u.s.a. bacteriocin antibiotic resistance plasmids ... · garden soil and identified as b. cereus or...
TRANSCRIPT

JOURNAL OF BACTERIOLOGY, Feb. 1978, p. 897-9030021-9193/78/0133-0897$02.00/0Copyright X 1978 American Society for Microbiology
Vol. 133, No. 2
Printed in U.S.A.
Bacteriocin and Antibiotic Resistance Plasmids in Bacilluscereus and Bacillus subtilis
K. BERNHARD, H. SCHREMPF, AND W. GOEBEL*Institut fur Genetik und Mikrobiologie, Universitat Wtrzburg, Rontgenring 11, 8700 Wiurzburg, Germany
Received for publication 29 August 1977
A number of plasmids have been isolated as covalently closed circular DNAsfrom strains of Bacillus cereus and B. subtilis. From 12 out of 15 strains of B.cereus, plasmids could be isolated. Most of the B. cereus strains contained twoor more plasmids. Their molecular weights ranged from 1.6 x 106 to 105 x 106.Bacteriocin production could be attributed to a 45 x 106-dalton plasmid (pBC7)from B. cereus DSM 336, and tetracycline resistance to a 2.8 x 106 plasmid(pBC16) from B. cereus GP7. Two streptomycin-resistant strains of B. subtilisharbored plasmids of 5.2 x 106 and 9 x 106, respectively, which were, however,not correlated with the antibiotic resistance. The plasmid carrying resistance totetracycline, pBC16, which was originally isolated from B. cereus, could besubsequently transformed in B. subtilis, where it is stably maintained.
Plasmids have been identified in many bac-teria (4, 7, 8; K. N. Timmis, S. N. Cohen, andF. Cabello, Prog. Med. Subcell. Biol., in press).A large variety of specific biochemical functions,such as fertility, resistance to antimicrobialdrugs, production of bacteriocins, production oftoxins, etc., have been attributed to these geneticelements.Most of the described plasmids in bacilli (11,
12, 17) are cryptic elements lacking geneticmarkers and are unsuitable for selection oftrans-formed colonies. We have undertaken a searchfor plasmids in Bacillus species, mainly Bacilluscereus and B. subtilis, to characterize their prop-erties and develop their potential use as vectorsfor gene cloning.
Since most Bacillus species populate the sameecosystems as Streptomyces and other producersof antibiotics, they may have acquired resistanceto the antibiotics being produced under naturalconditions.
Therefore plasmid-inherited functions, espe-cially antibiotic resistance, were examined andinvestigated in Bacillus species.
MATERIALS AND METHODSMedia and chemicals. PY medium contained 1%
peptone from casein, 0.5% yeast extract, and 0.5%NaCl. The pH value was adjusted to 7.0 by NaOH.HGP medium was PY medium supplemented by 0.5%glucose. PYS medium was PY medium containing 20%(wt/vol) sucrose. PY agar contained 1.5% agar. TGagar was a tris(hydroxymethyl)aminomethane (Tris)-buffered minimal medium (3).TES buffer contained 30 mM Tris, 50 mM NaCl,
and 5 mM ethylenediaminetetraacetic acid (pH 8.0).All chemicals, if not otherwise stated, were from
Merck, Germany. Ampicillin, tetracycline, and chlor-amphenicol were a gift from Bayer, Germany. Kana-mycin sulfate was purchased from Boehringer, Ger-many, and bacitracin from Serva, Germany. Filterdisks containing antibiotics were obtained from Pro-ductions Institut Pasteur, France. [methyl-3H]thymi-dine was obtained from the Radiochemical Centre,Amersham, U.K. Sodium dodecyl sulfate (SDS),ethidium bromide, acridine orange, polyethylene gly-col type 6000, pancreatic ribonuclease, and Pronase Pfrom Streptomyces griseus were obtained from Serva,Germany.
Isolation of B. cereus and B. subtilis from soilsamples. Sporeforming bacteria were enriched fromgarden soil and identified as B. cereus or B. subtilisby standard methods. The purified strains were thentested for antibiotic resistance and antibiotic or bac-teriocin production.
Tests for antibiotic resistance. Resistance toampicillin, streptomycin (Sm%), kanamycin (KMr), tet-racycline (Tcr), chloramphenicol, and bacitracin wastested by streaking the isolates on PY agar platescontaining one of these antibiotics. The final concen-trations were 25 jig of tetracycline per ml, 50 ug ofbacitracin per ml, and 100 ,ug/ml each for the otherantibiotics. Resistances to polymyxin, bacitracin, col-istin, oleandomycin, cephaloridin, erythromycin, gen-tamicin, neomycin, novobiocin, and spiromycin weretested with filter disks containing these antibiotics.
Tests for antibiotic and bacteriocin produc-tion. HGP plates were inoculated with strains to betested. A 0.1-ml sample of a stationary-phase cultureof the indicator strain was plated on a second HGPplate to give a bacterial lawn. Both plates were incu-bated at 30°C overnight. Then the lawn of indicatorbacteria was transferred to the plate inoculated withthe strain to be tested by replica plating. This platewas incubated for another 24 h. Formation of a clearzone in the bacterial lawn around the strain to betested indicated the production of a killing substance
897
on June 4, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

898 BERNHARD, SCHREMPF, AND GOEBEL
(bacteriocin or antibiotic). Indicator bacteria usedwere Micrococcus flavus, B. megaterium, B. thurin-giensis, B. cereus, and B. subtilis.
Isolation of plasmid DNA. Overnight cultures ofB. cereus in PYS medium were diluted sixfold withPYS medium and labeled by adding 1.5 /Ci of [methyl-3H]thymidine per ml. The cultures were incubated at30°C for another 6 h. Cleared lysates from thesecultures were prepared by the SDS-NaCl procedure(6) and subsequently centrifuged in cesium chloride-ethidium bromide gradients as described (15). Forpreparation of larger amounts of plasmid DNA, cellsfrom 350-ml cultures were lysed. Cleared lysates were
mixed with polyethylene glycol (50% stock solution)to a final concentration of 10% and kept on ice for atleast 3 h (9). Then the lysates were centrifuged in aBeckman JA-20 rotor at 8,000 rpm for 10 min. Thepellet was suspended in 5 ml of TES buffer. Afteraddition of 0.5 ml of pancreatic ribonuclease (500,tg/ml), the suspension was kept at 60°C for 30 min.
Preparation of lysates for agarose gel electro-phoresis. Cultures (30 ml) were harvested and di-gested with lysozyme. The protoplasts were rapidlyadded to 2.5 ml of lysis buffer (2.0% SDS, 50 mMpotassium ethylenediaminetetraacetic acid, 30 mMTris, 50 mM NaCI) at 60°C. When the cells were
lysed, 500 pl of Pronase P (1 mg/ml) was added, andthe lysate was incubated at 60°C for 3 h. The lysatewas kept on ice overnight and then cleared by centrif-ugation. The cleared supernatant was treated with250 ,ug of pancreatic ribonuclease at 60°C for 30 minand extracted twice with 50 mM Tris-saturatedphenol. LiCl was added to a final concentration of 1M, and the DNA was precipitated by adding 15 ml ofice-cold ethanol. The mixture was kept at -20°Covernight and centrifuged at 8,000 rpm for 10 min ina Beckman JA-20 rotor. The pelleted DNA was sus-
pended in 50 pl of TES buffer. Portions of 10 to 20 td
were used for agarose gel electrophoresis. Electropho-resis was performed on 0.7% agarose slab gels in Tris-phosphate buffer (36 mM Tris, 30 mM NaH2PO4, 10mM ethylenediaminetetraacetic acid, pH 7.5) (13).Transformation ofB. subtilis 168 trp with plas-
mid DNA and chromosomal DNA. Cells were madecompetent and transformed with chromosomal DNAby the method of Laird as described by Cahn andFox (3). Transformation with plasmid DNA was car-
ried out by mixing this DNA with competent cells ina total volume of 1 ml of Laird medium II. Afterincubation at 30°C for 30 min, double-concentratedHGP medium (1 ml) was added. These assays wereincubated at 30°C for another 90 min to allow expres-sion of plasmid-coded information. Then samples were
plated on PY agar containing antibiotics.Curing of antibiotic-resistant or antibi-
otic/bacteriocin-producing B. cereus and B. sub-tilis. PY medium (20 ml) containing the curing re-
agent was inoculated with 1% of a stationary cultureand incubated overnight at 30°C. The culture hadthen reached a cell density of 4 x 10' cells per ml.Appropriate dilutions of the culture were preparedand plated on PY agar, then incubated overnight at30°C. Antibiotic resistance or killing activity was thentested by replica plating. In curing experiments per-formed with B. cereus, the curing reagents were
used in the following concentrations: ethidium bro-mide and acridine orange at 0.02 and 0.04 yg/ml eachand SDS at 0.001% and 0.002%. With B. subtilis,ethidium bromide and acridine orange were used atconcentrations of 0.01 and 0.02 jig/ml, respectively.
Electron microscopy. Spreading of plasmid DNAfor electron microscopy was performed as describedby Ohtsubo et al. (14).Assays for digestion of plasmid DNA with re-
striction enzymes. The restriction enzymes were agenerous gift of H. Mayer, Gesellschaft fur Biotech-nologische Forschung Stockheim, Germany. Cleavageof plasmid DNA was performed as described (16).Calculation of the plasmid copy number. The
plasmid copy number was calculated from the amountof 3H radioactivity present in the covalently closedcircular (CCC) band of the cesium chloride-ethidiumbromide gradients compared with the amount of 'Hradioactivity in total DNA. The molecular weight ofthe chromosome of B. subtilis was taken as 2 x 109(10). It is assumed that the chromosome of B. cereushas a similar size.
RESULTSIsolation and characterization ofBacillus
strains. Bacillus strains were isolated by stan-dard procedures (2) from local soil samples.After purification, all strains except GP5 andGP13 were identified as B. cereus. Strains GP5and GP13 were identified as B. subtilis. In ad-dition, several B. cereus strains from the GermanType Culture Collection (DSM) were includedin the investigation.The strains were tested for resistance to the
following antibiotics: ampicillin, kanamycin,streptomycin, chloramphenicol, tetracycline,bacitracin, colistin, erythromycin, gentamicin,oleandomycin, polymyxin, cephaloridin, neo-mycin, novobiocin, and spiromycin.
All B. cereus strains were resistant to highlevels (up to 1 mg/ml) of ampicillin, colistin,and polymyxin, and were hemolytic. These prop-erties are therefore most likely chromosomallydetermined in B. cereus. In contrast, resistanceto kanamycin, tetracycline, bacitracin, or ce-phaloridin was observed in only a few strains.Two B. subtilis strains were isolated that wereresistant to streptomycin. Antibiotic resistancesother than those described above were not foundin the B. subtilis and B. cereus strains tested.Two B. cereus strains (30 and DSM 336) pro-duced a substance with killing activity for theother B. cereus strains. Since this activity seemsto be species specific, or at least restricted toclosely related species like B. thuringiensis andB. megaterium, it probably represents a bacte-nocin.
Strains DSM 31, GP1, and GP2 producedantibiotic substances of unknown structure ac-tive against M. flavus but not against B. cereus.
J. BACTERIOL.
on June 4, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

VOL. 133, 1978
B. cereus DSM 351 produced a killing substancethat was strongly active against B. thuringiensisand B. megaterium, but only weakly activeagainst some strains of B. cereus and inactiveagainst other strains of B. cereus.Isolation of CCC plasmid DNA from ba-
cilli. The B. cereus and B. subtilis strains de-scribed were analyzed by preparing cleared ly-sates that were subsequently centrifuged in ce-sium chloride-ethidium bromide gradients.Strains that showed plasmid DNA were culti-vated in the presence of [methyl-3H]thymidineto label the DNA.The labeled CCC DNA thus obtained was
further analyzed on neutral and alkaline sucrosegradients together with 32P-labeled standardCCC DNAs (ColEl, 4.2 x 106 daltons, andRldrd-19, 65 x 106 daltons) to determine themolecular weights of the plasmid species presentin the CCC fraction of the cesium chloride-ethid-ium bromide gradient. In addition, electron mi-croscopy studies were performed (Fig. 1) withthe plasmids oftwo strains, which were of specialinterest to further characterize their molecular
PLASMIDS OF BACILLUS SPECIES 899
properties. Table 1 summarizes the results ob-tained from these analyses. A large variety ofplasmids were found with molecular weightsranging from 1.6 x 106 to 105 x 106. Some ofthe strains contained only a single plasmid, butmost of them harbored two or even more plas-mids, one of which was often a rather smallextrachromosomal element (Table 1). The moststriking result was the frequent occurrence ofplasmids in B. cereus strains, since 12 out of 15strains contained plasmids.Correlation of biochemical functions
with plasmids. All B. cereus strains that hada non-common antibiotic resistance or were bac-teriocinogenic harbored plasmids. Strain DSM336 could be cured of its bacteriocinogenic activ-ity (kill+) by treatment with ethidium bromideor SDS (curing rate was 2% with ethidium bro-mide and 1.5% with SDS). None of the curedstrains harbored CCC plasmid DNA, suggestingthat this function is determined by the 45 x106-dalton plasmid of strain DSM 336. Strain30, another kill' strain harboring a 37 x 106-dalton plasmid, yielded also kill- colonies upon.
I p J
'.\
(";
fi
,' i, t. J
,/ \ .t
* s.s \
_ ,f s
_, v o_
A s.;tv :
.
(.Iijml
I-,m
FIG. 1. Electron micrographs of open circular molecules of plasmid DNA isolated from B. cereus GP7and B. subtilis GP5. (A) The small molecules represent pBC16 DNA; the larger molecules are referencemolecules of ColEl DNA (6.34 kilobases [14]). (B) Open circular molecules (monomers and oligomers) ofpBSI isolated from B. subtilis GP5.
on June 4, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

900 BERNHARD, SCHREMPF, AND GOEBEL
TABLE 1. Plasmids in B. cereus and B. subtilis
Strain Unusual property expressed Plasniids Mol wt (x 106) Copy no. Plasmid-corre-by the strain lated functions
B. cereus 21 Resistance to bacitracin pBC4 33 3-5pBC5 6.4 7-10
B. cereus 30 Production of bacteriocin pBC3, 37 1-2B. cereus 35 pBC1 105 1-2
pBC2 13 10-12B. cereus DSM 316 Production of antibiotic
substanceB. cereus DSM 3186 pBC6 8 7-8B. cereus DSM 3366 Production of bacteriocin pBC7 45 1-2 Production of
bacteriocinB. cereus DSM 3516 Production of antibiotic
substanceB. cereus DSM 3606 Resistance to cephaloridin pBC8 12.5 12B. cereus DSM 487b pBC9 50 1-2
pBC10 7.6 8B. cereus P1 pBCll 35 1-2
pBC12 5.3 8-10B. cereus GP1 Production of antibiotic
substanceB. cereus GP2 Production of antibiotic pBC13 59 1-2
substance pBC14 21 2-3B. cereus GP7 Tcr pBC15 23 1-2
pBC16 2.77 ± 0.12c 20 TcB. cereus GP11 Kni pBC17 40 1-2
pBC18 8.2 3-4B. cereus GP17 pBC19 32 1-2
pBC20 8 3-4pBC21 1.6 12-13
B. subtilis 168B. subtilis GP5 Smr pBS1 5.26 ± 0.13c 5-7B. subtilis GP6 Smr pBS2 9 1-2a Molecular weight of the plasmids was determined by sucrose gradient centrifugation.b These strains were obtained from the German Type Culture Collection (Deutsche Sammlung von Mikroor-
ganismen [DSM]), Gottingen, Germany.e As measured by electron microscopy.
treatment with ethidium bromide. However, allkill variants still contained CCC DNA, whichwas not further analyzed.
B. cereus strain GP7 is resistant to high dosesof tetracycline (up to 200 pug/ml) and carries twoplasmids of 23 x 106 and 2.9 x 106 daltons.Treatment with ethidium bromide led to theisolation of one Tc8 variant. Analysis of CCCDNA revealed that the sensitive strain had lostthe small plasmid pBC16 but retained the largerone. This suggests that pBC16 carries the infor-mation for Tcr. The kanamycin-resistant B. cer-eus strain GPll could be cured of antibioticresistance by treatment with SDS and ethidiumbromide. Streptomycin-sensitive colonies wereobtained from the two Smr B. subtilis strains,GP5 and GP13, which harbor rather small plas-mids of 4.6 x 1i6 and 9 x 106 daltons, respec-tively. However, these "cured" strains did notlose CCC DNA, since plasmid DNA could berecovered from cleared lysates after cesium chlo-ride-ethidium bromide and sucrose gradient cen-trifugation.
Transformation of plasmids pBC16 andpBS1 in B. subtilis 168. Competence of cellswas tested by transformation with chromosomalDNA isolated from a Trp+ variant of B. subtilis168, and selection was made for Trp+ transforn-ants. With pBC16 DNA, Tcr colonies were ob-tained at a frequency of about 10-l, whereasTrp+ transformants were found at a frequencyof about 10-3, as shown in Table 2. The level ofTcr in these B. subtilis transformants wasequally as high (200 ,ug/ml) as that of the paren-tal B. cereus strain GP7. Analysis of CCC DNAfrom the Tcr transformants by agarose gel elec-trophoresis (Fig. 2) showed that they all con-tained DNA having the same size and restrictionenzyme digest pattern as pBC16 isolated fromthe original B. cereus strain (Fig. 3). PlasmidpBC16 was rather stable in the new host. Uponextended growth for more than 100 generations,less than 0.1% tetracycline-sensitive colonieswere obtained. Competent cells of B. subtilis168 treated with equal amounts of pBC16 DNAfrom either B. subtilis or B. cereus yielded sim-
J. BACTERIOL.
on June 4, 2019 by guesthttp://jb.asm
.org/D
ownloaded from
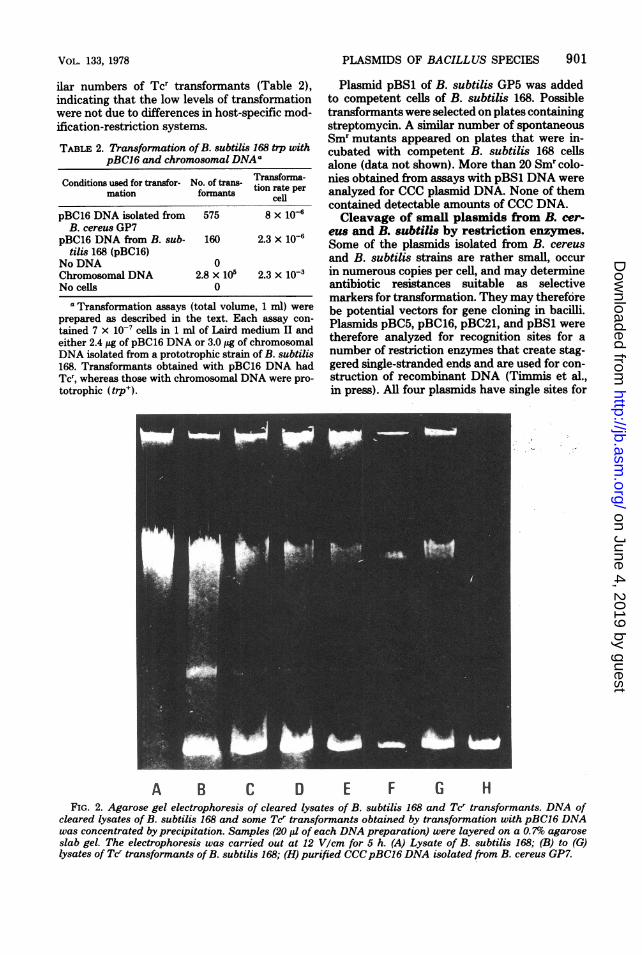
PLASMIDS OF BACILLUS SPECIES 901
ilar numbers of Tcr transformants (Table 2),indicating that the low levels of transformationwere not due to differences in host-specific mod-ification-restriction systems.
TABLE 2. Transformation ofB. subtilis 168 trp withpBC16 and chromosomal DNAa
Conditions used for transfor- No. of trans- Tranformamation formants tion rate per
cell
pBC16 DNA isolated from 575 8 x 10-6B. cereus GP7
pBC16 DNA from B. sub- 160 2.3 x 10-6tilis 168 (pBC16)
No DNA 0Chromosomal DNA 2.8 x 105 2.3 x 10-3No cells 0
a Transformation assays (total volume, 1 ml) wereprepared as described in the text. Each assay con-tained 7 x 1O-7 cells in 1 ml of Laird medium II andeither 2.4 /g of pBC16 DNA or 3.0 ug of chromosomalDNA isolated from a prototrophic strain of B. subtilis168. Transformants obtained with pBC16 DNA hadTcr, whereas those with chromosomal DNA were pro-totrophic (trp').
Plasmid pBS1 of B. subtilis GP5 was addedto competent cells of B. subtilis 168. Possibletransformants were selected on plates containingstreptomycin. A similar number of spontaneousSmr mutants appeared on plates that were in-cubated with competent B. subtilis 168 cellsalone (data not shown). More than 20 Smr colo-nies obtained from assays with pBS1 DNA wereanalyzed for CCC plasmid DNA. None of themcontained detectable amounts of CCC DNA.Cleavage of small plasmids from B. cer-
eus and B. subtilis by restriction enzymes.Some of the plasmids isolated from B. cereusand B. subtilis strains are rather small, occurin numerous copies per cell, and may determineantibiotic resistances suitable as selectivemarkers for transformation. They may thereforebe potential vectors for gene cloning in bacilli.Plasmids pBC5, pBC16, pBC21, and pBS1 weretherefore analyzed for recognition sites for anumber of restriction enzymes that create stag-gered single-stranded ends and are used for con-struction of recombinant DNA (Timmis et al.,in press). All four plasmids have single sites for
A B C D E F G HFIG. 2. Agarose gel electrophoresis of cleared lysates of B. subtilis 168 and Tcr transformants. DNA of
cleared lysates of B. subtilis 168 and some Tcr transformants obtained by transformation with pBC16 DNAwas concentrated by precipitation. Samples (20 p1 of each DNA preparation) were layered on a 0. 7%o agaroseslab gel. The electrophoresis was carried out at 12 V/cm for 5 h. (A) Lysate of B. subtilis 168; (B) to (G)lysates of Tcr transformants ofB. subtilis 168; (H) purified CCCpBC16 DNA isolated from B. cereus GP7.
VOL. 133, 1978
on June 4, 2019 by guesthttp://jb.asm
.org/D
ownloaded from
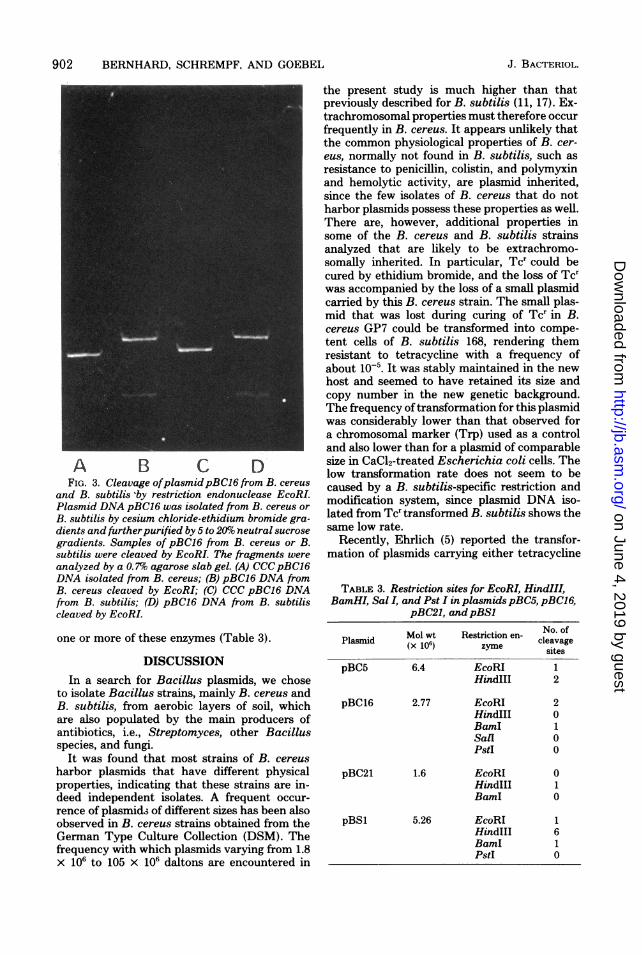
902 BERNHARD, SCHREMPF, AND GOEBEL
A B C DFIG. 3. Cleavage ofplasmidpBC16 from B. cereus
and B. subtilis 'by restriction endonuclease EcoRLPlasmid DNA pBC16 was isolated from B. cereus or
B. subtilis by cesium chloride-ethidium bromide gra-dients and furtherpurified by 5 to 20% neutral sucrose
gradients. Samples of pBC16 from B. cereus or B.subtilis were cleaved by EcoRL The fragments were
analyzed by a 0.7%o agarose slab gel. (A) CCCpBC16DNA isolated from B. cereus; (B) pBC16 DNA fromB. cereus cleaved by EcoRI; (C) CCC pBC16 DNAfrom B. subtilis; (D) pBC16 DNA from B. subtiliscleaved by EcoRI.
one or more of these enzymes (Table 3).
DISCUSSIONIn a search for Bacillus plasmids, we chose
to isolate Bacillus strains, mainly B. cereus andB. subtilis, from aerobic layers of soil, whichare also populated by the main producers ofantibiotics, i.e., Streptomyces, other Bacillusspecies, and fungi.
It was found that most strains of B. cereusharbor plasmids that have different physicalproperties, indicating that these strains are in-deed independent isolates. A frequent occur-
rence of plasmid.3 of different sizes has been alsoobserved in B. cereus strains obtained from theGerman Type Culture Collection (DSM). Thefrequency with which plasmids varying from 1.8X 106 to 105 x 106 daltons are encountered in
the present study is much higher than thatpreviously described for B. subtilis (11, 17). Ex-trachromosomal properties must therefore occurfrequently in B. cereus. It appears unlikely thatthe common physiological properties of B. cer-
eus, normally not found in B. subtilis, such as
resistance to penicillin, colistin, and polymyxinand hemolytic activity, are plasmid inherited,since the few isolates of B. cereus that do notharbor plasmids possess these properties as well.There are, however, additional properties insome of the B. cereus and B. subtilis strainsanalyzed that are likely to be extrachromo-somally inherited. In particular, Tcr could becured by ethidium bromide, and the loss of Tcrwas accompanied by the loss of a small plasmidcarried by this B. cereus strain. The small plas-mid that was lost during curing of Tcr in B.cereus GP7 could be transformed into compe-tent cells of B. subtilis 168, rendering themresistant to tetracycline with a frequency ofabout 10-5. It was stably maintained in the newhost and seemed to have retained its size andcopy number in the new genetic background.The frequency oftransformation for this plasmidwas considerably lower than that observed fora chromosomal marker (Trp) used as a controland also lower than for a plasmid of comparablesize in CaCl2-treated Escherichia coli cells. Thelow transformation rate does not seem to becaused by a B. subtilis-specific restriction andmodification system, since plasmid DNA iso-lated from Tcr transformed B. subtilis shows thesame low rate.
Recently, Ehrlich (5) reported the transfor-mation of plasmids carrying either tetracycline
TABLE 3. Restriction sites for EcoRI, HindIII,BamHI, Sal I, and Pst I in plasmids pBC5, pBC16,
pBC21, andpBS1
Mol wt Restriction en-No. of
Plasmid (x wt6) zyme cleavagesites
pBC5 6.4 EcoRI 1HindIII 2
pBC16 2.77 EcoRI 2HindIII 0BamI 1Sall 0PstI 0
pBC21 1.6 EcoRI 0HindIII 1BamI 0
pBS1 5.26 EcoRI 1HindlIl 6BamI 1PstI 0
J. BACTERIOL.
on June 4, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

PLASMIDS OF BACILLUS SPECIES
or chloramphenicol resistance from Staphylo-coccus aureus into the same B. subtilis host.The size of the Tcr S. aureus plasmid is similarto the tetracycline plasmid pBC16 found herein B. cereus. However, the recognition sites forvarious restriction enzymes seem to be different.Whereas pBC16 has one BamHI site, two EcoRIsites, and no HindIII site, the Tcr plasmid pT127from S. aureus has three HindIII sites (5). Alsothe described transformation rate is higher forthe B. cereus plasmid. The Tcr determined bypBC16 is relatively high, whereas pT127 rendersthe B. subtilis cells resistant only to low concen-trations of tetracycline. The properties of bothplasmids make them potential vectors for genecloning in B. subtilis.
ACKNOWLEDGMENTS
This work was supported by a grant from the DeutscheFroschungsgemeinschaft (SFB 105-Al).
H. Mayer is thanked for generous gifts of restriction en-zymes. We acknowledge the invaluable training by E. Ohtsuboin analysis of DNA by electron microscopy. We thank W.Oertel for many helpful discussions, J. Launer for technicalhelp with the electron microscope, and J. Collins for readingthe manuscript.
LITERATURE CITED1. Bouanchaud, D. H., M. R. Scavizzi, and Y. Chabbert.
1969. Elimination by ethidium bromide of antibioticresistance in enterobacteria and staphylococci. J. Gen.Microbiol. 54:417-425.
2. Buchanan, R. E., and N. E. Gibbons (ed.). 1974. Ber-gey's manual of determinative bacteriology, 8th ed. TheWilliams and Wilkins Co., Baltimore.
3. Cahn, F. H., and M. S. Fox. 1968. Fractionation oftransformable bacteria from competent cultures of Ba-cillus subtilis on renografin gradients. J. Bacteriol.95:867-875.
4. Cohen, S. N. 1976. Transposable elements and plasmidevolution. Nature (London) 263:731-738.
5. Ehrlich, S. D. 1977. Replication and expression of plas-mids from Staphylococcus aureus in Bacillus subtilis.Proc. Natl. Acad. Sci. U.S.A. 74:1680-1682.
6. Guerry, P., D. J. LeBlanc, and S. Falkow. 1973. Gen-eral method for the isolation of plasmid deoxyribonu-cleic acid. J. Bacteriol. 116:1064-1066.
7. Helinski, D. R., and D. B. Clewell. 1971. Circular DNA.Annu. Rev. Biochem. 40:899-941.
8. Helinski, D. R. 1973. Plasmid determined resistance toantibiotics: molecular properties of R-factors. Annu.Rev. Microbiol. 27:437-470.
9. Humphreys, G. O., G. A. Willshaw, and E. S. Ander-son. 1975. A simple method for the preparation of largequantities of pure plasmid DNA. Biochim. Biophys.Acta 383:457-463.
10. Kavenoff, R. 1972. Characterization of the B. subtilisW23 genome by sedimentation. J. Mol. Biol. 72:801-806.
11. Lovett, P. S., and M. G. Bramucci. 1975. Plasmid de-oxyribonucleic acid in Bacillus subtilis and Bacilluspumilus. J. Bacteriol. 124:484-490.
12. Lovett, P. S., E. J. Duvall, and K. M. Keggins. 1976.Bacillus pumilus plasmid pPL10: properties and inser-tion into Bacillus subtilis 168 by transformation. J.Bacteriol. 127:817-828.
13. Meyers, J. A., D. Sanchez, L. P. Elwell, and S. Fal-kow. 1976. Simple agarose gel electrophoretic methodfor the identification and characterization of plasmiddeoxyribonucleic acid. J. Bacteriol. 127:1529- 1537.
14. Ohtsubo, E., R. C. Deonier, H. T. Lee, and N. David-son. 1974. Electron microscope heteroduplex studies ofsequence repeats among plasmids of Escherichia coli.V. Deletion mutants of F'14. J. Mol. Biol. 89:565-650.
15. Radloff, R., W. Bauer, and J. Vinograd. 1967. A dyebuoyant density method for the detection and isolationof closed circular duplex DNA: the closed circular DNAin HeLa cells. Proc. Natl. Acad. Sci. U.S.A. 57:1514-1521.
16. Schrempf, H., and W. Goebel. 1977. Characterizationof a plasmid from Streptomyces coelicolor A3(2). J.Bacteriol. 131:251-258.
17. Tanaka, T., M. Kuroda, and K. Sakaguchi. 1977. Iso-lation and characterization of four plasmids from Ba-cillus subtilis. J. Bacteriol. 129:1487-1494.
903VOL. 133, 1978
on June 4, 2019 by guesthttp://jb.asm
.org/D
ownloaded from