tomato research report for 2006-2007 - citrus · tomato research report for 2006-2007 supported by...
TRANSCRIPT

Tomato Research Reportfor
2006-2007
Supported by the Florida Tomato Committee

Institute of Food and Agricultural Sciences (IFAS) 1022 McCarty HallFlorida Agricultural Experiment Station PO Box 110200Office of the Dean for Research Gainesville, FL 32611-0200
Tel: (352) 392-1784Fax: (352) 392-4965
The Foundation for The Gator Nation
MEMORANDUM
TO: The Florida Tomato Committee
FROM: George J. Hochmuth, Associate Dean for Research
SUBJECT: 2006-2007 Research Report
DATE: September 5, 2007
This report describes research by scientists in UF/IFAS/FAES that received support from the
Florida Tomato Committee during the past year. The Florida Tomato Committee support
combines with State and Federal resources to allow IFAS tomato scientists to have a strong and
productive research program focused on the tomato industry of Florida.
This is an extremely important partnership between the Tomato Industry and IFAS scientists.
We have worked together to identify the questions and needs for the industry. Together we have
prioritized the research to be accomplished. This year’s work ranged from food safety and
handling to tomato breeding and methyl bromide alternatives. Many of the results are
immediately applicable by the industry.
We hope to continue in our quest for improving production methods and product quality. We
are pleased to have the Florida Tomato Committee as a partner in programmatic support for
tomato research. On behalf of the scientists involved in tomato research, we thank you and
appreciate your support.
This report of research results is presented in electronic format. We hope you find the
information in the report useful to all facets of tomato production.
GJH:las
cc: Mark McLellan

INDEX
Research supported by FLORIDA TOMATO COMMITTEE
2006-2007 IFAS Research Reports
Page Title Investigator(s)
FOOD SAFETY AND HANDLING
1 Characterization of Genetic Mechanisms that Determine
Contamination of Tomatoes with Salmonella: from Field to the
Packing House
Max Teplitski
Keith R. Schneider
8 Refinements in the Chlorine Dioxide Gas Treatment of Tomatoes
for Controlling Microbial Contamination
Jerry A. Bartz
Keith R. Schneider
Steven A. Sargent
Jeffrey K. Brecht
METHYL BROMIDE ALTERNATIVES
11 Evaluation of Methyl Bromide Alternatives Potentially Useful in
Miami-Dade County
Waldemar Klassen
Aaron J. Palmateer
Merlyn Codallo
TOMATO BREEDING
18 Breeding Tomatoes for Florida John W. Scott
Waldemar Klassen
25 Breeding Tomatoes for Resistance to all Races of the Bacterial
Spot Pathogen
John W. Scott
Jeffrey B. Jones
30 Development of Premium Tomato Varieties with High Lycopene
and Fruity/Floral Flavor Characteristics
Elizabeth A. Baldwin
John W. Scott
Charles A. Sims
WHITEFLY AND DISEASE MANAGEMENT
36 Integrated Management of Whiteflies and TYLCV on Tomato David Schuster
Natalia Peres
48 Control of Bacterial Spot of Tomato in the Greenhouse and Field
with Kasugamycin
Kenneth L. Pernezny
Pam D. Roberts
Nikol Havranek
Nadia Abdallah
Rod Sytsma
56 Evaluating Factors Affecting Movement of the Silverleaf Whitefly
and Tomato Yellow Leaf Curl Virus
David J. Schuster
James A. Taylor
Craig D. Stanley
Jane E. Polston
Sabine Grunwald
64 Monitoring Resistance of the Silverleaf Whitefly to Insecticides David J. Schuster
Mark A. Mossler
Cover photos provided by: Phyllis R. Gilreath; Teresa Olczyk; Monica Ozores-Hampton; Steven A. Sargent; & Eric H. Simonne

Characterization of Genetic
Mechanisms that Determine
Contamination of Tomatoes
With Salmonella: From the Field
to the Packing House
Max Teplitski and Keith Schneider
Abstract
The first objective of this project was
to begin the characterization of differences
in the infection and colonizaiton of roma
and round tomatoes by a human pathogen,
Salmonella enterica. While some of the
studies are still in progress, we demonstrate
that Salmonella can colonize the interior of
tomatoes via the vascular system when taken
up through the roots. Salmonella was not
able to infect roma or round tomatoes with
intact skin, however the infection of
wounded tomatoes proceeded rapidly. Sur-
prisingly, even after 14 days of incubation,
the infected tomatoes did not show any signs
of spoilage or decay. This highlights the
need for ensuring microbiological safety of
tomatoes: even fruits with 109 cells of Sal-
monella will pass visual inspections. As we
hypothesized earlier, the infection of toma-
toes by Salmonella is an active process,
which required major virulence regulators.
The identification of these regulators now
offers an opportunity to specifically disrupt
these regulators with targeted BMP.
The second objective of the research
was to investigate potential mechanisms of
inhibiting biofilm formation by Salmonella
on surfaces, similar to those found in pack-
ing houses. By using the reporter system in
the Salmonella biofilm formation cascade,
we demonstrate that potent inhibitors of
biofilm formation are compounds producedd
by common spices, therefore these are gen-
erally recognized as safe. We followed up
on this original discovery and identified sev-
eral candidate compounds that have a potent
ability to disrupt the biofilm regulatory cas-
cade.
Introduction
This project funded by Florida To-
mato Council has two goals aimed at identi-
fying genetic mechanisms that determine the
ability of Salmonella to contaminate toma-
toes and persist as biofilms in packing
houses. Specifically, the first objective of
our proposed research focuses on the ability
of Salmonella mutants with deletions in
major virulence regulators to colonize
vegetative and reproductive organs of roma
and round tomatoes. We are also using tran-
scriptomic analysis to identify catabolic
genes that are activated during growth on
pulp of roma or round tomatoes. These ex-
periments are necessary to try to explain
why the majority of salmonellosis outbreaks
are associated with roma, and not round
commercial varieties of tomatoes. In an-
swering these questions, we also gain a
valuable insight into the dynamics and ki-
netics of tomato infection by human patho-
gens. The second objective of this proposal
is to identify activities that could disrupt
biofilm formation by Salmonella. Biofilms
are multicellular microbial aggregates that
form on biotic and abiotic surfaces and serve
as a recalcitrant, persistent reservoirs of in-
fections. The ultimate goal of this work is to
develop BMPs that specifically target and
disrupt the ability of Salmonella to contami-
nate tomatoes in the field and during proc-
essing.
The results of our studies (presented
and discussed below) contribute to helping
ensure microbiological safety of Florida
produce. We have developed an EDIS pub-
lication (http://edis.ifas.ufl.edu/SS458)
which describes sources and survival strate-
gies that human enteric pathogens rely upon
to persist on farms. The EDIS publication
also suggests management strategies for
eliminating sources of pathogens. We are
1

working on developing a similar EDIS pub-
lication, specifically for tomato growers and
processors. The data generated in the past
few months was central to our developing a
competitive USDA T-STAR proposal (the
project was highly ranked and suggested for
funding, however USDA T-STAR program
was terminated). We were able to recruit an
outstanding student, Mr. Ali Al-Agely, who
secured additional funding ($3,000) for this
project through PMCB Internship Program.
In addition to the PI’s, two other sci-
entists have contributed to this work. Part-
time technician, Mr. Dmitri Kaganov carried
out studies with Salmonella mutants. Mr.
Kaganov is a US Army veteran, and an ap-
plicant in the pre-nursing program. Mr. Ali
Al-Agely has carried studies on vascular
colonization of tomatoes, identified the con-
stitutive gfp construct and performed mi-
croscopy work described below. He is cur-
rently working on genomic analysis of bac-
terial catabolic pathways required for tomato
infection with Salmonella. Mr. Al-Agely is a
UF University Scholar, PMCB intern and a
prospective graduate student.
Current work and preliminary results
For all experiments described below,
tomato fruits were purchased from Albert-
sons or Publix. For hydroponic experiments,
segments of tomato plants were harvested in
the field and rooted in the laboratory in a
mineral Hoagland solution without use of
rooting hormones. We avoided the use of
surface-sterilized samples, exotic chemicals
or aseptically-grown plants to make sure that
normal field microflora and agrichemicals,
typically used in tomato horticulture, are
present in all our experiments.
Salmonella colonizes tomato plants
by invading the vascular system and by
multiplying in wounded fruits. Under-
standing routes of tomato infection by Sal-
monella is the first key question that we
wanted to address. To test whether wild type
S. enterica 14028 can infect tomato fruits,
saline-washed suspensions of the pathogen
were either spotted onto the intact surface of
round or roma tomatoes or spotted onto a
wound inflicted by pricking the fruit epi-
dermis with a sterile aluminum wire (1 mm
in diameter). After a week-long incubation
at 20oC, fruits were cored with a sterile core-
borer (1 cm in diameter). Cores of 3-5 cm
were recovered, epidermis was peeled off,
the rest of the core was then cut into 0.5 cm
segments. Each sample was homogenized in
sterile phosphate-saline buffer (PBS), and
then plated on XLD, a selective medium for
identification of Salmonella.
The results of these studies indicate
that Salmonella cannot infect tomato fruits
(round or roma) if the skin is intact.
Wounded tomatoes became heavily colo-
nized by the pathogen. When fruits were in-
fected with ~1,000 cells of Salmonella, after
7 days 108-10
9 colony forming units of the
pathogens were recovered. When infected
onto a wound, within a week wild type Sal-
monella was able to move and colonize up
to 10 mm of the tomato tissue. This is simi-
lar to the observation from Dr. Triplett’s
group that Salmonella and other human
pathogens readily colonize and grow within
alfalfa sprouts (Iniguez, 2005 #801). It ap-
pears, therefore, that preventing wounding
of tomatoes is tremendously important in
avoiding contamination with Salmonella.
Surprisingly, we note that tomatoes
that were infected with Salmonella did not
develop any visual symptoms typically as-
sociated with spoilage. Some of the experi-
ments with wound-infected tomatoes were
allowed to proceed for 14 days, and no signs
of spoilage were observed (data not shown) .
After 14 days of incubation, roma tomatoes
appeared wilted, while round tomatoes re-
mained turgid and did not show any symp-
toms of rot, decay or spoilage. It was sur-
prising that wounds were seldomly co-
colonized by spoilage organisms – when
2

tomato fruits samples were plated on XLD
medium only Salmonella was recovered. We
do not yet know whether Salmonella has
mechanisms of excluding other, competing
organisms or whether this is a coincidence.
Because tomatoes heavily infected with
Salmonella (up to 109 colony forming units
per sample) are indistinguishable from un-
infected fruits, it is important to avoid con-
tamination of tomatoes in order to prevent
large outbreaks of tomato-associated salmo-
nellosis.
We also tested the hypothesis that
Salmonella can infect tomatoes via the vas-
cular system. For these experiments, 15-20
cm explants of field-grown tomato plants
were rooted in Hoagland #2 solution for 2
weeks. As soon as adventious roots devel-
oped, the growth medium was seeded with
dilutions of the Salmonella inoculum. To
facilitate the identification and tracking of
Salmonella, we used a strain labeled with a
constitutively-expressed green fluorescent
protein (GFP). As shown in Fig.1, colonies
of Salmonella are clearly visible inside to-
mato roots, again consistent with the conclu-
sion of Iniguez et al., (2005) that Salmonella
are proficient endophytic colonizers.
To test whether root colonization
actually leads to translocation of the patho-
gen into above-ground parts, we developed a
“printing technique” which allows for a
quick identification of target bacteria inside
the plant tissues. Tomato stems were cut
lengthwise, from top down (to avoid cross-
contamination) and halves of stems were
printed onto XLD agar. After overnight in-
cubation, Salmonella colonies are visible on
XLD agar (Fig. 2). These results indicate
that Salmonella cannot only colonize root
tissues, but it can also be translocated into
the above-ground parts. The translocation
occured at all infection doses, and individual
salmonellae were even present in plants that
were infected with as few as 1,000 cells.
We are currently concluding our
studies on the ability of Salmonella to colo-
nize interior of fruits when infected via vas-
cular tissue or by contaminating stigmas of
flowers.
Wound infection by Salmonella is
an active process. It is unclear whether
wounding simply provides a route for entry
or whether wound exudates stimulate ex-
pression of the specific genes. The prefer-
ence of Salmonella for colonizing and in-
fecting plant wounds is similar to our obser-
vation that genes required for attachment
(the pef operon) are induced in Salmonella
during infection of sprouts (Fig. 3). The in-
volvement of the pef genes in attachment to
wounded plant tissues is similar to a better-
characterized interactions between Salmo-
nella and animal cells. For its attachment to
animal cells, Salmonella relies on extracel-
lular appendages with homologous functions
(Ledeboer et al., 2006; van der Velden et al.,
1998). The pef genes are regulated by a quo-
rum sensor SdiA (Smith and Ahmer, 2003).
As shown in Fig. 3, pefI-luxCDABE reporter
was induced during Salmonella infection of
a plant seedling in the sdiA-dependent man-
ner. The pef genes are only activated on
wounded serfaces of the seedling. This sug-
gests that Salmonella may rely on the same
virulence genes and regulators to colonize
and infect both plant and animal hosts.
Specific regulators contribute to
the ability of Salmonella to infect toma-
toes. We further tested the hypothesis that
known regulators of virulence are involved
in the ability of Salmonella to infect and
colonize tomatoes. Mechanisms controlling
virulence of Salmonella in animal models
have been well-documented, and novel
virulence mechanisms are characterized with
each new study. In this initial work, we fo-
cused on the best-characterized regulators of
virulence. A mutation in flhD, a gene re-
quired for motility and flagella production
led to 10x increase in its ability to colonize
3

fruits. Consistently with these observations,
in coinfection experiments (wild type +
mutant), however, the flhD mutant was
70±3% more proficient than the wild type in
colonizing the interior of the wounded fruit.
To further investigate the basis of this phe-
notype, we tested the phenotypes of flagel-
lated but nonmotile mutant (motA) and those
of nonflagellated mutant (fliF). The nonflag-
ellated strain behaved as the wild type, the
motA mutant with paralyzed flagella be-
haived like the flhD mutant. This indicates
that the observed phenotype is most likely
due to the presence of the flagellum itself,
not due to the regulatory functions of FlhD,
and independent of the motility.
A dramatic (10-fold) reduction in the
fitness was associated with the presence of
genes encoded by the Salmonella virulence
plasmid (pSLT-) and the ability to utilize
purine and aromatic aminoacids. A mutation
in hns (histone-like protein) also dramati-
cally reduced the ability of Salmonella to
infect tomatoes. A mutation in envZ, a sen-
sor of osmotic potential of the environment,
was not recovered from the coinfections,
suggesting, perhaps, that perceiving and re-
sponding to changes in the osmolarity are
crucial for Salmonella survival inside to-
matoes. Surprisingly, a mutation in rpoS, a
gene responsive to and controlling oxidative
stress, had no impact on the competitive
colonizaiton of the pathogen.
A deletion of hilA, a virulence
regulator abolishes hypersensitive re-
sponse. A mutant in hilA, a major regulator
of virulence and Type 3 secretion system
was at a 5x disadvantage when coinoculated
with the wild type. To further characterize
the contribution of hilA to the phenotype, the
corresponding mutant was tested for its
ability to elicit hypersensitive response in
tobacco. HilA is a major regulator of the
Type 3 secretion system encoded within
Salmonella pathogenicity island I (Bajaj et
al., 1995). HilA is central to the ability of
Salmonella to invade epithelial cells and
cause gastroenteritis (Bajaj et al., 1995).
When a suspension of the hilA mutant was
infiltrated into tobacco leaves, it did not
elicit hypersensitive response (HR) (Fig. 4).
Salmonella 14028 (wild type) elicited HR,
similar to a dedicated plant pathogen, Pseu-
domonas syringae (Fig. 4).
The second Objective of the proposal
was to focus on identifying and blocking
genes that lead to the persistence of Salmo-
nella in biofilms that form on surfaces in
packing houses. In our previous work, we
have identified several regulatory and
structural genes required for Salmonella
biofilm formation on a variety of abiotic sur-
faces. With this funding, we also discovered
that the presence of MgSO4 strongly pro-
motes biofilm formation by Salmonella. We
are investigating whether the addition of in-
expensive chelating agents may be one of
the mechanisms for disrupting the ability of
Salmonella to form biofilms on abiotic sur-
faces.
Extracts of spices and novel syn-
thetic chemicals inhibit the biofilm forma-
tion cascade. We have also screened ex-
tracts of common spices (rosemary, thyme,
ginger) and vegetables (garlic) to see
whether they contain compounds that spe-
cifically inhibit expression of genes required
for biofilm formation. The extracts of garlic
and ginger contained activities that specifi-
cally blocked expression of csrB-lacZ, a re-
porter that is tightly and specifically regu-
lated by the BarA/SirA system in Salmo-
nella enterica and biofilm formation
(Teplitski, 2006 #1003).
This is significant for several rea-
sons. BarA/SirA is a known key regulator of
both virulence and biofilm formation in all
known gamma-proteobacteria (Teplitski,
2004 #507). The discovery that spices pro-
duce compounds that block BarA/SirA de-
pendent regulation now presents an oppor-
tunity to search and identify compounds that
4

block this key regulator. Because BarA/SirA
inhibitory activity was isolated from com-
mon spices and vegetables, it indicates that
the activity is likely fit the “GRAS” (gener-
ally recognized as safe) criteria. Regulatory
approval for use of GRAS compounds in
human consumption is typically straightfor-
ward. We have initiated a collaboration with
Scripps Florida to identify synthetic inhibi-
tors of BarA/SirA, and therefore of biofilm
formation. Structures of candidate molecules
are shown in Fig. 5.
Because functional homologues of
these compounds are present in common
spices, it is likely that these compounds fit
the GRAS category and will find soon ap-
plications in food safety.
Conclusions
Our results to date indicate that:
a) Salmonella can infect tomatoes, both
round and roma, through several routes.
Salmonella can colonize roots, and cells of
Salmonella can be translocated via the vas-
cular system into aboveground parts. Salmo-
nella can infect both roma and round toma-
toes through wounds. Round tomatoes in-
fected with Salmonella appear healthy and
do not develop any visual signs of infection
for at least two weeks. Roma tomatoes tend
to wilt after 14 days of infection, although it
is not clear whether the wilting is the result
of wounding itself or the infection with Sal-
monella. Salmonella was not able to infect
roma or round tomatoes with intact skin.
b) The infection of tomatoes with Salmo-
nella is an active process, which requires
specific virulence and attachment genes.
Specific catabolic genes are required for
persistence inside the fruit. The identifica-
tion of these regulators will allow the devel-
opment of targeted BMPs for disrupting the
ability of pathogens to invade or persist
within the produce.
c) Biofilms formed by Salmonella and other
pathogens are a persistent and recalcitrant
source of infection in both the field and the
packing houses. Our screen of spices identi-
fied activities capable of inhibiting biofilm
formation. A follow-up ongoing screen
identified at least four candidate chemicals
with the potential to block biofilm formation
by Salmonella.
5

Fig. 1. Salmonella infects interior tissues of tomato roots. GFP-labeled Salmonella enterica 14028 was seeded into
the hydroponic solution. After incubation for 14 days, roots were harvested, washed, and vortexed extensively to
remove any loosely adhering bacteria. Roots and the infecting salmonellae were then visualized under the confocal
microscope. Microcolonies of Salmonella appear in green, cells walls of tomato epidermis and cortex are grey.
Fig. 2. Colonization of tomato roots with Salmonella leads to the translocation of the pathogen to above-ground
parts. When halves of the tomato stems were imprinted onto Salmonella detection medium, characteristic black
colonies indicate the presence of Salmonella inside the stems, as far as 20 cm from the roots. Even low doses of root
inoculum (103 cells/infection) led to the infection of the stems. Arrows point to the black colonies, indicative of
Salmonella. Infection doses listed at the bottom of the figure.
6

Fig. 3. pefI, a gene encoding a regulator of the fimbrial operon, is upregulated during plant colonization. Cultures of
pefI-luxCDABE reporter in sdiA+ (wild type) and sdiA- backgrounds were inoculated onto surfaces of aseptically
grown seedlings. The seedlings were incubated at room temperature, and luminescence of the sdiA-regulated pefI-
luxCDABE reporter was visualized under the Hamamtsu CCD camera. In the wild type (and not in the sdiA) back-
ground, the reporter was strongly expressed on the seedling at the infection sites (Teplitski and Ahmer,
unpublished). False color overlay indicates activation of the reporter (red>yellow>green>blue).
Fig. 4. A mutation in the virulence regulator hilA eliminates the ability of Salmonella to elicit HR, which indicates
that hilA is required for the ability of the bacteria to invade the plant. A plant pathogen, Pseudomonas syringae elic-
its HR (positive control). Negative control (buffer only) did not elicit HR. HR is an apoptosis-like plant defense re-
sponse, it appears as brown necrotic lesions. Please note, that red markings on the leaf were made with a permanent
marker to indicate sites of infection with each test strain.
Fig. 5. Structures of novel chemicals that inhibit Salmonella biofilm formation pathway.
7

Refinements in the Chlorine
Dioxide Gas Treatment of
Tomatoes for Controlling
Microbial Contamination—A
Progress Report
Jerry A. Bartz, Ph.D., Plant Pathology
Department
Keith Schneider, Ph.D., Food Science &
Human Nutrition Department
Steven A. Sargent, Ph.D., Horticultural
Sciences Department
Jeffrey K. Brecht, Ph.D., Horticultural
Sciences Department
University of Florida/IFAS Gainesville
Purpose: determine where the gas could
be applied to during commercial practice
• Concentrated on applying the gas
to fruit that had already been
boxed because:
– Fruit could be treated in
ripening rooms or trucks
– Field-packed fruit could
be treated.
• Results of initial treatments of
fruit in boxes were not encour-
aging—rates that controlled de-
cay when fruit alone were treated
in the chamber did not control
decay when fruit were enclosed
in boxes and then treated in a
sealed drum-liner plastic bag.
2.4
0.0
0.5
1.0
1.5
2.0
Stock Solution Moist Cardboard Dessicated Cardboard Cardboard in Aluminum foil Waxed Cardboard
Chamber type
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Source (ClO2)
Sink (ClO2-)
Total Cl acounted for
% of stock accounted for
Cl
(mg
) re
co
vere
d a
fter
2 h
rs.
Recovery as affected by the presence of corrugated cardboard
8

Treatment of wound-inoculated
tomatoes with 2 mg of ClO2 gas per
kg of fruit completely prevented the
development of bacterial soft rot if
the fruit were stored in a plastic
treatment chamber. By contrast,
100% percent of the control fruit de-
veloped soft rot lesions. When simi-
larly inoculated fruit were randomly
distributed among 11.3 kg of fruit in
a single commercial tomato box, the
application of up to 99 mg of ClO2
(roughly 9 mg/kg fruit) failed to re-
duce decay incidence below 50%.
Subsequently, samples of the box
enclosed in the treatment chamber
were found to inactivate up to 75%
of the chlorine dioxide that off-
gassed from an aqueous solution.
The chlorine dioxide oxidized
some of the tannins in the un-
bleached box surfaces. Conse-
quently, to treat tomatoes in a stan-
dard commercial box at an equiva-
lent of 2 mg/kg, one would have to
apply at least 10 mg ClO2/kg fruit.
Better control of bacterial soft rot in
tomatoes treated in a commercial
box was achieved by forcing more
air through the box of fruit. How-
ever, the high air-flow rate may not
be feasible in commercial packing-
houses or transportation vehicles.
Additionally, in the absence of the
cardboard, a 20 mg gas/kg fruit dos-
age was associated with severe phy-
totoxicity, which included shrinkage
of the stem scars and bleaching of
exposed corky areas. Thus, high
Treatment of tomatoes in standard tomatobox using high air-flow rates.
48/72 hrs v 10/10
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
sealed vented Average Flow + -
box treatment
% decay
Decay incidence among wound inoculated fruit treated with 4.3 mgClO2/kg for 2 hr while enclosed in a standard tomato box that is in-side a plastic drum liner. Vents on the box were sealed or not,whereas the fan was set on high. (+=inoculated fruit not treated.)
9

ClO2 doses may produce fruit quality
issues.
Because ClO2 gas is heavier than
air, uniform distribution of this gas
in a bulk container of fruit appears to
require some air movement. Whether
the air movement occurring in trucks
is sufficient is unclear. Most labora-
tory tests, including those summa-
rized above have been conducted
with a point source of ClO2 gas.
Multiple sources if properly dis-
persed above the fruit in a standard
container may reduce the amount
lost to reaction with the cardboard.
Tests are planned that will simulate
placing multiple small ClO2 produc-
ing packets on the underside of the
lid of a tomato box.
10

Evaluation of Methyl Bromide
Alternatives Potentially Useful in Mi-
ami-Dade County
Waldemar Klassen, Aaron J. Palmateer and
Merlyn Codallo
Tropical Research and Education Center Uni-
versity of Florida/IFAS
18905 SW 280 Street
Homestead, FL 33031
Currently the only registered alternative to MC-
33 (mixture of 67% methyl bromide and 33%
chloropicrin) fumigants that can be used by to-
mato growers in Miami-Dade County are
metam potassium, metam
sodium and chloropicrin, although 50%
iodomethane + 50% chloropicrin can be used
under an experimental use permit. Metam com-
pounds are somewhat erratic in their perform-
ance as soil sterilants, and the beds cannot be
planted for three weeks after treatment because
of phytotoxicity of lingering residues. Tomato
growers in Miami-Dade County are prohibited
from using 1,3-Dichloropropene/chloropicrin,
the preferred alternative elsewhere in Florida,
because there is no impervious layer separating
the root zone from the Biscayne aquifer, the
source of drinking water for Miami. 1,3-D ap-
plied in the tomato beds undoubtedly would
leach into this aquifer.
Iodomethane + chloropicrin has performed
consistently well in small plot field trials con-
ducted annually since 1995 by faculty of the
Tropical Research and Education Center. How-
ever, iodomethane + chloropicrin (MIDASR)
still lacks EPA registration. In addition the io-
domethane + chloropicrin at $10 per pound rep-
resents a ser-ious investment for growers oper-
ating on slim profit margins.
Clearly there is a need to identify several
more materials to serve as methyl bromide al-
ternatives in Miami-Dade County.
Deviation from original objectives:
1. In our original proposal, one of the
treatments listed was EDN (ethane di-
nitrile) , very promising fumigant that
has been evaluated in Australia during
the past four years. This material was to
be supplied by BOC (now part of the
Linde Group). Unfortunately the re-
quested canister of EDN has still not ar-
rived.
2. On the other hand, we were able to ob-
tain two formulations of dimethyl disul-
fide (DMDS), and we have included
these evaluations as the second study in
this project.
3. In accordance with our plans to measure
the effects of the various treatments in
suppressing the root knot nematode,
Meloidogyne incognita, we included
one sachet filled with galled tomato
roots in each plot. Unfortunately we did
not check the viability of this inoculum
just before installing the sachets in the
beds prior to treatment. When we re-
covered the sachets after the treatment,
all of the nematodes were dead even in
the untreated control. Therefore we
cannot report any data on the effective-
ness of the various treatments against
the root knot nematode.
Objectives of studies no. 1 and 2,
respectively:
In field trials of the following two sets of
treatments obtain data on efficacy against ma-
jor pathogens and pests, tomato yield and qual-
ity in order to determine the near optimum dose
rates of methyl bromide alternatives.
11

Study no. 1:
1. Shank-applied chloropicrin (PIC) at 100
pounds/acre (112.1 kg/ha) followed by
drip-applied metam potassium (KPAMR) at
60 gallons/acre (561 l./ha) + metalized
plastic mulch.
2. Preplant incorporated napropamide (Devri
nolR) at 2 pounds/acre + halosulfuron (San-
deaR) at 1 ounce/acre (70.1 gm/ha), fol-
lowed by fosthiazate (NemathorinR)
sprayed onto bed surface at 3/74 lbs/acre
(4.2 kg/ha), then shank applied chloropicrin
(PIC) at 100 pounds/acre (112.1 kg/ha) +
metalized plastic mulch.
3. Shank applied 50% iodomethane + 50%
chloropicrin (MIDASR at 150 pounds/acre
(168.1 kg/ha) + metalized plastic mulch.
4. Shank applied MC-33 at 175 lbs/acre
(196.2 kg/ha) + metalized plastic mulch.
5. Untreated control (metalized plastic mulch
only).
Study no. 2:
1. Non-treated control,
2. MC 67-33 + metalized plastic mulch.; 175
lbs acre-1
[196 kg ha-1
],
3. DMDS 100 + metalized plastic mulch.; 38
gal acre-1
[355 l ha-1
],
4. DMDS 100 + metalized plastic mulch.; 56
gal acre-1
[524 l ha-1
],
5. DMDS 100 + metalized plastic mulch.; 74
gal acre-1
[692 ha-1
],
6. DMDS 79-21 + metalized plastic mulch.;
38 gal acre-1
[355 l ha-1
],
7. DMDS 79-21 + metalized plastic mulch.;
56 gal acre-1
[524 l ha-1
], and
8. DMDS 79-21 + metalized plastic mulch.;
74 gal acre-1
[692 l ha-1
].
Note
MC-33 is a mixture of methyl bromide
(67%) and chloropicrin (33%), MIDASR is a
mixture of equal weights of iodomethane and
chloropicrin, DMDS is dimethyl disulfide,
DMDS 100 is the pure material, and DMDS
29-21 is a mixture of DMDS (79%) with chlo-
ropicrin (21%).
Materials and Methods
Site description
Both experimental sites were located at
Block 12 of the Tropical Research and Educa-
tion Center, University of Florida, Homestead,
Florida. The soil is a Krome very gravelly loam
(loamy-skeletal, carbonatic, hyperthermic
Lithic Udorthents) with 58.8% gravel (>2mm),
48.4% sand, 30.3% silt and 21.3% clay con-
tents, respectively. Soil organic carbon is from
17.8 – 26.2 g kg-1
, total nitrogen 1.1 – 1.8 g kg-
1 and soil is pH 7.6 – 8.1.
Experiment design and management
Both experiments were laid out in a ran-
domized complete block design with four repli-
cations for each treatment. Each plot consisted
of a segment of a raised bed of 39-ft (11.9 m)
in study no. 1, of and 40-ft (12.2 m) in study
no. 2. In study no. 1 and study no. 2 the treat-
ments were as described above.
For assaying the effectiveness of the treat-
ments, Fusarium oxysporum f. sp. lycopersici
and Rhizoctonia solani were increased on ster-
ile millet seed in 250 ml flasks. Flasks were
aseptically inoculated by adding a 5-mm-diam.
plug from the periphery of one-week-old cul-
tures growing on potato dextrose agar (PDA)
(Difco Laboratories, Detroit, MI). Each flask
was incubated for 7 days at 25 °C under cool-
light fluorescent illumination, and shaken daily
to thoroughly mix fungal propagules. Yellow
nutsedge tubers were procured from Azlin Seed
Service, Leland, MS. Each of these four types
of inoculum was placed in sachets formed from
nylon stocking and 100-cm long colored plastic
tape was tied to the end of each sachet with a
different color for each type of inoculum. The
sachets were loaded individually with one of
the following: 5 grams of Fusarium culture, 5
grams of Rhizoctonia culture, and 20 yellow
nutsedge tubers. The sachets containing yellow
12

nutsedge tubers were immersed in water for 24
hours. One sachet of each type was buried in
each plot at a depth of 15 cm and with the color
coded ribbon extending onto the surface of the
bed. Just before the tomato seedlings were
transplanted into the beds, the sachets were
recovered. The
F. oxysporum f. sp. lycopersici and R. solani
infested millet seeds were aseptically plated on
acidified potato dextrose agar (PDA) (Difco
Laboratories, Detroit, MI). The number of
viable colonies was counted and a percentage
of the total recovery was recorded. Percent
germination of the yellow nutsedge tubers was
recorded.
On February 21, 2007 raised beds 15-cm
high, 91-cm wide, and 182-cm between centers
were formed in a field of shallow gravelly soil.
Next, 1120 kg ha-1 of 6N:6P2O5:12K20 dry
fertilizer was rototilled into the soil, and the
raised beds were re-formed.
On March 5 and 6, 2007 the appropriate
plots on the raised beds with the above formu-
lation and at the above rates were applied in the
appropriate plots of the raised beds, with the
exception of metam potassium, which was ap-
plied through the drip lines on March 11, 2007.
Concurrent with the application of the fumi-
gants or sprays, two drip tapes (T-TAPE TSX
508-12-450, flow rate: 5.6 l/min/100 m at 5.6 m
pressure, drip spacing = 0.3 m, diameter: 16
mm; Agro Distributors, Homestead, FL) and
CanslitR metalized plastic mulch (Intergro Inc.,
Clearwater, FL) were applied. The T-tapes
connected with layflat hose for irrigation. Three
weeks after fumigation, the sachets were recov-
ered and taken to the laboratory for processing.
Immediately thereafter, tomato, cv. ‘Tygress’
seedlings (Six L Farms, Naples, FL) were
transplanted on the beds at 51 cm between
plants. For insect control, imidacloprid (Ad-
mireR) was applied through the drip tapes, and
a backpack sprayer was used to spray the plants
2 or 3 times per week with various tank mixes
containing several of the following: mancozeb
+ copper hydroxide (MankocideR), azox-
ystrobin (QuadrisR), pyriproxyfen (Knack
R),
imidacloprid (ProvadoR) and spinosad (Spin-
TorR). Populations of the silverleaf whitefly
(Bemisia tabaci [Gennadius], Biotype B) were
high and tomato yellow leaf curl virus infection
rate of susceptible tomatoes in nearby plots
planted with a susceptible tomato, ‘FL 47’ was
high. No ‘Tygress’ plants showed any symp-
toms of virus infection. Early blight, Alternaria
solani, appeared before flowering and was suc-
cessfully controlled until the first harvest, but
thereafter the disease became severe.
The tomato fruits were harvested from 24
plants per plot on June 1, June 13 and June 22,
2007. Fruits were graded following Florida
Tomato Committee Standards (Brown, 2000).
The fruits were separated into extra large, large,
medium and small after each harvest and the
marketable and yields and total number of
fruits were recorded.
Immediately after the final harvest, root
samples were taken from 6 tomato plants per
plot and the roots were rated for galling (data
not shown).
The data were subjected to analysis of vari-
ance and Duncan’s multiple range tests by
means of SAS (version 8.1, SAS Inst., Inc.,
Cary, NC, USA). When significant (P < 0.05 or
less) F values were found in the Analysis of
Variance (ANOVA), the means were separated
by Duncan’s Multiple Range Test (DMRT).
Results and Discussion
The levels of infection of the tomato roots
with the root knot nematode, Meloidogyne
incognita, in both experiments were extremely
light and not significantly different in any
treatment than in the control (data not shown).
Also the levels of root rot were uniformly light
in all of the plots (data not shown).
Results of Study no. 1 (Table 1):
Rhizoctonia solani populations in the sa-
chets (Table 1) were not significantly sup-
pressed by any of the fumigant treatments.
Fusarium oxysporum f. sp. Lycopersici
13

populations in the sachets were suppressed to
the greatest extent by treatment no.1 (shank-
applied chloropicrin followed by drip-applied
metam potassium + metalized plastic mulch),
followed by treatment no. 3 (shank applied
50% iodomethane + 50% chloropicrin +
metalized plastic mulch), but these population
levels were not statistically different than the
numerically higher levels following treatment
no.2 (pre-plant incorporated napropamide +
halosulfuron, followed by fosthiazate sprayed
onto the bed surface, then shank applied chlo-
ropicrin + metalized plastic mulch) and treat-
ment no.4 (shank applied MC-33 + metalized
plastic mulch).
Clearly, even chloropicrin (PIC) at 100
pounds/acre (112.1 kg/ha) alone or in combi-
nation with KPAM, and with the herbicides and
the nematicide in the 2nd
treatment is an insuf-
ficient dose to suppress Rhizoctonia solani
even moderately and to suppress F. oxysporum
f. sp. Lycopersici strongly. Also chloropicrin
(PIC) at 75 pounds/acre (84.1 kg/ha) in combi-
nation with an equal weight of iodomethane
and chloropicrin (PIC) at 57.8 pounds/acre
(64.7 kg/ha) in combination with 117.3
lbs/acres (131.4 kg/ha) of methyl bromide is an
insufficient dose to suppress Rhizoctonia solani
even moderately and to suppress F. oxysporum
f. sp. Lycopersici strongly.
The total marketable tomato yields were the
highest in treatment no. 1 (shank-applied chlo-
ropicrin followed by drip-applied metam potas-
sium + metalized plastic mulch) and treatment
no.3 (shank applied 50% iodomethane + 50%
chloropicrin + metalized plastic mulch). Fol-
lowed by treatment no.4 (shank applied MC-33
+ metalized plastic mulch), treatment no.2 (pre-
plant incorporated napropamide + halosulfuron,
followed by fosthiazate sprayed onto the bed
surface, then shank applied chloropicrin +
metalized plastic mulch) and the untreated
control, which had the lowest yield. Only the
chloropicrin + metam potassium and io-
domethane + chloropicrin treatments had sta-
tistically higher total marketable tomato yields
than the untreated control. Tomato yields in the
remaining treatments were not statistically dif-
ferent than in the untreated control, probably
because the field was not severely infested with
nutsedge, root knot nematode, Fusarium or
Rhizoctonia.
The total number of marketable tomato
fruits was the highest in the chloropicrin +
metam potassium treatment and this number
was statistically greater than in the untreated
control. However none of the numbers of fruits
obtained in the chemical soil treatment were
different statistically.
The best overall treatment effects in study
no. 1 trial were obtained with chloropicrin +
metam potassium (PIC + KPAMR) and io-
domethane + chloropicrin (MIDASR).
Results of Study no. 2 (Table 2):
Rhizoctonia solani populations in the sa-
chets were not significantly suppressed by any
of the fumigant treatments. Fusarium
oxysporum f. sp. Lycopersici populations were
significantly, but not drastically, suppressed by
all of the formulations containing chloropicrin.
However, Fusarium populations in the sachets
exposed to pure DMDS at the highest rate were
not significantly lower than in the untreated
control. All of the formulations containing
chloropicrin drastically suppressed Cyperus
esculentus L. populations in the sachets,
but not to a statistically significant greater
extent than by DMDS 100; 56 gal acre-1
[524 l ha-1
] or by DMDS 100; 74 gal acre-1
[692 ha-1
].
Clearly DMDS without chloropicrin had
only a mild effect against yellow nutsedge and
no effect against Fusarium and Rhizoctonia.
However DMDS in combination with chlo-
ropicrin was highly effective against yellow
nutsedge and moderately effective against
Fusarium. Chloropicrin weighs 13.79 lbs/gal at
20 oC, and DMDS weighs 8.85 lbs/gal at 20
oC,
In the DMDS-chloropicrin 79:21 (w/w) mix-
ture, chloropicrin make up 14.05% of the vol-
ume. The weight of chloropicrin applied per
14

acre in each of the three different application
rates of DMDS 79-21 was 76 lbs/acre (85.2
kg/ha), 112 lbs/acre (126 kg/ha) and 148
lbs/acre (166 kg/ha). However even at the
highest rate at which 560 pounds of DMDS is
combined with 148 pounds of chloropicrin
Fusarium was not suppressed to a greater ex-
tent than by MC-33 in which only 57.8
pounds/acre (64.7 kg/ha) is combined with
117.3 lbs/acres (131.4 kg/ha) of methyl bro-
mide. This suggests that Fusarium control
needs to be achieved by the development of
resistant tomato cultivars.
The total marketable tomato yield was the
highest in the DMDS 79-21; 74 gal acre-1
[692 l
ha-1
] treatment, but not significantly higher than
in the DMDS 79-21; 56 gal acre-1
[524 l ha-1
]
treatment. Tomato yields in the remaining
treatments were not statistically different than
in the untreated control, probably because the
field was not severely infested with nutsedge,
root knot nematode, Fusarium or Rhizoctonia.
The total number of marketable tomato
fruits was the highest in the DMDS 79-21; 74
gal acre-1
[692 l ha-1
] treatment but not signifi-
cantly higher than in the DMDS 79-21; 56 gal
acre-1
[524 l ha-1
] and in the DMDS 79-21; 56
gal acre-1
[524 l ha-1
] treatments, nor than in the
MC 67-33; 175 lbs acre-1
[196 kg ha-1
] and
DMDS 100; 38 gal acre-1
[355 l ha-1
] treat-
ments.
The best overall treatment effects in this 2nd
trial occurred with DMDS 79-21; 74 gal acre-1
[692 l ha-1
] followed by DMDS 79-21; 56 gal
acre-1
[524 l ha-1
]
Acknowledgements
The MC-33 was provided by Helena
Chemical Co., Florida City, FL. MIDAS was
donated by Arysta Life Science Corporation,
and fosthiazate was provided courtesy of Dr.
James P. Gilreath, GCREC, Wimauma. We are
deeply grateful to Tony Estes, Cerexagri-Nisso,
LLC for providing the DMDS formulations,
and to Mr. Phil Crumpacker, Hendrix and Dail,
Inc. for assistance in applying the fumigants.
Mr. Robert Stubblefield, Mr. Jose Castillo and
other members of the Center’s field crew pro-
vided invaluable expert assistance. This work
was financially supported in part by the Florida
Tomato Committee.
15
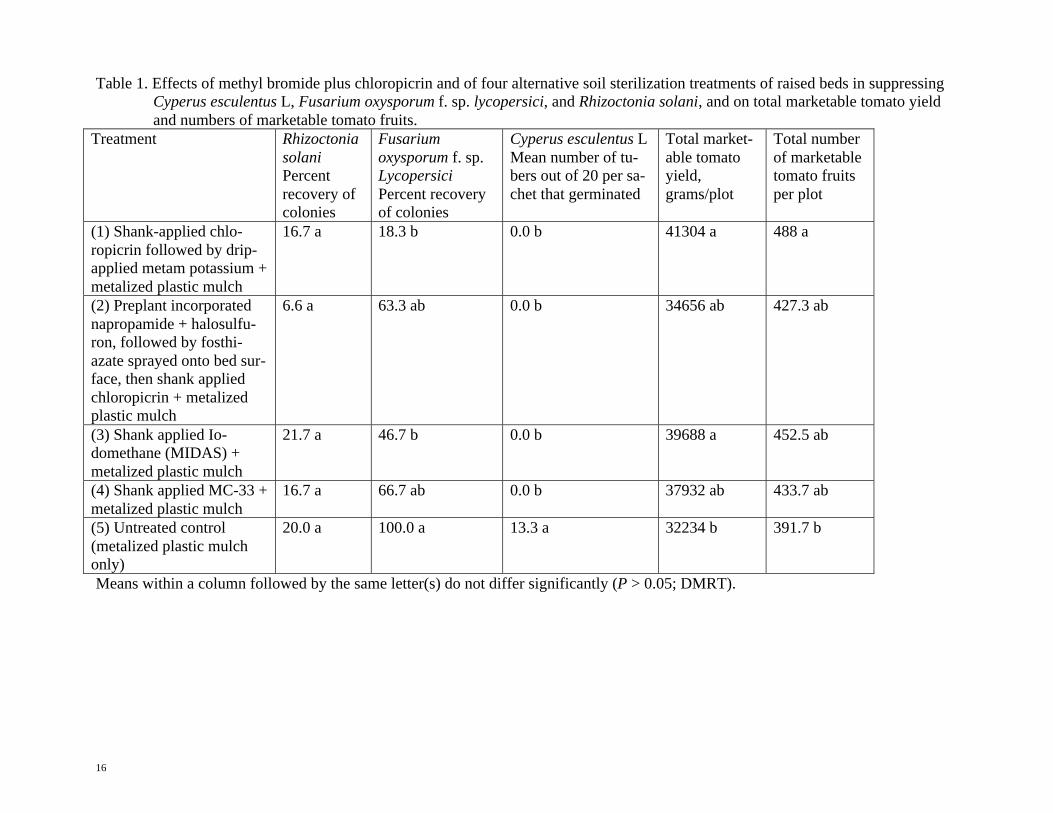
Table 1. Effects of methyl bromide plus chloropicrin and of four alternative soil sterilization treatments of raised beds in suppressing
Cyperus esculentus L, Fusarium oxysporum f. sp. lycopersici, and Rhizoctonia solani, and on total marketable tomato yield
and numbers of marketable tomato fruits.
Treatment Rhizoctonia
solani
Percent
recovery of
colonies
Fusarium
oxysporum f. sp.
Lycopersici
Percent recovery
of colonies
Cyperus esculentus L
Mean number of tu-
bers out of 20 per sa-
chet that germinated
Total market-
able tomato
yield,
grams/plot
Total number
of marketable
tomato fruits
per plot
(1) Shank-applied chlo-
ropicrin followed by drip-
applied metam potassium +
metalized plastic mulch
16.7 a 18.3 b 0.0 b 41304 a 488 a
(2) Preplant incorporated
napropamide + halosulfu-
ron, followed by fosthi-
azate sprayed onto bed sur-
face, then shank applied
chloropicrin + metalized
plastic mulch
6.6 a 63.3 ab 0.0 b 34656 ab 427.3 ab
(3) Shank applied Io-
domethane (MIDAS) +
metalized plastic mulch
21.7 a 46.7 b 0.0 b 39688 a 452.5 ab
(4) Shank applied MC-33 +
metalized plastic mulch
16.7 a 66.7 ab 0.0 b 37932 ab 433.7 ab
(5) Untreated control
(metalized plastic mulch
only)
20.0 a 100.0 a 13.3 a 32234 b 391.7 b
Means within a column followed by the same letter(s) do not differ significantly (P > 0.05; DMRT).
16

Table 2. Effects of fumigation of raised beds with DMDS alone, DMDS plus chloropicrin and methyl bromide plus chloropicrin in
suppressing Cyperus esculentus L, Fusarium oxysporum f. sp. lycopersici, Rhizoctonia solani, Meloidogyne incognita, and on
total marketable tomato yield and numbers of marketable tomato fruits.
Treatment Rhizoctonia
solani
Percent
recovery of
colonies
Fusarium
oxysporum f.
sp. lycopersiciy
Percent
recovery of
colonies
Yellow nutsedge,
Cyperus esculentus Lz
Mean number of tubers
out of 20 per sachet that
germinated
Total mar-
ketable to-
mato yield,
grams/plotz
Total num-
ber of mar-
ketable to-
mato fruits
per plotz
Non-treated control 42.5 a 100.0 a 16.5 a 26459 bc 292.0 bc
DMDS 100; 38 gal acre-1
[355 l ha-1
] 22.5 a 92.5 ab 3.8 b 25548 bc 315.8 abc
DMDS 100; 56 gal acre-1
[524 l ha-1
] 57.5 a 100.0 a 2.5 bc 21156 c 268.3 c
DMDS 100; 74 gal acre-1
[692 ha-1
] 20.0 a 100.0 a 1.8 bc 25083 bc 276.5 c
DMDS 79-21; 38 gal acre-1
[355 l ha-1
] 40.0 a 62.5 ab 0.3 c 26620 bc 320.5 abc
DMDS 79-21; 56 gal acre-1
[524 l ha-1
] 22.5 a 50.0 ab 0.0 c 32186 ab 369.8 ab
DMDS 79-21; 74 gal acre-1
[692 l ha-1
] 15.0 a 47.5 ab 0.0 c 34848 a 381.8 a
MC 67-33; 175 lbs acre-1
[196 kg ha-1
] 62.5 a 35.0 b 0.0 c 26994 bc 318.8 abczMeans within a column followed by the same letter(s) do not differ significantly (P > 0.01; DMRT).
yMeans within this column followed by the same letter(s) do not differ significantly (P > 0.05; DMRT).
17

Breeding Tomatoes for Florida
J. W. Scott
Gulf Coast Research & Education Center
University of Florida, IFAS
14625 CR 672
Wimauma, FL 33598
W. Klassen
Tropical Research & Education Center
University of Florida, IFAS
18905 SW 280th Street
Homestead, FL 33031
Abstract
High lycopene, Fusarium wilt resistant
hybrid ‘Fla. 8153’ was released in October
2006. This hybrid has performed consis-
tently well in taste panels over several years
and if harvested vine-ripe and branded could
compete well with greenhouse tomatoes in
supermarkets. Seed production is underway
and good volumes of seed should be available
in fall 2007 for the 2008 season. Fusarium
crown and root rot resistant hybrid Fla.
8413 has performed well on grower trials in
SW Florida over the past two years and has
also done well in GCREC trials. It is pres-
ently being considered for release pending
further testing. Fruit have a consistently
good marketable shape and firmness with
overall good fruit quality and a strong vine.
Tomato spotted wilt resistant hybrid Fla.
8367 yielded very well in the fall 2006 trial
at GCREC and is being considered for re-
lease. Spring data from NFREC is not yet
available. Seven of 9 stage 2 TYLCV resis-
tant hybrids were selected for further testing
while 10 new hybrids were selected for stage
2 testing in the fall. The spring TYLCV in-
oculated yield trial was grown under poor
conditions but some hybrids were selected
for further stage 2 testing.
Introduction
Improved varieties are needed to keep
the Florida tomato industry competitive in a
changing world. Improvements include the
areas of yield, pest resistance, and fruit
quality. Florida’s geography offers many
challenges for tomato improvement, since
high temperatures limit fruit set and are con-
ducive to disease and insect problems. There
are no neighboring states with similar lati-
tudes to most of Florida, so there is less help
in solving agricultural problems than is the
case in many states. Although private com-
panies have tomato breeding programs that
develop varieties for Florida, these compa-
nies are not set up to handle many of the
long-term, high risk projects that could
prove valuable in the future. The University
of Florida tomato breeding program aims at
such projects and works in partnership with
the private companies to deliver improved
varieties of benefit to the Florida tomato in-
dustry.
Much of the groundwork for the devel-
opment of heat-tolerant varieties was done
at the University of Florida (Scott et al.,
1986). ‘Solar Set’ was an important com-
mercial, heat-tolerant variety for 11 years
after it was released (Scott et al., 1989). The
improved heat-tolerance in ‘Solar Fire’, re-
leased in 2003, could be of benefit to grow-
ers in seasons with fewer hurricanes than in
2004 when it was being introduced. We now
have Fusarium wilt race 3 resistant varieties
using a resistance gene discovered in a wild
species by this program in the 1980's (Scott
and Jones, 1989; Scott and Jones, 1995).
Breeding lines with Fusarium crown and root
18

rot resistance were released in 1999 (Scott
and Jones, 2000) and UF breeding lines are
in commercial varieties ‘Sebring’, ‘Soraya’,
and ‘Crown Jewel.’ With the impending loss
of methyl bromide, these pathogens could
become more widespread. Other diseases
such as bacterial wilt and spotted wilt occur
in Florida, but cause far more damage in
other regions of the world. If these diseases
become more prevalent here, resistant varie-
ties will prove beneficial. Tomato yellow
leaf curl virus (TYLCV) is a serious threat to
Florida tomato production (Polston et al.,
1999) and some crops have sustained severe
losses in the last few years. Commercial
companies have released TYLCV resistant
varieties but none have yet been adapted by
Florida growers. Our project has been ongo-
ing since 1990 utilizing different resistance
genes derived from the wild species Ly-
copersicon chilense. It appears that four
genes have been introgressed, with two genes
needed in a variety to provide resistance.
These genes must be incorporated into both
parents of a hybrid since the resistance is
not dominant. This makes the breeding of
finished varieties more difficult but such hy-
brids are presently being evaluated and this
is discussed herein. Molecular markers
linked to the resistance genes are being de-
veloped that will accelerate future breeding
progress and these will be provided to all
tomato breeders for their use and benefit to
the Florida industry.
Tomatoes have met with dissatisfac-
tion in the marketplace. Essentially, this re-
lates to compromises that are made in pro-
viding fruit that will ship well. Solutions to
this problem are not simple. Research is
needed to provide tomatoes that will be
more acceptable to consumers. On the bright
side, Nugyen and Swartz, (1999) and others
have shown that lycopene, the red pigment
in tomato, has strong antioxidant properties
that reduce several cancers. Work in the
breeding program has been ongoing for 25
years with a crimson gene (ogc) that im-
proves internal tomato color and increases
lycopene content. Seven breeding lines with
this gene have been released to seed compa-
nies over the last 12 years. Crimson varieties
may be a boon to the Florida industry in the
near future. Genetic alteration of plant archi-
tecture might provide ground tomatoes that
reduce labor costs for staking and harvesting.
This could also be important for future pro-
duction in Florida. This is another long-term,
high risk project that is being pursued in our
program that is not likely to be attempted
by the private sector. Florida Tomato Com-
mittee funding has been imperative to the
operation of this breeding program, an in-
vestment that has and will continue to pay
dividends.
Objectives of this project were
1. To develop varieties or breeding lines
resistant to soil-borne pathogens.
2. To develop improved heat-tolerant
inbreds and hybrids.
3. To develop commercially acceptable
breeding lines and hybrids resistant
to TYLCV or tomato spotted wilt vi-
rus.
4. To improve fruit quality and post-
harvest characteristics.
5. To develop commercial hybrids for
the Florida tomato industry.
Objective 1
Methods. There were 68 lines
screened for Fusarium wilt race 3 and 113
19

lines screened for Fusarium crown rot in fall
2006 and spring 2007. There were 46 lines
inoculated with the bacterial wilt pathogen in
fall 2006. In spring 2006 32 lines were
screened for bacterial wilt resistance but the
inoculation failed. A replicated yield trial
comparing advanced inbreds was conducted
in fall 2006 and spring 2007 that included 1
line resistant to Fusarium wilt race 3 and 3
lines resistant to Fusarium crown rot. At
NFREC 3 Fusarium wilt race 3 resistant and
3 Fusarium crown rot resistant hybrids were
tested in fall 2006.
Results. Several hybrids with huge-
fruited Fla. 8293 as a parent looked good in
fall 2005 and spring 2006 trials. Fla. 8293
was supposed to be Fusarium crown rot re-
sistant but recent selections of it were sus-
ceptible. We have gone back to an earlier se-
lection and will soon know if this selection
carries resistance. Fla. 8293 is a parent in
hybrid Fla. 8413 that has looked good in
grower and GCREC trials. Several other
Fusarium crown rot resistant hybrids (Fla.
8412, Fla. 8415) have shown merit and are
being tested further for possible release.
Vines of these crown rot lines have looked
strong under some difficult growing condi-
tions. New crown rot resistant inbreds Fla.
8491, Fla. 8499, and Fla. 8497 have all
looked good in last year’s trials and are pres-
ently being tested in new hybrid combina-
tions. Fla. 8109 has a high level of bacterial
wilt resistance with large fruit and it has
been used in a number of crosses with elite
inbreds to improve horticultural type. Sev-
eral inbreds have been developed from these
crosses and many have huge fruit, some with
parental potential. Furthermore, some ap-
pear to have bacterial spot tolerance so this
will be watched carefully as new hybrids are
evaluated for commercial potential.
Objective 2
Methods. Heat-tolerant (HT) fruit
setting ability is being incorporated into all
phases of the breeding program. For instance
many lines being developed with bacterial
spot tolerance have heat tolerance as well. In
winter 2007 27 HT lines were evaluated at
TREC and in fall 2006 and spring 2002, 184
HT inbreds were evaluated at GCREC for
their performance. There were 9 HT hybrids
and 14 HT inbreds tested in fall 2006 and
spring 2007 yield trials. At NFREC in fall
2006, 4 HT hybrids were evaluated in ob-
servation trials.
Results. Inbred Fla. 8044 has been
mentioned as a line with outstanding HT in
past reports. It is a parent in tomato spotted
wilt resistant hybrids Fla. 8367 and Fla.
8485B mentioned in this report. It was also
crossed with Fla. 8109 (see objective 1) and
Fla. 8493 was developed. Fla. 8493 had the
highest marketable yield in both fall 2006
and spring 2007 GCREC yield trials and it is
being tested as a parent line at present. Fla.
8485B is a crimson, HT hybrid that did well
in yield trials during the last year. As indi-
cated the HT trait is now in several lines
listed under other projects so is more ubiqui-
tous than indicated here. Heat-tolerant hy-
brids with the proper horticultural charac-
teristics should provide Florida growers with
varieties that set marketable fruit more relia-
bly under stress conditions such as either
cool or high temperatures.
Objective 3
Methods. Geminivirus resistance.
20

There were 218 and 270 lines inoculated
separately with ToMoV and TYLCV, rated
for disease severity, and evaluated for horti-
cultural type in fall 2006 spring 2007, re-
spectively. There were 86 resistant hybrids
evaluated during the year in stage 1 testing.
Crosses were made to produce 91 hybrids,
some new and some to replenish seed. Nine
hybrids were in stage 2 testing in the fall and
7 of these were retested in the spring along
with 9 others that were advanced from stage
1 testing in the fall. Twelve inbreds were
tested in advanced trials during the year.
Spotted wilt resistance. There were 17 lines
with spotted wilt resistance evaluated in fall
2005 and spring 2006. Three hybrids were
evaluated in yield trials at GCREC and 5
were evaluated at NFREC. There were 22
F2’s with a new resistance source evaluated
in fall 2006. In the spring at NFREC 108 F3
lines with this new resistance were evalu-
ated. This new source has resistance to a
Hawaiian strain of the virus that overcomes
Sw-5 the resistance gene that is used in all
present commercial hybrids.
Selection for conventional (not the
new) resistance is done with sequence char-
acterized amplified region (SCAR) molecular
markers that eliminate the need to screen
with thrips and potentially spread the virus
in the west coast growing region.
Results. Geminivirus resistance.
The goal is to develop commercial quality
hybrids with resistance in both parents since
resistance in one parent has not been ade-
quate. Seven of 9 hybrids tested in fall 2006
were tested again in spring 2007. However,
irrigation problems in the spring prevented a
good assessment of hybrids in the trial. Sev-
eral hybrids will continue to be tested while
some others were too small fruited. In spring
2007, 149 selections were made and 5 F1's
were advanced to F2. In spring some new
inbreds with good characteristics were se-
lected and will next be tested as parents.
Molecular marker work is progressing and
markers are beginning to accelerate breeding
efforts for resistance to TYLCV although
more markers need to be elucidated. An im-
portant gene Ty-3 was described (Ji et al.,
2007) and a closely linked molecular marker
has been discovered. This will be licensed to
the tomato breeding community in the near
future. Ultimately we want to combine
TYLCV resistance with heat-tolerance, bac-
terial spot resistance and other desirable
traits in the same variety.
Spotted wilt resistance. Zeraim Ged-
era has seed available for grower testing of
released hybrid Fla. 7964. Fla. 8124C, a re-
sistant parent that is presently being tested
in several hybrid combinations, did well in
both spring and fall trials. Two hybrids with
the most promise for release are Fla. 8363
and Fla. 8367. These will be tested further,
especially at NFREC where these and other
hybrids are being tested this fall. Fla. 8367
did very well in the fall GCREC trial but it
could not be tested in the spring because of
seed lot germination problems.
Objective 4
Methods. Fruit quality and shelf life
are emphasized in all breeding projects. One
method to improve shelf-life is to develop
varieties with ultrafirm (UF) fruit. In the fall
and spring 18 UF inbreds were evaluated.
Another aspect of fruit quality is the devel-
opment of high lycopene varieties by using
the crimson (ogc) gene. This gene is widely
distributed in the various breeding projects.
21

New hybrids are continually being made and
evaluated.
There were 44 and 37 lines evaluated
primarily for improved flavor in the fall and
spring, respectively. Nineteen of these lines
were also evaluated at TREC. Part of this
work involves the incorporation of high sug-
ars from a cherry line into large-fruited lines.
The most interesting lines from all flavor
work are tested in flavor trials described in
the grant report on flavor with cooperator
Dr. Elizabeth Baldwin.
Another project is to develop toma-
toes that do not require staking by use of the
brachytic (br) gene that reduces plant height
and increases side shoots. These tomatoes
are called compact growth habit (CGH) to-
matoes. In winter 2007 92 CGH lines were
evaluated at TREC and in fall and spring 158
lines and 43 hybrids were evaluated at
GCREC. Jointless tomatoes are also receiv-
ing considerable attention with 46 and 58
lines evaluated at GCREC and TREC, re-
spectively. In the fall 65 jointless hybrids
were evaluated at GCREC. During the year
39 new jointless hybrids were made and 41
new CGH crosses were made with most be-
ing jointless.
Finally, a project has been underway
for a number of years to develop a superior
tomato cultivar or hybrid especially well-
suited for Miami-Dade County winter
growing conditions. This thrust began in
1999 when Dr. Jim Strobel selected a series
of ‘Sanibel’ plants from a planting at the
Knaus Berry Farm, Homestead. These
‘Sanibel’ lines were advanced through several
cycles of selection at Homestead,
Fletcher, NC and at Bracey, VA. The late
Dr. Leandro Ramos crossed some of these
lines to some large-fruited, crimson, jointed
pedicel lines provided by Dr. Randy
Gardner, NCSU. Subsequently some of
these lines have been crossed to multiple
disease-resistant, firm, large-fruited, crimson
lines from the breeding program at GCREC.
Our efforts have focused on selection
of vigorous determinate plants (equal to or
larger than ‘Sanibel’ in vine status), jointless,
large globe-shaped fruit with glossy red ex-
ternal color, very little yellow shoulder and
cracking, and tolerant to graywall. The inter-
nal structure of the fruit should be fleshy,
firm, uniformly crimson in color, with a stem
core extension of white tissue of _ inch or
less, and with superior flavor. Firmness and
shelf life characteristics should be such that
the breakers and pink fruit can withstand the
picking, packing and transportation proc-
esses; and arrive at the market in prime con-
dition with deep red internal and external
color–and have good flavor.
Results. Fla. 8153 was released in
October 2006 and will likely be sold under
the trademark Flora-Lee. Some quality data
from this hybrid has been described in the
reports with Liz Baldwin. This variety is a
premium tomato for the supermarket trade
and is meant to be harvested at the breaker
stage. The concept is to better compete with
greenhouse grown tomatoes. Fla. 8297 is a
crimson UF line with very good flavor that
looked good in several hybrid combinations
including the crimson HT F1 Fla. 8485. Fla.
8107, a CGH hybrid, was tested as a proto-
type for stake-less tomato production on
grower farms in Dade county. The plant
type was good and yields were generally
good. There was some off-shape fruit that
require improvement. The trial at TREC was
not ideal due to irrigation problems and a
22

severe infestation of early blight. Neverthe-
less, some lines did well and hopefully ge-
netic progress was made in selecting them.
Numerous selections with jointless stems
were made that would allow for mechanical
harvest.
Twenty-three of the best self-
pollinated lines and F1 hybrids of the im-
proved lines with ‘Sanibel’ lineage crossed to
the multiple disease-resistant, firm, large-
fruited, crimson lines developed in the
GREC and North Carolina programs were
evaluated in 2006-2007 in one grower’s field
near TREC and 6 lines were evaluated in a
second grower’s field 15-miles north of
TREC. Four of these lines appeared to be
especially promising. Subsequent selections
from these same lines and hybrids of these
lines with advanced jointless lines from
GCREC were evaluated at GCREC in the
spring and yielded very well. Testing will
continue in the 2007-2008 season.
Objective 5
Methods. Hybrids anticipated to
have commercial potential are evaluated for
horticultural type in replicated breeding
plots each season (stage 1). After the initial
evaluation, those selected for further (stage
2) testing are put in observation trials at
GCREC and NFREC, with some also being
tested at TREC. Those that perform well in
the observation trials are tested in state rep-
licated trials (at the above 3 centers and
IRREC) and in trials on grower farms (stage
3). For seed production, it is hoped that the
initial crossing produces enough seed for the
first two evaluations. When a hybrid moves
onto the advanced trial phase more seed is
produced at GCREC or sometimes by seed
companies.
Results. Seed was increased for 78
hybrids that are in stage 2 testing. In addi-
tion, 15 crosses were made for stage 1 hy-
brid testing for bacterial spot tolerance.
TYLCV tolerant, CGH, and jointless crosses
are mentioned elsewhere. Extra seed was
made for Fla. 8367 and Fla. 8413 that will be
moving to stage 3 testing.
Literature Cited:
Ji, Yuanfu, David J. Schuster, and Jay W.
Scott. 2007. Ty-3, a begomovirus resistance
locus near the Tomato yellow leaf curl virus
resistance locus Ty-1 on chromosome 6 of
tomato. Molecular Breeding (in press, avail-
able on line)
Nguyen, M.L. and S.J. Schwartz. 1999. Ly-
copene: Chemical and biological properties,
Food Technol. 53(2): 38-45.
Polston, J.E., R.J. McGovern, L.G. Brown.
1999. Introduction of Tomato yellow leaf
curl virus in Florida and implications for the
spread of this and other geminiviruses of
tomato. Plant Dis. 8384-988.
Scott, J. W., and J. P. Jones. 1989. Mono-
genic resistance in tomato to Fusarium ox-
ysporum f. sp. lycopersici race 3. Euphytica
40:49-53.
Scott, J. W., and John Paul Jones. 1995. Fla.
7547 and Fla. 7481 tomato breeding lines
resistant to Fusarium oxysporum f. sp.
lycopersici races 1, 2, and 3. HortScience
30(3):645-646.
23

Scott, J. W., and John Paul Jones. 2000. Fla.
7775 and Fla. 7781: Tomato breeding lines
resistant to Fusarium crown and root rot.
HortScience 35(6):1183-1184.
Scott, J. W., S. M. Olson, J. J. Bryan, T. K.
Howe, P. J. Stoffella, and J. A. Bartz. 1989.
Solar Set: A heat tolerant, fresh market to-
mato hybrid. Fla. Agric. Expt. Sta.
Circ. S-359 10p.
Scott, J. W., R. B. Volin, H. H. Bryan, and
S. M. Olson. 1986. Use of hybrids to de-
velop heat tolerant tomato cultivars. Proc.
Fla. State Hort. Soc. 99:311-314.
24

Breeding Tomatoes for Resistance to
All Races of the Bacterial Spot
Pathogen
J. W. Scott
Gulf Coast Research & Education Center
University of Florida
14625 CR 672
Wimauma, FL 33598
J. B. Jones
Plant Pathology Department
University of Florida
Gainesville, FL 23611
Abstract
There are 4 races of the bacterial spot
pathogen that infect tomato, but races T3 and
T4 are of primary concern in Florida. Race T3
is widespread, whereas the prevalence of T4 is
not known, although it has been identified
several times in South and West Florida. Par-
ent lines with T3 resistance have been devel-
oped and crosses made with horticulturally
advanced, susceptible parents to obtain hy-
brids that have commercial potential with in-
termediate resistance. The best of these, Fla.
8314, has had outstanding yields but fruit size
is slightly less than that of ‘Florida 47’. This
hybrid yielded well in trials last year but is not
being released because of the fruit size being
slightly small and due to the presence of race
T4 in Florida. No hybrids with T3 and T4
resistance are acceptable for release at least
one new hybrid looked very good in the spring
and it will be tested further. Several inbred
lines have looked good and crosses will be
made with these in attempt to find hybrids
acceptable for release. Genetic studies have
not been definitive as yet but indicate epistasis
is present. Three sources of resistance are be-
ing studied and combining resistance from
these sources may provide increased resistance
levels. Molecular marker work is underway to
identify and combine resistance genes to de-
termine if such combinations improve resis-
tance and ultimately to be able to breed for
resistance using marker assisted selection
which will accelerate the breeding process.
Introduction
Bacterial spot is still the most ubiquitous
disease problem of tomatoes in Florida. Four
tomato races have been discovered to date and
these belong to four species of Xanthomonas
(Xcv), the bacterial spot pathogen (Jones et
al., 2005). In Florida, there have been three
races; the original race (T1), the race discov-
ered in 1991 (T3) (Jones et al., 1995), and a
new race (T4) (Astua-Monge et al., 2000).
Race T3 has largely replaced race T1 in Florida
(Jones et al., 1998). Race T4 has mutated from
T3 and appears to be spreading. It has been
found in at least Dade, Manatee, and Hillsbor-
ough counties. It is not known how important
this race is now or might become should race
T3 tolerant varieties be commercially grown.
Race T2 was originally reported from Brazil,
but has now been isolated in some states in the
USA including Ohio but not Florida.
Our breeding project began in 1983 when
we found Hawaii 7998 was resistant to race T1
(Jones and Scott, 1986). In the early 1990’s
resistance to race T3 was discovered in several
accessions including Hawaii 7981 (Scott et al.,
1995), and this resistance has been incorporated
into advanced T1 resistant breeding lines. Lines
with resistance to T1 and T3 also had tolerance
to race T2, whereas lines with resistance to
either race alone were susceptible to T2 (Scott
25

et al., 2003). This suggests combining bacterial
resistance genes in a line may have unexpected
beneficial effects against other bacterial races.
We found PI 114490 was resistant to race T2 in
the summers of 1995 and 1996 and this was
further verified from 1997 through 2005 at
Wooster, Ohio. PI 114490 also was resistant to
race T1 and tolerant to race T3 (Scott et al.,
2003). In studies from 2003 and beyond we
found PI 114490 was resistant to race T4
(Scott et al., 2006; Yang et al., 2005). The gen-
eral resistance from PI 114490 needs to be
incorporated into advanced breeding lines. This
general resistance could be important, should
race T2 migrate into Florida, or it might also
protect against any new race which might
emerge in the future. We also found resistance
to races T3 and T4 in Fla. 8326 a breeding line
derived from PI 126932 (Scott et al., 2006). PI
128216 is also resistant to races T3 (Scott et
al., 1995) and race T4 (Scott et al., 2006). It is
desirable to develop genetic information on
these resistances to allow for more efficient to
breed for race T4 resistance and combine this
with T3 resistance. Varieties with T3 and T4
resistance would provide Florida tomato grow-
ers with protection from losses due to bacterial
spot infection and allow for minimal spraying
thus saving money and reducing environmental
concerns.
The objectives of this research were
1) To develop commercial hybrids
with tolerance to bacterial spot race
T3 and T4.
2) To develop inbreds with durable
resistance for use as parents in
commercial hybrids.
3) To determine genetic information
about raceT3 and T4 resistance.
Objective 1
Methods. During the fall and spring there
were 48 large-fruited hybrids in stage 1 testing
and 33 in stage 2 testing at GCREC. Eleven
hybrids were tested at NFREC in fall 2006 and
8 hybrids were tested in Fletcher, NC in sum-
mer 2006. There were 35 cherry F1’s and 6
plum F1’s tested in stage 1 at GCREC. There
were 65 crosses involving bacterial spot resis-
tant parent lines.
Results. Fla. 8314 has tolerance to T3 and
marketable yields have been very reliable over
several years of testing including 3 trials in the
last year. However, the fruit size is a little less
than that of ‘Florida 47’ and other commer-
cially grown varieties. Despite the reliable
marketable yields and good fruit firmness, it
was not felt that growers would adapt Fla.
8314 due to a lower percentage of 5 x 6 fruit.
The other issue is the presence of race T4 in the
state. For instance, Fla. 8314 had a lot of bacte-
rial infection in the spring 2007 trial at GCREC
due to T4 infection. Thus, although this hybrid
will continue to be tested it will not likely be
released. There are no prime candidates for
release with T3 and T4 tolerance at present.
Although several hybrids that have been tested
have had certain merits, none has shown enough
advantage to be released. In spring 2007 a new
stage 2 hybrid (8478 x 8208-no official hybrid
designation yet) did very well with a strong
vine and good marketable yield of extra-large
fruit. It will be tested further.
Objective 2
Methods. There were 128, 160, 12, and
44 breeding lines (F2 and more inbred) with
race T3 or race T3 and T4 resistance evalu-
26

ated in fall 2006, spring 2007, summer 2006
at Fletcher, NC, and winter 2007 (TREC),
respectively. Other lines developed from ge-
netic studies were tested as indicated under
objective 3. At GCREC, lines were inoculated
with race T3 in the fall 2006 and T4 in spring
2007. Race T4 has now been occurring natu-
rally at GCREC for about two years, thus we
have started to artificially inoculate with it.
Fifteen lines have been tested in advanced
yield trials at GCREC during the year.
Results. Breeding lines with more ad-
vanced horticultural characteristics typically
have moderate tolerance levels to race T4.
Emphasis is being placed on lines that show
less foliar disease to not only bacterial spot
but early blight as well. Of the lines tested in
replicated trials (stage 2 testing) Fla. 8401H,
Fla. 8495B, and Fla. 8594 looked good. The
former 2 are derived from the cross of Fla.
8233 with heat-tolerant line Fla. 7949B. A
sister line also looked nice at TREC last win-
ter. Three lines from the genetic studies (see
objective 3) looked impressive in the spring;
one line from the 8233 x 7776 cross and two
lines from the 8233 x 8326 cross. These will
be put into stage 2 testing and crosses will be
made to develop hybrids for objective 1. In-
bred Fla. 8496 appeared outstanding in NC
but hybrids with this line as a parent were
disappointingly small-fruited in spring trials.
Several early generation selections derived
from Fla. 8517 show some promise for plum
tomatoes. A line with T3 resistance only
looked outstanding with huge fruit at TREC
and will be used in crossing for improved hy-
brids.
Objective 3
Methods. We are working with T4 resis-
tant genes in breeding lines derived from 3
sources; PI 114490, PI 126932, and PI
128216. Fla. 8326 has resistance from PI
126932 as well as resistance to other races
from Hawaiian sources. Fla. 8233 has resis-
tance from PI 114490 and Hawaiian sources.
Fla. 8517 has resistance with PI 128216 and
PI 114490 in its background. A family was
previously developed with Fla. 8326 and sus-
ceptible parent Fla. 7946. This family and 5
F3’s were planted and inoculated with race T4
at Citra in summer-fall 2006 as in 2005. Also
36 F3’s between 8233 x 8326 were planted at
Citra. In the fall, families of 8233 and 8517
crossed with susceptible Fla. 7776 were
planted at GCREC along with 29 F4’s derived
from the 8233 x 8326 cross. In spring 2007,
families of 8233 and 8517 each crossed with
susceptible 7776 were grown along with 18
8233 x 8326 F3-F5’s, 7 8326 x 7946 F3’s, 19
8233 x 7776 F3’s, and 19 8517 x 7776 F3’s.
This summer families of 8233 x 7776, 8517 x
7776, and 8326 x 7946 are all planted at Citra
along with 65 F3-F5’s derived from the resis-
tant sources. All plants were inoculated with
race T4 and rated for disease severity. Selec-
tions were made for horticultural traits and
resistance in the F2 and later generations. A
Ph.D. student, supported from other funding,
is conducting this work for his dissertation.
He is also locating molecular markers linked
to the resistance genes that will accelerate the
bacterial spot resistance breeding progress
once identified.
Results. From the summer 2005 and
2006 experiments the F1 between Fla. 8326
and Fla. 7946 was intermediate between the
27

parents but skewed toward susceptibility.
This would indicate that if this resistance
alone was used one would need resistance in
both parents to have useful resistance in the
hybrid. Inheritance results varied between
experiments and there is some indication of
epistatic gene action. Inheritance from 8233
may also have epistasis which precludes es-
timating the number of genes in control and
heritability. There was not enough disease in
the 8517 family in the spring and no data
were obtained. We hope to get a good data set
from all 3 families in Citra 2007 to clarify in-
heritance of resistance from all the sources.
Analysis of data over the last several years
indicates that all 3 sources provide useful re-
sistance, but sometimes the levels are not as
high as desired under high-disease pressure. In
essence, we have various breeding lines with
fair resistance, but we hope to develop lines
with higher resistance levels. Combining lines
from different resistance sources may accom-
plish this. At present we do have some inter-
esting lines derived from 8326 and 8233, but
we have not proven if they have genes from
both sources yet. The molecular work will be
the only way to discern this. If good markers
are discovered they can also be used for
marker assisted selection which will accelerate
the breeding process. Once lines with resis-
tance genes from different sources are verified
we will find out if they do in fact have en-
hanced T4 resistance. They can also be tested
for races T1 and T2 in Ohio to determine if
they have broad spectrum resistance which
would likely mean the resistance is durable
and not likely to break down with the emer-
gence of new pathogen races.
Literature Cited
Astua-Monge, G., G.V. Minsavage, R.E.
Stall, Eduardo C. Vallejos, M.J. Davis, and J.
B. Jones. 2000. Xv4-avrxv4: A new gene -for-
gene interaction identified between Xantho-
monas campestris pv. vesicatoria race T3 and
the wild tomato relative Lycopersicon pennel-
lii. Molecular Plant-Microbe Interactions
13(12):1346-1355.
Jones, J.B., H. Bouzar, G. C. Somodi, R.E.
Stall, K. Pernezny, G. El-Morsy, and J.W.
Scott. 1998. Evidence for the preemptive na-
ture of tomato race 3 of Xanthomonas cam-
pestris pv. vesicatoria. In Florida. Phytopa-
thology. 88:33-38.
Jones, J.B., G.H. Lacey, H. Bouzar, G.V.
Minsavage, R.E. Stall, and N.W. Schaad.
2005. Bacterial spot- Worldwide distribution,
importance and review. Acta Horticulturae
695:27-33.
Jones, J.B. and J.W. Scott. 1986. Hypersensi-
tive response in tomato to Xanthomonas
campestris pv. vesicatoria. Plant Dis. 70:337-
339.
Jones, J.B., R.E. Stall, J.W. Scott, G.C. So-
modi, H. Bouzar, and N.C. Hodge. 1995. A
third tomato race of Xanthomonas campestris
pv. vesicatoria. Plant Dis. 79:395-398.
Scott, J.W., S.F. Hutton, J.B. Jones, D.M.
Francis, and S. A. Miller. 2006. Resistance to
bacterial spot race T4 and breeding for dura-
ble, broad-spectrum resistance to other races.
Rept. Tomato Genet. Coop. 56:33-36.
28

Scott, J.W., J.B. Jones, G.C. Somodi, and
R.E. Stall. 1995. Screening tomato accessions
for resistance to Xanthomonas campestris pv.
vesicatoria, race T3. HortScience 30:579-581.
Scott, J.W., D.M. Francis, S.A. Miller, G.C.
Somodi, and J.B. Jones. 2003. Tomato bacte-
rial spot resistance derived from PI 114490;
inheritance of resistance to race T2 and
relationship across three pathogen races. J.
Amer. Soc. Hort. Sci. 128(5):698-703.
Yang, W., S.A. Miller, J.W. Scott, J.B. Jones,
and D.M. Francis. 2005. Mining tomato ge-
nome sequence databases for molecular mark-
ers: Application to bacterial resistance and
marker assisted selection. Acta Horticulturae
695:241-249.
29

Development of Premium
Tomato Varieties with High
Lycopene and Fruity/Floral
Flavor Characteristics
Elizabeth A. Baldwin, USDA/ARS
Citrus & Subtropical Products Labora-
tory, Winter Haven, FL
J.W. Scott, University of Florida, Gulf
Coast Research & Education Center,
Wimauma, FL
Charlie Sims, Food Science Department,
University of Florida, Gainesville, FL
Abstract
Tomato fruit were grown in repli-
cated plots in Homestead, FL and the
University of Florida Gulf Coast Re-
search and Education Center in Wi-
mauma and harvested in March and June
of 2007, respectively. Fruit were ana-
lyzed by an experienced sensory panel
for overall flavor, sweetness, and acidity
as well as for chemical components in-
cluding color, soluble solids (SS), indi-
vidual sugars, titratable acidity (TA),
organic acids, vitamin C, and aroma
volatiles. In March some crimson hy-
brids, developed for the Homestead area
as possible Sanibel replacements, were
analyzed including 6153 a jointed com-
mercial hybrid from Syngenta, currently
grown on some acreage in Dade County.
There was little difference in flavor for
the March study on Homestead tomatoes
except that Sanibel and the Syngenta hy-
brid (6153) exhibited less acidity com-
pared to some of the crimson hybrids.
Line 9249 and 9251 were higher in SS
and sugars and the 6153 had a high
SS/TA ratio and low TA. In June, a
newly released crimson (high lycopene)
hybrid, ‘Fla 8153’ was harvested at table
ripe stage and rated along with tomatoes
typical of the Florida industry, including
some indeterminate pseudo-heirlooms
(heirloom combined with a parent sel-
ected for superior flavor) and non-
crimson good flavored hybrids or in-
breds developed over the last several
years for flavor. For the June fruit, the
pseudo-heirlooms and Fla. 8485 were
rated highest in flavor and the pseudo-
heirlooms for sweetness. Florida 47 was
rated lowest for flavor and sweetness
and low for acidity along with Fla. 8413
and the pseudo-heirloom Italianx.
The Florida tomato industry would
gain back lost market share with a pre-
mium tomato product that had good fla-
vor. Supermarkets like Publix are al-
ready demanding vine-ripe tomatoes for
their produce departments to enhance
flavor quality. Much of the Florida gas-
green industry currently goes to food
service. Past experienced sensory panels
have shown a preference for tomatoes
with a good sugar/acid balance such as
newly released ‘Fla. 8153’ that has the
added benefit of high levels of lycopene.
In this study, we continued to look at
genotypes with high lycopene back-
grounds, good horticultural characteris-
tics, a good sugar/acid balance, and
sometimes a fruity/floral flavor note that
would differentiate a potential release
from the rest of the fresh tomato market.
We tested some lines under south Flor-
ida conditions, including 6153, a jointed
commercial hybrid from Syngenta, cur-
rently grown on some acreage in Dade
County. We also tested pseudo-
heirlooms, which are heirlooms com-
bined with a parent selected for superior
flavor (Italianx = Fla. 8059 x Cour di
Bue di Albenga; Bulgaianx = Fla. 8214
x unknown Bulgarian land race). High-
lycopene tomatoes like ‘Fla. 8153’ may
garner health benefits due to the anti-
oxidant, anti-cancer activity of this pig-
30

ment as well as having good internal
color and flavor.
Methods
Tomato (Lycopersicon esculen-
tum Mill.) genotypes were grown at
grower plots in Homestead (March,
2005) and in a completely randomized
block design with two blocks and 5
plants per plot at the University of Flor-
ida Gulf Coast Research and Education
Center in Wimauma (June 2007).
Genotypes with superior flavor were
emphasized for comparison with stan-
dard varieties. All varieties were har-
vested at the table ripe stage.
Tomatoes were analyzed by an
experienced panel (30+ panelists) who
sampled tomato wedges representing at
least 4 fruit per cultivar. Sub-samples
were taken 3 times during the panel ses-
sion and prepared for analyses of color
and flavor components by measuring
solids, sugars, acids, color, titratable
acidity, vitamin C, and aroma com-
pounds with refractometer, HPLC,
chromometer, spectrophotometer, titra-
tor, and gas chromatograph (GC). Mean
differences were determined by
Duncan’s Multiple Range at < 0.1 for
sensory and using standard error bars for
chemical analyses.
Results
For March 2007, there was no
difference for overall flavor or sweet-
ness, but Sanibel and the Syngenta hy-
brid (6153) exhibited less acidity com-
pared to some of the crimson hybrids
(Table 1), which was reflected in the low
TA for these two lines (Fig. 1B where
circled lines were included in the sen-
sory panel). Other lines were tested
chemically that were not rated by the
sensory panel. Fla. 8583 is a cherry to-
mato crossed with a high sugar line, Fla.
8344 is a “fruity-floral” plum tomato
(selected for fruit-floral flavor), Fla.
8343 is the non “fruit-floral” iso-line to
Fla. 8343, Fla. 8570 is a small
fruited/high acid tomato, line 9255 was
selected for balanced flavor and Fla.
8572 is a crimson line. All of these lines
were selected by the breeder for high
flavor in the field. Lines F;a. 8583 and
Fla. 8344 were high in total sugars, su-
crose equivalents (indicator of sweet-
ness), individual sugars (glucose and
fructose, data not shown), Brix (SS),
SS/TA, and TA. The commercial hybrid
(6153) also had a high SS/TA ratio, but
low TA (Fig. 1A and B). Lines 832, Fla.
8570 and line 9255 were high in citric
acid, while lines 802, 9249 and Fla. 8572
were high in total ascorbic acid (TAA)
or vitamin C (Data not shown). Lines
802, Fla. 8344 and line 9255 exhibited
low hue, indicating more red color (Fig.
1C). These color ratings are an average
of peel and inner color. Fla. 8572, lines
9254 and 9255, Fla. 8344 and Fla. 8343
showed low chroma (low brightness).
31

Table 1. Experienced panel sensory ratings in March 2007. Ratings on a 1-9 scale where
higher numbers indicate more sweetness and acidity and preferred flavor quality.
Line Flavor Sweet Acidity
802 5.00 a 4.24 a 4.65 a
925 4.88 a 4.24 a 3.94 bc
800 4.82 a 4.10 a 4.82 a
803 4.76 a 4.15 a 4.53 ab
Sanibel 4.71 a 4.15 a 3.62 c
6153 4.21 a 3.74 a 3.32 c
Duncan's Alpha = 0.1
For June 2007, Bulgaianx, Fla. 8485 and
Italianx were rated highest for flavor and
Bulgaianx and Italianx for sweetness
(Table 2). Bulgarianx, Fla. 8485, Fla
8610, and Fla. 8153 were rated highest
for acidity. Italianx, Bulgarianx, Fla.
8485, and Fla. 8344 (not included in
sensory, indicated by rectangle in fig-
ures) exhibited the highest Brix (SS),
and Italianx and Fla. 8344 were high in
glucose, fructose (data not shown), and
total sugars (Fig 2A). This generally re-
flects the sensory results for sweetness.
Italianx and Fla. 8344 were highest for
SS/TA and Fla. 8344 for TA (Fig 2B).
Bulgarianx, Fla. 8485, Fla. 8610 and Fla.
8153 were highest in TA for those lines
included in the sensory panel, which re-
sulted in the high sensory rating for
acidity. Florida 47 was not particularly
low in sugars, but was low in TA and
received a poor overall flavor rating.
Volatile analysis is still underway, but
may shed further light on the low overall
flavor rating for Florida 47 compared to
the other lines. Line 8344 had the high-
est level of citric acid (data not shown),
while Florida 47 and Fla. lines 8059,
8214 and 8413 had the highest levels of
TAA (vitamin C, data not shown). Ital-
ianx, Fla. 8153, Bulgiaranx, and Fla.
8485 exhibited the lowest hue (were
most red, Fig 2C). Italianx, Bulgarianx
and Fla. 8344 showed the highest
chroma (most brightness).
32

Table 2. Experienced panel sensory ratings in June 2007. Ratings on a 1-9 scale where
higher numbers indicate more sweetness and acidity and preferred flavor quality.
Line Flavor Sweet Acidity
Bulgarianx 5.87 a 5.42 a 4.38 a
Fla. 8485 5.40 a 4.24 b 4.58 a
Italianx 5.38 a 5.36 a 3.51 b
Fla. 8610 4.60 b 4.07 b 4.51 a
Fla. 8153 4.49 b 4.36 b 4.11 a
Fla. 8413 4.00 bc 4.38 b 3.31 b
Florida 47 3.80 c 3.73 b 3.56 b
Duncan's Alpha = 0.1
In conclusion, the March tomato lines
compared similarly to Sanibel, not being
different in overall flavor or sweetness,
when grown under south Florida condi-
tions. Therefore, these are possible can-
didates to replace Sanibel in south Flor-
ida. For the June study, Italianx and
Bulgarianx, the pseudo-heirlooms, along
with Fla. 8485, exhibited good flavor
and sweetness, while all the lines tested
except Fla. 8413 were rated higher in
flavor compared to Florida 47. These
results were generally reflected in the
chemical data. The results for the other
lines, not included in the sensory panel,
will be of use to the breeder in making
selections for improved flavor.
33
Fig 1. March tomatoes.
Acids
0
2
4
6
8
1 0
1 2
1 4
1 6
1 8
2 0
Cultivars
%S
S/T
A,
TA
or
pH
S.S./T.A.
Titratable
Acidity x 10pH
B
Sugars
0
1
2
3
4
5
6
7
8
9
Cultivars
Perc
en
tTotal Sugars
Sucrose
Equivalence
Brix
A
Color
0
1 0
2 0
3 0
4 0
5 0
6 0
7 0
8 0
9 0
1 0 0
Ch
rom
am
ete
r valu
es
Chroma
Hue
Lycopene x
10
C
34

Fig. 2. June tomatoes.
Sugars
0 . 0
1 . 0
2 . 0
3 . 0
4 . 0
5 . 0
6 . 0
7 . 0
Cultivars
Perc
en
t
Total Sugars
Sucrose
EquivalenceBrix
A
Acids
0 . 0 0
1 . 0 0
2 . 0 0
3 . 0 0
4 . 0 0
5 . 0 0
6 . 0 0
7 . 0 0
Cultivars
% S
S/T
A,
TA
or
pH
S.S./T.A.
Titratable
Acidity x 10
pH
B
Color
0
1 0
2 0
3 0
4 0
5 0
6 0
Ch
rom
am
ete
r valu
es
Chroma
Hue
C
35

Integrated Management of
Whiteflies and TYLCV on Tomato
David Schuster (Entomology) &
Natalia Peres (Plant Pathology)
University of Florida, IFAS, GCREC,
14625 CR 672, Wimauma, FL 33598
Abstract
Experiments were conducted in the
fall 2006 and spring 2007 seasons at the
Gulf Coast Research and Education Center-
Balm to evaluate the singular and integrated
efficacies of Actigard™ and UV-reflective
aluminum mulch for managing the silverleaf
whitefly, Bemisia argentifolii Bellows &
Perring, and tomato yellow leaf curl virus
(TYLCV) on tomato. Treatments consisted
of plants growing on UV-reflective full-
metallized aluminum mulch or white virtu-
ally impermeable mulch with the tomato
plants either treated or not treated with Acti-
gard 50WG. Transplants for the Actigard
treatments were sprayed one week before
transplanting. After transplanting additional
foliar applications of Actigard were made
every two weeks in 2006 and weekly in
2007. Although some significant differences
among the treatments were observed in both
years for the numbers of whitefly adults and
immature life stages, neither the aluminum
mulch nor the Actigard treatments consis-
tently reduced or increased whitefly num-
bers. In 2006 when the whitefly population
was higher earlier in the crop, the plants
growing on the aluminum mulch tended to
have a lower percentage of plants with
symptoms of TYLCV than plants growing
on the white mulch. In addition, foliar appli-
cations of Actigard resulted in further re-
duction of the percentage of symptomatic
plants. In 2007 when the whitefly population
initially was low but increased to a higher
level later in the crop, the aluminum mulch
did not result in reduced incidence of
symptomatic plants but the combination of
aluminum mulch and Actigard did. The
early yield of extra large fruit was higher for
plants growing on the aluminum mulch
compared to plants growing on the white
mulch, although the difference was signifi-
cant only in 2006. Applications of Actigard
reduced the early yield of extra large fruit of
plants growing on the aluminum mulch in
2006 and of plants growing on the white
mulch in 2007. The results suggest that the
use of aluminum plastic mulch can result in
reduced incidence of TYLCV if viruliferous
whiteflies are present early in the crop be-
fore the plastic is covered by the plants. The
results further suggest that pre and post-
plant foliar applications of Actigard may
enhance this effect, but may reduce early
yield of extra large fruit. The rate of the pre-
transplant application of Actigard was dou-
ble the intended rate in these experiments
and could have contributed to the decreased
yields.
Introduction
Whiteflies and tomato yellow leaf curl
virus (TYLCV) are the major economic
pests impacting tomato production in south-
ern Florida during both the spring and fall
growing seasons. TYLCV causes significant
losses on tomatoes in Florida. The host
range of TYLCV is diverse, and some hosts
may not show any symptoms but may ac-
tively serve as a virus reservoir. TYLCV is
vectored by biotype B of Bemisia tabaci
(Genn.), also known as the silverleaf
whitefly B. argentifolii Bellows & Perring.
The host range of TYLCV and its vector
make epidemics extremely challenging to
manage in tomato production fields.
Controlling adult whiteflies with insecti-
cides is effective in preventing disease
transmission, but recently insecticides ap-
plied for control of whitefly adults have
been ineffective. In addition, whitefly bio-
type Q was recently detected in central FL.
36

Even though the biotype has not yet been
detected in tomato production fields, its es-
tablishment would further complicate con-
trol because the biotype is resistant to many
insecticides used to manage biotype B.
These include the nicotinoids (Admire Pro,
Platinum and Assail) as well as the insect
growth regulators (Courier, Knack).
Recently, an integrated management
program has been developed for managing
thrips and the Tomato spotted wilt virus
(TSWV) they transmit. Momol et al. (2004)
have shown during three years of research
that using UV-reflective plastic mulch re-
duces thrips populations and subsequent vi-
rus incidence about one half to two thirds.
The same results were obtained in a com-
mercial tomato field during 2000, 2001, and
2002. In 2005, an experiment was conducted
in southern Italy where biotype Q is present
(Momol et al., unpublished data). Acti-
gard™ (acibenzolar-S-methyl, Syngenta
Crop Protection, Greensboro, NC), a com-
pound inducing the systemic acquired resis-
tance (SAR) response, coupled with UV-
reflective mulches significantly reduced
TYLCV incidences in greenhouse tomatoes.
The SAR natural protection of plants
against pathogens is partly based on a vari-
ety of barriers already present in the plant
before the actual invasion. Plants can acti-
vate protective mechanisms upon detection
of invading pathogens. It is a concept similar
to immunization in warm blooded animals.
The SAR is expressed locally at the site of
primary inoculation and also systemically in
tissues remote from the initial treatment.
SAR has now been demonstrated in many
pathosystems. SAR is active against a broad
range of pathogens, including fungi, bacte-
ria, and viruses. Actigard is an inducer of
SAR against wheat powdery mildew and is
effective against certain diseases of rice and
tobacco. In Florida Louws et al. (2001)
found that Actigard showed activity against
bacterial spot of tomato and Momol et al.
(2004) found disease reduction in the
TSWV-tomato pathosystem. SAR inducers
in tomato show significant promise for the
future of disease management programs.
No single management tactic is effective
in reducing losses from whiteflies and
TYLCV. The objective of this study was to
determine the singular and integrated effica-
cies of Actigard and UV-reflective mulch
for managing whiteflies and TYLCV on to-
mato.
Materials and Methods
Experiments were conducted in the fall
2006 and spring 2007 seasons at the
GCREC-Balm. On Aug 15 and March 20
transplants of tomato cv. ‘FL 47’ were set
18 inches apart on raised beds of Myakka
fine sand soil covered with either white vir-
tually impermeable mulch (Vapor Shield™,
Canslit, Inc., Montreal, Quebec, Canada) or
UV-reflective full-metallized aluminum
mulch (Canslit, Inc., Montreal, Quebec,
Canada). Plots were three, 30 ft long rows
with rows separated by 30 ft of bare soil end
to end. Row spacing was 5 ft side. Plots
were irrigated by a drip irrigation system.
Treatments were replicated four times in a
randomized complete block design and con-
sisted of plants growing on UV-reflective or
white mulch treated or not treated with Ac-
tigard 50WG. Transplants for the Actigard
treatments were treated one week before
transplanting at a rate of 1 gm product/1000
plants. This rate was double the intended
rate of 2 gm/4000 plants. The applications
were made with a 2.5 gal, hand-held CO2-
powered sprayer fitted with a D-5 disk and
#45 core. The sprayer was operated at 60 psi
and delivered 3qt spray/1000 plants in 2006
and 2.75 pt spray/1000 plants in 2007. Ad-
ditional foliar applications of Actigard were
made in the field with a high clearance, self-
propelled sprayer operated at 200 psi and 3.4
mph. It was fitted with eight Albuz orange
nozzles per row and delivered 60 gpa (4
37

nozzles open) and 90 gpa (6 nozzles open.
In 2006 Actigard was applied at 0.75 oz
product/acre on 1, 15 and 29 September at
60 gpa and on 12 Oct at 90 gpa. In 2007
Actigard was applied at a concentration of
0.75 oz product/60 gal on March 21 and 30,
April 5, 19 and 26 at 60 gpa and on May 4 at
90 gpa.
Whitefly densities were assessed on the
middle 10 plants of the middle row of each
plot. The numbers of adults were counted
weekly by carefully turning the third leaf of
two stems of each plant. The numbers of
eggs, 1st, 2
nd & 3
rd, and 4
th instars were
counted weekly on the terminal leaflet from
the 7th
- 8th
leaf counting from the top of
each plant. All plants in each plot were ex-
amined weekly for plants with definite
symptoms of TYLCV. In 2006, plant tissue
was collected from selected plants with typi-
cal TYLCV symptoms to confirm the pres-
ence of TYLCV using PCR (polymerase
chain reaction). Reactions utilized primers
C473 and PTY1v2406 that amplify an 859
bp DNA product specific to TYLCV.
Results
In 2006, the silverleaf whitefly
population as evidenced by the numbers of
adults observed was low to moderate
throughout the trial (Table 1); however, the
population as evidenced by the numbers of
eggs was moderate early in the trial and low
later in the trial (Table 2). Significant differ-
ences in the numbers of adults were detected
on five of the sampling dates (Table 1). In
general, the application of Actigard in com-
bination with aluminum mulch resulted in
fewer adults than application of Actigard in
combination with white mulch. No signifi-
cant differences in the numbers of eggs were
observed on any sampling date (Table 2).
Differences in the numbers of 1st instars
were observed on three sampling dates, and,
on two of the dates, the application of Acti-
gard in combination with white mulch re-
sulted in more nymphs than with the alumi-
num mulch alone (Table 3). The numbers of
2nd
and 3rd
instars differed significantly on
the last three sampling dates (Table 4). Acti-
gard applications on either mulch did not
affect the numbers of nymphs on plants
compared to the respective mulches without
Actigard. The numbers of 4th
instars were
very low and did not differ significantly on
any sampling date (Table 5). Significant dif-
ferences in the numbers of sessile nymphs
were observed on October 23 and 30, but
Actigard applications on either mulch did
not affect the numbers of nymphs on plants
compared to the respective mulches without
Actigard (Table 6). The numbers of total
nymphs differed significantly only on the
last sampling date, when more nymphs were
observed on plants growing on white mulch
and treated with Actigard than on plants
growing on either mulch without Actigard
(Table 7). Applications of Actigard in com-
bination with mulch resulted in lower per-
centages of plants with symptoms of
TYLCV on five sampling dates compared to
the mulches without Actigard (Table 8). In
addition, a lower percentage of plants with
symptoms of TYLCV was observed on alu-
minum mulch compared to plants on white
mulch on all sampling dates except the last.
Visual observations of TYLCV correlated
with PCR with 100% of plants with typical
symptoms.
Yields of cull, small, medium, and
large fruit did not differ among any of the
treatments on either harvest date (Tables 9
and 10) or when combined over both harvest
dates (Table 11). Plants growing on the alu-
minum mulch without Actigard yielded
more extra large fruit than all other treat-
ments on October 31 (Table 9) and when
both harvests were combined (Table 11). No
visible phytotoxicity was observed for any
treatment.
In 2007, the silverleaf whitefly
population as evidenced by the numbers of
38

adults and eggs observed was low early in
the trial but increased to a very high level by
May (Tables 12 & 13). Significant differ-
ences in the numbers of adults were detected
only on May 22 when fewer adults were ob-
served on plants growing on white mulch
combined with Actigard (Table 12). No sig-
nificant differences in the numbers of eggs
were observed on any sampling date (Table
13). Differences in the numbers of 1st instars
were observed only on May 21 when the
application of Actigard in combination with
white mulch resulted in more nymphs than
with the white mulch alone (Table 14). The
numbers of 2nd
and 3rd
instars and sessile
nymphs did not differ significantly on any
sampling date (Tables 15 and 17). On May
14, there were significantly fewer 4th
instars
on plants treated with Actigard and growing
on the white mulch compared to plants not
treated with Actigard growing on the alumi-
num mulch (Table 16). The numbers of total
nymphs differed significantly on 7 May and
June 4, when fewer nymphs were observed
on plants growing on white mulch and
treated with Actigard than on plants growing
on the aluminum mulch without Actigard
(Table 18). On May 16 and June 15, appli-
cations of Actigard in combination with
aluminum mulch resulted in a lower per-
centage of plants with symptoms of TYLCV
compared to the white mulch with Actigard
(Table 19). On May 16, applications of Ac-
tigard on aluminum mulch resulted in a
lower percentage of plants with symptoms
of TYLCV compared to the aluminum
mulch alone. Applications of Actigard in
combination with either mulch resulted in
fewer Liriomyza leafmines compared to the
mulches alone (Table 20).
On June 4, plants growing on white
mulch without Actigard applications yielded
more cull fruit than plants growing on alu-
minum mulch with Actigard applications
(Table 21). Plants growing either on the
aluminum mulch or the white mulch without
Actigard applications yielded more extra
large fruit than plants growing on white
mulch with Actigard applications on 4 June
(Table 21) and when both harvests were
combined (Table 23). Yields of small, me-
dium and large fruit did not differ among
any of the treatments on either harvest date
(Tables 10 and 11) or when combined over
both harvest dates (Table 12). No visible
phytotoxicity was observed for any treat-
ment.
References Cited
Louws, F. J., M. Wilson, H. L. Campbell, D.
A. Cuppels, J. B. Jones, P. B. Shoe-
maker, F. Sahin and S. A. Miller.
2001. Field control of bacterial spot
and bacterial speck of tomato using a
plant activator. Plant Diseas 85:481-
488.
Momol, M. T., S. M. Olson, J. E. Funder-
burk, J. Stavisky and J. J. Marois.
2004. Integrated management of to-
mato spotted wilt on field-grown to-
matoes. Plant Disease 88:882-890.
39
Table 1. Effects of UV-reflective plastic mulch and foliar applications of Actigard on adults of the silverleaf whitefly on tomato in 2006.
No. silverleaf whitefly adults/20 leaves
Treatment* 23 Aug 31 Aug 8 Sept 15 Sept 21 Sept 27 Sept 5 Oct 13 Oct 19 Oct 27 Oct
UV-reflective mulch alone 1b 3a 2a 2a <1a 3ab 4b 4b 2a 1a
UV-reflective mulch + Actigard 0a 2a <1a 3a 1ab 2a 1a 3a 2a 1a
White mulch alone 1b 7a 3a 3a 2b 4ab 4b 7b 2a 1a
White mulch + Actigard 0a 5a 3a 6a 2b 6b 4b 4b 3a 1a
F3,9 3.65 2.73 1.67 0.86 3.22 3.29 5.01 4.16 0.24 0.23
P-value 0.06 0.11 0.24 0.50 0.07 0.07 0.03 0.04 0.86 0.87
*A "+" indicates that products were combined.
Means within a column followed by the same letter are not significantly different by the LSD (P=0.05).
Table 2. Effects of UV-reflective plastic mulch and foliar applications of Actigard on the silverleaf whitefly on tomato in 2006.
No. silverleaf whitefly eggs/10 leaflets
Treatment* 11Sept 18 Sept 26 Sept 2 Oct 9 Oct 16 Oct 23 Oct 30 Oct 6 Nov
UV-reflective mulch alone 7a 12a 3a 1a 7a 1a 0a <1a <1a
UV-reflective mulch + Actigard 7a 16a 9a 2a 4a 1a 1a 0a 1a
White mulch alone 12a 22a 5a 7a 11a 1a <1a 1a 0a
White mulch + Actigard 19a 20a 8a 6a 10a 1a <1a 1a 1a
F3,9 1.73 1.20 0.57 0.73 1.48 0.21 0.75 0.68 2.46
P-value 0.23 0.36 0.65 0.56 0.28 0.88 0.55 0.59 0.13
*A "+" indicates that products were combined.Means within a column followed by the same letter are not significantly different by the LSD (P=0.05).
Table 3. Effects of UV-reflective plastic mulch and foliar applications of Actigard on the silverleaf whitefly on tomato in 2006.
No. silverleaf whitefly 1st instars/10 leaflets
Treatment* 11Sept 18 Sept 26 Sept 2 Oct 9 Oct 16 Oct 23 Oct 30 Oct 6 Nov
UV-reflective mulch alone 2a 5a 10a 1a 3a 6a 5a 2a 1a
UV-reflective mulch + Actigard 3a 12ab 3a 4a 5a 12b 4a 4a 1a
White mulch alone 4a 18b 10a 1a 6a 8ab 5a 3a 1a
White mulch + Actigard 6a 23b 10a 3a 7a 8ab 3a 4a 5b
F3,9 0.92 3.43 0.84 1.48 1.33 4.37 0.47 0.67 3.28
P-value 0.47 0.07 0.50 0.28 0.33 0.04 0.71 0.59 0.07
*A "+" indicates that products were combined.Means within a column followed by the same letter are not significantly different by the LSD (P=0.05).
40
Table 4. Effects of UV-reflective plastic mulch and foliar applications of Actigard on the silverleaf whitefly on tomato in 2006.
No. silverleaf whitefly 2nd
and 3rd instars/10 leaflets
Treatment* 11Sept18
Sept
26
Sept 2 Oct 9 Oct 16 Oct23
Oct 30 Oct6
Nov
UV-reflective mulch alone 3a 10a 9a 1a 0a 4a 5ab 5ab 3a
UV-reflective mulch + Actigard 3a 8a 6a 4a <1a 10a 3a 2a 4ab
White mulch alone 11a 22a 11a 6a 1a 6a 7ab 11b 4ab
White mulch + Actigard 3a 17a 17a 3a 1a 4a 14b 10b 7b
F3,9 2.39 0.97 0.19 1.60 0.66 0.24 3.04 5.56 3.23
P-value 0.14 0.45 0.90 0.26 0.60 0.87 0.09 0.02 0.08
*A "+" indicates that products were combined.
Means within a column followed by the same letter are not significantly different by the LSD (P=0.05).
Table 5. Effects of UV-reflective plastic mulch and foliar applications of Actigard on the silverleaf whitefly on tomato in 2006.
No. silverleaf whitefly 4th instars/10 leaflets
Treatment* 11Sept18
Sept
26
Sept 2 Oct 9 Oct 16 Oct23
Oct 30 Oct6
Nov
UV-reflective mulch alone 0a 2a 1a 0a 0a 0a 0a 0a 0a
UV-reflective mulch + Actigard 0a 1a <1a 0a 0a 0a 0a 0a 0a
White mulch alone 0a 2a <1a 1a <1a 0a <1a <1a <1a
White mulch + Actigard 0a 5a 1a 0a 0a 0a 0a 0a 0a
F3,9 ---- 0.39 1.00 1.00 1.00 ---- 1.00 1.00 0.60
P-value ---- 0.76 0.44 0.44 0.44 ---- 0.44 0.44 0.63
*A "+" indicates that products were combined.
Means within a column followed by the same letter are not significantly different by the LSD (P=0.05).
Table 6. Effects of UV-reflective plastic mulch and foliar applications of Actigard on the silverleaf whitefly on tomato in 2006.
No. silverleaf whitefly sessile nymphs (2nd
, 3rd, and 4
th instars)/10 leaflets
Treatment* 11Sept18
Sept26
Sept 2 Oct 9 Oct 16 Oct23Oct 30 Oct
6Nov
UV-reflective mulch alone 3a 13a 10a 1a 0a 4a 5ab 5ab 3a
UV-reflective mulch + Actigard 3a 9a 6a 4a <1a 10a 3a 2a 4a
White mulch alone 11a 24a 11a 7a 1a 6a 8ab 11b 4a
White mulch + Actigard 3a 21a 18a 3a 1a 4a 14b 10b 8a
F3,9 2.39 0.58 0.30 2.03 0.61 0.24 2.84 6.50 2.53
P-value 0.14 0.64 0.82 0.18 0.62 0.87 0.10 0.01 0.12
*A "+" indicates that products were combined.Means within a column followed by the same letter are not significantly different by the LSD (P=0.05).
41

Table 7. Effects of UV-reflective plastic mulch and foliar applications of Actigard on the silverleaf whitefly on tomato in 2006.
Total no. silverleaf whitefly nymphs/10 leaflets
Treatment* 11Sept18
Sept
26
Sept 2 Oct 9 Oct 16 Oct23
Oct 30 Oct6
Nov
UV-reflective mulch alone 4a 19a 20a 1a 3a 10a 10a 7a 4a
UV-reflective mulch + Actigard 5a 21a 9a 8a 5a 22a 7a 6a 6ab
White mulch alone 15a 42a 21a 7a 7a 14a 12a 14a 5a
White mulch + Actigard 9a 44a 27a 6a 8a 12a 17a 14a 12b
F3,9 2.08 1.08 0.74 1.75 1.73 1.06 2.04 2.23 4.00
P-value 0.17 0.40 0.55 0.23 0.23 0.41 0.18 0.15 0.05
*A "+" indicates that products were combined.
Means within a column followed by the same letter are not significantly different by the LSD (P=0.05).
Table 8. Effects of UV-reflective plastic mulch and foliar applications of Actigard on the silverleaf whitefly on tomato in 2006.
% plants with symptoms of TYLCV
Treatment*12
Sept 22 Sept 27 Sept 5 Oct 11 Oct 19 Oct 27 Oct 1 Nov
UV-reflective mulch alone 0.4a 14.1b 28.2b 75.9b 88.9b 91.0b 97.0a 100.0
UV-reflective mulch + Actigard 0.0a 5.8a 12.0a 55.7a 69.4a 82.7a 95.5a 100.0
White mulch alone 2.1b 29.3c 40.3c 89.5c 97.5c 99.2c 100.0b 100.0
White mulch + Actigard 1.3ab 15.7b 24.2b 78.4b 90.3b 94.1b 99.6b 100.0
F3,9 3.58 46.93 20.92 16.28 32.08 22.08 6.08 ----
P-value 0.06 <0.0001 0.0002 0.0006 <0.0001 0.0002 0.02 ----
*A "+" indicates that products were combined.Means within a column followed by the same letter are not significantly different by the LSD (P=0.05).
Table 9. Effects of UV-reflective plastic mulch and foliar applications of Actigard on yield of tomato in 2006.
Yield/10 plants on 31 Oct
Culls Small Medium Large Extra large
Treatment* No. Wt. (lb) No. Wt. (lb) No. Wt. (lb) No. Wt. (lb) No. Wt (lb)
UV-reflective mulch alone 7a 2.7a 2a 0.3a 24a 5.8a 28a 8.9a 73a 35.8a
UV-reflective mulch + Actigard 5ab 1.8a 3a 0.5a 27a 6.3a 30a 9.0a 36b 15.2b
White mulch alone 4ab 1.3a 3a 0.4a 27a 6.2a 33a 10.1a 41b 18.2b
White mulch + Actigard 4b 1.2a 1a 0.2a 17a 3.9a 26a 8.4a 28b 12.4b
F3,9 3.69 2.15 1.53 2.06 2.39 2.33 0.57 0.33 4.16 4.85
P-value 0.06 0.16 0.27 0.18 0.14 0.14 0.65 0.80 0.04 0.03
*A "+" indicates that products were combined.Means within a column followed by the same letter are not significantly different by the LSD (P=0.05).
42

Table 10. Effects of UV-reflective plastic mulch and foliar applications of Actigard on yield of tomato in 2006.
Yield/10 plants on 14 Nov
Culls Small Medium Large Extra large
Treatment* No. Wt. (lb) No. Wt. (lb) No. Wt. (lb) No. Wt. (lb) No. Wt (lb)
UV-reflective mulch alone 2a 0.7a 4a 0.7a 31a 7.6a 24a 7.7a 4ab 1.8ab
UV-reflective mulch + Actigard 4a 1.2a 3a 0.4a 48a 12.5a 36a 11.7a 5a 2.0a
White mulch alone 2a 0.4a 3a 0.7a 29a 7.6a 17a 5.8a 0b 0.0b
White mulch + Actigard 2a 1.0a 4a 0.9a 32a 8.0a 21a 7.0a 2ab 0.9ab
F3,9 0.95 1.07 0.56 1.23 1.61 1.77 2.70 2.67 3.01 2.91
P-value 0.46 0.41 0.66 0.35 0.25 0.22 0.11 0.11 0.09 0.09
*A "+" indicates that products were combined.Means within a column followed by the same letter are not significantly different by the LSD (P=0.05).
Table 11. Effects of UV-reflective plastic mulch and foliar applications of Actigard on yield of tomato in 2006.
Total yield/10 plants
Culls Small Medium Large Extra large
Treatment* No.Wt.
(lb) No. Wt. (lb) No. Wt. (lb) No. Wt. (lb) No. Wt (lb)
UV-reflective mulch alone 9a 3.5a 6a 1.0a 55a 13.4a 52a 16.6a 76a 37.5a
UV-reflective mulch + Actigard 9a 2.9a 5a 0.9a 75a 18.8a 65a 20.7a 41b 17.2b
White mulch alone 6a 1.7a 6a 1.1a 55a 13.8a 50a 15.8a 41b 18.2b
White mulch + Actigard 5a 2.2 6a 1.0a 49a 11.8a 48a 15.4a 30b 13.2b
F3,9 1.78 1.57 0.42 0.74 1.61 1.81 0.24 1.00 4.39 5.19
P-value 0.22 0.26 0.75 0.55 0.25 0.22 0.35 0.44 0.04 0.02
*A "+" indicates that products were combined.
Means within a column followed by the same letter are not significantly different by the LSD (P=0.05).
Table 12. Effects of UV-reflective plastic mulch and foliar applications of Actigard on the silverleaf whitefly on tomato in 2007.
No. silverleaf whitefly adults/20 leaves
Treatment* 27 March 2 April 9 April 16 April 23 April 30 April 7 May 14 May 22 May 29 May
UV-reflective mulch alone 0a <1a 3a 1a 1a 5a 1a 29a 167a 352a
UV-reflective mulch + Actigard 0a <1° 4a 0a <1a 7a 6a 17a 155a 319a
White mulch alone 0a 0a 3a 0a <1a 8a 2a 20a 122ab 325a
White mulch + Actigard 0a 0a 4a <1a 1a 12a 1a 32a 79b 331a
F3,9 ---- 0.60 0.79 1.94 1.10 1.97 1.20 2.01 4.76 0.45
P-value ---- 0.63 0.53 0.19 0.40 0.19 0.36 0.18 0.03 0.72
*A "+" indicates that products were combined.Means within a column followed by the same letter are not significantly different by the LSD (P=0.05).
43

Table 13. Effects of UV-reflective plastic mulch and foliar applications of Actigard on the silverleaf
whitefly on tomato in 2007.
No. silverleaf whitefly eggs/10 leaflets
Treatment* 16 April 23 April 30 April 7 May 14 May 21 May 28 May
UV-reflective mulch alone 1a 10a 14a 26a 56a 54a 516a
UV-reflective mulch + Actigard 6a 11a 12a 36a 55a 81a 329a
White mulch alone 5a 10a 14a 27a 42a 183a 452a
White mulch + Actigard 5a 7a 16a 37a 80a 114a 216a
F3,9 0.84 0.93 0.14 0.51 0.19 1.96 2.50
P-value 0.51 0.46 0.93 0.68 0.90 0.19 0.13
*A "+" indicates that products were combined.Means within a column followed by the same letter are not significantly different by the LSD (P=0.05).
Table 14. Effects of UV-reflective plastic mulch and foliar applications of Actigard on the silverleaf whitefly on
tomato in 2007.
No. silverleaf whitefly 1st instars/10 leaflets
Treatment* 16 April 23 April 30 April 7 May 14 May 21 May 28 May 4 June
UV-reflective mulch alone <1a 1a 3a 25a 17a 29ab 60a 235a
UV-reflective mulch + Actigard 1a 3a 2a 17a 36a 49ab 64a 190a
White mulch alone 3a 2a 3a 11a 18a 22a 64a 200a
White mulch + Actigard 0a 0a 1a 15a 25a 99b 47a 197a
F3,9 2.31 1.31 1.90 1.43 0.87 3.19 0.29 0.96
P-value 0.14 0.33 0.20 0.30 0.49 0.08 0.83 0.45
*A "+" indicates that products were combined.Means within a column followed by the same letter are not significantly different by the LSD (P=0.05).
Table 15. Effects of UV-reflective plastic mulch and foliar applications of Actigard on the silverleaf whitefly on
tomato in 2007.
No. silverleaf whitefly 2nd
and 3rd instars/10 leaflets
Treatment* 16 April 23 April 30 April 7 May 14 May 21 May 28 May 4 June
UV-reflective mulch alone <1a 0a <1a 9a 15a 23a 65a 433a
UV-reflective mulch + Actigard 1a 1a <1a 4a 24a 33a 54a 278a
White mulch alone 1a 1a 5a 8a 12a 28a 63a 336a
White mulch + Actigard 1a 1a 1a 1a 11a 44a 74a 316a
F3,9 0.32 0.70 1.32 1.90 0.49 0.30 0.02 2.29
P-value 0.81 0.58 0.33 0.20 0.70 0.82 1.00 0.15
*A "+" indicates that products were combined.Means within a column followed by the same letter are not significantly different by the LSD (P=0.05).
44

Table 16. Effects of UV-reflective plastic mulch and foliar applications of Actigard on the silverleaf whitefly on
tomato in 2007.
No. silverleaf whitefly 4th instars/10 leaflets
Treatment* 16 April 23 April 30 April 7 May 14 May 21 May 28 May 4 June
UV-reflective mulch alone 0a 0a 0a <1a 3b 13a 27a 106a
UV-reflective mulch + Actigard 0a 0a 0a 0a 2ab 18a 35a 84a
White mulch alone 0a 0a 0a 0a 1ab 18a 26a 60a
White mulch + Actigard 0a 0a <1a 0a 0a 36a 32a 101a
F3,9 ---- ---- 1.00 1.00 3.44 0.45 0.83 2.45
P-value ---- ---- 0.44 0.44 0.07 0.72 0.51 0.13
*A "+" indicates that products were combined.Means within a column followed by the same letter are not significantly different by the LSD (P=0.05).
Table 17. Effects of UV-reflective plastic mulch and foliar applications of Actigard on the silverleaf whitefly ontomato in 2007.
No. silverleaf whitefly sessile nymphs (2nd
, 3rd, and 4
th instars)/10 leaflets
Treatment* 16 April 23 April 30 April 7 May 14 May 21 May 28 May 4 June
UV-reflective mulch alone <1a 0a <1a 10a 18a 36a 92a 538a
UV-reflective mulch + Actigard 1a 1a <1a 4a 26a 52a 89a 362a
White mulch alone 1a 1a 5a 8a 13a 46a 89a 396a
White mulch + Actigard 1a 1a 1a 1a 11a 80a 106a 417a
F3,9 0.32 0.70 1.11 2.15 2.25 0.07 0.31 2.28
P-value 0.81 0.58 0.40 0.16 0.15 0.97 0.82 0.15
*A "+" indicates that products were combined.Means within a column followed by the same letter are not significantly different by the LSD (P=0.05).
Table 18. Effects of UV-reflective plastic mulch and foliar applications of Actigard on the silverleaf whitefly ontomato in 2007.
Total no. silverleaf whitefly nymphs/10 leaflets
Treatment* 16 April 23 April 30 April 7 May 14 May 21 May 28 May 4 June
UV-reflective mulch alone 1a 1a 3a 34a 36a 64a 151a 773a
UV-reflective mulch + Actigard 1a 4a 2a 20ab 62a 101a 153a 552b
White mulch alone 4a 4a 8a 19ab 31a 68a 153a 596b
White mulch + Actigard 1a 1a 2a 17b 36a 179a 153a 613b
F3,9 1.05 0.87 1.13 3.23 1.30 0.92 0.32 4.79
P-value 0.42 0.49 0.39 0.08 0.33 0.47 0.81 0.03
*A "+" indicates that products were combined.Means within a column followed by the same letter are not significantly different by the LSD (P=0.05).
45

Table 19. Effects of UV-reflective plastic mulch and foliar applications of Actigard on the silverleaf whitefly on tomato in 2007.
% plants with symptoms of TYLCV
Treatment* 11 April 17 April 25 April 2 May 9 May 16 May 24 May 31 May 8 June 15 June
UV-reflective mulch alone 0.0a 0.4a 1.3a 2.5a 4.2a 11.3a 33.8a 52.5a 64.6a 75.0ab
UV-reflective mulch + Actigard 0.0a 0.0a 0.4a 0.8a 1.3a 3.0b 16.5a 36.2a 44.3a 52.4b
White mulch alone 0.0a 0.0a 0.0a 0.4a 2.5a 6.7ab 20.5a 43.0a 50.1a 57.7ab
White mulch + Actigard 0.4a 0.4a 0.8a 3.4a 4.2a 10.2a 34.4a 64.1a 70.5a 77.4a
F3,9 1.00 0.60 0.74 1.26 0.96 3.77 2.05 2.64 2.30 3.29
P-value 0.44 0.63 0.55 0.35 0.45 0.05 0.18 0.11 0.15 0.07
*A "+" indicates that products were combined.Means within a column followed by the same letter are not significantly different by the LSD (P=0.05).
Table 20. Effects of UV-reflective plastic mulch
and foliar applications of Actigard on Liriomyzaon tomato in 2007.
Treatment*
No. Lirio-
myzaleafmines/2min search
UV-reflective mulch alone 73a
UV-reflective mulch + Actigard 61bc
White mulch alone 70ab
White mulch + Actigard 51c
F3,9 8.09
P-value 0.006
*A "+" indicates that products were combined.Means within a column followed by the same
letter are not significantly different by the LSD(P=0.05).
Table 21. Effects of UV-reflective plastic mulch and foliar applications of Actigard on yield of tomato in 2007.
Yield/10 plants on 4 June
Culls Small Medium Large Extra large
Treatment* No. Wt. (lb) No. Wt. (lb) No. Wt. (lb) No. Wt. (lb) No. Wt (lb)
UV-reflective mulch alone 17ab 6.5a 4a 0.8a 28a 6.6a 58a 19.7a 95a 42.0a
UV-reflective mulch + Actigard 10b 3.7b 5a 0.8a 34a 8.3a 48a 16.4ab 75ab 32.9ab
White mulch alone 19a 7.3a 5a 0.7a 32a 7.6a 49a 16.3ab 84a 36.6a
White mulch + Actigard 14ab 4.8ab 4a 0.6a 30a 7.3a 42a 13.3b 55b 23.7b
F3,9 3.50 4.35 0.29 0.19 0.24 0.25 2.40 2.76 5.52 5.88
P-value 0.06 0.04 0.83 0.90 0.87 0.86 0.14 0.10 0.02 0.02
*A "+" indicates that products were combined.Means within a column followed by the same letter are not significantly different by the LSD (P=0.05).
46
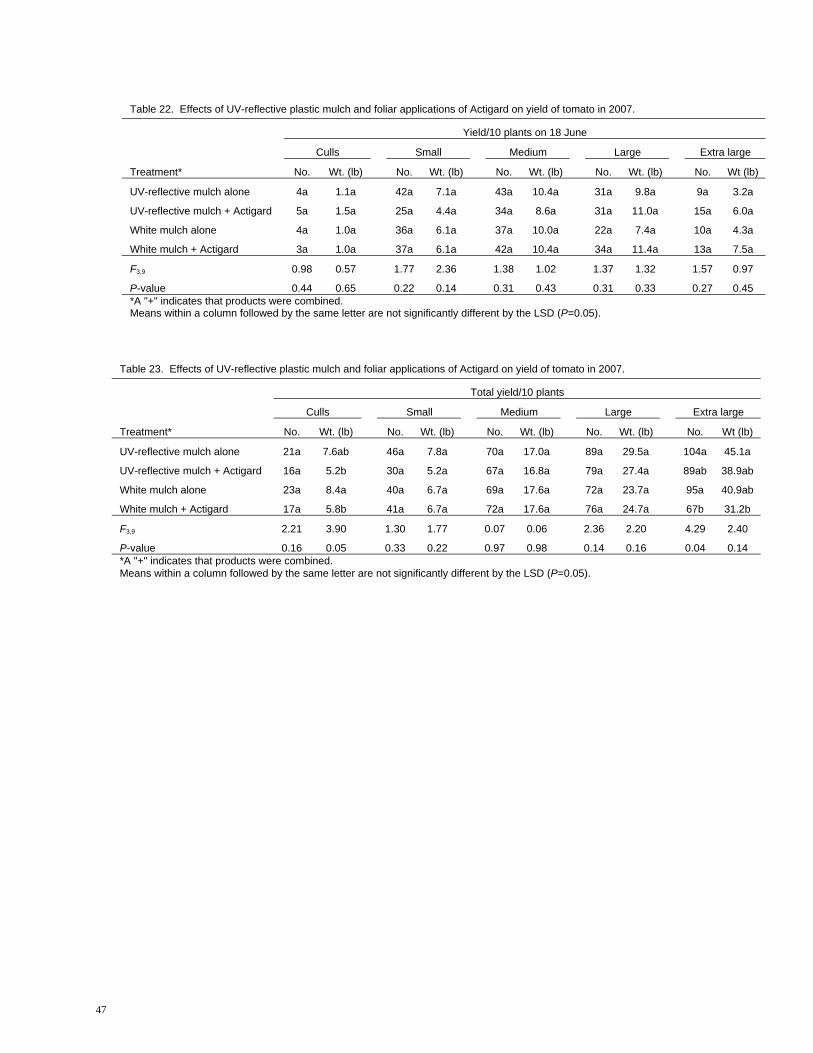
Table 23. Effects of UV-reflective plastic mulch and foliar applications of Actigard on yield of tomato in 2007.
Total yield/10 plants
Culls Small Medium Large Extra large
Treatment* No. Wt. (lb) No. Wt. (lb) No. Wt. (lb) No. Wt. (lb) No. Wt (lb)
UV-reflective mulch alone 21a 7.6ab 46a 7.8a 70a 17.0a 89a 29.5a 104a 45.1a
UV-reflective mulch + Actigard 16a 5.2b 30a 5.2a 67a 16.8a 79a 27.4a 89ab 38.9ab
White mulch alone 23a 8.4a 40a 6.7a 69a 17.6a 72a 23.7a 95a 40.9ab
White mulch + Actigard 17a 5.8b 41a 6.7a 72a 17.6a 76a 24.7a 67b 31.2b
F3,9 2.21 3.90 1.30 1.77 0.07 0.06 2.36 2.20 4.29 2.40
P-value 0.16 0.05 0.33 0.22 0.97 0.98 0.14 0.16 0.04 0.14
*A "+" indicates that products were combined.
Means within a column followed by the same letter are not significantly different by the LSD (P=0.05).
Table 22. Effects of UV-reflective plastic mulch and foliar applications of Actigard on yield of tomato in 2007.
Yield/10 plants on 18 June
Culls Small Medium Large Extra large
Treatment* No. Wt. (lb) No. Wt. (lb) No. Wt. (lb) No. Wt. (lb) No. Wt (lb)
UV-reflective mulch alone 4a 1.1a 42a 7.1a 43a 10.4a 31a 9.8a 9a 3.2a
UV-reflective mulch + Actigard 5a 1.5a 25a 4.4a 34a 8.6a 31a 11.0a 15a 6.0a
White mulch alone 4a 1.0a 36a 6.1a 37a 10.0a 22a 7.4a 10a 4.3a
White mulch + Actigard 3a 1.0a 37a 6.1a 42a 10.4a 34a 11.4a 13a 7.5a
F3,9 0.98 0.57 1.77 2.36 1.38 1.02 1.37 1.32 1.57 0.97
P-value 0.44 0.65 0.22 0.14 0.31 0.43 0.31 0.33 0.27 0.45
*A "+" indicates that products were combined.Means within a column followed by the same letter are not significantly different by the LSD (P=0.05).
47

Control of Bacterial Spot on
Tomatoes in the Greenhouse and
Field with Kasugamycin
Ken Pernezny, Nikol Hayranek, &
Nadia Abdullah
UF/IFAS, Everglades Research and
Education Center
University of Florida
3200 E. Palm Beach Rd.
Belle Glade 33430
Pam Roberts & Rod Sytsma,
IFAS, Southwest Florida REC
2686 Hwy. 29 N.
Immokalee, FL 34142
Introduction and background
Bacterial spot, caused by Xan-
thomonas perforans, continues to be one
of the most important diseases of toma-
toes in Florida. Losses of up to 50% in
large and extra-large fruit have been
documented when epidemics begin at
flowering (Pohronezny and Volin,
1983). Control is based on cultural prac-
tices and, typically, frequent sprays of
copper and mancozeb. However, wide-
spread occurrence of copper-resistant
strains often hamper management efforts
with copper (Marco and Stall, 1983; Po-
hronezny et al., 1992). Even the addition
of mancozeb is not sufficient to over-
come resistance to copper in some
strains (Pernezny et al., 2007).
Kasugamycin is a relatively new
antibiotic available to agriculture for
management of bacterial pathogens. It is
currently available for use on tomatoes
and peppers in Mexico but not in the
United States. In work reported to the
Florida Tomato Committee in 2006, we
found preliminary evidence that kasu-
gamycin significantly reduces bacterial
spot severity on tomato plants in the
greenhouse. Therefore, we initiated these
studies to determine the efficacy of ka-
sugamycin for control of bacterial spot
of tomato both in the greenhouse and the
field.
Materials and methods
Greenhouse experiments. Ex-
periments were conducted in an air-
conditioned greenhouse at Everglades
Research and Education Center in Belle
Glade. Maximum temperatures reached
28°C at midday. Tomato seed, cv. BHN
586, were planted in 15-cm-diameter
plastic pots in a commercial potting mix
(Fafard Soil Mix No. 2). After emer-
gence, plants were fertilized once per
week with a 20-20-20 soluble fertilizer
(1 g/L) (Agriliance, Inc.).
Plants were sprayed twice with
test chemicals at a 7-day interval before
inoculation, and twice at a 7-day interval
after inoculation. Kasugamycin was
tested with and without addition of cop-
per or captan (Table 1). Specific treat-
ments were Kasumin 2L (64 fl. oz./100
gal); Kasumin 2L + Kocide 2000 (1.5
lb/100 gal); Kasumin + Captan 50WP (3
lb/100gal); Kasuran 50WP (a pre-mixed
combination of kasugamycin and copper
hydroxide) (1.7 lb/100 gal); and a water-
sprayed control.
Xanthomonas perforans strains
T193, resistant to copper, was grown for
3 days at 28°C on glucose-nutrient agar.
Plates were flooded with sterile phos-
phate-buffered saline (Leben et al.,
1968), and resultant suspensions were
adjusted trubidimetrically to approxi-
mately 1 x 108 CFU/ml. Suspensions
were diluted to 106 CFU/ml for inocula-
tion. Applications of bacterial suspen-
sions were made by misting both adaxial
and abaxial leaf surfaces to run off using
a plastic spray bottle. Two drops of
Tween 80 were added to 660 ml of in-
oculum suspension to enhance wetting
48

of leaf surfaces. Plants were covered in
plastic bags for three days after
inoculation.
The experiment was a nested de-
sign with three plants nested within each
treatment and treatments assigned ran-
domly within each of four blocks. Dis-
ease ratings were made 16 days after in-
oculation and consisted of the percent-
age of leaf surface covered with lesions
combined with that lost from premature
defoliation (Pernezny et al., 1996). The
experiment was repeated. These tests
were conducted between December 2006
and late February 2007.
Field experiment. An experi-
ment was conducted at the Southwest
Florida Research and Education Center
to evaluate the efficacy of kasugamycin
for management of bacterial spot. To-
mato seedlings, cv. Hazera 3073, were
transplanted on March 6 2007 into a
field with soil type Immokalee fine sand.
Treatments were kasugamycin (Kasumin
2L, 64 fl.oz. /100 gal); kasugamycin +
copper hydroxide (Kocide 2000, 50%
Cu, 1.5 lb/Ac); kasugamycin + Captan
50WP (3 lb/100gal); a premixed product
of kasugamycin + copper hydroxide
(Kasuran 50WP, 1.7 lb/100gal); and an
untreated control.
Treatments were arranged in a
randomized complete block design with
four replications. Each plot consisted of
15 plants spaced 1.5 ft apart with 15 ft
between each plot. Beds were 2.7 wide
on 6-ft centers and covered with black
polyethylene film. Guidelines estab-
lished by the University of Florida,
IFAS, were followed for land prepara-
tion, fertility, irrigation, and weed and
insect management. Plants were sprayed
9 times at a 7-day interval on March 20
and 27, April3, 10, 17, and 24 and May
1, 8, and 15. All test materials were ap-
plied with a high-clearance sprayer des-
ignated specifically for applications in
staked tomato plots at a tractor speed of
2 mph and a pressure of 200 psi. A dou-
ble-drop boom equipped with six noz-
zles delivered a spray volume of 66
gal/acre.
Plots were inoculated on 12 and
27 April with a suspension of the bacte-
rial spot pathogen Xanthomonas vesica-
toria, race T1 and Xanthomonas per-
forans, race T3 at a final concentration
of 1 x 108 CFU/ml with a hand-pump
sprayer. A visual assessment was made
of the percentage of foliage covered with
lesions plus that lost due to premature
defoliation combined into one rating
(Pernezny et al., 1996) on April 26 and
May 21.
All fruit were harvested once on
May 23 and the number and weight of
marketable and nonmarketable (small,
misshapen, or diseased) fruit recorded.
Disease severity ratings and yield data
were subjected to analysis of variance,
followed by mean separation using
Waller’s method at P 0.05 (Statistical
Analysis System SAS Institute,
Cary, NC).
Samples of diseased leaf tissues
were taken from each replicated plot of
the following treatments: control, kasu-
gamycin, and kasugamycin + copper hy-
droxide on April 26 and May 21. Xan-
thomonas vesicatoria (perforans) strains
were isolated to determine their sensi-
tivity to kasugamycin in vitro. Briefly, a
flamed and cooled inoculation needle
with a small right-angle bend at the tip
was inserted through the margin of dis-
eased tissue and, without removal,
streaked onto duplicate plates of nutrient
agar supplemented with 50 g/ml of cy-
cloheximide. Plates were incubated at
28°C for 3 days. Single yellow colonies
characteristic of Xanthomonas were
49

restreaked from the isolation plates on
nutrient agar containing 0.5% glucose
(GNA). Cultures were restreaked as nec-
essary to ensure purity and stored in
sterile 15% aquous glycerol at -70°C.
A total of 12 strains were isolated
on each sampling plate. These strains
were streaked on two GNA plates con-
taining 100 ppm kasugamycin and two
control plates without kasugamycin
amendment. After 3-days incubation at
28°C, growth on GNA with and without
kasugamycin was qualitatively com-
pared as a measure of strain sensitivity
to kasugamycin.
Results
The kasugamycin treatments,
with the exception of the Kasuran (kasu-
gamycin + copper hydroxide premix)
reduced bacterial spot severity on toma-
toes in the greenhouse (Table 1). While
not striking, differences between the
control and treatments with kasugamycin
were often statistically significant and
represented as much as 33% less disease
in some of the better treatments. There
did not appear to be a big advantage
gained by addition of copper or captan to
kasugamycin; these combination treat-
ments were statistically equivalent to
sprays of kasugamycin alone (Table 1).
In the first field assessment made
on plants in the Immokalee trial, kasu-
gamycin-treated plots had significantly
lower disease ratings than the control
plots (Table 2). For example, kasugamy-
cin alone had a disease rating of 10.5%
compared to 23.8% in the control plots,
less than half as much foliar damage.
However, none of the treatments con-
taining kasugamycin were any better
than the long-time grower standard, cop-
per + mancozeb.
No differences were found in any
of the yield data collected (Table 3), in-
cluding total number and weight of all
fruit harvested, number and weight of
marketable and unmarketable fruit, and
nonmarketable fruit as a percentage of
the total weight harvested.
None of the 12 strains of X. vesi-
catoria (X. perforans) collected on April
26 showed any signs of kasugamycin
resistance (Table 4). However, at least
one strain isolated on May 21 from the
kasugamycin plots was highly resistant
and several others showed some growth
on kasugamycin-amended plates.
Discussion
Under relatively low-disease
pressure in the greenhouse and the field,
kasugamycin provided a level of control
of bacterial spot of tomato. In both in-
stances, the degree of control was about
the same as that observed with the in-
dustry standard of copper + mancozeb. If
registered for use on tomatoes in the
United States, it would give growers an
alternative chemical control option that
could be rotated with copper + man-
cozeb as part of an integrated program to
manage this disease. At high-disease
pressure as experienced after the second
inoculation in the field, kasugamycin, as
well as the grower standard, did not sig-
nificantly reduced disease levels. Jones
and Jones (1985) also found that avail-
able chemical control measures were
relatively ineffective when bacterial spot
epidemics were intense.
Addition of copper as a formu-
lated additive or a tank mix option with
kasugamycin did not improve the effi-
cacy of this antibiotic. Similarly, captan
had no synergistic effect when tank-
mixed with the kasugamycin. Most of
the disease control observed in treat-
ments containing kasugamycin had to
come from the kasugamycin itself.
50

Development of resistance is a
major concern any time antibiotics are
used routinely in the field. Rapid devel-
opment of resistance to streptomycin
rendered this once promising chemical
ineffective within a few seasons on sev-
eral vegetable crops (Stall and Thayer,
1962). This resistance may be relatively
long-lived even after streptomycin use
has been discontinued (Pohronezny et
al., 1994).
A total of nice applications of
kasugamycin were made to Immokalee
tomato plots. By the end of the experi-
ment, at least one strain of the bacterial
spot pathogen was found to be highly
resistant to kasugamycin in an in vitro
screening test. Some growth on kasu-
gamycin-amended plates was observed
with several other strains. This fairly
rapid development of resistance parallels
that experienced with streptomycin in
the past. If kasugamycin does get a full
label in the US, it will have to be used
very judiciously if it is to remain a viable
option in an integrated disease manage-
ment program for bacterial spot of
tomato.
Table 1. Ratings of bacterial spot of tomato in the greenhouse with kasugamycin at the
Everglades Research and Education Center, 2006-2007 x
Percent disease y
Treatment (formulation/100gal) Exp. 1 Exp. 2
Untreated (control).................................................. 35.2 a z
36.8 a
Kasuran 50WP (1.72 lb)………………………….. 35.2 a 36.2 a
Kasumin 2L (64 fl. oz)…………………………… 27.4 b 23.0 ab
Kasumin 2L (64 fl. oz) + Captan 50WP (3 lb)........ 23.8 b 29.3 b
Kasumin 2L (64 fl. oz) + Kocide 2000 (2 lb)……... 27.6 b 28.6 b
x Experiments consisted of three tomato plants, cv. BHN586, nested within each of four
blocks for each treatment. Plants were sprayed with chemicals twice at a 7-day interval
before inoculation with a suspension of Xanthomonas perforans, strain T193 and twice
at a 7-day interval after inoculation.y Ratings consisted of a visual estimate of the percentage of foliage lost prematurely or
covered with lesions combined into one rating.z Means followed by the same letter are not statistically different, according to Waller-
Duncan’s k-ratio t-test at P# 0.05.
51

Table 2. Rations of bacterial spot severity in field plots treated with kasugamycin at Im-
mokalee, spring 2007 w
Rating (% foliar disease)x
Treatment (formulation/100gal) April 26 May 21
Untreated (control).................................................. 23.8 a y
40.5 N.S.z
Kasuran 50WP (1.72 lb)………………………….. 17.3 ab 40.0
Kasumin 2L (64 fl. oz) + Kocide 2000 (2 lb)……... 16.0 ab 33.2
Kasumin 2L (64 fl. oz)…………………………… 10.5 b 42.0
Kasumin 2L (64 fl. oz) + Captan 50WP (3 lb)........ 10.3 b 35.2
Kocide 2000 (2 lb) + Manzate 75DF (2 lb)..……... 8.0 b 28.8
w Treatments were replicated 4 times in a randomized complete block design. All plants
were inoculated with a 1 x 108 CFU/ml suspension of the bacterial spot pathogen on
April 12 and 27. Plots were sprayed 9 times on a weekly basis using a tractor-mounted
high-clearance sprayer.x Ratings consisted of a visual estimate of the percentage of foliage lost prematurely or
covered with lesions combined into one rating.y Means followed by the same letter are not statistically different, according to Waller-
Duncan’s k-ratio t-test at P# 0.05.z N.S.= no significant difference found at P # 0.05.
52
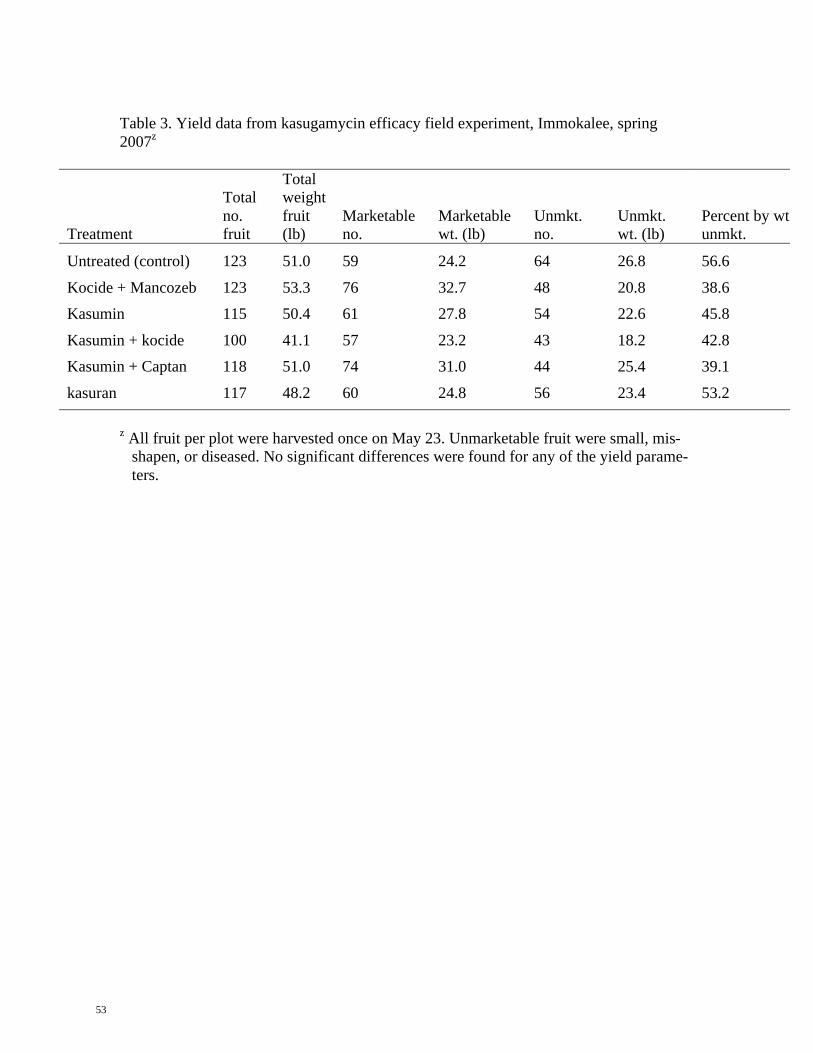
Table 3. Yield data from kasugamycin efficacy field experiment, Immokalee, spring
2007z
Treatment
Total
no.
fruit
Total
weight
fruit
(lb)
Marketable
no.
Marketable
wt. (lb)
Unmkt.
no.
Unmkt.
wt. (lb)
Percent by wt
unmkt.
Untreated (control) 123 51.0 59 24.2 64 26.8 56.6
Kocide + Mancozeb 123 53.3 76 32.7 48 20.8 38.6
Kasumin 115 50.4 61 27.8 54 22.6 45.8
Kasumin + kocide 100 41.1 57 23.2 43 18.2 42.8
Kasumin + Captan 118 51.0 74 31.0 44 25.4 39.1
kasuran 117 48.2 60 24.8 56 23.4 53.2
z All fruit per plot were harvested once on May 23. Unmarketable fruit were small, mis-
shapen, or diseased. No significant differences were found for any of the yield parame-
ters.
53

Table 4. Resistance of strains of Xanthomonas vesicatoria (perforans) to kasugamycin on
two sampling dates, Immokalee, spring 2007 y
Resistance estimatez
Treatment Rep
26 April 21 May
Untreated (control) 1
2
3
-
-
-
+
+
-
Kasumin 1
2
3
-
-
-
+++
++
+
Kasumin + Kocide 1
2
3
-
-
-
+
-
-
y Resistance based on growth of strains on plates of glucose-nutrient agar amended with
100 ppm kasugamycin compared to growth on GNA plates without kasugamycin.z Qualitative estimate of growth of bacteria on kasugamycin-amended plates were
equals no growth, + equals slight growth, and +++ equals very heavy growth. Plates
were streaked in duplicate.
Literature
Jones, J. B., Jones, J.P., 1985. The
effect of bactericides, tank mix-
ing time and spray schedule on
bacterial leaf spot of tomato.
Proc. Fla. State Hortic. Soc. 98,
244-247.
Leben, C. Daft, G.C. Schmitthenner,
A.F., 1968. Bacterial blight of
soybeans: Population levels of
Pseudomonas glycinea in rela-
tion to symptom development.
Plytopathology 50, 1143-1146.
Marco, G. M., Stall, R.E., 1983.
Control of bacterial spot of pep-
per initiated by strains of Xan-
thomonas campestris pv. vesica-
toria that differ in sensitivity to
copper. Plant Dis. 67, 779-781.
Pernezny, K., Datnoff, L. E., Muel-
ler, T., Collins, J.,1996. Losses in
fresh-market tomato production
in Florida due to target spot and
bacterial spot and the benefits of
protectant fungicides. Plant Dis.
80, 559-563.
Pernezny, K., Nagata, R., Havranek,
N., Sanchez, J. 2007. Compari-
son of two culture media for de-
termination of copper resistance
of Xanthomonas strains and their
usefulness for prediction of con-
trol with copper bactericides.
Crop Prot. 26 (in press).
Pohronezny, K., Sommerfeld, M.L.,
Raid, R.N., 1994. Streptomycin
54

resistance and copper tolerance
among strains of Pseudomonas
cichorii in celery seedbeds. Plant
Dis. 78, 150-153.
Pohronezny, K., Stall, R.E., Can-
teros, B.I, Kegley, M., Datnoff,
L.E., Subramanya, R., 1992.
Sudden shift in the prevalent race
of Xanthomonas campestris pv.
vesicatoria in pepper fields in
southern Florida. Plant Dis. 76,
118-120.
Pohronezny, K. Volin, R.B., 1983.
The effect of bacterial spot on
yield and quality of fresh market
tomatoes. HortScience 18, 69-70.
Stall, R. E., Thayer, P. L., 1962.
Streptomycin resistance of the
bacterial spot pathogen and con-
trol with streptomycin. Plant Dis.
Rep. 46, 389-392.
55

Evaluating Factors Affecting
Movement of the Silverleaf
Whitefly and Tomato Yellow Leaf
Curl Virus
David J. Schuster, James A. Taylor and
Craig D. Stanley
UF/IFAS, Gulf Coast Research and Educa-
tion Center, Wimauma
Jane E. Polston
UF/IFAS, Plant Pathology Department,
Gainesville
Sabine Grunwald
UF/IFAS, Soil and Water Science Depart-
ment, Gainesville
Abstract
The silverleaf whitefly, Bemisia ar-
gentifolii (also known as biotype B of the
sweetpotato whitefly, B. tabaci), is the key
insect pest of tomatoes in southern Florida.
Most damage associated with the whitefly is
due to the transmission of plant viruses, the
most damaging of which is tomato yellow
leaf curl virus (TYLCV). In the spring of
2006 and the spring of 2007, a 250-acre
field in Hillsborough County Florida was
used as a preliminary field site to determine
scouting techniques and monitor for silver-
leaf whiteflies and TYLCV. Whitefly adult
and nymphs density data and percent inci-
dence of plants with symptoms of TYLCV
data were collected on one geo-referenced
site per each 2.5 acres. Correlation analy-
ses indicated that early season silverleaf
whitefly adult counts made two and three
weeks previous to TYLCV incidence were
positively correlated. Reported is an evalua-
tion of geographic information systems
(GIS) and global positioning systems (GPS)
to map adult whitefly density and TYLCV
incidence. Also included are inverse dis-
tance weighted maps created by ArcMap
(ESRI, ArcGIS, ver. 9.x) graphically de-
picting TYLCV and adult whitefly densities.
Introduction
The silverleaf whitefly, Bemisia ar-
gentifolii (also known as biotype B of the
sweetpotato whitefly, B. tabaci), is an im-
portant economic pest in tropical and sub-
tropical climates. The silverleaf whitefly can
produce direct feeding damage in tomatoes,
but causes considerable yield loss in many
areas of the world due to its capability of
vectoring plant viruses. One important
sweetpotato whitefly vectored virus is to-
mato yellow leaf curl virus (TYLCV: genus
Begomovirus, family Geminiviridae).
TYLCV is vectored in a persistent manner
and causes damage to tomatoes worldwide.
Geographic information systems (GIS) and
global positioning systems (GPS) can be
used to monitor and predict the spread of
whiteflies and TYLCV, spatially and tempo-
rally.
The silverleaf whitefly has wide host
range and feeds by sucking plant juices from
the underside of leaves. Adults look like tiny
white moths and fly when the leaf is dis-
turbed. Eggs are light colored and are laid
on the underside of leaves and hatch in
around 16 to 38 days depending on envi-
ronmental conditions. Hot and dry condi-
tions can favor the whitefly and they can
disperse over long distances through wind
which can lead to whitefly/TYLCV man-
agement problems.
TYLCV can be a devastating disease
causing yield loss due to abscission of flow-
ers and severe plant stunting. Other symp-
toms include yellow (chlorotic) leaf edges,
upward leaf cupping, leaf mottling, and re-
duced leaf size. TYLCV was first recorded
in the Middle East, Africa, and Southeast
Asia. It has recently spread to the Caribbean
and into Florida by the late 1990s. TYLCV
has a broad host range from several families
including Fabaceae (common bean, lentil),
56

Malvaceae (cheeseweed), Solanaceae (to-
mato, pepper). TYLCV is not transmitted by
seed or mechanically. The TYLCV vector,
silverleaf whitefly acquires the virus as an
adult and is capable of harboring the virus
for several weeks. It takes an estimated 15-
30 minutes of feeding for the whitefly to ac-
quire the virus. After an incubation period in
the insect of about 24 hours TYLCV can be
spread to other plants. Symptom expression
in tomato is approximately 2-3 weeks.
GIS are software tools which allow
for storage, analysis, synthesis, and output
of spatial data. Georeferenced data which is
obtained by GPS can be incorporated into a
GIS, and provides the framework for geo-
statistical analysis. Historically, GIS has
been used for forest and rangeland entomol-
ogy, but recently it has been applied to man-
age insect pests in agricultural systems. Us-
ing GIS, entomologists have been able to
relate insect populations to biological vari-
ables and other features of the landscape.
Further applications include creating inter-
polation maps of insect counts at un-
sampled locations. Interpolation methods
include nearest neighbor, inverse distance
weighting, and spatial prediction models in-
cluding Kriging. GIS can also perform geo-
statistical analyses which can describe cor-
relations through space and/or time and
could be important for developing manage-
ment strategies for whiteflies and TYLCV.
Benefits of using GIS can include in-
creased visual expression of ecological vari-
ables, geostatistical analysis of spatiotempo-
ral data and applications into the manage-
ment of agricultural pests such as the sil-
verleaf whitefly/TYLCV as an area-wide
pest management system. Current objectives
include: analyze spatio-temporal data from
2006-2007 grower field site in Hillsborough
Co. Florida, evaluate correlations between
silverleaf whitefly and TYLCV, create in-
terpolation maps of un-sampled locations in
the grower’s field and acquire new field
sites to further expand research possibilities
on an area-wide scale.
Methods
In the spring of 2006 and the spring
of 2007, a 250 acre field in Hillsborough
County Florida was used as a preliminary
field site to determine proper scouting and
data collection techniques. Sampling in-
cluded adult whitefly counts (total number
of adults per 20 leaves on 6 plants), whitefly
nymphs (total number of nymphs on a ter-
minal leaflet per each of 6 plants), and
TYLCV incidence (visual inspection of 50
plants). Sample points were located 2.5
acres apart in a grid-like fashion within each
block and were georeferenced. Sampling
points were sampled twice weekly through-
out the season, although certain blocks were
planted and harvested earlier than other
blocks and were not sampled on every sam-
pling date, pesticide application kept scouts
from entering blocks, and rain events post-
poned scouting efforts.
In 2006, the grower’s field was di-
vided into blocks and the scouts sampled
randomly selected sites throughout each
block. Scouting was initiated on January 14
and continued biweekly until June 9. Data
points varied within blocks from each sam-
pling date, which created a problem using
temporal analysis data. Even though the data
points moved throughout the 2006 fall sea-
son, maps could be created and analyzed
spatially.
In 2007, the grower’s field was di-
vided into blocks but sampling points re-
mained constant throughout the season (i.e.,
the same 6 plants were scouted and the same
50 plants were evaluated for TYLCV inci-
dence). Scouting was initiated on February 2
and continued biweekly until May 25.
Spatiotemporal analysis of 2006-
2007 data was analyzed using ArcMap
(ESRI, ArcGIS, ver. 9.x). Maps were cre-
ated using ArcMap (ESRI, ArcGIS, ver. 9.x)
57

with inverse distance weighting (IDW)
which estimates predicted values based on
the distance they are away from a sampled
point. Correlations were conducted with
Pearson’s correlation analysis (SAS).
Results
From 2007, adult whitefly counts 2
weeks and 3 weeks prior to TYLCV inci-
dence were positively correlated over all
dates and sampled points (CORR = 0.71,
P = 0.0001, N = 346) (CORR = 0.68, P =
0.0001, N = 346), respectively. Other dates
were also positively correlated. Most of
these dates occurred during the early season
when adult whitefly numbers were low.
Correlations represented with a map
of interpolated values are the best way to
view georeferenced data without geostatisti-
cal analysis, as the correlations can tell us
something about the interpolated maps. Fig-
ures 1, 2, 3, and 4 show two dates of
TYLCV incidence overlaid with contour
maps of interpolated adult whitefly counts.
TYLCV incidence is the solid color scheme
with the adult whitefly density as the con-
tour or isolines. Figure 1a depicts the corre-
lation of TYLCV incidence on February 27,
2007 and adult whiteflies 21 days before
(CORR = 0.77, P = 0.0052, N = 11). Figure
1b graphically depicts the correlation of
TYLCV incidence on February 27, 2007 and
adult whiteflies 14 days before (CORR =
0.93, P = 0.0001, N = 11). Notice the hot
spots or bulls eyes of adult whiteflies that
aren’t represented by TYLCV as indicated
in Figure 1a and 1b by arrows. Figure 2a
describes the correlation of TYLCV inci-
dence on March 2, 2007 and adult whiteflies
21 days prior (CORR = 0.77, P = 0.0052, N
= 11). Figure 2b illustrates the correlation of
TYLCV incidence on March 2, 2007 and
adult whiteflies 17 days prior (CORR =
0.93, P = 0.0001, N = 11). Figure 3a shows
the correlation of TYLCV incidence on
April 13 and adult whiteflies 21 prior
(CORR = 0.43, P = 0.13). Figure 3b shows
the correlation of TYLCV incidence on
April 13 and adult whiteflies 14 days before
(CORR = 0.29, P = 0.38). Figure 4a shows
the correlation of TYLCV incidence on May
8 and adult whiteflies 20 days prior (CORR
= -0.23, P = 0.49). Figure 4b shows the cor-
relation of TYLCV incidence on May 8 and
adult whiteflies 14 days before (CORR = -
0.05, P = 0.87). Note the change of scale in
all figures.
Discussion
Early season scouting dates showed
the highest correlations. Also, correlations
between adult whitefly counts 2 and 3 weeks
before TYLCV incidence were most impor-
tant due to the length of time between in-
oculation and symptom expression. These
correlations varied in significance over dates
throughout the season and within short time
frames (i.e., significant at 14 days, not sig-
nificant at 17, and then significant at 21
days). Variation was inevitable as TYLCV
incidence data were held constant and bio-
logical factors such as weather, crop phe-
nology, or insecticide applications could in-
fluence the distribution of whiteflies. Also,
whitefly density may not have direct impact
on TYLCV spread as one viliferous whitefly
can spread TYLCV to multiple plants.
Figure 1a and 1b show whitefly hot-
spots (arrows) that aren’t represented by
TYLCV. This indicates that not all whitefly
populations within the field are viliferous.
These hotspots continue throughout the sea-
son. Also, notice that there are virus hot-
spots that aren’t depicted with adult
whiteflies. The viliferous whiteflies that
caused these virus hotspots could have been
killed/moved by insecticides, moved by
wind, or moved due to changing crop phe-
nology within the field. Note the scale of
TYLCV incidence in Figures 1 and 2 and
see how TYLCV incidence increased as the
epidemic progresses into April (Fig. 3). We
58

can see that the early season patch of
TYLCV (Figures 1 and 2) is not represented
in the later season (Figures 3 and 4), indi-
cating that the initial source of TYLCV and
whiteflies may not have impacted the larger
infection area around the northeast quadrant
of the field. This implicates that there was
more than one area of initial whitefly and
virus spread. The interpolated maps from the
end of the season suggest an influx of virus
coming from the northeast and moving
down and across the field. A more thorough
knowledge of the distribution of the virus in
the environment and/or movement of secon-
dary spread throughout the field will have to
be evaluated. Also, wind direction and speed
will be evaluated to determine their effect on
whitefly dispersal and subsequent virus
spread.
From the figures, we see the graphi-
cal mapping power of GIS, but also some of
the limitations in using it as a single analyti-
cal tool. We can see from the interpolated
maps created by IDW that bulls eyes are
created around data points. This is an inher-
ent problem with using IDW. It’s a good
first tool at viewing the data visually, but
IDW can be misleading. To solve this prob-
lem future research will be conducted using
spatial prediction models that can deal with
the statistical relationships of the data. These
methods include Kriging, which uses auto-
correlation or the statistical relationship
among measured points to create its inter-
polation maps. Kriging also has the ability to
create a standard error map that would give
us some measure of the accuracy of the pre-
diction surface. Kriging allows for the trans-
formation of the data if needed. In general,
Kriging allows for much more flexibility
over IDW in data presentation but is inher-
ently more complicated.
We will use the 2.5 acre sample size
distribution again for future research as it
gave us the spatial resolution needed to cre-
ate interpolation maps at the field scale. One
of the largest factors in deciding the amount
and intensity of sample size is the monetary
costs of those samples. Our sampling meth-
ods are time intensive and it takes two qual-
ity scouts a full day to take 125 samples or
one field 250 acres. To add more acreage
to our program, which would be required for
an area-wide management program, we
could either alter our scouting intensity and
reduce the resolution of created interpolation
and geostatistical analysis, or add scouts.
These are both problematic as the introduc-
tion of scouts increases variability in scout-
ing data and increases monetary costs, and
decreasing the scouted samples per area
would reduce resolution on the field scale in
our analysis. This has implications on how
we would conduct this research program for
managing whiteflies and TYLCV at the field
level or expansion into an area-wide man-
agement program.
There is importance in understanding
whitefly dispersal and TYLCV spread. GIS
gives us the power to both graphically visu-
alize and analyze the field data using geo-
statistics. By expanding our research with
more fields we might be able to add to our
current management program by better un-
derstanding where silverleaf whiteflies and
TYLCV originate and how they move
through the field.
59

Figure 1. Inverse distance weighting (IDW) maps of adult whitefly density overlaid on TYLCV
incidence from selected dates in 2007.
A
E
60
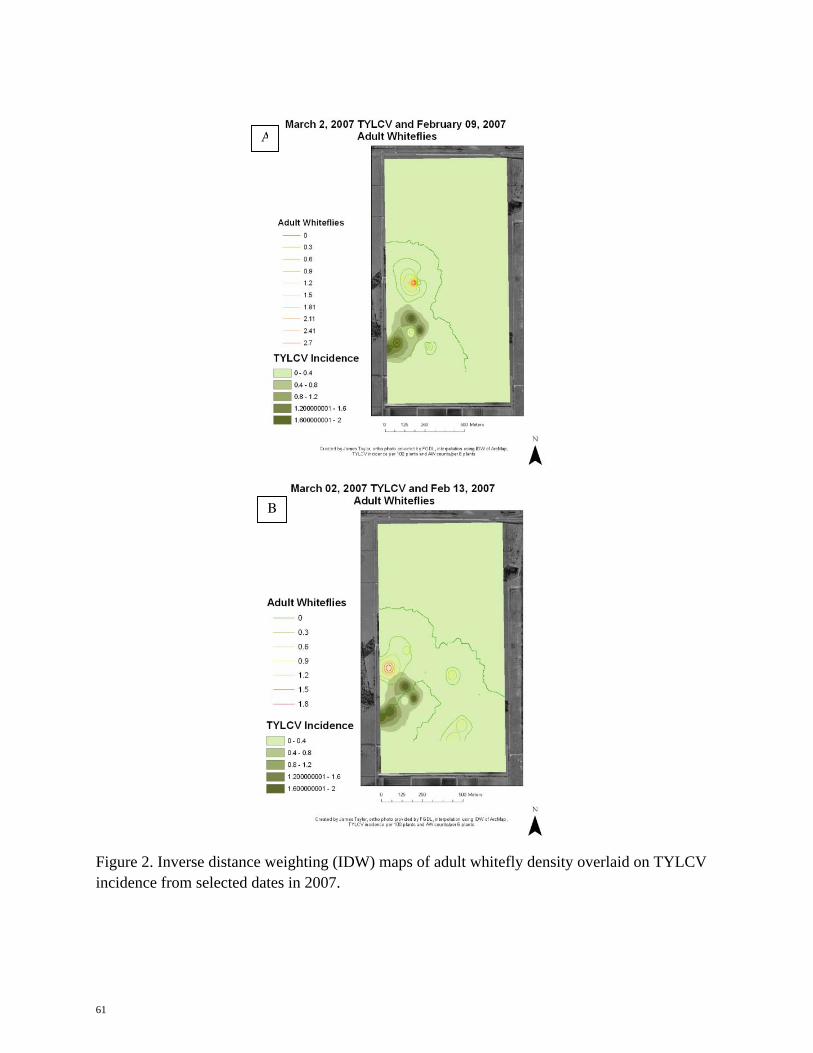
Figure 2. Inverse distance weighting (IDW) maps of adult whitefly density overlaid on TYLCV
incidence from selected dates in 2007.
A
B
61

Figure 3. Inverse distance weighting (IDW) maps of adult whitefly density overlaid on TYLCV
incidence from selected dates in 2007.
A
B
62

Figure 4. Inverse distance weighting (IDW) maps of adult whitefly density overlaid on TYLCV
incidence from selected dates in 2007.
A
B
63

Monitoring Resistance of the
Silverleaf Whitefly to Insecticides
David J. Schuster
UF/IFAS, Gulf Coast Research and Educa-
tion Center, Wimauma
Mark A. Mossler
UF/IFAS, Pesticide Information
Office, Gainesville
Abstract
Resistance monitoring for the neoni-
cotinoids Admire and Platinum for the sil-
verleaf whitefly (SLWF), Bemisia argenti-
folii Bellows & Perring was continued in
2007. Monitoring for resistance to the
neonicotinoids Assail and Venom, the py-
rethroids bifenthrin, and the chlorinated hy-
drocarbon endosulfan were initiated in 2007.
Resistance to the neonicotinoids was esti-
mated in the laboratory using a cut leaf peti-
ole bioassay method and resistance to the
pyrethroids and chlorinated hydrocarbons
was estimated using a residual vial bioassay
method. All bioassays were conducted using
adults reared from foliage infested with
nymphs that had been collected from each
crop field. Standard probit analyses were
used to estimate the LC50 values for a labo-
ratory colony and for each field population.
The relative susceptibility (RS50) of each
field population was calculated by dividing
the LC50 values of the field populations by
the LC50 value of the laboratory colony. The
average RS50 value for Admire for 2007 de-
creased from 2006 about 30% while that for
Platinum decreased about 60%. One popu-
lation was particularly high for Admire with
an RS50 value of 85.8. All populations
evaluated were susceptible to Venom and
Assail. Five of the six populations evaluated
for susceptibility to bifenthrin were highly
resistant with an average RS50 value of
about 141, while the seven populations
evaluated with endosulfan were susceptible
with an average RS50 value of 1.7. The re-
sults emphasize more than ever that growers
should strictly observe the guidelines for
managing the silverleaf whitefly, whitefly
vectored plant viruses, and insecticide re-
sistance.
Introduction
The silverleaf whitefly (SLWF), Be-
misia argentifolii Bellows & Perring [also
known as biotype B of the sweetpotato
whitefly, B. tabaci (Gennadius)] and tomato
yellow leaf curl virus (TYLCV) remain the
key pests of tomatoes in southern Florida.
Insecticides, particularly the neonicotinoids
(Admire Pro®, imidacloprid, Bayer Crop-
Science, Research Triangle Park, NC; As-
sail®, acetamiprid, Cerexagri Inc., King of
Prussia, PA; Platinum®, thiamethoxam,
Syngenta Crop Protection, Inc., Greensboro,
NC; and Venom®, dinotefuran, Valent
U.S.A. Corp., Walnut Creek, CA), remain
integral tools for the management of the
pests. Because of the potential of the
whitefly to develop resistance to the insecti-
cides, a program to monitor the susceptibil-
ity of field populations of the SLWF to Ad-
mire and Platinum using a cut leaf petiole
method was conducted from 2000 to 2006
(Schuster and Thompson 2001, 2004;
Schuster et al. 2002, 2003, 2006). Suscepti-
bility of the SLWF to Admire decreased
from 2000 to 2003, increased in both 2004
and 2005, and then decreased tremendously
in 2006. Susceptibility of the SWLF to
Platinum decreased from 2003 to 2005 and
then, as with Admire, susceptibility de-
creased dramatically in 2006. Because of the
reduced susceptibility indicated in 2006, the
resistance monitoring program was contin-
ued in 2007 and expanded to include the
other neonicotinoids Assail and Venom as
well as pyrethroids and chlorinated hydro-
carbons.
64

Materials and Methods
Resistance to the neonicotinoids was
estimated in the laboratory using a cut leaf
petiole bioassay method (Schuster and
Thompson 2001, 2004; Schuster et al. 2002,
2003, 2006). Cotton seedlings grown under
insect-free greenhouse conditions were used
at the two true leaf stages. The petioles of
the cotton leaves were cut and placed imme-
diately in the vials containing seven differ-
ent concentrations of insecticides (viz; 10.0,
5.0, 2.5, 1.25, 0.625, 0.3125, 0.15625 ppm
and control (de-ionized water). The petioles
were immersed individually in the insecti-
cide solutions for 24 hrs. After 24 hrs., ten
whitefly adults were confined on the under-
side of each leaf with clip cages (2 cm di-
ameter, 1 cm high) for another 24 hrs. The
mortality was assessed by tapping the ab-
domen of the whitefly adults and observing
the movements under a microscope. Mortal-
ity was defined as any adults that were un-
able to coordinate their movements and,
hence, was unable to walk or fly.
Resistance to the pyrethroids and
chlorinated hydrocarbons was estimated us-
ing a residual vial bioassay method. Bifen-
thrin was selected as a representative pyre-
throid and endosulfan as a chlorinated hy-
drocarbon. Serial dilutions of bifenthrin and
endosulfan as technical grade compounds
were made with methanol at 1000, 300, 100,
30, 10, 3, 1 and 0.3 ppm, and 0.24ml of each
dilution was pipetted individually into sepa-
rate 20 ml vials. The vials were placed on a
hot dog roller, which had the heating ele-
ment disconnected, and placed under a fume
hood. The vials were rotated for about 15-25
minutes until the methanol had evaporated.
The vials were capped and stored at room
temperature until use. To conduct the bioas-
says, 10 whitefly adults were carefully aspi-
rated into each vial of each concentration
and capped with a ventilated lid. The vials
were held upright in a temperature con-
trolled room for 6 hr when mortality was
determined. Mortality was defined as adults
that could move but were unable to fly
and/or leave the bottom of the vial when
gently touched with a fine paintbrush. There
were 3-4 replications (vials) for each con-
centration for each population.
All bioassays were conducted using
adults reared from foliage infested with
nymphs that had been collected from each
crop field. In some cases, the populations
were maintained in the laboratory for two to
three generations in order to obtain enough
adults to conduct the bioassays. Standard
probit analyses (SAS Institute 1989) were
used to estimate the LC50 values (the con-
centration estimated to kill 50% of the
population) for a laboratory colony and for
each field population. The laboratory colony
used as a susceptible standard in this study
has been in continuous culture since the late
1980s without the introduction of whiteflies
collected from the field and, therefore,
would be expected to be particularly sus-
ceptible to insecticides. The relative suscep-
tibility (RS50) of each field population com-
pared to the laboratory colony was calcu-
lated by dividing the LC50 values of the field
populations by the LC50 value of the labo-
ratory colony. Increasing values greater than
1 suggest decreasing susceptibility in the
field population. While values approaching
8 could indicate decreasing susceptibility of
the whiteflies, such variability is not unex-
pected when comparing field-collected in-
sects with susceptible, laboratory-reared in-
sects. Values of 10 or greater, especially
those of 20 or higher, are sufficiently high to
draw attention.
Results and discussion
The average RS50 value for Admire
for 2007 decreased from 2006 about 23%
while that for Platinum decreased about 60%
(Fig. 1). One population, NECollier, was
particularly high for Admire with an RS50
value of 85.8 (Table 1). This is the highest
60

RS50 ever identified in 8 years of monitor-
ing, especially considering that the popula-
tion had been reared for two generations (3rd
generation) in the lab without further expo-
sure to Admire. Research in the past has in-
dicated that reduced susceptibility declines
as the whiteflies are reared on successive
generations of plants not treated with Ad-
mire (Schuster and Thompson 2004). The
NECollier population was also higher for
Platinum. Some other populations were also
high for both Admire and Platinum includ-
ing Homestead, SWFREC (Southwest Flor-
ida Research & Education Center, Immoka-
lee), and TR 3. However, there were two
populations that were higher for Platinum
but not Admire (Apollo Beach and
TomG#2) and one that was higher for Ad-
mire but not Platinum (Parrish-1). These re-
sults may suggest that there isn’t cross toler-
ance between the two neonicotinoids but
that there may be simultaneous selection for
tolerance. Previous monitoring had sug-
gested a similar conclusion (Schuster and
Thompson 2004). All 10 populations evalu-
ated for susceptibility to Venom were sus-
ceptible, even some populations that were
higher for Admire and/or Platinum. The six
populations evaluated with Assail were sus-
ceptible, although some, especially NECol-
lier, were higher with either Admire and/or
Platinum.
Five of the six populations evaluated
for susceptibility to bifenthrin were highly
resistant, with RS50 values greater than 100,
even though five of six were evaluated in the
third generation (Table 1). All seven popu-
lations evaluated for susceptibility to endo-
sulfan were highly susceptible, with RS50
values all less than 3, although all but one
were evaluated in the third or fourth genera-
tion.
The results emphasize that, even
though average RS50 values for Admire and
Platinum declined from 2006 to 2007, there
still remain some populations that are prob-
lematic. At least one population, NECollier,
could be classified as resistant. With the ac-
tive ingredient of Admire, imidacloprid, no
longer under patent protection, the prolif-
eration of generic formulations of imidaclo-
prid can be expected and is already occur-
ring. This not only will likely cause prices
for products containing imidacloprid to de-
crease, but will also likely result in reduced
prices for the other neonicotinoids. Reduced
costs of neonicotinoids could cause growers
to use higher rates of the products and/or
tempt them to make multiple applications.
Because of this new threat to the sustain-
ability of the neonicotinoids coupled with
the high level of resistance observed for the
pyrethroid bifenthrin, growers are encour-
aged even more than ever to strictly observe
the guidelines for managing the silverleaf
whitefly, whitefly vectored plant viruses,
and insecticide resistance that have been
formulated and promoted by the University
of Florida, and representatives of the ag-
richemical, commodity, and consultant in-
dustries (Schuster et al. 2007). Chief among
these guidelines is the establishment of a
minimum two-month crop-free period dur-
ing the summer off-season and the creation
of a crop-free break in time and/or space
between fall and spring crops. Growers
should also use the pyrethroids judiciously.
If growers fail to adhere strictly to the
whitefly management guidelines, they can
only expect levels of tolerance/resistance to
increase and difficulty in managing
whiteflies and TYLCV to escalate.
Acknowledgments
The authors wish to express their ap-
preciation to Sabrina Spurgeon and Aaron
Shurtleff for conducting the 2007 bioassays;
to Phil Stansly, Dak Seal, Henry Yonce,
Sarah Hornsby and Leon Lucas for identi-
fying and/or collecting whitefly samples for
the 2007 monitoring; and to Bayer Crop-
Science, Cerexagri Inc., Syngenta Crop
61

Protection, Valent Agricultural Products and
the Florida Tomato Committee for providing
funding for the neonicotinoid resistance
monitoring. Appreciation also is expressed
to representatives of the Florida Tomato
Committee, Florida Fruit and Vegetable As-
sociation, Bayer CropScience, Syngenta
Crop Protection, Cerexagri Inc., Glades
Crop Care, Agricultural Crop Consulting,
Agri-Tech Services, KAC Agricultural Re-
search, and Integrated Crop Management,
and to UF/IFAS personnel Alicia Whidden,
Gene McAvoy, Jim Price and Phil Stansly
for their participation in the Resistance
Management Working Group and for their
many contributions to the whitefly and re-
sistance management recommendations.
References Cited
SAS Instititute Inc. 1989. SAS/STAT User’s
Guide, Version 6, Fourth Edition,
Vol. 2, SAS Institute Inc., Cary, NC.
Schuster, D. J. and S. Thompson. 2001.
Monitoring susceptibility of the sil-
verleaf whitefly to imidacloprid, pp.
16-18. In P. Gilreath and C. S.
Vavrina [eds.], 2001 Fla. Tomato In-
stitute Proc., Univ. Fla., Gainesville,
PRO 518.
Schuster, D. J. and S. Thompson. 2004. Sil-
verleaf whitefly resistance manage-
ment update, pp. 19-25. In P. Gil-
reath and W. H. Stall [eds.], Fla.
Tomato Institute Proc., Univ. Fla.,
Gainesville, PRO 521.
Schuster, D. J., S. Thompson, P. A. Stansly
and J. Conner. 2002. Update on in-
secticides for whitefly and leafminer
control, pp. 51-60. In P. Gilreath and
C. S. Vavrina [eds.], 2002 Fla. To-
mato Institute Proc., Univ. Fla., PRO
519.
Schuster, D. J., S. Thompson and P. R. Gil-
reath. 2003. What’s up with all these
whiteflies?, pp. 12-19. In P. Gilreath
and W. H. Stall [eds.], Fla. Tomato
Institute Proc., Univ. Fla., PRO 520.
Schuster, D. J., R. Mann and P. R. Gilreath.
2006. Whitefly resistance update and
proposed mandated burn down rule.
Pp. 24-28. In K. Cushman and P.
Gilreath [eds.], Fla. Tomato Institute
Proc., Univ. Fla., PRO 523.
Schuster, D. J., P. A. Stansly, J. E. Polston,
P.s R. Gilreath and E. McAvoy.
2007. Management of whiteflies,
whitefly-vectored plant virus, and in-
secticide resistance for vegetable
production in southern Florida. Univ.
Fla., IFAS, Fla. Coop. Ext. Serv.,
ENY-735 (IN695),
http://edis.ifas.ufl.edu
62

0
5
10
15
20
25
30
35
2000 2001 2002 2003 2004 2005 2006 2007
Avg
RS
50
9 (2) 14 (4) 10 (8)
6.7
9.9
14.7
3.7
3 (0) 11 (1)
6.1
2 (0)
2.5
29.9
7 (6)
Fig. 1. Monitoring relative susceptibility (RS 50 ) of whitefly adults from
nicotinoid -treated crops to Admire (A) or Platinum (B) using a laboratory
bioassay. Numbers at the top of the bars are the average RS 50 values and
the numbers inside the bar are the number of populations bioassa yed and
the number with RS 50 values >10 (in parentheses).
0
5
10
15
20
25
30
2003 2004 2005 2006 2007
27.2
6.1
2.01.85 (4)7 (1)3 (0)8 (0)
A
B
Avg
RS
50
14 (8)
23.1
16 (8)
11.1
63

Table 1. Results of resistance bioassays of silverleaf whitefly populations collected from west central, southwest and southeast Florida
to neonicotinoid, pyrethroid, and chlorinated hydrocarbon insecticides, Spring 2007.
Neonicotinoids Pyrethroid Chlorinated
Population Generation Admire Assail Platinum Venom Bifenthrin Endosulfan
site Crop Tested1
LC50 RS50 LC50 RS50 LC50 RS50 LC50 RS50 LC50 RS50 LC50 RS50
GCREC/Lab Tomato ---- 0.38 ---- 0.58 ---- 1.36 ---- 0.32 ---- 0.06 ---- 6.79 ----
GCREC/Lab2 Tomato ---- 0.56 ---- ---- ---- 1.70 ---- ---- ---- ---- ---- ---- ----
Apollo Beach Tomato 1st 2.75 7.3 ---- ---- 13.8 10.2 1.25 4.0 7.282
116.4 19.3 2.8
Collier-2 Tomato 2nd ---- ---- ---- ---- 25.4 18.7 ---- ---- ---- ---- ---- ----
F1 Tomato 2nd 11.73
30.9 ---- ---- 10.4 7.7 ---- ---- ---- ---- 10.13
1.5
FM Eggplant 1st 2.13 5.6 ---- ---- 6.58 4.8 ---- ---- ---- ---- 9.693
1.4
Homestead Tomato 2nd 10.73
28.3 ---- ---- 29.8 21.9 ---- ---- 1.873
29.8 9.073
1.3
HomesteadB Bean 2nd ---- ---- ---- ---- 4.37 3.2 ---- ---- ---- ---- ---- ----
HSRC Tomato 2nd ---- ---- ---- ---- 3.83 2.8 ---- ---- ---- ---- ---- ----
Myakka-1 Tomato 2nd 9.603
25.32 1.424
2.5 6.15 4.5 ---- ---- ---- ---- 10.04
1.5
Maykka-5 Tomato 1st ---- ---- 2.16 3.7 5.30 3.9 1.39 4.4 ---- ---- ---- ----
NECollier Tomato 2nd 32.53
85.8 0.683
1.2 31.1 22.9 ---- ---- 6.933
110.8 8.583
1.3
P 1&2 Potato 1st 1.01 1.8 1.602
2.8 24.82
18.2 1.192
3.8 ---- ---- ---- ----
P 9 Pepper 1st 3.20 5.70 ---- ---- 3.83 2.3 1.312
4.1 ---- ---- ---- ----
Parrish-1 Tomato 1st 18.12
47.8 ---- ---- 8.82 6.5 2.21 7.0 15.13
240.9 10.83
1.6
SWFREC Watermelon 2nd 12.6 33.2 0.773
1.3 29.7 21.8 2.242
7.1 14.63
233.6 11.283
1.7
SWHendry Tomato 2nd 11.2 29.6 2.28 3.9 ---- ---- ---- ---- 7.133
114.1 ---- ----
T 5 Tomato 1st 2.43 4.33 ---- ---- ---- ---- 1.702
5.4 ---- ---- ---- ----
T 6 Tomato 2nd ---- ---- ---- ---- 8.00 5.9 1.28 4.0 ---- ---- ---- ----
TG12N Tomato 2nd ---- ---- ---- ---- ---- ---- 1.61 5.1 ---- ---- ---- ----
TomG#2 Tomato 1st 2.08 5.5 ---- ---- 14.3 10.5 0.90 2.8 ---- ---- ---- ----
TR 3 Tomato 1st 4.67 12.3 ---- ---- 19.7 14.5 ---- ---- ---- ---- ---- ----
1The first generation would be those whitefly adults emerging from the foliage collected in the field. The second and third generations
were reared on tomato plants in the laboratory that had not been treated with neonicotinoid.2The populations were tested in the 2
nd generation.
3The population was tested in the 3
rd generation.
4The population was tested in the 4
th generation.
64