universidad del mar -...
TRANSCRIPT

Universidad del Mar
Campus Puerto Ángel
Comparación del patrón reproductivo entre Ophiocoma alexandri y Ophiocoma aethiops (Echinodermata: Ophiuroidea) y su relación con las
variables ambientales temperatura, luminosidad, horas de luz, pluviosidad, clorofilas y salinidad.
TESIS
Que como parte de los requisitos para obtener el grado de
Maestro en Ciencias: Ecología Marina
Presenta:
Biol. Catalina Aguilar Duarte
Dirigido por:
Francisco Benítez Villalobos. PhD.
Oaxaca, México 2011

Puerto Ángel, Oaxaca, septiembre del 2011
DR. EDGAR ROBLES ZAVALA JEFE DE LA DIVISIÓN DE ESTUDIOS DE POSTGRADO DE LA UNIVERSIDAD DEL MAR P R E S E N T E Después de haber analizado y evaluado la tesis: “Comparación del patrón reproductivo entre Ophiocoma alexandri y Ophiocoma aethiops (Echinodermata: Ophiuroidea) y su relación con las variables ambientales temperatura, luminosidad, horas de luz, pluviosidad, clorofilas y salinidad” que presenta la Bióloga Catalina Aguilar Duarte. Por este conducto, le comunicamos que la tesis si cumple con los requisitos académicos para que el (la) citado (a) tesista presente el correspondiente examen profesional. Sin más por el momento, quedamos de Usted.
A t e n t a m e n t e ______________________ ______________________ Dr. Eugenio Carpizo Ituarte Dr. Martín Brogger Subdirector del I.I.O -U.A.B.C Inv. Museo Arg. Cs. Naturales
Revisor Revisor ______________________ ______________________ MCs. Pablo Torres Hernández Dra. Tamara Rubilar Prof. Inv. Universidad del Mar Inv. Centro Nal. Patagónico
Revisor Universidad Nal. del Comahue Revisor
___________________________________ Dr. Francisco Benítez Villalobos
Director del Instituto de Recursos Universidad del Mar
Director de Tesis

Resumen Uno de los objetivos más importantes de la ecología, es explicar la evolución de las historias de vida incluidos los aspectos relacionados con la reproducción. En este trabajo se caracterizaron y compararon los patrones reproductivos de Ophiocoma aethiops y Ophiocoma alexandri, además se evaluaron las relaciones de los mismos con las variaciones de los factores ambientales que incluyeron temperatura, clorofila, pluviosidad, salinidad y horas de luz. Para caracterizar los patrones reproductivos se hicieron 12 muestreos mensuales en los que se coleccionaron aproximadamente 20 individuos de cada especie en cada muestreo. Los ejemplares fueron procesados para obtener los valores mensuales del índice gonadal (IG), frecuencias de tallas de los ovocitos y fecundidad real. Además se registraron los valores mensuales de las variables ambientales, las cuales fueron comparadas con las variaciones en el IG. Se identificaron y describieron las etapas reproductivas del ciclo de ambas especies. Se encontró una correlación significativa y positiva de ambos ciclos con las variables temperatura, horas de luz y pluviosidad, sin embargo el análisis de regresión lineal múltiple vinculó a estas de forma diferente con ambas especies. Así, la temperatura presentó un valor de correlación de 0.73 con el IG de O. alexandri y las variables horas de luz y pluviosidad presentaron un valor de 0.74 de correlación con el IG O. aethiops. Entre ambas especies se encontraron diferencias significativas con respecto a los valores de IG. Sin embargo, los ciclos de ambas especies tienen el mismo comportamiento temporal. Se describieron cuatro estadios reproductivos en el caso de las hembras y tres para los machos. Los resultados arrojaron información que concuerda con las temporadas reproductivas encontradas para las mismas especies en Panamá. Se establece la necesidad de estudios de laboratorio futuros, con relación a las causas próximas y últimas, y el desarrollo embrionario y larval, para continuar describiendo las historias de vida de O. aethiops y O. alexandri en lo relacionado con sus patrones de reproducción y desarrollo. Palabras clave: ofiuros, fecundidad, índice gónadal.

Dedicatoria
A mi familia que siempre estuvo presente en la distancia.

Agradecimientos Al CONACYT por brindarme una beca de dos años de manutención, gracias a la cual pude costear mi estadía en Puerto Ángel durante ese tiempo. Y a la UMAR por la beca de colegiatura. A mi mamá, quien me mantuvo con su apoyo económico en todos los meses que ya no contaba con la beca CONACYT. Al profesor Francisco Benítez PhD. quien fungió como mi asesor de tesis y se encargó de facilitarme los medios para mi trabajo. A los doctores Andrés López P, Edgar Robles Z y Ma Auxilio Esparza, quienes me brindaron su apoyo (materiales y logística) para la ejecución de los muestreos y el procesamiento de la muestra. A los maestros Antonio López S y Francisco Becerril, por su compañía en algunas inmersiones y a Imelda por el mismo motivo. A los revisores por sus comentarios oportunos que generaron un debate corto e interesante, que me permitió tener un punto de vista diferente sobre el grueso de este trabajo. También por el tiempo dedicado, la prontitud en las revisiones y la retroalimentación. A cada una de las personas que desinteresadamente colaboraron, a quienes estuvieron presentes y me brindaron su calor humano. A los amigos, allá en Colombia, quienes siempre me animaron a seguir adelante y desearon lo mejor para mi.

Índice
Introducción y antecedentes................................................................................................ 1
Definición del problema e hipótesis de investigación........................................................ 6
Características taxonómicas................................................................................................ 6
Objetivo general ................................................................................................................... 8
Objetivos específicos.......................................................................................................... 8
Área de estudio ..................................................................................................................... 9
Método................................................................................................................................. 10
Muestreo y procesamiento de las muestras ..................................................................... 10
Proceso histológico.......................................................................................................... 11
Estimación de parámetros reproductivos ........................................................................ 11
Resultados ........................................................................................................................... 14
Discusión ............................................................................................................................. 32
Conclusiones ....................................................................................................................... 37
Literatura Citada ............................................................................................................... 38

Índice de figuras Figura 1. Ophiocoma aethiops Lütken, 1859. A) vista dorsal, B) vista ventral.................... 7 Figura 2. Ophiocoma alexandri Lyman, 1860. A) vista dorsal, B) vista ventral.................. 7 Figura 3. Caracteres relevantes para la diferenciación de Ophiocoma aethiops y O.
alexandri (escudo oral, papilas dentales y espinas de los brazos). ...................... 8 Figura 4. Mapa de la Bahía Estacahuite (Oaxaca, México). En la parte superior se
encuentra el mapa de México y el punto indica la ubicación de la Bahía de Estacahuite. ........................................................................................................ 10
Figura 5. Promedios mensuales (+/- DS) del Índice Gonadal. A) Ophiocoma aethiops y
O. alexandri, B) O. aethiops hembras y machos por separado, C) O. alexandri hembras y machos por separado. ....................................................... 17
Figura 6. Promedios mensuales (+/- DS) de fecundidad para Ophiocoma aethiops y O.
alexandri. ........................................................................................................... 18 Figura 7. Frecuencia de tallas de ovocitos de Ophiocoma aethiops a lo largo del año de
muestreo. ............................................................................................................ 20 Figura 8. Frecuencia de tallas de ovocitos de Ophiocoma alexandri a lo largo del año
de muestreo. ....................................................................................................... 21 Figura 9. Promedios mensuales de las variables ambientales que presentaron
correlación significativa con el Índice Gonadal para ambas especies. A) pluviosidad, B) horas de luz, C) temperatura..................................................... 22
Figura 10. Etapas reproductivas de las hembras de Ophiocoma aethiops. A)
crecimiento, B) y C) maduración, D) desove, E) postdesove y reactivación. Clf (células simi-lipofuscina), CG (celoma genital), Og (ovogonias), OPv (ovocitos previtelogénicos), OV (ovocitos vitelogénicos), SE (saco externo), SI (saco interno) y PF (membrana del folículo)................................................. 25
Figura 11. Etapas reproductivas de las hembras de Ophiocoma alexandri. A) y B)
crecimiento, C) maduración, D) desove, E) postdesove y reactivación. CG (celoma genital), Og (ovogonias), OPv (ovocitos previtelogénicos), OV (ovocitos vitelogénicos), SE (saco externo) y SI (saco interno).. ...................... 26
Figura 12. Etapas reproductivas de los machos de Ophiocoma aethiops. A) y B)
crecimiento-maduración, C) evacuación, D) postevacuación. CG (celoma genital), Clf (células simi-lipofuscina), Csg (capa espermatogénica), FM

(fibras musculares), PF (pared del folículo), SE (saco externo), SI (saco interno) y Sz (espermatozoides)......................................................................... 27
Figura 13. Imágenes de las diferentes etapas reproductivas de los machos de
Ophiocoma alexandri. A) y B) crecimiento-maduración, C) evacuación, D) postevacuación. CG (celoma genital), Csg (capa espermatogénica), FM (fibras musculares), SE (saco externo), SI (saco interno) y Sz (espermatozoides). ............................................................................................. 28
Figura 14. Porcentaje acumulado de hembras de Ophiocoma aethiops en las diferentes
temporadas reproductivas a lo largo del año de muestreo. 1. Crecimiento gonadal, 2. Maduración gonadal, 3. Desove de la gónada y 4. Postdesove y reactivación de la gónada. .................................................................................. 29
Figura 15. Porcentaje acumulado de hembras de Ophiocoma alexandri en las diferentes
temporadas reproductivas a lo largo del año de muestreo. 1. Crecimiento gonadal, 2. Maduración gonadal, 3. Desove de la gónada y 4. Postdesove y reactivación de la gónada. .................................................................................. 30
Figura 16. Porcentaje acumulado de machos de Ophiocoma aethiops en las diferentes
temporadas reproductivas a lo largo del año de muestreo. 1. Crecimiento y maduración de la gónada, 2. Evacuación de la gónada y 3. Postevacuación de la gónada........................................................................................................ 30
Figura 17. Porcentaje acumulado de machos de Ophiocoma alexandri en las diferentes
temporadas reproductivas a lo largo del año de muestreo. 1. Crecimiento y maduración de la gónada, 2. Evacuación de la gónada y 3. Postevacuación de la gónada........................................................................................................ 31

Índice de tablas Tabla 1. Porcentaje de machos y hembras coleccionados durante el periodo de muestreo.
............................................................................................................................ 14 Tabla 2. Porcentaje acumulado para frecuencias de ovocitos previtelogénicos,
vitelogénicos y en reabsorción de Ophiocoma aethiops y O. alexandri. P (ovocitos previtelogénicos), V (Ovocitos vitelogénicos) y R (ovocitos no evacuados o en reabsorción) .............................................................................. 16
Tabla 3. Valores de los coeficientes de correlación (en negrilla las correlaciones
significativas) y p encontrados con el análisis de correlación de Spearman aplicado entre las variables ambientales y los promedios mensuales del Índice Gonadal de Ophiocoma aethiops y O. alexandri. ................................... 19
Tabla 4. Valores de la regresión lineal paso a paso para las variables que presentaron
correlación significativa con el Índice Gonadal de Ophiocoma aethiops y O. alexandri. Las no incluidas son aquellas que el modelo no tomó en cuenta por no ser significativas en la regresión. ............................................................ 19

1
Introducción y antecedentes Uno de los objetivos más importantes de la ecología, es explicar la evolución de las historias de vida (McEdward y Morgan 2001, Grange 2005; Benítez-Villalobos 2005); dentro de los rasgos más sobresalientes de las historias de vida de los organismos se encuentran varios aspectos, de los cuales se pueden señalar los relacionados con la reproducción. Una historia de vida se puede tomar como la forma en la que la selección natural modifica la reproducción y el desarrollo, generando los patrones que observamos en la naturaleza (McEdward y Morgan 2001, Benítez-Villalobos 2005), ya que estos resultan de la selección de una distribución de energía óptima entre la reproducción y el crecimiento somático (Grange 2005). En virtud a ello, es necesario entender los efectos de la selección natural para un buen conocimiento de los rasgos de las historias de vida de las especies (Grange 2005). Para que las historias de vida sean estudiadas se requiere la identificación de las presiones selectivas que pudieran explicar la presencia de los patrones observados e identificar que causa las diferencias en el éxito reproductivo (Grange 2005). El estudio de un patrón de variación fenotípica o genotípica y la distribución de energía entre los diferentes procesos fisiológicos y reproductivos, contribuye a la estructuración de las historias de vida (McEdward y Miner 2003). En el siglo pasado se establecieron dos de las reglas más importantes en lo que a la reproducción de los invertebrados marinos concierne. La primera es la de Orton (1920), que sugiere que la reproducción es controlada por la temperatura del mar, esperando que las especies que viven en ambientes isotermos (ej. aguas profundas y polares) presenten reproducción continua. La segunda es la de Thorson (1950), la cual propone que las especies de aguas profundas y polares deberían tener un desarrollo directo por el alto riesgo de mortalidad asociado a un periodo prolongado de vida larvaria. Esto implica que la frecuencia del desarrollo pelágico (planctotrófico y lecitotrófico), decrece de las regiones ecuatoriales a las polares y que la condición demersal o incubadora comienza a ser dominante (Poulin y Féral 1996, Gillespie y McClintock 2007). Sin embargo, hoy se sabe que dichos postulados no son estrictos dado que hay muchas situaciones en que no se sigue ninguna de las dos reglas (Poulin y Féral 1996, Ramirez-Llodra 2001, Grange 2005, Gillespie y McClintock 2007). Dentro de los parámetros reproductivos, el análisis de la fecundidad adquiere importancia dado que ésta, tiene una estrecha relación con la distribución de energía y con otros aspectos reproductivos, como el tamaño del huevo y la frecuencia de incubación, dado que la relación fecundidad - talla del huevo es uno de los rasgos más importantes ya que tiene una relación directa con la inversión parental en la reproducción (Grange 2005). Por ello, estos parámetros han sido de interés para la biología evolutiva y la ecología durante décadas (Grange 2005). En términos muy amplios, la fecundidad se refiere a la cantidad de descendencia producida por una hembra, la cual puede ser expresada como el número total de ovocitos, óvulos o embriones producidos en un periodo de tiempo (temporada reproductiva, años, tiempo de

2
vida, etc.) (Ramirez-Llodra 2001, Benítez-Villalobos 2005, Grange 2005). Generalmente es considerada como el número de óvulos de una hembra en una temporada reproductiva (Grange 2005). La evolución de la talla del óvulo en invertebrados ha sido relacionada con diferentes factores pre y postcigóticos. La talla óptima es determinada por la relación entre la cantidad de la progenie y el éxito reproductivo de la misma. Las presiones selectivas pueden afectar la evolución de la talla del óvulo en diferentes momentos del ciclo de vida de una especie: 1) antes de que la larva o el embrión entre a la columna de agua, 2) durante la fase planctónica y 3) justo antes de la metamorfosis y el asentamiento (Grange 2005). Así, la talla del óvulo es influenciada por diferentes presiones selectivas como el éxito de fecundación, el tiempo de desarrollo larvario y la mortalidad de la larva (Benítez-Villalobos 2005). La selección natural se considera como la fuerza que direcciona el nivel de aprovisionamiento del óvulo, seguida de estrategias reproductivas con alto éxito reproductivo (McEdward y Miner 2003). Éstas estrategias han sido divididas en dos grupos, según sea el desarrollo larvario, teniendo entonces especies lecitotróficas y planctotróficas (Grange 2005). Vance (1973) propuso un modelo teórico para predecir la talla optima del óvulo, con una distribución bimodal con forma de U, que va de una escala de cero a uno y corresponde a los dos tipos de desarrollo larvario; donde el contenido energético del óvulo está distribuido en un intervalo que va de entre cero (óvulos de desarrollo planctotrófico) a uno (óvulos lecitotróficos) (McEdward y Janies 1997, Grange 2005). Sin embargo el autor asumió que solo los extremos de la distribución confieren un máximo en la eficiencia reproductiva (Sewell y Young 1997). En el caso de los equinodermos, entre la estrategia planctotrófica y la lecitotrófica existen óvulos que presentan intervalos de tallas que se traslapan (Sewell y Young 1997), lo que evidencia la existencia de un continuo, en el que los tamaños intermedios pueden ubicarse en una región en la que la transición de un estado reproductivo al otro podría ser direccionada por las fluctuaciones del medio (McEdward y Miner 2003). Este modelo ha sido modificado con el tiempo y se han incluido otras variables como la disponibilidad de alimento y el grado de dependencia del mismo (planctotrófico obligado, facultativo o lecitotrófico), debido a que las observaciones evidenciaban que las tallas exclusivas de cada una de las estrategias no se presentaban en los invertebrados marinos (McEdward y Janies 1997) y que los análisis filogenéticos indicaban que han ocurrido transiciones de un tipo de desarrollo al otro en repetidas ocasiones (McEdward y Miner 2003). El rendimiento reproductivo (cantidad de gametos producidos por un individuo en una temporada reproductiva), el índice reproductivo (cantidad de gametos por unidad de peso de un organismo en una temporada reproductiva) y el esfuerzo reproductivo (cantidad de gametos producidos por un individuo por cantidad total de producción en una temporada) son indicadores importantes del ciclo reproductivo de las especies. La variación intraespecífica resulta de la variación del rendimiento y el índice reproductivo. Para que el

3
individuo comience la etapa reproductiva necesita una talla mínima que lo hace funcional en términos de su madurez sexual (Benítez-Villalobos 2005). La biología de poblaciones toma como generalización que entre mayor sea el tamaño corporal, es más alto el éxito reproductivo. Esto se basa en la evidencia de que la producción de gametos es directamente proporcional al tamaño corporal (Benítez-Villalobos 2005). Sin embargo esto ha sido discutido ampliamente. Levitan (1991) sugiere que la influencia de la densidad en el éxito de la fecundación y el rendimiento reproductivo en invertebrados de aguas someras no es importante. Los modelos más recurrentes en las teorías de las historias de vida explican la covariación de los rasgos reproductivos basados en parámetros demográficos y su relación con las variables ambientales. La vitelogénesis consume una gran proporción de la energía tomada por el organismo y sus requerimientos varían según la vía vitelogénica de la especie (Ramirez-Llodra 2001). Debido a esto, las especies de larga vida y baja producción de óvulos tienen estrategias vitelogénicas que son consistentes con un medio en el cual el alimento es continuo o predecible y relativamente estable (Gage y Tyler 1991, Benítez-Villalobos 2005). La ovogénesis en una especie se encuentra relacionada filogenéticamente a la morfología del ovario, las vías vitelogénicas (inherentes a cada especie) y las estructuras digestivas relacionadas con la transferencia de energía a los ovarios. La fecundidad y la calidad del desove están directamente relacionadas con el estado nutricional del adulto y la distribución de los recursos energéticos (Benítez-Villalobos 2005). Los patrones vitelogénicos de los organismos también están filogenéticamente relacionados y son especie-específicos. Éstos tienen mayores implicaciones en los rasgos reproductivos, como la tasa de producción de óvulos, fecundidad y desarrollo larvario (Ramirez-Llodra 2001) En estudios sobre reproducción de equinodermos han sido usados los términos de fecundidad potencial y real, haciendo referencia a todos los ovocitos en el ovario o sólo a los que están cerca del desarrollo máximo (Ramirez-Llodra 2001). Para el caso de las especies que liberan gametos, se han desarrollado métodos indirectos de estimación de la fecundidad, siendo algunos no destructivos. Sin embargo, cuando los ovocitos se desarrollan en estructuras bien definidas (gónadas), no es posible inducir el desove o los ovocitos maduros no son visibles a través de las paredes de la gónada, entonces es necesaria la aplicación del método histológico. Éste proporciona una estimación del número de óvulos que serán liberados en el próximo evento reproductivo, pero no precisa el número de óvulos producidos, puesto que el número real de óvulos disponibles para la fecundación puede ser diferente de la cantidad de ovocitos que se encuentran maduros en los ovarios (Ramirez-Llodra 2001). Los procesos de liberación de gametos y las estrategias reproductivas en los ofiuros son extremadamente diversos y su plasticidad reproductiva ha resultado de la adaptación a condiciones ambientales variables (Hendler 1991, Grange et al. 2004). El ciclo reproductivo de varios ofiuros ha sido examinado y publicado, pero el efecto de los factores externos en la gametogénesis nunca había sido confirmado experimentalmente. Yamashita

4
e Iwata (1983) llevaron a cabo un estudio en condiciones in situ e in vitro, en el cual confirmaron la relación que presenta la temperatura con la duración de la espermatogénesis de Amphipholis kochii. Actualmente se continúa relacionando los ciclos reproductivos con las variables fisicoquímicas in situ; no obstante, los efectos no se han monitoreado simultáneamente en el campo experimental. En los estudios relacionados con ciclos reproductivos en ofiuros, generalmente no han sido considerados los factores últimos como las condiciones biológicas y físicas que seleccionan para un óptimo esfuerzo reproductivo (Hendler 1979), sin embargo este tipo de estudios ha aumentado con mayor énfasis en ambientes extremos como las latitudes altas y las aguas profundas (Selvakumaraswamy y Byrne 2000, Ramirez-Llodra 2001, Williams y Bentley 2002, Falkner y Byrne 2003, Tominaga et al. 2004, Soong et al. 2005, Grange 2005, Byrne et al. 2008). La fluctuación de la temperatura es considerada como uno de los factores más importantes que pueden afectar la periodicidad en el ciclo reproductivo en áreas templadas. En condiciones relativamente isotermas influyen otros factores como la salinidad, productividad, mareas y corrientes (Hendler 1979). Pearse (1974) sostiene que la estacionalidad de los ciclos reproductivos de invertebrados es más marcada conforme la localidad se aleja del ecuador. Sin embargo Stephenson (1934), indica que en aguas templadas la más mínima fluctuación en temperatura puede hacer que las especies liberen los gametos en los meses más cálidos. Thorson (1950) propone que las densidades de las larvas planctotróficas son iguales durante todo el año, lo que implica la existencia de condiciones ambientales que las soporten. De este modo los factores últimos que restringen las temporadas reproductivas operarían más en adultos y juveniles que en las larvas. En 1910 H. L. Clark sugirió que las especies simpátricas necesitan que sus ciclos reproductivos no se traslapen como mecanismo de aislamiento (Hendler 1979). La comparación del índice gonadal con el análisis histológico de las gónadas de los ofiuros muestra que estas variables están estrechamente relacionadas y que dicho índice refleja los cambios de la gónada y el tejido durante el transcurso del ciclo (Hendler 1973). El incremento en la talla de la gónada indica el crecimiento y acumulación de gametos, mientras que la disminución de ésta muestra la liberación de ellos (Hendler 1973). Dentro del género Ophiocoma se han realizado más estudios para las especies del Atlántico que del Pacífico Americano. Hasta hoy se cuenta con algunos detalles de los ciclos reproductivos (Hendler 1979, 1991); sin embrago, falta mucha más información para elaborar una buena caracterización de los mismos. Grave (1898) observó especimenes de O. echinata liberando gametos en julio y estimó que O. wendti lo había hecho al menos dos meses antes. Mortensen (1931) observó liberación de gametos en O. echinata para las poblaciones del sur de Bermuda. Mladenov (1976) observó en las poblaciones más al sur de Barbados que el periodo reproductivo de O. echinata comprendía de marzo a octubre (Hendler 1979, 1991). Hendler (1979) afirmó que O. echinata presenta un periodo de liberación de gametos entre septiembre y diciembre y que O. wendti presenta una tendencia a liberarlos antes que O. echinata, además que el inicio de su periodo de desove es variable. Para el Pacífico Panameño el mismo autor describe un descenso precipitado en el índice gonadal de O.

5
aethiops y O. alexandri entre los meses de septiembre y diciembre, el cual es seguido de un periodo no interrumpido de crecimiento gonadal. Para éstas dos especies, la liberación de gametos de poblaciones submareales fue precedida por la de las intermareales. Contrastando los resultados hallados, el autor observó que el tiempo que el índice gonadal permanece bajo, es mayor en el Pacífico que en el Atlántico y además afirma que O. echinata y O. wendti muestran más irregularidades en la progresión del crecimiento gonadal, aunque los picos reproductivos son comparables entre las poblaciones inter y submareales; a esto se suma que O. pumila muestra una variación mensual en el índice reproductivo, indicando una liberación de gametos continua durante todo el año (Hendler 1979). Al hacer una comparación de los datos obtenidos por los diferentes investigadores acerca de los ciclos de Ophiocoma en el Atlántico, Hendler (1991) concluye que O. echinata y O. wendti en el Atlántico Panameño están fuera de ritmo con respecto a sus conespecíficos de otras regiones, ya que para Panamá la temporada reproductiva ocurre entre otoño e invierno y en localidades más al norte durante el verano. El autor atribuye las discontinuidades a las diferencias en las temporadas de lluvias entre estos lugares, ya que en Panamá se presentan durante otoño e invierno, mientras que en los sitios de distribución más al norte la temporada de lluvias ocurre durante los meses más cálidos del año. Para el género Ophiocoma no hay información de la talla de primera madurez sexual (Hendler 1991) y tampoco ha habido estudios posteriores a lo reportado por Hendler en 1991. Sin embargo se sabe que las especies de éste género producen óvulos de aproximadamente 0,1 mm de diámetro. En el caso de O. echinata y O. pumila, y otras especies de Ophiocoma con un patrón de desarrollo conocido, se ha observado que presentan una larva planctotrófica. Así, juzgando por el tamaño y abundancia de ovocitos, O. wenditi también tendría larvas ophiopluteus (Hendler 1979) las cuales podrían ser planctotróficas. Dada la diferencia en la relación de producción primaria entre el Atlántico y el Pacífico en Panamá y la similitud en el ciclo reproductivo de las especies de Ophiocoma presentes para ambos lugares, se ha observado que existe una alta probabilidad de que el alimento no sea un factor último en determinar la temporada reproductiva, sin embargo no se ha hecho hasta hoy ningún estudio que corrobore dicha aseveración de forma directa para ninguna de las especies de este género. Tampoco se ha encontrado que la temperatura máxima, la salinidad, los patrones de circulación y las turbulencias sean factores que determinen directamente la distribución de las larvas; debido a esto se ha sugerido que para las comunidades de Panamá es necesario considerar como factor último para la reproducción una combinación de las variables en lugar de una sola variable aislada (Hendler 1979), lo anterior, tanto para los ofiuroideos como para el resto de equinodermos que han sido estudiados (Mercier y Hamel 2009).

6
Definición del problema e hipótesis de investigación Debido al estado del arte en el estudio de la reproducción de ofiuros, y específicamente para las especies pertenecientes al Pacífico Americano, se evidencia la necesidad de ahondar en las investigaciones que contribuyan con el conocimiento de los ciclos reproductivos y su relación con las variables ambientales. Para esto se abordó el estudio de los ciclos reproductivos de O. aethiops y O. alexandri. En primera instancia se determinó si los eventos de liberación de gametos en ambas especies ocurren en la misma época del año. Luego, se identificó si existía alguna relación entre el patrón reproductivo encontrado para cada especie y las variaciones de los factores ambientales incluyendo temperatura, clorofila, pluviosidad, salinidad y horas de luz. Partiendo de lo anterior se pudo responder a las siguientes preguntas: ¿Cuál es el patrón reproductivo de O. aethiops y O. alexandri?, ¿Existe alguna diferencia entre los patrones de reproducción de O. aethiops y O. alexandri?, ¿Cómo se asocian los cambios en los parámetros ambientales (temperatura, clorofila, pluviosidad, salinidad y horas de luz) con los patrones reproductivos de O. aethiops y O. alexandri? De este modo la hipótesis establecida para dicha investigación fue la siguiente: si hay variación estacional significativa en el patrón reproductivo de las especies estudiadas a lo largo del tiempo de muestreo, entonces ésta debe tener una estrecha relación con los cambios en las variables ambientales y debe ser consistente en ambas especies.
Características taxonómicas Las dos especies de ofiuros objeto de este estudio pertenecen a la Familia Ophiocomidae Ljungman, 1867. Los miembros de ésta familia se distinguen por tener dos grupos de papilas orales y dentales en cada mandíbula. Dentro de los caracteres más sobresalientes del género Ophiocoma Agassiz, 1835, se encuentran el disco cubierto por gránulos, escamas tentaculares pequeñas y ovaladas y presentar espinas lisas y sólidas en los brazos. Las únicas especies del género reportadas para el Pacífico Americano son O. aethiops (Fig. 1) y O. alexandri (Fig. 2) y se encuentran distribuidas entre el Golfo de California y Ecuador (Hendler 1979) Además de las evidentes diferencias en talla y color (Fig 1 y 2), O. aethiops y O. alexandri se diferencian por la forma de los escudos orales, el número de espinas en los brazos y el arreglo que presentan en las papilas dentales (Fig. 1, 2 y 3).

7
Figura 1. Ophiocoma aethiops Lütken, 1859. A) vista dorsal, B) vista ventral.
Figura 2. Ophiocoma alexandri Lyman, 1860. A) vista dorsal, B) vista ventral.
A B
A B

8
Figura 3. Caracteres relevantes para la diferenciación de Ophiocoma aethiops y O. alexandri (escudo oral, papilas dentales y espinas de los brazos).
Objetivo general Determinar si existen similitudes entre los patrones reproductivos en las poblaciones de Ophiocoma alexandri y O. aethiops presentes en la Bahía Estacahuite (Oax. México) y establecer su relación con las variables ambientales (temperatura, clorofila, pluviosidad, luminosidad, salinidad y horas de luz).
Objetivos específicos
1. Determinar la proporción de sexos en las poblaciones muestreadas de O. aethiops y O.
alexandri. 2. Calcular el valor promedio mensual del índice gonadal para las poblaciones de O.
aethiops y O. alexandri, a lo largo de un año de muestreo. 3. Caracterizar la fecundidad de las poblaciones de O. aethiops y O. alexandri presentes
en la Bahía Estacahuite, a lo largo de un año de muestreo. 4. Describir el ciclo de maduración gonádica para O. aethiops y O. alexandri.

9
5. Establecer si existe estacionalidad en el ciclo reproductivo para cada una de las especies
estudiadas. 6. Relacionar los patrones reproductivos de las poblaciones de O. aethiops y O. alexandri
con las variables ambientales (temperatura, clorofila, pluviosidad, luminosidad, salinidad y horas de luz).
7. Determinar si existen diferencias entre los patrones reproductivos de O. aethiops y O.
alexandri; en el caso de encontrarlas, establecer cuáles son dichas diferencias.
Área de estudio La Bahía de Estacahuite (15°40´5.57”N – 96°28´53.61”O) se encuentra localizada en las costas de Oaxaca, México, a 2 Km. aproximadamente de la Bahía de Puerto Ángel, en el municipio de San Pedro Pochutla (Fig. 4). El área presenta un fondo marino de composición rocosa y de arena con una pequeña cobertura de parches de coral (observaciones in situ). La bahía se encuentra en una zona que presenta un clima cálido – subhúmedo, con una estación seca en invierno y presenta otra (canícula) en el periodo de lluvias en verano (García 1981). Se caracteriza por una temperatura promedio anual de 27.5°C (máx. 32°C – mín. 22.7°C), humedad relativa de 74% promedio anual, con una evaporación de 1843.1mm al año, precipitación anual de 1025.4mm y vientos con una velocidad máxima de 6.5 m/s (CONAGUA 2000). El régimen de mareas para la región es semidiurno mixto con una amplitud de 1.050m (BN100210 ligado al BNT 498-8 del INEGI).

10
Figura 4. Mapa de la Bahía Estacahuite (Oaxaca, México). En la parte superior se encuentra el mapa de México y el punto indica la ubicación de la Bahía de Estacahuite.
Método
Muestreo y procesamiento de las muestras
Se efectuaron muestreos mensuales desde mayo de 2009 a abril de 2010, en los cuales se recolectaron 20 individuos de cada especie por muestreo. Las muestras se recolectaron por medio de buceo autónomo (3-15m), de forma manual, hasta completar la colección. Las variables temperatura e intensidad lumínica fueron medidas con un data logger (HOBO Pendant MAN-UA-002), que se instaló en el área de muestreo durante el año de estudio, y calibrado para hacer registros de la temperatura (+/- 0.47°C) y la intensidad lumínica (+/- 1 lux) cada hora. Para las variables pluviosidad, horas de luz y clorofila se utilizaron bases de datos en línea (http://www.wunderground.com/ para las dos primeras y http://oceancolor.gsfc.nasa.gov/ para la última), donde se encontraron registrados los promedios mensuales o diarios para cada una de estas variables durante el año de muestreo. En el caso de la salinidad se usaron los datos instantáneos del día de muestreo tomados con un medidor multiparámetro (HANNA HI 9828).

11
Los organismos fueron trasladados al laboratorio, donde se sacrificaron bajando la temperatura del agua a 4°C, posteriormente se fijaron con formol al 10% (12h) y luego fueron preservados en alcohol al 70% (Grange et al. 2004), para su procesamiento posterior. Se midieron los diámetros (Vernier, Scala 22A +/- 0.05mm) de los discos de cada organismo y el peso de los mismos (sin brazos) (Fenaux 1970, Hendler 1979). Se hizo la disección por la cara aboral del disco, para ubicar las gónadas y hacer su extracción. Se sacó la mitad de las gónadas totales de cada individuo y se registró su peso. Todas las mediciones de peso se realizaron en una balanza OHAUS (+/- 0.001 g). El volumen de las góonadas se determinó por medio del método de diferencia de volúmenes. El volumen y el peso obtenidos se multiplicaron por dos para obtener los valores totales de cada organismo. La otra mitad de la gónada fue conservada como respaldo.
Proceso histológico
La deshidratación de las gónadas se hizo sumergiéndolas en etanol al 70% durante 12h y luego se efectuaron dos cambios de etanol al 96% y dos de etanol al 100%, haciendo el recambio cada dos horas. Posteriormente se aclararon con una solución HemoD por 2h. Al terminar éste tiempo se hizo el embebido en parafina líquida a 70°C por 4h (Histoquinete Leica TP1020), luego se incluyeron en moldes de plástico (Incluidor de Leica EG1160) y se dejaron en proceso de secado por un tiempo mínimo de 12h. Después de tener las muestras secas se cortaron en un micrótomo (Leica RM2145) a un espesor de 7µm (Fenaux 1970, Gage et al. 2004, Grange et al. 2004), el corte fue colocado en un portaobjetos y se dejó en la plancha de secado durante 8h. Terminado el secado de las muestras se continuó con la tinción de las mismas, ésta se llevó a cabo con el método Hematoxilina- Eosina (Fenaux 1970, Ramirez-Llodra 2001, Gage et al. 2004, Grange et al. 2004, Tominaga et al. 2004). Las placas fueron fotografiadas (microscopio Olympus CX21 dotado con una cámara Logitech HD Pro WebCam C910) para su posterior análisis.
Estimación de parámetros reproductivos
1. Índice Gonadal (IG): éste se estimó a partir del peso del organismo (disco) y el de
las gónadas de acuerdo con lo descrito por Ramirez-Llodra (2001), usando la fórmula:
100*PD
PGIG =
Donde: PG (peso de la gónada) PD (peso del disco)

12
Hace referencia al porcentaje del peso del disco ocupado por la gónada. Para hacer la comparación de las diferencias del índice a lo largo el año y entre sexos se aplicó un ANOVA de 2 vías (factores: mes y sexo). El mismo análisis se aplicó para efectuar la comparación de las variaciones del IG entre de ambas especies a lo largo del año.
2. Estimación de Fecundidad: para esto se estimó el volumen medio de los ovocitos y
su relación con el volumen de la gónada de cada hembra (fecundidad real), calculando el volumen de cada ovocito (OV). Para el cálculo del volumen primero se midió el diámetro Feret (µm) de cien ovocitos de cada individuo (Programa Image-J); evaluando sólo los ovocitos que fueron seccionados a través del núcleo (Gage et al. 2004, Grange et al. 2004, Benítez-Villalobos 2005). A partir de los patrones de tinción y características particulares se diferenciaron los ovocitos vitelogénicos de los previtelogénicos. Los cálculos se aplicaron de la siguiente manera:
3
**4 3ROV
π= Volumen medio del ovocito
( )PVV
VF
PVOVO
g
VO *+= Fecundidad real donde
vo
pvo
N
NP =
Donde: Vg = volumen de la gónada
Vpvo = volumen medio de ovocitos previtelogénicos Vvo = volumen medio de ovocitos vitelogénicos P = promedio de ovocitos previtelogénicos y vitelogénicos Nvo = número de ovocitos vitelogénicos Npvo = número de ovocitos previtelogénicos
Tomando la estimación de la fecundidad real de cada individuo se hizo la estimación del promedio mensual de ésta para cada especie.
Con la información de las tallas de ovocitos se elaboraron histogramas de frecuencias para observar las distribuciones de tallas para cada mes (Gage et al. 2004).
Por medio de observación al microscopio de los cortes histológicos, se propuso una clasificación y descripción de los estadios de desarrollo gonádico de acuerdo a las clasificaciones que se han propuesto en la literatura (Selvakumaraswamy y Byrne 1995, Stewart y Mladenov 1995, Falkner y Byrne 2003, Tominaga et al. 2004, Borges et al. 2009).
En el caso de las hembras los parámetros obtenidos (índice gonadal, fecundidad y frecuencia de ovocitos) fueron relacionados con las características histológicas observadas

13
y con gráficas de frecuencia acumulada de individuos en cada etapa reproductiva. Esto para corroborar la adecuada clasificación de los diferentes estadios de desarrollo gonádico propuestos.
En el caso de los machos se hizo la descripción de los estadios de desarrollo basada en las características de los testículos, el estado de las células reproductivas (espermatocitos, espermatogonias) y su relación con el índice gonadal. Además se midieron 50 cabezas de espermatozoides con el fin de determinar su tamaño medio.
La proporción de machos y hembras se analizó mediante una prueba de Chi cuadrado con modificación de continuidad de Yates. Para comparar las variaciones mensuales de los parámetros ambientales (temperatura, luminosidad, pluviosidad, horas luz, clorofila y salinidad) con las del índice gonadal en cada especie se hizo una Correlación de Spearman y un Análisis de Regresión Lineal Múltiple de acuerdo con lo descrito por Ramirez-Llodra (2001), Gage et al. (2004), Grange et al. (2004) y Benítez-Villalobos (2005).

14
Resultados Se coleccionaron 427 individuos adultos (tallas mayores a 1cm de diámetro del disco) a lo largo del año de muestreo (202 de O. aethiops y 225 de O. alexandri). A 37 de ellos no fue posible determinarles el sexo dada la etapa de desarrollo gonádico en que se encontraban (Tabla 1). La proporción total de sexos encontrada para ambas especies no difirió significativamente de la proporción esperada 1:1 (g.l=1, el valor de χ2 corregido de Yates fue 0.99 con p=0.327 y 1.44 con p=0.2008 para O. aethiops y O. alexandri respectivamente).
Tabla 1. Porcentaje de machos y hembras coleccionados durante el periodo de muestreo. O. aethiops O. alexandri
%Hembras %Machos %Hembras %Machos
mayo 71 29 33 67 junio 68 32 42 58 julio 55 45 27 73 agosto 67 33 50 50 septiembre 77 23 43 57 octubre 72 28 59 41 noviembre 47 53 44 56 diciembre 100 25 75 enero 65 35 57 43 febrero 14 86 33 67 marzo 57 43 35 65 abril 68 32 30 70 % anual 58 42 40 60
El índice gonadal (IG) presentó una media de 13.66 con una desviación estándar de 6.57 para O. aethiops y de 19.88 con 12.72 de desviación estándar en el caso de O. alexandri. Las variaciones mensuales de este índice a lo largo del año de muestreo (Fig. 5A) fueron significativas para ambas especies (F=9.999 y F=16.151 con p<0.001 para O. aethiops y O. alexandri). También se presentaron diferencias significativas en la comparación de las variaciones del IG entre ambas especies durante el año de estudio (F=4.255 y p<0.001). Al hacer la comparación múltiple de Tukey, dichas diferencias se encontraron dadas por los valores presentes en los meses de julio a noviembre y abril (p<0.05 y q= 3.656 – 5.012 – 8.816 – 5.530 – 2.952 de julio a noviembre respectivamente y q=4.542 de abril) ya que los otros meses no presentaron diferencias significativas. La comparación entre los IG por sexo no mostró diferencias significativas en O. alexandri (F=0.395 y p=0.957) (Fig. 5C), mientras que en O. aethiops sí hubo diferencias significativas en los valores del IG entre sexos (F=3.032 y p=0.002), estando dichas

15
diferencias determinadas por los valores presentados en los meses de junio, septiembre, octubre y abril (p<0.05 y q= 3.014 – 4.082 – 4.098 y 2.978 respectivamente) (Fig. 5B). Sin embargo, las diferencias en el valor del índice fueron alternantes en estos cuatro meses, de modo tal que los valores más altos no los conservó siempre el mismo sexo; en el mes de junio fue mayor el IG en hembras, septiembre en machos, octubre en hembras y finalmente en abril fue de nuevo mayor el valor de las machos. La fecundidad real presentó una media de 3.038*106 ovocitos vitelogénicos por hembra con una desviación estándar (DS) de 1.722*106 para O. aethiops. En el caso de O. alexandri se observó una media de 1.612*106 con DS 0.991*106. Al hacer la comparación de los valores de fecundidad (Fig. 6), ambas especies presentaron diferencias significativas a lo largo del año (F=4.837 y p<0.001 para O. aethiops y F=2.067 y p=0.038 en el caso de O. alexandri). De la misma manera fueron significativas las diferencias al comparar entre los valores de fecundidad real entre las dos especies (F= 1.926 y p=0.047), dichas diferencias se encontraron dadas por los meses de mayo y junio (p<0.005 con valores de q=3.023 en mayo y q=6.398 en junio); en estos la diferencia de la fecundidades promedio fue de tres a cuatro millones de ovocitos vitelogénicos por hembra. El valor medio mensual de las tallas de ovocitos osciló entre 32–58µm para O. alexandri y 47–63µm (Fig. 7 y 8) para O. aethiops, lo que evidencia la presencia de ovocitos vitelogénicos a lo largo de todo el año (ver descripción histológica). El diámetro Feret medio de los ovocitos presentó una variación significativa (ANOVA F= 12.974 y p<0.01 para O. aethiops y F= 7.377 y p<0.01). En términos generales, la cantidad de ovocitos de cada intervalo de talla fue cambiando a lo largo del año (Fig. 7 y 8); el intervalo comprendido por los ovocitos de 10-40µm, mismo que contenía en su mayoría ovocitos previtelogénicos, tuvo un stock permanente a lo largo del año. Del mismo modo, los ovocitos (en su mayoría vitelogénicos) que se encontraron en el intervalo comprendido entre 40-70µm para O. alexandri y 40-80µm para O. aethiops se pudieron encontrar durante casi todo el año. En el mes de diciembre, O. aethiops no tuvo ningún individuo que se pudiera clasificar como hembra; para el caso de O. alexandri no hubo ningún ovocito de las tallas anteriormente mencionadas. Sin embargo, el total de los ovocitos encontrados en esta última especie, para éste mes tenía un tamaño comprendido entre 20 y 40µm. Además, es de notar que el mes de diciembre fue el que tuvo más individuos que no pudieron ser sexados y un muy bajo porcentaje de hembras (O. alexandri). Al interior de la gónada, la cantidad de ovocitos fue mínima, evidenciando que sólo se trataba de un remanente de la temporada reproductiva. Los organismos con intervalos de tallas mayores a 70 y 80µm para O. alexandri y O. aethiops respectivamente, se encontraron ocupados por una cantidad mínima de ovocitos, la que en ningún mes superó el 5%; es decir, que en estas tallas solo estuvieron representadas por un remanente de ovocitos vitelogénicos. Al hacer un conglomerado de las frecuencias para los diferentes tipos de ovocitos (previtelogénicos, vitelogénicos y en reabsorción) (Tabla 2), se puede ver claramente cual

16
de estos tipos peresentó un mayor porcentaje y en que mes; de ello se destaca que O. aethiops tuvo en todos los meses una mayor cantidad de ovocitos vitelogénicos (excepto diciembre donde no hubo hembras identificadas en la muestra). El acumulado de frecuencias para O. alexandri, de forma similar, tuvo mayor presencia de ovocitos vitelogénicos casi todo el año con excepción de tres meses (julio, diciembre y enero) (Tabla 2). Tabla 2. Porcentaje acumulado para frecuencias de ovocitos previtelogénicos, vitelogénicos y en reabsorción
de Ophiocoma aethiops y O. alexandri. P (ovocitos previtelogénicos), V (Ovocitos vitelogénicos) y R (ovocitos no evacuados o en reabsorción)
O. alexandri O. aethiops
P V R P V R mayo 29 71 0 7 90 2 junio 49 51 0 14 85 1 julio 57 43 0 20 80 0 agosto 24 74 3 20 79 1 septiembre 6 84 10 6 90 4 octubre 7 88 4 5 91 4 noviembre 17 83 0 1 96 2 diciembre 100 0 0 - - - enero 76 24 0 38 54 8 febrero 27 73 1 32 68 0 marzo 23 71 5 27 73 0 abril 21 68 11 9 85 6

17
Tiempo (mes)
May
o
Juni
o
Julio
Ago
sto
Sep
tiem
bre
Oct
ubre
Nov
iem
bre
Dic
iem
bre
Ene
ro
Feb
rero
Mar
zo
Abr
il
IG P
rom
edio
(%
)
0
10
20
30
40
50
60
O. aethiopsO. alexandri
Tiempo (mes)
May
June
July
Aug
Sep
t
Oct
Nov
Dec Jan
Feb
Mar
Apr
IG P
rom
edio
(%
)
0
10
20
30
40
50
O. aethipos HembrasO. alethiops Machos
Tiempo (mes)
May
o
Juni
o
Julio
Ago
sto
Sep
tiem
bre
Oct
ubre
Nov
iem
bre
Dic
iem
bre
Ene
ro
Feb
rero
Mar
zo
Abr
il
IG P
rom
edio
(%
)
0
10
20
30
40
50
60
70
O. alexandri HembrasO. alexandri Machos
Figura 5. Promedios mensuales (+/- DS) del Índice Gonadal. A) Ophiocoma aethiops y O. alexandri, B) O. aethiops hembras y machos por separado, C) O. alexandri hembras y machos por separado.
A
B
C

18
Tiempo (mes)
May
o
Juni
o
Julio
Ago
sto
Sep
tiem
bre
Oct
ubre
Nov
iem
bre
Dic
iem
bre
Ene
ro
Feb
rero
Mar
zo
Abr
il
Fvo
Pro
med
io (
cant
idad
de
ovoc
itos
* 10
6 )
0
2
4
6
8
10
12
O. aethiops O. alexandri
Figura 6. Promedios mensuales (+/- DS) de fecundidad para Ophiocoma aethiops y O. alexandri. Para O. alexandri se puede observar que en junio fue un poco más alto el valor de ovocitos vitelogénicos y éstos disminuyeron en julio, aumentando el valor de los previtelogénicos; lo que evidencia un recambio de ovocitos para los meses siguientes, en los que vuelve a caer la cantidad de ovocitos previtelogénicos; viéndose así reflejado de forma consecuente un nuevo aumento de ovocitos vitelogénicos. Estos últimos al comenzar a disminuir, promueven el aumento del stock en los de las tallas anteriores. El patrón de recambio de ovocitos (P-V) a lo largo del año evidencia como hay continuamente producción de ovocitos previtelogénicos tras los eventos de desove (disminución de ovocitos vitelogénicos). Esta relación inversa en las cantidades es constante, de modo tal que al efectuar una correlación de Spearman entre los ovocitos previtelogénicos y vitelogénicos da un valor de -0.92 para un valor de p=0.000, lo cual soporta el carácter cíclico en dicha dinámica. Contrario a lo sucedido en el caso de O. alexandri, O. aethiops no presentó valores similares de frecuencia de ovocitos (P-V), el único que tuvo una diferencia pequeña en estos valores fue el encontrado en el mes de enero, donde ésta fue de 16%, pero continuó siendo mayor la cantidad de ovocitos vitelogénicos. Aquí se puede ver como al disminuir los valores de ovocitos vitelogénicos, aumentaron los previtelogénicos, pero no en las proporciones observadas en O. alexandri. Esto refleja una dinámica en la cual se mantiene una mayor cantidad de ovocitos en stock, solo que en este caso sería mayor de vitelogénicos que de previtelogénicos. Además, aquí la correlación de Spearman no fue significativa (p= 0.0705). Cabe recordar que el mes de diciembre fue sido excluido del análisis, debido a que aquí no fue posible identificar ninguna hembra y que este puede ser el mes donde se desarrollan la ovogonias necesarias para comenzar el proceso de ovogénesis y mantener ese nivel tan alto de ovocitos vitelogénicos. Las variables ambientales (Fig. 9) y el IG fueron comparadas a través de una correlación de Spearman, la cual fue significativa y positiva para ambas especies en los parámetros de

19
pluviosidad, horas de luz y temperatura (Tabla 3); las otras variables no presentaron correlación significativa. El ARLM hecho para O. aethiops fue resuelto en tres pasos. Se encontró que la variación total explicada por las variables seleccionadas (pluviosidad y horas luz) con respecto a la variable dependiente (IG) fue de 0.537, con un valor de correlación múltiple de 0.732 y p<0.05. Para O. alexandri éste análisis se resolvió en un paso, dado que toda la variación explicada del IG se dio en términos de la temperatura. Según el modelo dicha variación fue de 0.547; con un valor de correlación múltiple de 0.740 y p<0.05. Es decir, el aumento del índice gonadal (crecimiento de la gónada) se encontró directamente relacionado con el incremento en los valores de horas luz y pluviosidad en el caso de O. aethiops y con la temperatura en el caso de O. alexandri. El análisis de residuales mostró poca desviación sobre la distribución normal, lo que denota un modelo de predicción estadísticamente válido y confiable, aunque es de resaltar que en ambas especies la varianza explicada no fue mayor del 60%.
Tabla 3. Valores de los coeficientes de correlación (en negrilla las correlaciones significativas) y p encontrados con el análisis de correlación de Spearman aplicado entre las variables ambientales y los
promedios mensuales del Índice Gonadal de Ophiocoma aethiops y O. alexandri.
O. aethiops O. alexandri
Variable Coeficiente
de correlación
p
Coeficiente de
correlación p
Pluviosidad 0.673 0.0154 0.681 0.0139
Horas luz 0.685 0.0126 0.629 0.0263
Temperatura 0.643 0.0222 0.755 0.0336
Clorofila -0.364 0.257 -0.418 0.188
Salinidad 0.0182 0.946 -0.00606 0.973
Luminosidad 0.0350 0.904 0.189 0.542
Tabla 4. Valores de la regresión lineal paso a paso para las variables que presentaron correlación significativa con el Índice Gonadal de Ophiocoma aethiops y O. alexandri. Las no incluidas son aquellas que el modelo no
tomó en cuenta por no ser significativas en la regresión.
Variables
Intercepto Temperatura Pluviosidad Horas luz O. aethiops -44.31 No incluida β=0.475716 β=0.326140 O. alexandri -119.93 β=0.740155 No incluida No incluida

20
Figura 7. Frecuencia de tallas de ovocitos de Ophiocoma aethiops a lo largo del año de muestreo.
Mayon=453
0
10
20
30
40
50
10-19.9 20-29.9 30-39.9 40-49.9 50-59.9 60-60,9 70-79.9 80-89.9 90-99.9
Junion=1202
0
10
20
30
40
50
10-19.9 20-29.9 30-39.9 40-49.9 50-59.9 60-60,9 70-79.9 80-89.9 90-99.9
Julion=1388
0
10
20
30
40
50
10-19.9 20-29.9 30-39.9 40-49.9 50-59.9 60-60,9 70-79.9 80-89.9 90-99.9
Agoston=592
0
10
20
30
40
50
10-19.9 20-29.9 30-39.9 40-49.9 50-59.9 60-60,9 70-79.9 80-89.9 90-99.9
Septiembren=1013
0
10
20
30
40
50
10-19.9 20-29.9 30-39.9 40-49.9 50-59.9 60-60,9 70-79.9 80-89.9 90-99.9
Octubren=955
0
10
20
30
40
50
10-19.9 20-29.9 30-39.9 40-49.9 50-59.9 60-60,9 70-79.9 80-89.9 90-99.9
Noviembren=553
0
10
20
30
40
50
10-19.9 20-29.9 30-39.9 40-49.9 50-59.9 60-60,9 70-79.9 80-89.9 90-99.9
Eneron=773
0
10
20
30
40
50
10-19.9 20-29.9 30-39.9 40-49.9 50-59.9 60-60,9 70-79.9 80-89.9 90-99.9
Febreron=88
0
10
20
30
40
50
10-19.9 20-29.9 30-39.9 40-49.9 50-59.9 60-60,9 70-79.9 80-89.9 90-99.9
Marzon=559
0
10
20
30
40
50
10-19.9 20-29.9 30-39.9 40-49.9 50-59.9 60-60,9 70-79.9 80-89.9 90-99.9
Abriln=1283
0
10
20
30
40
50
10-19.9 20-29.9 30-39.9 40-49.9 50-59.9 60-60,9 70-79.9 80-89.9 90-99.9
Diámetro Feret de ovocito (µm)

21
Figura 8. Frecuencia de tallas de ovocitos de Ophiocoma alexandri a lo largo del año de muestreo.
Mayon=300
0
10
20
30
40
50
10-19.9 20-29.9 30-39.9 40-49.9 50-59.9 60-60.9 70-79.9 80-89.9
Junion=804
0
10
20
30
40
50
10-19.9 20-29.9 30-39.9 40-49.9 50-59.9 60-60.9 70-79.9 80-89.9
Julion=442
0
10
20
30
40
50
10-19.9 20-29.9 30-39.9 40-49.9 50-59.9 60-60.9 70-79.9 80-89.9
Agoston=1034
0
10
20
30
40
50
10-19.9 20-29.9 30-39.9 40-49.9 50-59.9 60-60.9 70-79.9 80-89.9
Septiembren=521
0
10
20
30
40
50
10-19.9 20-29.9 30-39.9 40-49.9 50-59.9 60-60.9 70-79.9 80-89.9
Octubren=1323
0
10
20
30
40
50
10-19.9 20-29.9 30-39.9 40-49.9 50-59.9 60-60.9 70-79.9 80-89.9
Noviembren=544
0
10
20
30
40
50
10-19.9 20-29.9 30-39.9 40-49.9 50-59.9 60-60.9 70-79.9 80-89.9
Diciembren=10
0
10
20
30
40
50
60
10-19.9 20-29.9 30-39.9 40-49.9 50-59.9 60-60.9 70-79.9 80-89.9
Eneron=135
0
10
20
30
40
50
10-19.9 20-29.9 30-39.9 40-49.9 50-59.9 60-60.9 70-79.9 80-89.9
Febreron=403
0
10
20
30
40
50
10-19.9 20-29.9 30-39.9 40-49.9 50-59.9 60-60.9 70-79.9 80-89.9
Marzon=429
0
10
20
30
40
50
10-19.9 20-29.9 30-39.9 40-49.9 50-59.9 60-60.9 70-79.9 80-89.9
Abriln=598
0
10
20
30
40
50
10-19.9 20-29.9 30-39.9 40-49.9 50-59.9 60-60.9 70-79.9 80-89.9
Diámetro Feret de ovocito (µm)

22
0,00
0,10
0,20
0,30
0,40
0,50
0,60
0,70
May
oJu
nio Julio
Agosto
Septie
mbre
Octubr
e
Noviem
bre
Diciem
bre
Enero
Febre
ro
Mar
zoAbr
il
Tiempo (mes)
Plu
vios
idad
(cm
)
10:04:48
10:33:36
11:02:24
11:31:12
12:00:00
12:28:48
12:57:36
13:26:24
May
oJu
nio Julio
Agosto
Septie
mbr
e
Octubr
e
Noviem
bre
Diciem
bre
Enero
Febre
ro
Mar
zoAbr
il
Tiempo (mes)
Tie
mpo
(ho
ras
de lu
z)
24
26
28
30
32
May
oJu
nio Julio
Agosto
Septie
mbr
e
Octubr
e
Noviem
bre
Diciem
bre
Enero
Febre
ro
Mar
zoAbr
il
Tiempo (mes)
Tem
pera
tura
(°C
)
Figura 9. Promedios mensuales de las variables ambientales que presentaron correlación significativa con el Índice Gonadal para ambas especies. A) pluviosidad, B) horas de luz, C) temperatura.
C
B
A

23
El aspecto macroscópico de la gónada, fue de un saco (epitelio delgado y transparente) que contiene numerosos sacos ovalados mucho más pequeños (folículos). Éstos pueden variar en su coloración, de blanco a beige en el caso de los machos y en tonos de rosa a violáceo en el caso de las hembras. Los folículos en algunas ocasiones pueden presentase con una o dos ramificaciones. Cada gónada se encontró de ambos lados de la hendidura bursatil, esto da un total de diez unidades discretas que pueden ser identificadas en el disco de un individuo. A nivel microscópico, la pared del folículo presentó dos capas de tejido, un saco externo y un saco interno, éstos se encuentran separados por el celoma genital (Fig. 10D, 11B, 13A). El saco externo corresponde al peritoneo visceral; en la pared del saco interno, también de naturaleza peritoneal, se observó la presencia de células musculares que lo delimitan (Fig. 13B), las cuales adquieren tonos de rosa debido a su naturaleza acidófila. Como resultado del análisis de los cortes histológicos que se realizaron, se observó un folículo ovárico en el cual el tejido germinal no fue fácil de apreciar (similar a lo que ocurre en varias especies). En etapas iniciales, las ovogonias se disponen repartidas en el lumen del folículo, mientras que en las etapas avanzadas se disponen de forma intercalada entre los ovocitos o repartidas en el lumen. Para las hembras de ambas especies estudiadas se encontró que el ciclo gonádico se puede dividir en cuatro etapas. Esto fue comparado con lo reportado por la literatura, donde generalmente se presentan cinco, de las cuales tres son similares y una presenta la unión de las otras dos. Así, para O. aethiops y O. alexandri se definieron los estadios de crecimiento, madurez, desove y postdesove; las cuales se describen a continuación:
1. Crecimiento gonadal: comienza con el desarrollo de ovogonias, las cuales se observan en tonalidades azulosas a violáceas (basófilas), acomodadas en la capa germinal del folículo ovárico; conforme avanza dicha etapa, algunas ovogonias de forma asincrónica comienzan el proceso de mitosis, lo cual da origen a los ovocitos previtelogénicos (primarios). Estos presentan una coloración violácea menos intensa que las ovogonias, además se puede ver en algunos de ellos la presencia del nucléolo (coloración mucho más intensa al interior del núcleo). Después del comienzo de la aparición de ovocitos previtelogénicos, de forma muy esporádica pueden encontrarse ovocitos que ya han iniciado el proceso de vitelogénesis, debido a que la ovogénesis no fue sincrónica en todas las células presentes en cada folículo (Fig. 10A, 11A y 11B). De esta forma, lo que se puede observar es una gónada que se encuentra ocupada en su mayor parte por ovogonias, las cuales pueden estar acompañadas de algunos ovocitos. En esta etapa el epitelio gonadal es grueso y en algunos casos se pueden observar espículas, la coloración es azul intenso, lo que indica una naturaleza basófila, a diferencia de las espículas que aunque no tiñen se pueden observar relativamente transparentes. En algunos casos se puede ver también células simi-lipofuscina que vienen desde la etapa final de la temporada anterior. Durante este periodo se puede observar cómo el espacio en el lumen se encuentra cada vez más reducido debido a la ocupación de éste por las ovogonias y los

24
ovocitos, dicha ocupación ocurre tanto de forma regular (de la periferia hacia el centro), como de forma irregular.
2. Maduración ovocitaria: al inicio de la etapa, el panorama es de un folículo con
pocas ovogonias remanentes, la mayoría de los ovocitos se encuentran en estadios previtelogénicos, mientras que los ovocitos vitelogénicos están presentes en una cantidad reducida. Ya casi no hay espacios libres dentro del lumen puesto que éste ha sido ocupado por los ovocitos. Conforme prosigue el desarrollo la actividad vitelogénica hace que los ovocitos previtelogénicos pasen a estados vitelogénicos cada vez más avanzados (mayor cantidad de vitelo en su interior), lo que se evidencia por un incremento en su talla. Además, la coloración de estos se torna de violácea a fucsia o rosa pálido, denotando un cambio en su naturaleza química (de basofilia a acidofilia) dado por la presencia de los gránulos de vitelo. Debido al aumento de tamaño de los ovocitos durante el proceso de vitelogénesis, el núcleo representa un menor tamaño con respecto al tamaño total de la célula (Fig. 10B, 10C y 11C). En el interior de folículo ovárico se puede observar el empaquetamiento propio de los ovocitos que se presenta con apariencia de malla, dentro de la cual se disponen los mismos. Además, la membrana del folículo y de la gónada se hacen cada vez más delgadas y desaparecen las espículas, quedando en muy pocos casos remanentes al final de este proceso. En general se observa una gónada con folículos llenos de ovocitos, en su mayoría vitelogénicos, lo cual no permite apreciar espacios vacíos en el lumen de los folículos.
3. Desove: el desove se evidencia a nivel gonadal por la presencia de espacios que
claramente se encontraban ocupados por un ovocito vitelogénico. A medida que avanza la etapa, no sólo se ven espacios de forma esporádica, sino que cada vez el arreglo de los ovocitos en el lumen del folículo es más laxo (Fig. 10D y 11D). No hay un patrón general de desocupación, dado que ocurre de la misma forma heterogénea que la maduración. La presencia de ovocitos previtelogénicos es cada vez más escasa debido a que estos terminan de madurar conforme avanza la etapa. Durante este proceso el ovario se comienza a colapsar lentamente; a medida que esto sucede comienza a aumentar la aparición de fagocitos, células que se aprecian de color azul intenso. En general en este estadio se puede observar una gónada que tiene la mayor parte de sus folículos con espacios entre los ovocitos, donde éstos últimos ya no se encuentran en un arreglo estrecho.
4. Postdesove y reactivación: comienza cuando ha terminado el último desove del
individuo, la gónada se encuentra vacía o con un remanente de ovocitos, los cuales pueden llegar a ser degradados o conservados para el siguiente periodo reproductivo. El lumen se encuentra ocupado parcialmente por células fagocíticas y en algunos casos hay células simi-lipofuscina, las cuales permanecen en ocasiones durante esta etapa y parte de la etapa de crecimiento del ciclo siguiente. Las membranas de la gónada y de los folículos son más evidentes debido a que estos se encuentran contraídos; es más evidente la presencia de numerosas espículas en la pared gonadal. Conforme va finalizando la actividad fagocítica, inicia de nuevo la actividad del epitelio germinal, la cual comienza con la producción de ovogonias dando así paso a

25
la primera etapa del ciclo siguiente (Fig. 10E y 11E). Aquí se observa en general folículos gonadales pequeños, con células fagocíticas en su interior y partículas amarillas que corresponden a las células simi-lipofuscina, una membrana gruesa y una mayor cantidad de tejido conjuntivo en la periferia y circundante.
Figura 10. Etapas reproductivas de las hembras de Ophiocoma aethiops. A) crecimiento, B) y C) maduración, D) desove, E) postdesove y reactivación. Clf (células simi-lipofuscina), CG (celoma genital), Og (ovogonias), OPv (ovocitos previtelogénicos), OV (ovocitos vitelogénicos), SE (saco externo), SI (saco interno) y PF (membrana del folículo).
A B
C
E
D

26
Figura 11. Etapas reproductivas de las hembras de Ophiocoma alexandri. A) y B) crecimiento, C) maduración, D) desove, E) postdesove y reactivación. CG (celoma genital), Og (ovogonias), OPv (ovocitos previtelogénicos), OV (ovocitos vitelogénicos), SE (saco externo) y SI (saco interno). En los testículos la espermatogénesis se desarrolla formando columnas hacia el lumen de los mismos. En estas columnas se pueden reconocer espermatogonias y columnas espermatogénicas (en ellas generalmente se reconocen espermatocitos y espermátidas, sin embargo en estas dos especies la diferenciación de dichas células se dificultó por su similitud en tamaño) (Fig. 12B, 13B), finalmente los espermatozoides ya maduros son liberados y acumulados en el lumen del testículo espermario (Fig. 12A). Las cabezas de los espermatozoides son ovaladas y tienen una diámetro promedio de 2.24µm +/-0.24 para
A B
C D
E

27
Ophiocoma aethiops y de 2.50µm +/_0.22 para O. alexandri, las colas fueron difícilmente observadas. Para los machos de ambas especies se hizo una caracterización del ciclo. Sin embargo en este caso sólo se encontraron tres etapas claramente identificables. Se observaron testículos que, con excepción de las pocas ocasiones en la que se encontraron vacíos; estaban en actividad de maduración o desove, observándose la actividad espermatogénica de manera constante. Considerando que la literatura reporta cinco fases para el ciclo espermatogénico, se tomaron las características más relevantes de estos y se compararon con lo observado. Con base en lo anterior se proponen tres etapas para el ciclo reproductivo de los machos de O. aethiops y O. alexandri, las cuales se describen a continuación:
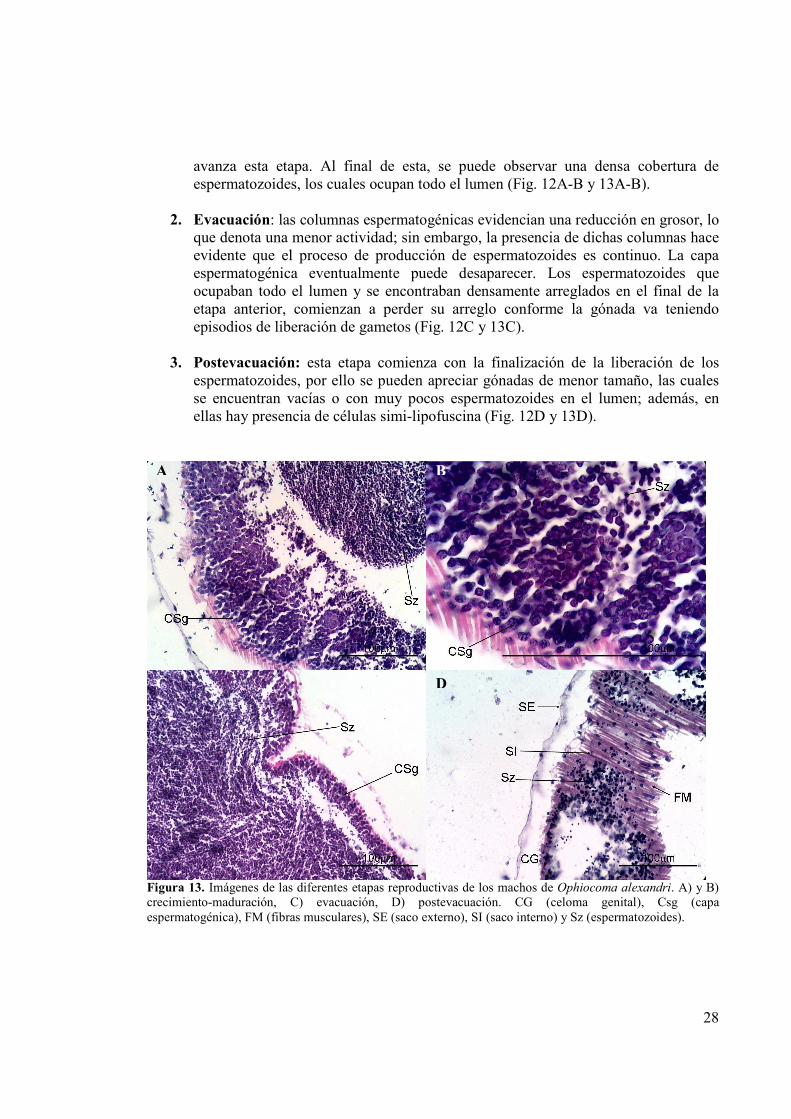
Figura 12. Etapas reproductivas de los machos de Ophiocoma aethiops. A) y B) crecimiento-maduración, C) evacuación, D) postevacuación. CG (celoma genital), Clf (células simi-lipofuscina), Csg (capa espermatogénica), FM (fibras musculares), PF (pared del folículo), SE (saco externo), SI (saco interno) y Sz (espermatozoides).
1. Crecimiento y maduración gonadal: en esta etapa se puede visualizar una capa espermatogénica gruesa, la cual se encuentra cercana al epitelio germinal; de ésta se desprenden los espermatozoides y se van separando para migrar al centro del testículo. Dado lo anterior, en el lumen se puede observar una agrupación de espermatozoides en el centro, la cual se va haciendo cada vez más densa conforme
A B
C D
28
avanza esta etapa. Al final de esta, se puede observar una densa cobertura de espermatozoides, los cuales ocupan todo el lumen (Fig. 12A-B y 13A-B).
2. Evacuación: las columnas espermatogénicas evidencian una reducción en grosor, lo
que denota una menor actividad; sin embargo, la presencia de dichas columnas hace evidente que el proceso de producción de espermatozoides es continuo. La capa espermatogénica eventualmente puede desaparecer. Los espermatozoides que ocupaban todo el lumen y se encontraban densamente arreglados en el final de la etapa anterior, comienzan a perder su arreglo conforme la gónada va teniendo episodios de liberación de gametos (Fig. 12C y 13C).
3. Postevacuación: esta etapa comienza con la finalización de la liberación de los
espermatozoides, por ello se pueden apreciar gónadas de menor tamaño, las cuales se encuentran vacías o con muy pocos espermatozoides en el lumen; además, en ellas hay presencia de células simi-lipofuscina (Fig. 12D y 13D).
Figura 13. Imágenes de las diferentes etapas reproductivas de los machos de Ophiocoma alexandri. A) y B) crecimiento-maduración, C) evacuación, D) postevacuación. CG (celoma genital), Csg (capa espermatogénica), FM (fibras musculares), SE (saco externo), SI (saco interno) y Sz (espermatozoides).
A B
C D

29
También se elaboraron gráficas del porcentaje de individuos en cada etapa reproductiva, tanto para hembras como para machos de cada una de las dos especies estudiadas (Fig. 14, 15, 16 y 17). En ellas se puede observar como se distribuyeron los estadios reproductivos a lo largo del año y en que porcentaje de individuos se presentaron. En el caso de las hembras de O. aethiops el tiempo de crecimiento gonadal para la población estuvo comprendido entre los meses de enero a marzo, la maduración gonadal de enero a septiembre, la temporada de desove de mayo a noviembre y el postdesove-reactivación de la gónada se presentó de noviembre a febrero y en agosto (Fig. 14). Ophiocoma alexandri varió un poco en los tiempos en que ocurren las diferentes etapas de desarrollo (Fig. 15), así, el crecimiento gonadal inició al igual que en O. aethiops pero terminó en mayo. La maduración gonadal ocurre desde febrero a julio teniendo un intervalo menor que el de la otra especie de Ophiocoma. La temporada de desove se distribuyó de abril a diciembre teniendo una duración igual que O. aethiops, sin embargo, las dos especies presentaron un mes de desfase. La ultima etapa de O. alexandri, el postdesove-reactivación inició en octubre y finalizó en diciembre, con un episodio más en febrero; esto ocurrió de igual manera que en O. aethiops, ya que esta también presentó un evento adicional. No obstante, esta última etapa no fue similar en su duración.
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
may
ojun
iojul
io
agos
to
sept
iembr
e
octub
re
novie
mbr
e
diciem
bre
ener
o
febr
ero
mar
zoab
ril
Tiempo (mes)
% d
e in
divi
duos
en
O. a
eth
iop
s e
n et
apa
repr
oduc
tiva
1 2 3 4 Figura 14. Porcentaje acumulado de hembras de Ophiocoma aethiops en las diferentes temporadas reproductivas a lo largo del año de muestreo. 1. Crecimiento gonadal, 2. Maduración gonadal, 3. Desove de la gónada y 4. Postdesove y reactivación de la gónada.

30
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
may
ojun
iojul
io
agos
to
sept
iembr
e
octu
bre
novie
mbr
e
diciem
bre
ener
o
febr
ero
mar
zo abril
Tiempo (mes)
% d
e in
divi
duos
de
O. a
lexa
ndri
en
etap
a re
prod
uctiv
a
1 2 3 4 Figura 15. Porcentaje acumulado de hembras de Ophiocoma alexandri en las diferentes temporadas reproductivas a lo largo del año de muestreo. 1. Crecimiento gonadal, 2. Maduración gonadal, 3. Desove de la gónada y 4. Postdesove y reactivación de la gónada.
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
mayo
junio
julio
agos
to
sept
iembr
e
octub
re
novie
mbre
diciem
bre
ener
o
febr
ero
marzo
abril
Tiempo (mes)
% d
e in
divi
duos
en
O. a
eth
iop
s e
n et
apa
repr
oduc
tiva
1 2 3 Figura 16. Porcentaje acumulado de machos de Ophiocoma aethiops en las diferentes temporadas reproductivas a lo largo del año de muestreo. 1. Crecimiento y maduración de la gónada, 2. Evacuación de la gónada y 3. Postevacuación de la gónada.

31
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
may
ojun
iojul
io
agos
to
sept
iembr
e
octu
bre
novie
mbre
diciem
bre
ener
o
febr
ero
mar
zoab
ril
Tiempo (mes)
% d
e in
divi
duos
en
O. a
lexa
nd
ri e
n et
apa
repr
oduc
tiva
1 2 3
Figura 17. Porcentaje acumulado de machos de Ophiocoma alexandri en las diferentes temporadas reproductivas a lo largo del año de muestreo. 1. Crecimiento y maduración de la gónada, 2. Evacuación de la gónada y 3. Postevacuación de la gónada. Para los machos, fue evidente como en ambas especies el tiempo de liberación de gametos permanece durante todo el año. La temporada de crecimiento-maduración se presentó en O. aethiops en diciembre y de febrero a abril; mientras que para O. alexandri ésta ocurrió de enero a julio y octubre. Finalmente los organismos en postevacuación se encontraron entre octubre y enero en el caso de O. aethiops y de octubre a febrero para O. alexandri. Lo anterior denota que en ambas especies es coincidente también el tiempo de postevacuación, el cual presentó solo diferencia de un mes.

32
Discusión Ophiocoma aethiops y O. alexandri son especies dioicas, las proporciones sexuales no presentaron desvío significativo a la esperada 1:1, lo cual concuerda con lo reportado en la literatura para ofiuros dioicos (Hendler 1991). Los índices gonadales presentaron una fluctuación anual que demuestra el cambio de tamaño de la gónada en proporción a su cuerpo; es decir, se observó claramente como se da un crecimiento de ella en los primeros meses del año. Esta observación es apoyada en el caso de las hembras por el aumento en las fecundidades, que también evidencia dicho crecimiento. Lo anterior concuerda con las observaciones hechas en los cortes histológicos, en las cuales coincide éste tiempo con los primeros estadios de desarrollo. Sin embargo, como se mencionó en los resultados, hubo concomitancia de estadios, pero esto no implicó que no haya un crecimiento dentro de un panorama general. Basándose en la fecundidad, el índice gonadal y las observaciones histológicas, se puede concluir que ambas especies tienen un ciclo anual, las hembras presentan una temporada de desove marcada, la cual oscila entre abril y diciembre, a diferencia de lo reportado para otros equinodermos estudiados en la zona, los cuales tienen una temporada de desove más corta (de junio a septiembre para Holothuria fuscosinerea y de julio a septiembre para Pharya Piramidatus) (Martinez-Garcia 2010, Gutierrez-Mendez 2011). Los machos presentaron disponibilidad de espermatozoides durante todo el año, ya que durante la etapa de evacuación la espermatogénesis fue continua, con excepción del tiempo en que la gónada entra en postevacuación; donde cesa temporalmente dicho proceso. La temporada de liberación de gametos de machos y hembras no coincide por completo, ya que el macho presenta una disponibilidad de espermatozoides durante todo el año. Debido a que el desarrollo de los ovarios fue variable, algunas hembras presentan ovocitos con desarrollo sincrónico y otros asincrónicos. Así mismo, se encontró en las hembras una concomitancia esporádica de un estadio gonadal anterior o siguiente, en folículos aislados, dentro de la gónada de individuos claramente clasificados en uno de los estadios de desarrollo. Se puede sugerir que el inicio de cada una de las etapas del ciclo puede variar al menos en un mes de diferencia a lo encontrado, sobre todo el comienzo de la temporada de desove. La única especie en que hubo sincronía en uno de los estadios fue O. alexandri, la cual tanto para machos como para hembras se presentó entre agosto y septiembre, cuando todos los ejemplares coleccionados se encontraban en la misma etapa, correspondiente a la evacuación de la gónada. Los folículos gonadales de Ophiocoma aethiops y O. alexandri son similares a las de otros ofiuroideos que también presentan estructura formada por dos sacos, separados por el celoma genital. Sobre el saco interno descansa el epitelio germinal, más evidente en los machos. Las membranas de los sacos externo e interno presentan células epiteliales y células mioepiteliales, una característica del epitelio celómico de los equinodermos (Byrne, 1989). La relación precisa entre los ovocitos y la membrana gonadal es difícil de

33
comprender debido a que los diferentes tipos de éstos no están espacialmente segregados; por esto, ésta relación ha sido pobremente documentada como lo menciona Hendler, (1991). Al hacer la extracción de las gónadas en algunas ocasiones se encontraron folículos más pequeños en la base de los más grandes (cerca de la hendidura bursatil), estas estructuras pueden ser gónadas en formación. Selvakumaraswamy y Byrne (1995) sostienen que lo anterior se puede deber a que las especies tengan folículos con un tiempo de vida limitado y éstos sean reemplazados eventualmente por nuevos folículos. Estas observaciones las hicieron en Ophionereis schayeri, y a su vez basaron su hipótesis en la planteada por Smiley (1988) quién la propuso con base en las observaciones hechas en el holoturoideo Stichopus californicus (Selvakumaraswamy y Byrne 1995). Probablemente dicho reemplazo ocurra de forma parcial o total ya que en algunos de los organismos en los que no se pudo determinar el sexo, el impedimento se debió a que no hubo presencia de folículos; a pesar de que los organismos eran individuos de tallas en las que no pueden ser considerados como juveniles. Lo anterior podría estar indicando un recambio de todos los folículos en la gónada. Sin embargo, dentro de los individuos no sexados hubo una porción que sí tenía folículos en las gónadas presentes (de tamaños muy pequeños), estas gónadas se observaron en el periodo de IG más bajo y de menor fecundidad, lo que sugiere que la liberación de gametos fue total en esos individuos (tanto machos como hembras). Dichas gónadas no fueron incluidas en ninguno de los sexos, dado que en ellas no se encontró ninguna traza de gametos no evacuados, sólo en algunas ocasiones células simi-lipofuscina. Falkner y Byrne (2003) sostienen que la lipofuscina es un pigmento que comúnmente es encontrado en tejidos de invertebrados, se acumula en estos tejidos como producto de un metabolismo de rompimiento; los autores sostienen que en este caso los productos de lipofuscina pueden ser el resultado principalmente del rompimiento de gametos. Además, para la época de su estudio en 2003 estos pigmentos no habían sido reportados para ningún equinodermo; ya que ellos hacen énfasis en que era el primer reporte para los ofiuros. Siendo los autores quienes acuñaron el término células simi-lipofuscina para denominar dichas partículas que presentan forma de célula, de tono amarillo a café encontradas en las placas de los cortes histológicos de las gónadas de los ofiuros. La actividad de liberación de gametos en algunos ofiuros es corta y fuertemente sincrónica, mientras que en otros es caracterizada por un periodo intenso de desove seguido de varios meses de baja actividad de expulsión de gametos (Falkner y Byrne 2003). Para una de las dos especies estudiadas, O. aethiops, se encontró coincidencia con el segundo patrón. Dicha asunción, esta basada en que el periodo de liberación de gametos encontrado es aproximadamente de siete meses y que la variación de la fecundidad promedio indica un pulso de desove más fuerte; en el cual, la cantidad de ovocitos baja abruptamente; en los pulsos siguientes la disminución es menos fuerte. Mientras tanto, O. alexandri no presentó evidencia de un pulso de desove tan fuerte, las cantidades de ovocitos disminuyeron de forma menos abrupta, para lo cual se podría sostener que esta especie si tiene un largo periodo de desove, pero que el mismo esta caracterizado por una actividad continua con pequeños pulsos de expulsión de gametos. Para ambas especies, el pulso de liberación de

34
gametos entre los meses de octubre y diciembre es el último del ciclo reproductivo, seguido por un periodo de recuperación en la fecundidad. La sincronización de la liberación de gametos puede contemplar varios factores físicos y biológicos relacionados con señales del ambiente, entre los que se pueden ver involucrados mecanismos endógenos de los relojes biológicos (Soong et al. 2005). Dado lo anterior, muchas especies en áreas tropicales tienen una sola temporada reproductiva al año al igual que las dos especies estudiadas en el presente trabajo. En algunos casos para especies con un amplio rango latitudinal, la reproducción puede ocurrir a lo largo del año en las poblaciones cercanas al ecuador, mientras que puede ser estacional en aquellas que se encuentran a mayor distancia de este (Giese y Pearse 1974). No obstante, la reproducción continua y largas temporadas de desove son características de una gran porción de especies de los ofiuros tropicales que han sido estudiados en detalle, muchos de los cuales producen un alto número de huevos (Mladenov y Emson 1984). Sin embargo, al ser moduladas dichas temporadas por las señales ambientales, varias especies filogenéticamente cercanas pueden encontrarse influenciadas de la misma manera por la misma combinación de factores, lo que hace necesarios mecanismos que eviten el entrecruzamiento (Soong et al. 2005). La duración del tiempo de reproducción es una respuesta a las presiones selectivas, las cuales reflejan condiciones favorables para que ésta sea exitosa, lo cual es conocido como factores próximos. Por otro lado, las causas últimas son usualmente asociadas con condiciones favorables para el desarrollo de las larvas o los juveniles. Esto en ambientes marinos ha sido catalogado como la regla de Crisp, la cual sostiene que los ciclos reproductivos pueden ser regulados de manera tal que la larva eclosione durante la temporada que es más favorable para la adquisición de plancton, en condiciones que prevalecen sobre una amplia parte de su rango (Giese y Pearse 1974). A pesar de ello, hay pocos datos que lo soporten y dichos datos son difíciles de obtener. Thorson (1950) argumenta que hay muchas especies de aguas templadas que se reproducen cuando el plancton necesario se encuentra disponible para las larvas. A parte de la alimentación existen otras condiciones que son importantes para las larvas, la más obvia de ellas es la temperatura, la cual puede ser necesariamente favorable para el desarrollo de la larva. Los depredadores son otro claro ejemplo de ellas; no obstante, estos pueden ejercer una presión selectiva crítica para la reproducción, no solo sobre las larvas sino también sobre juveniles o adultos (Giese y Pearse 1974). Los modelos de fecundidad sostienen que si el alimento es escaso, el éxito reproductivo baja para las especies cuya estrategia tiene huevos de tamaño pequeño y que éste incrementa con el aumento de talla de huevo, sin embargo cuando los niveles de alimento son altos, entonces el éxito reproductivo es alto para los huevos de tallas pequeñas (Grange et al. 2004, McEdward y Miner 2003). Esto implica que si las dos especies estudiadas tienen una estrategia planctotrófica, en la que los huevos son producidos en grandes cantidades y en tamaños pequeños, es porque los niveles de alimento son suficientes para satisfacer los requerimientos alimenticios de las larvas. Dado que la temporada de desove no coincide con los picos de clorofila (aumento de la productividad primaria) como se observa en los resultados al no tener correlación significativa; se evidencia lo anterior y por

35
lo tanto se propone que las larvas de Ophiocoma aethiops y O. alexandri aparentemente no dependen de esos pulsos de producción para su supervivencia. Esta observación también coincide con Hendler (1979) quien reporta para estas dos especies en Panamá un periodo de liberación de gametos similar al encontrado en el presente estudio. Además, el autor compara de igual forma el tiempo de la liberación con el aumento en la productividad primaria y encuentra que tampoco coinciden. A pesar de que se encontró una correlación significativa del ciclo reproductivo con la temperatura, las horas de luz (fotoperiodo) y las lluvias, y que el análisis de regresión lineal múltiple relacionó estas variables con ambas especies (de diferente forma), la cantidad de varianza explicada es menor del 60%, a diferencia de los estudios realizados para otros equinodermos en la zona, los cuales en la relación con los factores ambientales presentaron valores más altos en la varianza explicada (Martinez-Garcia 2010, Gutierrez-Mendez 2011). Lo anterior conduce a que estas variables no son tan influyentes en la determinación de dicho ciclo. Esto se ve de forma más clara cuando se compara los resultados con Hendler (1979), aunque el estudio de dicho autor fue en Panamá, los ciclos coinciden, conllevando a concluir que los patrones reproductivos no están determinados por una dinámica local. Lo anterior lleva a pensar que el patrón general subyacente de las temporadas reproductivas de las especies de Ophiocoma estudiadas se ha generado como resultado de su proceso evolutivo y en la actualidad tiene un alto componente genético, mientras que la dinámica local de los factores ambientales solo lo ajusta a una escala temporal a corto plazo, con posibles fluctuaciones interanuales, las cuales solo pueden ser evaluadas a través de estudios que se desarrollen en una escala temporal mas amplia. Dado que al interior de las gónadas solo se encontraron ovocitos vitelogénicos (primarios) se puede deducir que el final del proceso meiótico es llevado a cabo de la misma manera que ha sido reportado tanto para ofiuroideos como asteroideos. En los que las divisiones de maduración de los ovocitos no ocurren hasta poco tiempo antes del desove o inmediatamente después de este (Patent 1969, Hendler 1991). Lo anterior concuerda con Giese y Pearse (1974) que sostienen que también se presenta en otros invertebrados; en los que los huevos son liberados como ovocitos primarios (vitelogénicos) y la división meiótica es completada después del desove. Los autores hacen algunas aclaraciones entre los términos huevo y óvulo dado que las diferencias en la forma de determinar el proceso meiótico con respecto al desove y la fecundación ha causado confusiones considerables en la terminología (Giese y Pearse 1974); con base en lo anterior para este escrito se han tratado los conceptos de la misma manera. Al hacer la comparación entre la relación del índice gonadal y la fecundidad entre ambas especies se puede ver claramente como presentan una relación inversa, donde O. alexandri tiene mayores valores de índice gonadal y menores de fecundidad, mientras que en O. aethiops pasa lo contrario. Esto podría indicar que la especie que tiene mayor índice gonadal produce una menor cantidad de ovocitos y viceversa. El hecho de tener menos ovocitos en una gónada más pesada sugiere que dichos huevos son más densos comparado con los que se encuentran en una gónada con mayor número pero menos peso. Esta diferencia de densidades entre los ovocitos de ambas especies puede llegar a ser uno de los factores que contribuyen en el aislamiento reproductivo, ya que como se ha mencionado

36
anteriormente, estas dos especies son cogenéricas y sus periodos reproductivos están solapados. Probablemente la diferencia de flotación entre los gametos de ambas especies es un factor que hace que estos se ubiquen a diferentes alturas en la capa de agua, exponiéndose así a diferentes tipos de presión selectiva, como la dispersión por corrientes y los depredadores. Lo anterior es consecuente con lo reportado por Hendler (1991), quien afirma que la gravedad específica de los ovocitos es una característica que indudablemente influencia el éxito de fertilización y la dispersión del embrión, además el autor aclara que esto puede variar entre poblaciones y especies; sin embargo, los factores determinantes de la flotación de los ovocitos son desconocidos. De esto parte la necesidad de hacer estudios que conduzcan a conocer la naturaleza química de cada uno de los huevos y su atracción con los espermatozoides. Así, se podría determinar realmente cuáles son las diferencias entre ambas especies y cómo ellas puedan contribuir en el aumento del éxito reproductivo. Al confirmar dichas diferencias, estas se podrían considerar como un factor de aislamiento precigótico, el cual contribuiría a evitar entrecruzamiento. Dentro de las características más relevantes encontradas se observaron los valores correspondientes a la talla máxima de los huevos. Ophiocoma aethiops presentó huevos que llegan hasta 98µm y O. alexandri hasta 89µm de diámetro. Además, se pudo establecer un valor de fecundidad que alcanzó los valores de 6.413*106 para O. aethipos y de 3.312*106 para O. alexandri. Lo anterior es importante dado que coincide con las estrategias reproductivas que se caracterizan por bajos niveles de aprovisionamiento de huevo (estrategias de huevo pequeño), que presentan una alta fecundidad pero puede llegar a tener un proceso de desarrollo prolongado hasta la metamorfosis (McEdward y Miner 2003). En general las características de los patrones de desarrollo en ofiuroideos han sido establecidas, de modo tal que especies con huevos pequeños (70-200µm de diámetro), producen aproximadamente entre 19.000 y 2.6*106 huevos, tienen un desarrollo plantotrófico y requieren de 12 a 40 días aproximadamente (pueden ser más) para la metamorfosis (Schoener 1972, Hendler 1975, Mladenov 1979). Al asociar dicha información con lo encontrado para las dos especies de Ophiocoma estudiadas, se puede sostener que éstas presentan un desarrollo plantotrófico; lo cual ha sido confirmado por medio de observaciones personales. Sin embargo, se requieren más estudios para elucidar el patrón de desarrollo y las características específicas de cada una de las larvas de ambas especies. Esto es de gran importancia ya que el tiempo de desarrollo esta asociado con una tasa de mortalidad de las larvas cuando se encuentran en el plancton, la cual es muy alta generalmente (McEdward y Miner 2003).

37
Conclusiones Ophiocoma aethiops y Ophiocoma alexandri son especies que presentan un ciclo reproductivo anual, con una temporada de liberación de gametos marcada que va de mayo a noviembre para O. aethiops y de abril a diciembre en el caso de O. alexandri. Ambas especies presentaron una proporción de sexos 1:1. Las variables temperatura, horas de luz y pluviosidad fueron las únicas que tuvieron una correlación significativa positiva con los ciclos reproductivos, es decir que estas tienen influencia en ellos y forman parte de los factores próximos que afectan los mismos. La forma en que las variables ambientales contribuyen en las variaciones del ciclo aún no es clara y son necesarios estudios que conduzcan a dilucidar dicha relación. Ambas especies tienen un comportamiento similar en cuanto a la temporada de liberación de gametos, sin embargo presentan diferencias en los valores de fecundidad e índices gonadales. Es posible que los ovocitos de ambas especies presenten diferencia en las densidades, lo que conlleva a una diferencia de flotación, la cual probablemente contribuye como mecanismo de aislamiento reproductivo precigótico.

38
Literatura Citada
Benítez-Villalobos F, 2005. Reproduction and larval biology of North Atlantic asteroids related to the invasion of the deep sea. Thesis PhD. University of Southampton, 150 pp.
Borges M, Yokoyama L Q y Amaral A C Z, 2009. Gametogenic cycle of Ophioderma januarii, a
common Ophiodermatidae (Echinodermata: Ophiuroidea) in southeastern Brazil. Zoologia 26(1): 118–126.
Byrne M, 1989. Ultraestructure of the ovary and oogenesis in the ovoviviparous ophiuroid
Ophiolepis paucispina (Echinodermata). Biological Bulletin, 176: 79-95. Byrne M, Cisternas P y O’Hara T, 2008. Brooding of pelagic-type larvae in Ophiopeza spinosa:
Reproduction and development in a tropical Ophiodermatid brittlestar. Invertebrate Biology 127(1): 98–107.
CONAGUA, 2000. Informe de los promedios mensuales en algunas las variables meteorológicas
del período 1981 a 2000 para Puerto Ángel. SMN-CNA. http://smn.cna.gob.mx/observatorios/historica/ptoangel.pdf
Falkner I y Byrne M, 2003. Reproduction of Ophiactis resiliens (Echinodermata: Ophiuroidea) in
New South Wales with observations on recruitment. Marine Biology 143: 459–466. Fenaux F, 1970. Maturation of the gonads and seasonal cycle of the planktonic larvae of the
ophiuroid Amphiura chiajei Forbes. Biological Bulletin 138: 262–271. García E, 1981. Modificaciones al sistema de clasificación climática de Köppen. Instituto de
Geografía, UNAM. Tercera edición. México, D.F, 252 pp. Gage J D y Tyler P A, 1991. Deep-sea biology: A natural history of organisms at the deep-sea
floor. Cambridge University Press, Cambridge, 504 pp. Gage J D, Anderson R M, Tyler P A, Chapman R y Dolan E, 2004. Growth, reproduction and
possible recruitment variability in the abyssal brittle star Ophiocten hastatum (Ophiuroidea: Echinodermata) in the NE Atlantic. Deep-Sea Research I, 51: 849–864.
Giese A C y Pearse J S, 1974. Introduction: General principles. En: Giese A C y Pearse J S (Eds).
Reproduction of marine invertebrates. Vol 1. Academic Press, New York, pp 1–49. Gillespie J M y McClintock J B, 2007. Brooding in echinoderms: How can modern experimental
techniques add to our historical perspective? Journal of Experimental Marine Biology and Ecology, 342: 191–201.

39
Grange L J, Tyler P A, Peck Ll S y Cornelius N, 2004. Long-term interannual cycles of the gametogenic ecology of the Antarctic brittle star Ophionotus victoriae. Marine Ecology Progress Series, 278: 141–155.
Grange L J, 2005. Reproductive success in Antarctic marine invertebrates. Thesis PhD. University
of Southampton, 397 pp. Grave C, 1898. Embryology of Ophiocoma echinata, Agassiz. Preliminary note. Johns Hopkings
Univ. Circ 18: 6–7. Gutierrez-Mendez I S, 2011. Biología reproductiva de Holothuria (Stauropora) fuscocinerea
Jaeger, 1833 (Echinodermata: Holothuroidea) en Bahía La Entrega, Oaxaca. México. Tesis de Licenciatura. Universidad del Mar, 60 pp.
Hendler G, 1973. Northwest Atlantic amphiuroid brittlestars, Amphioplus abditus (Verril),
Amphioplus macilentus (Verril), and Amphioplus sepultus n.sp. (Ophiuroidea: Echinodermata): Systematics, zoogeography, annual periodicities, and larval adaptations. Ph.D. Thesis, University of Connecticut.
Hendler G, 1975. Adaptational significance of the patterns of ophiuroid development. American
Zoologist, 15: 691–715. Hendler G, 1979. Reproductive periodicity of Ophiuroids (Echinodermata: Ophiuroidea) on the
Atlantic and Pacific coasts of Panama. En: Stancyk S E (Ed.). Reproductive ecology of marine invertebrates. Univversity of South Carolina Press, pp. 145–156.
Hendler G, 1991. Echinodermata: Ophiuroidea. En: Giese A C, Pearse J S y Pearse V (Eds).
Reproduction of marine invertebrates. Vol VI. Echinoderms and Lophophorates. Boxwood Press, pp. 356–510.
Levitan D R, 1991. The importance of sperm limitation to the evolution of egg size in marine
invertebrates. American Naturalist, 141: 517–536. Martinez-Garcia M, 2010. Patrones reproductivos de Pharia pyramidatus (Gray, 1840)
(Echinodermata: Asteroidea) en Isla Montosa, Pacífico Tropical Mexicano. Tesis de Licenciatura. Universidad del Mar, 67 pp.
McEdward L R y Janies D A, 1997. Relationships among development, ecology and morphology
in the evolution of Echinoderm larvae and life cycles. Biological Journal of the Linnean Society, 60: 381–400.
McEdward L R y Miner B G, 2003. Fecundity-time models of reproductive strategies in marine
benthic invertebrates: Fitness differences under fluctuating environmental conditions. Marine Ecology Progress Series, 256: 111–121.

40
McEdward L R y Morgan K H, 2001. Interspecific relationships between egg size and the level of parental investment per offspring in echinoderms. Biological Bulletin, 200: 33–50.
Mercier A y Hamel J P, 2009. Endogenous and exogenous control of gametogenesis and spawning
in echinoderms. Advances in Marine Biology, 55: 1-291. Mladenov P V, 1976. Reproduction and larval development in ophiuroids (Echinodermata) of
Barbados, West Indies. M.Sc. Thesis, McGill University, 136pp. Mladenov P V, 1979. Unusual lecithotrophic development of the Caribbean brittle star Ophiothrix
oerstedi. Marine Biology, 55: 55–62. Mladenov P V y Emson R H, 1984. Divide and broadcast: Sexual reproduction in the West Indian
brittle star Ophiocomella ophiactoides and its relationship to fissiparity. Marine Biology, 81: 273–282.
Mortensen T, 1931. Contribution to the study of the development and larval forms echinoderms. I.
The development and larval forms of some tropical echinoderms. II. Observations on some Scandinavian echinoderm larvae. K. Danske Vidensk. Selsk. Skr. 4: 1–39
Orton J H, 1920. Sea temperature, breeding and distribution in marine animals. Journal of the
Marine Biological Association of the U.K., 12: 339–366. Patent D H, 1969. The reproductive cycle of Gorgonocephalus caryi (Echinodermata:
Ophiuroidea). Biological Bulletin, 136: 241–252. Pearse J S, 1974. Reproductive patterns of tropical reef animals: Three species of sea urchins. Proc.
Second Int. Coral Reef Symp. 1: 235–240. Poulin É y Féral J-P, 1996. Why are there so many species of brooding Antarctic echinoids?
Evolution, 50(2): 820–830. Ramirez-Llodra E Z, 2001. Reproductive patterns of deep-sea invertebrates related to phylogeny
and energy availability. PhD Dissertation. University of Southampton, 214 pp. Schoener A, 1972. Fecundity and possible mode of development of some deep-sea ophiuroids.
Limnology and Oceanography, 17(2): 193–199. Selvakumaraswamy P y Byrne M, 1995. Reproductive cycle of two populations of Ophionereis
schayeri (Ophiuroidea) in New South Wales. Marine Biology, 124: 85–97. Selvakumaraswamy P y Byrne M, 2000. Vestigial ophiopluteal structures in the lecithotrophic
larvae of Ophionereis schayeri (Ophiuroidea). Biological Bulletin, 198: 379–386. Sewell M A y Young C M, 1997. Are echinoderm egg size distributions bimodal? Biological
Bulletin, 193: 297–305.

41
Smiley S, 1988. The Dynamics of Oogenesis and the Annual Ovarian Cycle of Stichopus
californicus (Echinodermata: Holothuroidea). Biological Bulletin, 175: 79–93 Soong K, Chang D y Chao S M, 2005. Presence of spawn-inducing pheromones in two brittle stars
(Echinodermata: Ophiuroidea). Marine Ecology Progress Series, 292: 195–201. Stephenson A, 1934. The breeding of reef animals. Part II. Invertebrates other than corals. Great
Barrier Reef Exp. 1928–1929. Sci. Rpts. 3: 247–272. Stewart B G y Mladenov P V, 1995. Reproductive periodicity in the euryalinid snake star
Astrobrachion constrictum in a New Zealand fiord. Marine Biology, 123: 543–553. Thorson G, 1950. Reproduction and larval development of marine bottom invertebrates. Biological
Reviews, 25: 1–45. Tominaga H, Nakamura S y Komatsu M, 2004. Reproduction and development of the
conspicuously dimorphic brittle star Ophiodaphne formata (Ophiuroidea). Biological Bulletin, 206: 25–34.
Vance R R, 1973. On reproductive strategies in marine benthic invertebrates. American Naturalist,
47: 337–371. Williams M E y Bentley M G, 2002. Fertilization success in marine invertebrates: The influence of
gamete age. Biological Bulletin, 202: 34–42. Yamashita M y Iwata F, 1983. A quantitative analysis of the annual testicular cycle of the brittle-
star Amphipholis kochii by means of autoradiographic investigation. Biological Bulletin, 164: 327–340.