variations in cerebral organization as a function of … · variations in cerebral organization as...
TRANSCRIPT

See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/22471213
Variations in cerebral organization as a function of
handedness, hand posture in writing, and sex
Article in Journal of Experimental Psychology General · July 1978
DOI: 10.1037/0096-3445.107.2.119 · Source: PubMed
CITATIONS
185READS
333
2 authors, including:
Some of the authors of this publication are also working on these related projects:
Gender Differences View project
Interhemispheric Interaction View project
Jerre Levy
University of Chicago
83 PUBLICATIONS 4,704 CITATIONS
SEE PROFILE
All content following this page was uploaded by Jerre Levy on 13 July 2014.
The user has requested enhancement of the downloaded file.

Journal of Experimental Psychology: General1978, Vol. 107, No. Z, 119-144
Variations in Cerebral Organization as a Function ofHandedness, Hand Posture in Writing, and Sex
Jerre Levy and Marylou ReidUniversity of Pennsylvania
SUMMARY
During' the past century, it has become increasingly apparent that there is agreat deal of variation in the direction and degree of cerebral lateralization, aplurality of people having language and related functions strongly specialized tothe left hemisphere and visuospatial functions strongly specialized to the right, withsubstantial minorities manifesting various deviations from this pattern. In partic-ular, in 35%-50% of sinistrals and 1%-10% of dextrals, the right hemisphere isspecialized for linguistic skills, and in some unknown fraction of the two handed-ness groups, verbal and/or spatial abilities are, to varying extents, bilateralized.Levy (1973) suggested that the hand posture adopted during writing might be anindex of the lateral relationship between the dominant writing hand and the lan-guage hemisphere, a normal posture indicating' contralateral language specializa-tion, and an inverted posture indicating ipsilatcral language specialization. In thepresent investigation, two tachistoscopic tests of cerebral lateralization, one measur-ing spatial functions and one measuring verbal function, were administered to 73subjects classified by handedness, hand posture during writing-, and sex. Amongboth dextral and sinistral subjects with a normal writing posture, language andspatial functions were specialized to the contralateral and ipsilateral hemispheres,respectively, and lateral differentiation of the brain was strong'. The reverse wasseen in subjects having an inverted writing posture. In all groups, females wereless laterally differentiated than males. In 70 out of 73 subjects, the direction ofcerebral lateralization was accurately predicted by handedness and hand posture.The 3 subjects (2 females and 1 male) who failed to manifest the predicted rela-tions were all left-handers having an inverted hand posture. In this group, lateraldifferentiation was so weak that the reliability of the tachistoscopic tests was re-duced, and we attribute these three predictive failures to this cause. Thus, almostall of the variation in the lateral organization of the brain was accounted for byhandedness, hand posture, and sex.
Copyright 1978 by the American Psychological Association, Inc. 0096-3445/78/0702-0119$00.75
119

120 JERRK LEVY AND MARYLOU REID
Brain Asymmetry and PsychologicalI'unction
Although neurologists have been awarefor well over a century (Broca, 1861 ; Dax,1836/1865; Wernicke, 1874) that the hu-man brain is laterally specialized in function,only during the last decade or so have psy-chologists recognized the pervasive conse-quences for human psychological functionthat follow from this fact. This new interestin lateral dominance has been stimulated bythree factors : Sperry's work with split-brainpatients (see Sperry, 1974; Sperry, Gaz-zaniga, & Bogen, 1969) ; the development ofnew techniques for assessing lateral organ-ization of the brain in normal populations(Kimura, 1966, 1967; Witelson, 1974);and, the recognition that abilities, processes,and structures discovered by psychometri-cians, by physiological, cognitive, and clin-ical psychologists, and by those interestedin the phylogeny and ontogeny of behavior(often in the absence of any knowledge of orinterest in cerebral lateralization) now seemto relate directly to the asymmetry of the hu-man brain.
Until the 1930s, the left cerebral hem-isphere of right-handers was consideredto be the dominant hemisphere, not only forlanguage function but also for the control ofall higher (and sometimes lower) cognitiveprocesses. Lord Brain stated in 1962 thatthe left side of the brain was the seat ofunderstanding of sensory input (Brain,1962). Even as late as 1973, Eccles main-tained that the activities of the right hem-isphere do not directly give rise to consciousexperience and that the source of intentionalbehavior was in the left half-brain (Eccles,1973). Thus, for many years the concept ofcerebral asymmetry was interpreted to mean,not that there were two specialized hem-ispheres, each predominant in a set of func-tions for which the other was ill designed,but rather that man was a half-brained
This research was supported in part by NationalScience Foundation Grant RNS 75-23061.
Requests for reprints should he sent to JerreLevy, Department of Behavioral Sciences, Uni-versity of Chicago, Chicago, Illinois 60637.
species whose left hemisphere was the soleintegrative organ of thought, the right hem-isphere being merely a defective, parallelcircuit whose only function was to serve asan input-output unit of the computer onthe other side of the brain.
Obviously, had this view been correct, thefact of cerebral asymmetry would have hadlittle relevance to most psychologists, sinceany cortically integrated function they couldstudy—intelligence, verbal and visual mem-ory, speech and picture perception, writing,reading, drawing, learning, Gestalt percep-tion, reasoning, imagination, and so forth—would have been products of the left hem-isphere, which is hardly saying more thanthat they were products of the brain. Littleor no understanding of the neurologicalfoundations of human cognition could havebeen gained from purely behavioral investi-gation of neurologically normal populations.
Psychologists are currently devoting somuch attention to the psychological con-sequences of cerebral asymmetry becauseevidence gathered over the last 40 yearsdemonstrates beyond any question that theorgan of human thought consists of twocomplementary brains, each with its owncognitive (and possibly affective) processes,each superior in functions for which theother is inferior. It now appears that verballearning and memory, phonetic analysis,deductive reasoning, serial processing, verbalIQ, and symbolic thinking are all processesintegrated in the left hemisphere, whileimagistic learning and memory, perceptualsynthesis, analogical reasoning, parallelprocessing, performance IQ, and concretethinking are all processes integrated in theright hemisphere. Much of the evidencesupporting the above conclusions can befound in Dimond and Beaumont (1974).
However, the modern view of humanbrain organization is, in some ways, aslimited in its generality as was the classicalview. The available evidence shows thatthere is a great deal of individual variationin the extent to which the two hemispheresare laterally differentiated, the competencyof each hemisphere to perform its specializedfunctions, the direction of lateralization, the

VARIATIONS IN CEREBRAL ORGANIZATION 121
adequacy of what has been called the meta-control system (Levy & Trevarthen, 1976)—which selectively and adaptively activatesthe appropriate hemisphere in accordancewith task requirements—the extent to whichone or the other hemisphere tends to controlbehavior, and in the relationship betweenhandedness and side of cerebral dominancefor language. Clearly, to gain a solid under-standing of the relationship between neuro-logical and psychological organization, stud-ies cannot be limited to those segments ofthe population who vary little, if at all, inthe pattern of cerebral lateralization. At bestsuch studies can refine and clarify our under-standing of the nature of each hemisphere'sspecializations. They cannot, however, revealthe magnitude, nature, or causes of the co-variation between neural and cognitive or-ganization. In the sections that follow, someof the conclusions and speculations regard-ing the neuro-behavioral-cognitive associa-tions are briefly reviewed.
Handedness and Cerebral Lateralii
It has been known for a number of yearsthat brain organization in left- and right-handers differs. Left-handers are less later-ally differentiated than are right-handers(Goodglass & Quadfasal, 1954; Hecaen &De Ajuriaguerra, 1964; Hecaen & Sauguet,1971), as demonstrated by their (a) mani-festation of bilateral language in the Wadatest (Milner, Branch, & Rasmussen, 1966) ;(b) more frequent, but typically transientaphasia following lesions to either hem-isphere (Goodglass & Quadfasal, 1954;Luria, 1947/1970; Subirana, 1969); and(c) in normals, smaller sensory half-fielddifferences on both dichotic listening tests(Curry, 1967; Curry & Rutherford, 1967;Satz, Achenbach, Pattishall, & Fennell,1965) and tachistoscopic lateral field tests(Beaumont, 1974; Bryden, 1965). In addi-tion, the left-handed population is hetero-geneous with respect to the direction oflateralization, some 60 % or so having lan-guage functions in the left hemisphere andvisuospatial functions in the right, withabout 40% having the reversed pattern, as
inferred from neurological patients havingunilateral lesions, about 85% of whom aremale (Goodglass & Quadfasal, 1954;Roberts, 1969). In right-handers, only avery small fraction have reversed dominancerelations (see Levy, 1974). A number ofrecent studies have also demonstrated varia-tions with respect to the direction of lateral-ization in the normal sinistral population(Cohen, 1972; Satz et al., 1965; Zurif &Bryden, 1969).
In spite of the obvious association be-tween handedness and brain organization,there is little or no understanding of theunderlying basis of this relationship. Theheterogeneity within both left- and right-handers renders any simple model untenable.First, why is the degree of cerebral lateral-ization correlated with handedness? Second,why do some people have their dominanthand ipsilateral to the language hemisphere,and why is this so prevalent in left-handers?Third, how, in such people, is manual con-trol achieved?
With regard to the first two questions, somehave suggested that right-handedness and lefthemisphere dominance for language arespecies-specific traits and that any deviationsfrom this result from early brain injurywhich interfered with normal development(Bakan, Dibb, & Reed, 1973). Others havesuggested that the various phenotypes aredue to varying genotypes (Annett, 1964;Levy & Nagylaki, 1972). Neither of thesepositions would deny the possible modulat-ing effect of sociocultural factors in pheno-typic expression. However, the specificationof the precipitating causes of some phe-nomenon does not thereby constitute amechanistic explanation: The postulate thatbrain damage or genes lie at the source ofobserved brain—hand relationships providesno insight into the neurological mechanismsby which these relationships are realized.
With regard to the third question, somehave postulated that in those sinistrals withlanguage in the left hemisphere, writingmovements are programmed by the righthemisphere (see Hudson, 1975 ; Levy, 1977).Others have suggested that the right hem-isphere is specialized in these people for

122 JERRE LEVY AND MARYLOU REID
manual skill and that linguistic informationnecessary to program writing is transcallo-sally transmitted from the left to right hem-isphere, which directly controls the left hand(Geschwind, 1975). Finally, Levy (1974)has hypothesized that there are variationsamong individuals in the proportion ofcrossed and uncrossed pyramidal fibers andthat people in whom the uncrossed bundleis larger control the writing hand from thelanguage hemisphere via the ipsilateralpyramidal tract.
Of possible relevance to the issue of themanual control mechanism is the fact thatleft-handers are heterogeneous not only withrespect to the direction of cerebral lateraliza-tion but also with respect to their handposture in writing. The vast majority ofright-handers hold their pencil in a typicalfashion, the hand held below the line ofwriting, the pencil slanted upward, pointingtoward the top of the page. However, only50% ±20% of left-handers adopt this writ-ing posture, with the remainder either ro-tating their hands or the paper on whichthey are writing in such a way that the handis held above the line of writing and thepencil points toward the bottom of the page(see Figure 1).
The inversion of hand position in writinghas been interpreted as an adaptation to thenecessity of writing from left to right withthe left hand (Enstrom, 1966, 1971; Wahl,1955). However, contradicting this argu-ment are the observations that a significantfraction of sinistrals do not use an invertedhand position; that a large fraction of left-handed Israelis do invert, though right-handed Israelis usually do not (Shanon, inpress; Gur & Gur, Note 1) ; and, finally,that some small fraction of dextral writersalso invert their hands in writing (Levy,1974). Lewis Carroll, a mirror writer, ap-parently used the inverted position whenwriting with the right hand, as shown in aphotograph of him included in Barsley's(1967) book. Levy (1973) reasoned that ifhand inversion could not be accounted forby the interaction of the side of manualdominance and directionality of writing,then it must reflect aspects of central neuro-logical patterning.
Levy and Mandel (Levy, 1973, 1974)tested noninverted and inverted left-handersand noninverted right-handers with twotachistoscopic tests, one measuring verbaldominance and one measuring spatial dom-inance. They found results consistent with
SinistralNoninverted Writer
SinistralInverted Writer
DextralInverted Writer
DextralNoninverted Writer
UncrossedPyramidal Tract
Figure 1. Illustrations of the inverted and noninverted writing postures of left- and right-handersand postulated relationships among cerebral dominance for language, manual dominance forwriting, and hand posture.

VARIATIONS IN CEREBRAL ORGANIZATION 123
the view that inversion is indicative of alanguage-dominant hemisphere ipsilateral tothe dominant hand, while noninversion isindicative of a language hemisphere con-tralateral to the dominant hand, thoughnoise in the data left them open to otherpossible interpretations. Figure 1 depicts therelationships among cerebral dominance,manual dominance, and hand position inwriting as postulated by Levy (1973, 1974).
If the hypothesized relationship betweenhand position in writing and the location ofthe language hemisphere were confirmed,we would be faced with the problem of ex-plaining why hand position in writing shouldvary with the relation between manual dom-inance and cerebral dominance for language.The inverted hand posture during writing-would clearly require a different motor pro-gram than a noninverted hand position andwould produce a different kinesthetic feed-back pattern. If people with an ipsilateralrelation between the writing hand and lan-guage hemisphere control the hand, never-theless, from the contralateral half-brain, itis difficult to conceive of why an atypicalmotor program or atypical feedback infor-mation would be utilized or necessitated.If, on the other hand, manual control isachieved via the ipsilateral motor tracts, itmay be that the motor program and/or thekinesthetic feedback patterns remain in-variant regardless of whether it is a contra-lateral noninverted hand or an ipsilateral in-verted hand whose writing movements arebeing controlled. It may be of relevance inthis connection that simultaneous mirrormovements of one hand of the actions of theother (synkinesis) are observed in cases ofcallosal agenesis and in hemiplegics (Chaco& Blank, 1974; Ettlinger, Blakemore,Milner, & Wilson, 1972; Green, 1967;Marie & Foix, 1916).
This phenomenon, given the conditionsunder which it occurs, indicates that themotor programs for manual control in onehemisphere, typically controlling the con-tralateral hand, will also produce mirroredmovements in the ipsilateral hand, presum-ably via ipsilateral pathways, in the absenceof inhibiting signals from the other side of
the brain. Quite possibly the writing mo-tions of an inverted ipsilateral hand are, insome way, a mirror reflection of certainaspects of the writing motions of a nonin-verted contralateral hand. In any case, theproblem of understanding why some people,in contrast to the majority, write with thehand ipsilateral to the language hemisphereand how manual control is achieved in suchindividuals stands as a major challenge toneuropsychological research.
Cognitive Organisation and Handedness
In addition to neurological differences,there are also cognitive differences cor-related with handedness. That sinistrals anddextrals should display certain variationsin their cognitive patterns was postulatedby Levy (1969) on the following grounds.Work with split-brain patients strongly sug-gested that the cognitive capacities of theleft and right hemispheres were not onlydifferent, but that the processing strategiesutilized by one side of the brain seemed toexclude those utilized by the other. As anexample, the left hemisphere appeared toextract only those properties of a stimulusthat were needed to categorize and label it,whereas the right hemisphere appeared togenerate a Gestalt representation based on thestructural relations in the stimulus. Further,Levy concluded that the verbal-analyticoperational strategy of the left hemispherewas not merely optional but was obligatoryfor that side of the brain, whereas thespatial-synthetic strategy was obligatory forthe right side of the brain. Levy reasonedthat a bilaterally symmetric brain would belimited to verbal or to spatial processing, orthat each hemisphere would be partially or-ganized to serve each function and could,therefore, serve neither of them well.
Since some sinistrals, as discussed pre-viously, tend toward bilateralization of func-tion, we would expect that their perform-ance would be adequate or more than ade-quate in bilateralized functions but depressedin functions for which no fully specializedhemisphere exists. Those individuals inwhom language organization has invaded

124 JERRE LEVY AND MARYLOU REID
the visuospatial hemisphere would displaydefective visuospatial performance relative toverbal skills. Those in whom visuospatial or-ganization has invaded the language hem-isphere would manifest, in contrast, highervisuospatial than verbal skills.
Statistically, left-handed graduate sciencestudents could be predicted, therefore, toshow inferior performances on tests ofvisuospatial function, since they have beenadmitted to university study on the basis ofdemonstrated verbal competence. Levy(1969) compared sinistral and dexlral malegraduate students on difference scores be-tween the verbal and performance IQs ofthe Wechsler Adult Intelligence Scale. Shefound a verbal IQ minus performance TQdifference of 25 points for left-handers, butonly an 8-point difference for right-handers.Depression in spatial-perceptual test scoresof left-handed college students has also beenreported by Silverman, Adevai, and Mc-Gough (1966), James, Mifferd, and Wieland(1967), Miller (1971), and Nebes (1971).Also note that in the Levy (1969) study, whenthe Wechsler subtest scores were factoranalyzed to purify the verbal and perceptualfactors (Oilman, Note 2 ) , sinistrals as com-pared to dextrals were depressed on the per-ceptual factor and superior on the verbal fac-tor. Those left-handers with partial bilateral-ization of function may well be cognitivespecialists who, although impaired on one setof abilities, excel on another (Levy, 1974).Having two verbal hemispheres, for ex-ample, may reduce cognitive fatigue forverbal processing and/or may attenuate thedeterioration in performance that wouldoccur if the system controlling the selectiveactivation of the task-appropriate hemispherewere imperfect.
Among certain populations, such as artistsand architects, the reverse pattern of cog-nitive abilities should be seen in left-handers :reduced verbal abilities and possibly superiorspatial reasoning. We have found only onestudy in the literature relevant to this ques-tion. Peterson and Lansky (1974) foundthat the proportion of sinistrals increasedfrom the freshman to the senior year in aschool of architecture and that left-handed
architecture students, whether freshman orseniors, greatly surpassed right-handers ina complex maze construction task. Thoughno measures of verbal function were ob-tained, the proposed model predicts a largerdiscrepancy between verbal and spatialabilities in left-handers than in right-handers.
The currently available evidence regard-ing the cognitive differences in left- andright-handers can be integrated with ourknowledge of neurological variability, butclearly, many more data are needed beforedefinitive conclusions regarding the cogni-tive consequences of variations in brainorganization can be reached.
Sex Variations in Cerebral Organisation
Another source of variation in cerebralorganization has been related to sex differ-ences. Females as compared to males haveconsistently shown smaller sensory fielddifferences on dichotic listening (Harshman,1976) and tachistoscopic tests (Knox &Kimura, 1970). The relative inferiority offemales on tests of spatial relations, whichhas been widely reported across a variety ofcultures (Harris, 1975), also offers supportfor the notion of partial bilateralization ofverbal function in females. Since in mostright-handers, spatial reasoning and verbalcapacity are integrated by the right and lefthemispheres, respectively, various authors(Knox & Kimura, 1970; Levy, 1973; Rudel,Denckla, & Spalten, 1974; Witelson, 1976)have proposed that the right hemisphere infemales is less well organized for its special-ized functions than in males and is partiallyorganized for the opposite hemisphere'sspecialty. Harris (1975) has, in contrast,suggested that the male and female hem-ispheres are not differently organized butrather that females tend to use the verbalhemisphere for all types of tasks. If, how-ever, the dynamics of hemispheric function-ing differ in males and females, it must bepresumed that there are underlying organ-izational differences responsible for the dif-fering dynamics.
The causes of male-female differences in

VARIATIONS IN CEREBRAL ORGANIZATION 125
cognitive and neurological patterns are un-known, but regardless of the extent ofgenetic, hormonal, or sociocultural deter-mination, the nature of the sex differencesseen in psychological functioning can berelated quite directly to those in cerebrallateralization. A major question is whetherit is indeed correct that the spatial hem-isphere in females, whether left or right, isincompletely specialized. The data are justas consistent with the hypothesis that re-gardless of whether a female right hem-isphere is specialized for spatial or for verbalfunction, it is incompletely developed. Sincein about 95% of people spatial abilities areorganized in the right hemisphere, datafrom a random sample of the populationcould not discriminate between the "spatialdeficit" hypothesis and the "right hem-isphere deficit" hypothesis. A much closerlook at male-female differences in peoplewith mirror-reversed lateralization patternscould yield a definitive answer.
Neurobehavioral Relations
The complex relationships between cere-bral and cognitive organization, handedness,eyedness, the brain-hand control system,and sex-linked factors are yet to be under-stood. Nevertheless, from the previouslydiscussed studies and foregoing speculations,certain regularities in these relationships canbe perceived. Our attempt to understandthe causal chains underlying correlatedvariations in neurological and psychologicalorganization requires that we interrelate thehost of factors affecting either or both intoa comprehensible model.
Tn the present study, some aspects ofwhich have been described (Levy & Reid,1976), we investigated the relationshipsamong degree and direction of brain lateral-ization, handedness, hand position in writ-ing, sex, and performance levels on verbaland spatial tests of lateralization. Cerebralorganization was measured by means oftachistoscopic lateral field tests. In normalpopulations, as mentioned previously, suchtests have generally confirmed deductionsregarding cerbral asymmetry drawn fromstudies of neurological patients.
Nevertheless, the validity of sensory half-field tests has been questioned because asmany as 30% of right-handers have beenfound to display left-field superiorities onverbal tests (Berlin & Cullen, 1977). Ifthese results were taken as an indicant ofthe proportion of dextrals with right hem-isphere dominance for language, they wouldbe at extreme variance with the neurologicalliterature. Clearly, factors other than cerebralasymmetry can make a significant contribu-tion to the variance in lateral field differ-ences.
However, given what is known of dextralbrain organization and, in particular, of themale dextral brain, we can specify the cri-teria that tests must meet if they are to beconsidered valid indices of hemisphericlateralization. Essentially 100% of right-handed males from dextral families, unlessthey are brain damaged, are left hemispheredominant for language and right hemispheredominant for visuospatial integration. Since,depending on a variety of conditions havingnothing to do with hemispheric specializa-tion, a right or a left sensory field superior-ity may emerge on some presumptive testof hemispheric lateralization, at least twotests must be designed to assess the special-izations of opposite hemispheres. Based ontheoretical considerations, it is importantthat each test should depend, to the maxi-mum extent possible, 011 the operationalstrategies of one side of the brain and shouldbe resistent to solution by the strategies ofthe other side. If the two tests are valid, anygiven dextral male should display a rightfield superiority on the test designed toassess the specialties of the left hemisphereand a left field superiority on the test de-signed to assess the specialties of the righthemisphere. If more than a few percent ofsuch subjects manifest the inappropriatefield superiority on either test, a secondgroup of subjects should be tested. If in ex-cess of a very small percentage of thisgroup also displays an inappropriate fieldsuperiority on one or the other of the tests,the test is insufficiently valid and shouldbe modified. If, on the other hand, essen-tially all right-handed male subjects from

126 JERRE LEVY AND MARYLOU REID
right-handed families, excluding those whomthe experimenter has reason to believe maybe brain damaged, display opposite andappropriate field superiorities on the twotests, the tests may be considered to behighly valid indices of cerebral lateraliza-tion; in fact, no more stringent a validitycriterion for functional lateralization can bespecified.
In the present investigation, it was ofparticular importance that our measures ofcerebral lateralization have very high validitysince we wished to assess not only the direc-tion but also the degree of lateral differentia-tion in a sample of subjects, many of whomwe expected to have only a very slight de-gree of lateral specialization. Obviously, ifthe validity of our tests were not high evenin strongly lateralized subjects, it would beeven lower in weakly lateralized subjects.Consequently, we spent well over a year indeveloping two tests that meet the stringentvalidity criteria discussed previously, andwe feel justified in claiming that our testsactually index cerebral lateralization. In ourpilot subjects, consisting of familial right-handed males, 100% displayed opposite andappropriate field superiorities on our twotests of cerebral lateralization.
Method
Subjects
A total of 77 university undergraduates par-ticipated in the sudy on a voluntary basis. Twelvemale and 12 female subjects were included in eachof three groups denned according to handednessand hand posture in writing. In addition, 1 sub-ject, a female, constituted a fourth "group." Foursubjects were eliminated during initial thresholdtesting, bringing the total number of subjects to 73.Subjects were informed only that left- and right-handers were being tested on tasks of visual per-ception. No subject with known but uncorrectedvisual problems was included. Subjects were notpaid for their participation.
Procedures
Laterality qiiestionnaire. Before tachistoscopictesting began, each subject filled out a lateralityquestionnaire, describing his or her handednesspattern for eight common manual activities (e.g.,writing, combing hair, dealing cards, throwing aball). Subjects rated their own hand preferencefor each activity in terms of both the relative de-
gree of usage and of skill. A 5-point scale wasused for both the skill and usage indices, 1 rep-resenting extreme right-handed dominance and 5complete left-handed dominance for the activity.The form also contained questions pertaining tothe development of hand preference (i.e., whethera change had occurred in handedness or writingpattern since childhood) and familial history ofleft-handedness or ambilaterality. Subjects werealso asked to draw a profile of a human face.Some evidence has suggested (Crovitz, 1962) thatwhile right-handers typically draw left-facing pro-files, left-handers are heterogeneous with respectto profile direction. This drawing task was in-cluded here to see if profile orientation might provean additional behavioral measure indicating cere-bral dominance. Three tests for eye dominance(two administrations of the Asher test and twopointing tests) were also given. In the pointingtests, subjects fixated on a spot approximately6 ft (1.8 m) away, with their index finger oftheir dominant hand pointing to the spot. Theywere told to alternately close each eye whilemaintaining the pointing position and to reportwhen the finger seemed to move away from thespot. The test was repeated using the nondominanthand. Finally, each subject was asked to write asentence while the experimenter observed whetherthe pencil was held in an inverted or a nonivertedfashion (see Figure 1). An inverted writing pos-ture was classified according to one or more ofthe following criteria: (a) The writing hand wasrotated (to the right for left-handers or to theleft for right-handers) so that the hand was heldabove the line of writing, (b) The page on whichthe subject was writing was rotated (to the leftfor left-handers, to the right for right-handers)so that the hand was held to the side or abovethe line of writing, (c) The tip of the pencilpointed toward the bottom of the page. When thesheet was tilted, subjects were asked to rewritethe sentence after the page had been restored toa vertical position with respect to the table'sedge. If writing still proceeded in such a way thatthe pencil pointed toward the bottom of the page,hand posture was classified as inverted. A non-inverted writing posture was defined as one inwhich the hand was held below the line of writing,the pencil slanted upward, pointing toward thetop of the page.
In cases where some ambiguity remained in clas-sifying the hand position, subjects were asked towrite additional sentences on a page set vertical tothe table's edge by the experimenter. It was ob-served that some left-handed subjects, while start-ing to write in an ambiguous fashion, eventuallyadopted an inverted posture after writing a fewwords and maintained this hand position there-after. When hand posture remained ambiguous(two individuals), a potential subject was thankedfor coming to the laboratory and no further testingwas done. Potential subjects had been self-selectedby questionnaire or telephone interview regarding

VARIATIONS IN CEREBRAL ORGANIZATION 127
handedness and hand posture, possibly reducing thenumber of ambiguous cases seen.
On the basis of the dominant writing hand andthe hand position in writing, subjects were classi-fied either into the noninverted right-handed group(Group RN; » = 24), the noninverted left-handedgroup (Group LN; n = 24), the inverted right-handed subject (Subject RI, n = l ) , or the in-verted left-handed group (Group LI, n = 24).
Visual tests. After completing the questionnaire,the first tachistoscopic test (either the syllable testor dot location test) was administered.
The second test was given in a separate session,2-7 days later for most subjects. Task order wascounterbalanced across subjects in each sex andhandedness group. The general procedure for bothtasks was as follows. Subjects were told to fixateon a center point in the fixation field of anICONIX four-channel tachistoscope (model 6137and system 189 logic set) and when they wereready, to press a foot pedal which would initiatethe trial. After the pedal was pressed, the fixationfield remained on for an additional 300 msec, fol-lowed by a stimulus field presented either to theleft or right of fixation for the exposure durationdetermined for each subject. The stimulus fieldalso contained a number (0-9) at fixation. Subjectswere told to first report the number at fixation andthen to respond to the lateral stimulus appro-priately for each task (see task descriptions below).This method was employed to insure against eyemovements after fixation during stimulus presen-tation (see McKeever & Huling, 1971). The stimu-lus was always followed by a visual noise maskfor 200 msec. After every trial, each subject's re-sponses were recorded by the experimenter, whotold the subject to start the next trial when ready.
Stimuli were constructed with white Visigraphicdots (Vg in. 13.2 mm] diameter), numbers, andletters (%e if- [8 mm] high) on black posterboardcards. White stimuli on a black background werechosen to reduce the framing effects of the tachis-toscopic apparatus and thus to minimize any scan-ning tendencies that might be peculiar to a nar-rowly defined field. This was especially importantin the dot location test, where the stimuli in-cluded their own frame of reference, which mighthave competed with the tachistoscope's visual fieldboundaries. In addition, high-contrast borders,which would be produced by a tachistoscopic frameand a white background, would likely have di-verted the subject's attention away from the criticalstimuli.
Screening test. A screening test requiring sub-jects to respond to the presence or absence of alateral dot stimulus, without mask interference, wasadministered to insure that subjects demonstratedequal perceptual sensitivity in each visual field. Onhalf the trials, the dot was presented % in. (1.6cm; 1.0° of visual angle) away from fixation ineither the left or right visual field, while on theother half, no stimulus was presented. Exposuredurations were varied systematically in ascending
and descending scales until subjects displayed acriterion of 80% correct detections. Initial trialsbegan at 1 msec, and exposures were lengthened in1-msec increments every five trials. We thoughtthat any subject demonstrating unequal thresholdsfor the left and right fields on this simple detectiontask might be suffering from a subtle unilateraldysfunction in the visual processing regions of thebrain. In addition, if threshold asymmetries wereobtained, we would expect that performance onmore complex tasks would be influenced regardlessof the cognitive specializations required. Of the 77subjects tested, 73 yielded identical left and rightvisual field thresholds. In the other 4 subjects, thethreshold for one field was at least twice as longas that for the other. These subjects' data wereexcluded from further analyses.
Dot location test. A second dot detection testwas given, identical to the first except that a visualmask followed the stimulus for 200 msec and initialtrials began at 10 msec, incremented by S msecevery fifth trial. An additional 20 msec was addedto the threshold measure and determined the ex-posure duration for each subject on the subsequentdot location test. Exposure durations were set inthis way because pilot investigations had shown a50%-60% accuracy for dot location across visualfields with this stimulus duration.
The dot location test required subjects to locatethe correct position of a dot in a rectangle withinthe left or right visual field. The fixation fieldconsisted of two rectangular frames, 2.25 X 1.75 in.(5.72 X 4.45 cm), with the fixation point midwaybetween them. The medial borders of the frameswere located •i/w in. (4.8 mm; .3°) away fromfixation. Subjects were instructed that a single dotwould appear in 1 of 20 possible positions in eitherthe left or right rectangle and that their task wasto indicate in what position the dot had appeared.Location of the dots was reported from a cardmounted below the tachistoscope. To insure main-tenance of fixation, subjects were required to reporta single digit appearing at fixation during presen-tation of the dot stimulus. The fixation digit wasreported first, followed by the dot position. Sub-jects were encouraged to guess about the locationof the dot when they were not sure. The maskfor the dot detection and localization tasks wasidentical to the preexposure field specifications,only each rectangle was filled with a masking noise.Illumination levels in effective millilamberts forthe three fields were .28 for the fixation field, .28for the stimulus field, and .69 for the masking field.
The dot positions occupied a 5 row by 4 columnmatrix within each rectangular frame. Dot posi-tions ranged from .70° to no more than 2.7°laterally from fixation. Dots were presented inevery position once in each visual field for a totalof 40 trials. Position and visual field were ran-domized with respect to presentation. A 2-min restperiod was given after 20 trials.
Syllable test. In the syllable test, subjects re-ported a three-letter nonsense syllable flashed in

128 JERRE LEVY AND MARYLOU REID
either the left or right fields. Since the nondomi-nant hemisphere in commissurolomy patients hasshown at least some proficiency in the recognitionof meaningful words, but does not seem capableof dealing- with linguistic, material on a phonemicbasis (Levy & Trevarthen, 1977), nonsense sylla-bles were used instead of words in the presentexperiment in an attempt to induce maximal per-formance differences between language dominantand nondominant hemispheres.
All syllables had the form consonant-vowcl-consonant (e.g., gek, fap, tos). The consonantsused were d, g, p, k, t, s, and f, and the vowelswere a, e, and o. All letters were used an approxi-mately equal number of times in generating 60syllables on the stimulus list. Syllable letters werepresented in a vertical array, so that scanningwould proceed from top to bottom, thus counter-acting the left-right word or page-scanning tend-encies which have confounded previous results fromverbal lalerality tasks (Levy, 1974; White, 1969).The syllables appeared 1.25 in. (3.2 cm; approxi-mately 2°) to the left or right of the fixation num-ber. The syllables were \I/H in. (2.9 cm) in length.The mask for this test was a partial noise field,extending across bot'i visual fields. Effective illu-mination levels (in millilamberts) for the threefields were .41 for the fixation field, .46 for thestimulus field, and 1.19 for the masking field.
The higher illuminations in the syllable test thanin the dot location test resulted from the fact thatblack cards used for constructing the syllable testhad a somewhat higher reflectance than those usedin constructing the dot location test because wewere unable to secure a sufficient number of iden-tical black posterboards for making the tachisto-scopie cards.
Subjects were again required to report the fixa-tion number first and the syllable second. Subjectswere encouraged to report whole syllables ratherthan spelling out each letter. Each syllable waspresented once in each visual field, yielding 60trials per field. One order of stimulus presentation(which randomized the order of fields stimulated)was used for all subjects. Subjects were given arest period (3 min) after the 60th trial.
Before testing began, subjects received 18 letter-identitication trials in order to familiarize themwith the testing procedure and to determine expo-sure for the syllable test. In these trials, a singleletter was presented 2° from the fixation number,and the subject merely had to identify the letterpresented. Exposure for the letter stimulus wasfirst set at 50 msec and increased (or decreased)in 5-msec increments until threshold (80% cor-rect) was reached. Then 60 msec was added to eachsubject's threshold level for the syllable-identifica-tion task, since this linear addition yielded approxi-mately 50%-60% syllables correct across both vis-ual fields in pilot studies. Subjects also received12 practice trials with syllable stimuli not used inthe subsequent tests.
Scoring. Since subjects differed on exposure
durations for both tests of cerebral lateralization,the percent correct responses for each field for asubject on a given test was divided by his or herexposure duration. This score represents, in effect,the amount of information correctly processed perunit time. The mean scores of the syllable test andthe dot location test averaging across visual fieldswere set equal for Group RN in order that thepattern of test scores could be compared for othergroups and between sexes. Thus, each subject'sfield score on the syllable lest was the percentcorrect identifications divided by the exposure du-ration in milliseconds and multiplied by 300 ; thesame operation was applied to the dot location testexcept that the constant multiplier was 83.8, in-stead of 300. For Group RN, this yielded a meanverbal and spatial score of .67. Summing over allsubjects, the mean spatial score was .66, and themean verbal score was .65. Therefore, equalizationof the means for Group RN also produced approxi-mate equalization for the sample as a whole.
Results
Direction of Lateralization
An analysis of variance with three be-tween-group factors—sex (male, female),group (RN. LI, LN) , and task order (syl-lable test first, dot location test first)—andtwo within-group factors—lateral field (right,left) and task (syllable, dot location)—wascarried out, primarily to determine whetherthe triple interaction among group, lateralfield, and task was significant. Specifically,we wished to sec if Groups RN and LFwould show a right visual field (RVF) su-periority on the syllable test and a left visualfield (LVF) superiority on the dot locationtest, while Group LN would manifest thereverse. Subject RI was not included in thisor in any subsequent analyses.
The significant effects in this analysis aredepicted in Figure 2. Task order made nosignificant contribution to the variance, eitheras a main effect or in any of its interactions.The main effects of group, F(2, 60) = 5.95,p < .005, and sex, / ' ( I , 60) = 18.24, p <.001, were significant, showing that GroupsRN and LN had higher scores than didGroup LI and that males were superior tofemales.
The Group X Lateral Field X Task inter-action was highly significant, /;(4, 120) =80.35, p < .001, revealing that Groups RNand LI had higher RVF scores on the syl-

VARIATIONS IN CEREBRAL ORGANIZATION 129
lable test and higher LVF scores on thedot location test, with Group LN having thereverse. On the syllable test, Group RN alsooutperformed Group LI in the RVF hut notin the LVF. There were no differences be-tween these two groups in either field on thedot location test.
The Sex X Lateral Field X Task inter-action was significant, F(2, 120) = 4.82, p< .01, since the Field X Sex interaction wasnot significant on the syllable test (bothmales and females having higher RVFscores) but was significant on the dot loca-tion test (males having higher LVF" and fe-males having higher RVF scores). Thislatter result indicates that the RVF scores ofGroup LN females on the dot location testwere sufficiently high to outweigh the lowRVF scores of females in the other twogroups.
Finally, the Group X Sex X Lateral FieldX Task interaction was significant, /7(4,120) = 10.38, p < .001. This four-way inter-action was due to females' contributing asmaller percentage of the variance in theGroup X Lateral Field X Task interactionthan males.
Post hoc analyses were run on each groupseparately. In neither Group RN nor GroupLN was there a significant main effect ofsex, but in Group Lf, males were superiorto females, / ' ( I , 20) = 8.44, p < .01.
In none of the groups was there a signifi-cant Sex X Task or Sex X Field interac-tion, but in all groups the Sex X Field XTask interaction was significant: P(2, 40)= 21.36, p < .001 for Group R N ; F ( 2 , 40)= 5.54, p < .01 for Group LI; and F(2, 40)= 4.46, p < .025 for Group LN. In allgroups, the Field X Task interaction waslarger for males than females, and in allgroups the Sex X Field interaction was sig-nificant for the dot location test but not forthe syllable test. Specifically, on the clot lo-cation test, males in Groups RN and LI ex-ceeded females in the LVF but not in theRVF, showing that males and females didnot differ in their noncloniinant spatial hemi-sphere, but that females were depressed intheir dominant spatial hemisphere. The sameconclusion held for Group LN : Males sur-passed females in the RVF but not in the
Syllable Test Dot Location Test
1.0
0.8
0.4
Right Visual Field(RVF)
Left Visual FieldI LVF)
RN LI LN
Groups
RN LI LN
Groups
Figure 2. Right and left visual field scores on theverbal and spatial tests for right- and left-handerswith a noninverted writing posture (RN and LN,respectively), for left-handers with an invertedposture (LI) , and for males and females.
LVF, the fields contralateral and ipsilateral,respectively, to the dominant spatial hemi-sphere for this group.
Figure 3 depicts the difference scores be-tween fields and their standard errors as afunction of sex, group, and task. The datafor Subject RI are included to show thatshe had opposite field superiorities fromthose of Group RN.
Degree of Lateralization
In the foregoing analysis of variance, dif-ferences between the groups were due inpart to the opposite directions of lateraliza-tion for Groups RN and LI versus GroupLN and in part to differences in magnitudesof effects for different groups. In order todetermine whether the degree of field effects,ignoring direction, differed between groups,analyses of variance were run on each testseparately in which, instead of lateral field(right or left) being entered as a factor,

130 JERRE LEVY AND MARYLOU REID
(jROUP
LI
Rl
LN
RN
12 ?
12 <f
1 ?
0 «/
12 ?
12 J
12 ?
12 </47-
LI
Rl
LN
RN
-.6 -.4 -.2 0 .2 .4
RVF SCORE - LVF SCORE
SYLLABLE TEST
-.6 -.4 -.2 0 .2 .4
RVF SCORE - LVF SCORE
DOT LOCATION TEST
Figure 3. The differences between righ visual field (RVF) and left visual field (LVF) scoreswith standard errors of the differences for right- and left-handers with noninverted (GroupsRN and LN) and inverted (Subject Rl and Group LI) writing postures.
superior-inferior field (better or poorer)was entered as a factor. For each subject,the score of his or her better field, regard-less of whether it was the RVF or the LVFscore, was recorded as the superior fieldscore (SVF), and the score of the poorerfield was recorded as the inferior field score(TVF). There will, necessarily, be a maineffect of field in such an analysis because ofthe definition of the field factor.
In each of the two analyses, group, sex,and task order were entered as between-group factors and superior-inferior field asa within-group factor.
Task order made no significant contribu-tion to the variances in either analysis, eitheras a main effect or in any of its interactions,while field (SVF or IVF) was, of course,highly significant in both analyses.
Syllable test. On the syllable test, con-sistent with the first analysis of variance,both group, F(2, 60) = 9.78, p < .001, andsex, P(\, 60) = 13.20, p < .001, were sig-nificant. Group LI was inferior to Group RNat p < .05 and to Group LN at p < .01.Groups RN and LN did not differ. Malesand females did not differ in Group RN,
but in Groups LI and LN, males were su-perior (p < .05 and p < .01, respectively).
Of primary interest, the Group X Fieldeffect was also significant, F(2, 60) = 7.17,p < .005. In the IVF the groups only dif-fered at the p < .05 level, Group LI beinginferior to Groups RN and LN, these latternot differing. In the SVF the group effectwas larger (p < .01) and showed the samerelationships. Thus, Group LI displayed onlya minor depression in performance in theinferior verbal hemisphere, but a relativelylarge depression in the superior verbal hemi-sphere (see Figure 4), resulting in smallfield differences for Group LI.
There was a significant Sex X Field in-teraction, the difference in males and fe-males being larger in the SVF (p < .001)than in the IVF (p < .05), with males sur-passing females in both, again suggesting aspecific depression of superior hemisphereperformance. The triple Sex X Field XGroup interaction was not significant.
Dot location test. On the dot locationtest, the group effect was not significant, allgroups performing at similar levels. The sexeffect was, however, significant, F(l, 60) =

VARIATIONS IN CEREBRAL ORGANIZATION 131
7.20, p < .01, females scoring lower thanmales. The Sex X Group interaction wasnot significant.
As on the syllable test, both the Group XField and Sex X Field interactions weresignificant, F(2, 60) =9.16, p < .005 and/''(I, 60) = 33.02, p < .001, respectively. Inthe IVF, the groups did not differ, whilein the SVF, Group LN outperformed GroupRN ( / > < 0 . 1 ) and Group RN surpassedGroup LI (/> < .01). Thus, the magnitudeof field differences was greatest for GroupLN and least for Group LI. Males and fe-males differed in the IVF at only the p <.05 level; in the SVF they differed at thep < .001 level, males being superior to fe-males in both, and females, therefore, havingsmaller field differences. The Group X FieldX Sex interaction was not significant.
Figure 4 displays the significant relation-ships observed on both the syllable and dotlocation tests. The results of these analysesshow that in addition to the group differ-ences in direction of lateralization, there arealso group differences in degree of laterali-zation. Group LI, on both the syllable anddot location tests, was very significantly in-ferior to the other two groups in the SVF,but in the IVF showed only a minor defecton the syllable test and a nonsignificant su-periority on the dot location test. In the lat-ter task, also, Group LN surpassed the othertwo groups in the SVF but not in the IVF,indicating a stronger degree of lateralizationfor this group. The performance of females,relative to males, was similar to that ofGroup LI, relative to the other two groups.
Qualitative Differences in HemisphericProcessing
On the syllable test, in addition to accu-racy measures, the nature of incorrect re-sponses was analyzed. Though the subjects'responses on some trials bore no obvious re-lation to the stimulus, on other trials therewere partially correct responses; that is,either all three letters were correctly given,but in permuted order (e.g., keb for bek),or one or two letters were the same as thosein the stimulus (e.g., gek for bek). Theseresponse errors were examined to see whether
Syllable Test
_c•a
0.8
o 812a cu
. •&.2 ro
0.4
1.0
£ i£ 0.8"° "o
3 33"> .2 0.4
Superior Visual Field(SVF)
- Inferior Visual FieldIIVF)
V
Dot Location Test
/. V
r
RN LI LN
GroupsRN LI LN
Groups
Figure 4. Superior and inferior visual field scoreson the verbal and spatial tests for right- and left-handers with a noninverted writing posture (RNand LN, respectively), for left-handers with aninverted posture (LI) , and for males and females.
error patterns were associated with a par-ticular field or group.
One or Ivuo letters correct. Partially cor-rect responses in which one or two letterswere correct were partitioned into those inwhich the initial or the initial two letterswere correct (1C responses), those in whichthe final or final two letters were correct(FC responses), or those in which the firstand last letters were correct, a category ig-nored in further analyses.
For purposes of these analyses, based onpreceding results, we defined the right visualfield as the dominant language field for sub-jects in Groups RN and LI and the leftvisual field as the dominant language fieldfor subjects in Group LN and for SubjectRT.
In order to determine whether these par-tial errors varied as a function of the domi-nant or nondominant language field, weconstructed the following index of field dif-

132 JERRE LEVY AND MARYLOU REID
ferentiation (D score). Whenever an 1Cresponse was given to a stimulus in thedominant field, it was assigned a value of— 1 ; when it was given in response to astimulus in the nonclominant field, it wasgiven a value of +1. Score assignments forFC responses were reversed (+1 for domi-nant field, — 1 for nondominant field). Asubject's scores were then summed and di-vided by the total number of 1C and FCresponses. The number of 1C and FC scoresoccurring- in each field were also preserved.If TC and FC responses arc randomly givento stimuli in the two fields, the mean D scoreshould be zero. If D scores are positive, then1C responses are asymmetrically allocatedto nondominant field stimuli and/or FCresponses are asymmetrically allocated todominant field stimuli, and vice versa fornegative scores.
We predicted that all groups would havea positive mean D score on the followinggrounds. Partially correct responses in whichthe initial letters are correct (1C responses)would represent primacy memory, while FCresponses would represent recency memory.Work by Milncr (1959, 1966) and col-leagues (Milner & Teuber, 1968; Scoville &Milner, 1957) with the patient H.M., whosuffers from a total inability to consolidateany information into permanent memory,shows that he can, nevertheless, retain anyamount of verbal information within theshort-term memory span almost indefinitelyby means of verbal rehearsal. He is appar-ently unable, however, to utilize a rehearsalstrategy in order to remember nonverbaliz-able stimuli. In view of the many observa-tions that interference with rehearsal reducesthe recency effect in verbal recall, while leav-ing the primacy effect intact (see Kintsch,1970), this finding suggests that the visuo-spatial hemisphere is incapable of rehearsaland is compelled, therefore, to utilize alter-native storage mechanisms; consequently, itshould display a primacy effect but little,if any, recency effect. Based on the sameconsiderations, one would expect to findstrong recency effects displayed by the ver-bally dominant hemisphere, and in our in-vestigation, a predominance of FC responses.
Our hypothesis was strongly confirmed.
All subjects in Groups RN and LI, as wellas Subject RI, had positive D scores, and19 out of 24 LN subjects also had positiveD scores. The D scores were significantlypositive for all groups: £(23) =6.83, /> <.001 for Group R N ; f ( 2 3 ) = 5.89, p < .001for Group L I ; and / (23) = 1.96, p < .04for Group LN (one-tailed test for all groups).
We do not wish to suggest from these re-sults that the stimuli in the nondominantfield were phonetically analyzed and pro-nounced by the visuospatial hemisphere. Thefinal analysis of the syllables and the pro-nunciation of them was almost certainly car-ried out by the verbally specialized hemi-sphere. Lfowever, the information it receivedfrom the ipsilateral field via commissuralpathways must have been already partiallyabstracted and encoded by the nondominanthemisphere in accordance with its speciali-zations. This is to say that the nature ofinformation received through the cerebralcommissures is very different from thatwhich is transmitted directly from the con-tralateral field. The nondominant hemisphereappears to communicate only certain charac-teristics of information—those appropriatefor its specializations—to the opposite sideof the brain. The fact that there are nocallosal axons from neurons in direct pro-jection cortex is consistent with this con-clusion.
In order to sec whether the positive Dscores were due to FC responses to dominantfield stimuli, TC responses to nondominantfield stimuli, or to both, we examined the 1Cand FC scores for each field. These scoreswere computed as simple proportions of re-sponses in these categories relative to thetotal of 1C + FC responses in each field.
An analysis of variance with field (domi-nant, nondominant), group (RN, LI, andLN), and response type (1C, 17C) as factorswas performed on these proportions. A pre-liminary analysis had shown no main effectof sex and no effect of sex in any interac-tions, and the sexes were therefore com-bined. A main effect of response type wasfound, with 1C responses exceeding FC re-sponses, /'"(!, 66) = 259.72, /; < .001. Asignificant Response Type X Field interac-tion was observed, / ' ( I , 66) =47.98, p <
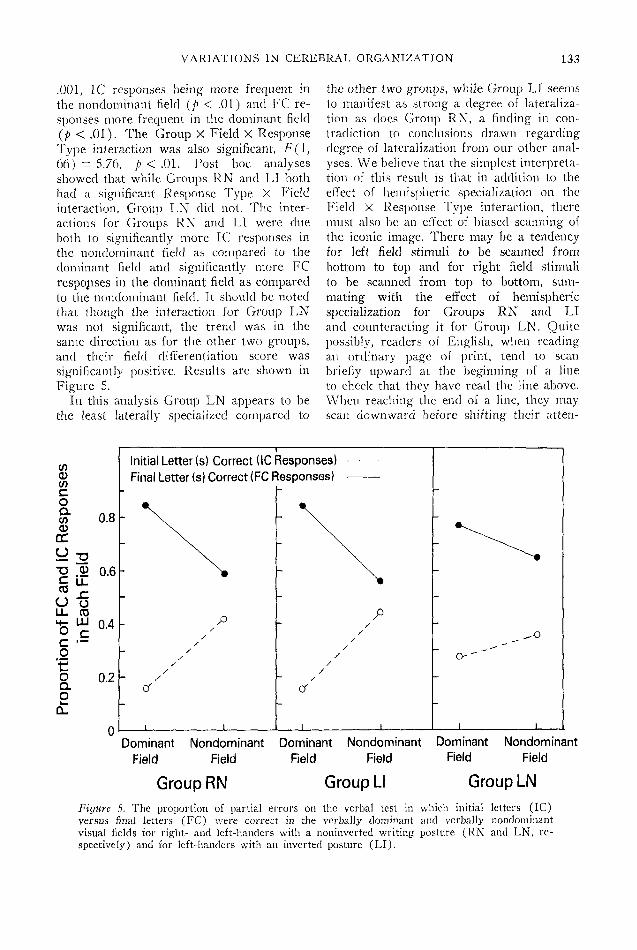
VARIATIONS IN CEREBRAL ORGANIZATION 133
.001, 1C responses being more frequent inthe nondominant field (p < .01) and FC re-sponses more frequent in the dominant field(p < .01). The Group X Field X ResponseType interaction was also significant, F(l,66) = 5.76, p < .01. Post hoc analysesshowed that while Groups RN and LI bothhad a significant Response Type X Fieldinteraction, Group LN did not. The inter-actions for Groups RN and LI were dueboth to significantly more 1C responses inthe nondominant field as compared to thedominant field and significantly more FCresponses in the dominant field as comparedto the nondominant field. It should be notedthat though the interaction for Group LNwas not significant, the trend was in thesame direction as for the other two groups,and their field differentiation score wassignificantly positive. Results are shown inFigure 5.
In this analysis Group LN appears to bethe least laterally specialized compared to
the other two groups, while Group LI seemsto manifest as strong a degree of lateraliza-tion as does Group RN, a finding in con-tradiction to conclusions drawn regardingdegree of lateralization from our other anal-yses. We believe that the simplest interpreta-tion of this result is that in addition to theeffect of hemispheric specialization on theField X Response Type interaction, theremust also be an effect of biased scanning ofthe iconic image. There may be a tendencyfor left field stimuli to be scanned frombottom to top and for right field stimulito be scanned from top to bottom, sum-mating with the effect of hemisphericspecialization for Groups RN and LIand counteracting it for Group LN. Quitepossibly, readers of English, when readingan ordinary page of print, tend to scanbriefly upward at the beginning of a lineto check that they have read the line above.When reaching the end of a line, they mayscan downward before shifting their atten-
tf)
Co
0)DC
— 32"O CDC
o-SU_ 034- LLIO c
_g
oQ.2a.
0.8
32 0.6
0.4
0.2
Initial Letter (s) Correct (1C Responses)Final Letter (s) Correct (FC Responses)
_L _L _L
Dominant Nondominant Dominant Nondominant Dominant NondominantField Field Field Field Field Field
Group RN Group LI Group LN
Figure 5. The proportion of partial errors on the verbal test in which initial letters (1C)versus final letters (FC) were correct in the verbally dominant and verbally nondominantvisual fields for right- and left-handers with a noninvcrted writing posture (RN and LN, re-spectively) and for left-handers with an inverted posture (LI).

134 JKRRE LEVY AND MARYLOU REID
tion to the beginning of the next line to beread. If this interpretation is correct, thennative readers of Hebrew or Arabic shoulddisplay a Group X Field X Response Typeinteraction in the opposite direction from theone we observed : Group LN should manifestthe larger Field X Response Type interac-tion and Groups RN and LI, the smaller
Permutation errors. Permutation errorswere next considered in instances when allthree letters in the syllable were correctlyreported, but in a permuted order. Empiricaland theoretical considerations led us to ex-pect that the proportion of permutation er-rors, relative to total within-field errors,would be greater for the nondominant thandominant field.
Unreported analyses (by the first author)of data gathered from split-brain patientsshowed that when a vertical array of threeforms, consisting of permutations of Xs andsquares (see Levy, Trevarthen, & Sperry,1972), was displayed to the right and lefthemispheres and subjects were required tochoose a matching pattern, the right hemi-sphere was impaired on mixed patterns. Thatis, when the pattern contained both Xs andsquares, the right hemisphere made signifi-cantly more errors than the left, as comparedwith stimulus patterns consisting of threeXs or three squares. This suggests that theright hemisphere, although excellent at im-agistic encoding and memory for visual stim-uli having what we would call "visual syn-tax," is poor at preserving the spatial order-ing of stimulus features when this orderingis not constrained by the requirements ofmeaning, learned associations, or laws of Ge-stalt organization. Theoretically, given whatwe know of right and left hemisphere spe-cialization, it is not unreasonable to supposethat imagistic memory is specific for preserv-ing an invariant in a set of spatial relations,while verbal memory is specific for preserv-ing the ordering of stimulus features.
The dependent variable for this analysiswas the ratio of permutation errors to totalerrors within a visual field. For Groups RNand LI, the dominant verbal field was de-fined as the right visual field and for GroupLN, as the left visual field. There is no rea-son for supposing that any kind of biased
scanning effects should differentially affectthe groups on the permutation ratios. If,therefore, Group LI is less laterally differen-tiated than the other groups, their field effect,if one exists, should be smaller. Further, ifspatial functions are bilateralized in thisgroup, their ratio should be higher than thatof the other groups in the dominant field,but if verbal functions are bilateralized theirratio should be lower than that of the othergroups in the nondominant field.
We found a significant effect of field in thepredicted direction, F(\, 66) = 19.75, p <.001, the nondominant field producing ahigher ratio. In addition, there was a maineffect of group, F(2, 66) = 5.50, p < .025,with Group LI having a significantly smallerratio than Group LN and a nonsignificantlysmaller ratio than Group RN, Groups RNand LN not differing. A Group X Fieldeffect was also found, F(2, 66) = 22.00,p < .001, Groups RN and LN showingequally large between-field differences (p <.01 for both) and Group LI having no sig-nificant difference between fields. The groupsdid not differ in the dominant field, butGroup LI had a significantly smaller pro-portion of permutation errors than GroupsRN and LN in the nondominant field, sug-gesting bilateralization of verbal function inGroup LI, An analysis of the absolute fre-quency of permutation errors in the twofields yielded results identical to those withproportional permutation errors. Results aresummarized in Figure 6.
Other Indices of Laterality
Each subject was given a mean score onthe handeclness questionnaire which repre-sented his or her total score divided by 16. Amean score of 1 would indicate maximumdcxtrality; a mean score of 3, complete am-bilaterality; and a mean score of 5, maxi-mum sinistrality. The absolute value of thedifference in a subject's score and 3 was anindex of the degree of manual lateralization,ignoring direction. On this index, Group RNwas more manually lateralized than eitherGroup LI or LN, these latter two not differ-ing. Thus, our measure of the degree ofmanual lateralization did not reflect the de-

VARIATIONS IN CEREBRAL ORGANIZATION 135
gree of cerebral lateralization. We suspectthat the assessment of the degree of manualasymmetry by means of a questionnaire re-flects, in part, the operation of cultural fac-tors. Right-handers are positively reinforcedfor the use of the right hand and tend tothink of themselves as fully right-handed. Tncontrast, left-handers are negatively rein-forced for use of the left hand, quite probablyattempt many activities with the right hand,and tend to think of themselves as partiallyskilled with both hands. It is quite conceiva-ble that actual measurements of the relativeskill of the two hands would reflect the de-gree of hemispheric asymmetry.
The groups were also compared on anindex of familial handedness. The index wascomputed as follows: A parent or siblingwho was left-handed or ambidextrous wasgiven a weighting of 1, and a left-handed orambidextrous grandparent, a weighting of .5.These weightings were summed and dividedby the total number of family members re-ported. Group LN had a familial index of.153, Group LI an index of .127, and GroupRN an index of .095. Although the grouporder of these indices (highest for sinistralswith right hemisphere language, lowest fordextrals with left hemisphere language) isconsistent with that reported by Varney andBenton (1975), the indices do not differ sig-nificantly among groups. The familial indexwas uncorrelatecl with scores on the syllableand dot location tests when group member-ship was partialed out.
On tests of eye dominance, all Group RNsubjects except one were right-eye dominanton the Asher Test, though there were oc-casional discrepancies on the pointing tests.Among male subjects in Groups LI and LN,half were left-eye and half were right-eyedominant. Of female LI subjects, eight wereright-eye and four were left-eye dominant.In Group LN, seven females were right-eye and five were left-eye dominant. Thedistributions significantly differed amonggroups, X
2(2) = 11.92, p < .005, Group RNdiffering from LN and LI, these latter notdiffering. There was no significant sex effect.Eye dominance was uncorrelated with scores
m 0.06
"55IT 0.05i
.tt 0.04
Perm
uta
tion
Err
ors/
o
°
o2 8
S
/°q\ i
'• \ /h
\ /
Verbally Nondominant Field
RN LI LN
GroupsFigure 6. The ratio of permutation errors to totalerrors on the verbal test in the verbally dominantand nondominant fields for right- and left-handerswith a noninverted writing posture (RN and LN,respectively) and for left-handers with an invertedposture (LI).
on the syllable and dot location tests whengroup membership was partialed out.
On profile drawing, 20 out of 24 RN sub-jects drew left-facing profiles. Subject RIdrew a right-facing profile. In Group LI,18 subjects drew right-facing profiles and6 drew left-facing profiles. In Group LN, 16right-facing and 8 left-facing profiles weredrawn. A significant group effect on thedistribution, X
2 (2) = 19.17, p < .001, re-flected the fact that right-handed subjectsdrew significantly more left-facing profilesthan left-handed subjects, and there was nodifference in the distributions of Groups LIand LN. Thus, hand posture in writing didnot predict the direction of profile drawing.
Discussion
This study provides strong confirmationfor the proposal that an inverted hand pos-ture is indicative of a language hemisphereipsilateral to the dominant hand and that thetypical, noninverted posture is indicative ofthe usual contralateral relation. This associa-

136 J ERRE LEVY AND MARYLOU REID
tion held for left-handers with an invertedposture, for one right-hander with an in-verted posture, and for both handednessgroups with the noninverted posture. Thefailure up until the present time to discoverany simple relationship between the direc-tions of cerebral and manual lateralizationwould seem to be explained by the hand-position variable.
Although it is conceivable that in individ-uals with the ipsilateral relationship, thecontrol of the writing hand is mediated viacommissural pathways through the contra-lateral, visuospatial hemisphere, it is ex-tremely unlikely that the ultimate manualcontrol signals for writing actually originatein that half of the brain. In the first place,if language integration occurs in the ipsi-lateral hemisphere, then, almost by defini-tion, the contralateral hemisphere would beincapable of programming writing. Second,the verbally specialized hemisphere appearsto be also specialized for the patterning ofcomplex sequential movements of the hands,fdeational and ideomotor apraxias are diag-nostic of lesions to the verbal hemisphere(De Ajuriaguerra, Hccaen, & Angelergues,1960), and during verbalization in right-handers, automatic gesturing occurs moreoften with the right than with the left hand(Kimura, 1973a, 1973b). Finally, left hem-isphere lesions in left-handers which resultin aphasic disorders and other symptomsdiagnostic of damage to the language hem-isphere also produce agraphia (Hecaen,Angelergues, & Douzens, 1963). Clearly, ifwriting were programed by the right hem-isphere in such people, no agraphia shouldbe seen, One is therefore forced to the con-clusion that writing movements, except pos-sibly in cases of unusual pathology, alwaysderive from motor programs originating inthe verbally specialized hemisphere.
We do not know whether the expressionof these programs is mediated via the ipsi-lateral pyramidal tracts or via commissuralpathways in people with an ipsilateral lan-guage hemisphere-writing hand relation.However, we do not think that the observa-tions by Flechsig (1876) and Yakovlev andRadic (1966) of 190 fetal and neonatal
brains are irrelevant; of the 190 specimens,4.21% (P.02R - 1.85%, P.,,,5 = 7.3470) hadonly a minority of pyramidal fibers crossingin the medullary decussation. Unless fibersin the direct pyramidal tract decussate at thelevel of synapse, we would expect that peoplewith a larger ipsilateral than contralateralmotor tract could better use the former forthe control of fine movement. It is also ofinterest that in about 7.5% of 210 specimensexamined by Mizuno, Nakamura, and Oka-moto (1968), in addition to the usual lateraland ventral pyramidal tracts, a third pyra-midal bundle, the ventrolateral pyramidaltract of Barnes, was also present bilaterally.Barnes' tract consists of uncrossed, ipsi-lateral fibers.
Since approximately 7% of the populationhave language hemispheres ipsilateral to thewriting hand, if such individuals controlwriting movements directly through the un-crossed motor pathways, we would expectthat approximately the same proportion ofthe population might have an unusually largefraction of their motor fibers running ipsi-laterally. On the other hand, if control isexerted via the cerebral commissures, thesimilarity in the functional and anatomicalproportions must be purely coincidental.Further, if writing is mediated transhem-ispherically, the question of what neurolog-ical organization compels such a complexcontrol system suggests no theoreticallysimple answer.
Nevertheless, this possibility cannot becompletely ruled out. Heilman, Coyle,Gonyea, and Geschwind (1973) describe thecase of a left-handed man who wrote withthe right hand. The patient suffered a righthemisphere lesion in association with lefthemiplegia, a total absence of aphasic symp-toms, and apraxia and agraphia of the righthand. The authors concluded that languagewas organized in the left hemisphere, whilemanual skill was organized in the right, andthey attributed the apraxic and agraphicsymptoms of the right hand to the destruc-tion of the praxic programs of the right sideof the brain, programs which had formerlydirected movements of the right hand byway of the left hemisphere. In view of the

VARIATIONS IN CEREBRAL ORGANIZATION 137
left hemiplegia, it would be difficult to main-tain that in this patient the majority ofmotor fibers were uncrossed; if they hadbeen, a right hemiplegia would have beenexpected. It should be kept in mind, how-ever, that this unusual symptom complexpertains to a single patient, that this patientmay have had unsuspected, long-standingbrain damage which resulted in a peculiarneurological reorganization, and that in thecases of Hecaen et al. (1963) agraphia inleft-handers was associated with aphasia,while absence of agraphia was associatedwith constructional apraxia.
In our view, the possibility that some7% or so of people control the writing handvia uncrossed pyramidal tracts stands as aviable hypothesis, though Geschwind (1975)believes that all instances of ipsilateral con-trol involve only the proximal muscles andonly the extrapyramidal system. If Gesch-wind is correct, this system must possess thecapacity to control writing, since in at leasttwo split-brain patients, the left hemispherecan control writing movements of the lefthand (Levy, Nebes, & Sperry, 1971).
In addition to the association betweenhand posture and the direction of hemi-spheric lateralization found in this study, wealso discovered that the degree of lateraldifferentiation of the brain was stronglyrelated to the hand-posture variable. In con-trast to Levy's (1974) hypothesis, based onsuggestions from the neurological literature,that sinistrals with right hemisphere lan-guage are less well laterally differentiatedthan those with left hemisphere language,we found that left-handers with right hem-isphere language were well lateralizecl,while those with left hemisphere languagewere weakly lateralizecl. These results con-form with other findings on normal popula-tions (Satz et al., 1965).
The discrepancy between conclusionsdrawn from neurological patients and thosedrawn from studies of normal samples isdue, perhaps, to a conceptual error in inter-preting neurological findings. Typically, if apatient suffers a unilateral lesion associatedwith aphasia, it is concluded that the dam-aged hemisphere is dominant for language
functions. However, the frequency of aphasiain sinistrals following unilateral brain dam-age is much too high to justify such a con-clusion. In any sample of such patients, ifhalf suffer left brain damage and half sufferright brain damage, then regardless of theproportions with left and right hemisphericdominance for language, assuming these tobe approximately equal in the two lesiongroups, there should be no more than 50%of the entire group who display aphasicsymptoms if aphasia only occurs with alesion to the language-dominant hemisphere.As Levy (1974) has discussed, transientaphasia develops in a considerably higherpercentage. The conclusion which one mustreach is that a substantial fraction of left-handers suffer a transient aphasia regardlessof whether it is their verbally dominant orverbally nondominant hemisphere which isdamaged.
One would guess that this occurs in thoseindividuals who are partially bilaterized forlanguage, namely, according to our data, left-handers with their main language hemi-sphere on the left. If such an individualwere to suffer a right hemisphere lesion, heor she might well become aphasic; theaphasia would most likely be quite mild,and recovery almost certain. The fact ofthe aphasia would lead many, if not most,neurologists to conclude that the right hem-isphere was dominant for language, and themildness of symptoms and the rapid re-covery would lead them to conclude thatlateralization was quite weak. While thelatter conclusion would be correct, the for-mer would be wrong. In view of these con-siderations, we believe the data obtainedfrom normal populations with regard to thisissue give a more valid reflection of realitythan do data obtained from neurologicalsamples.
The question arises, however, as to whypartial bilateralization of function should beassociated with an ipsilateral relation be-tween the writing hand and the verbal hem-isphere. There are three other known con-ditions in which bilateralization of functionoccurs. First, in infantile hemiplegics inwhom one hemisphere is nonfunctional from

138 JERRE LEVY AND MARYLOU REID
birth, the remaining hemisphere has beenfound to display relatively normal functionin both the verbal and spatial realm (Basser,1962), Second, investigations of people withagenesis of the corpus callosurn (Jeeves,1972; Netley, 1977; Saul & Sperry, 1968)reveal partial bilateralization of function.Finally, in young children, in spite of thefact that recent studies show the infant hem-ispheres to be laterally specialized (Dennis& Kohn, 1975; Kohn & Dennis, 1974a,1974b; Wada, Clarke, & Hamm, 1975), bothsides of the brain appear to be involved inlanguage integration to a much larger extentthan is the case in adults (Brown, 1975).Before the age of 5, transient aphasias occurin children regardless of whether it is theleft or right hemisphere which has beendamaged.
These findings suggest rather stronglythat at birth each hemisphere possesses notonly a program for its primary specializa-tion but also a secondary program for thespecializations of the other side of the brainand that, with maturation, the secondaryprograms, under the influence of callosallymediated inhibition, arc either blocked indevelopment or regress in functional ca-pacity. If one hemisphere or the corpuscallosurn is nonfunctional or missing frombirth, developmental blockage or functionalregression of the secondary programs docsnot occur, and the consequence is partialbilateralization of function in adulthood.
Adult infantile hemiplegics typically showeither a rather large discrepancy in verbaland spatial abilities or a depression in bothsets of capacities (Dennis & Kohn, 1975;Kohn & Dennis, 1974a, 1974b; Sperry,1974; Wilson, 1970). People with callosalagenesis display similar patterns of perform-ance (Ettlinger et al., 1972; Ferriss & Dor-sen, 1975 ; Lehmann & Lampe, 1970; Sperry,1974), suggesting that the maximum amountof cognitive power of a hemisphere is fixedand can be allocated in varying amounts toverbal or spatial functions. If this inferenceis correct, then complete bilateralization ofboth verbal and spatial capacities shouldreduce a hemisphere's ability for both, whileverbal bilateralization should leave verbal
abilities intact while reducing spatial in-tegration, with spatial bilateralization pro-ducing the reverse.
In the present study, the performancelevels of ipsilateral subjects were signifi-cantly worse than those of others on eitherthe dot location test, the syllable test, or onboth. Subject RT, although performing ex-tremely well on the clot location test, scoredlower on the syllable test than any of theother 72 subjects. Three of the 12 LI malesobtained unusually high scores on the dotlocation test and had an unusually large dis-crepancy with their scores on the syllabletest. One LI male outperformed all except2 dextral males on the syllable test, butscored worse than all dextral males on theclot location test. A 5th LI male subject wasin the dextral male range on both tests, anda 6th was in the male dextral range on thespatial test but worse than all dextral maleson the verbal test. The remaining 6 sub-jects were all below the male dextral rangeon the dot location test, and 3 of these wereworse than all except 1 male dextral on thesyllable test. Thus, among male LI subjects,all except 1 showed either extreme dis-crepancies in scores on the two tests or arather striking depression in scores on oneor both tests. Among female LI subjects, 1outperformed all dextral females except 1on the dot location test but displayed an ex-treme discrepancy with her score on thesyllable test. The remaining 11 LI femaleswere worse on the dot location test than 11out of 12 RN females. On the syllable test,all 12 LI females were below the mean ofRN females, and 5 scored lower than allRN female subjects. One sees, therefore,tha# in both male and female subjects withan ipsilateral relation between the languagehemisphere and writing hand, the perform-ance patterns are very similar to those seenin people with callosal agenesis.
It is not unreasonable to suggest that insome individuals, and specifically in our LIand RI subjects, there may be a partialfailure during some critical stage of embryo-genesis for axons to cross the midline of thenervous system. If the corpus callosum andthe pyramidal decussation are developing

VARIATIONS IN CEREBRAL ORGANIZATION 139
at this time, there may be a partial dysgene-sis of the callosum, as well as incompletedecussation of the pyramidal fibers, leadingboth to greater-than-average bilateralizationof function and to ipsilateral hand control.In some cases of callosal agenesis, there is ageneralized failure of midline development(Francois, Eggermont, Evens, Logghe, &DeBock, 1973). In support of the view thatagenesis of the corpus callosum reflectsinterference with midline crossing of thedeveloping axons, rather than a failure ofaxogenesis, is the observation that in casesof agenesis the callosal axons do, in fact,develop, but instead of forming commissuralconnections, grow posteriorly, ipsilateral totheir cell bodies (Tennyson, 1970).
The phenomenon of mirror movements inhemiplegics and patients with callosal agene-sis, discussed previously, is of relevance herealso. As pointed out in the introduction, itwould seem that such mirror movementsmust be mediated through the ipsilateral andcontralateral motor tracts of one hemisphere,and it is of some interest that simultaneousmirror writing is most often reported inleft-handers (Gordon, 1920). The LewisCarroll example (Barsley, 1967), mentionedearlier, suggests that such mirror writersmay typically have an inverted hand posturewhen writing with one or the other hand;Carroll was left-handed, a mirror writer,and adopted the inverted posture whenwriting with the right hand. It might benoted also that the frequency of left-handed-ness is extremely high in cases of agenesisof the callosum: 1 of 2 patients described bySaul and Gott (1976); 2 of 4 patientsdescribed by Ferriss and Dorsen (1975),with a 3rd (patient B) being able to writewith the left hand; 2 of 4 patients withtotal agenesis tested by Ettlinger et al.(1972), with a 3rd patient (T3) beingambilateral; and 2 of 9 patients of Lehmannand Lampe (1970), giving a total of 9 left-handed or ambilateral patients out of the 19examined (47%).
We recognize that our suggestion thatadequately functioning, normal college stu-dents may have partial dysgenesis of thecorpus callosum appears to be rather radical.
However, even total agenesis of the corpuscallosum, as Ferris and Dorsen (1975, p. 95)point out, "has been found in some individ-uals who function well in response to theordinary demands of life, and specific testsare required to measure subtle responsedifferences between them and normal in-dividuals to less obvious demands which areneither frequent nor crucial." Ettlinger,Blakemore, Milner, and Wilson (1974) re-port that patients with total callosal agenesisdo not even show cross-hemispheric match-ing defects. We might add, also, that inpatients with surgical division of all the neo-cortical commissures, including the anteriorcommissure, the symptoms are so slight thatit was not until the early 1960s that a "split-brain syndrome" could be identified, in spiteof diligent searching by earlier investigators(Akelaitis, 1943, 1944; Akelaitis, Risteen,Herren, & Van Wagenen, 1942). Clearly, ifcomplete absence of the callosum is not in-compatible with relatively normal function,even fewer symptoms would be expected inpeople with partial dysgenesis.
In addition to differences in the degreeof cerebral lateralization associated withhandedness and hand position, we found, asothers have reported (Kimura, 1969; Lans-dell, 1962; McGlone & Kertesz, 1973), con-sistent sex differences within all groups. Tosome extent on the syllable test and to amuch larger extent on the dot location test,females in Groups RN and LI displayedsmaller field differences than males. In con-trast, females in Group LN and Subject RIhad equally small field differences on thesyllable and dot location tests compared toGroup LN males and, further, absolute fielddifferences for these females were larger onthe spatial than verbal test. In general, theperformance scores of females were alsolower, but this may well be due to a gen-eralized superiority of males at processingvisual, as opposed to auditory, information(Chandra, 1972; Farrimond, 1967; Kubo,1928; Kimmel & Kimmel, 1965; Waag,Halcomb, & Tyler, 1973). Consistent withsuch an interpretation is the fact that theexposure thresholds for the simple dot de-tection task used to determine exposure

140 JERRE LEVY AND MARYLOU REID
durations on the dot location test werelower for males than females, i(70) = 1.70,/> < .05, one-tailed test.
The smaller field differences of femaleshave been interpreted by others as reflectingbilateralization of the measured function.However, although small field differences onsome test may reflect better-than-expectedperformance in the nondominant hemisphere(functional bilateralization), they can alsoreflect worse-than-cxpected performance inthe dominant hemisphere (incomplete spe-cialization). In fact, on theoretical grounds,bilateralization of, for example, verbal func-tion, ought to produce incomplete special-ization of the spatially dominant hemisphere.One would expect a significant, positive cor-relation between the magnitudes of field dif-ferences on a verbal and spatial test, but notnecessarily because both functions are bi-lateralized. One function may be bilateral-ized, producing incomplete specialization ofthe other.
When the results of the superior field-in-ferior field analysis are considered, wherethe nature of sex effects can be seen mostclearly, males surpassed females in bothfields on the syllable test, but the Sex XField interaction was significant, showing agreater male superiority in the superior thanin the inferior field. The same results wereseen on the dot location test: Males sur-passed females in both fields, the differencein sexes being greater in the superior thanin the inferior field. Though the reducedfield differences for females on both tests,as compared to males, was due to a depres-sion in dominant field performance, suggest-ing incomplete specialization rather thanbilateralization, we do not believe this is thewhole story. If the overall female inferiorityis due, as we suggested, to an advantage formales in visual processing, the performancesof females in their nondominant fields, beingbarely lower than those of males, may ac-tually reflect relatively enhanced perform-ance.
In females with left hemisphere language(Groups RN and LI), verbal performanceexceeded spatial performance, and field dif-ferences were smaller on the spatial test. Infemales with right hemisphere language
(Group LN and Subjects RI), spatial per-formance exceeded verbal performance, andfield differences were smaller on the verbaltest. Though the Task X Group effect wasnot significant, these associations appearedin all three groups. Of interest, also, is thefact that males in all three groups displayedthe opposite task superiority to females, anda male RI subject, tested on the WechslerAdult Intelligence Scale some years ago bythe first author, had a 35-point superiorityon verbal IQ as compared to performanceIQ. The consistent finding, in spite of non-significance, that females performed best intasks specialized to the left hemisphere,while males performed best in tasks special-ized to the right hemisphere, suggests thatleft hemisphere functions in females tend tobilateralize, producing incomplete special-ization of the right.
Recent data from Reid (Note 3) offerconfirmation for the idea that the left hem-isphere in females, regardles of what it pro-grams, is more highly specialized than theright, while the right hemisphere in malestends to greater specialization.
Reid developed two tactile tests of lateral-ization : one requires the visual selection of anonsense shape felt by the left or right handand assessing specialties of the spatial hem-isphere, and one requires a same-differentjudgment of two temporal patterns stimu-lating the right or left hand and assessingspecialties of the verbal hemisphere. Boysand girls of ages 5 and 8, 40 with right hem-isphere language (left-handed with a non-inverted posture) and 80 with left hemi-sphere language (noninverted right-handersand inverted left-handers), were tested onboth tests. In the older children, noninvertedleft-handers (LN) had a right-hand supe-riority on the nonsense shape test and a left-hand superiority on the temporal patterntest. Inverted left-handers (LI) and non-inverted right-handers (RN) displayed thereverse, though, as in our adult subjects,left-right differences were small in the in-verted left-handers. Of major interest werethe results in the younger children. In girlswith left hemisphere language, performancewas poor on the nonsense shape test andthere was no hand difference, but on the

VARIATIONS IN CEREBRAL ORGANISATION 141
temporal pattern test, a clear right-handsuperiority emerged. In girls with left hem-isphere spatial function (Group LN) , per-formance was poor on the temporal patterntest and there was no hand difference, buton the nonsense shape test, a significantright-hand superiority was seen. Boys inboth groups manifested precisely oppositeresults. Reid's data leave little doubt thatthe left hemisphere matures earlier than theright in girls and that the right hemispherematures earlier than the left in boys, regard-less of the specialties of the hemispheres.
We would guess that the earlier maturinghemisphere gains a functional advantagewhich is maintained throughout develop-ment and adulthood. It is quite possible,also, that the function showing earlier ma-turation receives greater reinforcement fromthe environment, both because the child mayseek out environmental experiences whichexercise his or her newly acquired skills andbecause adults and other children may pro-vide such experiences. That the postnatal en-vironment cannot be the sole source of male-female differences is almost certain in viewof the reversal of the typical cognitive pat-terns associated with sex in children withmirror-reversed functions of the cerebralhemispheres.
Although a recent report by Waber (1976)suggests that the neuropsychological organ-izations of males and females are a functionof the age of sexual maturation, since thesedifferences appear in children by age 5, wewould suggest that the age of puberty is notin itself causative but is, rather, a correlateof causative events occurring during prenataldevelopment—events, we suspect, that arerelated to the levels of various fetal sexhormones. In girls with Turner's Syndrome,a chromosomal abnormality in which onlyone X chromosome is present, there is atotal dysgenesis of the gonads and a totalabsence of gonadal hormones. Verbal abilityis at least average in these girls while spatialability is greatly depressed (Duane Alex-ander, Ehrhardt, & Money, 1966). It is quiteconceivable that absence of gonadal hormonesduring fetal development produces extremebilaterali nation of left hemisphere functionsand a profound interference with right hem-
isphere specialization. Indeed, Gordon (Note4) and Netley (1977) recently applied ver-bal dichotic lateralization tests to womenwith Turner's Syndrome, concluding fromtheir results that verbal functions werestrongly bilateralized. It is well known thatabsence of fetal sex hormones is associatedwith the development of female body mor-phology and female behavior, while the pres-ence of androgens, as well as progesterone,is associated with the development of malebody morphology and male behavior (Money& Ehrhardt, 1972). Since a strong degreeof lateral differentiation and a highly special-ized right hemisphere are characteristics ofmales, it may be that the extent to whichthese traits are displayed is also a functionof the levels of fetal sex hormones.
The various findings in the present in-vestigation suggest that many of the in-dividual differences in cognitive organiza-tion can be directly related to the directionand degree of cerebral lateralization and tothe extent to which each cerebral hemisphereis specialized for its particular primary func-tion. These differences, in turn, appear to beclosely associated with handedness, handposture in writing, and sex. Clearly, moreresearch is required to adequately specifythe relations between cognitive and neuro-logical patterns and to determine the mech-anisms responsible for the relations. How-ever, the results of the present study andthose of Reid are sufficiently orderly toprovide encouragement in this endeavor.
Reference Notes
1. Gur, R. C., & Gur, R. E. Personal communica-tion, September 1974.
2. Olttnan, P. K. Personal communication, May1970.
3. Reid, Marylou. Cerebral lateralisation in chil-dren: An ontogenetic and organismic analysis.Manuscript in preparation, 1978.
4. Gordon, H. Personal communication, October1976.
References
Akelaitis, A. J. Studies on the corpus callosum:VII. Study of language functions (tactile andvisual lexia and graphia) unilaterally followingsection of the corpus callosum. Journal of Ncu-ropathology and Experimental Neurology, 1943,3, 226-262.

142 JERRE LEVY AND MARYLOU REID
Akelaitis, A. J. A study of gnosis, praxis andlanguage following section of the corpus callosumand anterior commissure. Journal of Neuro-surgery, 1944, 1, 94-101.
Akelaitis, A. J., Risteen, W. A., Herren, R. Y.,& Van Wagenen, W. P. Studies on the corpuscallosum: III. A contribution to the study ofdyspraxia and apraxia following partial and com-plete section of the corpus callosum. Archivesof Neurology and Psychiatry, 1942, 47, 971-1007.
Annett, M. A. A model of the inheritance ofhandedness and cerebral dominance. Nature,1964, 204, 59-60.
Bakan, P., Dibb, G., & Reed, P. Handedness andbirth stress. Neuropsychologia, 1973, 3, 363-366.
Barsley, M. The other hand. New York: Haw-thorn Books, 1967.
Basser, L. S. Hemiplegia of early onset and thefaculty of speech with special reference to theeffects of hemispherectomy. Brain, 1962, 85, 427-460.
Beaumont, J. G. Handedness and hemisphere func-tion. In S. J. Dimond & J. G. Beaumont (Eds.),Hemisphere function in the human brain. NewYork: Wiley, 1974.
Berlin, C. I., & Cullen, J. K., Jr. Acoustic problemsin dichotic listening tasks. In S. J. Segalowitz& F. A. Gruber (Eds.), Language developmentand neurological theory. New York: AcademicPress, 1977.
Brain, W. R. Diseases of the nervous system (6thed.) London: Oxford University Press, 1962.
Broca, P. Remarques sur le siege de la faculte dulangage articule suives d'une observationd'aphemie. Bulletin Societie Anatomique [Paris],1861, 6, 330-357.
Brown, J. The neural organization of language:Aphasia and neuropsychiatry. In S. Arieti(Ed.), American handbook of psychiatry (Vol.4). New York: Basic Book, 1975.
Bryden, M. P. Tachistoscopic recognition, handed-ness and cerebral dominance. Neuropsychologia,1965, 3, 1-8.
Chaco, J., & Blank, A. Mirror movements inhemiparesis. Confinia Neurologica, 1974, 36, 1-4.
Chandra, S. An assessment of perceptual acuity inFiji: A cross-cultural study with Indians andFijians. Journal of Cross-Cultural Psychology1972, 3, 401-406.
Cohen, G. Hemispheric differences in a letterclassification task. Perception & Psychophysics1972, 11, 139-142.
Crovitz, H. F. On direction in drawing a person.Journal of Consulting Psychology, 1962, 26, 196.
Curry, F. K. W. A comparison of left-handed andright-handed subjects on verbal and nonverbaldichotic listening tasks. Cortex, 1967, 3, 343-352.
Curry, F. K. W., & Rutherford, D. R. Recognitionand recall of dichotically presented verbal stimuliby right and left handed persons. Neuropsy-chologia, 1967, 5, 119-126.
Dax, M. Lesions de la moitie gauche de 1'encephalecoincident avec 1'oubli des signes de la pensee.
Gazette Hebdomadaire de Medecine et deChirurgie, 1865, 2(2 erne serie), 2. (Read atMontpellier in 1836.)
De Ajuriaguerra, J., Hecaen, H., & Angelergues,R. Les apraxies: Varietes cliniques et lateralisa-tion lesionnelle. Revue Neurologique, 1960, 102,566-594.
Dennis, M., & Kohn, B. Comprehension of syntaxin infantile hemiplegics after cerebral hemide-cortication: Left-hemisphere superiority. Brainand Language, 1975, 5, 472-482.
Dimond, S., & Beaumont, J. G. (Eds.) Hemispherefunction in the human brain. New York: Wiley,1974.
Duane Alexander, B. S., Ehrhardt, A. A., &Money, J. Defective figure drawing, geometricand human, in Turner's syndrome. The Journalof Nervous and Mental Disease, 1966, 142, 161-167.
Eccles, J. C. Brain, speech, and consciousness.Die Naturwissenschaften, 1973, 60, 167-176.
Enstrom, E. A. The little turn that makes the bigdifference. Elementary English, 1966, 43, 865-868.
Enstrom, E. A. Writing hints for the strugglinglefty. Instructor, 1971, 81, 155.
Ettlinger, G., Blakemorc, C. B., Milner, A. D., &Wilson, J. Agenesis of the corpus callosum:A behavioural investigation. Brain, 1972, 95,327-346.
Ettlinger, G., Blakemore, C. B., Milner, A. D., &Wilson, J. Agenesis of the corpus callosum:Afurther behavioural investigation. Brain, 1974, 97,225-234.
Farrimond, T. Visual and auditory performancevariations with age: Some implications. Aus-tralian Journal of Psychology, 1967, 19, 193-201.
Ferriss, G. S., & Dorsen, M. M. Agenesis of thecorpus callosum: I. Neuropsychological studies.Cortex, 1975, 11, 95-122.
Flechsig, P. Die Leitungsbahnen in Gehirn undRiickenmark des Menschen. Leipzig: W. Engel-mann, 1876.
Francois, J., Eggermont, E., Evens, L., Logghc, N.,& DeBock, F. Agenesis of the corpus callosum,the median facial cleft syndrome and associatedocular malformations. American Journal ofOphthalmology, 1973, 76, 241-245.
Geschwind, N. The apraxias: Neural mechanismsof disorders of learned movement. AmericanScientist, 1975, 63, 188-195.
Goodglass, H., & Quadfasal, F. A. Languagelaterality in left handed aphasics. Brain, 1954,77, 521-548.
Gordon, H. Left-handedness and mirror-writing,especially among defective children. Brain, 1920,43, 313-367.
Green, J. B. An electromyographic study of mirrormovements. Neurology, 1967, 17, 91-94.
Harris, L. J. Sex differences in spatial ability:Possible environmental, genetic, and neurologicalfactors. In M. Kinsbourne (Ed.), Hemispheric

VARIATIONS IN CEREBRAL ORGANIZATION 143
asymmetries of function. Sprinfield, 111.: CharlesC Thomas, 197S.
Harshman, R. A. Sex, language, and the brain:Adult sex differences in lateralization. In D. O.Walter, L. Rogers, & J. M. Finzi-Fried (Eds.),Conference on human brain function: Brain in-formation service. Los Angeles: BRI Publica-tions Office, University of California, 1976.
Hecaen, H., Angelergues, R., & Douzens, L. A.Les agraphies. Neuropsychologia, 1963, 1, 179-208.
Hecaen, H., & De Ajuriaguerra, J. Les gauchers.Prevalence manuelle et dominance cerebrate.New York: Grune & Stratton, 1964.
Hecaen, H., & Sauguet, J. Cerebral dominance inleft-handed subjects. Cortex, 1971, 7, 19-48.
Heilman, K. M., Coyle, J. M., Gonyea, E. F., &Geschwind, N. Apraxia and agraphia in a left-hander. Brain, 1973, 96, 21-28.
Hudson, P. T. W. The genetics of handedness—A reply to Levy and Nagylaki. Neuropsychol-ogia, 1975, 13, 331-339.
James, W. E., Mifferd, R. B., & Wieland, B.Repetitive psychometric measures: Handednessand performance. Perceptual and Motor Skills,1967, 25, 209-212.
Jeeves, M. A. Further studies of the effects ofagenesis of the corpus callosum in man andneonatal sectioning of the corpus callosum inanimals. In J. Cernacek & F. Podivinsky (Eds.),Cerebral interhemispheric relations. Bratislava:Vydatelstvo Slovenskej Akademie Vied, 1972.
Kimmel, H. D., & Kimmel, E. Sex differences inthe adaptation of the GSR under repeated ap-plications of a visual stimulus. Journal of Ex-perimental Psychology, 1965, 70, 536-537.
Kimura, D. Dual functional asymmetry of thebrain in visual perception. Neuropsychologia,1966, 4, 275-285.
Kimura, D. Functional asymmetry of the brain indichotic listening. Cortex, 1967, 3, 163-173.
Kimura, D. Spatial localization in left and rightvisual fields. Canadian Journal of Psychology,1969, 23, 445-458.
Kimura, D. Manual activity during speaking: I.Right-handers. Neuropsychologia, 1973, 11, 45-50. (a)
Kimura, D. Manual activity during speaking: II.Left-handers. Neuropsychologia, 1973, 11, 51-55. (b)
Kintsch, W. Learning, memory, and conceptualprocesses. New York: Wiley, 1970.
Knox, C., & Kimura, D. Cerebral processing ofnonverbal sounds in boys and girls. Neuro-psychologia, 1970, 8, 227-238.
Kohn, B., & Dennis, M. Selective impairments ofvisuo-spatial abilities in infantile hemiplegicsafter right cerebral hemidecortication. Neuro-psychologia, 1974, 12, 505-512. (a)
Kohn, B., & Dennis, M. Somatosensory functionsafter cerebral hemidecortication for infantilehemiplegia. Neuropsychologia, 1974. 12, 119-130. (b)
Kubo, Y. On the results of the discriminationtests. Jido Kenkyu-sho Kuyo [Transactions ofthe Institute for Child Study], 1928, 9, 1-14.
Lansdell, H. A sex difference in effect of temporal-lobe neurosurgery on design preference. Nature,1962, 194, 852-8S4.
Lehmann, H. J., & Lampe, H. Observations on theinterhemispheric transmission of information in9 patients with corpus callosum defect. EuropeanNeurology, 1970, 4, 129-147.
Levy, J. Possible basis for the evolution of lateralspecialization of the human brain. Nature, 1969,224, 614-615.
Levy, J. Lateral specialization of the human brain.Behavioral manifestations and possible evolu-tionary basis. In J. Kiger (Ed.), The biology ofbehavior. Corvallis: Oregon State UniversityPress, 1973.
Levy, J. Psychobiological implications of bilateralasymmetry. In S. Dimond & J. G. Beaumont(Eds.), Hemisphere function in the human brain.New York: Wiley, 1974.
Levy, J. Cerebral lateralization and spatial ability.Behavior Genetics, 1976, 6, 171-188.
Levy, J. A reply to Hudson regarding the Levy-Nagylaki model for the .genetics of handedness.Neuropsychologia, 1977, 15, 187-190.
Levy, J., & Nagylaki, T. A model for the geneticsof handedness. Genetics, 1972, 72, 117-128.
Levy, J., Nebes, R. D., & Sperry, R. W. Expres-sive language in the surgically separated minorhemisphere. Cortex, 1971, 7, 49-58.
Levy, J., & Reid, M. Variations in writingposture and cerebral organization. Science, 1976,194, 337-339.
Levy, J., & Trevarthen, C. Meta-control of hem-ispheric function in human split-brain patients.Journal of Experimental Psychology: HumanPerception and Performance, 1976, 2, 299-312.
Levy, J., & Trevarthen, C. Perceptual, semantic,and phonetic aspects of elementary languageprocesses in split-brain patients. Brain, 1977, 100,105-118.
Levy, J., Trevarthen, C., & Sperry, R. W. Percep-tion of bilateral chimeric figures following hem-ispheric deconnexion. Brain, 1972, 95, 61-78.
Luria, A. R. [Traumatic aphasia]. The Hague:Mouton, 1970. (Originally published, 1947).
Marie, P., & Foix, C. Les syncinesies des hem-iplegiques. Revue Neurologique, 1916, 30, 145-161.
McGlone, J., & Kertesz, A. Sex differences incerebral processing of visuospatial tasks. Cortex,1973, 9, 313-320.
McKeever, W. F., & Huling, M. D. Bilateraltachistoscopic word recognition as a function ofhemisphere stimulated and interhemispherictransfer time. Neuropsychologia, 1971, 9, 281-288.
Miller, E. Handedness and the pattern of humanability. British Journal of Psychology, 1971, 62,111-112.
Milner, B. The memory defect in bilateral hip-

144 JERRE LEVY AND MARYLOU REID
pocampal lesions. Psychiatric Research Reports,1959, 11, 43-52.
Milner, B. Amnesia following operation on thetemporal lobes. In C. W. M. Whitty & O. L.Zangwill (Eds.), Amnesia. London: Butter-worth, 1966.
Milner, B., Branch, C., & Rasmussen, T. Evidencefor bilateral speech representation in some non-right-handers. Transactions of the AmericanNeiirological Association, 1966, 91, 306-308.
Milner, B., & Teuber, H. L. Alteration of percep-tion and memory in man : Reflections on methods.In L. Weiskrantz (Ed.), Analysis of behavioralchange. New York: Harper, 1968.
Mizuno, N., Nakamura, Y., & Okamoto, M. Thefrequency of occurrence of Barnes' ventrolateral'pyramidal tract. Journal of Comparative Neu-rology. 1968, 134, 1-8.
Money, J., & Ehrhardt, A. A. Man, woman, boyand girl. Baltimore: The Johns Hopkins Uni-versity Press, 1972.
Nebes, R. D. Handedness and the perception ofwhole-part relationship. Cortex, 1971, 7, 350-356.
Netley, C. Dichotic listening of callosal agenesisand Turner's Syndrome patients. In S. J. Segal-owitz & F. A. Gruber (Eds.), Language de-velopment and neurological theory. New York:Academic Press, 1977.
Peterson, J. M., & Lansky, L. M. Left-handednessamong architects: Some facts and speculation.Perceptual and Motor Skills, 1974, 38, 547-550.
Roberts, L. Aphasia, apraxia and agnosia in ab-normal states of cerebral dominance. In P. J.Vinken & G. W. Bruyn (Eds.), Handbook ofclinical neurology (Vol. 4). Amsterdam: North-Holland, 1969.
Rudel, R. G., Denckla, M. B., & Spalten, E. Thefunctional asymmetry of Braille letter learningin normal, sighted children. Neurology, 1974, 24,733-738.
Satz, P., Achenbach, K., Pattishall, E., & Fennell,E. Order of report, car asymmetry and handed-ness in dichotic listening tasks. Cortex, 1965, 1,377-396.
Saul, R. E., & Gott, P. S. Language and speechlateralization by amytal and dichotic listeningtests in agenesis of the corpus callosum. In D. O.Walter, L. Rogers, & J. M. Minzi-Fried (Eds.),Conference on human brain junction: Braininformation service. Los Angeles: BRI Publica-tions, University of California, 1976.
Saul, R., & Sperry, R. W. Absence of com-missuratomy symptoms with agenesis of thecorpus callosum. Neurology, 1968, 18, 307.
Scoville, W. B., & Milner, B. Loss of recent mem-ory after bilateral hippocampal lesions. Journalof Neurology, Ncurositrgery, and Psychiatry,1957, 20, 11-21.
Shanon, B. Writing positions in Americans andIsraelis. Neuropsychologia, in press.
Silver-man, A. J., Adevai, G., & McGough, W. E.Some relationships between handedness and
perception. Journal of Psychosomatic Research,1966, 10, 151-158.
Sperry, R. W. Lateral specialization in the sur-gically separated hemispheres. In F. O. Schmitt& F. G. Worden (Eds.), The neurosciencesthird study program. Cambridge, Mass.: MITPress, 1974.
Sperry, R. W., Gazzaniga, M. S., & Bogen, J. E.Intel-hemispheric relationships. The neocorticalcommissures; syndromes of hemispheric discon-nection. In P. J. Vinken & G. W. Bruyn (Eds.),Handbook of clinical neurology (Vol. 4).Amsterdam: North-Holland, 1969.
Subirana, A. Handedness and cerebral dominance.In P. J. Vinken & G. W. Bruyn (Eds.), Hand-book of clinical neurologv (Vol. 4). Amster-dam: North-Holland, 1969.
Tennyson, V. M. The fine structure of the develop-ing nervous system. In W. Himwich (Ed.), De-velopmental neurobiology. Springfield, 111.:Charles C Thomas, 1970.
Varney, N. R., & Benton, A. L. Tactile perceptionof direction in relation to handedness and familialhandedness. Neuropsychologia, 1975, 13, 449-454.
Waag, W. L., Halcomb, C. G., & Tyler, D. M.Sex differences in monitoring performance. Jour-nal of Applied Psychology, 1973, 58, 272-274.
Waber, D. P. Sex differences in cognition: Afunction of maturation rate? Science, 1976, 192,572-574.
Wada, J. A., Clarke, R., & Hamm, A. Cerebralhemispheric asymmetry in humans. Corticalspeech zones in 100 adult and 100 infant brains.Archives of Neurology, 1975, 32, 239-246.
Wahl, C. R. Left-handed writers. Instructor, 1955,64, 85.
Wernicke, C. Der aphasische Symptomenkomplex.Breslau: Cohn & Weigert, 1874.
White, M. J. Laterality differences in perception:A review. Psychological Bulletin, 1969, 72, 387-405.
Wilson, P. J. E. Cerebral hemispherectomy forinfantile hemiplegia: A report of 50 cases.Brain, 1970, 93, 147-180.
Witelson, S. F. Hemispheric specialization forlinguistic and nonlinguistic tactual perceptionusing a dichotomous stimulation technique. Cor-tex, 1974, 10, 13-17.
Witelson, S. Sex and the single hemisphere. Righthemisphere specialization for spatial processing.Science, 1976, 193, 425-427.
Yakovlev, P. I., & Radic, P. Patterns of decussa-tion of bulbar pyramids and distribution ofpyramidal tracts on two sides of the spinal cord.Transactions of the American Neurological As-sociation, 1966, 91, 366-367.
Zurif, E. B., & Bryden, M. P. Familial handednessand left-right difference in auditory and visualperception. Neuropsychologia, 1969, 7, 179-187.
Received April 5, 1977 •
View publication statsView publication stats