tissue respiration - · pdf filethe production of active form of oxygen ... limited by...
TRANSCRIPT

THE MINISTRY OF HEALTH OF UKRAINE
ZAPORIZHZHIA STATE MEDICAL UNIVERSITY
Tissue respirationand
Oxidative phosphorylation:
their mechanisms of duration;
factors for the regulation
Produced by ass.professor Krisanova N.V., 2017

Substrates of catabolic phase II
Metabolites of Krebs Cycle, etc)
Aerobic oxidation (+O2)
NADH FADH2 Donors
of electrons
ELECTRON TRANSPORT CHAIN
of the inner membrane of mitochondria

Substrate (SH2) oxidation catalyzed
by NAD+-linked dehydrogenase
SH2 SH
+++
N N
(Z) (E)
C
HH
R R
O
NH2 C
O
NH2
H
++
2 e
NAD+
NADH
-
NAD+-dehydrogenases are found in
cytoplasm and in the matrix of mitochondria

ELECTRON TRANSPORT CHAIN (ETC)
is the sequence of enzymes
and transporters of electrons
(CoQ, Fe-S-Proteins)
promoted the transfer of electrons
from NADH, FADH2
to molecular oxygen O2
During the transport of electrons there is
the duration of reactions with the type:
Reduced A+Oxidized B→Oxidized A + Reduced B

ETC contains four complexes:
Complex I
FMN and Iron-Sulfur (Fe-S)- centers
NADH-coenzyme Q
reductase
(NADH-dehydrogenase)
FAD and Fe-S- centers
Composition
of non-protein parts :
Complex IISuccinate dehydrogenase,
Cytochrome b560
Complex III
Cytochromes b, c1
Fe-S-proteinsHeme containing Fe 2+/ Fe 3+
Complex IV
Cytochrome C oxidase
(Cytochrome aa3)
Heme containing Fe 2+ /Fe 3+
Cu+/Cu 2+

Complex I (FMN and Fe-S-centers) location
NADH-
dehydrogenase
(NADH-coenzyme
Q reductase )
Matrix
Intermembrane
space
Inner membrane
Second name:

Complex I function at the moment
of HADH oxidation
N
N (E)
N
C
NH
C
(E)
R
O
O
N
HN
(Z)
NH
C
NH
C
R
O
O
2e+ +
CH3
CH3
CH3
CH3
2H+
FMN FMNH2
NADH+H+
NADH + H+ + FMN•E → NAD+ + FMNH2•E (1)
FMN• E – NADH-dehydrogenase (oxidized form)

O
O
CH3
OH3C
OH3C
R
OH
OH
CH3
OH3C
O
R
H3C
2H+
2e+ +
FMNH2*E
CoQ CoQH2
Fe-S-proteins
Matrix
Ubuquinone (CoQ) function at the
moment of oxidation of complex I
CoQ + FMNH2•E FMN•E + CoQH2 (2)

Complex III (Cytochromes b, c1,
heme containing Fe 2+ / Fe 3+ )

3+/2+
Heme structure represented
in cytochromes C and C1

The function of complex III
CoQH2 + 2 cyt b (Fe 3+ ) →2 cyt b(Fe 2+) +
+ CoQ + 2H+ (3)
2 cyt b(Fe2+)+2cyt c1(Fe3+)→2cyt b(Fe3+) +
+2 cyt c1(Fe2+) (4)
2 cyt c(Fe3+)+2cyt c1(Fe2+)→2cyt c(Fe2+) +
+ 2 cyt c1(Fe 3+) (5)

Complex IV
or
Cytocrome C
Oxidase
or
Cytochrome
aa3
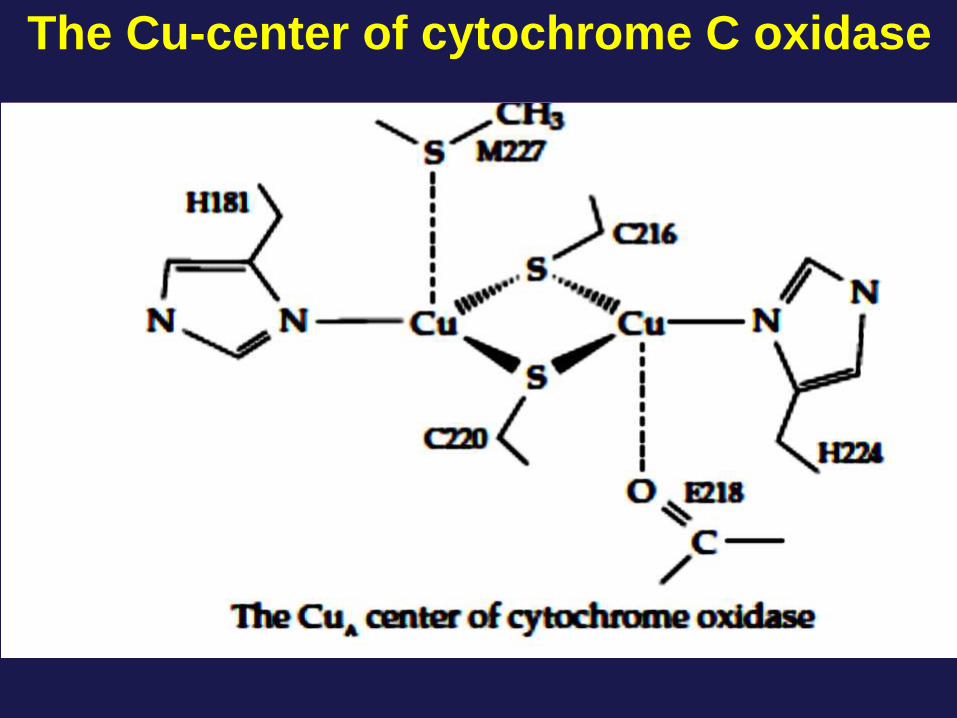
The Cu-center of cytochrome C oxidase

Linkage mechanism of
Oxygen to Cytochrome oxidase

The function of complex IV
cyt aa3(Fe3+, Cu2+) + 2 cyt c(Fe 2+ )
2 cyt c(Fe3+) + cyt aa3 (Fe 2+, Cu+)
(6)
cyt aa3 (Fe 2+, Cu+) + ½O2
cyt aa3(Fe3+, Cu2+)+ O2-
(7)
or:O
2-+ 2H
+ →H2O


Tasks dissolved due to the function
of Electron Transport Chain (ETC)
1. The transport of electrons from reduced derivatives of vitamins (NADH, FADH2) to molecular oxygen.
2. The release of energy at any step of oxidation in ETC.
3. The pumping of protons from the matrix to intermembrane space
4. The production of active form of oxygen - oxide-anion O2- and then H2O

Some cytotoxic particles may be
formed from molecular oxygen
during the tissue respiration
instead of oxide-anion:
Superoxide anion Ó2-
Superoxide radical Ó2
Hydrogen peroxide radical HOÓ
Hydrogen peroxide H2O2

The change of red-ox-potential of
each red-ox pair of ETC is estimated in
volts. This change becomes more and
more in the direction of electron
transport to oxygen. A part of released
energy is thermal energy, and another
one may be used for ATP synthesis in
mitochondria -
this process is named
oxidative phosphorylation.

Notions of chemiosmotic
coupling hypothesis proposed
by Peter Mitchell in 1961
1. The inner membrane is not
permeable for protons in any
site of membrane in the
direction from the
intermembrane space to the
matrix.

2.Some electron carriers act as
pumps, which cause directional
pumping of protons across the
inner membrane from the matrix to
the intermembrane space. As the
electrons move down the chain
protons are expelled, penetrating
from the matrix to the
intermembrane space.

3. Because proton is a charged
particle the flow of free energy
across the inner membrane is due
to the combination of
a concentration gradient (ΔpH),
and a charge gradient (ΔΨ).
The sum of them is electrochemical
potential of the inner membrane:
ΔμH+ = ΔpH + ΔΨ

4. The chemical gradient of protons
across the inner membrane of
mitochondria serves as the means of
coupling the energy flow of electron
transport to the formation of ATP
5. Protons pass back into the matrix at
a special site
where ATP synthetase resides
(proton channel - factor Fo).

The pumps for protons movement
from the matrix to the
intermembrane space
Complex I
Complex III
Complex IV
(proved in 1988)

Inhibitors for ATP synthetase (Factor F0):
Oligomycin and Dicyclohexylcarbodiimide (DCCD)
Fo
F1
ADP
H3PO4
*ATPATP
2H+
2H+
Intr
am
em
bra
ne
sp
ac
e
2H+
Inner membrane Matrix

ATP-synthetase
Matrix
Inner membrane
Intermembrane
space

6. Oxidative phosphorylation is the
synthesis of ATP in the inner membrane of
mitochondria catalyzed by ATP synthetase
due to energy produced by ETC enzymes
7. ATP formation from ADP and phosphoric
acid does not require energy.
ATP is obtained in the linked form with F1.
Energy is required for ATP removal
from F1. This action is due to the moment of
protons movement through the inner
membrane across the Fo as a channel for
protons.

8.The moment of protons movement
through the inner membrane across
the Fo as a channel is the moment
for coupling of phosphorylation (ATP
synthesis) with the change of
electrochemical potential of the
inner membrane (or the use of
energy produced by the ETC).

The P/O ratio signs how many molecules of
ATP is synthesized per 2 electrons
transferred across the ETC to one atom of
molecular oxygen
The P/O ratio
Three sites of coupling in ETC:
1 – site where electrons are transferred from
complex I to CoQ;
2 – site where electrons are transferred from
cytochrome b to cytochrome c1 ;
3 - site where electrons are transferred from
cytochrome c oxidase to molecular oxygen.

P/O ratio values
• Substrates that donate electrons to NAD+ yield P/O ratio of 3
• Substrates that donate electrons to FAD or FMN yield P/O ratio of 2
P/O ratio is changed:
1) under the influence of substances-uncouplers;
2) at the presence of inhibitors of respiratory chain complexes, etc.

Five states of respiratory control have been
defined in which the rate of respiration is
limited by different factors:State 1 is limited by availability of ADP and a sub-
strate (source of electrons).
State 2 is limited by the availability of substrate.
State 3 is limited by the capacity of the electron
chain itself, when ADP, oxygen and substrate are
saturating.
State 4 is limited by the availability of ADP.
State 5 is limited by availability of oxygen.
ADP/ATP transport is due to ATP/ADP translocase
system. Inhibitors of this system are:
1) Atractyloside; 2) Bongkrekic acid.


Inhibitors for complex I:
•Rotenone, an insecticide;
•Barbiturates (drugs for
sleepless treatment);
•Piericidin A, an antibiotic.
Result: NADH is accumulated in
the matrix!
P/O=0 for all NAD+-dependent
oxidation reactions

Inhibitors for complex II:
Malonic acid (1) – the competitive
inhibitor for succinate
dehydrogenase (SDH ase);
Carboxin (2); Thenoyl trifluoride
acetone (TTFA, 3).
Substances (2) and (3) block the
electron transport
from reduced SDHase to CoQ.
P/O = 0 for succinate oxidation.

Inhibitors for complex III:
•Dimercaprol (1);
•Antimycin A, an antibiotic (2)
P/O may be lower then 3 or 2
(respectively oxidized
substrate)

Inhibitors for complex IV:
Carbon monooxide which competes
with oxygen for its binding site on
Cytochrome aa3;
Cyanides block the heme-centers of
cytochrome aa3 binding by covalent
bonds;
Hydrogen sulfide;
Azides .
The death of biological system!

Uncouplersallow the transport of protons
across the inner membrane (they are
very lipophylic compounds), thus
collapsing the proton gradient
before it may be used for ATP
synthesis.
P/O = 0 in all cases !
The energy produced by ETC is
thermal energy, only!

•2,4 - Dinitrophenol (weight-loss drug
used in 1970s but was discontinued
because of its toxicity);
• Dicumarol that is anticoagulant;
•Thyroxin in high abnormal concen-
tration;
•Valinomycin, an antibiotic.
Uncouplers

39
THANK YOUFor
ATTENTION!