two new species of bats (chiroptera) for the fauna of ... · libya: rhinopoma hardwickii and...
TRANSCRIPT

MYOTIS Vol. 41-42 109 - 124 Bonn, November 2004
Two new species of bats (Chiroptera) for the fauna of Libya: Rhinopoma hardwickii and Pipistrellus rueppellii
Petr B e n d a , Vladimír H a n á k , Michal A n d r e a s , Antonín R e i t e r & Marcel U h r i n
A b s t r a c t . A colony of approx. 40 individuals of Rhinopoma hardwickii was found, and one individual of Pipistrellus rueppellii was netted, in the oasis of Al Jaghbub (= Giarabub; 29° 45’ N, 24° 31’ E), Cyrenaica, northeastern Libya, on 13 May 2002. Both species were recorded in this country for the first time and therefore the number of bat species in Libyan fauna has increased to 17. Some notes on the distribution and geographic variation of the two species in northern Africa are given. K e y w o r d s . Rhinopoma hardwickii, Pipistrellus rueppellii, distribution, variation, northern Africa.
Introduction According to current knowledge, the bat fauna of Libya is poor in comparison with the surrounding countries (Hutson et al. 2001). In the first compendium of Libyan mammals, Klaptocz (1909) mentioned three bat species only. In the Italian colonial era, four bat species were known from the whole country (Zavattari 1934, 1937). In the 1950s, Toschi (1954) and later Setzer (1957) mentioned seven bat species and the same number was also reported by Hufnagl (1972). After two extensive field expeditions, Hanák & Elgadi (1984) presented and/or summarised records of 13 bat species in Libya. Qumsiyeh (1983), Qumsiyeh & Schlitter (1982) and Spitzenberger (1982) added several records of other bat species to altogether 15 species of four families. Although the bat fauna of Libya still remains poorly known, two forms of bats were described from the present Libyan territory, viz., Vespertilio isabellinus Temminck, 1840 (terra typica: environs of Tripoli) and Pipistrellus deserti Thomas, 1902 (t. t.: Mursuk). During a field trip to Libya in 2002, we collected two bat species which were hith-erto not known to occur in the territory of Libya: the Lesser mouse-tailed bat, Rhinopoma hardwickii Gray, 1831 and the Rüppell’s bat, Pipistrellus rueppellii (Fischer, 1829). The number of bat species in the fauna of Libya has thus increased to 17, and the number of bat families to five.
Record Both bat species were collected in the oasis of Al Jaghbub (= Giarabub) (Tubruq Dist., Cyrenaica, northeastern Libya; 29° 45’ N, 24° 31’ E; approx. 30 m a. s. l.; Figs. 1, 2) on

110 Benda et al.
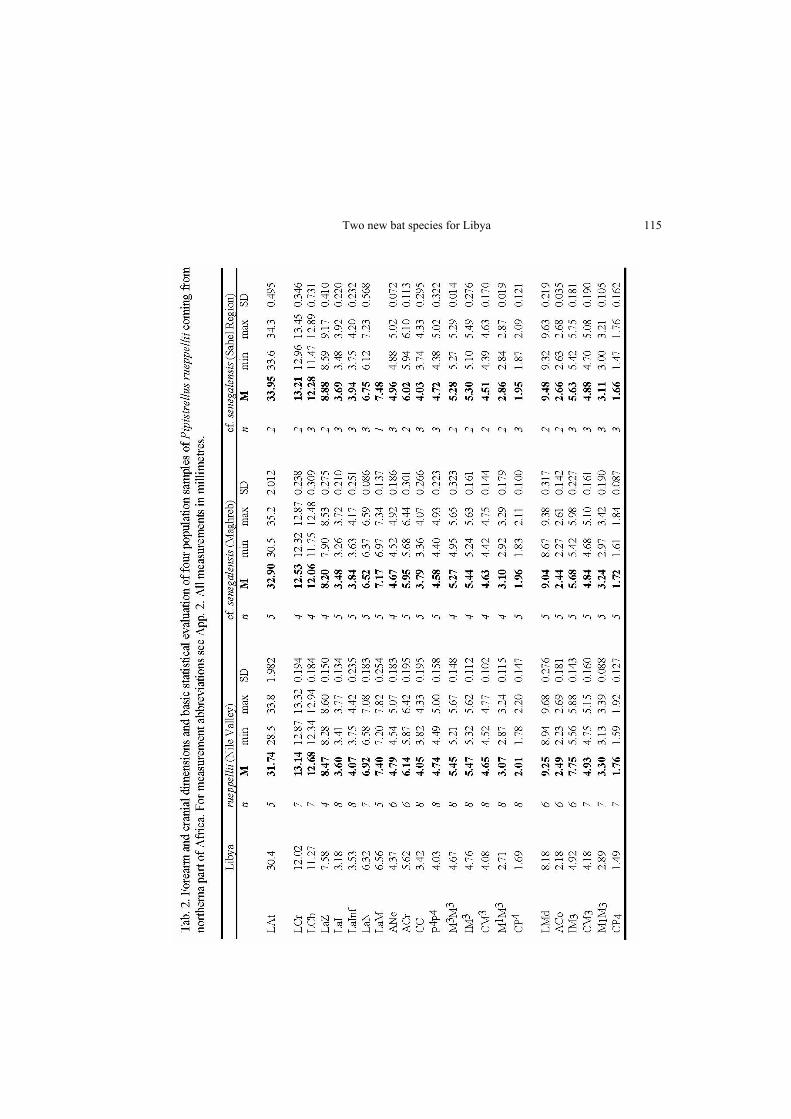
13 May 2002. The site is a relatively small oasis surrounded by sandy desert and ham-mada, situated approx. 250 km southwards from coast of the Mediterranean Sea, very close to the Libyan-Egyptian border. In the centre of the oasis, there is a small hill with the ruins of an Italian military fortress; this hill is surrounded by gardens and orchards irrigated by a grid of narrow channels. The oasis is found on the western margin of the Siwa Basin adjoining the Qattara Depression in northwestern Egypt; this region is a very important site in northern Africa from the biogeographical point of view, and has been studied since the colonial era (De Beaux 1928, Hayman 1949, Ranck 1968, etc.). A colony of approx. 40 individuals of Rhinopoma hardwickii was observed in dark rooms on the ground floor of the ruins of the Italian fortress; from this colony, seven adult males, three subadult males, and four adult females were collected. The individuals are deposited in the collection of the National Museum (Natural History), Prague (NMP 49864–49877, two complete alcohol specimens, 10 alcohol specimens with extracted skulls, and two skeletons). The observed and collected bats were very pale in comparison with the populations from the Nile Valley and the Middle East. In the Libyan mouse-tailed bats (Fig. 3), naked parts of skin range from creamy-white (muzzle, ears, margins of wing membranes) up to very pale greyish-brown (main parts of wing membranes). Distal parts of dorsal hairs are pale greyish-brown, proximal parts are whitish, ventral hairs are whitish. Basic measurements of the collected specimens are given in Tab. 1. An adult female of Pipistrellus rueppellii (Fig. 4) was mist-netted above a water channel between gardens in northern part of the oasis; the individual has been deposited at the same place (NMP 49878, alcohol specimen with extracted skull). Measurements of the specimen are included in Tab. 2.
Notes on distribution According to Van Cakenberghe & De Vree (1994), Rhinopoma hardwickii has the widest distribution of all species within the genus. Its range covers deserts and semi-deserts of northern Africa and southern Asia; in Africa the distribution extends southwards along the Nile as far as southern Sudan and central Ethiopia. The distribution of R. hardwickii in Africa was described in detail by several authors (Kock 1969, Koch-Weser 1984, Van Cakenberghe & De Vree 1994). There are many records scattered across Africa north of 15° N in both the marginal and central parts of the Sahara, and some records as far as 5° N in Sudan, and R. hardwickii has been known from all of the Saharan countries except for Libya. Most records come from the Nile Valley of Egypt and Sudan (Kock 1969, Koopman 1975, Qumsiyeh 1985); scattered records are known from Maghreb (Aellen & Strinati 1969, Kock 1969, Aulagnier & Thevenot 1986, Kowalski & Rzebik-Kowalska 1991), western Mauritania (Poulet 1970, Qumsiyeh & Schlitter 1981), and the Sahel belt from Mali to Sudan (Vogel opposite page: Fig. 1. Ruins of an Italian fortress in the centre of the oasis of Al Jaghbub, Cyrenaica, NE Libya, where the shelter of a colony of Rhiniopoma hardwickii was ob-served (photo by A. Reiter) (above). Fig. 2. The oasis of Al Jaghbub, Cyrenaica, NE Libya, view from Italian fortress (photo by A. Reiter) (below).

Two new bat species for Libya 111

112 Benda et al.

Two new bat species for Libya 113
opposite page: Fig. 3. A female of Lesser mouse-tailed bat (Rhinopoma hardwickii) col-lected in the oasis of Al Jaghbub (photo by A. Reiter) (above). Fig. 4. A female of Rüp-pell’s bat (Pipistrellus rueppellii) netted in the oasis of Al Jaghbub (photo by A. Reiter) (below). 1977, Koch-Weser 1984, Kock 1969). From the central Sahara, records come from south-eastern Algeria (Kock 1969, Vesmanis 1985), from central Niger (Thomas 1925, Kock 1969, Hill 1977, Vogel 1977), and from northern Chad (Van Cakenberghe & De Vree 1994). Sites of the Tunisian, Algerian and Chad records are the closest to the territory of Libya. Rhinopoma hardwickii was recorded in Libya for the first time in the northeastern part of the country, close to the Egyptian border. This finding interconnects abundant findings of the species in the Nile Valley with the infrequent records in Maghreb, from Tunisia to Morocco. The continuation of the distribution range along the African coast of the Mediterranean Sea including that of Libya was assumed previously by Kock (1969), Qumsiyeh & Knox Jones (1986), and Van Cakenberghe & De Vree (1994). In their pub-lished maps, these authors assumed that R. hardwickii occurs throughout the whole Lib-yan territory. Occurrence in other oases of continental Cyrenaica (Jalu, Tazirbu, Al Ku-frah, Al Awaynat), Tripolitania (Ghadames, Darj), or of Fezzan (Murzuq, Brak, Ubari, and Ghat regions) is presumable. These places have habitats very similar to those of the Al Jaghbub oasis and are situated close to other sites of records of R. hardwickii in north-ern Africa. Pipistrellus rueppellii is a species of mainly Afro-tropical distribution, but it also inhabits semi-arid regions of northern Africa and the Middle East. In the sub-Saharan Africa, it is widespread in steppe and semi-desert habitats from Senegal in the west to Ethiopia in the east and to South Africa in the south (Hayman & Hill 1971). In the north-ern, Saharan and supra-Saharan parts of Africa, it was recorded abundantly only along the Nile in Sudan and Egypt, from where about 20 findings are known (Kock 1969, Koopman 1975, Qumsiyeh 1985). From other regions of this part of Africa, records of Rüppell’s bat are very scarce; two notes per country come from Morocco (Aulagnier & Thevenot 1986, Arlettaz & Aulagnier 1988) and from Algeria (Hayman & Hill 1971, Gaisler & Kowalski 1986), only one record is known from Chad (Dorst 1963, Vielliard 1974) and Mauritania (Qumsiyeh & Schlitter 1981). In Tunisia, Mali, and Niger, this bat species remains unknown. Unlike the previous species, P. rueppellii was recorded in northern Africa in mar-ginal regions of the Sahara, while it is not known from the whole central Sahara (i.e. from the true continental desert) and from the central parts of the northern Sahara and the African Mediterranean (central and eastern Algeria, Tunisia, and Tripolitania). In Maghreb, this bat is known only from the border of the Sahara and the Atlas Mts. in the western part of the region (Arlettaz & Aulagnier 1988), and in northeastern Africa it had been known until now only from region of the Nile Valley (Kock 1969, Qumsiyeh 1985).

114 Benda et al.
Two new bat species for Libya 115
116 Benda et al.
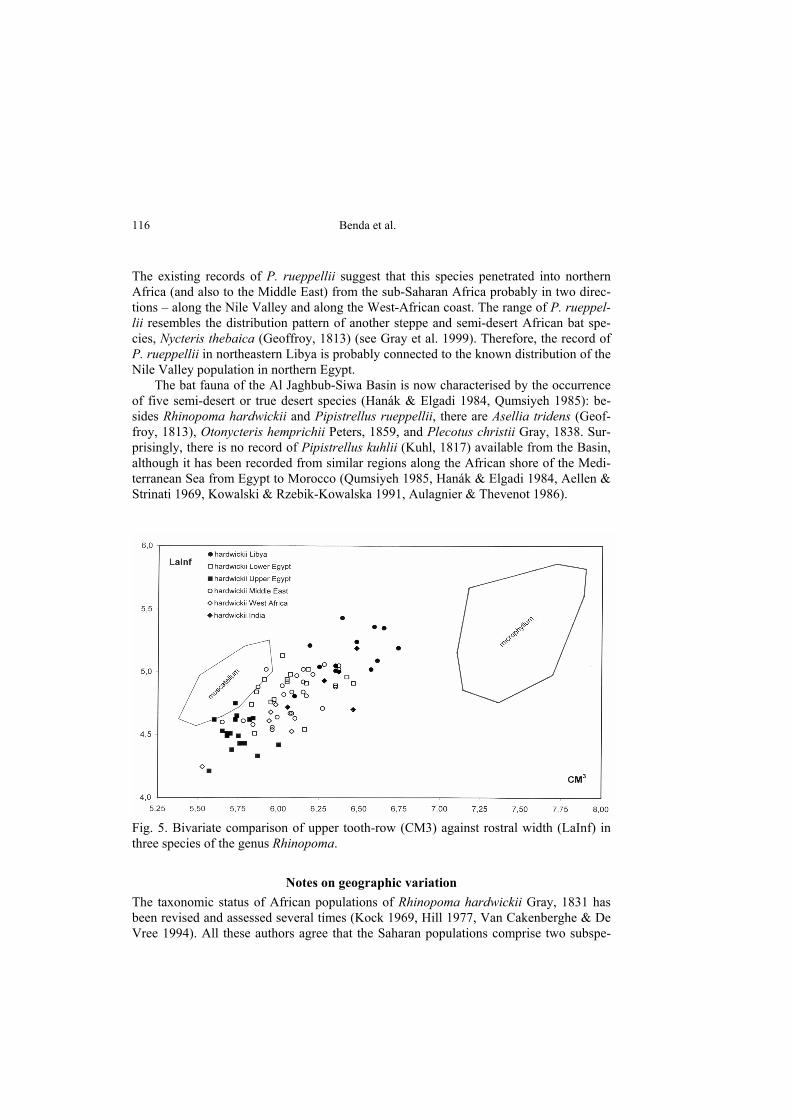
The existing records of P. rueppellii suggest that this species penetrated into northern Africa (and also to the Middle East) from the sub-Saharan Africa probably in two direc-tions – along the Nile Valley and along the West-African coast. The range of P. rueppel-lii resembles the distribution pattern of another steppe and semi-desert African bat spe-cies, Nycteris thebaica (Geoffroy, 1813) (see Gray et al. 1999). Therefore, the record of P. rueppellii in northeastern Libya is probably connected to the known distribution of the Nile Valley population in northern Egypt. The bat fauna of the Al Jaghbub-Siwa Basin is now characterised by the occurrence of five semi-desert or true desert species (Hanák & Elgadi 1984, Qumsiyeh 1985): be-sides Rhinopoma hardwickii and Pipistrellus rueppellii, there are Asellia tridens (Geof-froy, 1813), Otonycteris hemprichii Peters, 1859, and Plecotus christii Gray, 1838. Sur-prisingly, there is no record of Pipistrellus kuhlii (Kuhl, 1817) available from the Basin, although it has been recorded from similar regions along the African shore of the Medi-terranean Sea from Egypt to Morocco (Qumsiyeh 1985, Hanák & Elgadi 1984, Aellen & Strinati 1969, Kowalski & Rzebik-Kowalska 1991, Aulagnier & Thevenot 1986).
Fig. 5. Bivariate comparison of upper tooth-row (CM3) against rostral width (LaInf) in three species of the genus Rhinopoma.
Notes on geographic variation The taxonomic status of African populations of Rhinopoma hardwickii Gray, 1831 has been revised and assessed several times (Kock 1969, Hill 1977, Van Cakenberghe & De Vree 1994). All these authors agree that the Saharan populations comprise two subspe-

Two new bat species for Libya 117
cies, viz., R. hardwickii cystops Thomas, 1903, inhabiting the most arid parts of the cen-tral Sahara – southern Algeria, central Niger, northern Chad, and Upper Egypt – de-scribed from Luxor, Egypt (Thomas 1903), and R. hardwickii arabium Thomas, 1913 living in the marginal regions of the Sahara – Lower Egypt, Maghreb, western Maurita-nia, and the Sahel belt from Mali to Sudan, extralimitally also in Arabia and Persia – described from Wasil, Yemen (Thomas 1913). The specific and subspecific identification of the east-Libyan population of Rhi-nopoma would seem to be simple because the closest records come from the northern part of Egypt and therefore the form inhabiting Lower Egypt and the Nile Delta would be expected to occur in Libya. The Libyan individuals of Rhinopoma (Tab. 1) are larger and fit close to the sample of R. hardwickii from Lower Egypt and the Middle East (Fig. 5), i. e. to the large Afro-Asian form which is traditionally considered as R. h. arabium. They differ completely from the population of Upper Egypt, which belongs to R. h. cystops, and from all samples of R. microphyllum (Brünnich, 1782) (which is much big-ger) and R. muscatellum Thomas, 1903 (which is much smaller). From the simple com-parison based on measurements, and in accordance with the above mentioned authors, the subspecific identification of the Libyan sample seems to be R. hardwickii arabium.
Fig. 6. Bivariate plot of the first two canonical axes of all cranial measurements of Afri-can and Middle Eastern samples of Rhinopoma hardwickii. Nevertheless, a slightly more profound comparison of the Libyan Rhinopoma sample with other populations of R. hardwickii from Africa and the Middle East (Tab. 1) shows differ-ences between the compared samples both in cranial and external dimensions. The Libyan
118 Benda et al.
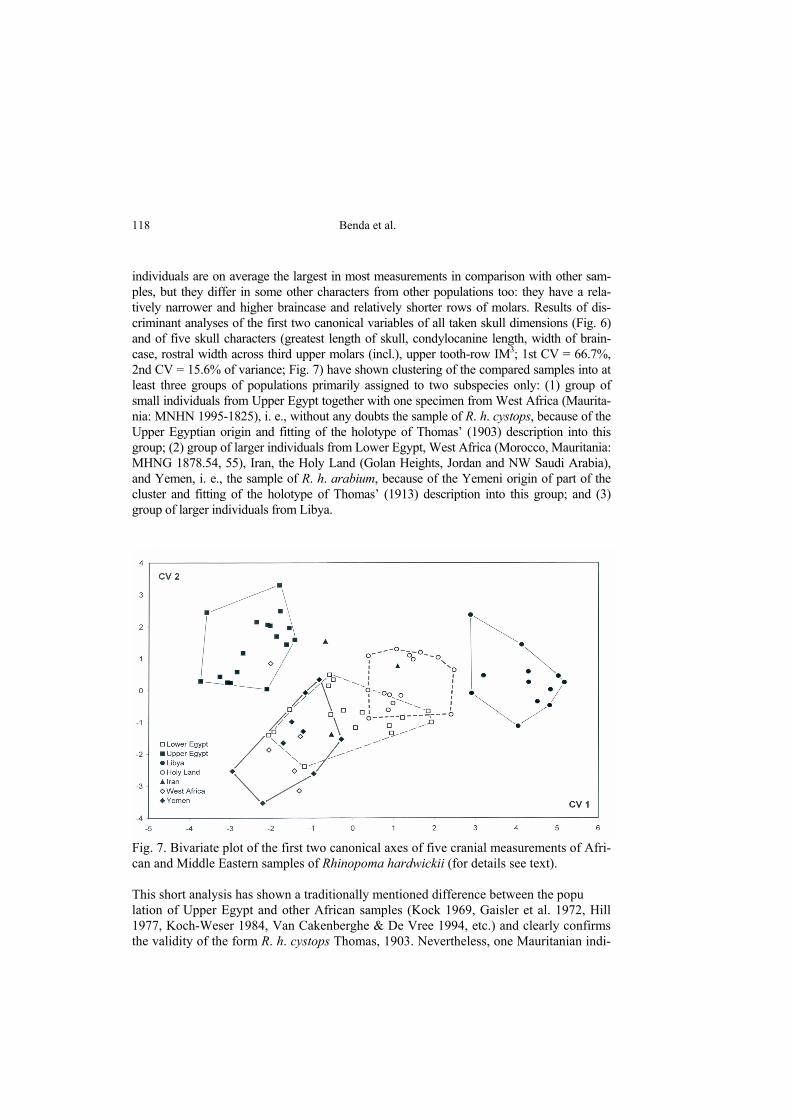
individuals are on average the largest in most measurements in comparison with other sam-ples, but they differ in some other characters from other populations too: they have a rela-tively narrower and higher braincase and relatively shorter rows of molars. Results of dis-criminant analyses of the first two canonical variables of all taken skull dimensions (Fig. 6) and of five skull characters (greatest length of skull, condylocanine length, width of brain-case, rostral width across third upper molars (incl.), upper tooth-row IM3; 1st CV = 66.7%, 2nd CV = 15.6% of variance; Fig. 7) have shown clustering of the compared samples into at least three groups of populations primarily assigned to two subspecies only: (1) group of small individuals from Upper Egypt together with one specimen from West Africa (Maurita-nia: MNHN 1995-1825), i. e., without any doubts the sample of R. h. cystops, because of the Upper Egyptian origin and fitting of the holotype of Thomas’ (1903) description into this group; (2) group of larger individuals from Lower Egypt, West Africa (Morocco, Mauritania: MHNG 1878.54, 55), Iran, the Holy Land (Golan Heights, Jordan and NW Saudi Arabia), and Yemen, i. e., the sample of R. h. arabium, because of the Yemeni origin of part of the cluster and fitting of the holotype of Thomas’ (1913) description into this group; and (3) group of larger individuals from Libya.
Fig. 7. Bivariate plot of the first two canonical axes of five cranial measurements of Afri-can and Middle Eastern samples of Rhinopoma hardwickii (for details see text). This short analysis has shown a traditionally mentioned difference between the popu lation of Upper Egypt and other African samples (Kock 1969, Gaisler et al. 1972, Hill 1977, Koch-Weser 1984, Van Cakenberghe & De Vree 1994, etc.) and clearly confirms the validity of the form R. h. cystops Thomas, 1903. Nevertheless, one Mauritanian indi-
Two new bat species for Libya 119
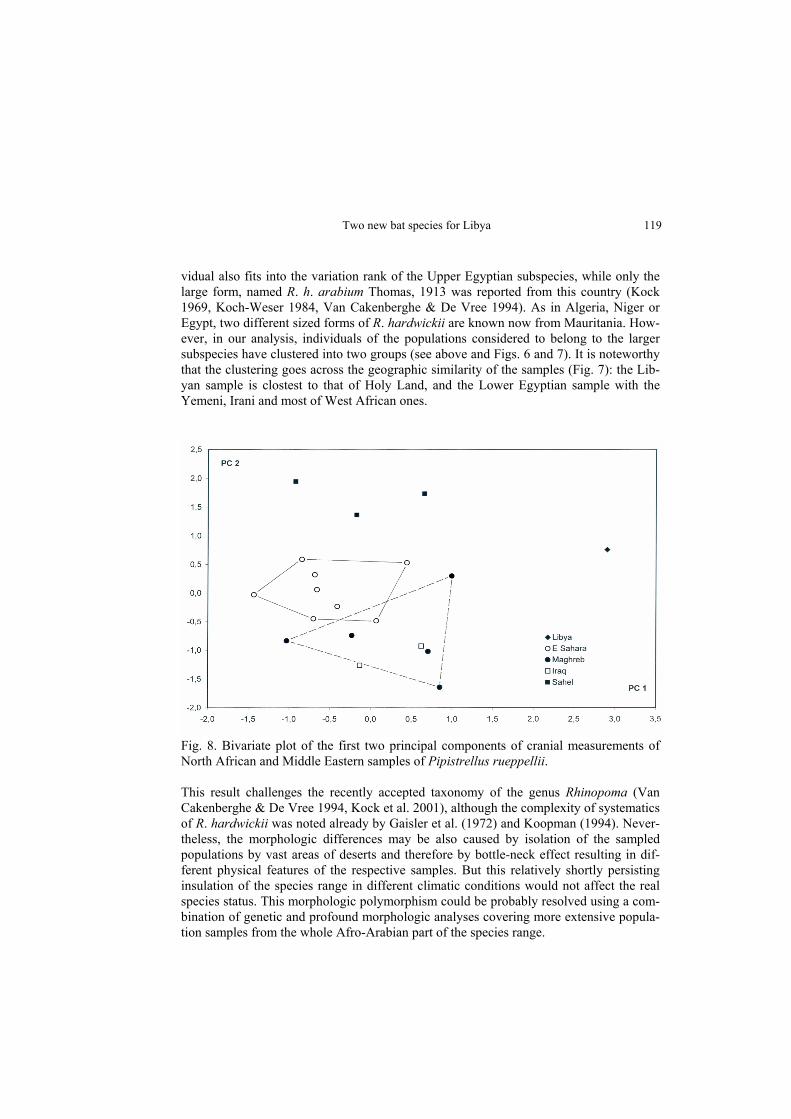
vidual also fits into the variation rank of the Upper Egyptian subspecies, while only the large form, named R. h. arabium Thomas, 1913 was reported from this country (Kock 1969, Koch-Weser 1984, Van Cakenberghe & De Vree 1994). As in Algeria, Niger or Egypt, two different sized forms of R. hardwickii are known now from Mauritania. How-ever, in our analysis, individuals of the populations considered to belong to the larger subspecies have clustered into two groups (see above and Figs. 6 and 7). It is noteworthy that the clustering goes across the geographic similarity of the samples (Fig. 7): the Lib-yan sample is clostest to that of Holy Land, and the Lower Egyptian sample with the Yemeni, Irani and most of West African ones.
Fig. 8. Bivariate plot of the first two principal components of cranial measurements of North African and Middle Eastern samples of Pipistrellus rueppellii. This result challenges the recently accepted taxonomy of the genus Rhinopoma (Van Cakenberghe & De Vree 1994, Kock et al. 2001), although the complexity of systematics of R. hardwickii was noted already by Gaisler et al. (1972) and Koopman (1994). Never-theless, the morphologic differences may be also caused by isolation of the sampled populations by vast areas of deserts and therefore by bottle-neck effect resulting in dif-ferent physical features of the respective samples. But this relatively shortly persisting insulation of the species range in different climatic conditions would not affect the real species status. This morphologic polymorphism could be probably resolved using a com-bination of genetic and profound morphologic analyses covering more extensive popula-tion samples from the whole Afro-Arabian part of the species range.

120 Benda et al.
Three to five subspecies of P. rueppellii are thought to occur in Africa (Hayman & Hill 1971, Koopman 1994). Traditionally, two forms are assigned to the north-African popu-lations, viz., P. rueppellii rueppellii (Fischer, 1829) and P. rueppellii senegalensis Dorst, 1960. Pipistrellus rueppellii was firstly found and described from Dongola, northern Sudan (Cretzschmar 1826, Fischer 1829), therefore the east-Saharan populations (these of Egypt, Sudan and Chad), belong to the nominotypical subspecies, P. r. rueppellii. The west-African populations, recorded from Algeria to Senegal are assigned to P. r. sene-galensis by Koopman (1994). Dorst (1960) described the western subspecies on the basis of a record of one female in northern Senegal which documented the occurrence of this species in western Africa for the first time. Nevertheless, as Hayman & Hill (1971) stated, the validity of the subspecies remains to be shown. Besides its geographical isola-tion, the main difference from the nomimotypical form of P. rueppellii mentioned by Dorst (1960) is large size of P. r. senegalensis in comparison with Sudanese individuals. The Libyan specimen of P. rueppellii is extremely small, and it is the smallest of all compared Rüppell’s bats (Tab. 2). From the comparison of particular measurements and from the principal component analysis of all measurements (Fig. 8) of several individuals of P. rueppellii from the northern part of Africa and the Middle East, an extensive size variation within both west- and east-Saharan populations of the species is evident. Never-theless, results of the PC analysis cluster all specimens into more or less one larger clus-ter with the exception of the Libyan specimen, which is extremely different from all other examined individuals mainly by its small size (PC 1). However, the systematic position of the Saharan and nearby sub-Saharan populations cannot be resolved because there is currently insufficient number of museum specimens. Furthermore, the taxonomic expression of geographical variation in P. rueppellii as a whole should be considered uncertain until adequately large samples from all populations of this species can be analysed (cf. Hayman & Hill 1971).
Acknowledgements The authors thank Jiří Gaisler and Jiří Chamr (Brno), Jacques Cuisin and Ronan Kirsch (Paris), Manuel Ruedi (Geneve), Pauline Jenkins and Daphne Hills (London) for kind providing the mu-seum material of bats. For fruitful discussions and for grammatical improvements we thank Mere-dith Happold and Eva Cepáková. The preparation of this study was supported by a grant from the Grant Agency of the Czech Rep. (206/02/D041), and by grants of the Ministry of Culture of the Czech Rep. (RK 01P03OMG006 and MK 0CEZ99F0201).
References Ael len , V. & P. S t r ina t i (1969): Liste des chiroptères de la Tunisie. – Revue suisse de zoologie
76: 421–431. Ar le t t az , R. & S. Aulagnie r (1988): Statut de trois espèces de chiroptères rares au Maroc:
Nycteris thebaica, Hipposideros caffer et Pipistrellus rueppelli. – Zeitschrift für Säugetierkun-de 53: 321–324.
Aulagnie r , S. & M. Thevenot (1986): Catalogue des mammiferes sauvages du Maroc. – Tra-vaux de l’Institute Scientifique, Série Zoologie 41: 1–164.

Two new bat species for Libya 121
Cre tzschmar , P. J. (1826): Säugethiere. – In: Atlas zu der Reise in nördlichen Afrika von Eduard
Rüppell. Ed. by E. Rüppell. Frankfurt am Mein: H. L. Brönner: 1–78. De Beaux , O. (1928): Risultati zoologici della Missione inviata dalla R. Societa Geografica
Italiana per l’esplorazione dell’oasi di Giarabub (1926–1927). – Annali del Museo Civico di Storia Naturale di Genova 53: 39–76.
Dors t , J. (1960): A propos de quelques chiroptères du Sénégal, et description d’une forme nouvel-le du genre Pipistrellus. – Bulletin du Muséum National d’Histoire Naturelle, Paris, 2. Sér., 31[1959]: 471–474.
Dors t , J. (1963): Présence de Pipistrellus rüppelli au Tchad. – Mammalia 27: 443. F i scher , J. B. (1829): Synopsis mammalium. – Stuttgardt: J. G. Cotta, 527 pp. Ga i s le r , J. & K. Kowalsk i (1986): Results of the netting of bats in Algeria (Mammalia: Chirop-
tera). – Věstník Československé společnosti zoologické 50: 161–173. Ga is le r J., G. Madkour & J. Pe l ikán (1972): On the bats (Chiroptera) of Egypt. – Acta scien-
tarium naturalium Academiae scientarium bohemoslovacae Brno 6(8): 1–40. Gray , P. A., M. B. Fen ton & V. Van Cakenberghe (1999): Nycteris thebaica. – Mammalian
Species 612: 1–8. Hanák , V. & A. E lgad i (1984): On the bat fauna (Chiroptera) of Libya. – Věstník
Československé společnosti zoologické 48: 165–187. Hay man, R. W. (1948): The Armstrong College Zoological Expedition to Siwa Oasis (Libyan
Desert) 1935. Mammalia. – Proceedings of the Egyptian Academy of Science 4: 36–42. Hay man, R. W. & J. E. Hi l l (1971): Part 2. Order Chiroptera. – In: The Mammals of Africa: an
Identification Manual. Ed. by J. Meester & H. W. Setzer. Washington: Smithsonian Institution Press: 1–73.
Hufnag l , E. (1972): Libyan Mammals. – London: The Oleander Press, 85 pp. Hutson , A. M., S. P. Micklenburgh & P. A. Racey (2001): Microchiropteran Bats. Global Status
Survey and Conservation Action Plan. – Gland and Cambridge: IUCN, 258 pp. Hi l l , J. E. (1977): A review of the Rhinopomatidae (Mammalia: Chiroptera). – Bulletin of the
British Museum Natural History (Zoology) 32: 29–43. Klap tocz , B. (1909): Beitrag zur der Säuger von Tripolis und Barka. – Zoologische Jahrbücher,
Abteilung für Systematik, Geographie und Biologie der Tiere 27: 237–272. Koch-Weser , S. (1984): Fledermäuse aus Obervolta, W-Afrika (Mammalia: Chiroptera). – Sen-
ckenbergiana biologica 64: 255–311. Kock , D. (1969) Die Fledermaus-Fauna des Sudan (Mammalia, Chiroptera). – Abhandlungen der
Senckenbergischen Naturforschenden Gesellschaft 521: 1–235 . Kock , D., M. M. Al -Jumai ly & A. K. Nasher (2001): On the genus Rhinopoma E. Geoffroy
1818, and a record of Rh. muscatellum Thomas 1903 from Yemen (Mammalia, Chiroptera, Rhinopomatidae). – Senckenbergiana biologica 81: 285–287.
Koopman, K. F. (1975): Bats of The Sudan. – Bulletin of the American Museum of Natural History 154: 354–443.
Koopman, K. F. (1994): Chiroptera: Systematics. Handbook of Zoology. Volume VIII. Mamma-lia. Part 60. – Berlin-New York: Walter de Gruyter, 224 pp.
Kowalsk i , K. & B. Rzebik-Kowalska (1991): Mammals of Algeria. – Wrocław: Zakład Narodowy im. Ossolińskich, 370 pp.
Poule t , A. R.. (1970): Les Rhinopomatidae de Mauritanie. – Mammalia 34: 237–243. Qumsiy eh , M. B. (1983): Occurrence and zoogeographical implications of Myotis blythi (Tomes,
1857) in Libya. – Mammalia 47: 429–430. Qumsiy eh , M. B. (1985): The bats of Egypt. – Special Publications, The Museum Texas Tech
University 23: 1–101.

122 Benda et al.
Qumsiy eh , M. B. & J. Knox Jones Jr. (1986): Rhinopoma hardwickii and Rhinopoma musca-
tellum. – Mammalian Species 263: 1–5. Qumsiy eh , M. B. & D. A. Sch l i t t e r (1981): Bat records from Mauritania, Africa. – Annals of
Carnegie Museum 50: 345–351. Qumsiy eh , M. B. & D. A. Sch l i t t e r (1982): The bat fauna of Jabal Al Akhdar, Northeast Li-
bya. – Annals of Carnegie Museum 51: 377–389. Ranck , G. L. (1968): The Rodents of Libya. Taxonomy, Ecology, and Zoogeographical Relati-
onships. – U. S. National Museum Bulletin 275: 264 pp + 8 plates. Se tze r , H. W. (1957): A review of Libyan mammals. – Journal of the Egyptian Public Health
Association 32: 41–82. Sp i t zenberger , F. (1982): Der Riesenabendsegler (Nyctalus lasiopterus) in Afrika nachgewie-
sen. – Zeitschrift für Säugetierkunde 47: 115–116. Thomas , O. (1903): On the Species of the Genus Rhinopoma. – Annals and Magazine of Natural
History, Ser. 7, 11: 496–499. Thomas , O. (1913): Some new Ferae from Asia and Africa. – Annals and Magazine of Natural
History, Ser. 8, 12: 88–92. Thomas , O. (1925): On the Mammals (other than Ruminants) collected by Captain Angus Bu-
chanan during his Second Saharan Expedition, and presented by him to the National Museum. – Annals and Magazine of Natural History, Ser. 9, 16: 187–197.
Tosch i , A. (1954): Elenco preliminare dei Mammiferi della Libia. – Supplemento alle Ricerche di Zoologia Applicata alla Caccia 2: 241–273.
Van Cakenberghe , V. & F. De Vree (1994): A revision of the Rhinopomatidae Dobson 1872, with the description of a new subspecies (Mammalia: Chiroptera). – Senckenbergiana biologica 73: 1–24.
Vesmanis , I. (1985): Kleinsäugertiere aus Algerien. – Zoologische Abhandlungen Staatliches Museum für Tierkunde Dresden 40: 125–152.
Vie l l i a rd , J. (1974): Les Chiroptères du Tchad. – Revue Suisse de zoologie 81: 975–991. Voge l , P. (1977): Neue Nachweise der Rhinopoma hardwickei (Chiroptera) aus Westafrika. –
Bonner zoologische Beiträge 28: 228–231. Zava t t a r i , E. (1934): Prodromo della fauna della Libia. – Pavia, 1234 pp. Zava t t a r i , E. (1937): I Vertebrati della Libia. – In: Festschrift für Prof. Dr. Embrik Strand. Vol.
II.: 526–560.
Petr Benda, Department of Zoology, National Museum (Natural History), Václavské nám. 68, CZ–115 79 Praha 1, Czech Republic; [email protected]; Vladimír Hanák, Department of Zoology, Charles University, Viničná 7, CZ–128 44 Praha 2, Czech Republic; Michal Andreas, Agency for Nature Conservation and Landscape Protection of the Czech Repub-lic, Kališnická 6, CZ–133 23 Praha 3, Czech Republic; Antonín Reiter, South Moravian Museum Znojmo, Přemyslovců 8, CZ–669 45 Znojmo, Czech Republic; Marcel Uhrin, National Park Muránska planina, J. Kráľa 12, SK–050 01 Revúca, Slovakia

Two new bat species for Libya 123
Appendix 1 List of comparative material Only skulls used. Abbreviations: m – male, f – female, ind. – sex not determined, BMNH – collec-tion of Natural History Museum, London, United Kingdom, IVB – collection of Institute of Verte-brate Biology, Brno, Czech Rep., MHNG – collection of Natural History Museum, Geneve, Swit-zerland, MNHN – collection of National Museum of Natural History, Paris, France, MUB – collec-tion of Department of Zoology, Masaryk University, Brno, Czech Rep., NMP – collection of Na-tional Museum (Natural History), Prague, Czech Rep. Rhinopoma microphyllum (Brünnich, 1782) India: 1 m (BMNH 11.12.11.1., holotype of Rhinopoma kinneari Wroughton, 1912), Bhuj, Cutch, 28 August 1911; – 3 f (MNHN 1985-1390, 1392, 1393), Fathepur Sikri (Agra), March 1961, leg. A. Brosset; – 1 f (MNHN 1985-1391), Sanchi, March 1961, leg. A. Brosset; – 3 ind. (MNHN 1985-1963–1965), Orcha, May 1961, leg. A. Brosset; – 1 ind. (MHNG 913.8), Sanchi, Bhopal, Oct. 1955, leg. Pittet. Afghanistan: 1 m (MHNG 952.82) Grande Grotte Moumlaï, Farah, 26 April 1958, leg. K. Lindberg; – 1 m (MHNG 952.77), Grotte Chamchir, Kandahar, 4 Dec. 1957, leg. K. Lindberg; – 6 m, 4 f (IVB AF 582, 606, 633, 634, 638–641, 643, 645), Chack-Naur, 7 April 1967, leg. J. Gaisler. Pakistan: 4 f (MNHN 1983-1630, 1631, 1997, 1999), Karachi, 21 June 1961, leg. M. A. El Hussein. Lebanon: 1 ind. (MNHN 1983-1962), Grotte de Litani, 1962, leg. Khairallah. Jordan: 1 f (NMP 47965), Tabaqat Fal, 4 July 1997, leg. P. Benda. Egypt: 1 ind. (MHNG 683.31), Theben, 1905. Mauritania: 1 f (MNHN 1995-1824), Guelb Moghrein, October 1968, leg. A. R. Poulet; – 1 ind. (MNHN 1995-3142), Fourt de Moudieria, 30 December 1994, leg. F. Colas. Senegal: 1 f (MNHN 1995-1823), Dakar, 2 December 1971, leg. R. Taufflieb. Rhinopoma hardwickii Gray, 1831 India: 1 m (BMNH i.e., holotype of Rhinopoma hardwickii Gray, 1831), India, leg. Hardwicke; – 1 m (NMHN 1902-502a), Genji Coromandel, Aug.–Sept. 1901, leg. M. Maindron; – 1 m (MNHN 1962-343b), Vijayanagar, Oct. 1960, leg. A. Brosset; – 2 f (MNHN 1985-1386, 1387), Badami, Nov. 1960, leg. A. Brosset; – 1 f (MNHN 1985-1388), Hampi, Nov. 1960, leg. A. Brosset. Iran: 2 f (NMP 48162, 48163), Izeh (Khuzestan Prov.), 12 October 1998, leg. A. Reiter & M. Uhrin; – 1 m (NMP 48157), Jelugir (Lorestan Prov.), 11 October 1998, leg. J. Obuch. Jordan: 9 m (NMP 47966–47974), Tabaqat Fal, 4 July 1997, leg. P. Benda. Saudi Arabia: 2 m, 2 f (BMNH 48.351.–354.), Al Bada, 300 mts, 24–27 March 1947, leg. G. Popov. Syria/Israel: 2 m (NMP 48268, 48269), Nimrod Fortress (Golan Heights), 18 July 1999, leg. P. Benda. Yemen: 5 m, 2 f (BMNH 13.6.19.1.–5., 13.6.19.7., 13.6.19.8., incl. holotype of Rhinopoma cystops arabium Thomas, 1913), Wasil, 4000 ft., 6 March 1913, leg. G. W. Bury; – 1 m, 1 f (BMNH 99.11.6.29, 99.11.6.32.), Myba, near aden, leg. W. Dodson. Egypt: 1 f (BMNH 2.1.17.2., holotype of Rhinopoma cystops Thomas, 1903), Luxor, leg. N. C. Ro-tschild; – 1 m, 7 f (IVB E-23, E-24, E-26, E-27, E-34–36, E-38), Cairo, Bar Kouky Mosque, 21 April 1969, leg. J. Gaisler; – 2 m, 10 f (IVB E-135–144, E-146, E-147), Dandara, Temple of Hathor, 27 April 1969, leg. J. Gaisler; – 1 m, 2 f (IVB E-81, E-82, E-84), Karnak, Temple, 26 April 1969, leg. J. Gaisler; – 1 m (IVB E-199), Luxor, Valley of the Kings, 30 April 1969, leg. J. Gaisler; – 4 m, 4 f (IVB E-40–42, E-49, E-50, E-55–57), Sakkara, Prison of Joseph, 23 April 1969, leg. J. Gaisler. Morocco: 2 f (MNHN 1954-360a, 360e), Tata, 25 April 1954, leg. J. Dorst.

124 Benda et al.
Mauritania: 1 f (MNHN 1995-1825), Guelb Moghrein, October 1968; – 2 m (MHNG 1878.54, 55), Atar, Grotte du Lapin, 1 March 1951, leg. P. L. Dekeyser & A. Villiers. Rhinopoma muscatellum Thomas, 1903 Iran: 3 m, 1 f (NMP 48443–48446), Hormoz I. (Hormozgan Prov.), 17 April 2000, leg. P. Benda & A. Reiter; – 1 m, 1 f (NMP 48463, 48464), Pol-e-Tang (Lorestan Prov.), 11 April 1977, leg. B. Pražan; – 8 m, 1 f (NMP 48164–48172), Izeh (Khuzestan Prov.), 12 October 1998, leg. A. Reiter & M. Uhrin; – 7 m, 2 f (NMP 48387–48395), Kahiri (Sistan-ve-Baluchestan Prov.), 11 April 2000, leg. J. Obuch & A. Reiter; – 1 m, 1 f (NMP 48421, 48422), Pir Sohrab (Sistan-ve-Baluchestan Prov.), 12 April 2000, leg. A. Reiter; – 1 f (NMP 48423), Tis (Sistan-ve-Baluchestan Prov.), 13 April 2000, leg. J. Obuch. Oman: 1 m (BMNH 94.3.9.17., holotype of Rhinopoma muscatellum Thomas, 1903), Wadi Bani Ruba, near Muscat, leg. Jayaker. Pipistrellus rueppellii (Fischer, 1829) Egypt: 1 ind. (MNHN 1985-1032), Egypt, 1829, leg. A. Lefébre; – 1 m (BMNH 92.9.9.20.), Luxor, leg. J. Anderson. Sudan: 1 f (MHNG 1626.22), Kerma, 40 km amont de la 3e chute, bord du Nil, Jan. 1983, leg. L. Chaix; – 2 m (BMNH 1.8.8.13.–14.), Khartoum, 24 January and 21 March 1901, leg. R. M. D. Hawker; – 1 m (BMNH 1.8.8.15.), Fashoda, leg. R. M. D. Hawker; – 1 f (BMNH 1.8.8.16.), Goz Abu Gumer, 17 May 1901, leg. R. M. D. Hawker; – 1 f (BMNH 15.3.6.63), Tongo, White Nile, leg. W. P. Lowe. Morocco: 1 f (NMP 90057), Oued Drâa, 5 km E Anagam, 31 August 2003, leg. P. Benda; – 2 m (NMP 90080, 90081), Cascades Bou Mazouz, 10 km NW Taourirt, 6 September 2003, leg. P. Benda; – 1 f (MHNG 1706.28), Aoufouss, 19–20 April 1985, leg. R. Arlettaz. Algeria: 1 m (MUB A 496), Abadla, Oued Guir, 20 July 1983, leg. J. Gaisler. Senegal: 1 m (MNHN 1995-2677), Parc National Le Djoudj, April 1972. Nigeria: 1 m (MHNG 1717.61), Baga-Kawwa, 31 May 1970, leg. J. Vielliard. Central African Rep.: 1 f (MNHN 1985-1035), Bangui, leg. A. Chippeaux. Iraq: 1 ind. (BMNH 19.3.1.3., holotype of Pipistrellus coxi Thomas, 1919), Beit Mahommad, Amara, near Chahala canal, 20 March 1918, leg. R. E. Cheesman; – 1 ind. (BMNH 20.5.22.1.), Busrah, Mesopotamia, leg. Christy. Appendix 2 Abbreviations of measurements used in Tabs. 1, 2 LC = head and body length; LCd = tail length; LAt = forearm length (incl. wrist); LA = auricle length; LT = tragus length; G = weight; LCr = greatest length of skull; LCb = condylobasal length of skull; LCc = condylocanine length of skull; LaZ = zygomatic width; LaI = width of interorbital constriction; LaInf = rostral width between foramina infraorbitalia; LaN = braincase width; ANc = braincase height; ACr = skull height (incl. tympanic bullae); LBT = length (largest diameter) of tympanic bulla; CC = rostral width between canines (incl.); P4P4 = rostral width between third upper premolars (incl.); M3M3 = rostral width between third upper molars (incl.); I1M3 = length of upper tooth-row between first incisor (I1) and last molar (M3) (incl.); CM3 = length of upper tooth-row (between CM3, incl.); M1M3 = length of upper molar-row; CP4 = length of upper unicuspidal tooth-row; LMd = mandible length; ACo = height of coronoid process of the mandible; I1M3 = length of lower tooth-row between first incisor (I1) and third molar (M3) (incl.); CM3 = length of lower tooth-row (between CM3, incl.); M1M3 = length of lower molar-row; CP4 = length of lower unicuspidal teeth-row.