thiobacillus - journal of bacteriologyjb.asm.org/content/85/3/595.full.pdf · isolation...
TRANSCRIPT

ISOLATION AND PROPERTIES OF AN IRON-OXIDIZING THIOBACILLUS
W. E. RAZZELL1 AND P. C. TRUSSELLBritish Columbia Research Council, University of British Columbia, Vancouver, Canada
Received for publication 9 October 1962
ABSTRACT
RAZZELL, W. E. (British Columbia ResearchCouncil, Vancouver, Canada) AND P. C. TRUS-SELL. Isolation and properties of an iron-oxidizingThiobacillus. J. Bacteriol. 85:595-603. 1963.-An organism isolated from acidic copper-leachingwaters has been shown to oxidize ferrous ions,sulfur, and metallic sulfides but exhibit peculiarresponses to thiosulfate. The name Thiobacillusferrooxidans has been used to describe it. A pHof 2.5 is optimal for growth on iron, sulfur, andmetallic sulfides, but cells free from iron can beobtained from growth at pH 1.6, and sulfurcultures adjusted to pH 5.5 readily attain a pHof 1.8. A stationary cultivation procedure ap-pears superior to percolation techniques forstudying the oxidation of finely divided metallicsulfides. Concentrations of soluble copper inexcess of 1 g per liter were obtained from chal-copyrite in less than 4 weeks. Chalcocite oxida-tion proceeded in the absence of iron. Sodiumchloride inhibits iron oxidation without pre-venting oxidation of metallic sulfides by theorganism.
Within the past few years, a number of re-ports have appeared on the isolation and proper-ties of acidophilic bacteria capable of oxidizingferrous ions. The first of these organisms, de-scribed in detail by Colmer, Temple, and Hinkle(1950) and Temple and Colmer (1951), was as-signed the name Thiobacillus ferrooxidans. Testswith finely ground pyrite (Temple and Del-champs, 1953; Bryner et al., 1954) revealedthat these bacteria could rapidly oxidize thesolid metallic sulfide and confirmed the observa-tion of Leathen, Braley, and McIntyre (1953)that T. thiooxidans could not. A similar organismwas described by Leathen and Braley (1954) andLeathen, Kinsel, and Braley (1956) and namedFerrobacillus ferrooxidans because, in contrast to
1 Present address: Syntex Institute for Molecu-lar Biology, Palo Alto, Calif.
T. ferrooxidans, it could not be shown to oxidizesulfur or thiosulfate. Still another iron oxidizerwas named F. .sulfooxidans by Kinsel (1960)because this organism could oxidize sulfur andiron. Apparent oxidation of thiosulfate by thisorganism was attributed to chemical generationof sulfur at low pH and subsequent biologicaloxidation of the sulfur.
Because their isolates oxidized ferrous ions,sulfur, and metallic sulfides, Bryner and Jameson(1958) suggested that the strains were similar toT. ferrooxidans. The isolates studied here likewisecorrespond to T. ferrooxidans and, until evidenceproves otherwise, have been considered isolatesof this described species.
Detailed biochemical studies on the growthand metabolism of T. ferrooxidans were madeby Silverman and Lundgren (1959a, b), who re-ported a Qo, (N) of 4,516 for freshly harvestedbacteria acting on ferrous sulfate, and calculatedan efficiency of 16.2 to 25% for CO2 fixation.Observations by Ljalikova (1958) on the effi-ciency of CO2 fixation indicated a maximum of24 to 34% with 3-day cultures and 13 to 15%with 14-day cultures. Beck (1960) was able todemonstrate sulfur oxidation in a Warburgapparatus by cells grown on a mixture of ironand sulfur, but the characteristics of sulfuroxidation by cells grown solely on iron, or studieson metallic sulfides, were not mentioned.The results reported here establish further the
ubiquitous distribution of T. ferrooxidans andthe conditions for oxidation of iron, sulfur, andmetallic sulfides by this organism.
MATERIALS AND METHODS
Media. Except for preliminary enrichmentand mineral-oxidation experiments with filter-sterilized mine water, the inorganic salts mediumof Silverman and Lundgren (1959a; medium9K, minus the FeSO4) was used throughout thiswork, with FeSO4, S, Na2S203, or metallicsulfides added as energy sources. Silica gelmedium for isolation studies was prepared by
595
on Septem
ber 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

RAZZELL AND TRUSSELL
mixing the free silicic acid (Leathen et al., 1956)with an equal volume of 9K inorganic saltsmedium containing 4% FeSO4 * 7H20. Platespoured with this mixture set firmly after auto-claving for 7 min at 15 psi. Although some browndiscoloration was observed occasionally, thisdid not interfere with colony formation. All pHadjustments were made with sulfuric acid orammonium hydroxide.
Analytical procedures. Total soluble iron (Fe++and Fe+) was determined spectrophotometri-cally with potassium thiocyanate (Sandell,1959). To a sample containing 5 to 60 ,ug oftotal iron in 4 ml of water were added 0.5 ml of6 N HCl and 0.05 ml of 0.6% hydrogen peroxide.After the solution had been thoroughly mixed,0.5 ml of 2% potassium thiocyanate was added,the solution remixed, and the absorption at440 m, determined immediately. A standardcurve was prepared by use of ferrous ammoniumsulfate. Soluble ferric ion was determined asabove, without peroxide, and the ferrous ion bydifference.
Soluble copper was measured spectropho-tometrically by extraction of the diphenyldithio-carbazone complex from 0.01 N hydrochloricacid into carbon tetrachloride (Sandell, 1959).
Sulfate was determined gravimetrically byprecipitation as barium sulfate.
Nitrogen content of bacteria, washed by re-peated centrifugations in distilled water (pH2.5 with sulfuric acid), was determined aftermicro-Kjeldahl digestion by direct nesslerization(using cell suspensions) or microdiffusion (Con-way, 1947).
Cultivation of bacteria. Special techniques wererequired for cultures on sulfur and metallicsulfides. Sulfur cultures were prepared by sprin-kling 0.5 g of flowers of sulfur on 50 ml of 9Kinorganic salts medium (pH 2.8) in 300-mlscrew-capped bottles. The bottles were sterilizedby steaming for 20 min on 3 successive days.Metallic sulfide cultures consisted of 50 ml of9K salts medium (pH 2.5) plus 0.5 g of minus325-mesh mineral (<44 A diam) autoclaved in300-ml bottles. These bottled media are referredto as "stationary cultures."
Percolators (Bryner et al., 1954) contained200 g of acid-washed Ottawa sand intimatelymixed with 2.5 to 10 g of mineral (usually -325mesh), supported on a pad of glass wool, plus
250 ml of 9K salts medium (pH 2.5). The perco-lator and medium were sterilized separately byautoclaving. The air passing through the perco-lators was saturated by bubbling through acolumn of water and then sterilized by passagethrough a column packed with sterile nonab-sorbent cotton.The effect of a variety of solutes on iron oxida-
tion by cell suspensions was rapidly determined,as follows. Cells were harvested (see below) fromiron cultures grown at pH 1.6. A 50-,uliterquantity of 9K salts medium containing the cellswas added to 0.33 ml of solution under test, atpH 2.5, plus 20 ,uliters of 9K salts medium con-taining 4% FeSO4 * 7H20, at 37 C. After aconvenient time, the reaction was stopped byadding 0.5 ml of 6 N HCl, and the ferric ioncontent determined as above. Controls were runat zero time and for a time corresponding to thelength of the actual determination, withoutbacteria.
RESULTS
Isolation of the organism. The strain of T.ferrooxidans used in most of the experimentswas isolated from percolator liquid after thepercolator containing chalcopyrite (CuFeS2,mineralogical grade) had been inoculated with5 ml of acidic mine water and the liquid circu-lated for 39 days. Loopfuls of the percolatingsolution were spread on silicic acid-ferroussulfate plates. From these plates, 22 colonieswere picked, and all yielded colonies on a silicicacid-ferrous sulfate subculture. Subcultures offive colonies have been maintained by biweeklytransfers in tubes containing 10 ml of Silvermanand Lundgren (1959a) medium.More recently, direct inoculations of ferrous
sulfate-9K medium with water from a number ofcommercial copper-leaching operations in BritishColumbia, Arizona, and from the Rio Tintomine, Huelva, Spain, yielded active iron-oxidiz-ing cultures after 120 hr of shaking at 35 C. Theorganism in each case has been isolated as aboveand has been a short, gram-negative rod, withthe properties of the original isolate maintainedin our laboratories.
Preparation of cell suspensions. Active cellswere obtained by resuspending colonies fromsilicic acid-FeSO4 plates in distilled water atpH 2.5, allowing the suspension to stand for
596 J. BACTERIOL.
on Septem
ber 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
VOL. 85, 1963 ISOLATION AND PROPERTIES OF T. FERROOXIDANS
30 min, and recovering bacteria from the super-natant by centrifugation at 500 X g for 10 min.The sedimented cells were suspended in a 1:10dilution of 9K salts solution (pH 2.5). Subse-quently, cells from liquid cultures were demon-strated to be equally active, so that inoculapropagated in shaken flasks of liquid 9K-FeSO4medium at pH 2.5 or 1.6 were used routinely inlater experiments.
Iron oxidation. The ability of the organisms touse as energy sources 1 % solutions of variousinorganic ferrous salts in the 9K salts medium(pH 2.5) was measured by the oxidation of theiron to the ferric state, In addition to ferroussulfate, ferrous ammonium sulfate and potas-sium ferrosulfate were oxidized, but ferrouschloride and ferrous nitrate were not. Sub-cultures from ferrous chloride and ferrous nitratecultures into ferrous sulfate medium did notgrow. The addition of sodium or ammoniumchloride to shake flasks containing ferrous sulfaterevealed that 0.14 M chloride was toxic to thesebacteria, as was nitrate (see below).
8 -
Ea.C.
+
0
IF- 0
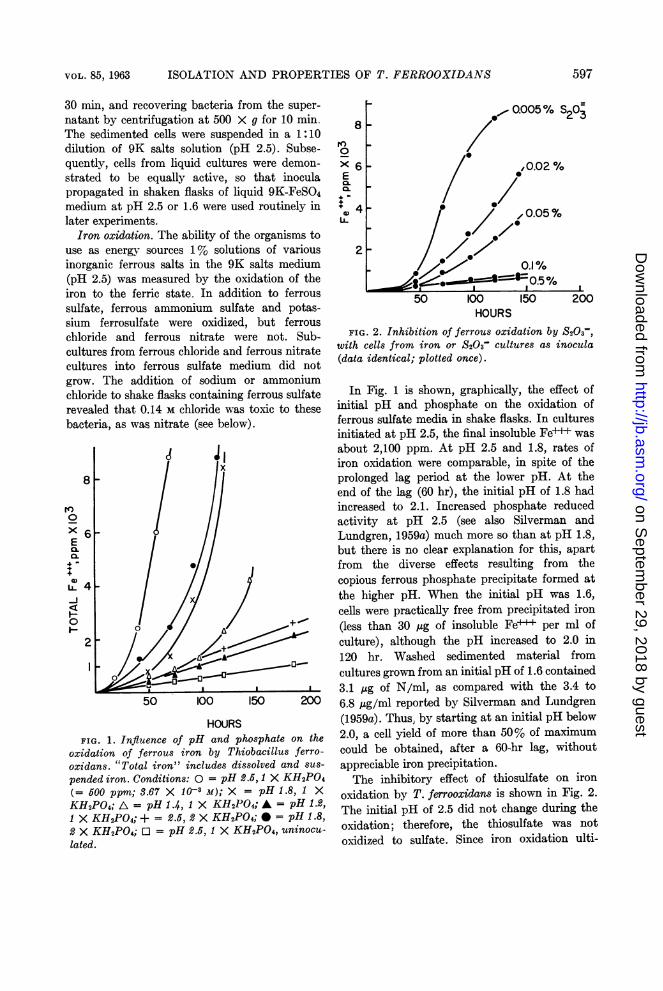
HOURSFIG. 1. Influence of pH and phosphate on the
oxidation of ferrous iron by Thiobacillus ferro-oxidans. "Total iron" includes dissolved and sus-
pended iron. Conditions: 0 = pH 2.5, 1 X KH2PO4(= 500 ppm; 3.67 X 10-3 M); X = pH 1.8, 1 XKH2PO4; A = pH 1.4, 1 X KH2PO4; * = pH 1.2,1 X KH2PO4; + = 2.5, 2 X KH2PO4; * = pH 1.8,2 X KH2PO4; O = pH 2.5, 1 X KH2PO4, uninocu-lated.
to0XEC0
LLU.
0.005% S2O3
,0.05%
50 100 150 200HOURS
FIG. 2. Inhibition of ferrous oxidation by S203-,with cells from iron or S203- cultures as inocula(data identical; plotted once).
In Fig. 1 is shown, graphically, the effect ofinitial pH and phosphate on the oxidation offerrous sulfate media in shake flasks. In culturesinitiated at pH 2.5, the final insoluble Fe-+ wasabout 2,100 ppm. At pH 2.5 and 1.8, rates ofiron oxidation were comparable, in spite of theprolonged lag period at the lower pH. At theend of the lag (60 hr), the initial pH of 1.8 hadincreased to 2.1. Increased phosphate reducedactivity at pH 2.5 (see also Silverman andLundgren, 1959a) much more so than at pH 1.8,but there is no clear explanation for this, apartfrom the diverse effects resulting from thecopious ferrous phosphate precipitate formed atthe higher pH. When the initial pH was 1.6,cells were practically free from precipitated iron(less than 30 ,ug of insoluble Fe+++ per ml ofculture), although the pH increased to 2.0 in120 hr. Washed sedimented material fromcultures grown from an initial pH of 1.6 contained3.1 ,ug of N/ml, as compared with the 3.4 to6.8 Ag/ml reported by Silverman and Lundgren(1959a). Thus, by starting at an initial pH below2.0, a cell yield of more than 50% of maximumcould be obtained, after a 60-hr lag, withoutappreciable iron precipitation.The inhibitory effect of thiosulfate on iron
oxidation by T. ferrooxidans is shown in Fig. 2.The initial pH of 2.5 did not change during theoxidation; therefore, the thiosulfate was notoxidized to sulfate. Since iron oxidation ulti-
597
I
on Septem
ber 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
RAZZELL AND TRUSSELL
mately occurred at lower thiosulfate levels, thecells were inhibited rather than killed. Separateexperiments with the rapid method of measuringiron oxidation rates, using cell suspensions,showed that thiosulfate immediately inhibitediron oxidation. The inhibitory molecule could bea metabolic product of the bacterial action on thethiosulfate, but is clearly not solely a reductionof Fe++ to Fe+ by thiosulfate, since even 0.1%S203= is not stoichiometrically equivalent to10,000 ppm of Fe+++.At pH 2.5, 0.43 to 0.68 ,ug of cell N was formed
per mg of Fe- oxidized. Calculations from dataobtained as described below indicate values forsulfur and chalcopyrite of 10 ,tg of N per mg of Sand 4.8 Ag of N per mg of CuFeS2, respectively,and probably reflect a larger number of energy-generating steps in electron transport from sulfuror sulfides. These results are at variance withthose of Beck (1960), who concluded that theenergy yields from iron and sulfur were approxi-mately the same.The effects of some compounds on iron oxida-
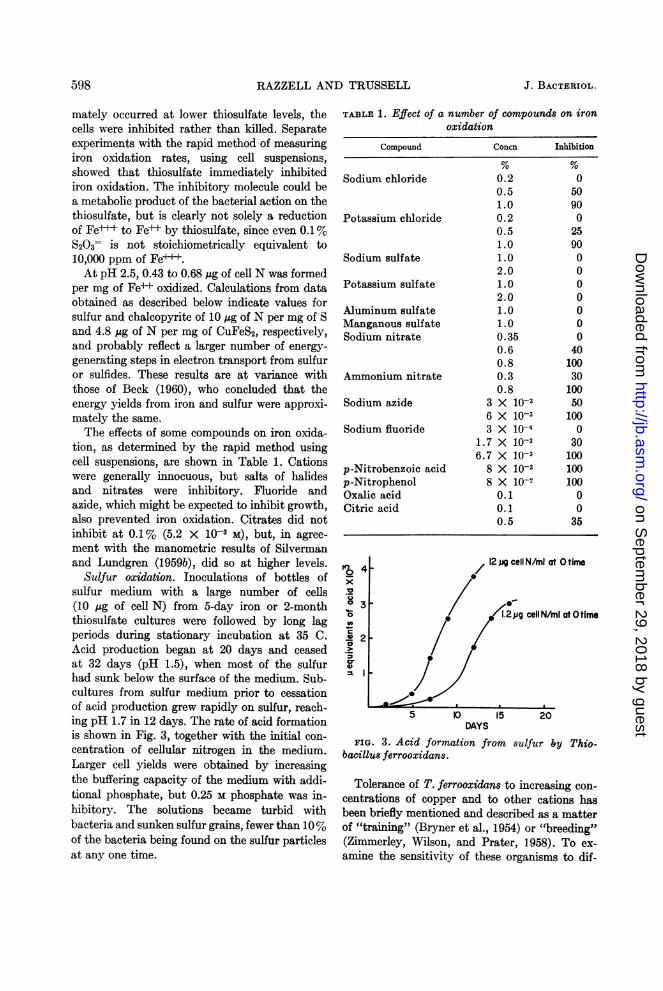
tion, as determined by the rapid method usingcell suspensions, are shown in Table 1. Cationswere generally innocuous, but salts of halidesand nitrates were inhibitory. Fluoride andazide, which might be expected to inhibit growth,also prevented iron oxidation. Citrates did notinhibit at 0.1% (5.2 X 103 M), but, in agree-ment with the manometric results of Silvermanand Lundgren (1959b), did so at higher levels.
Sulfur oxidation. Inoculations of bottles ofsulfur medium with a large number of cells(10 Mg of cell N) from 5-day iron or 2-monththiosulfate cultures were followed by long lagperiods during stationary incubation at 35 C.Acid production began at 20 days and ceasedat 32 days (pH 1.5), when most of the sulfurhad sunk below the surface of the medium. Sub-cultures from sulfur medium prior to cessationof acid production grew rapidly on sulfur, reach-ing pH 1.7 in 12 days. The rate of acid formationis shown in Fig. 3, together with the initial con-centration of cellular nitrogen in the medium.Larger cell yields were obtained by increasingthe buffering capacity of the medium with addi-tional phosphate, but 0.25 M phosphate was in-hibitory. The solutions became turbid withbacteria and sunken sulfur grains, fewer than 10%of the bacteria being found on the sulfur particlesat any one time.
TABLE 1. Effect of a number of compounds on ironoxidation
Compound
Sodium chloride
Potassium chloride
Sodium sulfate
Potassium sulfate
Aluminum sulfateManganous sulfateSodium nitrate
Ammonium nitrate
Sodium azide
Sodium fluoride
p-Nitrobenzoic acidp-NitrophenolOxalic acidCitric acid
Tc.D
a0o
0D-4-
0>ca
Concn
0.20.51.00.20.51.01.02.01.02.01.01.00.350.60.80.30.8
3 X 10-36 X 10-33 X 10-4
1.7 X 10-36.7 X 10-3
8 X 10-38 X 10-2
0.10.10.5
Inhibition
05090025900000000
401003010050100
030100100100
0035
5 IO 15 20DAYS
FIG. 3. Acid formation from sulfur by Thio-bacillus ferrooxidans.
Tolerance of T. ferrooxidans to increasing con-centrations of copper and to other cations hasbeen briefly mentioned and described as a matterof "training" (Bryner et al., 1954) or "breeding"(Zimmerley, Wilson, and Prater, 1958). To ex-amine the sensitivity of these organisms to dif-
598 J. BACTERIOL.
on Septem
ber 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

VOL. 85, 1963 ISOLATION AND PROPERTIES OF T. FERROOXIDANS
TABLE 2. Effect of ions on sulfur oxidation by Thiobacillus ferrooxidans
Incubation (days)*Addition Amt Concn
12 16 23 40
ppmt moles/liter
None - - 2.4 1.7 1.5Na2SO4 20,000 0.146 2.4 1.8 1.7NaCl 5,000 0.085 2.3 1.8 1.6
10,000 0.171 2.4 2.2 2.230,000 0.513 2.6 2.5 2.6 2.6 2.6
Cd++ 4,380 0.039 2.3 1.6 1.5Zn++ 2,280 0.035 2.5 1.9 1.9CaSO4 50,000 0.367 2.6 1.8 1.7Fe2(SO4)3 1,000 0.0025 2.1 1.5 1.5Fe2(S04)3, no inoculum 1,000 0.0025 2.8 2.7 2.7 2.7
Ni+t 223 0.0038 2.7 1.8 1.72,230 0.038 2.7 2.6 2.1 1.6
MnH 325 0.006 2.4 1.6 1.6Co++ 380 0.0064 2.8 2.7 2.7 2.7 2.7Cu++ 225 0.00355 2.7 2.7 2.7 2.7 2.7
Ag+ 210 0.00194 2.7 2.7 2.7 2.7 2.7KHAsO4 1,000 0.0056 2.8 2.7 2.7 2.7 2.7NaNO3 10,000 0.118 2.7 2.6 2.6 2.6 2.6Na2S203 200 0.0126 2.9 2.5 2.2 1.9 1.6
500 0.0316 3.4 3.3 3.5 3.5 3.6
* Results expressed as pH readings.
ferent cations, bottles of sulfur mediumcontaining the sulfates of various cations, andsome anions of interest, were inoculated withcells grown on sulfur, and the rate of decrease inpH compared with controls containing no addedions. The data (Table 2) show that cells grownon sulfur were insensitive to sodium but in-hibited by chloride; insensitive to cadmium,zinc, calcium, ferric iron, and low concentra-tions of nickel and manganese; and inhibited bycobalt, copper, silver, arsenate, and nitrate.Partial inhibition of acid formation by S203= wasapparent; determinations of cell N showed thatthis was not caused by concomitant alkali forma-tion but rather by lack of cell multiplication. Thesensitivity to relatively low concentrations ofcopper, cobalt, and manganese and to higherconcentrations of nickel could be overcome iflarger inocula were added, suggesting that asmall fraction of the original cell population wasinherently resistant to the ions. In separate ex-periments, subcultures of cells grown from largeinocula, through several subcultures in the
presence of high concentrations of copper, wereable to grow in concentrations of manganese orcobalt which were toxic to unselected cells, sug-gesting that they possessed a general mechanismfor resisting the action of many cations. The lackof accurate methods for counting colonies of thesebacteria precluded a more detailed study ofgenetic variations.Whereas cells grown on iron did not rapidly
initiate growth on sulfur, even from large inocula,small inocula of cells grown on sulfur (1.2 ,ug ofcell N or 8 X 107 cells) reached maximal growthin 200 ml of iron medium (9 mg of Fe+++/ml)in 1 week. This compared favorably with 3 daysfor an inoculum consisting of a similar numberof cells grown on iron.The pH limits within which T. ferrooxidans
will grow depend on the medium to some extent,since iron salts become insoluble above pH 4.Sulfur oxidation was difficult to measure at lowpH where media were poorly buffered. Silvermanand Lundgren (1959a) could not find a suitablebuffer for iron oxidation, and we found only
599
on Septem
ber 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
RAZZELL AND TRUSSELL
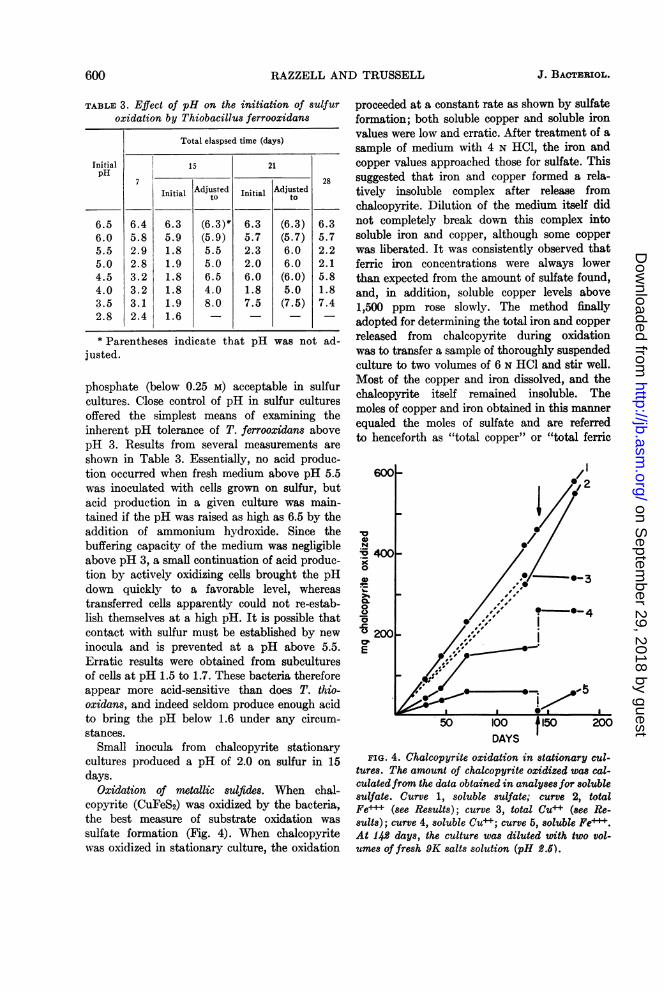
TABLE 3. Effect of pH on the initiation of sulfuroxidation by Thiobacillus ferrooxidans
Total elaspsed time (days)
Initial 15 21pH__
Initial Adjusted Initial Adjustedto to
6.5 6.4 6.3 (6.3)* 6.3 (6.3) 6.36.0 5.8 5.9 (5.9) 5.7 (5.7) 5.75.5 2.9 1.8 5.5 2.3 6.0 2.25.0 2.8 1.9 5.0 2.0 6.0 2.14.5 3.2 1.8 6.5 6.0 (6.0) 5.84.0 3.2 1.8 4.0 1.8 5.0 1.83.5 3.1 1.9 8.0 7.5 (7.5) 7.42.8 2.4 1.6 - _
* Parentheses indicate that pH was not ad-justed.
phosphate (below 0.25 M) acceptable in sulfurcultures. Close control of pH in sulfur culturesoffered the simplest means of examining theinherent pH tolerance of T. ferrooxidans abovepH 3. Results from several measurements areshown in Table 3. Essentially, no acid produc-tion occurred when fresh medium above pH 5.5was inoculated with cells grown on sulfur, butacid production in a given culture was main-tained if the pH was raised as high as 6.5 by theaddition of ammonium hydroxide. Since thebuffering capacity of the medium was negligibleabove pH 3, a small continuation of acid produc-tion by actively oxidizing cells brought the pHdown quickly to a favorable level, whereastransferred cells apparently could not re-estab-lish themselves at a high pH. It is possible thatcontact with sulfur must be established by newinocula and is prevented at a pH above 5.5.Erratic results were obtained from subculturesof cells at pH 1.5 to 1.7. These bacteria thereforeappear more acid-sensitive than does T. thio-oxidans, and indeed seldom produce enough acidto bring the pH below 1.6 under any circum-stances.
Small inocula from chalcopyrite stationarycultures produced a pH of 2.0 on sulfur in 15days.
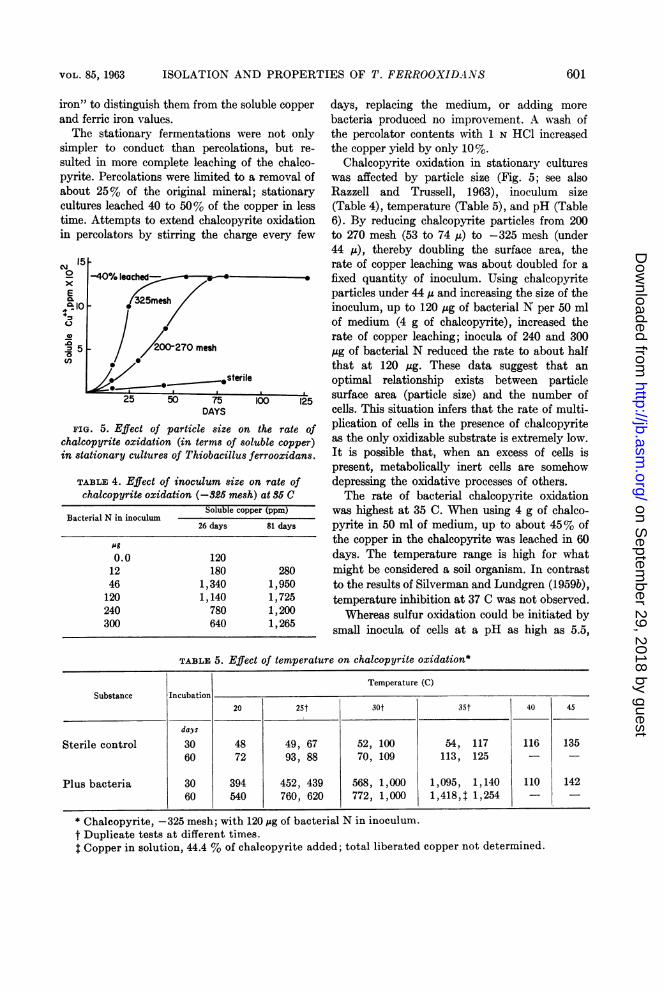
Oxidation of metallic sulfides. When chal-copyrite (CuFeS2) was oxidized by the bacteria,the best measure of substrate oxidation wassulfate formation (Fig. 4). When chalcopyritewas oxidized in stationary culture, the oxidation
proceeded at a constant rate as shown by sulfateformation; both soluble copper and soluble ironvalues were low and erratic. After treatment of asample of medium with 4 N HCI, the iron andcopper values approached those for sulfate. Thissuggested that iron and copper formed a rela-tively insoluble complex after release fromchalcopyrite. Dilution of the medium itself didnot completely break down this complex intosoluble iron and copper, although some copperwas liberated. It was consistently observed thatferric iron concentrations were always lowerthan expected from the amount of sulfate found,and, in addition, soluble copper levels above1,500 ppm rose slowly. The method finallyadopted for determining the total iron and copperreleased from chalcopyrite during oxidationwas to transfer a sample of thoroughly suspendedculture to two volumes of 6 N HCl and stir well.Most of the copper and iron dissolved, and thechalcopyrite itself remained insoluble. Themoles of copper and iron obtained in this mannerequaled the moles of sulfate and are referredto henceforth as "total copper" or "total ferric
600_ /I 2
X400 -
0~~~~~~~~
200
E ,,,"
FIG. 4. Chalcopyrite oxidation in stationary cul-tures. The amount of chalcopyrite oxidized was cal-culated from the data obtained in analyses for solublesulfate. Curve 1, soluble sulfate; curve 2, totalFe" (see Results); curve 3, total Cu+ (see Re-sults); curve 4, soluble Cu' ; curve 5, soluble Fe++.At 142 days, the culture was diluted with two vol-umes of fresh 9K salts solution (pH 2.5).
600 J. BACTERIOL.
on Septem
ber 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
VOL. 85, 1963 ISOLATION AND PROPERTIES OF T. FERROOXIDANS
iron" to distinguish them from the soluble copper
and ferric iron values.The stationary fermentations were not only
simpler to conduct than percolations, but re-
sulted in more complete leaching of the chalco-pyrite. Percolations were limited to a removal ofabout 25% of the original mineral; stationarycultures leached 40 to 50% of the copper in lesstime. Attempts to extend chalcopyrite oxidationin percolators by stirring the charge every few
clJ 150
xE
.*C 10
0)
=5 5a3)
-40% leached- -
325mesh
200-270 mesh
:7 ssterile
25 50 75 100 125DAYS
FIG. 5. Effect of particle size on the rate ofchalcopyrite oxidation (in terms of soluble copper)in stationary cultures of Thiobacillus ferrooxidans.
TABLE 4. Effect of inoculum size on rate ofchalcopyrite oxidation (-&25 mesh) at 86 C
Soluble copper (ppm)Bacterial N in inoculum
26 days 81 days
Ag
0.0 12012 180 28046 1,340 1,950120 1,140 1,725240 780 1,200300 640 1,265
days, replacing the medium, or adding more
bacteria produced no improvement. A wash ofthe percolator contents with 1 N HCl increasedthe copper yield by only 10%.
Chalcopyrite oxidation in stationary cultureswas affected by particle size (Fig. 5; see alsoRazzell and Trussell, 1963), inoculum size(Table 4), temperature (Table 5), and pH (Table6). By reducing chalcopyrite particles from 200to 270 mesh (53 to 74 ju) to -325 mesh (under44 Iu), thereby doubling the surface area, therate of copper leaching was about doubled for a
fixed quantity of inoculum. Using chalcopyriteparticles under 44 ju and increasing the size of theinoculum, up to 120 jug of bacterial N per 50 mlof medium (4 g of chalcopyrite), increased therate of copper leaching; inocula of 240 and 300jug of bacterial N reduced the rate to about halfthat at 120 ,ug. These data suggest that an
optimal relationship exists between particlesurface area (particle size) and the number ofcells. This situation infers that the rate of multi-plication of cells in the presence of chalcopyriteas the only oxidizable substrate is extremely low.It is possible that, when an excess of cells ispresent, metabolically inert cells are somehowdepressing the oxidative processes of others.The rate of bacterial chalcopyrite oxidation
was highest at 35 C. When using 4 g of chalco-pyrite in 50 ml of medium, up to about 45% ofthe copper in the chalcopyrite was leached in 60days. The temperature range is high for whatmight be considered a soil organism. In contrastto the results of Silverman and Lundgren (1959b),temperature inhibition at 37 C was not observed.Whereas sulfur oxidation could be initiated by
small inocula of cells at a pH as high as 5.5,
TABLE 5. Effect of temperature on chalcopyrite oxidation*
Temperature (C)Substance Incubation
20 25t 30t 35t 40 45
days
Sterile control 30 48 49, 67 52, 100 54, 117 116 13560 72 93, 88 70, 109 113, 125 -
Plus bacteria 30 394 452, 439 568, 1,000 1,095, 1,140 110 14260 540 760, 620 772, 1,000 1,418,4 1,254 -
* Chalcopyrite, -325 mesh; with 120 ,ug of bacterial N in inoculum.t Duplicate tests at different times.t Copper in solution, 44.4 % of chalcopyrite added; total liberated copper not determined.
601
on Septem
ber 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

~~~~RAZZELL AND TRUSSELLJ.Bc
biological chalcopyrite oxidation did not occurat pH 5. At pH 5, most of the cells were not at-tached to chalcopyrite particles, whereas, atpH 4 to 2, most cells were associated with thechalcopyrite particles.
Oxidation of acid-washed CuS (covellite) instationary culture by these bacteria was ap-parent, in spite of spontaneous chemical oxida-tion (Table 7). MNore than 80% of the mineralwas dissolved by bacteria in 6 months in thevirtual absence of iron (less than 12 ppm), andat a constant pH of 2.6.
Thiosulfate oxidation. Growth of the cells onthiosulfate was difficult to assess, owing to de-composition of thiosulfate below pH 4. Thepossibility that chemical formation of sulfur atlow pH provides an oxidizable substrate for thebacteria has been pointed out by Leathen andBraley (1955). Although growth in Starkey's0.5% thiosulfate medium has been erratic inthis laboratory, cultures have been maintained
TABLE 6. Effect of pH on chalcopyrite oxidation andcell settlement
Suspended Soluble copper (ppm)pH celgI
at 5 days 5 days 12 days 25 days
5 1.2 12 36 384 0.24 80 260 4903.5 0.36 115 280 5003.0 0.28 125 350 4802.5 0.28 190 350 5002.0 0.48 120 290 400
*The pH was adjusted daily by the addition ofdilute H2S04 or NH40H as required. Incubatedat room temperature.
t Stationary bottles, containing 50 ml of in-organic salts and inoculum but no chalcopyrite,contained 1.6 p.g of cell N/ml after standing thesame length of time as required to settle chalco-pyrite in experimental bottles.
in this medium by serial transfer for more than
1 year, and have retained their ability to oxidize
ferrous iron. Recent studies by Colmer (1962)
confirmed these observations. In contrast to T.
thiooxidans, which caused a drop in pH to 2.0
in 30 days, T. ferrooxidans caused a rise in pH
to 5.5 to 6.0 in 30 days, followed by a drop to
pH 3.5 to 4 in the subsequent month. Tests for
tetrathionate (Vishniac and Santer, 1957) were
strongly positive throughout the period of ele-
vated pH, and were still positive when all the
iodine-titrable thiosulfate had disappeared. As
a basis for further experimentation, it seems
reasonable to assume that tetrathionate is
rapidly formed from thiosulfate, thereby ac-
counting for the rise in pH, but is inhibitory to
our strains. Low concentrations of thiosulfate
inhibit sulfur and iron oxidation but, since this
action does not show any lag, it is probably direct
and not via tetrathionate.
DiscussioN
Isolation of the iron-oxidizing organism from
mine water proved difficult, since the obligate-
autotrophic nature of this organism precluded
its growth on agar media. Single-colony growth,
in our hands, was obtained only on silicic acid
plates.
The distribution of this organism is quite
widespread in nature but is dependent upon an
acidic environment in which ferrous iron, sulfur,or inorganic sulfide is present. Without doubt,
this organism has been associated closely with
the leaching of copper from chalcopyrite piles
at the Rio Tinto mine, Spain. It is interesting to
observe that the temperature, pH, and oxygen
levels found optimal for leaching in the present
studies correspond to those conditions arrived at
by decades of experience at this mine, as reported
by Taylor and Whelan (1943).
For laboratory experimental work, the sta-
tionary cultures have proven preferable to perco-
TABLE, 7. Oxidation of covellite (CuS) by Thiobacillus ferrooxidans
Culture ~~~~~~~~~~Soluble copper (ppm) at (days)9 48 76 126 126* 178 178*
Sterile control 624 795 1,200 1,000 1,330 1,100 1,650Inoculated 720 3,500 3,900 5,200 6,960 7,700 8,100
* Total copper, other than as CuS, determined by transferring 1 ml of entire suspension includingmineral to 2 ml of 6 N HCI and assaying the supernatant after 1 hr.
602 J. BACTERIOL.
on Septem
ber 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

VOL. 85, 1963 ISOLATION AND PROPERTIES OF T. FERROOXIDANS
lators, providing more reproducible results,more complete and more rapid leaching, andgreater convenience.The adaptability of these organisms to metallic
ions is quite surprising. For example, by serialtransfer it has been possible to show iron oxida-tion in a solution containing both 5% iron sulfateand 5% copper sulfate. A direct attack by thesebacteria on the metallic sulfides may be assumedto occur; oxidation of CuS has been shown toproceed in the virtual absence of iron, andBryner et al. (1954) also observed oxidation ofCu2S when iron was presumably absent.The ability of these organisms to resist in-
fluences commonly toxic to other forms of lifesuggests an opportunity to investigate mecha-nisms of cellular resistance to chemical damagewhich could hardly be duplicated with other cells.The apparently wide distribution of these bac-teria establishes them as a probable factor insoil and rock transformations at low pH, leadingto the solution of metallic sulfides which mayaffect plant and estuarine life, and which isknown to be of economic importance in the min-ing industry (Razzell and Trussell, 1963).
LITERATURE CITEDBECK, J. V. 1960. A ferrous-iron-oxidizing bac-
terium. I. Isolation and some general physio-logical characteristics. J. Bacteriol. 79:502-509.
BRYNER, L. C., J. V. BECK, D. B. DAVIS, AND
D. G. WILSON. 1954. Microorganisms inleaching sulfide minerals. Ind. Eng. Chem.46:2587-2592.
BRYNER, L. C., AND A. K. JAMESON. 1958. Micro-organisms in leaching sulfide minerals. Appl.Microbiol. 6:281-287.
COLMER, A. R. 1962. Relation of the iron oxidizer,Thiobacillus ferrooxidans, to thiosulfate. J.Bacteriol. 83:761-765.
COLMER, A. R., K. L. TEMPLE, AND M. E. HINKLE.1950. An iron-oxidizing bacterium from theacid drainage of some bituminous coal mines.J. Bacteriol. 59:317-328.
CONWAY, E. J. 1947. Microdiffusion analysis andvolumetric error. Crosby, Lockwood and Son,London.
KINSEL, N. A. 1960. New sulfur oxidizing ironbacterium: Ferrobacillus sulfooxidans sp. n.J. Bacteriol. 80:628-632.
LEATHEN, W. W., AND S. A. BRALEY. 1954. A new
iron-oxidizing bacterium: Ferrobacillus ferro-oxidans. Bacteriol. Proc., p. 44.
LEATHEN, W. W., AND S. A. BRALEY. 1955. In-terpretation of reactions in acid thiosulfatemedia. J. Bacteriol. 69:481.
LEATHEN, W. W., S. A. BRALEY, AND L. D. Mc-INTYRE. 1953. The role of bacteria in theformation of acid from certain sulfuriticconstituents associated with bituminous coal.II. Ferrous iron oxidizing bacteria. Appl.Microbiol. 1:65-68.
LEATHEN, W. W., N. A. KINSEL, AND S. A. BRALEY.1956. Ferrobacillus ferrooxidans: a chemo-synthetic autotrophic bacterium. J. Bacteriol.72:709-704.
LJALIKOVA, M. N. 1958. A study of the processof chemosynthesis by Thiobacillus ferro-oxidans. Mikrobiologiya 27:556-559.
RAZZELL, W. E., AND P. C. TRUSSELL. 1963.Microbiological leaching of metallic sulfides.Appl. Microbiol. 11:105-110.
SANDELL, E. B. 1959. Colorimetric determinationof traces of metals, 3rd ed. IntersciencePublishers, New York.
SILVERMAN, M. P., AND D. G. LUNDGREN. 1959a.Studies on the chemoautotrophic iron bacte-rium Ferrobacillus ferrooxidans. I. An im-proved medium and a harvesting procedurefor securing high cell yields. J. Bacteriol. 77:642-647.
SILVERMAN, M. P., AND D. G. LUNDGREN. 1959b.Studies on the chemoautotrophic iron bac-terium Ferrobacillus ferrooxidans. II. Mano-metric studies. J. Bacteriol. 78:326-331.
TAYLOR, J. H., AND P. F. WHELAN. 1943. Theleaching of cupreous pyrites and the precipita-tion of copper at Rio Tinto, Spain. Trans.Inst. Mining Met. 52:36-96.
TEMPLE, K. L., AND A. R. COLMER. 1951. Theautotrophic oxidation of iron by a new bac-terium: Thiobacillus ferrooxidans. J. Bac-teriol. 62:605-611.
TEMPLE, K. L., AND E. W. DELCHAMPS. 1953.Autotrophic bacteria and the formation ofacid in bituminous coal mines. Appl. Micro-biol. 1:255-258.
VISHNIAC, W., AND M. SANTER. 1957. The thio-bacilli. Bacteriol. Rev. 21:195-213.
ZIMMERLEY, S. R., D. G. WILSON, AND J. D.PRATER. 1958. Cyclic leaching process em-
ploying iron oxidizing bacteria. U.S. Patent2,829,964.
603
on Septem
ber 29, 2018 by guesthttp://jb.asm
.org/D
ownloaded from