afab volume 4 issue 2
DESCRIPTION
This journal is a peer reviewed scientific forum for the latest advancements in bacteriology research on a wide range of topics including food safety, food microbiology, gut microbiology, biofuels, bioremediation, environmental microbiology, fermentation, probiotics, and veterinary microbiology.TRANSCRIPT

Volume 4, Issue 22014
ISSN: 2159-8967www.AFABjournal.com

72 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 73
Sooyoun Ahn University of Florida, USA
Walid Q. Alali University of Georgia, USA
Kenneth M. Bischoff NCAUR, USDA-ARS, USA
Debabrata Biswas University of Maryland, USA
Claudia S. Dunkley University of Georgia, USA
Michael Flythe USDA, Agricultural Research Service
Lawrence Goodridge McGill University, Canada
Leluo Guan University of Alberta, Canada
Joshua Gurtler ERRC, USDA-ARS, USA
Yong D. Hang Cornell University, USA
Armitra Jackson-Davis Alabama A&M University, USA
Divya Jaroni Oklahoma State University, USA
Weihong Jiang Shanghai Institute for Biol. Sciences, P.R. China
Michael Johnson University of Arkansas, USA
Timothy Kelly East Carolina University, USA
William R. Kenealy Mascoma Corporation, USA
Hae-Yeong Kim Kyung Hee University, South Korea
Woo-Kyun Kim University of Georgia, USA
M.B. Kirkham Kansas State University, USA
Todd Kostman University of Wisconsin, Oshkosh, USA
Y. M. Kwon University of Arkansas, USA
Maria Luz Sanz MuriasInstituto de Quimica Organic General, Spain
Melanie R. Mormile Missouri University of Science and Tech., USA
Rama Nannapaneni Mississippi State University, USA
Jack A. Neal, Jr. University of Houston, USA
Benedict Okeke Auburn University at Montgomery, USA
John Patterson Purdue University, USA
Toni Poole FFSRU, USDA-ARS, USA
Marcos Rostagno LBRU, USDA-ARS, USA
Roni Shapira Hebrew University of Jerusalem, Israel
Kalidas Shetty North Dakota State University, USA
EDITORIAL BOARD

74 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
EDITOR-IN-CHIEFSteven C. RickeUniversity of Arkansas, USA
EDITORSTodd R. CallawayFFSRU, USADA-ARS, USA
Philip G. CrandallUniversity of Arkansas, USA
Janet Donaldson Mississippi State University, USA
Ok-Kyung KooKorea Food Research Institute, South Korea
MANAGING and LAYOUT EDITOREllen J. Van LooGhent, Belgium
TECHNICAL EDITORJessica C. ShabaturaFayetteville, USA
ONLINE EDITION EDITORC.S. ShabaturaFayetteville, USA
ABOUT THIS PUBLICATION
Agriculture, Food & Analytical Bacteriology (ISSN
2159-8967) is published quarterly.
Instructions for Authors may be obtained at the
back of this issue, or online via our website at
www.afabjournal.com
Manuscripts: All correspondence regarding pend-
ing manuscripts should be addressed Ellen Van Loo,
Managing Editor, Agriculture, Food & Analytical
Bacteriology: [email protected]
Information for Potential Editors: If you are interested
in becoming a part of our editorial board, please con-
tact Editor-in-Chief, Steven Ricke, Agriculture, Food &
Analytical Bacteriology: [email protected]
Advertising: If you are interested in advertising with
our journal, please contact us at advertising@afab-
journal.com for a media kit and current rates.
Reprint Permission: Correspondence regarding re-
prints should be addressed Ellen Van Loo, Managing
Editor, Agriculture, Food & Analytical Bacteriology
Ordering Print Copies: print editions of this journal
may be purchased and shipped internationally from
our website order form at www.afabjournal.com
Subscription Rates: Subscriptions are not available
at this time. To be advised when subscriptions plans
are made available, please join our newsletter at
www.afabjournal.com
Mailing Address: 2138 Revere Place . Fayetteville, AR . 72701 Website: www.AFABjournal.com
EDITORIAL STAFF

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 75
Antibiotic Use in Livestock ProductionBroadway, P. R., J. A. Carroll, and T. R. Callaway
76
Effects of Co-nutrients in Foods and Bioremediation in the Environment on Methylmercury
P. G. Crandall, C. A. O’Bryan
86
Alternative antimicrobial supplements that positively impact animal health and food safety Broadway, P. R., J. A. Carroll, and T. R. Callaway
109
Human Health Benefits of Isoflavones from Soybeansk. Kushwaha, C. A. O’Bryan, D. Babu, P. G. Crandall, P. Chen, and S.-O. Lee
122
REVIEW
Contribution of Chemical and Physical Factors to Zoonotic Pathogen Inactivation during Chicken Manure CompostingM.C. Erickson, J. Liao, X. Jiang, and M.P. Doyle
96
ARTICLES
Instructions for Authors147
Introduction to Authors
The publishers do not warrant the accuracy of the articles in this journal, nor any views or opinions by their authors.
TABLE OF CONTENTS

76 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
www.afabjournal.comCopyright © 2014
Agriculture, Food and Analytical Bacteriology
ABSTRACT
Antibiotic usage is a useful and commonly implemented practice in livestock and production ag-
riculture that has progressively gained attention in recent years from consumers of animal products due
to concerns about human and environmental health. Sub-therapeutic usage of antibiotics has led to a
concern that prophylactic supplementation leads to antimicrobial resistance, and this particular practice
has come under public scrutiny. The consumer and media misconceptions about antibiotic usage and pro-
duction strategies utilized in livestock production have caused a shift in consumer demands. Antibiotics
directly and indirectly affect the livestock industry by treating illness and promoting the overall health of
the animal, which may enhance production parameters such as growth and profitability. However, pending
legislation threatens to eliminate the current antibiotic usage strategies implemented by producers. This
review will address the historical and current use of antibiotics as it pertains to production animal agricul-
ture to summarize how antibiotics promote animal health and growth performance.
Keywords: Antibiotic, livestock, animal health, review
INTRODUCTION
Antibiotic usage in meat animal production is
a hotly debated issue in the livestock industry that
has acquired more attention as consumers seek to
place more “natural” and “safer” products on their
Correspondence: Todd Callaway, [email protected]: +1-979-260-9374 Fax: +1-979-260-9332.
table (Gilbert and McBain, 2003). Consumer percep-
tion can greatly influence food animal production as
has been recently observed for some common food
production practices; such as lean finely textured
beef (“pink slime”) which was removed from meat
formulations of producers due to negative media at-
tention and consumer perception (Flock, 2012). The
use of gestation crates in swine production has also
drawn increasing attention, leading to the refusal of
REVIEWAntibiotic Use in Livestock Production
P. R. Broadway1, J. A. Carroll2, and T. R. Callaway3
1Department of Animal and Food Sciences, Texas Tech University, Lubbock, TX2Livestock Issues Research Unit, Agricultural Research Service, USDA, Lubbock, TX
3Food and Feed Safety Research Unit, Southern Plains Agricultural Research Center, Agricultural Research Service, USDA, College Station, TX
“Proprietary or brand names are necessary to report factually on available data; however, the USDA neither guarantees nor warrants the standard of the product, and the use of the name by the USDA implies no approval of the product, or exclusion of others that may
be suitable.” USDA is an equal opportunity provider and employer
Agric. Food Anal. Bacteriol. 4: 76-85, 2014

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 77
some retailers and restaurants to purchase pork from
producers that utilize gestation crates (Food Safety
News, 2013). Furthermore, antibiotic usage in ani-
mals for health benefits and growth promotion has
continued to be a concern of the American consum-
er in recent years. In response to similar concerns,
the European Union (EU) banned sub-therapeutic
supplementation of animal feeds with antibiotics
(Pradella, 2006). Recently, the U.S. Food and Drug
Administration issued a guidance directive on the
judicious use of antibiotics in food animals, and this
measure has led some to believe that this is phase
one of an agenda to remove sub-therapeutic antibi-
otic use from livestock production.
The gastrointestinal tract of animals is populated
with a complex microbial ecosystem that is essen-
tial for the function, growth, and overall health of
the animal (Chaucheyras-Durand and Durand, 2010).
Many livestock producers currently utilize feeding
and production strategies, including the use of an-
tibiotics, that alter the microbial ecology of the gas-
trointestinal tract of the animal to benefit the overall
health and production efficiency of their animals. As
a bonus to the consumer, some of these strategies
may also help eliminate or reduce foodborne patho-
gens that may contaminate the food supply (Perl-
man, 1973). If and when sub-therapeutic antibiotic
use in food animals is banned in the U.S., alternative
strategies must be implemented to replicate these
positive effects in order for the livestock industry to
remain viable.
CURRENT USE OF ANTIBIOTICS IN LIVE-STOCK
Antibiotics are used in the livestock industry for
a variety of reasons including treatment of disease,
prophylaxis, as well as improving feed efficiency
and overall growth performance (Berge et al., 2005;
Brown et al., 1975). While antibiotics do not make
label claims that suggest alteration of growth param-
eters in livestock, the association between their use
and growth promotion has been reported in many
species such as cattle, swine, and poultry for over 50
years (Moore et al., 1946; Jukes et al., 1950; Rogers et
al., 1995; Salinas-Chavira et al., 2009). Performance
parameters can be quantitatively measured in a va-
riety of ways including, but not limited to: mortality,
weight gain, meat/milk quality, and feed efficiency.
While the mode of action by which antibiotics im-
prove feed efficiency has not been fully elucidated,
growth performance may be enhanced due to de-
creased inflammation in the small intestine (Feighner
and Dashkevicz, 1987; Eyssen and DeSomer, 1963).
To further explain how antibiotics may work in con-
junction to promote animal health and food safety,
McCracken and Gaskins (1999) indicated that the de-
velopment of the intestinal immune system occurs
in conjunction with the development of the normal
microflora of the animal; however chronic stimula-
tion of the immune system may decrease the amount
of protein available for growth (Gordon et al., 1963).
Studies comparing germ-free and conventionally
raised animals have demonstrated this phenomenon
and have reported alterations in immune function of
these animals in conjunction with the development
of the intestinal microflora (McCraken and Lorenz,
2001). Thinning of the intestinal epithelium in con-
junction with the use of antibiotics may be the result
of decreased microbial production of polyamines and
volatile fatty acids (VFAs) that enhance intestinal cell
growth and activity (Ferket et al., 2002). Ferket et al.
(2002) states that intestinal mucosal thinning that may
occur with the use of antibiotics may increase energy
availability for growth because the animal does not
have to maintain a larger intestinal mucosal layer.
Cattle
Antibiotics have been used for decades in cattle,
and some of the most commonly used antibiotics in
the feedlot setting are a class of compounds known
as ionophores (Russell and Strobel, 1989). Iono-
phores were approved for use in ruminants in the
1970s (Russell and Strobel, 1989). The ionophore
monensin was fed to chickens as a coccidiostat, and
the manure from these poultry houses was spread
on cattle pastures as a fertilizer. Cattle grazing these
pastures grew more rapidly than cattle grazing pas-

78 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
tures fertilized with manure from poultry houses
where the chickens were not fed monensin (Callaway,
2013). As a result, the ionophore monensin was di-
rectly incorporated into cattle rations beginning in
the 1970’s, and this compound has been reported
to enhance growth performance through a variety
of modifications of the ruminal microbiome (Raun et
al., 1976; Callaway et al., 2003). Ionophores primar-
ily inhibit bacteria with Gram positive physiology, in-
cluding lactic acid bacteria, and this improves growth
efficiency, average daily gain (ADG), reduces waste-
ful protein degradation (by hyperammonia produc-
ing bacteria), reduces methanogenesis, and reduces
ruminal acidosis via lower lactate production (Russell
and Strobel, 1988). Ionophores have been reported
to reduce liver abscesses by inhibiting epithelial ke-
ratinization caused by lactic acidosis and subsequent
Fusobacterium necrophorum infections (Nagaraja
and Chengappa, 1998; Lechtenberg et al., 1998).
While compounds such as ionophores alter the
microbial ecology of the gastrointestinal tract to pro-
mote overall health and performance, other antibi-
otics are used to treat specific bacterial disease and
illness. Some of these antibiotics may also elicit a
dual effect, promoting both health and performance
in the animals. Bovine respiratory disease (BRD) is
the most common and expensive disease present in
American cattle, and the use of antibiotics to treat/
prevent this disease is a great example of this dual
effect of antibiotics (Smith, 1998; Snowder et al.,
2006). Bovine respiratory disease is a complex dis-
ease caused by exposure to various viral (e.g., Infec-
tious Bovine Rhino-tracheitis, Bovine Viral Diarrhea,
Bovine Respiratory Syncytial Virus, and Parainfluenza
Virus) and/or bacterial (e.g.., Pasteurella hemolytica,
Pasteurella multocida, Haemophilussomnus, Myco-
plasmasp. and Actinomycespyogenes) pathogens.
Bovine respiratory disease may be mitigated in a
number of ways including vaccination, management
practices, and antibiotic treatments to prevent and/
or treat the disease. Addition of chloratetracycline
and sulfamethazine to treat enteritis, coccidiosis, and
bovine respiratory disease (BRD) in the ration of cattle
arriving at the feed lot was also reported to increase
ADG while decreasing the risk of bovine respiratory
disease for the first 28 days at the feedlot (Guillermo
and Berg, 1995; Smith et al., 1993). Another com-
monly used antibiotic in beef production is Tilmico-
sin which is a broad spectrum antibiotic used to treat
and prevent BRD. Tilmicosin works to inhibit protein
synthesis of bacteria such as Pasteurella hemolytica
that may lead to the onset of BRD. Treatment of cat-
tle upon arrival into feedlots with Micotil®, a solution
of Tilmicosin, was shown to decrease BRD symptoms
and increase dry matter intake (Galyean et al., 1995).
Antibiotics are also used in livestock to prevent
specific physiologic disorders such as ruminal lactic
acidosis, a common problem in grain fed cattle that
can be chronic or acute and range from moderate to
severe (Nagaraja and Titgemeyer, 2007; Slyter, 1976;
Muir et al., 1981; Nagaraja et al., 1982). Ruminal aci-
dosis is the accumulation of lactate in the rumen re-
sulting in a lowered pH that decreases animal growth
performance parameters, and leads to the devel-
opment of other health problems such as laminitis,
bloat, and liver abscesses (Nagaraja and Chengappa,
1998; Nocek, 1997; Enemark, 2008). In acute clinical
lactic acidosis, D-lactate is the acid primarily respon-
sible for this condition (Dunlop, 1965); however, the
role of lactate in sub-acute acidosis is not fully under-
stood (Enemark, 2009). The onset of acidosis is linked
with feeding readily fermentable carbohydrates that
are commonly associated with a high concentrate ra-
tion as would normally be fed in the cattle feedlot
or swine finishing production systems (Owens et al.,
1998; Russell and Hino, 1985).
Antibiotics/antimicrobials and other feedstuffs
have been reported to be effective strategies to pre-
vent the onset of ruminal acidosis (Owens et al.,1998;
Callaway et al., 2003). Antibiotics may decrease the
incidence of liver abscesses in cattle which may be
the result of ruminal acidosis and may predict carcass
performance (Rogers et al., 1995; Brown and Law-
rence, 2010). Virginamycin is an antibiotic used to
prevent necrotic enteritis in cattle and has also been
reported to increase the gain to feed ratio in cattle
(Salinas-Chavira et al., 2009). Rogers et al. (1995) re-
ported an increase in ADG and feed conversion, and
a decrease in liver abscesses in cattle fed virginamy-
cin.

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 79
Swine
As in ruminants, such as cattle, antibiotics are used
in swine production for many of the same reasons.
These pharmaceuticals are used in swine for both pro-
phylactic and treatment therapies, and in some cases,
these antibiotics can also effect performance parame-
ters. Jensen et al. (1955) reported increased gains and
feed conversion in swine fed the antibiotic aureomy-
cin. While aeuromycin was also initially reported to en-
hance reproductive performance in swine (Yestal et al.,
1952), subsequent work by Davey et al. (1955) reported
no difference in reproductive performance when swine
were fed various concentrations of the antibiotic. Via-
bility and performance of newborn and suckling piglets
was also unaffected when swine were supplemented
with aureomycin (Davey et al., 1955). Aureomycin was
further reported to increase profitability by increasing
belly weight and decreasing backfat thickness (Perry et
al., 1953). Zimmerman (1986) reported that antibiot-
ics such as chloratetracyline, furazolidone, lincomycin,
salinomycin, tylosin, and virginamycin may improve
average weight gain by approximately 15%. Addition-
ally, Zimmerman (1986) reported that combined use
of chloratetracycline, penicillin, and sulfamethazine
(2:1:2) increased ADG in starter pigs by 25%. Multiple
studies in swine also indicate that treatment by any of
the aforementioned antibiotics can increase farrowing
rate (Zimmerman, 1986; Ruiz et al., 1968; Anderson,
1969; Hays 1978). Litter size may also be increased with
the addition of a combination of antibiotics (Zimmer-
man, 1986; Ruiz et al., 1968; Hays 1978). The antibiotics
penicillin and streptomycin increased the growth rate
of swine fed to market weight (Bridges et al., 1952).
Penicillin and streptomycin used in conjunction are still
approved for use in the swine industry, as well as bo-
vine, equine, and ovine species, to treat bacteria such
as Arcanobacterium, Klebsiella pneumonia, Listeria
spp., Mannheimia haemolytica, Pasteurella, Staphy-
lococcus, and Salmonella (Norbrook Laboratories,
2013). Tylosin is another antibiotic approved for use in
swine that can be provided via intramuscular injection,
feed, or water, and is effective in preventing and con-
trolling porcine proliferative enteropathy (ileitis; Para-
dis, 2004; Marseller et al., 2001; McOrist et al., 1997).
Tylosin supplemented in the drinking water of swine
for 17 days decreased clinical signs of gastrointestinal
infection and promoted growth performance (Paradis
et al., 2004). Tylosin-supplemented swine showed no
clinical or pathological signs of proliferative enteropa-
thy (ileitis) after experimental infection with Lawsonia
intracellularis (McOrist et al., 1997). The mitigation of
disease in concert with enhanced growth and repro-
ductive performance as a result of antibiotic usage in
swine help make the use of antibiotics a profitable pro-
duction strategy (Zimmerman, 1986).
Poultry
Antibiotic usage is an extremely important as-
pect of poultry production and has been used in
production and researched extensively since the
1950s (Feighner and Dashkevicz, 1987). Antibiotics
used in poultry production are believed to be effec-
tive growth promotants due to the alterations they
induce in the microflora of the gastrointestinal tract
(Feighner and Dashkevicz, 1987). This theory is sup-
ported by experiments that report germ-free chick-
ens grow more efficiently than commercially raised
poultry, and germ-free animals do not grow faster
when given antibiotics with growth promoting capa-
bilities (Coates et al., 1963; Forbes and Pank, 1959).
In poultry, antibiotic feeding has been reported to
increase weight gain and feed conversion efficiency
(feed/gain; Feighner and Dashkevicz, 1987; Bunyan
et al., 1977). Feed efficiency has been reported to
be improved in poultry supplemented with antibiot-
ics by reducing microbial populations in competition
for nutrients and reduction of pathogenic bacteria
(Feighner and Dashkevicz, 1987; Eyssen and de-
Somer, 1963; Barnes et al., 1978). Studies have re-
ported that ammonia production by bacteria in the
GI tract of monogastrics may suppress growth (Dang
and Visek, 1960; Harbers et al., 1963; Visek, 1978).
Deconjugation of bile salts may also play a role in
growth suppression due to Streptococcus faecium
in the small intestine; however, the use of antibiotics
has been reported to reduce attachment of this bac-
terium to intestinal epithelia (Cole and Fuller, 1984;
Fuller et al., 1984; Fuller et al., 1983).

80 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
Table 1. Some antibiotics used in animal agriculture that may be used to promote overall animal health and impact pathogen colonization and shedding
Some Antibiotics used in Animal Agriculture By Species
Cattlea Swineb Poultryc Aquacultured
Amoxicillin Apramycin Ardacin Amoxicillin
Ampicillin Bacitracin Avilamycin Ampicillin
Enroflaxin Bambermycin Avoparcin Chloramphenicol
Erythromycin Carbadox Bacitracin manganese Cortimoxazole
Florfenicol Chloratetracycline Erythromycin Enroflaxin
Oxytetracycline Furazolidone Lincomycine Erythromycin
Penicillin Lincomycin Mocimycin Florfenicol
Sulfadimethoxine Nosiheptide Neomycin Furazolidine
Tilmicosin Salinomycin Nosiheptide Nitrofurans
Tylosin Tiamulin Penicillin Oxolinic acid
Tylosin Soframycin Oxytetracycline
Virginamycin Tetracycline Sarafloxacin
Tylosin Streptomycin
Virginamycin Sulphadizine
Trimethoprim-
Sulfamethoxazole
aCurrin and Whittier, 2009b Zimmerman, 1986cCastanon, 2007dDefoirdt et al., 2011

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 81
The intestinal epithelia in poultry and other spe-
cies play a large role in the growth capabilities of
animals, and antibiotics can alter the intestinal micro-
flora as well as the intestinal epithelia of animals to
promote growth. As mentioned previously, thinner
intestinal epithelia may result in more efficient nutri-
ent uptake and absorption (Eyessen and deSomer,
1963; Ford and Coates, 1971; Siddons and Coates,
1972; Sieburth et al., 1951). Also, antibiotics reduce
populations of bacteria in the intestines, thereby
making more nutrients available for animal growth
(Eyssen, 1962; Monson et al., 1954). When antibiotics
reduce the microbial population in the GI tract, they
may inherently reduce pathogens responsible for dis-
ease or subclinical infections (Eyssen and deSomer,
1963a; Eyssen and deSomer, 1963b; Eyssen and de-
Somer, 1967; Sieburth et al., 1951). The combination
of all these effects elicited by antibiotics provides a
possible explanation as to why antibiotics enhance
growth performance and feed efficiency.
Aquaculture
As in mammalian production, antibiotics also play
a critical role in the aquaculture industry. Diseases in
production aquaculture are estimated to cause losses
of approximately 3 billion dollars per year globally
(Subasinghe, et al., 2001). There are more than 100
known pathogens to fish; however, some of these are
opportunistic pathogens (Alderman and Hastings,
1998). One of the main bacterial culprits are Vibrio
bacteria (harveyi, cambellii, and parahaemolyticus;
Defoirdt et al., 2007). While these pathogens are
detrimental to the health of the aquaculture, some
bacteria such as Vibrio cholera and vulnificus, may
cause human disease as well (Thompson et al., 2004).
Some of the antibiotics used in aquaculture are chlor-
amphenicol, gentamycin, trimethorprim, tiamulin,
tetracyclines, quinolones, and sulfonamides (Table 1;
Defoirdt et al., 2007). Most of these antibiotics are in-
corporated into the feed of the aquaculture at speci-
fied dosages with required withdrawal times (Alder-
man and Hastings, 1998). However, countries around
the world have vastly different regulations regarding
the administration, dosage, withdrawal, and control
of antibiotics in aquaculture (Alderman and Hastings,
1998). As in many food-producing species, antibiotic-
resistant bacteria such as Aeromonas salmonicida, A.
hydrophila, Vibrio anguillarum, Pseudomonas fluores-
cens, Pasteurella piscida, and Edwardsiella tarda have
been documented in aquaculture species (Aoki, 1988).
SUMMARY
Antibiotics are an important part of agriculture
and food production originating from the cattle,
swine, poultry, and aquaculture industries, and much
research has been conducted to determine the ef-
ficacy and safety of these pharmaceuticals. These
compounds are used not only to treat disease, but
can also be used effectively as a prophylactic treat-
ment. Such strategies to control pathogens in food-
producing animals may, in some cases, improve
growth performance parameters while simultaneous-
ly promoting the overall health of the animal. Thus,
antibiotics are a critical player in the profitability of
agriculture in the U.S. and throughout the world and
play a vital role in feeding the ever growing world
population. However, an ever changing population
and shifts in consumer demand have placed pres-
sure on the agricultural industry and governments
to reduce and/or eliminate the use of antibiotics in
food production. While this potential change could
possibly be detrimental to current management
strategies, there are potential alternatives to antibi-
otics that have been extensively researched in live-
stock to promote health, performance, profitability,
and food safety.
REFERENCES
Alderman, D.J. and T.S. Hastings. 1998. Antibiotic
use in aquaculture: development of antibiotic re-
sistance- potential for consumer health risk. Int. J.
Food Sci. & Technol. 33:139-155.
Anderson, M.D. 1969. Effect of feeding furazilidone
to gilts and sows at breeding. Hess & Clark, Re-
search Digest. 7(5):1.

82 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
Aoki, T. 1988. Drug-resistant plasmids from fish
pathogens. Microbiol. Sci.. 5:219-223.
Barnes, E.M., G.C. Mead, C.S. Impey, and B.W. Ad-
ams. 1978. The effect of dietary bacitracin on the
incidence of Streptococcus faecalis subspecies
liquefaciencs and related streptococci in the intes-
tines of young chicks. Br. Poult. Sci. 19:713-723.
Berge, A.C.B., P. Lindeque, D.A. Moore, and W.M.
Sischo. 2005. A clinical trial evaluating prophy-
lactic and therapeutic antibiotic use on health and
performance of preweaned calves. J. Dairy Sci.
88:3036.
Bridges, J.H., I.A. Dyer, and W.C. Burkhart. 1952. Ef-
fects on penicillin and streptomycin on the growth
rate and bacterial count in the feces of pigs. J.
Anim. Sci. 11:474-479.
Brown, H., R.F. Bind, H.P. Grueter, J.W. McAskill, C.O.
Cooley, and R.P. Rathmacher. 1975. Tylosin and
chloratetracycline for the prevention of liver ab-
cesses, improved weight gains and feed efficiency
in feedlot cattle. J. Anim. Sci. 40:207-213.
Brown, T.R. and T.E. Lawrence. 2010. Association
of liver abnormalaties with carcass grading perfor-
mance and value. J. Anim. Sci. 88:4037-4043.
Bunyan, J., L. Jeffries, J.R. Sayers, A.L. Gulliver, and
K. Coleman. 1977. Antimicrobial substances and
chick growth promotion: the growth-promoting
activities of antimicrobial substances, including
fifty-two used either in therapy or as dietary addi-
tives. Br. Poult. Sci. 18:283-294.
Callaway, 2013. Personal communication about
monenesin on cattle pastures.
Callaway, T. R., T. S. Edrington, et al. (2003). Iono-
phores: Their use as ruminant growth promotants
and impact on food safety. Curr. Iss. Intest. Micro-
biol. 4:43-51.
Callaway, T.R., Edrington, T.S., Rychik, J.L., Geno-
vese, K.J., Poole, T.L., Jung, Y.S., Bischoff, K.M.,
Anderson, R.C., and Nisbet, D.J. 2003. Iono-
phores: Their use as ruminant growth promotants
and impact on food safety. Curr. Issues Intest. Mi-
crobiol. 4:43-51.
Castanon, J.I.R. 2007. History of the use of antibiotic
as growth promoters in European poultry feeds.
Poult. Sci. 86:2466-2471.
Chaucheyras-Durand, F. and Durand, H. 2010. Pro-
biotics in animal nutrition and health. Benef. Mi-
crobes. 1:3-9.
Coates, M.E., R. Fuller, G.F. Harrison, M. Lev, and
S.F. Suffolk. 1963. A comparison of the growth
of chicks in the Gustafsson germ-free apparatus
and in a conventional environment, with and with-
out dietary supplements of penicillin. Br. J. Nutr.
17:141-150.
Cole, C.B. and R. Fuller. 1984. Bile acid deconjuga-
tion and attachment of chicken gut bacteria; their
possible role in growth depression. Br. Poult. Sci.
25:227-231.
Currin, J.F. and D. Whittier. 2009. Recognition and
Treatment of Bovine Respiratory Disease Com-
plex. Virginia Cooperative Extension Publication.
Publication 400-008.
Dang, H.C. and W.J. Visek. 1960. Effects of urease
injection on body weights of growing rats and
chicks. Proc. Soc. Exp. Biol. Med. 105:164-167.
Davey, R.J., W.W. Green, and J.W. Stevenson. 1955.
The effect of aureomycin on growth and reproduc-
tion in swine. J. Anim. Sci. 14:507-512.
Defoirdt, T., N. Boon, P. Sorgeloos, W. Verstraete,
and P. Bossier. 2007. Alternatives to antibiotics to
control bacterial infections: luminescent vibriosis
in aquaculture as an example. Trends in Biotech-
nology. 25:472-479.
Defoirdt, T., P. Sorgeloos, and P. Bossier. 2011. Al-
ternatives to antibiotics for the control of bacte-
rial disease in aquaculture. Curr. Opin. Microbiol.
14:251-256.
Dunlop, R.H. and P.B. Hammond. 1965. D-lactic aci-
dosis of ruminants. Annals of the New York Acad-
emy of Science 119:1109-1132.
Enemark, J.M.D. 2009. The monitoring, prevention
and treatment of sub-acute ruminal acidosis (SARA):
a review. The Veterinary Journal 176:32-43.
Eyssen, H. 1962. The additive effects of nucleic
acids and antibiotics as individual growth promo-
tants for chicks. Poult. Sci. 41:1822-1828.
Eyssen, H. and P. deSomer. 1963. Effects of antibi-
otics on growth and nutrient absorption of chicks.
Poult. Sci. 42:1373-1379.
Eyssen, H. and P. deSomer. 1963b. The mode f ac-

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 83
tion of antibiotics in stimulating growth of chicks.
J. Exp. Med. 117:127-138.
Eyssen, H. and P. deSomer. 1967. Effects of Strep-
tococcus faecalis and a filterable agent on growth
and nutrient absorption in gnotobiotic chicks.
Poult. Sci. 46:323-333.
Eyssen, H., and P. deSomer. 1963a. Effects of antibi-
otics on growth and nutrient absorption of chicks.
Poult. Sci. 42:1373-1379.
Feighner, S.D. and M.P. Dashkevicz. 1987. Subther-
apeutic levels of antibiotics in poultry feeds and
their effects on weight gain, feed efficiency, and
bacterial cholytaurine hydrolase activity. Appl. En-
viron. Microbiol. 53:331-336.
Ferket, P.R., C.W. Parks, and J.L. Grimes. 2002. Ben-
efits of dietary antibiotic and mannanoligosaccha-
ride supplementation for poultry. Multi-state poul-
try meeting. May 14-16, 2002.
Flock, E. 2012. Pink slime removed from McDon-
ald’s burgers- but other food additives remain.
The Washington Post. Postted on 2/01/2012.
Available: http://www.washingtonpost.com/blogs/
blogpost/post/pink-slime-removed-from-mcdon-
alds-burgers--but-other-weird-food-additives-re-
main/2012/02/01/gIQAdfvAiQ_blog.html
Food Safety News. 2013. Pork producers agreet o
phase out gestation crates across all of Canada.
Published June, 4, 2013. Food Safety News. Avail-
able: http://www.foodsafetynews.com/2013/06/
gestation-crates-being-phased-out-across-cana-
da/#.UrHN4NJDtVc
Forbes, M. and J.T. Pank. 1959. Growth of germ-free
and conventional chicks: effect of diet, dietary pen-
icillin and bacterial environment. J. Nutr. 67:69-84.
Ford, D.J., and M.E. Coates. 1971. Absorption of
glucose and vitamins of the B complex by germ-
free and conventional chicks. Proc. Nutr. Soc.
30:10-11.
Fuller, R., C.B. Cole, and M.E. Coates. 1984. The
role of Streptococculs faecium in antibiotic-re-
lieved growth depression in chickens, p. 395-403.
In M. Woodbine (ed.), Antimicrobials and agricul-
ture. Butterworths, London.
Fuller, R., S.B. Houghton, and M.E. Coates. 1983.
The effect of dietary penicillin on the growth of
gnotobiotic chickens mono-associated with Strep-
tococcus faecium. Br. Poult. Sci. 24:111-114.
Galyean, M.L., S.A. Gunter, and K.J. Malcom-Callis.
1995. Effects of arrival medication with tilmicosin
phosphate on health and performance of newly
received beef cattle. J. Anim. Sci. 73:1219-1226.
Gilbert, P. and A.J. McBain. 2003. Potential impact
of increased use of biocides in consumer products
on prevalence of antibiotic resistance. Clinical Mi-
crobiology Reviews. 16:189-208.
Gordon, H.A., B.S. Wostmann, and E. Bruckner-Kar-
doss. 1963. Effects of microbial flora on cardiac
output and other elements of blood circulation.
Proc. Soc. Exp. Biol. Med. 114:301-304.
Guillermo, F.G. and J.L. Berg. 1995. Efficacy of a
feed-additive antibacterial combination for im-
proving feedlot cattle performance and health.
Canadian Vet. J. 36:223-229.
Harbers, L.H., A.P. Alvares, A.I. Jacobson, and W.J.
Visek. 1963. Effect of barbituric acid and chora-
tetracycline upon growth, ammonia concentration
and urease activit in the gastrointestinal tract of
chicks. J. Nutr. 80:75-79.
Hays, W.V., J.L. Krug, G.L. Cromwell, R.H. Dutt, and
D.D. Kratzer. 1978. Effect of lactation length and
dietary antibiotics on reproductive performance of
sows. J. Anim. Sci. 46:884.
Jensen, A.H., D.C. Aker, H.M. Maddock, G.C. Ashton,
P.G. Homeyer, E.O. Heady, and D.V. Catron. 1955.
Different protein levels with and without antibiot-
ics for growing-finishing swine: effect on growth
rate and feed efficiency. J. Anim. Sci. 14:69-81.
Jukes, T.H., E.L.R. Stokstad, R.R. Taylor, T.J. Combs,
H.M. Edwards, and G.B. Meadows. 1950. Growth
promoting effect of auromycin on pigs. Arch. Bio-
chem. 26:324-330.
Lechtenberg, K.F., T.G. Nagaraja, and M.M. Chen-
gappa. 1998. Antimicrobial susceptibility of Fu-
sobacterium necrophorum isolated from bovine
hepatic abscesses. Amer. J. Vet. Res. 59:44-47.
Marseller, T., N Winkelman, C. Gebhart, W. Weldon, R.
Muller, J. Weatherford, and L. Symanowski. 2001. Ef-
ficacy of intramuscular tylosin for the treatment and
control of porcine proliferative enteropathy caused
by Lawsonia intracellularis. Vet. Ther. 2:51-60.

84 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
McCraken, V.J. and H.R. Gaskins. 1999. Probiotcs
and the immune system. Pages 85-111 in Probiot-
ics: A critical review. G.W. Tannock, ed. Horizon
Scientific Press, Norfolk, UK.
McCraken, V.J. and R.G. Lorenz. 2001. The gastro-
intestinal ecosystem: a precarious alliance among
epithelium, immunity and microbiota. Cellu. Mi-
crob. 3:1-11.
McOrist, S., J. Morgan, M.F. Veenhuizen, K. Lawrence,
and H.W. Kroger. Oral administration of tylosin
phosphate for treatment and prevention of prolifera-
tive enteropathy in pigs. Am. J. Vet. Res. 58:136-169.
Monson, W.J., A.E. Harper, M.E. Winje, C.A. Elve-
hjem, R.A. Rhodes, and W.B. Sarles. 1954. A
mechanism of the vitamin sparing effect of antibi-
otics. J. Nutr. 52:627-636.
Moore, P.R., A. Evenson, T.D. Luckey, E. McCoy, E.A.
Elvehjem, and E.B. Hart. 1946. Use of suophasucci-
dine, streptothriocin, and streptomycin in nutrition
studies with the chick. J. Biol. Chem. 165:437-441.
Muir, L. A., E. L. Rickes, P.F. Duquette, and G.E. Smith
(1981). Prevention of induced lactic acidosis in cat-
tle by thiopeptin. J. Anim. Sci. 52:635-643.
Nagaraja, T. G., T. B. Avery, E.E. Barley, S.K. Roof, and
A.D. Dayton. (1982). Effect of lasalocid, monensin
or thiopeptin on lactic acidosis in cattle. J. Anim.
Sci. 54:649-658.
Nagaraja, T.G. and E.C. Titgemeyer. 2007. Ruminal
acidosis in beef cattle: the current microbiological
and nutritional outlook. J. Dairy Sci. 90:E17-E38.
Nagaraja, T.G. and M.M. Chengappa. 1998. Liver
abscesses in feedlot cattle: A review. J. Anim. Sci.
76:287-298.
Norbrook Laboratories. 2013. Drug information
sheet: available: http://www.norbrook.com/prod-
ucts/pen-strep-suspension-for-injection/
Owens, F.N., Secrist, D.S., Hill, W.J., and Gill, D.R.
1998. Acidosis in cattle: a review. J. Anim. Sci.
76:275-286.
Perlman, D. Advances in Applied Microbiology.
1973. Vol. 16. Academic Press, Inc. New York, NY.
Perry, T.W., W.M. Beeson, and B.W. Vosteen. 1953.
The effect of an antibiotic or a surfactant on the
growth and carcass composition of swine. J.
Anim. Sci. 12:310-315.
Pradella, G. 2006. Antibiotic ban in the European
Union: A pyrrhic victory? J. Vet. Pharmacol. Ther.
29:41.
Raun, A. P., C. O. Cooley, E.L. Potter, R.P. Rathmach-
er, and L.F. Richardson. (1976). Effect of monensin
on feed efficiency of feedlot cattle. J. Anim. Sci.
43:670-677.
Rogers, J.A., M.E. Branine, C.R. Miller, M.I. Wray, S.J.
Bartle, R.L. Preston, D.R. Gill, R.H. Pritchard, R.P.
Stilborn, and D.T. Bechtol. 1995. Effects of dietary
virginamycin on performance and liver abscess in-
cidence in feedlot cattle. J. Anim. Sci. 73:9-20.
Ruiz, M.E., V.C. Speer, V.W. Hays, and W.P. Switzer.
1968. Effect of feed intake and antibiotic on repro-
duction in gilts. J. Anim. Sci. 27:1602.
Russell, J.B. and H.J. Strobel. 1989. Effect of iono-
phores on ruminal fermentation. Appl. Environ.
Microbiol. 55:1-6.
Russell, J. B. and H. J. Strobel. 1988. Effects of ad-
ditives on in vitro ruminal fermentation: a com-
parison of monensin and bacitracin, another gram-
positive antibiotitc. J. Anim. Sci. 66:552-558.
Russell, J. B. and T. Hino 1985. Regulation of lactate
production in Streptococcus bovis: a spiraling ef-
fect that contributes to rumen acidosis. J. Dairy
Sci. 68:1712-1721.
Salinas- Chavira, J., J. Lenin, E. Ponce, U. Sanchez, N.
Torrentera, and R.A. Zinn. 2009. Comparative ef-
fects of virginamycin supplementation on charac-
teristics of growth-performance, dietary energetic,
and digestion of calf-fed Holstein steers. J. Anim.
Sci. 87:4101-4108.
Siddons, R.C. and M.E. Coates. 1972. The influence
of the intestinal microflora on disaccharidase ac-
tivities in the chick. Br. J. Nutr. 27:101-112.
Sieburth, J.M., J. Gutierrrez, J. McGinnis, J.R. Stern,
and B.H. Schneider. 1951. Effects of antibiotics on
intestinal microflora and on growth of turkeys and
pigs. Proc. Soc. Exp. Biol. Med. 76:15-18.
Slyter, L. L. (1976). Influence of acidosis on rumen
function. J. Anim. Sci. 43:910-929.
Smith, R.A. 1998. Impact of disease on feedlot per-
formace: a review. J. Anim. Sci. 76:272-274.
Smith, R.A., Gill, D.R., and M.T. Van Koevering. 1993.
Effects of tilmicosin of ceftiofur on health and per-

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 85
formance of stressed stocker catlle. Animal Sci-
ence Research Report, Oklahoma Agriculture Ex-
periment Station. pp. 308-311.
Snowder, G.D., L.D. Van Vleck, L.V. Cundiff, and G.L.
Bennett. 2006. Bovine respiratory disease in feed-
lot cattle: Environmental , genetic, and economic
factors. J. Anim. Sci. 84:1999-2008.
Subasinghe, R.P., P. Bueno, M.J. Phillips, C. Hough,
S.E. McGladdery, and J.R. Arther (Eds.) 2001.
Aquaculture development, health, and welfare. In
Aquaculture in the Third Millenium Technical Pro-
ceedings of the Conference on Aquaculture in the
Third Millenium (Subasinghe, R.P. et al., eds), pp.
167-191. Bangkok and FAO, NACA.
Thompson, F.L., T. Iida, and J. Swings. 2004. Bio-
diveristy of Vibrios. Microbiol. Mol. Biol. Rev. 68,
403-431.
Visek, W.J. 1978. The mode of growth promotion by
antibiotics. J. Anim. Sci. 46:1147-1469.
Yestal, C.M., W.M. Beeson, F.N. Andrews, L.M.
Hutchings, and L.P. Doyle. 1952. Effect of aureo-
mycin on the development and livability of new-
born pigs. Purdue Mimeo. AH 87
Zimmerman, D.R. 1986. Role of subtherapeutic lev-
els of antimicrobials in pig production. J. Anim.
Sci. 62:6-16.

86 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
www.afabjournal.comCopyright © 2014
Agriculture, Food and Analytical Bacteriology
ABSTRACT
Mercury, a potentially toxic metal, is present in the environment as a result of both natural processes and
from man-made sources. The amount of mercury mobilized and released into the biosphere has increased
significantly since the beginning of the industrial age. Inorganic mercury deposits in water and bottom sed-
iments where it is subject to bacterial conversion to methylmercury, which bioaccumulates in the aquatic
food chain with sometimes tragic consequences. This review discusses the production of methylmercury
in the environment and exposure to and health effects for humans. We also discuss current knowledge of
other nutrient interactions with methylmercury in the diet as well as possible methods for bioremediation
of methylmercury in the environment.
Keywords: Methylmercury, Minamata disease, mercury poisoning, biomagnification, bioaccumula-
tion, bioavailability, bioremediation
INTRODUCTION
The element mercury is a non-essential trace el-
ement that is toxic to humans and animals. At the
fifty-third meeting of the Joint FAO/WHO Expert
Committee on Food Additives (JECFA, 2000) an up-
date on the toxicity risks from methylmercury was
summarized and a provisional tolerable weekly in-
take of methylmercury for the general population
Correspondence: Philip G. Crandall, [email protected]: +1 -479-575-7686 Fax: +1-479-575-6936
(3.3 µg/kg body weight) was reaffirmed with the ad-
monition that pregnant women and nursing mothers
may be in a higher risk category. The US Environ-
mental Protection Agency has also calculated a ref-
erence dose (RfD) level for methylmercury, which is
EPA’s estimate of the maximum acceptable daily ex-
posure to humans that is not likely to cause harmful
effects during a lifetime. The RfD for methylmercury
was last revised by EPA 2001 and is currently 0.1 µg/
kg of body weight per day (Environmental Protec-
tion Agency, 2014), which is appreciably higher than
the JECFA recommendations. In the environment,
REVIEWEffects of Co-nutrients in Foods and Bioremediation
in the Environment on Methylmercury
P. G. Crandall, and C. A. O’Bryan
1 Department of Food Science, University of Arkansas, 2650 Young Ave., Fayetteville, AR 72704
Agric. Food Anal. Bacteriol. 4: 86-95, 2014

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 87
particularly lakes, waterways and wetlands, mercury
can be converted from its elemental state to a high-
ly toxic, organic compound called methylmercury
through biogeochemical interactions. Once ingest-
ed methylmercury can easily cross the blood-brain
and placental barriers and high levels of exposure
may cause severe and irreversible damage, partic-
ularly to the fetal central nervous system (Clarkson
and Magos, 2006). Methylmercury concentrations
in water, soil, and sediments are usually very low es-
pecially when compared to the less toxic inorganic
form (Zhang et al., 2010a; 2010b). However, methyl-
mercury can accumulate (bioaccumulation) and be
magnified (biomagnification) in aquatic food webs
and even some terrestrial plants, for instance rice
(Zhang et. al., 2010b), eventually posing a serious
threat to humans through the consumption of fish
and/or rice (Zhang et al., 2010a). See Figures 1 and 2
for additional information.
EXPOSURE TO METHYLMERCURY
The main source of methylmercury contamina-
tion to humans is fish, a highly nutritious food with
known benefits for human health. The Food and
Drug Administration just completed and published
a 10 year study on the levels of methylmercury con-
tamination in the domestic fish supply (Tables 1 and
2; FDA, 2013). Fish are also a vital cultural and eco-
nomic commodity for many communities around the
world. All fish, however, do not have similar amounts
of mercury because of bioaccumulation of methyl-
mercury through the many levels of the aquatic food
chain. Concentrations of total mercury vary widely
across fish and shellfish species, with the mean val-
ues differing by as much as 100-fold (Keating et al.,
1997). Methylmercury is bound to proteins, as well
as to free amino acids, that are components of mus-
cle tissues, and is not removed by any cooking or
Figure 1. Mercury enters the food chain via manmade and natural emissions and is transformed into methylmercury in the lakes and oceans where it accumulates in fish (Environmental Protec-tion Agency, 2014).
88 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
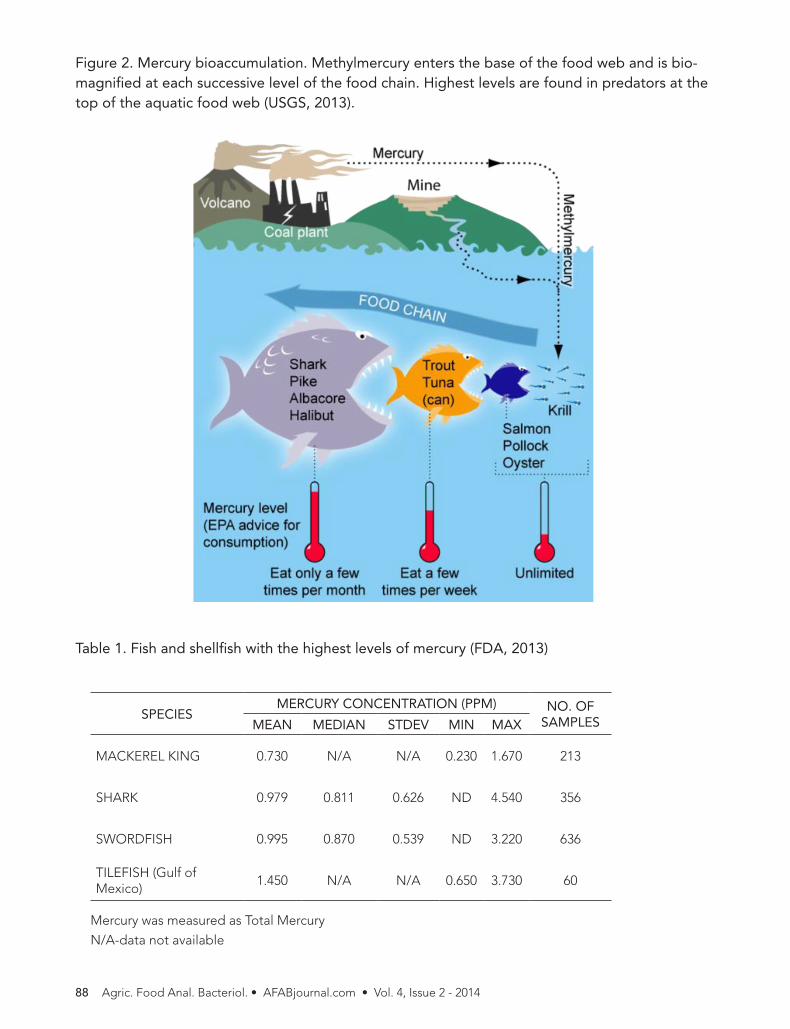
Figure 2. Mercury bioaccumulation. Methylmercury enters the base of the food web and is bio-magnified at each successive level of the food chain. Highest levels are found in predators at the top of the aquatic food web (USGS, 2013).
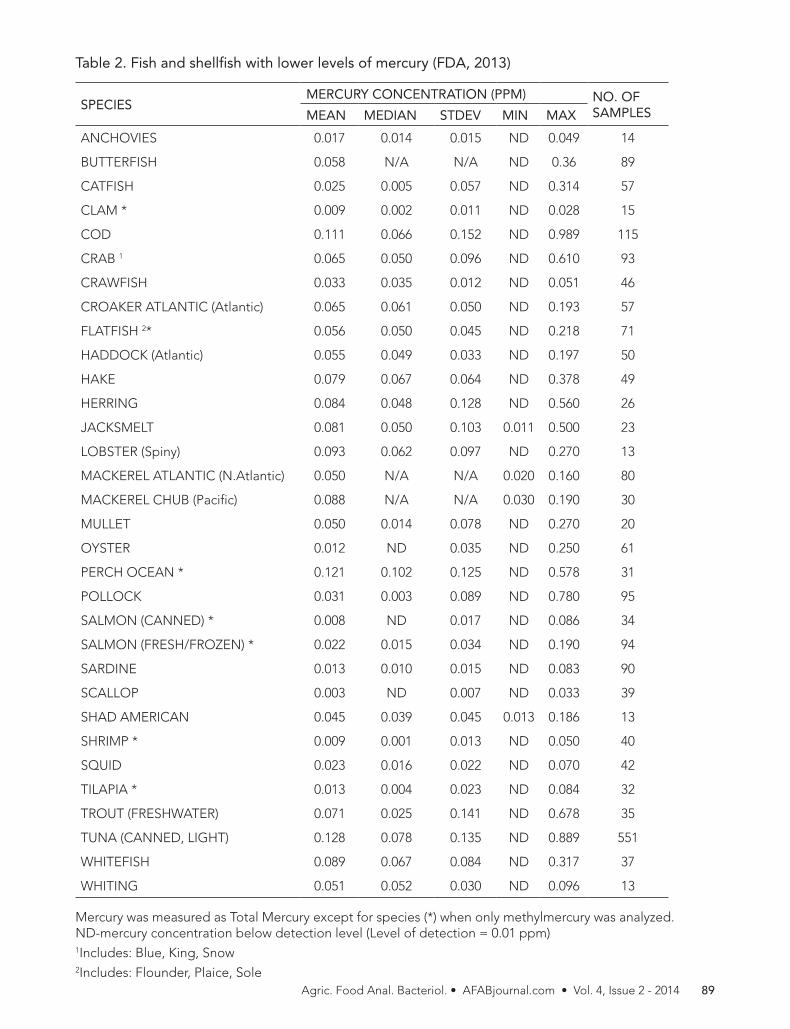
Table 1. Fish and shellfish with the highest levels of mercury (FDA, 2013)
SPECIESMERCURY CONCENTRATION (PPM) NO. OF
SAMPLESMEAN MEDIAN STDEV MIN MAX
MACKEREL KING 0.730 N/A N/A 0.230 1.670 213
SHARK 0.979 0.811 0.626 ND 4.540 356
SWORDFISH 0.995 0.870 0.539 ND 3.220 636
TILEFISH (Gulf of Mexico)
1.450 N/A N/A 0.650 3.730 60
Mercury was measured as Total Mercury
N/A-data not available
Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 89
Table 2. Fish and shellfish with lower levels of mercury (FDA, 2013)
SPECIESMERCURY CONCENTRATION (PPM) NO. OF
SAMPLESMEAN MEDIAN STDEV MIN MAX
ANCHOVIES 0.017 0.014 0.015 ND 0.049 14
BUTTERFISH 0.058 N/A N/A ND 0.36 89
CATFISH 0.025 0.005 0.057 ND 0.314 57
CLAM * 0.009 0.002 0.011 ND 0.028 15
COD 0.111 0.066 0.152 ND 0.989 115
CRAB 1 0.065 0.050 0.096 ND 0.610 93
CRAWFISH 0.033 0.035 0.012 ND 0.051 46
CROAKER ATLANTIC (Atlantic) 0.065 0.061 0.050 ND 0.193 57
FLATFISH 2* 0.056 0.050 0.045 ND 0.218 71
HADDOCK (Atlantic) 0.055 0.049 0.033 ND 0.197 50
HAKE 0.079 0.067 0.064 ND 0.378 49
HERRING 0.084 0.048 0.128 ND 0.560 26
JACKSMELT 0.081 0.050 0.103 0.011 0.500 23
LOBSTER (Spiny) 0.093 0.062 0.097 ND 0.270 13
MACKEREL ATLANTIC (N.Atlantic) 0.050 N/A N/A 0.020 0.160 80
MACKEREL CHUB (Pacific) 0.088 N/A N/A 0.030 0.190 30
MULLET 0.050 0.014 0.078 ND 0.270 20
OYSTER 0.012 ND 0.035 ND 0.250 61
PERCH OCEAN * 0.121 0.102 0.125 ND 0.578 31
POLLOCK 0.031 0.003 0.089 ND 0.780 95
SALMON (CANNED) * 0.008 ND 0.017 ND 0.086 34
SALMON (FRESH/FROZEN) * 0.022 0.015 0.034 ND 0.190 94
SARDINE 0.013 0.010 0.015 ND 0.083 90
SCALLOP 0.003 ND 0.007 ND 0.033 39
SHAD AMERICAN 0.045 0.039 0.045 0.013 0.186 13
SHRIMP * 0.009 0.001 0.013 ND 0.050 40
SQUID 0.023 0.016 0.022 ND 0.070 42
TILAPIA * 0.013 0.004 0.023 ND 0.084 32
TROUT (FRESHWATER) 0.071 0.025 0.141 ND 0.678 35
TUNA (CANNED, LIGHT) 0.128 0.078 0.135 ND 0.889 551
WHITEFISH 0.089 0.067 0.084 ND 0.317 37
WHITING 0.051 0.052 0.030 ND 0.096 13
Mercury was measured as Total Mercury except for species (*) when only methylmercury was analyzed. ND-mercury concentration below detection level (Level of detection = 0.01 ppm)1Includes: Blue, King, Snow2Includes: Flounder, Plaice, Sole

90 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
cleaning processes that do not destroy muscle tis-
sues. In addition to fish, rice cultivated in areas con-
taminated with mercury can contain relatively high
levels of methylmercury (Horvat et al., 2003; Zhang
et al., 2010b). Other food sources of methylmercury
have been reported including organ meats of ter-
restrial animals (Ysart et al., 2000), and chicken and
pork (Lindberg et al., 2004), probably as a result of
the use of fish meal as livestock feed. Persons living
in certain communities also have higher methylmer-
cury exposure because they consume the flesh of
fish-eating marine mammals (Grandjean et al., 1995;
Van Oostdam et al., 2005).
MERCURY IN THE ENVIRONMENT
Mercury is found in the environment in three forms,
elemental mercury, inorganic compounds and or-
ganic compounds; each form has specific solubility,
chemical reaction, and toxicity characteristics (Clark-
son, 2002; Goldman and Shannon, 2001). Elemental
mercury is released via degassing from the crust and
oceans of the earth, and the combustion of fossil fuels
releases elemental mercury to the environment (ATS-
DR, 1999). Additional mercury is released from indus-
trial waste; the total amount of mercury released each
year from all sources may add up to as much as 9000
tons each year (ATSDR, 1999; Trasande et al., 2005).
Mercury is deposited in surface waters from both
industrial and naturally-occurring atmospheric sourc-
es where it can attach to particles suspended in the
water. These particles eventually settle into the sedi-
ment where the mercury can be “methylated” dur-
ing a complex chemical process facilitated by anaer-
obic organisms, thus forming methylmercury. Many
factors dictate the occurrence rate of the methyla-
tion process. For example, studies have shown that
water with a lower pH and higher dissolved organic
carbon content generally results in higher levels of
methylation (United States Geological Survey, 2009).
Methylmercury is biomagnified in the aquatic food
chain from bacteria, to plankton, through macro-
invertebrates, to herbivorous fish, to fish-eating fish
(Wiener et al., 2003). Humans and other fish eating
mammals, such as otters and whales, which consume
fish from the top of the aquatic food chain receive
the methylmercury that has bioaccumulated through
this process (Mergler et al., 2007).
HEALTH EFFECTS OF METHYLMERCURY IN HUMANS
When methylmercury is ingested it is readily and
completely absorbed by the gastrointestinal tract.
Methylmercury is complexed with the amino acid
cysteine and with proteins and peptides contain-
ing cysteine; this complex is then recognized by
the amino acid transporting system of the body as
methionine, another essential amino acid (Kerper
et al., 1992). Because this complex is recognized by
the body as an essential amino acid, it is transport-
ed freely throughout the body including across the
blood–brain barrier and across the placenta, where
it is absorbed by the developing fetus. Since the
methylmercury is so strongly bound to proteins and
because the complex is recognized as an amino acid
it is not readily removed from food or from the body
(Carrier et al., 2001).
There are several studies that suggest that meth-
ylmercury causes developmental delays in children
exposed before birth, including attention defi-
cits, loss of IQ points and decreased performance
in tests of language skills and memory (Rice et al.,
2003). There is insufficient data to make a causal
link between pre-natal exposure to methylmercury
from the mother’s diet and autism in spite of the
expressed concerns of the public (van Wijngaarden
et al., 2013). In adults, ingestion of methylmercury
has been linked to increased risk of cardiovascular
disease including heart attack (Salonen et al., 1995;
Guallar et al., 2002; Choi et al., 2009), and there is
some evidence that methylmercury can cause auto-
immune diseases in sensitive individuals (Hultman
and Hansson-Georgiadis, 1999).
In addition to chronic exposure to methylmercury
there have been several episodes of acute expo-
sure in which large numbers of people were severely
poisoned by food contaminated with high levels of

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 91
methylmercury. The most widely known incident is
probably the dumping of industrial waste that re-
sulted in the pollution of water and fish and subse-
quent mass poisonings in Minamata and Niigata,
Japan (Harada, 1995). Another such episode took
place in Iraq in 1971; wheat treated with methylmer-
cury was shipped to Iraq as seed grain not intended
for human consumption. Due to a number of factors,
including foreign-language labeling and late distri-
bution within the growing cycle, this toxic grain was
consumed as food by Iraqi residents in rural areas.
The recorded death toll was 650 people, but figures
at least ten times greater have been suggested,
making this the largest mercury poisoning disaster
(Bakir et al., 1973). These episodes resulted in neu-
rological symptoms including loss of feeling, loss of
physical coordination, difficulty in speech, narrowing
of the visual field, hearing impairment, blindness,
and death. Children who had been exposed in-utero
through their mothers’ ingestion were also affected
with a range of symptoms including motor difficul-
ties, sensory problems and mental retardation.
REDUCING BIOACESSABILITY
The protective effect of selenium against methyl-
mercury toxicity has been hypothesized for a num-
ber of years (Pařízek and Oštádalová, 1967; Skerfv-
ing, 1978; Cuvinaralar and Furness, 1991; Raymond
and Ralston, 2004; Falnoga and Tusek-Znidaric, 2007;
Yang et al., 2008; Khan and Wang, 2009). The protec-
tive effects of selenium against methylmercury toxic-
ity in fetal brain development have now been con-
firmed but only in animal studies (Beyrouty and Chan,
2006; Sakamoto et al., 2013). Yang, et al. (2008) and
Khan and Wang (2009) have summarized the several
physiologic/biochemical mechanisms proposed to
explain the antagonism between methylmercury and
selenium. It seems likely that the molecular mecha-
nism involves the formation of insoluble, equimolar,
and biologically unavailable mercury selenide pre-
cipitates, since approximately 1:1 molar ratios of se-
lenium and mercury have been observed in marine
mammals, sea birds and humans (Chen et al., 2006;
Khan and Wang, 2009; Li et al., 2012) . Shim et al.
(2009) found that phytochemical rich foods, spe-
cifically green tea extract, black tea extract, and soy
protein significantly reduced mercury bioaccessibil-
ity by 82 to 92%, 88 to 91%, and 44 to 87%, respec-
tively. Wheat bran decreased bioaccessibility by 84%,
oat bran by 59 to 75% and psyllium by 15 to 31% at
amounts greater than 500 mg (Shim et al., 2009). Evi-
dence also exists that suggests the developmental
and cardiovascular toxicity of methylmercury may be
mediated by co-exposures to omega-3 fatty acids,
in particular docosahexaenoic acid (DHA) (Jin et al.,
2007). Nøstbakken et al. (2012) found that omega-3
lessened methylmercury toxicity, either by decreas-
ing programmed cell death (eicosapentaenoic acid)
or by reducing methylmercury uptake (DHA).
BIOREMEDIATION
Bioremediation is a waste management tech-
nique that involves the use of organisms to remove
or neutralize pollutants from a contaminated site.
The use of microbial biomass for bioremediation
of toxic metals has been pursued for a number of
years (Akthar et al., 1995, 1996; Akthar and Mohan,
1995; Gupta et al., 2000; Karna et al., 1999; Pethkar et
al., 2001; Puranik and Paknikar, 1997; Volesky, 1987).
Both live and non-living microbial biomass has been
studied for removal of toxic metal ions but many re-
searchers believe that non-living or processed bio-
mass is a better choice. Non-living biomass does not
have toxicity limitations as would living cells, nutri-
ents are not needed for growth of biomass and since
non-living biomass acts as an ion exchanger the pro-
cess is rapid (Paknikar et al., 2003).
The cell wall polymers of fungi are known to have
functional groups such as amino, amide, hydroxyl,
carboxyl, sulfhydryl and phosphate which have been
implicated in metal binding (Akhtar et al., 1995; Gup-
ta et al., 2000). Karunasagar et al. (2003) studied the
effectiveness of a biosorbent prepared from biomass
of Aspergillus niger for removal of methylmercury
from dilute solutions. They determined that removal
of methylmercury from spiked ground water samples

92 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
was efficient and not influenced by other ions, and
that the biosorbent was reusable for up to six cycles
without appreciable loss of binding capacity.
Live bacteria provide another means of methylmer-
cury bioremediation (Barkay, et al., 2003; Barkay and
Wagner-Dobler, 2005; Chen and Wilson, 1997; Miller,
1999; Nascimento and Chartone-Souza, 2003). Bac-
teria can break down mercury compounds through
the acquisition of a transferable genetic element
known as the mer operon (Omichinski2007). The
mer operon is a dedicated set of mercury-resistant
genes that are self-regulated by the DNA-binding
protein MerR; bacteria resistant to methylmercury
code for proteins that regulate mercury transport
(MerA, MerP, MerT) and mercury degradation (MerA
and MerB) (Osborn et al., 1996; Sahlman et al., 1997;
Silver and Phung, 1996; Wilson et al., 2000). Chien
et al. (2010) made the point that there are substrate
specificities among the MerB enzymes, elucidating
the necessity for selecting the appropriate bacterial
strains or MerB enzymes to apply them in bioreme-
diation engineering for cleaning up specific mercury
contaminants.
Meagher (2000) engineered MerA and MerB into
plants to remediate methylmercury contamination.
Their theory was that remediation using plants is
potentially more robust than bacterial remediation,
because plants use solar energy, have roots that
penetrate contaminated sediments, and accumu-
late a large aboveground biomass. There are actu-
ally a few well-characterized plant species used to
clean up contaminated wetland ecosystems (Mea-
gher, 2000). Plants such as cottonwood trees (Lyyra
et al., 2007) and tobacco (Heaton et al., 2005) have
been modified to express either MerB or both MerB
and MerA; the plants converted the methylmercury
to ionic mercury or elemental mercury, respectively;
however, the elemental mercury was released into
the atmosphere, where it may still pose a risk.
CONCLUSIONS
Bioremediation is considered to have advantages
over conventional techniques such as chemical pre-
cipitation, conventional coagulation, adsorption by
activated carbons, adsorption by natural materials,
ion exchange, or reverse osmosis. Mercury-resistant
bacteria possess the mer operon enabling them to
convert the toxic forms of mercury to nontoxic forms.
Those possessing the merB gene are more valuable
as they can detoxify methylmercury along with other
organic mercurial compounds and inorganic mercury
to nontoxic, volatile mercury. Bacteria harboring the
merB gene and genetically modified organisms pos-
sessing the mer operon including merB are promis-
ing tools for use in bioremediation of methlymercury.
However, the cons for the bacterial-based or plant-
based processes may include production of large
volumes of mercury-loaded biomass, the disposal of
which is problematic.
REFERENCES
Akthar, N. and P. M. Mohan 1995. Bioremediation of
toxic metal ions from polluted lake waters and in-
dustrial effluents by fungal biosorbent. Current Sci.
69: 1028–1030.
Akthar, N., K. S. Sastry, and P. M. Mohan 1995. Bio-
sorption of silver ions by processed Aspergillus ni-
ger biomass. Biotechnol. Letts. 17: 551–556.
Akthar, N., K. S. Sastry, and P. M. Mohan 1996. Mech-
anism of metal biosorption by fungal biomass. Bio-
metals 9: 21–28.
ATSDR, Agency for Toxic Substances and Disease
Registry. 1999. Toxicological profile for mercury:
TP-93/10. Atlanta, Georgia: Centers for Disease
Control.
Bakir, F., S. F. Damluji, L. Amin-Zaki, M. Murtadha, A.
Khalidi, N. Y. al-Rawi, S. Tikriti, H. I. Dahahir, T. W.
Clarkson, J. C. Smith, and R. A. Doherty 1973. Meth-
ylmercury poisoning in Iraq. Science. 181:230-241.
Barkay, T., S. M. Miller, A. O. Summers 2003. Bacte-
rial mercury resistance from atoms to ecosystems.
FEMS Microbiol. Rev. 27: 355-384.
Barkay, T., and I. Wagner-Dobler 2005. Microbial
transformations of mercury: potentials, challenges,
and achievements in controlling mercury toxicity in
the environment. Adv. Appl. Microbiol. 57: 1–52.

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 93
Beyrouty, P. and H. M. Chan 2006.Co-consumption
of selenium and vitamin E altered the reproduc-
tive and developmental toxicity of methylmercury
in rats. Neurotoxicol. Teratol. 28: 49– 58.
Carrier, G., M. Bouchard, R. C. Brunet, M. Caza 2001.
A toxicokinetic model for predicting the tissue dis-
tribution and elimination of organic and inorganic
mercury following exposure to methyl mercury in
animals and humans. II. application and validation
of the model in humans. Toxicol. Appl. Pharmacol.
171: 50–60.
Chen, S., and D. B. Wilson 1997. Construction and
characterization of Escherichia coli genetically
engineered for bioremediation of Hg2+-contami-
nated environments. Appl. Environ. Microbiol. 63:
2442–2445.
Chen, C. Y., H. W. Yu, J. J. Zhao, B. Li, L. Y. Qu, S. P.
Liu, P. Q. Zhang, and Z. F. Chai, 2006. The roles
of serum selenium and selenoproteins on mercury
toxicity in environmental and occupational expo-
sure Environ. Health Perspect. 114: 297– 301.
Chien, M. F., M. Narita, K. H. Lin, K. Matsui, C. C.
Huang, and G. Endo 2010. Organomercurials re-
moval by heterogeneous merB genes harboring
bacterial strains. J. Biosci. Bioeng. 110: 94-98.
Choi, A.L., P. Weihe, E. Budtz-Jørgensen, P. J. Jør-
gensen, J. T. Salonen, T. P. Tuomainen, K. Mura-
ta, H. P. Nielsen, M. S. Petersen, J. Askham, and
P. Grandjean 2009. Methylmercury exposure and
adverse cardiovascular effects in Faroese whaling
men. Environ. Health Perspect. 117: 367-372.
Clarkson, T. W. 2002. The three modern faces of mer-
cury. Environ. Health Perspect. 10:11–24.
Clarkson, T. W. and L. Magos 2006. The toxicology
of mercury and its chemical compounds Crit. Rev.
Toxicol. 36: 609– 662.
Cuvinaralar, M. L. A. and R. W. Furness 1991. Mercury
and selenium interaction—A review Ecotox. Envi-
ron. Safe. 21: 348– 364.
Environmental Protection Agency. 2014. Exposure to
methylmercury. Available at: http://www.epa.gov/
hg/exposure.htm
Falnoga, I. and M. Tusek-Znidaric 2007. Selenium–
mercury interactions in man and animals Biol.
Trace Elem. Res. 119: 212– 220.
FDA. 2013. Mercury levels in commercial fish and
shellfish (1990-2010). Available at: http://www.fda.
gov/food/foodborneillnesscontaminants/metals/
ucm115644.htm
Goldman, L. R., and M. W. Shannon 2001. Technical
report: mercury in the environment: implication for
pediatricians. American Academy of Pediatrics,
Committee on Environmental Health. Pediatrics
108:197–205.
Grandjean, P., P. Weihe, L.L. Needham, V.W. Burse,
D.G. Patterson, Jr, E.J. Sampson, P.J. Jorgensen,
and M. Vahter 1995. Relation of a seafood diet to
mercury, selenium, arsenic, and polychlorinated bi-
phenyl and other organochlorine concentrations in
human milk. Environ. Res. 71: 29–38.
Guallar, E., I. Sanz-Gallardo, P. van’t Veer, P. Bode, A.
Aro, J. Gómez-Aracena, J. D. Kark, R. A. Riemers-
ma, J. M. Martín-Moreno, and F. J. Kok 2002/ Mer-
cury, fish oils, and the risk of myocardial infarction.
New England J. Med. 347: 1747-1754.
Gupta, R., P. Ahuja, S. Khan, R. K. Saxena, and H.
Mohapatra 2000. Microbial biosorbents: meeting
challenges of heavy metal pollution in aqueous so-
lutions. Current Sci. 78: 967–973.
Harada, M. 1995. Minamata disease: methylmercury
poisoning in Japan caused by environmental pol-
lution. Crit. Rev. Toxicol. 25: 1-24.
Heaton, A. C. P., C. L. Rugh, N.-J. Wang, R. B. Mea-
gher 2005. Physiological responses of transgenic
merA-TOBACCO (Nicotiana tabacum) to foliar
and root mercury exposure. Water Air, Soil Pollut.
161:137-155.
Horvat, M., N. Nolde, V. Fajon, V. Jereb, M. Logar, S.
Lojen, R. Jacimovic, I. Falnoga, Q. Liya, J. Faganeli,
and D. Drobne 2003. Total mercury methylmercury
and selenium in mercury polluted areas in the prov-
ince Guizhou, China. Sci. Total Environ. 304: 231–256.
Hultman, P., and H. Hansson-Georgiadis 1999. Meth-
yl mercury-induced autoimmunity in mice. Toxicol.
Appl. Pharmacol. 154: 203–211.
Jin, X. E. Lok, G. Bondy, D. Caldwell, R. Mueller, K.
Kapal, C. Armstrong, M. Taylor, S. Kubow, R. Meh-
ta, and H. M. Chan 2007. Modulating effects of di-
etary fats on methylmercury toxicity and distribu-
tion in rats. Toxicol. 230: 22–44.

94 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
Karna, R.R., L. Uma, G. Subramanian, and P. M. Mo-
han 1999. Biosorption of toxic metal ions by alkali-
extracted biomass of a marine cyanobacterium,
Phormidium valderianum BDU 30501. World J. Mi-
crobiol. Biotechnol. 15: 729–732.
Karunasagar, D., J. Arunachalam, K. Rashmi, J. N. L.
Latha, and P. M. Mohan 2003. Biosorption of in-
organic and methyl mercury by a biosorbent from
Aspergillus niger. World J. Microbiol. Biotech-
nol.19: 291-295.
Keating, M.H., K.R. Mahaffey, R. Schoeny, G.E. Rice,
O.R. Bullock, R.B. Ambrose, J. Swartout and J.W.
Nichols 1997. Mercury study report to Congress,
Vol. III: fate and transport of mercury in the en-
vironment. Office of Air Quality Planning and
Standards and Office of Research and Develop-
ment, U.S. Environmental Protection Agency, EPA-
452/R-97–005. EPA, Washington, D.C.
Kerper, L.E., N. Ballatori, and T. W. Clarkson 1992.
Methylmercury transport across the blood–brain
barrier by an amino acid carrier. Am. J. Physiol. 262
(5 Pt. 2): R761–R765.
Khan, M. A. K. and F. Y. Wang 2009. Mercury–sele-
nium compounds and their toxicological signifi-
cance: Toward a molecular understanding of the
mercury–selenium antagonism Environ. Toxicol.
Chem. 28: 1567– 1577.
Li, Y.-F., Z. Dong, C. Chen, B. Li, Y. Gao, L. Qu, T.
Wang, X. Fu, Y. Zhao, and Z. Chai, 2012. Organic
selenium supplementation increases mercury ex-
cretion and decreases oxidative damage in long-
term mercury-exposed residents from Wanshan,
China Environ. Sci. Technol. 46: 11313– 11318.
Lindberg, A., K.A. Bjornberg, M. Vahter, and M. Ber-
glund 2004. Exposure to methylmercury in non-
fish-eating people in Sweden. Environ. Res. 96:
28–33.
Lyyra, S. R. B. Meagher, T. Kim, A. Heaton, P. Mon-
tello, R. S. Balish, and S. A. Merkle 2007. Coupling
two mercury resistance genes in Eastern cotton-
wood enhances the processing of organomercury.
Plant Biotechnol. J. 5: 254-262.
Meagher, R. B. 2000. Phytoremediation of toxic el-
emental and organic pollutants. Curr. Opin. Plant
Biol. 3: 153.
Mergler, D., H. A. Anderson, L. H. M. Chan, K. R. Ma-
haffey, M. Murray, M. Sakamoto, and A. H. Stern
2007. Methylmercury exposure and health effects
in humans: A worldwide concern. AMBIO 36: 3-11.
Miller, S. M. 1999. Bacterial detoxification of Hg(II)
and organomercurials. Essays Biochem. 34: 17–30.
Nascimento, A. M., and E. Chartone-Souza 2003.
Operon mer: Bacterial resistance to mercury and
potential for bioremediation of contaminated en-
vironments. Genetics Molec. Res. 2: 92–101.
Nøstbakken, O.J., L. Bredal, P. A. Olsvik, T. S. Huang,
and B. E. Torstensen. 2012. Effect of marine Omega
3 fatty acids on methylmercury-induced toxicity in
fish and mammalian cells in vitro. J. Biomed. Bio-
technol. 2012: Article ID 417652, 13 pages, http://
dx.doi.org/10.1155/2012/417652
Osborn, A. M., K. D. Bruce, D. A. Ritchie, and P. Strike
1996. The mercury resistance operon of the IncJ
plasmid pMERPH exhibits structural and regula-
tory divergence from other Gram-negative mer
operons. Microbiol. 142: 337–345.
Paknikar, K. M., A. V. Pethkar, and P. R. Puranik 2003.
Bioremediation of metalliferous wastes and prod-
ucts using inactivated microbial biomass. Indian J.
Biotechnol. 2: 426-443.
Pařízek, J., and I. Oštádalová 1967.The protective ef-
fect of small amounts of selenite in sublimate in-
toxication Cell. Mol. Life Sci. 23: 142– 143.
Pethkar, K.V., R. P. Gaikaiwari, and K. M. Paknikar
2001. Biosorptive removal of contaminating heavy
metals from plant extracts of medicinal plants.
Current Sci. 80: 1216–1218.
Puranik P.R. and K. M. Paknikar 1997. Biosorption of
lead and zinc from solutions using Streptoverticil-
lium cinnamoneum waste biomass. J. Biotechnol.
55: 113–124.
Raymond, L. J., and N. V.C. Ralston 2004. Mercury:
selenium interactions and health implications Sey-
chelles Med. Dent. J. 7 ( Special issue): 72– 77.
Rice, D. C., R. Schoeny, and K. Mahaffey 2003. Meth-
ods and rationale for derivation of a reference dose
for methylmercury by the U.S. EPA. Risk Analysis 23:
107–115.
Sahlman, L., W. Wong, and J. Powlowski 1997. A
mercuric ion uptake role for the integral inner

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 95
membrane protein, MerC, involved in bacterial
mercuric ion resistance. J. Biol. Chem. 272: 29518–
29526.
Sakamoto, M., A. Yasutake, A. Kakita, M. Ryufuku,
H. M. Chan, M. Yamamoto, S. Oumi, S. Kobayashi,
and C. Watanabe 2013.Selenomethionine protects
against neuronal degeneration by methylmercury
in the developing rat cerebrum Environ. Sci. Tech-
nol. 47: 2862– 2868.
Salonen, J. T., K. Seppänen, K. Nyyssönen, H. Kor-
pela, J. Kauhanen, M. Kantola, J. Tuomilehto, H.
Esterbauer, F. Tatzber, and R. Salonen 1995. Intake
of mercury from fish, lipid peroxidation, and the
risk of myocardial infarction and coronary, cardio-
vascular, and any death in Eastern Finnish men”.
Circulation 91: 645–655.
Shim, S. M., M. G. Ferruzzi, Y. C. Kim, E. M. Janle,
and C. R. Santerre 2009. Impact of phytochemical-
rich foods on bioaccessibility of mercury from fish.
Food Chem. 112: 46-50.
Silver, S., and L. T. Phung 1996. Bacterial heavy metal
resistance: New surprises. Ann. Rev. Microbiol. 50:
753–789.
Skerfving, S. 1978. Interaction between selenium
and methylmercury. Environ. Health Perspect. 25:
57–65.
Trasande, L., P. J. Landrigan, and C. Schechter 2005.
Public health and economic consequences of
methyl mercury toxicity to the developing brain.
Env. Health Persp. 113: 590-596.
U.S. Geological Survey 2009. Mercury in the en-
vironment. http://www.usgs.gov/themes/fact-
sheet/146-00/index.html Accessed 18 March 2014.
USGS. 2013. The South Florida mercury science pro-
gram. Available at: http://sofia.usgs.gov/publica-
tions/posters/merc_program/
Van Oostdam, J., S.G. Donaldson, M. Feeley, D. Ar-
nold, P. Ayotte, G. Bondy, L. Chan, E. Dewaily, C.
M. Furgal, H. Kuhnlein, E. Loring, G. Muckle, E.
Myles, O. Receveur, B. Tracy, U. Gill, and S. Kalhok.
2005. Human health implications of environmental
contaminants in Arctic Canada: a review. Sci. Total.
Environ. 351–352: 165–246.
van Wijngaarden, E., P. W. Davidson, T. H. Smith, K.
Evans, K. Yost, T. Love, S. W. Thurston, G. E. Wat-
son, G. Zareba, C. M. Burns, C. F. Shamlaye, and
G. J. Myers 2013. Autism spectrum disorder phe-
notypes and prenatal exposure to methylmercury.
Epidemiol. 24: 651-659.
Volesky, B. 1987 Biosorbents for metal recovery.
Trends Biotechnol. 5: 96–101.
Wiener, J.G., D. P. Krabbenhoft, G. H. Heinz, and A.
M. Scheuhammer 2003. Ecotoxicology of mercury
in Hoffman, D.J., B.A. Rattner, G.A. Burton, Jr., and
J. Cairns, Jr., eds., Handbook of Ecotoxicology,
2nd edition.: Boca Raton, Florida, CRC Press, p.
409-463.
Wilson, J. R., C. Leang A. P. Morby, J. L. Hobman,
and N. L. Brown 2000. MerF is a mercury transport
protein: Different structures but a common mech-
anism for mercuric ion transporters? FEBS Letts.
472: 78–82.
Yang, D. Y., Y. W. Chen, J. M. Gunn, and N. Belzile
2008. Selenium, mercury in organisms: Interactions
and mechanisms Environ. Rev. 16: 71– 92.
Ysart, G., P. Miller, M. Croasdale, H. Crews, P. Robb,
M. Baxter, C. de L’Argy, and N. Harrison 2000. 1997
UK Total Diet Study—dietary exposures to alumin-
ium, arsenic, cadmium, chromium, copper, lead,
mercury, nickel, selenium, tin, and zinc. Food Ad-
dit. Contam. 17: 775–786.
Zhang, H., X. Feng, T. Larssen, G. Qiu, and R. D. Vogt
2010a. In inland China, rice, rather than fish, is the
major pathway for methylmercury exposure Envi-
ron. Health Perspect. 118: 1183– 1188.
Zhang, H. X. Feng, T. Larssen, L. Shang, and P. Li
2010b. Bioaccumulation of methylmercury versus
inorganic mercury in rice (Oryza sativa L.) grain. En-
viron. Sci. Technol. 44: 4499– 4504.

96 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
www.afabjournal.comCopyright © 2014
Agriculture, Food and Analytical Bacteriology
ABSTRACT
Land application is a common method for disposal of manure and litter that accumulate during poul-
try production; however, zoonotic pathogens residing in the manure may contaminate either directly or
indirectly ready-to-eat produce crops. Aerobic composting of animal manure is a beneficial process treat-
ment that inactivates these pathogens. Although heat is considered to be the primary contributing factor
to inactivation, ammonia and volatile acids may also serve antimicrobial roles during composting. This
study was designed to determine the relative contributions of chemicals and heat to the inactivation of
Salmonella and Listeria monocytogenes in chicken manure-based compost mixtures formulated to give
initial carbon:nitrogen (C:N) ratios of 20:1, 30:1 and 40:1. The different initial C:N ratio formulations of the
compost mixtures had no effect on pH or the cumulative heat generated. In general, there was within all
compost mixtures an initial decline in pH followed by an increase in pH that coincided with an increase
in temperature. Levels of ammonia and volatile acids were higher in compost mixtures formulated to an
initial C:N ratio of 20:1 than in other C:N formulations. The inactivation rates of Salmonella and L. monocy-
togenes within 20:1 C:N formulations were higher than in other formulations. Regression models derived
from the data revealed that volatile acid levels, in addition to heat, played a major role in pathogen inac-
tivation. Therefore, it may be advantageous to formulate compost mixtures containing chicken litter to an
initial C:N of 20:1 to take advantage of the antimicrobial activity of volatile acids generated when sub-lethal
temperatures occur.
Keywords: manure, litter, composting, chicken, heat, ammonia, volatile acids, pH, Listeria monocy-togenes, Salmonella
Correspondence: M.C. Erickson, [email protected]: +1 -770-412-4742 Fax: +1-770-229-3216
Contribution of Chemical and Physical Factors to Zoonotic Pathogen Inactivation during Chicken Manure Composting
M.C. Erickson1*, J. Liao1, X. Jiang2, and M.P. Doyle1
1Center for Food Safety and Department of Food Science and Technology, University of Georgia, Griffin, GA
2Department of Food, Nutrition, and Packaging Sciences, Clemson University, Clemson, SC
Agric. Food Anal. Bacteriol. 4: 96-108, 2014

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 97
INTRODUCTION
Poultry production in the United States is a
major enterprise having had a combined value
in broiler, egg, and turkey production of $38 bil-
lion in 2012 (USDA, 2012). Although meat and
eggs are the major outputs from this enterprise,
a substantial amount of manure (generated from
layer and turkey operations) and litter (mixture of
manure, bedding material, wasted feed, feathers,
and soil generated from broiler operations) is also
produced. For example, the estimated tons of
manure produced from poultry operations in the
U.S. in 2007 was 81 million tons (US EPA, 2013).
To dispose of this waste, land application has of-
fered the best solution (Moore et al., 1998; Ritz and
Merka, 2013). Poultry manure can harbor zoonotic
pathogens such as Salmonella, Listeria monocy-
togenes, and Campylobacter (Chinivasagam et
al., 2010; Hutchison et al., 2004, 2005), and if ap-
plied to fields growing ready-to-eat produce, these
pathogens may contaminate those crops. Once
excreted from the animal, pathogen survival is de-
pendent on storage conditions (Goss et al., 2013;
Leifert et al., 2008; Ziemer et al., 2010). If left un-
disturbed, Williams and Benson (1978) determined
that Salmonella Typhimurium survived for at least
18 months in chicken litter at 11 or 25°C, and 13
days at 38°C. Decimal reduction times for S. Ty-
phimurium in poultry manure are not only affected
by storage temperature, but also by the type of
matrix, being greater in manure slurries compared
to manure piles (Himathongkham et al., 2000). Ac-
cording to USDA, only 5% of all U.S. cropland in
2006 was fertilized with manure, with most chicken
manure being applied to peanut and cotton fields
(MacDonald et al., 2009). Although this mode of
disposal would appear to have a minimal food
safety risk, natural waterways and irrigation ponds
in the Southeastern U.S. have been found contami-
nated with Salmonella and Campylobacter, espe-
cially after precipitation events (Gu et al., 2013b;
Haley et al., 2009; Luo et al., 2013) and likely oc-
curred from pathogen runoff of peanut and cotton
fields amended with poultry manure. These water
sources are frequently used to irrigate fields grow-
ing ready-to-eat produce (Gu et al., 2013a), hence
animal manure should be treated to inactivate
pathogens prior to land application.
A treatment that is often recommended to inac-
tivate vegetative bacterial pathogens in manures is
thermophilic aerobic composting. In this process,
manure is mixed with one or more carbon amend-
ments to produce a nutrient-rich environment
favorable for the metabolism of thermophilic mi-
croorganisms. The major factor responsible for in-
activating pathogens in such systems is heat gener-
ated by the metabolic activity of these thermophilic
microorganisms (Erickson et al., 2010; Wichuk and
McCartney, 2007)). As a result, process conditions
that are based on time and temperature have been
promulgated in regulations or guidelines world-
wide (Hogg et al., 2002). For example, guidelines
within the U.S. include either a minimum temper-
ature of 55°C for 3 days in aerated static piles or
in-vessel systems or 55°C for 15 days in windrow
systems (narrow trapezoidal elongated rows) dur-
ing which time the piles must be turned a minimum
of 5 times to ensure that all material is subjected to
the necessary thermal conditions (US EPA, 1999).
Although heat is the primary mechanism for in-
activating pathogens during aerobic composting,
temperature stratification within static piles can re-
sult in extended survival of pathogens at the sur-
face as well as extended survival of pathogens at
interior sites of piles composted during the winter
(Berry et al., 2013; Erickson et al., 2010; Shepherd
et al., 2007). In addition, exposure of the pathogen
to nonlethal heat or selected moisture conditions
could lead to metabolic alterations in the patho-
gen that makes them more resistant to the thermal
conditions encountered during the thermophilic
phase of composting (Chen et al., 2013; Shepherd
et al., 2010; Singh et al., 2011, 2012). As evidence of
this potential activity, Salmonella, Escherichia coli
O157:H7, and Listeria survived in poultry manure-
based compost piles when exposed to tempera-
tures above 55°C for more than 8 days (Hutchison
et al., 2005). Hence, other factors, either chemical
or biological, may provide a greater contribution

98 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
to pathogen inactivation under those conditions.
For example, accumulation of free ammonia in
poultry manure has been reported to contribute to
inactivation of S. Typhimurium and E. coli O157:H7
in poultry manure (Himathongkham et al., 2000).
Alternatively, volatile acids generated during the
early phase of composting in cow manure systems
formulated to have an initial carbon:nitrogen (C:N)
ratio of 20:1 were suggested to be bactericidal
agents effective against Salmonella but not Liste-
ria (Erickson et al., 2009a,b). Given that there are
differences in microbial and raw material composi-
tion between cow and chicken manure (De Bertoldi
et al., 1987; Lynch, 1987; Wang et al., 2007), it was
the objective of this study to determine the relative
contributions of heat, volatile acids, and ammonia
to the inactivation of Salmonella and L. monocyto-
genes in chicken manure-based compost mixtures
formulated to C:N ratios ranging from 20:1 to 40:1.
MATERIALS AND METHODS
Pathogen Strains and Their Preparation for Experimental Trials
Five strains of Listeria monocytogenes (101M,
12443, F6854, G3982, and H7550) from the culture
collection housed at the Center for Food Safety, Uni-
versity of Georgia were used for these studies. In
addition, three strains of Salmonella enterica serovar
Enteritidis (ME-18, H4639, and H3353) and one strain
of S. enterica serovar Newport (11590) were also
used from the culture collection. All strains had been
labeled with the green-fluorescent plasmid (GFP),
but for L. monocytogenes strains, the plasmid also
contained an erythromycin-resistant marker, whereas
Salmonella strains contained an ampicillin-resistant
marker. Previously, plasmid stability of these GFP-
labeled strains was reported to range from 8 to 52%
and 15 to 77% plasmid loss after 20 generations for
the L. monocytogenes and Salmonella strains, re-
spectively (Ma et al., 2011).
To prepare the pathogen strains for challenge
studies, frozen cultures of L. monocytogenes and Sal-
monella were thawed and individually streaked onto
plates containing brain heart infusion agar (Becton
Dickinson, Sparks, MD) with 8 µg/mL of erythromy-
cin (BHIA-E) and tryptic soy agar (Difco Laboratories,
Detroit, MI) with 100 µg/mL ampicillin (TSA-A), re-
spectively. Following incubation of plates at 37°C for
ca. 24 h, individual colonies were removed and sub-
sequently streaked onto a second plate and held at
37°C for an additional 24 h. From this second set of
plates, individual colonies of L. monocytogenes and
Salmonella were removed and inoculated into 10
ml of brain heart infusion broth (Becton Dickinson)
containing 8 µg/mL erythromycin (BHIB-E) and tryp-
tic soy broth containing 100 µg/mL ampicillin (TSB-
A), respectively. These suspensions were incubated
for ca. 24 h at 37°C with agitation (150 rpm) before
harvesting the bacteria by centrifugation (4,050 x g,
15 min, 4°C). The pelleted cells were washed three
times in 0.1% peptone water (Difco) and resediment-
ed by centrifugation before reconstituting in 0.1%
peptone water to an optical density at 630 nm of
ca. 0.5 that corresponded to a concentration of ca.
109 CFU/mL. The five strains of L. monocytogenes
were then combined in equal proportions to make
one 5-strain stock culture mixture, whereas the four
strains of Salmonella were combined for one 4-strain
stock culture mixture. Each of these stock culture
mixtures was then diluted 10-fold with deionized wa-
ter for mixtures of 108 CFU/mL that were used to spray
chicken litter. Immediately after preparation of the
spray mixtures, L. monocytogenes and Salmonella
was enumerated by plating serial dilutions (1:10) on
modified oxford medium (Acumedia Manufacturers,
Lansing, MI) containing 10 mg/mL buffered colistin
methanesulfonate, 20 mg/mL buffered moxalactam
solution, and 8 µg/mL erythromycin (MOX-E) and
TSA-A, respectively. Salmonella colonies emitted a
bright green fluorescence when plates were held un-
der a handheld UV light (365 nm) and the fluorescent
colonies were counted as Salmonella. Fluorescent L.
monocytogenes colonies were smaller than Salmo-
nella colonies and required a Leica MZ16 FA stereo
fluorescence microscope (Bannockburn, IL) for visu-
alization and counting.

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 99
Compost Ingredients, Preparation, and Experimental Design
Fresh chicken litter was collected from a broiler
production facility located in Orchard Hill, GA.
Batches were collected at different times for each
independent replicate trial. Following transport, the
litter was mixed thoroughly and a portion of the lit-
ter was removed for compositional analysis. The re-
mainder of the litter was frozen to kill insect eggs and
then held at -20°C until which time it was ready to be
composted. Wheat straw and cottonseed meal were
purchased from a local feed store and served as the
major carbon sources for the compost mixtures.
Chicken litter was added to a 28-L sanitized bowl
and sprayed manually using a spray bottle with both
the Salmonella and L. monocytogenes inocula for
populations approximating 107 CFU/g. This inocu-
lated mixture was then mixed with a Hobart mixer
(model D320:0.75 h.p.). Wheat straw, cottonseed
meal, and water were then added in such quantities
that compost mixtures had an initial moisture con-
tent of 60% and a C:N ratio of either 20:1, 30:1, or
40:1. Immediately after mixing, the compost mix-
tures were sampled for chemical and microbiologi-
cal analysis. The remainder of the compost mixture
was then placed in one of three bioreactors.
In this experimental study, three independent tri-
als were conducted wherein each trial consisted of
three bioreactor systems containing one compost
mixture each of the 20:1, 30:1 and 40:1 C:N ratio mix-
tures. The compost mixtures were composted for up
to 6 days and were sampled on days 1, 2, 3, and 6 to
measure microbiological and chemical parameters.
Composting Apparatus and Sampling
Bioreactors (46 cm high x 32 cm diameter) were
constructed from PVC plastic pipe. Tightly fitting
PVC covers had holes drilled into their center such
that the bottom cover hole allowed condensate to
drip into an attached bottle and the top cover hole
allowed compressed air (155 ml/min) to be delivered
to the system. Within the biochamber, a perforated
shelf was supported 5 cm above the bottom. Two
sampling ports (3 cm diameter) at heights of 6 to
9 cm and 10 to 13 cm above the PVC shelf and a
hole (0.5 cm diameter) at a height of 6.5 cm above
the shelf for insertion of a thermocouple wire were
drilled into the sides of the bioreactors. Bioreac-
tors were housed within a Precision 30 Mechani-
cal Convection incubator (Thermo Fisher Scientific,
Waltham, MA) that was maintained at a temperature
of 40°C. Trapped air in the incubator was vented to
a filtered exhaust system.
Compost material (ca. 5 kg) was placed into each
bioreactor after which a type T thermocouple wire
was inserted through the small hole to a site desig-
nated as the bottom center (16 cm from bioreactor
wall). An additional thermocouple was inserted to
a depth of 10 cm into the top center of the com-
post mixture. All thermocouples (two per bioreac-
tor) were connected to a multi-channel HotMux
data logger (DCC Corp., Pennsauken, NJ) that was
programmed to record temperatures at the 6 loca-
tions at 30-min intervals. Cumulative heat > 40°C
(degree-days) was calculated as the product of time
(days) and temperature (°C above the ambient in-
cubator temperature of 40°C). Oxygen levels in the
bioreactor system were measured on all sampling
days using a Demista OT-21 oxygen probe (Arling-
ton Heights, IL) prior to removing duplicate samples
(25 g) with a sanitized grabbing tool at both the bot-
tom center and top center locations.
Chemical and Microbiological Analyses
All compost ingredients (chicken litter, wheat
straw, and cottonseed meal) as well as the initial
compost mixtures were analyzed for carbon, nitro-
gen, and moisture contents. Carbon content was
determined on the basis of ash content obtained af-
ter combustion of samples at 550°C. The University
of Georgia’s Soil Testing Laboratory (Athens, Geor-
gia) was used for analysis of nitrogen content via a
macro-Kjeldahl method. Moisture levels were based
on residual weights of vacuum dried samples.
Ammonia concentrations in compost samples

100 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
(5 g) was determined with a phenol-hypochlorite
spectrophotometric procedure (Weatherburn, 1967),
whereas the Hach spectrophotometric Method 8196
test kit (Loveland, CO) as adapted by Montgomery
et al. (1962) was used to measure volatile acid con-
centrations in compost samples. Measurement of
pH was made with an Acumet Basic pH meter (Fisher
Scientific, Pittsburgh, PA) on compost samples (5 g)
dispersed in 250 ml of deionized water.
Salmonella and L. monocytogenes were enumer-
ated by direct plate counts (limit of detection was 2
log CFU/g) or detected by selective enrichment cul-
ture (limit of detection was 1 log CFU/g). In either
case, compost samples (5 g) placed in a Whirl-Pak
bag were first pummeled in a Stomacher 400 Circu-
lator (Seward Ltd., West Sussex, UK) for 1 min after
adding 45 mL of 0.1% peptone water. Diluted (1:10)
aliquots of this homogenate were applied to either
TSA-A plates to enumerate Salmonella or MOX-E
plates to enumerate L. monocytogenes. Enrichment
cultures of Salmonella and L. monocytogenes con-
sisted of adding 1 mL of the homogenate to 9 ml
of selective enrichment medium (TSB-A or BHIB-E,
respectively) and incubating this mixture for 24 h at
37°C. Aliquots of these enriched samples were then
streaked onto TSA-A or MOX-E plates to determine
the presence or absence of fluorescent Salmonella
or L. monocytogenes colonies, respectively.
Statistical Analyses
The StatGraphics Centurion XVI software, version
16.1.03 (StatPoint Technologies, Inc., Herndon, VA)
was used for statistical analysis of the collected data;
however, pathogen populations were first converted
to logarithmic values prior to conducting these op-
erations. When samples did not yield any colonies
during plate count enumeration but did have fluo-
rescent colonies on plates streaked from enrichment
cultures, a value of 1.0 log CFU/g, corresponding
to the limit of detection by enrichment culture, was
assigned to that sample. Otherwise, samples yield-
ing negative results for both plate counts and en-
richment cultures were assigned a value of 0.0 log
CFU/g. After conversion of enrichment culture data,
all data were subjected to general linear models
analysis of variance (GLM ANOVA) to determine the
significance of experimental variables over all sam-
pling times examined in the study. To differentiate
treatments at individual sampling times, the data
were subjected to one-way ANOVA and when sta-
tistical differences were observed (P < 0.05), sample
means were differentiated using the least significant
difference test. Multiple linear regression analysis
was also conducted on data from each sampling
day and treatment in an attempt to relate the total
pathogen loss in the mixtures to the independent
variables of pH, cumulative heat, and concentrations
of volatile acids and ammonia.
RESULTS AND DISCUSSION
Chicken litter, collected from broiler houses, was
mixed with wheat straw, cottonseed meal, and water
in combinations to give mixture treatments varying in
their initial C:N ratio. Following analysis of these ini-
tial compost mixtures, the C:N ratios that were mea-
sured for the 3 independent replicate trials averaged
20.6 ± 1.7, 32.4 ± 2.4, and 43.6 ±1.7, respectively.
Initial moisture contents in the 20:1, 30:1, and 40:1
C:N ratio formulations were 62.7 ± 1.9, 60.8 ± 1.7,
and 60.5 ± 2.8%, respectively. Continued monitor-
ing of moisture contents on days 2 and 6 revealed
that compost mixtures were generally above 40%
moisture during this time and thus aerobic microbial
activity would not have been inhibited (Rynk, 1992).
Oyxgen concentrations during composting were also
well above the 5% level that is considered to limit
aerobic microbial activity (Rynk, 1992).
All compost mixtures were initially characterized as
slightly alkaline (Table 1). After one day of compost-
ing, the pH of all mixtures had decreased from 1.5 to
2.2 units and declines were greater as the C:N ratio of
the compost formulation decreased. After this point in
time, the pH of all mixtures increased. Overall, there
were no significant differences in pH with the different
C:N ratio treatments throughout the composting pe-
riod (P < 0.05).

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 101
Table 1. pH (mean ± S.D.) in compost mixtures formulated with chicken litter, wheat straw, and cottonseed meal to different initial C:N ratios
Table 2. Volatile acid concentrations (mg/g, mean ± S.D.) in compost mixtures formulated with chicken litter, wheat straw, and cottonseed meal to different initial C:N ratios
1 Levels followed by a different letter are significantly different (P < 0.05)
Initial C:N ratio
Days 20:1 30:1 40:1
0 7.69 ± 0.48 c-e1 7.89 ± 0.19 d-f 7.79 ± 0.22 d-f
1 5.50 ± 0.39 a 5.96 ± 1.36 a 6.30 ± 1.37 ab
2 6.47 ± 1.68 a-c 7.52 ± 2.00 d 7.40 ± 1.80 cd
3 8.09 ± 0.95 d-f 7.37 ± 1.48 cd 7.26 ± 1.58 b-d
6 9.00 ± 0.22 f 8.55 ± 0.78 ef 8.12 ± 1.05 d-f
Initial C:N ratio
Days 20:1 30:1 40:1
0 5.49 ± 2.37 a-c1 4.27 ± 1.32 a 3.68 ± 1.32 a
1 11.68 ± 3.52 d-g 12.42 ± 3.26 fg 9.08 ± 4.04 c-e
2 12.08 ± 3.59 e-g 11.34 ± 4.91 d-g 8.79 ± 3.88 b-d
3 13.26 ± 6.88 g 9.76 ± 4.49 d-f 5.79 ± 4.28 ab
6 8.80 ± 4.20 b-d 4.79 ± 2.67 a 4.85 ± 2.99 a
1 Levels followed by a different letter are significantly different (P < 0.05)

102 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
These results were in contrast to those that were
observed when compost mixtures were formulated
to different C:N ratios with dairy manure as the ni-
trogen source (Erickson et al., 2009a, b). In those
studies, compost mixtures formulated to a C:N ratio
of 40:1 did not decline in pH during the first day of
composting.
Volatile acids, including acetate, butyrate, and
propionate, are produced during the early phases of
aerobic composting and digestion (Beck-Friis et al.,
2003; Ugwuanyi et al., 2005a,b) and may be poten-
tial contributors to the pH declines observed in this
study during the first day of composting of chicken
litter. This suggestion was corroborated by the ob-
served increase in volatile acid levels that occurred
in the chicken litter compost mixtures during the first
day (Table 2). The greatest increase in volatile acid
concentrations was observed in the 20:1 C:N com-
post mixtures, whereas the least increase occurred
in the 40:1 C:N compost mixture. Furthermore, as
composting continued, volatile acid levels declined
in all compost mixtures, but the decrease was slower
in 20:1 C:N compost mixtures than in the 40:1 C:N
compost mixtures. Generally, facultative anaerobic
microorganisms produce volatile acids in response
to low oxygen concentrations (Brinton, 1998); how-
ever, it would appear that the nutrient conditions
provided in the 20:1 C:N compost mixtures were
more conducive than the other compost mixture for-
mulations for generating such compounds.
Ammonia is another common byproduct pro-
duced during the degradation of chicken manure or
chicken litter (Bush et al., 2007; Himathongkham et
al., 2000). There were significant differences in the
ammonia concentrations of the different formula-
tions of the chicken compost mixtures (Table 3).
Specifically, when all sampling days were taken into
account, the 20:1 C:N ratio compost mixture had the
highest ammonia concentrations, whereas the low-
est levels were in the 40:1 C:N ratio compost mix-
tures (P < 0.05). In addition, during the composting
process, the ammonia concentrations were continu-
ously shifting, with maximal levels found in the 20:1
C:N mixtures on day 3, whereas maximal levels in
Table 3. Ammonia concentrations (µg/g, mean ± S.D.) in compost mixtures formulated with chicken litter, wheat straw, and cottonseed meal to different initial C:N ratios
Initial C:N ratio
Days 20:1 30:1 40:1
0 131.6 ± 24.7 a-c1 114.2 ± 23.8 ab 99.7 ± 30.2 ab
1 218.6 ± 94.3 c-e 215.9 ± 74.2 c-e 132.5 ± 23.4 b
2 271.0 ± 98.9 e 217.0 ± 56.4 c-e 145.3 ± 19.9 bc
3 379.4 ± 158.6 f 185.8 ± 47.1 b-d 123.9 ± 73.8 ab
6 246.6 ± 37.9 de 168.8 ± 216.7 bc 54.3 ± 57.3 a
1 Levels followed by a different letter are significantly different (P < 0.05)

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 103
Table 4. Cumulative metabolic heat > 40°C (degree-days1, mean ± S.D. )during composting of mixtures formulated with chicken litter, wheat straw, and cottonseed meal to different initial C:N ratios
Initial C:N ratio
Days 20:1 30:1 40:1
1 2.90 ± 2.00 a2 2.84 ± 2.36 a 3.48 ± 3.17 a
2 6.42 ± 4.53 ab 9.14 ± 10.67 a-c 9.16 ± 7.79 a-c
3 15.96 ± 10.02 cd 16.72 ± 16.09 b-d 19.05 ± 11.17 d
1 Accumulated product of temperature (°C above the ambient incubator temperature of 40°C) and
composting time (days) 2 Levels followed by a different letter are significantly different (P < 0.05)
Table 5. Fate of Salmonella and L. monocytogenes populations (log CFU/g, mean ± S.D.) in com-post mixtures formulated with chicken litter, wheat straw, and cottonseed meal to different initial C:N ratios
1 Populations for each pathogen followed by a different letter are significantly different (P < 0.05).
Salmonella L. monocytogenes
Day 20:1 C:N 30:1 C:N 40:1 C:N 20:1 C:N 30:1 C:N 40:1 C:N
0 7.43 ± 0.25 f 7.22 ± 0.26 f 7.38 ± 0.44 f 7.53 ± 0.17 g 7.21 ± 0.12 g 7.14 ± 0.13 g
1 3.64 ± 0.83 d 3.73 ± 1.96 d 5.31 ± 1.16 e 3.49 ± 1.22 e 3.79 ± 1.09 e 4.90 ± 0.97 f
2 1.58 ± 1.82 bc 0.48 ± 0.89 a 2.57 ± 2.33 c 1.21 ± 1.48 c 0.99 ± 0.88 bc 2.40 ± 1.83 d
3 0.28 ± 0.66 a 0.28 ± 0.66 a 1.82 ± 2.42 c 0.00 ± 0.00 a 0.28 ± 0.66 ab 1.12 ± 1.05 c
6 0.00 ± 0.00 a 0.00 ± 0.00 a 0.57 ± 0.84 ab 0.00 ± 0.00 a 0.00 ± 0.00 a 0.00 ± 0.00 a

104 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
both the 30:1 and 40:1 C:N mixtures were detected
on day 2. The increased generation of both ammo-
nia and volatile acids in compost mixtures initially
formulated to low C:N ratios agrees with the models
presented by Delgado-Rodríguez et al. (2010) that
demonstrated a higher level of volatile compounds
present during municipal solid waste composting at
low C:N ratios.
Temperatures during composting of the mixtures
were monitored throughout the period when sam-
ples were collected from the bioreactors. To assess
the cumulative heat exposure above the ambient
incubator temperature of 40°C, time-temperature
curves were integrated using 40°C as the baseline.
Results for the first 3 days of composting, expressed
as cumulative heat > 40°C (degree days), are pre-
sented in Table 4 and, although heat generation in
compost mixtures was slightly greater as the C:N ra-
tio increased, it was not significantly different (P >
0.05). Heat generation within each bioreactor was
fairly homogeneous, as location was not a significant
factor affecting the cumulative levels (P > 0.05). In
contrast, over the three independent replicate trials,
the level of heat accumulated in the compost mix-
tures was significantly different from each other (P <
0.05). As the manure source for each of these inde-
pendent trials was collected at separate times from
the broiler houses, a plausible explanation is that
the chicken litter had been collected in the houses
at different periods of time before being removed
for composting. Aged manure used in composting
mixtures produces less heat than fresh manure (Berry
et al., 2013; Li et al., 2008). Such variability in manure
age and subsequent variability in heat generation
in this study would likely have contributed to an in-
ability to detect a significant effect of C:N ratio on
heat generation. A similar situation was also likely
responsible for the inconsistent response of heat
generation in compost mixtures formulated to dif-
ferent C:N ratios when using dairy manure (Erickson
et al., 2009b). In that study, no statistical differences
occurred in the heat generated for the different C:N
formulations in the bioreactor trials inoculated with
E. coli O157:H7, whereas in bioreactor trials inocu-
lated with L. monocytogenes, 20:1 formulations were
statistically different for the 30:1 C:N formulations.
During composting of chicken litter with different
C:N ratio mixtures, pathogen levels were monitored
(Table 5). Using ANOVA on data collected shortly
after composting was initiated (days 1 and 2 only),
it was revealed that the C:N ratio had a significant
effect on inactivation of L. monocytogenes and Sal-
monella (P < 0.05). For both pathogens, the levels
of the pathogen were higher in mixtures formulated
to a C:N ratio of 40:1 than in those mixtures formu-
lated to either 20:1 or 30:1. Slower inactivation had
been observed previously for Salmonella in dairy
manure compost mixtures of formulations having a
C:N ratio of 40:1 compared to ratios of 20:1 and 30:1
(Erickson, 2009a), whereas the C:N ratio in dairy ma-
nure formulations did not affect the inactivation of
L. monocytogenes (Erickson et al., 2009b). Pathogen
inactivation, however, was not log-linear, but was
characterized as biphasic. Hence, to determine if
the C:N treatment affected inactivation during tail-
ing, the number of days to complete inactivation of
the pathogen was recorded for each replicate trial.
For Salmonella, the days to complete inactivation
ranged from 2 to 4, 2 to 5, and 3 to 8 for 20:1, 30:1,
and 40:1 C:N formulations, respectively, whereas the
days to complete inactivation of L. monocytogenes
ranged from 2 to 3, 2 to 4, and 4 to 5, respectively.
Given that only three values for each treatment were
available, ANOVA applied to the days to inactivation
data failed to reveal any significant effect by the C:N
ratio of the mixture for either pathogen (P > 0.05).
Despite this negative response, there is a trend of
increasing days to inactivation with an increasing ini-
tial C:N ratio of the compost mixture and if explored
in the future with a larger number of trial replicates,
could prove to be significant.
Comparison of Salmonella and L. monocytogenes
responses in the composting mixtures revealed no
significant differences in the rate of inactivation or
days to inactivation (Table 5, P > 0.05). The similar-
ity in responses contrast to those reported for com-
posting of rural sewage sludge with straw (Pourcher
et al., 2005) and composting of swine manure (Gre-
wal et al., 2007), in which L. monocytogenes persist-
ed for longer periods of time than Salmonella. Dif-

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 105
ferences in comparative pathogen response in this
study and others may have arisen due the different
isolates used or to the different formulations used
for composting.
To understand the contribution of potential chem-
ical and physical factors to pathogen losses during
composting on any one sampling day, models were
derived using backward stepwise regression. Both
cumulative heat and levels of volatile acids were fac-
tors included in those models, described below, and
explained 19.7% and 28.9 % of the variability in the
data for Salmonella and L. monocytogenes, respec-
tively.
Salmonella losses = 1.455 + (0.121*cumulative
heat>40°C) + (0.221* volatile acid concentration)
(P = 0.0123)
L. monocytogenes losses = 0.076 + (0.215*cumu-
lative heat>40°C) + (0.317* volatile acid concentra-
tion)
(P = 0.0018)
These models reveal that volatile acids, in addi-
tion to heat, have a bactericidal role in chicken litter
compost mixtures, particularly in those formulations
(i.e. 20:1 C:N compost mixtures) in which high con-
centrations of volatile acids are produced.
CONCLUSIONS
In summary, chicken litter was mixed with wheat
straw and cottonseed meal to give formulations hav-
ing initial C:N ratios of 20:1, 30:1 or 40:1. Although
the pH decreased in all formulations during the first
day of composting, the C:N ratio of the formulation
did not have a significant effect on pH (P > 0.05) nor
did it have a significant effect on the cumulated heat
generated in the mixtures during composting (P >
0.05). In contrast, the different C:N formulations did
have a significant effect on ammonia concentrations
and volatile acids produced during composting, with
the greatest amounts of these antimicrobials being
generated in the 20:1 C:N compost mixtures and the
least in the 40:1 C:N compost mixtures (P < 0.05).
Moreover, in the 20:1 C:N compost mixtures, the
inactivation rates of both Salmonella and L. mono-
cytogenes were higher as well as the days required
to achieve complete inactivation were in general
sooner, than in compost mixtures formulated to ei-
ther 30:1 or 40:1. Multiple linear regression models
that were derived from fitting pathogen losses to cu-
mulative heat and volatile acid levels were significant
and explained 20 to 29% of the variability in the data.
Hence, in conditions where heat may be insufficient
to inactivate pathogens (winter composting or at the
surface of unturned static compost piles), it may be
advantageous to formulate the initial C:N ratio of
chicken litter compost mixtures to values approach-
ing 20:1, as higher volatile acid concentrations in
these mixtures provide additional antimicrobial ac-
tivity.
ACKNOWLEDGEMENTS
The project was supported by the National Re-
search Initiative of the USDA Cooperative State Re-
search, Education, and Extension Service, grant #
2008-35201-18658.
REFERENCES
Beck-Friis, B., S. Smårs, H. Jönsson, Y. Eklind, and H.
Kirchmann. 2003. Composting of source-separat-
ed household organics at different oxygen levels:
Gaining an understanding of the emission dynam-
ics. Compost Sci. Util. 11:41-50.
Berry, E.D., P.D. Millner, J.E. Wells, N. Kalchayanand,
and M.N. Guerini. 2013. Fate of naturally occur-
ring Escherichia coli O157:H7 and other zoonotic
pathogens during minimally managed bovine
feedlot manure composting processes. J. Food
Prot. 76:1308-1321.
Brinton, W.F. 1998. Volatile organic acids in compost:
Production and odorant aspects. Compost Sci.
Util. 6(1):75-82.
Bush, D.J., M.H. Poore, G.M. Rogers, and C. Altier.

106 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
2007. Effect of stacking method on Salmonella
elimination from recycled poultry bedding. Biore-
source Technol. 98:571-578.
Chen, Z., J. Diao, C. Ionita, and X. Jiang. 2013. Ther-
mal inactivation of desiccation-adapted Salmonel-
la spp. in aged chicken litter. Appl. Environ. Micro-
biol. 79:7013-7020.
Chinivasagam, H.N., M. Redding, G. Runge, and P.J.
Blackall. 2010. Presence and incidence of food-
borne pathogens in Australian chicken litter. Br.
Poultry Sci. 51:311-318.
De Bertoldi, M., M.P. Ferranti, P. L’Hermite, and F.
Zucconi. 1987. Compost: Production, Quality and
Use. Elsevier Applied Science, New York, NY.
Delgado-Rodríguez, M., M. Ruiz-Montoya, I. Giral-
dez, I.O. Cabeza, R. López, and M.J. Díaz. 2010.
Effect of control parameters on emitted volatile
compounds in municipal solid waste and pine
trimmings composting. J. Environ. Sci. Health Part
A 45:855-862.
Erickson, M.C., J. Liao, L. Ma., X. Jiang, and M.P.
Doyle. 2009a. Inactivation of Salmonella spp. in
cow manure composts formulated to different
initial C:N ratios. Bioresource Technol. 101:1014-
1020.
Erickson, M.C., J. Liao, L. Ma., X. Jiang, and M.P.
Doyle. 2009b. Pathogen inactivation in cow ma-
nure compost. Compost Sci. Util. 17:229-236.
Erickson, M.C., J. Liao, G. Boyhan, C. Smith, L. Ma, X.
Jiang, and M.P. Doyle. 2010. Fate of manure-borne
pathogen surrogates in static composting piles of
chicken litter and peanut hulls. Bioresource Tech-
nol. 101:1014-1020.
Erickson, M.C., F. Critzer, and M.P. Doyle. 2010. Com-
posting criteria for animal manure. Georgetown
University Pew Charitable Trusts, Produce Safety
Project, Issue Brief. Available at: http://www.pe-
whealth.org/uploadedFiles/PHG/Content_Level_
Pages/Reports/PSP%20Issue%20Brief%20Series.
pdf. Accessed April, 2014.
Goss, M.J., A. Tubeileh, and D. Goorahoo. 2013. A
review of the use of organic amendments and the
risk to human health. Adv. Agron. 120:275-379.
Grewal, S., S. Sreevatsan, and F.C. Michel, Jr. 2007.
Persistence of Listeria and Salmonella during swine
manure treatment. Compost Sci. Util. 15:53-62.
Gu, G.Y., Z.Y. Luo, J.M. Cevallos-Cevallos, P. Adams,
G. Vellidis, A. Wright, and A.H.C. Van Bruggen.
2013a. Factors affecting the occurrence of Esch-
erichia coli O157 contamination in irrigation ponds
on produce farms in the Suwanee River watershed.
Can. J. Microbiol. 59:175-182.
Gu, G.Y., Z.Y. Luo, J.M. Cevallos-Cevallos, P. Adams,
A. Wright, and A.H.C. van Bruggen. 2013b. Occur-
rence and population density of Campylobacter
jejuni in irrigation ponds on produce farms in the
Suwannee River watershed. Can. J. Microbiol.
59:339-346.
Haley, B.J., D.J. Cole, and E.K. Lipp. 2009. Distribu-
tion, diversity, and seasonality of waterborne Sal-
monellae in a rural watershed. Appl. Environ. Mi-
crobiol. 75:1248-1255.
Himathongkham, S., H. Riemann, S. Bahari, S. Nu-
anualsuwan, P. Kass, and D.O. Cliver. 2000. Survival
of Salmonella typhimurium and Escherichia coli
O157:H7 in poultry manure and manure slurry at
sublethal temperatures. Avian Dis. 44:853-860.
Hogg, D., J. Barth, E. Faviono, M. Centemero, V.
Caimi, F. Amlinger, W. Devliegher, W. Brinton, and
S. Antler. 2002. Comparison of compost standards
within the EU, North America, and Australasia.
Main Report, The Waste and Resources Action
Programme, Banbury, Oxon, U.K. Available at:
http://www.compostingvermont.org/pdf/WRAP_
Comparison_of_Compost_Standards_2002.pdf.
Accessed April, 2014.
Hutchison, M.L., L.D. Walters, S.M. Avery, B.A. Synge,
and A. Moore. 2004. Levels of zoonotic agents in
British livestock manures. Lett. Appl. Microbiol.
39:207-214.
Hutchison, M.L., L.D. Walters, S.M. Avery, F. Munro,
and A. Moore. 2005. Analyses of livestock produc-
tion, waste storage, and pathogen levels and prev-
alences in farm manures. Appl. Environ. Microbiol.
71:1231-1236.
Leifert, C., K. Ball, N. Volakakis, and J.M. Cooper.
2008. Control of enteric pathogens in ready-to-eat
vegetable crops in organic and ‘low-input’ produc-
tion systems: a HACCP-based approach. J. Appl.
Microbiol. 105:931-950.

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 107
Li, X.J., R.H. Zhang, and Y.Z. Pang. 2008. Character-
istics of dairy manure composting with rice straw.
Bioresource Technol. 99:359-367.
Luo, Z., G. Gu, M.P. Adams, G. Vellidis, A. VanBrug-
gen, M. Danyluk, and A. Wright. 2013. Distribu-
tion and genetic diversity of Salmonella enterica
isolated from irrigation water in the Suwanee Riv-
er Watershed. J. Food Prot. Online Supplement
76S:104. (Abstr.)
Lynch, J.M. 1987. Lignocellulolysis in composts. In:
M. De Bertoldi, M.P. Ferranti, P. L’Hermite, and F.
Zucconi, Eds. Compost: Production, Quality and
Use. Elsevier, New York. pp. 178-179.
Ma, L, G. Zhang, and M.P. Doyle. 2011. Green fluo-
rescent protein labeling of Listeria, Salmonella,
and Escherichia coli O157:H7 for safety-related
studies. PLoS One 6:e18083.
MacDonald, J.M., M.O. Ribaudo, M.J. Livingston, J.
Beckman, and W. Huang. 2009. Manure use for fer-
tilizer and for energy: Report to Congress. USDA-
ERS Publication No. AP-037. 53 pp. http://www.
ers.usda.gov/ersDownloadHandler.ashx?file=/me-
dia/156155/ap037_1_.pdf. Accessed Jan, 2014.
Montgomery, H.A.C., J.F. Dymock, and N.S. Thom.
1962. The rapid colorimetric determination of or-
ganic acids and their salts in sewage-sludge liquor.
Analyst 87:949-955.
Moore, P.A., Jr., T.C. Daniel, A.N. Sharpley, and C.W.
Wood. 1998. In: Wright, R.J., W.D. Kemper, P.D.
Millner, J.F. Power, and R.K. Korcak, Eds. Poultry
manure management. Agricultural uses of munici-
pal, animal, and industrial byproducts. USDA-ARS:
Conservation Research Report #44. pp. 60-77.
Pourcher, A.-M., P. Morand, F. Picard-Bonnaud, S. Bil-
laudel, S. Monpoeho, M. Federighi, V. Ferré, and
G. Moguedet. 2005. Decrease of enteric micro-
organisms from rural sewage sludge during their
composting in straw mixture. J. Appl. Microbiol.
99:528-539.
Ritz, C.W., and W.C. Merka. 2013. Maximizing poultry
manure use through nutrient management plan-
ning. UGA Extension Bulletin 1245.
Rynk, R. 1992. On-farm Composting Handbook.
Northeast Regional Agricultural Engineering Ser-
vice, Ithaca, NY.
Shepherd, M.W., Jr., P. Liang, X. Jiang, M.P. Doyle,
and M.C. Erickson. 2007. Fate of Escherichia coli
O157:H7 during on-farm dairy manure-based com-
posting. J. Food Prot. 70:2708-2716.
Shepherd, M.W., Jr., R. Singh, J. Kim, and X.P. Jiang.
2010. Effect of heat-shock treatment on the sur-
vival of Escherichia coli O157:H7 and Salmonella
enterica Typhimurium in dairy manure co-compos-
ted with vegetable wastes under field conditions.
Bioresource Technol. 101:5407-5413.
Singh, R., J. Kim, M.W. Shepherd, Jr., F. Luo, and X.
Jiang. 2011. Determining thermal inactivation of
Escherichia coli O157:H7 in fresh compost by sim-
ulating early phases of the composting process.
Appl. Environ. Microbiol. 77:4126-4135.
Singh, R., J. Kim and X. Jiang. 2012. Heat inactiva-
tion of Salmonella spp. in fresh poultry compost by
simulating early phase of composting process. J.
Appl. Microbiol. 112:927-935.
Ugwuanyi, J.O., L.M. Harvey, and B. McNeil. 2005a.
Effect of digestion temperature and pH on treat-
ment efficiency and evolution of volatile fatty acids
during thermophilic aerobic digestion of model
high strength agricultural waste. Bioresource
Technol. 96:707-719.
Ugwuanyi, J.O., L.M. Harvey, and B. McNeil. 2005b.
Effect of aeration rate and waste load on evolution
of volatile fatty acids and waste stabilization dur-
ing thermophilic aerobic digestion of a model high
strength agricultural waste. Bioresource Technol.
96:721-730.
United States Department of Agriculture [USDA].
2013. Poultry – Production and Value. 2012 Summa-
ry. Available at: http://usda.mannlib.cornell.edu/
usda/current/PoulProdVa/PoulProdVa-04-29-2013.
pdf. Accessed April, 2014
United States Environmental Protection Agency [US
EPA]. 1999. Standards for the use or disposal of
sewage sludge (40 CFR parts 403 and 503). Revised
August 4, 1999. http://www.epa.gov/EPA-WA-
TER/1999/August/Day-04/w18604.htm. Accessed
Jan, 2014.
United States Environmental Protection Agency [US
EPA]. 2013. Literature review of contaminants in
livestock and poultry manure and implications for

108 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
water quality. http://water.epa.gov/scitech/cec/
upload/Literature-Review-of-Contaminants-in-
Livestock-and-Poultry-Manure-and-Implications-
for-Water-Quality.pdf. Accessed Jan, 2014.
Wang, C.-M., C.-L. Shyu, S.-P. Ho, and S.-H. Chiou.
2007. Species diversity and substrate utilization
patterns of thermophilic bacterial communities in
hot aerobic poultry and cattle manure composts.
Microbial Ecol. 54:1-9.
Weatherburn, M.W. 1967. Phenol-hypochlorite reac-
tion for determination of ammonia. Anal. Chem.
39:971-974.
Wichuk, K.M. and D. McCartney. 2007. A review of
the effectiveness of current time-temperature
regulations on pathogen inactivation during com-
posting. J. Environ. Engr. Sci. 6:573-586.
Williams, J.E., and S.T. Benson. 1978. Survival of Sal-
monella Typhimurium in poultry feed and litter at 3
temperatures. Avian Dis. 22:742-747.
Ziemer, C.J., J.M. Bonner, D. Cole, J. Vinjé, V. Con-
stantini, S. Goyal, M. Gramer, R. Mackie, X.J. Meng,
G. Myers, and L.S. Saif. 2010. Fate and transport of
zoonotic, bacterial, viral, and parasitic pathogens
during swine manure treatment, storage, and land
application. J. Anim. Sci. 88:E84-E94.

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 109
www.afabjournal.comCopyright © 2014
Agriculture, Food and Analytical Bacteriology
ABSTRACT
Recently, a vast array of potential antibiotic alternatives have been introduced and researched in the
livestock industry as a means to provide livestock producers with products that will positively impact animal
health and performance. Some of these products may be used in conjunction with current antibiotic usage
strategies, and some of these products may be used to completely replace some antibiotics in livestock
production. These innovative antibiotic alternatives include direct fed microbials (DFM), yeast extracts,
bacteriocins, bacteriophages, phytochemicals, and various acids. Many of these products have the ability
to promote animal health and improve growth performance simultaneously, and some of these compounds
may additionally enhance food safety through pre-harvest pathogen reduction. Antibiotic alternatives may
be essential tools for livestock production in the future should legislation arise that inhibits prophylactic
usage of conventional antibiotics and as a means to appeal to shifting consumer demands. Furthermore, it
is also possible that these alternatives can be used as an additional supplement to incorporate into current
practices and strategies in livestock production to maximize the potential to enhance both animal health
and growth performance. This review will discuss potential alternative antimicrobial supplements in animal
agriculture and their impact on animal health, performance and pathogen reduction.
Keywords: Antibiotic, livestock, animal health, review
Correspondence: Todd Callaway, [email protected]: +1-979-260-9374 Fax: +1-979-260-9332.
REVIEWAlternative Antimicrobial Supplements
That Positively Impact Animal Health and Food Safety
P. R. Broadway1, J. A. Carroll2, and T. R. Callaway3
1Department of Animal and Food Sciences, Texas Tech University, Lubbock, TX2Livestock Issues Research Unit, Agricultural Research Service, USDA, Lubbock, TX
3Food and Feed Safety Research Unit, Southern Plains Agricultural Research Center, Agricultural Research Service, USDA, College Station, TX
“Proprietary or brand names are necessary to report factually on available data; however, the USDA neither guarantees nor warrants the standard of the product, and the use of the name by the USDA implies no approval of the product, or exclusion of others that may
be suitable.” USDA is an equal opportunity provider and employer
Agric. Food Anal. Bacteriol. 4: 109-121, 2014

110 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
INTRODUCTION
Internet news sources and social media play in-
creasing roles in publicizing livestock production
practices and influence public opinion and percep-
tion, both fairly and unfairly. Recently, internet me-
dia sources have published propaganda to condemn
the use of antibiotics in meat and poultry (Consumer
Reports, 2012, 2013). Consumer-driven changes to
the market due to shifting consumer demands mean
that the beef, pork and poultry industries are faced
with increasing challenges to profitability and must
search for alternatives to remain profitable while
still remaining environmentally friendly. Thus, the
animal production industry needs potential alter-
native strategies to supplement or replace current
antibiotic implementation practices. Furthermore,
as consumer purchasing trends continue to gravi-
tate towards “natural” and “organic” products, the
livestock industry needs to be equipped to survive
in an environment where sub-therapeutic concentra-
tions of antibiotics supplemented as feed additives
are absent. Consumer demand and public concern
have previously been shown to influence antibiotic
legislation. For example, the European Union (EU)
banned sub-therapeutic supplementation of animal
feeds with antibiotics (Pradella, 2006). Recently, the
U.S. Food and Drug Administration (FDA) issued a
guidance to further regulate antibiotic usage in food
animals. While this directive does not eliminate the
use of antibiotics, some believe this is the beginning
of the end for antibiotic usage in food animals. For-
tunately, substantial research has been conducted
regarding the use of antibiotics in food animals, and
many alternatives have been proposed and evalu-
ated in regard to meeting the shifting consumer
demands without impinging on the health, welfare,
profitability, or wholesomeness of the food supply.
The gastrointestinal (GI) tract of animals is popu-
lated with a complex microbial ecosystem that is es-
sential for the function, growth, and overall health of
the animal (Chaucheyras-Durand and Durand, 2010);
therefore, any potential alternatives to antibiotics
must support this symbiotic relationship. Evidence
across multiple species generally supports the posi-
tive effects antibiotics exert on growth promotion,
growth rates, and overall animal health, thus produc-
ers have adopted the use of these pharmaceuticals
over the years as a strategy to increase profitabil-
ity, performance and animal health. Antibiotics are
typically defined as compounds that inhibit bacteria
(antibacterials), while antimicrobials are compounds
that inhibit microorganisms. Many livestock produc-
ers currently utilize antibiotic/antimicrobial feeding
and production strategies that alter the microbial
ecology of the GI tract of the animal to benefit the
overall production efficiency of their animals, as well
as strategies that can eliminate or reduce foodborne
pathogens that may contaminate the food supply
(Perlman 1973). However, these positive effects must
be replicated by strategies that fill in the gap left if
and when antibiotic use in food animals in banned in
the U.S. While strategies such as genetic selection,
selective breeding, and other management prac-
tices are utilized to promote health and profitability
along with antibiotics, this review will focus primarily
on feed additives.
ALTERNATIVES TO ANTIBIOTICS IN LIVE-STOCK PRODUCTION
Due to recent developments in feed additives,
direct fed microbials (DFM), and pharmaceuticals,
producers now have multiple options available to
enhance the natural microbial ecology of the animal,
to prevent illness and improve production efficiency.
While numerous alternatives to antibiotic use have
been researched in animal production systems (Table
1), to date, there has been no “silver bullet” identi-
fied, nor is one likely to be found. While it has been
demonstrated that many of the currently used antibi-
otics work across species, the results associated with
the use of antibiotic alternatives have been inconsis-
tent at times with varied results generated from spe-
cies to species. Additionally, most alternatives have
been evaluated as potential prophylactic treatments,
and they may not be effective in treating actual illness
or disease. Often times, the cost of antibiotic alter-
natives are offset by increases in performance; how-

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 111
ever, depending on different management and pro-
duction strategies, these alternatives have not been
demonstrated to be as effective as antibiotics. Each
strategy has specific advantages and limitations in-
cluding: effectiveness, production stage/system, ani-
mal age, animal type, changes in performance vari-
ables, cost, and labor. All of these variables must be
explored before implementing antibiotic alterative(s)
in a livestock production setting.
Direct-Fed Microbials (DFM)
One such strategy that may be a potential alterna-
tive to antibiotics is known as competitive enhance-
ment. Callaway et al. (2008) defines competitive
enhancement strategies as introducing live cultures
of bacteria or fungi into the GI tract that provide a
competitive advantage to commensal organisms
that can, in turn, exclude pathogenic bacteria (e.g.,
supplementing with a probiotic, or addition of a pre-
biotic). Probiotic supplements fed to livestock are
defined by Chaucheyras-Durand and Durand (2010)
as “live microorganisms that possess the ability to
evoke positive health benefits (at appropriate dos-
age concentrations) in the animal to which the micro-
organism was administered”; however, when used to
treat or prevent disease, this definition is not identi-
cal to the definition provided by the Food and Drug
Administration (FDA). Prebiotics are defined as feed
ingredients that benefit the host by selectively stim-
ulating the growth or activity of bacteria (Gibson and
Table 1. Alternatives to antibiotics that may be used in food animals that may be used to pro-mote overall animal health and/or impact pathogen colonization and shedding
DFM1 Bacteriophages Phytochemicals Acids
Bacillus Finylase Citrullene Acetic acid
Lactobacillus Citrus pulp Caproic acid
Lactococcus Curcumin Formic acid
Streptococcus Eugenol Fumarate
Yeast Flavonoids Malate
Yeast cell wall Limonene Propionic acid
Linalool Sodium Chlorate
Piperin
Thymol
1Direct-Fed Microbials

112 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
Roberfroid, 1995). A combination of prebiotic ingre-
dients is referred to as synbiotics (Patterson and Bur-
kholder, 2003). Fuller et al. (1989) explained that the
fully developed GI tracts use its microbial ecosystem
symbiotically to fill voids in microbial populations
and thereby inhibit the colonization of pathogens.
However, even when the GI tract is fully developed
and contains a healthy microflora, some pathogens
may be able to attach and cause illness. When a GI
flora ecosystem is fully developed and the system is
in a well-balanced state, the bacteria attach to the
intestinal epithelium and can possibly physically in-
hibit the binding and colonization of pathogens that
may be introduced into the gut (Collins and Gibson,
1999; Lloyd et al., 1977). Numerous compounds are
produced by bacteria in the gut such as volatile fatty
acids (VFA) that may inhibit the growth of and/or kill
pathogens introduced to the animal (Wolin, 1969;
Walsh et al., 2008).
Direct-fed microbials have recently gained inter-
est from the livestock feed industry as a tool to in-
crease performance while simultaneously serving as
a replacement for antibiotics (Ghorbani et al., 2002).
These DFM products have been shown to enhance
the formation of a healthy microbial community
within the GI tract of the animal (Fuller, 1999). As
with antibiotics, addition of DFM in the diet of dairy
cows has been shown to reduce the risk of ruminal
acidosis (Nocek et al., 2000). While the mode of ac-
tion that reduces acidosis with the inclusion of lac-
tate producing bacteria is not fully understood, the
phenomena may be the result of changes in fermen-
tation and microbial populations (Ghorbani et al.,
2002). Enhanced performance parameters such as
feed efficiency and ADG have also been reported
in feedlot cattle supplemented with DFM (Swinney-
Floyd et al., 1999; Rust et al., 2000).
Some bacteria and DFM feed additives produce
compounds called bacteriocins which are proteins
synthesized by bacteria that inhibit the growth of
other bacteria occupying the same environmental
niche (Jack et al., 1995). Such bacteriocins have been
isolated from the rumen of cattle (Wells et al., 1997;
Russell and Mantovani, 2002). Bacteriocins, such as
those commonly produced by Lactococcus bacte-
ria have the ability to inhibit pathogens such as E.
coli (Russell and Mantovani, 2002) that pose a risk
to human health. Brashears et al. (2003) reported
that supplementing feedlot cattle with a Lactobacil-
lus DFM was effective in reducing E. coli O157:H7
in fecal samples as well as reducing prevalence at
slaughter.
As in cattle, Lactobacillis has been shown to in-
hibit multiple strains of Salmonella in poultry (Jin et
al., 1996). A combination of L. acidophilus and Strep-
tococcus faecium cultures were shown to reduce col-
onization of Campylobacter jejuni (Morishita et al.,
1997). Reductions in E. coli and Salmonella strains
have also been observed when poultry were supple-
mented with Bacillus subtilis spores (LaRagione et
al., 2001; Laragione and Woodward, 2003). While
bacteriocins may be beneficial in most cases, some
bacteria may exhibit resistance to specific bacterio-
cins (Russell and Mantovani, 2002). Colicin E1, a bac-
teriocin produced by E. coli, has been shown to be
effective against pathogens that may be present in
young swine that contribute to diarrheal symptoms
(Cutler et al., 2007). Reduction/elimination of these
pathogens through the use of Colicin E1 may be an
effective tool to improve health and performance
(Cutler et al., 2007). In addition to pre-harvest patho-
gen reduction applications, Colicin E1 has also been
used successfully when applied directly to beef car-
casses to inhibit E. coli O157:H7 (Patton et al., 2008).
Researching probiotic supplementation in swine
has yielded inconsistent results (Turner et al., 2001).
Some studies have shown no differences in the
growth and performance of swine fed Lactobacilli
(Harper et al., 1983), while other studies reported en-
hanced growth and profitiability (Jasek et al., 1992;
Gombo et al., 1995). Bacillus spp. feed supplemen-
tation has also been shown to decrease incidence
of disease, reduce E. coli shedding and improve
feed efficiency in swine (Bonomi, 1992; Kyriakis et
al., 1999; Succi et al., 1995). Other probiotics that
have exhibited positive effects in growing and finish-
ing swine are Streptococcus spp. (Turner et al., 2001).
Multiple studies with Streptococcus cultures have
suggested that supplementation with these probi-
otics enhance growth and feed conversion in swine

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 113
(Kumpercht and Zobac, 1998; Roth and Kirchgessner,
1986; Underahl, 1983).
Another probiotic that may be utilized in swine is
a yeast culture. Yeast cultures fed to swine have been
reported to increase growth performance (Bertin et
al., 1997; Maloney et al. 1998; Matthew et al., 1998).
Yeast DFM may enhance digestion and maintain the
microbial ecosystem of the GI tract in swine (Van
Heugten et al., 2003), thereby making yeast DFM a
possible antibiotic alternative during the weaning
phase of young swine by inhibiting colonization of
pathogens and improving performance (Anderson
et al., 1999). Yeasts have also been used in cattle
as antibiotic alternatives (Jouany and Margavi, 2007).
For example, supplementation of live yeast probi-
otics in dairy cattle has been reported to increase
milk production and dry matter intake (Jouany, 2006;
Sniffen et al., 2004; Stella et al., 2007).
Yeast cell wall products are another type of feed
supplement that have been fed to livestock as a
means to improve animal performance, and to elimi-
nate the pathogenicity of certain bacteria. Yeast cell
wall components have been reported to function as
immunomodulators that activate immune compo-
nents such as macrophages, neutrophils, and other
immunocompetent cells (Eicher et al., 2006; Onder-
donk et al., 1992; Seljelid et al., 1987). Approximate-
ly half of the yeast cell wall is composed of biological
response modifiers (Bohn and BeMiller, 1995), and
these components have antibacterial (Kogan et al.,
1989), antimutagenic (Kogan et al., 2005), antioxi-
dant, and antitumor (Khalikova et al., 2005) activities
that may promote animal health. Kogan and Kocher
(2007) suggested that yeast cell wall polysaccharides
may prevent bacterial attachment of pathogens to
the mucosal epithelium in swine. Multiple studies
conclude that yeast products may protect swine from
bacterial infections while improving performance
parameters such as weight gain (Lemieux et al.,
2003; Rozeboom et al., 2005). Yeast cell wall prod-
ucts have also been shown to improve metabolism
in heifers during an endotoxin immune challenge
without degradation of carcass tissues (Burdick San-
chez et al., 2013). With respect to animal health and
food safety, yeast products have been reported to
help alleviate infections caused by E. coli (Buts et
al., 2006), Salmonella (Mahzounieh et al., 2006), and
Clostridium (Katz, 2006) in lab animals. In poultry,
supplementation with yeast cell wall products has
yielded inconsistent results with respect to growth
performance and pathogen reduction (Griggs and
Jacob, 2005).
Bacteriophages
Bacteriophages are viruses found commonly in
the GI tract and environment that prey specifically on
bacteria, including pathogenic bacteria. Bacterio-
phages can bind to specific bacterial receptors, in-
ject phage DNA, take control of that cell, reproduce,
and release new phages that lyse (or rupture) the host
bacterial cell, thus resulting in bacterial cell death
(Guttman et al., 2004 Kutter and Sulakvelidze, 2005).
However, there are limitations to the use of bacte-
riophages. Just as bacteria can become resistant to
antibiotics, bacteria may also become resistant to
bacteriophages (Sklar and Joerger, 2000; Smith and
Huggins, 1982; Smith et al., 1987). Another concern
with the use of a bacteriophage is the passage of the
supplement through the GI tract. Factors such as pH,
viscosity, and microbial populations may influence
the survivability and effectiveness of bacteriophage
therapies (Hurley et al., 2008). Hurley et al. (2008)
reported no reduction in fecal Salmonella shedding
when fed to 28-day old chickens. Sklar and Joerger,
(2000) reported minimal reductions in Salmonella
populations of chickens when fed bacteriophages,
and these researchers hypothesized that the intra-
cellular nature of Salmonella may prevent phage at-
tachment. When isolated and fed to feedlot cattle,
researchers concluded that bacteriophages could
be beneficial in a pre-harvest pathogen reduction in-
tervention strategy to combat food pathogens such
as E. coli (Callaway et al., 2008; Johnson et al., 2008).
Additionally, phage therapy has also been reported
successful in pathogen reduction in swine (Wall et
al., 2010) and sheep (Bach et al., 2003). In fact, Roz-
ema et al. (2009) reported that supplementation of
feedlot cattle with bacteriophages for the control
of E. coli O157:H7 shedding may be an alternative

114 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
pre-harvest intervention that could be utilized to
promote food safety in finishing cattle (Dini and De
Urraza 2010). Also a product called Finalyse® has
been developed for use as a hide spray to reduce E.
coli O157:H7 on hides of cattle before they enter the
slaughter plant, and FDA has issued no objection or-
der to the use of phage (Coffey et al., 2011).
Phytochemicals
Phytochemicals, such as citrus pulp, are being ex-
plored as replacements for sub-therapeutic antibiot-
ics supplementation in animal feeds. Citrus pulp and
peel are low cost byproduct feedstuffs with good
nutritive values (high TDN) that have been used in
beef and dairy production for many years (Arthing-
ton et al., 2002). Citrus pulp and peel contain es-
sentials oils, including but not limited to citrullene,
linalool, and limonene, that are bactericidal and can
alter the microbial ecology of the GI tract (Lota et
al., 2002; Fisher and Phillips, 2006; Viuda-Martos et
al., 2008). Recent studies have demonstrated that
feeding citrus pulp and peel to cattle, sheep, and
swine can reduce populations of E. coli O157:H7 and
Salmonella Typhimurium (Nannapaneni et al., 2008;
Callaway et al., 2008; Callaway et al., 2011). While
citrus pulp may have antipathogenic effects in cattle,
Broadway et al. (2013) reported that there were only
minimal changes to the bacterial ecology in the ru-
men of cattle supplemented with dried citrus pulp,
thereby concluding that citrus pulp may be used an
alternative agent to prevent colonization and shed-
ding of foodborne pathogens without significantly
altering ruminal microbial ecology and digestibility.
Essential oils have also been researched in poultry
to control pathogens (Griggs and Jacob, 2005). Thy-
mol, eugenol, curcumin, and piperin are some of the
essential oils found in thyme, clove, turmeric, and
black pepper, and these products have been shown
to inhibit enteridis causing Clostridium perfringens
in poultry (Mitsch, et al., 2004). Other pathogens
such as Salmonella and E. coli have been reduced,
in vitro, when cultured with thymol (Marino et al.,
1999; Karapinar and Aktug, 1987; Helander et al.,
1998) and thyme (Aktug and Karapinar, 1986). Other
phytochemicals including garlic (Singh and Shukla,
1984), cinnamon (Hernandez et al., 2004) and black
pepper (Dorman et al., 2000) have been reported to
reduce pathogen populations and enhance metabo-
lism in some cases.
Phytochemicals found in the seaweed such as
A. nodosum have been reported to reduce E. coli
prevalence of feces and on the hides of cattle at har-
vest (Behrends et al., 2000). Feeding this same com-
pound to swine was reported to enhance growth pa-
rameters but was unsuccessful in treating Salmonella
infections (Turner, 2001). Other plant extracts called
Saponins have been reported to alter ruminal micro-
flora (Killeen et al., 1998), and have been reported to
prevent the growth of E. coli (Sen et al., 1998).
Other naturally occurring phytochemicals, such as
flavonoids, may be included in a production strat-
egy to promote overall animal health and decrease
pathogen shedding into the food supply (Holiman
et al., 1996; Mandalari et al., 2007). Flavonoids are
found in plant tissues and bark, and display some an-
tioxidant capability (Pietta, 2000). Flavonoids have
also been reported to decrease the viability of patho-
genic bacteria such as E. coli and Salmonella, as
well as Candida albicans and Sacchromyces species
(Mandalari et al., 2007; Sohn et al., 2004; Friedman,
2007). Therefore, flavonoids may be an alternative to
antibiotics that could positively impact the health of
the animal while promoting food safety through the
reduction of pathogenic microorganisms.
Acids
Inorganic and organic acids, as well as inorganic
compounds, are another potential natural alterna-
tive that could be incorporated into livestock pro-
duction. Acids are used to eliminate foodborne
pathogens in food production, and they also may be
beneficial in live animals to decrease or eliminate the
presence of pathogens and improve digestion. Or-
ganic acids may target the cell wall and membrance
and interfere with bacterial metabolism, and inter-
nalization of dissociated acid components into the
cytoplasm also alters pH and interferes with cellular
metabolism (Ricke, 2003). However, the mechanisms

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 115
by which acids are bacteriocidal have not been ful-
ly elucidated and may target a variety of bacterial
components and metabolic processes (Ricke, 2003).
Organic acids such as lactic acid in diets have been
reported to decrease the incidence of pathogens
in livestock (Byrd et al. 2001). Other acids such as
propionate have been used to improve the ruminal
fermentation, and research pertaining to malate/fu-
marate reported increased lactic acid utilization by
Megasphaera and Selenomonas ruminantium which
resulted in similar impacts as those seen when utiliz-
ing ionophores (Martin and Nisbet, 1990; Nisbet and
Martin, 1990; Nisbet and Martin, 1993; Waldrip and
Martin, 1993). Propionic acid and formic acid fed
in combination has also been researched as a feed
supplement in broilers, and the acid was reported to
decrease populations of Salmonella in experimental-
ly-infected birds (Hinton and Linton, 1988). Similarly,
caproic acid was also shown to decrease Salmonella
Enteridis in chickens (Van Immerseel et al. 2004). In
addition to Salmonella inhibition, supplementing a
combination of organic acids such as formic, acetic,
and propionic acids were reported to reduce the
growth of Campylobacter (Chaveerach et al., 2004).
Another antibiotic alternative that may reduce
the prevalence of foodborne pathogens is sodium
chlorate, an organic product shown to inhibit nitrate
reductase positive bacteria. Callaway et al. (2002),
and Anderson et al. (2000, 2002) reported that E. coli
populations could be reduced without significant
changes in the microbial ecology of the rumen of
cattle. Byrd et al. (2003) reported that water treat-
ment with sodium chlorate was able to decrease the
prevalence of S. Typhimurium in broilers. Addition-
ally, sodium chlorate has been reported to decrease
E. coli O157:H7 populations in the GI tract of inocu-
lated swine (Anderson et al., 2001a), Sodium chlo-
rate has also been reported to decrease Salmonella
in swine prior to harvest (Anderson et al., 2001b).
Other studies have reported chlorate to inhibit the
survival of Salmonella while not interfering with po-
tentially beneficial species in the GI tract (Anderson
et al., 2001a,b; Byrd et al., 2003; Jung et al., 2003);
however, chlorate is still awaiting FDA approval for
use in food animals.
CONCLUSION
As more increasingly negative attention surrounds
the use of antibiotics in livestock production, and as
consumers increase the pressure for future legisla-
tion regarding livestock production practices, more
research and product development is needed to find
suitable alternatives to the use of antibiotic supple-
mented feed for growth promotion and health ben-
efits to meet the demand of an ever growing and
evolving consumer population. There are many
currently available or near-market ready products
that can assist livestock producers in the control of
bacterial populations within their animals to assist in
the maintenance of growth and production perfor-
mance parameters while simultaneously improving
the safety of food products for the consumer. How-
ever, many factors must be taken into consideration
when selecting a particular product as an alternative
to antibiotics, and there are advantages and limita-
tions to each product. Therefore, producers must
select product(s) that best suit their specific opera-
tional needs and are economically feasible.
REFERENCES
Aktug, S.E. and M. Karpinar. 1986. Sensitivity of
some common food-poisoning bacteria to thyme,
mint, and bay leaves. Int. J. Food Microbiol. 3:349-
354.
Anderson, D.B., V.J. McCracken, R.I. Aminov, J.M.
Simpson, R.I. Mackie, M.W.A. Verstegen, and H.R.
Gaskins. 1999. Gut microbiology and growth-
promoting antibiotics in swine. Pig News Info.
20:115-122.
Anderson, R. C, S. A. Buckley, T. R. Callaway, K. J.
Genovese, L. F. Kubena, R. B. Harvey, and D. J.
Nisbet. 2001a. Effect of sodium chlorate on Salmo-
nella Typhimurium concentrations in the weaned
pig gut. J. Food Prot. 64:255–258.
Anderson, R. C., S. A. Buckley, L. F. Kubena, L. H.
Stanker, R. B. Harvey, and D. J. Nisbet, 2000. Bacte-
ricidal effect of sodium chlorate on Escherichia coli

116 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
O157:H7 and Salmonella typhimurium DT104 in ru-
men contents in vitro. J. Food Prot. 63:1038-1042.
Anderson, R. C., T. R. Callaway, T. J. Anderson, L. F.
Kubena, N. K. Keith, and D. J. Nisbet. 2002. Bac-
tericidal effect of sodium chlorate on Escherichia
coli concentrations in bovine ruminal and fecal
contents in vitro. Microb. Ecol. Health D. 14:24–29.
Anderson, R.C., S.A. Buckley, T.R. Callaway, K.J.
Genovese, L.F. Kubena, R.B. Harvey, and D.J. Nis-
bet. 2001. Effect of sodium chlorate on Salmo-
nella Typhimurium concentrations in the weaned
pig gut. J. Food Prot. 64:255-258.
Anderson, R.C., T.R. Callaway, S.A. Buckley, T.J. An-
derson, K.J. Genovese, C.L. Sheffield, and D.J.
Nisbet. 2001. Effect of oral sodium chlorate ad-
ministration on Escherchia coli O157:H7 in the gut
of experimentally infected pigs. Int. J. Food Mi-
crobiol. 71:125-130.
Arthington, J.D., W.E. Kunkle, and A.M. Martin. 2002.
Citrus pulp for cattle, in the Veterinary Clinics of
North America – Food Animal Practice, G. Rogers
and M. Poore, Editors. W.B. Saunders Company:
Philidelphia, PA, p. 317-328.
Bach, S.J., T.A. McAllister, D.M. Veira, V.P.J. Gannon,
and R.A. Holley. 2003. Effect of bacteriophage
DC22 on Escherichia coli O157:H7 in an artificial
rumen system (Rusitec) and inoculated sheep.
Anim.Res. 52:89-101.
Behrends, L.L., J.R. Blanton, M.F. Miller, K.R. Pond,
and V.G. Allen. 2000. Tasco supplementation
in feedlot cattle: Effects on pathogen loads. J.
Anim. Sci. 78:106 (Abs.).
Bertin, G., M. Brault, M. Baud, M. Mercier, and J.
Tournot. 1997. Saccharomyces cervisiae I-1079,
microbial feed additive: Zootechnical effects on
piglets. In Digestive Physiology in Pigs. EAAP
Publ. No 88. P. 446. EEAP, Paris, France.
Bohn, J.A. and J.N. BeMiller. 1995. (1-3) β-D-Glucans
as biological response modifiers: a review of struc-
ture-functional activity relationships. Carbohydr.
Polym. 28:3-14.
Bonomi, A. 1992. Probiotics in pig breeding. Re-
sults from the use of Bacillus subtilis and Bacillus
licheniformis. Experimental contribution. Riv. Soc.
Ital. Sci. Aliment. 21:481.
Brashears, M.M, M.L. Galyean, G.H. Loneragan, J.E.
Mann, and K. Killinger-Mann. 2003. Prevalence
of Escherichia coli O157:H7 and performance by
beef feedlot cattle given Lactobacillus Direct-Fed
Microbials. J. Food Prot. 66:748-754.
Broadway, P.R., T.R. Callaway, J.A. Carroll, N.C.
Burdick, J.R. Donaldson, R.J. Rathmann, B.J. John-
son, J.T. Cribbs, L.M. Durso, D.N. Miller, D.J. Nis-
bet, and T.B. Schmidt. Evaluation of the ruminal
bacterial diversity of cattle fed diets containing cit-
rus pulp pellets (CPP) using bacterial tag-encoded
FLX amplicon pyrosequencing (bTEFAP). Agri.
Food & Anal. Bacteriol. 2:297-308.
Burdick Sanchez, N.C., T.R. Young, J.A. Carroll, J.R.
Corley, R.J. Rathmann, and B.J. Johnson. Yeast
cell wall supplementation alters the metabolic re-
sponses of crossbred heifers to an endotoxin chal-
lenge. Innate Immun. Accepted 29 Jan. 2013.
Buts, J.P., N. Dekeyser, C. Stilmant, E. Delem, F.
Smets, and E. Sokal. 2006. Saccharomyces bou-
lardii produces in rat small intestine a novel pro-
tein phosphatase that inhibits E. coli endotoxin by
dephosphorylation. Pediatric Research. 60:24-29.
Byrd, J. A., B. M. Hargis, D.J. Caldwell, R.H. Bailey,
K.L. Herron, J.L. McReynolds, R.L. Brewer, R.C. An-
derson, K.M. Bischoff, T.R. Callaway, and L.F. Kube-
na. 2001. Effect of lactic acid administration in the
drinking water during preslaughter feed withdraw-
al on Salmonella and Campylobacter contamina-
tion of broilers. Poultry Sci. 80:278-283.
Byrd, J.A., R.C. Anderson, T.R. Callway, R.W. More,
K.D. Knape, L.F. Kubena, R.L. Ziprin, and D.J. Nis-
bet. 2003. Effect of experimental chlorate product
administration in the drinking water on Salmonella
typhimurium contamination of broilers. Poultry
Sci. 82:1403-1406.
Callaway, T. R., J. A. Carroll, J. D. Arthington, C. Pratt,
T. S. Edrington, R. C. Anderson, M. L. Galyean, S.
C. Ricke, P. Crandall, and D. J. Nisbet. 2008. Cit-
rus products decrease growth of E. coli O157:H7
and Salmonella Typhimurium in pure culture and in
fer¬mentation with mixed ruminal microorganisms
in vitro. Foodborne Path. Dis. 5:621-627.
Callaway, T. R., J. A. Carroll, J. D. Arthington, T.
S. Edrington, M. L. Rossman, M. A. Carr, N. A.

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 117
Krueger, S. C. Ricke, P. Crandall, and D. J. Nisbet.
2011a. Escherichia coli O157:H7 populations in ru-
minants can be reduced by orange peel product
feeding. J. Food Prot. 74:1917-1921.
Callaway, T. R., R. C. Anderson, K. J. Genovese, T. L.
Poole, T. J. Anderson, J. A. Byrd, L. F. Kubena, and
D. J. Nisbet. 2002. Sodium chlorate supplementa-
tion reduces E. coli O157:H7 populations in cattle.
J. Anim. Sci. 80:1683–1689.
Callaway, T.R. , T.S. Edrington, A.D. Brabban, R.C.
Anderson, M.L. Rossman, M.J. Engler, M.A. Carr,
K.J. Genovese, J.E. Keen, M.L. Looper, E.M. Kut-
ter, and D.J. Nisbet. 2008. Bacteriophage isolat-
ed from feedlot cattle can reduce Escherichia coli
O157:H7 populations in ruminant gastrointestinal
tracts. Foodborne Path. Dis. 5:183-191.
Callaway, T.R., T.S. Edrington, R.C. Anderson, J.A.
Byrd, M.H. Kogut, R.B. Harvey, and D. J. Nis-
bet. 2008. Using antimicrobial cultures, bacte-
riocins, and bacteriophages to reduce carriage
of foodborne pathogens in cattle and swine.
Foodborne Path. and Dis. 5:205-224.
Chaveerach, P.L., J.A. Lipman, and F. van Knapen.
2004. Antagonistic activities of several bacteria on
in vitro growth of 10 strains of Campylobacter je-
juni/coli. Int. J. Food Microbiol. 90:43-50.
Coffey, B. L. Rivas, G. Duffy, A. Coffey, R.P. Ross, and
O. McAuliffe. 2011. Assessment of Escherichia
coli O157:H7-specific bacteriophages e11/2 and
e4/1c in model broth and hide environments. Int.
J. Food Microbiol. 147:188-194.
Collins, D.M. and G.R. Gibson. 1999. Probiotics,
prebiotics, and synbiotics: approaches for mod-
ulating the microbial ecology of the gut. Am. J.
Clin. Nutr. 69:1052S-1057S.
Consumer Reports. 2012. Antibiotics are widely
used by U.S. meat industry. Available: http://www.
consumerreports.org/cro/2012/06/antibiotics-are-
widely-used-by-u-s-meat-industry/index.htm
Consumer reports. 2013. Use of antibiotics on
healthy animals must stop. Available: http://www.
consumerreports.org/cro/2013/10/antibiotic-resis-
tant-superbugs-salmonella-outbreak/index.htm
Cutler, S.A., S.M. Loneragan, N.Cornick, A.K. John-
son, and C.H. Stahl. Dietary inclusion of Colicin
E1 is effective in preventing postweaning diarrhea
caused by F18-positive Escherichia coli in pigs. An-
timicrobial Agents and Chemotherapy. 51:3830-
3835.
Dini, C. and P. J. De Urraza. 2010. Isolation and selec-
tion of coliphages as potential biocontrol agents
of enterohemorrhagic and Shiga toxin-producing
E. coli(EHEC and STEC) in cattle. J. Appl. Micro-
biol. 109:873-887.
Dorman, H.J.D. and S.G. Deans. 2000. Antimicro-
bial agents from plants: Antibacterial activity of
plant volatile oils. J. Appl. Microbiol. 88:308-316.
Eicher, S.D., C.A. McKee, J.A. Carroll, and E.A. Pajor.
2006. Supplemental vitamin C and yeast cell wall
β-glucan as growth enhancers in newborn pigs and
as immunomodulators after an endotoxin chal-
lenge after weaning. J. Anim. Sci. 84:2352-2360.
Fisher, K., and C.A. Phillips. 2006. The effect of lem-
on, orange and bergamot essential oils and their
components on the survival of Campylobacter
jejuni, Escherichia coli O157, Listeria monocyto-
genes, Bacillus cereus and Staphylococcus aureus
in vitro and in food systems. J. Appl. Microbiol.
101:1232-1240.
Friedman, M. 2007. Overview of antibacterial, anti-
toxin, antiviral, and antifungal activities of tea flavo-
noids and teas. Mol. Nutr. Food Res. 51:116-134.
Fuller, R. 1989. Competitive exclusion and microl-
fora management: strategy for the swine industry.
J. Swine Health Prod. 7:229-232.
Fuller, R. 1999. Probiotics for farm animals. In: G.W.
Tannock (ed.) Probiotics – A Critical Review. P. 15.
Horizon Scientific Press, Wymondham, England.
Ghorbani, G.R., D.P. Morgavi, K.A. Beauchemin, and
J.A.Z. Leedle. 2002. Effects of bacterial direct-fed
microbials on ruminal fermentation, blood vari-
ables, and the microbial populations of feedlot
cattle. J. Anim. Sci. 80:1977-1985.
Gibson, G.R. and M.B. Roberfoid. 1995. Dietary
modulation of the human colonic microbiota:
introducing the concept of prebiotics. J. Nutr.
125:1401-1412.
Gombo, S., J. Tossenberger, and C. Szabo. 1995. Ef-
fects of probiotics and yeast culture on the perfor-
mance of pigs and dairy cows. Krmiva. 37:13.

118 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
Griggs, J.P. and J.P. Jacob. 2005. Alternatives to an-
tibiotics for organic poultry production. J. Appl.
Poult. Res. 14:750-756.
Guttman, B., R. Raya, and E. Kutter. Basic phage biol-
ogy. In: Bacteriophages: Biology and Applications.
Kutter E and Sulakvelidze A (eds). New York: CRC
Press, 2004, pp. 29–66.
Harper, A.F., E.T. Kornegay, K.L. Bryant, and H.R.
Thomas. 1983. Efficacy of virginamycin and a
commercially-available Lactobacillus probiotic in
swine diets. Anim. Feed Sci. Technol. 8:69-76.
Helander, I.M., H.L. Alakomi, K. Latva-Kala, T. Mattila-
Sandholm, I. Pol, E.J. Smid, L.G.M. Gorris, and A.
von Wright. 1998. Characterization of the action o
fselected essential oil components on gram-nega-
tive bacteria. J. Agric. Food Chem. 46:3590-3595.
Hernandez, F., J. Madrid, V. Garcia, J. Orengo, and
M.D. Megias. 2004. Influence of two plant extracts
on broilers performance, digestibility, and diges-
tive organ size. Poult. Sci. 83:169-174.
Hinton, M., and A.H. Linton. 1988. Control of Sal-
monella infections in broiler chickens by the acid
treatment of their feed. Vet. Rec. 123:416-421.
Holiman, P.C.H., M.G.L. Hertog, and M.B. Katan.
1996. Analysis and health effects of flavonoids.
Food Chem. 57:43-46.
Hurley, A., J.J. Maurer, and M.D. Lee. 2008. Using
bacteriophages to modulate Salmonella coloniza-
tion of the chicken’s gastrointestinal tract: lessons
learned from in silico and in vivo modeling. Avian
Diseases. 52:599-607.
Jack, R. W., J. R. Tagg, and B. Ray. (1995). Bacte-
riocins of gram-positive bacteria. Microbiol. Mol.
Biol. Rev. 59:171-200.
Jasek, S., R. Kalinowska, D. Knecht, and R. Pawiak.
1992. Effect of Biogen probiotic additon on repro-
duction results and physiological indices in pigs.
Effect of Biogen T dietary additive on fattening
performance and slaughter value. Roczniki Nau-
kowe Zootechniki. 31:239.
Jin, L.Z., Y.W. Ho, N. Abdullah, M.A. Ali, and S. Jalal-
udin. 1996. Antagonistic effects on intestinal Lac-
tobacillus isolates on pathogens of chicken. Lett.
Appl. Microbiol. 23:67-71.
Johnson, R.P., C.L. Gyles, W.E. Huff, S. Ojha, G.R. Huff,
N.C. Rath, and A.M. Donoghue. 2008. Bacterio-
phages for prophylaxis and therapy in cattle, poul-
try, and pigs. Anim. Health Res. Rev. 9:201-215.
Jouany, J.P., 2006. Optimizing rumen functions in
the close-up transition period and early lactation
to dry matter intake and energy balance in cows.
Anim. Repro. Sci. 96:250-264.
Jouany, J.P and D.P Morgavi. 2007. Use of ‘natural’
products as alternative to antibiotic feed additives
in ruminant production. Animal. 1:1143-1466.
Jung, Y.S., R.C. Anderson, J.A. Byrd, T.S. Edrington,
R.W. Moore, T.R. Callaway, J. McReynolds, and D.J.
Nisbet. 2003. Reduction of Salmonella Typhymuri-
um in experimentally challenged broilers by nitrate
adaptation and chlorate supplementation in drink-
ing water. J. Food Prot. 66:660-663.
Karapinar, M. and S.E. Aktug. 1987. Inhibition of
foodborne pathogens by thymol, eugenol, menthol
and anethole. Int. J. Food Microbiol. 4:161-166.
Katz, J.A. 2006. Probiotics for the prevention of anti-
biotic associated diarrhea and Clostridium difficile
diarrhea. J. Clin. Gastroenterology. 40:249-255.
Khalikova, T.A., S.Ya., Zhanaeva, T.A. Korolenko, V.I.
Kaledin, and G. Kogan. 2005. Regulation of activ-
ity of cathepsins B, L, and D in murine lymphosar-
coma model at a combined treatment with cyclo-
phosphamide and yeast polysaccharide. Cancer
Lett. 223:77-83.
Killeen, G.F., C.A. Madigan, C.R. Connolly, G.A.
Walsh, C.Clark, M.J. Hynes, B.F. Timmins, P. James,
D.R. Headon, and R.F. Power. 1998. Antimicrobial
saponins of Yucca schidigera and the implications
of the invitro properties for their in vivo impact. J.
Agric. Food Chem. 46:3178-3186.
Kogan, G., L. Masler, J. Sandula, J. Navarova, T. Trn-
ovec. 1989. Recent results on the structure and
immunomodulating activities of yeast glucan. In:
Crescenzi, V., I.C.M. Dea, S. Paoletti, SS. STvala,
and I.W. Sutherland (Eds.). Biomedical and Bio-
technological Advances in Industrial Polysaccha-
rides. Gordon and Breach Science Publishers.
New York. Pp. 251-258.
Kogan, G. A Stasko, K. Bauerova, M. Polovka, L.
Soltes, V. Brezova, J. Navarova, and D. Mihalova.
2005. Antioxidant properties of yeast (1-3) β-D-

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 119
glucan studied by electron paramagnetic reso-
nance spectroscopy and its activity in the adjuvant
arthritis. Carbohydr. Polym. 61:L 18-28.
Kogan, G. and A. Kocher. 2007. Role of yeast cell
wall polysaccharides in pig nutrition and health
protection. Livestock Sci. 109:1-3.
Kumprecht, I. and P. Zobac. 1998. Study of the ef-
fect of a combined preparation containing Entero-
coccus faecium M-74 (Streptococcus faecium) and
mannan-oligosaccharides in diets for weanling
piglets. Czech J. Anim. Sci. 43:477-481.
Kutter, E. ,and A. Sulakvelidze. Bacteriophages: Biol-
ogy and Applications. New York: CRC Press, 2005.
Kyriakis, S.C., V.K. Tsiloyiannis, J. Vlemmas, K. Sar-
ris, A.C. Tsinas, C. Alexopoulos, and L. Jansegers.
1999. The effect of probiotic LSP 122 on the con-
trol of post-weaning diarrhea syndrome of piglets.
Res. Vet. Sci. 67:223-228.
La Ragione, R.M., G. Casula, S.M. Cutting, and M.J.
Woodward. 2001. Bacillus subtilis spores compet-
itively exclude Escherchia coli O78:K80 in poultry.
Vet. Microbiol. 79:133-142.
La Ragione, R.M. and M.J. Woodward. 2003. Com-
petitive exclusion by Bacillus subtillis spores of
Salmonella enteric serotype Enteridis and Clos-
tridium perfringens in young chickens. Vet. Micro-
biol. 94:245-256.
LeMieux, M., L.L. Southern, T.D. Bidner. 2003. Ef-
fect of mannan oligosaccharides on growth per-
formance of weanling pigs. J. Anim. Sci. 81:2482–
2487.
Lloyd, A.B., R.B. Cumming, and R.D. Kent. 1977.
Prevention of Salmonella typhimurium infection in
poultry by pre-treatment of chickens and pults with
intestinal extracts. Aust. Vet. J. 53:82-87.
Lota, M.L., D. de Rocca Serra, F. Tomi, C. Jacque-
mond, J. Casanova. 2002. Volatile components of
peel and leaf oils of lemon and lime species. J.
Agric. Food Chem. 50:796-805.
Mahzounieh, M., I. Karimi, T.Z. Salehi, and R. Mar-
janian. 2006. The preventive effect of Saccharo-
myces bouldarii in pathogenesis of Salmonella ty-
phimurium in experimentally infected rats.
Maloney, C.A., J.D. Hancock, R.H. Hines, H. Cao, C.S.
Nemecek, and J.S. Park. 1998. Effects of a heat-
stable yeast product in pelleted diets for weanling
pigs. Kansas Agric. Exp. Stn. Rep. Prog. No. 819. P.
62. Kansas State Swine Day Report.
Mandalari, G, R. N. Bennet, G.Bisignano, D. Trom-
betta, A. Saija, C. B. Faulds, M.J. Gasson, A. Nar-
bad. 2007. Antimicrobial activity of flavonoids
extracted from bergamot (Citrus bergamia Risso)
peel, a byproduct of the essential oil industry. J.
App. Microbiol. 103:2056-2064.
Marino, M., C. Bersani, and G. Comi. 1999. Antimi-
crobial activity of the essential oils of Tymus vulgar-
is L. measured using a bioimpedometric method.
J. Food Prot. 62:1017-1023.
Martin, S.A., and D.J. Nisbet. 1990. Effects of Asper-
gillus oryzae fermentation extract on fermentation
of amino acids, bermudagrass and starch by mix-
ing ruminal microorganisms in vitro. J. Anim. Sci.
68:2142-2149.
Mathew, A.G., S.E. Chattin, C.M. Robbins, and D.A.
Golden. 1998. Effects of a direct-fed yeast culture
on enteric microbial populations, fermentation ac-
ids, and performance of weanling pigs. J. Anim.
Sci. 76:2138-2145.
Mitsch, P., K. Zitterl-Eglseer, B. Kohler, C. Gabler, R.
Losa, and I. Zimpernik. 2004. The effect of two dif-
ferent blends of essential oil components on the
proliferation of Clostridium perfringens in the in-
testines of broiler chickens. Poult. Sci. 83:669-675.
Morishita, T.Y., P.P. Aye, B.S. Harr, C.W. Cobb, and
J.R. Clifford. 1997. Evaluation of an avian specific
probiotic to reduct the colonization and shedding
of Campylobacter jejuni in broilers. Avian Dis.
41:850-855.
Nannapaneni, R., A. Muthaiyan, P. G. Crandall,
M. G. Johnson, C.A. O’Bryan, V. I. Chalova, T. R.
Cal¬laway, J. A. Carroll, J. D. Arthington, D. J. Nis-
bet, and S. C. Ricke. 2008. Antimicrobial activity of
commercial citrus-based natural extracts against
Escherichia coli O157:H7 isolates and mutant
strains. Foodborne Path. Dis. 5:695-699.
Nisbet, D.J. and S.A. Martin. 1990. Effect of dicar-
boxylic acids and Aspergillus oryzae fermentation
extract on lactate uptake by the ruminal bacterium
Selenomonas ruminantium. Appl. Environ. Micro-
biol. 56:3515-3518.

120 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
Nisbet, D.J. and S.A. Martin. 1993. Effects of fuma-
rate, L-malate, and Aspergillus oryzae fermenta-
tion extract on D-lactate utilization by the ruminal
bacterium Selenomonas ruminantium. Curr. Mi-
crobiol. 26:133-136.
Nocek, J.E., W.P. Kautz, J.A.Z. Leedle, and J.G. All-
man. 2000. Altering diurinal pH and in situ diges-
tion in dairy cows with ruminal supplementatioin of
direct-fed microbials (DFM) and yeast. J. Dairy Sci.
83(Suppl. 1):1242 (Absr.).
Onderdonk, A.B., R.L. Cisneros, P. Hinkson, and
G. Ostroff. 1992. Anti-infective effect of poly-
beta 1-6 glucotriosyl-beta 1-3-glucopyranose
glucan in vivo. Infect. Immun. 60:1642-1647.
Patterson, J.A. and K.M. Burkholder. 2003. Applica-
tion of prebiotics and probiotics in poultry produc-
tion. J. Poult. Sci. 82:627-361.
Patton, B.S., S.M. Lonergan, S.A. Culter, C.H. Stahl,
and J.S. Dickinson. 2008. Application of Colicin
E1 as a prefabrication intervention strategy. J.
Food Prot. 71:2519-2522.
Pietta, P.G. 2000. Flavonoids as antioxidants. J. Nat.
Prod. 63:1035-1042.
Ricke, S.C. 2003. Perspectives on the use of organic
acids and short chain fatty acids as antimicrobials.
Pout. Sci. 82:632-639.
Roth, F.X., and M. Kirchgessner. 1986. Nutritive ef-
fects of Streptococcus faecium (Strain M 74) in
starter pigs. Landwirtsch. Forsch. 39:198.
Rozeboom, D.W., D.T. Shaw, R.J. Tempelman, J.C.
Miguel, J.E. Pettigrew, and A. Connolly. 2005. Ef-
fects of mannan oligosaccharide and an antimicro-
bial product in nursery diets on performance of
pigs reared on three different farms. J. Anim. Sci.
83:2637–2644.
Rozema, E., T. Stephens, S.J. Bach, E.K. Okine, R.P.
Johsnon, K. Stanford, and T.A. McAllister. 2009.
Oral and rectal administration of bacteriophages
for control of Escherichia coli O157:H7 in feedlot
cattle. J. Food Prot. 72:241-250.
Russell, J.B. and H.C. Mantovani. 2002. The bacte-
riocins of ruminal bacteria and their potential as an
alternative to antibiotics. J. Mol. Microbiol. Bio-
technol. 4:347-355.
Rust, S. R., K. Metz, and D. R. Ware. 2000. Effect of
Bovamine rumen culture on the performance and
carcass characteristics of feedlot steers. p 22–26.
Mich. Agric. Exp. Sta. Beef Cattle, Sheep and For-
age Sys. Res. Dem. Rep. No. 569, East Lansing.
Seljelid, R.L., T. Rassmussen, O. Larm, and J. Hoff-
man. 1987. The protective effect of β1-3D-glucan-
derivated platic beads against Escherichia coli in-
fection in mice. Scan. J. Immunol. 25:55-60.
Sen, S., H.P.S. Makkar, S. Muetzel, and K. Becker.
1998. Effect of Quillaja saponaria saponins and
Yucca schidigera plant extract on growth of Esch-
erichia coli. Lett. Appl. Microbiol. 27:35.
Singh, K.V. and N.P. Shulka. 1984. Activity on mul-
tiple resistant bacteria of garlic (Allium Stivum) ex-
tract. Fitoterapia. 55:313-315.
Sklar, I.B. and R.D. Joerger. 2000. Attempts to uti-
lize bacteriophage to combat Salmonella enteric
enteridis infection in chickens. J. Food Safety.
21:15-29.
Smith, H.W. and M.B. Huggins. 1982. Successful
treatment of experimental Eschercichia coli infec-
tion in mice using phage: its general superiority
over antibiotics. J. Gen. Microbiol. 128:307-318.
Smith, H.W., M.B. Huggins, and K.M. Shaw. 1987.
The control of experimental Escherichia coli di-
arrhea in calves by means of bacteriophages. J.
Gen. Microbiol. 133:1111-1126.
Sniffen, C.J., Chaucheryas-Durand, F., De Ondarza,
M.B., and Donaldson, G. 2004. Predicting the im-
pact of a live yeast strain on rumen kinetics and ra-
tion formulation. In: proceeding of the 19th Annual
Southwest Nutrition and Management conference,
24-25 Februrary 2004, Tempe, AZ, pp. 53-59.
Sohn, H-Y, Son, KH, Kwon, C-S, Kang, SS. 2004. An-
timicrobial and cytotoxic activity of 18 prenylated
flavonoids isolated from medicinal plants: Morus
alba L., Morus mongolica Schneider, Broussnetia
papyrifera (L.) Vent, Sorphora flavescens Ait and
Echinosophora koreensis Nakai. Phytomedicine.
11:666-672.
Stella, A.V., R. Paratte, L. Valnegri, G. Cigalino, G.
Soncini, E.Chevaux, V. Dell’Orto, and G. Savoini.
2007. Effect of the administration of live Saccha-
romyces cerevisiae on milk production, milk com-
position, blood metabolites, and fecal flora in early

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 121
lactating dairy goats. Small Ruminant Res. 67:7-13.
Succi, G., A. Sandrucci, A. Tamburini, A. Adami, and
V. Cavazzoni. 1995. Effects of using a new strain
of Bacillus coagulans as a probiotic on the perfor-
mance of piglets. Riv. Suinicol. 36:59.
Swinney-Floyd, D., B.A. Gardiner, F.N. Owens, T. Reh-
berger, and T. Parrott. 1999. Effects of inoculation
with strain P-63 alone or in combination with Lac-
tobacillus acidophilus LA53545 on performance of
feedlot cattle. J. Anim. Sci. 77:77 (Abstr.).
Turner, J.L., S.S. Dritz, and J.E. Minton. 2001. Re-
view: Alternatives to conventional antimicrobials
in swine diets. The Professional Animal Scientist.
17:217-226.
Turner, J.L. 2001. Effects of natural alternatives to
conventional antimicrobials on growth perfor-
mance and immune function of nursery pigs during
n acute enteric disease challenge with Salmonella
typhimurium, Ph.D. dissertation. Kansas State Uni-
versity, Manhattan, KS.
Underahl, N.R. 1983. The effect of feeding Strepto-
coccus faecium upon Escherichia coli induced diar-
rhea in gnotobiotic pigs. Prog. Food Nutr. Sci. 7:5.
Van Heugeten, E., D.W. Funderburke, and K.L. Dor-
ton. 2003. Growth performance, nutrient digest-
ibility, and fecal microflora in weanling pigs fed live
yeast. J. Anim. Sci. 81:1004-1012.
Van Immerseel, F., J. de Buck, F. Boyen, L. Bohez, F.
Pasmans, J. Volf, M. Sevcik, I. Rychlik, F. Haeseb-
rouck, and R. Ducatelle. 2004. Medium-chain fatty
acids decrease colonization and invasion through
hilA suppression shortly after infection of chickens
with Salmonella enteric serovar Enteridis. Appl.
Environ. Microbiol. 70:3582-3587.
Viuda-Martos, M., Y. Ruiz-Navajas, J. Fernandez-
Lopez, and J. Perez-Alvarez. 2008. Antibacterial
activity of lemon (Citrus lemon L.), mandarin (Cit-
rus reticulata L.), grapefruit (Citrus paradisi L.) and
orange (Citrus sinensis L.) essential oils. J. Food
Safety. 28:567-576.
Waldrip, H.M. and S.A. Martin. 1993. Effects of an As-
pergillus oryzae fermentation extract and other fac-
tors on lactate utilization by the ruminal bacterium
Megasphaera elsdenii. J. Anim. Sci. 71:2770-2776.
Wall, S.K., J. Zhang, M.H. Rostango, and P.D. Ebner.
2010. Phage therapy to reduce preprocessing Sal-
monella infections in market-weight swine. Appl.
Environ. Microbiol. 76:48-53.
Walsh, M.C., G.E. Gardiner, O.M. Hart, P.G. Law-
lor, M. Daly, B. Lynch, B.T. Richert, S. Radcliffe, L
Giblin, C. Hill, G.F. Fitzgerald, C. Stranton, and P.
Ross. 2008. Predominace of a bacteriocin-produc-
ing Lactobacillus salivarius component of a five-
strain probiotic in the porcine ileum and effects on
host immune phenotype. FEMS Microbiol. Ecol.
64:317-327.
Wells, J. E., D. O. Krause, et al. 1997. A bacterio-
cin-mediated antagonism by ruminal lactobacilli
against Streptococcus bovis. FEMS Microbiol.
Ecol. 22:237-243.
Wolin, M.J. 1969. Volatile fatty acids and the inhi-
bition of Escherichia coli growth by rumen fluid.
Appl. Microbiol. 17:83-87.

122 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
www.afabjournal.comCopyright © 2014
Agriculture, Food and Analytical Bacteriology
ABSTRACT
Isoflavones are a group of chemicals that are found in legumes, predominantly in soybean and soy prod-
ucts. Soy isoflavones have been a component of the diet of certain populations for centuries. Many health
claims have been made for isoflavones including: cancer prevention, alleviation of menopausal symptoms,
positive effects on bone health and a lowering of blood lipids leading to lowered susceptibility to cardio-
vascular disease. However, because of their estrogenic activity some negative effects of isoflavones have
been postulated. This review examines the literature associated with benefits as well as the negative effects
of consumption of soy isoflavones. Results in some studies are limited or conflicting, but when viewed in
its entirety, the current literature supports the safety of isoflavones as typically consumed in diets based on
soy containing products.
Keywords: Isoflavones, soybeans, soy products, health benefits, cancer prevention, bone metabo-
lism, blood lipids
INTRODUCTION
Soybeans are legumes, plants that form root nod-
ules containing nitrogen-fixing soil bacteria (Rhizo-
bia) in a symbiotic relationship. The soybean plant
releases chemical signals, called isoflavonoids, to
attract the nitrogen fixing bacteria (Rolfe, 1988). Iso-
flavonoids, also known as isoflavones, are produced
by the same pathway that produces flavonoids, the
Correspondence: Philip G. Crandall, [email protected]: +1 -479-575-7686 Fax: +1-479-575-6936
phenylpropanoid pathway. The phenylpropanoid
pathway begins with phenylalanine and naringenin
being converted into the isoflavone genistein by two
enzymes, isoflavone synthase and a dehydratase,
that are found only in legumes (Deavours and Dixon,
2005). Naringenin chalcone, another intermediate
is converted to daidzein by the sequential action of
two other legume-specific enzymes, chalcone reduc-
tase and type II chalcone isomerase as well as iso-
flavone synthase (Deavours and Dixon, 2005). Within
the soybean, isoflavones are bound to a sugar mol-
ecule (glycosidic form) but fermentation or digestion
REVIEWHuman Health Benefits of Isoflavones From Soybeans
K. Kushwaha1, C. A. O’Bryan1,3, Dinesh Babu1§, P. G. Crandall1,3, P. Chen2, and S.-O. Lee1
1 Department of Food Science, University of Arkansas, 2650 Young Ave., Fayetteville, AR 72704
2Crop, Soil and Environmental Sciences, University of Arkansas, Fayetteville, AR 727013Center for Food Safety, University of Arkansas, Fayetteville, AR 72704
§Present address: Food Safety Toxicology, College of Pharmacy, University of Louisiana at Monroe, Monroe, LA 71209
Agric. Food Anal. Bacteriol. 4: 122-142, 2014
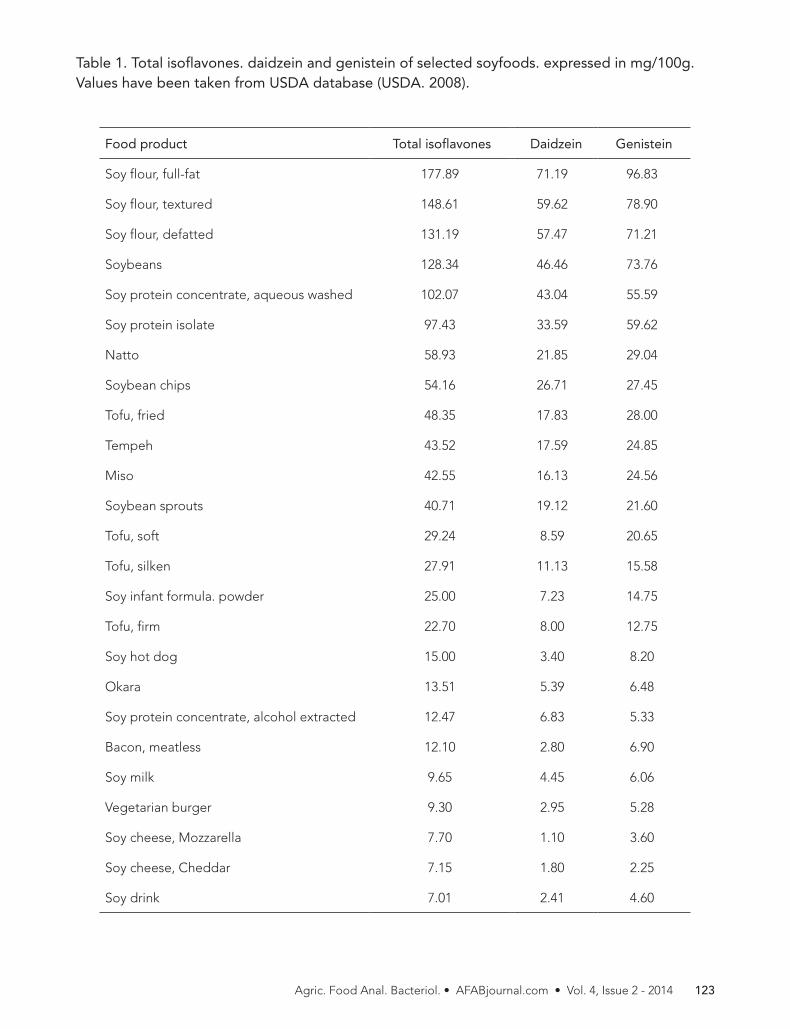
Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 123
Table 1. Total isoflavones. daidzein and genistein of selected soyfoods. expressed in mg/100g. Values have been taken from USDA database (USDA. 2008).
Food product Total isoflavones Daidzein Genistein
Soy flour, full-fat 177.89 71.19 96.83
Soy flour, textured 148.61 59.62 78.90
Soy flour, defatted 131.19 57.47 71.21
Soybeans 128.34 46.46 73.76
Soy protein concentrate, aqueous washed 102.07 43.04 55.59
Soy protein isolate 97.43 33.59 59.62
Natto 58.93 21.85 29.04
Soybean chips 54.16 26.71 27.45
Tofu, fried 48.35 17.83 28.00
Tempeh 43.52 17.59 24.85
Miso 42.55 16.13 24.56
Soybean sprouts 40.71 19.12 21.60
Tofu, soft 29.24 8.59 20.65
Tofu, silken 27.91 11.13 15.58
Soy infant formula. powder 25.00 7.23 14.75
Tofu, firm 22.70 8.00 12.75
Soy hot dog 15.00 3.40 8.20
Okara 13.51 5.39 6.48
Soy protein concentrate, alcohol extracted 12.47 6.83 5.33
Bacon, meatless 12.10 2.80 6.90
Soy milk 9.65 4.45 6.06
Vegetarian burger 9.30 2.95 5.28
Soy cheese, Mozzarella 7.70 1.10 3.60
Soy cheese, Cheddar 7.15 1.80 2.25
Soy drink 7.01 2.41 4.60

124 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
results in the release of the sugar molecule from the
isoflavone, leaving the isoflavone as an aglycone
(Zubik and Meydani, 2003). Soy isoflavone glyco-
sides include genistin, daidzin, and glycitin, while the
aglycones include genistein, daidzein, and glycitein
(Figure 1). The typical composition for soybeans is
40% diadzin/diadzein, 50% genistin/genestein and
10% glycetin/glycitein (Murphy et al., 1999).
People from countries that consume large
amounts of soy foods are reported to have an im-
proved chronic disease burden compared with coun-
tries consuming very little soy (Boring et al., 1994;
Thom et al., 1992). The average isoflavone intake
from soy food by Asian women ranges from 25 to
50 mg/day (Messina et al., 2006) compared to non-
Asian women who take in less than 2 mg/day (Van
Erp-Baart et al., 2003; de Kleijn et al., 2001). Health
benefits attributed to soy isoflavones include low-
ering blood pressure ( Hooper et al., 2008), preven-
tion of coronary heart disease (Nagata et al., 1998;
Smit et al., 1999), better bone health ( Ho et al., 2001;
Mei et al., 2001; Horiuchi et al., 2000; Tsuchida et al.,
1999) relief of menopausal symptoms (Messina,1999)
and decreased risk of certain types of cancer such as
breast and prostate (Severson et al., 1989; Jacobsen
et al., 1998; Lee et al., 1991; Wakai et al., 1999). Since
isoflavones are capable of exerting estrogen-like ef-
fects they are often referred to as phytoestrogens
(Lampe, 2003). Since isoflavones structurally closely
resemble esterogenic steroids of animals they are
able to bind to both estrogen receptors alpha (ERα)
and beta (ERβ) (Figure 2) (Kuiper et al., 1997; Kuiper
et al., 1998). This review will look at the evidence, pro
and con, regarding the health benefits of isoflavones
derived from soy.
CONSUMPTION OF SOYFOODS
Soybeans and soy products represent one of the
richest and cheapest sources of protein (Codina et
al., 2003). Soybeans have been used as a major part
of the diet in Asian countries and some other parts
of the world for more than 5,000 years. Soy foods
Figure 1. Chemical structures of soybean isoflavones (Naya and Imai, 2013).
Reproduced with permission of Nayga and Imai (2013), available from http://www.intechopen.com/books/
soybean-bio-active-compounds/recent-advances-on-soybean-isoflavone-extraction-and-enzymatic-modifi-
cation-of-soybean-oil

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 125
eaten in the Asian countries are often fermented by
microorganisms; for example miso which is added to
soups and stews in Japan, soy paste used in Korea
and tempeh, with a meat-like texture in Indonesia
(Synder and Kwon, 1987a). Soy sauce is perhaps the
most familiar soy product which is made either by
a long fermentation process or by acid hydrolysis;
this acid hydrolyzed product contains no isoflavones
(Luh, 1995). Soy milk is made by extracting the pro-
teins and lipids in soybeans with boiling water; soy-
milk may then be curdled to prepare tofu, which is
pressed to remove water and can be fried or added
to numerous other dishes (Snyder and Kwon, 1987a).
Soybean-containing foods have become more
popular in the United States, especially after October
1999 when the Food and Drug Administration (FDA)
approved a health claim for soy proteins for reduc-
ing heart disease (FDA, 1999). However, soy foods in
the United States are generally quite different from
the forms of soy consumed in Asia. Soybeans grown
in the United States are utilized mostly as a source
of edible oil. After extraction the defatted soy flour,
which is high in protein, is used in many bread and
cake products, particularly in doughnuts (Snyder and
Kwon, 1987b). Alternatively the soy flour is washed
with water to remove soluble carbohydrates creating
soy protein concentrate, which is even higher in pro-
tein. When the soy flour is extracted with hot, aque-
ous 65% alcohol it forms a different type of soy pro-
tein concentrate which contains no isoflavones. Both
of these soy protein concentrates can be extruded
to form textured soy protein, a meat-like product.
The proteins in soy flour may be solubilized with a
mild alkaline extraction followed by a precipitation
at a low pH to produce soy protein isolate (SPI). This
SPI is widely found in canned foods or is used by
athletes as a source of protein. There are also new
“soy” foods such as soy cheese, soy ice cream and
soy yogurt.
EFFECT OF PROCESSING ON ISOFLA-VONE CONTENT OF FOODS
Processing has a substantial influence on the
amount and form of isoflavones in soyfood products.
During the course of processing, some isoflavones
may be lost and the chemical composition may also
change. Soybeans for oil production have the hulls
removed and the remainder of the beans after the
fat is removed is pressed into flakes and then ground
into soy flour. The isoflavone content of the soy flour
Figure 2. Comparison between isoflavone and estrogen molecule showing similarity in conforma-tion (Setchell and Cassidy, 1999).
Reproduced with permission of American Society for Nutrition

126 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
is about the same as whole soybeans, indicating that
defatting and milling causes little loss or transforma-
tion of isoflavones; soy oils contain only traces of iso-
flavones because of the highly polar nature of the
isoflavones leading to an inability to partition into
the lipophilic oil during soy oil extraction (Coward
et al., 1998; Setchell, 1998). Production of soy pro-
tein concentrate using alcohol washing causes the
loss of most of the isoflavones, whereas a substantial
amount of isoflavones are retained after water wash-
ing (Wang and Murphy, 1996). More isoflavones are
retained in SPI than in soy protein concentrate (Wang
and Murphy, 1996). Fermented soybean products
such as miso and natto are reported to have higher
isoflavone content than in products such as soymilk
and tofu, which is thought to be due to the action of
bacteria during fermentation (Fukutake et al., 1996).
Extruded soy products such as cereals have a lower
amount of isoflavones due primarily to the heat and
loss of moisture during the extrusion (Mahungu et
al., 1999).
ISOFLAVONE METABOLISM
After the soybean is eaten, the glycosidic forms of
the isoflavones undergo hydrolysis due to the action
of the brush border and bacterial β-glucosidases to
remove the sugar moiety; the aglycone form is then
either absorbed or undergoes further metabolism
by intestinal bacteria in the large bowel (Chen et
al., 2003; Setchell et al., 2003). The isoflavone daid-
zein is usually metabolized to dihydrodaidzein or O-
desmethylangolensin (Bowey et al., 2003; Setchell,
1998; Yuan et al., 1995; Zubik and Meydani, 2003). In
a small number of persons daidzein may also be me-
tabolized in the intestine to equol, a metabolite that
has greater estrogenic activity than daidzein (Muth-
yala et al., 2004). Equol exists in 2 stereoisomers,
R or S, which differ significantly from each other in
terms of their binding affinities with estrogen recep-
tor (ER) (Muthyala et al., 2004). The S isomer has a
high binding affinity for both receptors but prefers
ERβ, whereas the R isomer binds weakly and prefers
ERα; however, both isomers have a higher affinity for
both ERs than does the precursor daidzein (Muth-
yala et al., 2004). Human gut microflora metabolize
daidzein to produce only the S isomer (Setchell et
al., 2005). Studies that measured urinary equol ex-
cretion after soy consumption indicated that only
about 33% of individuals from Western populations
metabolize daidzein to equol (Setchell et al., 2002).
The prevalence of equol producers appears to be
higher in Asian populations than in non-Asian (Arai
et al., 2000; Wu et al., 2006; Cassidy et al., 2006) and
appears to be linked to the gastrointestinal normal
flora in these individuals (Setchell et al., 2002).
The role of gut microflora in the production of
equol was elucidated in experiments with germ-
free rats which, when fed daidzein, did not produce
equol; when the rats were inoculated with fecal flora
from equol producers they were able to produce
equol from daidzein (Bowey et al., 2003; Axelson
and Setchell, 1981). A number of bacterial species
capable of converting daidzein to the S isomer of
equol in vitro have been isolated from both food
and human gut flora including, Lactococcus garviae
from Italian cheese (Fortina et al., 2007), 6 strains of
bacteria belonging to the Coriobacteriaceae fam-
ily from tofu brine (Abiru et al., 2013), Eggerthella
spp strain YY791 (Yokoyama and Suzuki, 2008) and
YY7918 (Yokoyama et al., 2011), and Slackia isoflavo-
niconvertens (Schroder et al., 2013) from human gut
flora. Many researchers have proposed that equol
producers may have improved disease risk patterns
as compared with non-producers (Fujioka et al.,
2004; Kurahashi et al., 2008; Lampe, 2009; Magee,
2011; Setchell et al., 2002; Wu et al., 2007). There
is much evidence that suggests that equol produc-
ers have a lower breast cancer risk as compared with
non-producers (Atkinson et al., 2003; Duncan et al.,
2000; Falk et al., 2005; Ursin et al., 1999).
DISEASE PREVENTION ACTIVITIES
Isoflavones and Bone Health
Loss of bone mass, known as osteoporosis, poses
a major human health threat by contributing to bone

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 127
fractures in those afflicted. Worldwide, osteoporosis
causes more than 8.9 million fractures annually, re-
sulting in an osteoporotic fracture every 3 seconds
(Johnell and Kanis, 2006). Osteoporosis is estimated
to affect 200 million women worldwide; approxi-
mately 10% of women aged 60, 20% of women aged
70, 40% of women aged 80 and 67% of women aged
90 are afflicted (Kanis, 2007). One of the means by
which isoflavones promote bone health is hypoth-
esized to be because of their affinity for ERβ since
bone tissue contains large amounts of ERβ (Mes-
sina, 1999). Arjmandi et al. (1998) found that there
was a significant increase in insulin-like growth fac-
tor 1 (IGF-1) mRNA in isoflavone treated groups
compared to control groups; since IGF-1 mRNA
stimulates bone formation, this is another possible
mechanism for the positive role of isoflavone in bone
health. Isoflavones have also been postulated to
prevent osteoporosis because of their similar struc-
ture to ipriflavones, which inhibit bone resorption in
humans (Tsuda et al., 1986; Brandi, 1997).
There is inconsistent data available on the ben-
eficial effect of soy isoflavones on bone density in
human studies due to the small subject number and
short duration of the soy consumption, a larger ef-
fect on bone density was observed in animal models
using higher doses (Arjmandi et al. 1998). From epi-
demiological studies as well as clinical trials, Messina
et al., (2004) showed that Asian women who take in
more soy isoflavones have higher bone mineral den-
sity and have a low rate of hip fracture compared to
non-Asians, and concluded that isoflavones reduce
bone loss in postmenopausal women.
Isoflavones and Cancer Prevention
Cancer affects persons of every socioeconomic
level and every area of the world; cancer accounts for
one in every eight deaths worldwide – more than HIV/
AIDS, tuberculosis, and malaria combined (American
Cancer Society, 2013). An estimated 14.1 million new
cancer cases and 8.2 million cancer-related deaths
occurred in 2012 (International Agency for Research
on Cancer, 2013). The most common causes of can-
cer death were cancers of the lung, liver, and stom-
ach (International Agency for Research on Cancer,
2013). It is estimated that more than two-thirds of hu-
man cancers could be prevented by modification of
lifestyle including dietary modification (Haque et al.,
2010). The U.S. National Cancer Institute has been
actively investigating the anticancer effects of soy-
beans since 1991 (Messina and Barnes, 1991).
Akiyama et al., (1987) demonstrated that genistein
was a specific inhibitor of a tyrosine-specific protein
kinase, an enzyme that is often overexpressed in
cancer cells. Constantinou et al. (1990) found that
genistein suppressed growth and induced differen-
tiation in leukemia cells. Later, genistein was found
to inhibit multiple protein tyrosine kinases relevant
to cancer cell proliferation (Bektic et al., 2005). Ra-
biau et al. (2010) treated human prostate cancer cells
with genistein or daidzein and found that they down
regulated growth factors involved in proliferation of
new blood vessels in tumors. In a study reported in
2011, prostate cancer patients scheduled for radical
prostatectomy were randomly assigned to receive a
placebo or 30 mg genistein daily for 3 to 6 weeks
before surgery. Among the patients who received
genistein, serum prostate specific antigen (PSA) lev-
els decreased by 7.8%, whereas serum PSA levels
increased by 4.4% in patients who received the pla-
cebo (Lazarevic et al., 2011).
Consumption of soy isoflavones is higher in Asian
diets as compared to Western; daily consumption of
soy isoflavones in Japan ranges from 26 to 54 mg,
compared to 0.5 to 3 mg in the United States (Naga-
ta, 2010). Breast cancer incidence increased by more
than 20% between 2008 and 2012 and mortality from
breast cancer increased by 14%; it is the most com-
mon cause of cancer deaths among women and the
most frequently diagnosed cancer in women in 140
of 184 countries (International Agency for Research
on Cancer, 2013). Some epidemiologic evidence
suggests that soy consumption early in life and
through puberty reduces breast cancer risk (Mes-
sina and Hilakivi, 2009) and this has been supported
by animal studies which suggest that soy intake is
protective at specific stages of development but not
at other points (Warri et al., 2008). Lamartiniere et
al., (2000) briefly exposed young rodents to dietary

128 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
supplements of genistein and found that it reduced
mammary cancers indicating early intake of soy iso-
flavones protects against breast cancer. Animal stud-
ies demonstrated that when carcinogenic rodents
were fed with isoflavone-rich soy protein or isolated
isoflavones, mammary carcinogenesis was inhibited
by 25 to 50% (Magee and Rowland, 2004; Messina
and Loprinzi, 2001) indicating that isoflavones do
have an antiesterogenic effect. Pisani et al., (1999)
and Kitamura et al. (2002) reported low risk of breast
cancer in the Asian countries where soy is commonly
consumed as isoflavone might exert an antiestero-
genic effect on breast tissue.
Adults who consumed large amounts of soy as ad-
olescents have been determined to have a lower risk
for breast cancer as compared to adults who did not
consume large amounts of soy in adolescence (Shu
et al., 2001). Wu et al. (1996) found that among Asian
Americans, tofu consumption protected against both
pre- and postmenopausal breast cancer. Another
study reported by Chinese epidemiologists wherein
adolescents had high soy consumption which re-
sulted in a 50% reduction in adult breast cancer risk
whereas adult intake did not impact these findings
(Shu et al., 2001). Similarly, a U.S. case-control study
involving Asian Americans reported that high soy
consumption during both adolescence and adult-
hood was associated with a one-third reduction in
risk whereas high adult intake alone was not protec-
tive (Wu et al., 2002). Both dosage and timing of
exposure to soy isoflavones appear relevant to their
potential chemopreventive effect. Two meta-analy-
ses (Dong and Qin, 2011; Wu et al., 2008) found soy
intake to be significantly associated with reduced
risk of breast cancer in Asian but not Western human
populations, which may be explained by both higher
soy intake among Asians and their tendency to con-
sume soy from an early age.
Several studies have shown mixed results regard-
ing the effect of isoflavones supplements on the
proliferation of breast cells in breast cancer patients.
In 2007, a Japanese collaborative cohort study sug-
gested that consumption of soy foods such as tofu,
boiled beans, and miso soup has no protective ef-
fects against breast cancer (Nishio et al., 2007). In
multiple trials no effects on breast proliferation or
mammographic density were observed for isofla-
vones and considerable epidemiologic data shows
either no effect or only a modest protective role of
soy/isoflavone intake on breast cancer risk (Messina
and Wood, 2008). Studies conducted by Caan et al.,
(2011) and Guha et al., (2009) reported that soy con-
sumption had no adverse effects on breast cancer
survivors. Furthermore, they suggest that soy con-
sumption at levels comparable to those among Asian
populations does not detract from the benefits of
tamoxifen therapy, and may even offer some protec-
tion against recurrence and cancer-related death.
The American Cancer Society stated that con-
sumption of soy foods would not decrease survival
nor increase recurrence of cancers, but there was
not enough evidence to make a statement about
isoflavone supplements (Rock et al., 2012). However,
in another study conducted by a U.S.- Chinese re-
search team, researchers monitored and measured
intake of soy isoflavones over the course of seven
years; they determined that soy isoflavones signifi-
cantly reduced risk of cancer recurrence in patients
who consumed at least 10 mg of isoflavones (ap-
proximately 3 g of soy protein) per day (Nechuta et
al., 2012). Bloedon et al. (2002) and Allred et al. (2001)
found that soy protein and isoflavones stimulated
the growth of mammary tumors in ovariectomized
mice implanted with estrogen-sensitive breast can-
cer cells. In contrast Zhou et al., (2004) demonstrated
that isoflavones could inhibit the growth of tumors in
mice when intact ovaries were implanted with these
same types of cells.
Messina et al., (2006) suggested that a daily dose
of 120 mg isoflavones may be useful in prostate can-
cer prevention, but recommended consumption of
soy foods rather than isolated isoflavones supple-
ments, because other soy components such as soy
protein, fiber and saponins may offer additional
health benefits. Nagata et al., (2007) studied the ef-
fect of dietary isoflavone against prostate cancer in
Japanese males and found that inclusion of dietary
isoflavone might be an effective dietary protective
factor against prostate cancer in Japanese men. In
this study male subjects in the highest category of

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 129
isoflavones intake (greater than 90 mg/day) exhib-
ited a 58% lower prostate cancer risk than male sub-
jects in the lowest category (less than 30 mg/day). In
one study, early-stage prostate cancer patients were
randomly assigned to receive a soy protein supple-
ment (60 mg/day isoflavones) or a placebo daily for
12 weeks. Patients who received the soy protein sup-
plement exhibited greater decreases in total serum
PSA and free testosterone than did patients who re-
ceived the placebo, but these differences were not
statistically significant (Kumar et al., 2004). In two
small studies prostate cancer patients were fed soy
isoflavone and in these studies there appeared to be
a decrease in the rate of the rising serum PSA con-
centration associated with prostate tumor growth
(Fischer et al., 2004; Hussain et al., 2003). A trial of
soy milk supplementation (141 mg/day isoflavones)
in men with PSA recurrent prostate cancer found
that PSA levels increased by an average of 20%
over a 12-month period compared to a 56% yearly
increase prior to the study (Pendleton et al., 2008).
Hamilton-Reeves and coworkers at the University of
Minnesota, (2007) examined the potential protective
effect of soy protein isolate, with low and high lev-
els of isoflavones, on prostate cancer risk in men at
high risk for developing the advanced form of pros-
tate cancer. They found that soy protein isolate con-
sumption suppressed androgen receptor expression
in the prostate and could be beneficial in prevent-
ing prostate cancer. Kumar et al., (2007) treated 53
prostate cancer patients with 80 mg purified isofla-
vones or a placebo for 12 weeks. Although plasma
isoflavones increased with no observed clinical tox-
icity, there was no modulation of serum sex hormone
binding globulin, total estradiol, or testosterone in
the isoflavone-treated group compared to placebo.
The study establishes the need to explore other po-
tential mechanisms by which prolonged and consis-
tent purified isoflavone consumption may modulate
prostate cancer risk.
A meta-analysis of eight studies performed by
Yan and Spitznagel (2009) found that isoflavone con-
sumption was associated with a reduction in risk of
prostate cancer, but the association was not statisti-
cally significant. Similar results were reported by Mi-
yanaga et al., (2012) wherein isoflavones had no influ-
ence on the level of PSA, but biopsies showed that
isoflavone intake reduced the incidence of prostate
cancer, but that this difference was not statistically
significant. Several epidemiologic studies have also
shown no association between high consumption of
fermented soy foods and prostate cancer (Hwang et
al., 2009). Isoflavones are purported to slow prostate
cancer growth and cause cancer cells to die (Fot-
sis et al., 1993). Supplementation with soy protein
or soy isoflavone decreased the markers of cancer
development and progression in prostate cells in-
cluding PSA, testosterone, and androgen receptor
in patients with prostate cancer (Kumar et al., 2004;
Dalais et al., 2004) or in men at high risk for develop-
ing advanced prostate cancer (Hamilton-Reeves et
al., 2007).
Isoflavones and Cardiovascular Disease
In 2010, cardiovascular disease (CVD) was the lead-
ing cause of death responsible for 597,689 deaths in
the U.S. (CDC, 2013). An estimated 30% of all global
deaths in 2008 were from CVD (WHO, 2011a); of these
deaths, an estimated 7.3 million were due to coro-
nary heart disease and 6.2 million were due to stroke
(WHO, 2011b). In order to reduce coronary heart dis-
ease it is recommended that saturated fat should be
replaced with polyunsaturated fatty acids. Soy foods
are ideal for this replacement since they contain the
omega-6 polyunsaturated fatty acid (PUFA) linoleic
acid, which comprises about 55 percent of the total
fat in soybeans and which reduces blood cholesterol
levels (Slavin et al., 2009; Jenkins et al., 2002). Jen-
kins et al. (2011) recommended a diet supplemented
with cholesterol-lowering foods including soyfoods
like soymilk and soy meat alternatives, oats, nuts and
plant sterols for adults with high cholesterol. This diet
lowered low density lipoprotein (LDL) cholesterol by
13.8% compared with a decrease of only 3% in those
that followed a standard low saturated fat diet. Other
epidemiological studies have also suggested that
Asian populations consuming large amounts of soy
have lower rates of cardiovascular disease than West-
ern populations (Zhang et al. 2003).

130 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
Research suggests that soy protects against ath-
erosclerosis by lowering cholesterol, or by increas-
ing blood levels of nitric oxide which helps in blood
vessel dilation, inhibits oxidative damage caused by
cholesterol and prevents the adhesion of white cells
to the vascular wall. (Cena and Steinberg, 2011). A
combination of soy protein and isoflavones appear
to exhibit the strongest hypocholesterolemic effects
compared to isolated soy protein or soy isoflavones
alone (Mortensen et al., 2009). Research studies
suggest that soy protein decreases postprandial
triglyceride levels, which is increasingly viewed as
important for reducing coronary heart disease risk
(Zhan and Ho, 2005). Whole soy can promote a 3%
to 5% reduction in blood cholesterol (Lichtenstein et
al. 2006; Zhan and Ho 2005). Henrotin et al., (2003)
concluded that soy protein led to increased blood
levels of L-arginine (the amino acid that the human
body uses to produce nitric oxide) and nitric oxide
metabolites. Isoflavones increase endothelial nitric
oxide production, enhancing vasodilation and im-
proving blood flow (Taku et al., 2007; Zhan and Ho,
2005). Specific findings include beneficial effects on
lipids and lipoproteins, with a decline in total cho-
lesterol (9%), LDL cholesterol (13%), and triglycer-
ides (11%) and an increase in high density lipopro-
tein (HDL) cholesterol (2.4%) (Anderson et al., 1995).
Isoflavones found in soybean and soy foods provide
cardiovascular health benefits by neutralizing free
radicals that cause oxidative damage to cells thus
improving arterial elasticity, a vascular function that
normally decreases with age, helping to reduce LDL
cholesterol levels (Steinberg et al. 2003). Otherwise,
these free radicals within blood vessels can oxidize
circulating LDL cholesterol, starting a cascade of in-
flammatory events that ultimately increases the risk
of developing heart disease. In a study conducted
by Candy (1996) 61 middle-aged men that had been
diagnosed as having a high risk of developing cor-
onary disease were asked to consume soy protein
(20 g) and soy isoflavone (80 mg) for five weeks;
those consuming soy showed significant reductions
in both diastolic and systolic blood pressure com-
pared to those who were given a placebo diet con-
taining olive oil. In the mid-1990s, a meta-analysis of
29 clinical trials found that compared to animal pro-
tein, soy protein significantly reduced blood levels
of several lipids (total cholesterol, LDL cholesterol
and triglycerides) (Anderson et al. 1995). There is ad-
ditional data suggesting isoflavones have indepen-
dent coronary benefits. In several studies isoflavones
have been shown to enhance endothelial function
(Walker et al., 2001; Squadrito et al., 2002, 2003) and
systemic arterial elasticity (Nestel et al., 1997, 1999);
both of these measures are considered to be indi-
cators of coronary health (Bonetti et. al., 2003; Her-
rington et al., 2004).
In several research studies it has been shown that
on average, soy protein lowers LDL cholesterol ap-
proximately 4% (Sacks et al., 2006; Zhan and Ho,
2005). Each one percent reduction in cholesterol low-
ers coronary heart disease risk at least 2% (Law et al.,
1994). A meta-analysis conducted by Weggemans
and Trautwein (2003) found that soy protein slightly
raised HDL cholesterol levels leading tham to con-
clude that as a result of the changes in lipid levels,
soy could reduce heart disease risk by as much as 20
percent. Furthermore, there is evidence to suggest
that soyfoods may decrease blood pressure (Rivas et
al., 2000) and increase LDL cholesterol particle size
(Desroches et al., 2004). Dong et al., (2011) analyzed
27 clinical (human intervention) studies and found
that on average, soy lowered blood pressure about
2 ½ points. Li et al. (2010) studied the effect of oral
isoflavone supplementation on vascular endothelial
function in postmenopausal women and concluded
that isoflavone helps to improve endothelial func-
tion.
Zhan and Ho (2005) reported that the inclusion of
soy in the diet can decrease blood levels of LDL cho-
lesterol. A meta-analysis of 7 studies found that soy
protein that contained enriched isoflavones, and in
comparison with animal protein without isoflavones,
were associated with a significant decrease in serum
total cholesterol (0.32 mmol/L or 5.69%) in the hyper-
cholesterolemic subcategory and LDL cholesterol
(0.18 mmol/L or 4.98%) in the total human popula-
tion. It was also reported in this study that a signifi-
cant increase in serum HDL cholesterol (0.04 mmol/L
or 3.00%) occurred in the total population. Similarly

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 131
a study by Steinberg et al., (2003) demonstrated that
consuming intact soy protein and isoflavone may im-
prove vascular function better than the consumption
of either component alone. Gardner et al., (2007)
compared cholesterol levels between those who
drank soymilk and those who drank 1% dairy milk as
part of an overall diet containing moderate fat (35%
of calories). In only four weeks, the soymilk group
exhibited a 5% reduction in LDL cholesterol, a sta-
tistically significant advantage over those who con-
sumed dairy milk.
ROLE OF ISOFLAVONES IN REDUCING MENOPAUSAL SYMPTOMS AND HOT FLASHES
Hot flashes are the primary reason that women
seek medical attention for menopausal symptoms
(Tice et al., 2003). A low incidence of hot flashes and
other menopausal symptoms in Japanese women
is believed to be due to the estrogen-like effects of
soy isoflavones (Kuiper et al., 1998; Lock, 1992; 1994).
Women fed with soy flour (45 g) daily demonstrated
a reduction (40%) in menopause symptoms (Allaoua
et al., 2005). Conversely, other studies have reported
no beneficial effect of soy intake on menopausal
symptoms. A meta-analysis of 25 trials involving
2,348 participants published between 1966 and 2004
concluded that soy phytoestrogens did not improve
hot flashes or other menopausal symptoms (Krebs
et al., 2004). The isoflavone daidzein has been re-
ported to reduce hot flashes in menopausal women.
The chemical structure of daidzein is very similar to
the human body’s own estrogen. A study was con-
ducted by Khaodhiar et al., (2008) on 190 women
ranging in age from 38 to 60 years in various stages
of menopause, who had 4 to 14 hot flashes daily. The
women were given either one or two concentrations
of diadzein rich isoflavone-aglycone. The number of
hot flashes in the diadzein rich isoflavone groups was
reduced by 52% and 51% at the end of 12 weeks,
while the placebo group experienced a 39% reduc-
tion. A few studies using higher doses of isoflavone
(50 to 80 mg/day), enrolling women with more vaso-
motor symptoms at baseline (4 to 7 symptoms/day)
and with larger sample sizes, have exhibited mildly
beneficial effects on self-reported frequency and se-
verity of vasomotor symptoms (Albertazzi et al. 1998,
Washburn et al. 1999). Han et al., (2002) reported a
26% decrease in hot flash frequency in a group who
consumed 100 mg/day of isoflavone as compared to
a group who receive a placebo. Isoflavones from soy
have also received attention as a possible alterna-
tive to conventional hormone replacement therapy
(HRT) (Brandi, 1999; Eden, 2001; Elkind-Hirsch, 2001;
Glazier and Bowman, 2001; Vincent and Fitzpatrick,
2000). Since there is a chemical similarity to the fe-
male sex hormone estrogen, isoflavones have been
used in studies for relief of menopausal symptoms
(Adlercreutz et al., 1992; North and Sharples, 2001).
A ROLE FOR ISOFLAVONES IN OBESITY
Obesity is a state of excessive fat accumulation
in the body, especially in abdominal adipose tissue,
and is closely linked to metabolic disorders, which
include diabetes, cardiovascular disease, nonal-
coholic fatty liver disease, dyslipidemia, and other
health problems (Després et al., 2008; Lois et al.,
2008;). Nearly 20% of women aged 55 to 65 years
suffer an increase in glucose tolerance and insulin
resistance, thought to be due to estrogen defciency
(Gaspard et al., 1995; Tufano et al., 2004). The effect
of soy isoflavone supplementation on postmeno-
pausal women has yielded mixed responses in sev-
eral clinical trials due to differences in dose, duration
of isoflavone supplementation, body weight, physi-
cal status of individual, and variability of experimen-
tal designs (Zhang et al., 2013). Some researchers
have suggested a reduction in body weight (Sites et
al., 2007; Gardner et al., 2001), fasting blood glucose
(Villa et al., 2009; Crisafulli et al., 2005), blood fat lev-
els (Reynolds et al, 2006) and insulin level (Villa et al.,
2009; Crisafulli et al., 2005) as possible causes. How-
ever, other researchers did not reach this same con-
clusion (Charles et al., 2009; Khaodhiar et al., 2008).

132 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
EFFECT OF ISOFLAVONES ON THYROID FUNCTION
Concerns have been expressed that soy iso-
flavone intake adversely affects thyroid function
(Messina and Redmond, 2006). The thyroid gland
releases two primary hormones T4 and T3 (thyrox-
ine and trio-iodothyronine, respectively) in a ratio
of roughly 80:20 in response to the signal sent by
thyroid stimulating hormone (TSH). Several studies
have reported the interaction of soy isoflavones with
thyroid function. Isoflavones lead to immune dys-
function by causing potent stimulation of T cell and
B cell mediated immunity due to induced structural
changes in thyroid peroxidase (Chen and Rogan,
2004). A study in premenopausal women (Duncan et
al., 1999) demonstrated a decline in free T3 levels
with a high isoflavone diet. Another study conduct-
ed with post-menopausal women demonstrated a
rise in T4 with 56 mg isoflavones/day and a rise in
T3 and TSH with 90 mg isoflavones/day at 6 months
compared with controls, but these alterations were
considered clinically non-significant (Persky et al.,
2002). Several other studies revealed non-significant
changes in thyroid profile with isoflavones in meno-
pausal women (Borchers et al., 2008; Duncan et al.,
1999; Teas et al., 2007). The precise reason for such
reported variations is unclear although differences
in doses, dosage forms, isoflavone composition or
the duration of treatment may all be considered im-
portant factors. Milerová et. al., (2006) reported on a
study that looked at thyroid hormones and thyroid
autoantibodies, along with blood levels of daidzein
and genistein. The study focused on children with-
out overt thyroid disease, who were not iodine de-
ficient. They found a significant positive association
of genistein with thyroglobulin autoantibodies and a
negative correlation with thyroid volume. They con-
cluded that even small differences in soy phytoes-
trogen intake may influence thyroid function, which
could be important when iodine intake is insufficient.
Studies by Huang et al., (2005) and Xiao et al.,
(2004) have shown that isoflavones suppress the
binding ability of hepatic thyroid hormone recep-
tor to the thyroid hormone response element of the
target genes. Similarly, in vitro studies by Divi et al.,
(1996; 1997) have shown that isoflavones inhibit thy-
roid peroxidase. Genistein and daidzein block thy-
roid peroxidase-catalyzed tyrosine iodination by act-
ing as alternate substrates (Divi et al., 1997). Chang
and Doerge (2000) reported that consumption of soy
could cause goiter only in animals or humans con-
suming diets marginally adequate in iodine or who
were predisposed to develop goiter and in most
cases dietary supplementation with adequate iodine
can reverse the disorders (Schone et al., 1990). How-
ever, study by Poirier et al., (1999) reported that even
the feeding of genistein to rats, which as an animal
species are very sensitive to goitrogenic agents,
does not disrupt normal thyroid functioning.
CONCLUSIONS
Consumers in the U. S. have become aware of
the potential health benefits of soy, and as a result
consumption in the U. S. has increased although it
still remains far below that in Asia. There is a great
deal of evidence to support the beneficial effects
of soy isoflavones in the prevention of bone loss in
postmenopausal women. Although isoflavones have
been demonstrated to positively impact the bio-
markers of prostate cancer, their potential benefits
have not been substantiated in clinical trials. Benefi-
cial effects of isoflavones for relieving menopause
symptoms and prevention of breast cancer have not
been proven and the antithyroid actions of soyfoods
appear to be consistent in both animals and hu-
mans. The inconsistency of results from animal and
human studies may be partially due to variation in
the bioavailability of the isoflavones. Varying levels
and duration of isoflavone consumption have also
been shown to be of importance in whether soy-
foods have a beneficial role. In addition, the role of
the gut microflora may be particularly important in
the production of equol from daidzein, since equol
may actually confer more health benefits than daid-
zein. Much more research needs to be carried out in
this area, in order to understand how soy can have
health benefits in the broader population.

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 133
ACKNOWLEDGEMENTS
The writing of this review was supported in part by
a grant from the Arkansas Soybean Board.
REFERENCES
Abiru, Y., T. Ueno, and S. Uchiyama. 2013. Isolation
and characterization of novel S-equol–producing
bacteria from brines of stinky tofu, a traditional fer-
mented soy food in Taiwan. Int. J. Food Sci. Nutr.
64:936–943.
Adlercreutz, H., E. Hamalainen, S. Gorbach, and
B. Goldin 1992. Dietary phtyoestrogens and the
menopause in Japan. Lancet. 339:1233-1233.
Akiyama, T., J. Ishida, S. Nakagawa, H. Ogawara, S.
Watanabe, N. Itoh, M. Shibuya, and Y. Fukami 1987.
Genistein, a specific inhibitor of tyrosinespecific
protein kinases. J. Biol. Chem. 262:5592–5595.
Albertazzi, P., F. Pansini, G. Bonaccorsi, L. Zanotti, E.
Forini, and D. De Aloysia, 1998. The effect of di-
etary soy supplementation on hot flushes. Obstet.
Gynecol. 91:6-11.
Allaoua, A., I. B. Joyce, and B. Denis 2005. Soybean
isoflavones efficacy of extraction conditions and
effect of food type on extractability. Food Res. Int.
38:1199-1204.
Allred, C. D., K. F. Allred, Y. H. Ju, S. M. Virant and
W. G. Helferich 2001. Soy diets containing varying
amounts of genistein stimulate growth of oestro-
gen-dependent (MCF-7) tumors in a dose-depen-
dent manner. Cancer Res. 61:5045–5050.
American Cancer Society. 2013. Cancer facts and
figures, 2013. http://www.cancer.org/acs/groups/
content/@epidemiologysurveilance/documents/
document/acspc-036845.pdf Accessed 11 March
2014.
Anderson, J. W., B. M. Johnstone, and M. E. Cook-
Newell 1995. Meta-analysis of the effects of soy
protein intake on serum lipids. N. Engl. J. Med.
333:276–282.
Arai, Y., S. Watanabe, M. Kimira, K. Shimoi, R. Mo-
chizuki, and N. Kinae. 2000. Dietary intakes of
flavonols, flavones and isoflavones by Japanese
women and the inverse correlation between quer-
cetin intake and plasma LDL cholesterol concen-
tration. J. Nutr.130:2243–2250.
Arjmandi, B. H., R. Birnbaum, N. V. Goyal, M. J.
Getlinger, S. Juma, L. Alekel, C. M. Hasler, M. L.
Drum, B. W. Hollis, and S. C. Kukreja. 1998. Bone-
sparing effect of soy protein in ovarian hormone-
deficient rats is related to its isoflavone content.
Am. J. Clin. Nutr. 68:1364S-1368S.
Atkinson, C., H.E. Skor, F.E. Dawn, D. Scholes, C.
Chen, K. Wahala, S. M. Schwartz, and J. W. Lampe
2003. Urinary equol excretion in relation to 2-hy-
droxyestrone and 16alpha-hydroxyestrone con-
centrations: an observational study of young to
middle-aged women. J. Steroid Biochem. Mol.
Biol. 86:71–77.
Axelson, M. and K.D. Setchell. 1981. The excretion
of lignans in rats—evidence for an intestinal bacte-
rial source for this new group of compounds. FEBS
Lett. 123:337–342.
Bektic, J., R. Guggenberger, I. E. Eder, A. E. Pelzer, A.
P. Berger, G. Bartsch, and H. Klocker 2005. Molecu-
lar effects of the isoflavonoid genistein in prostate
cancer. Clin. Prostate Cancer 4:124-129.
Bloedon, L. T., A. R. Jeffcoat, W. Lopaczynski W, M.
J. Schell, T. M. Black, K. J. Dix, B. F. Thomas, C. Al-
bright, M. G. Busby, J. A. Crowell, and S. H. Zeisel
2002. Safety and pharmacokinetics of purified soy
isoflavones:single-dose administration to post-
menopausal women. Am. J. Clin. Nutr. 76:1126-
1137.
Bonetti, P. O., L. O. Lerman, and A. Lerman 2003. En-
dothelial dysfunction: a marker of atherosclerotic
risk. Arterioscler. Thromb. Vasc. Biol. 23:168-175.
Borchers, T. R., B. Chew, J. S. Park, M. McGuire, L.
Fournier, and K. Beerman 2008. Effects of dietary
and supplemental forms of isoflavones on thyroid
function in healthy postmenopausal women. Top.
Clin. Nutr. 23:13–22.
Boring, C. C., T. S. Squires, T. Tong, and S. Mont-
gomery 1994. Cancer statistics 1994. CA - Cancer
J. Clin. 44:7-26.
Bowey, E., H. Adlercreutz, and I. Rowland. 2003.
Metabolism of isoflavones and lignans by the gut
microflora: a study in germ-free and human flora

134 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
associated rats. Food Chem. Toxicol. 41: 631–636
Brandi, M. L. 1999. Phytoestrogens and menopause.
Environ. Toxicol. Pharmacol. 7:213-216.
Caan, B. J., L. Natarajan, B. Parker, E. B. Gold, C.
Thomson, V. Newman, C. L. Rock, M. Pu, W. Al-De-
laimy, and J. P. Pierce 2011. Soy food consumption
and breast cancer prognosis. Cancer Epidemiol.
Biomarkers Prev. 20:854-858.
Candy, O. 1996. The role of soy in medicine, Part 1.
Palais des Congress, 15-17 Sept., 1996.
Cassidy, A., J. E. Brown, A. Hawdon, M. S. Faughnan,
L. J. King J. Millward J, L. Zimmer-Nechemias, B.
Wolfe, K. D. Setchell. 2006. Factors affecting the
bioavailability of soy isoflavones in humans after
ingestion of physiologically relevant levels from
different soy foods. J. Nutr. 136:45–51.
CDC, Centers for Disease Control and Prevention.
2013. Leading causes of death. http://www.cdc.
gov/nchs/fastats/lcod.htm Accessed 11 March
2014.
Cena, E. R. and F. M. Steinberg. 2011. Soy may help
protect against cardiovascular disease. California
Agric. 65:118-123.
Chang, H. C., and D. R. Doerge 2000. Dietary genis-
tein inactivates rat thyroid peroxidase in vivo with-
out an apparent hypothyroid effect. Toxicol. Appl.
Pharmacol. 168:244–252.
Charles, C., J. Yuskavage, O. Carlson, M. John, A.
S. Tagalicud, M. Maggio, D. C. Muller, J. Egan,
and S. Basaria 2009. Effects of high-dose isofla-
vones on metabolic and inflammatory markers
in healthy postmenopausal women. Menopause.
16:395–400.
Chen, A., and W. J. Rogan 2004. Isoflavones in soy
infant formula: a review of evidence for endo-
crine and other activity in infants. Ann. Rev. Nutr.
24:33–54.
Chen, J., H. Lin, and M. Hu. 2003. Metabolism of
flavonoids via enteric recycling: Role of intestinal
disposition. J. Pharmacol. Exp. Ther. 304:1228-
1235.
Codina, R., Ardusso, L., Lockey, R. F., Crisci, C. and
Medina I. 2003. Allergenicity of varieties of soy-
bean. Allergy 58:1293-1298.
Constantinou, A., K. Kiguchi, and E. Huberman 1990.
Induction of differentiation and DNA strand break-age in human HL-60 and K-562 leukemia cells by genistein. Cancer Res. 50:2618–2624.
Coward, L., M. Smith, M. Kirk, and S. Barnes. 1998. Chemical modification of isoflavones in soy foods during cooking and processing. Am. J. Clin. Nutr. 68(suppl):1486S–1491S.
Crisafulli, A., D. Altavilla, H. Marini, A. Bitto, D. Cuci-notta, N. Frisina, F. Corrado, R. D’Anna, G. Squad-rito, E. B. Adamo, R. Marini, A. Romeo, F. Cancel-lieri, M. Buemi, and F. Squadrito 2005. Effects of the phytoestrogen genistein on cardiovascular risk factors in postmenopausal women. Menopause. 12:186–192.
Dalais, F. S., A. Meliala, N. Wattanapenpaiboon, M. Frydenberg, D. A. Suter, W. K. Thomson, and M. L. Wahlqvist 2004. Effects of a diet rich in phy-toestrogens on prostate-specific antigen and sex hormones in men diagnosed with prostate cancer. Urology. 64:510–515.
de Kleijn, M. J., Y. T. van der Schouw, P. W. Wilson, H. Adlercreutz, W. Mazur, D. E. Grobbee and P. F. Jacques 2001. Intake of dietary phytoestrogens is low in postmenopausal women in the United States: the Framingham study. J. Nutr.131:1826–1832.
Deavours, B.E. and R. A. Dixon. 2005. Metabolic en-gineering of isoflavonoid biosynthesis in alfalfa. Plant Physiol. 138:2245–2259.
Després, J. P., I. Lemieux, J. Bergeron, P. Pibarot, P. Mathieu, E. Larose, J. Rodés-Cabau, O. F. Ber-trand, and P. Poirier 2008. Abdominal obesity and the metabolic syndrome: contribution to global cardiometabolic risk. Arterioscler. Thromb. Vasc. Biol. 28:1039-1049.
Desroches, S., J. F. Mauger, L. M. Ausman, A. H. Lich-tenstein, and B. Lamarche 2004. Soy protein favor-ably affects LDL size independently of isoflavones in hypercholesterolemic men and women. J. Nutr. 134:574-579.
Divi, R. L., H. C. Chang and D. R. Doerge 1997. Anti-thyroid isoflavones from soybean: isolation, char-acterization , and mechanisms of action. Biochem. Pharmacol. 54:1087-1096.
Divi, R. L. and D. R. Doerge 1996 .Inhibition of thy-roid peroxidase by dietary flavonoids. Chem. Res.

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 135
Toxicol. 96:16-23.Dong, J. Y., X. Tong, Z. W. Wu, P. C. Xun, K. He, and L.
Q. Qin 2011. Effect of soya protein on blood pres-sure: a meta-analysis of randomised controlled tri-als. Br. J. Nutr. 106:317-326.
Dong, J. Y. and L. Q. Qin 2011. Soy isoflavones con-sumption and risk of breast cancer incidence or recurrence: a meta-analysis of prospective studies. Breast Cancer Res. Treat. 125:315-323.
Duncan, A. M., B. E. Merz, X. Xu, T. C. Nagel, W. R. Phipps, and M. S. Kurzer 1999. Soy isoflavones exert modest hormonal effects in premenopausal women. J. Clin. Endocrinol. Metab. 84:192–197.
Duncan, A.M., B.E. Merz-Demlow, X. Xu, W.R. Phipps, and M.S. Kurzer 2000. Premenopausal equol ex-cretors show plasma hormone profiles associated with lowered risk of breast cancer. Cancer Epide-miol. Biomarker. Prev. 9:581–586.
Eden, J. A. 2001. Managing the menopause: phyto-oestrogens or hormone replacement therapy? Ann. Med. 33:4-6.
Elkind-Hirsch, K. 2001. Effect of dietary phytoestro-gens on hot flushes: Can soy-based proteins sub-stitute for traditional estrogen replacement thera-py? Menopause 8:154-156.
Falk, R.T., T.R. Fears, X. Xu, R.N. Hoover, M.C. Pike,
A.H. Wu, A. M. Nomura, L. N. Kolonel, D. W. West,
D. W. Sepkovic, H. L. Bradlow and R. G. Ziegler
2005. Urinary estrogen metabolites and their ratio
among Asian American women. Cancer Epidemi-
ol. Biomarkers Prev. 14:221–226.
FDA, Food and Drug Administration. 1999. Food
labeling: Health claims; soy protein and coronary
heart disease. Fed Regist. 64(206):57700-57733.
Fischer, L., C. Mahoney, A. R. Jeffcoat, M. A. Koch,
B. E. Thomas, J. L. Valentine, T. Fortina, M.G., G.
Ricci, R. Foschino, C. Picozzi, P. Dolci, G. Zeppa, L.
Cocolin, and P. L. Manachini 2007. Phenotypic typ-
ing, technological properties and safety aspects of
Lactococcus garvieae strains from dairy environ-
ments. J. Appl. Microbiol. 103:445–453.
Fotsis, T., M. Pepper, H. Adlercreutz, G. Fleischmann,
T. Hase, R. Montesano, and L. Schweigerer (1993).
Genistein, a dietary –derived inhibitor of in vitro
angiogenesis. Proc. Natl. Acad. Sci. 90:2690-2694.
Fujioka, M., M. Uehara, J. Wu, H. Adlercreutz, K. Su-
zuki, K. Kanazawa, K. Takeda, K. Yamada, and Y.
Ishimi 2004. Equol, a metabolite of daidzein, in-
hibits bone loss in ovariectomized mice. J. Nutr.
134:2623–2627.
Fukutake, M., M. Takahashi, K. Ishida, H. Kawamura,
T. Sugimura, and K. Wakabayashi. 1996. Quantifi-
cation of genistein and genistin in soybeans and
soybean products. Food Chem. Toxicol. 34:457–
461.
Gardner, C. D., M. Messina, A. Kiazand, J. L. Morris,
and A. A. Franke 2007. Effect of two types of soy
milk and dairy milk on plasma lipids in hypercho-
lesterolemic adults: a randomized trial. J. Am. Coll.
Nutr. 26:669-677.
Gardner, C. D., K. A. Newell, R. Cherin, and W. L.
Haskell 2001. The effect of soy protein with or with-
out isoflavones relative to milk protein on plasma
lipids in hypercholesterolemic postmenopausal
women. Am. J. Clin. Nutr. 73:728–735.
Gaspard, U. J., J. M. Gottal, and F. A. van den Brule
1995. Postmenopausal changes of lipid and glu-
cose metabolism: a review of their main aspects.
Maturitas. 21:171–178.
Glazier, M.G. and M. A. Bowman 2001. A review of
the evidence for the use of phytoestrogens as a
replacement for traditional estrogen replacement
therapy. Arch. Intern. Med. 161:1161-1172.
Guha, N., M. L. Kwan, Q. P. Quesenberry Jr, E. K.
Weltzien, A. L. Castillo, and B. J. Caan 2009. Soy
isoflavones and risk of cancer recurrence in a co-
hort of breast cancer survivors: The Life after Can-
cer Epidemiology study. Breast Cancer Res. Treat.
118:395-405.
Hamilton-Reeves, J. M., S. A. Rebello, W. Thomas,
J. W. Slaton, M. S. Kurzer 2007. Isoflavone-rich
soy protein isolate suppresses androgen recep-
tor expression without altering estrogen recep-
tor-beta expression or serum hormonal profiles
in men at high risk of prostate cancer. J. Nutr.
137:1769–1775.
Han, K. K., J. M. Soares Jr, M. A. Haidar, G. R. de
Lima, and E. C. Baracat 2002. Benefits of soy isofla-
vone therapeutic regimen on menopausal symp-
toms. Obstet. Gynecol. 99:389–394.

136 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
Haque, N., U. Salma, T. R. Nurunnabi, A. K. Haque,
I. J. Mukti, S. Pervin, and R. Nahar R. 2010. Life-
style related causes of cancer and chemopre-
vention through phytonutrients. Pak. J. Biol. Sci.
13:916-26.
Henrotin, Y. E., C. Sanchez, M. A. Deberg, N. Pic-
cardi, G. B. Guillou, P. Msika, and J. Y. Reginster
2003. Avocado/soybean unsaponifiables in-
crease aggrecan synthesis and reduce catabolic
and proinflammatory mediator production by hu-
man osteoarthritic chondrocytes. J. Rheumatol.
8:1825-1834.
Herrington, D. M., W. V. Brown, L. Mosca, W. Da-
vis, B. Eggleston, W. G. Hundley, and J. Raines
2004. Relationship between arterial stiffness and
subclinical aortic atherosclerosis. Circulation
110:432-437.
Ho, S. C., S. G. Chan, Q. Yi, E. Wong, and P. C.
Leung 2001. Soy intake and the maintenance of
peak bone mass in Hong Kong Chinese women.
J. Bone Miner. Res. 16:1363-1369.
Hooper, L., P.A. Kroon, E.B. Rimm, J. S. Cohn, I. Har-
vey, K. A. Le Cornu, J. J. Ryder, W. L. Hall, and A.
Cassidy 2008. Flavonoids, flavonoid-rich foods,
and cardiovascular risk: a meta-analysis of random-
ized controlled trials. Am. J. Clin. Nutr. 88:38–50.
Horiuchi, T., T. Onouchi, M. Takahashi, H. Ito, and
H. Orimo 2000. Effect of soy protein on bone me-
tabolism in postmenopausal Japanese women.
Osteoporos. Int. 11:721-724.
Huang, W., C. Wood, M. R. L’Abbé, S. Gilani, K. A.
Cockell and C. W., Xiao 2005. Soy protein isolate
increases hepatic thyroid hormone receptor con-
tent and inhibits its binding to the target genes in
rats. J. Nutr. 135:1631–1635.
Hussain, M., M. Banerjee, F. H. Sarkar, Z. Djuric, M. N.
Pollak, D. Doerge, J. Fontana, S. Chinni, J. Davis,
J. Forman, D. P. Wood & O. Kucuk 2003. Soy iso-
flavones in the treatment of prostate cancer. Nutr.
Cancer 47:111-117.
Hwang, Y. W., S. Y. Kim, S. H. Jee, Y. N. Kim, and C.
M. Nam 2009. Soy food consumption and risk of
prostate cancer: a meta-analysis of observational
studies. Nutr. Cancer 61:598-606.
International Agency for Research on Cancer. 2013.
Latest world cancer statistics. http://www.iarc.fr/
en/media-centre/pr/2013/pdfs/pr223_E.pdf Ac-
cessed 11 March 2014.
Jacobsen, B. K., S. F. Knutsen, and G. E. Fraser 1998.
Does high soy milk intake reduce prostate cancer
incidence? The Adventist Health Study (United
States). Cancer Causes Control 9:553-557.
Jenkins, D. J., C. W. Kendall, D. Faulkner, E. Vidgen,
E. A. Trautwein, T. L. Parker, and A. Marchie. 2011.
Effect of a dietary portfolio of cholesterol-lowering
foods given at 2 levels of intensity of dietary ad-
vice on serum lipids in hyperlipidemia: epidemiol-
ogy of soy exposures and breast cancer risk. JAMA
306:831-839.
Johnell, O. and J. A. Kanis 2006. An estimate of the
worldwide prevalence and disability associated with
osteoporotic fractures. Osteoporos Int. 17:1726.
Kanis, J. A. 2007. WHO Technical Report, University
of Sheffield, UK: 66.
Khaodhiar, L., H. A. Ricciotti, L. Li, W. Pan, M. Schick-
el, J. Zhou, G. L. Blackburn 2008. Daidzein-rich
isoflavone aglycones are potentially effective in re-
ducing hot flashes in menopausal women. Meno-
pause. 15:125–132.
Kitamura, A., H. Iso, M. Iida, Y. Naito, S. Sato, D. R.
Jacobs, M. Nakamura, T. Shimamoto, and Y. Koma-
chi 2002. Trends in the incidence of coronary heart
disease and stroke and the prevalence of cardio-
vascular risk factors among Japanese men from
1963 to 1994. Am. J. Med. 112:104-109.
Krebs, E. E., K. E. Ensrud, R. MacDonald, and T. J.
Wilt 2004. Phytoestrogens for treatment of meno-
pausal symptoms: a systematic review. Obstet. Gy-
necol. 104:824–836.
Kuiper, G. G., B. Carlsson, K. Grandien, E. Enmark,
J. Haggblad, S. Nilsson, J. A. Gustafsson 1997.
Comparison of the ligand binding specificity and
transcript tissue distribution of estrogen receptors
alpha and beta. Endocrinol. 138:863-870.
Kuiper, G. G., J. G. Lemmen, B. Carlsson, J. C. Cor-
ton, S. H. Safe, P. T. van der Saag, B. van der Burg,
and J. A. Gustafsson 1998. Interaction of estrogen-
ic chemicals and phytoestrogens with estrogen re-
ceptor beta. Endocrinol. 139:4252-4263.
Kumar, N. B., A. Cantor, K. Allen, D. Riccardi, K.

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 137
Besterman-Dahan, J. Seigne, M. Helal, R. Salup,
and J. Pow-Sang 2004. The specific role of isofla-
vones in reducing prostate cancer risk. Prostate
59:141-147.
Kumar, N. B., J. P. Krischer, K. Allen, D. Riccardi, K.
Besterman-Dahan, R. Salup, L. Kang, P. Xu, and
J. Pow-Sang 2007. Phase II randomized, placebo-
controlled clinical trial of purified isoflavones in
modulating steroid hormones in men diagnosed
with localized prostate cancer. Nutr. Cancer
59:163-168.
Kurahashi, N., M. Iwasaki, M. Inoue, S. Sasazuki, and
S. Tsugane 2008. Plasma isoflavones and subse-
quent risk of prostate cancer in a nested case-con-
trol study: the Japan Public Health Center. J. Clin.
Oncol. 26:5923–5929.
Lampe, J. W. 2003. Isoflavonoid and lignan phytoes-
trogens as dietary biomarkers. J. Nutr. 133 Suppl
3:956S-964S.
Lampe, J.W. 2009. Is equol the key to the efficacy of
soy foods? Am. J. Clin. Nutr. 89:1664S–1667S.
Law, M. R., N. J. Wald, and S. G. Thompson 1994.
By how much and how quickly does reduction in
serum cholesterol concentration lower risk of isch-
aemic heart disease? BMJ. 308:367-372.
Lazarevic, B., G. Boezelijn, L. M. Diep, K. Kvern-
rod, O. Ogren, H. Ramberg, A. Moen, N. Wes-
sel, R. E. Berg, W. Egge-Jacobsen, C. Hammar-
strom, A. Svindland, O. Kucuk, F. Saatcioglu, K.
A. Taskèn, and S. J. Karlsen 2011. Efficacy and
safety of short-term genistein intervention in
patients with localized prostate cancer prior to
radical prostatectomy: a randomized, placebo-
controlled, double-blind Phase 2 clinical trial.
Nutr. Cancer 63:889-898.
Lamartiniere, C. A., Y. X. Zhao, and W. A. Fritz WA.
Genistein: mammary cancer chemoprevention, in
vivo mechanisms of action, potential for toxicity
and bioavailability in rats. J. Women’s Cancer 2000;
2:11-9.
Lee, H. P., L. Gourley, S. W. Duffy, J. Esteve, J. Lee,
and N. E. Day 1991. Dietary effects on breast-can-
cer risk in Singapore. Lancet 337:1197-1200.
Li, S. H., X. X. Liu, Y. Y. Bai, X. J. Wang, K. Sun, J.
Z. Chen, and R. T. Hui RT 2010. Effect of oral iso-
flavone supplementation on vascular endothelial
function in postmenopausal women: a meta-analy-
sis of randomized placebo-controlled trials. Am. J.
Clin. Nutr. 91:480-486.
Lichtenstein, A. H., L. J. Appel, M. Brands M, et al.
2006. Diet and lifestyle recommendations revision
2006: A scientific statement from the American
Heart Association Nutrition Committee. Circula-
tion 114:82–96.
Lock M. 1992. Contested meanings of the meno-
pause. Lancet 337:1270-1272.
Lock, M. 1994. Menopause in cultural context. Exp.
Gerontol. 29:307-317.
Lois, K., J. Young, and S. Kumar 2008. Obesity; epi-
phenomenon or cause of metabolic syndrome?
Int. J. Clin. Pract. 62:932–938.
Luh, B. S. 1995. Industrial production of soy sauce. J.
Indust. Microbiol. 14:467-471.
Magee, P.J. 2011. Is equol production beneficial to
health? Proc. Nutr. Soc. 70:10–18.
Magee, P. J., and I. T. Rowland 2004. Phyto-oestro-
gens, their mechanism of action: current evidence
for a role in breast and prostate cancer. Br. J. Nutr.
91:513-531.
Naya, M. and M. Imai. 2013. Recent advances
on soybean isoflavone extraction and enzy-
matic modification of soybean oil in Soybean
- Bio-Active Compounds, Prof. Hany El-Shemy
(Ed.), ISBN: 978-953-51-0977-8, InTech, DOI:
10.5772/52603. Available from: http://www.
intechopen.com/books/soybean-bio-active-
compounds/recent-advances-on-soybean-iso-
flavone-extraction-and-enzymatic-modification-
of-soybean-oil
Mahungu, S. M., S. Diaz-Mercado, J. Li, M. Schwenk,
K. Singletary, and J. Faller. 1999. Stability of iso-
flavones during extrusion processing of corn/soy
mixture. J. Agric. Food Chem. 47:279–284.
Mei, J., S. S. Yeung, and A. W. Kung 2001. High di-
etary phytoestrogen intake is associated with high-
er bone mineral density in postmenopausal but
not premenopausal women. J. Clin. Endocrinol.
Metab. 86:5217-5221.
Messina, M. 1999. Legumes and soybean overview
of their nutritional profiles and health effects. Am.

138 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
J. Clinical Nutr. 70(suppl.):439-450.
Messina, M., and S. Barnes. 1991. The role of soy
products in reducing risk of cancer. J. Natl. Cancer
Inst. 83:541-546.
Messina, M., and L. Hilakivi-Clarke 2009. Early intake
appears to be the key to the proposed protective
effects of soy intake against breast cancer. Nutr.
Cancer 61:792-728.
Messina, M., S. Ho, and D. L. Alekel 2004. Skeletal
benefits of soy isoflavones: a review of theclini-
cal trial and epidemiologic data. Curr. Opin. Clin.
Nutr. Metab. Care 7:649-658.
Messina, M., O. Kucuk, and J. W. Lampe 2006.
An overview of the health effects of isoflavones
with an emphasis on prostate cancer risk and
prostate-specific antigen levels. J. AOAC Int.
89:1121-1134.
Messina, M. J. and C. L. Loprinzi 2001. Soy for breast
cancer survivors: a critical review of the literature.
J. Nutr. 131(11 Suppl):3095S-3108S.
Messina, M., and G. Redmond 2006. Effects of soy
protein and soybean isoflavones on thyroid function
in healthy adults and hypothyroid patients: a review
of the relevant literature. Thyroid. 16:249-258.
Messina, M., C. Nagata, and A. H. Wu 2006. Estimat-
ed Asian adult soy protein and isoflavone intakes.
Nutr. Cancer 55:1-12.
Messina, M. J. and C. E. Wood. 2008. Soy isoflavones,
estrogen therapy, and breast cancer risk: analysis
and commentary. Nutr. J. 7:17 doi:10.1186/1475-
2891-7-17.
Milerová, J., J. Cerovská, V. Zamrazil, R. Bílek, O.
Lapcík, and R. Hampl 2006. Actual levels of soy
phytoestrogens in children correlate with thyroid
laboratory parameters. Clin. Chem. Lab. Med.
44:171-174.
Miyanaga, N., H. Akaza, S. Hinotsu, T. Fujioka, S.
Naito, M. Namiki, S. Takahashi, Y. Hirao, S. Horie,
T. Tsukamoto, M. Mori, and H. Tsuji 2012. Prostate
Cancer Chemoprevention Study: An investigative
randomized control study using purified isofla-
vones in men with rising prostate-specific antigen/
Cancer Sci. 103:125-130.
Mortensen, A., S. E. Kulling, H. Schwartz, I. Rowland,
C. E. Ruefer, G. Rimbach, A. Cassidy, P. Magee, J.
Millar, W. L. Hall, F. Kramer Birkved, I. K. Sorensen,
and G. Sontag 2009. Analytical and compositional
aspects of isoflavones in food and their biological
effects. Mol. Nutr. Food Res. 53(Suppl 2):S266–309.
Murphy, P. A., T. Song, G. Buseman, K. Barua, G. R.
Beecher, D. Trainer, and J. Holden 1999. Isofla-
vones in retail and institutional soy foods. J. Agric.
Food Chem. 47:2697-2704.
Muthyala, R.S., Y.H. Ju, S. Sheng, L.D. Williams, D.R.
Doerge, B.S. Katzenellenbogen, W. G. Helferich,
and J. A. Katzenellenbogen. 2004. Equol, a natu-
ral estrogenic metabolite from soy isoflavones:
convenient preparation and resolution of R- and
S-equols and their differing binding and biologi-
cal activity through estrogen receptors alpha and
beta. Bioorg. Med. Chem. 12:1559–1567.
Nagata, C. 2010. Factors to consider in the associa-
tion between soy isoflavone intake and breast can-
cer risk. J. Epidemiol. 20:83-89.
Nagata, Y., T. Sonoda, M. Mori, N. Miyanaga, K. Oku-
mura, K. Goto, S. Naito, K. Fujimoto, Y. Hirao, A.
Takahashi, T. Tsukamoto, H. Akaza 2007. Dietary
isoflavones may protect against prostate cancer in
Japanese men. J. Nutr. 137:1974–1979.
Nechuta, S. J., B. J. Caan, W. Y. Chen, W. Lu, Z. Chen,
M. L. Kwan, S. W. Flatt, Y. Zheng, W. Zheng, J. Pier,
and X. O. Shu 2012. Soy food intake after diagnosis
of breast cancer and survival: an in-depth analysis
of combined evidence from cohort studies of US
and Chinese women. Am. J. Clin. Nutr. 96:123–132.
Nestel, P. J., T. Yamashita, T. Sasahara, S. Pomeroy,
A. Dart, P. Komesaroff , A. Owen, and M. Abbey
1997. Soy isoflavones improve systemic arterial
compliance but not plasma lipids in menopausal
and perimenopausal women. Arterioscler. Thromb.
Vasc. Biol. 17:3392-3398.
Nestel, P. J., S. Pomeroy, S. Kay, P. Komesaroff, J.
Behrsing, J. D. Cameron, and L. West 1999. Isofla-
vones from red clover improve systemic arterial
compliance but not plasma lipids in menopausal
women. J. Clin. Endocrinol. Metab. 84:895-898.
Nishio, K., Y. Niwa, H. Toyoshima, K. Tamakoshi,
T. Kondo, H. Yatsuya, A. Yamamoto, S. Suzuki, S.
Tokudome, Y. Lin, K. Wakai, N. Hamajima, and A.
Tamakoshi 2007. Consumption of soy foods and

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 139
the risk of breast cancer: findings from the Japan
Collaborative Cohort (JACC) Study. Cancer Causes
Control. 18:801–808.
North, F. M., and K. Sharples 2001. Changes in the
use of hormone replacement therapy in New Zea-
land from 1991-1997. N. Z. Med. J. 114:250-253.
Pendleton, J. M., W. W. Tan, S. Anai, M. Chang, W.
Hou, K. T. Shiverick and C. J. Rosser 2008. Phase
II trial of isoflavone in prostate-specific antigen re-
current prostate cancer after previous local therapy.
BMC Cancer 8:132 doi:10.1186/1471-2407-8-132.
Poirier, L.A., D. R. Doerge, D. W. Gaylor et al. 1999.
An FDA review of sulfamethazine toxicity. Regul.
Toxicol. Pharmacol. 30:217-222.
Rabiau, N., M. Kossaï, M. Braud, N. Chalabi, S. Satih,
Y. J. Bignon, and D. J. Bernard-Gallon 2010. Genis-
tein and daidzein act on a panel of genes implicat-
ed in cell cycle and angiogenesis by polymerase
chain reaction arrays in human prostate cancer cell
lines. Cancer Epidemiol. 34:200-206.
Reynolds, K., A. Chin, K. A. Lees, A. Nguyen, D. Bu-
jnowski, and J. He 2006. A meta-analysis of the
effect of soy protein supplementation on serum
lipids. Am. J. Cardiol. 98:633–640.
Rivas, M., R. P. Garay, J. F. Escanero, P. Cia Jr., P. Cia,
and J. O. Alda 2002. Soy milk lowers blood pres-
sure in men and women with mild to moderate es-
sential hypertension. J. Nutr. 132:1900-1902.
Rock, C. L., C. Doyle, W. Demark-Wahnefried, J.
Meyerhardt, K. S. Courneya, A. L. Schwartz, E. V.
Bandera, K. K. Hamilton, B. Grant, M. McCullough,
T. Byers, and T. Gansler 2012. Nutrition and physi-
cal activity guidelines for cancer survivors. CA Can-
cer J. Clin. 62:242-274.
Rolfe, B.G. 1988. Flavones and isoflavones as induc-
ing substances of legume nodulation. Biofactors.
1:3–10.
Sacks, F. M., A. Lichtenstein, L. Van Horn, W. Harris,
P. Kris-Etherton, and M. Winston 2006. Soy protein,
isoflavones, and cardiovascular health: an Ameri-
can Heart Association Science Advisory for profes-
sionals from the Nutrition Committee. Circulation.
113:1034-1044.
Schone, F., G. Jahreis, R. Lange, W. Seffner, B. Grop-
pel, A. Hennig, and H. Ludke 1990. Effect of vary-
ing glucosinolate and iodine intake via rapeseed
meal diets on serum thyroid hormone level and
total iodine in the thyroid in growing pigs. Endo-
crinol. Exp. 24:415–427.
Schroder, C., A. Matthies, W. Engst, M. Blaut, and
A. Braune 2013. Identification and expression of
genes involved in the conversion of daidzein and
genistein by the equol-forming bacterium Slackia
isoflavoniconvertens. Appl. Environ. Microbiol.
79:3494–3502.
Setchell, K. D. 1998. Phytoestrogens: the biochemis-
try, physiology, and implications for human health
of soy isoflavones. Am. J. Clin. Nutr. 68:1333S–
1346S.
Setchell, K. D., N. M. Brown, and E. Lydeking-Olsen
2002. The clinical importance of the metabolite
equol-a clue to the effectiveness of soy and its iso-
flavones. J. Nutr. 132:3577-3584.
Setchell, K.D.R. and A. Cassidy. 1999. Dietary isofla-
vones: Biological effects and relevance to human
health. J. Nutr. 129:758-767.
Setchell, K.D., C. Clerici, E.D. Lephart, S.J. Cole, C.
Heenan, D. Castellani, B. E. Wolfe, L. Nechemias-
Zimmer, N. M. Brown, T. D. Lund, R. J. Handa, and
J. E. Heubi 2005. S-equol, a potent ligand for es-
trogen receptor beta, is the exclusive enantiomeric
form of the soy isoflavone metabolite produced by
human intestinal bacterial flora. Am. J. Clin. Nutr.
81:1072–1079.
Severson, R. K., A. M. Nomura, J. A. Grove, and G.
N. Stemmermann 1989. A prospective study of
demographics, diet, and prostate cancer among
men of Japanese ancestry in Hawaii. Cancer Res.
49:1857-1860.
Shu, X. O., F. Jin, Q. Dai, W. Wen, J. D. Potter, L. H.
Kushi, Z. Ruan, Y. T. Gao, and W. Zheng 2001. Soy-
food intake during adolescence and subsequent
risk of breast cancer among Chinese women. Can-
cer Epidemiol. Biomarkers Prev. 10:483–488.
Sites, C. K., B. C. Cooper, M. J. Toth, A. Gastaldell,
A. Arabshahi, and S. Barnes 2007. Effect of a daily
supplement of soy protein on body composition
and insulin secretion in postmenopausal women.
Fertil. Steril. 88:1609–1617.
Slavin, M., W. Kenworthy, and L. L. Yu. 2009. Anti-

140 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
oxidant properties, phytochemical composition,
and antiproliferative activity of Maryland-grown
soybeans with colored seed coats. J. Agric. Food
Chem. 57:11174-11185.
Smit, E., F. J. Nieto, C. J. Crespo and P. Mitchell 1999.
Estimates of animal and plant protein intake in us
adults: results from the Third National Health and
Nutrition Examination Survey, 1988–1991. J. Am.
Diet. Assoc. 99:813-820.
Synder, H.E. and T.W. Kwon. 1987a. Oriental soy food
products in Soybean Utilization. Van Nostrand Re-
inhold: New York, pp. 218–241.
Synder, H.E. and T.W. Kwon. 1987b. Processing of
soybeans in Soybean Utilization. Van Nostrand Re-
inhold: New York, pp. 74–144.
Squadrito, F., D. Altavilla, A. Crisafulli, A. Saitta, D.
Cucinotta, N. Morabito, R. D’Anna, F. Corrado, P.
Ruggeri, N. Frisina, and G. Squadrito 2003. Effect
of genistein on endothelial function in postmeno-
pausal women: a randomized, double-blind, con-
trolled study. Am. J. Med. 114:470-476.
Squadrito, F., D. Altavilla, N. Morabito, R. D’Anna, F.
Corrado, P. Ruggeri, G. M. Campo, G. Calapai, A.
P. Caputi, and G. Squadrito 2002. The effect of the
phytoestrogen genistein on plasma nitric oxide
concentrations, endothelin-1 levels and endothe-
lium dependent vasodilation in postmenopausal
women. Atherosclerosis 163:339-347.
Steinberg, F. M., N. L. Guthrie, A. C. Villablanca, K.
Kumar, and M. J. Murray. 2003. Soy protein with
isoflavones has favorable effects on endothelial
function that are independent of lipid and antioxi-
dant effects in healthy postmenopausal women.
Am. J. Clin. Nutr. 78:123–130.
Taku, K., K. Umegaki, Y. Sato, Y. Taki, K. Endoh, and
S. Watanabe. 2007. Soy isoflavones lower serum
total and LDL cholesterol in humans: A meta-anal-
ysis of 11 randomized controlled trials. Am. J. Clin.
Nutr. 85:1148–1156.
Teas, J., L. E. Braverman, M. S. Kurzer, S. Pino, T. G.
Hurley, and J. R. Hebert 2007. Seaweed and soy:
companion foods in Asian cuisine and their effects
on thyroid function in American women. J. Med.
Food. 10:90–100.
Thom, T. J., F. H. Epstein, J. J. Feldmen, P. E. Leaver-
ton, and M. Wolz 1992. Total morbidity and mor-
tality from heart disease, cancer and stroke from
1950 to 1987 in 27 countries. National Institutes of
Health Publication No. 92–3088, Bethesda, MD.
Tice, J. A., B. Ettinger, K. Ensrud, R. Wallace, T. Black-
well, and S. R. Cummings 2003. Phytoestrogen
supplements for the treatment of hot flashes: The
isoflavone clover extract (ICE) study: A randomized
controlled trial. JAMA 290:207-214.
Tsuchida, K., S. Mizushima, M. Toba, and K. Soda
1999. Dietary soybeans intake and bone mineral
density among 995 middle-aged women in Yoko-
hama. J. Epidemiol. 9:14-19.
Tsuda, M., T. Kitazaki, T. Ito, and T. Fujita 1986. The
effect of ipriflavone (TC-80) on bone resorption in
tissue culture. J. Bone Miner. Res. 1:207-11.
Tufano, A., P. Marzo, R. Enrini, L. Morricone, F. Ca-
viezel, and B. Ambrosi 2004. Anthropometric, hor-
monal and biochemical differences in lean and
obese women before and after menopause. J. En-
docrinol. Invest. 27:648–653.
Ursin, G., S. London, F.Z. Stanczyk, E. Gentzschein,
A. Paganini-Hill, R.K. Ross and M. C. Pike 1999.
Urinary 2-hydroxyestrone/16alpha-hydroxyestrone
ratio and risk of breast cancer in postmenopausal
women. J. Natl. Cancer Inst. 91:1067–1072.
USDA. 2008. USDA database for the isoflavone con-
tent of selected foods. Available at: http://www.
ars.usda.gov/SP2UserFiles/Place/12354500/Data/
isoflav/Isoflav_R2.pdf Accessed 28 April 2014.
van Erp-Baart, M. A., H. A. Brants, M. Kiely, A. Mul-
ligan, A. Turrini, C. Sermoneta, A. Kilkkinen, and L.
M. Valsta 2003. Isoflavone intake in four different
European countries: the VENUS approach. Br. J.
Nutr. 89 (Suppl 1):S25-30.
Villa, P., B. Costantini, R. Suriano, C. Perri, F. Macri,
L. Ricciardi, S. Panunzi, and A. Lanzone 2009. The
differential effect of the phytoestrogen genistein
on cardiovascular risk factors in postmenopausal
women: relationship with the metabolic status. J.
Clin. Endocrinol. Metab. 94:552–558.
Vincent, A. and L. A. Fitzpatrick 2000. Soy isofla-
vones: Are they useful in menopause? Mayo Clin.
Proc. 75:1174-1184.
Wakai, K., Y. Ohno, K. Genka, K. Ohmine, T. Kawamu-

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 141
ra, A. Tamakoshi, Y. Lin, T. Nakayama, K. Aoki,
and S. Fukuma 1999. Risk modification in lung
cancer by a dietary intake of preserved foods
and soyfoods: findings from a case-control study
in Okinawa, Japan. Lung Cancer 25:147-159.
Walker, H. A., T. S. Dean, T. A. Sanders, G. Jackson,
J. M. Ritter, and P. J. Chowienczyk 2001. The phy-
toestrogen genistein produces acute nitric oxide-
dependent dilation of human forearm vasculature
with similar potency to 17b- Estradiol. Circulation
103:258-262.
Wang, H. J. and P. A. Murphy. 1996. Mass balance
study of isoflavones during soybean processing. J.
Agric. Food Chem. 44:2377–2383.
Warri, A., N. M. Saarinen, S. Makela, and L. Hilakivi-
Clarke 2008. The role of early life genistein expo-
sures in modifying breast cancer risk. Br. J. Cancer
98:1485-1493.
Washburn, S., G. L. Burke, T. Morgan, and M. An-
thony 1999. Effect of soy protein supplementation
on serum lipoproteins, blood pressure, and meno-
pausal symptoms in perimenopausal women.
Menopause 6:7-13.
Weggemans, R. M., and E. A. Trautwein 2003. Rela-
tion between soy-associated isoflavones and LDL
and HDL cholesterol concentrations in humans: a
meta-analysis. Eur. J. Clin. Nutr. 57:940-946.
WHO. 2011a. Global status report on noncommu-
nicable disaeses 2010. Geneva, World Health Or-
ganization. http://www.who.int/nmh/publications/
ncd_report2010/en/ Accessed 11 March 2014.
WHO. 2011b. Global atlas on cardiovascular disease
prevention and control. Geneva, World Health Orga-
nization. http://www.who.int/cardiovascular_diseases/
publications/atlas_cvd/en/ Accessed 11 March 2014.
Wu, A. H., P. Wan, J. Hankin, C. C. Tseng, M. C. Yu,
and M. C. Pike 2002. Adolescent and adult soy in-
take and risk of breast cancer in Asian-Americans.
Carcinogenesis 23:1491-1496.
Wu, A. H., M. C. Yu, C. C. Tseng, and M. C. Pike 2008.
Epidemiology of soy exposures and breast cancer
risk. Br. J. Cancer. 98:9–14
Wu, A. H., R. G. Ziegler, P. L. Horn-Ross, A. M. No-
mura, D. W. West, L. N. Kolonel, J. F. Rosenthal,
R. N. Hoover, and M. C. Pike 1996. Tofu and risk of
breast cancer in Asian-Americans. Cancer Epide-
miol. Biomark. Prev. 5:901–906.
Wu, J., J. Oka, J. Ezaki, T. Ohtomo, T. Ueno, S. Uchi-
yama, T. Toda, M. Uehara, and Y. Ishimi 2007. Pos-
sible role of equol status in the effects of isofla-
vone on bone and fat mass in postmenopausal
Japanese women: a double-blind, randomized,
controlled trial. Menopause 14: 866–874
Wu, J., J. Oka, M. Higuchi, I. Tabata, T. Toda, M. Fu-
jioka, N. Fuku, T. Teramoto, T. Okuhira, T. Ueno, S.
Uchiyama, K. Urata, K. Yamada, and Y. Ishimi 2006.
Cooperative effects of isoflavones and exercise
on bone and lipid metabolism in postmenopausal
Japanese women: a randomized placebo-con-
trolled trial. Metabolism. 55:423–433.
Xiao, C. W., M. R. L’Abbé, S. Gilani, G. Cooke, I.
Curran, and S. A. Papademetriou 2004. Dietary
soy protein isolate and isoflavones modulate he-
patic thyroid hormone receptors in rats. J. Nutr.
134:743–749.
Yan, L., and E. L. Spitznagel 2009. Soy consumption
and prostate cancer risk in men: a revisit of a meta-
analysis. Am. J. Clin. Nutr. 89:1155-1163.
Yokoyama, S. and T. Suzuki 2008. Isolation and char-
acterization of a novel equol-producing bacterium
from human feces. Biosci. Biotechnol. Biochem.
72:2660–2666.
Yokoyama, S., K. Oshima, I. Nomura, M. Hattori,
and T. Suzuki 2011. Complete genomic se-
quence of the equol-producing bacterium Eg-
gerthella sp. strain YY7918, isolated from adult
human intestine. J. Bacteriol. 193:5570–5571.
Yuan, J.M., Q.S. Wang, R.K. Ross, B.E. Hender-
son, and M.C. Yu 1995. Diet and breast cancer
in Shanghai and Tianjin, China. Br. J. Cancer
71:1353–1358.
Zhan, S., and S. C. Ho 2005. Meta-analysis of the
effects of soy protein containing isoflavones on
the lipid profile. Am. J. Clin. Nutr. 81:397–408.
Zhang, X., X. O. Shu Y. T. Gao, G. Yang, Q. Li, H.
Li, F. Jin, and W. Zheng. 2003. Soy food con-
sumption is associated with lower risk of coro-
nary heart disease in Chinese women. J. Nutr.
133:2874–8.
Zhang, Y. B., W. H. Chen, J. J. Guo, Z. H. Fu, C. Yi, M.

142 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
Zhang, and X. L. NA 2013. Soy isoflavones supple-
mentation could reduce body weight and improve
glucose metabolism in non-Asian postmenopausal
women-A meta- analysis. Nutr. 29:8-14.
Zhou, J. R., L. Yu, Z. Mai, and G. L. Blackburn 2004.
Combined inhibition of estrogen dependent hu-
man breast carcinoma by soy and tea bioactive
components in mice. Int. J. Cancer 108:8-14.
Zubik, L. and M. Meydani 2003. Bioavailability of
soybean isoflavones from aglycone and gluco-
side forms in American women Am. J. Clin. Nutr.
77:1459– 1465.

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 143

144 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 145
The publishers do not warrant the accuracy of the articles in this journal, nor any views or opinions by their authors.
VOLUME 4 ISSUE 1
Preventing Post-Processing Contamination in a Food Nugget Processing Line When Lan-guage Barriers ExistJ. A. Neal, C. A. O’Bryan and P. G. Crandall
20
A Personal Hygiene Behavioral Change Study at a Midwestern Cheese Production PlantJ. A. Neal, C. A. O’Bryan and P. G. Crandall
13
Behavioral Change Study at a Western Soup Production Plant
C. A. O’Bryan, J. A. Neal, and P. G. Crandall
27
Salmonella in Cantaloupes: You Make Me Sick!B. A. Almanza
35
The Hurricane Sandy DilemmaB. A. Almanza
43
Introduction Special IssueP. G. Crandall
8
Case Studies
Instructions for Authors61
Introduction to Authors
Intellect-u-ale: A Smart Approach to Quality Assurance in a Micro-BreweryA. J. Corsi, M. Goodman, and J. A. Neal
50

146 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 147
MANUSCRIPT SUBMISSION
Authors must submit their papers electronically
([email protected]). According to instruc-
tions provided online at our site: www.afabjournal.
com. Authors who are unable to submit electroni-
cally should contact the editorial office for assistance
by email at [email protected].
INSTRUCTIONS TO AUTHORS
• Aerobic microbiology
• Aerobiology
• Anaerobic microbiology
• Analytical microbiology
• Animal microbiology
• Antibiotics
• Antimicrobials
• Bacteriophage
• Bioremediation
• Biotechnology
• Detection
• Environmental microbiology
• Feed microbiology
• Fermentation
• Food bacteriology
• Food control
• Food microbiology
• Food quality
• Food Safety
• Foodborne pathogens
• Gastrointestinal microbiology
• Microbial education
• Microbial genetics
• Microbial physiology
• Modeling and microbial kinetics
• Natural products
• Phytoceuticals
• Quantitative microbiology
• Plant microbiology
• Plant pathogens
• Prebiotics
• Probiotics
• Rumen microbiology
• Rapid methods
• Toxins
• Veterinary microbiology
• Waste microbiology
• Water microbiology
CONTENT OF MANUSCRIPT
We invite you to consider submitting your re-
search and review manuscripts to AFAB. The jour-
nal serves as a peer reviewed scientific forum for to
the latest advancements in bacteriology research
on Agricultural and Food Systems which includes
the following fields:

148 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
With an open access publication model of this
journal, all interested readers around the world can
freely access articles online. AFAB publishes origi-
nal papers including, but not limited to the types
of manuscripts described in the following sections.
Papers that have been, or are scheduled to be, pub-
lished elsewhere should not be submitted and will
not be reviewed. Opinions or views expressed in pa-
pers published by AFAB are those of the author(s)
and do not necessarily represent the opinion of the
AFAB or the editorial board.
MANUSCRIPT TYPES
Full-Length Research Manuscripts
AFAB accepts full-length research articles con-
taining four (4) figures and/or tables or more. AFAB
emphasizes the importance of sound scientific ex-
perimentation on any of the topics listed in the focus
areas followed by clear concise writing that describes
the research in its entirety. The results of experi-
ments published in AFAB must be replicated, with
appropriate statistical assessment of experimental
variation and assignment of significant difference.
Major headings to include are: Abstract, Introduc-tion, Materials and Methods, Results, Discussion (or Results and Discussion), Conclusion, Acknowl-edgements (optional), Appendix for abbreviations (optional), and References.
Manuscripts clearly lacking in language will be re-
turned to author without review, with a suggestion
that English editing be sought before the paper is
reconsidered. AFAB offers a fee based language
service upon request. Please contact [email protected] for more information about our fees
and services.
Rapid Communications
Under normal circumstances, AFAB aims for re-
ceipt-to-decision times of approximately one month or less. Accepted papers will have priority for publi-
cation in the next available issue of AFAB. However,
if an author chooses or requires a much more rapid
peer review, the journal editorial office has the capa-
bility to shorten the review timing to one week or less.
Any type of manuscript whether it be a full length
manuscript, brief communication or review paper can
be submitted as a rapid communication. There will be
additional costs for processing and page charges will
be double the normal rate. Authors who choose this
option must select Rapid Communications as the pa-
per type when submitting the paper and the editors
will judge whether a rapid review is possible and let
the author know immediately.
Brief Communications
Brief communications are short research notes giv-
ing the results of complete experiments but are con-
sidered less comprehensive than full-length articles
with three (3) figures and/or tables or less. Manuscripts
should be prepared with the same subheadings as full
length research papers. The running head above the
title of the paper is “Brief Communications.”
Unsolicited Review Papers
Review papers are welcome on any topic listed in
the focus section and have no page limits. Reviews
are assessed the same pages charges as all other
manuscripts. All AFAB guidelines for style and form
apply. Major headings to include are: Abstract, In-troduction, Main discussion topics and appropri-ate subheadings, Conclusions, Acknowledgements (optional) and References. Review papers shorter
than 20 pages of double spaced text and references
will be considered mini-reviews with the subhead-
ing above the title on the first page. The running
head above the title of the paper is either “Review”
or “Mini-review”.
Solicited Review Papers
Solicited reviews will have no page limits. The
editor-in-chief will send invitations to the authors
and then review these contributions when they are
submitted. Nominations or suggestions for potential
timely reviews are welcomed by the editors or edito-

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 149
rial board members and should be sent to submit@
afabjournal.com. There will be no page charges for
solicited review papers but the solicitation must origi-
nate from the editor-in-chief or one of the editors. Re-
quests from authors will automatically be classified as
unsolicited review papers. The running head above
the title of the paper will be “Invited Review.”
Conference and Special Issues Reviews
AFAB welcomes opportunities to publish papers
from symposia, scientific conference, and/or meet-
ings in their entirety. Conference organizers need
simply to contact AFAB at [email protected]
and a rapid decision is guaranteed. If in agreement,
the conference organizers must guarantee delivery
of a set number of peer reviewed manuscripts within
a specified time and submitted in the same format
as that described for unsolicited review papers. Con-
ference papers must be prepared in accordance with
the guidelines for review articles and are subject to
peer review. The conference chair must decide
whether or not they wish to serve as Special Issue
Editor and conduct the editorial review process. If
the conference chair/organizer chooses to serve as
special issue editor, this will involve review of the pa-
pers and, if necessary, returning them to the authors
for revision. The conference organizer then submits
the revised manuscripts to the journal editorial of-
fice for further editorial examination. Final revisions
by the author and recommendations for acceptance
or rejection by the chair must be completed by a
mutually agreed upon date between the editor and
the conference organizer. Manuscripts not meeting
this deadline will not be included in the published
symposium proceedings but if submitted later can
still be considered as unsolicited review papers. Al-
though offprints and costs of pages are the same
as for all other papers, the symposium chair may be
asked to guarantee an agreed upon number of hard
copies to be purchased by conference attendees. If
the decision is not to publish the symposium as a
special issue, the individual authors retain the right
to submit their papers for consideration for the jour-
nal as ordinary unsolicited review manuscripts.
Book Reviews
AFAB publishes reviews of books considered to
be of interest to the readers. The editor-in-chief ordi-
narily solicits reviews. Book reviews shall be prepared
in accordance to the style and form requirements of
the journal, and they are subject to editorial revision.
No page charges will be assessed solicited reviews
while unsolicited book reviews will be assigned the
regular page charge rate.
Opinions and Current Viewpoints
The purpose of this section will be to discuss, cri-
tique, or expand on scientific points made in articles
recently published in AFAB. Drafts must be received
within 6 months of an article’s publication. Opinions
and current perspectives do not have page limits.
They shall have a title followed by the body of the
text and references. Author name(s) and affiliation(s)
shall be placed between the end of the text and list
of references. If this document pertains to a par-
ticular manuscript then the author(s) of the original
paper(s) will be provided a copy of the letter and of-
fered the opportunity to submit for consideration a
reply within 30 days. Responses will have the same
page restrictions and format as the original opinion
and current viewpoint, and the titles shall end with
“Opinions.” They will be published together. Letters
and replies shall follow appropriate AFAB format
and may be edited by the editor-in-chief and a tech-
nical editor. If multiple letters on the same topic are
received, a representative set of opinions concern-
ing a specific article will be published. A disclaimer
will be added by the editorial staff that the opinion
expressed in this viewpoint is the authors alone and
does not necessarily represent the opinion of AFAB
or the editorial board.
COPYRIGHT AGREEMENT
The copyright form is published in AFAB as space
permits and is available online (www.afabjournal.com).

150 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
AFAB grants to the author the right of re-publication
in any book of which he or she is the author or edi-
tor, subject only to giving proper credit to the original
journal publication of the article by AFAB. AFAB re-
tains the copyright to all materials accepted for pub-
lication in the journal. If an author desires to reprint
a table or figure published from a non-AFAB source,
written evidence of copyright permission from an au-
thority representing that source must be obtained by
the author and forwarded to the AFAB editorial office.
PEER REVIEW PROCESS
Authors will be requested to provide the names
and complete addresses including emails of five (5) potential reviewers who have expertise in the research
area and no conflict of interest with any of the authors.
Except for manuscripts designated as Rapid Commu-
nication each reviewer has two (2) weeks to review
the manuscript, and submit comments electronically
to the editorial office. Authors have three (3) weeks
to complete the revision, which shall be returned to
the editorial office within six (6) weeks after which the
authors risk having their manuscript removed from
AFAB files if they fail to ask the editorial office for
an extension by email. Deleted manuscripts will be
reconsidered, but they will have to be processed as
new manuscripts with an additional processing fee as-
sessed upon submission. Once reviewed, the author
will be notified of the outcome and advised accord-
ingly. Editors handle all initial correspondence with
authors during the review process. The editor-in chief
will notify the author of the final decision to accept or
reject. Rejected manuscripts can be resubmitted only
with an invitation from the editor or editor-in chief. Re-
vised versions of previously rejected manuscripts are
treated as new submissions.
PRODUCTION OF PROOFS
Accepted manuscripts are forwarded to the edito-
rial office for technical editing and layout. The manu-
script is then formatted, figures are reproduced, and
author proofs are prepared as PDFs. Author proofs
of all manuscripts will be provided to the correspond-
ing author. Author proofs should be read carefully and
checked against the typed manuscript, because the
responsibility for proofreading is with the author(s).
Corrections must be returned by e-mail. Changes
sent by e-mail to the technical editor must indicate
page, column, and line numbers for each correction
to be made on the proof. Corrections can also be
marked using “track changes” in Microsoft Word or
using e-annotation tools for electronic proof correc-
tion in Adobe Acrobat to indicate necessary chang-
es. Author alterations to proofs exceeding 5% of the
original proof content will be charged to the author. All
correspondence of proofs must be agreed to by the
editorial office and the author within 48 hours or proof
will be published as is and AFAB will assume no re-
sponsibility for errors that result in the final publication.
PUBLICATION CHARGES
AFAB has two publication charge options: conven-
tional page charges and rapid communication. The
current charge for conventional publication is $25 per printed page in the journal. There is no additional
charge for the publication of pages containing color
images, micrographs or pictures. For authors who
wish to have their papers processed as a rapid com-
munication, authors will pay the rapid communication
fee when proofs are returned to the editorial office
in addition to twice the conventional page charges.
Charges for rapid communications are $1000 per manuscript for guaranteed peer review within one
week and $100 per journal page.
HARD COPY OFFPRINTS
If you are wishing to obtain a physical hard copy of
the AFAB journal, offprints are available in any quan-
tity at an additional charge: $100/page for black-white
and $150/page for color prints. You may order your
offprints at any time after publication on our website.
Scientific conference organizers may be expected to
agree to a set number of offprints as a part of their
agreement with AFAB.

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 151
MANUSCRIPT CONTENT REQUIREMENTS
Preparing the Manuscript File
Manuscripts must be written in grammatically
correct English. AFAB offers a fee based language
service upon request ([email protected]).
Manuscripts should be typed double-spaced, with
lines and pages numbered consecutively. All docu-
ments must be submitted in Microsoft Word (.doc or
.docx, PC or Mac). All special characters (e.g., Greek,
math, symbols) should be inserted using the sym-
bols palette available in this font. Tables and figures
should be placed in separate sections at the end of
the manuscript (not placed in the text). Failure to fol-
low these instructions will cause delays of the pro-
cessing and review of the manuscript.
Title Page
At the very top of the title page, include a title of
not more than 100 characters. Format the title with
the first letter of each word capitalized. No abbre-
viations should be used. Under the title, the authors
names are listed. Use the author’s initials for both first
and middle names with a period (full-stop) between
initials (e.g., W. A. Afab). Underneath the authors, a
list affiliations must be listed. Please use numerical
superscripts after the author’s names to designate
affiliation. If an authors address has changed since
the research was completed, this new information
must be designated as “Current address:”. The cor-
responding author should be indicated with an aster-
isk e.g., * Corresponding author. The title page shall
include the name and full address of the correspond-
ing author. Telephone and e-mail address must also
be provided for the corresponding author, and email-addresses must be provided for all authors.
Editing
Author-derived abbreviations should be defined
at first use in the abstract and again in the body of
the manuscript. If abbreviations are extensive au-
thors may need to provide a list of abbreviations
at the beginning of the manuscript. In vivo, in vitro
and bacterial names must be italicized (obligatory).
Authors must avoid single sentence paragraphs and
merge such paragraphs appropriately. Authors must
not begin sentences with “Figure or Table shows…”
as these are inanimate objects and cannot “show”
anything. When number are reported in text or in ta-
bles, always put a zero in front of decimal numbers:
“0.10” instead of “.10”.
MANUSCRIPT SECTIONS
Abstract
The abstract provides an abridged version of the
manuscript. Please submit your abstract on a sepa-
rate page after the title page. The abstract should
provide a justification of your work, objectives, meth-
ods, results, discussion and implications of study or
review findings . Your abstract must consist of com-
plete sentences without references to other work or
footnotes and must not exceed 250 words. On the
same page as your abstract, please provide at least ten (10) keywords to be used for linking and index-
ing. Ideally, these keywords should include signifi-
cant words from the title.
Introduction
The introduction should clearly present the foun-
dation of the manuscript topic and what makes the
research or the review unique. The introduction
should validate why this topic is important based on
previously published literature, and the relevance of
the current research. Overall goals and project ob-
jectives must be clearly stated in the final sentence
of the last paragraphs of the introduction.
Materials and Methods
Information on equipment and chemicals used
must include the full company name, city, and state
(country if outside the United States or Province if
in Canada) [i.e., (Model 123, ACME Inc., Afab, AR)].

152 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
Variability, Replication, and Statistical Analysis
To properly assess biological systems indepen-
dent replication of experiments and quantification
of variation among replicates is required by AFAB.
Reviewers and/or editors may request additional
statistical analysis depending on the nature of the
data and it will be the responsibility of the authors
to respond appropriately. Statistical methods com-
monly used in the bacteriology do not need to be
described in detail, but an adequate description
and/or appropriate references should be provided.
The statistical model and experimental unit must
be designated when appropriate. The experimen-
tal unit is the smallest unit to which an individual
treatment is imposed. For bacterial growth stud-
ies, the average of replicate tubes per single study
per treatment is the experimental unit; therefore,
individual studies must be replicated. Repeated
time analyses of the same sample usually do not
constitute independent experimental units. Mea-
surements on the same experimental unit over time
are also not independent and must not be consid-
ered as independent experimental units. For analy-
sis of time effects, assess as a rate of change over
time. Standard deviation refers to the variability
in the biological response being measured and is
presented as standard deviation or standard error
according to the definitions described in statistical
references or textbooks.
Results
Results represent the presentation of data in
words and all data should be described in same
fashion. No discussion of literature is included in
the results section.
Discussion
The discussion section involves comparing the
current data outcomes with previously published
work in this area without repeating the text in the
results section. Critical and in-depth dialogue is
encouraged.
Results and Discussion
Results and discussion can be under combined or
separate headings.
Conclusions
State conclusions (not a summary) briefly in one
paragraph.
Acknowledgments
Acknowledgments of individuals should include
institution, city, and state; city and country if not U.S.;
and City or Province if in Canada. Copies being re-
viewed shall have authors’ institutions omitted to re-
tain anonymity.
References
a) Citing References In Text
Authors of cited papers in the text are to be pre-
sented as follows: Adams and Harry (1992) or Smith
and Jones (1990, 1992). If more than two authors of
one article, the first author’s name is followed by the
abbreviation et al. in italics. If the sentence structure
requires that the authors’ names be included in pa-
rentheses, the proper format is (Adams and Harry,
1982; Harry, 1988a,b; Harry et al., 1993). Citations to a
group of references should be listed first alphabeti-
cally then chronologically. Work that has not been
submitted or accepted for publication shall be listed
in the text as: “G.C. Jay (institution, city, and state,
personal communication).” The author’s own un-
published work should be listed in the text as “(J.
Adams, unpublished data).” Personal communica-
tions and unsubmitted unpublished data must not
be included in the References section. Two or more
publications by the same authors in the same year
must be made distinct with lowercase letters after
the year (2010a,b). Likewise when multiple author ci-
tations designated by et al. in the text have the same
first author, then even if the other authors are differ-
ent these references in the text and the references
section must be identified by a letter. For example

Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014 153
“(James et al., 2010a,b)” in text, refers to “James,
Smith, and Elliot. 2010a” and “James, West, and Ad-
ams. 2010b” in the reference section.
b) Citing References In Reference Section
In the References section, references are listed in
alphabetical order by authors’ last names, and then
chronologically. List only those references cited in the
text. Manuscripts submitted for publication, accepted
for publication or in press can be given in the refer-
ence section followed by the designation: “(submit-
ted)”, “(accepted)’, or “(In Press), respectively. If the
DOI number of unpublished references is available,
you must give the number. The year of publication fol-
lows the authors’ names. All authors’ names must be
included in the citation in the Reference section. Jour-
nals must be abbreviated. First and last page num-
bers must be provided. Sample references are given
below. Consult recent issues of AFAB for examples
not included in the following section.
Journal manuscript:
Examples:
Chase, G., and L. Erlandsen. 1976. Evidence for a
complex life cycle and endospore formation in the
attached, filamentous, segmented bacterium from
murine ileum. J. Bacteriol. 127:572-583.
Jiang, B., A.-M. Henstra, L. Paulo, M. Balk, W. van
Doesburg, and A. J. M. Stams. 2009. A typical
one-carbon metabolism of an acetogenic and
hydrogenogenic Moorella thermioacetica strain.
Arch. Microbiol. 191:123-131.
Book:
Examples:
Hungate, R. E. 1966. The rumen and its microbes
Academic Press, Inc., New York, NY. 533 p.
Book Chapter:
Examples:
O’Bryan, C. A., P. G. Crandall, and C. Bruhn. 2010.
Assessing consumer concerns and perceptions
of food safety risks and practices: Methodologies
and outcomes. In: S. C. Ricke and F. T. Jones. Eds.
Perspectives on Food Safety Issues of Food Animal
Derived Foods. Univ. Arkansas Press, Fayetteville,
AR. p 273-288.
Dissertation and thesis:
Maciorowski, K. G. 2000. Rapid detection of Salmo-
nella spp. and indicators of fecal contamination
in animal feed. Ph.D. Diss. Texas A&M University,
College Station, TX.
Donalson, L. M. 2005. The in vivo and in vitro effect
of a fructooligosacharide prebiotic combined with
alfalfa molt diets on egg production and Salmo-
nella in laying hens. M.S. thesis. Texas A&M Uni-
versity, College Station, TX.
Van Loo, E. 2009. Consumer perception of ready-to-
eat deli foods and organic meat. M.S. thesis. Uni-
versity of Arkansas, Fayetteville, AR. 202 p.
Web sites, patents:
Examples:
Davis, C. 2010. Salmonella. Medicinenet.com.
http://www.medicinenet.com/salmonella /article.
htm. Accessed July, 2010.
Afab, F. 2010, Development of a novel process. U.S.
Patent #_____
Author(s). Year. Article title. Journal title [abbreviated].
Volume number:inclusive pages.
Author(s) [or editor(s)]. Year. Title. Edition or volume (if
relevant). Publisher name, Place of publication. Number
of pages.
Author(s) of the chapter. Year. Title of the chapter. In:
author(s) or editor(s). Title of the book. Edition or vol-
ume, if relevant. Publisher name, Place of publication.
Inclusive pages of chapter.
Author. Date of degree. Title. Type of publication, such
as Ph.D. Diss or M.S. thesis. Institution, Place of institu-
tion. Total number of pages.

154 Agric. Food Anal. Bacteriol. • AFABjournal.com • Vol. 4, Issue 2 - 2014
Abstracts and Symposia Proceedings:
Fischer, J. R. 2007. Building a prosperous future in
which agriculture uses and produces energy effi-
ciently and effectively. NABC report 19, Agricultural
Biofuels: Tech., Sustainability, and Profitability. p.27
Musgrove, M. T., and M. E. Berrang. 2008. Presence
of aerobic microorganisms, Enterobacteriaceae and
Salmonella in the shell egg processing environment.
IAFP 95th Annual Meeting. p. 47 (Abstr. #T6-10)
Vianna, M. E., H. P. Horz, and G. Conrads. 2006. Op-
tions and risks by using diagnostic gene chips. Pro-
gram and abstracts book , The 8th Biennieal Con-
gress of the Anaerobe Society of the Americas. p.
86 (Abstr.)
Data Presentation in Tables and Figures
Figures and tables to be published in AFAB must
be constructed in such a fashion that they are able
to “stand alone” in the published manuscript. This
means that the reader should be able to look at
the figure or table independently of the rest of the
manuscript and be able to comprehend the experi-
mental approach sufficiently to interpret the data.
Consequently, all statistical analyses should be very
carefully presented along with variation estimates
and what constitutes an independent replication
and the number of replicates used to calculate the
averages presented in the table or figure.
Each table and figure must be on a separate
page in the submitted paper. In addition, you will
need to submit all data for charts, tables and
figures in native format when possible (e.g., Mi-
crosoft Excel, Powerpoint). Photographs should
be submitted as high-resolution (600 dpi) .jpg or
tif. files. All figures should be clearly presented with
well defined axis and units of measurement. Sym-
bols, lines, and bars must be made distinct as “stand
alone” black and white presentations. Stippling,
dashed lines etc. are encouraged for multiple com-
parison but shades of gray are discouraged. Color
images, micrographs, pictures are recommended
and there is no additional fee for their submission.
AFAB Online Edition is Now Available!
www.AFABjournal.com
• Free Access
• Print PDFs
• Flip Through Issues
• Search Article Archives
• Order Reprints
• Submit a Paper


Online Publication: www.AFABjournal.com