vav proteins as signal integrators for multi-subunit immune-recognition receptors
TRANSCRIPT

476 | JULY 2002 | VOLUME 2 www.nature.com/reviews/immunol
R E V I E W S
networks that interact in complex and poorly under-stood ways to integrate the cell cycle, actin cytoskeleton,gene transcription and cell motility.
Elucidating the mechanisms by which receptor-mediated tyrosine-kinase activation is coupled to thesedisparate functional outcomes is an important area ofinvestigation. One class of signalling molecules that hasthe potential to orchestrate these different processes isthe VAV family of guanine-nucleotide exchange factors(GEFs). The importance of Vav proteins in the genera-tion and maturation of receptor-regulated effector func-tions is evident from genetic studies in mice (TABLE 1).Moreover, recent studies in cell lines that lack specificVav isoforms have revealed previously unrecognizedroles for these proteins in the regulation of cell survivaland the function of the signalling complex at the acti-vated MIRR (the SIGNALOSOME). Taken together, thesedata highlight the importance of VAV-family membersfor the regulation of haematopoietic-cell signalling.
The VAV familyVAV proteins belong to the DBL family of RHO–GEFs,of which there are now more than forty members1.Three structurally related VAV genes have been identi-fied in mammals2. VAV1 is expressed primarily in cellsof the haematopoietic system, whereas VAV2 and VAV3
Protective immunity requires interaction between com-ponents of the innate and acquired immune systems in such a way as to ensure that subsequent exposure tothe same antigen elicits a robust and decisive defence.Antigen-mediated engagement of specific multi-subunit immune-recognition receptors (MIRRs) on thecell surface of lymphocytes activates signalling cascades.These result in either the deletion of self-reactive cells orthe proliferation of non-self-reactive cells, which leadsto the generation of effector and memory functions thatdefine the immune response. Typically, these responsesinvolve changes in cellular proliferation, differentiation,lifespan and anatomical localization. Antigen receptorsdo not have intrinsic enzymatic function, but they areassociated with non-polymorphic subunits that includeimmunoreceptor tyrosine-based activation motifs(ITAMs). After engagement of the antigen receptor,the ITAMs are tyrosine phosphorylated by activatedSRC-family protein-tyrosine kinases (PTKs). Thephosphorylated ITAMs serve as docking sites for thetandem SRC-homology 2 (SH2)-domain-containingPTKs SYK and ZAP70 (ζ-chain-associated proteinkinase, 70 kDa), which are then activated by means oftyrosine phosphorylation by SRC-family PTKs. The acti-vation of SYK and ZAP70 leads to the phosphorylationof several molecules, which, in turn, activate signalling
*Lymphocyte Signalling andDevelopment Laboratory,Molecular ImmunologyProgramme, The BabrahamInstitute, Babraham,Cambridge CB2 4AT, UK.‡Division of DevelopmentalOncology Research and theDepartment of Immunology,Mayo Clinic, Rochester,Minnesota 55905, USA.e-mails: [email protected];[email protected] doi:10.1038/nri840
VAV PROTEINS AS SIGNALINTEGRATORS FOR MULTI-SUBUNITIMMUNE-RECOGNITION RECEPTORSMartin Turner* and Daniel D. Billadeau‡
In recent years, substantial progress has been made towards the identification of intracellularsignalling molecules that couple multi-subunit immune-recognition receptors (MIRRs) to theirvarious effector functions. Among these, the VAV proteins have been observed to have a crucialrole in regulating some of the earliest events in receptor signalling. VAV proteins function, in part,as guanine-nucleotide exchange factors (GEFs) for the RHO/RAC family of GTPases. This reviewfocuses on the role of VAV proteins in the regulation of lymphocyte development and function,and emphasizes the regulatory roles that these proteins have through both GEF-dependent and -independent mechanisms.
SIGNALOSOME
A putative, stable signallingcomplex, which consists ofBTK, BLNK, BCAP,VAV1,VAV2,PLC-γ2 and PI3K, that isproposed to regulate the level of intracellular calcium andsubsequent downstream events.
NATURE REVIEWS | IMMUNOLOGY VOLUME 2 | JULY 2002 | 477
R E V I E W S
increased oncogenic potential and enhanced transcrip-tional regulation10–12. Although these data might indi-cate that the mutated VAV proteins fail to recruit anegative regulator of VAV function, the results areprobably the result of failure of the phenylalanineresidue to stabilize the autoinhibitory interaction thatis required to maintain VAV1 in an inactive state.
VAV proteins have been shown to interact directly,by means of their SH2 domain, with several receptortyrosine kinases13 and co-receptors, and indirectly withactivated receptors, by binding to adaptor proteins13,14.Several tyrosine-phosphorylated molecules have beenshown to interact with the SH2 domain of VAV pro-teins, including the adaptor proteins SLP76 (SH2-domain-containing leukocyte protein of 76 kDa) in T cells15,16 and B-cell linker (BLNK; SLP65/BASH) in B cells17. The importance of SLP76 in mediating thephosphorylation of VAV1 is highlighted by the observa-tion that thymocytes that are reconstituted with amutant form of SLP76 — in which tyrosines 112, 128and 145 have been mutated — are unable to phospho-rylate VAV1 after T-cell receptor (TCR) crosslinking18.Co-receptors, such as CD28 on T cells and CD19 on B cells, also participate in the activation of VAV proteins.CD28 engagement19,20 or CD3–CD28 crosslinking21
enhances the tyrosine phosphorylation and membranerecruitment of VAV1 (REF. 22), although it seems that theVAV SH2 domain might not interact physically withCD28. By contrast, both VAV1 and VAV2 have beenshown to interact, by means of their SH2 domain, withthe tyrosine phosphorylated Tyr391-Glu-Glu-Promotif in the cytoplasmic tail of CD19 (REFS 23–25). B-cellreceptor (BCR) and CD19 co-ligation is strongly syn-ergistic for the induction of VAV1 and VAV2 tyrosinephosphorylation24,25, and VAV1 is poorly phosphory-lated after activation of CD19-deficient B cells26.
have broader patterns of expression. DBL-homology(DH)-domain-containing GEFs promote the activationof RHO–GTPases by catalysing the exchange of GDPfor GTP. RHO–GTPases are important components ofintracellular signalling pathways in many cells; theyaffect diverse cellular responses, such as the regulation ofgene transcription, cell proliferation, survival, migrationand differentiation3. In addition to the DH domain —which mediates GEF activity — VAV proteins containdomains that are also found in other intracellular sig-nalling molecules (BOX 1), including adaptor proteins.So, as discussed later, VAV proteins might also regulatesignalling cascades in a GEF-independent manner.
Coupling VAV to activated receptorsThe recruitment of VAV proteins to activated receptorson lymphocytes and their subsequent tyrosine phos-phorylation require an intact SH2 domain. Several tyro-sine kinases, including LCK, FYN, HCK and SYK, havebeen shown to phosphorylate the VAV family ofGEFs4–8. As the GEF activity of VAV proteins is activatedby tyrosine phosphorylation, the recruitment of VAVproteins to MIRRs provides a causal link between theactivation of PTKs by MIRRs,VAV tyrosine phosphory-lation and the regulation of RHO/RAC GTPases.Recently, nuclear magnetic resonance (NMR) structuralstudies have indicated the mechanism by which tyrosinephosphorylation regulates VAV GEF activity9. It wasshown that, when it is unphosphorylated, tyrosine 174within the acidic region of VAV1 interacts with a pocketwithin the DH domain, which inhibits GEF activity bypreventing the DH-domain–GTPase interaction (FIG. 1).Phosphorylation of Tyr174 disrupts this interaction,thereby restoring GEF activity. Mutation of this andother regulatory tyrosines in the acidic region of VAV1to phenylalanine results in a hyperactive protein that has
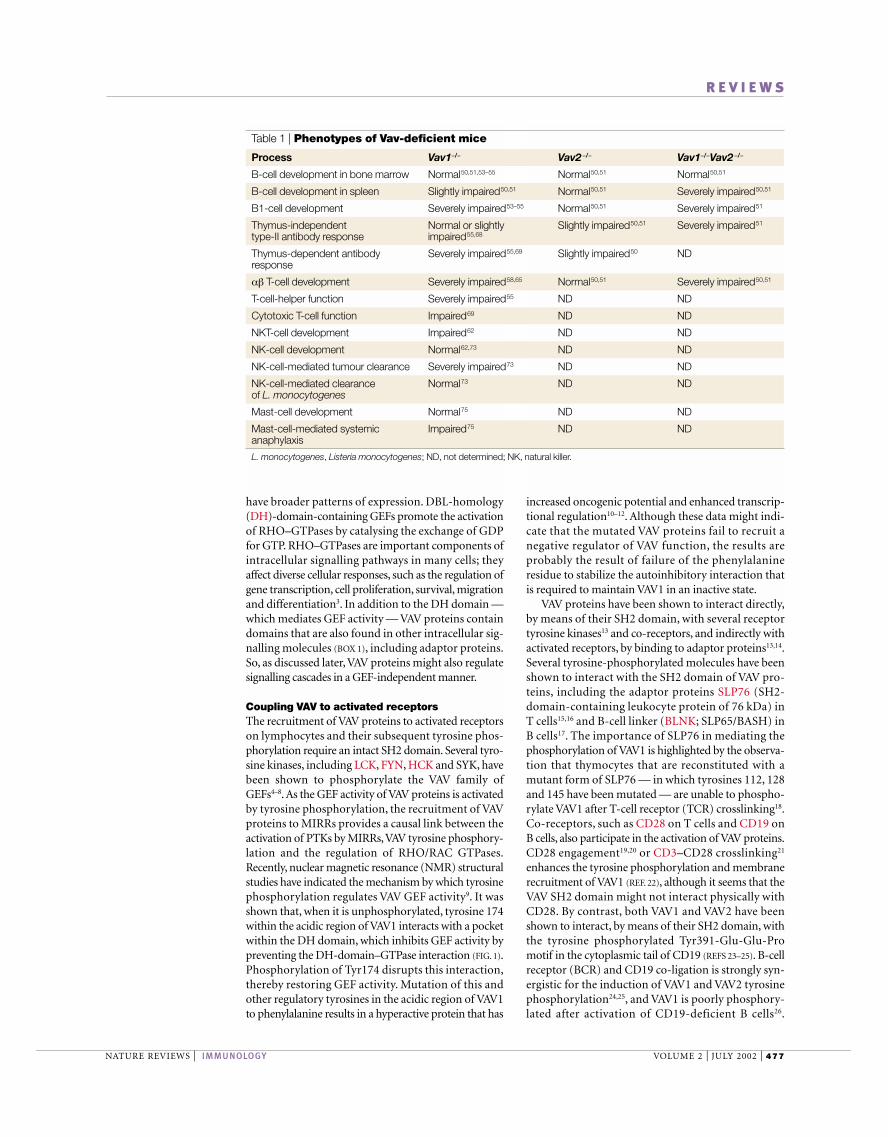
Table 1 | Phenotypes of Vav-deficient mice
Process Vav1−/− Vav2 −/− Vav1−/−Vav2 −/−
B-cell development in bone marrow Normal50,51,53–55 Normal50,51 Normal50,51
B-cell development in spleen Slightly impaired50,51 Normal50,51 Severely impaired50,51
B1-cell development Severely impaired53–55 Normal50,51 Severely impaired51
Thymus-independent Normal or slightly Slightly impaired50,51 Severely impaired51
type-II antibody response impaired55,68
Thymus-dependent antibody Severely impaired55,69 Slightly impaired50 NDresponse
αβ T-cell development Severely impaired58,65 Normal50,51 Severely impaired50,51
T-cell-helper function Severely impaired55 ND ND
Cytotoxic T-cell function Impaired69 ND ND
NKT-cell development Impaired62 ND ND
NK-cell development Normal62,73 ND ND
NK-cell-mediated tumour clearance Severely impaired73 ND ND
NK-cell-mediated clearance Normal73 ND NDof L. monocytogenes
Mast-cell development Normal75 ND ND
Mast-cell-mediated systemic Impaired75 ND NDanaphylaxis
L. monocytogenes, Listeria monocytogenes; ND, not determined; NK, natural killer.

478 | JULY 2002 | VOLUME 2 www.nature.com/reviews/immunol
R E V I E W S
Box 1 | Structural domains of VAV proteins
The three mammalian VAV proteins have an identicaldomain structure34,66,108–111.
Calponin-homology (CH) domainA sequence motif of ~ 100 amino acids that has beenidentified in several actin-binding proteins. It has beensuggested that VAV might bind actin through thisdomain, but, in general, proteins that bind actin bymeans of CH domains contain two CH domains, andboth domains are required for actin binding. The roleof the CH domain in the function of VAV seems to beextremely complex, because it is involved not only inthe regulation of VAV guanine-nucleotide exchangefactor (GEF) activity, but also in Ca2+ mobilizationdownstream of multi-subunit immune-recognitionreceptors (MIRRs).
Acidic (Ac) motifA motif of ~ 50 amino acids that contains three highlyconserved tyrosines (Y) that are involved in theautoinhibition of VAV GEF activity (FIG. 1). These tyrosine residues are preceded by several negatively charged residuesand conform to a tyrosine-phosphorylation motif — Ile/Val-Tyr-Xaa-Xaa-Leu/Ile — that is recognized by both SRC-and SYK-family protein tyrosine kinases (PTKs). Tyrosine 174 is the regulatory tyrosine in VAV1. The regulatorytyrosines in the VAV2 and VAV3 acidic regions have yet to be identified.
DBL-homology (DH) domainA conserved domain of ~ 200 amino acids that is found in all members of the DBL-domain-containing RHO–GEFfamily. This domain interacts physically with RAC and RHO–GTPases and promotes the exchange of GDP for GTP.The introduction of point mutations at highly conserved residues within the DH domain inhibits GEF activity. GEF-deficient VAV proteins are no longer able to transform cells and are unable to regulate previously identified VAV-mediated events downstream of MIRRs (for example, gene transcription, enhanced cellular cytotoxicity and activationof JUN N-terminal kinases).
Pleckstrin-homology (PH) domainA domain that is present in many signalling molecules that bind phosphatidylinositol 3-kinase (PI3K)-generated lipidproducts, such as phosphatidylinositol-4,5-bisphosphate (PtdInsP
2) and phosphatidylinositol-3,4,5-trisphosphate
(PtdInsP3). In all members of the DBL family, the PH domain is located immediately distal to the DH domain. The
PH domain in DBL-family members has been shown to have a role in the regulation of GEF activity by means ofintramolecular interactions with the DH domain and the binding of phosphatidylinositols. The mechanism by whichthe PH domain controls the function of VAV is controversial (see text for details).
Zinc-finger (ZF) domainThis cysteine-rich motif is similar to those that are present in c-RAF and members of the atypical protein kinase C(PKC) family. Mutational inactivation of this domain has been show to abolish VAV GEF activity in vitro and theactivation of VAV-mediated events that require GEF activity in vivo.
Proline-rich (PR) regionA short amino-acid stretch that is rich in proline residues that can be recognized by SH3 domains and WW motifs.The conserved PR region in VAV proteins seems to interact with the amino-terminal SH3 domain of the samemolecule. This exposes a hydrophobic interface on the SH3 domain that is then recognized by the carboxy-terminalSH3 domain of growth-factor-receptor-bound protein 2 (GRB2).
SRC-homology 3 (SH3) domainThis domain interacts with proline-rich stretches in a left-handed polyproline type-II helix. The specificity and affinityof the SH3 domain for its ligand is influenced by the amino acids that flank the polyproline stretch. Recent evidenceindicates that SH3 domains might make contacts with other sequence motifs — aside from proline-rich stretches —that involve interaction with hydrophobic interfaces.
SRC-homology 2 (SH2) domainThis domain interacts with specific phosphotyrosine (pY) residues in the context of 3–6 amino acids that are locatedcarboxy-terminal to the pY. The preferred binding motif for the VAV1 SH2 domain is pTyr-Xaa-Glu-Pro, where Xaa can be Met, Leu or Glu. The observation that VAV2 and VAV3 can interact with an overlapping set of tyrosinephosphorylated molecules through their SH2 domains indicates that they might also preferentially recognize thepTyr-Xaa-Glu-Pro motif; however, this remains to be tested formally.
BLNK, B-cell linker; SLP76, SH2-domain-containing leukocyte protein of 76 kDa; ZAP70, ζ-chain-associated proteinkinase, 70 kDa.
Ca2+ regulation?
ZFPHDHAcCH SH3SH2SH3PR
GEF activity
BindsRAC/RHO–GTPases
Interaction withPI3K-generatedlipid products
Regulatorytyrosines
Recruitment tophosphorylatedtyrosines
Interactionwithproline-richmotifs
NefBLNKSLP76CD19ZAP70SYK
GRB2BTKITK
YYY

NATURE REVIEWS | IMMUNOLOGY VOLUME 2 | JULY 2002 | 479
R E V I E W S
Domains that regulate VAV GEF activityAlthough the tyrosine phosphorylation of VAV proteinsis essential for their activation, it is clear that several otherdomains participate in the regulation of GEF activity.Truncation of the first 65 amino acids of the VAV1calponin-homology (CH) domain, or removal of theVAV3 CH domain results in a constitutively active GEF,which indicates that the CH domain is involved in main-taining VAV proteins in an inactive conformation33,34. It isnot known whether the CH domain is involved in an asyet undescribed intramolecular interaction that stabilizesthe binding of the autoinhibitory tyrosines to the DHdomain. By contrast, the oncogenic potential of VAV2 isonly unleashed after deletion of both the CH and acidicdomains, which indicates that, unlike for VAV1 andVAV3, the CH domain of VAV2 might not participate inlocking the GEF in an inactive conformation6.
The pleckstrin-homology (PH) domain has also beenshown to affect the GEF activity of VAV proteins, but itsmechanism of action is controversial. It is a generalassumption that PH domains bind phosphatidylinositol3-kinase (PI3K)-generated lipid products and, thereby,function to mediate the membrane recruitment ofPH-domain-containing proteins. Indeed, in an in vitrobinding assay, the interaction of the VAV1 PH domainwith phosphatidylinositol-3,4,5-trisphosphate (PtdInsP
3)
was shown to modestly enhance the GEF activity ofVAV1, whereas binding to phosphatidylinositol-4,5-bisphosphate (PtdInsP
2) was inhibitory35. The activa-
tion of VAV1 by PtdInsP3
seemed to be a consequenceof the promotion of VAV1 tyrosine phosphorylation byPtdInsP
3(REF. 35). Such a model predicts that the inhibi-
tion of PI3K activity should inhibit VAV tyrosine phos-phorylation. Although this prediction was borne outfor FYN-mediated phosphorylation of VAV1 in areconstituted system, the inducible phosphorylation ofVAV1 by SYK in the same system was normal36. So, theactivation of VAV1 by receptors that associate with SYKmight be PI3K independent. Consistent with this view,VAV1 phosphorylation in CD3- and CD28-stimulatedthymocytes was PI3K dependent37, whereas in mast cellsthat were stimulated by means of the high-affinityreceptor for IgE (FcεRI)38 or in BCR-stimulated B cells39,PI3K inhibition did not reduce the extent of VAV1 tyro-sine phosphorylation. Moreover, the activation of RACdownstream of the BCR or FcεRI did not require PI3Kactivation, which implies that, in these cell types, theregulation of RAC by VAV is PI3K independent38,39.Taken together, these data highlight the complexity ofthe mechanisms by which the PH domain can influenceVAV GEF activity.
Surprisingly, the PH domain of VAV3 is not requiredfor its ability to function as a GEF in an in vitro assay orits ability to induce F-actin polymerization or morpho-logical changes in transfected fibroblasts34. However, itis clear that the PH domain is important for some VAV-modulated activities, because deletion of or inactivatingpoint mutations in the VAV1 or VAV2 PH domainsresult in proteins that fail to enhance TCR-induced genetranscription or cell-mediated killing by natural killer(NK) cells11,40,41. It is of note that the VAV1 PH domain
It is thought that CD19 activates VAV-family membersthrough the recruitment of LYN27. The significance ofbinding of VAV to CD19 is highlighted by the observa-tions that CD19 signal transduction is less efficient in B cells that lack VAV1, and that the Tyr391Phe muta-tion of CD19 abolishes both VAV1 binding and CD19-regulated responses24.
Additional methods for the recruitment of VAV pro-teins might exist. For example, Jurkat T cells that lackthe adaptor protein linker for activation of T cells(LAT) have reduced VAV1 tyrosine phosphorylationafter TCR crosslinking28. LAT is a small transmembraneprotein that is localized to lipid rafts by PALMITOYLATION14.The cytoplasmic domain of LAT contains numeroustyrosines that are phosphorylated after TCR cross-linking, but none of the phosphorylated tyrosines is inthe Tyr-Xaa-Glu-Pro motif that would be recognizedby the VAV SH2 domain29,30. There are, however, sev-eral potential growth-factor-receptor-bound protein 2(GRB2)-binding sites within the LAT cytoplasmic tail.This is important in light of the observation that thecarboxy-terminal SH3 domain of GRB2 has beenshown to bind the amino-terminal SH3 domain ofVAV1 (REFS 31,32). This interaction might allow therecruitment of VAV1 to tyrosine phosphorylated LATthrough association with GRB2. However, a require-ment for the physical association of GRB2 and VAVproteins in the regulation of lymphocyte responsesremains to be tested. The above data highlight mecha-nisms by which the activation of VAV proteins is cou-pled to MIRRs under normal circumstances. Clearly,the aberrant recruitment of VAV molecules to the cellmembrane could, potentially, have a negative impacton cellular functions (BOX 2).
PALMITOYLATION
The post-translational additionof C16 palmitates to cysteineresidues by a thioester bondtargets proteins to specificmembrane microdomains.
P
GEFactivity
P
P
Ca2+
regulation?
Phosphatase?
SRC or SYKkinase
Inactive (closed) Active (open)
DH
AcY
Y
Figure 1 | Mechanism of action of VAV proteins. There are three highly conserved tyrosine (Y)residues within the acidic (Ac) region of VAV proteins. These tyrosines conform to the Ile/Val-Tyr-Xaa-Xaa-Leu/Ile tyrosine phosphorylation motif for SRC- and SYK-family kinases. The structuralmechanism that controls the activation of VAV proteins was revealed by nuclear magneticresonance (NMR) studies, which showed that the acidic region of VAV1 — in particular, Tyr174 —makes several contacts with side chains of amino acids within the DBL-homology (DH) domain ofVAV1 that are involved in GTPase binding and guanine-nucleotide exchange factor (GEF) activity9.After phosphorylation of Tyr174, the tyrosine is released from the binding pocket, which allowsaccess of the DH domain to its substrate. The fact that all three mammalian VAV proteins, as wellas Drosophila melanogaster vav, contain these highly conserved tyrosines within the acidic regionindicates that this mechanism of VAV regulation might be conserved.

480 | JULY 2002 | VOLUME 2 www.nature.com/reviews/immunol
R E V I E W S
and Vav1−/−Vav2−/− mice, normal numbers of immatureB cells are found in the bone marrow, which indicatesthat the early developmental transitions occur inde-pendently of these proteins. However, Vav1−/−Vav2 −/−
immature B cells fail to complete their maturation intorecirculating B cells50,51. In these mice, many splenic B cells remain immature, express high levels of immuno-globulin M and do not have the increased surfaceexpression of IgD that is characteristic of mature B cells50,51. In Vav1−/−, Vav2−/− and Vav1−/−Vav2−/− mice,peripheral B cells are impaired in their in vitro prolifera-tive and Ca2+ responses after BCR stimulation50,51. Thisimpairment is particularly evident when surface IgM iscrosslinked under conditions of low avidity. In Vav1−/−
B cells, BCR-elicited induction of the expression ofcyclin D2 is blocked, and this might account for thedefective replicative capacity of the cells52. Additionally,Vav1−/− and Vav1−/−Vav2 −/− mice have substantiallyreduced numbers of peritoneal B cells51,53–55. These B-cell defects are similar to those that have beendescribed for deficiencies of various other signallingmolecules (FIG. 2). So, VAV proteins seem not to beimportant in bone-marrow B-cell development but,rather, are required for the peripheral maturationand/or survival of B cells.
T-cell maturation. An early checkpoint that occurs in T-cell development selects for cells that have success-fully rearranged the TCR β-locus and are able toexpress a pre-TCR56. Signalling through the pre-TCRpromotes a proliferative burst and ALLELIC EXCLUSION, andallows further differentiation. Studies of gene-targetedmice have shown that the efficient transition of T cellsthrough this checkpoint requires Vav1, but not Vav2(REFS 49,51,53,54,57,58). Importantly, complementation ofthe Vav1 deficiency at this checkpoint by an active alleleof Rac1 (REF. 59), but not RhoA60, provides evidence that itis the GEF activity of Vav1 towards Rac-family membersthat is crucial for the transition from double-negative todouble-positive thymocytes. In non-TCR-transgenicVav1−/− mice, some single-positive T cells and naturalkiller T (NKT) cells do develop, presumably becausetheir receptors have sufficient signalling capacity in theabsence of Vav1 (REFS 58,61,62). However, these peripheralT cells have impaired function and signalling61–64. UsingTCR-transgenic,Vav1-deficient mice, it was found thatVav1 has a role in the positive- and negative-selectionevents that are imposed on double-positive thymo-cytes58,65. The development of TCR-transgenic T cellswas absolutely dependent on Vav1. The decreased effi-ciency of positive selection is likely to be an importantreason for the reduced number of T cells in Vav1−/− mice.However, it remains to be seen if TONIC SIGNALLING throughthe TCR, which is thought to be required for the survivalof mature T cells, is also dependent on Vav1.
Several lines of evidence indicate that Vav2 and Vav3cannot compensate functionally for the loss of Vav1.First, both proteins are expressed in T cells and are tyro-sine phosphorylated after TCR crosslinking25,34,40,66.Second, when Vav1−/− mice are crossed with mice thatlack Cbl-b — an important negative regulator of Vav
does not conform to a recently defined PtdInsP3-specific
binding motif42,43, and it is not able to function in vivo ina yeast assay that is designed to detect PH domains,which indicates that it has very low affinity for PtdInsP
3
(REF. 44). Intriguingly, the PH domain of the proto-oncogene product DBL was found to target DBL to notonly the plasma membrane, but also the actincytoskeleton45. It is clear that the PH domain is impor-tant in regulating the function of VAV1 and VAV2, butthe underlying mechanism is still unclear and mightinvolve functions in addition to binding to lipids, suchas protein–protein interactions.
The zinc-finger (ZF) domain also regulates the GEFactivity of VAV proteins. The mutation of highly con-served cysteine resides within the ZF domain of all threeVAV proteins has been shown to inhibit GEF activity in vitro and VAV-regulated biological processes thatrequire GEF function, including gene transcription, acti-vation of JUN N-terminal kinases (JNKs), cell-mediatedkilling and cellular transformation11,34,46–48. Interestingly,it has been shown for VAV3 that the mechanism bywhich the ZF domain regulates GEF activity involvesdirect interaction with GTPases. The ZF domain ofVAV3 was found to interact directly with RHOA34 and itmight participate in binding and/or stabilizing theGEF–GTPase interaction during the exchange reaction34.It is not known whether VAV1 and VAV2 ZF domainsbind RHO GTPases, but the presence of the DH–PH–ZFcassette in all three VAV proteins indicates a cognatemechanism for controlling GEF activity.
VAV proteins: physiological importanceThere is compelling evidence from studies of gene-targeted mice to indicate that VAV proteins have rolesin the development and function of many types ofimmune cell (TABLE 1).
B-cell maturation. The development of B cells proceedsthrough a well-defined set of stages49. In Vav1−/−, Vav2−/−
ALLELIC EXCLUSION
This process by which thesuccessful rearrangement andexpression of an antigen-receptor subunit preventsrearrangement at the other allele.
TONIC SIGNALLING
A survival signal that arises as aconsequence of antigen-receptorexpression that is insufficient togive rise to cell activation.
Box 2 | Aberrant membrane recruitment of VAV molecules by HIV
The HIV-1-encoded Nef protein is a virulence factor that is required for efficient viralreplication and progression to AIDS. Nef is a myristoylated membrane-bound proteinthat downmodulates the level of expression of CD4, T-cell receptor (TCR)–CD3complexes, MHC class I antigens and CD28 on the cell surface. In addition, theexpression of Nef has been found to profoundly influence intracellular signallingcascades, which results in the activation of SRC-family kinases, RAC1/CDC42 and p21-activated kinase-1 (PAK1), and, also, in the activation of nuclear factor of activatedT cells (NFAT) and protection from apoptotic signals. It has been found that the Nefprotein functions as an adaptor-like molecule; it nucleates SRC-kinase–VAV–PAK1complexes by means of the physical interaction of these signalling proteins with definedmotifs that are present in the Nef protein117. In this way, Nef can regulate VAV activity byits recruitment to a Nef-bound SRC kinase and the RAC1 target PAK1. It remains to bedetermined which Nef-mediated signalling cascades are controlled by VAV proteins. It isof interest that the J.Vav1 cell line — a derivative of the Jurkat T-cell line, in which VAV1has been deleted by homologous recombination — was found to have enhancedsusceptibility to FAS ligand and oxidative stress, which indicates that VAV1 regulates aprotective pathway in T cells (Y. Cao et al., unpublished observations). It is tempting tospeculate that the activation of VAV1 by Nef contributes to the protection fromapoptosis that is associated with the expression of Nef.

NATURE REVIEWS | IMMUNOLOGY VOLUME 2 | JULY 2002 | 481
R E V I E W S
B-cell and T-cell responses. The immune responses ofVav1−/− mice to several model antigens have been tested.Responses to THYMUS-DEPENDENT ANTIGENS are severelyimpaired in Vav1−/− mice55,68 owing to a lack of T-helper-cell function. Vav1−/− T cells proliferate only slightly afterin vitro stimulation with antibodies that are specific forCD3 or CD28, whereas antigen-specific Vav1−/− T cellsproliferate normally in vivo55. Therefore, the failure toproduce cytokines, such as interleukin-4 (IL-4), that arenecessary for B-cell differentation might account for thedeficient helper-cell function55. Studies of the antiviralresponses of Vav1−/− mice have revealed that primaryresponses are defective, but that primed T cells producenormal secondary cytotoxic T lymphocyte (CTL)responses69. These data indicate that Vav1 is not requiredfor the expansion or formation of memory CD8+ CTLpopulations, but they might be consistent with arequirement for Vav1 for the effector function of pri-mary CTLs. The response of Vav2-deficient mice tothymus-dependent antigens is also defective, althoughit has not been established whether this is due todefects in the function of B cells, T cells or both lin-eages50. However, it is known that Vav1−/− B cells canundergo CLASS SWITCHING when T-cell help is provided55.B cells from Vav1−/− mice respond relatively well totype-2 THYMUS-INDEPENDENT ANTIGENS that highlycrosslink BCRs, leading to B-cell activation, whereasthe response of Vav2−/− mice is mildly impaired50,51,55.Vav1−/−Vav2−/− mice are further impaired, which indi-cates that there is redundancy between these two pro-teins51. These data highlight the important role of Vav1in T-cell function and reveal that both Vav1 and Vav2are crucial for B-cell function.
Natural-killer-cell function. NK cells are lymphocytesthat express various receptors that trigger cytotoxickilling and the release of cytokines without previoussensitization70, and they might be an early first line ofdefence against viral infection and tumour develop-ment. Genetic studies that used the overexpression ofVav1 (REF. 46) or Vav2 (REF. 40), or antisense oligonu-cleotides to Vav1 (REF. 71) have provided evidence thatboth of these GEFs are able to positively regulate NK-cell-mediated killing. These studies, together with exper-iments that used dominant-negative forms of Rac1 andRhoA46,72, indicate that the GEF function of Vav isimportant for the induction of killing. Importantly,Vav1−/− mice are defective in their ability to cleartumours in vivo and are defective in natural killing andantibody-dependent cellular cytotoxicity (ADCC) in vitro62,73. Interestingly, not all NK-cell functions requiresignalling through Vav1, as interferon-γ (IFN-γ) produc-tion by NK cells and the ability to resolve infections withListeria monocytogenes is normal in Vav1−/− mice73.Mechanistic studies have indicated that Vav1−/− NK cellsretain the capacity to form conjugates with target cells,but are defective in their release of cytotoxic granules46,73.Taken together with the observation that dominant-negative Rac1 decreases both NK-cell conjugate forma-tion and the polarization of cytolytic granules towardsthe target46, a pathway that involves Vav1 and Rac1
proteins — Vav2 becomes hyper-phosphorylated,which is indicative of activation, but T-cell develop-ment remains impaired67. Surprisingly, some aspects ofthe Vav1-deficient phenotype are reversed in thismouse, including TCR-induced activation of Cdc42and actin polymerization. This is, presumably, accom-plished through the activation of other Vav-familymembers; however, this is clearly still insufficient topromote normal T-cell development and activation.Finally, Vav2 deficiency does not affect T-cell develop-ment, and the combined deficiency of both Vav1 andVav2 has little additional effect on T-cell developmentbeyond that observed in the absence of Vav1 (REFS 50,51).These data point to an important role for Vav1, but notVav2, in T-cell development and activation. However,determining the potential role of Vav3 in both B-celland T-cell development and function will requiregenetic studies in animals.
THYMUS-DEPENDENT ANTIGENS
Antigenic stimuli that requirethe function of thymus-derivedlymphocytes to generate ahumoral immune response.
CLASS SWITCHING
The somatic recombinationprocess by whichimmunoglobulin isotypes areswitched from IgM to IgG orIgA.
THYMUS-INDEPENDENT
ANTIGENS
Antigenic stimuli that promotehumoral immune responses inthe absence of thymus-derivedlymphocytes.
BLNK
p110
Receptor
Co-receptor
LYN
SYKPLC-γ2
BTK
p85
GRB2SOS
RAC
RAS
RAS GRP
? ?
Amplification
DAG
InsP3 PtdInsP2
PtdInsP3 PtdInsP2
Ca2+
VAV
?
Figure 2 | VAV and the signalosome. The B-cell phenotype of Vav1−/−Vav2−/− mice closelyresembles that of XID mice that lack functional Bruton’s tyrosine kinase (Btk)112. A point mutation inthe pleckstrin-homology (PH) domain of Btk, which results in the inability of Btk to interact withphosphatidylinositol-3,4,5-trisphosphate (PtdInsP3), is the underlying defect in XID mice113. As aconsequence of this, Btk cannot be recruited to membrane-associated signalling complexes.Mutations of the p85α regulatory subunit of phosphatidylinositol 3-kinase (PI3K), phospholipase-Cγ2 (PLC-γ2) and B-cell linker (BLNK; also known as SLP65 or BASH) also give rise to defects inperipheral B-cell maturation that are similar to those that are found in Vav1−/−Vav2−/− mice112,114. All of these proteins might interact physically to form a signalling complex known as the‘signalosome’, of which the best characterized function is the regulation of PLC-γ2 after antigen-receptor ligation. The ability of VAV proteins to regulate phosphatidylinositol 3-kinase (PI3K) activityplaces VAV proteins in a central position in such a model. A similar multi-protein complex mightfunction in T cells (not shown), as the phenotype of Vav1−/− T cells closely resembles that of micethat are deficient in the Tec kinases Rlk and Itk115. The model also offers an explanation for thefailure of mitogen-activated protein kinase (MAPK) activation after T-cell-receptor crosslinking64.Decreased activation of PLC-γ2 would affect not only the production of InsP3, but alsodiacylglycerol (DAG), in the membrane, which then could impinge on the RAS–MAPK pathway byaffecting the activation of the DAG-dependent RAS exchange factor, RAS GRP116. VAV proteinsmight regulate PI3K through direct interactions, the activation of RAC or the activation of RAS,which binds to the p110 catalytic subunit of PI3K. GRB2, growth-factor-receptor-bound protein 2.

482 | JULY 2002 | VOLUME 2 www.nature.com/reviews/immunol
R E V I E W S
In addition to the activation of NFAT/AP1,VAV-familymembers have been observed to enhance gene transcrip-tion from many isolated elements in the interleukin-2(IL2) promoter, including NF-κB, AP1 and the CD28-responsive element (CD28RE)66,84–86. J.Vav1 cells — aderivative of Jurkat T cells, in which VAV1 has beendeleted by homologous recombination — have a severedefect in TCR-mediated IL2 gene transcription, the regu-lation of isolated transcriptional elements (such as AP1,NFAT/AP1, NF-κB and CD28RE) and JNK activity thatcan only be ameliorated by the reintroduction of a GEF-competent VAV1 (Y. Cao et al., unpublished observa-tions). These data indicate a central role for the GEFactivity of VAV proteins in the regulation of transcription.
The mechanism by which VAV proteins regulatetranscription factors has not been fully elucidated; how-ever, several recent experiments have begun to piece thepuzzle together. The activation of T cells from Vav1−/−
mice failed to promote the degradation of IκBα, whichimplicates Vav1 as a regulator of IκB kinase (IKK) — anenzyme that phosphorylates IκBα and triggers itsdegradation64. Several reports have indicated that Vav1might regulate protein kinase-Cθ (PKCθ), which is akey regulator of NF-κB. In mouse thymocytes, Vav1 isconstitutively associated with PKCθ65. In response toTCR and CD28 co-engagement, PKCθ translocates tothe cell membrane and has been found to associate withthe IKK complex, where it presumably activates IKK87,88.In addition, overexpression studies in Jurkat T cells haveindicated a role for VAV1 and its GTPase target RAC1 inthe membrane recruitment and activation of PKCθ,and the optimal activation of the JNK pathway21,41,82,89,90.Interestingly, the recruitment of PKCθ to the mem-brane and its activation after antibody-mediated CD3and CD28 co-crosslinking is not impaired in J.Vav1 cells(Y. Cao et al., unpublished observations), which indi-cates that either VAV1 is not the sole regulator of PKCθ,or that VAV2 and VAV3 might compensate for the lossof VAV1. Consistent with previous data, the J.Vav1 cellsare unable to activate JNKs after anti-CD3, or anti-CD3and anti-CD28 crosslinking, which implicates VAV1 asan obligate regulator of JNK activation in this cell line.Surprisingly, peripheral T cells from PKCθ-deficientmice are deficient in NF-κB activation, but not JNKactivity91. Therefore, it remains to be determined howVAV1 regulates NF-κB in J.Vav1 cells, if it does not do sothrough PKCθ.
VAV proteins and activation of TEC-family kinases. VAVproteins seem to regulate calcium mobilization in sev-eral ways, and sustained levels of cytoplasmic Ca2+ arecrucial for NFAT function92. The observations that Vav-deficient T cells57,58,61,63,64, B cells39,50,51 and mast cells75
have a defect in Ca2+ mobilization downstream ofMIRRs and that the overexpression of Vav proteinspotentiates antigen-receptor-stimulated calciumrelease11,25 provides compelling evidence of a role forVav proteins in the regulation of Ca2+ mobilization.Initial studies indicated a role for RAC1-mediated acti-vation of phosphatidylinositol-4-phosphate 5-kinase(PIP5K) in regulating the level of PtdInsP
2, the substrate
seems to be of crucial importance for NK-cell-mediatedcytotoxicity.
Mast-cell degranulation. Mast cells are another impor-tant cellular component of the innate immune system74.These cells can sense antigen indirectly through thebinding of antigen that has been complexed by IgE toFcεRI on the cell surface. The activation of mast cells byantigen stimulates the release of inflammatory media-tors in the form of secretory-granule contents, cytokinesand arachadonic-acid metabolites. Studies of Vav1−/−
mice have shown that mast-cell development seems tobe normal. However, the animals were defective in theirability to undergo SYSTEMIC ANAPHYLAXIS75. This effect couldbe accounted for by a requirement for Vav1 for optimalsignalling through FcεRI75.
GTPase-dependent aspects of VAV signallingThe activation of multiple GTPases is one mechanismby which VAV proteins control cellular responses.Several in vitro and in vivo studies have shown a distinct,yet overlapping, spectrum of GTPase targets for differ-ent VAV-family members. VAV1 is primarily a GEF forRAC1, RAC2 and RHOG, whereas VAV2 and VAV3 actprimarily on RHOA, RHOB and RHOG GTPases6,34,48.The ability of VAV-family members to mediate the acti-vation of CDC42 in vitro is controversial6,34,76–78. A recentstudy has identified the structural requirements of theRAC- and RHO-family members that allow bindingand GTP exchange by VAV-family proteins, excludingCDC42 (REF. 79). In addition, consistent with the in vitroexperiments, the activation of Rac1 is deficient in thymo-cytes from Vav1−/− mice and in Vav3−/− DT40 B cells39,80.Although it has been reported that the activation ofCdc42 in Vav1−/− T cells is defective, it is not clear whetherthis indicates that Vav1 has a direct effect on thisGTPase67. Given that GTPase signalling networks controldiverse biological processes, we consider those functionsof VAV proteins that require GTPase activation.
VAV, the cytoskeleton and immunological synapses. Thecytoskeleton is crucial for the assembly of signalling-path-way components and formation of the IMMUNOLOGICAL
SYNAPSE between lymphocytes and target cells. Vav1 hasbeen shown to regulate actin polymerization61,63 andlipid-raft polarization, as well as the concentration ofMHC-class-II–TCR complexes at the APC–T-cell inter-face in Vav1-deficient T cells67,81,82. These studies impli-cate VAV proteins in the signalling events that depend oncytoskeletal reorganization; however, it is unlikely thatthe formation of an immunological synapse is requiredfor signal transduction, because this event occurs aftersignal transduction83.
VAV-mediated control of gene transcription. VAV-dependent gene transcription in both B and T cellsrequires GTPase activation. The ability of VAV1 andVAV2 to activate NFAT/AP1 transcriptional elementscan be blocked by dominant-negative GTPases, whichis consistent with the notion that the GEF activity ofVAV1 is important for transcriptional activation25,41,84.
SYSTEMIC ANAPHYLAXIS
Acute hypersensitivity shockthat occurs after the exposure ofsensitized animals to antigen.
IMMUNOLOGICAL SYNAPSE
A structure that is formed at thecell surface between a T cell andan antigen-presenting cell; alsoknown as the supra-molecularactivation cluster (SMAC).Important molecules involved inT-cell activation — includingthe T-cell receptor, numeroussignal-transduction moleculesand molecular adaptors —accumulate at this site.Mobilization of the actincytoskeleton of the cell isrequired for immunological-synapse formation.

NATURE REVIEWS | IMMUNOLOGY VOLUME 2 | JULY 2002 | 483
R E V I E W S
activate PI3K seems to be Rac-dependent, becauseBCR-mediated PI3K activation could be restored inVav3−/− DT40 cells by wild-type Vav3, but not a GEF-deficient Vav3 (REF. 39). These studies indicate a GEF-dependent role for VAV proteins in the activation of PI3K.In addition, a deficiency in the generation of PtdInsP
3
might not only adversely affect the activation of TEC-family kinases and the subsequent tyrosine phosphoryla-tion and activation of PLC-γ isoforms, but might alsoaffect the proper localization and activation of otherlipid-binding-domain-containing molecules. The possi-bility that VAV proteins might regulate the activity ofTEC-family kinases through additional mechanisms,such as direct interaction100–104, has not been excluded.When considered together, these new observations placeVAV proteins in a central position in MIRR signal trans-duction (FIG. 2), as VAV proteins are required for theproper activation of lipid and protein-tyrosine kinases, aswell as PLC.
VAV: more than just an exchange factor?GEF-independent functions of VAV proteins contributeto signal transduction downstream of MIRRs, but theymust be precisely coordinated with GEF-dependentfunctions to generate the appropriate biologicalresponse. In particular, the control of intracellular cal-cium levels by VAV proteins might be mediated, in part,by GEF-independent pathways. VAV-mediated regula-tion of PI3K activity is not the only mechanism bywhich VAV proteins control PLC-γ1 activity. So, the inhi-bition of PI3K activity in thymocytes reduced PLC-γ1phosphorylation only fractionally compared with thereduction of phosphorylation that is seen in Vav1−/− thy-mocytes80. The analysis of signalling-complex formationin Vav1−/− thymocytes indicates that the inducible associ-ation of PLC-γ1 with Slp76/Gads is defective80.Althoughthis interaction is thought to be mediated by the SH3domain of PLC-γ1 binding Slp76/Gads14, these dataindicate that Vav1 is important for the formation and/orstabilization of this complex. The importance of thisinteraction for PLC-γ1 activity is indicated by the obser-vation that mutations in Slp76 that prevent PLC-γ1binding lead to a reduction in the level of TCR-inducedphosphorylation105. The treatment of thymocytes with aPI3K inhibitor had little effect on this interaction, whichshows that PI3K activation is not required. It is notknown whether this property of Vav1 requires GEFactivity in thymocytes, although it is of note that GEF-deficient Vav1 was able to restore the tyrosine phospho-rylation, but not full activation, of PLC-γ1 in Vav1−/−
mast cells75. Taken together, these data indicate that VAVproteins might regulate PLC-γ isoforms by means ofboth GEF-dependent and -independent mechanisms.Indeed, PLC-γ1 has been found to be associated withVav1, but the functional significance of the interactionremains to be evaluated106.
Also consistent with the suggestion of a GEF-inde-pendent function of VAV proteins is the observationthat truncation mutants of the CH domain orCH–acidic region are still able to activate GTPases andAP1 transcription-factor elements21,47,86,90,91, but are
for phospholipase-Cγ (PLC-γ) proteins24,93. In thismodel, exhaustion of the PLC-γ substrate would lead toreduced InsP
3production, and calcium release would be
curtailed. Several recent studies indicate that VAV pro-teins control Ca2+ flux by means of the regulation ofPLC-γ activation. So, it has been observed that the levelof tyrosine phosphorylation of PLC-γ1 and PLC-γ2 inVav1−/− mast cells is diminished after FcεRI crosslink-ing75. Furthermore, in Vav3−/− DT40 B cells and in B cellsfrom Vav1−/−Vav2−/− mice, BCR-stimulated PLC-γ2phosphorylation and lipase activity are reduced39.Importantly, these studies analysed PLC-γ2 activityunder conditions of excess substrate, revealing an intrin-sic defect in enzyme activation. Moreover, by using atransgenic TCR on a β
2-microglobulin-deficient back-
ground — in which Vav1−/− double-positive thymocytesare poised to undergo signalling events that mimic posi-tive and negative selection — it has been shown that thephosphorylation of PLC-γ1 on residue Tyr783, the siteof activation for PLC-γ1, requires Vav1 function80. Thefailure to activate the PLC-γ isoforms is owing, in part,to the defective activation of the Tec-family PTKs Itkand Tec in thymocytes and Btk in B cells. Consistentwith the defect in PLC-γ1 and PLC-γ2 activation in Vav-deficient cells, TCR-inducible phosphorylation of Itkand Tec80, and BCR-stimulated phosphorylation of Btkis defective in the absence of Vav-family members39.Furthermore, it was shown — using monoclonal anti-bodies specific for individual sites of tyrosine phospho-rylation on Btk that are required for its activation94 —that the activation of the kinase activity of Btk requiresthe function of Vav proteins (E. Clayton et al., unpub-lished observations). Additionally, Btk phosphorylationin trans by upstream kinases requires Vav proteins. It is,however, important to point out that the deficiency inVav proteins does not lead to a global defect in tyrosinephosphorylation, but seems to specifically affect a path-way that leads to the activation of Tec-family PTKs andtheir downstream substrates.
TEC-family PTKs contain a PH domain that hasbeen shown to be absolutely required for enzymaticactivity95. The above data indicate that there is a VAV-regulated pathway that leads to the activation of TEC-family PTKs. The finding that VAV proteins becomeassociated with immune complexes of the p85 regula-tory subunit of PI3K in stimulated cells indicates thatVAV and PI3K activity could, potentially, be coordi-nately regulated23,96–98. Indeed, RAC GTPases canenhance the catalytic activity of PI3Ks through bindingto the regulatory p85 subunit of the p85–p110 het-erodimer39,93,99. In view of the data that indicate thatPI3K might positively regulate VAV, it was, therefore,surprising to find that BCR-stimulated activation ofPI3K was defective in DT40 B cells and mouse B cellsand that the activation of PI3K by FcεR1 crosslinkingwas defective in mast cells that lack VAV proteins39,75.Furthermore, phosphorylation of protein kinase B —which is a good indicator of PI3K activation — isdefective in Vav1−/− double-positive thymocytes, whichindicates that Vav is an upstream regulator of PI3Kactivation80. Moreover, the ability of Vav proteins to

484 | JULY 2002 | VOLUME 2 www.nature.com/reviews/immunol
R E V I E W S
Indeed, a Vav1 mutant that has Tyr→Phe mutations atall three tyrosines within the acidic region does hyper-activate TCR-induced NFAT/AP1-mediated gene tran-scription and result in sustained Ca2+ fluxes in theJurkat T-cell line11. In summary, these data indicatethat CH-domain-mediated regulation of Ca2+ flux,combined with DH-domain-mediated activation ofGTPases, provides optimal signals for the activation ofNFAT/AP1 transcriptional elements. Although Vav1mutants that lack CH domains have been reported tohave altered subcellular localization in fibroblasts, it isnot clear if this is simply a result of the profoundcytoskeletal changes that result from the activation ofGTPases86,107. Clearly, identifying the mechanism bywhich the CH domain regulates Ca2+ mobilization willbe an area of intense interest in the future.
Concluding remarksThe data that have been discussed in this review high-light the importance of VAV-family proteins in theregulation of immune-cell development and activa-tion. Clearly, there are many unresolved issues,including the role of VAV3 in the immune responseand of VAV proteins in non-lymphoid cells. Themechanism by which the CH domain of VAV proteinscontrols Ca2+ fluxes, and the contribution of thenumerous modular domains of VAV proteins to theireffector functions are still not understood fully.Mouse models and cell lines that lack VAV proteinswill continue to be an extremely useful tool for thedisection of VAV-regulated signalling pathways. Inparticular, structure–function studies of VAV proteinsin cell lines and in vivo should provide mechanisticinsights into the mechanism of MIRR function andthe physiology of the immune response. In addition,the knowledge that we gain from studies carried outin the haematopoietic system might give us an insightinto the role of VAV proteins in cells outside of theimmune system.
unable to activate NFAT/AP1 (REFS 11,25,85). Comparedwith full-length VAV1, the mutant that lacks the CHdomain is unable to potentiate Ca2+ release in Jurkat11 orJ.Vav1 cells (Y. Cao et al., unpublished observations)after TCR crosslinking. In addition, wild-type VAV2, butnot a mutant that lacked the CH–acidic region, wasfound to enhance Ca2+ mobilization in B cells25. Finally,a full-length VAV1 protein that had a point mutation inthe CH domain was unable to regulate NFAT/AP1 tran-scriptional elements in T cells11. All of these mutant pro-teins were fully able to modulate TCR-inducedNFAT/AP1 activity in the presence of the Ca2+ ionophoreionomycin or if co-transfected with activatedcalcineurin11. These data point to a role for the CHdomain of VAV proteins in calcium regulation that is dis-tinct from their ability to regulate GTPases. Consistentwith this, the overexpression of constitutively activeGTPases in Jurkat T cells failed to enhance NFAT/AP1gene transcription downstream of the TCR unless co-transfected with an activated form of calcineurin thatfunctions independently of Ca2+ (REF. 3).
The ability of VAV proteins to control the activationof RHO GTPases, as well as the level of intracellular Ca2+,could contribute to the activation of several effectorfunctions. As mentioned above, the CH domain isinvolved in the autoinhibition of GEF activity.We specu-late that the autoinhibition of VAV proteins throughintramolecular interactions between the CH domainand acidic region will also prevent the CH domain fromregulating the level of Ca2+. Therefore, after recruitmentto the activated receptor, the VAV protein is tyrosinephosphorylation in the acidic region, which results innot only the activation of its GEF activity, but also theability of the CH domain to regulate Ca2+ (FIG. 1).Dephosphorylation of the tyrosine(s) within the acidicregion by receptor-recruited tyrosine phosphatases wouldpromote autoinhibition. In such a model, a VAV proteinthat is unable to become autoinhibited would be hyper-active in terms of both GEF function and Ca2+ signalling.
1. Bishop, A. B. & Hall, A. Rho GTPases and their effectorproteins. Biochem. J. 348, 241–255 (2000).
2. Bustelo, X. R. Vav proteins, adaptors and cell signalling.Oncogene 20, 6372–6381 (2001).
3. Henning, S. & Cleverley, S. Small GTPases in lymphocytebiology — Rho proteins take center stage. Immunol. Res.20, 29–42 (1999).
4. Deckert, M., Tartare-Deckert, S., Couture, C. & Altman, A.Functional and physical interactions of Syk family kinaseswith the Vav proto-oncogene. Immunity 5, 591–604 (1996).
5. Han, J. et al. Lck regulates Vav activation of members of theRho family of GTPases. Mol. Cell. Biol. 17, 1346–1353 (1997).
6. Schuebel, K. E., Movilla, N., Rosa, J. L. & Bustelo, X. R.Phosphorylation-dependent and constitutive activation ofRho proteins by wild-type and oncogenic Vav-2. EMBO J.17, 6608–6621 (1998).
7. Michel, F., Grimaud, L., Tuosto, L. & Acuto, O. Fyn and ZAP-70 are required for Vav phosphorylation in T cellsstimulated by antigen-presenting cells. J. Biol. Chem. 273,31932–31938 (1998).
8. Huang, J., Tilly, D., Altman, A., Sugie, K. & Grey, H. M. T-cell receptor antagonists induce Vav phosphorylation byselective activation of Fyn kinase. Proc. Natl Acad. Sci. USA97, 10923–10929 (2000).
9. Aghazadeh, B., Lowry, W. E., Huang, X.-Y. & Rosen, M.Structural basis for relief of autoinhibition of the Dblhomology domain of proto-oncogene Vav by tyrosinephosphorylation. Cell 102, 625–633 (2000).
This paper describes a unique mechanism for theactivation of the enzymatic function of VAV1.
10. Lopez-Lago, M., Lee, H., Cruz, C., Movilla, N. & Bustelo, X. R.Tyrosine phosphorylation mediates both activation anddownmodulation of the biological activity of Vav. Mol. Cell.Biol. 20, 1678–1691 (2000).
11. Billadeau, D. D., Mackie, S. M., Schoon, R. A. & Leibson, P. J.Specific subdomains of Vav differentially affect T-cell andNK-cell activation. J. Immunol. 164, 3971–3981 (2000).
12. Kuhne, M. R., Ku, G. & Weiss, A. A guanine nucleotideexchange factor-independent function of Vav-1 intranscriptional activation. J. Biol. Chem. 275, 2185–2190(2000).
13. Bustelo, X. R. Regulatory and signalling properties of the Vavfamily. Mol. Cell. Biol. 20, 1461–1477 (2000).
14. Koretzky, G. A. & Myung, P. S. Positive and negativeregulation of T-cell activation by adaptor proteins. NatureRev. Immunol. 1, 95–107 (2001).
15. Tuosto, L., Michel, F. & Acuto, O. p95vav associates withtyrosine-phosphorylated SLP-76 in antigen-stimulated T cells. J. Exp. Med. 184, 1161–1166 (1996).
16. Tartare-Deckert, S. et al. Vav2 activates c-Fos serumresponse element and CD69 expression but negativelyregulates nuclear factor of activated T cells and interleukin-2gene activation in T lymphocytes. J. Biol. Chem. 276,20849–20857 (2001).
17. Fu, C. & Chan, A. C. Identification of two tyrosinephosphoproteins, pp70 and pp68, which interact with
phospholipase Cγ, Grb2 and Vav after B-cell antigenreceptor activation. J. Biol. Chem. 272, 27362–27368(1997).
18. Myung, P. S. et al. Differential requirement for SLP-76domains in T-cell development and function. Immunity 15,1011–1026 (2001).
19. Nunes, J. A., Collette, Y., Truneh, A., Olive, D. & Cantrell, D. A.The role of p21ras in CD28 signal transduction: triggering ofCD28 with antibodies, but not the ligand B7-1, activatesp21ras. J. Exp. Med. 180, 1067–1076 (1994).
20. Klasen, S., Pages, F., Peyron, J.-F., Cantrell, D. A. & Olive, D.Two distinct regions of the CD28 intracytoplasmic domainare involved in the tyrosine phosphorylation of Vav andGTPase activating protein-associated p62 protein. Int.Immunol. 10, 481–489 (1998).
21. Hehner, S. P., Hofmann, T. G., Dienz, O., Droge, W. &Schmitz, M. L. Tyrosine-phosphorylated Vav1 as a point ofintegration for T-cell-receptor- and CD28-mediatedactivation of JNK, p38 and interleukin-2 transcription. J. Biol.Chem. 275, 18160–18171 (2000).
22. Raab, M., Pfister, S. & Rudd, C. E. CD28 signalling viaVav/SLP-76 adaptors: regulation of cytokine transcriptionindependent of TCR ligation. Immunity 15, 921–933 (2001).
23. Weng, W. K., Jarvis, L. & LeBien, T. W. Signaling through CD19 activates Vav/mitogen-activated proteinkinase pathway and induces formation of aCD19/Vav/phosphatidylinositol 3-kinase complex in humanB-cell precursors. J. Biol. Chem. 269, 32514–32521 (1994).

NATURE REVIEWS | IMMUNOLOGY VOLUME 2 | JULY 2002 | 485
R E V I E W S
24. O’Rourke, L. et al. CD19 as a membrane-anchored adaptorprotein of B lymphocytes: costimulation of lipid and proteinkinases by recruitment of Vav. Immunity 8, 635–645 (1998).
25. Doody, G. M. et al. Vav-2 controls NFAT-dependenttranscription in B but not T lymphocytes. EMBO J. 19,6173–6184 (2000).
26. Sato, S., Jansen, P. J., & Tedder, T. F. CD19 and CD22expression reciprocally regulates tyrosine phosphorylation ofVav protein during B-lymphocyte signalling. Proc. Natl Acad.Sci. USA 94, 13158–13162 (1997).
27. Fujimoto, M. et al. CD19 regulates Src family protein tyrosinekinase activation in B lymphocytes through processiveamplification. Immunity 13, 47–57 (2000).
28. Finco, T. S., Kadlecek, T., Zhang, W., Samelson, L. E. &Weiss, A. LAT is required for the TCR-mediated activation ofPLCγ1 and the Ras pathway. Immunity 9, 617–626 (1998).
29. Zhang, W., Sloan-Lancaster, J., Kitchen, J., Trible, R. &Samelson, L. LAT: the ZAP-70 tyrosine kinase substrate thatlinks T-cell receptor to cellular activation. Cell 92, 83–92(1998).
30. Songyang, Z. et al. Specific motifs recognized by the SH2domains of Csk, 3BP2, fps/fes, GRB-2, HCP, SHC, Syk andVav. Mol. Cell. Biol. 14, 2777–2785 (1994).
31. Ye, Z. S. & Baltimore, D. Binding of Vav to Grb2 throughdimerization of Src-homology 3 domains. Proc. Natl Acad.Sci. USA 91, 12629–12633 (1994).
32. Nishida, M. et al. Novel recognition mode between Vav andGrb2 SH3 domains. EMBO J. 20, 2995–3007 (2001).
33. Katzav, S., Cleveland, J. L., Heslop, H. E. & Pulido, D. Lossof the amino-terminal helix–loop–helix domain of the Vavproto-oncogene activates its transforming potential. Mol.Cell. Biol. 11, 1912–1920 (1991).
34. Movilla, N. & Bustelo, X. R. Biological and regulatoryproperties of Vav-3, a new member of the Vav family ofoncoproteins. Mol. Cell. Biol. 19, 7870–7885 (1999).
35. Han, J. et al. Role of substrates and products of PI 3-kinasein regulating activation of Rac-related guanosinetriphosphates by Vav. Science 279, 558–560 (1998).
36. Das, B. et al. Control of intramolecular interactions betweenthe pleckstrin homology and Dbl homology domains of Vavand SOS1 regulates Rac binding. J. Biol. Chem. 275,15074–15081 (2000).
37. Fang, D. & Liu, Y.-C. Proteolysis-independent regulation ofPI3K by Cbl-b-mediated ubiquitination in T cells. NatureImmunol. 2, 870–875 (2001).
38. Djouder, N. et al. Rac and phosphatidylinositol 3-kinaseregulate the protein kinase B in FcεRI signalling in RBL 2H3mast cells. J. Immunol. 166, 1627–1634 (2001).
39. Inabe, K. et al. Vav3 modulates B-cell receptor responses byregulating phosphoinositide 3-kinase activation. J. Exp.Med. 195, 189–200 (2002).References 38 and 39 show that the activation of VAVis not dependent on PI3K.
40. Billadeau, D. D., Mackie, S. M., Schoon, R. A. & Leibson, P. J.The Rho family guanine nucleotide exchange factor Vav-2regulates the development of cell-mediated cytotoxicity. J. Exp. Med. 192, 381–391 (2000).
41. Villalba, M. et al. A novel functional interaction between Vavand PKCθ is required for TCR-induced T-cell activation.Immunity 12, 151–160 (2000).
42. Ferguson, K. M. et al. Structural basis for discrimination of 3-phosphoinositides by pleckstrin homology domains. Mol.Cell. 6, 373–384 (2000).
43. Lietzke, S. E. et al. Structural basis of 3-phosphoinositiderecognition by pleckstrin homology domains. Mol. Cell. 6, 385–394 (2000).
44. Isakoff, S. J. et al. Identification and analysis of PH domain-containing targets of phosphatidylinositol 3-kinase using anovel in vivo assay in yeast. EMBO J. 17, 5374–5387(1998).
45. Zheng, Y. Dbl family guanine nucleotide exchange factors.Trends Biochem. Sci. 26, 724–732 (2001).
46. Billadeau, D. D. et al. The Vav–Rac pathway in cytotoxiclymphocytes regulates the generation of cell-mediatedkilling. J. Exp. Med. 188, 549–559 (1998).
47. Crespo, P. et al. Rac-1-dependent stimulation of theJNK/SAPK signaling pathway by Vav. Oncogene 13,455–460 (1996).
48. Crespo, P., Schuebel, K. E., Ostrom, A. A., Gutkind, J. S. &Bustelo, X. R. Phosphotyrosine-dependent activation ofRac-1 GDP/GTP exchange by the Vav proto-oncogeneproduct. Nature 385, 169–172 (1997).
49. Meffre, E., Casellas, R. & Nussenzweig, M. C. Antibodyregulation of B-cell development. Nature Immunol. 1, 379–385 (2000).
50. Doody, G. M. et al. Signal transduction through Vav-2participates in humoral immune responses and B-cellmaturation. Nature Immunol. 2, 542–547 (2001).
51. Tedford, K. et al. Compensation betwenn Vav1 and Vav2 inB-cell receptor development and antigen-receptorsignalling. Nature Immunol. 2, 548–555 (2001).References 50 and 51 describe the redundant andnon-redundant roles of Vav proteins in lymphocytesusing mouse models.
52. Glassford, J. et al. Vav is required for cyclin D2 induction andproliferation of mouse B lymphocytes activated via theantigen receptor. J. Biol. Chem. 276, 41040–41048 (2001).
53. Tarakhovsky, A. et al. Defective antigen receptor-mediatedproliferation of B and T cells in the absence of Vav. Nature374, 467–470 (1995).
54. Zhang, R., Alt, F. W., Davidson, L., Orkin, S. H. & Swat, W.Defective signalling through the T- and B-cell antigenreceptors in lymphoid cells lacking the vav proto-oncogene.Nature 374, 470–473 (1995).
55. Gulbranson-Judge, A. et al. Defective immunoglobulin class switching in Vav-deficient mice is attributable tocompromised T-cell help. Eur. J. Immunol. 29, 477–487(1999).
56. Fehling, H. J. & von Boehmer, H. Early αβ T-celldevelopment in the thymus of normal and genetically alteredmice. Curr. Opin. Immunol. 9, 263–275 (1997).
57. Fischer, K. et al. Defective T-cell receptor signalling andpositive selection of Vav-deficient CD4+CD8+ thymocytes.Nature 374, 474–477 (1995).
58. Turner, M. et al. A requirement for the Rho-family GTPexchange factor Vav in positive and negative selection ofthymocytes. Immunity 7, 451–460 (1997).
59. Gomez, M., Tybulewicz, V. & Cantrell, D. A. Control of pre-T-cell proliferation and differentiation by the GTPase Rac-1.Nature Immunol. 1, 348–352 (2000).
60. Corre, I., Gomez, M., Vielkind, S. & Cantrell, D. A. Analysis ofthymocyte development reveals that the GTPase RhoA is apositive regulator of T-cell receptor responses in vivo. J. Exp.Med. 194, 903–913 (2001).
61. Fischer, K. D. et al. Vav is a regulator of cytoskeletalreorganisation mediated by the T-cell receptor. Curr. Biol. 8, 554–562 (1998).
62. Chan, G., Hanke, T. & Fischer, K.-D. Vav1 regulates NK T-cell development and NK-cell cytotoxicity. Eur. J. Immunol.31, 2403–2410 (2001).
63. Holsinger, L. J. et al. Defects in actin-cap formation in Vav-deficient mice implicate an actin requirement for lymphocytesignal transduction. Curr. Biol. 8, 563–572 (1998).This reference, together with reference 62, was thefirst to document cytoskeletal impairment in Vav1-deficient T cells.
64. Costello, P. S. et al. The Rho-family GTP exchange factorVav is a critical transducer of T-cell receptor signals to thecalcium, ERK and NF-κB pathways. Proc. Natl Acad. Sci.USA 96, 3035–3040 (1999).
65. Kong, Y. Y. et al. Vav regulates peptide-specific apoptosis inthymocytes. J. Exp. Med. 188, 2099–2111 (1998).
66. Moores, S. L. et al. Vav family proteins couple to diverse cell-surface receptors. Mol. Cell. Biol. 20, 6364–6373 (2000).
67. Krawczyk, C. et al. Cbl-b is a negative regulator of receptorclustering and raft aggregation in T cells. Immunity 13,463–473 (2000).
68. Bachmann, M. F. et al. The guanine-nucleotide exchangefactor Vav is a crucial regulator of B-cell receptor activationand B-cell responses to non-repetitive antigens. J. Immunol.163, 137–142 (1999).
69. Penninger, J. M. et al. The oncogene product Vav is a crucialregulator of primary cytotoxic T-cell responses but has noapparent role in CD28-mediated co-stimulation. Eur. J.Immunol. 29, 1709–1718 (1999).
70. Raulet, D. H., Vance, R. E. & McMahon, C. W. Regulation ofthe natural killer cell receptor repertoire. Annu. Rev. Immunol.19, 291–331 (2001).
71. Galandrini, R., Palmieri, G., Piccloi, M., Fragati, L. & Santoni, A.Role for the Rac1 exchange factor Vav in the signallingpathways leading to NK-cell cytotoxicity. J. Immunol. 162,3148–3152 (1999).
72. Jiang, K. et al. Pivotal role of phosphoinositide-3 kinase inregulation of cytotoxicity in natural killer cells. NatureImmunol. 1, 419–425 (2000).
73. Colucci, F. et al. Functional dichotomy in NK-cell signalling:Vav1-dependent and -independent mechanisms. J. Exp.Med. 193, 1413–1424 (2001).
74. Wedemeyer, J., Tsai, M. & Galli, S. J. Roles of mast cells ininnate and acquired immunity. Curr. Opin. Immunol. 12,624–631 (2000).
75. Manetz, T. S. et al. Vav1 regulates phospholipase Cγactivation and calcium responses in mast cells. Mol. Cell.Biol. 21, 3763–3774 (2001).This paper was the first to document a defect in theproduction of PtdInsP3 in Vav1-deficient mast cells.
76. Liu, B. P. & Burridge, K. Vav2 activates Rac1, Cdc42 andRhoA downstream from growth-factor receptors but notβ1-integrins. Mol. Cell. Biol. 20, 7160–7169 (2000).
77. Abe, K. et al. Vav2 is an activator of Cdc42, Rac1 and RhoA.J. Biol. Chem. 275, 10141–10149 (2000).
78. Marignani, P. A. & Carpenter, C. L. Vav2 is required for cellspreading. J. Cell. Biol. 154, 177–186 (2001).
79. Movilla, N., Dosil, M., Zheng, Y. & Bustelo, X. R. How Vavproteins discriminate the GTPases Rac1 and RhoA fromCdc42. Oncogene 20, 8057–8065 (2001).
80. Reynolds, L. F. et al. Vav1 transduces T-cell receptor signalsto the activation of phospholipase Cγ1 via phosphoinositide-3-kinase-dependent and -independent pathways. J. Exp.Med. 195, 1103–1114 (2002).Together with references 39 and 75, this studydocuments the impairment of the PI3K–Tec axis inVav-deficient cells.
81. Wulfing, C., Bauch, A., Crabtree, G. R. & Davis, M. M. The Vav exchange factor is an essential regulator in actin-dependent receptor translocation to thelymphocyte–antigen-presenting cell interface. Proc. NatlAcad. Sci. USA 97, 10150–10155 (2000).
82. Villalba, M. et al. Vav1/Rac-dependent actin cytoskeletonreorganisation is required for lipid-raft clustering in T cells. J. Cell. Biol. 155, 331–338 (2001).
83. Lee, K. et al. T-cell receptor signalling precedesimmunological synapse formation. Science 295, 1539–1542(2002).
84. Kaminuma, O., Deckert, M., Elly, C., Liu, Y. & Altman, A.Vav–Rac1-mediated activation of the c-Jun N-terminalkinase/c-Jun/AP-1 pathway plays a major role in stimulationof the distal NFAT site in the interleukin-2 gene promoter.Mol. Cell. Biol. 21, 3126–3136 (2001).
85. Wu, J., Katzav, S. & Weiss, A. A functional T-cell receptorsignaling pathway is required for p95vav activity. Mol. Cell.Biol. 15, 4337–4346 (1995).
86. Abe, K., Whitehead, I. P., O’Bryan, J. P. & Der, C. J.Involvement of NH2-terminal sequences in the negativeregulatiuon of Vav signalling and transforming activity. J. Biol.Chem. 274, 30410–30418 (1999).
87. Lin, X., O’Mahoney, A., Geleziunas, R. & Greene, W. C.Protein kinase Cθ participates in NF-κB/Rel activationinduced by CD3/CD28 costimulation through selectiveactivation of IκBβ (IKKβ). Mol. Cell. Biol. 20, 2933–2940(2000).
88. Coudronniere, N., Villalba, M., Englund, N. & Alyman, A. NF-κB activation induced by CD28 costimulation is mediatedby PKCθ. Proc. Natl Acad. Sci. USA 97, 3394–3399 (2000).
89. Teramoto, H., Salem, P., Robbins, K., Bustelo, X. & Gutkind, J.Tyrosine phosphorylation of the Vav proto-oncogeneproduct links FcεRI to the Rac1–JNK pathway. J. Biol.Chem. 272, 10751–10755 (1997).
90. Song, J. S. et al. Tyrosine phosphorylation of Vav stimulatesIL-6 production in mast cells by a Rac/JNK-dependentpathway. J. Immunol. 163, 802–810 (1999).
91. Sun, Z. et al. PKC-θ is required for TCR-induced NF-κBactivation in mature but not immature T lymphocytes.Nature 404, 402–407 (2000).
92. Bakash, S. & Burakoff, S. J. The role of calcineurin inlymphocyte activation. Semin. Immunol. 12, 405–415 (2000).
93. Tolias, K. F., Cantley, L. C. & Carpenter, C. L. Rho familyGTPases bind to phosphoinositide kinases. J. Biol. Chem.270, 17656–17659 (1995).
94. Nisitani, S., Kato, R. M., Rawlings, D. J., Witte, O. N. &Wahl, M. I. In situ detection of activated Bruton’s tyrosinekinase in the Ig signalling complex by phosphopeptide-specific monoclonal antibodies. Proc. Natl Acad. Sci. USA96, 2221–2226 (1999).
95. Lewis, C. M., Broussard, C., Czar, M. J. & Schwartzberg, P. L.Tec kinases: modulators of lymphocyte signalling anddevelopment. Curr. Opin. Immunol. 13, 317–325 (2001).
96. Shigematsu, H. et al. Role of the vav proto-oncogeneproduct (Vav) in erythropoietin-mediated cell proliferationand phosphatidylinositol 3-kinase activity. J. Biol. Chem.272, 14334–14340 (1997).
97. Bertagnolo, V., Marchisio, M., Violina, S., Caramelli, E. &Capitani, S. Nuclear association of tyrosine-phosphorylatedVav to phospholipase Cγ1 and phosphoinositide 3-kinaseduring granulocytic differentiation of HL-60 cells. FEBS Lett.441, 480–484 (1998).
98. Zeng, L. et al. Vav3 mediates receptor protein tyrosinekinase signaling, regulates GTPase activity, modulates cellmorphology and induces cell transformation. Mol. Cell. Biol.20, 9212–9224 (2000).
99. Bokoch, G. M., Vlahos, C. J., Wang, Y., Knaus, U. G. &Traynor-Kaplan, A. E. Rac GTPase interacts specifically withphosphatidylinositol 3-kinase. Biochem. J. 315, 775–779(1996).

486 | JULY 2002 | VOLUME 2 www.nature.com/reviews/immunol
R E V I E W S
100. Sato, S. et al. IL-5 receptor-mediated tyrosinephosphorylation of SH2/SH3-containing proteins andactivation of Bruton’s tyrosine and Janus 2 kinases. J. Exp.Med. 180, 2101–2111 (1994).
101. Guinamard, R., Fougereau, M. & Seckinger, P. The SH3domain of Bruton’s tyrosine kinase interacts with Vav,Sam68 and EWS. Scand. J. Immunol. 45, 587–595 (1997).
102. Bunnell, S. C. et al. Identification of Itk/Tsk Src homologydomain ligands. J. Biol. Chem. 271, 25646–25656 (1996).
103. Machide, M., Mano, H. & Todokoro, K. Interleukin-3 anderythropoietin induce association of Vav with Tec kinasethrough Tec homology domain. Oncogene 11, 619–625(1995).
104. Takahashi-Tezuka, M. et al. Tec tyrosine kinase links thecytokine receptors to PI-3 kinase probably through JAK.Oncogene 14, 2273–2282 (1997).
105. Yablonski, D., Kadlecek, T. & Weiss, A. Identification of aphospholipase Cγ1 (PLC-γ1) SH3 domain-binding site inSLP-76 required for T-cell receptor-mediated activation ofPLC-γ1 and NFAT. Mol. Cell. Biol. 21, 4208–4218 (2001).
106. Ramos Morales, F., Druker, B. J. & Fischer, S. Vav binds toseveral SH2/SH3-containing proteins in activatedlymphocytes. Oncogene 9, 1917–1923 (1994).
107. Kranewitter, W. J. & Gimona, M. N-terminally truncated Vavinduces the formation of depolymerization-resistant actinfilaments in NIH 3T3 cells. FEBS Lett. 455, 123–129 (1999).
108. Katzav, S., Martin, Z. D. & Barbacid, M. Vav, a novelhuman oncogene derived from a locus ubiquitously
expressed in hematopoietic cells. EMBO J. 8, 2283–2290(1989).
109. Adams, J. M., Houston, H., Allen, J., Lints, T. & Harvey, R.The hematopoietically expressed vav proto-oncogeneshares homology with the dbl GDP–GTP exchange factor,the bcr gene and a yeast gene (CDC24) involved incytoskeletal organization. Oncogene 7, 611–618 (1992).
110. Schuebel, K. E. et al. Isolation and characterization ofmurine Vav-2, a member of the Vav family of proto-oncogenes. Oncogene 13, 363–371 (1996).
111. Trenkle, T., McClelland, M., Adlkofer, K. & Welsh, J. Majortranscript variants of VAV3, a new member of the VAV familyof guanine nucleotide exchange factors. Gene 245,139–149 (2000).
112. Fruman, D. A., Satterthwaite, A. B. & Witte, O. N. Xid-likephenotypes: a B-cell signalosome takes shape. Immunity13, 1–3 (2000).
113. Rawlings, D. J. et al. Mutation of unique region of Bruton’styrosine kinase in immunodeficient XID mice. Science 261,358–361 (1993).
114. DeFranco, A. L. Vav and the B-cell signalosome. NatureImmunol. 2, 482–484 (2001).
115. Schaeffer, E. M. et al. Requirement for Tec kinases Rlk andItk in T-cell receptor signaling and immunity. Science 284,638–641 (1999).
116. Ebinu, J. O. et al. RasGRP links T-cell receptor signalling toras. Blood 95, 3199–3203 (2000).
117. Fackler, O. T., Luo, W., Geyer, M., Alberts, A. S. & Peterlin, B. M. Activation of Vav by Nef induces cytoskeletal
rearrangements and downstream effector functions. Mol.Cell. 3, 729–739 (1999).
AcknowledgementsWe thank L. Clayton for assistance with the figures, P. Leibson forcritical review of the manuscript and R. T. Abraham, T. Kurosaki,W. Swat and V. J. L. Tybulewicz for the communication of resultsbefore publication.
Online links
DATABASESThe following terms in this article are linked online to:Entrez: http://www.ncbi.nlm.nih.gov/Entrez/HIV-1 | Listeria monocytogenes | NefInterPro: http://www.ebi.ac.uk/interpro/CH domain | DH domain | ITAM | PH domain | SH2 domain | SH3domain | WW motifLocusLink: http://www.ncbi.nlm.nih.gov/LocusLink/AP1 | β2-microglobulin | BLNK | Btk | Cbl-b | CD3 | CD4 | CD19 |CD28 | CDC42 | Cdc42 | cyclin D2 | DBL | FAS ligand | FcεRI |FYN | Gads | GRB2 | HCK | IFN-γ | IκBα | IKK | IL-2 | IL-4 | Itk | LAT | LCK | NFAT | NF-κB | PAK1 | PI3K | PIP5K | PKCθ | PLC-γ1 |PLC-γ2 | RAC1 | Rac1 | RAC2 | RHOA | RhoA | RHOB | RHOG |Rlk | SLP76 | Slp76 | SRC | SYK | Tec | vav | VAV1 | Vav1 | VAV2 |Vav2 | VAV3 | Vav3 | ZAP70Access to this interactive links box is free online.