university of groningen acetylcholine beyond ... filechapter 7 murine precision‐cut lung slices as...
TRANSCRIPT

University of Groningen
Acetylcholine beyond bronchoconstriction: a regulator of inflammation and remodelingKistemaker, Loes
IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite fromit. Please check the document version below.
Document VersionPublisher's PDF, also known as Version of record
Publication date:2015
Link to publication in University of Groningen/UMCG research database
Citation for published version (APA):Kistemaker, L. (2015). Acetylcholine beyond bronchoconstriction: a regulator of inflammation andremodeling. [S.l.]: [S.n.].
CopyrightOther than for strictly personal use, it is not permitted to download or to forward/distribute the text or part of it without the consent of theauthor(s) and/or copyright holder(s), unless the work is under an open content license (like Creative Commons).
Take-down policyIf you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediatelyand investigate your claim.
Downloaded from the University of Groningen/UMCG research database (Pure): http://www.rug.nl/research/portal. For technical reasons thenumber of authors shown on this cover page is limited to 10 maximum.
Download date: 26-02-2019

CHAPTER 7
MURINE PRECISION‐CUT LUNG SLICES AS A MODEL TO STUDY
TGF‐β AND BRONCHOCONSTRICTION‐INDUCED REMODELING
Loes E.M. Kistemaker
I. Sophie T. Bos
Cécile M. Bidan
Jürgen Wess
Huib A.M. Kerstjens
Herman Meurs
Reinoud Gosens

Chapter 7
136
Abstract
Rationale
Asthma is a chronic obstructive airway disease, characterized by airway remodeling.
Airway remodeling is conventionally thought to be the result of airway inflammation, with
inflammatory cells giving rise to the production of pro‐remodeling factors, such as TGF‐β.
However, new insights suggest that airway remodeling might also or alternatively be a
consequence of bronchoconstriction, which triggers TGF‐β release via biomechanical
activation. Previously, we developed a model to study this phenomenon, using guinea pig
lung slices treated with bronchoconstrictors in culture. As the mouse offers species
advantages with respect to the availability of molecular and genetic tools, we aimed to
develop a similar model using murine precision cut lung slices. In addition, we aimed to
investigate the involvement of the M3 receptor in bronchoconstriction‐induced
remodeling.
Methods
Airway constriction in response to methacholine was assessed in lung slices from wild‐
type and M3R‐/‐ mice. In subsequent experiments, lung slices from wild‐type and M3R
‐/‐
mice were stimulated with TGF‐β1 (2 ng/ml) or methacholine (10 μM) for two days to
induce airway remodeling. Thereafter, slices were used for gene expression analysis,
Western Blot analysis, 2‐photon imaging, or for constriction experiments.
Results
Slices were viable for at least three days. Methacholine induced a dose dependent airway
constriction in wild‐type mice, with a maximal effect of 80%. In M3R‐/‐ mice, a 35% airway
constriction was observed. TGF‐β induced a 2‐fold increase in the mRNA expression of
fibronectin, collagen Iα1 and α‐sm‐actin in both wild‐type and M3R‐/‐ mice. Similar findings
were observed on the protein level. 2‐Photon imaging revealed that collagen and α‐sm‐
actin expression was localized around the airways. No effect of methacholine on
parameters of remodeling was observed. Stimulation with TGF‐β or methacholine did not
alter airway constriction induced by the thromboxane A2 agonist U46619 compared to
control conditions in both wild‐type and M3R‐/‐ mice.
Conclusions
Murine lung slices are suitable to study TGF‐β‐induced airway remodeling, which was
comparable in wild‐type and M3R‐/‐ mice. Methacholine did not induce remodeling in
murine lung slices, suggesting that the mouse, in contrast to the guinea pig, is not a good
model to study bronchoconstriction‐induced remodeling. Despite effects of TGF‐β on the
expression of fibronectin, collagen Iα1 and α‐sm‐actin, this did not translate into changes
in airway contractility. Further studies, using a pharmacological approach, are needed to

Lung slices as a model to study remodeling
137
investigate the potential involvement of the M3 receptor in bronchoconstriction‐induced
remodeling.
Introduction
Asthma is a chronic obstructive airway disease, which is characterized by structural
changes in the airways, referred to as airway remodeling. Airway remodeling
encompasses airway smooth muscle thickening and excessive deposition of extracellular
matrix proteins (1). These airway structural changes are considered to be an important
component of airflow limitation in asthma and may contribute to an accelerated decline in
lung function (2, 3). In addition, asthma is characterized by airway hyperresponsiveness,
which consists of an excessive airway narrowing in response to bronchoconstricting
stimuli, and by airway inflammation (4).
It is believed that airway remodeling is the result of ongoing airway inflammation. For
example, eosinophils are a rich source of the pro‐remodeling factor TGF‐β (5). However,
the severity of airway remodeling in patients with asthma is not related to the severity of
inflammation, and treatment with corticosteroids, which modify airway inflammation,
does not affect the decline in lung function or the natural history of asthma (6‐8).
New insights suggest that airway remodeling might also or alternatively be a consequence
of bronchoconstriction. In patients with asthma, repeated challenges with methacholine,
inducing bronchoconstriction, induced features of airway remodeling, without affecting
eosinophilic inflammation. Interestingly, these effects could be prevented when
methacholine was combined with the β2‐agonist albuterol, suggesting that
bronchoconstriction is importantly involved (9). Furthermore, in a mouse model of
asthma, we demonstrated that ovalbumin‐induced remodeling is prevented in muscarinic
M3 receptor deficient (M3R‐/‐) mice, in which bronchoconstriction is almost completely
abolished, without affecting the inflammatory response (chapter 6). Together, these
studies indicated that airway remodeling is not merely induced by inflammation, but may
also be a consequence of mechanical forces applied to the airways during
bronchoconstriction.
The notion that mechanical forces could promote airway remodeling is also supported by
in vitro evidence. For example, compression of epithelial cells increases the activation of
TGF‐β (10), the epithelial thickness (11), and the expression of fibronectin and collagen in
a co‐culture system with fibroblasts (12). Moreover, in airway smooth muscle cells,
muscarinic receptor stimulation and mechanical strain induces TGF‐β activation (13) and

Chapter 7
138
myosin expression (14). Clearly, mechanical forces have an impact on multiple cell types in
the airway, and might thereby enhance remodeling of the airways.
Recently we developed a model to study bronchoconstriction‐induced remodeling ex vivo,
using guinea pig precision‐cut lung slices (PCLS). Oenema et al. demonstrated that
stimulation of guinea pig PCLS with methacholine or other bronchoconstrictors induces
airway remodeling (15). The effects of methacholine were comparable to the effects of
TGF‐β, and were shown to be mediated via TGF‐β (15). In this study, we aimed to develop
a model to study bronchoconstriction‐induced remodeling using murine PCLS as the
mouse offers species advantages with respect to the availability of molecular and genetic
tools. In addition, we aimed to investigate the potential involvement of the M3 receptor in
these responses, for which PCLS from M3R‐/‐ mice were used.
Methods
Animals
Homozygous, inbred, specific‐pathogen‐free breeding colonies of M3R‐/‐ mice and
C57Bl/6NTac wild‐type mice with the same genetic background were obtained from
Taconic (Cambridge City, Indiana, USA). The M3R‐/‐ mice used were on a 129 Sv/J
background and backcrossed for at least 10 generations onto the C57Bl/6NTac
background. M3R‐/‐ mice did not differ from their wild‐type littermates in overall health,
fertility and longevity (16). Animals were housed conventionally under a 12‐h light‐dark
cycle and received food and water ad libitum. All experiments were performed in
accordance with the national guidelines and approved by the University of Groningen
Committee for Animal Experimentation (DEC5463I and DEC6792A).
Precision‐cut lung slices
Mouse PCLS were prepared, according to a protocol described previously for guinea pig
PCLS by our lab (15). Male mice (6‐8 weeks old) were euthanized by intraperitoneal
pentobarbital injection (400 mg/kg, hospital pharmacy, University Medical Center
Groningen), after which the lungs were filled with 1.5 ml low melting‐point agarose
solution (1.5% final concentration (Gerbu Biotechnik GmbH, Wieblingen, Germany) in
CaCl2 (0.9 mM), MgSO4 (0.4 mM), KCl (2.7 mM), NaCl (58.2 mM), NaH2PO4 (0.6 mM),
glucose (8.4 mM), NaHCO3 (13 mM), Hepes (12.6 mM), sodium pyruvate (0.5 mM),
glutamine (1 mM), MEM‐amino acids mixture (1:50), and MEM‐vitamins mixture (1:100),
pH=7.2). The agarose was solidified for 15 minutes by placing the lungs on ice. Lungs were
harvested and individual lobes were sliced at a thickness of 250 µm in medium composed
of CaCl2 (1.8mM), MgSO4 (0.8 mM), KCl (5.4 mM), NaCl (116.4 mM), NaH2PO4 (1.2 mM),

Lung slices as a model to study remodeling
139
glucose (16.7 mM), NaHCO3 (26.1 mM), Hepes (25.2 mM), pH = 7.2, using a tissue slicer
(Compresstome™ VF‐300 microtome, Precisionary Instruments, San Jose CA, USA).
Thereafter, slices were kept at 37°C in a humidified atmosphere of 5% CO2 and washed
every 30 minutes for four times to remove the agarose and cell debris in medium
composed of CaCl2 (1.8mM), MgSO4 (0.8 mM), KCl (5.4 mM), NaCl (116.4 mM), NaH2PO4
(1.2 mM), glucose (16.7 mM), NaHCO3 (26.1 mM), Hepes (25.2 mM), sodium pyruvate
(1mM), glutamine (2 mM), MEM‐amino acids mixture (1:50), MEM‐vitamins mixture
(1:100,) penicillin (100 U/mL) and streptomycin (100 µg/mL), pH = 7.2. Average diameter
of the airways was 179 µM ± 82 µM.
Mitochondrial activity assay
Viability of lung slices was assessed by a mitochondrial activity assay performed after
culturing the slices for different time periods in Dulbecco’s Modification of Eagle’s
Medium (DMEM) supplemented with sodium pyruvate (1 mM), non‐essential amino acid
mixture (1:100), gentamycin (45 µg/mL), penicillin (100 U/mL), streptomycin (100 µg/mL)
and amphotericin B (1.5 µg/mL) at 37°C‐5% CO2. Slices were incubated with Hanks’
balanced salt solution (pH 7.4), containing 10% Alamar blue solution (BioSource,
Camarillo, CA), for 30 min at 37°C‐5% CO2. Mitochondrial activity was assessed by
conversion of Alamar blue into its reduced form, as described previously (17).
Treatment of lung slices
Lung slices were incubated in Dulbecco’s Modification of Eagle’s Medium (DMEM)
supplemented with sodium pyruvate (1 mM), non‐essential amino acid mixture (1:100),
gentamycin (45 µg/mL), penicillin (100 U/mL), streptomycin (100 µg/mL) and
amphotericin B (1.5 µg/mL) at 37°C‐5% CO2 in a 12‐well plate, using 3 slices per well. The
slices were treated with vehicle (CTR), 2 ng/ml TGF‐β1 (R&D systems, Abingdon, UK) or 10
μM methacholine (MCh; ICN Biomedicals, Zoetermeer, the Netherlands) for two days.
After two days, slices were collected for contraction studies, gene expression analysis,
Western Blot analysis, or for 2‐photon imaging.
Airway narrowing studies
Airway narrowing studies were performed on naïve lung slices and on slices treated with
vehicle, TGF‐β or methacholine (MCh) for two days. On naïve lung slices, dose response
curves for MCh (10‐9M ‐ 10‐3M) were recorded, as well as constriction after a single dose of
MCh (10‐4M), after which the airways were dilated with the bitter taste receptor agonist
chloroquine (10‐3M, Sigma‐Aldrich, Zwijndrecht, The Netherlands). On slices treated with
vehicle, TGF‐β or MCh for two days, dose response curves towards the thromboxane A2
agonist U‐46619 (10‐10M ‐ 10‐5M, Sigma‐Aldrich) were recorded. As described previously, a

Chapter 7
140
nylon mesh and metal washer were used to keep the lung slice in place (18). Lung slice
images were captured in time‐lapse (1 frame per 2 seconds) using an inverted phase
contrast microscope (Eclipse, TS100; Nikon). Airway luminal area was quantified using
image acquisition software (NIS‐elements; Nikon).
Gene expression analysis
Total RNA was extracted from lung slices by automated purification using the Maxwell 16
instrument and the corresponding Maxwell 16 LEV simply RNA tissue kit (Promega,
Madison, USA) according to the manufacturer's instructions. Equal amounts of total mRNA
were then reverse transcribed and cDNA was subjected to real‐time qPCR (Westburg,
Leusden, The Netherlands). Real time PCR was performed with denaturation at 94°C for 30
seconds, annealing at 59°C for 30 seconds and extension at 72°C for 30 seconds for 40
cycles followed by 10 minutes at 72°C. Real‐time PCR data were analyzed using the
comparative cycle threshold (Ct: amplification cycle number) method. The amount of
target gene was normalized to the endogenous reference gene 18S ribosomal RNA. The
specific forward and reverse primers used are listed in table 1.
Western Blot analysis
Lung slices were washed once with ice‐cold phosphate‐buffered saline (PBS, composition:
NaCl (140 mM), KCl (2.6 mM), KH2PO4 (1.4 mM), Na2HPO4 (8.1 mM), pH 7.4), followed by
lysis using ice‐cold SDS‐lysis buffer (Tris‐HCl (62.5 mM), SDS (2 %), NaF (1 mM), Na3VO4 (1
mM), aprotinin (10 μg/mL), leupeptin (10 μg/mL), pepstatin A (7 µg/mL) at pH 8.0).
Protein amounts were determined by Pierce and equal amounts of protein were
separated on SDS polyacrylamide gels, transferred onto nitrocellulose membranes,
followed by standard immunoblotting techniques. All bands were normalized either to
lamin AC or to tubulin.
Table 1. Primers used for qRT‐PCR analysis.
Gene Primer sequence NCBI accession
number
Fibronectin Forward – ACCACCCAGAACTACGATGC
Reverse – GGAACGTGTCGTTCACATTG
NM_010233.2
Collagen Iα1 Forward – CACCCTCAAGAGCCTGAGTC
Reverse – GTTCGGGCTGATGTACCAGT
NM_007742.3
α‐sm‐actin Forward – CTGACAGAGGCACCACTGAA
Reverse – CATCTCCAGAGTCCAGCACA
X13297.1
18S Forward – AAACGGCTACCACATCCAAG
Reverse – CCTCCAATGGATCCTCGTTA
NR_003278

Lung slices as a model to study remodeling
141
2‐photon imaging
An immunofluorescent antibody staining was used to visualize α‐smooth muscle‐actin,
and 2‐photon imaging was used to visualize collagen. For α‐smooth muscle‐actin staining,
lung slices were washed two times with cytoskeleton buffer (CB: MES (10 mM), NaCl (150
mM), EGTA (5 mM), MgCl2 (5 mM), and glucose (5 mM) at pH=6.1) and fixed with CB
containing 3% paraformaldehyde (PFA) for 15 min. Thereafter, slices were incubated with
CB buffer containing 3% PFA and 0.3% Triton‐X‐100 for 5 min, followed by two washes
with CB buffer. Lung slices were blocked for 1 h in cyto‐TBS (Tris‐base (20 mM), NaCl (154
mM), EGTA (2.0 mM) and MgCl2 (2.0 mM), pH=7.2), supplemented with BSA (1%) and
normal donkey serum (2%). Thereafter, lung slices were stained with rabbit anti‐alpha
smooth muscle actin antibody (Abcam, Cambridge, UK) overnight at 4°C. After washing 3
times with cyto‐TBS containing 0.1% Tween‐20 (cyto‐TBS‐T) for 10 min, incubation with
the secondary antibody (FITC) was performed during 2 hour at room temperature. Lung
slices were then washed 3 times for 15 min in cyto‐TBS‐T, followed by 3 washes with ultra‐
pure water. The slides were mounted with ProLong Gold anti‐fade reagent (Invitrogen,
Breda, The Netherlands). The slides were then analyzed using a fluorescence confocal
microscope (Leica) equipped with a multiphoton laser. The α‐sm‐actin staining was excited
at 100 nm and collected at 500 nm, whereas the collagen bundles excited at 820 nm
naturally emitted a second harmonic generation signal collected around 410 nm.
Statistical analysis
Data are presented as mean ± s.e. of the mean. Statistical differences between means
were calculated using a paired Student’s t‐test with two‐tailed distribution. Differences
were considered significant at p<0.05.
Results
Viability of lung slices
Viability of the lung slices was assessed by a mitochondrial activity assay after 1, 2 and 3
days of culturing. As depicted in figure 1, no significant differences were observed in
mitochondrial activity for up to three days after culturing. This indicates that the lung
slices are viable for at least three days and can be used for experiments within this period.
Methacholine‐induced constriction
MCh induced a dose‐dependent airway constriction in PCLS from wild‐type mice, with a
maximal effect of 80% airway closure compared to the initial airway lumen (figure 2A). In
PCLS from M3R‐/‐ mice, the airways were more sensitive to MCh compared to wild‐type
PCLS, but the maximal airway narrowing was only 35% (figure 2A).

Chapter 7
142
Figure 1. Viability of lung slices is preserved for three days. Mitochondrial activity, assessed by conversion of
Alamar blue into its reduced form, was not significantly different in the first three days of culture. Data represent
mean ± s.e. of the mean of 4 animals, 2 slices per animal.
As depicted in figure 2B, MCh induced 40% airway closure in PCLS from wild‐type mice
after a single dose of MCh (10‐4M), and airway dilation was obtained after subsequent
addition of chloroquine (10‐3M). In contrast to PCLS from wild‐type animals, in PCLS from
M3R‐/‐ mice there was a lag time after addition of MCh before the airway constriction
started. However, the constriction rate was higher, and a plateau phase was reached
earlier in slices from M3R‐/‐ mice compared to wild‐type mice. The maximal effect of MCh
in PCLS from M3R‐/‐ mice was 30% closure, and the airways similarly dilated after addition
of chloroquine (figure 2B). Oscillations in airway constriction were observed in the PCLS
from M3R‐/‐ mice, as is demonstrated in figure 2C for a single experiment in which strong
oscillations were observed. The oscillations can also be observed in the plateau phase of
constriction in figure 2B; however, the effect of the strong oscillations observed in single
experiments is leveled out by averaging multiple experiments.

Lung slices as a model to study remodeling
143
Figure 2. Methacholine‐induced airway constriction in PCLS from wild‐type and M3R
‐/‐ mice. A: Airway
constriction in response to increasing concentrations of methacholine (MCh) in PCLS from wild‐type and M3R‐/‐
mice. Data represent mean ± s.e. of the mean, n=8 for wild‐type mice, n=4 for M3R‐/‐ mice, 2‐3 airways were
quantified for each animal. B: Airways were constricted with MCh (10‐4M) and after 10 minutes dilated with
chloroquine (Cq, 10‐3M). Data represent mean of 8 mice, 3‐7 airways were quantified for each animal. For clarity
reasons, error bars are omitted. C. A single trace of airway contraction in an M3R‐/‐ mouse, in which strong
oscillations can be observed.
Remodeling in PCLS from wild‐type animals
In order to assess remodeling effects in PCLS from wild‐type animals, slices were treated
with vehicle, TGF‐β or MCh for two days, and expression of the extracellular matrix
proteins fibronectin and collagenIα1, and the contractile protein α‐sm‐actin, was assessed.
Stimulation with TGF‐β for two days induced a 1.7‐fold increase in fibronectin mRNA
expression (figure 3A), a 2.4‐fold increase in collagen Iα1 mRNA expression (figure 3B) and

Chapter 7
144
a 2.0‐fold increase in α‐sm‐actin mRNA expression (figure 3C) compared to vehicle control.
Similar findings were observed on the protein level for all markers (1.8‐3.2 fold increase,
data not shown). In addition, to evaluate bronchoconstriction‐induced remodeling, the
effects of MCh were assessed. As shown in figure 3, MCh did not induce an increase in the
mRNA expression of fibronectin, collagenIα1 or α‐sm‐actin.
Figure 3. TGF‐β, and not methacholine, induces remodeling in PCLS from wild‐type mice. Lung slices from wild‐
type mice were treated with TGF‐β (2 ng/ml) or methacholine (MCh, 10 µM) for two days. mRNA expression of
fibronectin (A), collagen Iα1 (B) and α‐sm‐actin (C) was analyzed. Data represent mean ± s.e. of the mean, n=8
animals, * p<0.05; ** p<0.01 compared to vehicle treated controls (CTR).
Lung slices as a model to study remodeling
145
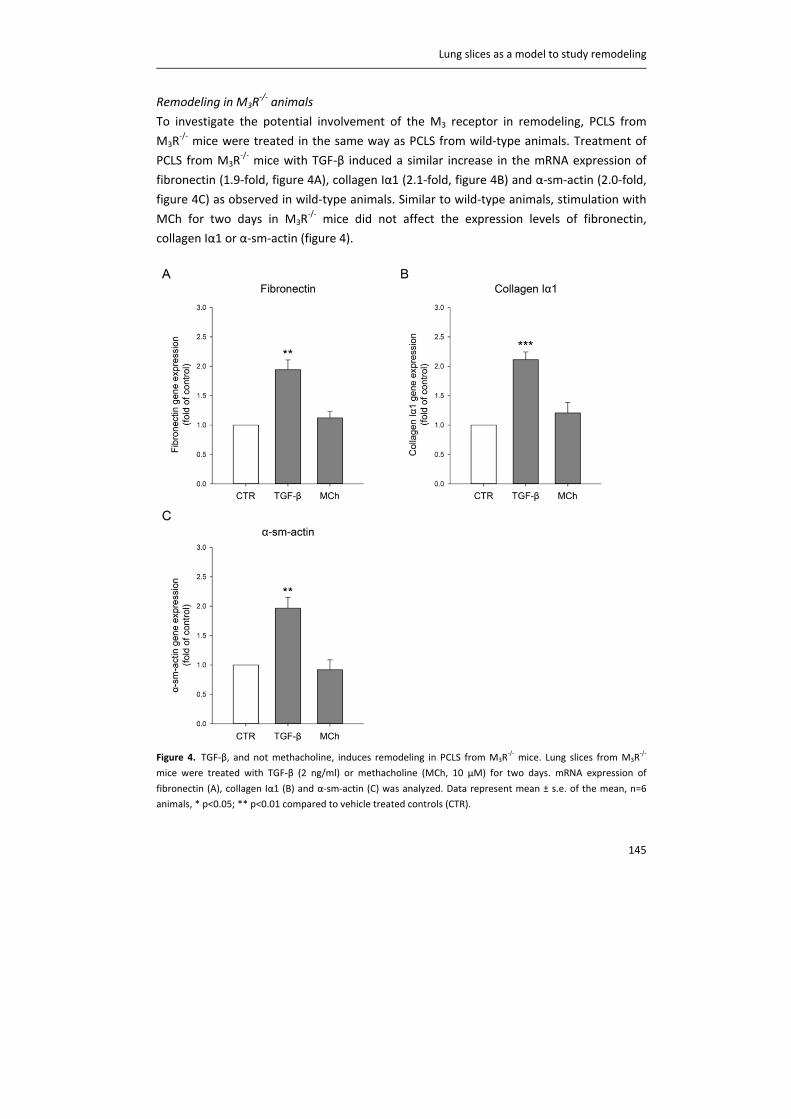
Remodeling in M3R‐/‐ animals
To investigate the potential involvement of the M3 receptor in remodeling, PCLS from
M3R‐/‐ mice were treated in the same way as PCLS from wild‐type animals. Treatment of
PCLS from M3R‐/‐ mice with TGF‐β induced a similar increase in the mRNA expression of
fibronectin (1.9‐fold, figure 4A), collagen Iα1 (2.1‐fold, figure 4B) and α‐sm‐actin (2.0‐fold,
figure 4C) as observed in wild‐type animals. Similar to wild‐type animals, stimulation with
MCh for two days in M3R‐/‐ mice did not affect the expression levels of fibronectin,
collagen Iα1 or α‐sm‐actin (figure 4).
Figure 4. TGF‐β, and not methacholine, induces remodeling in PCLS from M3R‐/‐ mice. Lung slices from M3R
‐/‐
mice were treated with TGF‐β (2 ng/ml) or methacholine (MCh, 10 µM) for two days. mRNA expression of
fibronectin (A), collagen Iα1 (B) and α‐sm‐actin (C) was analyzed. Data represent mean ± s.e. of the mean, n=6
animals, * p<0.05; ** p<0.01 compared to vehicle treated controls (CTR).
Chapter 7
146
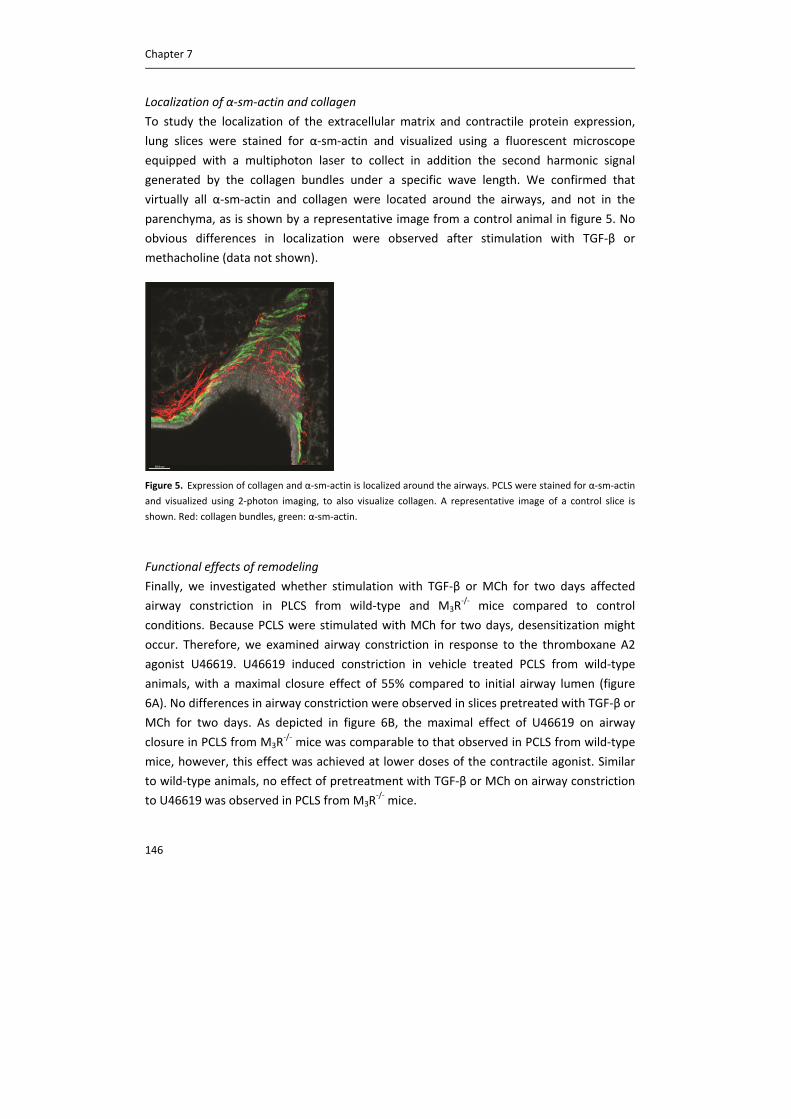
Localization of α‐sm‐actin and collagen
To study the localization of the extracellular matrix and contractile protein expression,
lung slices were stained for α‐sm‐actin and visualized using a fluorescent microscope
equipped with a multiphoton laser to collect in addition the second harmonic signal
generated by the collagen bundles under a specific wave length. We confirmed that
virtually all α‐sm‐actin and collagen were located around the airways, and not in the
parenchyma, as is shown by a representative image from a control animal in figure 5. No
obvious differences in localization were observed after stimulation with TGF‐β or
methacholine (data not shown).
Figure 5. Expression of collagen and α‐sm‐actin is localized around the airways. PCLS were stained for α‐sm‐actin
and visualized using 2‐photon imaging, to also visualize collagen. A representative image of a control slice is
shown. Red: collagen bundles, green: α‐sm‐actin.
Functional effects of remodeling
Finally, we investigated whether stimulation with TGF‐β or MCh for two days affected
airway constriction in PLCS from wild‐type and M3R‐/‐ mice compared to control
conditions. Because PCLS were stimulated with MCh for two days, desensitization might
occur. Therefore, we examined airway constriction in response to the thromboxane A2
agonist U46619. U46619 induced constriction in vehicle treated PCLS from wild‐type
animals, with a maximal closure effect of 55% compared to initial airway lumen (figure
6A). No differences in airway constriction were observed in slices pretreated with TGF‐β or
MCh for two days. As depicted in figure 6B, the maximal effect of U46619 on airway
closure in PCLS from M3R‐/‐ mice was comparable to that observed in PCLS from wild‐type
mice, however, this effect was achieved at lower doses of the contractile agonist. Similar
to wild‐type animals, no effect of pretreatment with TGF‐β or MCh on airway constriction
to U46619 was observed in PCLS from M3R‐/‐ mice.

Lung slices as a model to study remodeling
147
Figure 6. Pretreatment of PCLS with TGF‐β or methacholine does not affect U46619‐induced constriction. Lung
slices from wild‐type and M3R‐/‐ mice were pretreated with vehicle (CTR), TGF‐β (2 ng/ml) or methacholine (MCh,
10 µM) for two days. Airway constriction in response to increasing concentrations of U46619 was assessed in
wild‐type mice (A) and in M3R‐/‐ mice (B). Data represent mean ± s.e. of the mean of 4 animals, 2‐3 airways were
quantified for each animal.
Discussion
We hypothesized that mouse PCLS would be a good model to study bronchoconstriction‐
induced airway remodeling and its functional consequences, and aimed to investigate the
involvement of the M3 receptor in this response. However, although TGF‐β induced an
increase in the expression of fibronectin, collagen and α‐sm‐actin in the airways of wild‐
type mice and M3R‐/‐ mice, no effect of methacholine on airway remodeling was observed.
Previously, it has been shown by our lab that methacholine can induce airway remodeling
in guinea pig PCLS (15). Therefore, species differences could underlie the lack of effect of
methacholine in mouse PCLS. It is known that maximal contraction towards methacholine
is higher in guinea pig PCLS compared to mouse PCLS, and that the relative amount of
airway smooth muscle in the airways is considerably higher in the guinea pig compared to
the mouse. Moreover, whereas guinea pig PCLS stay contracted over 48 hours while
continuously stimulated with methacholine, our unpublished observations indicate that
contraction in response to methacholine is rapidly lost in mouse PCLS (not shown).
Therefore, the mouse PCLS preparation may not be a good model to study
bronchoconstriction‐induced remodeling, which might be explained by species differences
between guinea pigs and mice.

Chapter 7
148
TGF‐β did induce features of airway remodeling in mouse PCLS, which is in line with
previous studies (15, 19). However, despite the clear effect of TGF‐β on the expression of
fibronectin, collagen Iα1 and α‐sm‐actin, this did not translate into changes in airway
contractility. Generally, it is assumed that airway remodeling affects airway contractility.
Here, we were not able to demonstrate such a direct link. Chew et al. demonstrated that
although ovalbumin challenge, leading to airway remodeling, induced airway
hyperresponiveness in vivo, airway constriction of mouse PCLS in response to
acetylcholine ex vivo was not altered (20). Using a similar approach, Donovan et al.
demonstrated airway hyperresponiveness to methacholine in vivo and in isolated trachea
in vitro after ovalbumin challenge, however, in PCLS, airway narrowing was slower and the
potency of methacholine was reduced in ovalbumin‐challenged mice compared to
controls (21). Although PCLS come closer to the in vivo situation than single cells or
smooth muscle strips, it may still be possible that mechanical differences exist. It might
therefore be difficult to demonstrate a direct link between airway remodeling and airway
contractility.
Muscarinic receptor stimulation can augment TGF‐β‐induced expression of contractile
proteins and extracellular matrix proteins in airway smooth muscle cells (22‐24).
Moreover, Oenema et al. demonstrated in guinea pig PCLS that methacholine can induce
remodeling via enhanced release of TGF‐β (15). Furthermore, the muscarinic antagonist
tiotropium has been shown to inhibit ovalbumin‐induced TGF‐β release, and also airway
remodeling, in a mouse model of asthma (25). Morevoer, TGF‐β release is decreased in
M3R‐/‐ animals (chapter 3). Methacholine has also been shown to increase TGF‐β and
collagen levels in biopsies of asthma patients (9). Therefore, methacholine‐induced
bronchoconstriction, and subsequent TGF‐β release, may enhance airway remodeling.
Since we have shown here that murine PCLS may not be an adequate model to study
bronchoconstriction‐induced remodeling, future studies to investigate mechanisms
involved in this response and the potential involvement of the M3 receptor are better
performed using guinea pig or human lung slices.
In our study, we found limited contraction towards methacholine in M3R‐/‐ mice, which is
in line with previous studies using lung slices from M3R‐/‐ mice (26). Although the M3
receptor is the dominant receptor in mediating airway smooth muscle contraction (27), it
has been shown that after knock out of the M3 receptor, some contraction can be elicited
via M2 receptors, and that airway contraction is only completely abolished after knock out
of both M2 and M3 receptors (26, 28, 29). Therefore, the observed contraction in the
M3R‐/‐ mice might be mediated via M2 receptors in our study. Clearly, methacholine‐
induced contraction is regulated via at least partially different mechanisms in M3R‐/‐

Lung slices as a model to study remodeling
149
animals than in wild‐type animals. Not only is there a lag time in effect, the contraction in
M3R‐/‐ mice also seemed to be sensitive to lower doses of methacholine compared to
contraction in WT animals. Using airway smooth muscle strips, Kume et al. demonstrated
that contraction in wild‐type animals at low concentrations of methacholine could be
completely inhibited by the M2 antagonist AF‐DX 116 and by pertussis toxin, whereas
contraction at higher concentrations was only partially inhibited by pertussis toxin, and
almost completely mediated via M3 receptors (30). Similar findings were observed using
ileal smooth muscle strips, in which 80% of the contraction was inhibited at low
concentrations of agonist by pertussis toxin, whereas contraction at higher concentrations
was not altered by pertussis toxin and completely mediated via M3 receptors (28). This
might explain the effect of methacholine at low concentrations via M2 receptors after
knock‐out of the M3 receptors compared to wild‐type animals. Future studies are needed
to elucidate the exact mechanisms underlying airway contraction in M3R‐/‐ mice. For
example, experiments under calcium free conditions, or pretreatment with an M2 receptor
selective antagonist or pertussis toxin, to uncouple Gi proteins, might help to explain the
mechanisms involved.
Finally, strong oscillations in airway constriction were observed in M3R‐/‐ mice after a
single dose of methacholine, which was not observed in wild‐type animals. Previously, we
have shown that levels of fibronectin and collagen are lower in the airways of M3R‐/‐ mice
(chapter 3). Decreased levels of these extracellular matrix proteins might result in altered
mechanical stability of the airways and thereby affect the capacity of the airway to
maintain a sustained contraction in M3R‐/‐ mice. Altered mechanical stability might also
explain the enhanced contraction rate in M3R‐/‐ mice compared to wild‐type mice after a
single dose of methacholine. Moreover, this might explain the enhanced airway
constriction in M3R‐/‐ mice compared to wild‐type mice observed at low concentrations of
methacholine, but also at low concentrations of U46619. Increased sensitivity for both
methacholine and U46619 might suggest that this is not a receptor driven process, but is
rather caused by intrinsic differences in the airways of M3R‐/‐ mice compared to wild‐type
mice, involving reduced mechanical stability.
In conclusion, murine PCLS are suitable to study TGF‐β‐induced airway remodeling, but are
most likely not an appropriate model to study bronchoconstriction‐induced remodeling.
This is in contrast to guinea pig PCLS, which are well suited to study this response.

Chapter 7
150
Acknowledgements
We would like to thank the Netherlands Lung Foundation (grant: 3.2.08.014) and
Boehringer Ingelheim (contract number: 43054530) for financial support. Part of the work
has been performed at the University Medical Center Groningen Imaging and Microscopy
Center (UMIC), which is sponsored by NWO‐grants 40‐00506‐98‐9021 and 175‐010‐2009‐
023.

Lung slices as a model to study remodeling
151
References
(1) Jeffery PK. Remodeling in asthma and chronic obstructive lung disease. Am J Respir Crit Care Med
2001;164:S28‐S38.
(2) An SS, Bai TR, Bates JH, Black JL, Brown RH, Brusasco V, Chitano P, Deng L, Dowell M, Eidelman
DH, Fabry B, Fairbank NJ, Ford LE, Fredberg JJ, Gerthoffer WT, Gilbert SH, Gosens R, Gunst SJ,
Halayko AJ, Ingram RH, Irvin CG, James AL, Janssen LJ, King GG, Knight DA, Lauzon AM, Lakser OJ,
Ludwig MS, Lutchen KR, Maksym GN, Martin JG, Mauad T, McParland BE, Mijailovich SM, Mitchell
HW, Mitchell RW, Mitzner W, Murphy TM, Pare PD, Pellegrino R, Sanderson MJ, Schellenberg RR,
Seow CY, Silveira PS, Smith PG, Solway J, Stephens NL, Sterk PJ, Stewart AG, Tang DD, Tepper RS,
Tran T, Wang L. Airway smooth muscle dynamics: a common pathway of airway obstruction in
asthma. Eur Respir J 2007;29:834‐860.
(3) Pare PD, Roberts CR, Bai TR, Wiggs BJ. The functional consequences of airway remodeling in
asthma. Monaldi Arch Chest Dis 1997;52:589‐596.
(4) Lemanske RF,Jr, Busse WW. Asthma: clinical expression and molecular mechanisms. J Allergy Clin
Immunol 2010;125:S95‐102.
(5) Minshall EM, Leung DY, Martin RJ, Song YL, Cameron L, Ernst P, Hamid Q. Eosinophil‐associated
TGF‐beta1 mRNA expression and airways fibrosis in bronchial asthma. Am J Respir Cell Mol Biol
1997;17:326‐333.
(6) Benayoun L, Druilhe A, Dombret MC, Aubier M, Pretolani M. Airway structural alterations
selectively associated with severe asthma. Am J Respir Crit Care Med 2003;167:1360‐1368.
(7) Guilbert TW, Morgan WJ, Zeiger RS, Mauger DT, Boehmer SJ, Szefler SJ, Bacharier LB, Lemanske
RF,Jr, Strunk RC, Allen DB, Bloomberg GR, Heldt G, Krawiec M, Larsen G, Liu AH, Chinchilli VM,
Sorkness CA, Taussig LM, Martinez FD. Long‐term inhaled corticosteroids in preschool children at
high risk for asthma. N Engl J Med 2006;354:1985‐1997.
(8) Murray CS, Woodcock A, Langley SJ, Morris J, Custovic A, IFWIN study team. Secondary
prevention of asthma by the use of Inhaled Fluticasone propionate in Wheezy INfants (IFWIN):
double‐blind, randomised, controlled study. Lancet 2006;368:754‐762.
(9) Grainge CL, Lau LC, Ward JA, Dulay V, Lahiff G, Wilson S, Holgate S, Davies DE, Howarth PH. Effect
of bronchoconstriction on airway remodeling in asthma. N Engl J Med 2011;364:2006‐2015.
(10) Ressler B, Lee RT, Randell SH, Drazen JM, Kamm RD. Molecular responses of rat tracheal
epithelial cells to transmembrane pressure. Am J Physiol Lung Cell Mol Physiol 2000;278:L1264‐72.
(11) Choe MM, Sporn PH, Swartz MA. An in vitro airway wall model of remodeling. Am J Physiol Lung
Cell Mol Physiol 2003;285:L427‐33.
(12) Swartz MA, Tschumperlin DJ, Kamm RD, Drazen JM. Mechanical stress is communicated
between different cell types to elicit matrix remodeling. Proc Natl Acad Sci U S A 2001;98:6180‐
6185.
(13) Tatler AL, John AE, Jolly L, Habgood A, Porte J, Brightling C, Knox AJ, Pang L, Sheppard D, Huang
X, Jenkins G. Integrin alphavbeta5‐mediated TGF‐beta activation by airway smooth muscle cells in
asthma. J Immunol 2011;187:6094‐6107.

Chapter 7
152
(14) Wahl M, Eddinger TJ, Hai CM. Sinusoidal length oscillation‐ and receptor‐mediated mRNA
expression of myosin isoforms and alpha‐SM actin in airway smooth muscle. Am J Physiol Cell
Physiol 2004;287:C1697‐708.
(15) Oenema TA, Maarsingh H, Smit M, Groothuis GM, Meurs H, Gosens R. Bronchoconstriction
Induces TGF‐beta Release and Airway Remodelling in Guinea Pig Lung Slices. PLoS One
2013;8:e65580.
(16) Yamada M, Miyakawa T, Duttaroy A, Yamanaka A, Moriguchi T, Makita R, Ogawa M, Chou CJ,
Xia B, Crawley JN, Felder CC, Deng CX, Wess J. Mice lacking the M3 muscarinic acetylcholine receptor
are hypophagic and lean. Nature 2001;410:207‐212.
(17) Gosens R, Baarsma HA, Heijink IH, Oenema TA, Halayko AJ, Meurs H, Schmidt M. De novo
synthesis of {beta}‐catenin via H‐Ras and MEK regulates airway smooth muscle growth. FASEB J
2010;24:757‐768.
(18) Rosner SR, Ram‐Mohan S, Paez‐Cortez JR, Lavoie TL, Dowell ML, Yuan L, Ai X, Fine A, Aird WC,
Solway J, Fredberg JJ, Krishnan R. Airway contractility in the precision‐cut lung slice after
cryopreservation. Am J Respir Cell Mol Biol 2014;50:876‐881.
(19) Halwani R, Al‐Muhsen S, Al‐Jahdali H, Hamid Q. Role of transforming growth factor‐beta in
airway remodeling in asthma. Am J Respir Cell Mol Biol 2011;44:127‐133.
(20) Chew AD, Hirota JA, Ellis R, Wattie J, Inman MD, Janssen LJ. Effects of allergen on airway
narrowing dynamics as assessed by lung‐slice technique. Eur Respir J 2008;31:532‐538.
(21) Donovan C, Royce SG, Esposito J, Tran J, Ibrahim ZA, Tang ML, Bailey S, Bourke JE. Differential
effects of allergen challenge on large and small airway reactivity in mice. PLoS One 2013;8:e74101.
(22) Goldsmith AM, Bentley JK, Zhou L, Jia Y, Bitar KN, Fingar DC, Hershenson MB. Transforming
growth factor‐beta induces airway smooth muscle hypertrophy. Am J Respir Cell Mol Biol
2006;34:247‐254.
(23) Oenema TA, Mensink G, Smedinga L, Halayko AJ, Zaagsma J, Meurs H, Gosens R, Dekkers BG.
Cross‐talk between transforming growth factor‐beta(1) and muscarinic M(2) receptors augments
airway smooth muscle proliferation. Am J Respir Cell Mol Biol 2013;49:18‐27.
(24) Oenema TA, Smit M, Smedinga L, Racke K, Halayko AJ, Meurs H, Gosens R. Muscarinic receptor
stimulation augments TGF‐beta1‐induced contractile protein expression by airway smooth muscle
cells. Am J Physiol Lung Cell Mol Physiol 2012;303:L589‐97.
(25) Ohta S, Oda N, Yokoe T, Tanaka A, Yamamoto Y, Watanabe Y, Minoguchi K, Ohnishi T, Hirose T,
Nagase H, Ohta K, Adachi M. Effect of tiotropium bromide on airway inflammation and remodelling
in a mouse model of asthma. Clin Exp Allergy 2010;40:1266‐1275.
(26) Struckmann N, Schwering S, Wiegand S, Gschnell A, Yamada M, Kummer W, Wess J,
Haberberger RV. Role of muscarinic receptor subtypes in the constriction of peripheral airways:
studies on receptor‐deficient mice. Mol Pharmacol 2003;64:1444‐1451.
(27) Roffel AF, Elzinga CR, Van Amsterdam RG, De Zeeuw RA, Zaagsma J. Muscarinic M2 receptors in
bovine tracheal smooth muscle: discrepancies between binding and function. Eur J Pharmacol
1988;153:73‐82.
(28) Unno T, Matsuyama H, Sakamoto T, Uchiyama M, Izumi Y, Okamoto H, Yamada M, Wess J,
Komori S. M(2) and M(3) muscarinic receptor‐mediated contractions in longitudinal smooth muscle
of the ileum studied with receptor knockout mice. Br J Pharmacol 2005;146:98‐108.

Lung slices as a model to study remodeling
153
(29) Ehlert FJ. Pharmacological analysis of the contractile role of M2 and M3 muscarinic receptors in
smooth muscle. Receptors Channels 2003;9:261‐277.
(30) Kume H, Mikawa K, Takagi K, Kotlikoff MI. Role of G proteins and KCa channels in the muscarinic
and beta‐adrenergic regulation of airway smooth muscle. Am J Physiol 1995;268:L221‐9.
