gene transcription due to 504
mechanical stress were determined in A. koa. Prolonged mechanical stimulation led to stunted 505
stem length and thickened stem diameter with a greater number of xylem cells. Anthocyanin and 506
lignin were accumulated in the stems of mechanically stressed A. koa, indicating that mechanical 507
stress induces the phenylpropanoid biosynthesis pathway. The gene expression analyses showed 508
upregulation of genes in a wide range of cellular mechanisms in response to mechanical stress, 509
suggesting complex signaling pathways of thigmomorphogenesis. Genes for calcium signaling, 510
such as calmodulin and MAPK, ethylene and ABA metabolisms, and stress-related 511
transcriptional regulation were highly expressed. Two of the most rapidly upregulated genes 512
encoding a putative RING finger protein and a zinc finger protein may be related to both abiotic 513
and biotic stresses, such as drought and disease resistance. Surprisingly and interestingly, genes 514
for disease resistance, which are generally induced as a result of biotic stress, were also induced 515
by mechanical stress alone. This result suggests that trees may develop disease resistance in 516
response to mechanical stress applied by strong wind and heavy rain in natural forests. It also 517
suggests that a simple mechanical stress may be used to identify genetic and biochemical factors 518
that influence disease resistance; these molecules may be useful as potential markers for seedling 519
selection in A. koa improvement programs. 520
521
ACKNOWLEDGEMENTS 522
Page 24 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
25
We would like to acknowledge Tyler Jones and Nick Dudley from HARC for providing 523
A. koa seeds. We would also like to acknowledge Bradley Porter for his useful discussion. This 524
work was supported by the USDA National Institute of Food and Agriculture, the McIntire-525
Stennis Grant HAW00597-M, managed by the College of Tropical Agriculture and Human 526
Resources (CTAHR). KI was supported by a Monsanto Graduate Fellowship. 527
Page 25 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
26
REFERENCES
Ambawat, S., Sharma, P., Yadav, N.R., and Yadav, R.C. 2013. MYB transcription factor genes as regulators for plant responses: An overview. Physiol Mol Biol Plants. 19(3), 307–321. doi:10.1007/s12298-013-0179-1.
Andersen, C.L., Jensen, J.L., and Ørntoft, T.F. 2004. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 64(15): 5245–5250. doi:10.1158/0008-5472.CAN-04-0496.
Anten, N.P.R., Casado-Garcia, R., and Nagashima, H. 2005. Effects of mechanical stress and plant density on mechanical characteristics, growth, and lifetime reproduction of tobacco plants. Am. Nat. 166(6): 650–660. doi:10.1086/497442.
Arteca, J.M., and Arteca, R.N. 1999. A multi-responsive gene encoding 1-aminocyclopropane-1-carboxylate synthase (ACS6) in mature Arabidopsis leaves. Plant Mol. Biol. 39(2): 209–219. doi:10.1023/A:1006177902093.
Baker, P.J., Scowcroft, P.G., and Ewel, J.J. 2009. Koa (Acacia koa) ecology and silviculture. USDA Forest Service, General Technical Report - Pacific Southwest Research Station, PSW-GTR-211.
Bakshi, M., and Oelmüller, R. 2014. WRKY transcription factors: Jack of many trades in plants. Plant Signal. Behav. 9(1): 1–18. doi:10.4161/psb.27700.
Beyl, C.A., and Mitchell, C.A. 1977. Automated mechanical stress application for height control of greenhouse chrysanthemums. Hort. Scient. 12: 575-577.
Biddington, N.L., and Dearman, A.S. 1985. The effect of mechanically induced stress on the growth of cauliflower, lettuce and celery seedlings. Ann. Bot. 55(1): 109–119.
Biro, R., and Jaffe, M. 1984. Thigmomorphogenesis: ethylene evolution and its role in the changes observed in mechanically perturbed bean plants. Physiol. Plant. 62(3): 289–296. doi:10.1111/j.1399-3054.1984.tb04575.x.
Biro, R., Hunt, E.R., Erner, Y., and Jaffe, M.J. 1980. Thigmomorphogenesis: changes in cell division and elongation in the internodes of mechanically-perturbed or ethrel-treated bean plants. Ann. Bot. 45(6): 655-664.
Bögre, L., Ligterink, W., Meskiene, I., Barker, P., Heberle-Bors, E., Huskisson, N., and Hirt, H. 1997. Wounding induces the rapid and transient activation of a specific MAP kinase pathway. Plant Cell 9(1): 75–83. doi:10.1105/tpc.9.1.75.
Botella, J.R., and Arteca, R.N. 1994. Differential expression of two calmodulin genes in response to physical and chemical stimuli. Plant Mol. Biol. 24(5): 757–766. doi: 10.1007/BF00029857.
Botella, J.R., Arteca, R.N., and Frangos, J.A. 1995. A mechanical strain-induced 1-aminocyclopropane-1-carboxylic acid synthase gene. Proc. Natl. Acad. Sci. USA. 92(5): 1595–1598. doi:10.1073/pnas.92.5.1595.
Botella, J.R., Arteca, J.M., Somodevilla, M., and Arteca, R.N. 1996. Calcium-dependent protein kinase gene expression in response to physical and chemical stimuli in mungbean (Vigna
radiata). Plant Mol. Biol. 30(6): 1129–1137. doi:10.1007/BF00019547. Boubakri, H., Poutaraud, A., Wahab, M.A., Clayeux, C., Baltenweck-Guyot, R., Steyer, D.,
Marcic, C., Mliki, A., and Soustre-Gacougnolle, I. 2013. Thiamine modulates metabolism of the phenylpropanoid pathway leading to enhanced resistance to Plasmopara viticola in grapevine. BMC Plant Biol. 13(1): 31. doi:10.1186/1471-2229-13-31.
Page 26 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
27
Bourgeade, P., Boyer, N., de Jaegher, G., and Gaspar, T. 1989. Carry-over of thigmomorphogenetic characteristics in calli derived from Bryonia dioicainternodes. Plant Cell Tiss. Organ Cult. 19(3): 199–211. doi:10.1007/BF00043347
Braam, J., and Davis, R.W. 1990. Rain-, wind-, and touch-induced expression of calmodulin and calmodulin-related genes in Arabidopsis. Cell 60(3): 357–364. doi:10.1016/0092-8674(90)90587-5.
Braam, J. 2005. In touch: plant responses to mechanical stimuli. New Phytol. 165(2): 373–89. doi:10.1111/j.1469-8137.2004.01263.x.
Chehab, E.W., Eich, E., and Braam, J. 2009. Thigmomorphogenesis: a complex plant response to mechano-stimulation. J. Exp. Bot. 60(1): 43–56. doi:10.1093/jxb/ern315.
Constabel, C.P., Yoshida, K., Walker, V. 2014. Diverse Ecological Roles of Plant Tannins: Plant Defense and Beyond. In: (Romani, A., Lattanzio, V., QuideauS., eds.) Recent Advances in Polyphenol Research, Volume 4. John Wiley & Sons, Ltd, Chichester, UK. doi: 10.1002/9781118329634.ch5.
Coutand, C., Julien, J.L., Moulia, B., Mauget, J.C., and Guitard, D. 2000. Biomechanical study of the effect of a controlled bending on tomato stem elongation: global mechanical analysis. J. Exp. Bot. 51(352): 1813–1824. doi:10.1093/jexbot/51.352.1813.
de Jaegher, G., Boyer, N., and Gaspar, T. 1985. Thigmomorphogenesis in Bryonia dioica: changes in soluble and wall peroxidases, phenylalanine ammonia-lyase activity, cellulose, lignin content and monomeric constituents. Plant Growth Regul. 3(2): 133-148. doi:10.1007/BF01806053
de Jaegher, G., Boyer, N., Bon, M.C., and Gaspar, T. 1987. Thigmomorphogenesis in Bryonia
dioica: early events in ethylene biosynthesis pathway. Biochem Physiol Pflanz 182(1): 49–56. doi:10.1016/S0015-3796(87)80037-9.
Dudley, N.S., and Yamasaki, J. 2000. A Guide to Determining Wood Properties of Acacia koa. Hawaii Agriculture Research Center, Forestry Report 3.
Elevitch, C.R., Wilkinson, K.M., and Friday, J.B. 2006. Acacia koa (koa) and Acacia koaia (koai‘a). In Species Profiles for Pacific Island Agroforestry. Edited by C.R. Elevitch. Permanent Agriculture Resources, Holualoa, HI. Available from www.traditionaltree.org [accessed 7 April 2013].
Erner, Y., and Jaffe, M.J. 1982. Thigmomorphogenesis: the involvement of auxin and abscisic acid in growth retardation due to mechanical perturbation. Plant Cell Physiol. 23(6): 935–941.
Guzmán, P. 2012. The prolific ATL family of RING-H2 ubiquitin ligases. Plant Signal. Behav. 7(8): 1014–21. doi:10.4161/psb.20851.
Evert, R.F. 2006. The Protoplast: Endomembrane System, Secretory Pathways, Cytoskeleton, and Stored Compounds. In: Esau's Plant Anatomy: Meristems, Cells, and Tissues of the Plant Body: Their Structure, Function, and Development, Third Edition. John Wiley & Sons, Inc., Hoboken, NJ, USA. doi: 10.1002/0470047380.ch3.
Harrington, R.A., Fownes, J.H., Meinzer, F.C., and Scowcroft, P.G. 1995. Forest growth along a rainfall gradient in Hawaii: Acacia koa stand structure, productivity, foliar nutrients, and water- and nutrient-use efficiencies. Oecologia 102(3): 277–284. doi:10.1007/BF00329794.
Hepworth, D., and Vincent, J. 1999. The growth response of the stems of genetically modified tobacco plants (Nicotiana tabacum “Samsun”) to flexural stimulation. Ann. Bot. 83(1): 39–43. doi:10.1006/anbo.1998.0784.
Page 27 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
28
Hon, D.N.S., Minemura, N. 2000. Color and Discoloration. In: (Hon, D.N.S., Minemura, N., eds.) Wood and cellulosic chemistry, second edition, revised and extended. Marcel Dekker Inc., New York, NY. pp. 385-442.
Ishihara, K., Lee, E.K.W., Rushanaedy, I., and Borthakur, D. 2015. Illumina-based de novo transcriptome analysis and identifications of genes involved in the monolignol biosynthesis pathway in Acacia koa. Am. J. Bioinforma. 4(1): 7–27. doi:10.3844/ajbsp.2015.7.27.
Ishihara, K., Lee, E.K.W., and Borthakur, D. 2016. An improved method for RNA extraction from woody legume species Acacia koa A. Gray and Leucaena leucocephala (Lam.) de Wit. Int. J. Forestry Wood Sci. 3(1): 31-35.
Jacobs, M.R. 1954. The effect of wind sway on the form and development of Pinus radiata. Austral J. of Bot. 2(1): 35-51.
Jaffe, M.J., and Biro, R. 1979. Thigmomorphogenesis: the effect of mechanical perturbation on the growth of plants, with special reference to anatomical changes, the role of ethylene, and interaction with other environmental stresses. In Stress physiology of crop plants. Edited by H Mussel and R.C. Staples. John Wiley and Sons, New York, NY. pp. 25–29.
Jaffe, M.J. 1973. Thigmomorphogenesis: The response of plant growth and development to mechanical stimulation. Planta. 114(2): 143–157. doi: 10.1007/BF00387472.
Johnson, K.A., Sistrunk, M.L., Polisensky, D.H., and Braam, J. 1998. Arabidopsis thaliana responses to mechanical stimulation do not require ETR1 or EIN2. Plant Physiol. 116(2): 643–9.
Kamphuis, L.G., Williams, A.H., Küster, H., Trengove, R.D., Singh, K.B., Oliver, R.P., and Ellwood, S.R. 2012. Phoma medicaginis stimulates the induction of the octadecanoid and phenylpropanoid pathways in Medicago truncatula. Mol. Plant Pathol. 13(6): 593–603. doi:10.1111/j.1364-3703.2011.00767.x.
Koller, D. 2011. The restless plant. Edited by E.V. Volkenburgh. Harvard University Press, Cambridge, MA.
Lata, C., and Prasad, M. 2011. Role of DREBs in regulation of abiotic stress responses in plants. J. Exp. Bot. 62(14): 4731-48. doi:10.1093/jxb/err210.
Lee, D., Polisensky, D.H., and Braam, J. 2005. Genome-wide identification of touch- and darkness-regulated Arabidopsis genes: a focus on calmodulin-like and XTH genes. New Phytol. 165(2): 429–44. doi:10.1111/j.1469-8137.2004.01238.x.
Lev-Yadun, S., and Flaishman, M.A. 2001. The effect of submergence on ontogeny of cambium and secondary xylem and on fiber lignification in inflorescence stems of Arabidopsis. IAWA J. 22(2): 159–169. doi:10.1163/22941932-90000275
Livak, K.J., and Schmittgen, T.D. 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25(4): 402–408. doi:10.1006/meth.2001.1262.
Loudat, T.A., and Kanter, R. 1996. The economics of commercial koa culture in Hawaii. In Koa: A Decade of Growth Proceedings of the Symposium, Honolulu, HI, 18-19 November 1996. Edited by L. Ferentinos and D. Evans. Hawaii Forest Industry Association, Honolulu, HI. pp. 124–147.
Mayer, M.J., Narbad, A., Parr, A.J., Parker, M.L., Walton, N.J., Mellon, F.A., and Michael, A.J. 2001. Rerouting the plant phenylpropanoid pathway by expression of a novel bacterial enoyl-CoA hydratase/lyase enzyme function. Plant Cell 13(7): 1669–1682. doi:10.1105/TPC.010063.
McHale, L., Tan, X., Koehl, P., and Michelmore, R.W. 2006. Plant NBS-LRR proteins: adaptable guards. Genome Biol. 7: 212. doi:10.1186/gb-2006-7-4-212.
Page 28 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
29
Mitchell, C.A., Severson C.J., Wott J.A., and Hammer P.A. 1975. Seismomorphogenic regulation of plant growth. J. Amer. Soc. Hort. Sci. 100: 161–165.
Mittler, R., Kim, Y., Song, L., Coutu, J., Coutu, A., Ciftci-Yilmaz, S., Lee, H., Stevenson, B., and Zhu, J.K. 2006. Gain- and loss-of-function mutations in Zat10 enhance the tolerance of plants to abiotic stress. FEBS Lett. 580(28-29): 6537–6542. doi:10.1016/j.febslet.2006.11.002.
Mizoguchi, T., Irie K., Hirayama, T., Hayashida, N., Yamaguchi-Shinozaki, K., Matsumoto, K., Shinozaki, K.. 1996. A gene encoding a mitogen-activated protein kinase kinase kinase is induced simultaneously with genes for a mitogen-activated protein kinase and an S6 ribosomal protein kinase by touch, cold, and water stress in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA. 93(2): 765-9.
Negi, V.S., Pal, A., Singh, R., and Borthakur, D. 2011. Identification of species-specific genes from Leucaena leucocephala using interspecies suppression subtractive hybridisation. Ann. Appl. Biol. 159(3): 387–398. doi: 10.1111/j.1744-7348.2011.00506.x.
Novaes, E., Kirst, M., Chiang, V., Winter-Sederoff, H., and Sederoff, R. 2010. Lignin and biomass: A negative correlation for wood formation and lignin content in trees. Plant Physiol. 154: 555-561. doi:10.1104/pp.110.161281
Nuruzzaman, M., Sharoni, A.M., and Kikuchi, S. 2013. Roles of NAC transcription factors in the regulation of biotic and abiotic stress responses in plants. Front Microbiol. 4:248. doi:10.3389/fmicb.2013.00248.
Oh, S.A., Kwak, J.M., Kwun, I.C., and Nam, H.G. 1996. Rapid and transient induction of calmodulin- encoding gene(s) of Brassica napus by a touch stimulus. Plant Cell Rep. 15(8): 586–590. doi: 10.1007/BF00232458.
Paul-Victor, C., and Rowe, N. 2011. Effect of mechanical perturbation on the biomechanics, primary growth and secondary tissue development of inflorescence stems of Arabidopsis thaliana. Ann. Bot. 107(2): 209–218. doi:10.1093/aob/mcq227.
Porra, R. 2002. The chequered history of the development and use of simultaneous equations for the accurate determination of chlorophylls a and b. Photosynth. Res. 73(1): 149–156. doi:10.1023/a:1020470224740.
Porter, B.W., Zhu, Y.J., Webb, D.T., and Christopher, D. a. 2009. Novel thigmomorphogenetic responses in Carica papaya: touch decreases anthocyanin levels and stimulates petiole cork outgrowths. Ann. Bot. 103(6): 847–58. doi:10.1093/aob/mcp009.
Pruyn, M.L., Ewers III, B.J., and Telewski, F.W. 2000. Thigmomorphogenesis: changes in the morphology and mechanical properties of two Populus hybrids in response to mechanical perturbation. Tree Physiol. 20(8): 535–540. doi:10.1093/treephys/20.8.535.
Purwar, S. 2012. Enzymes of phenylpropanoid metabolism involved in strengthening the structural barrier for providing genotype and stage dependent resistance to karnal bunt in wheat. Am. J. Plant Sci. 03(02): 261–267. doi:10.4236/ajps.2012.32031.
Rossel, J.B., Wilson, P.B., Hussain, D., Woo, N.S., Gordon, M.J., Mewett, O.P., Howell, K.A., Whelan, J., Kazan, K., and Pogson, B.J. 2007. Systemic and intracellular responses to photooxidative stress in Arabidopsis. Plant Cell 19(12): 4091–4110. doi:10.1105/tpc.106.045898.
Rushanaedy, I., Jones, T.C., Dudley, N.S., Liao, R.J.F., Agbayani, R., and Borthakur, D. 2012. Chitinase is a potential molecular biomarker for detecting resistance to Fusarium oxysporum in Acacia koa. Trop. Plant Biol. 5(3): 244–252. doi:10.1007/s12042-012-9108-7.
Saidi, I., Ammar, S., Demont-Caulet, N., Thévenin, J., Lapierre, C., Bouzid, S., and Jouanin, L. 2009. Thigmomorphogenesis in Solanum lycopersicum: morphological and biochemical
Page 29 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
30
responses in stem after mechanical stimulation. Plant Sci. 177(1): 1–6. doi:10.1016/j.plantsci.2009.03.002.
Seo, S., Okamoto, M., Seto, H., Ishizuka, K., Sano, H., and Ohashi, Y. 1995. Tobacco MAP kinase: a possible mediator in wound signal transduction pathways. Science 270(5244): 1988–92. doi:10.1017/CBO9781107415324.004.
Sims, D.A., and Gamon, J.A. 2002. Relationships between leaf pigment content and spectral reflectance across a wide range of species, leaf structures and developmental stages. Remote Sens. Environ. 81(2-3): 337–354. doi:10.1016/S0034-4257(02)00010-X.
Skyba, O., Douglas, C.J., Mansfield S.D. 2013. Syringyl-rich lignin renders poplars more resistant to degredation by wood decay fungi. Appl. Environ, Microbiol. 79(8):2560-2571. doi:10.1128/AEM.03182-12.
Smoth, V.C., and Ennos, A.R. 2003. The effects of air flow and stem flexure on the mechanical and hydraulic properties of the stems of sunflowers Helianthus annuus L. J. Exp. Bot. 54(383):845–849. doi:10.1093/jxb/erg068.
Tatsuki, M., and Mori, H. 1999. Rapid and transient expression of 1-aminocyclopropane-1-carboxylate synthase isogenes by touch and wound stimuli in tomato. Plant Cell Physiol. 40(7): 709–15. doi:10.1093/oxfordjournals.pcp.a029597.
Telewski, F.W. 1989. Structure and function of flexure wood in Abies fraseri. Tree Physiol. 5(1): 113–21. doi:10.1093/treephys/5.1.113.
Telewski, F.W. 2006. A unified hypothesis of mechanoperception in plants. Am. J. Bot. 93(10): 1466–76. doi:10.3732/ajb.93.10.1466.
Telewski, F.W., and Jaffe, M.J. 1986a. Thigmomorphogenesis: Field and laboratory studies of Abies fraseri in response to wind or mechanical perturbation. Physiol. Plant. 66(2): 211–218. doi:10.1111/j.1399-3054.1986.tb02411.x.
Telewski, F.W., and Jaffe, M.J. 1986b. Thigmomorphogenesis: anatomical, morphological and mechanical analysis of genetically different sibs of Pinus taeda in response to mechanical perturbation. Physiol. Plant. 66: 219–26. doi:10.1111/j.1399-3054.1986.tb02412.x.
Telewski, F.W., and Pruyn, M.L. 1998. Thigmomorphogenesis: a dose response to flexing in Ulmus americana seedlings. Tree Physiol. 18: 65–68. doi:10.1093/treephys/18.1.65.
Temmei, Y., Uchida, S., Hoshino, D., Kanzawa, N., Kuwahara, M., Sasaki, S., and Tsuchiya, T. 2005. Water channel activities of Mimosa pudica plasma membrane intrinsic proteins are regulated by direct interaction and phosphorylation. FEBS Lett. 579(20): 4417–22. doi:10.1016/j.febslet.2005.06.082.
Tretner, C., Huth, U., and Hause, B. 2008. Mechanostimulation of Medicago truncatula leads to enhanced levels of jasmonic acid. J. Exp. Bot. 59(10): 2847–2856. doi:10.1093/jxb/ern145.
Uppalapati, S.R., Marek, S.M., Lee, H.-K., Nakashima, J., Tang, Y., Sledge, M.K., Dixon, R. a, and Mysore, K.S. 2009. Global gene expression profiling during Medicago truncatula-Phymatotrichopsis omnivora interaction reveals a role for jasmonic acid, ethylene, and the flavonoid pathway in disease development. Mol. Plant. Microbe. Interact. 22(1): 7–17. doi:10.1094/MPMI-22-1-0007.
VanEtten, H.D., Matthews, D.E., and Matthews, P.S. 1989. Phytoalexin detoxification: importance for pathogenicity and practical implications. Annu. Rev. Phytopathol. 27: 143–164. doi:10.1146/annurev.py.27.090189.001043.
Vanholme R., Demedts B., Morreel K., Ralph J., Boerjan W. 2010. Lignin biosynthesis and structure. Plant Physiol 153(3): 895-905. doi:10.1104/pp.110.155119.
Page 30 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
31
Voelker, S.L., Lachenbruch, B., Meinzer F.C., Strauss S.H. 2011. New Phytol. 189:1096-1109. doi: 10.1111/j.1469-8137.2010.03572.x.
Visnovitz, T., Világi, I., Varró, P., and Kristóf, Z. 2007. Mechanoreceptor cells on the tertiary pulvini of Mimosa pudica L. Plant Signal. Behav. 2(6): 462–6. doi:10.4161/psb.2.6.4743.
Vogt, T. 2010. Phenylpropanoid biosynthesis. Mol. Plant 3(1): 2–20. doi:10.1093/mp/ssp106. Volkov, A.G., Adesina, T., Markin, V.S., and Jovanov, E. 2008. Kinetics and mechanism of
Dionaea muscipula trap closing. Plant Physiol. 146(2): 694–702. doi:10.1104/pp.107.108241. Volkov, A.G., Foster, J.C., Ashby, T.A., Walker, R.K., Johnson, J.A., and Markin, V.S. 2010.
Mimosa pudica: Electrical and mechanical stimulation of plant movements. Plant. Cell Environ. 33(2): 163–73. doi:10.1111/j.1365-3040.2009.02066.x.
War, A.R., Paulraj, M.G., Ahmad, T., Buhroo, A.A., Hussain, B., Ignacimuthu, S., et al. 2012. Mechanisms of plant defense against insect herbivores. Plant Signal. Behav. 7:1306–20. doi: 10.4161/psb.21663.
Whitehead, F.H., and Luti, R. 1962. Experimental studies on the effect of wind on plant growth and anatomy: I. Zea mays. New Phytol. 61(1): 56–58. doi:10.1111/j.1469-8137.1962.tb06273.x
Wilkinson, K.M., Elevitch, C.R. 2005. Propagation protocol for koa (Acacia koa Gray). Native Plants J. 6(3): 228-232. doi: 10.2979/NPJ.2005.6.3.228.
Wurzinger, B., Mair, A., Pfister, B., and Teige, M. 2011. Cross-talk of calcium-dependent protein kinase and MAP kinase signaling. Plant Signal. Behav. 6(1): 8–12. doi:10.4161/psb.6.1.14012.
Yanagida, J.F., Friday, J.B., Illukpitiya, P., Mamiit, R.J., and Edwards, Q. 2004. Economic value of Hawaii’s forest industry in 2001. In Economic Issues 7. College of Tropical Agriculture and Human Resources. University of Hawai’i at Manoa.
Zhao, J., Pang, Y., Dixon, R.A. 2010. The mysteries of proanthocyanidin transport and polymerization. Plant Physiol. 153:437-443. doi: 10.1104/pp.110.155432.
Page 31 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
32
TABLES
Table 1. Fold change of the genes at 10, 30, and 60 min following stress treatment, comparing stressed to unstressed determined using NanoString nCounter analysis. Acc. No. Putative function Expression fold change
10 min 30 min 60 min
Ratio SE Ratio SE Ratio SE
Protein degradation (4) GBYE01027655 RING finger protein, ALT family 10.68 0.72 38.06 2.38 18.81 1.47 GBYE01011012 RING finger protein 5.93 0.33 15.73 0.65 7.20 0.46 GBYE01014767 RING finger protein, ALT family 1.87 0.18 3.41 0.22 3.41 0.16 GBYE01007181 E3 ubiquitin ligase 1.81 0.15 3.50 0.11 4.48 0.15 Transcriptional regulation (23) GBYE01007979 AP2 domain class transcription factor 2.45 0.33 3.66 0.30 3.58 0.29 GBYE01025728 AP2 domain-containing transcription 2.90 0.14 18.74 1.93 31.43 4.86 GBYE01041899 AP2/ERF domain-containing protein 2.49 0.34 6.24 0.92 3.57 0.54 GBYE01036775 AP2/ERF domain-containing protein 2.69 0.23 6.66 0.57 3.54 0.33
GBYE01014032 Auxin-induced protein (1.36) 0.29 4.74 0.69 3.81 0.48 GBYE01048118 DRE binding protein 6.33 0.33 18.15 0.88 4.06 0.45 GBYE01071204 DRE binding protein 7.95 1.40 56.91 6.13 66.68 11.17 GBYE01008127 Ethylene-responsive element-binding protein 3.71 0.24 8.63 0.40 6.08 0.60 GBYE01017658 Ethylene-responsive transcription factor 4.61 0.33 26.58 2.11 21.16 2.05 GBYE01005208 Ethylene-responsive transcription factor 2.12 0.24 5.65 0.27 8.92 0.55 GBYE01026684 MYB transcription factor 1.40 0.04 6.09 0.59 11.36 1.06 GBYE01030298 MYB transcription factor 1.47 0.08 5.55 0.47 9.98 0.80 GBYE01024450 NAC domain protein 1.68 0.18 10.27 1.10 26.38 3.46 GBYE01010007 NAC domain protein 2.44 0.49 6.88 0.46 7.91 0.72 GBYE01039799 WRKY transcription factor 5.25 0.76 58.71 5.25 37.01 5.82 GBYE01016099 WRKY transcription factor 2.13 0.25 15.47 1.03 14.31 1.81 GBYE01050872 WRKY transcription factor 2.19 0.40 11.64 1.10 13.39 1.24 GBYE01005028 WRKY transcription factor 1.68 0.15 3.68 0.21 3.51 0.30 GBYE01041303 WRKY transcription factor 4.72 1.04 70.75 10.92 55.66 11.37 GBYE01004300 WRKY transcription factor 2.15 0.26 9.32 0.82 14.86 1.62
GBYE01019260 Zinc finger protein 11.42 0.87 45.00 2.23 31.36 2.93 GBYE01019654 Zinc finger protein 5.21 0.30 27.14 1.02 13.22 1.34 GBYE01022591 Zinc finger protein 2.93 0.21 7.41 0.48 7.99 0.71 Disease resistance (3) GBYE01072738 CC-NBS-IRR resistance protein 8.72 2.17 19.15 2.31 10.29 1.60 GBYE01078745 NBS-LRR disease-resistance protein 5.08 0.80 15.17 2.13 5.32 1.19 GBYE01050160 NBS-LRR disease-resistance protein 3.93 0.15 10.41 0.56 6.09 0.47 Signal transduction (12) GBYE01006842 Calcium-binding EF-hand protein 1.99 0.03 7.25 0.28 8.12 0.65 GBYE01006511 Calcium-binding protein 1.72 0.06 7.12 0.68 15.96 1.25 GBYE01005328 Calcium-binding protein 1.59 0.09 4.13 0.30 5.16 0.44 GBYE01000487 Calmodulin 1.23 0.05 2.47 0.08 3.67 0.19 GBYE01007399 Calmodulin binding protein 2.30 0.11 7.49 0.36 5.66 0.53 GBYE01012536 Lectin protein kinase family protein 2.13 0.26 7.22 0.64 9.31 1.13 GBYE01007796 LysM type receptor kinase 2.53 0.22 12.02 0.97 12.29 0.77 GBYE01008850 Mitogen-activated protein kinase 5.00 0.40 24.10 1.24 23.69 2.03 GBYE01006033 Mitogen-activated protein kinase 4.31 0.31 23.17 1.05 23.20 1.94 GBYE01052680 Octicosapeptide/Phox/Bem1p (PB1) 6.60 0.45 8.67 0.34 8.56 0.98 GBYE01013735 Protein phosphatase 2C 4.89 0.28 19.97 0.74 20.03 1.57 GBYE01026109 Protein phosphatase 2C 4.71 0.35 20.52 0.70 7.30 1.18 Carbohydrate metabolism (3) GBYE01003680 UDP-D-glucuronic acid 4-epimerase 3.14 0.09 10.34 0.66 11.14 0.82 GBYE01007604 UDP-glucosedehydrogenase 1.40 0.06 4.87 0.25 6.25 0.72 GBYE01001472 UDP-glucuronate 5-epimerase 2.80 0.11 10.83 0.71 11.61 0.87 Hormone metabolism (2) GBYE01025591 1-aminocyclopropane 1-carboxylate synthase 3.51 0.67 17.56 1.58 16.44 1.88 GBYE01014926 Abscisic acid 8'-hydroxylase 2.62 0.18 29.96 1.37 35.80 3.87 Protein transport and sorting (2) GBYE01041529 Syntaxin 1.43 0.18 8.38 0.63 9.91 1.39 GBYE01007398 Syntaxin 1.97 0.18 5.15 0.18 5.33 0.54 Others (2) GBYE01000347 Lignin biosynthetic peroxidase (0.91) 0.15 1.69 0.29 3.38 0.57 GBYE01000925 Metal ion binding protein 2.65 0.32 6.57 0.38 5.69 0.31
NOTE: n = 3 for each treatment, where n represents 10 A. koa stems; p < 0.05 unless in prentices; p ≥ 0.05 for those in prentices.
Page 32 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
33
FIGURE LEGENDS
Fig. 1. Phenotypical quantifications of A. koa seedlings following two and six months of the mechanical stress treatment. The stressed and unstressed plants are labeled as S and U, respectively: (A) Visual growth differences between stressed and unstressed after two months of treatment and (B) six months of treatment; (C) quantification of the stem thickness of stressed and unstressed; (D) quantification of the stem heights of stressed and unstressed; (E) quantification of the maximum leaflet length of stressed and unstressed; (F) quantification of the root dry weight of stressed and unstressed; In (C-F) n = 39 for each of the 2-month-old stressed and unstressed groups, and n = 15 for each of the 6-month-old stressed and unstressed groups. An asterisk (*) indicate significant changes in the stressed compared to unstressed by two-tail t-test (p < 0.05), and error bars represent standard errors.
Fig. 2. Quantifications of anthocyanin and lignin from stems of A. koa seedlings following two months of mechanical stress treatment. The stressed and unstressed plants are labeled as S and U, respectively: (A) Visual stem anthocyanins in stressed and unstressed; (B) quantification of anthocyanin concentration in stems of stressed and unstressed; (C) quantification of lignin composition in hypocotyls of stressed and unstressed. In (B), n = 14 for each of the stressed and unstressed group; in (C), n = 3 for each of the stressed and unstressed group, where n represents a pool of 10 hypocotyls. An asterisk (*) indicate significant changes in the stressed compared to unstressed by two-tail t-test (p < 0.05), and error bars represent standard errors. Fig. 3. Microscopic analysis of cross-sections of xylem in two-month-old stressed and unstressed A. koa seedlings; (A) stem cross-section of the stressed; (B) stem cross-section of the unstressed; (C) quantification of average xylem cell size in the stressed and unstressed; (D) quantification of xylem cell density in the stressed and unstressed A. koa seedlings. In (C) and (D), n = 3 for each of the stressed and unstressed group. An asterisk (*) indicate significant changes in the stressed compared to unstressed by two-tail t-test (p < 0.05), and error bars represent standard errors. Fig. 4. Microarray analysis of 4,000 genes that may be related to disease resistance and wood formation and development. A scatter plot of log2 of the signal intensities comparing the gene expressions in stressed (S) and unstressed (U). Each group had two biological replicates (n = 2). In stressed, n represents pools of stems from 30 plants collected at 10, 30, and 60 min after the treatment (10 plants for each time point); in unstressed, n represents pools of 20 unstressed stems. The two lines represent a cutoff for > 2-fold change in expression; those genes induced more than 2-fold were analyzed in detail. Fig. 5. Functional categorization of genes determined to be upregulated by mechanical stress through the microarray analysis. Sixty sequences shown to be upregulated by mechanical stress more than twofold were categorized into functional groups according to their closest annotations obtained from Blastx.
Page 33 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
Ishihara, et al. 2016
P a g e | 34
SUPPLEMENTARY DATA
Table S1. Probe sequences used for the NanoString nCounter analysis. Table S2. Primer sequences used for the qRT-PCR analysis. Table S3. Stability values of candidate reference genes for qRT-PCR analysis determined by NormFinder. Table S4. Microscopic analysis of cross-sections of xylems in two-month-old stressed, two- and three-month-old unstressed A. koa seedlings. Table S5. Microarray analysis of sequences induced by mechanical stress in A. koa.
Fig. S1. Verification of expression levels obtained from the NanoString nCounter analysis by qRT-PCR.
Page 34 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research
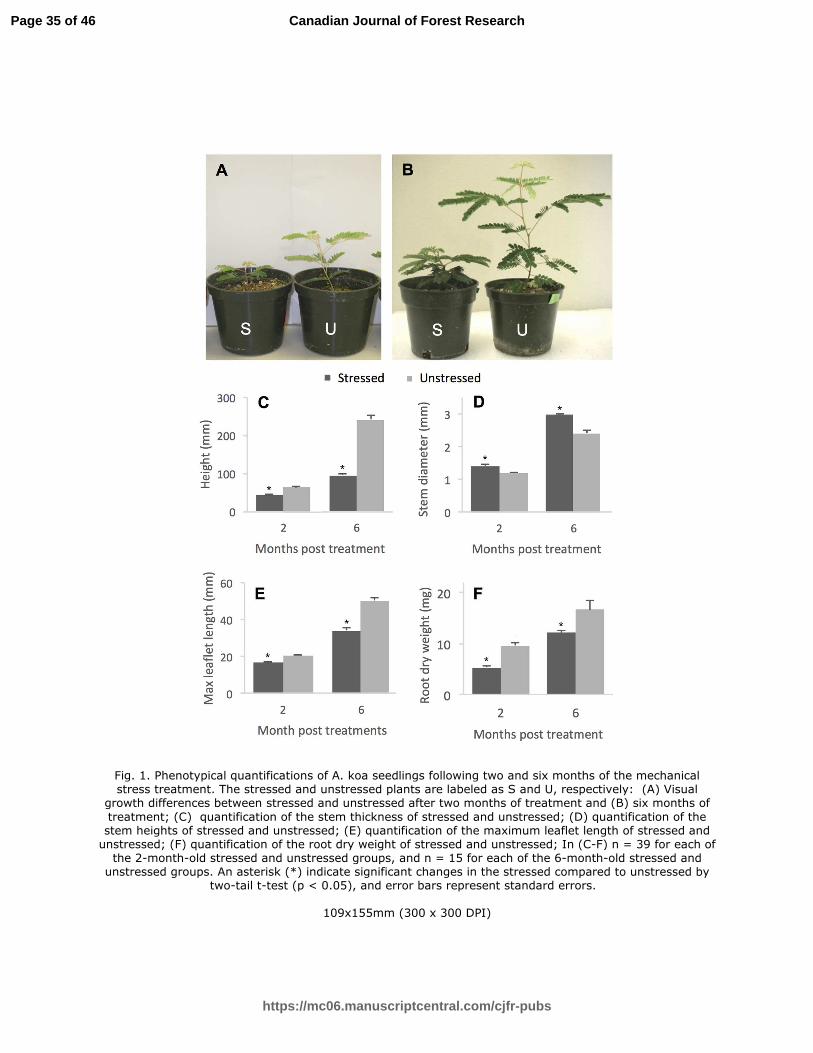
Draft
Fig. 1. Phenotypical quantifications of A. koa seedlings following two and six months of the mechanical stress treatment. The stressed and unstressed plants are labeled as S and U, respectively: (A) Visual
growth differences between stressed and unstressed after two months of treatment and (B) six months of
treatment; (C) quantification of the stem thickness of stressed and unstressed; (D) quantification of the stem heights of stressed and unstressed; (E) quantification of the maximum leaflet length of stressed and unstressed; (F) quantification of the root dry weight of stressed and unstressed; In (C-F) n = 39 for each of the 2-month-old stressed and unstressed groups, and n = 15 for each of the 6-month-old stressed and unstressed groups. An asterisk (*) indicate significant changes in the stressed compared to unstressed by
two-tail t-test (p < 0.05), and error bars represent standard errors.
109x155mm (300 x 300 DPI)
Page 35 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
Fig. 2. Quantifications of anthocyanin and lignin from stems of A. koa seedlings following two months of mechanical stress treatment. The stressed and unstressed plants are labeled as S and U, respectively: (A) Visual stem anthocyanins in stressed and unstressed; (B) quantification of anthocyanin concentration in
stems of stressed and unstressed; (C) quantification of lignin composition in hypocotyls of stressed and unstressed. In (B), n = 14 for each of the stressed and unstressed group; in (C), n = 3 for each of the stressed and unstressed group, where n represents a pool of 10 hypocotyls. An asterisk (*) indicate
significant changes in the stressed compared to unstressed by two-tail t-test (p < 0.05), and error bars represent standard errors.
155x43mm (300 x 300 DPI)
Page 36 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
Fig. 3. Microscopic analysis of cross-sections of xylem in two-month-old stressed and unstressed A. koa seedlings; (A) stem cross-section of the stressed; (B) stem cross-section of the unstressed; (C)
quantification of average xylem cell size in the stressed and unstressed; (D) quantification of xylem cell
density in the stressed and unstressed A. koa seedlings. In (C) and (D), n = 3 for each of the stressed and unstressed group. An asterisk (*) indicate significant changes in the stressed compared to unstressed by
two-tail t-test (p < 0.05), and error bars represent standard errors.
419x106mm (300 x 300 DPI)
Page 37 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
Fig. 4. Microarray analysis of 4,000 genes that may be related to disease resistance and wood formation and development. A scatter plot of log2 of the signal intensities comparing the gene expressions in stressed (S) and unstressed (U). Each group had two biological replicates (n = 2). In stressed, n represents pools of 30
stems collected at 10, 30, and 60 min after the treatment; in unstressed, n represents pools of 20 unstressed stems. The two lines represent a cutoff for > 2-fold change in expression; those genes induced
more than 2-fold were analyzed in detail.
103x94mm (300 x 300 DPI)
Page 38 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
Fig. 5. Functional categorization of genes determined to be upregulated by mechanical stress through the microarray analysis. Sixty sequences shown to be upregulated by mechanical stress more than twofold were
categorized into functional groups according to their closest annotations obtained from Blastx.
109x71mm (300 x 300 DPI)
Page 39 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
Table S1. Probe sequences used for the NanoString nCounter analysis AccessionNo. ProbeA ProbeB
GBYE01000072 CAATATCGATTGTGATTCCTCTTTCACGCTCAGCCTTGAGCTTGTCCAGCCTCAAGACCTAAGCGACAGCGTGACCTTGTTTCA
CGAAAGCCATGACCTCCGATCACTCGGCGTCAATGACTGTGCAGTAGTACTTGGTGGTCTCGAATTTCCAGAGAG
GBYE01000611 AGAGAGAGAACAGCCTGGATGGCAACATACATGGCAGGGACATTAAAGGTCATCCTCTTCTTTTCTTGGTGTTGAGAAGATGCTC
CGAAAGCCATGACCTCCGATCACTCCACCATCACCAGAGTCCAAAACAATACCGGTTGTACGACCACTAGCATAT
GBYE01000815 CTGCTCTCGACCTCGAGAGTTATGGTCTTCCCTGTAAGGGTTTTAACGAACACAATTCTGCGGGTTAGCAGGAAGGTTAGGGAAC
CGAAAGCCATGACCTCCGATCACTCGGATGCCTTCCTTGTCTTGAATCTTAGCCTTAACATTGTCGATGGTGTCG
GBYE01027655 GAGCGGCGGCGGAGTCTTCGTAGAATACAAGGTGAGAGACTGCGTGAGCGCTGTTGAGATTATTGAGCTTCATCATGACCAGAAG
CGAAAGCCATGACCTCCGATCACTCGAGAGACATGAGTAGAGATGAGTCAAGACCGCGCGTTGAGGCCACGACAG
GBYE01019654 GACTCTCCACCTCTTGCTCTACACCGATCTTCTTCATCCCAGAATAATCGCAAAGACGCCTATCTTCCAGTTTGATCGGGAAACT
CGAAAGCCATGACCTCCGATCACTCGGTAAAAAGCAAACGTCTTCTCTTATTACCGGCGGCAGCCACCGGCAACG
GBYE01019260 TCTCTTGGAACGCTTGCGCTTGGTCCATGACTCCAAGGAGTGAAGATTGGCGAACCTAACTCCTCGCTACATTCCTATTGTTTTC
CGAAAGCCATGACCTCCGATCACTCCGAGATACTCTTCCTCGGAGGTGCATGACGAGGGATGGTCGAGACGGGA
GBYE01017658 TGCGGAAGAGTGCCAGGAAACACGGTAGCCGGCAACGGGAACTGCTGCTGCCAATTTGGTTTTACTCCCCTCGATTATGCGGAGT
CGAAAGCCATGACCTCCGATCACTCCCGCCATGCTTGCACGGAAGACCGCGTCGAAGTAAAACGCCTGATTCACC
GBYE01026109 CAAACCCCACGGCATAAATCAACATAGCCACCTGTGGTCTCAATTCTGTCCTTTCGGGTTATATCTATCATTTACTTGACACCCT
CGAAAGCCATGACCTCCGATCACTCAATACCGATCTCCAATTCCTCTGGAAACGGCTAGGGACCCCTGAATCCTC
GBYE01048118 AGAGTTGGCTGCTCGAAACCAATCAAGTTTTGGTTGCTGAATAACCCGTCCAACAGCCACTTTTTTTCCAAATTTTGCAAGAGCC
CGAAAGCCATGACCTCCGATCACTCGTTGAAACTGGTCGGTCTGAGACAGAGTCTGGTTGTTGAGCCCAAAAGGA
GBYE01041303 TCATTCTCTAGTGCCAAGGAAGGCAAACTGTAACTTTCTGCTGGTGTCATCACCGTGTGGACGGCAACTCAGAGATAACGCATAT
CGAAAGCCATGACCTCCGATCACTCGCGTATTTGGAGATAGCAAGTCTGTTTGTGAAAGGTTCCTCCAGAAGGGA
GBYE01039799 ACTAGGTAATACTGGCCCCGTGGGAAGAGAATTGGTATTTGAGTTTGATGCCTGGAGTTTATGTATTGCCAACGAGTTTGTCTTT
CGAAAGCCATGACCTCCGATCACTCTCATCTCTGAAAGGACTTACGTTGACAGATATTGGAGACTCTGGTGGCAA
GBYE01011012 CGGCGGAACCCTTGTCTGCTGAAATCGTAAGAAGTAGGAGCTGAATCGGGCAGATAAGGTTGTTATTGTGGAGGATGTTACTACA
CGAAAGCCATGACCTCCGATCACTCGAACAATAACAGGGTTAAAGGGAGATCTGTTGCCTGCGTTGCGGCGTCTT
GBYE01078745 ATGACAAATATCTTTTGAGGAGTCCCCAAGTTCAAAGTTTCTAGATTCCACTTCCTTCCTGTGTTCCAGCTACAAACTTAGAAAC
CGAAAGCCATGACCTCCGATCACTCTATAAAGATGCCTTAGTCTCTTGAGCTTCCACATTTCAATGTTGAAGTTT
GBYE01016099 TTGTGTTGTGAAGCGAATTTGTGAAACTCGATTTGCCGGGATAGCTGGTCCATAAAATTGGTTTTGCCTTTCAGCAATTCAACTT
CGAAAGCCATGACCTCCGATCACTCCATGTCAAATGGGAATGGCTCTTGACTTGCAGATGTTGTTGTTGATTGAC
GBYE01007796 CTTTGTAGATTTTAACGGCGGCATCGTCTCCTTTAAACGTTGCTCTGTATCTGGTCAAGACTTGCATGAGGACCCGCAAATTCCT
CGAAAGCCATGACCTCCGATCACTCGTTGGTGTGGTTGAGTCGCTTAAGAATGCTGATCTCTTTCGAGACGTCGC
GBYE01072738 ATCTGTTCAATCTTCTGTTTAATATCTCAATCTCCTCGATTACAGGAGTTCTTTCGTTGGGACGCTTGAAGCGCAAGTAGAAAAC
CGAAAGCCATGACCTCCGATCACTCACCATCTTCCAGCAATCTCATCAACACAATAGTTTCTCTTCCATATTCTA
GBYE01022591 CTCTTCTTCTCCGATCTTCACACTCTTCCCGTCTGCCAGCAGACCTGCAATATCAAAGTTATAAGCGCGT
CGAAAGCCATGACCTCCGATCACTCATGAACAAACGAGGCTTCTTGGCGGCGGCCATCGGTAACGGGCTCTCCAC
GBYE01013735 TCGGTTTCATCCTCAAGGCCGCCAAAACAGCGTTCCAGACTTCCATAAAACCTGCCAATGCACTCGATCTTGTCATTTTTTTGCG
CGAAAGCCATGACCTCCGATCACTCAGAAATCACCAGAGGCGTGTGGCTTCAAGTCCGAGTGCCAGAGAAGACCG
GBYE01014926 AGATCCTCTCTGTATAAAATCTCATCTTTTCCGAAGATCGAGAGCAGAGCCAAACTGGAGAGAGAAGTGAAGACGATTTAACCCA
CGAAAGCCATGACCTCCGATCACTCTAATTGGCATTGAGTTGTAACCTCTCTCCAGAATGTAGTAGCAGCGTTTC
GBYE01008127 AGTCACTCGAAGCTGCGCTTCCCATTTTGGTCTTAACGTCGACCGACAAACGATTGCTGCATTCCGCTCAACGCTTGAGGAAGTA
CGAAAGCCATGACCTCCGATCACTCGAGATTAATGGCTTTCTTCGGCTGGCAATCAACGACGGAGGAGGAATCGG
GBYE01008850 TCAGTATCCATAAGTTCCGTGGTCATGTAGACATCGGTAAACTCTCTACGCTGAGGCTGTTAAAGCTGTAGCAACTCTTCCACGA
CGAAAGCCATGACCTCCGATCACTCGGCAGTGCTCCTCCGAGAGATTTTGATTTGAACGGATGATGTGGTGAAGA
GBYE01006033 TCATCAACTCCATGAAGATGCAACCTACAGACCACACATCTATGGCAGAGCTAGGACGCAAATCACTTGAAGAAGTGAAAGCGAG
CGAAAGCCATGACCTCCGATCACTCTAAGCGCATCTGATGCACATGATCCTTGCCGGGAAATAGAGGCTTTTTAT
GBYE01000925 TTCTGATAATAGTTCTGATAGGCTTTAAGAAGATCAGCGACTTGTTCCTTCCACGCGATGACGTTCGTCAAGAGTCGCATAATCT
CGAAAGCCATGACCTCCGATCACTCTGGCATAATAAGTAGGATGTTGAGGATAGTAGTATTGTGTTTTCATCTGA
GBYE01025591 TGGGATGAAACGGATCGTTTTCGTAAGCCTTCCAGCCATCAAAGTACGGGCATTTGGAATGATGTGTACTGGGAATAAGACGACG
CGAAAGCCATGACCTCCGATCACTCGAGCTGATTTTCAGCCAGACCCATCTGAATAACCCCATTAGAGTTTTTGG
GBYE01024450 CTGGGTGTTGTATTTCTCAATCACACCTTTCTTGTTGTAAATCCGGCACACACAAGAATCCCTGCTAGCTGAAGGAGGGTCAAAC
CGAAAGCCATGACCTCCGATCACTCGTTTCGTGCTCATCCACATCTGAGGAAACAAACCCGCTCGATTTCTGATC
GBYE01003680 AGGTGCATTACGTGCGTGAAGGCAACGGTGTCGAAAAGCTTCGCCAGAAGCTTGACGTAGATTGCTATCAGGTTACGATGACTGC
CGAAAGCCATGACCTCCGATCACTCCATAGGAGTTTGGATTCTCAATAGCATACCTAACACCGGCTTGAGCCGCC
GBYE01050160 TACTCTCAGATATGCATAGTTCTCTCAGAAGATCATGGATGCAACATGTCCTTACAGATCGTGTGCTCATGACTTCCACAGACGT
CGAAAGCCATGACCTCCGATCACTCTTCCTCCAAAATGTCTACATCCCTACAAACTTCAAAAGCTTTCTCGTCCC
GBYE01005208 CGCCCCAATTGTCCGTCAAACACGGGTACAAGCTGCTGAAACTGGAGCTCCTTGGAGGAGTTGATAGTGGTAAAACAACATTAGC
CGAAAGCCATGACCTCCGATCACTCCGTAGACCACCATGTCTTCGGAATCGTCCTCCTTGAGCGGCAATT
GBYE01041529 GCTTTACGAAGGAAAGTCTCACTCTCACCTGTAGAAATCAGGAGGTCGATCCTACGTATATATCCAAGTGGTTATGTCCGACGGC
CGAAAGCCATGACCTCCGATCACTCGAATTTCATTGATGGTGTCCAGTATTCTTCCTCTACCTTGTTCTTGGATG
GBYE01050872 AATTTTATCCTTTGTATCAAATTCCTCTGTTTTAACTTCAACCTCTGCTCCAGCAAGAAGGAGTATGGAACTTATAGCAAGAGAG
CGAAAGCCATGACCTCCGATCACTCTCTTCATTCTCAGATCCTATTGACGCGTAAGGGAAGCAAAAGGGTTCGAA
GBYE01001472 TTCCTCTTCGCCTTCACCTTCAAGTGACTCTCAAGGATACTCACTAATTTCACCCCTCCAAACGCATTCTTATTGGCAAATGGAA
CGAAAGCCATGACCTCCGATCACTCTATTGGCGTGTGTGAACGGAACGTCGCCGTTTCCGGGCATGTCTACGATC
GBYE01052680 AGCAGTTGTGACAGATAAGGCGATAGGCCATCGAATAAGGCAAGGACTCGCCCGAAGCAATACTGTCGTCACTCTGTATGTCCGT
CGAAAGCCATGACCTCCGATCACTCAAACTTAGGGCTCCCGCTATGGACACCGATTGGATATGCAACCGGCGGTG
GBYE01026684 GAATCTGAATCTTCTGTGACTCCTTCAAATCGGGTCCCTGGGTGTACAGACCGGGAATCGGCATTTCGCATTCTTAGGATCTAAA
CGAAAGCCATGACCTCCGATCACTCCAACTTCTCAGACTGCCTTTGTACCTTGGCCTTAAAACCAGGAGGACTCG
GBYE01025728 GATACCCATGATCCCCAGCTTCTCATTCTCACTCCCCGATCTTCATAACGGACAAACTGAACGGGCCATT
CGAAAGCCATGACCTCCGATCACTCCAGCCATATTCTTGTTTTCTGGTTAGGTGCTCTGATCTCT
GBYE01006842 TAACACGGATCTCAATTCGTCGACGGTGATGAAGCCGTCGCTGTTTTGGTCGCTATGCAGACGAGCTGGCAGAGGAGAGAAATCA
CGAAAGCCATGACCTCCGATCACTCTTCCTGCAATCTTCGACCGCTCTTCCCTGTTTCAACCCAAGCGACGA
Page 40 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
Table S1 (cont.). Probe sequences used for the NanoString nCounter analysis AccessionNo. ProbeA ProbeB
GBYE01014767 CACACAACTTCGGACTCCGCCAGTTCAGGATCGGCCTCCACAGAATGCATTCGCAACCATGTGAAGTAATGTGAGCGTACTT
CGAAAGCCATGACCTCCGATCACTCCGATCGAGACCCTCGCGTTGTGCGGTTCTGTTTCACCGTCTGATTAAGG
GBYE01012536 CAAAGTCTCCCAACTTGGCCGTGAAGTCTGAGTCCAACAATATGTTGCTGCACCAGTTAGCGTGGCGTATACCATGTTGTTAACA
CGAAAGCCATGACCTCCGATCACTCTTTTGTTGAGTGGTGAGTTTCATCTCTCTTCTGAATGGTACGAGCCAGAC
GBYE01000421 GGTTCGCAGTGAAGAGTGCCAATAGGAGAAATAGAGAAGGCAACAAGGCCCCTGAATCAATAGAACAATATCAGTTATGGCGGTG
CGAAAGCCATGACCTCCGATCACTCGTTGCCCATCATGCCATAACAAACACCGATCTGAGCATCTGAGATGTGAA
GBYE01004300 GACTTCCTTTTCTTTGATGGGATGTTGGTGGTGTTAAGATCCTTTTGAGGCGGTTGTTAATATGACAGGCCGCTAAAGACGTTCT
CGAAAGCCATGACCTCCGATCACTCCCATGGGATTACTATTAGAGTTGTTGTTGGTAGTGGTGATGCTGGTATCA
GBYE01021991 CATTAGGCTCGTTTATGGTAACCCAATACTTCACTCTATCCCCAAACTTCCCGTCTCAGATGAGTGGGTTAATCAATCAAGTATG
CGAAAGCCATGACCTCCGATCACTCGCATCTTGAAGGAGGCCAAATTCCTTTTCTGTACCCTCTAATTACTGCTA
GBYE01071204 CCCACTTGCCAGAGTTCCTCCTCCGAATACCGCGGCTGACACATTAGTAACGTCGGCAAGCACTTAGTCG
CGAAAGCCATGACCTCCGATCACTCTCTCGTCTTCTTATTGGGCTCCCTCACTTCACACA
GBYE01007399 TTCAGGAAGAGACAGTATAGGCACAGTTTGGCTTGGTAGAGAATCAGAACCGTGAACCAGATTATGTATGGACGCGCAATAGATA
CGAAAGCCATGACCTCCGATCACTCGCAGCAGCATCAAGCTCACTTTTGGGTCTTGGTGAAGAGAAAATTACTTC
GBYE01006511 AAAGGCCTCTCTCAACTCCTCGGAATCGCAATCCGGCTCGCACGCCGAACCATACGAAATTTGAGCAAGCAATTGAAGGCTTAGA
CGAAAGCCATGACCTCCGATCACTCTTAAGTTCCTCCGCCGAGATTTTCCCGTCGTGATCCGTGTCAAAAATGGC
GBYE01007979 AAGTGTGTTTCCTTAGCATATCGACGATCTCAGCTTTCGAATGCGCGTTGCTATCAGCTAATAGGGTCGGCTCAACAGTGTATCC
CGAAAGCCATGACCTCCGATCACTCTCCACCGTTAATCCGTTGTTTCGCTTGAACTGTTCCAGCTCATCATTGT
GBYE01005028 TAAGCACGAGGACAAGGGTTTCCCTTCGCCATTTTCTGTCCGTATTTTCGCTATCAATTCGTGACCCCGATCATCCAGTCCAGAA
CGAAAGCCATGACCTCCGATCACTCGTTGTACTTGTTTTCGAACTGGGCAACCGACGGCCATGGTGCATCTATAG
GBYE01000327 ACACCGCCGCTGACGTCGTCGCAGTTCTCGGCCAGCTTGAGCTCTAGGCCCAAAACGACCTTAATGGTCA
CGAAAGCCATGACCTCCGATCACTCTGAGCATCTTATTGAAGGTGTCCTTGGAGATGACGCTGCCGAGGGCATTT
GBYE01010007 ATTTTGTCGGTTCCAGTGGCTTTCCAGTACCCAGACTAGCCCAGATCCTACGAGATGAGCTACGTAACTA
CGAAAGCCATGACCTCCGATCACTCAGAAAACGAGAGCTTTCTTTATGCCAACCTTTCGACCAGCGGTGGTGATG
GBYE01041899 TGTTCCCAGCCAGAGGCGAGTCCTGTTTCGAGGAAGACGAATCTCCAAATGCACTCTATATGGAGGGAGAGTAGCTGGAT
CGAAAGCCATGACCTCCGATCACTCTTAAAGGCCTCGCGATCATAAGCCAGGGCAGCATCTTCAGCTGTGTCGAA
GBYE01007604 AAACAAATGCAGGCTTCCTCATGTTGTCGTAGATTCTCTGGTAATCCAGACCTGGTCTAGGTATCTAATTCGTGGGTCGGGTACT
CGAAAGCCATGACCTCCGATCACTCGAAGCCAATCTTCCTCAGCTTCCCAACATCCATAATGTTTCTTCCATCAA
GBYE01005328 CTAATCTTGCCGTCGCGGTCAAAGTCAAGGACATCGAATGCTGGTCGAAACATTAGCTCGGATGCTATCAGCTTGCGCCTATTAT
CGAAAGCCATGACCTCCGATCACTCCATCCCTGCCGGCGTAGAATGCCCGAAGATCGTCGCGG
GBYE01030298 GGTGTGAAGGACCGGTGCTCCACGTCCGGCGACAGCACGATCTGTATTTTGCACCTTTCGCTATGCTGAG
CGAAAGCCATGACCTCCGATCACTCCGAACTCTTTGTGGGCCTTGACGATGATCTCGTCTTCCTCC
GBYE01036775 AACACCTGCTGCTGATTTTGCGTCGCCTGAAACGGTGGGCGAAACGCCGGCTGTGTCCGTCTATACGCATACTGGTCCACATATA
CGAAAGCCATGACCTCCGATCACTCGGGCTAAGAATTGTGGGTACACCATGTTTGGCTGTTGGGACCCAAAAGAC
GBYE01000487 CTTCATCAGTAAGCTTCTCCCCAAGATTGGTCATCACATGGCGGAGCTCGCATGTTGGAGTTAACGGAGACCCGCCATCGTTTAC
CGAAAGCCATGACCTCCGATCACTCTATTTGGCCATCACCATCAACATCAGCCTCTCGAATCATCTCGTCAACTT
GBYE01016418 ATCAGCAATATGGGCCTCAAATTCAGAATCCAGCAACACATTACTAGCCTCGCTCATTTTGAACATACGATTGCGATTACGGAAA
CGAAAGCCATGACCTCCGATCACTCTGAGTCGACACATGAGAGGTCGAAGTATCAATCCTCCTAGCGAGACCAAA
GBYE01014032 TCCTCAGGCGAACATTTCCTTTTGCTTCCAACATTGCAACGTTTCTCTGGCCTATGCATCATGTGCCTCACTAGGACATCATGCT
CGAAAGCCATGACCTCCGATCACTCCTGACTGGTTATGATCACTTTTAGCGTCACTTCTATTGCCTGTTGATTCC
GBYE01007181 TATTCTCATCGCACAGTGAGAGATTCAAAATCGTGGTGACGCCGTACTCTCCTAAATTGGGAAAAAAGGTTTTAGCTATTGATGG
CGAAAGCCATGACCTCCGATCACTCGAGTGCTCGAACCAGAGGCTTTATTGCTCCTGACGAAACGATAAGCTCCT
GBYE01002764 AGATTCCCGCACAACCTTAGAGAAGAACTTCTGTGCATGGCTTCGAATCTCTTCAGTTAAAGGCTATCTTGCTCCGCTCGTTCTC
CGAAAGCCATGACCTCCGATCACTCCGAGGAGGTGGTATGTCAATTGGCTTTATAGAACTTTCAGTGCTACCCTC
GBYE01013774 AAGCAGCTCGAGCAACTATGGAACTTGTGATTGGTGGAATTATCTTACCGCTTAAAGCTATCCACGAATGTCAAAAATGTGGTTT
CGAAAGCCATGACCTCCGATCACTCGTACTCTATTGTCGATAGGATTTGCTCAACGATCTCTGTTTTCTTACCAA
GBYE01007398 GTGGACGCCATTTCGATGACATGATGGTCCGGCGATGGCTGCTCCCGAATGTATAATGCTGACGTTCTTGCTTTTGGC
CGAAAGCCATGACCTCCGATCACTCCACGTTGTCGAAGAACTTGTCGAGGTTGACACCGCCGGTGGGCGTC
GBYE01000347 TCGGGCTTCTCGGGTAAGAGCTTTACTTATTTCGCTGCGAACAATGGCCTCCTATTGAAGCAATCCTCTCCCCAATACTTAAAAA
CGAAAGCCATGACCTCCGATCACTCCCATTGACGAAGCAGTCGTGGAAGAAGAGACGGAGGATGGAGGCACCGAG
Page 41 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
Table S2. Primer sequences used for qRT-PCR analysis. Acc.no. Putativefunction Forwardprimer(5à3) Reverseprimer(5à3)GBYE01027655 RINGfingerprotein CACGCAGTCTCTCACCTTGT CATTCAACCTGCTCGGTGTGGBYE01048118 DREbindingprotein CTTCTTCCATTCCTCTCCT TTGAAACTGGTCGGTCTGGBYE01041303 WRKYtranscriptionfactor AATGCTCCTAAATCTCC CCTCTTGTAATGGCTCGTGBYE01019654 Zincfingerprotein GCCTTCAACTATGACCAAGAA GATGAGACAGAGAGCGAGGTGBYE01078745 NBS-LRRdisease-resistanceprotein TGGAATCTACACACCCTATCTT GCTTTGGGAATACACCTTTC
Page 42 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
Table S3. Stability values of candidate reference genes for qRT-PCR analysis determined by NormFinder. Genenames 10min 30min 60mintubulin 0.473 0.432 0.397ef1a 0.061 0.396 0.31718S 0.048 0.123 0.132ubq 0.073 0.322 0.2575.8S 0.255 0.301 0.267actin 0.165 0.535 0.520Thebestcombination* 0.039 0.157 0.153
*18S and ef1a for 10 min, 18S and 5.8S for 30 and 60 min
Page 43 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
Table S4. Microscopic analysis of cross-sections of xylem in two-month-old stressed and two- and three-month-old unstressed A. koa seedlings*. 2-month-old,stressed 2-month-old,unstressed 3-month-old,unstressedXylemcelldensity(cells/104µm2)
71.48±2.83a 49.07±3.85b 42.97±3.49b
Xylemcellsize(µm2)
140.43±5.63c 207.68±17.74d 235.97±19.52d
*Letters (a, b, c, d) indicate significant differences (P < 0.05) by t-test. Error bars represent standard error.
Page 44 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research
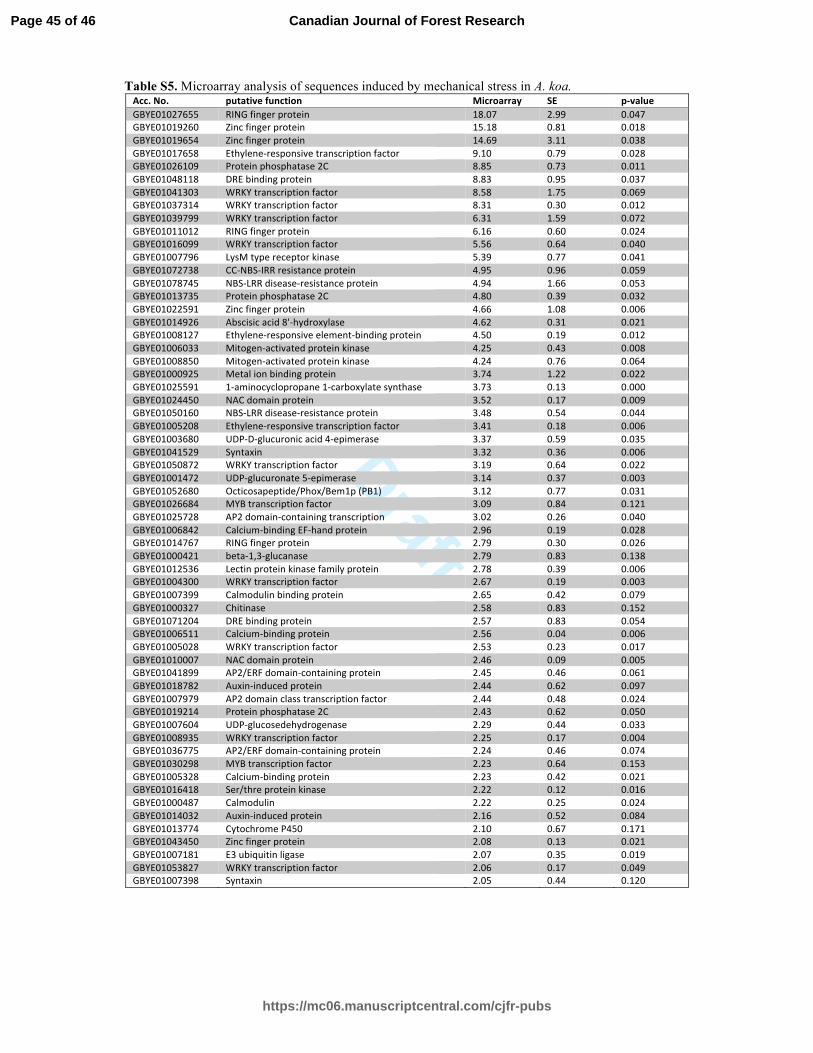
Draft
Table S5. Microarray analysis of sequences induced by mechanical stress in A. koa.
Acc.No. putativefunction Microarray SE p-valueGBYE01027655 RINGfingerprotein 18.07 2.99 0.047GBYE01019260 Zincfingerprotein 15.18 0.81 0.018GBYE01019654 Zincfingerprotein 14.69 3.11 0.038GBYE01017658 Ethylene-responsivetranscriptionfactor 9.10 0.79 0.028GBYE01026109 Proteinphosphatase2C 8.85 0.73 0.011GBYE01048118 DREbindingprotein 8.83 0.95 0.037GBYE01041303 WRKYtranscriptionfactor 8.58 1.75 0.069GBYE01037314 WRKYtranscriptionfactor 8.31 0.30 0.012GBYE01039799 WRKYtranscriptionfactor 6.31 1.59 0.072GBYE01011012 RINGfingerprotein 6.16 0.60 0.024GBYE01016099 WRKYtranscriptionfactor 5.56 0.64 0.040GBYE01007796 LysMtypereceptorkinase 5.39 0.77 0.041GBYE01072738 CC-NBS-IRRresistanceprotein 4.95 0.96 0.059GBYE01078745 NBS-LRRdisease-resistanceprotein 4.94 1.66 0.053GBYE01013735 Proteinphosphatase2C 4.80 0.39 0.032GBYE01022591 Zincfingerprotein 4.66 1.08 0.006GBYE01014926 Abscisicacid8'-hydroxylase 4.62 0.31 0.021GBYE01008127 Ethylene-responsiveelement-bindingprotein 4.50 0.19 0.012GBYE01006033 Mitogen-activatedproteinkinase 4.25 0.43 0.008GBYE01008850 Mitogen-activatedproteinkinase 4.24 0.76 0.064GBYE01000925 Metalionbindingprotein 3.74 1.22 0.022GBYE01025591 1-aminocyclopropane1-carboxylatesynthase 3.73 0.13 0.000GBYE01024450 NACdomainprotein 3.52 0.17 0.009GBYE01050160 NBS-LRRdisease-resistanceprotein 3.48 0.54 0.044GBYE01005208 Ethylene-responsivetranscriptionfactor 3.41 0.18 0.006GBYE01003680 UDP-D-glucuronicacid4-epimerase 3.37 0.59 0.035GBYE01041529 Syntaxin 3.32 0.36 0.006GBYE01050872 WRKYtranscriptionfactor 3.19 0.64 0.022GBYE01001472 UDP-glucuronate5-epimerase 3.14 0.37 0.003GBYE01052680 Octicosapeptide/Phox/Bem1p(PB1) 3.12 0.77 0.031GBYE01026684 MYBtranscriptionfactor 3.09 0.84 0.121GBYE01025728 AP2domain-containingtranscription 3.02 0.26 0.040GBYE01006842 Calcium-bindingEF-handprotein 2.96 0.19 0.028GBYE01014767 RINGfingerprotein 2.79 0.30 0.026GBYE01000421 beta-1,3-glucanase 2.79 0.83 0.138GBYE01012536 Lectinproteinkinasefamilyprotein 2.78 0.39 0.006GBYE01004300 WRKYtranscriptionfactor 2.67 0.19 0.003GBYE01007399 Calmodulinbindingprotein 2.65 0.42 0.079GBYE01000327 Chitinase 2.58 0.83 0.152GBYE01071204 DREbindingprotein 2.57 0.83 0.054GBYE01006511 Calcium-bindingprotein 2.56 0.04 0.006GBYE01005028 WRKYtranscriptionfactor 2.53 0.23 0.017GBYE01010007 NACdomainprotein 2.46 0.09 0.005GBYE01041899 AP2/ERFdomain-containingprotein 2.45 0.46 0.061GBYE01018782 Auxin-inducedprotein 2.44 0.62 0.097GBYE01007979 AP2domainclasstranscriptionfactor 2.44 0.48 0.024GBYE01019214 Proteinphosphatase2C 2.43 0.62 0.050GBYE01007604 UDP-glucosedehydrogenase 2.29 0.44 0.033GBYE01008935 WRKYtranscriptionfactor 2.25 0.17 0.004GBYE01036775 AP2/ERFdomain-containingprotein 2.24 0.46 0.074GBYE01030298 MYBtranscriptionfactor 2.23 0.64 0.153GBYE01005328 Calcium-bindingprotein 2.23 0.42 0.021GBYE01016418 Ser/threproteinkinase 2.22 0.12 0.016GBYE01000487 Calmodulin 2.22 0.25 0.024GBYE01014032 Auxin-inducedprotein 2.16 0.52 0.084GBYE01013774 CytochromeP450 2.10 0.67 0.171GBYE01043450 Zincfingerprotein 2.08 0.13 0.021GBYE01007181 E3ubiquitinligase 2.07 0.35 0.019GBYE01053827 WRKYtranscriptionfactor 2.06 0.17 0.049GBYE01007398 Syntaxin 2.05 0.44 0.120
Page 45 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research

Draft
Fig. S1. Verification of expression levels obtained from the NanoString nCounter analysis by qRT-PCR. The gene expression levels were verified in five sequences at three different time points 10, 30, and 60 min following the mechanical stress treatment. It shows the correlation between the data obtained from the two methods.
0.00
10.00
20.00
30.00
40.00
50.00
60.00
70.00
80.00
90.00
RINGfingerprotein DREbindingprotein WRKYtranscriptionfactor
Zincfingerprotein NBS-LRRdisease-resistanceprotein
Foldch
ange
Putativefunctions
10minnCounter
10minqRT-PCR
30minnCounter
30minqRT-PCR
60minnCounter
60minqRT-PCR
Page 46 of 46
https://mc06.manuscriptcentral.com/cjfr-pubs
Canadian Journal of Forest Research