tumormicroenvironmentmacrophageinhibitoryfactor ... ·...
TRANSCRIPT

Tumor Microenvironment Macrophage Inhibitory FactorDirects the Accumulation of Interleukin-17-producingTumor-infiltrating Lymphocytes and Predicts FavorableSurvival in Nasopharyngeal Carcinoma Patients*□S
Received for publication, March 30, 2012, and in revised form, August 1, 2012 Published, JBC Papers in Press, August 14, 2012, DOI 10.1074/jbc.M112.367532
Jiang Li‡§1,2, Hao-Yuan Mo¶1, Geng Xiong‡, Lin Zhang§, Jia He§, Zhou-Feng Huang¶, Zhi-Wei Liu‡, Qiu-Yan Chen¶,Zi-Ming Du‡, Li-Min Zheng‡§, Chao-Nan Qian¶�3, and Yi-Xin Zeng‡
From the ‡State Key Laboratory of Oncology in South China, Departments of ¶Nasopharyngeal Carcinoma and §Biotherapy, SunYat-sen University Cancer Center, Guangzhou 510060, China and the �Laboratory of Cancer and Developmental Cell Biology, VanAndel Research Institute, Grand Rapids, Michigan 49503
Background: Tumor microenvironments affect the progression of cancers.Results:We demonstrated that Th17 cells were accumulated in tumor tissues, and the tumor-derived MIF induced Th17 cellaccumulation and had clinical relevance in NPC.Conclusion: The cytokine MIF regulates intratumoral Th17 cell expansion and has prognostic value for NPC patients.Significance: The tumor microenvironment influences the clinical prognosis of NPC patients.
The accumulation of an intratumoral CD4� interleukin-17-producing subset (Th17) of tumor-infiltrating lymphocytes(TILs) is a general characteristic in many cancers. The relation-ship between the percentage of Th17 cells and clinical prognosisdiffers among cancers. The mechanism responsible for theincreasing percentage of such cells inNPC is still unknown, as istheir biological function. Here, our data showed an increase ofTh17 cells in tumor tissues relative to their numbers in normalnasopharynx tissues or in the matched peripheral blood of NPCpatients. Th17 cells in tumor tissue produced more IFN� thandid those in the peripheral blood of matched NPC patients andhealthy controls. We observed high levels of CD154, G-CSF,CXCL1, IL-6, IL-8, and macrophage inhibitory factor (MIF) outof 36 cytokines examined in tumor tissue cultures. MIF pro-moted the generation and recruitment of Th17 cells mediatedby NPC tumor cells in vitro; this promoting effect was mainlydependent on themammalian target of rapamycin pathway andwas mediated by the MIF-CXCR4 axis. Finally, the expressionlevel of MIF in tumor cells and in TILs was positively correlatedin NPC tumor tissues, and the frequency of MIF-positive TILswas positively correlated with NPC patient clinical outcomes.
Taken together, our findings illustrate that tumor-derived MIFcan affect patient prognosis, which might be related to theincrease of Th17 cells in the NPC tumor microenvironment.
Undifferentiated nasopharyngeal carcinoma (NPC),4 whichis associated with Epstein-Barr virus (EBV) infection, accountsfor over 95% of NPC in China (1–3). NPC tumor progression isusually accompanied by an increase of tumor-infiltrating lym-phocytes (TILs) and chronic inflammatory response due to thepresence on the tumor cells of EBV type II latent antigens,including latentmembrane proteins 1 and 2 (LMP1 andLMP2),EBV nuclear protein 1 (EBNA1), and BARF0 (4, 5). The tumormicroenvironment includes tumor cells and nonmalignantstromal cells such as fibroblasts, tumor-associated macro-phages, and lymphocytes (6). Th17 cells, a CD4� interleukin-17-producing subset of TILs, have been identified in bothhumans and mice by the secretion of proinflammatory cyto-kines, including the interleukins IL-17A, IL-17F, and IL-22.Crucial for the development of Th17 cells are the cytokinesIL-1�, IL-6, and transforming growth factor (TGF) �, and thetranscription factors STAT3 and retinoic acid-related orphanreceptor retinoic acid-related orphan receptor �� (7, 8).The clinical predictive value of Th17 cells differs among var-
ious cancers such as colon, liver, and ovarian cancer (9–11).The origin of Th17 cells and their biological function in humantumor microenvironments are still under investigation. Mech-anistic studies have revealed that many cytokines and chemo-kines, including CCL5, CCL17, CCL20, CCL22, MCP-1, and
* This work was supported by General Program Grants 30872981 and81172164 (to J. L.), State Key Program Grant 81030043 (to C.-N. Q.) from theNational Natural Science Foundation of China, Guangdong Province Nat-ural Science Foundation Grant 0151008901000156 (to J. L.), National HighTechnology Research and Development Program of China (863 Program)Grant 20060102A4002 (to C.-N. Q.), and Major State Basic Research Pro-gram (973 Project) of China Grant 2006CB910104 (to Y. X. Z.).
□S This article contains supplemental Figs. S1–S4, Tables S1–S3, and Methods.1 Both authors contributed equally to this work.2 To whom correspondence may be addressed: Dept, of Biotherapy, Sun Yat-
sen University Cancer Center, 651 Dongfeng East Rd., Guangzhou 510060,China. Tel.: 86-20-8734-3174; Fax: 86-20-8734-3392; E-mail: [email protected].
3 To whom correspondence may be addressed: Dept. of Nasopharyngeal Car-cinoma, Sun Yat-sen University Cancer Center, 651 Dongfeng East Rd.,Guangzhou 510060, China. Tel.: 86-20-8734-3457; Fax: 86-20-8734-3624;E-mail: [email protected].
4 The abbreviations used are: NPC, nasopharyngeal carcinoma; Th17 cells,CD4� interleukin-17-producing cells; MIF, macrophage inhibitory factor;TIL, tumor-infiltrating lymphocytes; PBMC, peripheral blood mononuclearcells; ISO-1, (SR)3-(4-hydroxyphenyl)-4,5-dihydro-5-isoxazole acetic acidmethyl ester; CsA, cyclosporin A; IHC, immunohistochemistry; NFAT,nuclear factor of activated T cell; mTOR, mammalian target of rapamycin;NP, nasopharyngeal; EBV, Epstein-Barr virus; Rh, recombinant human.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 287, NO. 42, pp. 35484 –35495, October 12, 2012© 2012 by The American Society for Biochemistry and Molecular Biology, Inc. Published in the U.S.A.
35484 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 287 • NUMBER 42 • OCTOBER 12, 2012
by guest on August 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from

IL-6, are expressed at a high level within the tumor microenvi-ronment in association with the accumulation of Th17 cells (3,11–14). However, the mechanistic relationships in NPCbetween the accumulation of Th17 cells, the cytokines releasedfrom tumor environments, and tumorigenesis and progressionremain unknown.In this study, our results indicated an elevated percentage of
Th17 cells in the NPC tumor microenvironment. We haveshown for the first time that microenvironment-derivedmacrophage inhibitory factor (MIF) promoted the generationand migration of Th17 cells mediated by tumor cells, and thepromotion effect of MIF on the generation and recruitment ofTh17 cells was mainly dependent on the mTOR pathway andwas mediated by the MIF-CXCR4 axis. Most importantly, theexpression of MIF in TILs predicted an improved clinical out-come in NPC. Taken together, our data provide novel evidencethat tumor-derived cytokines affect the development of cancerand its prognosis, and they are associated with Th17 cell subsetexpansion within the tumor microenvironment. These resultsdeepen our understanding of the inflammatory mechanismsinvolved in NPC progression.
EXPERIMENTAL PROCEDURES
Human Samples and Cell Lines—Tumor biopsy tissues andperipheral blood were collected from 21 newly diagnosed NPCpatients at Sun Yat-sen University Cancer Center (supplemen-tal Table 1) in 2009 and 2010, and peripheral blood was col-lected from 21 age-matched healthy individuals. The tumortissues were divided and cultured for a short period in RPMI1640 complete medium with a low dose (20 IU/ml) of IL-2 toobtain sufficient lymphocytes, afterwhich the cells were used togenerate TILs or in fluorescence-activated cell sorting (FACS).Peripheral blood mononuclear cells (PBMCs) from blood sam-ples taken fromNPC patients and healthy individuals were iso-lated and then frozen for FACS analyses. One hundred andeight paraffin-embedded tumor specimens from NPC patientswere collected in a previous study by our group, and detailedpatient information was provided (15). This study was con-ducted in accordancewith theHelsinki Declaration; all patientsand healthy controls signed a consent form approved by theResearch Ethics Committee of the Sun Yat-sen University Can-cer Center.An EBV-transformed LCL line and the NPC tumor cell lines
C666 (EBV�) and CNE2 (EBV�) were maintained in RPMI1640 medium containing 10% fetal bovine serum (FBS). Thenormal nasopharyngeal (NP) cell line NP69 was maintained inkeratinocyte-SFM medium (Invitrogen).Tumor-infiltrating Lymphocyte Culture—TILs were isolated
from NPC biopsy tissues and cultured in RPMI 1640 mediumcontaining 10% FBS supplemented with 2 mM L-glutamine, 1�M 2-mercaptoethanol, and recombinant human IL-2 (300IU/ml). They were grown using a rapid expansion protocol, asdescribed previously (16).Flow Cytometry and Antibodies—The expression of T cell
markers was analyzed by FACS after surface or intracellularstaining with anti-human-specific antibodies conjugated withfluorescent molecules. These human antibodies includedanti-CD4, anti-CCR6, anti-IL-17, anti-IFN�, anti-IL-2, anti-IL-
10, anti-IL-4, anti-TGF�, anti-CCR7, anti-CD45RO, anti-CD45RA, anti-CTLA-4, and anti-GITR.Thesewere conjugatedwith fluorescein isothiocyanate (FITC), phycoerythrin, allo-phycocyanin, or phycoerythrin-Cy7 (BD Biosciences or eBio-science). Recombinant human IL-2, TGF�, and IL-1� werepurchased from R&D Systems. Intracellular staining for IL-17and other cytokines was performed on T cells stimulated withphorbol 12-myristate 13-acetate and ionomycin for 4 h inRPMI1640 medium, and cytokine secretion was blocked by the addi-tion of brefeldin A (10 �g/ml, Sigma). After washing, cells werestained with anti-CD4, then were fixed and permeabilized withPerm/Fix solution (eBiosciences), and finally were stainedintracellularly with anti-IL-17A or with fluorescence-conju-gated antibodies for other cytokines. All stained cells were ana-lyzed on an FC500 flow cytometer, and the obtained data wereanalyzed with CXP software (Beckman Coulter).Co-culture for Generation of IL-17-producing Cells in Vitro—
Naive CD4� T cells were cultured in T cell medium containing100 IU/ml IL-2 and 10% FBS at a concentration of 1 � 105cells/well in a 48-well plate and were stimulated with plate-bound OKT3 (1 �g/ml). The naive CD4� T cells were co-cul-tured with irradiated NPC tumor cells or irradiated C666 cellstransfected with scrambled siRNA or pooled MIF-siRNAs at a1:1 ratio or in the presence of RhIL-1� (25 ng/ml), RhTGF� (3ng/ml), or RhIL-1� and RhTGF� together for 7 days. Half themediumwas replacedwith freshmedium on days 3 and 6. After7 days, the percentage of Th17 cells was determined by FACSanalysis. The experiments were repeated in triplicate. TheRhMIF (R&D Systems), the MIF small molecule antagonist(SR)3-(4-hydroxyphenyl)-4,5-dihydro-5-isoxazole acetic acidmethyl ester (ISO-1, Sigma), cyclosporin A (CsA, Sigma), orrapamycin (Enzo Life Science) was added to the medium atvarious concentrations in a subset of the samples as reportedbefore (17, 18).Cytokine Proteomic Profiling—NPC tumor cell lines (2.5 �
105 per well) or single cell suspensions from collagenase typeIV-digested NPC or normal NP tissues were cultured in 6-wellplates in 2%FBSRPMI 1640medium for 48 h. The supernatantswere then collected for detection of cytokines. Cytokines in theculture medium were measured using the Human CytokineArray Panel A (Proteome Profiler; R&D Systems). All cytokineprotein array analyses were performed according to the manu-facturer’s instructions. Positive controls were paired spotslocated in the upper left, lower right, and lower left corners ofeach array.Migration Assay—Migration assays of TILs from NPC
patients were performed using 24-well Transwell chemotaxisplates (5-mm pore size; Corning Costar) as described previ-ously (14). TILs were induced to migrate with supernatants fromvariouscell lines. Insomecases, theMIFsmallmoleculeantagonistISO-1(50–100 �M) or neutral anti-CXCR4 antibody (5–10 �M)(19)was added 150minbefore themigration assay. Themigrationof the Th17 cell subset was evaluated based on the percentage ofTh17cells in the inner andouter chambers.Theexperimentswererepeated in triplicate.Immunochemistry—MIF immunohistochemistry (IHC) was
carried out using a primary monoclonal mouse anti-humanMIF antibody (1:100 dilution, ab55445, Abcam), per manufac-
Clinical Relevance of Tumor-derived Cytokine MIF in NPC
OCTOBER 12, 2012 • VOLUME 287 • NUMBER 42 JOURNAL OF BIOLOGICAL CHEMISTRY 35485
by guest on August 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
turer’s instructions. The slides were scored independently bytwo pathologists blinded to the clinicopathological data of theNPC patients. The level of MIF expression in the tumor cellswas scored based on staining intensity (score 0–3) and area(score 1–4), as described previously (20); the final expressionlevel was scored as the product of the staining intensity and areascores. The level of MIF expression in lymphocytes wasobtained by counting the positively and negatively stained lym-phocytes in five separate �400 high power microscopic fieldsand calculating the mean percentage of positively stained lym-phocytes among the total lymphocytes per field. Mouse anti-human IgG1 (1:200 dilution, DAKO, Glostrup, Denmark) wasused as a negative control in this study.Statistics—All analyses were carried out with SPSS 13.0.
Numerical data are presented as the means � S.E. A standardtwo-tailed Student’s t test and paired Student’s t test were used forcomparison of numerical data, and p values less than 0.05 wereconsidered significant in this study. The median of the MIFexpression level in tumor cells or in lymphocytes was used as acutoff subgroup for MIF immunohistochemical variables in ourdata.ThePearson�2 testwascarriedout toassess the relationshipsamong IHC variables. The survival rate was estimated using theKaplan-Meier method and tested by log-rank analysis. A Coxregressionmodel was applied for multivariate analyses.
RESULTS
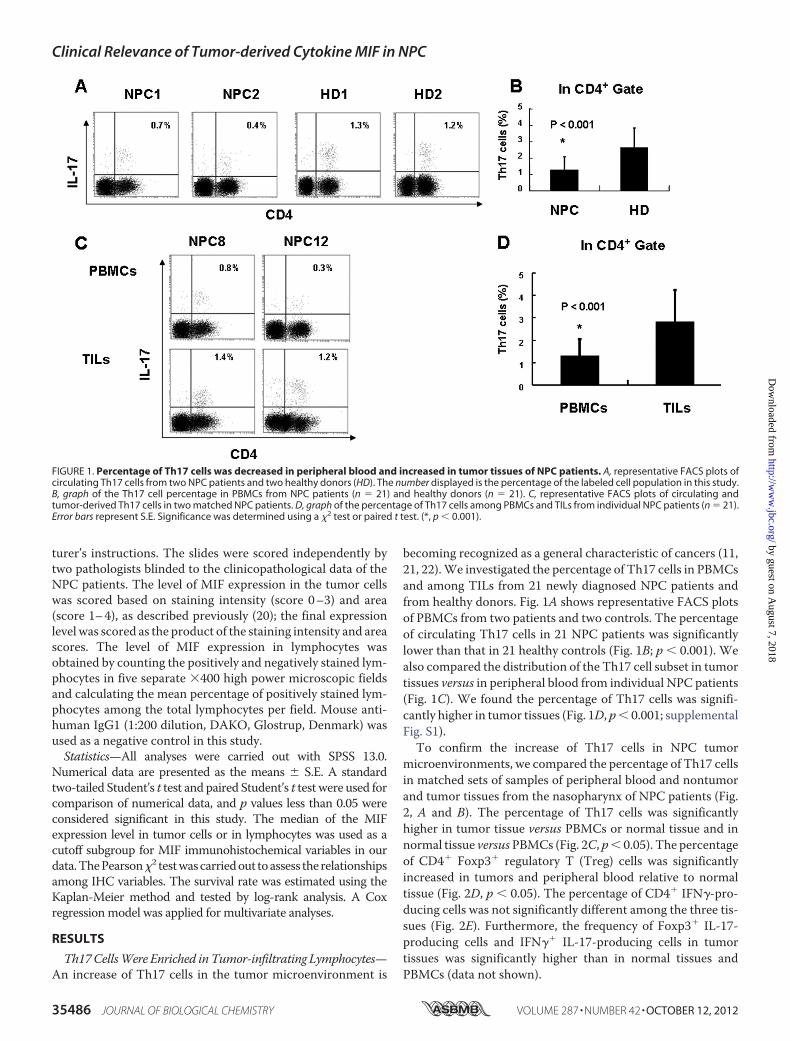
Th17CellsWere Enriched inTumor-infiltrating Lymphocytes—An increase of Th17 cells in the tumor microenvironment is
becoming recognized as a general characteristic of cancers (11,21, 22).We investigated the percentage of Th17 cells in PBMCsand among TILs from 21 newly diagnosed NPC patients andfrom healthy donors. Fig. 1A shows representative FACS plotsof PBMCs from two patients and two controls. The percentageof circulating Th17 cells in 21 NPC patients was significantlylower than that in 21 healthy controls (Fig. 1B; p � 0.001). Wealso compared the distribution of the Th17 cell subset in tumortissues versus in peripheral blood from individual NPC patients(Fig. 1C). We found the percentage of Th17 cells was signifi-cantly higher in tumor tissues (Fig. 1D, p� 0.001; supplementalFig. S1).To confirm the increase of Th17 cells in NPC tumor
microenvironments, we compared the percentage of Th17 cellsin matched sets of samples of peripheral blood and nontumorand tumor tissues from the nasopharynx of NPC patients (Fig.2, A and B). The percentage of Th17 cells was significantlyhigher in tumor tissue versus PBMCs or normal tissue and innormal tissue versusPBMCs (Fig. 2C, p� 0.05). The percentageof CD4� Foxp3� regulatory T (Treg) cells was significantlyincreased in tumors and peripheral blood relative to normaltissue (Fig. 2D, p � 0.05). The percentage of CD4� IFN�-pro-ducing cells was not significantly different among the three tis-sues (Fig. 2E). Furthermore, the frequency of Foxp3� IL-17-producing cells and IFN�� IL-17-producing cells in tumortissues was significantly higher than in normal tissues andPBMCs (data not shown).
FIGURE 1. Percentage of Th17 cells was decreased in peripheral blood and increased in tumor tissues of NPC patients. A, representative FACS plots ofcirculating Th17 cells from two NPC patients and two healthy donors (HD). The number displayed is the percentage of the labeled cell population in this study.B, graph of the Th17 cell percentage in PBMCs from NPC patients (n � 21) and healthy donors (n � 21). C, representative FACS plots of circulating andtumor-derived Th17 cells in two matched NPC patients. D, graph of the percentage of Th17 cells among PBMCs and TILs from individual NPC patients (n � 21).Error bars represent S.E. Significance was determined using a �2 test or paired t test. (*, p � 0.001).
Clinical Relevance of Tumor-derived Cytokine MIF in NPC
35486 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 287 • NUMBER 42 • OCTOBER 12, 2012
by guest on August 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Phenotypic Features and Cytokine Profiles of Circulating andTumor-infiltrating Th17Cells inNPCPatients—To explore thebiological properties of Th17 cells, we analyzed their pheno-typic markers on cells from circulating blood and from thetumormicroenvironment. As shown in Fig. 3A, among PBMCs,the Th17 cells were only found among CD45RO� andCD45RA� memory cells, and the Th17 cells expressed a highlevel of CCR6 and some CCR7 but lacked expression of CTLA4and GITR. Among TILs, the Th17 cells were also found onlyamong CD45RO� and CD45RA� memory cells and expresseda high level of CCR6, a low level of CCR7, some CTLA4, and alow level of GITR.We also evaluated the profiles of cytokines, including IL-2,
IFN�, IL-4, IL-10, TGF�, and GrB, released by the Th17 cellsubset among PBMCs and TILs fromNPC patients and amongPBMCs from healthy controls. All Th17 cells expressed highlevels of IL-2 and low levels of IL-4, IL-10, TGF�, and granzyme
B (Fig. 3B). Interestingly, the Th17 cells among TILs secreted alarge amount of IFN�, significantly more than from the Th17cells among the PBMCs ofNPCpatients or healthy donors (p�0.001).Generation and Migration of Th17 Cells Were Promoted by
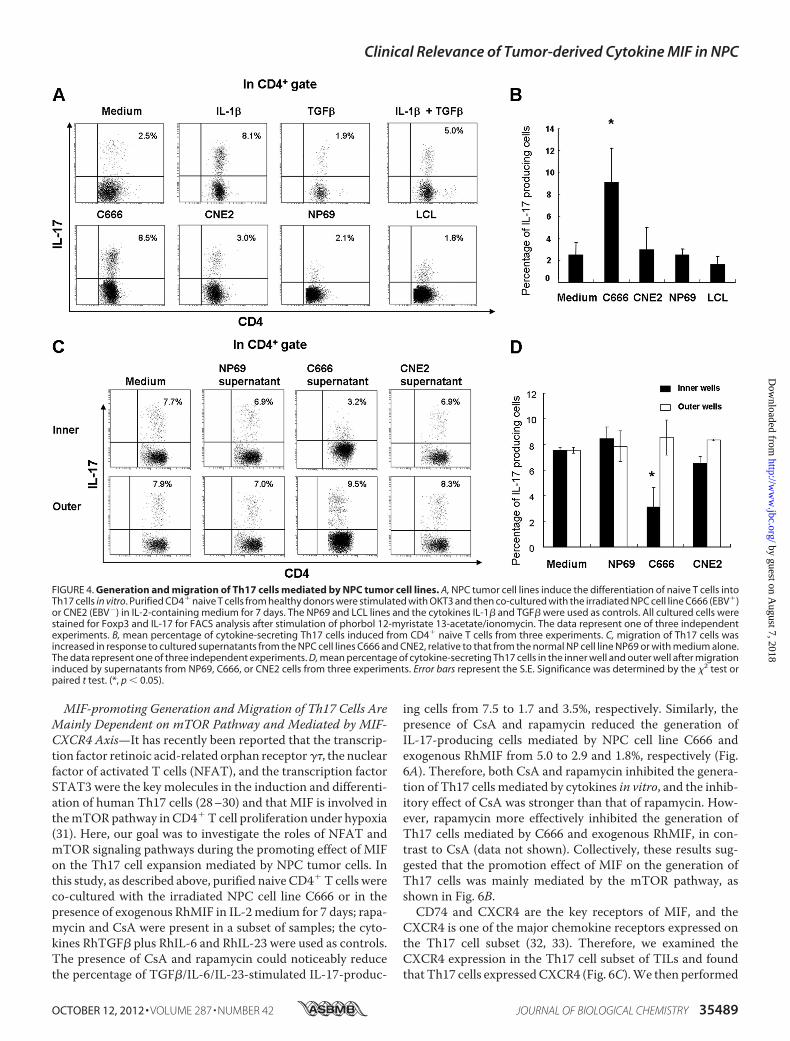
NPC Tumor Cell Lines in Vitro—The mechanism for the accu-mulation of Th17 cells in the NPC tumor microenvironmenthas been elusive, although tumor cells and immune cells oftencontribute to the induction of immune tolerance and inflam-mation at tumor sites (14, 23–25). To address whether NPCtumor cells could induce the generation or migration of Th17cells in vitro, we first investigated the induction of Th17 celldifferentiation from CD4� T cells. Purified naive CD4� T cellswere co-cultured with the irradiated NPC cell lines CNE2(EBV�) or C666 (EBV�), the normal NP cell line NP69, theEBV-transformed lymphoid blast cell line LCL, or in the pres-ence of cytokines. Cells were cultured for 7 days in medium
FIGURE 2. Distribution of different lymphocyte subsets in PBMC and in normal nasopharynx (NILs) and nasopharynx tumor tissues (TILs) fromindividual NPC patients. Representative FACS plots of Th17 cells and Foxp3� Treg (A) and IFN�-producing cells in total CD4� cells from two NPC patients outof five studied (B) are shown. Graphs for total CD4� cells of the percentage of Th17 cells (C), CD4� Foxp3� cells (D), and IFN�-producing cells in samples fromNPC patients (n � 5) (E) are shown. Significance was determined using a paired t test (p � 0.05).
Clinical Relevance of Tumor-derived Cytokine MIF in NPC
OCTOBER 12, 2012 • VOLUME 287 • NUMBER 42 JOURNAL OF BIOLOGICAL CHEMISTRY 35487
by guest on August 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from

containing IL-2. RhTGF�, RhIL-1�, or both were included ascontrols. The resulting percentage of IL-17-producing T cellswas evaluated by FACS.We found that naive CD4� T cells co-cultured with C666
cells, with cytokine IL-1�, or with a combination of IL-1�and TGF� exhibited a higher percentage of IL-17-producingcell differentiation in vitro relative to the other co-cultures(Fig. 4, A and B); these experiments were repeated threetimes.Next, we examined whether the supernatants of NPC
tumor cells could attract the migration of Th17 cells fromTILs or PBMCs (Fig. 4C). A higher percentage of Th17 cellswas present in the outer well of a Transwell assay of TILscultured with supernatants from the NPC C666 (p � 0.05) orCNE2 cell lines, indicating that the cytokines released fromNPC cell lines attracted Th17 cells more strongly than thosefrom normal NP cells or media alone (Fig. 4D). The super-natants from C666 and CNE2 cells also attracted the migra-tion of Th17 cells from a PBMC population (data not shown).Collectively, these data suggest that NPC tumor-derivedcytokines and chemokines may induce Th17 cell generationand chemotaxis.High Levels of MIF and Other Cytokines Released from NPC
Tumor Tissues and Cell Lines—To explore whether tumor-de-rived cytokines may affect NPC tumor microenvironments, weevaluated cytokine profiles of nontumor and tumor tissuesfrom an NPC patient and of the NPC cell lines C666 (EBV�)
and CNE2 (EBV�), using a Proteome Profiler Array asdescribed under “Experimental Procedures.” The 36 analyzedcytokines are listed in Table 1. CD154, G-CSF, CXCL1, CD54,IL-6, IL-8, MIF, serpin E1, and SDF-1 were detectable in one ormore of the culture supernatants, as shown in Table 1. G-CSF,CXCL1, IL-6, IL-8, MIF, and serpin E1 were detectable in thesupernatant of normal NP tissues. Overall, MIF was the onlycytokine found at a high level in all the supernatants (supple-mental Fig. S2).MIF Promotes the Generation and Migration of Th17 Cells
Mediated by Tumor Cells—Based on the high expression levelsof MIF in the NPC tumor microenvironment and on reportsthatMIFmay be involved in the generation of IL-17-producingcells in mice (26, 27), we investigated whether MIF is involvedin the generation and recruitment of Th17 cells in tumors. In aculture of CD4� naive T cells with C666 NPC cells, the pres-ence of the MIF small molecule inhibitor ISO-1 reduced thegeneration of Th17 cells (p � 0.05); in addition, the generationof Th17 cells also noticeably decreased when CD4� naive Tcells were co-cultured with C666 transfected with siRNAsagainst MIF (Fig. 5, A and B; supplemental Fig. S3). In a Tran-swell chemotaxis assay, ISO-1 and the supernatants fromMIF-siRNA-transfected C666 cells noticeably decreased themigration ofTh17 cells to the outerwell (p� 0.05) (Fig. 5,C andD; supplemental Fig. S3). The ability ofMIF directly to promotethe migration of Th17 cells in an NPC TIL population was alsodemonstrated (supplemental Fig. S4).
FIGURE 3. Characterization and cytokine expression profiles of the Th17 cell subset among PBMCs and TILs from NPC patients. A, T cell surface markerswere detected in Th17 cells from NPC patients. T cells were stimulated with phorbol 12-myristate 13-acetate/ionomycin for 4 h, then stained with fluorescence-conjugated antibodies against the markers shown, and analyzed by flow cytometry. B, graph of the percentage of cytokine-secreting Th17 cells among PBMCsand TILs from NPC patients (n � 5) and PBMCs from healthy donors (n � 5). Error bars represent the S.E. Significance was determined by the �2 test or pairedt test. (*, p � 0.001).
Clinical Relevance of Tumor-derived Cytokine MIF in NPC
35488 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 287 • NUMBER 42 • OCTOBER 12, 2012
by guest on August 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from
MIF-promoting Generation and Migration of Th17 Cells AreMainly Dependent on mTOR Pathway and Mediated by MIF-CXCR4 Axis—It has recently been reported that the transcrip-tion factor retinoic acid-related orphan receptor��, the nuclearfactor of activated T cells (NFAT), and the transcription factorSTAT3 were the key molecules in the induction and differenti-ation of human Th17 cells (28–30) and that MIF is involved inthemTORpathway in CD4�T cell proliferation under hypoxia(31). Here, our goal was to investigate the roles of NFAT andmTOR signaling pathways during the promoting effect of MIFon the Th17 cell expansion mediated by NPC tumor cells. Inthis study, as described above, purified naive CD4�T cells wereco-cultured with the irradiated NPC cell line C666 or in thepresence of exogenous RhMIF in IL-2medium for 7 days; rapa-mycin and CsA were present in a subset of samples; the cyto-kines RhTGF� plus RhIL-6 and RhIL-23 were used as controls.The presence of CsA and rapamycin could noticeably reducethe percentage of TGF�/IL-6/IL-23-stimulated IL-17-produc-
ing cells from 7.5 to 1.7 and 3.5%, respectively. Similarly, thepresence of CsA and rapamycin reduced the generation ofIL-17-producing cells mediated by NPC cell line C666 andexogenous RhMIF from 5.0 to 2.9 and 1.8%, respectively (Fig.6A). Therefore, both CsA and rapamycin inhibited the genera-tion of Th17 cells mediated by cytokines in vitro, and the inhib-itory effect of CsA was stronger than that of rapamycin. How-ever, rapamycin more effectively inhibited the generation ofTh17 cells mediated by C666 and exogenous RhMIF, in con-trast to CsA (data not shown). Collectively, these results sug-gested that the promotion effect of MIF on the generation ofTh17 cells was mainly mediated by the mTOR pathway, asshown in Fig. 6B.CD74 and CXCR4 are the key receptors of MIF, and the
CXCR4 is one of the major chemokine receptors expressed onthe Th17 cell subset (32, 33). Therefore, we examined theCXCR4 expression in the Th17 cell subset of TILs and foundthat Th17 cells expressedCXCR4 (Fig. 6C).We then performed
FIGURE 4. Generation and migration of Th17 cells mediated by NPC tumor cell lines. A, NPC tumor cell lines induce the differentiation of naive T cells intoTh17 cells in vitro. Purified CD4� naive T cells from healthy donors were stimulated with OKT3 and then co-cultured with the irradiated NPC cell line C666 (EBV�)or CNE2 (EBV�) in IL-2-containing medium for 7 days. The NP69 and LCL lines and the cytokines IL-1� and TGF� were used as controls. All cultured cells werestained for Foxp3 and IL-17 for FACS analysis after stimulation of phorbol 12-myristate 13-acetate/ionomycin. The data represent one of three independentexperiments. B, mean percentage of cytokine-secreting Th17 cells induced from CD4� naive T cells from three experiments. C, migration of Th17 cells wasincreased in response to cultured supernatants from the NPC cell lines C666 and CNE2, relative to that from the normal NP cell line NP69 or with medium alone.The data represent one of three independent experiments. D, mean percentage of cytokine-secreting Th17 cells in the inner well and outer well after migrationinduced by supernatants from NP69, C666, or CNE2 cells from three experiments. Error bars represent the S.E. Significance was determined by the �2 test orpaired t test. (*, p � 0.05).
Clinical Relevance of Tumor-derived Cytokine MIF in NPC
OCTOBER 12, 2012 • VOLUME 287 • NUMBER 42 JOURNAL OF BIOLOGICAL CHEMISTRY 35489
by guest on August 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from

an experiment in a Transwell system to determine whether theneutral antibody blocking CXCR4 could abort the effect ofMIFon themigration of Th17 cells in vitro. As shown in Fig. 6,D andE, the supernatants of C666 cell lines enhanced themigration ofTh17 cells relative to control medium, which was consistentwith Fig. 4C. However, the migration of Th17 cells was signifi-cantly inhibited when the different concentrations of the neu-tral antibody against CXCR4 were added in the supernatants ofthe C666 cell line.Expression Level ofMIF in Tumor-infiltrating Lymphocytes Is
an Independent Prognostic Factor for NPC Outcome—Weexamined the expression of MIF by immunohistochemistry inparaffin-embedded tumor specimens from 108 NPC patients.MIF could be localized to the cytoplasm of the tumor cells andto the cytoplasm of lymphocytes around tumor tissues (Fig.7A). Thirty one of 108 tumor specimens showed no MIFexpression in the tumor cells; the others displayed varied levelsof MIF expression. The MIF-positive lymphocytes per highlight microscope field in the 108 NPC tumor specimens rangedfrom 0 to 99% of total lymphocytes. Statistical analysis of the108 specimens demonstrated that the percentage of MIF-posi-tive lymphocytes was positively correlated with the level ofMIFexpression in the tumor cells (Fig. 7B).We divided the NPC patients into subgroups based on the
median expression level of MIF in tumor cells or lymphocytes.A high level of MIF expression in lymphocytes predicted a pos-itive clinical outcome for NPC patients (Fig. 7, C and D). Mul-tivariate analysis indicated that MIF was an independent prog-nostic factor for disease-free survival and overall survival of
NPC patients (supplemental Table 2), but we found no associ-ation between MIF expression in tumor cells or lymphocytesand clinicopathological characteristics (supplemental Table 3)and no association betweenMIF expression in tumors andNPCpatient survival (data not shown).
DISCUSSION
The idea that the immune system can control cancer hasbeen suggested for over a century. In recent years, the “cancerimmunoediting” hypothesis has been proposed, based on the2001 discovery that the immune system can control the qualityand quantity of cancer (6). The intratumor immune responsesin humans, such as the distribution of special TIL subsets andtumor-derived cytokines, could be used to predict patient prog-nosis (6, 34). In recent years, the distribution and clinical rele-vance of Th17 cells in cancer have been studied (10, 11, 22, 35).The accumulation ofTh17 cells in tumor tissues has been foundto be a feature ofmany cancers, but the percentage of Th17 cellsamong the circulating lymphocytes changes based on diseaseprogression and cancer type (10, 11, 36–38).In this study, we determined the distribution and functional
features of Th17 cells in peripheral blood and tumor tissuesfrom nasopharyngeal carcinoma patients, and we investigatedthe ability of tumor cells to promote the generation and migra-tion of this subset. We also analyzed the cytokines and chemo-kines released from the NPC tumor microenvironment anddetermined the clinical relevance of MIF expression in tumor-infiltrating lymphocytes.
TABLE 1Cytokines and chemokines released from NPC tumor cell lines, NPC tissue, or normal NP tissue in vitro
Cytokine Alternative nomenclature C666 (EBV�) CNE2 (EBV�) NPC tissue Normal NP tissue
C5a Complement component 5a � � � �CD154 CD40 ligand � � � �G-CSF CSF�, CSF-3 � � � �GM-CSF CSF�, CSF-2 � � � �GRO� CXCL1 � � � �I-309 CCL1 � � � �CD54 sICAM-1 � � � �IFN-� Type II IFN � � � �IL-1� IL-F1 � � � �IL-1� IL-1F2 � � � �IL-1ra IL-F3 � � � �IL-2 � � � �IL-4 � � � �IL-5 � � � �IL-6 � � � �IL-8 CXCL8 � � � �IL-10 � � � �IL-12p70 � � � �IL-13 � � � �IL-16 LCF � � � �IL-17 � � � �IL-17E � � � �IL-23 � � � �IL-27 � � � �IL-32� � � � �IP-10 CXCL10 � � � �I-TAC CXCL11 � � � �MCP-1 CCL2 � � � �MIF GIF, DER6 � � �� ��MIP-1� CCL3 � � � �MIP-1� CCL4 � � � �Serpin E1 PAI-1 � � � �RANTES CCL5 � � � �SDF-1 CXCL12 � � � �TNF� TNFSF 1A � � � �sTREM-1 � � � -
Clinical Relevance of Tumor-derived Cytokine MIF in NPC
35490 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 287 • NUMBER 42 • OCTOBER 12, 2012
by guest on August 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from

In 21 patients with NPC (19/21 in late-disease stage, supple-mentalTableS1),we foundadecreasedpercentageofTh17cells inthe peripheral blood of NPC patients relative to that of healthyindividuals.Moreover, a larger percentage ofTh17 cellswas foundamong TILs relative to the peripheral blood or normal nasophar-ynx tissues of NPC patients. It has been reported that the propor-tion of circulating Treg and Th17 cells is increased in early stagepatients, although the percentage of circulating Th17 cellsdecreased and that of circulating Treg cells increased in advancedpatients with gastric cancer (22). Our results also showed adecreased percentage of Th17 cells in PBMCs fromNPC patients(most at an advanced disease stage) relative to healthy controls.The clinical relevance of Th17 cells has been reported for
many kinds of cancers, but the results have sometimes beenopposed. For example, the number of Th17 cells positively pre-dicted the clinical outcome for ovarian cancer patients but neg-atively predicted the clinical outcome of patients with hepato-cellular carcinoma (10, 11). In a previous study, we could notdetermine the relationship between the density of Th17 cellsand the clinical outcome in NPC patients (39).The biological function of the Th17 cell subset in cancer
remains largely unknown. In this study, we analyzed the generalcharacteristics and functional features of theTh17 cell subset inPBMCs andTILs.Our results showed that Th17 cells expressed
high levels of the chemokine receptor CCR6 and some CCR7.Most of the Th17 cells came from the CD45RO� memory Tcell population. Th17 cells from NPC patients expressedsome CTLA4 but no GITR. These results are consistent withobservations in other cancers and provide evidence thatTh17 cells can home to tumor tissues (8, 14, 35, 40). Cytokineprofiles indicated that Th17 cells secrete high levels of IL-2and low levels of IL-4, TGF�, and GrB. Interestingly, theTh17 cells in TILs secreted significantly higher levels ofIFN� than did PBMCs from NPC patients and healthy con-trols (Fig. 3B). This result indicates that Th17 cells may havean anti-tumor function in NPC tumor tissues by secretingTh1 cytokines such as IFN� and IL-2, similar to Th17 cells inovarian cancer (11).The increased percentage of Th17 cells in tumor tissues is
associatedwith cytokines and chemokines released from tumormicroenvironments (14, 23, 41–44). This study shows for thefirst time that the EBV� C666 cells co-cultured with CD4�
naive T cells promoted the generation of Th17 cells, in compar-ison with the CNE2 (EBV�), LCL, or NP69 cells. Furthermore,the supernatants fromC666 and CNE2 cultures could promotethe migration of Th17 cells in vitro.Six cytokines were found in the culture medium of NPC
tumor tissues, with MIF being found in the greatest amount.
FIGURE 5. Role of MIF in the generation and migration of Th17 cells. A, generation of Th17 cells was decreased by the presence of either ISO-1 or siRNAagainst MIF. B, graph of IL-17-positive cells as a percentage of Th17 cells; data are from three independent experiments. Error bars represent S.E.; *, p �0.05. C, ISO-1 noticeably inhibited the migration of Th17 cells exposed to the supernatant of the C666 cell line, and the migration of Th17 cells exposedto supernatant from MIF-siRNA-treated C666 cells was also decreased; data are representative of three independent experiments. D, percentage of Th17cells migrating to the outer well; data are from three independent experiments. Error bars represent S.E.. *, p � 0.05. Significance was determined byperforming paired t test.
Clinical Relevance of Tumor-derived Cytokine MIF in NPC
OCTOBER 12, 2012 • VOLUME 287 • NUMBER 42 JOURNAL OF BIOLOGICAL CHEMISTRY 35491
by guest on August 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from

MIF, which is often secreted by naive CD4� T cells, can stimu-late IL-17-producing cells in the lymph nodes of mice (27, 45).However, the role of MIF in expanding the Th17 cell subsetin human tumors is not known. Our data showed that MIFwas involved in the generation andmigration of Th17 cells inNPC and that this activity was blocked by the MIF inhibitorISO-1 or by siRNAs targetingMIF-transfected NPC cell linesin vitro. Furthermore, our results for the first time indicatedthat the effect of MIF on the generation and recruitment ofTh17 cells was mainly dependent on the mTOR signalingpathway and mediated by the MIF-CXCR4 axis but inde-pendent of NFAT pathway. These results were consistentwith recent reports that the effect of MIF on stimulating theexpression of IL-17 in lymph node cells of mice was depen-dent on the MAPK-JAK2/STAT3 pathway but not the NF��or NFAT pathway (27, 46). Others also have found that MIFcan up-regulate IL-6, which was also involved in the gener-
ation of Th17 cells, IL-8, and TNF� in patients with Vibriovulnificus (47).Overexpression of MIF has been identified in tumor cells of
many types of cancers, including NPC (48–51). Here, wedetected the expression ofMIF in NPC tumor tissues by IHC inparaffin-embedded sections from 108 patients. MIF wasexpressed in tumor cells and in lymphocytes around NPCtumor tissues, and there was a correlation between the expres-sion levels of MIF in tumor cells and the MIF-positive lympho-cytes in tumor tissues. Moreover, we found that high expres-sion of MIF in TILs was associated with improved NPC patientoutcome, and MIF was an independent prognostic factor forNPC patients by multivariate analysis.High expression of MIF in NPC tumor tissues is associated
with increased microvessels and increased lymph node metas-tasis of NPC tumors, whereas the angiogenesis and lymph nodestatus exhibited in relation to patient survival has been identi-
FIGURE 6. Signaling pathways of MIF involved in the generation and migration of Th17 cells. A, generation of Th17 cells induced by cytokines or C666 cellsand MIF was decreased by the presence of rapamycin or CsA in the culture media at different levels; data are representative of three independent experiments.B, MIF promoting the generation of Th17 cells in vitro was dependent on mTOR signaling pathway. C, expression of CXCR4 on IL-17-positive and -negative cellpopulation. D, specific neutralizing antibody against CXCR4 significantly inhibited the migration of Th17 cells exposed to the supernatant of the C666 cell line;data are representative of three independent experiments. E, percentage of Th17 cells migrating to the outer well; data are from three independent experi-ments. Error bars represent S.E. *, p � 0.05. Significance was determined by performing paired t test.
Clinical Relevance of Tumor-derived Cytokine MIF in NPC
35492 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 287 • NUMBER 42 • OCTOBER 12, 2012
by guest on August 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from

fied as an independent prognostic factor ofNPCbymultivariateanalysis (52). MIF is overexpressed in human rhabdomyosar-coma cell lines, and it prevents rhabdomyosarcoma cells fromresponding to chemoattractants secreted outside of the grow-ing tumor (e.g. SDF-1), thereby preventing the release of cellsinto the circulation. Moreover, MIF inhibits the recruitment ofcancer-associated fibroblasts to growing tumors in an CXCR2/CD74-dependent manner; therefore, therapeutic inhibition ofMIF in rhabdomyosarcoma may accelerate metastasis andtumor growth (50). In drug-resistant human colon cancers, theMIF-CXCR4 axis mediates the invasive and metastatic pheno-type of the cancer (53, 54), but MIF does not play a prominentrole in tumor progression in gastric adenocarcinomas (55). Col-lectively, these findings indicate that MIF has multiple func-tions in tumor microenvironments. The data obtained in thisstudy are the first indicating a protective function ofMIF in thetumor microenvironment and involvement in promoting theNPC tumor cell-mediated induction and migration of Th17cells.In conclusion, our data provide novel evidence of the
accumulation of Th17 cells in the NPC tumor microenviron-ment and of the biological functions of the Th17 cells in NPCtumor tissues, such as secretion of Th1 cytokines. We haveshown for the first time that overexpression of MIF in thetumor microenvironment is involved in the induction andmigration of human Th17 cells in vitro, which mainlydepends on the mTOR pathway and is mediated by the MIF-CXCR4 axis. Finally, we found that there is a clinical rele-vance to tumor-derived MIF expression levels in NPC
patients. These data provide novel insights into inflamma-tory responses in NPC tumor progression.
Acknowledgments—We thank Rong-Fu Wang (Baylor College ofMedicine, Houston, TX) for valuable comments for this manuscriptand members of the Zeng laboratory for helpful discussions. We alsothank David Nadziejka of Grand Rapids, MI, for critical reading andtechnical editing of the manuscript.
REFERENCES1. Pathmanathan, R., Prasad, U., Chandrika, G., Sadler, R., Flynn, K., and
Raab-Traub, N. (1995) Undifferentiated, nonkeratinizing, and squamouscell carcinoma of the nasopharynx. Variants of Epstein-Barr virus-in-fected neoplasia. Am. J. Pathol. 146, 1355–1367
2. Cao, S. M., Simons, M. J., and Qian, C. N. (2011) The prevalence andprevention of nasopharyngeal carcinoma in China. Chin. J. Cancer 30,114–119
3. Wee, J. T., Ha, T. C., Loong, S. L., andQian, C. N. (2010) Is nasopharyngealcancer really a “Cantonese cancer”? Chin. J. Cancer 29, 517–526
4. Niedobitek, G., Agathanggelou, A., andNicholls, J.M. (1996) Epstein-Barrvirus infection and the pathogenesis of nasopharyngeal carcinoma. Viralgene expression, tumor cell phenotype, and the role of the lymphoidstroma. Semin. Cancer Biol. 7, 165–174
5. Vasef, M. A., Ferlito, A., and Weiss, L. M. (1997) Nasopharyngeal carci-noma, with emphasis on its relationship to Epstein-Barr virus. Ann. Otol.Rhinol. Laryngol. 106, 348–356
6. Schreiber, R. D., Old, L. J., and Smyth,M. J. (2011) Cancer immunoediting.Integrating immunity’s roles in cancer suppression and promotion. Sci-ence 331, 1565–1570
7. Yu, H., Kortylewski, M., and Pardoll, D. (2007) Cross-talk between cancerand immune cells. Role of STAT3 in the tumor microenvironment. Nat.Rev. Immunol. 7, 41–51
FIGURE 7. Increased expression of MIF in TILs predicts improved patient survival. A, immunohistochemical staining shows varied intensities of MIFexpression in the cytoplasm of tumor cells or tumor-associated lymphocytes (�40). B, percentage of MIF-positive lymphocytes around tumor cells wasincreased with the expression levels of MIF in tumor cells (R � 0.69, p � 0.0001). C and D, samples from NPC patients (n � 108) were divided into two groupsbased on positive or negative expression of MIF in NPC tumor cells. Disease-free survival and overall survival were significantly increased with increasedexpression of MIF in TILs, as displayed in Kaplan-Meier plots of overall survival.
Clinical Relevance of Tumor-derived Cytokine MIF in NPC
OCTOBER 12, 2012 • VOLUME 287 • NUMBER 42 JOURNAL OF BIOLOGICAL CHEMISTRY 35493
by guest on August 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from

8. Wang, L., Yi, T., Kortylewski, M., Pardoll, D. M., Zeng, D., and Yu, H.(2009) IL-17 can promote tumor growth through an IL-6-Stat3 signalingpathway. J. Exp. Med. 206, 1457–1464
9. Lv, L., Pan, K., Li, X. D., She, K. L., Zhao, J. J., Wang,W., Chen, J. G., Chen,Y. B., Yun, J. P., andXia, J. C. (2011) The accumulation and prognosis valueof tumor-infiltrating IL-17-producing cells in esophageal squamous cellcarcinoma. PLoS One 6, e18219
10. Zhang, J. P., Yan, J., Xu, J., Pang, X. H., Chen, M. S., Li, L., Wu, C., Li, S. P.,and Zheng, L. (2009) Increased intratumoral IL-17-producing cells corre-latewith poor survival in hepatocellular carcinomapatients. J. Hepatol.50,980–989
11. Kryczek, I., Banerjee, M., Cheng, P., Vatan, L., Szeliga,W.,Wei, S., Huang,E., Finlayson, E., Simeone, D., Welling, T. H., Chang, A., Coukos, G., Liu,R., andZou,W. (2009) Phenotype, distribution, generation, and functionaland clinical relevance of Th17 cells in the human tumor environments.Blood 114, 1141–1149
12. Dürr, C., Pfeifer, D., Claus, R., Schmitt-Graeff, A., Gerlach, U. V., Graeser,R., Krüger, S., Gerbitz, A., Negrin, R. S., Finke, J., and Zeiser, R. (2010)CXCL12 mediates immunosuppression in the lymphoma microenviron-ment after allogeneic transplantation of hematopoietic cells. Cancer Res.70, 10170–10181
13. Liu, J., Zhang, N., Li, Q., Zhang, W., Ke, F., Leng, Q., Wang, H., and Chen,J. (2011) Tumor-associated macrophages recruit CCR6� regulatory Tcells and promote the development of colorectal cancer via enhancingCCL20 production in mice. PLoS One 6, e19495
14. Su, X., Ye, J., Hsueh, E. C., Zhang, Y., Hoft, D. F., and Peng, G. (2010)Tumor microenvironments direct the recruitment and expansion of hu-man Th17 cells. J. Immunol. 184, 1630–1641
15. Zhang, Y. L., Li, J., Mo, H. Y., Qiu, F., Zheng, L. M., Qian, C. N., and Zeng,Y. X. (2010) Different subsets of tumor-infiltrating lymphocytes correlatewith NPC progression in different ways.Mol. Cancer 9, 4
16. Dudley, M. E., Wunderlich, J. R., Shelton, T. E., Even, J., and Rosenberg,S. A. (2003) Generation of tumor-infiltrating lymphocyte cultures for usein adoptive transfer therapy for melanoma patients. J. Immunother. 26,332–342
17. Deleted in proof18. Kopf, H., de la Rosa, G. M., Howard, O. M., and Chen, X. (2007) Rapamy-
cin inhibits differentiation of Th17 cells and promotes generation ofFoxP3� T regulatory cells. Int. Immunopharmacol. 7, 1819–1824
19. Baribaud, F., Edwards, T. G., Sharron,M., Brelot, A., Heveker, N., Price, K.,Mortari, F., Alizon, M., Tsang, M., and Doms, R. W. (2001) Antigenicallydistinct conformations of CXCR4. J. Virol. 75, 8957–8967
20. Wee, A., Teh, M., and Raju, G. C. (1994) Clinical importance of p53 pro-tein in gallbladder carcinoma and its precursor lesions. J. Clin. Pathol. 47,453–456
21. The Selected Cancers Cooperative Study Group (1990) The association ofselected cancers with service in the United States military in Vietnam. III.Hodgkin disease, nasal cancer, nasopharyngeal cancer, and primary livercancer. Arch. Intern. Med. 150, 2495–2505
22. Maruyama, T., Kono, K., Mizukami, Y., Kawaguchi, Y., Mimura, K., Wa-tanabe, M., Izawa, S., and Fujii, H. (2010) Distribution of Th17 cells andFoxP3� regulatory T cells in tumor-infiltrating lymphocytes, tumor-draining lymph nodes and peripheral blood lymphocytes in patients withgastric cancer. Cancer Sci. 101, 1947–1954
23. Miyahara, Y., Odunsi, K., Chen, W., Peng, G., Matsuzaki, J., and Wang,R. F. (2008) Generation and regulation of human CD4� IL-17-producingT cells in ovarian cancer. Proc. Natl. Acad. Sci. U.S.A. 105, 15505–15510
24. Moo-Young, T. A., Larson, J. W., Belt, B. A., Tan, M. C., Hawkins, W. G.,Eberlein, T. J., Goedegebuure, P. S., and Linehan, D. C. (2009) Tumor-derived TGF-�mediates conversion of CD4�Foxp3� regulatory T cells ina murine model of pancreas cancer. J. Immunother. 32, 12–21
25. Shen, X., Li, N., Li, H., Zhang, T., Wang, F., and Li, Q. (2010) Increasedprevalence of regulatory T cells in the tumor microenvironment and itscorrelation with TNM stage of hepatocellular carcinoma. J. Cancer Res.Clin. Oncol. 136, 1745–1754
26. Addis, M. F., Tanca, A., Pagnozzi, D., Crobu, S., Fanciulli, G., Cossu-Rocca, P., and Uzzau, S. (2009) Generation of high quality protein extractsfrom formalin-fixed, paraffin-embedded tissues. Proteomics 9, 3815–3823
27. Stojanovic, I., Cvjeticanin, T., Lazaroski, S., Stosic-Grujicic, S., andMiljkovic, D. (2009) Macrophage migration inhibitory factor stimulatesinterleukin-17 expression and production in lymph node cells. Immunol-ogy 126, 74–83
28. Harris, T. J., Grosso, J. F., Yen, H. R., Xin, H., Kortylewski, M., Albesiano,E., Hipkiss, E. L., Getnet, D., Goldberg, M. V., Maris, C. H., Housseau, F.,Yu, H., Pardoll, D. M., and Drake, C. G. (2007) Cutting edge. An in vivorequirement for STAT3 signaling in TH17 development and TH17-de-pendent autoimmunity. J. Immunol. 179, 4313–4317
29. Delgoffe, G. M., Kole, T. P., Zheng, Y., Zarek, P. E., Matthews, K. L., Xiao,B., Worley, P. F., Kozma, S. C., and Powell, J. D. (2009) The mTOR kinasedifferentially regulates effector and regulatory T cell lineage commitment.Immunity 30, 832–844
30. Radojcic, V., Pletneva, M. A., Yen, H. R., Ivcevic, S., Panoskaltsis-Mortari,A., Gilliam, A. C., Drake, C. G., Blazar, B. R., and Luznik, L. (2010) STAT3signaling in CD4� T cells is critical for the pathogenesis of chronic sclero-dermatous graft-versus-host disease in a murine model. J. Immunol. 184,764–774
31. Gaber, T., Schellmann, S., Erekul, K. B., Fangradt, M., Tykwinska, K.,Hahne, M., Maschmeyer, P., Wagegg, M., Stahn, C., Kolar, P., Dziurla, R.,Löhning, M., Burmester, G. R., and Buttgereit, F. (2011) Macrophage mi-gration inhibitory factor counter-regulates dexamethasone-mediatedsuppression of hypoxia-inducible factor-1� function and differentially in-fluences human CD4� T cell proliferation under hypoxia. J. Immunol.186, 764–774
32. Schober, A., Bernhagen, J., and Weber, C. (2008) Chemokine-like func-tions of MIF in atherosclerosis. J. Mol. Med. 86, 761–770
33. Lim, H.W., Lee, J., Hillsamer, P., and Kim, C. H. (2008) Human Th17 cellsshare major trafficking receptors with both polarized effector T cells andFOXP3� regulatory T cells. J. Immunol. 180, 122–129
34. Swann, J. B., Vesely, M. D., Silva, A., Sharkey, J., Akira, S., Schreiber, R. D.,and Smyth, M. J. (2008) Demonstration of inflammation-induced cancerand cancer immunoediting during primary tumorigenesis. Proc. Natl.Acad. Sci. U.S.A. 105, 652–656
35. Wang, C., Kang, S. G., Lee, J., Sun, Z., and Kim, C. H. (2009) The roles ofCCR6 in migration of Th17 cells and regulation of effector T-cell balancein the gut.Mucosal Immunol. 2, 173–183
36. Liu, J., Duan, Y., Cheng, X., Chen, X., Xie, W., Long, H., Lin, Z., and Zhu,B. (2011) IL-17 is associated with poor prognosis and promotes angiogen-esis via stimulating VEGF production of cancer cells in colorectal carci-noma. Biochem. Biophys. Res. Commun. 407, 348–354
37. Ratajczak, P., Janin, A., Peffault de Latour, R., Leboeuf, C., Desveaux, A.,Keyvanfar, K., Robin, M., Clave, E., Douay, C., Quinquenel, A., Pichereau,C., Bertheau, P., Mary, J. Y., and Socié, G. (2010) Th17/Treg ratio in hu-man graft-versus-host disease. Blood 116, 1165–1171
38. Tosolini, M., Kirilovsky, A., Mlecnik, B., Fredriksen, T., Mauger, S., Bin-dea, G., Berger, A., Bruneval, P., Fridman, W. H., Pagès, F., and Galon, J.(2011) Clinical impact of different classes of infiltrating T cytotoxic andhelper cells (Th1, th2, treg, and th17) in patients with colorectal cancer.Cancer Res. 71, 1263–1271
39. Zhang, Y. L., Li, J., Mo, H. Y., Qiu, F., Zheng, L. M., Qian, C. N., and Zeng,Y. X. (2010) Different subsets of tumor infiltrating lymphocytes correlatewith NPC progression in different ways.Mol. Cancer 9, 4
40. Zou, W., and Restifo, N. P. (2010) T(H)17 cells in tumor immunity andimmunotherapy. Nat. Rev. Immunol. 10, 248–256
41. Evans, H. G., Gullick, N. J., Kelly, S., Pitzalis, C., Lord, G. M., Kirkham,B.W., and Taams, L. S. (2009) In vivo activatedmonocytes from the site ofinflammation in humans specifically promote Th17 responses. Proc. Natl.Acad. Sci. U.S.A. 106, 6232–6237
42. Kimura, A., Naka, T., and Kishimoto, T. (2007) IL-6-dependent and -in-dependent pathways in the development of interleukin 17-producing Thelper cells. Proc. Natl. Acad. Sci. U.S.A. 104, 12099–12104
43. Paintlia, M. K., Paintlia, A. S., Singh, A. K., and Singh, I. (2011) Synergisticactivity of interleukin-17 and tumor necrosis factor-� enhances oxidativestress-mediated oligodendrocyte apoptosis. J. Neurochem. 116, 508–521
44. Ye, Z. J., Zhou,Q., Gu, Y. Y., Qin, S.M.,Ma,W. L., Xin, J. B., Tao, X.N., andShi, H. Z. (2010)Generation and differentiation of IL-17-producingCD4�
T cells in malignant pleural effusion. J. Immunol. 185, 6348–6354
Clinical Relevance of Tumor-derived Cytokine MIF in NPC
35494 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 287 • NUMBER 42 • OCTOBER 12, 2012
by guest on August 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from

45. Park, S. K., Cho, M. K., Park, H. K., Lee, K. H., Lee, S. J., Choi, S. H., Ock,M. S., Jeong, H. J., Lee, M. H., and Yu, H. S. (2009) Macrophage migrationinhibitory factor homologs of anisakis simplex suppress Th2 response inallergic airway inflammation model via CD4�CD25�Foxp3� T cell re-cruitment. J. Immunol. 182, 6907–6914
46. Bachand, A.M.,Mundt, K. A.,Mundt, D. J., andMontgomery, R. R. (2010)Epidemiological studies of formaldehyde exposure and risk of leukemiaand nasopharyngeal cancer. A meta-analysis. Crit. Rev. Toxicol. 40,85–100
47. Chuang, C. C., Chuang, Y. C., Chang,W. T., Chen, C. C., Hor, L. I., Huang,A. M., Choi, P. C., Wang, C. Y., Tseng, P. C., and Lin, C. F. (2010) Macro-phage migration inhibitory factor regulates interleukin-6 production byfacilitating nuclear factor-�B activation duringVibrio vulnificus infection.BMC Immunol. 11, 50
48. Chen, Y. C., Zhang, X. W., Niu, X. H., Xin, D. Q., Zhao, W. P., Na, Y. Q.,and Mao, Z. B. (2010) Macrophage migration inhibitory factor is a directtarget of HBP1-mediated transcriptional repression that is overexpressedin prostate cancer. Oncogene 29, 3067–3078
49. Krockenberger, M., Engel, J. B., Kolb, J., Dombrowsky, Y., Häusler, S. F.,Kohrenhagen, N., Dietl, J., Wischhusen, J., and Honig, A. (2010) Macro-phage migration inhibitory factor expression in cervical cancer. J. CancerRes. Clin. Oncol. 136, 651–657
50. Tarnowski, M., Grymula, K., Liu, R., Tarnowska, J., Drukala, J., Ratajczak,J., Mitchell, R. A., Ratajczak, M. Z., and Kucia, M. (2010) Macrophage
migration inhibitory factor is secreted by rhabdomyosarcoma cells, mod-ulates tumor metastasis by binding to CXCR4 and CXCR7 receptors, andinhibits recruitment of cancer-associated fibroblasts. Mol. Cancer Res. 8,1328–1343
51. Fang, W., Li, X., Jiang, Q., Liu, Z., Yang, H., Wang, S., Xie, S., Liu, Q., Liu,T., Huang, J., Xie,W., Li, Z., Zhao, Y.,Wang, E.,Marincola, F.M., and Yao,K. (2008) Transcriptional patterns, biomarkers, and pathways character-izing nasopharyngeal carcinoma of Southern China. J. Transl. Med. 6, 32
52. Liao, B., Zhong, B. L., Li, Z., Tian, X. Y., Li, Y., and Li, B. (2010) Macro-phagemigration inhibitory factor contributes angiogenesis by up-regulat-ing IL-8 and correlates with poor prognosis of patients with primary na-sopharyngeal carcinoma. J. Surg. Oncol. 102, 844–851
53. Nishihira, J., Ishibashi, T., Fukushima, T., Sun, B., Sato, Y., and Todo, S.(2003)Macrophagemigration inhibitory factor (MIF). Its potential role intumor growth and tumor-associated angiogenesis. Ann. N.Y. Acad. Sci.995, 171–182
54. Takahashi, N., Nishihira, J., Sato, Y., Kondo, M., Ogawa, H., Ohshima, T.,Une, Y., and Todo, S. (1998) Involvement ofmacrophagemigration inhib-itory factor (MIF) in the mechanism of tumor cell growth. Mol. Med. 4,707–714
55. Xia, H.H., Yang, Y., Chu, K.M., Gu,Q., Zhang, Y. Y., He,H.,Wong,W.M.,Leung, S. Y., Yuen, S. T., Yuen, M. F., Chan, A. O., andWong, B. C. (2009)Serum macrophage migration-inhibitory factor as a diagnostic and prog-nostic biomarker for gastric cancer. Cancer 115, 5441–5449
Clinical Relevance of Tumor-derived Cytokine MIF in NPC
OCTOBER 12, 2012 • VOLUME 287 • NUMBER 42 JOURNAL OF BIOLOGICAL CHEMISTRY 35495
by guest on August 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Liu, Qiu-Yan Chen, Zi-Ming Du, Li-Min Zheng, Chao-Nan Qian and Yi-Xin ZengJiang Li, Hao-Yuan Mo, Geng Xiong, Lin Zhang, Jia He, Zhou-Feng Huang, Zhi-Wei
Predicts Favorable Survival in Nasopharyngeal Carcinoma PatientsAccumulation of Interleukin-17-producing Tumor-infiltrating Lymphocytes and
Tumor Microenvironment Macrophage Inhibitory Factor Directs the
doi: 10.1074/jbc.M112.367532 originally published online August 14, 20122012, 287:35484-35495.J. Biol. Chem.
10.1074/jbc.M112.367532Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
Supplemental material:
http://www.jbc.org/content/suppl/2012/08/14/M112.367532.DC1
http://www.jbc.org/content/287/42/35484.full.html#ref-list-1
This article cites 54 references, 20 of which can be accessed free at
by guest on August 7, 2018
http://ww
w.jbc.org/
Dow
nloaded from