universidad de chile · cÁscara de manzana sobre helicobacter pylori: ... sfrbm south american...
TRANSCRIPT

UNIVERSIDAD DE CHILE
FACULTAD DE CIENCIAS QUIMICAS Y FARMACEUTICAS
“ACTIVIDAD DE EXTRACTOS RICOS EN POLIFENOLES DE
CÁSCARA DE MANZANA SOBRE Helicobacter pylori: ESTUDIOS
IN VITRO E IN VIVO”
Tesis presentada a la Universidad de Chile para optar al grado de Doctor en Farmacología por:
EDGAR RAFAEL PASTENE NAVARRETE
Directores de Tesis
Dr. Hernán Speisky C.
Dr. Guillermo Figueroa G.
SANTIAGO-CHILE
2010

2
UNIVERSIDAD DE CHILE FACULTAD DE CIENCIAS QUÍMICAS Y FARMACÉUTICAS
INFORME DE APROBACIÓN
TESIS DE DOCTORADO
Se informa a la Dirección de Postgrado de la Facultad de Ciencias Químicas y Farmacéuticas que la Tesis de Doctorado presentada por el candidato:
EDGAR RAFAEL PASTENE NAVARRETE
Ha sido aprobada por la Comisión Informante de Tesis como requisito para optar al grado de Doctor en Farmacología, en el examen de defensa de Tesis rendido el día _____________________ Directores de Tesis: Dr. Hernán Speisky C. ___________________________ Dr. Guillermo Figueroa G. ___________________________ Comisión Informante de Tesis: Dra Inés Contreras (Presidente) ___________________________ Dra. Apolinaria García C. ___________________________ Dr. Aurelio San Martín ___________________________ Dr. Romilio Espejo ___________________________

3
PUBLICACIONES
Edgar Pastene, Miriam Troncoso, Guillermo Figueroa, Julio Alarcón and Hernán
Speisky. Association between Polymerization Degree of Apple Peel Polyphenols
and Inhibition of Helicobacter pylori Urease. J. Agric. Food Chem. 2009, 57, 416-
424.
Edgar Pastene, Hernán Speisky, Miriam Troncoso, Julio Alarcón, and Guillermo
Figueroa. In vitro Inhibitory Effect of Apple Peel Extract on the Growth of
Helicobacter pylori and Respiratory Burst Induced on Human Neutrophils. J. Agric.
Food Chem. 2009, 57, 7743-7749.
Edgar Pastene, Hernán Speisky, Apolinaria García, Jessica Moreno, Miriam
Troncoso and Guillermo Figueroa. In vitro and In vivo Effects of Apple Peel
Polyphenols against Helicobacter pylori. J. Agric. Food Chem. 2010, 58, 7172-
7179.
PRESENTACIONES A CONGRESOS
Edgar Pastene, G. Figueroa, A. García, M. Troncoso, y H. Speisky. In vitro and in
vivo Protective Effect of an Apple Peel Extract on The oxidative Damage Induced
by Helicobacter pylori. Free Radicals and Antioxidants in Chile 2009, VI Meeting of
SFRBM South American Group. September 2009, Santiago, Chile. Poster.
Edgar Pastene: Generación in situ de moléculas activas derivadas de polifenoles
durante el proceso inflamatorio: Efecto de la inducción del estallido respiratorio de
neutrófilos humanos por Helicobacter pylori sobre antioxidantes polifenólicos.
Primer Congreso Internacional de Farmacobotánica, 2009, Chillán, Chile.
Presentación oral.

4
Edgar Pastene; Troncoso, M.; Figueroa, G.; Alarcón, J.; Speisky, H. Efecto de los
polifenoles de cáscara de manzana sobre el estallido respiratorio de neutrófilos
humanos inducido por Helicobacter pylori no-opsonisado. XVIII Congreso ALF,
Reunión Anual Sociedad de Farmacología de Chile, XXIII Sociedad Chilena de
Ciencias Fisiológicas, III Congreso Iberoamericano de Farmacología. Coquimbo,
Chile, octubre de 2008. Poster.
Edgar Pastene; J. Alarcón. Role of Natural Products in Drug Design (Rol de los
productos Naturales en el diseño de drogas). XXIX Congreso Anual de la
Sociedad de Farmacología de Chile, Iquique, Chile, septiembre de 2007.
Presentación oral.
PATENTES
Solictud N°1344-08. Procedimiento de preparación de un extracto enriquecido en
polifenoles a partir de cáscara de manzana; extracto seco enriquecido en polifenoles,
composición farmacéutica y/o alimenticia; y uso de la preparación en la prevención y/o
tratamiento de alteraciones y enfermedades digestivas asociadas con la infección por
Helicobacter pylori”. Fecha publicación diario oficial: 6 de noviembre de 2008. Estado de
tramitación: en examen pericial.

5
FINANCIAMIENTO
Beca CONICYT (Apoyo Tesis Doctoral Nº 24080071)
Recursos Intramurales Laboratorio de Antioxidantes (INTA, Universidad de Chile)
Recursos Intramurales Laboratorio de Microbiología y Probióticos (INTA, Universidad de
Chile).

6
ÍNDICE GENERAL
Página PUBLICACIONES 2
PRESENTACIONES A CONGRESOS 2
FINANCIAMIENTO 4
ÍNDICE GENERAL 5
ÍNDICE DE FIGURAS 9
ÍNDICE DE TABLAS 13
RESUMEN 14
ABSTRACT 18
ABREVIATURAS 22
1. INTRODUCCION 27
1.1. Epidemiología y principales manifestaciones clínicas de la infección por
H. pylori
27
1.2. Estrategias de supervivencia de H. pylori: Evasión de la respuesta
inmune e inflamatoria
28
1.3. Rol de la ureasa, VacA y CagA 30
1.3.1. Ureasa 30
1.3.2. Toxina VacA 31
1.3.3. Citotoxina CagA 32
1.4. H. pylori y evasión del estrés oxidativo 33
1.5. Efectos de la respuesta inmuno-celular y del estrés oxidativo sobre el
huésped
34
1.6. Terapias de erradicación de H. pylori 35
1.7. Efecto de los polifenoles sobre H. pylori 37
1.7.1. Antocianos, flavonoides, flavan-3-oles, procianidinas y taninos
hidrolizables
40
1.7.2 La cáscara de manzana como fuente de polifenoles 44
1.7.3 Actividad antioxidante y potencial actividad anti-H. pylori de polifenoles
presentes en la cáscara de manzana
47

7
2 HIPÓTESIS DE TRABAJO 52
3 OBJETIVO GENERAL 53
4 OBJETIVOS ESPECÍFICOS 53
5 MATERIALES Y MÉTODOS 54
5.1. Reactivos y solventes 54
5.2. Preparación del extracto rico en polifenoles de cáscara de manzana
(APPE)
54
5.3. Determinación cuantitativa de azúcares por HPTLC 55
5.4. Determinación del perfil de compuestos fenólicos en el extracto de
cáscara de manzana mediante RP-HPLC-DAD-FLD
55
5.5. Determinación del grado de polimerización de las procianidinas
presentes en el extracto de cáscara de manzana
56
5.5.1 HPLC en Fase Normal (NP-HPLC) 56
5.5.2. RP-HPLC-FLD de los productos de tiólisis con tolueno-tiol 56
5.5.3. RP-HPLC-FLD de los productos de tiólisis con cisteamina 57
5.5.4. RP-HPLC-FLD de los productos de floroglucinólisis 57
5.6. Análisis del perfil de compuestos fenólicos en el extracto de cáscara de
manzana mediante RP-HPLC-ESI-MS/MS
58
5.7. Fraccionamiento de APPE para la obtención de los polifenoles de alto y
bajo peso molecular
60
5.8. Efecto del extracto crudo de cáscara de manzana (APPE) sobre la
actividad de la enzima ureasa extraída de Jack bean y H. pylori
61
5.9. Influencia de los extractos de cáscara de manzana sobre parámetros
cinéticos de la enzima ureasa de Jack bean
63
5.10. Detección “in-gel” de la actividad anti-ureasa de Jack bean 63
5.11. Protocolo modificado para la detección “in-gel” de la actividad anti-
ureasa de H. pylori
64
5.12. Efecto de APPE sobre los niveles intracelulares de ATP de H. pylori
como indicador de viabilidad
65
5.13. Detección de la citotoxicidad temprana de los extractos de cáscara de
manzana utilizando como indicador la reducción de resazurina
66
5.14. Efecto de los extractos de cáscara de manzana sobre la respiración de 67

8
H. pylori medido por la sonda MitoXpress
5.15. Obtención de los neutrófilos humanos 67
5.16. Efecto de los extracto de cáscara de manzana sobre la Inducción del
estallido respiratorio por PMA, fMLP y H. pylori
68
5.17. Análisis de la estabilidad de los polifenoles de APPE en fluido gástrico
simulado
69
5.18. Cultivo de células HeLa para el estudio del efecto inhibitorio de los
polifenoles sobre la actividad de la toxina VacA
70
5.19. Obtención de sobrenadantes enriquecidos en VacA a partir de cultivos
de H. pylori
70
5.20. Determinación de proteínas 71
5.21. Ensayo de vacuolación 71
5.22. Ensayo de anti-adhesion de H. pylori cultivos de células HeLa 71
5.23. Determinación de la unión inespecífica y el ingreso de polifenoles de
APPE en células HeLa
72
5.24. Animales 72
5.25. Preparación del inoculo para el estudio in vivo 73
5.26. Protocolo experimental 73
5.27. Evaluación histopatológica 73
5.28. Niveles de MDA 74
5.29. Determinación cuantitativa de H. pylori por PCR de tiempo real (real-time
PCR)
74
5.30. Expresión de resultados y análisis estadístico 75
6. RESULTADOS 76
6.1. Preparación del extracto enriquecido en polifenoles de cáscara de
manzana libre de azúcares (APPE)
76
6.2. Análisis de la composición de los extractos de cáscara de manzana
mediante RP-HPLC-DAD-FLD y NP-HPLC-FLD
77
6.3. Determinación del grado de polimerización por degradación con los
nucleófilos tolueno--tiol, cisteamina y floroglucinol
80
6.4. Análisis del perfil de compuestos fenólicos en el extracto de cáscara de
manzana mediante RP-HPLC-ESI-MS/MS
84

9
6.5. Efecto del extracto de cáscara de manzana sobre la viabilidad de H.
pylori
88
6.5.1. Determinación de la actividad anti-H. pylori de APPE mediante
procedimientos convencionales
88
6.5.2 Viabilidad de H. pylori en periodos cortos de contacto APPE-bacteria
utilizando como indicadores la reducción de resazurina y los niveles de
ATP
89
6.6 Efecto de los extractos de cáscara de manzana sobre la actividad de la
ureasa de Jack bean
92
6.7. Reactivación por DTT de la actividad ureolítica de la ureasa Jack bean
pre-incubada con extracto APPE
94
6.8. Influencia de los extractos de cáscara de manzana sobre parámetros
cinéticos de la enzima ureasa de Jack bean
96
6.9. Efecto de APPE la actividad de la ureasa extraída de H. pylori 97
6.10. Detección “in-gel” de la actividad anti-ureasa de Jack bean 100
6.11. Efecto inhibitorio in vitro del extracto de cáscara de manzana (APPE)
sobre el estallido respiratorio de neutrófilos inducido por PMA, fMLP y H.
pylori no-opsonizado
103
6.12. Efecto inhibitorio de APPE sobre la actividad de la toxina VacA en
células HeLa
109
6.13. Efecto de APPE sobre la adhesión de H. pylori a cultivos de células
HeLa
114
6.14. Efecto de APPE sobre la adhesión de H. pylori a la mucosa gástrica de
ratones
116
6.15. Efecto de APPE sobre los niveles de MDA en la mucosa gástrica de
ratones infectados por H. pylori
119
7 DISCUSIÓN 121
7.1. Obtención y caracterización del extracto de cáscara de manzana APPE 122
7.2. Los polifenoles de APPE poseen un efecto anti-H. pylori asociado con su
tamaño molecular
124
7.2.1 Efecto de los polifenoles de APPE sobre la viabilidad de H. pylori 124
7.2.2. Efecto de los polifenoles de APPE sobre la actividad ureásica de H. 126

10
pylori
7.3 APPE inhibe la producción de especies reactivas del oxígeno en
neutrófilos activados por PMA, fMLP y H. pylori no-opsonizados
128
7.4 APPE inhibe el efecto vacuolante de VacA en células HeLa 132
7.5 APPE interfiere en el proceso de adhesión de H. pylori a cultivos de
células HeLa
134
7.6 APPE impide la colonización de H. pylori en ratones C57BL6J utilizados
como modelo de infección a corto plazo
135
8. CONCLUSIONES 140
9. REFERENCIAS 142
ÍNDICE DE FIGURAS
Página
Figura 1. Rol de la ureasa en la colonización de la mucosa gástrica por
H. pylori. ----------------------------------------------------------------------------------- 31
Figura 2. Historia natural de la infección por H. pylori: relación entre
patología y presentación clínica. ---------------------------------------------------- 38
Figura 3. Estructura de los principales polifenoles presentes en la
manzana. --------------------------------------------------------------------------------- 47
Figura 4. Potenciales sitios de acción de los polifenoles de la cáscara de
manzana como agentes citoprotectores y anti-H. pylori ----------------------- 52
Figura 5. Esquema de fraccionamiento del extracto APPE para la
obtención de sus constituyentes de alto y bajo peso molecular. ------------ 62
Figura 6. Esquema de preparación de una membrana sensible a ureasa
para la investigación de inhibidores. ----------------------------------------------- 66

11
Figura 7. Perfiles RP-HPLC-DAD de APPE y cáscara de manzana fresca ------------- 79
Figura 8. TLC en fase normal de fracciones ACUO purificada por
sephadex LH-20 ------------------------------------------------------------------------- 80
Figura 9. Separación de procianidinas de APPE mediante NP-HPLC-FLD ------------ 80
Figura 10. Depolimerización de las procianidinas de APPE utilizando los
agentes nucleofílicos floroglucinol, cisteamina y tolueno--tiol. ------------- 82
Figura 11. Cromatogramas representativos de la depolimerización de
APPE usando diferentes nucleófilos ----------------------------------------------- 83
Figura 12. Estrategia de identificación de los aductos formados entre
cisteamina y flavan-3-oles de APPE ----------------------------------------------- 84
Figura 13. Perfiles de RP-HPLC-UV y ESI/MS para APPE --------------------------------- 88
Figura 14. Titulación de H. pylori viables usando resazurina ------------------------------- 91
Figura 15. Efecto del extracto APPE sobre los niveles intracelulares de
ATP y el consumo de oxígeno de H. pylori (ATCC 43504) ------------------- 93
Figura 16. Efecto del extracto crudo de cáscara de manzana APPE
sobre la actividad ureasa de Jack bean como función del
tiempo de preincubación -------------------------------------------------------------- 94
Figura 17 Gráficas semi-log de concentración de extracto crudo de
cáscara de manzana APPE, versus efecto inhibitorio de la
enzima ureasa de Jack bean a diferentes tiempos de
preincubación. --------------------------------------------------------------------------- 95

12
Figura 18. Reactivación con DTT de la ureasa Jack bean inhibida por el
extracto APPE --------------------------------------------------------------------------- 96
Figura 19. Efecto del DTT sobre la inhibición de la actividad de ureasa
por el extracto APPE ------------------------------------------------------------------- 97
Figura 20. Inhibición de ureasa por APPE, HMW y LMW. (A) Gráficos
semi-log de la inhibición de la ureasa de H. pylori por los
polifenoles de cáscara de manzana. (B) Gráficos semi-log de
la inhibición de ureasa de Jack bean por los polifenoles de
cáscara de manzana ------------------------------------------------------------------ 100
Figura 21. Efectos de APPE y sus fracciones sobre la actividad de
ureasa detectados “en-gel” ---------------------------------------------------------- 102
Figura 22. Gráfico representativo del efecto de distintas concentraciones
del extracto APPE en g GAE mL-1 sobre la producción
intracelular de ROS en neutrófilos estimulados por PMA (200
nM) ---------------------------------------------------------------------------------------- 105
Figura 23. Efecto de los APPE sobre la producción intracelular y
extracelular de ROS en neutrófilos estimulados con PMA ------------------- 106
Figura 24. Efecto de los extractos de cáscara de manzana sobre la
producción extracelular de ROS en neutrófilos estimulados
con fMPL --------------------------------------------------------------------------------- 107
Figura 25. Efecto de la primación de neutrófilos con H. pylori sobre la
producción intracelular de ROS mediada por PMA. --------------------------- 108
Figura 26. Efecto de los extractos de cáscara de manzana sobre la
producción de quimioluminiscencia inducida por H. pylori no-
opsonizado en neutrófilos ------------------------------------------------------------ 109

13
Figura 27. Inhibición de la toxicidad (vacuolación) de VacA sobre células
HeLa -------------------------------------------------------------------------------------- 113
Figura 28. Estabilidad de los polifenoles de APPE en condiciones ácidas
que simulan el medio gástrico. ------------------------------------------------------ 115
Figura 29. APPE inhibe la adherencia de H. pylori a células HeLa ---------------------- 116
Figura 30. APPE impide la colonización de la mucosa gástrica de
ratones C57BL6/J por H. pylori ----------------------------------------------------- 119
Figura 31. (A), Índice de gastritis en ratones infectados con H. pylori y
tratados con APPE 150 mg/kg/día y 300 mg/kg/día. (B),
Apariencia macroscópica de los estómagos de ratones
después del tratamiento con APPE. (Barra = 1cm) ---------------------------- 120
Figura 32. APPE inhibe la formación MDA y la inflamación en el
estómago de ratones infectados con H. pylori ---------------------------------- 121
Figura 33. Efecto antioxidante de los polifenoles de APPE en neutrófilos
estimulados por H. pylori. ------------------------------------------------------------ 131
Figura 34. En células de epitelio gástrico, la regulación hacia arriba de la
expresión del supresor de la señalización por citoquinas
(SOCS)-2 y SOCS-3 por prebióticos y probióticos contrarresta
los efectos inflamatorios de H. pylori. --------------------------------------------- 137
Figura 35. Blancos moleculares de APPE en la primera etapa del
proceso de colonización de la infección por H. pylori. ------------------------ 140

14
ÍNDICE DE TABLAS
Página
Tabla 1. Contenido de polifenoles presentes en la cáscara de manzanas Granny
Smith y Red Delicious (142)............................................................................ 46
Tabla 2. Partidores para PCR en tiempo real ................................................................ 76
Tabla 3. Composición general del extracto de cáscara de manzana Granny Smith
enriquecido mediante Sepabeads SP-850 (APPE). ........................................ 77
Tabla 4. Composición polifenólica de cáscara fresca y el extracto APPE obtenido
de la manzana Granny Smith. ......................................................................... 85
Tabla 5. Datos obtenidos del análisis de APPE por HPLC-ESI/MS (polaridades
positiva y negativa). ........................................................................................ 87
Tabla 6. Efecto de APPE y sus fracciones sobre dos cepas de H. pylori. .................... 90
Tabla 7. Efecto de APPE sobre la viabilidad de H. pylori determinada mediante el
ensayo de microtitulación de la reducción de resazurina. ............................... 91
Tabla 8. IC 50 calculadas para cada periodo de preincubación del extracto APPE
con ureasa Jack bean. .................................................................................... 95
Tabla 9. Contenido total de polifenoles, Grado promedio de polimerización y
actividad inhibitoria de la ureasa de cáscara fresca de manzana, APPE y
sus fracciones de alto y bajo peso molecular. ................................................. 99
Tabla 10. Resumen de los resultados obtenidos en la evaluación del efecto
antioxidante de APPE en neutrófilos activados por H. pylori (ATCC
43504), fMLP and PMA. ................................................................................ 110

15
RESUMEN
Helicobacter pylori (H. pylori) infecta la mucosa gástrica de la mitad de la población
mundial, y es el único microorganismo conocido capaz de sobrevivir exitosamente en el
estómago humano. Muchos estudios han establecido a H. pylori como agente etiológico
del cáncer gástrico, linfoma del tejido linfoide asociado a la mucosa (MALT), y la úlcera
péptica. En efecto, en 1994, la Agencia Internacional para la Investigación sobre el
Cáncer (IARC) de la Organización Mundial de la Salud (OMS), definió a H. pylori como un
agente carcinógeno definitivo (tipo I) en el cáncer gástrico. Es importante destacar que en
Chile la prevalencia de la infección por H. pylori normalizada para la población mundial de
entre 20 y 60 años es aproximadamente de un 80%. Este alto nivel de infección por H.
pylori explicaría por qué Chile tiene la mayor tasa de mortalidad por cáncer gástrico en
América, y se ubica entre los cinco primeros países del mundo con respecto a la
mortalidad por cáncer gástrico. Sin embargo, la gran mayoría de las personas infectadas
con H. pylori permanece asintomática. El tratamiento de la infección por H. pylori requiere
el uso de una combinación de antibióticos con un inhibidor de la bomba de protones. La
literatura muestra un aumento en la aparición de cepas resistentes a los antibióticos, por
lo que hoy en día es importante buscar sustancias no-antibióticas con actividad anti-H.
pylori. Entre éstas, un número importante de diferentes productos del reino vegetal han
sido probados, incluyendo antimicrobianos pertenecientes a diferentes grupos de
fitoquímicos, tales como los aceites esenciales y los polifenoles. Los polifenoles son
componentes ubicuos de plantas medicinales y nutricionales. Estos compuestos, no sólo
actuarían como antioxidantes, sino que también podrían ejercer actividades
antimicrobianas en diferentes porciones del tracto digestivo. Numerosos antecedentes
sugieren que el consumo regular de alimentos o bebidas ricas en polifenoles, podría
ayudar a prevenir y atenuar el daño a la mucosa gástrica inducido por H. pylori. En
relación con esto último, las manzanas poseen muchas propiedades beneficiosas para la
salud humana asociadas con su alto contenido en compuestos fenólicos. Entre las
diversas variedades, las manzanas Granny Smith se exportan tanto como producto fresco
como para la producción de jugo concentrado. Aunque una gran parte de estas
exportaciones considera el uso de la fruta entera, en la última década, los productos de
manzana deshidratada han tenido un aumento significativo. Puesto que la deshidratación

16
requiere la eliminación de la cáscara, se llega a acumular importantes cantidades de ésta,
las que constituyen un residuo agroindustrial que genera problemas a las empresas con
las autoridades de la CONAMA. Algunas variedades de manzana podrían tener entre un
40 a 50% de los polifenoles totales del fruto en la cáscara. De hecho, la concentración de
polifenoles en la cáscara puede triplicar al observado en la pulpa. Los principales grupos
de polifenoles de la manzana incluyen los glucósidos de flavonoides, ésteres fenólicos de
ácidos carboxílicos, dihidrochalconas, catequinas y procianidinas. Los polifenoles de la
cáscara de manzana podrían actuar contra H. pylori no sólo como agentes
antimicrobianos, sino también neutralizando algunos de sus factores de virulencia.
También estos compuestos podrían reducir el daño inducido por las especies reactivas
del oxígeno y nitrógeno (ERON) generadas durante el proceso inflamatorio asociado a la
infección. En esta tesis se realizó un estudio farmacológico de los efectos in vitro e in vivo
de un extracto enriquecido en polifenoles de cáscara de manzana sobre H. pylori.
La preparación del extracto de cáscara de manzana (APPE) se llevó a cabo utilizando
adsorción de los polifenoles en la resina Sepabeads SP-850. De esta forma, los
polifenoles de los extractos acuosos de cáscara de manzana fueron adsorbidos en forma
exitosa. Los polifenoles fueron recuperados con etanol y concentrados en rotavapor,
llegando a un rendimiento de 3.50 ± 0.7 g por 1000 g de cáscara de manzana fresca. El
contenido de polifenoles totales fue de ~ 60%, con un perfil HPLC-DAD casi idéntico al
obtenido en la cáscara fresca de manzana Granny Smith. El análisis específico de los
compuestos polifenólicos de APPE obtenidos por RP-HPLC, permitieron identificar la
presencia de: (1) ácido clorogénico, (2) procianidina B1, (3) procianidina B2, (4) (-)-
epicatequina, (5) procianidina C1, ( 6) rutina, (7) hiperósido, (8) isoquercitrina (9),
quercetina-3-O-pentósido, (10), quercetina-3-O-pentósido, (11) quercitrina, (12) floretina-
O-xiloglucósido, y (13) floridzina. Para confirmar la identidad de los 13 principales
componentes se desarrolló un método de separación por HPLC-ESI/MS/MS, el cual
permitió obtener la huella digital de APPE caracterizada por el perfil de polifenoles.
Usando RP-HPLC, se logró una muy buena separación de los glucósidos quercetina y
algunas procianidinas desde los monómeros hasta los trímeros. Mediante RP-HPLC no
fue posible separar los oligómeros superiores a los trímeros, probablemente debido al
gran número de isómeros que interfieren con la separación. Por lo tanto, para el análisis

17
de oligómeros de prociandinas superiores de los trímeros se consideró a la NP-HPLC
como la técnica más adecuada. Así, con el uso de la NP-HPLC, las procianidinas desde
los monómeros a los undecámeros fueron separadas según su grado de polimerización,
siendo éste el método de elección para la separación de oligómeros. Con el objetivo de
fraccionar los polifenoles de APPE de acuerdo a su tamaño molecular, se utilizó una
combinación de cromatografía en columnas de Sephadex LH-20 y Toyopearl HW-40.
Como resultado de esta estrategia, se obtuvieron las fracciones de polifenoles de bajo
peso molecular (LMW) y alto peso molecular (HMW). La determinación del grado de
polimerización (mDP) de APPE, HMW y LMW se llevó a cabo mediante depolimerización
por floroglucinólisis y tiólisis con cisteamina y tolueno--tiol. La depolimerización generada
por dichos nucleófilos permitió obtener perfiles donde claramente se identificó como
principal unidad de extensión y de término a la epicatequina. Por medio del estudio de los
productos de despolimerización y los perfiles de NP-HPLC, fue posible confirmar que en
su mayoría LMW posee unidades monoméricas (mDP = 1). APPE posee un mDP = 2-4,
mientras que para HMW mDP = 8-10.
Se evaluó el efecto del extracto APPE y las fracciones LMW y HMW sobre la viabilidad de
H. pylori, la actividad de la ureasa y el estallido respiratorio de neutrófilos. Los resultados
sugieren, en base a los valores de sus IC50, que la actividad inhibitoria de la ureasa
mostrada por APPE estuvo asociada principalmente a la presencia de polifenoles de alto
peso molecular (HMW) y, en menor grado, con de la presencia de los componentes de
bajo peso molecular (LMW). Tal inhibición fue concentración-dependiente, tiempo-
independiente, reversible y competitiva. Por el contrario, el efecto de APPE tanto sobre la
viabilidad de H. pylori, como sobre la producción de las especies reactivas del oxígeno
(ERO) por parte de los neutrófilos, se asoció principalmente con los componentes de
LMW. APPE pudo inhibir el estallido respiratorio de neutrófilos inducido por H. pylori, PMA
y fMLP en forma concentración-dependiente. Este efecto se observó tanto en el interior
como en el exterior de los neutrófilos. Este resultado indicó que APPE podría atenuar el
daño a la mucosa gástrica asociado a las ERO producidas por los neutrófilos,
particularmente cuando H. pylori despliega sus mecanismos de evasión.
Se investigaron los efectos inhibitorios de APPE contra la infección por H. pylori, y la
vacuolación inducida por VacA. APPE disminuyó significativamente la vacuolación en

18
células HeLa con un valor de IC50 de 450 g GAE / mL. APPE también mostró un efecto
anti-adherente in vitro contra H. pylori. Se observó una inhibición significativa de la
adhesión de H. pylori con una reducción del 20-60% para concentraciones de entre 0.250
y 5 mg GAE / mL. En un modelo a corto plazo de infección (ratones C57BL6 / J), dos
niveles de dosis de APPE (150 y 300 mg / kg / día) mostraron un efecto inhibitorio sobre el
asentamiento de H. pylori. Después de 7 días de cultivo, no fue posible recuperar
bacterias con la morfología típica de H. pylori, tanto en los grupos tratados con APPE
como en los controles no infectados. Paralelamente, el análisis por PCR de tiempo real de
los estómagos de animales tratados con APPE, no mostró diferencias significativas con el
control no infectado. Estos resultados concordaron con la visible reducción de la carga de
H. pylori observada en las evaluaciones histológicas. Pese al corto plazo de interacción
huésped-patógeno y el bajo índice de gastritis observado en los animales infectados, los
polifenoles de APPE mostraron un efecto anti-inflamatorio sobre la gastritis asociada a la
presencia de H. pylori, reduciendo los niveles de malondialdehído.
En esta tesis se ha demostrado que APPE podría ejercer sus efectos anti-H. pylori
mediante múltiples mecanismos. Así, los polifenoles de alto peso molecular inhibieron la
actividad de la ureasa, el proceso de adhesión de las bacterias a la mucosa gástrica y la
actividad de la toxina VacA. Por otro lado, las propiedades anti-H. pylori y atrapadoras de
ERO fueron ligadas a la presencia de los polifenoles de bajo peso molecular. Como era
de esperar, el efecto antioxidante de APPE y sus propiedades anti-VacA, probablemente
cumplan un papel en la disminución del daño inflamatorio in vivo causados por H. pylori a
la mucosa gástrica de los ratones infectados. En resumen, en este estudio se observó que
APPE puede prevenir eficazmente las primeras etapas del proceso de colonización
gástrica por H. pylori y suprimir algunas de sus consecuencias patológicas más relevantes
como la inflamación.

19
ABSTRACT
Helicobacter pylori (H. pylori) infects the gastric mucosa of half of the world´s population,
and is the only microorganism known to successfully inhabit the human stomach. Many
studies have established H. pylori as an etiologic agent of gastric cancer, mucosa-
associated lymphoid tissue lymphoma (MALT), and peptic ulcer. In fact, in 1994, the
International Agency for Research on Cancer (IARC) of the World Health Organization
(WHO) defined H. pylori as a definite carcinogen in gastric cancer (type I). Importantly, in
Chile the prevalence of H. pylori infection standardised for world population for ages 20 to
60 is approximately 80%. This high level of H. pylori infection may explain why Chile has
the highest rate of gastric cancer mortality in the Americas, and is among the top five
countries in the world with regard to gastric cancer mortality. However, the vast majority of
people infected with H. pylori has no symptoms and will never develop problems. The
treatment of H. pylori infection requires the use of two or more antibiotics combined with a
proton pump inhibitor. The occurrence of strains resistant to antibiotics has been to
increasing, and it is nowadays important to search for new alternatives to antibiotics with
anti-H. pylori activity. Among these, a number of different products from vegetable origin
have been tested, including antimicrobials belonging to phytochemical such as essential
oils and polyphenols. Polyphenols are ubiquitous constituents of medicinal and nutritional
plants. Such compounds, not only would act as antioxidants, but also they could exert
antimicrobial activities in different portions of the digestive tract. Growing evidence
suggests that regular intake of foods or polyphenol-rich beverages might help to prevent
and attenuate H. pylori-induced damage to the gastric mucosa. Regarding the latter,
apples possess many beneficial properties for the human health related to their high
content in phenolic compounds. Among several varieties, Granny Smith apples are
exported both as fresh product and as juice concentrates. Although a great part of such
exports considers the whole fruit, during the past decade, exports of value added
dehydrated apple products have had a significant increase. Since dehydration requires
peel removal, important amounts of apple peel accumulate to be largely classified as an
agro-industrial waste. Some apple varieties concentrate in the peel from 40 to 50% of the
total fruit polyphenols. In fact, the amount of polyphenols in the apple peel could be up to

20
three times higher than that found in the pulp. The principal classes of whole apple
polyphenols include flavonoid glycosides, phenol carboxylic acid esters,
dihydrochalcones, catechins, and procyanidins.
Apple peel polyphenols might act against H. pylori exerting not only an antimicrobial
action but also neutralizing some of its virulence factors. These compounds could also
reduce the damage induced by the Reactive Oxygen and Nitrogen Species (RONS)
generated during the inflammatory process associated to the infection. In this thesis a
pharmacological study was undertaken considering the in vitro and in vivo effects of an
apple peel polyphenols-enriched extract upon H. pylori.
The preparation of apple peel extract (APPE) was carried out using the adsorption resin
Sepabeads SP-850. Thus, the polyphenols from the aqueous extracts of apple peel were
successfully adsorbed. Recovery of the polyphenols was carried out with ethanol and
concentrated through evaporative rotation with a yield of 3.50± 0.7g from 1000 g of fresh
apple peel. Total polyphenolic content was ~60 % with a HPLC-DAD profile remarkably
similar to that obtained in the fresh peel of Granny Smith apples. Specific analysis of
APPE polyphenolic compounds by RP-HPLC allowed to identify the presence of (1)
chlorogenic acid, (2) procyanidin B1, (3) procyanidin B2, (4) (−)-epicatechin, (5)
procyanidin C1, (6) rutin, (7) hyperoside, (8) isoquercitrin, (9) quercetin-3-O-pentoside,
(10) quercetin-3-O-pentoside, (11) quercitrin, (12) phloretin-O-xyloglucoside, and (13)
phloridzin. To confirm the identity of the 13 main components of APPE a fingerprint profile
method was developed using HPLC-ESI/MS/MS method. Using RP-HPLC, the extremely
good separation of quercetin glycosides and individual procyanidins from monomers to
trimers was achieved. RP-HPLC method was not able to separate oligomers higher than
trimers probably due to the higher number of isomers interfering with the separation.
Hence, NP-HPLC was better suited for analysis of procyandin oligomers higher than
trimers. Using NP-HPLC, procyanidin monomers through undecamers were separated by
the degree of polymerization and the separation of higher oligomers was preferable. With
the aim to fractionate APPE polyphenols according to their molecular size, a combination
of Sephadex LH-20 and Toyopearl HW-40s column chromatography was used. As result
of this strategy, low molecular weight (LMW) and high molecular weight (HMW)
polyphenolic fractions were obtained.

21
Determination of mDP of APPE, HMW and LMW was carried out by depolymerization via
phloroglucinolysis and thiolysis with cysteamine and toluene--thiol. Depolymerization with
nucleophile generated profiles where it is clearly seen that the main extension and terminal
unit was epicatechin. By means of the study of depolymerization products and NP-HPLC
profiles, it was possible to confirm that LMW possesses mostly monomeric units (mDP =
1). APPE possesses an mDP = 2-4, while for HMW mP = 8-10.
The effect of APPE, LMW y HMW against H. pylori viability, urease activity and neutrophil
respiratory burst was evaluated. On the basis of the IC50 ranking order, the results
suggested that the urease-inhibitory activity displayed by APPE is associated, primarily,
with the presence of HMW polyphenols and, to a lower degree, with the presence of
monomeric components (LMW). Such urease inhibition was concentration-dependent,
time-independent, reversible and competitive. On the contrary, inhibitory effect for both
viability and reactive oxygen species (ROS) production by stimulated neutrophils was
associated mainly with LMW components. Thus, APPE inhibited the respiratory burst of
neutrophils induced by H. pylori, PMA and fMLP in concentration-dependent form. This
effect was observed on both the interior and exterior of the neutrophil. This result suggests
that APPE have an attenuating effect on the damage to gastric mucosa caused by
neutrophil generated ROS and, particularly, when H. pylori display its evasion mechanism.
The inhibitory effects of APPE against H. pylori infection, and VacA-induced vacuolation
were investigated. APPE significantly prevented vacuolation in HeLa cell with an IC50
value of 450 g GAE/mL. APPE also displayed an in vitro anti-adhesive effect against H.
pylori. A significant inhibition was observed with a 20-60% reduction of H. pylori
attachment at concentrations between 0.250 and 5 mg GAE/mL. In a short-term infection
model (C57BL6/J mice), two levels of APPE doses (150 and 300 mg/kg/day) showed an
inhibitory effect on H. pylori attachment. After 7 days of culture, it was not possible to
recover bacteria with the typical H. pylori morphology neither in the groups treated with
APPE nor in the controls. Real-time PCR analysis of the stomachs from animals treated
with APPE did not show significant differences with non-infected control mice. These
results were in agreement with the visible reduction in H. pylori load observed in the
histology evaluations. Although the short-term pathogen-host interaction period and the

22
low gastritis score observed in the infected animals, APPE showed an anti-inflammatory
effect on the H. pylori-associated gastritis lowering malondialdehyde levels.
In this thesis it has been demonstrated that APPE could exert their anti-H. pylori effects by
multiple mechanism. So, high molecular weight polyphenols inhibit the urease activity, the
process of adherence of the bacteria to the gastric mucosa and the activity of VacA toxin.
On another side, low molecular weight polyphenols were involved in the anti-H. pylori and
ROS scavenging properties of APPE. As expected, the antioxidant and anti-VacA
properties of APPE probably played a role in diminishing the in vivo inflammatory damage
caused by H. pylori to the mice gastric mucosa. In summary, in this study it was observed
that APPE can effectively prevent the initial steps in the H. pylori colonization process and
suppress some of their most relevant pathological consequences such as mucosal
inflammation.

23
ABREVIATURAS
AhpC = Alquil hidroperóxido reductasa de H. pylori
AlpA = Proteína externa de membrana (OMP) de H. pylori
con función de adhesina.
AlpB = Proteína externa de membrana (OMP) de H. pylori
con función de adhesina.
APPE = Extracto de cáscara de manzana enriquecido en
polifenoles
ATP = Trifosfato de adenosina
BabA = Adhesina que se une al antígeno de Lewis B
Bax = Proteína X asociada al gen Bcl-2 (familia pro-
apoptótica).
Bcl-2 = Proteínas de la familia de las B-cell lymphoma 2
anti-apoptóticas.
BSA = Albúmina sérica de bovino
C57BL/6J = ratón de la cepa Black número 6
CagA = Citotoxina de 120-128 kDa asociada al gen cagA
CAT = Catalasa
c-Met = Receptor tirosin-kinasa del Factor de Crecimiento
de Hepatocitos.
DAF = Decay accelerating factor (CD55)
DMEM = Medio Dulbecco modificado de Eagle
DNA = Ácido desoxirribonucleico

24
DTT = Ditiotreitol
EDTA = Ácido etilendiaminotetraacético
EGFR = Receptor del Factor de crecimiento epidermal
ENA-78 = Proteína Epitelial Activadora de Neutrófilos 78
EPIYA = motivos con repeticiones Glu-Pro-Ile-Tyr-Ala
ERK = Kinasas reguladas por señales extracelulares
ERON = Especies reactivas del oxígeno y el nitrógeno
ESR = Electron Spin Resonance
fMLP =N-formil-L-metionil-L-leucil-L-fenilalanina
GPI = Glicofosfatidilinositol
GRO- = Growth regulated concogene-alpha; actualmente
CXCL-1 o ligando 1 de la quimioquina (motivo C-X-
C).
GSH = Glutatión
HMW = Fracción con polifenoles de alto peso molecular
HopZ = (Helicobacter outer membrane porins), proteína Z
de la membrana externa de H. pylori.
H. pylori = Helicobacter pylori
HpaA = Adhesina A de H. pylori con estructura tipo
lipoproteína.
HPTLC = High performance Thin Layer Chromatohgraphy
HRP = Peroxidasa de rábano
IFNγ = Interferón gamma

25
IL-2 = Interleuquina 2
IL-4 = Interlequina 4
IL-5 = Interleuquina 5
IL-8 = Interlequina 8
iNOS = Óxido Nítrico Sintasa inducible
JNK = c-Jun N-Terminal Kinasas
kDa = Kilo daltons
LDL = Lipoproteína de baja densidad
LMW = Fracción con polifenoles de bajo peso molecular
LPS = Lipopolisacárido
MALT = Linfoma del tejido linfoide asociado a las mucosas
MAP = Proteínas activadas por mitógenos
MDA = Malondialdehido
mg = miligramos
MIC = Concentración mínima inhibitoria
min = minuto
mL = mililitros
MMP-2 = Metaloproteinasa de matriz 2
MMP-7 = Metaloproteinasa de matriz 7
MMP-9 = Metaloproteinasa de matriz 9
MOI = Multiplicidad de infección
MPO = Mieloperoxidasa
NADPHox = NADPH oxidasa

26
NF-B = Factor Nuclear kappa B
nmol = nanomoles
Nod1 = (Nucleotide-binding oligomerization domain-
containing protein 1), es un receptor intracelular que
reconoce patrones moleculares asociados a
patógenos.
NP-HPLC-FLD = Cromatografía de alta eficiencia en fase normal
con detección por fluorescencia
p38 = Kinasa de la familia de las proteínas activadas por
mitógenos (MAP).
PAI = Isla de Patogenicidad
pb = Pares de base
PM = Peso molecular
PMA = 12-miristato-13-acetato de forbol
PPI = Inhibidor de la bomba de protones
rocF = Gen que codifica para la expresión de la Arginasa
de H. pylori
ROS = Especies reactivas del oxígeno (ERO)
RP-HPLC-DAD = Cromatografía de alta eficiencia en fase reversa
con detección por arreglo de diodos.
RP-HPLC-ESI/MS = Cromatografía de alta eficiencia en fase reversa
con detección por espectrometría de masas con
ionización por electro-spray.

27
RP-HPLC-FLD = Cromatografía de alta eficiencia en fase reversa
con detección por fluorescencia.
rpm = revoluciones por minuto
RPTP = Receptor de proteína tirosina fosfatasas
SD = Desviación estándar
SDS = Dodecil sulfato de sodio
SHP-2 = Tirosina fosfatasa con dominios citoplasmáticos
SH2
SOCS = Supresor de la señalización por citoquinas
SOD = Superóxido dismutasa
T4SS = Sistema de secreción tipo IV
TBARS = Sustancias reactivas al ácido tiobarbitúrico
TNF- = Factor de Necrosis Tumoral alfa
Trx1 = Tioredoxina 1
UBT = Urea Breath Test (Test de urea en el aliento)
UFC = Unidades formadoras de colonias
VacA = Vacuolating cytotoxin A
ZO-1 = Tight junction protein 1 (zona occludens 1)
g = microgramos
L = microlitros
mol = micromoles

28
1. INTRODUCCION
1.1. Epidemiología y principales manifestaciones clínicas de la infección por
Helicobacter pylori. Helicobacter pylori (H. pylori) fue identificado por primera vez por
Marshall y Warren en 1982 como una bacteria flagelada, microaerófila, Gram-negativa
de forma bacilar y espiralada. H. pylori se multiplica en forma óptima en el rango de
35-37 ºC, a pH neutro, es exigente y de lento desarrollo in vitro (3-5 días) y destaca
por su alta capacidad para colonizar el epitelio gástrico humano (1, 2). Es la única
bacteria capaz de habitar en forma persistente la mucosa gástrica humana. En los
países no desarrollados la infección es corrientemente adquirida en la niñez y en
ausencia de tratamiento puede persistir toda la vida. H. pylori es un microorganismo
cosmopolita, estimándose que más de la mitad de la población mundial esta infectada
(3). H. pylori puede ser aislado en personas tanto sintomáticas como asintomáticas. En
individuos infectados en forma crónica, el H. pylori constituiría el principal agente
etiológico de enfermedades como la gastritis crónica, la úlcera péptica, el
adenocarcinoma y el linfoma asociado a la mucosa gástrica (MALToma) (4). El curso
de la infección es muy variable y depende tanto de factores propios de la bacteria
como de la inmunidad del huésped. La infección no sólo altera en forma directa la
mucosa gástrica, sino que, además, modifica a nivel del antro gástrico la secreción de
polipéptidos gastrointestinales con acción hormonal (como gastrina), generando
alteraciones en la fisiología gástrica y la posterior ulceración de la mucosa. Por
ejemplo, la infección con H. pylori genera una hipergastrinemia conducente a una
mayor producción de HCl por parte de las células secretoras parietales del cuerpo
gástrico. Dichos efectos ocasionan un aumento en el número y masa de células
parietales, siendo la hiperclorhidria gástrica causa de una elevada carga de ácido en el
duodeno. Tan solo un 10-15% de los individuos infectados desarrolla úlcera péptica
que se localiza preferentemente a nivel antral. En los casos de gastritis crónica, tal
condición puede llevar al desarrollo de metaplasia gástrica como mecanismo protector
(5, 6). Por el contrario, Individuos con úlceras duodenales causadas por H. pylori
tienen una menor probabilidad de desarrollar posteriormente adenocarcinoma gástrico.
En cambio, aquellos pacientes infectados por H. pylori que exhiben una baja

29
producción de ácido tienen una mayor probabilidad de desarrollar gastritis del cuerpo
estomacal, lo que los predispone a desarrollar posteriormente una úlcera gástrica. En
base a los antecedentes epidemiológicos disponibles, en 1994 la Agencia
Internacional para la Investigación del Cáncer clasificó al H. pylori como un
carcinógeno humano clase I (7). El cáncer gástrico ocupa el segundo lugar entre las
causas de muerte por neoplasias a nivel mundial, y en Chile representa la primera
causa de muerte por tumores malignos, esta tendencia se ha estabilizado a partir de la
década de los 80 con una tasa de mortalidad cruda en alrededor de 20 por cada
100.000 habitantes (8).
1.2. Estrategias de supervivencia de H. pylori: Evasión de la respuesta inmune e
inflamatoria. En la interacción huésped-H. pylori, tanto la forma espiralar como los
flagelos del H. pylori, facilitan su movimiento hacia la mucosa gástrica. Posteriormente,
H. pylori se une a ciertos receptores de las células del epitelio gástrico (9-11), a través
de un número redundante de adhesinas del tipo BabA, AlpA, AlpB, HpaA y HopZ,
desencadenando la producción de interleuquinas, como IL-8 (pro-inflamatoria) y de
otras quimioquinas, como la ENA-78 (péptido activador de neutrófilos derivado de
células epiteliales) y el GRO- (oncogén regulado por crecimiento). Un factor
endógeno que, adicionalmente facilita la unión de H. pylori a las células epiteliales, es
la glicoproteína denominada DAF (Decay-accelerating Factor, 70 kDa), la cual en
presencia de H. pylori puede ser up-regulada (12-14). Tras su adhesión a la mucosa,
H. pylori secreta toxinas de naturaleza peptídica, como son la VacA y la CagA
(propias del H. pylori), ambas son capaces de actuar localmente sobre el epitelio
blanco. Si bien H. pylori logra asociarse al epitelio gástrico con bastante éxito, una vez
ahí debe lidiar con la primera línea de defensa constituida por la inmunidad innata.
Este sistema consta de componentes de barrera (epitelios, defensinas y linfocitos T
intraepiteliales), células efectoras circulantes (neutrófilos, macrófagos y células NK) y
diversas proteínas efectoras circulantes (complemento, colectina, factores de
coagulación, proteína C-reactiva y citoquinas como TNF, IL-1, 10, 12, 15). Aunque
algunos de estos componentes poseen receptores capaces de reconocer patrones
moleculares asociados a patógenos (PAMPs), que llevan a la destrucción del agente

30
extraño, H. pylori es capaz de vulnerarlos eficazmente. Por ejemplo, el TLR4 (un
receptor del tipo Toll), que normalmente reconoce a los LPS bacterianos para activar a
NF-B, genera frente al LPS de H. pylori una respuesta que si bien es proinflamatoria,
es notablemente más débil en comparación con la de los LPS de otras bacterias
entéricas (menor reconocimiento). Del mismo modo, en contraste con las flagelinas
secretadas por otros patógenos Gram negativos, como Salmonella o E. coli, que
activan en forma marcada la respuesta pro-inflamatoria mediada por los receptores
TLR5, la flagelina presente en H. pylori no es secretada y por ende se considera no
inflamatoria (15, 16). A su vez, H. pylori es capaz de interferir con algunas de las
respuestas inmunes locales; por ejemplo, inhibiendo la producción de óxido nítrico por
parte de macrófagos y “down-regulando” la expresión de receptores a quimioquinas en
neutrófilos infiltrantes. En forma adicional a la respuesta inmune innata, la infección
por H. pylori induce una respuesta inmune caracterizada por la infiltración de la
mucosa con neutrófilos, linfocitos y otras células inflamatorias, y por la producción
local y sistémica de anticuerpos y de otros mediadores de la respuesta celular. Si bien
las respuestas celulares y humorales son importantes en la patogénesis de H. pylori,
éstas, como tal, son inefectivas para erradicar la infección. Aún más, como resultado
de la interacción de linfocitos, neutrófilos, macrófagos y mastocitos, la respuesta
inmuno-celular es conducente a una amplificación de la inflamación inicial generada
en respuesta al patógeno. Tras ser atraídas a la lesión, las células infiltrantes liberan
mediadores como citoquinas, eicosanoides, y activadores de la cascada del
complemento, capaces de perpetuar la inflamación. Junto a dichos mediadores, las
células referidas liberan al medio una diversidad de especies reactivas del oxígeno y
nitrógeno (ERON), generando una condición de estrés oxidativo crónico en el epitelio
afectado. Las respuestas del huésped permiten la activación y diferenciación de
linfocitos Th0 (CD4+), los que según el patrón de citoquinas presentes en el medio y
de las condiciones inmunes del individuo, logran diferenciarse en Th1, mediando una
celular, o bien Th2 con una humoral. La celular es mediada a través de citoquinas tipo
Th1 –tales como IFN, IL-2 y TNF-, mientras que las citoquinas tipo Th2 –como la IL-4
e IL-5- promueven una respuesta de tipo humoral. Recientemente se ha comenzado a
comprender cómo –a través de mecanismos que permiten a H. pylori evadir la

31
respuesta inmune- la bacteria logra perpetuar la inflamación de la mucosa gástrica Un
ejemplo de esto último lo constituye la habilidad que tienen ciertas cepas de H. pylori
(60190; CagA+; VacA+) para inducir a través de la vía intrínseca (mitocondrial) y no la
extrínseca la apoptosis de linfocitos T (17).
1.3. Rol de la ureasa, VacA y CagA.
1.3.1. Ureasa: Entre los mecanismos más conocidos que dan cuenta de la persistencia y
patogenicidad del H. pylori en la mucosa gástrica destaca su alta actividad ureasa,
una enzima de 600 kDa, codificada por siete genes (ureA a ureG), que representa
cerca del 5% del total de sus proteínas celulares (18).
Figura 1. Rol de la ureasa en la colonización de la mucosa gástrica por H. pylori. Los números denotan etapas sucesivas en la colonización de la mucosa del huésped. La adhesión (1) es seguida por la formación y actividad de ureasas extracelulares (círculos negros) (2). Lo anterior lleva a agregación (3) y daño a las células de la mucosa (4). Adaptado de la referencia (19).
La importancia de la ureasa reside en que eleva el pH citoplasmático de H. pylori,
ayudando a tamponar el medio ácido circundante (periplasma), y favoreciendo la
permanencia de H. pylori en el inhóspito pH ácido del estómago (Figura 1). La
hidrólisis de urea actúa como tampón ya que genera amonio y carbamato, y este
último se descompone dando lugar a otra molécula de amonio y una de ácido

32
carbónico. Adicionalmente, el hidróxido de amonio (rápidamente generado a partir del
amonio) ayuda al H. pylori a movilizarse a través de mucus gástrico ya que disminuye
su viscosidad y facilita el movimiento flagelar.
1.3.2. Toxina VacA: Los genes que codifican por esta toxina, a diferencia de CagA, están
en todas las cepas de H. pylori. VacA puede ser transferida a las células epiteliales ya
sea por secreción o por contacto, siendo este último mecanismo una forma más
eficiente (20). La toxina puede unirse a varios receptores presentes en células
epiteliales (21-23) como son el receptor para proteína-tirosina-fosfatasa (RPTP) y ,
el receptor para factor de crecimiento epidermal (EGFR) (24), y una proteína asociada
a lipid rafts mediante una modificación tipo glicosilfosfatidilinositol (GPI) (25). La
importancia de la unión de VacA a la membrana plasmática de la célula eucariótica
reside en su capacidad para formar hexámeros capaces de inducir (en la superficie de
las células epiteliales del estómago) poros selectivos para aniones (Cl-) y pequeñas
moléculas no cargadas, como la urea. Esta propiedad de VacA le confiere actividad
urea-permeasa, la cual puede ser una ayuda importante para la supervivencia de la
bacteria en medio ácido (26). Luego de su unión a la membrana plasmática, VacA es
internada por endocitosis (observado in vitro). Los efectos que produce la
vacuolización dependen de su activación por ácido clorhídrico (27, 28). El efecto de
vacuolación in vivo parece ser no tan relevante (29). En altas concentraciones, VacA
también puede activar la vía intrínseca de la apoptosis generando poros en la
membrana mitocondrial con la consiguiente liberación de Citocromo-c (30, 31).
Recientemente, el interés por VacA se ha centrado en sus efectos inmunosupresores.
Por ejemplo, VacA puede interferir con la presentación de antígenos in vitro, lo cual
ocurre en compartimientos similares a los endosomas tardíos (32). Además, VacA
también puede inhibir específicamente la activación y proliferación de linfocitos T (33,
34). Esta observación ha llevado a especular que su efecto sería prevenir la
eliminación temprana de cepas de H. pylori CagA+. En los últimos años VacA se ha
convertido en uno de los candidatos a blanco molecular de algunos polifenoles (35,
36).

33
1.3.3. Citotoxina CagA: Ciertas cepas de H. pylori son productoras de la citotoxina CagA
(120-145 kDa), una proteína codificada dentro del denominado “isla de patogenicidad
cag” o cagPAI. Ésta última es responsable de la expresión del sistema de secreción
tipo IV (T4SS), que permite que el patógeno transloque en la mucosa del huésped
diversos componentes como 1) peptidoglicano, el cual es reconocido por Nod1,
llevando a la activación de NF-B y 2) CagA, que es el producto codificado en la
región génica terminal de la isla (37). En células epiteliales CagA es fosforilada en
residuos de tirosina (motivos EPIYA) por kinasas de la familia Src. CagA fosforilada
multimeriza, lo que le permite unir y activar a la fosfatasa SHP-2 (proteína-tirosina
fosfatasa con homología Src2), la cual lleva a una amplia desfosforilación de proteínas
del huésped y múltiples efectos celulares (38-41). CagA se une a la proteína zona
occludens-1 (ZO-1), afectando las uniones intercelulares y con ello causa pérdida de la
integridad epitelial (42). Esta toxina es capaz de transactivar el receptor de crecimiento
epidermal (EGFR) y otros similares, lo que lleva a señalización vía ERK y finalmente a
efectos pro-proliferativos mediados por AP-1 (activador de proteína-1) (43). Las vías
ERK, JNK (c-jun N-terminal kinasa) y MAP (proteína activada por mitógeno) son
también activadas por la forma fosforilada de CagA (44, 45). Algunas consecuencias
de la presencia de PAI y que son independientes de la traslocación de CagA, incluyen
la producción de moléculas proinflamatorias como IL-8 y MMP-7 (metaloproteinasa
derivada de la matriz epitelial con propiedades procarcinogénicas). Finalmente, CagA
se asocia con el receptor c-Met comprometiendo la motilidad de las células epiteliales.
La existencia de un sistema T4SS funcional y su relación con la activación del receptor
c-Met en la invasión de células del epitelio gástrico ha sido comprobada (46, 47). La
fosforilación de c-Met lleva a un aumento en las metaloproteinasas MMP-9 y MMP-2
que degradan componentes de matriz favoreciendo la generación de lesiones en el
epitelio gástrico (48). Estos efectos, observados en líneas celulares, pueden diferir de
la situación in vivo, donde es difícil predecir la preponderancia de cada uno. La
actividad de la citotoxina CagA está directamente relacionada con la presencia de
mayor proliferación del H. pylori en el epitelio (49), mayor inflamación de la mucosa
gástrica (50) y estas dos condiciones a su vez con la mayor probabilidad de que el
huésped desarrolle cáncer gástrico (51, 52). En definitiva la presencia de infección con

34
cepas que producen esta proteína se considera como un marcador de riesgo para la
aparición de cáncer gástrico (53). Se estima que intervenciones destinadas a disminuir
la densidad bacteriana en la mucosa contribuirán directamente a disminuir la
inflamación y con ello las probabilidades de que un individuo infectado crónicamente
(asintomático) se mantenga en el grupo de alto riesgo de desarrollar carcinogénesis
gástrica.
1.4. H. pylori y evasión del estrés oxidativo: Mención aparte merecen las enzimas con
que H. pylori está equipado para combatir las especies reactivas del oxígeno y de
nitrógeno liberadas por células del sistema inmune del huésped (54). Entre ellas, la
más abundante es la alquilhidroperóxido reductasa (AhpC), una enzima que dada su
alta expresión en H. pylori evita la acumulación de lipoperóxidos al interior del
patógeno. Cepas de H. pylori mutantes para AhpC presentan una aumentada
sensibilidad a diversas condiciones de estrés oxidativo (55), sobre-expresan la
proteína activadora de neutrófilos (56), y presentan una menor actividad catalasa (57).
Junto a lo anterior, H. pylori dispone de actividad superóxido dismutasa para la
remoción de radicales superóxido generados por células del sistema inmune (58), y es
capaz de aumentar la expresión de enzimas antioxidantes como la metionina sulfo-
reductasa, necesaria para corregir la modificación oxidativa de proteína en sus
residuos metionina (59). H. pylori induce la migración de los neutrófilos desde los
capilares a la lamina propria y a la zona glandular de la mucosa, especialmente en las
proximidades de los cuellos glandulares, donde se encuentran las células germinales.
Los neutrófilos son activados por factores citotóxicos del propio H. pylori (como N-
formilmetionil-leucil-fenilalanina) y por la IL-8 procedente de las células del epitelio tras
la adhesión bacteriana (60). Aún más relevante es que H. pylori posee la propiedad
única de alterar el reclutamiento de las diferentes subunidades de NADPH oxidasa en
los neutrófilos (61). Este mecanismo le permite desviar la producción de anión
superóxido hacia el exterior, evitando su acumulación en el fagosoma. De acuerdo a
dicho mecanismo, H. pylori actuaría concertadamente con los neutrófilos para producir
ulceración gástrica y además evadir su destrucción una vez fagocitado. Consistente
con lo anterior, se ha observado que la densidad de neutrófilos infiltrantes se

35
correlaciona directamente con la habilidad de H. pylori para causar daño a la mucosa
gástrica. Por otra parte, el daño provocado por las ERON sobre las células del epitelio
gástrico permite que H. pylori pueda acceder a una fuente adicional de nutrientes.
Respecto a la capacidad que tienen los macrófagos activados de la mucosa gástrica
para producir óxido nítrico, especie reactiva que en conjunto con otras persigue la
eliminación del patógeno, cabe destacar que H. pylori puede, limitar los efectos
bactericidas del NO., tanto in vitro como in vivo, gracias a que expresa (gen rocF) una
actividad arginasa capaz de “sifonar” la L-arginina lejos de la iNOS del huésped (62,
63). Un beneficio adicional asociado a la alta actividad arginasa de H. pylori es la
generación de urea (y ornitina) durante la hidrólisis de arginina. Si bien la arginasa
puede ser inactivada por diversas ERON (64), y desnaturalizada por altas
concentraciones de urea, H. pylori utiliza una tioredoxina (Trx1) como chaperona para
recuperar la actividad catalítica de la arginasa. Por tanto, se considera que el sistema
arginasa/Trx1 actuaría en forma concertada como sistema de adaptación y defensa
contra el estrés oxidativo resultante de la respuesta inmuno-celular.
1.5. Efectos de la respuesta inmuno-celular y del estrés oxidativo sobre el
huésped: Sumado al daño directo inducido por las ERON sobre el epitelio gástrico, los
eventos que llevan al proceso inflamatorio anteriormente descrito, conducen a
aumentar in vivo (en mucosa gástrica y a nivel sistémico) diversos parámetros de daño
oxidativo a lípidos (reflejado como aumento en TBARS e isoprostanos) (65, 66) y al
DNA (aumentada excreción de 8-hidroxi-guanidina) (67-69). La respuesta inmuno-
celular produce, además, ciertos eicosanoides biológicamente activos a través de un
mecanismo de peroxidación del ácido araquidónico catalizada por radicales del
oxígeno (65). Se ha planteado que el daño que en etapas tempranas (niñez y
adolescencia) genera H. pylori sobre el ADN estaría asociado con el desarrollo de
varios tipos de cáncer en la etapa adulta (70). Sin embargo, tal extrapolación no puede
ser directa debido a que el desarrollo del cáncer en respuesta a un agente
carcinogénico puede tomar entre 20 a 40 años. No obstante, se debe recordar que las
ERON son una de las causales más plausibles cuando se trata de explicar el
mecanismo involucrado en el desarrollo de cáncer y H. pylori (71, 72). Por otra parte,

36
se ha demostrado también un aumento de ácido hipocloroso (HOCl) en mucosa
gástrica de individuos infectados por H. pylori (73). El HOCl es un potente pro-oxidante
generado a partir de H2O2 (en presencia de CI-) por la enzima mieloperoxidasa (MPO),
liberada por los neutrófilos activados. El HOCl es capaz de reaccionar rápidamente
con el NH3 generado a nivel local por la enzima ureasa, para formar monocloroamina
(NH2Cl). Este último compuesto es altamente lipofílico y por tanto capaz de atravesar
membranas biológicas, lo que facilita la oxidación de macromoléculas al interior de las
células epiteliales. Se ha observado, además, que el NH2Cl es capaz de inducir la
infiltración y activación de neutrófilos en la mucosa gástrica de manera similar a lo
observado por H. pylori (73). A su vez, se ha encontrado que ciertos mecanismos
endógenos de defensa antioxidante como el glutatión (GSH) y ácido ascórbico, están
disminuidos en la mucosa de pacientes infectados por este agente (74-76). Tanto los
niveles de GSH como los de la enzima GSH-transferasa son significativamente
menores en la mucosa gástrica de pacientes infectados, que en pacientes H. pylori (-)
(77-79). Lo anterior conlleva una mayor producción local de ERON, las que al
acumularse pueden -vía regulación de ciertos genes, e induciendo daño oxidativo al
ADN- aumentar el riesgo de desarrollo de cáncer gástrico (67, 80). Cabe mencionar
que bajo condiciones desfavorables (por ejemplo, de estrés oxidativo), H. pylori es
transformado desde la forma bacilar original a la cocoide. Sólo esta última es capaz de
generar radicales hidroxilo, reconocidos como la especie de mayor reactividad hacia
sustratos biológicos (81). Se ha descrito que en mucosa gástrica de sujetos
asintomáticos, la erradicación de H. pylori mediante terapia tradicional (vide infra)
conduce a una normalización de los niveles de actividad de ciertas enzimas
antioxidantes (como iNOS y CAT) (82).
1.6. Terapias de erradicación de Helicobacter pylori: Se debe precisar que el
tratamiento de erradicación está reservado para aquellos individuos infectados y
sintomáticos con antecedentes de enfermedad ulceropéptica activa, linfoma MALT o
cáncer gástrico temprano. El tratamiento antibacteriano comprende la combinación de
dos o tres antibióticos, a los que se suma un inhibidor de la bomba de protones (PPI) y
bloqueadores H2 (83, 84). El consenso de Maastrich (85) sugiere que el tratamiento

37
de primera línea sea en base a una triterapia que incluya 2 antibióticos con diferentes
mecanismos de acción más un agente PPI (o un bloqueador H2). Se sabe que los PPI
poseen un efecto antimicrobiano directo sobre H. pylori, pudiendo inhibir en forma
irreversible a la ureasa y alterar su respiración. Como es de esperar, el empleo de
múltiples drogas aumenta el costo del tratamiento. En el esquema de tratamiento
habitual se usa una combinación de amoxicilina, claritromicina y/o metronidazol, más
un PPI, en un esquema de administración cada 12 horas durante 10-15 días. Este
esquema poseen una eficacia variable que va desde 50% hasta 85% de éxito (86).
Las tasas de reinfección con H. pylori son especialmente elevadas en países
Latinoamericanos (87), aunque no en Chile donde las cifras al año 1997 no superaron
el 4% (88). El incumplimiento (o cumplimiento sólo parcial) de la terapia prolongada (7-
15 días) la deserción de los pacientes causa alivio temporal de los síntomas clínicos
pero impide la erradicación del patógeno. Ambos factores son relevantes puesto que
aumentan la probabilidad de que aparezcan cepas resistentes de H. pylori. Se ha
estimado que entre un 10-50 % de los sujetos H. pylori (+) ya no responden a
metronidazol (89). Debido a que su uso lo ha convertido en uno de los antibióticos más
comunes para esta infección, es aún más preocupante la creciente resistencia
desarrollada a claritromicina (90, 91). En Chile, el costo asociado de la combinación de
antimicrobianos y PPIs, es otro factor que limita el cumplimiento del régimen
terapéutico. Esto último es particularmente importante dado que la prevalencia de la
infección por H. pylori es sustancialmente mayor en grupos de individuos de bajo nivel
socioeconómico (92, 93). Adicionalmente, grupos de individuos con hábitos higiénicos
deficientes ven incrementado su riesgo de re-infección (94, 95). Como se ilustra en la
Figura 2, debido a que la gran mayoría de los individuos colonizados por H. pylori,
infectados durante la infancia, permanece asintomáticos (85 - 90%), no es posible
identificar con exactitud cuáles son los factores determinantes que con los años
llevarían a algunos de éstos a desarrollar cuadros clínicos de mayor gravedad (96).
Sin embargo, los antecedentes epidemiológicos apuntan a que los individuos
tempranamente infectados poseen un alto riesgo (3 a 6 veces más que los no
infectados), de desarrollar una patología como el adenocarcinoma gástrico (97, 98).
Por lo tanto, aunque la erradicación no es recomendada por la OMS en individuos

38
asintomáticos, es muy deseable tratar de prevenir la infección o contar con agentes
que incorporados en los alimentos (como suplementos o nutraceúticos), puedan
contribuir a reducir el riesgo asociado a los elevados niveles de H. pylori en la mucosa
gástrica de dichos individuos (99). Como se ha discutido, el mayor conocimiento de los
factores de riesgo de infección, de aquellos que favorecen la colonización, virulencia, y
supervivencia de H. pylori, así como de aquellos que definen el daño al epitelio
resultante de la interacción H. pylori-respuesta del huésped, abre la posibilidad de
explorar nuevas estrategias de intervención.
Figura 2. Historia natural de la infección por H. pylori: relación entre patología y presentación clínica.
1.7. Efecto de los polifenoles sobre Helicobacter pylori: Dado que el estrés oxidativo
asociado a la respuesta inmune que sucede a la presencia de este patógeno
representa uno de los principales mecanismos de daño al epitelio gástrico, parece muy
relevante disponer de eficientes sistemas antioxidantes (tanto endógenos como
dietarios) en la zona colonizada por H. pylori. Sin embargo, las estrategias de

39
supervivencia con que cuenta H. pylori contra el estrés oxidativo, hacen surgir la
interrogante de cuán beneficiosa puede ser la administración o ingesta de
antioxidantes. Los antecedentes aquí revisados sugieren que estos compuestos
podrían tener un efecto protector, no sólo para el huésped (citoprotegiendo el epitelio),
sino también sobre H. pylori (contribuyendo a sus defensas antioxidantes).
Teóricamente, al aumentar la capacidad de H. pylori para “lidiar” con las ERON
generadas por el sistema inmune del huésped, se aumentaría también la posibilidad
de que la colonización del epitelio por H. pylori persista en el tiempo. No obstante,
estudios en los cuales se han testeado extractos de plantas ricos en compuestos
antioxidantes dan cuenta de efectos anti-H. pylori tanto in vitro como in vivo (100, 101).
La actividad anti-H. pylori de dichos preparados parece, sin embargo, estar asociada a
determinadas especies vegetales de uso médico y/o alimenticio, y por consiguiente
pudiera estar limitada a sólo algunas familias fitoquímicas. Ejemplos de esto lo
constituyen diferentes bayas (berries ricos en polifenoles), algunas crucíferas (brócoli
rico en glucosinolatos), propóleos y especies aromáticas (con aceites esenciales). Sin
embargo, dentro de los grupos de metabolitos secundarios más estudiados destacan
los polifenoles. La importancia de estos compuestos en salud humana ha sido
ampliamente revisada (102-105). No obstante, lo disperso de la información relativa
con su absorción, metabolismo, distribución y excreción ha llevado a muchos
investigadores a poner en duda los efectos de los polifenoles a nivel sistémico. De
esta forma, se ha sugerido que el principal sitio donde estos compuestos
podrían ejercer su acción antioxidante, sería el tracto gastrointestinal (106).
Muchos estudios en humanos destacan que sólo algunos polifenoles son absorbidos
en el intestino delgado. Cabe destacar que la mayor parte de los que logran llegar a
nivel sistémico lo hacen conjugados por glucuronidación, sulfo-conjugación y
metilación, y siempre en niveles plasmáticos extremadamente bajos. Sólo entre un 5 y
un 10% de los polifenoles absorbidos circulan en forma no-conjugada. Por lo tanto, es
muy difícil realizar estudios reales de farmacocinética, y las muestras de plasma u
orina habitualmente deben ser sometidas a hidrólisis con -glucoronidasa y/o sulfatasa
para liberar las agliconas. Tales estudios pseudos-farmacocinéticos han llevado a
concluir que los niveles de polifenoles en plasma son bajos, muy variables y con

40
máximos transitorios (Tmáx 1 - 2.5 horas). Es improbable que los conjugados
sobrepasen concentraciones de 10 M en total, o de 1 M en el caso de las agliconas.
La concentración de antioxidantes en plasma u orina representan valores
intracelulares pero por tratarse H. pylori de una bacteria extracelular es muy
importante conocer sus niveles en el espacio extracelular en la cercanía de la mucosa
que es el sitio de interacción entre la bacteria y el huésped en el fenómeno
fundamental de adhesividad mediado por una o mas adhesinas específicas. De
acuerdo a antecedentes de la literatura, los compuestos polifenólicos poseen
reconocida actividad bactericida, dada probablemente por mecanismos inespecíficos
que no necesariamente guardan relación con su actividad anti-H. pylori o antioxidante
(citoprotectora) sobre las células del epitelio gástrico (107). Una de las hipótesis que
se ha aceptado como paradigma, es que los polifenoles ejercen parte de su actividad
antimicrobiana mediante interacción no específica con componentes de la membrana
plasmática (108-110). Al respecto, se observó un efecto concentración-dependiente al
evaluar el daño provocado por algunos polifenoles en los modelos de integridad
liposomas y de alteraciones en la morfología de H. pylori (110). Este mecanismo es
relevante para taninos hidrolizables del tipo gálico (telimagrandina I y II), para los
cuales se determinó valores de concentración mínima inhibitoria (MIC) hasta 7 - 8
veces inferiores a los obtenidos con procianidinas del tipo B. Sin embargo, se debe
precisar que la mayoría de estos estudios categorizan la actividad de estos
compuestos sobre la base de aproximaciones in vitro y con antioxidantes en estado
puro no con mezclas de ellos tal como sucede con productos naturales, sin considerar
por ejemplo, que muchos polifenoles pueden alterar significativamente el pH del
periplasma (ácidos tánico, gálico y caféico entre otros). El hecho que sólo algunas de
las especies vegetales ricas en polifenoles muestren efectos anti-H. pylori genera la
interrogante de: en qué medida se vinculan mecanísticamente la actividad anti-H.
pylori con la actividad antioxidante de aquellos polifenoles que sí se muestran activos
sobre el patógeno. Al respecto, como se menciona más adelante, dependiendo del
grupo fitoquímico presente en la especie estudiada, existiría una asociación “no
necesariamente obligada” entre la actividad antioxidante de algunos polifenoles y su
actividad anti-microbiana (111, 112). Debido a que, entre los fitoquímicos bio-activos,

41
los polifenoles son los de mayor presencia en la dieta humana, se han publicado
varios estudios que relacionan su consumo con un efecto sobre H. pylori. La mayoría
de dichos estudios, exceptuando aquellos realizados con té verde y cranberry, han
contemplado la evaluación de extractos obtenidos a partir especies vegetales de uso
regional, no existentes en Chile. Considerando que la aplicación de las triterapias es
muy costosa, que presenta un alto nivel de reacciones adversas, y que junto a la
aparición de resistencia, su uso masivo sería controversial en poblaciones con
infección endémica por H. pylori, se hace crecientemente necesario investigar el
potencial que pudieran tener nutracéuticos ricos en antioxidantes naturales que
pudieran usarse en individuos colonizados asintomáticamente y con alto riesgo de
desarrollar cáncer gástrico. Una interesante opción considera la exploración de
productos naturales (con antecedentes de inocuidad de uso o consumo), que pudieran
ofrecer una favorable relación costo-efecto y aplicabilidad en sujetos infectados con
alto riesgo. Al respecto, se ha descrito que el uso combinado de extractos de jengibre,
propóleos y té verde con claritromicina y metronidazol posee un efecto sinérgico o
aditivo, inhibiendo la multiplicación y el desarrollo de resistencia en cepas de H. pylori
con diferente grado de sensibilidad (113, 114).
Tal como se describe más adelante (sección 1.7.1), una serie de estudios en los
cuales se ha intentado aislar e identificar los polifenoles presentes en plantas y
alimentos, sugieren que la actividad anti-H. pylori de los concentrados polifenólicos
estaría asociada, mayormente, a los antocianos, flavonoides, taninos hidrolizables
(galotaninos y elagitaninos) y taninos condensados (procianidinas) como los sub-
grupos más activos (35).
1.7.1. Antocianos, flavonoides, flavan-3-oles, procianidinas y taninos hidrolizables: Los
antocianos son aquellos polifenoles que confieren el color a las frutas, flores, cereales
y algunos tubérculos. Uno de los productos que más concentra antocianos es el jugo
de cranberry (Vaccinium macrocarpon). Este último ha sido estudiado, particularmente,
por sus propiedades antimicrobianas. Estudios clínicos han demostrado que el
consumo sostenido de jugo de cranberry previene las (re)infecciones del tracto urinario
en mujeres (115-119). Se ha sugerido que los polifenoles de cranberry actuarían

42
evitando la adhesión de Escherichia coli al uroepitelio (120, 121). Recientemente, las
propiedades antimicrobianas de cranberry han sido extendidas a H. pylori. Aunque los
antocianos contribuyen en forma importante a la capacidad antioxidante del cranberry,
se ha demostrado que la inhibición de la adhesión de H. pylori, a la mucosa gástrica
humana se debería, más bien, a sus constituyentes de alto peso molecular
(procianidinas) (122). Basado en la evidencia arrojada por este último estudio, Zhang
et al. (123) realizaron un ensayo clínico aleatorizado en una población adulta de Linqu
(China), placebo-controlado y doble ciego, donde se demostró que la administración
de 250 mL de jugo de cranberry (2 veces al día por 90 días) promueve la supresión de
H. pylori en un 14% de la población (test UBT negativo). Estudios in vitro han
demostrado que el jugo de cranberry asociado con otros extractos (Vitis vinifera;
arándano y orégano) inhibe la multiplicación in vitro de H. pylori en forma sinérgica,
correlacionándose la mayor presencia de polifenoles, con una mayor capacidad
antioxidante de la “compleja” mezcla ensayada (124, 125). Utilizando
bioprocesamiento en estado sólido (con los hongos Rhizopus oligosporus y Lentinus
edodes), los mismos investigadores lograron aumentar la extracción de los polifenoles
de la pomasa de cranberry, obteniendo un producto con mayor efecto anti-H. pylori
(126). En una aproximación similar, otros investigadores (111) encontraron una
actividad anti-H. pylori en extractos (ricos en quercetina y estructuras tipo bifenilos) de
desechos bioprocesados de la piña (Ananas cosmosus). Sin embargo, en este último
estudio no se logró correlacionar la actividad anti-H. pylori con la capacidad
antioxidante de los extractos empleados; estos últimos concentraban compuestos con
una mayor presencia de grupos fenólicos (asociados a las propiedades antioxidantes
de los polifenoles). En un reciente estudio donde se evaluó la actividad antimicrobiana
de 12 tipos de berries nórdicos sobre una serie de patógenos humanos, se encontró
que H. pylori y Bacillus cereus fueron las bacterias más sensibles. Interesantemente,
aunque se observó que los niveles de polifenoles disminuyeron durante el
almacenamiento en frío, la actividad antimicrobiana no fue modificada
significativamente (127). Por otra parte, en atención a su alta concentración de
polifenoles, se ha investigado también una posible acción anti-H. pylori en diferentes
tipos de té. Estudios in vitro realizados con infusiones al 5% de té verde (Lung Chen,

43
cv) demostraron la inhibición de la multiplicación de H. pylori (128), proponiendo que
dicha actividad residiría en la presencia de un flavan-3-ol conocido como galato de
epigalocatequina (EGCG). El mismo equipo de investigadores, realizó posteriormente
un estudio prospectivo entre dos grupos de individuos, infectados (n = 42) y no
infectados (n = 30) con H. pylori, comparando el efecto del consumo de té con la
presencia de H. pylori en biopsias gástricas. Se concluyó que habría una relación
inversa y significativa entre el consumo de té y la infección por H. pylori (129). A la luz
de los antecedentes dados por la etnomedicina, ocasionalmente avalados por estudios
experimentales de validación farmacológica, se han buscado diversos modelos para
explicar los mecanismos que posiblemente subyacerían a la acción anti-H. pylori de
los polifenoles presentes en los recursos etnomédicos y alimenticios anteriormente
referidos. Dentro de los mecanismos propuestos, se ha explorado la relación entre el
tipo de estructura de los polifenoles y su efecto como inhibidores de la actividad de
VacA (medida como actividad vacuolizante, flujo de urea transmembrana en células
HeLa) (35, 130). Así, el resveratrol, la morina, el ácido tánico y el piceatanol parecen
compartir características estructurales que sugieren la existencia de interacciones
específicas molécula-molécula entre ciertos polifenoles y VacA. Otros polifenoles, con
alta actividad antioxidante (incluso mayor con presencia de grupos fenólicos), como el
ácido elágico y la miricetina, se mostraron total o parcialmente inactivos hacia VacA,
respectivamente. Estos últimos compuestos están presentes en cantidades variables
en muchos alimentos de origen vegetal y se les puede encontrar en altas
concentraciones en el vino, cerveza, chocolate amargo y té verde. Recientemente, un
antecedente adicional respecto a la existencia o bien ausencia de relación entre
actividad antioxidante de los polifenoles y actividad anti-H. pylori (medida como
inhibición de la vacuolación y de la multiplicación de H. pylori) emergió del estudio de
13 diferentes tipos de estructuras flavonoideas, todas con propiedades antioxidantes
(112). Dentro de los compuestos evaluados, sólo quercetina y naringenina (dos
aglicones) inhibieron la vacuolación inducida por H. pylori en células HeLa. Como
evidencia adicional en favor de la asociación no obligada entre actividad antioxidante y
efecto anti-H. pylori, Shin et al., (112) demostraron también que antioxidantes de
estructuras tan disímiles como el ácido ascórbico, el glutatión, la epicatequina y el

44
Trolox® (análogo hidrosoluble del -tocoferol) tienen en común el ser inefectivos como
inhibidores de VacA. Entre estos últimos antioxidantes, se ha evaluado la acción
inhibitoria del crecimiento de H. pylori para epicatequina y vitamina C, siendo activa
sólo esta última tanto in vivo como in vitro por un mecanismo aún no esclarecido pero
que, en todo caso no estaría asociado al efecto del pH (131-133). La literatura informa
que otro hecho relevante es que algunos polifenoles inhiben la activación de
procaspasa-3 a caspasa-3 inducida por VacA, sin cambios en la expresión de las
proteínas Bax y Bcl-2 (proteínas antiapoptóticas). Por lo tanto, quercetina puede
proteger a las células gástricas de la apoptosis inhibiendo la acción vacuolante de la
toxina VacA de H. pylori. Si bien estas propiedades antioxidantes y citoprotectoras
para los compuestos señalados son reconocidas, llama la atención que en otros
modelos se observe una capacidad de inducir muerte y/o arresto de la proliferación en
líneas celulares inmortalizadas derivadas de tumores (134, 135). Esta
polifuncionalidad por parte de algunos compuestos fenólicos (EGCG por ejemplo), es
evidente si se considera que además, éstos podrían generar ambientes oxidativos
diferenciales que permitan proteger a las células del huésped de las ERON y por otro
lado promover la apoptosis de las células tumorales (136). Una hipótesis
recientemente planteada para explicar el por qué sólo ciertos antioxidantes
polifenólicos se muestran activos contra H. pylori, considera el potencial efecto
prooxidante que muestran ciertos polifenoles. La forma en que los polifenoles podrían
actuar como generadores de especies reactivas del oxígeno fue abordada por varios
investigadores (137-140), y utilizando como modelo de polifenol, catequinas de té
verde (cuya propiedad bactericida es conocida), los autores demostraron que a pH 7-8
(o superiores), tal tipo de estructura es capaz de generar cantidades significativas de
peróxido de hidrógeno. De acuerdo a los autores, la generación de peróxido de
hidrógeno podría explicar el efecto bactericida de los flavan-3-oles. La habilidad de las
catequinas para generar peróxido de hidrógeno sería favorecida por el arreglo de
grupos hidroxilo en este tipo de moléculas, lo que permitiría la disociación del H+ en
solución y un electrón en el fenol que reduce al oxígeno, generándose en
consecuencia anión superóxido. El anión superóxido posteriormente sufre reducción
por la catequina (ej. EGCG), lo que lleva a la formación de O2 2- ; adicionalmente, el

45
protón se combina con el superóxido generando H2O2. Este mecanismo no sólo
explicaría la generación de H2O2 producida por catequinas puras. También se ha
observado la producción de dicho oxidante en infusiones de té negro, té verde y té
Oolong en las cuales se pudo determinar concentraciones de H2O2 correspondientes a
1.5 x 10-4, 2,4 x 10-4 y 0.87 x 10-4 mol/L, respectivamente. Estos niveles de H2O2
serían suficientes para ejercer una acción bactericida en cepas Gram positivas y
negativas (138). Si bien es improbable que la generación de H2O2 ocurra en el medio
gástrico, podría especularse que el mayor pH del periplasma del H. pylori, dado por la
actividad ureasa, supondría un “ambiente” favorable a la formación de superóxido en
presencia de ciertos polifenoles. Por otra parte, cabe mencionar que cepas de
Lactobacilos entéricos, capaces de producir bacteriocinas, ácido láctico y acético, son
también capaces de generar concentraciones bactericidas de H2O2 (141).
Interesantemente, los autores observaron que el co-cultivo de H. pylori con
Lactobacillus entéricos produce de la inhibición de la multiplicación de H. pylori.
1.7.2. La cáscara de manzana como fuente de polifenoles: Frente al riesgo de
desarrollo de patologías gástricas crónicas de la infección por H. pylori, los polifenoles
podrían beneficiar potencialmente al huésped, actuando a través de diversos
mecanismos de acción. Por una parte, ejerciendo una acción directa sobre el agente
patógeno (reduciendo su viabilidad), y por la otra, actuando en forma indirecta sobre
su virulencia (inhibiendo las acciones mediadas por VacA). Adicionalmente, los
polifenoles podrían beneficiar al huésped, al actuar como antioxidantes, protegiendo a
las células del epitelio gástrico (acción citoprotectora) contra el daño oxidativo inducido
por ERON. Finalmente, la acción citoprotectora de los polifenoles podría, a su vez,
verse complementada por una acción anti-inflamatoria de estos compuestos sobre el
epitelio infectado (142). En la perspectiva de proveer polifenoles a la superfice del
estómago como una estrategia dirigida a controlar el riesgo de infección por H. pylori,
los berries, el chocolate, el té y el vino tinto destacan como posibles fuentes de dichos
compuestos. Sin embargo, más allá de las eventuales limitaciones de acceso, el
consumo de dichos productos, supone, a menudo, la co-ingesta obligada de calorías,
grasas, sustancias estimulantes y alcohol. Frente a esto, la obtención de extractos

46
cuya composición “concentre” los compuestos polifenólicos presentes en las fuentes
naturales de origen (materia prima), constituye una alternativa dirigida a “asegurar” en
un preparado (suplemento alimenticio o producto nutracéutico) la presencia de
compuestos polifenólicos en una dosificación y composición estandarizada. En la
medida en que existan antecedentes científicos suficientes, el uso de dichos extractos
pudiera responder a la intención de “asegurar” la ingesta regular de una dosis efectiva,
es decir, relevante a los efectos que se pretenden lograr tras el consumo de dichos
extractos. En general, la búsqueda de materias primas destinadas a la extracción de
estos polifenoles debe estar dirigida a fuentes abundantes, baratas, y para las cuales
se pueda aplicar procesos tecnológicamente viables y limpios. En ese sentido, en esta
propuesta se destaca a la cáscara de manzana como una materia prima que cumple
tales características. En efecto, dependiendo de la variedad, la cáscara de manzana
contiene entre 40 y 50% de los polifenoles presentes en la manzana entera, y su
concentración de polifenoles es a lo menos 3 veces superior a la presente en la pulpa
de dicho fruto. Chile es el principal exportador de manzanas del hemisferio sur,
seguido por Nueva Zelanda y Sudáfrica. Si bien la mayor parte de dichas
exportaciones implica el fruto entero, en los últimos años, ha aumentado
considerablemente la producción y exportación de manzana bajo la forma de producto
deshidratado.
Tabla 1. Contenido de polifenoles presentes en la cáscara de manzanas Granny Smith y Red Delicious (143).
Compuestos
Granny Smith (mg/kg base
fresca)
Red Delicious (mg/kg base
fresca)
antocianidinas 246 – 312 585 – 1037 floridzina 104 -159 12
procianidina B2 558 – 574 433 – 659 procianidina B1 127 – 172 173 – 241 procianidina B3 11 – 14 70 – 124 (-)-epicatequina 246 – 312 248 – 481
ácido clorogénico 113 – 157 6 – 60 rutina 136 – 504 390 – 414
glicósidos de quercetina
67 – 146 164 – 179

47
Actualmente, Chile es el principal exportador mundial de manzanas deshidratadas, con
el 30% del volumen total (144). Durante el procesamiento de la manzana para la
obtención de un producto deshidratado, el fruto es pelado, y la cáscara resultante es
esencialmente considerada un desecho industrial (145, 146). En relación a su
composición antioxidante, la Tabla 1 resume los principales polifenoles presentes en
la cáscara de manzana. Destaca de ésta, la alta presencia de epicatequina y sus
derivados diméricos (procianidinas del tipo B), que pueden llegar a representar
aproximadamente dos tercios de los polifenoles totales (147).
Dependiendo de la variedad, la cáscara de la manzana puede contener entre 2 y hasta
6 veces más compuestos fenólicos que la pulpa (148, 149).
Figura 3. Estructura de los principales polifenoles presentes en la manzana.
De este modo, se ha estimado que la contribución de cada grupo fenólico a la
actividad antioxidante total de la cáscara de manzana sería la siguiente: flavan-3-oles
Manzana
flavonoles
flavanoles
dihidrochalconas
hidroxicinamatos
Ácido clorogénico
Floridzina Floretina
Rutina Quercetina
Epicatequina
Procianidina B2

48
(71%), glicósidos de quercetina (12%), cianidina-3-galactósido (10%), ácidos fenólicos
derivados del ácido hidroxicinámico (7%) y dihidrochalconas (0.1%). Las principales
estructuras de estos compuestos se pueden apreciar en la Figura 3. En cambio en la
pulpa la actividad antioxidante ha sido atribuida a: flavan-3-oles (69%), ácidos
fenólicos derivados del ácido hidroxicinámico (31%) y glicósidos de quercetina y (150)
dihidrochalconas (0.1%) (151-157).
1.7.3. Actividad antioxidante y potencial actividad anti-H. pylori de polifenoles
presentes en la cáscara de manzana: Consistente con la alta concentración de
polifenoles en la cáscara de manzana, extractos de ésta son particularmente efectivos
como atrapadores de los radicales hidroxilo, superóxido y DPPH, en comparación con
los extractos elaborados en base a la pulpa (158, 159). En virtud de tal actividad los
extractos de cáscara de manzana son potentes inhibidores de la modificación
oxidativa de LDL humana in vitro (160), y efectivos en reducir la proliferación de
células cancerosas de hígado, colon y mamas (145, 146, 161, 162). Recientemente,
un estudio centró su atención en la evaluación de una posible actividad antimicrobiana
de extractos de cáscara de manzana (163). Si bien dicho estudio revela que los
extractos de cáscara de manzana exhiben cierta actividad anti-E. coli, anti- P.
aeruginosa y anti-S. aureus (entre otras), éste no incluyó al H. pylori en la evaluación.
Como resultado de ello, el potencial efecto que tendría un extracto de manzana sobre
H. pylori, así como el de sus componentes, sólo podría deducirse a partir de evidencia
indirecta. Tal como se describió anteriormente, muchos de los polifenoles que están
presentes en la cáscara de manzana (Tabla 1), en forma aislada han mostrado
diferentes grados de actividad anti-H. pylori. Cabe notar, sin embargo, que la evidencia
de que un determinado componente de un extracto es activo contra H.pylori no
necesariamente debe ser interpretada como evidencia de que el extracto, como tal,
compartirá también la actividad observada para el componente aislado. Si bien son
escasos los estudios dirigidos a evaluar la actividad anti-H. pylori de los compuestos
(aislados) presentes en la cáscara de manzana, para algunos de los polifenoles
presentes en ésta, se ha demostrado una actividad anti-ureasa y/o anti-VacA. Este es
el caso para los flavonoides (derivados de quercetina), las procianidinas (tipo-B), y la

49
floridzina que, si bien están presentes en diversos frutos, su concentración es
particularmente alta en la cáscara de manzana. En el caso de los flavonoides
mencionados, se ha descrito que dichos compuestos inhiben la vacuolación inducida
por H. pylori en células HeLa y que adicionalmente tendrían una moderada actividad
inhibitoria de la ureasa (112). En el caso de las procianidinas, utilizando extractos de
Vitis vinifera ricos en procianidinas tipo B y C, Lee et al. (164) observó que dichos
compuestos son particularmente activos inhibiendo la ureasa (ya a una concentración
de 0.1 mg/mL). En el caso de la floridzina, se ha encontrado que esta chalcona
promueve una acción anti-H. pylori inhibiendo la actividad formadora de poros de la
toxina VacA. La actividad anti-VacA (IC50= 273 M) de floridzina es, sin embargo,
extremadamente baja respecto a la del ácido tánico (IC50= 2,7 M), observado como el
polifenol más activo en dicho estudio. Respecto al ácido tánico, aunque ausente en
manzana, recientemente se ha establecido que éste (en asociación con galato de N-
propilo) es muy efectivo en inhibir la gastritis en un modelo de ratón infectado por H.
pylori o por VacA (130).
Adicionalmente, se ha comprobado que ciertos polifenoles de alto peso molecular
(procianidinas oligoméricas extraídas desde las brácteas del lúpulo), entre los cuales
algunos guardan una estrecha relación estructural con los polifenoles presentes en la
cáscara de manzana, son capaces de formar complejos con VacA in vitro (165, 166).
Yahiro observó que la interacción entre las proantocianidinas oligoméricas (con un
grado de polimerización promedio equivalente a 22 unidades de catequina) y la toxina
VacA constituye un potencial mecanismo de neutralización de virulencia de la bacteria.
Dichos compuestos fueron efectivos bloqueando la unión de VacA a sus receptores
RPTP y RPTP, inhibiendo la unión no específica de VacA a la membrana celular,
disminuyendo la vacuolación celular in vitro y atenuando en forma significativa la
gastritis inducida en ratones por la administración de VacA. De la misma forma, se
demostró que la administración de extractos de vino tinto y/o de té verde a ratones a
los cuales se les indujo gastritis por infección con H. pylori o por administración de la
toxina VacA, tienen un claro efecto gastroprotector, sugiriendo que la toxina sería un
potencial blanco molecular de los polifenoles presentes en los extractos utilizados
(167). Interesantemente, se ha observado que relativo a extractos de pulpa de

50
manzana, extractos de pulpa de membrillo tienen un efecto antiulcerogénico
claramente mayor sobre la injuria gástrica inducida por etanol y HCl (168). Los autores
sugieren que la diferencia en la efectividad anti-ulcerogénica de dichos extractos
residiría en la notablemente mayor concentración de procianidinas con un alto grado
de polimerización promedio en la pulpa del membrillo (DPm = 29) relativo a la pulpa
de manzana (DPm = 3). Respecto a esto último, cabe destacar que a diferencia de la
pulpa, la cáscara de manzana es particularmente concentrada en procianidinas de alto
grado de polimerización (DPm = 15 -190) (169). En relación a esto último, Saito et al.,
(170) habían reportado previamente que extractos de semilla de Vitis vinifera, así
como sus procianidinas (de alto PM), tienen un efecto antiulcerogénico; los autores
postularon que parte de dicho efecto se debería a la fuerte unión de estos compuestos
a proteínas presentes en la mucosa gástrica, permitiendo la formación de una barrera
protectora con potencial actividad antioxidante y antiinflamatoria local. En efecto,
recientemente se ha establecido que extractos de pulpa de manzana, ricos en
polifenoles, promueven una acción antioxidante y citoprotectora en cultivos primarios
de células de mucosa gástrica (MKN-28) sometidas a un sistema xantina-xantino
oxidasa, como generador de radicales anión superóxido (171). El efecto protector de
dichos extractos ha sido observado también en un modelo in vivo de injuria a la
mucosa gástrica inducida por la administración de indometacina (171).
Interesantemente, el daño oxidativo y las lesiones a la mucosa gástrica inducidas por
indometacina son exacerbadas por la infección con H. pylori (74). En otro estudio, se
ha establecido que extractos de cáscara de manzana inhiben, tanto in vitro como in
vivo, la activación de AP-1 y la transformación neoplásica (172). Tal como se señaló
anteriormente (sección 2.2.2), la activación del complejo AP-1 es uno de los eventos
claves en la promoción de tumores mediada por la citotoxina CagA de H. pylori. Al
respecto, se ha sugerido que los polifenoles de la cáscara de manzana podrían inhibir
la activación de AP-1 interfiriendo con la señalización vía MAP kinasas, ERK y JNK.
Mediante experimentos de ESR, Ding et al. (172) confirmaron el efecto estabilizador
de los extractos de cáscara de manzana sobre los radicales OH. y O2.-. Esto último
sería particularmente relevante dado que, ERKs, JNKs y p38 son moléculas activadas
en respuesta a estímulos oxidantes. Finalmente, en estudios adicionales conducidos

51
en ratones, los mismos autores demostraron que los extractos inhiben también la
inducción de tumores por TPA (12-O-tetradecanoil-forbol-13-acetato). En virtud de
aquellos antecedentes que indican que algunos polifenoles, que se encuentran
también en la cáscara de manzana, muestran una interesante actividad anti-H. pylori
(ya sea promoviendo una acción antimicrobiana directa, o bien indirectamente
afectando la virulencia y/o sobrevivencia de la bacteria), en la Figura 4 se
esquematiza los potenciales blancos moleculares o sitios de acción que los extractos
de cáscara de manzanas ricos en compuestos polifenólicos, pudieran tener como
agentes destinados a complementar el manejo de la infección por H. pylori y/o de sus
consecuencias. En observaciones preliminares realizadas en nuestro laboratorio, se
detectó una alta presencia de polifenoles en la cáscara de manzana, con potencial
anti-H. pylori in vitro. La decisión de estudiar los efectos de extractos de cáscara de
manzana sobre H. pylori responde, adicionalmente, a la oportunidad que supone para
Chile la vasta disponibilidad agro-industrial de cáscara de manzana. Curiosamente, a
pesar de concentrar entre un 40 y 50% de los polifenoles presentes en el fruto entero,
la cáscara de manzana es esencialmente “desaprovechada” en nuestro país ya que, a
nivel de consumidor es descartada, y a nivel agro-industrial considerada un “desecho
mayor” del proceso de deshidratado de manzanas para su exportación (146). En la
actualidad, sustentado en algunos de los antecedentes científicos anteriormente
mencionados, es posible encontrar varios productos desarrollados en base a
polifenoles de la manzana (http//www.applepolyphenols.com). En el marco de esta
propuesta, nos planteamos elaborar extractos de cáscara de manzana cuya
composición sea relevante al propósito de evaluar en forma sistemática el potencial
que sus polifenoles tendrían sobre la viabilidad y virulencia de H. pylori in vitro, y sobre
las consecuencias que conlleva una infección con H. pylori in vivo. Se espera que los
resultados de las investigaciones planteadas más adelante contribuyan a disminuir la
densidad de colonización y que por tanto impacten positivamente en el curso crónico
de la infección gástrica por H. pylori.

52
Figura 4. Potenciales sitios de acción de los polifenoles de la cáscara de manzana como agentes citoprotectores y anti-H. pylori. Los números representan los efectos: (1), anti-H. pylori (efectos sobre su viabilidad y adherencia a la mucosa); (2), cito-protector de la mucosa; (3), anti-VacA, (4), anti-ureasa; (5), antioxidante (producción de ROS por neutrófilos).

53
2. HIPÓTESIS DE TRABAJO
Fundamentación:
La cáscara de manzana, considerada actualmente un desecho agro-industrial, es
una muy buena fuente de polifenoles biológicamente activos. Como tal, constituye un
recurso potencialmente aprovechable para la elaboración de extractos ricos en
polifenoles destinados a ser utilizados como productos nutracéuticos e ingredientes
funcionales. Entre diversos polifenoles, la cáscara de la manzana concentra
procianidinas, flavonoides (derivados de quercetina), chalconas (floridzina) y ácidos
fenólicos (clorogénico). En esta propuesta se evaluó tanto in vitro como in vivo el
efecto de un extracto polifenólico de cáscara de manzana (APPE) sobre el patógeno
H. pylori.
Las hipótesis planteadas en este proyecto son:
1. El extracto de cáscara de manzana, rico en polifenoles con actividad antioxidante,
posee una actividad deletérea contra cepas patógenas de H. pylori. Planteamos
que tal actividad está asociada a una acción directa (bactericida o bacterioestática)
de los polifenoles sobre H. pylori.
2. Se plantea que los extractos a estudiar promueven (en un modelo celular) una
acción protectora contra el daño inducido por Helicobacter pylori, a través de la
inhibición de los mecanismos de virulencia de la bacteria lo que promueve un
efecto protector contra la injuria a la mucosa provocada (en un modelo animal) por
las ERON generadas durante el proceso inflamatorio que acompaña a la infección.
Postulamos que esta última acción de los extractos esta asociada a las
propiedades antioxidantes de sus polifenoles.

54
3. OBJETIVO GENERAL
Evaluar el efecto de extractos de cáscara de manzana ricos en polifenoles sobre
la viabilidad de Helicobacter pylori, sobre el daño provocado in vitro por la
bacteria en cultivos de células HeLa e in vivo en la mucosa gástrica de ratones
C57BL/6J infectados.
4. OBJETIVOS ESPECÍFICOS
Específicos:
Preparar extractos secos de cáscara de manzana, mediante procesos extractivos y
de concentración de los polifenoles presentes en la materia prima fresca.
Caracterizar dichos extractos en términos de su composición química, mediante
técnicas espectroscópicas y cromatográficas.
Evaluar los extractos obtenidos en términos de sus propiedades bactericidas o
bacteriostáticas sobre cepas aisladas y de referencia de H. pylori., así como el
efecto inhibidor de los extractos sobre la actividad ureasa de H. pylori.
Evaluar el efecto inhibitorio in vitro de extractos de cáscara de manzana sobre la
inducción del estallido respiratorio (producción de especies reactivas del oxígeno)
en neutrófilos activados por H. pylori, no opsonizado.
Evaluar el efecto inhibitorio de los extractos de cáscara de manzana sobre el daño
inducido por la toxina VacA en células HeLa (vacuolización y viabilidad). Utilizando
el mismo modelo celular, evaluar el potencial del extracto sobre la adherencia de H.
pylori.
Evaluar el efecto protector y erradicador de H. pylori derivado de la administración
oral del extracto de cáscara de manzana sobre daño a la mucosa gástrica de
ratones C57BL/6J. Evaluar el efecto de dos dosis del extracto sobre marcadores
moleculares de infección (PCR en tiempo real), indicadores de daño
histopatológico (índice de gastritis), y parámetros bioquímicos de inflamación y de
daño oxidativo (niveles de MDA).

55
5. MATERIALES Y MÉTODOS
5.1. Reactivos y solventes. Ácido gálico, ácido clorogénico, ácido cafeíco, (+)-
catequina, (-)-epicatequina, floridzina, quercetina, quercetina 3-O-rutinósido,
procianidina B1 y B2, y el reactivo de Folin-Ciocalteu fueron adquiridos de Sigma-
Aldrich (St. Louis, MO, USA). Los glicósidos de flavonoides (quercetina 3-O-
galactósido, quercetina 3-O-glucósido y quercetina 3-O-ramnósido fueron
adquiridos de Roth (Karlsruhe, Alemania). Todos los solventes fueron grado
HPLC adquiridos de Merck (Darmstadt, Alemania).
5.2. Preparación del extracto rico en polifenoles de cáscara de manzana (APPE).
Las cáscaras frescas de manzana Granny Smith madura fueron rápidamente
congeladas a -20 ºC hasta el momento de su empleo. Para la extracción a escala
del laboratorio, a 400 g se le agregaron 1200 mL de agua a 80 ºC con agitación
manual. Una vez alcanzados los 60-65 ºC, la mezcla fue homogenizada con
Ultraturrax a 20.000 rpm. Luego de la etapa de homogenización, el extracto
acuoso se obtuvo por maceración durante 30 min con posterior separación de los
sólidos insolubles a través de una malla de acero inoxidable (tamiz de 50 m). El
proceso se repitió dos veces más con igual cantidad de agua y los tres líquidos
resultantes fueron reunidos. Los extractos fueron aplicados sobre una columna
empacada con 400 g de la resina granular Sepabeads SP-850, previamente
acondicionada con agua. Luego, la columna fue lavada con 5 volúmenes de
agua, hasta eliminación completa de, sales, aminoácidos, proteínas y los
azúcares simples (glucosa, fructosa y sacarosa) y poliméricos (pectinas).
Finalmente el material polifenólico fue recuperado por elución con 200 - 500 mL
de etanol absoluto. El eluato hidro-alcohólico fue concentrado a presión reducida
(< 40°C) para obtener un polvo seco de aspecto amarillo-parduzco. El polvo
(APPE) fue almacenado en desecador bajo vacío durante 24-48 h para luego ser
envasado en un recipiente ámbar y guardado a -80 °C hasta el momento de su
empleo.

56
5.3. Determinación cuantitativa de azúcares por HPTLC. La concentración de
azúcares se determinó de acuerdo a la metodología de Zúñiga-Feest et al. (173).
Las muestras fueron aplicadas a placas de HPTLC de silicagel F-254 de 10 x 10
cm pretratadas con una solución de fosfato 0.1M en metanol 50% y activadas a
100 °C por 30 min. Para la aplicación se utilizó un sistema automatizado de
siembra ATS IV (Camag, Suiza), en bandas de 3 mm. Los patrones de glucosa,
sacarosa y fructosa fueron disueltos adecuadamente en etanol 70%. Las placas
fueron sometidas a desarrollo múltiple (4 veces) con acetonitrilo/agua (85:15 v/v)
hasta 5 cm con etapas de secado intermedio de 10 min a 40 °C, utilizando la
cámara automatizada AMD (Camag, Suiza). Las placas fueron posteriormente
reveladas con el reactivo difenidramina/anilina en ácido fosfórico a 100 °C por 5
min. La evaluación densitométrica se llevó a cabo en los sistemas reprostar 2.0 y
Scanner III (Camag, Suiza), utilizando el sofware Cats 3.0.
5.4. Determinación del perfil de compuestos fenólicos en el extracto de cáscara
de manzana mediante RP-HPLC-DAD-FLD. Para el análisis de los polifenoles
de cáscara de manzana se utilizó dos sistemas cromatográficos. El primero de
ellos, que se empleó para el análisis específico de las procianidinas y flavan-3-
oles por RP-HPLC-FLD, consistió de un sistema Agilent 1100 HPLC equipado
con autosampler, bomba HPLC cuaternaria y detector de fluorescencia. La
columna usada fue una Kromasil KR 100-5C18 (250 x 4.6 mm I.D., 5 m tamaño
de partícula). La fase móvil consistió en una mezcla binaria de (A) ácido
trifluoracético 0.1 % en agua y (B) ácido trifluoracético 0.1 % en acetonitrilo. Las
separaciones se efectuaron aplicando un gradiente lineal a 25 ºC con un flujo de
0.8 mL/min de la siguiente manera: 0-20 min 8-20% B; 20-40 min, 20-50 % de B;
seguido de un tiempo de re-equilibrio de 5 minutos. La detección por
fluorescencia se realizó con una excitación de 280 nm y una emisión de 320 nm.
Para el análisis cuantitativo de flavonoides se utilizó un segundo sistema con
detección por arreglo de diodos RP-HPLC-DAD, Lachrom Merck. Las condiciones
cromatográficas utilizadas fueron las mismas optimizadas para el sistema RP-
HPLC-FLD.

57
5.5. Determinación del Grado de Polimerización de las procianidinas presentes
en el extracto de cáscara de manzana.
5.5.1. HPLC en Fase Normal (NP-HPLC): El análisis del grado de polimerización de
las procianidinas del extracto de cáscara de manzana se llevó a cabo mediante
HPLC en fase normal con detección por fluorescencia (NP-HPLC-FLD),
mediante procedimientos previamente publicados (174, 175). Las separaciones
fueron realizadas utilizando un sistema Agilent 1100 HPLC equipado con
autosampler, bomba HPLC cuaternaria y detector de fluorescencia. La columna
usada fue una Lichrospher 100 Diol (250 x 4.0 mm I.D., 5 m tamaño de
partícula). La fase móvil consistió en una mezcla binaria de (A) acetonitrilo:
ácido acético (98:2, v/v) y (B) metanol: agua: ácido acético (95: 3: 2, v/v). Las
separaciones se efectuaron aplicando un gradiente lineal a 25 ºC con un flujo
de 0.6 mL/min de la siguiente manera: 0-35 min 0-40% B; 35-50 min, 40% B
isocrático; 50-60 min 40-0% B, seguido de un tiempo de re-equilibrio de 5
minutos. La detección por fluorescencia se realizó con una excitación de 230
nm y una emisión de 320 nm.
5.5.2. RP-HPLC-FLD de los productos de tiólisis con tolueno-tiol. Para la
estimación del grado de polimerización del extracto crudo se realizó la
depolimerización en presencia de tolueno-tiol, utilizando metodologías
previamente publicadas (176-178). El análisis de los productos de tiólisis se
realizó mediante RP-HPLC-FLD (Agilent 1100 Series). La fase móvil A fue ácido
trifluoracético 0.1 % (v/v) y B ácido trifluoracético 0.08 % en acetonitrilo (4:1,
v/v). El programa de gradiente fue: solvente B 0-50% en 40 min, seguido de una
fase de lavado y re-acondicionamiento de la columna. El flujo se mantuvo
constante a 1.0 mL/min. La columna utilizada fue Kromasil (250 x 4.6 mm; 5
m; Eka Chemicals AB, Bohus, Suiza). La detección se realizó por
fluorescencia utilizando una longitud de de onda de excitación de 280 nm y de
emisión de 310 nm. La tiólisis fue realizada en triplicado para cada muestra. La

58
determinación cuantitativa de (-)-epicatequina, (+)-catequina y los productos de
tiólisis se realizó por estandarización externa.
5.5.3. RP-HPLC-FLD de los productos de tiólisis con cisteamina. Para la
degradación con cisteamina, la muestra (10 mg) fue disuelta en metanol (1 mL).
Una alícuota de la muestra (200 L) se agrega a la mezcla de tiólisis
consistente en clorhidrato de cisteamina (50 mg) y HCl al 37 % (20 L) disueltos
en metanol (930 L). La mezcla (400 L) se mantiene a 65 °C por 15 min.
Posteriormente, la reacción se detiene con una solución de TFA al 0.1 % (1.2
mL). Para la separación de los adúctos con cisteamina se utilizó una columna
Kromasil (250 x 4.6mm; 5 m; Eka Chemicals AB, Bohus, Suecia), con un flujo
de 1.0 mL/min. Los solventes utilizados fueron: ácido acético 2.5 % (solvente
A) y acetonitrilo/ácido acético al 2.5 % (solvente B) en proporción 80:20 v/v. El
programa de gradiente fue: 5 – 50 % de solvente B en 35 min, 50 - 100% de B
en 40 min, seguido de una etapa de lavado y re-acondicionamiento de la
columna. La 4-(2-aminoetiltio)-epicatequina y 4-(2-aminoetiltio)-catequina
fueron obtenidas de acuerdo al procedimiento publicado previamente (177,
179). Los gráficos de calibración para (-)-epicatequina y (+)-catequina fueron
lineales en el rango de 2 – 20 g/mL y 10 – 60 g/mL respectivamente; los
coeficientes de correlación de las curvas fueron 0,9999 y 0,9997
respectivamente. Previo a la tiólisis, se determina el aporte de (-)-epicatequina y
(+)-catequina.
5.5.4. RP-HPLC-FLD de los productos de floroglucinólisis. La depolimerización en
presencia de floroglucinol (floroglucinólisis) se utilizó como procedimiento de
elección para la estimación del grado de polimerización medio (DPm) y el
contenido de procianidinas de acuerdo al procedimiento de Karonen (180).
Brevemente, 10 mg de APPE fueron disueltos en 2 mL de una solución de HCl
0.1 M en metanol que contiene 50 mg/mL de floroglucinol y 10 mg/mL de ácido
ascórbico. La mezcla de reacción fue incubada a 50°C por 20 min. La reacción

59
se detuvo agregando 10 mL de una solución acuosa de acetato de sodio 40
mM. Para el análisis de la cáscara fresca de manzana, 100mg fueron extraídos
con 1.0 mL de acetona al 70% (v/v) conteniendo 10 mg/mL de ácido ascórbico.
Las muestras fueron agitadas en un vortex por 1minuto y luego sonicadas por
20 min a temperatura ambiente. El extracto se obtuvo por centrifugación a
14.000 x g por 10 min. Una alícuota de 100 L de dicho extracto fue evaporada
a sequedad bajo corriente de nitrógeno previo a la floroglucinólisis. Los
productos de floroglucinólisis fueron analizados mediante RP-HPLC tal como se
describió anteriormente. La determinación cuantitativa de (-)-epicatequina, (+)-
catequina y los productos de degradación se llevó a cabo por estandarización
externa. Las curvas de calibración para (-)-epicatequina y (+)-catequina
mostraron un rango lineal desde 2 – 20 g/mL (r2 = 0.9999) y 10 – 60 g/mL (r2
= 0.9997), respectivamente. El contenido de procianidinas fue calculado
sumando la masa de todas las subunidades (excluyendo la porción de
floroglucinol de los adúctos). Para calcular el grado de polimerización DPm, la
suma de las subunidades de extensión fue dividida por la suma de las
subunidades terminales. El medio sin degradar fue utilizado para cuantificar el
contenido normal de (-)-epicatequina y (+)-catequina en los extractos y
fracciones.
5.6. Análisis del perfil de compuestos fenólicos en el extracto de cáscara de
manzana mediante RP-HPLC-ESI-MS/MS. La muestra (APPE) se examinó en
un sistema LC-MS que consistió de una bomba HPLC Agilent 1100 (Agilent
Technologies, Inc.) acoplada a un espectrómetro de masas de tipo electrospray-
trampa iónica Esquire 4000 ESI-IT (Bruker Daltonik GMBH, Alemania). Para el
control del sistema HPLC se utilizó el programa ChemStattion for LC 3D Rev. A.
10.02 (Agilent Technologies, Inc., CA-USA) y para el control del espectrómetro se
utilizó el programa esquireControl %.2 (Bruker Daltonik GMBH, Alemania). Para
la separación HPLC se utilizó una columna de 300 x 4 mm, 5 m (Supelcosil LC-
18, Supelco-USA), a la salida de la cual se conectó un split que dividió el flujo
hacia el detector UV y al espectrómetro de masas. Para la separación, el

60
volumen inyectado de muestra fue de 20 L y la corrida se realizó a temperatura
ambiente utilizando el siguiente programa de gradiente: solvente (A) ácido
fórmico 0,1 % en agua (v/v) y solvente (B) ácido fórmico 0,1 % (v/v) en
acetonitrilo. Las separaciones se efectuaron aplicando un gradiente lineal con un
flujo de 1 mL/min de la siguiente manera: 0-25 min 10-30% B; 25-30 min, 30-75
% de B; seguido de un tiempo de re-equilibrio de 5 min (hasta 10% de B). El
proceso de ionización se realizó mediante electrospray utilizando nitrógeno a una
temperatura de nebulización de 325 °C, presión de nebulización de 30 psi y flujo
de gas nebulizador de 10 L/min. Las condiciones espectrométricas para polaridad
positiva fueron: voltaje de ionización, 3000 V; offset de placa terminal (end plate),
-500 V; skimmer 1, 30.0 V; skimmer 2, 6.0 V; offset de salida de capilar, 90.0 V;
salida de capilar, 90.0 V; diferencia de octupolos, 2.40 V; trap drive, 55.0; voltaje
de lente 1, -5.0 V; voltaje de lente 2, -60.0 V. Y las condiciones de operación para
polaridad negativa fueron: voltaje de ionización, 3000 V; offset de placa terminal
(end plate), -500 V; skimmer 1, -30.0 V; skimmer 2, -6.0 V; offset de salida de
capilar, -60.0 V; salida de capilar, -90.0 V; diferencia de octupolos, -2.40 V; trap
drive, 55.0; voltaje de lente 1, 5.0 V; voltaje de lente 2, 60.0 V. La adquisición de
espectros se realizó para el intervalo 50-1500 m/z, utilizando scan estándar y
resolución normal (velocidad de scan 10300 m/z/s; ancho de peak 0.6
FWHM/m/z). La trampa de iones se utilizó bajo modo de control de carga de
iones (ICC activado), un tiempo máximo de acumulación de 200 ms. La
disociación inducida por colisión (CID) se efectuó mediante helio presente en la
trampa. La fragmentación se realizó utilizando los siguientes parámetros:
SmartFrag, 30-200%; amplitud de fragmentación, 1.00 V; tiempo de
fragmentación, 40ms; ancho de aislamiento MS(n), 4 m/z; promedios MS(n), 5;
número de iones precursores AutoMS(2), 5 señales; y la intensidad umbral
AutoMS(2) se fijó previo a cada inyección. Adicionalmente, para la selección de
iones precursores se utilizó modo de exclusión activa (análisis dependiente de
datos) bajo los siguientes parámetros: exclusión, 2 espectros; tiempo de
exclusión, 1 min.

61
5.7. Fraccionamiento de APPE para la obtención de los polifenoles de alto y
bajo peso molecular. Para obtener fracciones que agrupen los polifenoles de
APPE en aquellos de bajo y alto PM se siguió el esquema de la Figura 5. Un
gramo de extracto APPE seco fue desgrasado en aparato Soxhlet utilizando n-
hexano. El polvo se secó a temperatura ambiente hasta eliminación del solvente
orgánico para luego ser disuelto en agua destilada (50 mL). La disolución acuosa
fue filtrada y luego extraída con acetato de etilo (4 veces). Los productos de
extracción con acetato de etilo fueron reunidos y concentrados a presión reducida
(< 40°C), para obtener un polvo amarillo de sabor amargo (EAE = 180 mg). El
remanente acuoso (AQUO) fue reservado. El extracto EAE fue fraccionado en
una columna de Toyopearl HW-40s (30 x 2.5 cm, Tosoh, Tokio), utilizando un
gradiente lineal desde 5% a 100 % de metanol y luego hasta 60 % de acetona
para elución del material polimérico retenido, obteniendo nueve fracciones. Las
fracciones I-VI que contienen los compuestos de bajo peso molecular fueron
concentradas y secadas bajo presión reducida (LMW). La fracción AQUO fue
purificada mediante una columna de Sephadex LH-20 (30 mm i.d. x 300 mm)
previamente acondicionada con agua durante 24 horas (181). La muestra acuosa
se aplicó en la columna para posteriormente iniciar la purificación mediante un
gradiente escalonado de agua-metanol (incrementos de 20 %) hasta 100 % de
metanol y luego acetona al 70%. El material oligomérico y polimérico se recuperó
de la columna en estas dos últimas fracciones. La fracción polimérica se terminó
de enriquecer en columna de Toyopearl HW-40s. Las fracciones de igual
composición (TLC en fase normal) fueron reunidas. Todas las placas fueron
reveladas con una solución de dimetilaminocinamaldehido (DMACA) al 1% en
etanol/HCl concentrado (1:1). Para todos los extractos obtenidos se determinó el
contenido de polifenoles totales como equivalentes de ácido gálico (mg GAE por
gramo de extracto seco). La ecuación de la curva de calibración fue y = 0.091x +
0.0229 (r2 = 0.9951).

62
Figura 5. Esquema de fraccionamiento del extracto APPE para la obtención de sus constituyentes de alto y bajo peso molecular.
5.8. Efecto del extracto crudo de cáscara de manzana (APPE) sobre la actividad
de la enzima ureasa extraída de Jack bean y H. pylori. La investigación
preliminar del potencial inhibitorio de ureasas bacterianas, considera el empleo
de la enzima obtenida de Jack bean (Canavalia ensiformis), la cual es mas
barata, abundante y se puede obtener comercialmente con alta pureza. A los
criterios anteriores se suma el hecho que el sitio activo de la mayoría de las
ureasas es altamente conservado. Por lo tanto, para la evaluación preliminar del
extracto de cáscara de manzana Granny Smith, se empleó la ureasa Jack bean
(Tipo IV, Sigma). La generación de amoniaco se evidenció mediante el ensayo
del rojo fenol desarrollado originalmente por van Slyke (182) y modificado por
Tanaka (183). Este indicador de pH puede detectar cambios entre pH 6,8 a 7,7 lo
que se manifiesta como un viraje desde el color amarillo al rojo intenso. Desde el
punto de vista espectroscópico, este cambio concuerda con la aparición de una
banda de absorción intensa a 570 nm. La base del método considera que la
ureasa al tener disponibilidad de urea a pH 6,8 (buffer fosfato), rápidamente

63
puede generar amoniaco y elevar el pH hasta 7,7. Matemáticamente esta
variación de la absorbancia en el tiempo (dA/dt), corresponde a la pendiente en la
porción recta de cada curva según lo recomendado por Hu y Mobley (184), la
que se relaciona linealmente con la actividad enzimática remanente en el medio
de incubación (rango 0.0065-0.4 UI; y = 0.0043x + 0.0066, r2 = 0.9971), lo que
permite estimar los porcentajes de inhibición. El método de Tanaka tiene la
ventaja de que permite estudiar la actividad de la enzima en distintas condiciones
de pre-incubación con los inhibidores y además, potencial sitio de interacción de
éstos con la proteína, o si existe un efecto regenerador de la actividad por la
adición de agentes protectores como el DTT a la enzima previamente inhibida. La
ureasa de H. pylori se extrajo de suspensiones bacterianas obtenidas de colonias
desarrolladas en placas de agar sangre, las cuales se lavaron 3 veces con PBS
frío. Los lisados se obtuvieron por centrifugación (14000 x g por 15 min) del
producto de varios ciclos (8 en total) de congelamiento (-70°C)/
descongelamiento y aplicación de ultrasonido (60 sec). Los sobrenadantes
reunidos fueron desalados mediante una columna de Sephadex G-25 (1 x 5 cm)
para luego ser concentrados por ultrafiltración usando una membrana de 300 kDa
Amicom (Millipore) (185). Para estabilizar la ureasa, se agregó un volumen igual
de glicerol y almacenó a – 20°C. La concentración de proteínas totales se
determinó mediante el micro-método de Bradford (Bio-Rad), utilizando soluciones
de BSA para la calibración. Para el ensayo de actividad de ureasa de H. pylori, se
utilizó el método del rojo fenol sugerido por Schwartz (186) con leves
modificaciones. De las suspensiones o lisados de H. pylori (1.5 mg proteínas/mL)
se tomó una alícuota de 10 µL y mezcló con 10 µL de buffer o soluciones con
distintas concentraciones de los extractos en placas de 96 pocillos. Posterior al
periodo de preincubación (0 – 1 horas) se agregó 100 L de una solución que
contiene urea 250 mM y 0,001 % de rojo fenol disueltos en buffer fosfato pH 6,8,
EDTA 10 mM y se incubó a 37 °C por un máximo de 120 min. La inhibición se
estimó procediendo de la misma manera que se describió para la enzima de Jack
bean.

64
5.9. Influencia de los extractos de cáscara de manzana sobre parámetros
cinéticos de la enzima ureasa de Jack bean. Para el estudio del efecto sobre
parámetros cinéticos de la enzima ureasa (Km y Vmax), se utilizó un ensayo para
la determinación del amonio producido por la enzima ureasa, basado en el
protocolo original de Weatherburn (187). Este ensayo presenta mayor
sensibilidad, la cual es necesaria para determinar la actividad de la enzima a
concentraciones más bajas de sustrato que las empleadas en el ensayo del rojo
fenol. En este ensayo se incuba alícuotas de la preparación enzimática (5 a 10
L, conteniendo entre 0,6 a 1 g de proteína), por 10 min a temperatura ambiente
(25°C) en 1 mL de amortiguador fosfato (fosfato 100 mM, EDTA 10 mM; pH 7.5),
conteniendo distintas concentraciones de urea (0.2 hasta 15 mM).
Posteriormente, se extrae una alícuota de 90 L y mezcla con 150 L de reactivo
fenol-nitroprusiato (20 g fenol más 200 mg de nitroprusiato de sodio en un litro de
agua bidestilada), 150 L de hipoclorito alcalino (370 mM NaOH; 80 mM
Na2HPO4; 13 mM NaOCl, pH 12), y 750 L de agua destilada y se incuba por 10
min a 37 °C. La absorbancia se registra a 630 nm (producto coloreado azul de
indofenol). La cantidad de amonio producido durante los 10 min se calcula a partir
de una curva de calibración construida con soluciones de NH4Cl (rango 30 a 250
M, y = 0,001x – 0,0022; r2 = 0,996). Finalmente, a partir de las curvas de
velocidad inicial (mol NH4/min/mg de proteína) versus mM de urea, se obtiene la
Vmax y la Km utilizando el software Graph Pad Prisma. Para la representación de
los resultados se construye las gráficas de doble recíproco (Lineweaver-Burke).
5.10. Detección “in-gel” de la actividad anti-ureasa de Jack bean. La actividad de
la ureasa también fue estudiada por electroforesis (“in-gel”), en condiciones no-
denaturantes según protocolo sugerido por Mobley et al. (188). Para ello se
preparó geles de poliacrilamida al 8% sin SDS. La ureasa fue incubada
previamente con los extractos (30 min) e inmediatamente cargada en los geles
(20 l por pocillo). La electroforesis fue realizada con el buffer de corrida Tris-
glicina a pH 8.8 a un voltaje de 150 V por 1 h. La detección de la actividad fue

65
realizada posteriormente lavando los geles con un amortiguador acetato 5 M frío
y luego con varios traspasos en una solución de rojo cresol al 0.02 % y EDTA
0.1% hasta que el gel adquirió un color amarillo. Una vez drenado, el gel fue
cubierto con una solución de urea al 1.5 % y se observó la aparición de bandas
púrpura correspondientes a la elevación del pH producto de la generación de
amoniaco por la ureasa activa. El gel es escaneado y la imagen documentada
como archivo jpg.
5.11. Protocolo modificado para la detección “in-gel” de la actividad anti-ureasa
de H. pylori. Se desarrolló un nuevo ensayo para la detección “en-gel” de la
actividad de la ureasa, el cual consistió en interaccionar los componentes del
extracto una vez corrido el gel de acrilamida al 7 %, para la separación de la
ureasa nativa (Figura 6). Antes de incubar con el extracto, el gel con la ureasa
separada fue neutralizado con un amortiguador acetato pH 6,5. La incubación
con el extracto se realizó durante 1 hora para posteriormente drenar y lavar el
exceso de extracto. El gel fue puesto en contacto con una lámina de agarosa (0,5
mm) preparada al 0,8 % en una solución de urea 150 mM, EDTA 0,01 % y el
indicador rojo cresol al 0,01% en amortiguador fosfato pH 6,8. Para la detección
de la actividad ureolítica, la lámina fue puesta en contacto con el gel de
acrilamida y escaneada a diferentes tiempos (cada 5 min durante 30 min) hasta la
aparición de las bandas de color púrpura. A diferencia del método desarrollado
por Mobley, éste nuevo procedimiento permite estudiar el efecto de los
inhibidores sin que se generen artefactos como la agregación de la enzima o
pérdida del inhibidor durante los procesos de corrida electroforética y lavados
intermedios.

66
Figura 6. Esquema de preparación de una membrana sensible a ureasa para la investigación de inhibidores.
5.12. Efecto de APPE sobre los niveles intracelulares de ATP de H. pylori como
indicador de viabilidad. Como una medida indicativa de la carga energética
intrabacteriana, se midió los niveles intracelulares de ATP mediante el sistema
luciferina-luciferasa mediante el protocolo adaptado de Schweinitzer et al., (189).
La detección de ATP se basa en una reacción catalizada por la enzima luciferasa
que genera fotones. La preparación bacteriana fue obtenida a partir de cultivos
tempranos en fase de crecimiento mid-log como suspensiones con una
absorbancia (600 nm) de 0.3 (~ 108 UFC/mL) y 0.03 (~107 UFC/mL). Las células
fueron tomadas directamente de las placas de cultivo en fase de crecimiento
exponencial y cuidadosamente suspendidas en caldo Brucella sin ser lavadas.
Las bacterias fueron incubadas por 30 min en presencia del extracto de cáscara
de manzana APPE a 37 ºC antes de medir los niveles de ATP. Para la
determinación de los niveles de ATP el reactivo Bac Titer-Glo (mezcla de reactivo
luciferina-luciferasa ATP-dependiente, (Promega Inc., Madison, WI), fue
agregado directamente sobre la suspensión de bacterias vivas en una relación
1:1, después de los periodos de incubación con los extractos. Para permitir la lisis
de las bacterias y el inicio de la reacción enzimática, la mezcla se deja incubar

67
por 5 min a 37 ºC, para luego registrar la emisión de fotones mediante el
multilector Synergy HP (Biotek Instruments, Vermont, USA). Para la
cuantificación de ATP se construyó curvas de calibración entre 3 x 10-5 M y 3 x
10-10 M.
5.13. Detección de la citotoxicidad temprana de los extractos de cáscara de
manzana utilizando como indicador la reducción de resazurina. Para la
evaluación del efecto temprano de los extractos sobre la viabilidad de H. pylori
se utilizó el colorante resazurina, el cual a diferencia del MTT no requiere ser
extraído y genera un producto de intensa fluorescencia (resofurin), lo que permite
la detección de un menor número de bacterias viables que cuando se usa
reactivos coloreados (190). Para la estimación de la actividad anti-H. pylori se
basó en el protocolo de Takahashi et al., (191) modificado posteriormente por
Kamkaen et al., (192). En breve, en placas blancas de 96 pocillos se preparó
diluciones de los extractos de cáscara de manzana a partir de una solución stock
de 10 mg GAE/mL. Las placas fueron secadas al vacío en un desecador. H. pylori
TX 136 fue cultivado a 37 °C por 48 horas bajo atmósfera microaerofílica en caldo
Brucella suplementado con un 0.1 % de ciclodextrina. La pureza del cultivo se
verificó por tinción de Gram. La absorbancia del cultivo a 600 nm, se ajustó a
0.002 unidades, equivalente a 0.8 x 106 UFC/mL. De este cultivo se agregó 100
L a cada pocillo de las placas preparadas con el extracto y la placa se incubó
con agitación a 37 °C en condiciones de microaerofilia durante periodos de 30, 60
y 120 min. En cada tiempo se retiró una de las placas y agregó 10 L de una
solución de resazurina al 0.01 % en PBS 1X estéril. Rápidamente (< 5 min), a
cada pocillo se le agrega 50 L de aceite mineral con el fin de impermeabilizar la
muestra al efecto del oxígeno ambiental durante el periodo de lectura. Las placas
luego fueron incubadas a 37 °C dentro del multilector Synergy HT (Biotek),
durante 2 horas, registrando en forma continua el aumento de la fluorescencia
originada por bacterias viables usando como longitud de onda de excitación de
530 nm y de emisión 590 nm. El grado de citotoxicidad de los extractos se calcula
como el porcentaje del control sin extractos y sustrayendo la posible

68
autofluorescencia de cada muestra o control (pocillos con extracto y/o con H.
pylori, pero sin rezasurina).
5.14. Efecto de los extractos de cáscara de manzana sobre la respiración de H.
pylori medido por la sonda MitoXpress. El efecto de los extractos sobre la
respiración de H. pylori fue evidenciado mediante la determinación del consumo
de oxígeno. Dado que el uso del electrodo Clark tiene importantes limitaciones
para este tipo de estudio, recientemente el empleo de la sonda MitoXpress
(Luxcell, Irlanda), ha sido validado para dicho fin, mostrando resultados
comparables a los obtenidos mediante el método polarográfico, aunque con
mayor sensibilidad y High throughput (193, 194). El método puede ser aplicado a
preparaciones como mitocondria aislada, células eucarióticas, tejidos y bacterias
(195). La sonda MitoXpress se reconstituye en 1 mL de agua para dar una
solución de 1 M. Doscientos microlitros de H. pylori (104 - 107) cultivado en
medio líquido (caldo Brucella) son agregados a los pocillos de una placa con los
extractos y la sonda en una concentración final de 100 nM. Rápidamente, la placa
es sellada con una capa de aceite mineral (50 L), para minimizar el efecto del
oxígeno atmosférico sobre la viabilidad del microorganismo. La placa es leída a
37 °C cada 30 min hasta 24 h, utilizando el multilector Synergy HP (Biotek) en la
modalidad de fluorescencia Time-Resolved. En este ensayo las distintas
concentraciones del extracto son agregadas conjuntamente con la suspensión de
bacterias.
5.15. Obtención de los neutrófilos humanos: De voluntarios sanos se obtuvo sangre
venosa (6 mL) en tubos heparinizados. La sangre se diluyó a la mitad con PBS 1
X. La sangre diluida (6 mL) se agrega cuidadosamente sobre una mezcla de 3
mL de Histopaque 1119 (inferior) y 3 mL de Histopaque 1077 (superior). Los
tubos se centrifugan a 700 x g por 30 min a temperatura de 25 ºC. Los neutrófilos
se separan en la segunda interfase, son colectados y lavados dos veces con 10
mL de PBS 1 X, centrifugando a 200 x g por 10 min. Los neutrófilos se cuentan
en una cámara de Neubauer, y su viabilidad se confirma utilizando el ensayo de

69
exclusión de azul de Trypan. La preparación de esta manera contiene más de un
95 % de neutrófilos con una viabilidad superior al 98 %. Para el ensayo los
neutrófilos son suspendidos en amortiguador HBSS con 1 mM de CaCl2, 1.5 mM
de MgCl2 y 10 mM de glucosa.
5.16. Efecto de los extracto de cáscara de manzana sobre la Inducción del
estallido respiratorio por PMA, fMLP y H. pylori. La quimioluminiscencia del
luminol (sonda permeable) y su isómero Isoluminol (sonda impermeable) fue
medida en microplacas blancas de 96 pocillos (Nunc, fondo plano) basado en la
modificación de protocolos previamente publicados (196-200). Las sondas fueron
preparadas disolviendo 18 mg de 5-amino-2,3-di-hidro-1,4-ftalazinediona (luminol,
Sigma) y 6-amino-2,3-dihidro-1,4-ftalazinediona (isoluminol, Sigma) en 100 mL de
PBS 1 X (pH 7.4). Una alícuota de 100 L de la suspensión de neutrófilos (1 x 105
células por pocillo) se mezcló con 10 L de la solución stock de luminol o
isoluminol. Para el estudio de la producción intracelular de ROS, se utilizó luminol
más una mezcla de 50 U/mL de superóxido dismutasa (SOD, Sigma) y 2000
U/mL de catalasa (CAT, Sigma). Para el estudio de la producción extracelular de
ROS la luminiscencia del isoluminol (impermeable) se amplificó agregando 4
U/mL de horseradish peroxidasa (HRP, Calbiochem). Después de medir la
luminiscencia por 10 min a 37 °C, se agregó los distintos activadores y la lectura
se continuó entre 15 a 60 minutos, dependiendo del estímulo. Para estudiar el
efecto de los extractos de cáscara de manzana, distintas concentraciones de
éstos últimos fueron preincubadas con los neutrófilos previo a la activación con
PMA (200 nM); fMLP (1 M) y H. pylori no-opsonizado (MOI 25:1, o bien 2,5 x 106
UFC/mL) (61). La señal de luminiscencia fue integrada y los resultados se
expresaron como porcentaje de inhibición de la producción de ROS en relación al
control no tratado con los extractos. El efecto de primación de los neutrófilos por
incubación con H. pylori se comprobó por incubación durante 30 minutos con la
bacteria (107 y 105 UFC) y luego activación con PMA.

70
5.17. Análisis de la estabilidad de los polifenoles de APPE en fluido gastrico
simulado. Para evaluar el efecto de pH ácido sobre la estabilidad de los
glicósidos de flavonoides y procianidinas de APPE se utilizó procedimientos
previamente publicados (201, 202) con algunas modificaciones. APPE (250 mg)
fue disuelto en 25 mL de solución fluido gástrico artificial sin pepsina (2.0 g NaCl
y 3 mL HCl concentrado, diluido a un 1 L, pH ~ 2), e incubado por 4 h a 37°C con
agitación (100 rpm). Para el análisis de flavonoides se tomó alícuotas de 200 L a
los 15, 30, 60, 120, 180 y 240 min, se filtraron por membrana de 0.45 m y
analizaron por RP-HPLC-DAD con el sistema cromatográfico publicado
previamente (201), usando quercetina como estándar de calibración. Para HPLC
se utilizó un sistema Lachrom (Merck, Alemania), equipado con una columna
Kromasil KR100-5C18 de 250 x 4.6 mm, 5 m, (Eka Chemicals AB, Bohus,
Sweden). La fase móvil fue: solvente A (agua con 0.1 % TFA, v/v) y solvente B
(acetonitrilo con 0.1 % TFA). El programa de gradiente utilizado fue: 0-25 min:
10-30% de B; 25-30 min: 30-75 % de B; 30-35 min; 75-10% de B con un flujo de 1
mL/min. La estabilidad de las procianidinas fue evaluada por NP-HPLC de
acuerdo a un método previamente publicado (203) con una columna de 250 x 4.6
mm (5 m) Lichrospher 100 Diol column (Merck, Darmstadt, Germany). El
sistema de solventes utlizado fue: solvente A (acetonitrilo/ácido acetico, 98:2 v/v)
y solvente B (metanol/agua/ácido acético, 95:3:2 v/v). Las procianidinas fueron
eluídas empleando el siguiente programa de gradiente: 0-35 min, 0 - 40% de B;
35-55 min; isocrático 40 % de B; 55-60 min, 40-0% de B con 5 min de re-
acondicionamiento de la columna y un flujo de 0.6 mL/min. La separación fue
monitoreada por detección de fluorescencia a 230 nm y 321 nm como longitudes
de onda de excitación y emisión, respectivamente. La suma del área de los peaks
entre 16-50 min fue comparada con el aumento de los monómeros (epicatequina,
tR = 9.5 min), la cual se considera un marcador de hidrólisis. La estabilidad de los
flavonoides fue monitoreada específicamente a 355 nm. En esas condiciones los
glicósidos de flavonoides eluyen entre los 12-20 min. La disminución del área
total en ésta zona se interpreta como indicador de hidrólisis/inestabilidad, lo que

71
se comprueba observando el aumento paralelo del área del peak de quercetina
(tR = 24 min). La cuantificación de quercetina fue llevada a cabo usando
estandarización externa.
5.18. Cultivo de células HeLa para el estudio del efecto inhibitorio de los
polifenoles sobre la actividad de la toxina VacA. Las Células HeLa (ATCC)
fueron cultivadas en placas de 55 cm2 (1 - 2 x 105 células por cm2), hasta 80 %
de confluencia, utilizando MEM 90 % con 10 % de SFB, 2 mM GlutaMAX, 100
U/mL de penicilina, y 100 g/mL de estreptomicina. El medio fue cambiado cada
3 días. Para liberar las células se empleó 0.5 mg/mL de tripsina con 0.2 mg/mL
EDTA en PBS. Para los experimentos, las células fueron sembradas en placas de
6-12 ó 24 pocillos 24 a 48 h antes de los ensayos. La viabilidad de las células
HeLa fue determinada por el procedimiento de exclusión de azul de trypan.
5.19. Obtención de sobrenadantes enriquecidos en VacA a partir de cultivos de
H. pylori: Para la obtención de los sobrenadantes enriquecidos en VacA, se
utilizó la cepa de H. pylori TX 136 que fue cultivada de la forma habitual en caldo
Brucella hasta 48 h (37°C, 10 % O2, 6 % CO2). Posterior a la incubación, el medio
fue centrifugado (14000 x g) y los sobrenadantes esterilizados por filtración a
través de una membrana de acetato de celulosa de 0.22 m (112). Las proteínas
del sobrenadante fueron precipitadas con una solución de sulfato de amonio
saturada al 50 % con agitación. El precipitado se dejó reposar durante 12 h a 4
°C y luego se centrifugó. El pellet fue resuspendido en 60 mM Tris, pH 7.5 con
100 mM NaCl, para ser concentrado 40 veces utilizando una membrana de
ultrafiltración con un cut-off de 10 kD (Centricom). Para la determinación de
proteínas se utilizó el método de Bradford con BSA como estándar. La presencia
de VacA se comprobó mediante western blot (26). Un caldo Brucella sin inocular
fue procesado de la misma manera para ser empleado como control negativo de
vacuolación.

72
5.20. Determinación de proteínas: Para la determinación de proteínas en los
sobrenadantes se mezcló 5 L de muestra con 200 L de reactivo de Bradford
(Bio-Rad protein assay) y 795 L de agua. Las muestras se leyeron a 595 nm.
Como estándar de calibración se utilizó BSA (204).
5.21. Ensayo de vacuolación. Las células HeLa fueron incubadas por 16 horas a 37
°C en 5 % de CO2 con (i) VacA de H. pylori, (ii) caldo Brucella no-inoculado o (iii)
APPE. La vacuolación de las células HeLa fue evaluada cuantitativamente
mediante el ensayo de captación de rojo neutro según metodología previamente
publicada (205) y por microscopia óptica de acuerdo a Harris et al., (206). Se
definió como célula viable aquella que presentó contornos citoplasmáticos y
nucleares bien definidos, la vacuolación se definió como la presencia de una o
más vacuolas intracitoplasmáticas en células viables. Para cada determinación se
evaluó 100 células HeLa y se expresó en porcentaje de células vacuoladas como
el promedio ± la desviación estándar de la media y comparadas por la prueba
estadística de ANOVA.
5.22. Ensayo de adhesion de H. pylori a cultivos de células HeLa. Las propiedades
anti-adherentes de APPE fueron evaluadas utilizando una modificación del
protocolo de Lee et al., (207). Brevemente, las células HeLa (100 L) fueron
sembradas en placas de 6 pocillos (Corning, Cambridge, M.A., USA) a una
densidad de 5 x 104 células/mL. Después de alcanzar confluencia en 24 horas,
las monocapas celulares fueron lavadas tres veces con PBS. La unión no
específica fue bloqueada incubando los cultivos con BSA 0.5 %. Una suspensión
de H. pylori fue lavada dos veces con PBS para luego ajustar su densidad a
OD600nm = 1.0 (1 x 108 UFC/mL). De esta suspensión se tomó 50 L e incubó con
distintas concentraciones de APPE durante 30 min. Posteriormente, 100 L de la
mezcla (H. pylori, APPE) fueron agregados a los cultivos de células HeLa e
incubados por 1 h. La detección de las bacterias adherentes se evidenció
utilizando el reactivo rojo fenol 0.03 %, urea 2 % (pH 5.2) y leyendo el cambio de

73
absorbancia a 560 nm en un lector de ELISA (Tecan, Männedorf, Suiza). El
porcentaje de H. pylori adheridos se calculó utilizando la siguiente ecuación: %
de adhesión = 100 - [(ODexperimental – ODnegativo)/ (ODpositivo – ODnegativo) x 100]. El
control negativo contiene sólo células HeLa y el positivo contiene células HeLa y
bacterias (100 % de adhesión).
5.23. Determinación de la unión inespecífica y el ingreso de polifenoles de APPE
en células HeLa: Se utilizó una modificación del procedimiento de (171), para
determinar la unión inespecífica e ingreso de los polifenoles de manzana en los
cultivos de células HeLa. Las muestras procesadas fueron las mismas utilizadas
en los ensayos de adhesión. El medio fue aspirado y las células lavadas con PBS
1 X. Se agregó nuevo medio de cultivo MEM 90% y FBS 10% y se dejó incubar
las células por 2 h. Posterior a la incubación, las células fueron lavadas y
despegadas mediante tripsinización según el procedimiento de rutina. Los pellets
obtenidos por centrifugación (600.000 células/mL) se lavaron exhaustivamente
con amortiguador fosfato salino para luego ser extraídas con 300 μL de metanol
(0.01 % BHT) en un baño de ultrasonido por 30 min a 4ºC. El extracto final se
centrifugó a 13.000g. El sobrenadante y los pellets se analizaron por HPLC de
acuerdo a las condiciones señaladas en la sección 5.4 (RP-HPLC-DAD-FLD),
expresando los resultados como ng/106 células. Todos los ensayos se realizaron
en triplicado.
5.24. Animales. Para la evaluación in vivo se utilizó ratones C57BL/6 de 5 semanas de
vida. Los ratones fueron adquiridos en el ISP (Instituto de Salud Pública,
Santiago, Chile). Los animales fueron depositados en jaulas de plástico bajo
ciclos de luz-oscuridad de 12 horas, alimentados con dieta estándar consistente
en pellets para roedores y acceso ad libitum a agua autoclavada. El alimento para
los ratones se mantuvo almacenado en un lugar especialmente destinado para
ello, seco y ventilado. La aprobación para éste estudio fue obtenida previamente
a los experimentos de parte del comité de ética animal, y todos los

74
procedimientos cumplieron con la guía de cuidados y manipulación de animales
de laboratorio en la Universidad de Concepción.
5.25. Preparación del inóculo para el estudio in vivo. Se utilizó la cepa de H. pylori
(ATCC 43504, cagA+, vacA+, American Type Culture Collection, Rockville, MD,
USA), la cual fue obtenida de placas agar-sangre (5 días de cultivo) y luego
subcultivadas en caldo Brucella (Becton Dickinson, Cockeys-ville, MD, USA) con
SFB 10% v/v a 37 °C por 40 h bajo condiciones de microaerofilia (6 % CO2) y alta
humedad y agitación moderada (150 rpm).
5.26. Protocolo experimental. Veintiocho ratones C57BL/6 (20-25 g) fueron divididos
en 4 grupos de acuerdo al esquema de la Figura 1. Antes de las inoculaciones, a
los animales se les limitó el acceso a comida por 24 h. El inóculo usado fue de 1
x 109 UFC/mL (en caldo Brucella), el cual fue administrado en forma oro-gástrica
con un catéter de teflón (diámetro externo = 0.1cm). Se realizó 4 inoculaciones de
0.3 mL cada una con intervalos de dos días. Previo al inicio del experimento y
para monitorear el progreso de la infección por H. pylori, cada semana se
sacrificó un animal de un grupo infectado. Los estómagos obtenidos de estos
animales fueron utilizados para cultivo, PCR de tiempo real y examen histológico.
A partir del día 8 se inició la administración del extracto por vía orogástrica (200
l) durante 20 días. Al final del tratamiento los animales fueron mantenidos sin
alimento toda la noche y sacrificados por dislocación cervical. Los segmentos
antrales y el cuerpo fueron removidos, lavados con suero salino 0.9 % y
seccionados. Para la evaluación histopatológica parte de los segmentos fueron
almacenados en una mezcla de PBS con 10 % de formol. El resto de las
muestras fueron almacenadas a -20 °C hasta su empleo para el análisis por PCR
de tiempo real y la determinación de los niveles de MDA.
5.27. Evaluación histopatológica. Las muestras de cuerpo y de antro fijadas en
formalina e incluidas en parafina, fueron teñidas con hematoxilina-eosina, y
posteriormente se avaluó en forma semicuantitativa las siguientes cinco

75
características: 1) grado de inflamación, 2) infiltración neutrofílica, 3) atrofia
glandular, 4) metaplasma intestinal y 5) densidad de H. pylori (tinción de Warthy
Starry). Dentro de cada una de las cinco características se utilizó el sistema
actualizado de Sydney (208), que incluye las categorías de “ausente”, “leve”,
“moderado” y “severo”. Para lograr mayor afinamiento de los resultados se uso la
subdivisión recomendada por Chen et. al., (209). Para ello, se aplicó la escala de
0-6: 0 (ausente); 1-2 (leve), 3-4 (moderado), 5-6 (severo).
5.28. Niveles de MDA: Los niveles de MDA fueron determinados utilizando el protocolo
de Bergamo et al., (210). Para ello, los tejidos fueron sonicados con 10
volúmenes de amortiguador fosfato 50 mM para luego ser des-proteinizados con
TCA frío al 10%. Las muestras se sometieron a ultrasonido por 30 min. Después
de agitar durante 3 min, se centrifugó a 10.000 x g por 5 min. Al sobrenadante se
le agregó 700 L de ácido tiobarbitúrico (en amortiguador acetato 2M pH 3,
desgasificado y purgado con nitrógeno). La mezcla desgasificada se incubó por
30 min a 90ºC. Posteriormente se enfrió y centrifugó a 10.000 x g por 5 min. De
esta mezcla se inyectó 20 μL en sistema HPLC con detector de fluorescencia. La
separación del adúcto MDA-TBA se realizó en forma isocrática con la fase móvil
compuesta de metanoly amortiguador fosfato 50 mM (pH 6.8) en una proporción
60 + 40 v/v. La cuantificación de MDA se realizó por calibración con MDA
estándar preparado a partir de un patrón de tetrametoxipropano (Sigma) y
expresando los resultados como nmol MDA/g de tejido.
5.29. Determinación cuantitativa de H. pylori por PCR de tiempo real (real-time
PCR). El ADN genómico fue extraído desde las muestras de estómagos de ratón
de acuerdo al protocolo de Downset et al., (211) usando un Kit comercial (Wizard
® SV Genomic DNA Purification System Promega). Como controles negativos de
amplificación se usó agua libre de RNAsa/DNAsa y Lactobacillus. Como control
positivo, se usó el ADN genómico de la cepa de referencia de H. pylori ATCC
43504. Para la preparación de TaqMan® Master Mix (Roche) se mezcló: 4 L de
TaqMan Universal master mix, 9.1L de agua grado PCR, 0.5 L de la sonda

76
TaqMan 10 M, 0.7 L del partidor 16S forward 10 M, 0.7 L del partidor 16S
reverse 10 M y 5 L del ADN templado proveniente de la muestra. La
amplificación y detección se realizaron en el sistema LightCycler (Roche
Diagnostics). El ADN fue amplificado usando los siguientes parámetros:
calentamiento a 95°C por 10 min seguido por 50 ciclos de denaturación (95°C, 10
s), annealing (55 °C, 10 s), y extensión (72 °C, 8 s). Las secuencias de los
partidores para 16S rRNA se muestran en la Tabla 2.
5.30. Expresión de resultados y análisis estadístico. Los resultados presentados
corresponden al promedio ± SD o SEM del número de experimentos
independientes indicados. En algunos casos las figuras o imágenes
corresponden a resultados representativos de cada grupo de experimentos
repetidos al menos 3 veces en forma independiente. La significancia estadística
de los datos se evaluó mediante las pruebas t-test o ANOVA. Se consideró como
límite de significancia estadística, valores de P < 0.05. Como post-test a las
pruebas de hipótesis se utilizó la prueba de comparaciones múltiples de Tukey.
Tabla 2. Partidores para PCR en tiempo real
Partidor Target gene
Secuencia Tamaño del
producto
Accesión N°
Hp 16S rRNA TGCGAAGTGGAGCCAATCTT (forward)
GGAACGTATTCACCGCAACA (reverse)
119 pb U00679

77
6. RESULTADOS
6.1. Preparación del extracto enriquecido en polifenoles de cáscara de manzana
libre de azúcares (APPE).
De acuerdo a lo planteado en el Objetivo 1, en la Tabla 3 se presenta el resultado del
análisis de la composición general del extracto enriquecido en polifenoles mediante
Sepabeads SP-850 (APPE). Una de las principales ventajas de usar este tipo de soporte
cromatográfico es la remoción de la mayoría de los azúcares simples y pectinas presentes
en los extractos acuosos. La concentración de polifenoles totales observada inicialmente
en los extractos acuosos fluctúa entre un 3.0 a 5.0 mg por cada 100 g de cáscara fresca
(82 ± 8 % de humedad). Mediante esta técnica, la recuperación de los polifenoles
presentes en la cáscara fresca fue superior al 95 %. El enriquecimiento que se obtiene
como resultado del proceso de captura y remoción de los polifenoles se desprende de la
Tabla 3 y corresponde a un rango de 58-62 % de equivalentes de ácido gálico (GAE) por
cada 100 g de extracto seco, determinado con el reactivo de Folin-Ciocalteau.
Tabla 3. Composición general del extracto de cáscara de manzana Granny Smith enriquecido mediante Sepabeads SP-850 (APPE).
Composición % en el extracto*
sacarosa Nsd glucosa Nsd fructosa Nsd
Lípidos y ceras 4.0 Oligosacáridos neutros 15-20
Pectinas < 0.1 Fenoles totales 58-62
Otros 6-13
TOTAL 100
Nsd: No se detectó
*Cálculo en base seca

78
6.2. Análisis de la composición de los extractos de cáscara de manzana mediante
RP-HPLC-DAD-FLD y NP-HPLC-FLD:
Para comprobar que el proceso de extracción y concentración no genera distorsiones en
el perfil de polifenoles, el extracto APPE se analizó detalladamente por HPLC y comparó
con un extracto de cáscara fresca de la variedad Granny Smith obtenido con acetona 70
%. El resultado de éste análisis se presenta en la Figura 7a. Los perfiles de HPLC para la
cáscara fresca y APPE muestran gran similitud cualitativa para los 13 compuestos
identificados por co-inyección con estándares y detectados por UV y fluorescencia. En la
Figura 7c se muestra el resultado de la etapa final de purificación de las fracciones ACUO
y EAE que lleva a la obtención de las fracciones HMW y LMW (TLC, carril 4 y 5). Para
lograr dicha separación, la fracción ACUO fue limpiada mediante cromatografía en
Sephadex LH-20, dando lugar a las fracciones F1-F7 presentadas en la placa TLC de la
Figura 8. Las fracciones F6 y F7, con un elevado contenido de procianidinas de alto peso
molecular, fueron reunidas y se les denominó HMW (High Molecular Weight). La fracción
EAE (último carril de la TLC a la derecha) presenta principalmente glicósidos de
flavonoides y una mancha característica (Rf = 0.7) de epicatequina. Nótese que todos los
compuestos derivados de flavan-3-oles (epicatequina en este caso), forman un cromóforo
de color azul con DMACA que permite diferenciarlos de los flavonoides que permanecen
de color amarillo-anaranjado. La fracción EAE fue posteriormente purificada por Toyopearl
HW40s para la completa eliminación de procianidinas de alto peso molecular. Esta última
fracción purificada fue denominada LMW (Low Molecular Weight). Como se observa en la
Figura 7b, mediante RP-HPLC analítica sólo se pudo separar algunas procianidinas
(monómeros, dímeros y trímeros). De la TLC presentada en la Figura 7c, se desprende
que los polifenoles de bajo peso molecular (LMW) y de alto peso molecular (HMW), fueron
agrupados exitosamente mediante la combinación de cromatografía en columna de
Sephadex LH-20 y Toyopearl HW-40s para futuros bio-ensayos. Los polifenoles de LMW
se visualizan como manchas bien separadas con Rfs 0.5. Se debe notar que el reactivo
DMACA permite revelar selectivamente los flavan-3-oles (monómeros y polímeros), los
cuales después de 15 -20 min aparecen como manchas de color verde-azul. Como se
observa en la Figura 8, los flavan-3-oles (monómeros y dímeros) también están presentes

79
en la fracción LMW conjuntamente con los glicósidos de quercetina (manchas pardo-
anaranjadas) previamente identificadas por RP-HPLC-DAD (Figura 7 a). A escala
analítica, las procianidinas oligoméricas y poliméricas se lograron separar con mayor
eficiencia mediante NP-HPLC-FLD, tal como se muestra en la Figura 9 A. En las Figuras
9 B y C se muestra la separación en fase diol de las fracciones HMW y LMW,
confirmando la agrupación de los compuestos de APPE según su tamaño molecular.
Figura 7. Perfiles RP-HPLC-DAD de APPE y cáscara de manzana fresca: (a) comparación entre los perfiles de HPLC para caácara fresca y APPE monitoreados a 280 nm mostrado los peaks (1) ácido clorogénico (2) procianidina B1, (3) procianidina B2, (4) (-)-epicatequina, (5) procianidina C1, (*) desconocido, (6) rutina, (7) hiperósido, (8) isoquercitrina, (9) quercetina-3-O-pentósido, (10) quercetina-3-O-pentósido, (11) quercitrina, (12) floretina-O-xiloglucósido y (13) floridzina. (b) Inserto: ampliación de la zona de flavan-3-oles y algunas procianidinas de APPE región detectadas por RP-HPLC-FLD con excitación de 280 nm y emisión de 310 nm. La numeración utilizada para los polifenoles es la misma usada en el trazado de RP-HPLC-DAD. (c) Separación cromatográfica mediante TLC de las muestras reveladas con el reactivo DMACA. Carril 1, (-)-epicatequina; Carril 2, cáscara de manzana fresca; Carril 3, APPE; Carril 4, LMW; Carril 5, HMW y Carril 6, rutina.
Cáscara fresca
APPE

80
Figura 8. TLC en fase normal de fracciones ACUO purificada por sephadex LH-20. La fase móvil utilizada fue tolueno/acetona/ácido fórmico (3+6+1 v/v). (A) placa cromatográfica observada a la luz visible; (B) placa cromatográfica revelada con el reactivo DMACA. En (B), los compuestos de color amarillo corresponden a flavonoides y los de color azul a derivados de flavan-3-oles (monómeros, oligómeros y polímeros).
Figura 9. Separación de procianidinas de APPE mediante NP-HPLC-FLD. (A) APPE, (B) LMW y (C) HMW. Los números indicados al lado de los peaks indican el grado de polimerización de las procianidinas del tipo B.

81
6. 3. Determinación del grado de polimerización por degradación con los nucleófilos
tolueno--tiol, cisteamina y floroglucinol.
Para obtener mayor información sobre la naturaleza química de las procianidinas de
APPE, éstas fueron degradadas en presencia de tres agentes nucleofílicos: tolueno--
tiol, cisteamina y floroglucinol. Tal como se esquematiza en la Figura 10, los tres
agentes son capaces de depolimerizar las procianidinas generando adúctos entre el
nucleófilo y las unidades de extensión de las procianidinas, liberando a su vez un mol de
la unidad de término. Con los tres reactivos fue posible calcular el grado de
polimerización utilizando el procedimiento descrito en la sección de material y métodos.
Al observar los perfiles cromatográficos de los productos de la depolimerización con
tolueno--tiol, se aprecia claramente que la principal unidad de término y extensión es la
epicatequina con un tR = 8.0 min (Figura 11 A). En la Figura 11 B se observa dos
peaks a un at tR = 13.5 min y 17.0 min, correspondientes a la mezcla de catequina /
cisteamil-catequina y epicatequina / cisteamil-epicatequina, respectivamente. Para lograr
una adecuada identificación de los productos de tiólisis con cisteamina, se aprovechó la
naturaleza iónica de los adúctos formados entre el nucleófilo y el polifenol. Así, usando
una mini-columna de SP-Sepharosa (1 g) se logró la separación por intercambio iónico
de los cisteamil-derivados con carga positiva (179). Los derivados neutros (flavan-3-oles
correspondientes a unidades de término), se retuvieron por separado en una columna de
Amberlite XAD-16 (10 g). De esta manera, se logró elucidar que los peaks están
compuestos por una mezcla del flavan-3-ol y su cisteamil derivado (Figura 12).
Debido a los problemas experimentales (cisteamina) y de seguridad (tolueno--tiol)
presentados por los dos métodos mencionados, se recurrió a un tercer procedimiento
basado en la degradación de las procianidinas en presencia de un exceso de
floroglucinol (Figura 11 C). Mediante éste, fue posible obtener similares resultados, en
un periodo más corto de análisis y con mínimos problemas de toxicidad. A diferencia de
la tiólisis con cisteamina, la interconversión de epicatequina a catequina no es una
reacción importante en este método, siendo las concentraciones de catequina las
mismas que las observadas en las muestras sin degradar. Por lo anterior, los valores de
grado de polimerización (DPm) presentados en la Tabla 9 (sección 6.9) corresponden a

82
los calculados utilizando el proceso de floroglucinólisis. Dichos valores no fueron
diferentes de los obtenidos con los otros dos nucleófilos. Considerando todos estos
resultados, se sugiere que en los cuatro extractos (cáscara fresca, APPE, HMW y LMW),
las procianidinas estarían formadas principalmente por epicatequina, y en menor grado
por catequina, tal como ha sido informado previamente para la cáscara de manzana
(212). En la Tabla 4 se presenta el resultado del análisis cuantitativo de todos los
polifenoles de la cáscara de manzana y APPE y que permiten justificar el 100 % de los
polifenoles determinados por el método de Folin-Ciocalteau.
Figura 10. Depolimerización de las procianidinas de APPE utilizando los agentes nucleofílicos
floroglucinol, cisteamina y tolueno--tiol.
NH3+
Unidades de extensión
Unidad de término
Floroglucinol
Cisteamina
Tolueno--tiol
Procianidina Floroglucinol-aducto
Cisteamil-aducto
Benciltioeter-aducto

83
Figura 11. Cromatogramas representativos de la depolimerización de APPE usando diferentes nucleófilos (A), RP-HPLC de APPE antes (izquierda) y después (derecha) de la tiólisis con
tolueno--tiol; (B), RP-HPLC de APPE antes (izquierda) y después (derecha) de la tiólisis con cisteamina; (C), RP-HPLC de APPE antes (izquierda) y después (derecha) de la degradación con floroglucinol.
tolueno--tiol
cisteamina
floroglucinol

84
Figura 12. Estrategia de identificación de los aductos formados entre cisteamina y flavan-3-oles de APPE. SP-Sepharosa se utilizó como soporte para retener los aductos de carácter catiónico, mientras que la Amberlite XAD-16 se empleó para retener los flavan-3-oles neutros liberados por tiólisis. En los insertos (cuados grises) se muestra las corridas de RP-HPLC-FLD de los eluatos obtenidos con cada columna.

85
Tabla 4. Composición polifenólica de cáscara fresca y el extracto APPE obtenido de la manzana Granny Smith.
Compuesto Manzana fresca Polifenol (%)
a
APPE Polifenol (%)
a
Flavonoides
rutina 3.57 ± 0.50 3.80 ± 0.76
hiperósido 21.08 ± 1.20 19.70 ± 2.20
isoquercitrina 6.25 ± 2.00 8.80 ± 1.81
quercetina-3-O-pentósidosb 18.01 ± 1.54 17.40 ± 3.54
quercitrina 9.01 ± 1.88 7.80 ± 2.01
quercetina 0.09 ± 0.03 0.14 ± 0.08
flavonoides 58.01 ± 1.19 57.64 ± 1.73
Flavan-3-ol monómeros
epicatequina 6.20 ± 0.10 5.60 ± 1.89
catequina Nd 0.05 ±0.01
flavan-3-ol monomeros 6.20 ± 0.10 5.65 ± 0.95
Procianidinas c 22.09 ± 3.00 24.38 ± 2.78
Dihidrochalconas
floridzina 13.05 ± 3.22 11.34 ± 4.12
floretin-2´-xiloglucósido 0.10 ± 0.05 0.14 ± 0.06
dihidrochalconas 13.15 ± 1.64 11.48 ± 2.09
Ácidos fenol carboxilicos
ácido clorogénico 0.55 ± 0.01 0.80 ±0.06
ácido cafeíco Nd 0.05 ±0.01
ácidos carboxílicos 0.55 ± 0.01 0.85 ± 0.04
TOTAL 100 ± 1.19 100 ± 1.52
Nd = No detectado a Los datos son el promedio de tres determinaciones por RP-HPLC y expresados como porcentaje del total
de polifenoles medidos por el método de Folín-Ciocalteau. b Calculado en base a rutina medida por RP-HPLC.
c Estimado por RP-HPLC posterior a la floroglucinólisis.
6. 4. Análisis del perfil de compuestos fenólicos en el extracto de cáscara de
manzana mediante RP-HPLC-ESI-MS/MS.
La muestra APPE se analizó por LC-MS, utilizando el programa de separación descrito en
materiales y métodos. En la Figura 13 A se muestra el cromatograma registrado a 280
nm, mientras que en las Figuras 13 B y C se presenta el cromatograma ion base (BPC,
base peak chromatogram) proporcionado por el espectrómetro de masas adquirido en
polaridad negativa y positiva. Las señales m/z más importantes observadas para cada
peak cromatográfico en las diferentes polaridades examinadas se resumen en la Tabla 5.
Se indica la probable identidad de las señales m/z observadas, para lo cual se utilizó la

86
herramienta ESI-MS Adducts (http://fiehnlab.ucdavis.edu/staff/kind). Para las señales m/z
precursoras se destacan sus fragmentaciones más importantes (fragmentos MSn).
El peak 1 (tR 7.7 min) correspondería al ácido clorogénico (fórmula, C16H18O9; masa
nominal M1, 354 g/mol). Se realizó un análisis complementario de monitoreo de
reacciones múltiples (MRM, multiple reaction monitoring) con el fin de confirmar la
presencia de compuestos específicos. Así, del cromatograma BPC para la señal m/z 353
(especie desprotonada [M-H]-) se tiene un peak principal para un tiempo de retención de
7.7 min. La fragmentación indicó las señales m/z 191 y m/z 135, que coinciden con las
descritas en la literatura para el ácido clorogénico (213).
En base a la literatura el peak 2 correspondería a procianidina B1 (fórmula, C30H26O12;
masa nominal M2, 578 g/mol). Se observa en polaridad negativa la señal m/z 577 que
correspondería a la forma desprotonada (especie [M2-H]-). La fragmentación observada
en polaridad negativa indicó principalmente las señales m/z 425, m/z 289 que coinciden
con las descritas en literatura (214-216). En base a la literatura el peak 3 correspondería
procianidina B2 (fórmula, C30H26O12; masa nominal M3, 578 g/mol). Se observa en
polaridad positiva la señal m/z 579 que corresponde a la forma protonada (especie
[M3+H]+) y en polaridad negativa la señal m/z 577 que correspondería a la forma
desprotonada (especie [M3-H]-). La fragmentación observada en polaridad negativa
indicó principalmente las señales m/z 425, m/z 407, m/z 451 y m/z 290 que coinciden
con las descritas en literatura (214-216). El peak 4 correspondería a epicatequina
(fórmula, C15H14O6; masa nominal M4, 290 g/mol) y se puede observar la señal m/z 291
y m/z 289 en polaridad positiva y negativa respectivamente. La fragmentación de m/z
291 indicó las señales m/z 123, m/z 139, m/z 273, coincidentes con las descritas en
literatura (217). Por MRM se confirmó en polaridad negativa que el peak 4 corresponde a
epicatequina, ya que en esta modalidad el cromatograma BPC para m/z 289 (especie
desprotonada [M-H-] tiene un peak principal para un tiempo de retención de 10.6 min. La
fragmentación dio origen a las señales m/z 245, m/z 205, m/z 137 y m/z 161, que
coinciden con las descritas en literatura (215). El peak 5 correspondería a procianidinas
C1 (fórmula, C45H38O18; masa nominal M5, 866 g/mol). Se observan las señales m/z 867
y m/z 866 en polaridad positiva y negativa respectivamente. La fragmentación de m/z
866 en polaridad negativa (originando las señales m/z 739, m/z 713, m/z 695, m/z 577 y

87
m/z 287) permitió la confirmación de su identidad (214). El peak 6 correspondería a
rutina (fórmula, C27H30O16; masa nominal M6, 610 g/mol). Se observa en polaridad
positiva la señal m/z 611 y en polaridad negativa la señal m/z 609 que corresponden a la
forma protonada y desprotonada respectivamente. Además, se observan algunos
adúctos que confirman la masa principal. Las fragmentaciones observadas en polaridad
positiva y negativa confirman su identidad (216, 218).
Tabla 5. Datos obtenidos del análisis de APPE por HPLC-ESI/MS (polaridades positiva y negativa).
Compuesto Peak n°
tR (min)
MSn m/z (positivo)
MSn m/z (negativo)
ácido clorogénico 1 7.7 --- 353, 191, 135, 101 procianidina B1 2 8.5 --- 577, 425, 289 procianidina B2 3 9.4 579 577, 425, 407, 451,
289 (-) epicatequina 4 10.9 291, 123, 139, 165,
273 289, 245, 205, 137, 161
procianidina C1 5 11.8 867, 579, 713, 533, 695
865, 695, 577, 713, 287
quercetina-3-O-rutinósido 6 15.9 611, 303, 465, 449 609, 301, 343, 255, 179
quercetina-3-O-galactósido 7 16.7 465, 303 463, 301, 179, 151, 343
quercetina-3-O-glucósido 8 17.1 465, 303 463, 301, 179, 151 quercetina-3-O-pentósido 9 18.1 435, 303 433, 301, 151, 179 quercetina-3-O-pentosido 10 19.3 435, 303 434, 301, 151, 179 quercetina-3-O-ramnósido 11 19.7 449 447, 301, 179, 151,
271 floretina-O-xiloglucósido 12 20.5 ---- 567, 435, 273 floretina-2-O-glucósido 13 22.1 ---- 435, 273, 167, 301
El peak 7 correspondería a hiperósido (fórmula, C21H20O12; masa nominal M7, 464 g/mol).
Se observa la señal m/z correspondiente a la forma protonada (m/z 465) y la señal m/z
correspondiente a la forma desprotonada (m/z 463). Sus fragmentaciones en positivo y
negativo confirman su identidad (152, 215, 219-221). El peak 8 correspondería a
isoquercitrina (fórmula, C21H20O12; masa nominal M8, 464 g/mol). Se observan en
polaridad positiva y negativa las señales m/z 465 y m/z 463 respectivamente, además de
adúctos que confirman la masa. Sus fragmentaciones observadas en polaridad positiva y
negativa confirman su identidad (152, 218-220). Los peaks 9 y 10 corresponden a

88
quercetina-3-O-pentósido (fórmula, C20H18O11, masa nominal M9 y M10, 434 g/mol). Se
observan sus señales m/z 435 y m/z 433, además de adúctos que confirman la presencia
de la masa nominal 434. Sus fragmentaciones en polaridad negativa (m/z 301)
confirmarían sus identidades (222, 223).
Figura 13. Perfiles de RP-HPLC-UV y ESI/MS para APPE. (A) Perfiles HPLC de APPE registrados a 280 nm mostrando los peaks de (1) ácido clorogénico (2) procianidina B1, (3) procianidina B2, (4) (-)-epicatequina, (5) procianidina C1, (6) rutina, (7) hiperósido, (8) isoquercitrina, (9) quercetina-3-O-pentósido, (10) quercetina-3-O-pentósido, (11) quercitrina, (12) floretina-O-xiloglucósido y (13) floridzina. (B). Cromatograma de iones totales (TIC) en full scale y en el modo de iones negativo. (C), Cromatograma de iones totales (TIC) en full scale y en el modo de iones positivo.
El peak 11 correspondería a quercitrina (fórmula, C21H20O11; masa nominal M11, 448
g/mol). Se observan las señales m/z 449 y m/z 447 en polaridad positiva y negativa
respectivamente. La fragmentación en polaridad negativa confirma su identidad (213,
215). El peak 12 correspondería a floretina-O-xiloglucósido (masa nominal M12, 568

89
g/mol), sin embargo, no se observó las señales protonadas/desprotonadas ni adúctos que
confirmaran su masa. El peak 13 correspondería a floridzina (fórmula, C21H24O10; masa
nominal M13, 436 g/mol), en positivo se observan sólo señales de adúctos y en negativo
la señal m/z 436 correspondiente a la forma desprotonada. La comparación de las
fragmentaciones observadas para m/z 436 (m/z 273 y m/z 167) con literatura confirman su
identidad (216, 224).
6.5. Efecto del extracto de cáscara de manzana sobre la viabilidad de Helicobacter
pylori.
6.5.1 Determinación de la actividad anti-H. pylori de APPE mediante procedimientos
convencionales.
Los resultados presentados en esta sección se enmarcan en el Objetivo 2 de ésta tesis.
Se comparó la actividad anti-H. pylori de APPE, la fracción rica en material polimérico de
alto peso molecular (HMW), y la fracción de bajo peso molecular, obtenida desde el
extracto acetato de etilo (LMW). Interesantemente, la fracción de bajo peso molecular
(LMW; MIC = 89.5 g/mL) fue más efectiva contra H. pylori que la fracción polimérica
(HMW; MIC > 1500 mg/mL) (Tabla 6). Este resultado, al menos en parte, sugiere que la
actividad anti-H. pylori residiría en compuestos de bajo peso molecular. Como se
mencionó en la sección anterior, los componentes de bajo peso molecular (glicósidos de
quercetina principalmente) alcanzan una concentración de aproximadamente un 58 % del
total de polifenoles del extracto mientras que, la epicatequina y las procianidinas dan
cuenta de un 30 % del total de polifenoles. El valor de MIC para APPE fue de 112.5 g
GAE/mL para las dos cepas ensayadas. Este valor fue coincidente con el de la
concentración mínima bactericida (MBC). Sin embargo, el valor de MIC para APPE fue
más alto que aquellos determinados para los antibióticos amoxicilina y eritromicina. En el
presente estudio, los discos impregnados con 600 g de ácido tánico no produjeron halos
de inhibición en las dos cepas ensayadas.

90
Tabla 6. Efecto de APPE y sus fracciones sobre dos cepas de H. pylori.
H. pylori
ATCC 43504
H. pylori
TX136
H. pylori
ATCC 43504
H. pylori
TX136
Muestra
haloa
(mm)
haloa
(mm)
MIC 90
g/mL
MIC 90
g /mL
APPE 11 12 112.5b 112.5b
LMW 15 18 89.5b 89.5b
HMW Rc R >1500b >1500b
ácido tánico R R 900.0 900.0
amoxicilina 54 58 0.06 0.02
eritromicina 40 45 0.15 0.20
aEl halo de inhibición fue medido usando discos de 6 mm impregnados con 600 g de GAE
APPE,LMW, y HMW; 600 g de ácido tánico; 25 g de amoxicilina, y 15 g de eritromicina. bg
GAE/mL de APPE, cR = resistente.
6.5.2. Viabilidad de H. pylori en periodos cortos de contacto APPE-bacteria
utilizando como indicadores la reducción de resazurina y los niveles de ATP.
El uso del resazurina para la estimación de la viabilidad de H. pylori permitió realizar
curvas de titulación para las cuales la señal de fluorescencia del producto generado por
reducción del reactivo fue proporcional al número de UFC/mL (Figura 14 A y B). A partir
de estas curvas se seleccionó un inóculo de 107 UFC/ml para realizar los ensayos de
toxicidad temprana que se presentan en la Tabla 7. Las IC 50 obtenidas, para los tiempos
de 15, 30 y 120 min fueron de 63,6 g GAE/mL; 50,8 g GAE/mL y 32,8g GAE/mL,
respectivamente. La evaluación sólo se realizó hasta 2 horas de incubación, puesto que a
tiempos mayores de 4 horas la viabilidad de la bacteria se vio comprometida (< 80%) y no

91
fue posible obtener reproducibilidad en los resultados. Por lo anterior, no es posible
realizar una estimación de concentraciones MIC que pueda ser comparada con los
métodos convencionales recomendados para H. pylori. De este modo, la información que
entrega el método desarrollado sólo permite afirmar que el efecto bactericida o
bacteriostático de APPE se manifiesta tempranamente.
Figura 14. Titulación de Helicobacter pylori viables usando resazurina. (A) Distintas diluciones de un cultivo de 48 horas fueron hechas utilizando Caldo Brucella estéril. Los pocillos fueron sellados con una capa de aceite mineral e incubados durante 2 horas a 37 °C, con lectura constante de la fluorescencia generada por reducción del resazurina. (B) representación semi-log del número de bacterias versus señal de fluorescencia corregida a las 2 horas.
Tabla 7. Efecto de APPE sobre la viabilidad de H. pylori determinada mediante el ensayo de microtitulación de la reducción de resazurina.
Tiempo (min) IC50 g GAE/mL a
(g APPE/mL) b
15 63.6 (106.0)
30 50.8 (84.7)
120 32.8 (54.6)
a Contenido total de polifenoles determinado mediante el reactivo de Folin-Ciocaletau. b Peso seco de APPE

92
Adicionalmente, para comprobar el efecto temprano de APPE sobre H. pylori, se decidió
evaluar la viabilidad de éste último determinando sus concentraciones intracelulares de
ATP, después de 1 hora de incubación con APPE. Como se observa en la Figura 15 A,
APPE promueve una depletación de ATP en H. pylori en forma concentración-
dependiente. La concentración que provoca una disminución del 50 % en los niveles de
ATP fue aproximadamente 70 g GAE/mL. Este valor se encuentra cercano a las
concentraciones MIC y como se verá más adelante a las de IC50 para la actividad anti-
ureasa de H. pylori. Interesantemente, aunque se requirieron altas concentraciones de
APPE (1800 g/mL), fue posible suprimir completamente la síntesis de ATP. Si la
inhibición de la síntesis de ATP se debe a un efecto inhibitorio de la respiración celular por
parte de APPE y además involucra al pool insensible a lansoprazol, dicho efecto debería
verse reflejado en una disminución del consumo de oxígeno. En efecto, utilizando la
sonda MitoXpress, que es altamente sensible a la concentración de oxígeno se logró
detectar el consumo de oxígeno de títulos de H. pylori tan bajos como 102 UFC/mL. En la
Figura 15 B, se muestra el resultado de la aplicación de esta metodología en la detección
de la respiración de H. pylori viables desde 104 hasta 107 UFC/mL. Utilizando éste último
título (el mismo empleado en el experimento de niveles de ATP), se evaluó el efecto de
concentraciones de APPE similares a las usadas en los experimentos de MIC90 (25, 50,
75 y 150 g GAE/mL). Tal como se observa en la Figura 15 C, APPE inhibió el consumo
de oxígeno de H. pylori en forma-concentración dependiente. Dada la alta sensibilidad del
método se pudo obtener resultados en menor tiempo (24 horas) que con los métodos
convencionales, en los que se requiere de hasta 5 días de incubación.
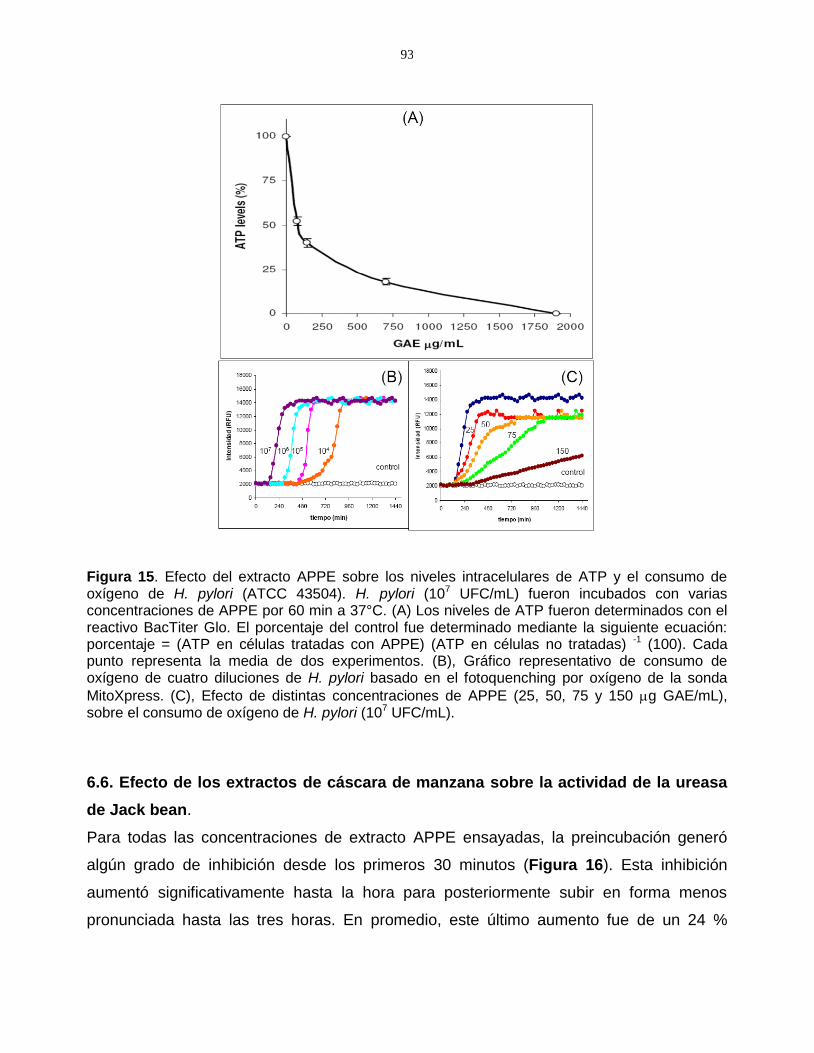
93
Figura 15. Efecto del extracto APPE sobre los niveles intracelulares de ATP y el consumo de oxígeno de H. pylori (ATCC 43504). H. pylori (107 UFC/mL) fueron incubados con varias concentraciones de APPE por 60 min a 37°C. (A) Los niveles de ATP fueron determinados con el reactivo BacTiter Glo. El porcentaje del control fue determinado mediante la siguiente ecuación: porcentaje = (ATP en células tratadas con APPE) (ATP en células no tratadas) -1 (100). Cada punto representa la media de dos experimentos. (B), Gráfico representativo de consumo de oxígeno de cuatro diluciones de H. pylori basado en el fotoquenching por oxígeno de la sonda
MitoXpress. (C), Efecto de distintas concentraciones de APPE (25, 50, 75 y 150 g GAE/mL), sobre el consumo de oxígeno de H. pylori (107 UFC/mL).
6.6. Efecto de los extractos de cáscara de manzana sobre la actividad de la ureasa
de Jack bean.
Para todas las concentraciones de extracto APPE ensayadas, la preincubación generó
algún grado de inhibición desde los primeros 30 minutos (Figura 16). Esta inhibición
aumentó significativamente hasta la hora para posteriormente subir en forma menos
pronunciada hasta las tres horas. En promedio, este último aumento fue de un 24 %

94
(rango 0 - 58.6%). Cuando se elaboró las curvas de concentración versus efecto
inhibitorio de la ureasa, se observo una concentración-dependencia para todos los
tiempos de preincubación ensayados (Figura 17). Las diferencias encontradas en las IC50
para cada tiempo de preincubación, sólo se hacen significativas si se comparan con el
tiempo cero (Tabla 8, IC50 = 85.3 g GAE /mL).
0
10
20
30
40
50
60
70
80
90
100
0 0,5 1 1,5 2 2,5 3
t (horas)
Inhib
ició
n (
%)
Figura 16. Efecto del extracto crudo de cáscara de manzana APPE, sobre la actividad ureasa de Jack bean como función del tiempo de preincubación. La actividad de la enzima se determinó posterior a la preincubación con el extracto de cáscara de manzana a tiempos 0, 0.5, 1, 2 y 3
horas. Las concentraciones finales de extracto empleadas fueron de 10 (), 20 (), 30 (▲), 60
(●), 150 (●), 300 (●) y 500 (┼) g GAE/mL.

95
0.0 0.5 1.0 1.5 2.0 2.5 3.00
25
50
75
100
0 hrs
0,5 hrs
1 hrs
2 hrs
3 hrs
Log[GAE] (g/mL)
% In
hib
ició
n
Figura 17 Gráficas semi-log de la concentración de APPE versus el efecto inhibitorio de la enzima ureasa de Jack bean a diferentes tiempos de preincubación.
Tabla 8. IC50 calculadas para cada periodo de preincubación del extracto APPE con ureasa Jack bean.
Tiempo (horas)
IC 50
(gGAE/mL)
0 85,3 0,5 55,7 1 54,8 2 44,0 3 41,5
6.7. Reactivación por DTT de la actividad ureolítica de la ureasa Jack bean pre-
incubada con extracto APPE.
En la Figura 18 se observa el efecto de la adición de DTT a la enzima previamente
incubada con tres niveles de concentración de APPE. Luego de adicionar el DTT la
actividad inhibitoria a las 2 horas decayó hasta un nivel similar al determinado a los 30
min. Este resultado sugiere por primera vez que un porcentaje de la población de tioles de
la enzima inactivada podría haber sido modificado por algunos componentes del extracto,
por ejemplo por oxidación de grupos –SH a especies que pueden ser revertidas por DTT

96
(-SOH, -SO2H y –SO3H). Según Krajewka et al., (225), si la oxidación progresa y se
generan los derivados sulfénicos, sulfínicos y sulfónicos no sería posible restaurar
completamente la actividad de la enzima. Adicionalmente, si los inhibidores son capaces
de generar la arilación de estos mismos tioles, el DTT tampoco puede revertir la
inhibición. En la Figura 18 se observa el comportamiento de las curvas de inhibición de
ureasa a las tres horas de preincubación, cuando se agrega DTT desde el comienzo
(tiempo = 0). Sólo para la concentración de 40 g GAE/mL de APPE, el DTT logró revertir
la inhibición en <10 %.
0
10
20
30
40
50
60
70
80
90
100
0 0,5 1 1,5 2
t (horas)
Inhi
bici
ón (%
)
Figura 18. Reactivación con DTT de la ureasa Jack bean inhibida por el extracto APPE. La ureasa
(0.4UI/mL final) fue incubada con tres niveles de concentración del extracto (40 ◆, 80 ■ y 160 ▲,
g GAE/mL), por 1 hora, después de lo cual se adicionó 800 M de DTT (final). La recuperación de la actividad fue determinada 1 hora después.
Adición de DTT

97
0
10
20
30
40
50
60
70
80
90
100
10 100 1000
gGAE/mL
Inh
ibic
ión
(%
)
Figura 19. Efecto del DTT sobre la inhibición de la actividad de ureasa por el extracto APPE. La actividad de ureasa fue ensayada a las tres horas después de la preincubación con el extracto y el
DTT. DTT M/mL: 0 (); 150 (X); 400 (■) y 800 (▲).
6.8. Influencia de los extractos de cáscara de manzana sobre parámetros cinéticos
de la enzima ureasa de Jack bean.
En la Figura 20C se representa las gráficas de Lineweaver-Burke para la enzima ureasa
Jack bean en ausencia y presencia de dos concentraciones del extracto de cáscara de
manzana (5 y 2.5 g GAE/mL final). De acuerdo a los antecedentes obtenidos de la
Figura 16, para el estudio cinético el tiempo de la preincubación se fijó arbitrariamente en
1 hora, que es el tiempo en que la inhibición comenzó a estabilizarse. El extracto mostró
un efecto inhibitorio de tipo competitivo, esto es con una modificación de la Km sin
inhibidor (0.77 g/mL) y sin cambios en la Vmáx. Las concentraciones de extracto de 2.5
y 5 g GAE/mL generaron un cambio en la Km de 2.5 y 3.3 gGAE/mL, respectivamente
(Figura 20 C).

98
6.9. Efecto de APPE sobre la actividad de la ureasa extraída de Helicobacter pylori.
El efecto inhibitorio de la ureasa de H. pylori fue ensayado tanto para APPE como para las
fracciones obtenidas según el esquema de fraccionamiento de la Figura 5. En la Figura
20 se observa que la inhibición de la actividad de ureasa de H. pylori es dependiente de la
concentración de extracto APPE. Dado que la inhibición de la ureasa de H. pylori por
parte de los extractos se manifiesta tempranamente, al igual que la observada con la
ureasa de Jack Bean, las curvas presentadas en la Figura 20 A-B, fueron obtenidas sin
pre-incubación. Para APPE la IC50 de 516 g/mL fue aproximadamente 6 veces superior a
la obtenida previamente para la ureasa de Jack Bean (IC50 = 180 g/mL). Este valor
puede ser explicado por la presencia de otras proteínas que acompañan a la ureasa en la
preparación enzimática (VacA, Hsp60). Estas proteínas de H. pylori, potencialmente
ofrecen sitios de unión adicionales para los polifenoles. Los resultados hasta aquí
presentados no permiten descartar que la inhibición de la ureasa por parte del extracto
pueda ser consecuencia de una interacción inespecífica. Como la baja concentración de
proteína no permite observar un precipitado visible que refleje un fenómeno de agregación
de la ureasa, fue necesario determinar qué componentes polifenólicos del extracto APPE
están asociados a la inhibición de ureasa.

99
Tabla 9. Contenido total de polifenoles, grado promedio de polimerización y actividad inhibitoria de la ureasa de cáscara fresca de manzana, APPE y sus fracciones de alto y bajo peso molecular.
Extracto/Fracción TPC (mg GAE/g
dw) a
mDPb IC50 Jack Bean
ureasac
(g/mL)
IC50 H. pylori ureasac
(g/mL)
Cáscara de manzana fresca
3,50 ± 0.8 2,9±0,2 Nd Nd
APPE 610 ±12 3,0± 0,5 180 516
LMW 480 ± 8 1,0 ±0,2 594 800
HMW 520 ± 10 9,5± 2 103 119
a Valores promedio ± S.D. (n = 3) expresados en miligramos equivalentes de ácido gálico (GAE) por gramo de material seca (cáscara o extracto seco). b Los números representan la relación entre los moles de los aductos flavan-3-oles-floroglucinol y los moles de las unidades monoméricas liberadas por depolimerización. c La actividad inhibitoria de la ureasa fue determinada después de la pre-incubación con el extracto o sus fracciones. Nd: No determinado.

100
Figura 20. Inhibición de ureasa por APPE, HMW y LMW. (A) Gráficos semi-log de la inhibición de la ureasa de H. pylori por los polifenoles de cáscara de manzana. (B) Gráficos semi-log de la inhibición de ureasa de Jack bean por los polifenoles de cáscara de manzana. APPE -o- ; HMW -
-; LMW -□- (C) Inserto: gráficos de Lineweaver-Burke mostrando el doble recíproco de la
actividad 1/ureasa y 1/substrato. Los datos en presencia de 0(); 2,5 (▲) and 5 (■) GAE (g/mL) de APPE son presentados como el promedio de cuatro determinaciones.
0.5 1.5 2.5 3.50
10
20
30
40
50
60
70
80
90
100
log [GAE] (g/mL)
Inh
ibitio
n (
%)
-2.0 -1.5 -1.0 -0.5 0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5 4.0 4.5 5.0 5.5 6.0 6.5 7.0
1
2
3
0 GAE (g/mL)
2.5 GAE (g/mL)
5.0 GAE (g/mL)
1/urea (M)
1/V
max (
mo
l/N
H4/m
in/m
g)
0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5 4.00
10
20
30
40
50
60
70
80
90
100
logGAE (g/mL)In
hib
itio
n (
%)
(A) (B)
(C)

101
Así, HMW y LMW fueron ensayadas mediante el método de rojo fenol para evaluar su
efecto sobre la actividad de ureasa de H. pylori. En la Figura 20 A y B se presenta las
curvas concentración-efecto para los extractos aludidos, en comparación con el extracto
de origen (APPE). Las concentraciones IC50 para HMW; LMW y APPE fueron 119; 800 y
516 g GAE/mL, respectivamente. Estos resultados sugieren que la principal contribución
al efecto inhibitorio de la actividad ureasa de H. pylori, estaría dada por la presencia de los
polifenoles de alto peso molecular y en menor grado a los polifenoles monoméricos. La
misma relación entre el grado de polimerización y la actividad inhibitoria se puede
observar para la ureasa de Jack Bean. Las tres curvas de inhibición obtenidas para
APPE, HMW y LMW con ambas enzimas poseen una pendiente similar, lo que apoya la
hipótesis que se trataría del mismo tipo de compuesto. En el caso del extracto LMW no
fue posible ensayar a concentraciones mayores debido a limitaciones en la solubilidad. En
la Tabla 9 se resume la relación existente entre el grado de polimerización determinado
por floroglucinólisis para APPE, HMW y LMW y la inhibición de la ureasa.
6.10. Detección “in-gel” de la actividad anti-ureasa de Jack bean.
En la Figura 21 A se observa el resultado de la electroforesis en geles de poliacrilamida
de muestras de ureasa incubadas con las fracciones HMW y LMW, en las mismas
condiciones del experimento de la sección 6.9. Las muestras fueron diluidas antes de ser
cargadas en el gel, de manera de tener 3 g de proteína por pocillo. Las fotografías se
tomaron a los 10 min de la aparición de las bandas correspondientes a la ureasa control.
La electroforesis en geles de poliacrilamida de los lisados de H. pylori se realizó sin
preincubación con los extractos, debido a que en ensayos previos, la inhibición de la
ureasa se observó tempranamente.
Las 2 bandas con actividad ureolítica observadas corresponden al hexámero (545 kD) y el
trímero (272 kD) que normalmente se disocian sin perder su actividad. Como se observa
en la Figura 21 A, los componentes de HMW y LMW producen una inhibición aparente de
la actividad ureasa. Con este experimento no puede descartarse la posibilidad de que, en
presencia del extracto, la enzima realmente haya sufrido una modificación de su
movilidad electroforética agregándose sin poder ingresar al gel. En efecto, en los pocillos

102
correspondientes a las concentraciones mayores de HMW, se observa actividad ureásica
en el agregado proteico. Por lo tanto, el resultado de esta evaluación no se correlaciona
con lo observado en los experimentos de inhibición de la actividad ureasa por los
métodos del rojo fenol y fenol-hipoclorito (Berthelot). En la Figura 21 A, la presencia de
los oligómeros de procianidinas en la fracción HMW es predominante a diferencia de
LMW, donde éstos últimos sólo se encuentran presentes en trazas. Sin embargo, al
realizar la detección con azul de Coomassie se comprobó que la disminución de la
actividad se debe principalmente a que hay menos ureasa. También se observa que el
extracto afecta a las otras bandas de proteínas presentes en el lisado de dos cepas de H.
pylori.
Figura 21. Efectos de APPE y sus fracciones sobre la actividad de ureasa detectados “en-gel”. Electroforesis en gel bajo condiciones no-denaturantes de la ureasa extraída de H. pylori en poliacrilamida al 7 %. (A) Antes de la electroforesis, las fracciones LMW y HMW fueron incubadas con la ureasa de H. pylori tal como se describe en materiales y métodos. Carril 1 = ureasa control;
Carriles 2, 3 y 4 = LMW = HMW = 100-400-800 GAE g/mL. (B) Efectos de APPE y sus fracciones sobre la actividad de la ureasa de H. pylori utilizando el procedimiento modificado en-gel. Para detectar el efecto de APPE, LMW y HMW sobre la actividad de la ureasa de H. pylori en geles no-denaturantes, se utilizó una membrana de agarosa-urea-rojo de cresol. Los mismos geles teñidos con azul de Coomassie son presentados en el panel inferior. Las figuras presentadas son representativas de tres experimentos independientes.

103
Respecto de los resultados de este experimento se debe precisar las diferencias
metodológicas con la determinación en placas de 96 pocillos. En este último ensayo, la
enzima permanece continuamente expuesta a los polifenoles testeados. Por otro lado, la
evaluación de la actividad ureasa “en-gel” sugiere la posibilidad cierta que después de la
incubación de la enzima con los polifenoles, éstos puedan ser removidos desde los
complejos proteína-polifenol durante las etapas convencionales que siguen a la corrida
electroforética. Por ejemplo, la detección “en-gel” de la actividad enzimática requiere tener
el gel en un rango óptimo de pH entre 6-7 después de la corrida, por lo que éste debe ser
lavado exhaustivamente con un tampón acetato (pH 6.8), como condición necesaria que
permita la subsecuente adición del indicador rojo de cresol. Tal como se indicó, después
de llevar a cabo dichos procedimientos obligados, se observó que para los tres extractos
probados (APPE, HWM y LMW), la enzima sujeta a PAGE mostró, en todos los casos,
una actividad ureasa cualitativamente comparable a la mostrada por el control sin
inhibidor. En atención a tales consideraciones, no parece razonable comparar los
resultados “en-gel” con aquellos obtenidos en el método en pocillo. Dado que los
resultados en pocillo (cinética enzimática) sugiere una inhibición de tipo reversible y
competitiva (Figura 20), se concluye que la detección de la actividad inhibitoria de “en-
gel” no sería el mejor procedimiento para evaluar este tipo de inhibidores. Por lo tanto, se
procedió a modificar el protocolo original, para lo cual se sometió la preparación
enzimática (ureasa de H. pylori) a una separación por PAGE (no denaturante). Tiras de
gel, en los cuales la presencia de la actividad ureásica previamente fue establecida,
fueron inicialmente incubadas con concentraciones crecientes (expresadas como GAE) de
APPE, HMW y LMW, para luego ser removidas de esas soluciones. Tal como se muestra
en la Figura 5 (sección de Materiales y Métodos), las tiras obtenidas fueron
posteriormente cubiertas con una lámina de agarosa que contiene el sustrato (urea) y el
indicador rojo de cresol, lo que permitió revelar en forma semi-cuantitativa, la actividad
ureasa remanente en el gel. Con este procedimiento, esencialmente se evitó las etapas
de lavado del gel, referidas previamente como una de las posibles causas de la
reactivación de la ureasa. Como se observa en la Figura 21B, cuando se ensayó la
concentración de 10 g GAE/mL, HMW mostró un claro efecto inhibitorio sobre la ureasa

104
de H. pylori. Por otro lado, para la misma concentración, APPE solo mostró un grado de
inhibición moderado. Cuando la concentración fue elevada a 100 g GAE/mL ambas
preparaciones promovieron una inhibición casi total de la ureasa.
El efecto inhibidor de dichos preparados no es atribuible a las diferencias en la
concentración de proteínas cargadas en los geles (Figura 21 B, panel inferior). Los
resultados de la aplicación de este enfoque metodológico no sólo son complementarios a
los resultados de concentración-dependencia obtenidos previamente "en-pocillo", sino
también proporcionan pruebas de que el efecto inhibidor de los polifenoles presentes en
HMW y APPE es observable en una preparación de ureasa aislada mediante PAGE, y
que dicho efecto es todavía evidente cuando se ha quitado la solución que contiene los
polifenoles. Curiosamente, en este último ensayo, la concentración necesaria para lograr
la inhibición de la ureasa total fue sustancialmente menor que la estimada en el ensayo en
el pocillo. La observación que, en comparación con APPE, se requiere una concentración
menor de HMW para alcanzar un grado similar de inhibición de la ureasa (Figura 21 B),
es coherente con la propuesta de que las macromoléculas con un mayor grado de
polimerización (probablemente procianidinas), son las responsables de las propiedades
inhibitorias de la ureasa.
6.11. Efecto inhibitorio in vitro del extracto de cáscara de manzana (APPE) sobre el
estallido respiratorio de neutrófilos inducido por PMA, fMLP y H. pylori no-
opsonizado.
Los resultados de esta sección se enmarcan en el Objetivo 3. En la Figura 22 se observa
el efecto del extracto APPE sobre la producción intracelular de ERO inducida por PMA.
Dada la alta polaridad de los polifenoles de APPE, resulta intrigante el hecho que éstos
fueran capaces de inhibir la producción intracelular de ERO en forma concentración-
dependiente (Figura 23 A). PMA es un activador directo de las proteína kinasas (PKCc)
que no requiere unión a receptor. Como resultado de su activación, hay movilización de
calcio y ensamblaje del complejo NADPH oxidasa. Lo anterior genera primeramente ERO
al interior del neutrófilo y posteriormente un flujo hacia el exterior. Dado que en este

105
experimento la producción de ERO en el exterior esta suprimida por la mezcla SOD/CAT,
el efecto que se observa corresponde a la disminución de las ERO exclusivamente en el
interior del neutrófilo. El rango de concentración de APPE empleado, no comprometió la
viabilidad de los neutrófilos durante el tiempo que duró el ensayo, por lo que la
disminución de la producción de ERO es atribuible al efecto antioxidante de APPE.
Figura 22. Gráfico representativo del efecto de distintas concentraciones del extracto APPE en g GAE mL-1 sobre la producción intracelular de ERO en neutrófilos estimulados por PMA (200 nM). La quimioluminiscencia del luminol se amplificó con una mezcla de 50 U/mL de SOD y 2000 U/mL de CAT.
Al examinar la producción extracelular de ERO con la sonda impermeable isoluminol-HRP
se observó un efecto concentración-dependiente con una IC50 de 1.1 g/mL, la cual es
aproximadamente 30 veces inferior a la determinada para el extracto APPE cuando se
mide su efecto a nivel intracelular. Lo anterior puede deberse a que en neutrófilos
0 10 20 300
1000
2000
3000
PMA 200 nmol L-1 APPE 10 g GAE mL -1 + PMA APPE 20 g GAE mL -1 + PMA
APPE 40 g GAE mL -1 + PMA APPE 80 g GAE mL -1 + PMA APPE 160 g GAE mL -1 + PMA
time (min)
RL
U/m
in

106
humanos, PMA estimula la producción de ERO a nivel intracelular más que extracelular,
por lo tanto el desafió oxidativo para los polifenoles es menor en el exterior del neutrófilo
que adentro de éste (226).
Figura 23. Efecto de los APPE sobre la producción intracelular y extracelular de ERO en neutrófilos estimulados con PMA. Los neutrófilos aislados se diluyeron a 1 x 105 PMNL/pocillo fueron incubados durante 10 minutos con extracto APPE previo a la estimulación con PMA (200 nmol L-1). Las mediciones fueron realizadas mediante la amplificación de luminiscencia de luminol
(75 mol L-1) a 37 ºC. A y B gráficas semi-log de concentración de los extractos versus porcentaje de inhibición del estallido respiratorio inducido por PMA medidos por los sistemas lumninol/SOD/CAT (2000 y 50 U) e isoluminol/HRP (4 U) respectivamente.
1.5 2.5 3.5 4.50
25
50
75
100
log [GAE ng/mL]
Inh
ibic
ión
(%
)
1.0 1.5 2.0 2.50
25
50
75
100
log [GAE g mL-1]
Inh
ibit
ion
(%
)
B) Extracelular
IC 50 = 1.1 g/mL
A) Intracelular
IC 50 = 30 g/mL

107
(A) (B)
Figura 24. Efecto de los extractos de cáscara de manzana sobre la producción extracelular de ERO en neutrófilos estimulados con fMPL. (A) Los neutrófilos aislados fueron incubados durante 10 minutos con luminol, HRP (4 U) y con el extracto de cáscara de manzana previo a la estimulación con fMLP. Las mediciones fueron realizadas mediante la amplificación de
luminiscencia de isoluminol (75mol L-1) a 37 ºC. (B) Gráfica semi-log de concentración de los extractos en ng GAE mL-1 versus el porcentaje de inhibición del estallido respiratorio inducido por fMPL.
Como se observa en la Figura 24 (A), la máxima señal de luminiscencia inducida en los
neutrófilos por el activador fMLP alcanzó 1200 RFU/sec (menos de la mitad de la
observada a nivel intracelular). No obstante, aunque marginal, la producción extracelular
de ERO estimulada por fMLP, pudo ser amplificada utilizando el sistema isoluminol-HRP.
La Figura 24 (B), muestra que el extracto APPE es capaz de inhibir la producción
extracelular de ERO con una IC 50 de 64 ng/mL. Esta baja concentración inhibitoria
calculada para APPE, sugiere un efecto directo sobre la unión del fMLP a su receptor de
0 1 2 3 40
25
50
75
100
log [ng GAE mL-1]
Inh
ibic
ión
(%
)
IC 50 = 64 ng mL-1
Extracelular
7ng/mL 14 ng/mL 37ng/mL 59 ng/mL 75 ng/mL 149 ng/mL 235 ng/mL 530 ng/mL 2571 ng/mL
control
0 5 10 150
250
500
750
1000
1250
1500
time (min)
RF
U

108
transmembrana, alterando los eventos de señalización que llevan a la activación de la
NADPH oxidasa, como por ejemplo la activación de PKC.
Figura 25. Efecto de la primación de neutrófilos con Helicobacter pylori sobre la producción
intracelular de ERO mediada por PMA. Los neutrófilos (1 x 105 /100 L por pocillo) fueron incubados en presencia o ausencia de H. pylori (107 y 105) durante 15 minutos y posteriormente activados con PMA (10 nmol L-1), en presencia SOD/CAT.
Se sabe que ciertos factores solubles del H. pylori son capaces de primar a los neutrófilos
haciéndolos más sensibles al estímulo con agentes como el PMA y el fMLP para la
generación de ERO y quimioquinas (199, 227, 228). Para comprobar la funcionalidad de
los neutrófilos, y si H. pylori es capaz de generar un estallido respiratorio mayor, el efecto
de primación ocasionado por incubación con H. pylori no opsonizado es presentando en la
Figura 25. En efecto se observa que al aumentar la MOI, el estallido respiratorio aumentó
en amplitud y se hizo más rápido.
(A)
0 10 20 30 40 500
500
1000
1500
2000
2500
3000
3500
4000
4500
time (min)
RF
U
PMA
HP 10 7
HP 10 5
PMA

109
(B)
0 1 2 30
25
50
75
100
log[g GAE mL-1]
Inh
ibit
ion
(%
)
Figura 26. Efecto de los extractos de cáscara de manzana sobre la producción de quimioluminiscencia inducida por H. pylori no-opsonizado en neutrófilos. (A). Determinación de la quimiluminiscencia en neutrófilos estimulados por H. pylori no-opsonizado a través de los sistemas luminol/SOD/CAT (intracelular) e isoluminol/HRP (extracelular). (B) Gráfica semi-log para los sistemas luminol/SOD/CAT (triángulos cerrados) e isoluminol/HRP (triángulos abiertos). Los neutrófilos aislados se diluyen a 1 x 10 5 PMNL/pocillo fueron incubados durante 10 minutos con extracto de cáscara de manzana previo a la primación con H. pylori (30 min; 1 x 108 UFC mL-1). Los datos corresponden al promedio de tres experimentos independientes ± SD realizados en triplicado.

110
Tabla 10. Resumen de los resultados obtenidos en la evaluación del efecto antioxidante de APPE en neutrófilos activados por H. pylori (ATCC 43504), fMLP and PMA.
Estímulo Luminol-SOD/CAT
IC50 g/mLa
(µmol/L)b
Isoluminol-HRP
IC50 g/mLa
(µmol/L)b
H. pylori 31.4 (29.5) 7.1 (6.7)
fmLP Nd 0.064 (0.061)
PMA 30.0 (26.9) 1.1 (1.0)
a Concentración de polifenoles totales calculados como equivalentes de ácido gálico (g GAE/mL), determinados según el método de Folin-Ciocalteau.
b Concentración de flavonoides totales expresada como equivalentes de rutina (mol RutE/L), determinados por HPLC-DAD.
Nd. No detectado.
En la Figura 26 A-B se observa el efecto inhibitorio concentración-dependiente del
extracto APPE sobre la producción intracelular y extracelular de ERO estimulada por H.
pylori no opsonizado. La IC50 de 31.4 g/mL fue similar a la encontrada para PMA. Por
otro lado, APPE fue capaz de inhibir en forma concentración-dependiente la producción
extracelular de ERO en neutrófilos estimulados por H. pylori, con un valor de IC50 de 7.1
g/mL. En ausencia de estímulo, APPE no indujo la producción de ERO en neutrófilos
(datos no presentados) ni comprometió la viabilidad celular. Como se muestra en la Tabla
10, los valores de IC50 obtenidos con los tres activadores fueron similares a aquellos
publicados previamente en términos de su equivalencia en rutina (RutE mol/L), cuando
otros autores han ensayado con polifenoles puros (229-231).
6.12. Efecto inhibitorio de APPE sobre la actividad de la toxina VacA en células
HeLa.
Los resultados de esta sección se enmarcan en el Objetivo 4 de la tesis. La citotoxina
VacA induce la formación de canales específicos que en la membrana plasmática de
eucariontes permiten el paso de aniones, esto último se asocia con la liberación de
bicarbonato y otros iones orgánicos, los que facilitan la multiplicación adecuada de la

111
bacteria (26). Los polifenoles extraídos de diferentes fuentes pueden actuar como agentes
anti-VacA, lo que se ha demostrado por una disminución de la vacuolación en células
susceptibles como HeLa y AGS (112, 166). El extracto APPE enriquecido en polifenoles
fue capaz de inhibir la vacuolación en células HeLa tratadas con sobrenadantes de H.
pylori (500 g proteinas/mL). Dicha inhibición fue concentración-dependiente con una IC50
de 450 g GAE/mL (Figura 27 A-E). Tal como se muestra en las fotografías
representativas de la Figura 27 A-D, las células HeLa tratadas con los sobrenadantes de
H. pylori e incubadas por 16 horas en presencia de APPE presentaron un índice de
vacuolación significativamente inferior (P < 0.05) al de las células tratadas sólo con el
sobrenadante, y ninguna diferencia con el control no tratando. Los resultados anteriores
sugieren que, en el rango de concentraciones utilizado, el efecto vacuolante de VacA
puede ser contrarestado por APPE. Puesto que algunos flavonoides como la rutina y
quercetina se les atribuyen un cierto efecto antivacuolante, para tratar de explicar tal
inhibición, se estimó la concentración de equivalentes de rutina en APPE a partir de los
datos de RP-HPLC-DAD. Así, la concentración de rutina equivalente a la IC 50 de APPE
fue de 28 M. Como se explicará en la discusión de esta tesis, dicha concentración, por si
sola no sería suficiente para sustentar la inhibición de la vacuolación observada. Además,
la concentración de quercetina en APPE determinada por RP-HPLC-DAD resultó
indetectable. Sin embargo, frente a la posibilidad que los glicósidos de quercetina sufran
hidrólisis durante su permanencia en el medio ácido gástrico, no se puede descartar que
“in-situ” se logren concentraciones suficientes de quercetina que inhiban la vacuolación.
Como una forma de predecir esta potencial liberación de quercetina, se incubó APPE
durante 4 horas en una solución que simuló el medio ácido gástrico. Tal como se observa
en la Figura 28 A, el monitoreo de la estabilidad de los glicósidos de quercetina indicó
que después de 4 horas la hidrólisis sólo afectó en un 3 % a dichos flavonoides,
encontrándose 0.20 M de quercetina al final de la incubación, la cual fue identificada por
su patrón de fragmentación mediante LC-MS (Figura 28 C). Por lo tanto, es posible que la
quercetina liberada desde APPE, por si sola no de cuenta del efecto anti-vacuolante de
APPE. Así, es probable que otros compuestos de APPE, como las procianidinas y las
chalconas contribuyan a dicho efecto tanto in vitro como in vivo. Al respecto, la principal

112
chalcona identificada en APPE es la floridzina (un glucósido de floretina), la cual se
encuentra en una concentración equivalente a la IC 50 de APPE de 117 M. Ésta última,
es aproximadamente la mitad de la IC 50 necesaria para inhibir la acción de VacA
reportada previamente para floridzina pura (35). Como se indicó anteriormente, algunos
polifenoles como las procianidinas pueden unirse a VacA para neutralizar sus efectos. En
APPE, las procianidinas dan cuenta de aproximadamente el 24 % del total de polifenoles
presentes en APPE. Previamente, Yahiro et al., (166) demostraron que es éste el tipo de
compuestos al unirse eficientemente a VacA forman un complejo que impide su
interacción con los receptores de membrana RPTP y RPTP. Dicho mecanismo
representa una forma distinta de neutralizar a la toxina VacA que se sumaría a los efectos
de los polifenoles descritos anteriormente. En el estudio, fue evaluada una serie de
extractos ricos en polifenoles, dentro de los cuales el más efectivo fue el extracto de
brácteas de Lúpulo. En efecto, 10 g/well (en un volumen final 100L) de éste extracto
disminuyeron la vacuolación de células susceptibles en un 90 %. En el mismo estudio, 10
g/well (volumen final 100 L) de un extracto de manzana disminuyó la vacuolación en un
50 %. En el presente estudio, dicha cantidad se alcanza cuando se utiliza
aproximadamente 100 g de polifenoles totales (GAE). Tal concentración que no fue
capaz de inhibir la vacuolación en células HeLa (Figura 27 E). La razón para esta
discrepancia de valores puede estar en parte en las diferentes condiciones
experimentales y aspectos asociados al modelo celular empleado. No obstante, el
extracto APPE usado en el presente estudio posee un mayor contenido de flavonoides
glicósidos (58 %) que el de procianidinas (24 %), las cuales además poseen un grado de
polimerización promedio DPm = 3. Interesantemente, las procianidinas de APPE se ven
afectadas por las condiciones ácidas del estómago, sufriendo una depolimerización que
alcanza a un 27 % a las 4 horas (Figura 27 B). Así, con los resultados presentados en
este estudio, no es posible predecir completamente si el efecto del pH sobre la estabilidad
de las procianidinas de APPE tendrá un impacto negativo sobre su actividad neutralizante
de VacA in vivo.

113
Figura 27. Inhibición de la toxicidad (vacuolación) de VacA sobre células HeLa. Fotografías representativas de: (A), Células HeLa incubadas sin sobrenadantes de H. pylori; (B) tratadas con
sobrenadantes de H. pylori; (C) tratadas con sobrenadantes de H. pylori y 100 g GAE/mL de
APPE; (D) tratadas con sobrenadantes de H. pylori y 500 g GAE/mL de APPE; (E) índices de
vacuolación en células HeLa tratadas con los sobrenadantes de H. pylori y 100 - 550 g GAE/mL de APPE. Inserto: gráfico lineal del porcentaje de vacuolación versus la concentración de APPE. Los valores son expresados como el promedio ± S.D. (n = 3); * P<0.05 (versus control). Barra = 10
m.

114
0 min
240 min
time (min)
0
10
20
30
40
50
60
70
80
90
100
0 50 100 150 200 250
time (min)
hydro
lysis
(%
)
0
10
20
30
40
50
60
70
80
90
100
0 50 100 150 200 250
time (min)
De
po
lim
eri
za
tio
n (
%)
(A) (B)
(D) (E)
(C)
UV Chromatogram, 355 nm
1
2
3
4
5
6
7 8
1
2
3
4
5
6
7 8

115
Figura 28. Estabilidad de los polifenoles de APPE en condiciones ácidas que simulan el medio gástrico. (A), análisis de los glicósidos de flavonoides presentes en APPE por RP-HPLC-DAD antes (medio) y después (fondo), después de 4 horas de incubación en fluído gástrico artificial a 37 °C. El cromatograma de arriba corresponde a un estándar de quercetina. Los cromatogramas RP-HPLC-DAD fueron registrados a 355 nm mostrando los peaks 1-8, cuya identidad corresponde a la señalada en la Tabla 5. (B), Los flavan -3-oles y procianidinas de APPE fueron analizados por NP-HPLC-FLD, antes (trazado negro) y después (trazado en gris) de 4 horas de incubación en fluido gástrico artificial a 37 °C. Los cromatogramas NP-HPLC-FLD fueron registrados a una longitud de onda de excitación de 230 nm y una de emisión de 321 nm. (C), Espectro de masa de quercetina (peak 8) in modalidad de iones positivos (m/z 302). (D) y (E), degradación de glicósidos de quercetina y procianidinas de APPE en ambiente de acidez gástrica simulada.
6.13. Efecto de APPE sobre la adhesión de H. pylori a cultivos de células HeLa.
La adhesión del H. pylori a las células HeLa fue evaluada por microscopia óptica y usando
la actividad residual de la ureasa, después de la incubación con APPE. Como se muestra
en la Figura 29 A-D, APPE fue capaz de de disminuir la adhesión bacteriana con una IC50
= 5.3 mg GAE/mL. Contrario a lo que pudiera pensarse, la actividad residual de ureasa no
es un reflejo de las propiedades inhibitorias de APPE sobre dicha enzima. Dado que la
actividad inhibitoria de APPE es reversible y competitiva, las etapas de lavado exhaustivo
de la células HeLa previamente al ensayo, deberían bastar para remover cualquier traza
de APPE. Para confirmar esto último, se determinó el grado adsorción inespecífica de
APPE a las membranas de células HeLa y el posible ingreso de los polifenoles a estas
últimas. La presencia de polifenoles se analizó mediante RP-HPLC-DAD, tanto en los
pellets celulares como en los lisados, no encontrándose trazas detectables de los
constituyentes característicos de APPE (datos no presentados). Estos resultados sugieren
que en los cultivos celulares, la actividad residual de ureasa refleja solamente aquellas
bacterias que permanecen adheridas. Interesantemente, la propiedad anti-adherente
observada para los polifenoles de APPE impide la interacción directa entre H. pylori y la
mucosa del huésped, lo cual contribuiría a evitar el efecto deletéreo no sólo de VacA sino
que también de otras toxinas que requieren contacto directo como es el caso de CagA.

116
Figura 29. APPE inhibe la adherencia de H. pylori a células HeLa. La actividad de la ureasa residual fue determinada por el método del rojo fenol-urea. Imágenes representativas de: (A), células Hela sin H. pylori; (B) con H. pylori; (C) con H. pylori y 5 mg GAE/mL de APPE; (D) con H. pylori y 10 mg GAE/mL of APPE. (E), Propiedades anti-adhesivas, después de la preincubación con APPE (0.03-10 mg GAE/mL) y H. pylori seguido de la adición a las células HeLa. El control positivo sin inhibidor fue usado para establecer el 100 % de adhesión. Inserto: gráfico lineal del porcentaje de adhesión versus la concentración de APPE. Los resultados son expresados como al
media ± SEM (n = 3); * P<0.05 (versus control). Bar = 10 m.

117
6.14. Efecto de APPE sobre la adhesión de H. pylori a la mucosa gástrica de ratones
C57BL/6.
Los resultados presentados en esta sección se enmarcan en el Objetivo 5. En
concordancia con los resultados reportados previamente en otros modelos murinos (232,
233), en el modelo experimental de infección con H. pylori a corto plazo empleado en esta
tesis, se encontró que las bacterias se localizaron preferentemente en el antro pilórico en
lugar del cuerpo (resultados que se desprenden del análisis cuantitativo por PCR en
tiempo real). La presencia de H. pylori tanto en el cuerpo como en el antro, también fue
confirmada paralelamente por cultivo tradicional (durante 7 días). Durante todo el periodo
experimental (28 días, datos no presentados), no se observó indicios de toxicidad aguda o
pérdida de peso significativa. En las Figuras 30 A-E se muestra imágenes
representativas de la detección de H. pylori mediante la tinción de Warthin-Starry (W-S),
para el antro pilórico de animales no infectados (30 A), infectados (30 B) y tratados con
APPE (30 C-D). Utilizando esta tinción, el H. pylori se observa como un bacilo curvado de
color marrón, localizado en la superficie de la mucosa epitelial, a menudo asociado con
mucus y restos de alimento (30 B). Por lo tanto, con el modelo de infección a corto plazo
fue posible limitar el asentamiento de aquellas bacterias localizadas principalmente en la
superficie de la mucosa antral. Como se mencionó anteriormente, para alcanzar un alto
grado de colonización probablemente se requiera de un modelo de infección a largo
plazo. Dichos modelos habitualmente requieren el uso de cepas de H. pylori adaptadas a
murinos, como es el caso de la cepa de Sydney 1 (SS1), la cual coloniza en forma
eficiente y masiva el estómago del ratón C57BL/6, llegando a producir niveles apreciables
de gastritis, similares a los observados en humanos (234). Por ejemplo, en estos animales
se ha reportado que después de un seguimiento post-infección de hasta 12 meses, la
infección se consolida y llega incluso a promover el desarrollo de adenocarcinoma (235).
El análisis por PCR en tiempo real de las muestras de estómagos de animales tratados
con APPE (150 y 300 mg/kg/d) no mostró diferencias significativas comparadas con el
grupo de animales control no tratados (Figura 30 F). Estos resultados están en
concurrencia con la reducción de la carga de H. pylori observada en la evaluación

118
histológica. Como se observa en el detalle de la Figura 30 E, las bacterias en su mayoría
se ubican en la superficie epitelial.
Adicionalmente, los resultados fueron corroborados en las muestras obtenidas desde los
animales control y aquellos tratados con APPE. Después de 7 días de cultivo no fue
posible recuperar bacterias con la morfología típica de H. pylori. Como se muestra en la
Figura 31 A, los índices de gastritis sugieren que APPE protege significativamente la
mucosa gástrica de los ratones infectados con H. pylori. Sin embargo, en la Figura 31 B
se observa que entre los diferentes grupos de animales, los estómagos no presentaron
signos macroscópicos de deterioro o injuria. Además, el examen histológico de las
porciones antrales de los dos grupos tratados con APPE (Figuras 31 A-B) no mostró
diferencias significativas con los estómagos de animales infectados y no infectados. En
las Figuras 31 C-D se presenta la imagen representativa de la porción antral de un ratón
en el cual excepcionalmente se desarrolló un grado notorio de infiltración leucocitaria. En
relación con esto último, nuestro protocolo de infección a corto plazo permite reproducir
las primeras etapas del proceso de colonización por H. pylori. Por lo tanto, el score de
gastritis básicamente esta influenciado por la densidad de bacterias adherentes en la
mucosa. En efecto, como se mostró en la Figura 30 B, después de 28 días post-infección,
H. pylori permanece adherido a la superficie epitelial pero, la mucosa no evidenció signos
macroscópicos de inflamación. Además, en todos los grupos de ratones, hubo ausencia
de infiltración leucocitaria y de bacterias adheridas al epitelio del cuerpo estomacal. Esto
último es coherente con el hecho que en nuestro modelo, la infección por H. pylori
selectivamente se localizó en el antro.

119
Figura 30. APPE impide la colonización de la mucosa gástrica de ratones C57BL6/J por H. pylori. (A), Imagen representativa de un corte histológico de la porción antral de un ratón no infectado utilizando la tinción de Warthin-Starry de. (B), antro de un ratón infectado por H. pylori después de 28 días post-inoculación, observándose bacterias en la superficie del epitelio gástrico (flecha). (C-D), porción antral de ratones tratados con 150 y 300 mg/kg/día de APPE, respectivamente. (E), Detalle de un foco de infección (flecha) en el antro de un ratón infectado (tinción H&E). (F), Detección cuantitativa de H. pylori mediante PCR en tiempo real en ratones infectados, y tratados
con 150 y 300mg/kg/día de APPE durante 20 días. Barra= 100 m.

120
Figura 31. (A), Índice de gastritis en ratones infectados con H. pylori y tratados con APPE 150 mg/kg/día y 300 mg/kg/día (n = 7). (B), Apariencia macroscópica de los estómagos de ratones después del tratamiento con APPE. (Barra = 1cm).
6.15. Efecto de APPE sobre los niveles de MDA en la mucosa gástrica de ratones
infectados por H. pylori.
Pese al corto periodo de interacción huésped-patógeno y al bajo índice de gastritis
observado para los animales infectados, en la Figura 32 se observa que en éstos, los
niveles de MDA detectados en el antro fueron significativamente mayores al de los
controles no infectados (ANOVA, P < 0.001). En las etapas iniciales de la colonización por

121
H. pylori, los niveles de MDA son un reflejo de cambios oxidativos que ocurren como un
evento primario que conduce a daño a la mucosa (66). En esta fase el extracto APPE tuvo
un efecto protector altamente significativo (P = 0.001) sobre los animales infectados con
H. pylori. Como era de esperar, no se observó diferencias significativas para los niveles
de MDA en el cuerpo de animales infectados.
Figura 32. APPE inhibe la formación MDA y la inflamación en el estómago de ratones infectados con H. pylori. (A), niveles de MDA en muestras de antro de ratones infectados y no infectados. (B), secciones representativas (antro) teñidas con H&E de una mucosa gástrica normal de un animal no infectado y no tratado con APPE. (C), Imagen representativa (H&E) mostrando una mucosa (antro) de un ratón infectado con H. pylori y no tratado con APPE. Se observa una leve Infiltración leucocitaria con neutrófilos y células mononucleares. (D), Organización linfo-folicular (flechas) en la submucosa antral del mismo animal infectado. (E), Detalle de neutrófilos PMNL
(cabezas de flecha) observados en el mismo ratón infectado con H. pylori (Barra = 100 m)

122
7. DISCUSIÓN
Los aportes más relevantes de esta tesis son: a) Se obtuvo un extracto de cáscara de
manzana (APPE) utilizando agua como solvente y un proceso de enriquecimiento de sus
polifenoles mediante el uso de una resina adsorbente (Sepabeads SP-850). Dicha
estrategia permitió conservar el perfil polifenólico observado en la materia prima fresca a
la forma de un extracto seco y estable. La caracterización química del extracto se llevó a
cabo exitosamente a través de la aplicación de procedimientos de separación
cromatográfica (HPLC y TLC) utilizando diferentes formas de detección (UV, DAD, FLD,
ESI-MS/MS y revelado con DMACA) y comparación con muestras auténticas que sirvieron
como estándares de los polifenoles; b) El extracto APPE mostró propiedades bactericidas
y bacteriostáticas sobre dos cepas de H. pylori. Dicho efecto se relacionó con la presencia
de polifenoles de bajo peso molecular (LMW) los cuales disminuyeron la viabilidad de la
bacteria en forma concentración y tiempo-dependientes; c) La fracción de alto peso
molecular (HMW) mostró un potente efecto inhibitorio de la actividad de la ureasa de H.
pylori así como, de la ureasa de Jack bean. Dicho efecto fue concentración-dependiente,
reversible y competitivo; d) El extracto APPE fue capaz de inhibir la producción de
especies reactivas del oxígeno (ERO) por parte de neutrófilos estimulados con H. pylori
no-opsonizado, PMA y fMLP. Se determinó que tal efecto estaría asociado a los
compuestos de bajo peso molecular (principalmente glicósidos de quercetina) los cuales
inhibieron en forma concentración-dependiente la producción tanto intra como extracelular
de radicales libres; e) En cultivos de células HeLa, APPE fue capaz de inhibir en forma
concentración-dependiente el efecto vacuolante de la toxina VacA de H. pylori; f) APPE
tuvo un efecto inhibitorio moderado sobre la adherencia de H. pylori a cultivos de células
HeLa; y g) En un modelo de infección a corto plazo, utilizando ratones C57BL/6J, la
administración oral durante 20 días de dos dosis de APPE, interfirió con las etapas
iniciales de la colonización por la bacteria y tuvo un efecto protector sobre daño
inflamatorio a la mucosa gástrica de los animales inoculados.

123
7.1. Obtención y caracterización del extracto de cáscara de manzana (APPE).
El proceso de obtención de extractos polifenólicos a partir de materiales vegetales como
el residuo agro-industrial empleado en esta tesis (cáscara de manzana), a menudo
contempla las etapas de extracción con solventes orgánicos, concentración y secado.
Alternativamente, el uso de enzimas como pectinasas y celulasas ha demostrado su
utilidad en mejorar el tenor de extracción de polifenoles (236). Ambas estrategias tienen
ciertas desventajas. Se destaca el hecho que el empleo de enzimas y solventes aumenta
sustancialmente el costo del proceso. Además, en ciertos casos los solventes representan
un impacto negativo sobre el medioambiente y/o limitan la aplicación final del producto,
toda vez que se debe garantizar la ausencia de residuos de aquellos que no son
aceptados como sustancias GRAS (Generally Recognized As Safe= Aceptados
Generalmente Como Seguros). El problema descrito anteriormente ha motivado la
constante búsqueda de nuevas estrategias tecnológicas que permitan la obtención de los
extractos polifenólicos en ausencia de solventes u otros agentes químicos. Al respecto, se
ha propuesto procesos alternativos como son el empleo de fluidos supercríticos, agua en
estado sub-crítico o enriquecimiento mediante “foaming-fractionation” (237-240). La
mayoría de estos procesos requiere una infraestructura sofisticada y personal técnico
altamente entrenado. Sumado a lo anterior, los rendimientos obtenidos normalmente no
superan a los que se reportan para los procesos convencionales y a veces llevan a
productos con perfiles químicos distintos a los observados en la materia prima original.
Esto último no es trivial si se piensa en las potenciales aplicaciones de estos extractos en
la industria de alimentos, donde no siempre se desea que el proceso de extracción sea
demasiado selectivo. De tal modo, es probable que durante el proceso de extracción se
concentre algunos metabolitos secundarios a tal nivel que potencialmente aparezcan
efectos biológicos impredecibles. En el presente estudio, el extracto de cáscara fresca de
manzana de la variedad Granny Smith posee un perfil polifenólico en el cual abundan los
flavonoides (glucósidos de quercetina) por sobre los flavan-3-oles y procianidinas. Dicho
perfil se conservó después de las etapas de extracción acuosa y adsorción con la resina
Sepabeads SP-850. La información recogida de la literatura señala que, con excepción de

124
ciertas variedades, la situación habitual es encontrar predominio de los flavan-3-oles y
procianidinas por sobre el nivel de los glucósidos de quercetina (147, 152, 176, 241). Sin
embargo, la explicación más lógica para el origen de esta diferencia con lo señalado en la
literatura reside en el hecho que la composición química de los frutos (particularmente de
polifenoles antioxidantes) varía considerablemente de región en región. Al respecto, Chile
posee ciertas ventajas derivadas de su particular clima y terroir que constituyen elementos
determinantes en la producción de elevados niveles de antioxidantes polifenólicos. Esto
es evidente para ciertas especies que se cultivan en otras regiones del planeta.
Comparativamente, el más claro ejemplo de tal situación es el de Vitis vinífera (242-244),
y que se ha reflejado en el creciente interés por los vinos Chilenos, por parte de los
consumidores extranjeros. Desde el punto de vista analítico, los resultados de la
composición cualitativa estuvieron dentro de lo esperado. Tal como se señaló
anteriormente, APPE posee glicósidos de quercetina y procianidinas como metabolitos
secundarios predominantes. La comparación con estándares y el uso de técnicas
acopladas contribuyó substancialmente a identificar gran parte de dichos polifenoles.
Dada la falta de estándares certificados de alta pureza, la caracterización de las
procianidinas requiere de técnicas adicionales. Así, la degradación utilizando agentes
nucleofílicos permitió no sólo obtener información relativa al grado de polimerización de
APPE (DPm = 3) sino que además, confirmar que en este extracto, tales procianidinas
están formadas principalmente por epicatequina. Sin embargo, llama la atención que la
tiólisis con cisteamina genere el aducto de catequina. Debido a que la catequina
originalmente está presente sólo en trazas en APPE y la cáscara fresca de la manzana
Granny Smith, los resultados sugieren que en el proceso de tiólisis, la totalidad de la
catequina podría originarse a partir de unidades terminales. Dicha conclusión podría ser
errónea debido a que la cisteamina, a pesar de sus ventajas analíticas (baja toxicidad),
promueve la epimerización de epicatequina a catequina (180). Por otro lado, en la tiólisis
realizada con tolueno–tiol, la tendencia a la isomerización de epicatequina es menor y
los aductos pueden resolverse de mejor manera. Sin embargo, dichos aductos son
bastante lipofílicos y eluyeron a tiempos de retención muy elevados (> 1h). Por el
contrario, los aductos generados con el nucleófilo floroglucinol eluyen a tiempos de
retención más cortos. A diferencia de la tiólisis, la floroglucinólisis además resultó ser el

125
procedimiento de elección dado que el reactivo posee baja toxicidad. Para completar el
análisis de APPE quisimos obtener información sobre la distribución oligomérica de las
procianidinas (hasta los undecámeros), la cual no puede inferirse de los resultados de la
depolimerización. Para tal fin se aplicó NP-HPLC-FLD en fase diol, una herramienta que
poco a poco ha desplazado el uso de fases normales de silicagel. En efecto se confirma
que APPE posee procianidinas oligoméricas distribuidas entre los dímeros y undecámeros
y con pocos indicios de material polimérico. Este último generalmente eluye como un
“cerro” pobremente resuelto al final del cromatograma obtenido por NP-HPLC-FLD. La
aparición de picos múltiples en este tipo de trazado, corresponde a verdaderos “clusters”
de procianidinas. Lo anterior indica la co-existencia de diferentes rotámeros asociados a
cada oligómero, tal como lo han descrito otros autores (174, 175, 245, 246). Tal aspecto
podría ser relevante para explicar los efectos anti-ureasa y anti-VacA de APPE y que
serán discutidos en las secciones ulteriores.
7.2. Los polifenoles de APPE poseen un efecto anti-H. pylori asociado con su
tamaño molecular.
7.2.1. Efecto de los polifenoles de APPE sobre la viabilidad de H. pylori.
Los polifenoles de APPE afectaron la viabilidad de H. pylori en forma concentración-
dependiente. Además, la viabilidad fue disminuyendo a medida el aumentó el tiempo de
contacto entre APPE-H. pylori. Más aún, se observó que gran parte de la actividad anti-H.
pylori reside en la fracción de bajo peso molecular. Estos hallazgos, en conjunto con los
datos de literatura sugieren claramente que los glicósidos de quercetina presentes en
APPE serían los responsables de la actividad anti-H. pylori, puesto que los flavan-3-oles
como la epicatequina poseen una actividad bactericida moderada. Por ejemplo,
previamente se reportó una MIC90 para epicatequina de 800-1600 g/mL, mientras que
para el galato de epicatequina (EGCG) este valor fue de 83-167 g/mL (128). Entre los
requisitos estructurales que determinan un aumento considerablemente de la actividad
anti-H. pylori de los polifenoles, se puede mencionar a la presencia de un grupo metoxilo
en C-4´ en las flavanonas. Se ha establecido que la presencia azúcares en C-3 y de
hidroxilos adicionales en el anillo B afecta negativamente la actividad bactericida de
dichos compuestos. Así por ejemplo, se ha descrito que las flavanonas ponciretina (4'-

126
metoxi-5,7-dihidroxiflavanona); hesperitina (3',5,7-Trihidroxi-4'-metoxiflavanona), y
naringenina (4',5,7-trihiroxiflavanona), poseen MIC de 10, 20 y 40 g/mL contra H. pylori,
respectivamente (247). Utilizando una aproximación experimental similar a la de la
presente tesis (viabilidad por resazurina), (248) otros autores estudiaron el efecto de
baicaleina (flavona) y ácido glicirretínico (sapogenina), extraídos de la regaliz (Glicirriza
glabra = Orozuz = Licorice). Ambos compuestos mostraron un marcado efecto inhibitorio,
cuando en el ensayo se incorporó ácido L-láctico como inductor de multiplicación para H.
pylori. Así, en el caso de baicaleina, el 100% de inhibición se alcanzó con
concentraciones entre 0.5-1.0 g/mL mientras que, en ausencia de L-láctico, las
concentraciones necesarias fueron siempre mayores a 100 g/mL. Lo anterior permite
explicar en parte por qué en nuestras condiciones experimentales (medio sin L-láctico),
las concentraciones necesarias para inhibir la proliferación de H. pylori fueron mayores
que en el ensayo turbidimétrico.
La tiempo-dependencia observada en este ensayo, podría estar asociada a la polaridad
de los constituyentes de APPE. Esta propiedad es determinante para su permeabilidad en
la membrana bacteriana. La mayoría de los polifenoles identificados en APPE posee una
baja solubilidad en lípidos, sin embargo, si el tiempo de contacto es suficiente, el ingreso
de algunos flavonoides derivados de quercetina y chalconas podría verse favorecido. En
efecto, numerosos estudios realizados con flavonoides, concluyen que la mayor
solubilidad en lípidos se asocia con un efecto bactericida más potente (101, 112, 249-
252). Recientemente, se ha propuesto un nuevo mecanismo mediante el cual ciertos
flavonoides de baja polaridad pueden ejercer un efecto anti-H. pylori. Así, la actividad
bactericida de quercetina, apigenina y (S)-sakuranetina, ha sido atribuida a su capacidad
de inhibir en forma competitiva a la dehidratasa HpFabZ (253). La principal función de
HpFabZ es participar en la fase de elongación durante la biosíntesis de ácidos grasos
saturados e insaturados (254). Dicha proteína también ha sido identificada como el blanco
de otros productos naturales como la juglona (hidroxi-1,4-naftoquinona), la cual es capaz
de inhibirla en forma competitiva (255, 256).
Cómo una manera de entender de qué forma APPE afecta la viabilidad de H. pylori, se
observó que sus constituyentes provocaron una disminución de la síntesis de ATP. En un
estudio similar, Nagata et al. (257), reportaron el efecto concentración-dependiente del

127
lansoprazol sobre los niveles de ATP en H. pylori. Sin embargo, aun cuando la actividad
respiratoria fue suprimida completamente a partir de concentraciones de 100 g/mL de
lansoprazol, no fue posible disminuir los niveles de ATP más allá de un 20 %. Los autores
concluyeron que existe una fracción de ATP insensible a lansoprazol y que
probablemente no esté ligado a la actividad respiratoria de H. pylori. En la presente tesis,
el efecto temprano del extracto APPE sobre la viabilidad de H. pylori también fue
comprobado mediante la caída de los niveles de ATP, luego de un periodo de incubación
de 1 hora. Adicionalmente, al comparar el consumo de oxígeno del control incubado en
ausencia de APPE, la mayor concentración utilizada (150 g GAE/mL), a las 24 horas
inhibió la respiración en aproximadamente un 80 % (P < 0.05). Dicha concentración está
muy cerca de la determinada como MIC90 por el ensayo turbidimétrico.
Si bien es cierto el efecto anti-H. pylori de los flavonoides es considerado moderado,
recientemente éstos se han utilizado como templado para el desarrollo de nuevos
derivados con mayor potencia. Al respecto, un derivado resultante del acoplamiento entre
antibiótico-antioxidante (metronidazol-genisteína) mostró una actividad anti-H. pylori muy
potente (MIC = 0.39 g/mL) (258).
7.2.2. Efecto de los polifenoles de APPE sobre la actividad ureásica de H. pylori.
La estructura, número y tipo de de subunidades, peso molecular y secuencia
aminoacídica de la ureasa depende de su origen (259). Las ureasas bacterianas
contienen tres subunidades, α, β y γ. Sus pesos moleculares están en el rango de 68–
73 kDa (α), 8–17 kDa (β) y 8–12 kDa (γ). Las ureasas bacterianas son principalmente
contienen tres subunidades: (αβγ)3. En contraste, la ureasa de H. pylori posee sólo dos
tipos de subunidad: α (60–66 kDa) y β (26–31 kDa). Sin embargo, los elementos
estructurales básicos son trímeros: (αβ)3 los cuales forman un complejo tetraédrico con
una estequiometría ((αβ)3)4, y cuyo peso molecular es de 1100 kDa (260, 261). La ureasa
vegetal extraída de Jack bean es una molécula homohexamérica (α6). La subunidad
contiene 840 aminoácidos y su peso molecular es de 90.77 ± 0.05 kDa (262).
Independiente de sus diferencias estructurales, las secuencias aminoacídicas de los sitios
activos son básicamente similares en todas las ureasas conocidas. La consecuencia de
ese hecho es que el mecanismo catalítico es el mismo. El sitio activo esta siempre

128
localizado en la subunidad α y contiene un centro catalítico con dos átomos de níquel (II)
separados por una distancia de 3.5 a 3.7 Å (263). En la ureasa de H. pylori (forma no
inhibida), ambos átomos están coordinados por His136, His138, Kcx219, His248, His247,
Asp362 y moléculas de agua, que se emplea en la reacción de hidrólisis.
Como el mecanismo de catálisis es el mismo, APPE pudo inhibir a la ureasa obtenida de
dos fuentes, Jack bean y H. pylori. Para ambas se observó una inhibición de tipo
concentración-dependiente, tiempo-independiente, reversible y competitiva. En similares
condiciones, otros autores (183) han observado que normalmente algunos inhibidores
como el ácido acetohidroxámico (AHA), pueden alcanzar hasta un 80 % de inhibición,
después de una hora de pre-incubación. En contraste, los mismos autores observaron
que el efecto inhibitorio de algunas -hidroxicetonas sintéticas presenta una mayor
dependencia de la preincubación, alcanzando grados de inhibición entre un 70 y 90 %
recién a las tres horas. No obstante esta similitud en cuanto al porcentaje de inhibición,
las -hidroxicetonas requieren al menos 30 a 60 min de preincubación, mientras que el
APPE manifiesta un efecto inhibitorio muy rápido. Lo anterior sugeriría que algunos
componentes del extracto interaccionan mediante mecanismos adicionales que llevan a la
inhibición más eficaz de la enzima. Las ureasas son enzimas ricas en tioles. Por ejemplo,
el contenido total de tioles en la ureasa Jack bean ha sido estimado en 15 grupos por
subunidad. Sin embargo, solo 6 de las 15 cisteínas son accesibles a reactivos selectivos
para ellos. Uno de esas 6 cisteínas, Cys592, está localizada en el brazo móbil que cierra el
sitio activo de la ureasa. Esta cisteína juega un papel crucial en la actividad catalítica
(264). Recientemente, una serie de estudios han demostrado cómo la presencia de
ciertos grupos funcionales como los quinónicos y tiosulfinatos en los inhibidores
irreversibles de ureasa, afecta el grado de oxidación de los tioles de la ureasa Jack bean
(265-269). En determinadas condiciones, los compuestos que presentan estos grupos
funcionales, pueden inducir la oxidación de los tioles de la ureasa de una forma similar al
efecto oxidante de H2O2. La reversibilidad del efecto inhibitorio de APPE por DTT fue
limitada, lo que hace suponer que la oxidación de tioles no es un mecanismo relevante
para los polifenoles. En efecto, la modificación química de Cys592 resulta en inactivación
de la enzima. Sin embargo, la modificación covalente de las otras 5 cisteínas, las cuales
son más reactivas que Cys592, no afectan la actividad de la enzima.

129
Por otro lado, la unión de los polifenoles a la ureasa podría llegar a afectar su actividad sin
alterar químicamente el centro catalítico. Bartolomé et al., (270) estudiaron el efecto de la
unión de distintos polifenoles con BSA sobre el perfil de elución cromatográfica (Sephadex
G-50). Los investigadores observaron un aumento significativo en el volumen de elución
de los complejos BSA-polifenol, en particular con las formas poliméricas que poseen
grupos –OH en posición orto. La posibilidad de un aumento en el peso molecular por
unión de los polifenoles a la ureasa fue considerada al interpretar los primeros resultados
obtenidos con los geles no-denaturantes. Cuando se empleó polifenoles monoméricos
puros (rutina, quercetina, catequina y ácido cafeíco) en el rango de concentración 20 a
8500 g/mL, no se observó efecto inhibitorio sobre la ureasa de H. pylori. Por el contrario,
la fracción de alto peso molecular (HMW) mostró un elevado efecto inhibitorio formando
agregados inactivos con la enzima. Por otro lado, la fracción de bajo peso molecular
mostró una actividad inhibitoria significativamente menor que la HMW. El nuevo
procedimiento de detección “en gel”, permitió comprobar que tal interacción inhibidor-
enzima tiene como consecuencia la inactivación de la ureasa y que un exceso de sustrato
puede regenerar en gran parte dicha actividad. Si la inactivación de la ureasa
comprometiera la estabilidad de sus tioles (por ejemplo, la Cys592), la actividad no habría
podido ser recuperada. Por lo tanto, en lo que respecta al efecto sobre la ureasa de H.
pylori podemos afirmar que éste residiría en los oligómeros y polímeros (procianidinas) de
APPE.
7.3. APPE inhibe la producción de especies reactivas del oxígeno en neutrófilos
activados por PMA, fMLP y H. pylori no-opsonizados.
APPE fue capaz inhibir la producción intracelular de ERO en neutrófilos activados por
PMA. Cualquiera sea el caso (captación directa de las ERO, la inhibición de las PKC u
otras enzimas), se requeriría del ingreso de los polifenoles al neutrófilo o alguno de sus
metabolitos generados durante el estallido respiratorio (productos de oxidación y/o
hidrólisis). Otra conjetura que se desprende del método utilizado, es que al incluir
SOD/CAT en el sistema se disminuye la depleción de antioxidantes polifenólicos de APPE
en el medio extracelular con lo que aumenta su probabilidad de ingreso al neutrófilo
(Figura 33, número 4). Por otro lado, el fMLP representa una forma más fisiológica de

130
inducción del estallido respiratorio en comparación con el PMA. El fMLP es un péptido que
en el neutrófilo humano estimula la producción de ERO principalmente a nivel intracelular
(226, 271), mediante su interacción con un receptor acoplado a proteína G (FPRs). La
evaluación del efecto inhibitorio de la producción de ERO por parte de los polifenoles,
utilizando el modelo celular del presente trabajo, fue tempranamente reportado por
Limasset (272). En dicho estudio se realizó un análisis multivariado que comprendió la
evaluación de 34 polifenoles sobre la producción de ERO en neutrófilos humanos
estimulados por PMA, fMLP y zimozano opsonizado (OZ). Como resultado, se concluyó
que dependiendo del tipo de estímulo para la producción de ERO, la inhibición mostrada
por ciertos polifenoles puede incluso llegar a ser opuesta, lo que supone diferentes
mecanismos de acción de dichas sustancias. Así por ejemplo, el origen de dicha
dicotomía pareciera emerger cuando se compara moléculas apolares como la crisina,
chalcona, flavona y galangina, con moléculas glicosiladas en la posición C-7 (hesperidina
y naringina) y monohidroxiladas en el anillo B. En otro trabajo, Krol et al., (273) estudiaron
14 flavonoides, confirmando que el grupo hidroxilo en posición C-3, dos grupos hidroxilos
en el anillo B y la presencia de un doble enlace entre C-2 y C-3, son de vital importancia
para la inhibición de la producción de ERO en neutrófilos. Los autores también sugirieron
que la liposolubilidad sería uno de los factores preponderantes en la actividad antioxidante
de los flavonoides. Posteriormente, Limasset y colaboradores (274), evaluaron un grupo
de 18 polifenoles constituido por flavonoides y algunos de sus metabolitos putativos como
lo son los ácidos fenólicos. Estos últimos mostraron una actividad inhibitoria de la
producción de ERO (inducida por PMA y fMLP) menos potente y específica que los
flavonoides parentales. El efecto de cuatro flavonoides (miricetina, quercetina, canferol y
galangina), sobre la producción de ERO en neutrófilos de conejo mediante la estimulación
de dos tipos de receptores del complemento (FcγR, CR) también ha sido reportado (275).
Interesantemente, la inhibición de la producción de ERO estuvo directamente asociada a
la liposolubilidad, inversamente correlacionada con el grado de hidroxilación del anillo B y
resultó ser independiente del tipo de receptor estimulado. En varios de los estudios
mencionados y otros más recientes, se han reportado similares hallazgos para algunos de
los mismos flavonoides presentes en el extracto APPE (135, 275, 276).

131
Figura 33. Efecto antioxidante de los polifenoles de APPE en neutrófilos estimulados por H. pylori. (1), APPE puede actuar uniéndose a macromoléculas de H. pylori involucradas en la inducción del estallido respiratorio. (2), APPE inhibe a la MPO y/o capta el HOCl producido en el medio extracelular. (3), APPE inhibe la producción de anión superóxido generado externamente por la NADPH oxidasa ensamblada en forma anómala en la membrana del neutrófilo. (4), Constituyentes de APPE y/o sus metabolitos inhiben la producción intracelular de APPE ingresando al neutrófilo o interfiriendo con la activación de receptores de membrana asociados con la fosforilación de proteínas efectoras (PKC).
En forma paralela a su capacidad de atrapar superóxido, algunos flavonoides como la
quercetina y luteolina, inhiben en forma concentración-dependiente, la fosforilación de
proteínas que sigue a la estimulación de los neutrófilos con fMLP (Figura 32, número 4)
(135). No obstante, el fMLP es utilizado para activar la respuesta de neutrófilos de la
misma manera que las bacterias, se debe considerar que en la interacción H. pylori-
neutrófilo, los receptores de fMLP tipo 1 (FPLR1) no son los únicos involucrados (228).
Satin et al. (277), reportaron que la proteína HP-NAP es uno de estos factores que
estimula a la NADPH oxidasa en neutrófilos aunque en una forma más tardía que fMLP.
Específicamente los autores observaron que la máxima producción de ERO se alcanzó
entre los 40 - 60 minutos después de la incubación. Muy importante es el hecho que H.
pylori pueda evadir el efecto bactericida de las ERO al estimular en forma sostenida la

132
producción extracelular de ERO en neutrófilos, ocasionando daño al huésped (61). Lo
anterior se explicaría por el hecho que H. pylori, provoca un ensamblaje anómalo del
complejo NADPH oxidasa en la membrana celular y no en el fagosoma. Específicamente,
mientras el flavocitocromo b558 puede ser adquirido por los fagosomas, las sub-unidades
p47phox o p67phox no pueden ser reclutadas eficientemente (61). Los factores que
determinan este tipo de evasión no están del todo identificados y sólo se sabe que son
moléculas ubicadas en la superficie de H. pylori, sensibles al calor y resistentes al
tratamiento con formalina. En el presente estudio se utilizó la cepa ATCC 43504, la cual
posee los factores activadores de neutrófilos, LPS, ureasa, HP-NAP y HP (2-20). Sin
embargo, se ha establecido que estos últimos factores no son requeridos para la estimular
el estallido respiratorio de neutrófilos inducido por H. pylori. De igual manera, el resultado
de investigaciones preliminares indican que las toxinas VacA y CagA tampoco son
necesarias para la activación de neutrófilos (278). Otros autores sugieren que la adhesina
SabA podría cumplir un rol preponderante como factor activador de neutrófilos por parte
de H. pylori (279). Por lo tanto, el hecho que APPE pueda inhibir la producción intracelular
de ERO, no necesariamente sugiere que los polifenoles podrían ingresar al neutrófilo, sino
que también podrían bloquear algunos de los factores presentes en H. pylori involucrados
en el la inducción del estallido respiratorio (Figura 33, número 1).
Como se señaló anteriormente, la producción extracelular de ERO inducida por H. pylori
resulta ser de mayor relevancia en el origen del daño tisular. Por lo tanto, se determinó el
efecto de APPE sobre la producción extracelular utilizando la quimioluminiscencia de
isoluminol amplificada por HRP. En el sistema isoluminol-HRP la producción de ERO se
relaciona directamente con la actividad de la NADPH oxidasa, y por lo tanto a la cantidad
de anión superóxido generado. Como se indicó previamente, debido a la alteración en la
localización de la NADPH oxidasa, el flujo extracelular de ERO inducido por H. pylori sería
en parte responsable del daño a la mucosa gástrica (Figura 33, número 3). APPE posee
una importante concentración de glicósidos de quercetina (58 % del total de polifenoles).
Los principales flavonoides identificados en APPE fueron la rutina, hiperósido,
isoquercitrina, y quercitrina. Tal como se indicó anteriormente, la presencia de grupos
fenólicos en C-5, C-7, C-3´ y C-4´ de los derivados de quercetina favorece la actividad
antioxidante. Por otro lado, para estos glicósidos no se ha podido establecer si la

133
sustitución con diversos azúcares en C-3 posee algún efecto sobre la capacidad de inhibir
la producción ERO en neutrófilos activados con H. pylori. El HOCl es la principal ERO
producida fisiológicamente por los neutrófilos a partir del peróxido de hidrógeno formado
por autodismutación del anión superóxido. El HOCl es capaz de reaccionar rápidamente
con tioles como la cisteína y el glutatión, así como con aminoácidos azufrados como la
metionina (280-282).
Interesantemente, la sustitución glicosídica en C-3 no parece afectar la capacidad
captadora de HOCl por parte de los flavonoides como sí ocurre con la capacidad de
atrapar anión superóxido. Más aún, dicha substitución es un requisito indispensable para
la captación eficiente de esta ERO (283). Así, si bien el OH en C-3 es fundamental para
la inhibición de la xantina oxidasa y la captación de anión superóxido, la presencia de un
azúcar en dicha posición para la rutina, acelera el grado de cloración del flavonoide
haciéndolo mejor sustrato que la genina libre (quercetina). En el caso de la rutina, lo
anterior se traduce en una elevada efectividad para captar el HOCl, con una IC50
reportada de 0.60 mol/L. En APPE los glicósidos de quercetina representan
aproximadamente un 58 % del total de polifenoles. La IC50 expresada en equivalentes de
rutina y que fue observada para la estimulación con H. pylori fue de 6.7 mol RutE/L.
Considerando los datos de la Tabla 4, del total de glicósidos de quercetina, la rutina
representa el 6.59 %. Como los valores de la Tabla 10 están expresados en equivalentes
de rutina, al recalcular basándose sólo en el aporte de rutina, la IC50 obtenida ahora es de
0.44 mol. Dicho valor está en el mismo orden que lo reportado en literatura y sugiere que
gran parte del efecto antioxidante observado en el estallido respiratorio inducido por H.
pylori, se debe a la capacidad de los flavonoides glicosídicos de APPE para neutralizar el
HOCl.
7.4. APPE inhibe el efecto vacuolante de VacA en células HeLa.
La inhibición de la vacuolación inducida por los sobrenadantes de H. pylori sobre células
HeLa se atribuye a la presencia de los compuestos de APPE (IC50 de 450 g GAE/mL).
Cuando se utilizó concentraciones de APPE mayores a 600 g GAE/mL la viabilidad
celular comenzó a verse comprometida y el índice de vacuolación inducida por los

134
sobrenadantes de H. pylori fue aumentando, llegando incluso hasta un 200 % con 7000
g GAE/mL (Datos no presentados). Lo anterior podría deberse a la aparición de efectos
citotóxicos ligados a la presencia de altas concentraciones de los polifenoles y que han
sido reportados anteriormente (284-286). El efecto de los polifenoles sobre la actividad de
VacA se ha tratado de explicar mediante dos mecanismos. El primero de ellos se basaría
en la interacción directa VacA-polifenol, lo que impediría la integración de la toxina a la
membrana plasmática. Lo anterior se ha demostrado para las procianidinas oligoméricas
de lúpulo y vino tinto, las cuales fueron capaces de unirse a VacA e inhibir sus efectos
tanto in vitro como in vivo (130, 167). El grado de polimerización de las procianidinas es
un aspecto estructural que resulta central para explicar la capacidad de neutralizar la
toxina VacA. En efecto, Yahiro et al. (166), demostraron que las procianidinas con mayor
tamaño molecular son más eficientes como neutralizadoras de VacA. Por ejemplo, los
polifenoles de alto peso molecular obtenidos de brácteas de lúpulo, fueron
aproximadamente 5 veces más efectivos que aquellos presentes en la fracción de bajo
peso molecular obtenida del mismo órgano de la planta. Para la fracción de alto peso
molecular del extracto de brácteas de lúpulo se ha reportado estructuras con niveles de
polimerización promedio DPm = 10 – 30. Al respecto, en esta tesis se determinó para
APPE un DPm = 3, lo que sugiere que existiría otros compuestos actuando mediante
mecanismos adicionales. Así, es posible que los glicósidos de quercetina y otros
polifenoles de bajo peso molecular también interfieran con la vacuolaciación inducida por
la toxina VacA. No obstante, algunos estudios han permitido demostrar que dentro de los
polifenoles, los glicósidos de flavonoides poseen un efecto moderado sobre la actividad
de VacA. Así, por ejemplo se ha reportado que la IC50 de rutina necesaria para inhibir la
vacuolación en células HeLa fue mayor que 400 M, mientras que para quercetina este
valor fue de 46 M (112). De igual manera, el efecto de los flavonoides sobre la actividad
de VacA medida en términos de su capacidad de inhibir la formación de canales y el flujo
de urea en células HeLa ha sido descrito en detalle (35). Respecto de esto último, en el
caso de la rutina, una concentración de 200 M sólo fue capaz de inhibir en un 15 % el
flujo de urea, mientras que la formación de canales inducida por VacA fue inhibida por
rutina con una IC50 = 106 M. Coherente con los hallazgos de Shin et al., (112), para

135
quercetina, el valor reportado de IC50 fue de 19 M, lo que sugiere que la glicosilación
pudiera tener un efecto negativo sobre la propiedad de los flavonoides de inhibir los
efectos funcionales de VacA. Al estudiar la potencial liberación de quercetina desde el
pool de glicósidos presentes en APPE, se encontró que a las 4 horas sólo un 3 % de
éstos sufre hidrólisis. Similares resultados fueron reportados por Goh et al., para
glicósidos de flavonoides de Gingko biloba (202). Lo anterior sugiere que, en ausencia de
alimentos, los flavonoides glicósidos presentes en APPE pueden mantenerse estables
hasta las 4 horas. No obstante la baja concentración de quercetina liberada, no se puede
descartar que ésta pueda actuar en forma aditiva con las procianidinas inhibiendo el
efecto vacuolante de VacA. Tampoco se debe descartar la presencia de chalconas como
la floridzina. Para ésta última, y aunque es un polifenol minoritario, se calculó un valor
equivalente en APPE de IC 50 = 117 M. Según lo reportado en literatura (35), si se
considera el aporte de la floridzina como molécula aislada, con dicha concentración se
podría lograr una inhibición de un 25 %. A diferencia de las actividades discutidas en las
secciones 7.2 y 7.3, los resultados de este ensayo sugieren que, por lo menos en lo que
respecta a la propiedad anti-VacA de APPE, los tres principales grupos de polifenoles que
contribuyen serían las procianidinas, flavonoides y chalconas.
7.5. APPE interfiere en el proceso de adhesión de H. pylori a cultivos de células
HeLa.
La adhesión de H. pylori a las células epiteliales del huésped, además de constituir la
primera etapa de colonización y ser un requisito para iniciar el proceso de infección,
también puede servir para inducir la expresión de otros factores de virulencia bacteriana.
Así, se ha reportado que alrededor del 50 % de la toxina VacA producida H. pylori
permanece inserta en la membrana plasmática de la bacteria, por lo tanto dicha fracción
puede inducir la formación de canales y vacuolas en forma más eficiente que la fracción
secretada al medio extracelular (20). Por tal motivo, resulta especialmente relevante que
APPE pueda limitar la adhesión de H. pylori. No obstante, en nuestro modelo no se puede
descartar del todo, que el efecto anti-adherente sea en parte consecuencia de la actividad
anti-microbiana y anti-ureasa de APPE. Este último aspecto fue descartado analizando la
presencia de polifenoles en los pellets celulares y sus lisados. Se observó que la pre-

136
incubación de H. pylori con 5 mg GAE/mL de APPE impidió la adhesión de la bacteria
después de 1 h de incubación con las células HeLa. Tal concentración es bastante mayor
que las observadas en los ensayos anteriores lo que sugiere que el efecto anti-adherente
de APPE es menos relevante que las actividades anti-ureasa y antioxidantes. En efecto,
algunos autores sugieren que la actividad anti-adherente de determinados extractos se
asocia a la presencia de compuestos de alto peso molecular como polisacáridos, y en
menor grado a las procianidinas (122, 287-292). Tal como se ha indicado anteriormente,
en APPE la concentración de este tipo de compuestos (polisacáridos y procianidinas) es
menor que la de los glicósidos de flavonoides. Más aún, los oligosacáridos que dan
cuenta del 40 % del peso seco de APPE corresponden a azúcares neutros (Tabla 3).
Respecto de esto último, se ha descrito que los polisacáridos ácidos (ricos en ácidos
glucurónico y galacturónico), poseen propiedades anti-adherentes sobre H. pylori
significativamente mayores que los polisacáridos neutros (292-298) con IC50 del orden de
los 2.5 mg/mL.
7.6. APPE impide la colonización de H. pylori en ratones C57BL6J utilizados como
modelo de infección a corto plazo.
De acuerdo a los resultados obtenidos in vitro se puede inferir que en un modelo animal,
APPE puede actuar impidiendo la colonización bacteriana:
a) afectando directamente la viabilidad de la bacteria y por lo tanto, su motilidad.
b) alterando su capacidad de colonizar la mucosa gástrica a través de la inhibición de la
ureasa, bloqueo de adhesinas, o
c) citoprotegiendo la mucosa gástrica en forma directa o a través de la inhibición del daño
por las ERO generadas durante el proceso inflamatorio.
Por lo tanto, no es posible concluir que la ausencia de genes específicos para H. pylori en
los estómagos de los animales tratados con APPE solamente sean un reflejo de su
actividad bactericida. Las propiedades enumeradas podrían estar asociadas con el efecto
de varios componentes de APPE que estén actuando sinérgicamente. En esta tesis,
varias de esas propiedades han logrado ser ligadas a grupos específicos de los

137
polifenoles contenidos en APPE. Así por ejemplo, es claro que los componentes de alto
peso molecular de APPE contribuirán a evitar el asentamiento de H. pylori en la mucosa
gástrica, inhibiendo eficientemente a la ureasa y neutralizando a la toxina VacA (164,
299). En sintonía con los resultados de esta tesis, las propiedades anti-adhesivas y anti-
VacA de las mismas procianidinas extraídas de otras fuentes, también han sido
reportadas (130, 166, 167, 289).
Figura 34. En células de epitelio gástrico, la regulación hacia arriba de la expresión del supresor de la señalización por citoquinas (SOCS)-2 y SOCS-3 por prebióticos y probióticos contrarresta los efectos inflamatorios de H. pylori. H. pylori puede inducir tanto señales inflamatorias como anti-inflamatorias, cuyo balance puede ser importante en la mantención de la homeostasis. La IL-8,
iNOS, TNF- y la COX-2 son mediadores conocidos por ser transcritos a través de factores de
transcripción redox-sensibles, como las vías de transducción de señales del NF-B y las MAPK. En esta situación, los prebióticos y probióticos pueden ejercer acciones anti-inflamatorias a través de la activación de la señalización de SOCS llevando a la expresión temprana y robusta de SOCS-2 y SOCS-3, a la cual además contribuye la inactivación de JAK2. En conjunto, las señales anti-inflamatorias mediadas por la activación de SOCS pueden ser una de las formas en que los prebióticos y los probióticos protegen contra la infección por H. pylori. Adaptado de la referencia (291).

138
En el presente trabajo se estudió el efecto protector de APPE mediante la observación
histológica y la determinación del MDA como marcador de daño por lipoperoxidación.
Dado que la mayoría de los animales no presentaron signos de infiltración leucocitaria,
sólo la determinación de MDA por HPLC-FLD resultó lo suficientemente sensible para
detectar diferencias incipientes entre los grupos de animales infectados y los tratados con
APPE. La actividad gastroprotectora de un extracto de manzana había sido reportada
previamente (171) para animales en los cuales se indujo un daño indirecto a la mucosa
gástrica por indometacina. Como principal conclusión, los autores asociaron dicha
protección a las propiedades antioxidantes del extracto de manzana. Por otro lado Yahiro
et al., (166), demostraron que el efecto inhibitorio sobre la ulceración e inflamación
inducida por la administración oral de VacA en ratones, se debería a la capacidad anti-
VacA de un extracto de lúpulo. Igualmente, Ruggiero el al. (167), reportaron el efecto de
extractos de té verde y vino tinto sobre el daño provocado tanto por la inoculación gástrica
de H. pylori como la administración de la toxina VacA en ratones BALB/c. Para ambos
productos el efecto protector también se asoció con su capacidad de neutralizar VacA in
vivo. La importancia de las sustancias de alto peso molecular también ha sido evidenciada
en el trabajo de Takabayashi et al., (300), quienes demostraron que el té negro (fracción
no-dializable) fue capaz de disminuir la densidad de colonización por H. pylori, así como la
inflamación inducida en gerbos de Mongolia. Recientemente, Brown et al. (301),
reportaron que el efecto de extractos de semilla y piel de uva sobre la viabilidad de H.
pylori estaría asociado al tipo de polifenol (estructura) y no al contenido total de éstos. En
otro trabajo, Akai et al., (302), estudiaron el efecto protector del té verde sobre la
proliferación y apoptosis inducida por H. pylori en ratones BALB/c. Los investigadores
observaron que sólo en los animales infectados hubo un efecto pro-apoptótico y
antiproliferativo en células de la mucosa, el que fue asociado a la administración de té
verde. La inhibición de la expresión de la citoquina pro-inflamatoria KC coincidió con una
la disminución de un 50% en la colonización por H. pylori.
En la Figura 34, se presenta un mecanismo propuesto recientemente (303), donde se
plantea una vía alternativa, mediante la cual los probióticos y probablemente los
prebióticos, pueden disminuir la inflamación a través del aumento en la expresión de las
moléculas supresoras de la señalización por citoquinas (SOCS-2, -3). Los autores

139
sugieren que el uso de sustancias que actúen aumentando la señalización anti-
inflamatoria (expresión de SOCS-2, -3 e inhibición de las JAK2), más que la atenuación
de la señalización inflamatoria, es una mejor estrategia para el control de la infección.
Dado que la vía que conduce al cáncer en la infección por H. pylori (Figura 2), se inicia
con la secuencia: inflamación – atrofia - metaplasia intestinal – displasia -
adenocarcinoma, los autores sugieren que el control de la inflamación es una estrategia
más relevante que la erradicación de la bacteria. En ese sentido el uso de prebióticos y
probióticos posee ventajas importantes asociadas al bajo costo, mejor tolerancia y
posibilidad de uso por periodos más prolongados.
Dado que en el estudio in vivo realizado en esta tesis no se evaluó las sustancias aisladas
de APPE, es difícil extrapolar los efectos anti-VacA y anti-adherente observados in vitro
para polifenoles aislados. Tal como se indicó anteriormente, el hecho que APPE sea
capaz de disminuir eficientemente la adherencia de H. pylori en etapas tempranas de
colonización, plantea la necesidad de esclarecer si en tal efecto participan otros
constituyentes no-polifenólicos del extracto, los cuales alcanzan a un 40 % del peso de
APPE. Dichos oligosacáridos neutros, si bien es cierto no poseen la potencia de sus
análogos ácidos, pueden aportar propiedades antiadherentes que se sumen a la de las
procianidinas de APPE. Al respecto, tal idea es reforzada por los hallazgos de Yang et al.
(304), quienes demostraron que las catequinas del té verde pueden disminuir
significativamente la colonización por H. pylori en ratones BALB/c cuando se asocian con
un reconocido agente antiadherencia como el ácido siálico. Adicionalmente, se ha descrito
recientemente que para lograr la erradicación de H. pylori con polifenoles puros como la
quercetina, se requiere de dosis elevadas (305). Por ejemplo, para lograr la erradicación
de H. pylori y protección del daño inflamatorio a la mucosa de guinea pigs, se ha tenido
que utilizar quercetina en dosis de 200 mg/kg/día durante 15 días. Lo anterior reafirma
que los ingredientes funcionales en los cuales co-existen varias sustancias bioactivas
pueden, a través de múltiples mecanismos, llegar a ser más efectivos en la prevención de
la infección por H. pylori y sus complicaciones. En la Figura 35, se resume de qué
manera APPE limita las primeras etapas que llevan al establecimiento de la colonización
por H. pylori (Figura 35, número 1). La capa de mucus posee un espesor de 100 m y un
elevado nivel de recambio por lo que la direccionalidad otorgada por la morfología y los

140
flagelos de H. pylori resultan vitales para el éxito de la adhesión a la mucosa (306). Como
ya se discutió anteriormente, la actividad ureasa también es muy importante para permitir
la multiplicación de la bacteria. Las adhesinas esquematizadas en la Figura 35, son
macromoléculas externas asociadas a la membrana de H. pylori, tienen expresión
variable, elevada redundancia y sólo en algunos casos poseen receptores conocidos.
Esta complejidad y combinatoria puede explicar por qué se requieren concentraciones
mayores de APPE para inhibir la adhesión de H. pylori comparativamente con las
encontradas para las otras actividades.
Figura 35. Blancos moleculares de APPE en la primera etapa del proceso de colonización de la infección por H. pylori. La actividad ureasa y los flagelos de H. pylori facilitan su orientación hacia el cuarto inferior de la capa de mucus sobre el epitelio por lo que su inhibición contribuye a disminuir el asentamiento de la bacteria. Varias moléculas de adhesión putativas, como BabA, SabA, antígeno de Lewis, y otras proteínas externas de membrana (OMP), están disponibles para adherirse a receptores conocidos o desconocidos del epitelio huésped. CagL de H. pylori que expresa el motif RGD, también sirve como puente de adherencia al epitelio gástrico que expresa
la integrina 51. Adaptado de referencia (297). El efecto anti-inflamatorio de APPE observado in vivo puede estar asociado a la presencia
mayoritaria de flavonoides derivados de quercetina y otros fenoles de bajo peso
molecular. En efecto, los resultados in vitro sugieren que la actividad antioxidante de

141
APPE sería consecuencia de la inhibición de la generación y/o atrapamiento de los ERO
producidas por los neutrófilos activados por la bacteria (307). Como resultado de los
estudios in vivo en la presente tesis se ha demostrado que el extracto APPE ejerce sus
efectos anti-H. pylori evitando el asentamiento del patógeno y protegiendo la mucosa
gástrica en las primeras etapas del daño inflamatorio. Así, en el modelo de infección a
corto plazo, la administración de dos dosis de APPE (150 y 300 mg/kg/día por 20 días),
fue suficiente para evitar el proceso de adhesión inicial de H. pylori a la mucosa antral de
ratones C57BL6/J. Como era de esperar, las propiedades anti-vacuolantes y antioxidantes
de los polifenoles de APPE permitieron disminuir el incipiente daño inflamatorio in vivo.
8. CONCLUSIONES
1. Se preparó un extracto seco (APPE) a partir de cáscara de manzana. En APPE se
logró una concentración de aproximadamente 60 % de polifenoles totales, cuyo
perfil reprodujo el de la cáscara fresca, siendo predominantes los glicósidos de
quercetina y las procinidinas derivadas de (-)-epicatequina. Del total de
polifenenoles, un 58 % correspondió a flavonoides y un 24% a procianidinas..
2. Los polifenoles de bajo peso molecular (LMW) de APPE fueron capaces de inhibir
la multiplicación de H. pylori en forma concentración-dependiente, comprometiendo
tempranamente la viabilidad de la bacteria.
3. APPE inhibió en forma eficiente a la ureasa obtenida de dos fuentes (Jack bean y
H. pylori). En ambos casos la inhibición fue concentración-dependiente, de tipo
competitiva y reversible. El fraccionamiento bio-dirigido de APPE permitió
esclarecer que la inhibición de la ureasa subyace principalmente en los
compuestos de alto peso molecular (HMW), como las procianidinas de tipo
oligoméricas y poliméricas.
4. APPE fue capaz de inhibir el estallido respiratorio en neutrófilos activados por los
agentes PMA, fMLP y H. pylori no-opsonizado. En todos los casos se observó un
efecto antioxidante concentración-dependiente y que abarcó tanto a las ERO

142
generadas al interior como en el exterior del neutrófilo. El tipo de sustancias
responsables de la actividad antioxidante corresponde a polifenoles de bajo peso
molecular, específicamente los glicósidos de quercetina. La IC50 necesaria para
que dichos compuestos fueran capaces de inhibir en un 50 % producción
extracelular de ERO fue del orden de 6 mol RutE/L (cinco veces menor a la
concentración requerida para suprimir la producción intracelular).
5. APPE inhibió en forma concentración-dependiente la vacuolación inducida por la
toxina VacA de H. pylori en células HeLa. Dicho efecto es la resultante de la
contribución de distintos grupos de polifenoles. Por un lado las procianidinas
oligoméricas son capaces de actuar directamente, formando complejos inactivos
con VacA. Por otro lado, los derivados de quercetina y chalconas interfieren con los
efectos funcionales de VacA, disminuyendo la formación de vacuolas.
Adicionalmente, se observó que concentraciones mayores de APPE además,
limitaron la adhesión de H. pylori a cultivos de células HeLa.
6. En un modelo de infección a corto plazo utilizando ratones C57BL/6J, la
administración de dos dosis de APPE por 20 días permitió la erradicación de H.
pylori. Dicho efecto se acompañó de una disminución del daño oxidativo e
inflamatorio (niveles de MDA), observado en las primeras etapas de colonización
de la mucosa gástrica (antral).

143
9. REFERENCIAS
1. Marshall, B.; Royce, H.; Annear, D.; Goowin, C.; Pearman, J.; Warren, J., Original
isolation of Campylobacter pyloridis from human gastric mucosa. Microbios Lett.
1984, 25, 83-88.
2. Warren, J., Unidentified curved bacilli on gastric epithelium in chronic active
gastritis. Lancet 1983, (i), 1293.
3. An international association between Helicobacter pylori infection and gastric
cancer. The EUROGAST Study Group. Lancet 1993, 341, (8857), 1359-1362.
4. Blaser, M. J.; Atherton, J. C., Helicobacter pylori persistence: biology and disease.
J. Clin. Invest. 2004, 113, (3), 321-333.
5. Hamlet, A.; Olbe, L., The influence of Helicobacter pylori infection on gastric
emptying, gastrin release, acid secretion, duodenal acid load and duodenal bulb pH
following neutral and acidified liquid peptone meals in man. Gastroenterology 1996,
110, (4), A128-A128.
6. Khulusi, S.; Badve, S.; Patel, P.; Lloyd, R.; Marrero, J. M.; Finlayson, C.; Mendall,
M. A.; Northfield, T. C., Pathogenesis of gastric metaplasia of the human
duodenum: Role of Helicobacter pylori, gastric acid, and ulceration.
Gastroenterology 1996, 110, (2), 452-458.
7. IARC Working Group on the Evaluation of Carcinigenesis Risks to Humans.
Helicobacter pylori. In Schistosomes, liver flukes and Helicobacter pylori. IARC
Monographs in the Evaluation of Carcinogenesis Risks in Humans 1994, 61, (IARC,
Lyon), 177-241.
8. Parkin, D. M.; Bray, F.; Ferlay, J.; Pisani, P., Global cancer statistics, 2002. Ca-a
Cancer Journal for Clinicians 2005, 55, (2), 74-108.

144
9. Hessey, S. J.; Spencer, J.; Wyatt, J. I.; Sobala, G.; Rathbone, B. J.; Axon, A. T. R.;
Dixon, M. F., Bacterial adhesion and disease-activity in Helicobacter associated
chronic gastritis. Gut 1990, 31, (2), 134-138.
10. Logan, R. P. H., Adherence of Helicobacter pylori. Aliment. Pharm. Therap. 1996,
10, 3-15.
11. Hessey, S. J.; Spencer, J.; Wyatt, J. I.; Sobala, G.; Rathbone, B. J.; Axon, A. T. R.;
Dixon, M. F., Bacterial adhesion and disease activity in Helicobacter associated
chronic gastritis. Gut 1990, 31, (2), 134-138.
12. O'Brien, D. P.; Israel, D. A.; Krishna, U.; Romero-Gallo, J.; Nedrud, J.; Medof, M.
E.; Lin, F.; Redline, R.; Lublin, D. M.; Nowicki, B. J.; Franco, A. T.; Ogden, S.;
Williams, A. D.; Polk, D. B.; Peek, R. M., The role of decay-accelerating factor as a
receptor for Helicobacter pylori and a mediator of gastric inflammation. J. Biol.
Chem. 2006, 281, (19), 13317-13323.
13. O'Brien, D. P.; Krishna, U.; Romero-Gallo, J.; Nedrud, J.; Medof, M. E.; Lin, F.;
Redline, R.; Lublin, D. M.; Nowicki, B. J.; Israel, D.; Franco, A. T.; Peek, R. M.,
Decay accelerating factor (DAF) is a cellular receptor for Helicobacter pylori.
Gastroenterology 2006, 130, (4), A127-A127.
14. O'Brien, D. P.; Romero-Gallo, J.; Schneider, B. G.; Chaturvedi, R.; Delgado, A.;
Harris, E. J.; Krishna, U.; Ogden, S. R.; Israel, D. A.; Wilson, K. T.; Peek, R. M.,
Regulation of the Helicobacter pylori cellular receptor decay-accelerating factor. J.
Biol. Chem. 2008, 283, (35), 23922-23930.
15. Gewirtz, A.; Krishna, U.; Israel, D.; Yu, Y. M.; Peek, R. M., Helicobacter pylori
evades Toll-like receptor 5 innate immunity. Gastroenterology 2003, 124, (4), A592-
A592.
16. Gewirtz, A. T.; Yu, Y. M.; Krishna, U. S.; Israel, D. A.; Lyons, S. L.; Peek, R. M.,
Helicobacter pylori flagellin evades toll-like receptor 5-mediated innate immunity. J.
Infect. Dis. 2004, 189, (10), 1914-1920.

145
17. Ganten, T. M.; Aravena, E.; Sykora, J.; Koschny, R.; Mohr, J.; Rudi, J.; Stremmel,
W.; Walczak, H., Helicobacter pylori-induced apoptosis in T cells is mediated by the
mitochondrial pathway independent of death receptors. Eu. J. Clin. Invest. 2007, 37,
(2), 117-125.
18. Rivas-Traverso, F.; Hernández, F., Helicobacter pylori: Factores de virulencia,
patología y diagnóstico. Rev. Biomed. 2000, 11, 187-205.
19. Khan, S.; Karim, A.; Iqbal, S., Helicobacter urease: Niche construction at the single
molecule level. J. Bioscience. 2009, 34, (4), 503-511.
20. Ilver, D.; Barone, S.; Mercati, D.; Lupetti, P.; Telford, J. L., Helicobacter pylori toxin
VacA is transferred to host cells via a novel contact-dependent mechanism. Cell.
Microbiol. 2004, 6, (2), 167-174.
21. Yahiro, K.; Niidome, T.; Kimura, M.; Hatakeyama, T.; Aoyagi, H.; Kurazono, H.;
Imagawa, K.; Wada, A.; Moss, J.; Hirayama, T., Activation of Helicobacter pylori
VacA toxin by alkaline or acid conditions increases its binding to a 250-kDa receptor
protein-tyrosine phosphatase beta. J. Biol. Chem. 1999, 274, (51), 36693-36699.
22. Yahiro, K.; Wada, A.; Nakayama, M.; Kimura, T.; Ogushi, K.; Niidome, T.; Aoyagi,
H.; Yoshino, K.; Yonezawa, K.; Moss, J.; Hirayama, T., Protein-tyrosine
phosphatase alpha, RPTP alpha, is a Helicobacter pylori VacA receptor. J. Biol.
Chem. 2003, 278, (21), 19183-19189.
23. Yahiro, K.; Wada, A.; Yamasaki, E.; Nakayama, M.; Nishi, Y.; Hisatsune, J.;
Morinaga, N.; Sap, J.; Noda, M.; Moss, J.; Hirayama, T., Essential domain of
receptor tyrosine phosphatase beta (RPTP beta) for interaction with Helicobacter
pylori vacuolating cytotoxin. J. Biol. Chem. 2004, 279, (49), 51013-51021.
24. Seto, K.; Hayashi-Kuwabara, Y.; Yoneta, T.; Suda, H.; Tamaki, H., Vacuolation
induced by cytotoxin from Helicobacter pylori is mediated by the EGF receptor in
HeLa cells. Febs Lett. 1998, 431, (3), 347-350.

146
25. Kuo, C. H.; Wang, W. C., Binding and internalization of Helicobacter pylori VacA via
cellular lipid rafts in epithelial cells. Biochem. Bioph. Res. Co. 2003, 303, (2), 640-
644.
26. Tombola, F.; Morbiato, L.; Del Giudice, G.; Rappuoli, R.; Zoratti, M.; Papini, E., The
Helicobacter pylori VacA toxin is a urea permease that promotes urea diffusion
across epithelia. J. Clin. Invest. 2001, 108, (6), 929-937.
27. McClain, M. S.; Schraw, W.; Ricci, V.; Boquet, P.; Cover, T. L., Acid activation of
Helicobacter pylori vacuolating cytotoxin (VacA) results in toxin internalization by
eukaryotic cells. Mol. Microbiol. 2000, 37, (2), 433-442.
28. Ricci, V.; Sommi, P.; Fiocca, R.; Necchi, V.; Romano, M.; Solcia, E., Extracellular
pH modulates Helicobacter pylori-induced vacuolation and VacA toxin
internalization in human gastric epithelial cells. Biochem. Bioph. Res. Co. 2002,
292, (1), 167-174.
29. Papini, E.; Satin, B.; de Bernard, M.; Molinari, M.; Arico, B.; Galli, C.; Telford, J. R.;
Rappuoli, R.; Montecucco, C., Action site and cellular effects of cytotoxin VacA
produced by Helicobacter pylori. Folia Microbiol. 1998, 43, (3), 279-284.
30. Willhite, D. C.; Cover, T. L.; Blanke, S. R., Cellular vacuolation and mitochondrial
cytochrome c release are independent outcomes of Helicobacter pylori vacuolating
cytotoxin activity that are each dependent on membrane channel formation. J. Biol.
Chem. 2003, 278, (48), 48204-48209.
31. Galmiche, A.; Rassow, J.; Doye, A.; Cagnol, S.; Chambard, J. C.; Contamin, S.; de
Thillot, V.; Just, I.; Ricci, V.; Solcia, E.; Van Obberghen, E.; Boquet, P., The N-
terminal 34 kDa fragment of Helicobacter pylori vacuolating cytotoxin targets
mitochondria and induces cytochrome c release. EMBO J. 2000, 19, (23), 6361-
6370.

147
32. Molinari, M.; Galli, C.; de Bernard, M.; Norais, N.; Ruysschaert, J. M.; Rappuoli, R.;
Montecucco, C., The acid activation of Helicobacter pylori toxin VacA: Structural
and membrane binding studies. Biochem. Bioph. Res. Co. 1998, 248, (2), 334-340.
33. Gebert, B.; Fischer, W.; Weiss, E.; Hoffmann, R.; Haas, R., Helicobacter pylori
vacuolating cytotoxin inhibits T lymphocyte activation. Science 2003, 301, (5636),
1099-1102.
34. Sundrud, M. S.; Torres, V. J.; Unutmaz, D.; Cover, T. L., Inhibition of primary
human T cell proliferation by Helicobacter pylori vacuolating toxin (VacA) is
independent of VacA effects on IL-2 secretion. P. Natl. Acad. Sci. USA. 2004, 101,
(20), 7727-7732.
35. Tombola, F.; Campello, S.; De Luca, L.; Ruggiero, P.; Del Giudice, G.; Papini, E.;
Zoratti, M., Plant polyphenols inhibit VacA, a toxin secreted by the gastric pathogen
Helicobacter pylori. Febs Lett. 2003, 543, (1-3), 184-189.
36. Tombola, F.; Carlesso, C.; Szabo, I.; de Bernard, M.; Reyrat, J. M.; Telford, J. L.;
Rappuoli, R.; Montecucco, C.; Papini, E.; Zoratti, M., Helicobacter pylori vacuolating
toxin forms anion-selective channels in planar lipid bilayers: Possible implications
for the mechanism of cellular vacuolation. Biophys. J. 1999, 76, (3), 1401-1409.
37. Yamazaki, S.; Yamakawa, A.; Ito, Y.; Ohtani, M.; Higashi, H.; Hatakeyama, M.;
Azuma, T., The CagA protein of Helicobacter pylori is translocated into epithelial
cells and binds to SHP-2 in human gastric mucosa. J. Infect. Dis. 2003, 187, (2),
334-337.
38. Stein, M.; Bagnoli, F.; Halenbeck, R.; Rappuoli, R.; Fantl, W. J.; Covacci, A., c-
Src/Lyn kinases activate Helicobacter pylori CagA through tyrosine phosphorylation
of the EPIYA motifs. Mol. Microbiol. 2002, 43, (4), 971-980.
39. Higashi, H., Disturbance of intracellular signaling by Helicobacter pylori infection.
Seikagaku 2003, 75, (9), 1243-1246.

148
40. Higashi, H.; Nakaya, A.; Tsutsumi, R.; Yokoyama, K.; Fujii, Y.; Ishikawa, S.;
Higuchi, M.; Takahashi, A.; Kurashima, Y.; Teishikata, Y.; Tanaka, S.; Azuma, T.;
Hatakeyama, M., Helicobacter pylori CagA induces Ras-independent
morphogenetic response through SHP-2 recruitment and activation. J. Biol. Chem.
2004, 279, (17), 17205-17216.
41. Ren, S. M.; Higashi, H.; Lu, H. S.; Azuma, T.; Hatakeyama, M., Structural basis and
functional consequence of Helicobacter pylori CagA multimerization in cells. J. Biol.
Chem. 2006, 281, (43), 32344-32352.
42. Amieva, M. R.; Vogelmann, R.; Covacci, A.; Tompkins, L. S.; Nelson, W. J.; Falkow,
S., Disruption of the epithelial apical-junctional complex by Helicobacter pylori
CagA. Science 2003, 300, (5624), 1430-1434.
43. Naumann, M.; Wessler, S.; Bartsch, C.; Wieland, B.; Covacci, A.; Haas, R.; Meyer,
T. F., Activation of activator protein 1 and stress response kinases in epithelial cells
colonized by Helicobacter pylori encoding the cag pathogenicity island. J. Biol.
Chem. 1999, 274, (44), 31655-31662.
44. Meyer-ter-Vehn, T.; Covacci, A.; Kist, M.; Pahl, H. L., Helicobacter pylori activates
mitogen-activated protein kinase cascades and induces expression of the proto-
oncogenes c-fos and c-jun. J. Biol. Chem. 2000, 275, (21), 16064-16072.
45. Mitsuno, Y.; Maeda, S.; Yoshida, H.; Hirata, Y.; Ogura, K.; Akanuma, M.; Kawabe,
T.; Shiratori, Y.; Omata, M., Helicobacter pylori activates the proto-oncogene c-fos
through SRE transactivation. Biochem. Biophys. Res. Co. 2002, 291, (4), 868-874.
46. Oliveira, M. J.; Costa, A. C.; Costa, A. M.; Atherton, J. C.; Machado, J. C.; Seruca,
R.; Mareel, M.; Leroy, A.; Figueiredo, C., Helicobacter pylori induces gastric
epithelial cell invasion in a c-Met and cag PAI-dependent manner. Helicobacter
2006, 11, (4), 331-331.
47. Oliveira, M. J.; Costa, A. C.; Costa, A. M.; Henriques, L.; Suriano, G.; Atherton, J.
C.; Machado, J. C.; Carneiro, F.; Seruca, R.; Mareel, M.; Leroy, A.; Figueiredo, C.,

149
Helicobacter pylori induces gastric epithelial cell invasion in a c-Met and type IV
secretion system-dependent manner. J. Biol. Chem. 2006, 281, (46), 34888-34896.
48. Kundu, P.; Mukhopadhyay, A. K.; Patra, R.; Banerjee, A.; Berg, D. E.; Swarnakar,
S., Cag pathogenicity island-independent up-regulation of matrix
metalloproteinases-9 and -2 secretion and expression in mice by Helicobacter pylori
infection. J. Biol. Chem. 2006, 281, (45), 34651-34662.
49. Atherton, J. C.; Cao, P.; Peek, R. M.; Blaser, M. J.; Cover, T. L., Mosaicism in vaca,
the vacuolating cytotoxin gene of Helicobacter-pylori, and genetic-linkage of vaca
subtypes with CagA, the cytotoxin-associated gene. Gastroenterology 1995, 108,
(4), A774-A774.
50. Peek, R. M., Microbes and microbial toxins: Paradigms for microbial-mucosal
interactions - IV. Helicobacter pylori strain-specific activation of signal transduction
cascades related to gastric inflammation. American Journal of Physiology-
Gastrointestinal and Liver Physiology 2001, 280, (4), G525-G530.
51. Oliveira, A. G.; Santos, A.; Guerra, J. B.; Rocha, G. G.; Rocha, A. M. C.; Oliveira, C.
A.; Cabral, M.; Nogueira, A.; Queiroz, D. M. M., babA2- and cagA-positive
Helicobacter pylori strains are associated with duodenal ulcer and gastric
carcinoma in Brazil. J. Clin. Microbiol. 2003, 41, (8), 3964-3966.
52. Zhou, J. C.; Zhang, J. Z.; Xu, C. P.; He, L. H., CagA genotype and variants in
Chinese Helicobacter pylori strains and relationship to gastroduodenal diseases. J.
Med. Microbiol. 2004, 53, (3), 231-235.
53. Akhter Y, A. I., Devi SM, Ahmed N., The co-evolved Helicobacter pylori and gastric
cancer: trinity of bacterial virulence, host susceptibility and lifestyle Infect. Agent.
Cancer 2007, 2, (2), 5.
54. Wang, G.; Alamuri, P.; Maier, R. J., The diverse antioxidant systems of Helicobacter
pylori. Mol. Microbiol. 2006, 61, (4), 847-860.

150
55. Olczak, A. A.; Olson, J. W.; Maier, R. J., Oxidative-stress resistance mutants of
Helicobacter pylori. J. Bacteriol. 2002, 184, (12), 3186-3193.
56. Olczak, A. A.; Seyler, R. W.; Olson, J. W.; Maier, R. J., Association of Helicobacter
pylori antioxidant activities with host colonization proficiency. Infect. Immun. 2003,
71, (1), 580-583.
57. Wang, G.; Maier, R. J., An NADPH quinone reductase of Helicobacter pylori plays
an important role in oxidative stress resistance and host colonization. Infect. Immun.
2004, 72, (3), 1391-1396.
58. Comtois, S. L.; Gidley, M. D.; Kelly, D. J., Role of the thioredoxin system and the
thiol-peroxidases Tpx and Bcp in mediating resistance to oxidative and nitrosative
stress in Helicobacter pylori. Microbiol-Sgm 2003, 149, 121-129.
59. Alamuri, P.; Maier, R. J., Methionine sulfoxide reductase in Helicobacter pylori:
Interaction with methionine-rich proteins and stress-induced expression. J.
Bacteriol. 2006, 188, (16), 5839-5850.
60. Mooney, C.; Keenan, J.; Munster, D.; Wilson, I.; Allardyce, R.; Bagshaw, P.;
Chapman, B.; Chadwick, V., Neutrophil activation by Helicobacter pylori. Gut 1991,
32, (8), 853-857.
61. Allen, L. A. H.; Beecher, B. R.; Lynch, J. T.; Rohner, O. V.; Wittine, L. M.,
Helicobacter pylori disrupts NADPH oxidase targeting in human neutrophils to
induce extracellular superoxide release. J. Immun. 2005, 174, (6), 3658-3667.
62. Peek, R. M., Events at the host-microbial interface of the gastrointestinal tract IV.
The pathogenesis of Helicobacter pylori persistence. American Journal of
Physiology-Gastrointestinal and Liver Physiology 2005, 289, (1), G8-G12.
63. Peek, R. M., Pathogenesis of Helicobacter pylori infection. Springer Semin. Immun.
2005, 27, (2), 197-215.

151
64. McGee, D. J.; Kumar, S.; Viator, R. J.; Bolland, J. R.; Ruiz, J.; Spadafora, D.;
Testerman, T. L.; Kelly, D. J.; Pannell, L. K.; Windle, H. J., Helicobacter pylori
thioredoxin is an arginase chaperone and guardian against oxidative and nitrosative
stresses. J. Biol. Chem. 2006, 281, (6), 3290-3296.
65. Davi, G.; Neri, M.; Falco, A.; Festi, D.; Taraborelli, T.; Ciabattoni, G.; Basili, S.;
Cuccurullo, F.; Patrono, C., Helicobacter pylori infection causes persistent platelet
activation in vivo through enhanced lipid peroxidation. Arterioscl. Throm. Vas. 2005,
25, (1), 246-251.
66. Drake, I. M.; Mapstone, N. P.; Schorah, C. J.; White, K. L. M.; Chalmers, D. M.;
Dixon, M. F.; Axon, A. T. R., Reactive oxygen species activity and lipid peroxidation
in Helicobacter pylori associated gastritis: relation to gastric mucosal ascorbic acid
concentrations and effect of H. pylori eradication. Gut 1998, 42, (6), 768-771.
67. Smoot, D. T.; Elliott, T. B.; Verspaget, H. W.; Jones, D.; Allen, C. R.; Vernon, K. G.;
Bremner, T.; Kidd, L. R.; Kim, G. S.; Groupman, J. D.; Ashktorab, H., Influence of
Helicobacter pylori on reactive oxygen-induced gastric epithelial cell injury.
Carcinogenesis 2000, 21, (11), 2091-2095.
68. Ladeira, M. S. P.; Rodrigues, M. A. M.; Salvadori, D. M. F.; Queiroz, D. M. M.;
Freire-Maia, D. V., DNA damage in patients infected by Helicobacter pylori. Cancer
Epidem. Biom. 2004, 13, (4), 631-637.
69. Siomek, A.; Rytarowska, A.; Szaflarska-Poplawska, A.; Gackowski, D.; Rozalski, R.;
Dziaman, T.; Czerwionka-Szaflarska, M.; Olinski, R., Helicobacter pylori infection is
associated with oxidatively damaged DNA in human leukocytes and decreased
level of urinary 8-oxo-7,8-dihydroguanine. Carcinogenesis 2006, 27, (3), 405-408.
70. Hatakeyama, M., Oncogenic mechanisms of the Helicobacter pylori CagA protein.
Nat. Rev. Cancer 2004, 4, (9), 688-694.

152
71. Kawanishi, S.; Hiraku, Y., Oxidative and Nitrative DNA damage as Biomarker for
Carcinogenesis with special reference to inflamation. Antioxid. Redox Sign. 2006, 8,
(5,6), 1047-1058.
72. Schottenfeld, D.; Beebe-Dimmer, J., Chronic inflammation: A common and
important factor in the pathogenesis of neoplasia. Ca-a Cancer J. Clin. 2006, 56,
(2), 69-83.
73. Matthews, G. M.; Butler, R. N., Cellular mucosal defense during Helicobacter pylori
infection: A review of the role of glutathione and the oxidative pentose pathway.
Helicobacter 2005, 10, (4), 298-306.
74. Arend, A.; Loime, L.; Roosaar, P.; Soom, M.; Krista, L.; Sepp, E.; Aunapuu, A.;
Zilmer, K.; Selstam, G.; Zilmer, M., Helicobacter pylori sustantially increases
oxidative stress in indometacin-exposed rat gastric mucosa. Medicina (Kaunas)
2005, 41, (4), 343-347.
75. Ernst, P., Review article: the role of inflammation in the pathogenesis of gastric
cancer. Alim. Pharmacol. Ther. 1999, 13, 13-18.
76. Naito, Y.; Yoshikawa, T., Molecular and cellular mechanisms involved in
Helicobacter pylori-induced inflammation and oxidative stress. Free Radical Bio.
Med. 2002, 33, (3), 323-336.
77. Jung, H.; Lee, K.; Chu, S.; Yi, S., Reactive oxygen species activity, mucosal
lipoperoxidation and glutathione in Helicobacter pylori-infected gastric mucosa. J
Gastroenterol Hepatol 2001, 16, 1336-1340
78. Verhulst, M.; Oijen, A.V.; Roelofs, H.; Peters, W.; Jansen, J., Antral glutathione
concentration and glutathione S-transferase activity in patients with and without
Helicobacter pylori. Digest Dis Sci 2000, 45, 629-632.
79. Shirin, H.; Pinto, J. T.; Liu, L. U.; Merzianu, M.; Sordillo, E. M.; Moss, S. F.,
Helicobacter pylori decreases gastric mucosal glutathione. Cancer Lett. 2001, 164,
(2), 127-133.

153
80. Pignatelli, B.; Bancel, B.; Esteve, J.; Malaveille, C.; Calmels, S.; Correa, P.,
Helicobacter pylori Inducible nitric oxide synthase, anti-oxidant enzymes and
Helicobacter pylori infection in gastritis and gastric precancerous lesions in humans.
Eur. J. Cancer Prevent. 1998, 439-447
81. Nakamura, A.; Park, A. M.; Nagata, K.; Sato, E. F.; Kashiba, M.; Tamura, T.; Inoue,
M., Oxidative cellular damage associated with transformation of from a bacillary to a
coccoid form. Free Radical Bio. Med. 2000, 28, (11), 1611-1618.
82. Felley, C.; Pignatelli, B.; Melle, G. V.; Crabtree, J.; Stolte, M.; Diezi, J.; Corthesy-
Theulaz, I.; Michetti, P.; Bancel, B.; Patricot, L.; Ohshima, H.; Felley-Bosco, E.,
Oxidative stress in gastric mucosa of asymptomatic humans infected with
Helicobacter pylori: effect of bacterial eradication. Helicobacter 2002, 7, (6), 342-
348.
83. Hentschel, E.; Brandstatter, G. Dragosics, B.; Hirschl, A. M.; Nemec, H.; Schutze,
K.; Taufer, M.; Wurzer, H., Effect of ranitidine and amoxicillin plus metronidazol on
the eradication of Helicobacter pylori and the recurrence of duodenal ulcer. New
Engl. J. Med. 1993, 328, (5), 308-312.
84. de Boer, W. A.; Van Etten, R.; Van de Wouw, B. A. M.; Schneeberger, P. M.; Van
Oijen, A.; Jansen, J., Bismuth-based quadruple therapy for Helicobacter pylori - a
single triple capsule plus lansoprazole. Aliment. Pharm.Ther. 2000, 14, (1), 85-89.
85. Current European concepts in the management of Helicobacter pylori infection. The
Maastricht Consensus Report. European Helicobacter pylori Study Group. Gut
1997, 41, 8-13.
86. O´Morain, C., Teatment of Helicobacter pylori infection. US Gastroenterology
Review 2006, 1-5.

154
87. Correa, P.; van Doorn, L. J.; Bravo, J. C.; Ruiz, B.; Bravo, L. E.; Realpe, J. L.,
Unsuccessful treatment results in survival of less virulent genotypes of Helicobacter
pylori in Colombian patients. Am. J. Gastroenterol. 2000, 95, (2), 564-566.
88. Figueroa, G.; Acuna, P.; Troncoso, M.; Portell, D. P.; Toledo, M. S.; Valenzuela, J.,
Helicobacter pylori infection in Chile. Clin. Infect. Dis. 1997, 25, (5), 983-989.
89. Sorberg, M.; Hanberger, H.; Nilsson, M.; Bjorkman, A.; Nilsson, L. E., Risk of
development of in vitro resistance to amoxicillin, clarithromycin, and metronidazole
in Helicobacter pylori. Antimicrob. Agents Chem. 1998, 42, (5), 1222-1228.
90. Gonzalez, C.; Garcia, A.; Daroch, F.; Kawaguchi, F.; Solar, H.; Rivera, N.; Vega, E.,
In vitro antimicrobial susceptibility of Helicobacter pylori strains isolated in Chile.
Rev. Med. Chile 2001, 129, (6), 643-646.
91. Kobayashi, I.; Saika, T.; Muraoka, H.; Murakami, K.; Fujioka, T., Helicobacter pylori
isolated from patients who later failed Helicobacter pylori eradication triple therapy
readily develop resistance to clarithromycin. J. Med. Microbiol. 2006, 55, (6), 737-
740.
92. Moayyedi, P.; Axon, A. T.; Feltbower, R.; Duffett, S.; Crocombe, W.; Braunholtz, D.;
Richards, I. G.; Dowell, A. C.; Forman, D., Relation of adult lifestyle and
socioeconomic factors to the prevalence of Helicobacter pylori infection. Int. J.
Epidemiol. 2002, 31, (3), 624-631.
93. Suerbaum, S.; Michetti, P., Medical progress: Helicobacter pylori infection. New
Engl. J. Med. 2002, 347, (15), 1175-1186.
94. Valenzuela, J., Helicobacter pylori. The microbiological revolution. Rev. Med. Chile
1999, 127, (8), 891-893.
95. Figueroa, G.; Troncoso, M.; Toledo, M. S.; Faundez, G.; Acuna, R., Prevalence of
serum antibodies to Helicobacter pylori VacA and CagA and gastric diseases in
Chile. J. Med. Microbiol. 2002, 51, (4), 300-304.

155
96. Farthing, M. J. G., Helicobacter pylori infection: an overview. Brit. Med. Bull. 1998,
54, (1), 1-6.
97. Ernst, P. B.; Gold, B. D., The disease spectrum of Helicobacter pylori: The
immunopathogenesis of gastroduodenal ulcer and gastric cancer. Annu. Rev.
Microbiol. 2000, 54, 615-640.
98. Mcmanus, T., Helicobacter pylori: an emergin infectious disease. Nurs. Pract. 2000,
25, (8), 42-46.
99. Rowland, M.; Drumm, B., Clinical significance of Helicobacter infection in children.
Brit. Med. Bull. 1998, 54, (1), 95-103.
100. Nostro, A.; Cellini, L.; Di Bartolomeo, S.; Di Campli, E.; Grande, R.; Cannatelli, M.
A.; Marzio, L.; Alonzo, V., Antibacterial effect of plant extracts against Helicobacter
pylori. Phytother. Res. 2005, 19, (3), 198-202.
101. Ustun, O.; Ozcelik, B.; Akyon, Y.; Abbasoglu, U.; Yesilada, E., Flavonoids with anti-
Helicobacter pylori activity from Cistus laurifolius leaves. J. Ethnopharmacol. 2006,
108, (3), 457-461.
102. Ross, J.; Kasum, C., Dietary Flavonoids: Bioavailability, Metabolic Effects, and
Safety. Annu. Rev. Nutr. 2002, 22, 19-24.
103. Kim, H.; Hee, O.; Sung, M., Effects of Phenol-Depleted and Phenol-Rich Diets on
Blood Markers os Oxidative Stress, and Urinary Excretion of Quercetin and
Kaempferol in Healthy Volunteers. Journal of the Americal College of Nutrition
2003, 22, 217-223.
104. Kris-Etherton, P.; Lefever, M.; Beecher, G.; Gross, M.; Keen, C.; Etherton, T.,
Bioactive Compounds in Nutrition and Health-Research Methodologies for
Establishing Niological Function: The Antioxidant and Anti-Inflammatory Effects of
Flavonoids an Atherosclerosis. Annu. Rev. Nutr. 2004, 24, 511-538.

156
105. Ramassamy, C., Emerging role of polyphenolic compounds in the treatment of
neurodegenerative diseases: A review of their intracellular targets. Eu. J.
Pharmacol. 2006, 545, (1), 51-64.
106. MN, M. C., Diet-Derived Phenols in Plasma and Tissues and their Implications for
Health. Planta Med. 2004, 70, 1103-1114.
107. Puupponen-Pimia, R.; Nohynek, L.; Meier, C.; Kahkonen, M.; Heinonen, M.; Hopia,
A.; Oksman-Caldentey, K. M., Antimicrobial properties of phenolic compounds from
berries. J. Appl. Microbiol. 2001, 90, (4), 494-507.
108. Mori, A.; Nishino, C.; Enoki, N.; Tawata, S., Antibacterial activity and mode of action
of plant flavonoids agaisnt Proteus vulgaris and Staphylococcus aureus.
Phytochemistry 1987, 26, 2231-2234.
109. Haraguchi, H.; Tanimoto, K.; Tamura, Y.; Mizutani, K.; Kinoshita, T., Mode of
antibacterial action of retrochalcones from Glycyrrhiza inflata. Phytochemistry 1998,
48, (1), 125-129.
110. Funatogawa, K.; Hayashi, S.; Shimomura, H.; Yoshida, T.; Hatano, T.; Ito, H.; Hirai,
Y., Antibacterial activity of hydrolyzable tannins derived from medicinal plants
against Helicobacter pylori. Microbiol. Immunol. 2004, 48, (4), 251-261.
111. Correia, R. T. P.; McCue, P.; Vattem, D. A.; Magalhaes, M. M. A.; Macedo, G. R.;
Shetty, K., Amylase and Helicobacter pylori inhibition by phenolic extracts of
pineapple wastes bioprocessed by Rhizopus oligosporus. J. Food Biochem. 2004,
28, (5), 419-434.
112. Shin, J. E.; Kim, J. M.; Bae, E. A.; Hyun, Y. J.; Kim, D. H., In vitro inhibitory effect of
flavonoids on growth, infection and vacuolation of Helicobacter pylori. Planta Med.
2005, 71, (3), 197-201.
113. Nostro, A.; Cellini, L.; Di Bartolomeo, S.; Cannatelli, M. A.; Di Campli, E.; Procopio,
F.; Grande, R.; Marzio, L.; Alonzo, V., Effects of combining extracts (from propolis

157
or Zingiber officinale) with clarithromycin on Helicobacter pylori. Phytother. Res.
2006, 20, (3), 187-190.
114. Yanagawa, Y.; Yamamoto, Y.; Hara, Y.; Shimamura, T., A combination effect of
epigallocatechin gallate, a major compound of green tea catechins, with antibiotics
on Helicobacter pylori growth in vitro. Curr. Microbiol. 2003, 47, (3), 244-249.
115. Avorn, J.; Monane, M.; Gurwitz, J. H.; Glynn, R. J.; Choodnovskiy, I.; Lipsitz, L. A.,
Reduction of bacteriuria and pyuria after ingestion of cranberry juice. J. Amer. Med.
Assoc. 1994, 271, (10), 751-754.
116. Henig, Y. S.; Leahy, M. M., Cranberry juice and urinary-tract health: Science
supports folklore. Nutrition 2000, 16, (7-8), 684-687.
117. Stothers, L., A ramdomized trial to evaluate effectiveness and cost effectiveness of
naturophatic cranberry products as profilaxis against urinary tract infection in
women. Can. J. Urol. 2002, 9, 1558-1562.
118. Di Martino, P.; Agniel, R.; David, K.; Templer, C.; Gaillard, J. L.; Denys, P.; Botto,
H., Reduction of Escherichia coli adherence to uroepithelial bladder cells after
consumption of cranberry juice: a double-blind randomized placebo-controlled
cross-over trial. World J. Urol. 2006, 24, (1), 21-27.
119. Kontiokari, T.; Sundqvist, K.; Nuutinen, M.; Pokka, T.; Koskela, M.; Uhari, M.,
Randomised trial of cranberry-lingonberry juice and Lactobacillus GG drink for the
prevention of urinary tract infections in women. Brit. Med. J. 2001, 322, (7302),
1571-1573.
120. Howell, A. B.; Foxman, B., Cranberry juice and adhesion of antibiotic-resistant
uropathogens. J. Amer. Med. Assoc. 2002, 287, (23), 3082-3083.
121. Howell, A. B.; Reed, J. D.; Krueger, C. G.; Winterbottom, R.; Cunningham, D. G.;
Leahy, M., A-type cranberry proanthocyanidins and uropathogenic bacterial anti-
adhesion activity. Phytochemistry 2005, 66, (18), 2281-2291.

158
122. Burger, O.; Ofek, I.; Tabak, M.; Weiss, E.; Sharon, N.; I, N., A high molecular mass
constituent of cramberry juice inhibits Helicobacter pylori adhesion to human gastric
mucus. FEMS Immunol Med. Mic. 2000, 29, 295-301.
123. Zhang, L.; Ma, J. L.; Pan, K. F.; Go, V. L. W.; Chen, J. S.; You, W. C., Efficacy of
cranberry juice on Helicobacter pylori infection: A double-blind, randomized
placebo-controlled trial. Helicobacter 2005, 10, (2), 139-145.
124. Lin, Y. T.; Vattem, D.; Labbe, R. G.; Shetty, K., Enhancement of antioxidant activity
and inhibition of Helicobacter pylori by phenolic phytochemical-enriched alcoholic
beverages. Process Biochem. 2005, 40, (6), 2059-2065.
125. Vattem, D. A.; Lin, Y. T.; Ghaedian, R.; Shetty, K., Cranberry synergies for dietary
management of Helicobacter pylori infections. Process Biochem. 2005, 40, (5),
1583-1592.
126. Vattem, D. A.; Lin, Y. T.; Shetty, K., Enrichment of phenolic antioxidants and anti-
Helicobacter pylori properties of cranberry pomace by solid-state bioprocessing.
Food Biotechnol. 2005, 19, (1), 51-68.
127. Nohynek, L. J.; Alakomi, H. L.; Kahkonen, M. P.; Heinonen, M.; Helander, K. M.;
Oksman-Caldentey, K. M.; Puupponen-Pimia, R. H., Berry phenolics: Antimicrobial
properties and mechanisms of action against severe human pathogens. Nutrition
and Cancer-an International Journal 2006, 54, (1), 18-32.
128. Yee, Y. K.; Koo, M. W. L., Anti-Helicobacter pylori activity of Chinese tea: in vitro
study. Aliment. Pharm. Ther. 2000, 14, (5), 635-638.
129. Yee, Y. K.; Koo, M. W. L.; Szeto, M. L., Chinese tea consumption and lower risk of
Helicobacter infection. J. Gastroen. Hepatol. 2002, 17, (5), 552-555.
130. Ruggiero, P.; Tombola, F.; Rossi, G.; Pancotto, L.; Lauretti, L.; Del Giudice, G.;
Zoratti, M., Polyphenols reduce gastritis induced by Helicobacter pylori infection or
VacA toxin administration in mice. Antimicrob. Agents Ch. 2006, 50, (7), 2550-2552.

159
131. Zhang, H. M.; Wakisaka, N.; Maeda, O.; Yamamoto, T., Vitamin C inhibits the
growth of a bacterial risk factor for gastric carcinoma: Helicobacter pylori. Cancer
1997, 80, (10), 1897-1903.
132. Mabe, K.; Yamada, M.; Oguni, I.; Takahashi, T., In vitro and in vivo activities of tea
catechins against Helicobacter pylori. Antimicrob. Agents Ch. 1999, 43, (7), 1788-
1791.
133. Wang, X.; Willen, R.; Wadstrom, T., Astaxanthin-rich algal meal and vitamin C
inhibit Helicobacter pylori infection in BALB/cA mice. Antimicrob. Agents Ch. 2000,
44, (9), 2452-2457.
134. Wang, W. Q.; Heideman, L.; Chung, C. S.; Pelling, J. C.; Koehler, K. J.; Birt, D. F.,
Cell-cycle arrest at G2/M and growth inhibition by apigenin in human colon
carcinoma cell lines. Mol. Carcinogen. 2000, 28, (2), 102-110.
135. Lu, J. C.; Feng, X. Z.; Sun, Q. S.; Lu, H. W.; Manabe, M.; Sugahara, K.; Ma, D. S.;
Sagara, Y.; Kodama, H., Effect of six flavonoid compounds from Ixeris sonchifolia
on stimulus-induced superoxide generation and tyrosyl phosphorylation in human
neutrophils. Clin. Chim. Acta 2002, 316, (1-2), 95-99.
136. Yamamoto, T.; Hsu, S.; Lewis, J.; Wataha, J.; Dickinson, D.; Singh, B.; Bollag, W.
B.; Lockwood, P.; Ueta, E.; Osaki, T.; Schuster, G., Green tea polyphenol causes
differential oxidative environments in tumor versus normal epithelial cells. J.
Pharmacol. Exp. Ther. 2003, 307, (1), 230-236.
137. Aoshima, H.; Okita, Y.; Hossain, S. J.; Fukue, K.; Mito, M.; Orihara, Y.; Yokoyama,
T.; Yamada, M.; Kumagai, A.; Nagaoka, Y.; Uesato, S.; Hara, Y., Effect of 3-O-
octanoyl-(+)-catechin on the responses of GABA(A) receptors and Na+/glucose
cotransporters expressed in Xenopus oocytes and on the oocyte membrane
potential. J. Agric. Food Chem. 2005, 53, (6), 1955-1959.

160
138. Arakawa, H.; Maeda, M.; Okubo, S.; Shimamura, T., Role of hydrogen peroxide in
bactericidal action of catechin. Biological & Pharmaceutical Bulletin 2004, 27, (3),
277-281.
139. Arakawa, H.; Kanemitsu, M.; Tajima, N.; Maeda, M., Chemiluminescence assay for
catechin based on generation of hydrogen peroxide in basic solution. Anal. Chim.
Acta. 2002, 472, (1-2), 75-82.
140. Aragawa, M.; Shigemitsu, T.; Suyama, K., Production of hydrogen peroxide by
polyphenols and polyphenol-rich beverages under quasi-physiological conditions.
Biosci. Biochem. Biotech. 2003, 67, (12), 2632-2640.
141. Fernandez, M. F.; Boris, S.; Barbes, C., Probiotic properties of human Lactobacilli
strains to be used in the gastrointestinal tract. J. Appl. Microbiol. 2003, 94, (3), 449-
455.
142. Kim, H.; Son, K.; Chang, H.; Kang, S., Anti-inflammatory plant flavonoids and
cellular action mechanisms. J. Pharmacol. Sci. 2004, 96, 229-245.
143. Escarpa, A.; Gonzalez, M. C., High-performance liquid chromatography with diode-
array detection for the determination of phenolic compounds in peel and pulp from
different apple varieties. J. Chromtogr. A 1998, 823, (1-2), 331-337.
144. Bruzone, A., Manzana deshidratada: Análisis de cadena alimentaria. En:
http://www.alimentosargentinos.gov.ar/0-
3/revistas/r_30/cadenas/manzana_deshidratada.htm 2006.
145. Wolfe, K.; Wu, X. Z.; Liu, R. H., Antioxidant activity of apple peels. J. Agric. Food
Chem. 2003, 51, (3), 609-614.
146. Wolfe, K. L.; Liu, R. H., Apple peels as a value-added food ingredient. J. Agric.
Food Chem. 2003, 51, (6), 1676-1683.

161
147. Vanzani, P.; Rossetto, M.; Rigo, A.; Vrhovsek, U.; Mattivi, F.; D'Amato, E.; Scarpa,
M., Major phytochemicals in apple cultivars: Contribution to peroxyl radical trapping
efficiency. J. Agric. Food Chem. 2005, 53, (9), 3377-3382.
148. Hassimotto, N. M. A.; Genovese, M. I.; Lajolo, F. M., Antioxidant activity of dietary
fruits, vegetables, and commercial frozen fruit pulps. J. Agric. Food Chem. 2005,
53, (8), 2928-2935.
149. Lata, B.; Przeradzka, M.; Binkowska, M., Great differences in antioxidant properties
exist between 56 apple cultivars and vegetation seasons. J. Agric. Food Chem.
2005, 53, (23), 8970-8978.
150. Schieber, A.; Hilt, P.; Streker, P.; Endreβ, H.; Rentschler, C.; Carle, R., A new
process for the combined recovery of pectin and phenolic compounds from apple
pomace. Innovative Food Sci. Emerg. Technol 2003, 4, 99-107.
151. Boyer, J.; Brown, D.; Liu, R. H., Uptake of quercetin and quercetin 3-glucoside from
whole onion and apple peel extracts by Caco-2 cell monolayers. J. Agric. Food
Chem. 2004, 52, (23), 7172-7179.
152. Alonso-Salces, R. M.; Ndjoko, K.; Queiroz, E. F.; Ioset, J. R.; Hostettmann, K.;
Berrueta, L. A.; Gallo, B.; Vicente, F., On-line characterisation of apple polyphenols
by liquid chromatography coupled with mass spectrometry and ultraviolet
absorbance detection. J. Chromatogr. A 2004, 1046, (1-2), 89-100.
153. Veberic, R.; Trobec, M.; Herbinger, K.; Hofer, M.; Grill, D.; Stampar, F., Phenolic
compounds in some apple (Malus domestica Borkh) cultivars of organic and
integrated production. Journal of the Science of Food and Agriculture 2005, 85,
(10), 1687-1694.
154. Chinnici, F.; Bendini, A.; Gaiani, A.; Riponi, C., Radical scavenging activities of
peels and pulps from cv. golden delicious apples as related to their phenolic
composition. J. Agric. Food Chem. 2004, 52, (15), 4684-4689.

162
155. McGhie, T. K.; Hunt, M.; Barnett, L. E., Cultivar and growing region determine the
antioxidant polyphenolic concentration and composition of apples grown in New
Zealand. J. Agric. Food Chem.2005, 53, (8), 3065-3070.
156. Lister, C. E.; Lancaster, J. E.; Sutton, K. H.; Walker, J. R. L., Developmental
changes in the concentration and composition of flavonoids in skin of red and green
apple cultivar. Journal of the Science of Food and Agriculture 1994, 64, (2), 155-
161.
157. Tsao, R.; Yang, R.; Xie, S.; Sockovie, E.; Khanizadeh, S., Which polyphenolic
compounds contribute to the total antioxidant activities of apple? J. Agric. Food
Chem. 2005, 53, (12), 4989-4995.
158. Ding, M.; Lu, Y. J.; Bowman, L.; Huang, C. S.; Leonard, S.; Wang, L. Y.; Vallyathan,
V.; Castranova, V.; Shi, X. L., Inhibition of AP-1 and neoplastic transformation by
fresh apple peel extract. J. Biol. Chem. 2004, 279, (11), 10670-10676.
159. Kondo, S.; Tsuda, K.; Muto, N.; Ueda, J., Antioxidative activity of apple skin or flesh
extracts associated with fruit development on selected apple cultivars. Sci. Hortic.
Amsterdam. 2002, 96, (1-4), 177-185.
160. Pearson, D. A.; Tan, C. H.; German, J. B.; Davis, P. A.; Gershwin, M. E., Apple
juice inhibits human low density lipoprotein oxidation. Life Sci. 1999, 64, (21), 1913-
1920.
161. Eberhardt, M. V.; Lee, C. Y.; Liu, R. H., Nutrition - Antioxidant activity of fresh
apples. Nature 2000, 405, (6789), 903-904.
162. Olsson, M. E.; Gustavsson, K. E.; Andersson, S.; Nilsson, A.; Duan, R. D., Inhibition
of cancer cell proliferation in vitro by fruit and berry extracts and correlations with
antioxidant levels. J. Agric. Food Chem. 2004, 52, (24), 7264-7271.
163. Alberto, M.; Canavosio, M.; Nadra, M. Antimicobial effect of polyphenols from apple
skins on human bacterial pathogens. Electron. J. Biotechno. 2006, 9, (3), 205-209.

163
164. Lee, J.; Um, H.; K., H.; Park, C.; Kang, D.; Surh, Y., Natural urease inhibitors.
Cancer Prev. Res. 2006, 11, (2), 129-136.
165. Friedman, M., Overview of antibacterial, antitoxin, antiviral, and antifungal activities
of tea flavonoids and teas. Mol. Nutr. Food Res. 2007, 51, (1), 116-134.
166. Yahiro, K.; Shirasaka, D.; Tagashira, M.; Wada, A.; Morinaga, N.; Kuroda, F.; Choi,
O.; Inoue, M.; Aoyama, N.; Ikeda, M.; Hirayama, T.; Moss, J.; Noda, M., Inhibitory
effects of polyphenols on gastric injury by Helicobacter pylori VacA toxin.
Helicobacter 2005, 10, (3), 231-239.
167. Ruggiero, P.; Rossi, G.; Tombola, F.; Pancotto, L.; Lauretti, L.; Del Giudice, G.;
Zoratti, M., Red wine and green tea reduce H. pylori- or VacA-induced gastritis in a
mouse model. World J. Gastroentero. 2007, 13, (3), 349-354.
168. Hamauzu, Y.; Inno, T.; Kume, C.; Irie, M.; Hiramatsu, K., Antioxidant and
antiulcerative properties of phenolics from Chinese quince, quince, and apple fruits.
J. Agric. Food Chem. 2006, 54, (3), 765-772.
169. Shibusawa, Y.; Yanagida, A.; Isozaki, M.; Shindo, H.; Ito, Y., Separation of apple
procyanidins into different degrees of polymerization by high-speed counter-current
chromatography. J. Chromatogr. A 2001, 915, (1-2), 253-257.
170. Saito, M.; Hosoyama, H.; Ariga, T.; Kataoka, S.; Yamaji, N., Antiulcer activity of
grape seed extract and procyanidins. J. Agric. Food Chem. 1998, 46, (4), 1460-
1464.
171. Graziani, G.; D'Argenio, G.; Tuccillo, C.; Loguercio, C.; Ritieni, A.; Morisco, F.;
Blanco, C. D.; Fogliano, V.; Romano, M., Apple polyphenol extracts prevent
damage to human gastric epithelial cells in vitro and to rat gastric mucosa in vivo.
Gut 2005, 54, (2), 193-200.
172. Ding, S. Z.; Denning, T. L.; Wang, J.; Boldogh, I.; Mitra, S.; Crowe, S. E.,
Expression of a redox-sensitive DNA repair enzyme, apurinic/apyrimidinic

164
endonuclease-1 (APE-1)in human gastric epithelial cells and its upregulation by
Helicobacter pylori and reactive oxygen species. Gastroenterology 2000, 118, (4),
4824.
173. Zuniga-Feest, A.; Inostroza, P.; Vega, M.; Bravo, L.; Corcuera, L., Sugars and
enzyme activity in the grass Deschampsia antarctica. Antarctic Sci 2003, 15, 483-
491.
174. Kelm, M. A.; Johnson, J. C.; Robbins, R. J.; Hammerstone, J. F.; Schmitz, H. H.,
High-performance liquid chromatography separation and purification of cacao
(Theobroma cacao L.) procyanidins according to degree of polymerization using a
diol stationary phase. J. Agric. Food Chem. 2006, 54, (5), 1571-1576.
175. Robbins, R. J.; Leonczak, J.; Johnson, J. C.; Li, J. L.; Kwik-Uribe, C.; Prior, R. L.;
Gu, L. W., Method performance and multi-laboratory assessment of a normal phase
high pressure liquid chromatography-fluorescence detection method for the
quantitation of flavanols and procyanidins in cocoa and chocolate containing
samples. J. Chromatogr. A 2009, 1216, (24), 4831-4840.
176. Guyot, S.; Marnet, N.; Laraba, D.; Sanoner, P.; Drilleau, J. F., Reversed-phase
HPLC following thiolysis for quantitative estimation and characterization of the four
main classes of phenolic compounds in different tissue zones of a French cider
apple variety (Malus domestica var. Kermerrien). J. Agric. Food Chem. 1998, 46,
(5), 1698-1705.
177. Torres, J. L.; Lozano, C., Chromatographic characterization of proanthocyanidins
after thiolysis with cysteamine. Chromatographia 2001, 54, (7-8), 523-526.
178. Contreras-Domínguez, M.; S, S. G.; Marnet, N.; Petit, J. L.; Perraud-Gaime, I.;
Roussos, S.; Augur, C., Degradation of procyanidins by Aspergillus fumigatus:
identification of a novel aromatic ring cleavage product. Biochimie 2006, 12, 1899-
1908.

165
179. Selga, A.; Torres, J. L., Efficient preparation of catechin thio conjugates by one step
extraction/depolymerization of pine (Pinus pinaster) bark procyanidins. J. Agric.
Food Chem. 2005, 53, (20), 7760-7765.
180. Karonen, M.; Leikas, A.; Loponen, J.; Sinkkonen, J.; Ossipov, V.; Pihlaja, K.,
Reversed-phase HPLC-ESI/MS analysis of birch leaf proanthocyanidins after their
acidic degradation in the presence of nucleophiles. Phytochem. Analysis. 2007, 18,
(5), 378-386.
181. Jerez, M.; Tourino, S.; Sineiro, J.; Torres, J. L.; Nunez, M. J., Procyanidins from
pine bark: Relationships between structure, composition and antiradical activity.
Food Chem. 2007, 104, (2), 518-527.
182. Slyke, D. v.; Archibald, R., Manometric, titrimetric, and colorimetric methods for
measurement of ureasa activity. J. Biol. Chem. 1944, (154), 623-642.
183. Tanaka, T.; Kawase, M.; Tani, S., alpha-hydroxyketones as inhibitors of urease.
Bioorgan. Med. Chem. 2004, 12, (2), 501-505.
184. Hu, L. T.; Mobley, H. L. T., Purification and N-terminal analysis of urease from
Helicobacter pylori. Infect. Immun. 1990, 58, (4), 992-998.
185. Xiao, Z. P.; Shi, D. H.; Li, H. Q.; Zhang, L. N.; Xu, C.; Zhu, H. L., Polyphenols based
on isoflavones as inhibitors of Helicobacter pylori urease. Bioorgan. Med. Chem.
2007, 15, (11), 3703-3710.
186. Schwartz, J. T.; Allen, L. A. H., Role of urease in megasome formation and
Helicobacter pylori survival in macrophages. J. Leukocyte Biol. 2006, 79, (6), 1214-
1225.
187. Weatherburn, M., Phenol-hypochlorite reaction fro determination of ammonia. Anal.
Chem. 1967, 39, 971-974.

166
188. Mobley, H.; Warren, J., Urease-positive bacteriuria and obstruction of long-term
urinary catheters. J. Clin Microbiol 1987, 25, 2216-2217.
189. Schweinitzer, T.; Mizote, T.; Ishikawa, N.; Dudnik, A.; Inatsu, S.; Schreiber, S.;
Suerbaum, S.; Aizawa, S. I.; Josenhans, C., Functional characterization and
mutagenesis of the proposed behavioral sensor TlpD of Helicobacter pylori. Journal
of Bacteriology 2008, 190, (9), 3244-3255.
190. Sarker, S. D.; Nahar, L.; Kumarasamy, Y., Microtitre plate-based antibacterial assay
incorporating resazurin as an indicator of cell growth, and its application in the in
vitro antibacterial screening of phytochemicals. Methods 2007, 42, (4), 321-324.
191. Takahashi, T.; Matsumoto, T.; Nakamura, M.; Matsui, H.; Kiyohara, H.; Sasakawa,
C.; Yamada, H., A novel in vitro infection model of Helicobacter pylori using mucin-
producing murine gastric surface mucous cells. Helicobacter 2004, 9, (4), 302-312.
192. Kamkaen, N.; Matsuki, Y.; Ichino, C.; Kiyohara; Yamada, H.; Linn, H., The isolation
of the anti-Helicobacter pylori compounds in seeds of Arctium lappa. The
Pharmaceutical and Health Science Journal 2006, 1, (2), 12-18.
193. Hynes, J.; Floyd, S.; Soini, A.; O´Connor, R.; Papkovsky, D., Fluorescence-based
cell viability screening assays using water-soluble oxygen probes. J. Biomol.
Screening 2003, 8, (264-272), 264.
194. Will, Y.; Hynes, J.; Ogurtsov, V. I.; Papkovsky, D. B., Analysis of mitochondrial
function using phosphorescent oxygen-sensitive probes. Nature Protocols 2006, 1,
(6), 2563-2572.
195. O´Mahony, F.; Papkovsky, S., Rapid Hight-Throughtput Assessment of aerobic
bacteria in complex samples by fluorescence-based oxygen respirometry. Appl.
Environ. Microb. 2006, 72, (2), 1279-1287.
196. Suzuki, M.; Miura, S.; Mori, M.; Kai, A.; Suzuki, H.; Fukumura, D.; Suematsu, M.;
Tsuchiya, M., Rebamipide, a novel antiulcer agent, attenuates Helicobacter pylori-

167
induced gastric mucosal cell injury associated with neutrophi-derived oxidants. Gut
1994, 35, (10), 1375-1378.
197. Lundqvist, H.; Dahlgren, C., Isoluminol-enhanced chemiluminescence: A sensitive
method to study the release of superoxide anion from human neutrophils. Free
Radical Bio. Med. 1996, 20, (6), 785-792.
198. Teneberg, S.; Jurstrand, M.; Karlsson, K. A.; Danielsson, D., Inhibition of
nonopsonic Helicobacter pylori-induced activation of human neutrophils by
sialylated oligosaccharides. Glycobiology 2000, 10, (11), 1171-1181.
199. Shimoyama, T.; Fukuda, S.; Liu, Q.; Nakaji, S.; Fukuda, Y.; Sugawara, K.,
Helicobacter pylori water soluble surface proteins prime human neutrophils for
enhanced production of reactive oxygen species and stimulate chemokine
production. J. Clin. Pathol. 2003, 56, (5), 348-351.
200. Viera, J.; Katarína, D.; Radomír, N.; Racková, L.; Májeková, M.; Dagmar, H., The
combined luminol/isoluminol chemiluminescence method for differentiating between
extracellular and intracellular oxidant production by neutrophils. Redox Report
2006, 11, (3), 110-116.
201. Spencer, J. P. E.; Chaudry, F.; Pannala, A. S.; Srai, S. K.; Debnam, E.; Rice-Evans,
C., Decomposition of cocoa procyanidins in the gastric milieu. Biochem. Biophys.
Res. Co. 2000, 272, (1), 236-241.
202. Goh, L. M. L.; Barlow, P. J., Flavonoid recovery and stability from Ginkgo biloba
subjected to a simulated digestion process. Food Chem. 2004, 86, (2), 195-202.
203. Gu, L. W.; Kelm, M.; Hammerstone, J. F.; Beecher, G.; Cunningham, D.; Vannozzi,
S.; Prior, R. L., Fractionation of polymeric procyanidins from lowbush blueberry and
quantification of procyanidins in selected foods with an optimized normal-phase
HPLC-MS fluorescent detection method. J. Agric. Food Chem. 2002, 50, (17),
4852-4860.

168
204. Bradford, M., A rapid and sensitive method for the quantitation of microgram
quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem
1976, 72, 248-254.
205. Cover, T. L.; Puryear, W.; Perezperez, G. I.; Blaser, M. J., Effect of urease on HeLa
cell vacuolation induced by Helicobacter pylori cytotoxin Infect. Immun. 1991, 59,
(4), 1264-1270.
206. Harris, P. R.; Cover, T. L.; Crowe, D. R.; Orenstein, J. M.; Graham, M. F.; Blaser,
M. J.; Smith, P. D., Helicobacter pylori cytotoxin induces vacuolation of primary
human mucosal epithelial cells. Infect. Immun. 1996, 64, (11), 4867-4871.
207. Lee, J. H.; Shim, J. S.; Chung, M. S.; Lim, S. T.; Kim, K. H., In Vitro Anti-Adhesive
Activity of Green Tea Extract against Pathogen Adhesion. Phytother. Res. 2009, 23,
(4), 460-466.
208. Dixon, M. F.; Genta, R. M.; Yardley, J. H.; Correa, P.; Batts, K. P.; Dahms, B. B.;
Filipe, M. I.; Haggitt, R. C.; Haot, J.; Hui, P. K.; Lechago, J.; Lewin, K.; Offerhaus, J.
A.; Price, A. B.; Riddell, R. H.; Sipponen, P.; Solcia, E.; Watanabe, H., Classification
and grading of gastritis - The updated Sydney System. American Journal of
Surgical Pathology 1996, 20, (10), 1161-1181.
209. Chen, X. Y.; van der Hulst, R. W. M.; Bruno, M. J.; van der Ende, A.; Xiao, S. D.;
Tytgat, G. N. J.; Ten Kate, F. J. W., Interobserver variation in the histopathological
scoring of Helicobacter pylori related gastritis. J. Clin. Pathol. 1999, 52, (8), 612-
615.
210. Bergamo, P.; Fedele, E.; Balestrieri, M.; Abrescia, P.; Ferrara, L., Measurement of
malondialdehyde levels in food by high-performance liquid chromatography with
fluorometric detection. J. Agric. Food Chem. 1998, 46, (6), 2171-2176.
211. Downsett, S.; Archila, L.; Segreto, V.; González, C.; Silva, A.; Vsatola, K.; Bartizek;
Kowolik, M., Helicobacter pylori Infection in Indigenous Families of Central America:

169
Serostatus and Oral and Fingernail Carriage. J. Clin. Microbiol. 1999, 37, 2456-
2460.
212. Guyot, S.; Marnet, N.; Drilleau, J. F., Thiolysis-HPLC characterization of apple
procyanidins covering a large range of polymerization states. J. Agric. Food Chem.
2001, 49, (1), 14-20.
213. Hevesi, T.; Blazics, B.; Kéry, A., Polyphenol composition and antioxidant capacity of
Epilobium species. J. Pharm. Biomed. Anal 2009, 49, (1), 26-31.
214. Monagas, M.; Garrido, I.; Lebron-Aguilar, R.; Bartolome, B.; Gomez-Cordoves, C.,
Almond (Prunus dulcis (Mill.) D.A. Webb) skins as a potential source of bioactive
polyphenols. J. Agric. Food Chem. 2007, 55, (21), 8498-8507.
215. Sun, J. Q.; Liang, F.; Bin, Y.; Li, P.; Duan, C. Q., Screening non-colored phenolics
in red wines using liquid chromatography/ultraviolet and mass spectrometry/mass
spectrometry libraries. Molecules 2007, 12, (3), 679-693.
216. Bystrom, L. M.; Lewis, B. A.; Brown, D. L.; Rodriguez, E.; Obendorf, R. L.,
Characterisation of phenolics by LC-UV/Vis, LC-MS/MS and sugars by GC in
Melicoccus bijugatus Jacq. 'Montgomery' fruits. Food Chem. 2008, 111, (4), 1017-
1024.
217. Tsimogiannis, D.; Samiotaki, M.; Panayotou, G.; Oreopoulou, V., Characterization
of flavonoid subgroups and hydroxy substitution by HPLC-MS/MS. Molecules 2007,
12, (3), 593-606.
218. LaTorre, G.; Saitta, M.; Vilasi, F.; Pellicano, T.; Dugo, G., Direct determination of
phenolics compounds in Sicilian wines by liquid chromatography with PDA and MS
detection. Food Chem. 2006, 94, (4), 640-650.
219. Lin, L. Z.; He, X. G.; Lindenmaier, M.; Yang, J.; Cleary, M.; Qiu, S. X.; Cordell, G.
A., LC-ESI-MS study of the flavonoid glycoside malonates of red clover (Trifolium
pratense). J. Agric. Food Chem. 2000, 48, (2), 354-365.

170
220. Wang, J.; Sporns, P., MALDI-TOF MS analysis of food flavonol glycosides. J. Agric.
Food Chem. 2000, 48, (5), 1657-1662.
221. Bilia, A. R.; Eterno, F.; Bergonzi, M. C.; Mazzi, G.; Vincieri, F. F., Evaluation of the
content and stability of the constituents of mother tinctures and tinctures: The case
of Crataegus oxyacantha L. and Hieracium pilosella L. J. Pharmaceut. Biomed.
2007, 44, (1), 70-78.
222. Innocenti, M.; Michelozzi, M.; Giaccherini, C.; Ieri, F.; Vincieri, F. F.; Mulinacci, N.,
Flavonoids and biflavonoids in tuscan berries of Juniperus communis L.: Detection
and quantitation by HPLC/DAD/ESI/MS. J. Agric. Food Chem. 2007, 55, (16), 6596-
6602.
223. Tiberti, L. A.; Yariwake, J. H.; Ndjoko, K.; Hostettmann, K., On-line LC/UV/MS
analysis of flavonols in the three apple varieties most widely cultivated in Brazil. J.
Brazil. Chem. Soc. 2007, 18, (1), 100-105.
224. Hilt, P.; Schieber, A.; Yildirim, C.; Arnold, G.; Klaiber, I.; Conrad, J.; Beifuss, U.;
Carle, R., Detection of phloridzin in strawberries (Fragaria x ananassa Duch.) by
HPLC-PDA-MS/MS and NMR spectroscopy. J. Agric. Food Chem. 2003, 51, (10),
2896-2899.
225. Krajewska, B.; Zaborska, W., Double mode of inhibition-inducing interactions of 1,4-
naphthoquinone with urease: Arylation versus oxidation of enzyme thiols. Bioorg.
Med. Chem. 2007, 15, (12), 4144-4151.
226. Bylund, J.; Samuelsson, M.; Collins, L. V.; Karlsson, A., NADPH-oxidase activation
in murine neutrophils via formyl peptide receptors. Exp. Cell Res. 2003, 282, (2),
70-77.
227. Noorgaard, A.; Andersen, L.; Nielsen, H., Neutrophil degranulation by Helicobacter
pylori proteins. Gut 1995, 36, 354-357.

171
228. Bylund, J.; Karlsson, A.; Boulay, F.; Dahlgren, C., Lipopolysaccharide-induced
granule mobilization and priming of the neutrophil response to Helicobacter pylori
peptide Hp(2-20), which activates formyl peptide receptor-like 1. Infect. Immun.
2002, 70, (6), 2908-2914.
229. Dales-Rakotoarison, D.; Greisser, B.; Trtotin, F.; Brunet, C.; Luyckx, M.; Dine, T.;
Bailleul, F.; Cazin, M.; Cazin, J., Effects of Rosa canina fruits extract on neutrophil
respiratory burst. Phytother. Res. 2002, 12, (2), 157-161.
230. Kanashiro, A.; Kabeya, L. M.; Polizello, A. C. M.; Lopes, N. P.; Lopes, J. L. C.;
Lucisano-Valim, Y. M., Inhibitory activity of flavonoids from Lychnophora sp on
generation of reactive oxygen species by neutrophils upon stimulation by immune
complexes. Phytother. Res. 2004, 18, (1), 61-65.
231. Souza, J. G.; Tomei, R. R.; Kanashiro, A.; Kabeya, L. M.; Azzolini, A.; Dias, D. A.;
Salvador, M. J.; Lucisano-Valim, Y. M., Ethanolic crude extract and flavonoids
isolated from Alternanthera maritima: Neutrophil chemilummescence inhibition and
free radical scavenging activity. Z. Naturforsch. C. 2007, 62, (5-6), 339-347.
232. Wirth, H. P.; Beins, M. H.; Yang, M. Q.; Tham, K. T.; Blaser, M. J., Experimental
infection of Mongolian gerbils with wild-type and mutant Helicobacter pylori strains.
Infect. Immun. 1998, 66, (10), 4856-4866.
233. Akada, J. K.; Ogura, K.; Dailidiene, D.; Dailide, G.; Cheverud, J. M.; Berg, D. E.,
Helicobacter pylori tissue tropism: mouse-colonizing strains can target different
gastric niches. Microbiology-Sgm 2003, 149, 1901-1909.
234. Zheng, Q.; Chen, X. Y.; Shi, Y.; Xiao, S. D., Development of gastric
adenocarcinoma in Mongolian gerbils after long-term infection with Helicobacter
pylori. J. Gastroen. Hepatol. 2004, 19, (10), 1192-1198.
235. Day, A. S.; Jones, N. L.; Policova, Z.; Jennings, H. A.; Yau, E. K.; Shannon, P.;
Neumann, A. W.; Sherman, P. M., Characterization of virulence factors of mouse-

172
adapted Helicobacter pylori strain SS1 and effects on gastric hydrophobicity.
Digest. Dis. Sci. 2001, 46, (9), 1943-1951.
236. Kim, Y. J.; Kim, D. O.; Chun, K.; Shin, D. H.; Jung, H.; Lee, C. Y.; Wilson, D. B.,
Phenolic extraction from apple peel by cellulases from Thermobifida fusca. J. Agric.
Food Chem. 2005, 53, (24), 9560-9565.
237. Alonso-Salces, R. M.; Korta, E.; Barranco, A.; Berrueta, L. A.; Gallo, B.; Vicente, F.,
Pressurized liquid extraction for the determination of polyphenols in apple. J.
Chromatogr. A 2001, 933, (1-2), 37-43.
238. Saleh, Z.; Stanley, R.; Nigam, M., Extraction of Polyphenolics from Apple Juice by
Foam Fractionation. International Journal of Food Engineering 2006, 2, (2), art 2.
239. Garcia-Marino, M.; Rivas-Gonzalo, J. C.; Ibanez, E.; Garcia-Moreno, C., Recovery
of catechins and proanthocyanidins from winery by-products using subcritical water
extraction. Anal. Chim. Acta 2006, 563, (1-2), 44-50.
240. Adil, I. H.; Cetin, H. I.; Yener, M. E.; Bayindirli, A., Subcritical (carbon dioxide plus
ethanol) extraction of polyphenols from apple and peach pomaces, and
determination of the antioxidant activities of the extracts. J. Supercrit. Fluid. 2007,
43, (1), 55-63.
241. Tsao, R.; Yang, R.; Christopher, J.; Zhu, Y.; Zhu, H. H., Polyphenolic profiles in
eight apple cultivars using high-performance liquid chromatography (HPLC). J.
Agric. Food Chem. 2003, 51, (21), 6347-6353.
242. McDonald, M. S.; Hughes, M.; Burns, J.; Lean, M. E. J.; Matthews, D.; Crozier, A.,
Survey of the free and conjugated myricetin and quercetin content of red wines of
different geographical origins. J. Agric. Food Chem. 1998, 46, (2), 368-375.
243. Faustino, R. S.; Sobrattee, S.; Edel, A. L.; Pierce, G. N., Comparative analysis of
the phenolic content of selected Chilean, Canadian and American Merlot red wines.
Mol. Cell. Biochem. 2003, 249, (1-2), 11-19.

173
244. Fernandez, K.; Kennedy, J. A.; Agosin, E., Characterization of Vitis vinifera L. cv.
Carmenere grape and wine proanthocyanidins. J. Agric. Food Chem. 2007, 55, (9),
3675-3680.
245. Gu, L. W.; House, S. E.; Wu, X. L.; Ou, B. X.; Prior, R. L., Procyanidin and catechin
contents and antioxidant capacity of cocoa and chocolate products. J. Agric. Food
Chem. 2006, 54, (11), 4057-4061.
246. Tarascou, I.; Ducasse, M.; Dufourc, E.; Moskau, D.; Fouquet, E.; Laguerre, M.;
Pianet, I., Structural and conformational analysis of two native procyanidin trimers.
Magn. Reson. Chem. 2006, 45, (2), 157-166.
247. Kim, J. M.; Shin, J. E.; Bae, E. A.; Han, M. J.; Kim, D. H., Inhibitory effect of
ponciretin on Helicobacter pylori VacA toxin-induced vacuolation in HeLa cells. J.
Microbiol. Biotechn. 2006, 16, (1), 46-51.
248. Matsumoto, T.; Takahashi, T.; Yamada, H., A novel approach for screening of new
anti-Helicobacter pylori substances. Biol. Pharm. Bull. 2008, 31, (1), 143-145.
249. Hashimoto, T.; Aga, T.; Tabuchi, A.; Shibuya, T.; Fukuda, S., Anti-Helicoabcter
pylori compounds in Brazilian Propolis. Nat. Med 1998, 52, (6), 518-520.
250. Bae, E. A.; Han, M. J.; Kim, D. H., In vitro anti-Helicobacter pylori activity of some
flavonoids and their metabolites. Planta Med. 1999, 65, (5), 442-443.
251. Isobe, T.; Doe, M.; Morimoto, Y.; Nagata, K.; Ohsaki, A., The anti-Helicobacter
pylori flavones in a Brazilian plant, Hyptis fasciculata, and the activity of
methoxyflavones. Biol. Pharm. Bull. 2006, 29, (5), 1039-1041.
252. Zhu, Y.; Zhang, P.; Yu, H.; Li, J.; Wang, M.; Zhao, W., Anti-Helicobacter pylori and
thrombin inhibitory components from Chinese Dragon´s Blood, Dracaena
cochinchinensis. J. Nat. Prod. 2007, 70, (10), 1570-1577.

174
253. Zhang, L.; Kong, Y. H.; Wu, D. L.; Zhang, H. T.; Wu, J.; Chen, J.; Ding, J. P.; Hu, L.
H.; Jiang, H. L.; Shen, X., Three flavonoids targeting the beta-hydroxyacyl-acyl
carrier protein dehydratase from Helicobacter pylori: Crystal structure
characterization with enzymatic inhibition assay. Protein Sci. 2008, 17, (11), 1971-
1978.
254. Zhang, L.; Liu, W. Z.; Hu, T. C.; Du, L.; Luo, C.; Chen, K. X.; Shen, X.; Jiang, H. L.,
Structural basis for catalytic and inhibitory mechanisms of beta-hydroxyacyl-acyl
carrier protein dehydratase (FabZ). J. Biol. Chem. 2008, 283, (9), 5370-5379.
255. Kong, Y. H.; Zhang, L.; Yang, Z. Y.; Han, C.; Hu, L. H.; Jiang, H. L.; Shen, X.,
Natural product juglone targets three key enzymes from Helicobacter pylori:
inhibition assay with crystal structure characterization. Acta Pharm. Sinic. 2008, 29,
(7), 870-876.
256. Park, B. S.; Lee, H. K.; Lee, S. E.; Piao, X. L.; Takeoka, G. R.; Wong, R. Y.; Ahn, Y.
J.; Kim, J. H., Antibacterial activity of Tabebuia impetiginosa Martius ex DC
(Taheebo) against Helicobacter pylori. J. Ethnopharmacol. 2006, 105, (1-2), 255-
262.
257. Nagata, K.; Sone, N.; Tamura, T., Inhibitory activities of lansoprazole against
respiration in Helicobacter pylori. Antimicrob. Agents Ch. 2001, 45, (5), 1522-1527.
258. Li, H. Q.; Xu, C.; Li, H. S.; Xiao, Z. P.; Shi, L.; Zhu, H. L., Metronidazole-flavonoid
derivatives as anti-Helicobacter pylori agents with potent inhibitory activity against
HPE-Induced interleukin-8 production by AGS cells. Chemmedchem 2007, 2, (9),
1361-1369.
259. Mobley, H. L. T.; Island, M. D.; Hausinger, R. P., Molecular biology of microbial
ureases Microbiol. Rev. 1995, 59, (3), 451-480.
260. Jabri, E.; Carr, M. B.; Hausinger, R. P.; Karplus, P. A., The crystal structure of
urease from Klebsiella aerogenes. Science 1995, 268, (5213), 998-1004.

175
261. Benini, S.; Rypniewski, W. R.; Wilson, K. S.; Miletti, S.; Ciurli, S.; Mangani, S., A
new proposal for urease mechanism based on the crystal structures of the native
and inhibited enzyme from Bacillus pasteurii: why urea hydrolysis casts two nickels.
Structure 1999, 7, (2), 205-216.
262. Mamiya, G.; Takishima, K.; Masakuni, M.; Kayumi, T.; Ogawa, K.; Sekita, T.,
Complete amino acid sequence of jack bean urease. P. JPN. Acad. B-Phys. 1985,
Series B61, 395-398.
263. Ciurli, S.; Benini, S.; Rypniewski, W. R.; Wilson, K. S.; Miletti, S.; Mangani, S.,
Structural properties of the nickel ions in urease: novel insights into the catalytic and
inhibition mechanisms. Coordin. Chem. Rev. 1999, 192, 331-355.
264. Takishima, K.; Suga, T.; Mamiya, G., The structure of Jack bean urease-The
complete amino acid sequence, limited proteolysis and reactive cysteine residues
Eur. J. Biochem. 1988, 175, (1), 151-165.
265. Kot, M., Inhibition of jack bean urease by p-benzoquinone: elucidation of the role of
thiols and reversibility of the process. J. Enzym. Inhib. 2006, 21, (6), 697-701.
266. Krajewska, B.; Zaborskaa, W., Double mode of inhibition-inducing interactions of
1,4-naphthoquinone with urease: Arylation versus oxidation of enzyme thiols
Bioorg. Med. Chem. 2007, 15, (12), 4144-4151.
267. Krajewska, B.; Zaborska, W., Jack bean urease: The effect of active-site binding
inhibitors on the reactivity of enzyme thiol groups. Bioorg. Med. Chem. 2007, 35,
(5), 355-365.
268. Zaborska, W.; Krajewska, B.; Kot, M.; Karcz, W., Quinone-induced inhibition of
urease: Elucidation of its mechanisms by probing thiol groups of the enzyme.
Bioorg. Chem. 2007, 35, (3), 233-242.

176
269. Zaborska, W.; Karcz, W.; Kot, M.; Juszkiewicz, A., Modification of jack bean urease
thiols by thiosulphinates contained in garlic extract: DTNB titration studies. Food
Chem. 2009, 112, (1), 42-45.
270. Bartolome, B.; Estrella, I.; Hernandez, M. T., Interaction of low molecular weight
phenolics with proteins (BSA). J. Food Sci. 2000, 65, (4), 617-621.
271. Fu, H. M.; Bylund, J.; Karlsson, A.; Pellme, S.; Dahlgren, C., The mechanism for
activation of the neutrophil NADPH-oxidase by the peptides formyl-Met-Leu-Phe
and Trp-Lys-Tyr-Met-Val-Met differs from that for interleukin-8. Immunology 2004,
112, (2), 201-210.
272. Limasset, B.; Ledoucen, C.; Dore, J. C.; Ojasoo, T.; Damon, M.; Depaulet, A. C.,
Effects of flavonoids on the release of reactive oxygen species by stimulated human
neutrophils-Multivariate analysis of structure-activity-relatioships (SAR) Biochem.
Pharmacol. 1993, 46, (7), 1257-1271.
273. Krol, W.; Czuba, Z.; Scheller, S.; Paradowski, Z.; Shani, J., Structure-activity
relationship in the ability of flavonols to inhibit chemiluminescence. J.
Ethnopharmacol. 1994, 41, (1-2), 121-126.
274. Limasset, B.; Ojasoo, T.; le Doucen, C.; Dore, J. C., Inhibition of
chemiluminescence in human PMNs by monocyclic phenolic acids and flavonoids.
Planta Med. 1999, 65, (1), 23-29.
275. Moreira, M. R.; Kanashiro, A.; Kabeya, L. M.; Polizello, A. C. M.; Azzolini, A.; Do
Amaral, A. T.; Curti, C.; Oliveira, C. A.; Lucisano-Valim, Y. M., Flavonoids effects in
neutrophil respiratory burst triggered by Fc-gamma and complement receptors:
influence of B-ring hydroxylation pattern and physicochemical properties. Free
Radical Res. 2006, 40, S107-S107.
276. Chen, G.; Lu, H. W.; Wang, C. L.; Yamashita, K.; Manabe, M.; Meng, Z. Y.; Xu, S.
X.; Kodama, H., Effect of five flavonoid compounds isolated from leaves of
Diospyros kaki on stimulus-induced superoxide generation and tyrosyl

177
phosphorylation of proteins in human neutrophils. Clin. Chim. Acta 2002, 326, (1-2),
169-175.
277. Satin, B.; Del Giudice, G.; Della Bianca, V.; Dusi, S.; Laudanna, C.; Tonello, F.;
Kelleher, D.; Rappuoli, R.; Montecucco, C.; Rossi, F., The neutrophil-activating
protein (HP-NAP) of Helicobacter pylori is a protective antigen and a major
virulence factor. J. Exp. Med. 2000, 191, (9), 1467-1476.
278. Danielsson, D.; Farmery, S. M.; Blomberg, B.; Perry, S.; Rautelin, H.; Crabtree, J.
E., Co-expression in Helicobacter pylori of cagA and non-opsonic neutrophil
activation enhances the association with peptic ulcer disease. J. Clin. Pathol. 2000,
53, (4), 318-321.
279. Unemo, M.; Aspholm-Hurtig, M.; Ilver, D.; Bergstrom, J.; Boren, T.; Danielsson, D.;
Teneberg, S., The sialic acid binding SabA adhesin of Helicobacter pylori is
essential for nonopsonic activation of human neutrophils. J. Biol. Chem. 2005, 280,
(15), 15390-15397.
280. Arnhold, J.; Mueller, S.; Arnold, K.; Sonntag, K., Mechanism of inhibition of
chemiluminescence in the oxidation of luminol by sodium-hypochlorite. J. Biolum.
Chemilum. 1993, 8, (6), 307-313.
281. Winterbourn, C. C.; Brennan, S. O., Characterization of the oxidation products of
the reaction between reduced glutathione and hypochlorous acid. Biochem. J.
1997, 326, 87-92.
282. Peskin, A. V.; Winterbourn, C. C., Kinetics of the reactions of hypochlorous acid and
amino acid chloramines with thiols, methionine, and ascorbate. Free Radical Bio.
Med. 2001, 30, (5), 572-579.
283. Selloum, L.; Djelili, H.; Sebihi, L.; Arnhold, J., Scavenger effect of flavonols on
HOCl-induced luminol chemiluminescence. Luminescence 2004, 19, (4), 199-204.

178
284. Mori, A.; Nishino, C.; Enoki, N.; Tawata, S., Citotoxicity of plant flavonoids against
HeLa cells. Phytochemistry 1988, 27, (4), 1017-1020.
285. Plochmann, K.; Korte, G.; Koutsilieri, E.; Richling, E.; Riederer, P.; Rethwilm, A.;
Schreier, P.; Scheller, C., Structure-activity relationships of flavonoid-induced
cytotoxicity on human leukemia cells. Arch. Biochem. Biophys. 2007, 460, (1), 1-9.
286. Szliszka, E.; Czuba, Z. P.; Jernas, K.; Krol, W., Dietary flavonoids sensitize HeLa
cells to tumor necrosis factor-related apoptosis-inducing ligand (TRAIL). Int. J. Mol.
Sci. 2008, 9, (1), 56-64.
287. Wittschier, N.; Faller, G.; Hensel, A., Pelargonium sidoides extract EPs 7630
inhibits adhesion of Helicobacter pylori to human gastric mucosa. Planta Med.
2006, 72, (11), P239.
288. Wittschier, N.; Faller, G.; Hensel, A., An extract of Pelargonium sidoides (EPs 7630)
inhibits in situ adhesion of Helicobacter pylori to human stomach. Phytomedicine
2007, 14, (4), 285-288.
289. Rohdewald, P.; Beil, W., In vitro inhibition of Helicobacter pylori growth and
adherence to gastric mucosal cells by Pyenogenol (R). Phytother. Res. 2008, 22,
(5), 685-688.
290. Beil, W.; Kilian, P., EPs (R) 7630, an extract from Pelargonium sidoides roots
inhibits adherence of Helicohacter pylori to gastric epithelial cells. Phytomedicine
2007, 14, 5-8.
291. Wittschier, N.; Lengsfeld, C.; Vorthems, S.; Stratmann, U.; Ernst, J. F.; Verspohl, E.
J.; Hensel, A., Large molecules as anti-adhesive compounds against pathogens. J.
Pharm. Pharmacol. 2007, 59, (6), 777-786.
292. Wittschier, N.; Faller, G.; Hensel, A., Aqueous extracts and polysaccharides from
Liquorice roots (Glycyrrhiza glabra L.) inhibit adhesion of Helicobacter pylori to
human gastric mucosa. J. Ethnopharmacol. 2009, 125, (2), 218-223.

179
293. Woo, J. S.; Ha, B. H.; Kim, T. G.; Lim, Y. G.; Kim, K. H., Inhibition of Helicobacter
pylori adhesion by acidic polysaccharide isolated from Artemisia capillaris. J.
Microbiol. Biotechn. 2003, 13, (6), 853-858.
294. Lee, J. H.; Park, E. K.; Uhm, C. S.; Chung, M. S.; Kim, K. H., Inhibition of
Helicobacter pylori adhesion to human gastric adenocarcinoma epithelial cells by
acidic polysaccharides from Artemisia capillaris and Panax ginseng. Planta Med.
2004, 70, (7), 615-619.
295. Lee, J. H.; Shim, J. S.; Lee, J. S.; Kim, J. K.; Yang, I. S.; Chung, M. S.; Kim, K. H.,
Inhibition of pathogenic bacterial adhesion by acidic polysaccharide from green tea
(Camellia sinensis). J. Agric. Food Chem. 2006, 54, (23), 8717-8723.
296. Ascenio, F.; Gama, N. L.; De Philippis, R.; Ho, B., Effectiveness of Cyanothece spp.
and Cyanospira capsulata exocellular polysaccharides as antiadhesive agents for
blocking attachment of Helicobacter pylori to human gastric cells. Folia Microbiol.
2004, 49, (1), 64-70.
297. Lengsfeld, C.; Deters, A.; Faller, G.; Hensel, A., High molecular weight
polysaccharides from black currant seeds inhibit adhesion of Helicobacter pylori to
human gastric mucosa. Planta Med. 2004, 70, (7), 620-626.
298. Loke, M. F.; Lui, S. Y.; Ng, B. L.; Gong, M.; Ho, B., Antiadhesive property of
microalgal polysaccharide extract on the binding of Helicobacter pylori to gastric
mucin. Fems Immunol. Med. Mic. 2007, 50, (2), 231-238.
299. Pastene, E.; Troncoso, M.; Figueroa, G.; Alarcon, J.; Speisky, H., Association
between Polymerization Degree of Apple Peel Polyphenols and Inhibition of
Helicobacter pylori Urease. J. Agric. Food Chem. 2009, 57, (2), 416-424.
300. Takabayashi, F.; Harada, N.; Yamada, M.; Murohisa, B.; Oguni, I., Inhibitory effect
of green tea catechins in combination with sucralfate on Helicobacter pylori infection
in Mongolian gerbils. J. Gastroenterol. 2004, 39, (1), 61-63.

180
301. Brown, J. C.; Huang, G. H.; Haley-Zitlin, V.; Jiang, X. P., Antibacterial Effects of
Grape Extracts on Helicobacter pylori. Appl. Environ. Microbiol. 2009, 75, (3), 848-
852.
302. Akai, Y.; Nakajima, N.; Ito, Y.; Matsui, T.; Iwasaki, A.; Arakawa, Y., Green tea
polyphenols reduce gastric epithelial cell proliferation and apoptosis stimulated by
Helicobacter pylori infection. J. Clinical Biochem. Nutr. 2007, 40, (2), 108-115.
303. Lee, J.; Paek, N.; Kwon, O.; Hahm, K., Anti-inflammatory actions of probiotics
through activating suppressor of cytokine signaling (SOCS) expression and
signaling in Helicobacter pylori infection: A novel mechanism. J. Gastroenterol.
Hepatol. 2010, 25, 194-202.
304. Yang, L. C.; Shun, C. T.; Chien, C. T.; Wang, T. H., Effective Prevention and
Treatment of Helicobacter pylori Infection Using a Combination of Catechins and
Sialic Acid in AGS Cells and BALB/c Mice. J. Nutr. 2008, 138, (11), 2084-2090.
305. Gonzalez-Segovia, R.; Quintanar, J. L.; Salinas, E.; Ceballos-Salazar, R.; Aviles-
Jimenez, F.; Torres-Lopez, J., Effect of the flavonoid quercetin on inflammation and
lipid peroxidation induced by Helicobacter pylori in gastric mucosa of guinea pig.
Journal of Gastroenterol. 2008, 43, (6), 441-447.
306. Sheu, B.; Yang, H.; Yeh, Y.; Wu, J., Helicobacter pylori colonization of the human
gastric epitehlium: A bug´s firts step is a novel target for us. J Gastroenterol.
Hepatol. 2010, 25, 26-32.
307. Pastene, E.; Speisky, H.; Troncoso, M.; Alarcon, J.; Figueroa, G., In Vitro Inhibitory
Effect of Apple Peel Extract on the Growth of Helicobacter pylori and Respiratory
Burst Induced on Human Neutrophils. J. Agric. Food Chem 2009, 57, (17), 7743-
7749.