an investigation into the mechanisms involved in the
TRANSCRIPT

Copyright is owned by the Author of the thesis. Permission is given for a copy to be downloaded by an individual for the purpose of research and private study only. The thesis may not be reproduced elsewhere without the permission of the Author.

AN INV EST IGATION INTO THE MECHAN ISMS INVOLVED IN THE DEPRES S ION OF
OVULATION RATES IN EWES GRAZING OESTROGENIC LUCERNE
A thes i s p r e s en t ed in pa rt ial ful f il ment of the requ iremen t s for
the degree o f Ma s t er of Agr icul ture Sc ienc e
in An ima l S c i ence at
Ma s sey Un iver s i ty
BERNARD JOSEPH McLEOD
1 9 7 8
Ot-\�1-0R

A B S T R A C T
The r eproduc t ive per f o rmance o f 123 Romney and Border Leic e s t er
x Romney ewe s wa s compar ed a f t e r they gra zed o e s t rogenic l uc erne or
non -oestrogen ic ryegra s s / c lover pa s t ur e .
i i
T o synchron ize oestrus pr ior to t h e exper imen tal t r ea tmen t ,
progestagen-impr egna t ed in t ravag inal sponges wer e inser t ed in a l l ewes
for 14 days . In Exper imen t I , wh ich invo lved 42 Romn ey and Border
Leic ester x Romney ewes , the an imal s gra z ed trea tmen t pa stures for one
comp l e t e oes trous cyc l e . B lood samples wer e taken a t interva l s
thro ughout t he cyc l e and l u t ein i z ing ho rmone concen t ra t ions wer e
d e t e rmined . Ovula t ion r a t e s and the numbers o f fol l ic l es presen t on
the surface of the ovaries wer e r ec orded a t laparotomy , three days po s t -
oes trus .
In Exper iment I I , which invo lved 81 Romney ewes , oes trogen ic lucerne
or non-oes trogenic ryegra s s / c lover pa s tu r e wa s grazed for a comp l e t e
o e s trous c yc l e , or t ea tmen t s wer e int erchanged i n mid-cyc l e . Al l ewes
wer e slaugh tered three days po s t -oes trus and their r eproduc t ive trac t s wer e
r ecovered and individual ly id en t i f i ed . Ovula tion r a t e s and the
numbers o f fo l l ic les on t he sur face of t he ovar i e s were r ecorded .
Af t er sec t ioning the ovar ies , a l l fo l l icles of a d iame ter gr eater t han
2.0mm wer e recorded . Sec t ions o f t i ssue from t he vag ina , c ervix , u t erus
and fa llopian tubes of each ewe were moun t ed , sta ined wi th ha ema toxyl in
and eosin , and the height of their ep i thel ial cel l s mea sur ed .
The inge s t ion o f oes trogenic lucerne for one comp l e t e oes trous
c yc l e depr e s s ed ovula t ion r a t e s by 29 per c en t (0.67 ovula t ions per ewe)
in Exper imen t I and by 22 percent (0.40 ovulat ion s per ewe) in Exper iment
II. The inge s t ion o f o es trog enic luc erne for pa r t o f the o e s t ro u s c yc l e ,

depr essed ovulat ion r a t e s only i f i t wa s consumed for the l a t t er
hal f o f t h e cyc l e .
i i i
O e s t ro genic luc erne d id no t sign i f ican tly in f luence t h e s ec r e t ion
of l u t e in i z ing hormone over the pre-ovula tory per iod , or a t any o ther
s tage of t h e oestrous cyc l e .
The r e wer e no s ignificant d i f f erenc es in t he to tal numb ers o f
ovar ian f o l l icles presen t , i n the numbers o f fol l ic l es o n the sur fac e
o f the ova r ie s , o r in the numbers o f large f o l l ic l e s ( wi t h a d iame ter
o f gr ea t er than 3 .5mm) p r e s en t , b e tween the trea tmen t group s .
The inges t ion of o e s t ro g enic lucerne did no t inc r ea s e sign i f ican tly ,
the h eight o f epi thel ia l c el l s in gen i ta l trac t t issue , when compared
with tha t in ewes graz ing non -oest rogenic ryegra s s / c l ov er pa s ture .
S imilar l y , there was no sign i f icant d i f f erence in u terine we ight s .
Th is i s in contrast to pub l i shed da ta where luc erne i s f ed to speyed
ewe s .

iv
A C K N 0 W L E D G E M E N T S
The author is spe c ia l ly indeb ted to his supervisor
Dr . M . F. McDona l d for his invaluab l e gui dance and a s s i s tance in
expe r ime n t a l work and adv ice dur ing the preparat ion o f this manu s c r ip t .
Special thanks are due t o D r . K . R. Lapwood for assis t ance with
radio immunoassays and for his he l p fu l d i s cuss ions and adv ice ; t o
Mr . M . J . Bir t les for a s s i s t ance with histo logica l preparat ions ; and
to Dr . R . W . Ke l ly , Inve rmay Agricu l t ural Research Centre , for the
coume s tan assays . Special apprec iat ion for t e chnical assis tance is
due to Mr. C . Mu i r , Mr . M. Wyc h e r l ey and Mr . B . Tha t cher and t o a l l
those who assisted with the co l l e c t ion o f b lood samp l e s , especi a l l y
my f e l l ow s t udents o f reprod u c t ive physio logy , Ma rk Fishe r , S t ua r t
Mc C u t cheon , Bi l l McMi l lan and Al a s t a i r Gibson .
Acknowl edgement i s mad e t o Mr . P.H. Whi t ehead and farm s t a f f
and to Miss J . McKegg for the typ ing o f t h i s manus crip t .
Financial assistance f rom the D . J . McGowan Tru s t and the
J . Hur ley Trus t , is gra t e f u l ly acknowledged .
Final ly , special t hanks a r e due t o my fami ly for s upport and
encouragement throughout this s t udy .

v.
P R E F A C E
This inv e s t iga t ion wa s c onduc t ed a t the Fer t il i ty Cen tr e , par t
o f the Depa r tmen t of S heep Hu sband ry , Ma s s ey Univer s i t y . The
experimen tal work wa s c a r r i ed out in April and May of 1 9 77 and represen t s
or ig ina l r es earch b y t h e au thor under the supervis ion o f Dr . M . F .
McDona ld , Reader , Sheep Husbandry Depa r tmen t , Ma s s ey Univer s i ty .

T A B L E 0 F C 0 N T E N T S
ABSTRACT • . . .
ACKNOWLEDGEMENTS
PREFACE
LIST OF TABLES
LIST OF FIGURES
INTRODUCTION . .
CHAPTER I
REVIEW OF LITERATURE .
(A) PHYTO-OES TROGENS
( i ) Phyto-oestrogen s in Pas ture Plants
( i i ) Fa c tors Inf luenc ing Phy to-oest rogen Levels
(iii) The Me tabo l ism o f Phy to-o e s t rogens in Sheep
( iv ) Phyto-oes trogens and Repro duc t ive Fa i lure in Sheep . . . . . . . . . . . . . . .
( B ) THE MECHANI SM O F ACTION OF OES TROGENS
( C ) EXOGENOUS OESTROGENS AND REPRODUCTION IN THE EWE
( i ) The Oes t rous Cyc l e
( i i ) Gonado trophin S ec r e t ion
( ii i ) The Reproduc t ive Trac t
( iv) Oes t rogen Recep t o r s
( D ) FOLLICLE MATURATION I N THE EWE
( i ) Pa t terns o f Fo l l ic l e G rowth and Atresia
( i i ) Hormonal Con trol o f Fo l l i c l e Ma tura t ion
Page
ii
iv
V
ix
xi
xii
1
1
1
4
6
8
12
13
13
1 5
1 8
19
2 0
20
21

( E )
( F )
OVULATI ON I N THE EWE . .
( i )
( i i )
( i i i )
Fac to r s Influenc ing Ovulat ion Ra tes
P o s tulated Mechanisms of Ovula t ion
Blockade of Ovulation
THE PURPOSE AND SCOPE OF THE INVESTIGATION
CHAPTER I I
MATERIALS AND METHODS .
(A) EXPERIMENTAL PLAN
( B )
( C )
( D )
( E )
( F )
( G )
( H )
( I )
LUCERNE AND CONTROL PASTURES
AN IMALS
OESTRUS SYNCHRONI ZATI ON
HORMONE ANALY S ES .
CERVICAL MUCUS ANALY S I S
UTERINE AND OVARIAN WEIGHTS
FOLLI CLE POPULATI ONS .
REPRODUCTIVE TRACT HISTOLOGY
( J ) ANALY S I S OF DATA
RESULTS . . . . . . .
CHAPTER Ill
(A) LUCERNE OESTROGENIC POTENCY
( i ) Luc erne Coume s tan Con t ent
( i i ) Daily Coumes tan Intake
( B ) EWE LIVEWEIGHTS AND OESTRUS
( i ) Ewe Livewe igh t s . . . .
( i i ) Oes trus Synchroni z a t ion
( i i i ) Oe s t rus-Ovulat ion Interval
Page
25
25
2 6
2 8
29
30
3 0
3 1
3 1
3 2
3 2
33
34
3 4
3 4
3 4
36
36
36
36
37
37
39
39

(C) LUTEINIZ ING HORMONE S ECRETION
( i ) Dur ing Oes t rous Cyc le
( i i) Pre-Ovul atory LH Peak
(D) OVULAT ION RATES
( i ) Experiment I
( i i ) Experimen t I I
( i i i ) Corpora Lu tea Diame t e r
(E) FOLLICLE POPULATIONS
( i ) Surface Fol l i c l e s
( i i ) To tal Fo llicle Popula t ions
(F) OVARIAN AND REPRODUCTIVE TRACT RESPONSES
( i ) Ovar ian and Ut erine Weigh t s
( i i ) Tra c t His tology
(G) SUMMARY . . . . . .
DI SCUSS ION
(A) OES TROGENIC LUCERNE
( B ) OES TRUS PHENOMENA .
CHAPTER IV
( i ) Synchron iza t ion of Oes trus
( ii ) Oes trus To Ovula t ion Int erval
( C ) LUTEINIZING HORMONE S ECRETION
( D ) DEPRESS ION O F OVULATION RATES
(E) FOLL ICLE POPULATIONS
(F) P ROGESTERONE S ECRETION
( G ) COUMESTANS AND THE REPRODUCTIVE TRACT
( H ) CONCLUS IONS
BIBLIOGRAPHY . .
Page
4 0
4 0
4 0
4 3
4 3
4 3
4 5
4 5
4 5
4 8
4 8
4 8
5 2
5 3
5 5
5 5
5 5
55
56
5 7
5 9
6 1
6 3
64
6 5
7 1

Tabl e 1 - 1
Tabl e 1-2
Tab l e 3- 1
Table 3-2
Table 3-3
Table 3-4
Table 3-5
Table 3-6
Table 3-7
Table 3-8
Tabl e 3-9
Tabl e 3- 1 0
Tabl e 3- 1 1
L I S T 0 F T A B L E S
The relat ive po t enc ies o f phyto-oes t rogens
admin i s tered orally or intramuscularl y .
Oestrogenic luc erne and repro duc t ive
per formanc e
Lucerne coume s tan conten t
The daily coume s t an intake (mg/day) o f ewes
inges t ing 1 kg ( DM) of lucerne .
Mean ewe l iveweights (kg) . .
Mean plasma LH concentrations (ng/ml ) during
the oes trous cyc l e , in ewes graz ing pas ture or
Page
3
1 1
3 6
3 7
3 8
oe s t rogenic luc e rne . 4 1
P re-ovulatory LH concentrations (ng/ml ) in ewes
graz ing pas ture or o e s t ro genic luc erne . 42
Mean ovula t ion ra t e s for ewe s grazing pas ture
o r oes trogenic luc erne - Experiment I . . 43
Mean ovulation ra t e s for ewe s grazing pas ture
o r oes t ro genic lucern e - Experimen t II
Compar i sons o f the d i f f er enc e s in ovulation rates
be tween t r ea tmen t group s - Experimen t II
Mean corpo ra lutea d iame ter (mm) three days
po s t-oe s trus
Surface f o l l i c l e s o f greater than 3 . 5mm d iamet e r,
corpora lutea and follicles p e r ovula t ion for
ewes grazing pas ture o r o e s t ro genic lucern e -
44
44
4 5
Exp erimen t I . . . . . . . . . . . . . . .. 46
Sur face f o l l i c l e s of grea t er t han 2 . 0mm
d iamet er , corpora lutea and fol licl e s p e r
ovula t ion f o r ewes graz ing pas ture o r
o e s t rogenic lucerne - Experiment I • • • • • • 4 7

Tabl e 3- 12
Tab l e 3- 1 3
Tab l e 3-14
Tab l e 3- 15
Tabl e 3- 1 6
Tabl e 3- 1 7
Page
Ovar ian fol l ic l e populat ions of ewes graz ing
pasture or oestrogenic l ucerne - Experimen t I I 4 9
To tal number o f f o l l i c l e s o f greater t han
3 . 5mm diameter , corpo ra l u t ea and f o l l i c l e s
p e r ovula t ion - Exper imen t I I
To tal numb er o f f o l l i c l e s o f grea ter than
2.0mm diameter , corpo ra l u t ea and f o l l i c l e s
p e r ovulat ion - Experiment I I
Fo l l i c l e s grea t er than 2.0mm d iame t er , p e r
gram o f ova rian t issue
Ovarian and uterine we ight s (g) of ewes
grazing pa s ture or o e s t ro gen ic lucerne
Ep ithel ial cell he igh t (�) o f reproduc t ive
t rac t t issue f rom ewe s graz ing pa s ture or
o e s t rogenic lucerne . . . . . . . . . . .
50
5 1
48
52
52

Fig . 1 - 1
Fig . 1 - 2
Fig . 1 - 3
Fig . 2 - 1
Fi g . 2-2
Fig . 3- 1
F i g . 3-2
Fig . 3-3
Fig . 3-4
Fig . 3-5
Fig . 3-6
Fig . 3-7
L I S T 0 F F I G U R E S
Phyto -o e s t rogens . . . . . . .
The metabol ism of fo rmononet in in sheep
Po stulated mechan ism o f o e s t rogen a c t ion
wi thin tar get cel l s
Exper imen tal plan - Exper imen t I .
Exper imen tal plan - Exper imen t I I
D i s t r ibut ion o f t he onset o f oes t rus-
Exp er imen t I . . . . . . . . .
Dis t r ibut ion o f t he onset o f oestrus-
Exper imen t II . . . . . .
Onset of Oest rus-LH peak int erval
LH secre tion over the oes trous cyc l e
Pr e-ovula tory L H s ecretion
Ovulat ion rates
Po s t-ovula t ion fo l l i c l e popula tions
f refers to following page
Page
f 3
f 6
f 1 2
f 3 0
f 30
f 39
f 39
f 39
f 4 1
f 4 2
f 44
f 51

INTRODUCTION
Luc erne (medicago sa t iva) has become incr eas ingly impor tant as a
forage crop in tho s e par t s o f New Zeal and which exper i ence dry summer
cond i t ion s . I t s abil i ty to p roduc e high yields o f fo rage , o f high
nutr i t ive value , whether grazed in situ or f ed as hay or silage , has
led to i t s popul arity . In these areas luc erne produc t ion frequently
exceeds tha t o f convent ional pa s ture s .
xii
Because i t produces forage o f high nut r i t ive value in late summer
and autumn , lucerne would appear t o be ideal for ' f l ushin g ' ewe s prior
to ma t ing . However , lucerne has b een shown to con tain oes t ro genic
compound s which impa ir the reproduc tive performance of ewes . The phyto-
oe s t ro gens , the coumes tans , and to a le sser degree the isoflavones , may
reach high concen t r a t ions in lucerne l ea f ma terial , and the coumes tans
have been shown to lower lamb ing perc en tages . This d epres s ion in lamb ing
percentage is rela t ed to a redu c t ion in ovulat ion r a t e s .
An unders tand ing o f tho se mechanisms involved in the luc erne -
induced depression o f ovulat ion rates would be o f cons iderable advantage .
I t may enab l e management pra c t i c e s to be devised , tha t would al low lucerne
to be u t il i z ed in achieving opt imum ewe l iveweights prior to tupping ,
wit hout incurr ing any lo ss in r eproduc t ive performance .

C H A P T E R I
REVIEW OF LITERATURE

(A) PHYTO-OESTROGENS
C H A P T E R I
REVI EW OF LITERATURE
The t erm ' o estrogen ' originally re ferred to c ertain s tero id
hormones secre t ed by the ovary . The dev elopmen t o f the Al l en -Do isy
l.
vaginal smear t es t in 1923 provid ed a wider definition o f o e s t ro genic i ty .
' An o e s t ro gen i s any pure chemical subs tance which , when inj ec ted into
adul t ovar iec tomized mice or ra t s , produc es co rmf ica t ion of the vag ina ,
s imilar to tha t occurring during oes trus in the normal in t a c t an imal '
(Al l en and Da i sy , 1923) .
The o ccurrenc e o f o e s t ro genic subs tances in plan t s was f i r s t
repo r t ed i n 1 9 26 ( Dorhn e t al . , 19 26; Fellner , 1926; Lo ewe , e t a l . , 19 27) ani
the f ir s t oes trogen to be isol a t ed and iden t ified in the p lan t world wa s
o e s t rone , extrac ted from the oil palm ( Butenandt and Jacob i, 1 9 3 3 ) .
Cons equen tly several compoun ds clo s ely resemb l ing the na tural s t eroid
o e s t ro gens wer e isola ted f rom a number of plan t s. An o e s triol-l ike
compo und wa s ext rac ted f rom wil low ca tkins (Skarzynski , 19 3 3), and
f rom l iquorice roo t ( Costello and Lynn , 1 9 50) . A c r eeper c l a imed by
the Chine se to possess aphro d i s iac propert ies was shown to con tain mire
oestral ( Jones and Pope , 1961) and plan t s used for con t racep t ive
purpo ses in Ind ia and China contain ed p so ral idin ( Da t tagup ta et al . , 19 60) .
( i ) Phyto-Oest rogens in Pasture Plan t s
When B enne t t s , Underwood and Shier ( 1 94 6) demon strated a n in f er t il i ty
syndrome a f f ec t ing ewes gra z ing sub terranean clover pas ture ( Tr i f o l ium
sub t e rraneum L var . Dwalganup) , they provided s trong presump t ive eviden c e
o f t h e presence o f a n oes t ro gen . This was conf irmed by Curnow e t a l . ,
( 19 4 8) who showed the o e s t ro genic a c t ivity o f e ther extrac t s o f
sub t erranean clover ; and suppo r t ed ind irec t ly b y Eas t e t a l . , ( 1949)
with the d emons t ra t ion o f a p ro te c t ive a c t ion o f androg en inj ec t ions t o

2 .
sheep grazing clover pastures . Oestrogenic activity was also attributed
to red clover ( Pope, 1954), lucerne ( Coop and Clark, 19 60) and red clover
and lucerne hay ( Cheng et al . , 1953).
In 19 51 Bradbury and White succeeded in isolating two isoflavones
from subterranean clover extract; 7-hydroxy-4 '-methoxy isoflavone
( formononetin) and 5:7: 4'-trihydroxy isoflavone ( genistein). Genistein
was also isolated from red clover ( Pope and Wright, 1 954; Curnow and
Rossiter, 1955; Wong, 19 62), and from white clover and lucerne
( Guggolz et al . , 19 61). Subsequently the isoflavones 7 : 4 '-dihydroxy
isoflavone ( diadzein), 5 : 7-dihydroxy-4'-methoxy isoflavone ( biochanin A),
and 5 : 7 : 3'-trihydroxy-4'-methoxy isoflavone were isolated from both
subterranean and red clovers (Livingstone et al . , 19 6 1). These isoflavones
were all shown to be oestrogenic ( Bradbury and White, 1954; Moule et al . ,
19 6 3 ), and their synthetic analogues were shown to be of equal oestrogenic
potency ( Cheng et al., 1 9 54 ) .
Bickoff et al . , ( 1 957; 1 958) isolated a crystalline substance of
oestrogenic nature from ladino clover, strawberry clover and alfalfa
( lucerne). This compound w�s not an isoflavone but a benzofurocoumarin
derivative - a class later named 'coumestans' . Bickoff proposed the
name 'coumestrol' for this oestrogenic compound which proved to be 30
times more potent than genistein ( Bickoff et al., 1958) . Coumestrol
and the non-oestrogenic 4'-methoxycoumestrol, have since been detected
in red clover, subterranean clover and burr clover ( Lyman et al. , 1959).
In addition, six other coumestans have been isolated from lucerne
( Bickoff et al . , 19 64) and from ladino clover ( Bickoff et al . , 19 6 5) .
The coumestans were later sho\vn to be biogenetically related to the
isoflavones ( Grisebach and Barz, 19 6 3 ) .
Until recently the most commonly used method o f measuring the
oestrogenic potency of a phyto-oestrogen has been the mouse uterine

3 .
response. However difficulties have often arisen in attempting to
correlate results obtained with mice and other laboratory animals, to
the results obtained under field conditions with farm animals (Morley
et al . , 1 9 6 8) . Bickoff and eo-workers ( 19 6 2 ) compared the oestrogenic
potencies of orally fed phyto-oestrogens and steroid oestrogens, with
injected admin::istration in mice. Braden et al . , (19 6 7 ) made similar
comparisons using ovariectomized ewes . The comparative potencies are
shown in Table 1- 1 .
TABLE 1-1: THE RELATIVE POTENCIES OF PHYTO-OESTROGENS ADMINISTERED
ORALLY OR INTRAMUSCULARLY
OESTROGEN MOUSE EWE
ORAL I/M ORAL
Diethylstilboestrol 1 00, 000 100, 000 1 00, 000
Oestrone 6, 900 6, 900 6,900
Coumestrol 3 5 3 5 9
Genistein 1 . 5 1 . 5 0 . 0 7
Diadzein 0.4 0 . 4 0 . 02
Biochanin A 1 . 0 1 . 0 0 . 05
Formononetin negligible negligible 0 . 06
from Bickoff et al., (1 9 6 2), Braden, et al . , (1 9 6 7) and
Wong and Flux, ( 1 9 6 2 )
I/M
1 00, 000
6 , 900
35
1 . 5
0 . 4
1 . 0
negligible
Despite the relatively low potencies of the phyto-oestrogens, they
are present in certain forages in sufficient quantities to influence
animal physiology . The structures of the phyto-oestrogens are shown
in FIG 1 - 1.
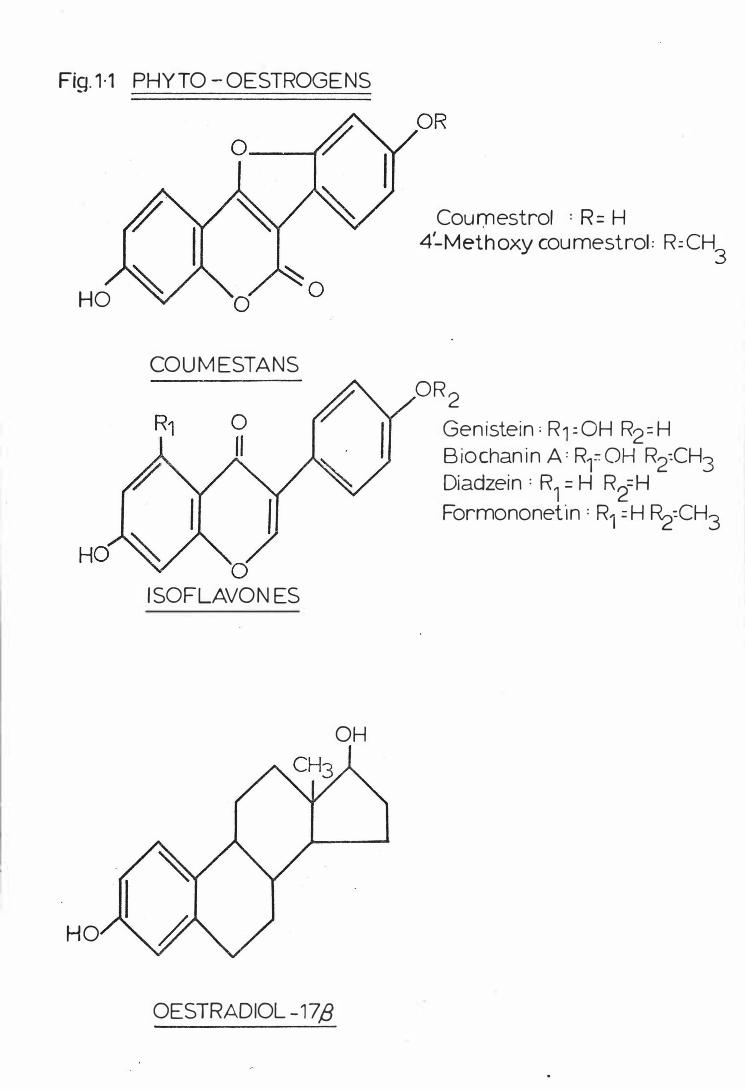
I· I
F ig. 1·1 PHYTO- OESTROGE NS
HO
HO
HO
COU M ESTANS
0 ISOF LAVON ES
OESTRADIOL -17/3
OH
Cournestrol : R = H 4'-Methoxy coumestrol: R:CH
3
OR 2
Genistein; R1=0H R2=H Biochanin A: RfOH R2:CH3 Diadzein : R1 = H R2=H Formononef1n : R1 = H �=CH3

4 .
(ii) Factors Influencing Phyto-oestrogen Levels
Investigations have shown that there is considerable variation in
the oestrogenic potency of pasture plants and that this is influenced by
both environmental and genetic factors .
GENETIC : Bradbury and White (19 54) tabulated over 50 different
species of plants reported as oestrogenic . There is variation between
species in the ratio of coumestans to isoflavones . The coumestans are
the dominant source of oestrogenicity in lucerne (Bickoff et al . , 19 6 2 ),
ladino clover (Guggolz et al . , 19 61), the annual medics (Bennett et al . ,
196 7b; Francis and Millington, 19 65a; Millington et al . , 19 64a) and
white clover (Wong et al . , 19 71) . The isoflavones account for most of
the oestrogenicity of subterranean clover (Millington et al . , 19 64b) and
red clover (Wong, 196 2 ) .
Wide variations in oestrogen levels are also evident between
varieties and between lines within a species; in lucerne (Francis and
Millington, 19 6 Sa; Stob et al., 19 5 7 ; Stuthman et al. , 19 6 7 ),
subterranean clover (Francis and Millington, 19 64; Davies and Dudzinski,
19 6 5 ) and red clover (Francis et al. , 19 6 7 ) . Francis and Millington
19 6 Sb) treated subterranean clover seed with a mutagenic agent and
produced a number of mutant plants of very low oestrogenicity .
ENVIRONMENTAL: Nutrient deficiencies influence the oestrogen
content of pasture plants . Oestrogenic activity has been shown to
increase significantly when phosphate deficiencies occur in subterranean
clover (Alexander and Rossiter, 19 52) and in red clover (Rossiter and Beck,
19 6 6 b; Butler et al . , 1967) . Concentrations of formononetin were most
affected although diadzein and biochanin A levels also increased (Rossiter
and Beck, 1966b) . Severe deficiencies in sulphur (Rossiter, 1970;
Rossiter and Barrow, 1972), and in nitrogen (Rossiter, 19 6 9) can almost
double formononetin levels .
Pieterse and Andrews (1956) and Kohler and Bickoff (1961) found
that coumestrol levels increased proportionally with maturation of lucerne

5 .
plants. Francis and Millington ( 1965a) demonstrated a similar trend
in annual medics .
Oestrogenicity is reported to fall dramatically after the harvesting
and drying of subterranean clover (Francis and Millington, 1965c), and
of lucerne (Bickoff et al. , 1959 ; 19 60a; Livingstone et al., 196 1) .
Commercial dehydration has a similar effect (Francis and Millington,
19 65a). In contrast, lucerne silage is more potent than the green
lucerne from which it is produced (Pieterse and Andrews, 1956) .
There are conflicting reports on the effects of high temperatures
( Coleman et al . , 1965; Loper and Hanson 19 64; Rossiter and Beck, 196 6a),
season (McLean, 19 67; Squires, 19 66) and location (Millington et al . ,
1964a) , on oestrogen levels of pasture plants .
Bickoff et al . , (1960b) noted that a disease-i�fected sample of
white clover contained considerably more oestrogenic activity than
comparable disease-free samples. Subsequently, a strong correlation
has been reported between pathogenic attack of lucerne and of white
clover, and coumestrol levels (Bennett et al., 19 6 7 a; Wong and Latch,
197 1). Two leaf-spotting organisms shown to be implicated are
Lepto�aerulina briosiana (Loper and Hanson, 19 64), the fungal organism
of 'Pepper spot' disease; and Pseudopeziza medicaginis ( Loper and
Hanson, 19 64), the c2usal organism of 'common leafspot' . Loper et al . ,
(19 6 7 ) found that coumestrol content was very high in the necrotic lesions
of infected leaves, and that selection of plants for disease resistance,
or the use of fungicidal agents, markedly reduced coumestrol levels .
Infestation of lucerne by aphids has also been shown to cause a build-up
of coumestans ( Hanson et al . , 1965; Loper, 1968) .
The accumulation of coumestrol following pathogenic attack may be
implicated in some non-specific disease resistance mechanism . There is
considerable data showing that phenolic compounds are formed in plants
subjected to pathogenic attack or physical damage (Farkas and Kiraly, 19 62).
The 'Phytoalexin Theory' ( Ingham, 1972) proposes that a group of phenolics ·

6 .
theBhY.to-alexins - which exhibit antifungal properties, are produced
by plant cells as a result of pathogenic attack . Coumestans appear not
to have antifungal properties, but they are structurally similar to
Fhyto-alexins. It is suggested that both these compounds arise from a
common biosynthetic pathway (Ingham, 1972) .
In summary, it can be stated that although several environmental
factors may contribute to wide variation in the oestrogenicity of
pasture plants, the presence of pathogenic disease is the single most
influencing factor .
(iii) The Metabolism of Phyto-oestrogens in Sheep .
When considering the biological effects of phyto-oestrogens, of
particular importance are the metabolic changes that they undergo within
the animal .
The presence of a microsomal enzyme system in a number of animal
species, including sheep, which catalyses the demethylation of iso-
flavones has been reported (Nilsson, 196 3 ) . It was demonstrated that the
major reaction occurring to biochanin A and formononetin incubated in
sheep rumen liquor, is demethylation to genistein and diadzein
respectively (Nilsson, 19 61) . There was n o evidence to suggest more
extensive metabolism in vivo than was demonstrated in vitro (Batterham
et al . , (1965) . Subsequent investigations using radioactive isotopes
14 ( C) confirmed that the products of metabolism of formononetin were
diadzein and 7: 4'-dihydroxy isoflavone (equol) (Nilsson,et al . , 19 6 7;
Batterham �. , 19 71) . About 70 percent of the formononetin ingested
by sheep is converted to urinary equol, together with a small amount of
diadzein, desmethylangolensin (Batterham et al . , 1971), 0-methyl equal
and angolensim (Cox and Braden, 1974). The pathways of formononetin
metabolism in sheep are shown in Fig. 1-2.
Formononetin is of negligible oestrogenic potency if administered
to sheep via intramuscular or intravenous pathways, whereas a much greater
oestrogenic activity is apparent if administered intra-rumina�ly

/. I
Flg. 1·2 THE METABOLISM OF FORMONONETIN IN SHEEP
/
CH HO OH 3
Angolensin
CH HO OH 3
HO
Desmethyl Angolensin
HO
0 Formononetin
0 Diad zein
Equal
OH
OH
--•major pathway
c::===:> minor(or possible) pathway

7.
Braden et al., 1 9 67). In contrast, equol proves to be oestrogenically
active when administered intramuscularly (Shutt and Braden, 1 968).
Furthermore high levels of equol are excreted in the urine of sheep
grazing oestrogenic clovers (Shutt et al., 1 9 6 7; 1 9 70). Equol is the
predominant phyto-oestrogen 1n digesta and excretia of sheep grazing
clovers with high formononetin content (Shutt et al., 1 9 70). Formononetin
consequently is now considered to be the most important isoflavone 1n
terms of oestrogenic activity, although it has little oestrogenic
activity itself. Differences in oestrogenicity between strains of
subterranean clover have been related to their formononetin content
(Lindsay and Francis, 1 9 6 8; Davies et al., 1 9 70). 0-methyl equol and
desmethyl angolensin are somewhat less oestrogenic than equol (Micheli
et al., 1 9 6 2 ).
Little is known of the metabolism of coumestans in sheep. The
demethylation of 4'-methoxy coumestrol to coumestrol has been demonstrated
� � (Shutt et al., 1 9 6 9 ) and in vitro ( Adler and Weitzkin-Neiman,
1 9 70).
Braden et al., (1 967) were unable to find any obvious metabolic
products in the urine or plasma of sheep given coumestrol intra-ruminally.
Cayen and Common (1 9 6 5), feeding tritiated coumestrol to chickens, were
unable to detect any radioactive ur1nary equol, suggesting different metabolic
pathways for the coumestans than for the isoflavones.
Less than 1 percent of the daily intake of phyto-oestrogens are
excreted as such in the faeces or urine ( Shutt et al., 1 9 70). The phyto-
oestrogens and their metabolites are absorbed, largely from the rumen,
and circulate in the blood. Plasma of sheep grazing oestrogenic clover
contains water-soluble conjugates of diadzein, genistein and biochanin
A - mainly in the form of glucuronides ( Shutt et al., 1 9 6 7; Lindner,
1 9 6 7; Beck and Knox, 19 7 1 ).
Plasma from sheep fed synthetic coumestrol or medic hay, contains

8 .
water-soluble conjugates of coumestrol and free coumestro� but 4 -methoxy
coumestrol is not present (Shutt et al . , 19 6 9 ) .
Levels o f conjugated phyto-oestrogens, both isoflavones and
coumestans, are considerably higher than levels of free phyto-oestrogens
(Wong and Cox, 19 72). As well as conjugates of glucosiduronates,
relatively high concentrations of sulpho-conjugates have been isolated
from sheep plasma (Wong and Cox, 1971).
(iv) Phyto-oestrogens and Reproductive Failure in Sheep
(a) Isoflavones
CLOVER DISEASE - The classical infertility syndrome associated
with ewes exposed for prolonged periods to oestrogenic subterranean clover
pastures, is a permanent depression in fertility . This infertility
syndrome, which became known as 'clover disease', was first reported by
Bennetts, Underwood and Shier in 194 6 . They described dramatic decreases
in lambing percentages (by up to 90%), a high incidence of dystocia,
uterine prolapse months after parturition or even in unbred ewes, and
copious milk secretion in virgin and non-pregnant ewes (Bennetts et al. ,
194 6; Bennet, 19 6 6 ) . In wethers, enlargement of the teats and induction
of lactation is common; and in ewes, the pathological condition, cystic
glandular hyperplasia, is almost always found in the cervix and uterus,
(Bennetts et al. , 194 6 ; Barrett et al . , 1961) . 'Clover disease' has
a high degree of permanence, persisting for periods of several years, if
not for the lifetime of the ewe, after the ewes are removed from oestrogenic
pastures (Schinckel, 194 8; Underwood and Shier, 1951; Barrett, et al. , 196 5;
Turnbull et al . , 196 6) .
Subsequent investigations have indicated that the main cause of
infertility is the failure of ova to be fertilized (Turnbull et al. , 1966;
Lightfoot et al. , 19 6 7; Kaltenbach and Davies, 1970; Lightfoot et al. ,
1974 ; Wrath and Lightfoot, 1976) . The establishment of a reservoir
of spermatozoa in the cervix immediately after mating, is necessary for

9 .
normal fertility in sheep ( Mattner, 196 6), and sperm movement within
the tract of 'clover disease' effected ewes is impaired . The numbers of
sperm within the cervix (Turnbull et al., 19 6 6 ; Lightfoot et al . , 19 6 7;
Kaltenbach and Davies, 1970; Lightfoot et al . , 1974), within the
fallopian tubes (Turnbull et al . , 1966; Lightfoot et al . , 19 6 7;
Kaltenbach and Davies, 1970), and adhering to the zona pellucida of
recovered ova (Lightfoot et al . , 1974 ; Wrath and Lightfoot, 1976), are
much lower than in 'normal' animals .
The rheological properties of cervical mucus are important in the
establishment and retention of a population of sperm in the cervix
of the ewe ( Gibbons and Mattner, 196 6 ), and the visco-elasticity of
cervical mucus is decreased in ewes affected by 'clover disease' ( Smith,
1971; Adams, 1976a) . Adams (1977) showed that spinbarkheit of cervical
mucus could be related to lowered reproductive performance of ewes
exposed to oestrogenic pasture.
Evidence for a higher than normal incidence of embryonic mortality
(Turnbull et al . , 19 6 6 ; Kaltenbach and Davies, 1970), diminished ovarian
activity ( Lightfoot et al . , 1967; Hearnshaw et al. , 1972; Adams, 1973;
1976 b), anovulatory oestrus (Firth et al . , 1977) and abnormal cycle
lengths (Underwood and
equivocal .
Shier, 1951; Obst and Seamark, 1970) is
TEMPORARY INFERTILITY - When ewes grazed oestrogenic red clover over
the mating period only, a depression in the incidence of multiple births
and in conception rates was evident ( Morley et al . , 1964; Clark, 1965) .
However, fertility and fecundity were restored to normal levels within
five weeks of removal of ewes from oestrogenic pasture (Morley et al. ,
19 6 6 ) . Chang (1963) noted a similar transient loss o f fertility.
There is conflicting evidence as to whether the depression of
multiple births is due to a depression in ovulation rate. Morley et al . ,

( 1964) and Holst and Braden, ( 1972) found no evidence of depressed
ovulation rates . However, recent work has shown ovulation rates to be
depressed in ewes mated on oestrogenic subterranean clover ( Lightfoot
and Wrath, 1974; Wrath and Lightfoot, 19 76) .
( b) Coumestans
In the early 1960's Coop and Clarke first reported a depression in
the lambing percentage of ewes grazing lucerne at the time of mating.
A similar response was reported in ewes mated on white clover ( Sanger
and Bell ( 1959 ) . Coop and Clarke ( 1960) noted a dramatic decrease
in twinning rate and a small, non-significant increase in the incidence
of barrenness .
A number of subsequent trials have confirmed their f indings,
showing reductions in lambing percentages of up to 4 0 percent (Kelly
10 .
et al . , 19 76a; McCutcheon, 1976; Scales and Hoss, 1976; Scales et al . ,
1977). These trials are summarized in Table 1-2.
It was established that the decrease in lambing percentage, was due
to a decrease in the number of multiple births and not to increased
barrenness ( Scales and Moss, 1976; McCutcheon, 1976; Scales et al. ,
1977) . Furthermore, this depression in multiple birth rate was related
to a reduction in ovulation rate and not to increased embryonic mortality
( McCutcheon, 1976; Jagusch et al. , 1977; Scales et al. , 1977).
It is apparent that ewes of a higher inherent ovulation rate are
most affected by coumestans ( See Table 1-2) .
There have been conflicting reports on the ability o f the coumestans
to inhibit or delay oestrus, and to induce anovulatory oestrus or
follicular abnormalities. Kelly et al . , ( 1976a) reported a high
incidence of anovulatory oestrus and prolonged oestrus cycles, when they
administered extremely high levels ( c 1000 ppm) of coumestans . Moderately
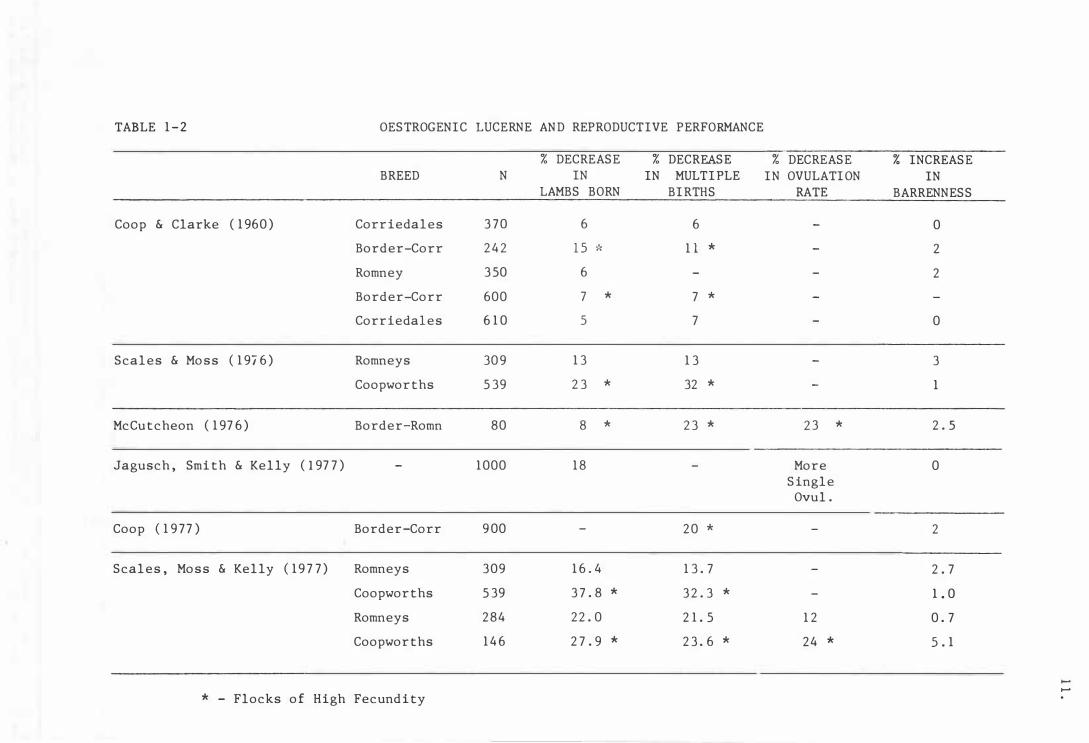
TABLE 1-2 OESTROGENIC LUCERNE AND REPRODUCTIVE PERFORMANCE
% DECREASE % DECREASE % DECREASE % INCREASE BREED N IN IN MULTIPLE IN OVULATION IN
LAMBS BORN BIRTHS RATE BARRENNESS
Coop & Clarke (1960) Corriedales 370 6 6 - 0
Border-Corr 24 2 15 -le 1 1 * - 2
Romney 350 6 - - 2
Border-Corr 600 7 * 7 *
Corriedales 610 5 7 - 0
Scales & Moss (1976) Romneys 309 13 13 - 3
Coopworths 539 23 * 32 * - 1
McCutcheon (1976) Border-Romn 80 8 * 2 3 * 2 3 * 2 . 5
Jagusch, Smith & Kelly (1977) - 1000 18 - More 0 Single
Ovul .
Coop (1977) Border-Corr 9 00 - 20 * - 2
Scales, Moss & Kelly (1977) Romneys 309 16.4 13 . 7 - 2 . 7
Coopworths 539 37 . 8 * 32. 3 * - 1.0
Romneys 284 22. 0 21. 5 12 0 . 7
Coopworths 146 27 . 9 * 23 . 6 * 24 * 5 . 1
...... ......
* - Flocks of High Fecundity

12 .
high levels of coumestan,such as occur in field conditions (100-400ppm),
apparently do not induce such abberations (Scales et al . , 1977) .
(B) THE MECHANISM OF ACTION OF OESTROGENS
The current model for oestrogen action in target tissues indicates
that after entering a target cell, the hormone binds with high affinity
to a cytoplasmic protein - the 'oestrogen receptor' - to form a complex .
This complex undergoes rapid translocation to the nucleus, where it
probably interacts with chromatin acceptor sites (Gorski and Gannon,
1 9 76). Considerable evidence suggests that the presence of the receptor-
oestrogen complex in the nucleus is a prerequisite for oestrogen-induced
stimulation (Jenson and De Sombre, 1973 ) . The nature of the oestrogen-
induced biosynthetic events, depends not only on the quantity of receptor
complexes within the nucleus, but also on the length of time the complexes
stay within the nucleus (Anderson et al., 1 975; Lan and Katzenellenbogen,
1976). Thus a single injection of oestriol, a weak oestrogen, can
initiate only early biosynthetic events (0-6 hours) and is ineffective
in stimulating later events (20-4 8 hours) , presumably because of its high
rate of dissociation from the receptor and its rapid clearance (Anderson
et al. , 1975) . However, oestriol can be as effective as oestradiol
in stimulating later responses if high circulating levels are maintained
(Clark et al . , 1977) .
When oestriol is administered in acute doses, it acts as an oestrogen
antagonist . It appears to compete with oestradiol for receptors (Clark
et al. , 1977). Oestrone apparently competes with oestradiol also
(Wise and Payne, 1975).
A number of non-steroidal compounds act as anti-oestrogens, preventing
the full expression of oestrogenic steroids on target tissues. Anti-
oestrogens bind to oestrogen receptors (Capony and Rochefort, 1977) and
induce translocation of receptors into the nucleus in the absence of
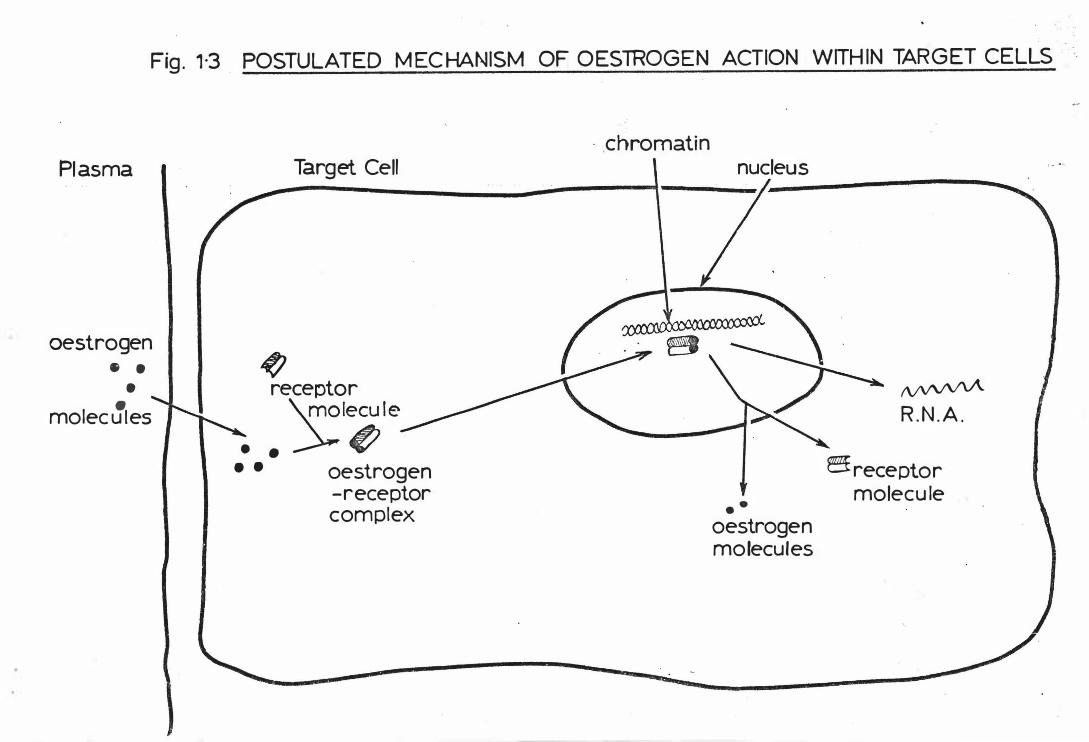
Pl asma
oestrogen 5 •
• • molecules
Fig. 1·3. POSTULAT ED MECHANISM OF O ESTROGEN ACTION WITHIN TARGET C ELLS
Target Cel l
� receptor \.molecule
. . �� • • oestrogen
-receptor c omplex
· .chromatin nucleus
•• oestrogen molecules
N'fVV'A R.N.A .
�receptor molecule

oestrogen, while inhibiting the translocation of receptor-oestrogen
complexes ( Ruh and Ruh, 19 74). Receptor synthesis may also be
blocked ( Katzenellenbogen et al . , 19 7 7 ) .
Paradoxically, in the absence of oestrogens, anti-oestrogens can
stimulate some 'oestrogenic ' responses ( Fergusson and Katzenellenbogen,
19 7 7) . The effect of a n anti-oestrogen is not identical in all target
tissues, but differs between uterus, pituitary and hypothalamus
( Luine and McEwen, 19 7 7 ) .
The postulated mechanism of action of oestrogens on target cells is
outlined in Fig . 1- 3 .
( C) EXOGENOUS OESTROGENS AND REPRODUCTION IN THE EWE
( i) The Oestrous Cycle
13 .
Behavioural oestrus can be induced in anoestrous ewes by the
administration of exogenous oestrogens (Moore and Robinson, 19 57; Morley
et al. , 19 6 3 ; Piper and Foote, 19 6 8; Coding et al . , 19 70; Radford
et al . , 19 7 0; Scaramuzzi et al . , 19 7 1a) . The sensitivity of ewes to
exogenous oestrogen is related to plasma progesterone levels (Moore
and Robinson, 1957 ; Scaramuzzi et al . , 19 7lb), and to breeding season
( Reardon and Robinson, 1961; Fletcher and Lindsay, 19 7 1a) . The implication
of oestrogens with behavioural oestrus has been confirmed in experiments
involving the admin�tration of antisera to endogenous oestrogens .
Scaramuzzi ( 19 7 5) and Fairclough et al. , ( 19 7 6) prevented the occurrence
of behavioural oestrus by immunizing sheep against oestradiol . Cox
and Wilson ( 19 7 6) repeated this result with anti-oestradiol and anti
oestrone sera, but found that immunization against phyto-oestrogens did
not prevent oestrus.
Scaramuzzi et al . , ( 19 7 la) and Land et al. , ( 19 7 2) found a linear
relationship between the dose of exogenous oestrogen and the duration
of the induced oestrus. Land et al . , (197 2) suggested that a breed

14 .
difference in sensitivity to exogenous oestrogens exists; with highly
fecund breeds being more sensitive in relation to this oestrus response .
Scaramuzzi et al . , ( 1972) found a refractoriness to oestrogen, with ewes
showing a decline in response to repeated daily injections of oestradiol
benzoate. Morley et al . , (19 6 3) administered stilboestrol daily to
cycling ewes prior to and during mating, and reported decreased conception
rates, and an increase in the incidence of silent heats .
Oestradiol injections given after day 8 of the oestrous cycle, result
in premature luteolysis in intact ewes (Stormshak et al. , 19 69; Hawk and
Bolt, 1970; Bolt and Hawk, 197 1), but not in hysterectomized ewes
(Stormshak et al . , 19 6 9) . This luteolytic effect of oestrogen is possibly
mediated through the stimulation of uterine prostaglandin F2a
Cox et al., ( 19 74) and Barcikowski et al . , ( 1974) reported increased
PG F2a secretion coinciding with endogenous oestrogen peaks on days 14 - 15
of the oestrous cycle . However, there was no coincidental PGF2a rise with
oestrogen peaks on days 4-5. Increases in PGF2a secretion following the
infusion of physiological doses of oestradiol into uterine plasma, have
been noted (Barcikowski et al . , 1974 ; Ford et al. , 1975).
Obst and Seamark ( 1970) reported that plasma progesterone levels in
sheep grazing oestrogenic clover, fell to 'oestrus' levels earlier than
in sheep grazing pasture (days 14- 15 compared with days 17- 18) . This
supports the findings of Ligttfoot and Wrath (1974) who reported that corpora
lutea of clover-affected ewes weighed less than those of normal ewes.
However, Hearnshaw et al . , ( 1972) and Smith (1975) found clover infertile
sheep to have normal progesterone secretion patterns .
Recently Newsome and Kitts (1977) measured receptor-bound oestrogen
levels in ewes grazing lucerne and found that receptor-bound endogenous
oestrogen levels were lower and more uniform than those of ewes grazing
pasture.

15 .
(ii) Gonadotrophin Secretion
LUTEINIZING HORMONE - A relationship between steroid oestrogens and
the pre-ovulatory LH surge was suggested as early as 19 34 , when Hohlweg
found that oestrogen injections induced ovulation in rats . This
'Hohlweg effect ' is also evident in the ewe (Hammond, 1 94 5).
Moore et al . , (19 6 9) and Cox et al. , (19 71) demonstrated that the
pre-ovulatory release of LH during oestrus in the ewe is primarily a
response to high levels of circulating oestrogens the previous day.
However, although oestradiol injections administered to cyclic ewes on
days 3 -4 of the oestrous cycle induced a pre-ovulatory type LH surge,
injections of a similar dose level on days 10-12 did not (Bolt et al . ,
19 7 1). Much higher dose levels are required at this �ime (Howland
et al . , 1 9 7 1) . These variations in response to exogenous oestrogens
would appear to be related to plasma progesterone levels, however, there
are conflicting reports on the ability of progesterone to block this
oestrogen-induced LH surge (Yuthasastrakosolet al., 19 7 4 ).
This pre-ovulatory-type LH surge can also be induced in anoestrous ewes
(Goding et al . , 19 6 9 ; Radford et al . , 19 7 0; Beck and Reeves, 19 7 3)
and much smaller dose levels of exogenous oestrogens are required
(Howland et al . , 197 1 ) .
Exogenous oestrogens exert a somewhat more complex influence on
ovariectomized ewes . Ovariectomy, and the subsequent decrease in plasma
oestrogen levels, results in an elevation of basal LH levels in anoestrous
ewes (Reeves et al . , 1 9 7 2; Diekman and Malvern, 1 9 7 3) . The administration
of exogenous oestrogen to ovariectomised ewes induces a biphasic response
in circulating LH levels - an initial depression followed by a large
elevation, similar to pre-ovulatory levels (Radford et al. , 196 9; Goding
et al . , 1 9 7 0; Scaramuzzi et al . , 1 9 7 1 b; Howland and Palmer, 1 9 7 3) .
Cen�ral nervous system blockade by sodium pentobarbitone anaesthesia,
prevents only the second phase of the LH response (Radford and Wallace,
1 9 7 4) . This indicates a non-nervous site for the inhibitory action, and a

16 .
CNS site for the stimulatory action on LH release.
Not only is there variation in the sensitivity of ewes to exogenous
oestrogen between the breeding season and the anoestrotts period, but there
is also variation in sensitivity between breeds . Finnish Landrace
ewes are less sensitive than Scottish Blackface ewes ( Land et al . , 19 76)
to the negative-feedback effects of oestrogen, and this is suggested as
a possible reason for variation in ovulation rates between
breeds.
The ingestion of phyto-oestrogens disrupts these responses to
exogenous oestrogens. Hearnshaw et al . , (1972) found that ovariectomized
ewes affected by 'clover disease' failed to produce any LH response to
oestradiol injections. Synthetic Gonadotrophin releasing hormone ( Gn-RH)
does evoke the pre-ovulatory type LH response ( Findlay et al . , 1973) .
This suggests that prolonged exposure to isoflavones impairs hypothalamic
receptors . Hearnshaw et al. , (1977) postulated that 'temporary infertility'
associated with isoflavones may also be related to the disruption of
normal LH secretion patterns . They found that feeding oestrogenic clover
to ovariectomized ewes, caused an initial depression then a rapid
elevation of LH levels . Thus the response to oestrogenic clover paralleled
the response obtained by administration of steroid oestrogens . There
are no reports on the effects of coumestans on LH secretion patterns .
FOLLICLE STIMULATING HORMONE - Unilateral ovariectomy of cyclic
ewes ( Findlay and Cumming, 1977) or bilateral ovariectomy of cyclic or
anoestrous ewes ( Salamonsen et al. , 1973), causes basal FSH levels to rise.
This suggests that basal levels of FSH are regulated by oestrogen through
a negative feedback system. As unilateral ovariectomy invokes a much
greater response in FSH than in LH levels ( Findlay and Cumming, 1977),
it is suggested that FSH is more sensitive to negative feedback effects
from the ovary than is LH - a situation which is also apparent during the

17 .
pre-ovulatory period in the intact ewe ( Salamonsen et al. , 1973; Baird and
Scaramuzzi, 1976 ) .
Infusion of oestradiol-178 into anoestrous ewes causes a dramatic
increase in FSH release, coincident with LH release ( Jonas et al. , 1973;
Salamonsen et al. , 19 73; Reeves et al. , 19 74 ; Pant, 1977) and
in a similar pattern of secretion to that which follows administration of
Gn-RH, except for an increased time lag ( Jonas et al . , 1973).
The addition of oestradiol to ovine pituitary cultures, even at
extremely low concentrations, causes a dramatic decrease in FSH
synthesis ( Miller et al., 1977a; Steinberger and Chowdhury, 19 77).
However, the adminstration of oestradiol in vivo did not alter pituitary
FSH levels ( Steinberger and Chowdhury, 1 977).
There are no reports on the effects of phyto-oestrogens on FSH
secretion patterns.
GONADOTROPHIN RELEASING HORMONE - In 1964 Bognadove postulated that
the variation in LH secretion throughout the oestrous cycle, was due to
a change in pituitary responsiveness to Gn-RH. Gn-RH induces a greater
change in serum LH ( Reeves et al., 1971a) and FSH ( Hooley et al. , 1974 ) during
an eight hour period on day 1, than at any other stage of the cycle .
However, some workers have failed to find the variation in response
( Foster and Crighton, 1974). A strong correlation between the oestradiol:
progesterone ratio and the LH response to Gn-RH has been demonstrated
( Thimonier et al . , 1974 ; Pelletier, 1 976) . When progesterone levels are
high, as in pregnancy, both FSH and LH responses to Gn-RH are reduced
( Chamley � ·, 19 74 a; 1974 b) . Similarly, daily progesterone injections
suppress FSH and LH responses to Gn-RH ( Hooley et al., 1974 ) .
The administration of exogenous steroid oestrogens also affects
pituitary responsiveness. Oestradiol benzoate admi�ration to anoestrous
ewes, increases the LH response to Gn-RH ( Reeves et al . , 1971b). More

precise studies have shown that oestrogens have a biphasic effect on
pituitary responsiveness. When given a few hours (4 -6) prior to Gn-RH,
oestrogens depress the LH response, but an amplified effect of Gn-RH
is observed if the time between oestrogen and Gn-RH treatments is
extended ( Pelletier and Signoret, 19 7 0; Libertum et al . , 19 7 4 ) . From
18 .
changes in the FSH : LH ratio, it would appear that the inhibitory effect
of oestrogen, influences FSH more than LH secretion ( Libertum et al . , 19 7 4 ).
( iii) The Reproductive Tract
CERVICAL MUCUS SECRETION - Copious mucus secretion by the ovine
cervix is generally attributed to oestrogen. Administration of oestradiol
benzoate to ovariectomized ewes increases cervical mucus secretion (Moore
and Robinson, 1957 ) and the response is linear (Allison, 19 7 2 ) . Rexroad
and Barb, ( 19 7 7 ) found that this increase in mucus production was not
accompanied by any change in spinnbarkheit, chloride content or percentage
dry matter .
The ingestion of phyto-oestrogens has also been shown to increase
cervical mucus production for both isoflavones ( Lindsay and Francis,
19 68; Smith, 19 7 1; Kelly et al. , 19 7 6 b) and for coumestans (Kelly et al . ,
197 6a) . Adams ( 1 9 7 6 a) disputed that an increase in the amount of mucus
produced did in fact occur, but that the visco-elasticity of the mucus was
decreased by phyto-oestrogens . He showed that 'clover disease' ewes
had lower spinnbarkheit (Adams, 19 7 7 ). The sensitivity of ewes to
exogenous steroid oestrogens is diminished by prolonged exposure to phyto
oestrogens (Adams, 19 7 8) .
OVA TRANSPORT AND FERTILIZATION - Exogenous oestrogens induce an
acceleration in ova transport through the fallopian tubes ( Chang, 19 6 6;
Restall, 19 6 6 ) . Holst and Braden ( 19 7 2) found that phyto-oestrogens
produced a similar effect .

1 9 .
The administration of oestradiol increases the ra�e and strength
of uterine contr actions mar kedly (Hawke, 1 9 7 5) and also increases the
number of contraction 'waves ' moving towards the oviduc� (Hawke, 1 9 7 4) .
This improves sperm transpor t within the female tract, increasing the
numbers of sper m found within the cervix , uterus and oviducts (Hawke
and Cooper , 1 9 7 5) . This is in contradict i on to the effects of the
isoflavones on sper m tr anspor t . In sheep affected by clover disease,
the number of sperm entering the cervix is considerably lower than normal
(Turnbull et al . , 1 9 66 ) . Apparently changes in the rheological properties
of cervical mucus which depress sper m transport, outweigh any advantages from
incr eased uter ine motility.
Exogenous oestrogens apparently have no direct effects on fertilization
or on embr yo survival (Hawke and Cooper , 1 9 7 6) .
(iv) Oestrogen Receptor s
Oestr ogen r eceptor levels are r eplenished in the cytoplasm of target
tissues by mechanisms which are par tly dependant on pr otein synthesis.
Oestrogens appear to be the most potent regulator of the dynamics of
oestrogen-r eceptor turnover (Cidlowski and Muldoon, 1 9 7 6; 1 9 7 8) .
Receptor concentr ations in ovine endometrium vary over the oestrous cycle.
The pre-ovulator y surge of oestradiol promotes maximal r eceptor synthesis
(Koligian and Stor mshak, 1 977b) , hence the concentr ation of receptors
is greatest at oestrus (Miller et al., 1 9 7 7 b) . The concentration of
receptors declines after oestrus, being lowest at the time of maximum
progesterone synthesis (Miller et al . , 1 977b) .
A single injection of exogenous oestradiol results in 6 0- 7 5 percent
of cytoplasmic r eceptors being translocated into the nucleus (Sutherland
and Mester, 1 976) , the degree of translocation differing between target
tissues (Morris, 1 9 7 6 ) . Continuous administration of exogenous oestrogen
incr eases the concentration of cytoplasmic oestrogen r eceptors (Koligian
and Stor mshak, 1 9 77a; Murphy et al. , 1 977 ) , but progesterone suppresses

2 0 .
this oestrogen-induced incr ease (Hseuh e t al . , 19 7 6 ) .
Shemesh e t al . , (197 2) d emonstrated that phyto-oestrogens also
bind to oestrogen receptors. Both coumestrol and genistein compete with
oestradiol for receptors. C hamley et al. , (19 74c) found a greater
concentration of oestradiol-receptor complexes in ' clover disease ' ewes.
This suggests that phyto-oestrogens are also able to ind uce the synthesis
of receptors .
( D) FOLLICLE MATURATION IN THE EWE
(i) Patterns of Follicle Growth and Atresia
Follicula r growth is d e scribed as a continuum . It proceeds at
all times, at all ages, uninterrupted by pregnancy, anoestrous or other
periods of non-ovulation (Peters et al . , 19 75; Matton et al . , 19 7 7 ) .
In the ewe, the numbers of follicular growth ' waves ' during the
oestrous cycle have been variously reported as one (Hutchinson and Robertson,
1966), two (Brand and De Jong, 19 7 3), and three or four (Smeaton and
Robertson, 197 1 ) . Recently Turnbull e t al . , (19 7 7 a) disputed the existence
of any ' wave s ' of fol�icular growth, and suggested that follicles are
growing and regressing as�chronously at any given time of the oestrous
cycle .
The initiation of growth of primordial follicles is a continuous
process in ewes (Peters et al. , 19 7 5), and the number of ' initiated '
follicles which finally ovulate is d etermined by the rate of atresia
d uring their growth phase (Hay and Moor, 19 7 5b; Turnbull et al . , 19 7 7 a) .
Turnbull et al. , (1977 a) estimated follicle growth rates by the
mitotic ind ex of granulosa and thecal cells . They found that irrespective
of the stage of the cycle , the growth rate was slow in follicles of up to
0 . 4mm d iameter, after which it accelerated until it reached a maximum in
follicle s of about 0 . 7mm d iameter. This fast rate was maintained until

the follicles r eac hed a diameter of about 2mm and then declined by about
50 per c ent . In sheep, those follicles destined to ovulate would have
a diameter greater than 0.5mm on day 6-7 of the cyc le, although not all
follicles of this diameter would ovulate . In Merinos with an ovulation
rate of 1 to 2, about 3 -4 follicles enter the rapid growth phase each
day (Turnbull et al . , 19 7 7 a) . In single ovulator breeds, follic l es of
a diameter gr eater than lmm have a slower gr owth rate than do follicles
of a similar size range in multiple ovulator breeds (Turnbull et al . ,
21 .
19 7 7 b) . Thus for the same total number of follicles present, the multiple
ovulator will have a greater number capable of ovulation, when compared
with a single ovulator ( Dufour and Matton, 19 7 7 ; Dufour, 19 7 6). The
number of follicles whic h ac tually do ovulate, is apparently determined
within the 3 days prior to oestrus ( Land, 19 7 3 ; Findlay and Cumming,
19 7 7 ; Bherer et al . , 19 7 7).
( ii) Hormonal Contr ol of Follic le Growth and Atresia
GONADOTROPHINS - The initiation of growth of primor dial follicles
appears to be independant of gonadotrophin levels ( Lunenfield et al. ,
19 7 5; Peter s et al., 19 7 5). Two intra-ovarian fac tors have been
demonstr ated to influenc e the initiation of follic ular growth; ( a)
the size of the pool of non-growing follicles - a reduction in pool size
causes fewer follic les to start growing (Krar up et al . , 1 9 69) ( b) the
degeneration of large follicles - large follicles apparently release a
substanc e that suppresses growth initiation ( Peters et al. , 197 3).
There is evidence that the administr ation of gonadotrophins ( PMSG)
to ewes, will increase the number of ' initiated ' follicles entering the
rapid growth phase, prevent atresia in large follicles ( Hay and Moor,
1 9 7 5b; Turnbul l et al. , 19 7 7 a), and inc rease ovulation rates ( Cumming
and McDonald, 19 6 7 ) . Conversely, the reduction of FSH and LH levels by
the administration of antiser a, ac utely prevents the development of

small follicles to Graafian follicles (Welchen and Dullaar� 19 7 6), and
even the temporary absence of LH results in f ollicle atresia (Schwartz,
19 7 4 ).
Although a direct relationship has not been established between
FSH levels and ovulation rate (Findlay and Gumming, 19 7 6a), there is
considerable indirect evidence for such a relationship . Unilateral
ovariectomy results in compensatory fol licular growth and in ovulation
rates, in the remaining ovary (Sundaram and Stob, 19 6 7 ; Mallampati and
Casida, 19 70; Dufour et al . , 197 1a; Land, 19 73; Findlay and Gumming,
19 7 7 ) . This compensatory growth is apparent only if hemicastration is
carried out prior to 3 days before the onset of oestrus. There is a
transient increase in FSH, and to a lesser degree in LH, levels after
hemicastration. which coincides with falling oestradiol levels (Ramir ez
and Sawyer, 19 74; Findlay and Cumming , 19 7 7 ). Oestrogen replacement
therapy suppresses both the rise in FSH levels and compensation in
follicular growth and ovulation following hemicastration (Ramirez and
2 2 .
Sawyer, 19 74). 'Clover disease ' ewes also fail to show expected ovarian
compensatory hypertrophy (Adams, 19 7 6b).
The increase in ovulation rates in ewes fed lupin grain (Lightfoot
et al . � 19 7 6) is also accompanied by increased FSH levels (Brien et al . ,
1 9 7 6 ) .
Trounson et al . , (197 4 ) reported that lambs selected on multiple
birth characteristics of their dam, had a greater number of primordial
follicles than did control lambs . They suggested that this may be related
to higher endogenous gonadotrophin levels . However, Land et al . , (19 7 3)
and Echter nkamp and Laster, (197 6) were unable to find any relationship
between ovulation rates and endogenous LH levels. Bindon et al . , ( 19 7 1)
reported a greater ovarian response to exogenous gonadotrophins by high
fecundity ewes than by low fecundity ewes . Land et al . , (19 7 2, 19 7 6)
and Land ( 19 7 6 ) have shown that high fecundity breeds are less sensitive
to the effects of exogenous oestrogens on gonadotrophin release.

23 .
STEROID HORMONES All follicles, including those in various stages
of atresia, secrete a range of steroids including oestrogens, androgens
and progestins (Seamark et al ., 19 74 ) . The amounts and precise pattern
of steroid secretion of an individual follicle varies according to its
developmental stage . The smaller follicles produce predominantly
androgens, whereas the larger follicles produce both oestrogens and
androgens (Seamark, 19 7 8 ) . Atretic and non-atretic small follicles are
equally active steroidogenically .
Ovarian oestrogen secretion rises from baseline levels on day 13 of
the cycle to a peak on day 15, and then declines rapidly at oestrus
(Moor et al . , 19 7 1; Moor 19 74 ; Edwards, 19 7 4 ) . These high levels of
oestrogens are secreted almost entirely by the largest one or two non
atretic follicles (Bj ersing et al . , 19 7 2 ; Moor, 19 7 3 ; Baird et al . , 19 7 5 ;
Hay and Moor, 19 7 5 b) . The enzyme 6 5 3 8 hydroxysteroid dehydrogenase
plays a key role in oestrogen synthesis (Moor et al., 197 1) and although
it ' s activity appears to be confined to the thecal cells (Hay and Moor,
19 7 5a), interaction between thecal and gr anulosa cells is apparently
required for oestrogen production (Moor, 19 7 7 ). It is not known what
initiates this surge in oestrogen production, but exogenous gonadotrophin
(PMSG) induces increased activity of this enzyme (Moor et al . , 19 7 1) .
The pre-ovulatory rise in LH terminates this surge in oestrogen production
(Moor, 19 74 ) . The Graafian follicle is extremely sensitive to minor
changes in basal LH concentrations (Baird et al . , 19 7 6 ) and variation in
the secretion of oestradiol- 17 8 probably reflects the response to small
pulses in LH (Wheeler et al . , l 9 7 7 a).
As a consequence of the pre-ovulatory changes in circulating gonado
trophins, oestrogen production by the large non-atretic follicles is
reduced and replaced by an increase in the release of androgens and
progestins . Twelve to 1 8 hours prior to ovulation, the large follicles

24.
produce little o r no oestrogen, but large amounts of pro gesterone
and 2 0« pregn-4 -en-3-one ( Seamark, 1978) . It has been suggested that
progesterone may have an effect on the ovary independent of its influence
via the hypothalamo-pituitary axis ( Rexroad and Casida, 1977). Du four
et al., ( 19 71b) reported that follicle growth was more rapid in ovaries
that co ntain a corpus lut eum . Inj ection o f progesterone int o one ovary
of anoestrous ewes, with concurrent systemic administration of PMSG,
results in more rapid follicular gro wth, although o estro gen secretion is
unchanged, ( Rexroad and Casida, 1977) and in a greater number of fo llicles
and corpora lutea in the inj ected, than in the co ntrol ovary ( Harned and
Casida, 1971) . Progestero ne may act to alter time-dependent changes in
follicle size, thus enabling some follicles to gro w while o thers undergo
atresia.
HORMONE RECEPTORS - It has been proposed that the selectio n of those
follicles destined to ovulate, is a co nsequence o f both sequential changes
in follicular hormone receptor levels and in circulating pituitary
gonado tro phin levels ( Carson et al., 1978) .
As follicular growth proceeds, FSH receptor levels decrease and LH
recepto r levels increase within granulosa cells ( Channing, 1970; Carson
et a l . , 1978) , but remain unchanged within thecal cells (Weiss and Armstrong ,
1977; Carso n et al . , 19 78) . FSH has been sho wn to increase numbers o f
FSH receptors; o estradiol to increase oestradiol receptors; and FSH
and oestradiol in concert to increase LH receptors, in granulosa cells
( Richards and Midgley, 1976). Termination of follicular growth by
LH-induced luteinizatio n is associated with the loss of receptors for FSH
and oestradiol, and the subsequent increase in LH receptors (Richards
and Midgley, 1976 ) .

2 5.
(E) OVULATION IN THE EWE
(i) Factors Influencing Ovulation Rates
Heape (189 9) first reported that heavy ewes produced more lambs than
light ewes. This is related to a higher production of ova (Wallace, 19 61;
Allen and Lamming, 19 6 1; Killeen, 19 6 7 ; Alli son, 19 68). This correlation
between bodyweight and ovulation rate has been described by Edey (19 68)
who found a linear relationship within a particular weight range.
Ducker and Boyd (19 7 7 ) demon strated that it is the body condition element
of ewe liveweight, not body size, that influences ovulation rates.
However, Cumming (19 7 7 ) showed that liveweight alone was as accurate
a pred i ctor of ovulation rate as both liveweight and bod y condition.
S everal workers have reported evidence of a d ynamic effect of flushing,
independen t of bod ywei ght, on ovulation rate (Coop, 196 6a; 19 6 6b;
Mcinnes and Smi th, 19 6 6; Killeen 19 6 7) although others have found no
effect (Hulet et al. , 19 68; Fletcher, 1 9 7 1 ; Cumming, 19 7 7 ) .
There is consid erable variation i n fecund i ty between breeds and between
strain s within a breed. Ewes of higher fecundity (Trounson _et al.,
19 7 4 ) and their daughters (McDonald and Chang, 1 9 6 6) ovulate a greater
number of follicles, an d a higher proportion of the Graafian follicles
that d evelop (Bradford et al., 19 7 1; Turnbull et al., 19 7 7 b). Fecund ity
and ovulation rates are not related to LH secretion throughout the
cycle (Echternkamp and Laster, 1 9 7 6 ) nor to LH peaks 1 Land et al . , 19 7 3) .
There i s some ind irect evid en ce that FSH levels around days 12 -14 are
important in determining ovulation rates at the subsequent oestrus,
(Cumming and McDonald, 19 6 7 ; Brien et al. , 19 7 6; Findlay and Cumming
19 7 6 b; 19 7 7 ) but there are no reports of a d irect relationship between
FSH levels and fecundi ty. Liveweight or flushing apparently d oes n ot
influence FSH levels (Findlay and Cumming, 19 7 6a).

2 6 .
Wheeler et al., (1 9 7 7 b) showed that mean secretion rates of
oestr adiol- 1 7 S are significantly higher in ewes with high ovulation rates,
and Land et al. , ( 1 9 7 6 ) found such ewes less sensitive to exogenous
oestradiol-induc ed depression of ovulation rates . Ewes of high fec undity
also show a greater r esponse to PMSG-induced superovul ation (Eastwood
and Mc Donald, 1 9 7 5).
It has been suggested that the length of the interval from the onset
of oestrus to the time of ovulation may influenc e ovulation rate.
Land (1 970) and Hanrahan and Quirke ( 1 9 7 5 ) showed a positive relationship
between the duration of oestrus and ovulation rate. Curnrning et al.,
( 1 9 7 3 ) demonstrated a high degree of constancy in the inter val from the
pre-ovulator y LH peak to ovulation. Lindsay et al., ( 1 9 7 5) found that
the presence of rams during oestrus induced ovulation earlier by advancing
the time of the LH surge. There appears to be a consider able variation
in the interval from the onset of oestrus to the LH surge (Lamming
�rs corn . , 1 9 78).
(ii) Postulated Mechanisms of Ovulation
The precise process, or processes, of ovulation r emain obscure
although considerable effort has been put into identifying them. Two
major types of ovulator y mechanisms have been suggested; those
involving ' enzyrnatic breakdown' of follicular tissue and those involving
the 'bur sting' of the follicle due to pressure changes .
There is substantial evidence suggesting that the collagenous
connective tissue of the wall of the Graafian follicle is degraded near
the time of ovulation (Espey, 1974 ) . Collagenolytic enzymes have been
detected within ovarian follicles; both neutral and acid proteases,
capable of degrading collagen, are produced in mature Graafian follicles
(Espey and Coons , 1 976).

2 7 .
Although the growth and maturation of follicles occurs without a
measurable increase in hydrostatic pressure (Ro dbard, 1 9 6 8 ) a follicular
contractile process has been demonstrated. Electron mic roscopy shows
many c ells in the theca externa of mature sheep follicles, with c ytoplasmic
filaments and dense bodies c harac teristic of smooth musc le cells (O ' S hea,
19 7 1) . Strip s of follic ular tissue exhibit classic al contrac tile
activity, (O ' Shea and Phillip s, 19 74) and autonomic innervation, both
adrenergic and cholinergic, is assoc iated with follicles (Bahr et al.,
19 74; Owman et al., 1 9 7 5 ; Walles et al., 19 7 6; 19 7 7 ) . Per fused
ovarian tissue has been shown to be elec tric ally active (Tojo et al.,
19 75).
It has been suggested that prostaglandin Fz� is involved in the
induction of ovulation (Tsafriri et al. , 19 72). Conc entrations of
prostaglandins inc rease within the developing follicle late in the
oestrous cyc le , most dramatic ally in those follic les whic h eventually
ovulate (Yang et al . , 19 7 4 ) . This rise i n prostaglandin levels is
apparently induc ed by the surge in endogenous oestradiol which oc curs
at days 14- 15 of the cycle, but not by earlier oestradiol peaks
(Barcikowski et al., 19 74 ; Cox et al . 19 74) .Infusion of physiologic al doses
of oestradiol into the uterus on days 14-15 will induce comparable
p rostaglandin peaks (Barcikowski et al., 19 7 4 ).
Systemic or intrafollicular injection of prostaglandin Fz� induces
ovulation (Phi et al., 1 9 7 7 ) and similarly p erfusion of follicles ' in
vitro' enhances ovulation (Hamada et al . , 1 9 7 7 ) . This induction of
ovulation is bloc ked by known prostaglandin Fz� antagonists suc h as
p rostaglandin Ez and indomethacin (Carlson et al. , 1 9 7 4 ) .
Y ang et al., ( 1 9 7 4 ) have suggested that prostaglandin Fz« may be
involved in the synthesis, release or ac tivation of collagenolytic
enzymes and Tojo et al., ( 1 9 7 5 ) has shown that prostaglandin F2�

2 8 .
accelerates the electrical activity of perfused ovarian tissue .
Intrafollicular inj ection of LH ( and in some experiments, of FSH)
will induce ovulation ( Jones and Nal bandov, 1972). Most evidence
indicates that LH is the sole agent required to trigger ovulation, while
both FSH and LH are required for the development, maturation and
l uteinization of follicular tissue ( Rondell, 1 974). It has been well
established that LH induces increased synthesis of cyclic AMP and
subsequently, increased protein synthesis ( Robison et al. , 1 971) .
Prostaglandins are capable of increasing cAMP formation ( Marsh, 1971) and
conversely anti-prostaglandin agents block the stimulatory action of LH
on cAMP synthesis (Kuehl et al . , 1970) . Protein synthesis appears to be
an essential part of the metabolic processes supporting ovulation, as
inhibitors of protein synthesis inhibit LH-induced ovulation (Lipner and
Greep, 197 1 ).
( iii) Blockade of Ovulation
It has been well established that indomethacin blocks ovulation in
ewes ( Carlson et al. , 1974; Armstrong et al . , 19 7 4 ; Lewis and Warren,
1975). Indomethacin blocks the LH-induced increase in prostaglandin
F2� levels in ovarian follicles (Armstrong et al. , 1974 ) and inhibits the
oestradiol-induced LH release in anoestrous ewes ( Carlson et al . , 1974).
Similarly the administration of inhibitory analogue s of LH-RH will also
effectively block ovulation ( Phelps et al . , 1977).
This complete blockade of ovulation is not apparent in ewes ingesting
coumestans . Oestrogenic lucerne reduces the number of ovulations, but
seldom prevents ovulation completely .

(F) THE PURPOSE AND SCOPE OF THE INVESTIGATION
29 .
It has been clearly established that the ingestion of oestrogenic
lucerne by ewes, depresses ovulation rates . Because of the important
r ole of the gonadotrophins in the maturation and ultimate ovulatio n of
ovarian follicles , it is possible that the coumestans could alter the
pattern of gonadotrophic secretion . FSH is invol ved in the development
of Graafian follicles, and LH facilitates the final stages of matur ation
and the induction of ovulation. The coumestan-induced depression of
ovulation rate may arise from a decreased pituitary output of these
gonadotrophins, or fr om changes in the pattern of their release, particularly
near the time of oestrus .
The present trials investigated the effects of oestrogenic lucerne,
fed over the oestrous cycle, on LH secretion patterns, and on the timing
of the pre-ovulator y LH-surge in r elation to oestr us and ovulation
(Experiment I) . Ovarian activity was recorded as a measur e of gonadotrophic
function . In Experiment II, the effects of oestrogenic lucerne on
ovarian activity, were studied when lucer ne was fed early or late in
the oestrous cycle, with the view of determining the critical time
involved in the depression of ovulation rates.

C H A P T E R I I
MATERIALS AND METHODS

30.
C H A P T E R I I
MATERIALS AND METHODS
(A) EXPERIMENTAL PLAN
Two separate trials were carried out to investigate the effects
of oestrogenic lucerne on cycling ewes . In the firs t experiment
plas ma gonadotrophin concentrations were measured over a complete
oestrous cycle, and in the second experiment the effects of coumes tan
intake either early or late in the oestrous cycle were recorded. The
experimental procedures and cale nder of events are set out in
FIGS. 2-1 and 2-2.
EXPERIMENT I - 22 mixed aged Romney ewes and 20 mixed aged Border
Leicester x Romney ewes were randomly as signed to two groups with equal
numbers of each breed. At the detection of oestrus following
synchro� ation, each group grazed either oestrogenic lucerne or non
oestrogenic ryegrass / clover pasture.
Blood samples were collected from all ewes by j ugular venepuncture.
Samples were taken daily for 1 7 days, from day 7 of experimental oestrous
cycle. On predicted day 16 of the cycle, blood samples were taken at
4 -hourly intervals until the detection of oestrus; then at 2-hourly
intervals for 24 hours and at 4 -hourly intervals for the subsequent
1 2 hours.
All ewes underwent laparotomy 3 days post-oestrus at which time
corpora lutea and s urface follicles were recorded.
Over the period of intensive blood sampling, the animals were kept
in s mall holding paddocks and yards adjacent to laboratory facilities
(Mas s ey University Fertility Centre) . Ewes in the lucerne group were
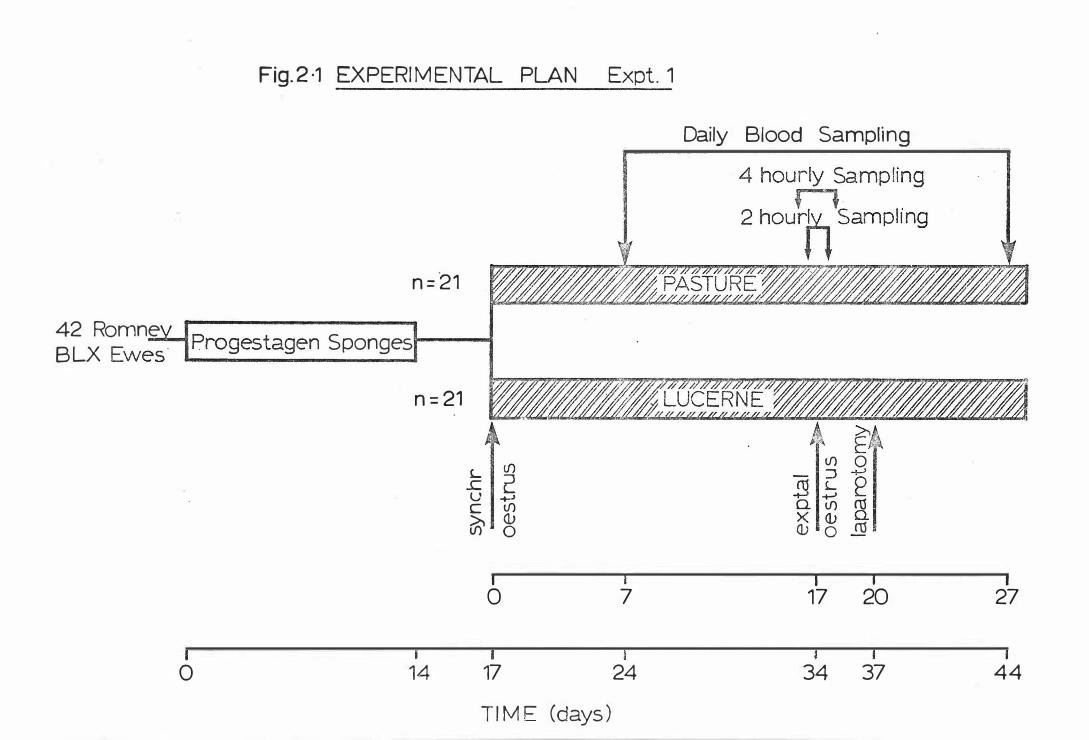
Fig. 2 ·1 EXPERI M ENTAL PLAN Expt . 1
Daily Blood Sampling
n = 21
42 Romne B LX Ewe�Progestagen Sponges
n = 21
L � � -5 b c (/) � gs
� E ..,
(/) 0 - ::J +-' � L e o. t) CO X <lJ Q. (lJ 0 CO
1 0 7 17 20 27
.-�--�-�-��1-----�---- I I -1 0 14 17 24 34 37 44
T I M F (days )
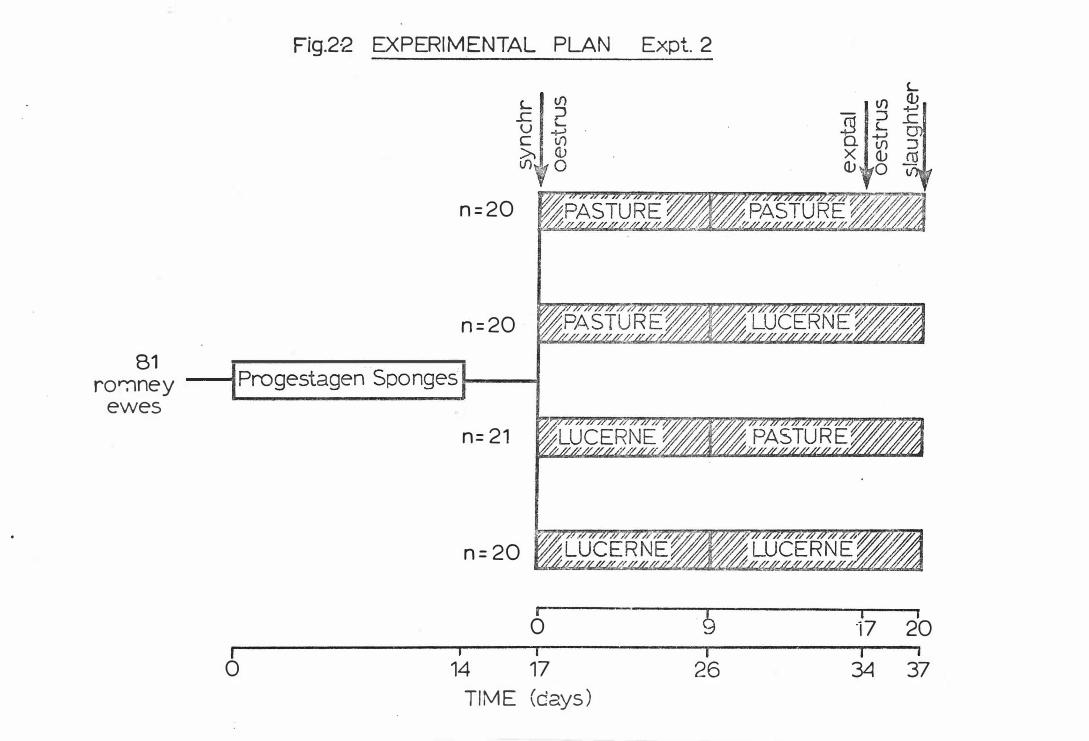
Fig.2·2 EXPERI M ENTAL PLAN
n = 20
ri = 20
L llf)
-5 2 c
�
Expt. 2
L - 1.1) Q;
_£912
1:
a +..J X 1.1) <lJ <lJ 0
81 ror.1ney IProgestagen Sponges & 1
ewes
n = 2 1
n = 2 0
.----0
.--------------,;-------r--- -·� 1 7 20
0 14 17 26 3L1 37
TI M E (days )

31.
f ed f resh lucerne cut daily f rom the crop .
EXPERIMENT II - 81 mixed aged Romney ewes were rando mly assigned to
f o ur groups. At the detection o f oestrus f ollowing synchronizatio n,
the gro ups were assigned to o ne o f four grazing regimes; past ure
thro ughout t he o estro us cycle, lucerne thro ughout the cycle, or these
treatments were interchanged at midcycle. In the lat t er two groups
treatments were changed on day 9 of t he experimental oestro us cycle.
All ewes were slaught ered 3 days po st -oestrus at the lo cal f reezing
works (Bort hwicks - CWS, Longburn) . Repro ductive tracts were recovered
within 1 5 minutes o f slaughter and individually identified.
(B) LUCERNE AND CONTROL PASTURES
The lucerne crop involved in t hese experiments consisted o f t wo
hectares o f Wairau lucerne on a Rangitikei Loamy sand . The paddo ck
was sub-divided into f our plo ts of approximately 0. 5ha . Experiment al
grazing co mmenced eight weeks after the lucerne had been previo usly
harvested, at which time plant heights ranged from 6 0 to 90cms .
Approximately 25 percent o f plants were in flower and there was
moderat ely heavy f ungal infectio n o f plant tissue (more t han 5 lesio ns
per leaf ) .
A three hectare ryegrass/ whit e clover pasture on a Manawatu Fine
Sandy Loam adj acent to the lucerne stand was grazed by contro l animals .
Leaf samples o f lucerne, and o f white clover, were taken weekly
and assayed for coumestan cont ent . This involved random select io n o f
material from 4 0 plants f ro m each plo t .
(C) ANIMALS
The animals utilized were 103 mixed-age Romney ewes and 20 mixed
aged F1
Border Leicest er X Romney ewes, drawn f rom several f lo cks

maintained on Massey University farms. All ewes were identified by
numbered brass eartags and by midside brands, which were applied at
the commencement of the trial .
32 .
All ewes were weighed prior to, and at the completion of oestrus
synchronization treatment, as well as at the completion of the trial.
Ewes in Experiment I were also weighed two weeks prior to synchronization
treatment . Attempts were made to carry out all liveweight recordings
at the same hour of the day, in order to mi nimise variations in gutfill .
(D) OESTRUS SYNCHRONIZATION
In order to synchronize oestrous cycles, all ewes were subj ected to
1 4 -day treatments with polyurethane intravaginal sponges containing
40mg of methyl-acetoxy-progesterone (M . A . P . ) . Vasectomized rams fitted
with Sire Sine harnesses were used to detect oestrus following pessary
removal .
(E) HORMONE ANALYSES
COUMESTAN S - Lucerne and clover samples were subjected to ethanol
extraction (Kelly, 1 9 7 6) and extracts were forwarded to Dr . R . W . Kelly,
Invermay Agricultural Research Centre, for analysis of coumestan
content.
PLASMA GONADOTROPHINS - Blood samples were assayed for ovine
luteinizing hormone by radioimmunoassay, following the double-antibody
technique of Niswender et. al. ,(19 6 9 ) , modified according to Barrell ( 1 9 7 6 ) .
This assay utilized the following materials: rabbit anti-ovine LH
serum (pool 15 courtesy Dr G . D . Niswender) , NIH-LH-Sl8 as assay
standards, and highly purified ovine LH (LER- 1 374A) for radioiodination.
Assay standards and samples were assayed in triplicate, with hypophysectomized

sheep plasma added to s tanda rd curve tubes . Mean assay s ens i t iv i ty
wa s 0 . 2 ng /ml , and the intra-and inter-assay coef f ic ien t s o f var ia t ion
( CV ) wer e 1 0 . 3 and 23% respec t ively .
At t he t ime o f planning o f these exper iments , a supply of FSH-
an t i s era became ava ilabl e . On the assump t ion tha t this a s say woul d
be i n opera t ion , F S H analyses were to be inc luded i n exper imen tal
proc edur es . However , with t he immuno chemical s ava ilab l e , b inding o f
iod ina t ed FSH wa s inadequa t e , and c ons equen t l y this a s say was omi t ted
from plasma analyses .
(F) CERVICAL MUCUS ANALY SIS
Cerv ica l mucus samples were taken f rom a l l ewes in Exp er imen t I
a t blood col l ec t ion time s over the oestrus pe riod . The rams used
for d e t e c t ion of oes trus were f i t t ed with canvas ' a pron s ' to prevent
penil e in tromiss ion and sub sequent con tamina t ion o f cervical mucus .
3 3 .
Mucus samp l e s were collec ted , wi th the a id o f a speculum , using absorbent
co t ton swab s .
Cerv ica l mucus samples were a s s essed for chlo ride ion con t ent by
the me thod o f Turnbull e t a l . , ( 1 96 7 ) . S i lver chroma t e was prec ip i t a t ed
onto s t rips of Wha tman No . 1 chroma t ogra phy paper , by impregna t ing the
paper , first with 0 . 2 7 5 N s i lver n i t rate and then with 0 . 1 7 5 po t a s s ium
chroma t e . Chl oride ions from the mucus samples produc e whi t e to bu f f -
coloured spo t s , which a r e compared to sodium chloride s tandards . Oes t rus
is a s so c ia t ed wi th a rise in chloride concentrations , and ovula t ion with
a sudden fal l in chloride l evel s .

(G) UTERINE AND OVARIAN WE IGHTS
34 .
Af t er co l l ec t ion a t slaugh t er , u t er i were separa t ed f rom the trac t
by tran s ec t ion immed ia tely an t erior to the c ervix , dried with paper
towe l s and weighed .
Ovaries f rom each ewe were weighed s eparately . Where par t o f the
uterus , or par t of an ovary , wa s l o s t a t s l aughter , tha t animal wa s
omi t t ed from t he analys i s .
(H) FOLL ICLE POPULATIONS
Af t er co l l ec t ion at s laughter , the ovaries wer e preserved in
Bouins f luid un t i l s ec t ioning and a s s e s sment o f fol l i c l e popula t ions .
To a s s e s s the t o ta l ovarian fo l l icular populat ion , a l l ovaries were
sec t ioned in to lmm s l ic e s . Fol l ic l e diame t er wa s c a l c u l a t ed a s the
mean of two measuremen t s taken a t r ight ang les to each o ther . Coun t s
were made o f a l l an tral foll i c l e s with a dimaeter o f 2mm o r greater .
( I ) REPRODUCTIVE TRACT HISTOLOGY
At the t ime of recovery of the reproduc t ive trac t , l cm s ec t ions
of vagina , c ervix , u t eru s and fal lop ian tube were taken and pres erved
in Bouin s Fluid for later his tological examina t ion .
Sec t ions o f 6� thicknes s , wer e taken o f all trac t s pec imens and
s ta ined wi th Ha ema toxyl in and Eo s in . The mean height o f ep i thel ia l
c el l s wa s cal culated fo r a l l sec tion s , by taking 2 5 s epara te mea surements
from each sec t ion .
(J) ANALY S I S OF DATA
Analyses o f d is c r e t e da ta - numb ers o f ovu lations and f o l l i c l e
2 po pulat ions - u til i z ed Chi s ta t is t ic .
Da ta invo lving :

Ewe l ivewe igh t s
LH conc entrat ion i n per ipheral p lasma
Ovar ian weights
Epi theaal c ell height s
were analysed by ana lys i s o f var iance ( Snedeco r and Cochran e , 1 9 6 7 ) .
3 5 .
Da t a from 8 ewe s were excluded from the ana lyses o f LH
conc en tra tions and ovulat ion ra t e s owing to either fa ilure o f o e s t r u s
synchroni za t ion ( 4 ewes) or failure t o detec t the pre-ovula tory LH peak
(4 ewes ) . Da ta f rom 9 ewes wer e excluded f rom Exp t . II due to fa ilure
o f hea t d e t ec t ion for a var iety o f r ea sons ; Sponges lo s t (4 ewes ) ,
bl ind tracts ( 2 ewes ) , s ilent hea t ( 2 ewes ) and pregnancy ( 1 ewe ) .
Data from a fur ther 1 2 ewe s were exc luded from reproduc t ive tract
analyses , due to cys t ic hyperpla s ia ( 1 ewe) , anovula tory o e s t rus (2 ewes ) ,
or incomp l e t e recovery o f the tract a t s laughter ( 9 ewes ) .

C H A P T E R I I I
RESULTS
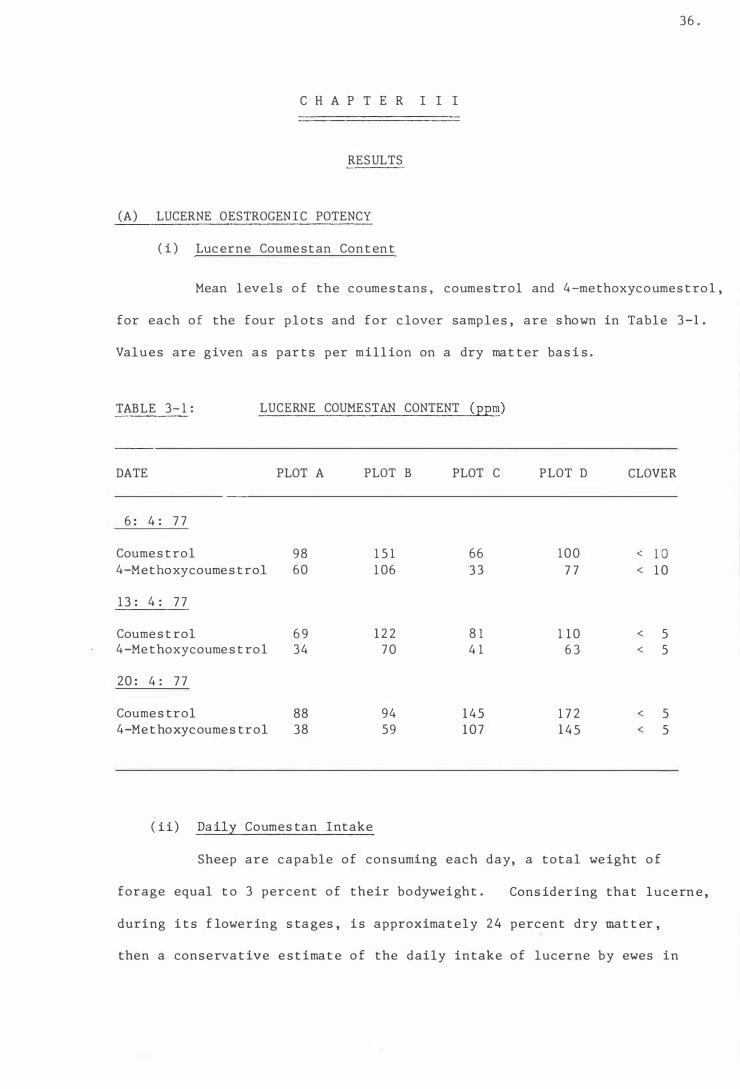
36 .
C H A P T E R I I I
RESULTS
(A) LUCERNE OESTROGEN IC POTENCY
( i ) Luc erne Coume stan Con t en t
Mean l evels o f the coume s tans , coumes trol and 4 -me thoxycoumes t ro l ,
for each o f the four p l o t s and for c lover samp l e s , are s hown in Tabl e 3 - 1 .
Values are g iven a s par t s per mill ion on a dry ma t t er ba s i s .
TABLE 3- 1 : LUCERNE COUMESTAN CONTENT (ppm)
DATE PLOT A PLOT B PLOT C
6 : 4 : 7 7
Coume s t rol 98 1 5 1 66 4-Methoxycoume s t rol 60 1 06 3 3
1 3 : 4 : 7 7
Coume s t rol 6 9 1 2 2 8 1 4 -Me thoxycoume s t rol 34 7 0 4 1
20 : 4 : 7 7
Coume s trol 88 94 1 4 5 4-Me t hoxycoume s trol 38 59 1 0 7
( i i) Da ily Coumes tan Intake
PLOT D CLOVER
100 < 10 7 7 < 1 0
1 1 0 < 5 6 3 < 5
1 7 2 < 5 1 4 5 < 5
Sheep are capable o f consuming each day, a to tal we ight o f
forage equal t o 3 percent o f their bo dyweigh t . Cons ider ing tha t lucern e ,
during i t s f lower ing s ta ges , i s approxima t ely 2 4 percent dry mat t er ,
then a conserva t ive e s t ima t e o f the daily in take o f lucerne by ewes in

this tria l , would b e 1 kg o f dry ma t t er . The mean daily intake o f
coume s tans , for ewe s i n both exper imen t s , bas ed on a dai ly intake o f
1 kg o f dry ma t t e r , are shown in Tabl e 3-2 .
TABLE 3-2 . THE DAILY COUMESTAN INTAKE (mg/ day) OF EWES
INGEST ING 1 KG ( DM) OF LUCERNE
EXPERIMENTAL STAGE OF OESTROUS CYCLE ( DAY S ) GROUP
0-9 1 0- 1 7 1 8- 2 5
EXPERIMENT I Pas ture 2 2 1 Lucerne 1 1 7 2 5 2 1 7 3
EXPERU1ENT I I Pas ture 1 1 Luc erne 1 9 2 2 1 7 Past-Luc 1 2 1 7 Luc-Pa st 1 9 2 1
The coumes tan intake o f ewes gra z ing ryegras s / c lover pas tu r e , is
based on the as sump t ion tha t whi t e c l over accounts for 10 percent of
daily dry ma t ter in take .
(B ) EWE LIVEWEIGHTS AND OESTRUS
( i ) Ewe Liveweigh t s
Mean l iveweights o f ewes in both exp e rimen t s a r e l i s t ed in
Tab l e 3-3 . There wer e no significant d i f f e rences be tween t r ea tment
groups a t any o f the weight recordings . S imilarly there were no
signi f ican t di f f e renc es in the rate of weight gain b e tween weighings .
3 7 .
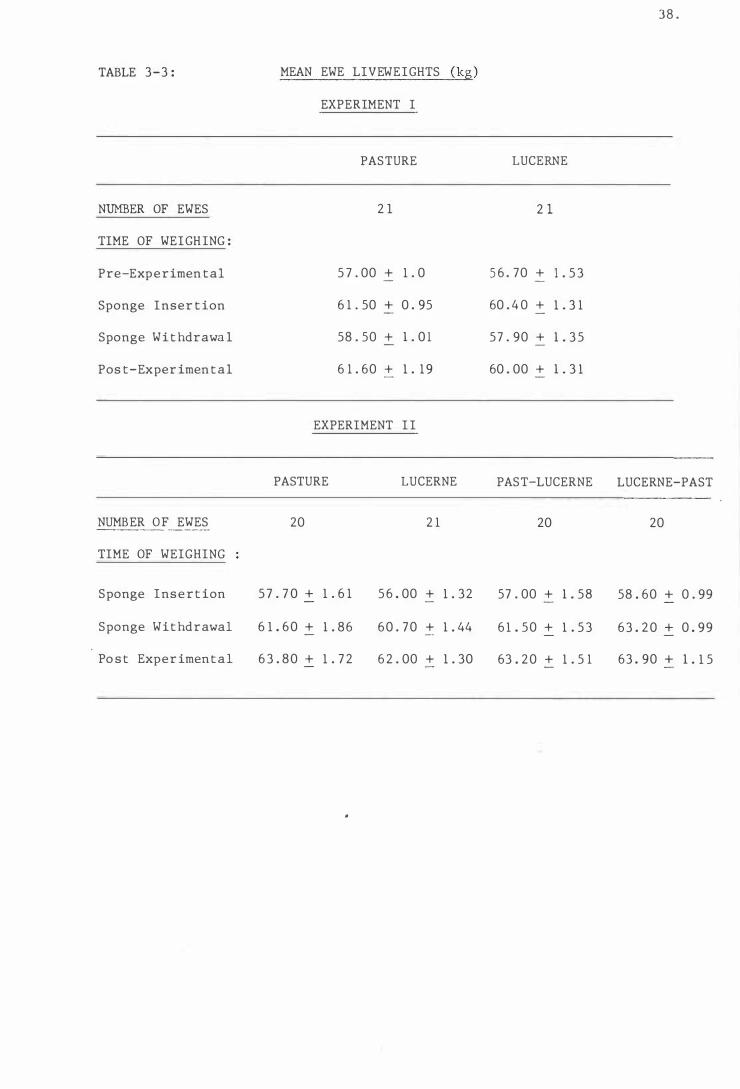
TABLE 3-3 :
NUMBER OF EWES
TIME OF WEIGHING :
Pre-Experimen ta1
Sponge Inser t ion
Sponge Withdrawa l
Pos t-Exper imen tal
NUMBER OF EWES
TIME OF WEIGHING
Sponge Ins e r t ion
Sponge Withdrawal
Po s t Exper imen tal
MEAN EWE LIVEWEIGHTS ( kg)
EXPERIMENT I
PASTURE
2 1
5 7 . 00 + 1 . 0
6 1 . 50 + 0 . 95
58 . 50 + l . 0 1
6 1 . 60 + l . 1 9
EXPERIMENT I I
PASTURE LUCERNE
20 2 1
5 7 . 7 0 + l . 6 1 5 6 . 00 + l . 3 2
6 1 . 6 0 + l . 8 6 6 0 . 7 0 + l . 44
6 3 . 8 0 + l . 7 2 6 2 . 00 + l . 30
38 .
LUCERNE
2 1
5 6 . 70 + 1 . 5 3
60 . 4 0 + l . 3 1
5 7 . 90 + l . 3 5
60 . 00 + l . 3 1
PAST-LUCERNE LUCERNE-PAST
20 20
57 . 00 + l . 5 8 58 . 60 + 0 . 99
6 1 . 5 0 + l . 5 3 6 3 . 2 0 + 0 . 9 9
63 . 2 0 + l . 5 1 63 . 90 + l . 1 5

3 9 .
( i i ) Oe s trus Synchroni za t ion
The d i s tribu tion of the onse t of oes trus for bo th the synchronized
and the exper imen t a l oe s trus ar e shown i n FIGS 3-1 and 3-2 .
In Expe r imen t I , 9 3 perce n t of the ewes were d e t ec ted i n oes trus
over a four day per iod fol lowing sponge wi thdrawal . E i gh ty two percent
of the ewe s were de tec ted in heat over a corre s pond ing f ou r day period in
the f o l lowing oes trou s cyc l e .
In Exper imen t II , 83 . 3 percent o f the ewe s were detec ted over a f our
day period for the synchroni zed oes trus and 7 6 . 5 percent in the fo l l owing
oe stru s . Inspe c t ion o f the reproduc t ive trac t s later revea led tha t a
number of ewes we re phy s i o l og i ca l ly unab l e to exhib i t oes trus . Two ewe s had
b l ind r eproductive tra c t s w i th only vag i na l t i s s ue present , and one ewe
was found to be pre gnan t . A fur ther four ewe s had l o s t s po nges dur ing
synchron i za t ion . Only two ewe s exhibi ted ' s i l ent ' hea t s , one in the
pa s ture group and one i n the lucerne-pa s ture group .
Th ere were no s igni f i cant d i f f ere nce s in oes trus synchr ony a t the se cond
oes trous cyc le be tween ewe s gra z i ng l u cerne and ewe s gra z ing pa s ture .
S imi l a rly there we re no d i f f erence s �n mean cyc le length .
( i i i ) Oe s t rus-Ovu l a t ion Int erval
The in terva l b e tween the ons e t of oes trus and the pre-ovu l a tory LH
peak , wa s extreme ly variab l e b e tween anima l s , i n both trea tment group s .
Some anima l s were r e co rded as only enter i ng oes trus , af ter the LH peak
had occurred . The i n terva l length f or ind ividual sheep � s shown i n
FIG 3-3 . Ther e were no s igni f i cant d i f ference s be tween ewes gra z i ng
lucerne and ewes graz ing pas tur e in the mean i n terval dura t ion .
The re were no apparent pat terns shown in the ch loride content o f
cerv i ca l mucus samp l e s co l le c ted . Ch l o r i d e concentr a t ions appeared t o
r i s e and fa l l rand omly . Consequen t ly , the t ime o f ovu l a t ion c ou l d no t
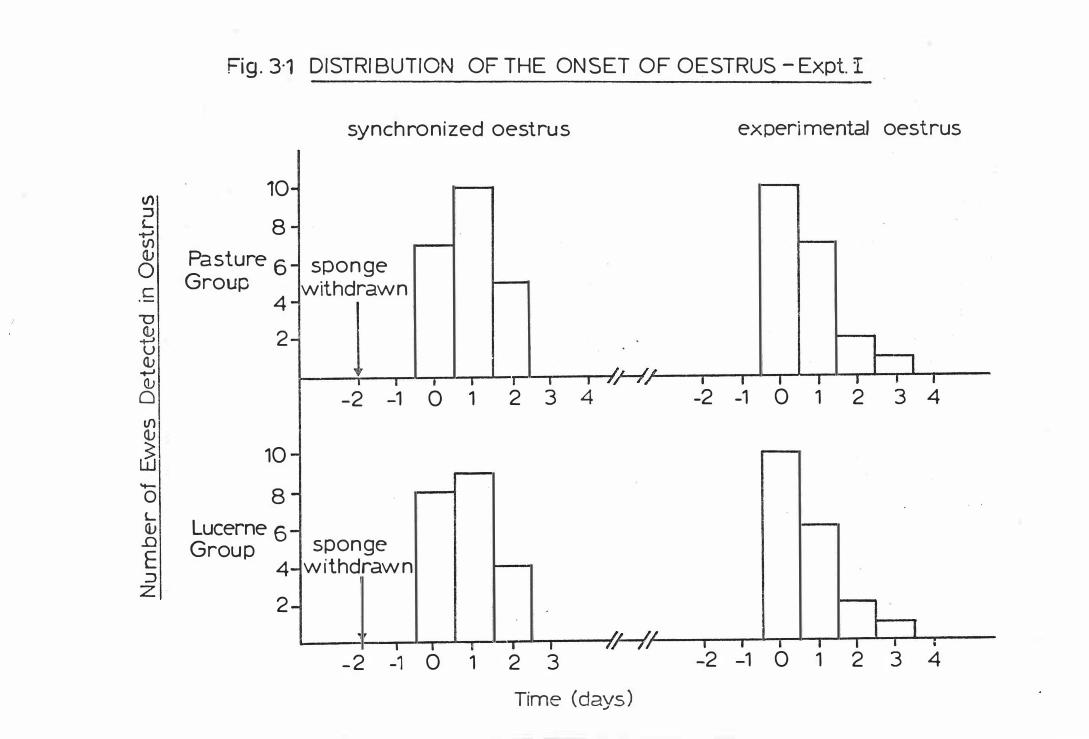
tl) ' :J �I tll QJ 0
"'0 �I u QJ � (!;
0 tll QJ �! -0 L (])
.0 E .:J
Z _
Fig . 3·1 D I STRI BUTION OF THE ON S ET O F O E STRUS - Expt. I
synch ro n i zed oestru s experi mental oestrus
10 .
8 Pa sture 6
Group 4 2
· spon ge withdrawn
-
. 1 I I I I I I I � , I . .
-2 -1 0 1 2 3 4 -2 -1 0 1 2 3 4
10
8 Lucerne 6 Grou p
4 spon ge
w ithdraw n
2 �
-. I I I I I, lf-!J _, I . . '
- 2 -1 0 1 2 3 -2 -1 0 1 2 3 4
Time (days )
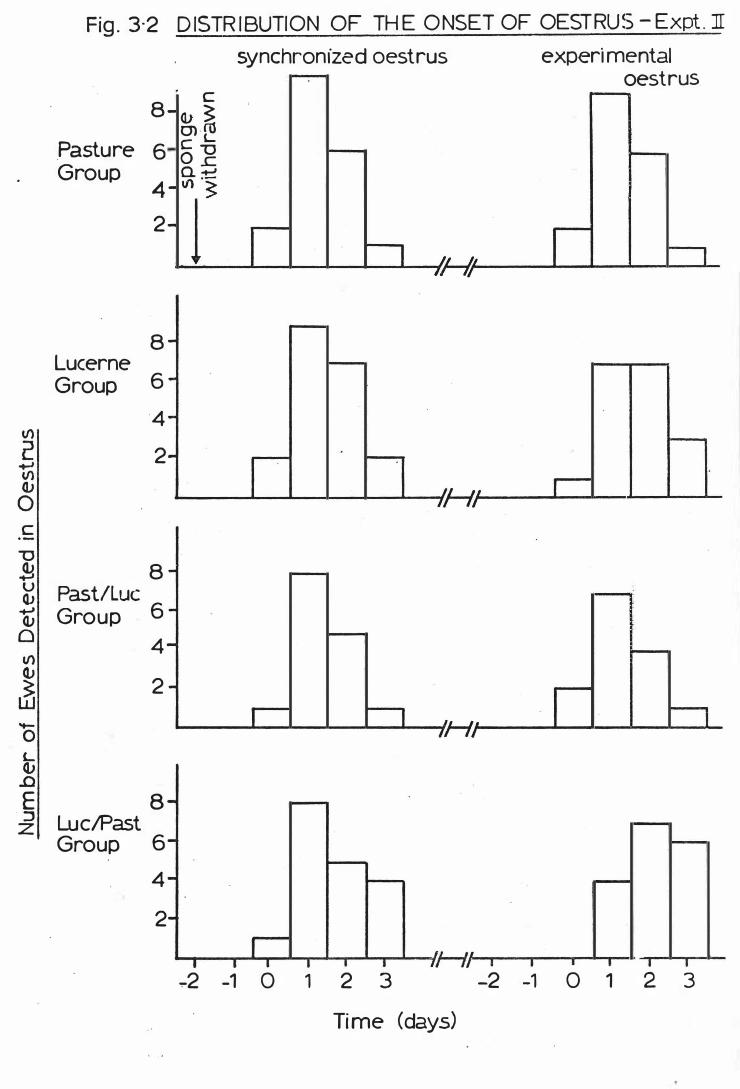
Fig. 3·2 D ISTR I BUTION OF TH E O NSET OF OEST RU S - Expt . TI
Cl) 2
+oJ Cl) Q) 0 c u
8
Pasture 6 Group
4 2
8 Lucerne Group 6
4 2
$ 8 � Past/Luc t Group 6 0
-0 L Q)
.0
4
2
E 8 .2_ Luc/Past Group 6
4
2
c - Q) �
rn · ru L _ c u O .c n .......
� �( -
-
-
..
..
...
-
-
-
-
-
..
synchronized oestrus
l 7.
7
I 11 7
.,
L r
.. I
experimental oestrus.
I
I I I I I I I I. 'L , I I I I I rl �2 -1 0 1 2 3 -2 -1 0 1 2 3
Time (days)
I
I
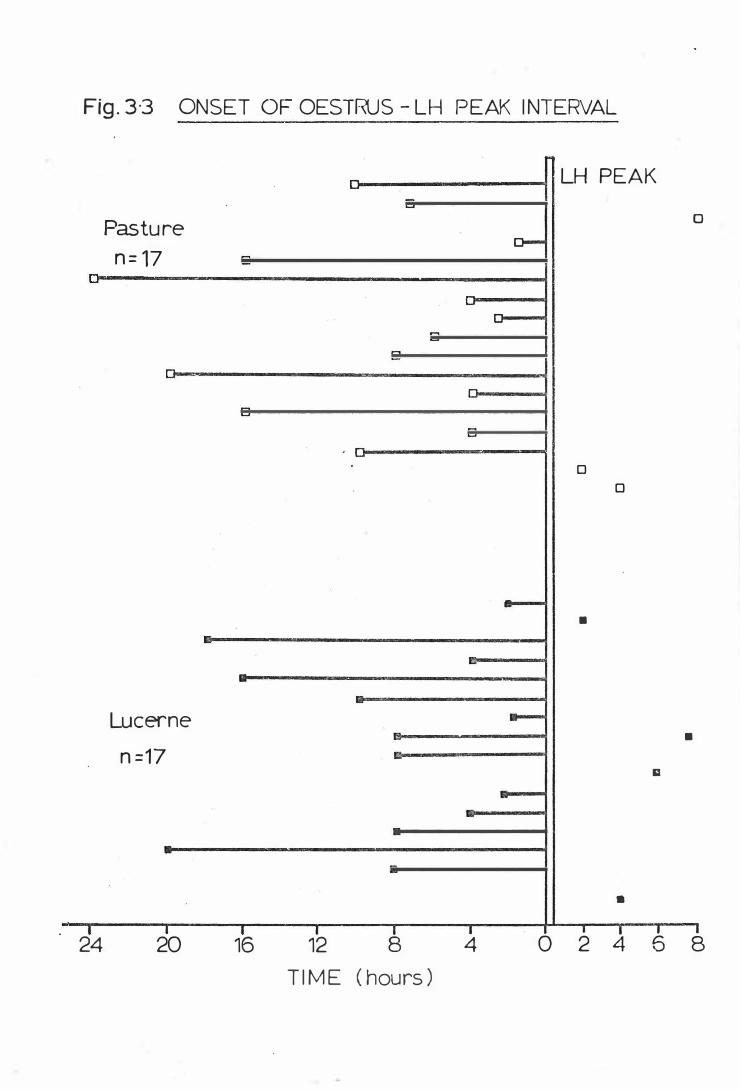
F ig. 3-3 ONSET OF OESTRUS - LH PEAK I NTERVAL
...... LH PEAK . �
Pastu re o--
n = 17 -
�
-......
....... ......
,....
......
-
0 0
•
Lucerne -
n =17 El
•
I I I I I I I I I
0
•
I 24 20 16 12 8 4 0 2 4 6 8
T I M E ( hours )

be e s t ima ted .
( C ) LUTE INIZING HORMONE SECRETION
( i ) Dur ing the Oe s trous Cycle
Mean dai l y concentra t ions o f lu t e in i z i ng hormone i n per ipheral
40 .
p lasma o f ewe s f r om both the pas ture and the lu cerne groups o f Exper iment I
are l i s t ed in Table 3-4 . Analyses o f var i ance b e tween the groups were
made for each samp le day . There we re no s ignif i can t d i f ferenc e s i n LH
secret ion at any s t age of the oe s trous cyc l e . Mean plasma LH c oncent r a t ions
r anged f rom 0 . 5 to 1 . 5 ng /ml over the cyc l e , exc lud i ng t he pre-ovu l a tory
peak . The pa t te rns o f secr e t ion are shown i n F I G . 3-4 .
( i i ) The Pre-Ovula tory LH Peak
Mean concen t r a t ion s of lute ini z i ng hormone in pe r i pher a l p l asma ,
measured a t 2-hourly in terva l s f rom the ons e t of oe s trus , are shown in
Tab le 3-5 . The pre-ovu la tory LH peaks we re inc luded in th i s pe r i od .
Ana l y s es of var i ance be tween the lucerne and the pas ture group s , a t each
sample t ime , f a i led to show any s i gn i f icant d i f f erenc es . The pre-ovu l a tory
LH secr e t ion pa t terns are shown in FIG 3 - 5 .

4 1 .
TABLE 3-4 : MEAN PLASMA LH CONCENTRATIONS (ng/ml ) DURING THE OESTROUS
CYCLE IN EWES GRAZING PAS TURE OR OESTROGENI C LUCERNE
DAY - 7 -6 -5 -4 - 3 -2 - 1 0 + 1
PASTURE 0 . 5 5 9 0 . 4 5 2 0 . 4 35 0 . 4 4 5 0 . 7 5 6 1 0 . 94 9 1 . 0 3 8 7 . 0 5 0 . 5 9 1
(MeaniSE ) +0 . 1 7 +0 . 1 2 +0 . 1 1 +0 . 1 3 +0 . 1 2 +0 . 2 6 +0 . 32 +3 . 9 2 +0 . 1
LUCERNE 0 . 4 0 6 5 0 . 5 7 3 0 . 307 1 0 . 409 0 . 5 7 3
Mean±SE) +0 . 1 3 +0 . 14 +0 . 1 1 +0 . 1 3 +0 . 5
DAY +2 +3 +4 +5 +6
PASTURE 0 . 9 7 5 0 0 . 7 6 5 0 . 859 0 . 7 4 9 0 . 7 4 9
MeaniS E) +0 . 3 6 +0 . 1 7 +0 . 2 +0 . 2 1 +0 . 1 9
LUCERNE 1 . 5648 l . 037 1 . 04 1 0 . 5 7 1 0 . 3 9 9
.(Mean±SE ) +0 . 4 1 +0 . 3 2 +0 . 2 9 +0 . 1 6 +0 . 1 2
1 . 24 7 1 . 1 6 1 9 1 . 1 4 1 . 04
+0 . 3 2 +0 . 2 5 +6 . 5 5 +0 . 1 1
+7 +8 +9 + 1 0
0 . 58 5 0 . 5 34 0 . 4 34 0 . 54 6
+0 . 1 6 +0 . 1 4 +0 . 1 1 +0 . 1 6
0 . 6 7 3 0 . 4 08 0 . 4 3 3 0 . 5 5 6
+0 . 1 2 +0 . 1 1 +0 . 1 4 +0 . 1 3
( Day 0 o f o e s t rous cyc l e co rrespond s t o t ime of pre-ovula tory LH peak)
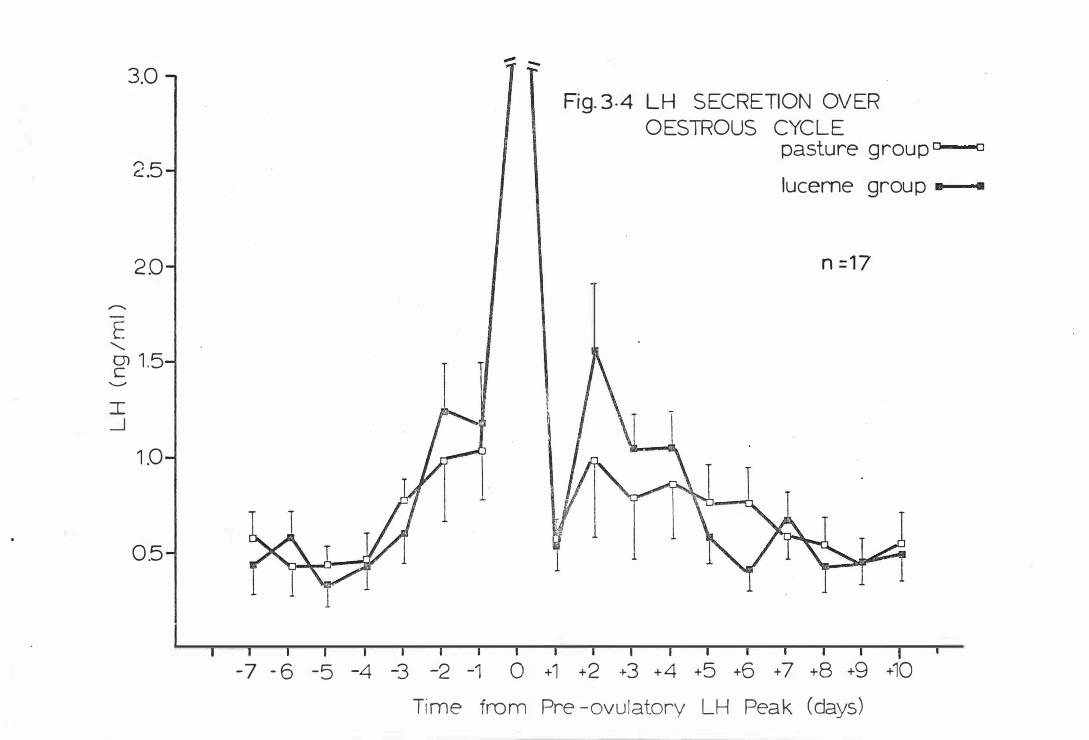
3.0
2 .5
2 0 ........
E .......... � 1 .5 ..____.
I _j
1 .0
0 .5
_,.,. -
1
Fig. 3 .4 L H S ECRETION OVER O ESTROUS CYC L E
pasture g roup o o
luceme group • •
n = 17
I I I I I I I T--T--uul - 1- ,------.----r-.---r--�--.---T----. -7 - 6 -5 -4 -3 -2 -1 0 +1 + 2 +3 + 4 +5 +6 +7 +8 +9 +10
Ti me fmm Pre - ovulatory LH Pea k (days)

TABLE 3-5 :
PASTURE
LUCERNE
PASTURE
LUCERNE
PASTURE
LUCERNE
PRE-OVULATORY PLASMA LH CONCENTRAT IONS (ng/ml) IN EWES
GRAZING PASTURE OR OESTROGENIC LUCERNE
HOUR- 1 2 - 1 0 -8 - 6 -4 -2 0 +2
1 . 1 3 2 . 4 3 1 . 8 5 6 . 04 7 . 9 9 29 . 20 8 7 . 05 4 0 . 2 6
+0 . 3 7 +0 . 59 +0 . 3 5 +2 . 3 9 +2 . 54 +4 . 84 +3 . 9 2 5 . 8 2
1 . 7 1
+0 . 4
+4
1 . 9 6 3 . 4 1 2 . 5 1 1 8 . 9 1
+0 . 6 7 +0 . 9 9 +0 . 4 8 +3 . 9 3
+6 +8 + 1 0 + 1 2
3 1 . 3 8 1 3 . 3 7 8 . 68 2 . 70 l . 40
39 . 99
6 . 30
+ 1 4
0 . 84
9 1 . 1 4
+6 . 5 5
+ 1 6
0 . 7 5
4 6 . 2 3
+5 . 24
+ 1 8
0 . 9 0
+4 . 1 1 +5 . 8 9 +2 . 6 8 +0 . 5 7 +0 . 2 6 +0 . 1 8 +0 . 1 9 +0 . 38
2 6 . 1 9 1 1 . 6 3 3 . 2 4 2 . 4 0 1 . 1 1 0 . 8 3 0 . 7 2 0 . 5 1
+3 . 7 4 +3 . 5 6 +0 . 3 7 +0 . 2 5 +0 . 2 8 +0 . 1 8 +0 . 2 0 +0 . 08
+20
l. 1 9
+0 . 38
0 . 4 1
+22 +24
0 . 5 1 0 . 2 5
0 . 24 +0 . 06
0 . 44 0 . 6 6
+0 . 06 +0 . 09 +0 . 1 9
4 2 .
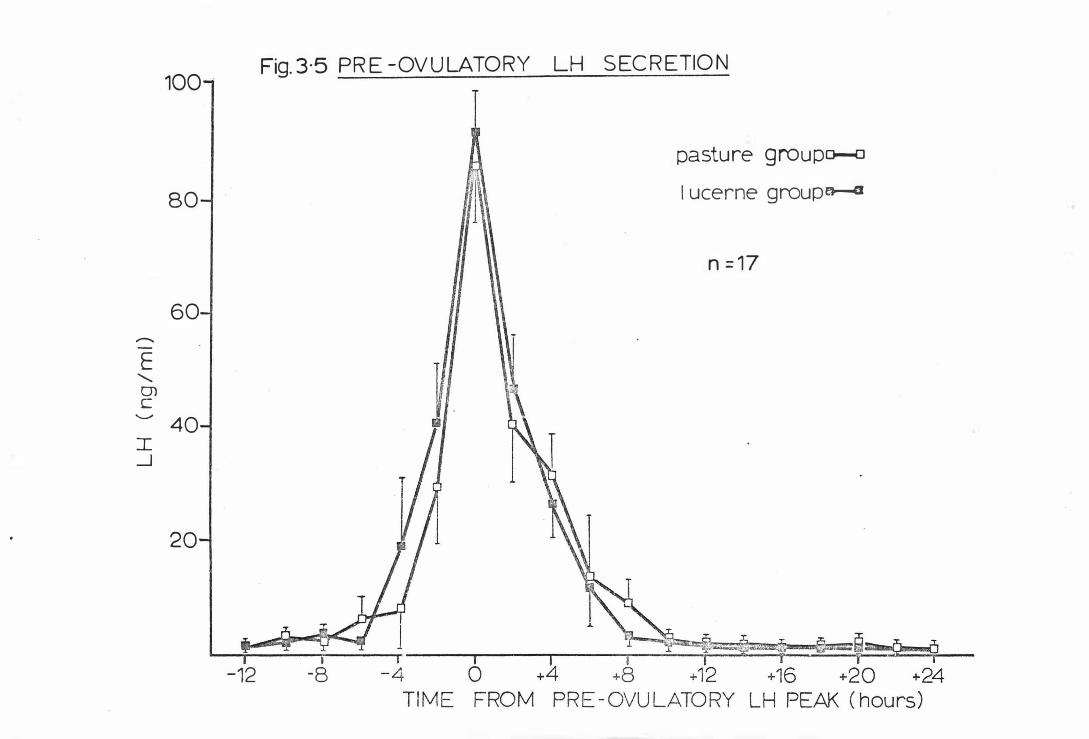
............
E ' 0) c
100 .
80
60
'-' 40 I _j
20
Fig. 3·5 PR E -OV U LATORY LH S EC R ETIO N
-12 -8
r ,, ]'1 pa sture groupo-o
I ucerne group �
n = 1 7
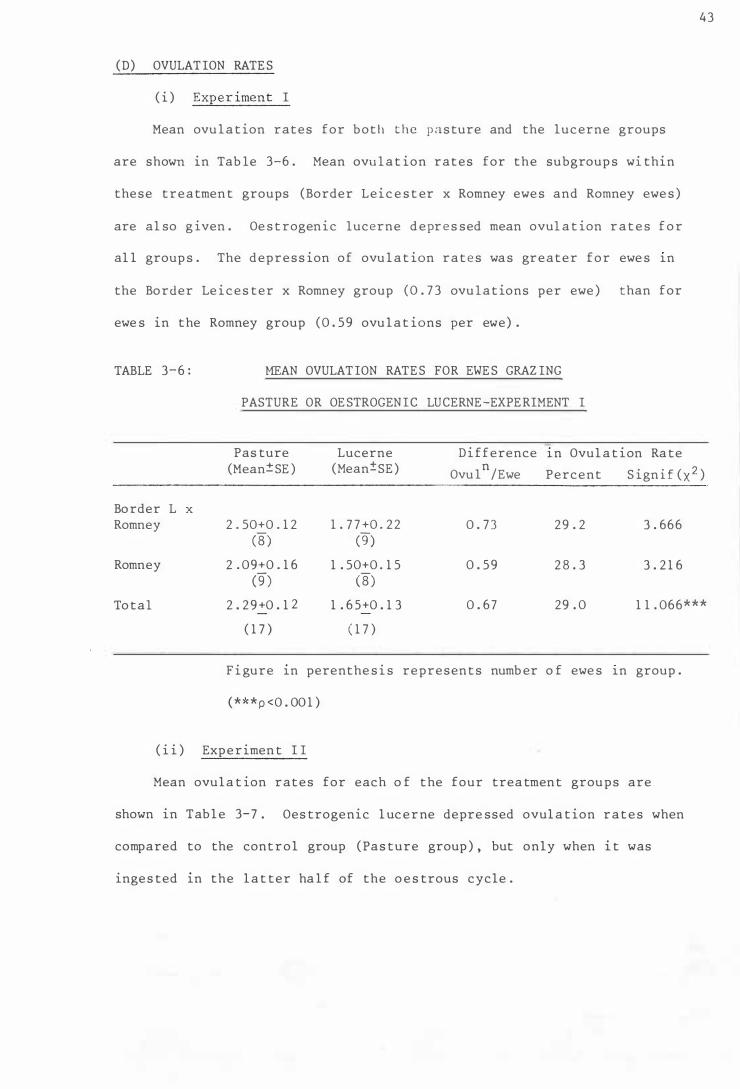
(D) OVULAT ION RATE S
( i ) Expe r iment I
Mean ovu l a t ion rates for both t h e p a s ture and the luc erne groups
are shown in Tab le 3-6 . Mean ovu l a t ion rates for the subgroup s wi thin
these t r e a tmen t groups (Border Lei c e s t er x Romney ewe s and Romney ewe s )
are al so g iven . Oe s t rogen ic lucerne d epr e s sed mean ovul a t ion r a t e s f o r
al l group s . The d epress ion of ovu l a t ion r a t e s wa s gr eater f o r ewe s �n
the Bo rder Le i c e s ter x Romney group ( 0 . 7 3 ovu l a t ions per ewe ) than for
ewe s in the Romney group ( 0 . 5 9 ovu l a t ions per ewe) .
TABLE 3- 6 : MEAN OVULAT ION RATE S FOR EWE S GRAZ ING
PASTURE OR OE STROGEN IC LUCERNE -EXPE RIMENT I
Pas ture Lucerne D i f f erence �n Ovul a t ion Ra te
43
(Mean± SE ) (Mean± SE ) n Ovu l /Ewe Percent S ign i f (x 2 )
Bo rder L X
Romney 2 . 50+0 . 1 2 1 . 7 7 +0 . 2 2 0 . 7 3 29 . 2 3 . 66 6 (8) (9)
Romney 2 . 09+0 . 1 6 1 . 50+0 . 1 5 0 . 5 9 2 8 . 3 3 . 2 1 6 (§) (8)
To tal 2 . 2 9 +0 . 1 2 1 . 6 5+0 . 1 3 0 . 6 7 29 . 0 1 1 . 066***
( 1 7 ) ( 1 7 )
F i gure �n perenthe s i s represen t s numb er o f ewe s �n group .
( * * * p <0 . 00 1 )
( i i ) Exp e r imen t I I
Mean ovul a t ion rates for each o f the four trea tmen t grou p s are
shown in Tab l e 3- 7 . Oes trogenic l uc e r ne depre ssed ovul a t ion r a t e s when
compared to the cont r o l group (Pas ture group ) , but only when i t was
inge s ted �n the l a t ter ha l f of the o e s trous cycle .

TABLE 3-7 : MEAN OVULATION RATES FOR EWES GRAZING
PASTURE OR OESTROGENI C LUCERNE-EXPERIMENT I I
TREATMENT OVULATION RATE
(MEAN.±_S E )
Pas ture ( P ) 1 . 7 5 + 0 . 1 3
Luc erne ( L ) l . 3 5 + 0 . 12
Pas ture-Lucerne (PL) 1 . 1 0 + 0 . 1 0
Luc erne-Pas ture (LP ) l . 5 5 + 0 . 12
n .
1 6
1 7
1 0
1 8
Compar isons o f the d i f f er ences in ovulat ion ra t e s b e tween
t r ea tmen t s are shown in Table 3-8 .
TABLE 3-8 : COMPARI S ONS OF THE DIFFERENCES IN OVULATION RATES
BETWEEN TREATMENTS - EXPERIMENT II
P . V L .
P . V PL .
P . V LP .
L . V PL .
LP . v L .
LP . v PL .
( n . s .
Diff er enc es Ovul at ions/ Ewe
0 . 4 0
0 . 6 5
0 . 20
0 . 2 5
0 . 2 0
0 . 4 0
no t s ignif ican t
Percen t x2 Diff erences S ta t i s t ic
2 2 . 9 3 . 34 5 2
37 . 1 8 . 54 6 7
1 1 . 4 0 . 6243
1 8 . 5 2 . 1 09 6
1 2 . 9 1 . 9983
2 9 . 0 5 . 5934
S ignif icance
tt
* *
n . s .
n . s .
n . s .
*
* p < 0 . 05 ** p < O . O l # O . l > p > 0 . 05 )
There we re no s ignif icant d i f f erenc es in ovula t ion ra t e s be tween
the two groups which gra z ed pa s t ure in the lat t er ha l f of the cyc l e
(Pasture and Luc erne-Pas ture) , o r be tween the two gro up s which grazed
44 .
luc erne in the la t t er hal f of the c yc l e ( Lucerne and Pas tur e -Luc erne ) .
However , t her e were d i f f erences in ovula t ion rate b etween the two t r ea t -
men t -in terchange group s .

tf) w � 0:::
z 0 � _J �
6 � <.0 M .rn LL.
1 1 c
l .. I I
I
<D �
I
1 1 c
0 .C\J
1 I
-
CO -1 � L
1 1 I . c
('. � L 1 1 I c
-1 .
1"- I .,...
l l L c l .<I
$2 L -1 1 1 J . c
-1 .
I o. �
_.!
lSVd I :Jn""l
:J nl / lS Vd
_ 3 Nt1 3:Jnl
3 � n1svd
3 N cf3:Jnl _
3 CJ nlS'v'd
I
(\J
1)_ X w

( i ii ) Corpora Lu tea Diameter
The d iameter o f each co rpus lut eum wa s calcula t ed as the mean o f
two measuremen t s taken a t r ight angles a t the widest po in t . The
mean corpus lut eum d iame ter in ewes f rom each trea tmen t group , a r e
shown i n Tabl e 3-9 . There wer e no s ign if icant dif f erences b e tween
trea tmen t groups .
TABLE 3-9 : MEAN CORPORA LUTEA DIAMETER (mm) THREE DAYS
POST-OESTRUS
PASTURE LUCERNE
5 . 9 7 + 0 . 3 7 5 . 59 + 0 . 2 5
( E) FOLLI CLE POPULATIONS
( i ) S ur face Fol l ic l e s
PASTURE-LUCERNE LUCERNE-PAS TURE
6 . 05 + 0 . 3 8 5 . 8 5 + 0 . 2 5
4 5 .
In Exp er imen t I , the number o f unrup tured fo l l ic l es with a d iame t er
grea t er than 3 . 5mm tha t wer e present on t he surface o f the ova r i e s , was
recorded a t laparotomy . Th is wa s comb ined with the number o f co rpora
lutea present to g ive an e s t ima t e o f the to tal number o f large f o l l i c l e s
present a t t h e t ime o f ovula t ion . From this ' es t ima t e ' o f total fol l ic l e
number s , t h e rat io o f the number o f large f o l l icles pr esen t per ovula t ion
is calcul a t ed . The se da ta are shown in Tabl e 3-1 0 .
I n Exper imen t I I , all unrup tured sur fac e fol l i c l e s with a d iamet er
o f grea t er than 2mm were r ecorded on recovery o f the ovaries a t s laugh t er .
Da ta f rom this exper imen t i s shown in Tab l e 3 - 1 1 .
The r emoval of the ova r i es in Exper iment I I enab led sma l l f o l l i c les
to be measu red w i th more accuracy than could be ach ieved in s i tu in
Exper iment I .
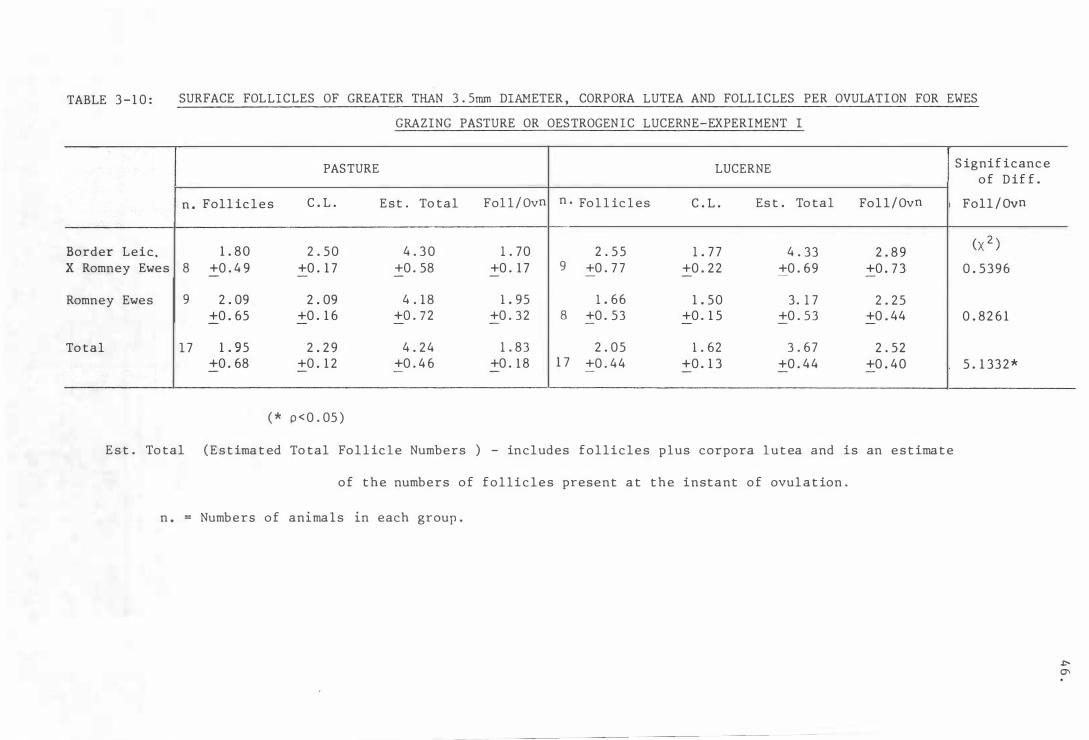
TABLE 3 - 1 0 : SURFACE FOLLICLES OF GREATER THAN 3 . 5mrn DIAMETER , CORPORA LUTEA AND FOLLICLES PER OVULATION FOR EWES
GRAZING PASTURE OR OESTROGEN IC LUCERNE-EXPERIMENT I
PASTURE LUCERNE
n . Foll ic l e s C . L . E s t . To tal Fol l / Ovn n · Fo l l ic l es C . L . Es t . To tal Fol l / Ovn
Border Leic, 1 . 8 0 2 . 5 0 4 . 3 0 1 . 7 0 2 . 5 5 1 . 7 7 4 . 33 2 . 8 9 X Romney Ewes 8 +0 . 4 9 +0 . 1 7 +0 . 58 +0 . 1 7 9 +0 . 7 7 +0 . 2 2 +0 . 6 9 +0 . 7 3
- - -
Romney Ewes 9 2 . 09 2 . 09 4 . 1 8 l . 95 l . 6 6 l . 5 0 3 . 1 7 2 . 2 5 +0 . 65 +0 . 1 6 +0 . 7 2 +0 . 3 2 8 +0 . 5 3 +0 . 1 5 +0 . 5 3 +0 . 44 -
To tal 1 7 1 . 9 5 2 . 2 9 4 . 2 4 1 . 8 3 2 . 0 5 l . 62 3 . 6 7 2 . 5 2 +0 . 68 +0 . 1 2 +0 . 4 6 +0 . 1 8 1 7 +0 . 4 4 +0 . 1 3 +0 . 4 4 +0 . 4 0 - - - - -
------
( * p < 0 . 05 )
E s t . To tal ( E s t ima t ed To tal Fo l l ic l e Numbers ) - includes follicles p lus corpora l u t ea and is an e s t ima t e
o f t h e numbers o f f o l l i c l e s present a t t h e instant o f ovul a t ion .
n . = Numbers o f anima l s in each group .
S ignif icanc e o f D if f .
Foll / Ovn
<x z ) 0 . 5 3 9 6
0 . 8 2 6 1
5 . 1 33 2 *
.t:-0'\

TABLE 3- 1 1 : S URFACE FOLLICLES OF GREATER THAN 2 . 0mm DIAMETER , CORPORA LUTEA AND FOLLICLES PER OVULATION FOR EWES GRAZING
PASTURE OF OESTROGENIC LUCERNE - EXPERIMENT I I
PASTURE
Surf Fo llicle Co rpora Lu t ea Est ima ted To tal Fo l l icles /Ovn
5 . 08 + 0 . 46 1 . 7 5 + 0 . 1 3 6 . 8 3 + 0 . 6 1 3 . 9 0 + 0 . 32
(n = 1 6 )
PASTURE-LUCERNE
LUCERNE
Surf Fo l l ic l e Co rpora Lut ea Estima t ed To tal Fo l l ic les/ Ovn
4 . 5 1 + 0 . 3 9 1 . 3 5 + 0 . 1 2 5 . 8 6 + 0 . 53 4 . 34 + 0 . 4 2
( n 1 7 )
LUCERNE-PASTURE
Surf Fol l i c l e Corpora Lu tea E s t imat ed To tal Fol l i c l e s / Ovn I Sur f Fo l l icle Corpora Lu t ea Es t ima t ed Total Foll ic l e s / Ovn
4 . 0 7 + 0 . 4 3 1 . 1 + 0 . 1 5 . 1 7 + 0 . 58 4 . 7 0 + 0 . 52 I 3 . 8 1 + 0 . 3 1 1 . 5 5 + 0 . 1 2 5 . 3 6 + 0 . 55 3 . 4 6 + 0 . 3 9
( n = 10) (n 1 8 )
n = Number o f anima l s i n each group .
.1:'-..J

4 8 .
( i i ) To tal Foll icle Popula t ions
The total numbers of follicles present , wi th a d iame ter grea ter
than 2rnrn , wer e r ecorded a f t er d i s sec t ion o f the ovar ies . Ther e were
no s ignif icant d i f f e renc e s , nor any apparent trend s , evident in t o t a l
fol l i c l e popula t ions be tween t r ea tmen t s ( Table 3-1 2 ) .
E s t ima t es o f the t o tal numbers o f follicles presen t a t the in s tan t
of ovulat ion wer e mad e , using the same proc edure as wa s u s ed in surface
foll i c l e es t ima t e s . S imilar ly , the total numb er o f f o l l i c l e s presen t
per ovula t ion was cal cula t ed . These da ta are shown in Tab le 3- 1 3
for f o l l icles with a d iame t er o f grea t er t han 3 . 5rnm , and in Table 3 - 1 4
for f o l l icles w i t h a d iamet er o f grea t er t han 2 . 0mrn .
To tal fo l l i c l e popula t ions a r e shown in F IG . 3-7 .
There we re no s ign i f icant d i f ferenc es be tween treatmen t group s , in
the mean numb er of foll icles o f a d iame t er grea ter than 2 . 0mm produced
per gram of ovar ian t i ssue ( Tab le 3- 1 5 ) .
TABLE 3 - 1 5 : FOLLICLES OF GREATER THAN 2 . 0rnm DIAMETER PER GRAM
OF OVARIAN TISSUE
PAS TURE LUCERNE PASTURE-LUCERNE
1 . 9 3 + 0 . 3 1 1 . 8 4 + 0 . 2 6 2 . 2 3 + 0 . 3 4
(F) OVARIAN AND REPRODUCTIVE TRACT RESPON S E
( i ) Ovar ian and U t e r ine Weigh t s
LUCERNE-PASTURE
2 . 0 2 + 0 . 29
Mean ovar ian and u t erine weight s for each trea tmen t gr oup are
shown in Table 3 - 1 6 . Ther e wer e no s ign if icant d i f ferenc e s in organ
weigh t s be tween group s .
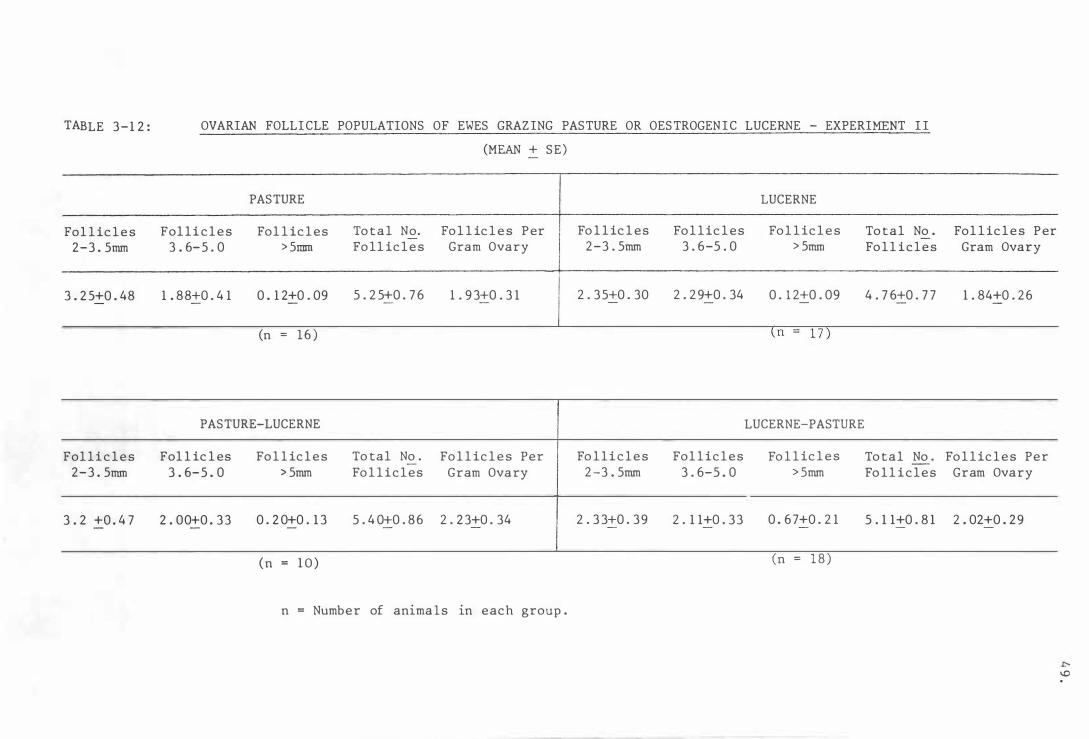
TABLE 3 - 1 2 :
Fol l ic l e s 2-3 . 5mrn
3 . 2 5+0 . 48
Fol l i c l e s 2-3 . 5mm
3 . 2 +0 . 4 7
OVARIAN FOLLICLE POPULATIONS OF EWES GRAZ ING PASTURE OR OES TROGEN IC LUCERNE - EXPERIMENT I I
(MEAN + SE)
Fol l i c l e s 3 . 6- 5 . 0
1 . 8 8+0 . 4 1
PASTURE
Fol l ic l es > 5mrn
0 . 1 2+0 . 09
(n = 16 )
PASTURE-LUCERNE
Fol l ic l es 3 . 6- 5 . 0
2 . 00+0 . 3 3
Fo l l ic l e s > 5mrn
0 . 2 0+0 . 1 3
(n = 1 0 )
To tal No. Fo l l ic l e s P er Fo ll ic l e s Gram Ovary
5 . 2 5+0 . 7 6 1 . 9 3+0 . 3 1
To tal No . Fo l l ic l e s Per Fo l l icles Gram Ovary
5 . 4 0+0 . 8 6 2 . 2 3+0 . 34
n = Number of anima l s in each group .
Fo l l ic l es 2-3 . 5mrn
2 . 3 5+0 . 30
Fo l l i c l e s 2 -3 . 5mrn
2 . 3 3+0 . 3 9
Fo l l ic l es 3 . 6-5 . 0
2 . 2 9+0 . 34
Fol l icles 3 . 6-5 . 0
2 . 1 1+0 . 3 3
LUCERNE
Fo l l ic l e s > 5mm
0 . 1 2+0 . 09
�n 17)
LUCERNE-PASTURE
Fo l l ic l e s > 5mrn
0 . 6 7+0 . 2 1
(n = 1 8 )
To tal No . Fol l ic l e s P er Fo l l icles Gram Ovary
4 . 7 6+0 . 7 7 1 . 84+0 . 2 6
To tal No . Fo l l ic l e s Per Fo l l icles Gram Ovary
5 . 1 1+0 . 8 1 2 . 02+0 . 2 9
� \0
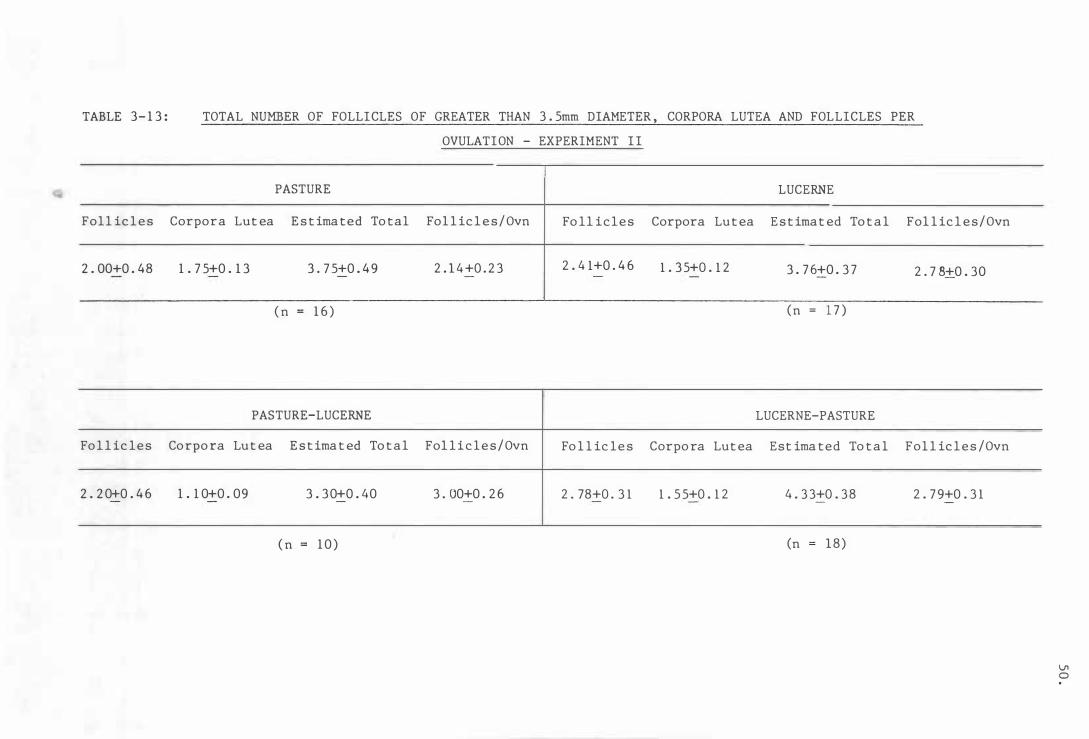
TABLE 3- 1 3 : TOTAL NUMBER OF FOLLICLES OF GREATER THAN 3 . 5mm DIAMETER , CORPORA LUTEA AND FOLLICLES PER
OVULATION - EXPERIMENT I I
PASTURE
Foll ic l e s Corpora Lut ea E s t ima ted To tal Fol l ic l e s / Ovn
2 . 00+0 . 48 1 . 7 5+0 . 1 3 3 . 7 5+0 . 4 9 2 . 1 4 +0 . 2 3
( n = 1 6 )
PASTURE-LUCERNE
Fo l l ic les Co rpo ra Lu t ea E s t ima t ed To tal Fol l i c l e s / Ovn
2 . 2 0+0 . 4 6 1 . 1 0+0 . 09 3 . 3 0+0 . 40 3 . 00+0 . 2 6
( n 1 0 )
LUCERNE
Fo ll icles Corpora Lu t ea E s t ima t ed To t a l Fol l ic l e s / Ovn
2 . 4 1+0 . 4 6 1 . 3 5+0 . 1 2 3 . 7 6+0 . 3 7 2 . 7 8+0 . 30
(n = 1 7 )
LUCERNE-PASTURE
Fo l l ic le s Corpo ra Lut ea Est ima ted To tal Fol l icles /Ovn
2 . 78+0 . 3 1 1 . 5 5+0 . 1 2 4 . 3 3+0 . 3 8 2 . 7 9+0 . 3 1
(n 1 8 )
V1 0
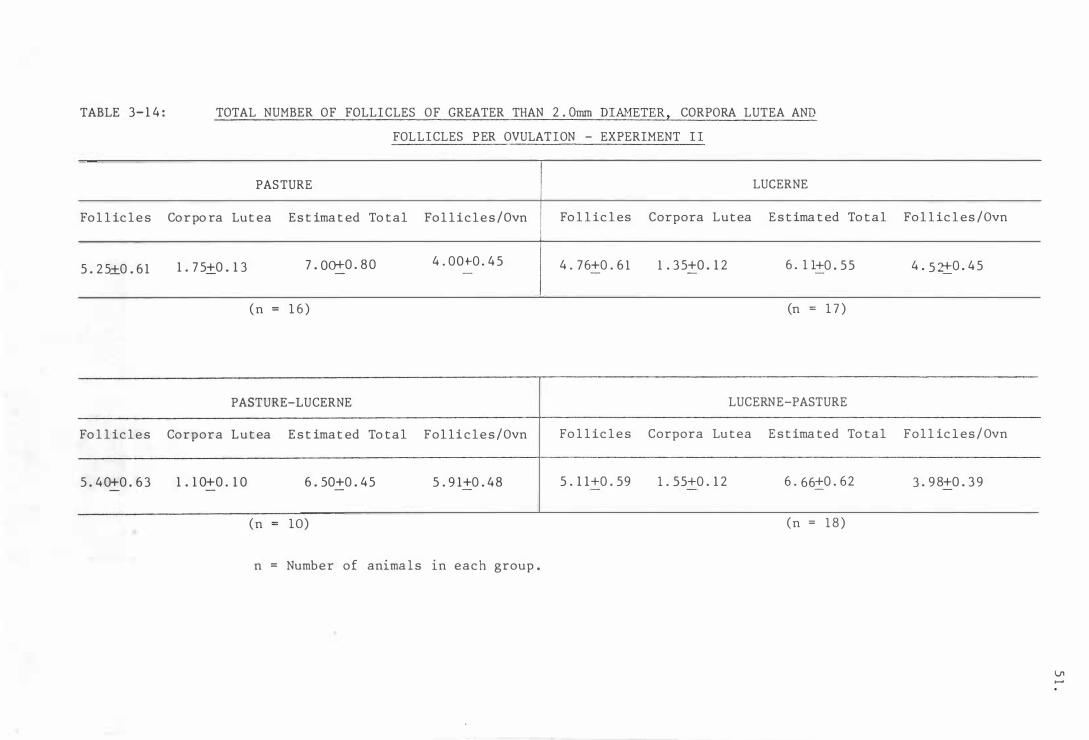
TABLE 3- 1 4 : TOTAL NUMBER OF FOLLICLES OF GREATER THAN 2 . 0mm DIAMETER, CORPORA LUTEA AND
FOLLICLES PER OVULATION - EXPERIMENT I I
PASTURE
Fo l l ic l e s Co r po ra Lut ea Es t ima t ed To tal Fo l l ic l e s /Ovn
5 . 2 5+0 . 6 1 1 . 7 5+0 . 1 3 7 . 0o+0 . 8 0 4 . 00+0 . 4 5
( n 1 6 )
PASTURE-LUCERNE
Fo l l ic l e s Corpora Lutea E s t ima t ed To tal Fol l i c l e s /Ovn
5 . 4 0+0 . 6 3 1 . 1 0+0 . 1 0 6 . 50+0 . 4 5 5 . 9 1+0 . 4 8
(n 10)
n = Number of anima l s in each group .
LUCERNE
Fo l l ic l es Corpora Lutea E s t ima t ed To tal Fo l l ic l es /Ovn
4 . 7 6+0 . 6 1 1 . 3 5+0 . 1 2 6 . 1 1+0 . 5 5 4 . 5 �0 . 4 5
(n = 1 7 )
LUCERNE-PASTURE
Fo l l ic l e s Corpora Lu tea E s t ima ted To tal Fol l ic l es / Ovn
5 . 1 1+0 . 59 1 . 55+0 . 1 2 6 . 66±0 . 6 2 3 . 9 8+0 . 3 9
(n = 1 8 )
V1 .......
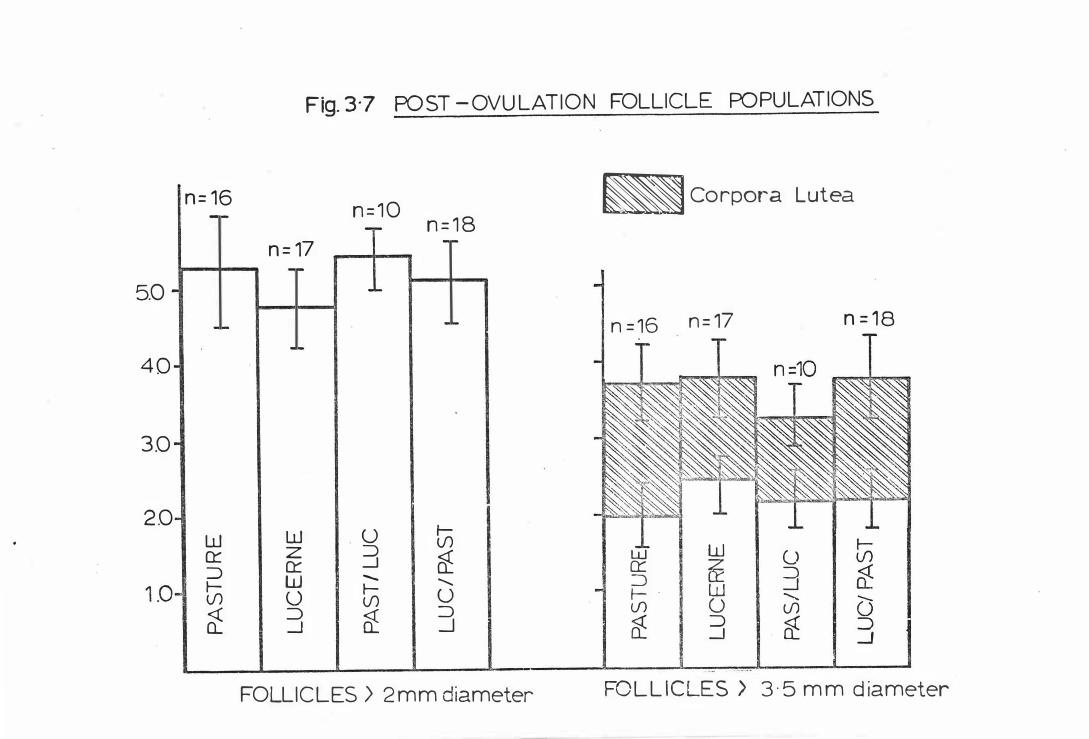
n = 16
5.0
4 0
3.0
2 .0 w 0::: �
1 .0-1 1-lf) �
Fig. 3·7 PO ST - OVU LAT I O N FOLLICLE POPULAT IONS
n = 17
w z 0::: w u ::) _j
n = 1 0 n = 1 8
u 1-lf) � & _J ........... 1- ......... lf) u � :J
_j I
FOLLIC LES > 2 mm diameter
� Corpora Lutea
n = 16 n = 17
....... w 0:: ::::J f- · lf) �
w
� w 0 � _J
- - - -
n = 1 8
(_) 3 ......... lf) 0
. � ::) _J
FO L L I C LES > 3 · 5 m m d iameter

TABLE 3 - 1 6 :
Ova rian Weight
5 2 .
OVARIAN AND UTERINE WEIGHTS (g) IN EWES GRAZING
PASTURE OR OESTROGEN IC LUCERNE
PASTURE LUCERNE PASTURE-LUG LUCERNE-PAST
(Mean ± SE) 2 . 7 2 + 0 . 1 6 2 . 58 + 0 . 14 2 . 4 2 + 0 . 1 9 2 . 52 + 0 . 1 2
U t erine We ight (Mean ± SE) 5 9 . 06 + 2 . 64 6 3 . 7 2 + 1 . 9 0 60 . 3 8 + 4 . 5 1 6 7 . 74 + 2 . 7 5
( i i ) Reproduc t ive Tra c t H i s to logy
Ther e were no s ignif ican t d i f f erenc es be tween groups in t he height
of epithel ial c el l s , in any of t he trac t t i s sue examined . Mean
ep i the l i a l c e l l heigh t s a r e s hown in Tabl e 3 - 1 7 .
TABLE 3 - 1 7 :
TISSUE
Vagina
Cervix
Uterus
EP ITHELIAL CELL HEIGHT ( � ) OF REPRODUCTIVE TRACT TIS SUE
F ROM EWES GRAZING PASTURE OR OES TROGEN IC LUCERNE
PASTURE
92 . 7 6 + 8 . 54
2 6 . 09 + l . 0 9
2 7 . 3 5 + 0 . 98
LUCERNE PASTURE-LUG LUCERNE-PAST
7 9 . 1 3 + 5 . 6 9 9 9 . 88 + 4 . 1 9 94 . 9 2 + 7 . 7 9
24 . 86 + 1 . 1 1 2 5 . 1 + 1 . 2 5 2 6 . 2 1 + 0 . 83
2 7 . 84 + 0 . 7 8 2 5 . 62 + 1 . 00 2 5 . 6 5 + 0 . 9 9
Fa llop ian Tube 24 . 4 9 ± 0 . 8 9 2 4 . 58 + 0 . 5 7 24 . 4 3 + 0 . 9 6 2 3 . 0 1 + 0 . 99

(G) SUMMARY
5 3 .
1 . The coumes tan l evels in the lucerne were modera t ely high and t h e
daily intake o f t h e ewes would have ea s ily exc e eded tha t required
to induce known oe stro genic respons es .
2 . Oes trogenic luc erne did no t e f f ec t proges tagen-induc ed synchroni
za t ion of oes trus , or the mean leng th o f oestrous cyc l es .
3 . The length o f the in t erval f rom the on set of o e s t rus to the pre
ovula tory surge of LH va ried ma rkedly be tween anima l s . The
coumes tans ha d no d e t ec table influenc e .
4 . The mea suremen t of the chlor ide ion con ten t o f c ervical mucus could
not be util i z ed to de t ermine the t ime o f ovul a t ion . Consequen t l y
t h e length o f t h e int e rval f rom the LH peak t o ovulat ion could
not be determin ed .
5 . The inges t ion o f coumes tans d id no t s ign i f icantly a l t er the sec r e t ion
o f lut einizing hormone . Secretion pa t t erns throughout the c yc l e
and over the pr e-ovulatory LH surge were unchanged .
6 . Oestrogen ic luc erne depressed ovulat ion ra tes by up to 2 9 per c en t ,
but only when lucerne wa s consumed for the la tter hal f o f the
o e s t rous cyc l e , wa s the depres sion o f ovulation rate s ta t i s t ic a l l y
s i gnif icant .
7 . The higher the ' na tura l ' ovu la t ion rate o f ewes , the grea t er � s
t h e coumestan-induced d epr e s s ion o f ovul at ion ra t e .
8 . The number s of fol l ic le s present in the ovar i e s were unchanged ,
a l though the number o f fo l l ic l es tha t ovu l a t ed wa s lowered by the
inge s t ion o f coume s tans .
9 . The mean diame t er o f c orpora lutea wa s not a f f e c t ed by the
inges t ion o f coumes tans .

54 .
10. The mean uterine we igh t s and ova r ian weight s o f ewes gra z ing
o e s t rogenic luc e rn e , d id no t d i f f e r s i gnif icantly f rom t ho s e o f
ewes gra z ing non-oe s t rogenic ryegr a s s / clover pa stur e , when
compared three days po s t -oes t rus .
1 1 . The mean h e igh t o f epi thel ial c e l l s in genital t ra c t t i ssue , d id no t
d i f f er be tween ewes gra z ing o e s tro genic lucerne and ewes graz ing
non-o e s t rogenic ryegra s s / c lover pa stur e , when compared three days
post-oes trus .

C H A P T E R I V
DI SCUSS ION

C H A P T E R I V
D I S CUSS ION
(A) OESTROGENIC LUCERNE
S evere infec t ion o f lucerne leaves by the fungi Pseudope z i za
0 med icagin is and Lepto spha erul ina brio s iana (Loper and Hans n , 1 9 64 ) ,
bo th o f which are common fungal pa thogens o f luc erne in New Z ea land
5 5 .
(Close , 1 9 6 7 ) , produc ed ma t er ia l of h igh coumes tan con t en t . A s imilar
degree of oes t rogenic po t ency wa s apparent under similar cond i t ions
with this lucerne stand in a p r evious t r ial (Mc Cutcheon , 1 9 7 6 ) .
The e s t ima tes o f daily in t ake of coumes tans sugges t t ha t p la sma
coumes tan conc en tra t ions woul d be we ll in exc e s s o f the l evels nec essary
to induce o e s t rogenic respon s e s ( Shu t t e t al . , 1 9 6 9 ) . A recen t trial
( Smith et al . , unpubl i shed da ta ) ha s s hown tha t as l i t t l e a s 2 5ppm
coume s t an in the feeds tuff i s suf f ic ient to suppre s s ovula t ion ra t e ,
and t he degree o f suppression i s inc r eased by a n in crease in coumes tan
conten t .
( B ) OESTRUS PHENOMENA
( i ) Oes t rus Synchron iza t ion
P roges t agen trea tment r e sul ted in a h igh propo r t ion o f ewes
s howing oes t ru s over a four day period , following sponge withdrawa l .
Some ewes wer e incapable o f r es ponding to proges tagen t r ea tmen t , owing
to hav ing bl ind trac t s or pregnant u t er i , or to lo s in g s ponges dur ing
synchron i z a t ion . I f t he s e an ima l s are d i sr egarded , t hen 9 3 p er c en t o f
ewes in Experimen t I and 9 1 percent o f ewes i n Exper imen t I I were
detec t ed in hea t over the synchron ized oes t rus . A h igh degree o f

s ynchrony was mai ntained a t the f o l l owing oes trus , wi th 8 2 percent
( Experimen t I) and 85 percent (Exper iment II) o f the ewes �n oes trus
over a comparab le four day per iod . The h igher propor t ion of ewe s
undetec ted in the se cond o e s t rou s cyc le � n Exper iment I , may re f l e c t
a n inadequacy in the per iod o f oes trus d e te c t ion , rather than any
b io logical phenomenon . The skewed d i s tr ibu t ion of the ons e t of
oestrus (FIG . 3- 1 ) sugge s ts tha t some ewe s may have been detec ted i f
rams were introdu ced ear l i er .
Oe s trogenic lucerne d id not a f f e c t the degree of oes trus
synchroni zat ion a t tai ned , or th e inc idence of ' s i l ent heats ' , a l th ough
behav ioura l oes trus ha s been r e l a ted to p l a sma oes trogen leve l s (Piper
56 .
and Foote , 1 9 6 8 ; God ing e t al . , 1 9 7 0 ; Scaramu z z i et al . , l 9 7 lb ) .
Conversely treatment o f cyc l ing ewe s w i th anti sera to endogenou s oes trogens
prevent s the o c currence of behav ioural o e s trus ( Scaramu z z i , 1 9 7 5 ;
Fairc lough e t al . , 1 9 7 6 ) , bu t the adm ini s t r a t ion o f anti-sera to
phy to-oe s trogens does no t prevent oes trus (Cox and Wi l son, 1 9 7 6 ) .
( i i ) The Oes trus-Ovu l a t ion Interva l
The wide var iat ion in the leng th o f the in terval from the ons e t
of oes tru s to the pre-ovu la tory LH peak , be tween indiv idua l ewes , i s
�n agr eement wi th ob servat ions by Lammin g , ( pers . corn . 1 9 7 8 ) . Lamming
sugges ted tha t th is is due to b io l ogical var ia tion ra ther than to
inadequate oes tru s de tec tion . Such wi de var iat ion b e tween ind ividu a l s
wou ld ma sk any d i f f erences induced b y lucerne .
The admi ni s trat ion of exogenous s tero id oe s trogens increases oes trus
dur a t ion ( S caramu z z i et al . , l 9 7 l a ; Land et al . , 197 2 ) . There is a
l i near r e l a t ionship b e tween oes trogen d ose and the durati on o f the i nduced
oes trus , however , ewe s exhib i t a r efrac tor i ne s s to repeated admin i s tra tion
o f oest rogen ( Scaramu z z i e t al . , 1 9 7 2 ) . Land et al . , ( 1 9 7 2 ) sugge s ted that
highly f ecund b reeds were more sen s i t iv e to exo genou s o e s t ro gens in

5 7
r e l a t ion t o thi s o e s t ru s r e s ponse . There i s no ev idence t o sugge s t that
coume s tan s a l ter the durat ion of o e s t ru s .
The f a i l ure of the cerv ical muc u s chloride te s t �n e s t ima t ing the
t ime of ovu l a t ion may hav e b een the r e su l t of poor s amp l ing technique s .
The e rra t i c changes in chl or ide concen trations may have been due to
var iat ion in the quan t i ty of mucus col le ct ed a t each san1p l ing . Turnbu l l
e t a l . , ( 1 9 6 7 ) and Li nd say & Franc i s ( 1 9 6 9 ) sugges t ed that chl o r ide
concentra t ions o f cerv ical mucu s may be u t il ized a s an a s say for phy to -oes t roge
intake in ewes . In this tr ial there were no d if ferenc e s de tec ted �n mucu s
chlor ide content , be tween ewe s in the lucerne and pas ture groups .
In conc lusion, these exper imen t s showed tha t there wa s con s iderab l e
var i a t ion � n the length o f the int erval from the onse t of oes trus t o the
pre-ovula tory LH peak . We were unab l e to determine the l ength of the
in terval f rom the pre-ovu l a tory LH peak to ovul a t ion . However Curnrning
et a l . , ( 1 9 7 3 ) found a high degree of cons tancy in the l ength of the
int erval f rom LH peak to ovul a t ion , in ewe s of wid e l y d i f fer ing age s and
under a number of trea tmen t s . Th i s would sugge s t tha t the re i s cons iderab le
var iat ion be tween anima l s i n the in terval f rom the onse t of o e s trus to
the pre-ovu latory LH peak .
(C ) LUTE IN IZ ING HORMONE SECRET ION
The d epres sion in l eve l s o f receptor-bound endogenou s oes trogens o f
ewe s graz ing lucerne (Newsome and Ki t t s , 1 9 7 7 ) sugge s t s tha t t o t a l endogenous
o e s tro gen conc en tra t ions may al so b e depre s s ed . A r e l a t ionsh ip be tween
s teroid o e s trogens and l u t e in i z ing hormone secret ion ha s c l e a r l y b een
e s tab l i s hed .
Ovar iec tomy , and the sub s equent f a l l in p l asma o e s trogen l evel s , r e su l t s
� n an e l eva t ion o f basal L H l ev e l s ( Reeve s , e t al . , 1 9 7 2 ; D iekrnan and
Ma l vern , 1 9 7 3 ) . The adminis trat ion of exogenous o e s trogens to ovar iec tomized
ewe s induces a biphas ic re sponse ; an init ial depr e s s ion in LH concentra t ions ,
f o l lowed by a large e l eva t ion s im i l ar to pre-ovul a tory l eve l s ( Rad f o rd e t a l . ,
1969 ; God ing et a l . , 1 9 7 0 ; Howl and and Palmer , 1 9 7 3 ) . Feed ing o e s tro genic

58 . c l over , of high i s of l avone content , to ov a r i ec tomi zed ewe s m 1 m 1 c s the
admin istra t ion of exogenous s t eroid oestrog ens on LH secre t ion
(Hearnshaw e t a l . , 1 9 7 7 ) .
In the presen t t r ia l s there were no inc rea ses in basal LH l evel s
in ewes graz ing oestrogenic l uc erne . Presumably , the depre s s ion o f
endogenous o e s t rogen level s , i f i t did occur , was no t o f suf f ic i en t
magni tude t o a f f ec t L H secret ion , o r al ternative ly , t h e coumestans
compensatro for l ower ed endogenous oes tro gen l evel s .
I f the coumes tans d o lower endo genous o e s t rogen l eve l s , i t is
f ea s ible that they may a f f ec t the pre-ovu l a t o ry LH surge as this is
pr imar ily a response to high l ev e l s o f c ircula t ing oes tro gen s o f the
p r evious day (Moore e t a l . , 1 9 69 ; Cox e t a l . , 1 9 7 1 ) . Oes t rad iol
inj ec t ions on days 3 -4 (Bol t e t al . , 1 9 7 1 ) o r on days 1 0- 1 2 ( How land
e t a l . , 1 9 7 1 ) of the oestrous cyc l e , wil l induce ' extra ' pre-ovul a tory
type LH surges in cyc l i c ewes . In the presen t trial such ex tra pre-
ovu l a tory LH p eaks were no t induced by the inge s t ion of coumes t ans ,
no r wa s the ' t rue ' pr e-ovula tory LH peak af fected .
The LH pro f il e s from ewe s graz ing o e s t rogenic l uc erne in this
t r i a l , failed to demon s t ra t e changes in e i t her the nega t ive-feedback
e f f e c ts of oes trogens on ba sal LH l evel s , o r their po s i t ive-f eedback
e f f e c t s on pre-ovulatory LH surges . I t is po ssible that either o r
both o f these o e s t rogenic e f f e c t s d id occu r , b u t were no t appa r en t .
To obta in accura t e quan t i t a t ive secre t ion p a t t erns, the in t erval b e tween
sampl ing mu s t be less than the hal f - l i f e o f the hormone (LH � l i f e is
approximately 2 0 minut es ) . S ampl ing a t such f requent in t erva l s would
demand the use of indwel l ing catheters and would a l so res t r ic t the
numb ers o f animal s used .
With the numbers o f anima l s involved in this tr ial , any maj o r
changes in L H secre tion pat t erns should have been detec ted with 2 -hourly

s amp l ing . Therefore i t i s conc luded tha t i f oes trogen i c lucerne d id
af f e c t secr e t ion pa t terns o f LH , the changes were no t e x tens ive .
(D) DEPRE S S ION OF OVULAT ION RATE S
Ewe l ivewe igh t s were recorded pr ior to , du r ing and a t the
comp l e t ion of the exper imen t s . There were no s i gni f i cant d i f f erence s
in ewe l ivewe igh t s be tween group s a t any of these t ime s , whi ch wou ld
ind i ca t e the abs ence of a ' body condi t i on ' e f f e c t on ovu l a t ion r a te
compar i sons . S imilarly the random a s s i gnment of ewe s to each trea tmen t
group shou l d have minimi sed any gene t i c d i f ferences in ovu l a t ion r a t e s
be twe en group s . Therefore i t may be a s sumed tha t any maj or var ia t ions
59 .
in ovu l a t ion r a t e s were due to the trea tment - the i nge s t ion o f oes t r ogeni c
lucerne .
In bo th experiment s the i nge s t ion o f oe s t rogen ic lucerne depre s s ed
ovu l a t ion rate s . Thi s i s 1 n agr eemen t wi th the ob s erva t i ons o f o ther
worker s ( Sc a l e s and Mo s s , 1 9 7 6 ; McCu tcheo n , 1 9 7 6 ; S c a l e s , et al . , 1 9 7 7 ) .
The mar g inal lev e l s of s i gni f i cance b e twe en groups ref l e c t s the
d i f f i cu l ty in ana l y s i ng d i screte d a ta ( number of ovu l a t i o n s ) wi th
re l a t ively sma l l numbers of anima l s . Th i s i s clearly ev ident f r om
Experiment I , when the depr e s s i on of ovu la t i on rate wi thin the br eed
subgroups (Romney and Border Le i ce s ter x Romney) i s cons idered . Al though
the degree to wh i ch ovu l a t ion r a t e s were de pre s sed in the s e group s ( 29 . 2
percent and 28 . 3 percent respe c t ively) i s comparab l e to the depre s s i o n
of ovu l a t i on r a te when a l l ewe s i n Experiment I ar e cons idered ( 29 percen t )
i t i s no t s ta t i s t i c a l l y s ignif i can t . However , when a l l ewe s are
cons idered , th i s s ame degree o f ovu l a t ion rate depr e s s i on 1s h i ghly
s igni f i can t ( p < O . OO l ) .

60 .
Tho se ewe s of higher f e cund i ty invo lved in the se trial s , were
a f f ec ted mo s t by the ingest ion o f coumes tans . In Exp e r iment I , mean
ovu l a t ion rate o f Bo rder Leice s ter x Romney ewe s is depressed by 0 . 7 3
ovu l a t i ons pe r ewe compared to 0 . 5 ovu l a t ions per ewe for Romney ewe s .
In a sim i lar manner , the ovu l a t ion r a te o f the contro l anima l s (Pas ture
gr oup ) wa s depre ssed une qua l ly b e tween Exper iment I and Expe r iment I I .
The con trol ewe s of Exper iment I had a higher inherent ovu l a t ion rate
than those o f Exper imen t II ( 2 . 29 ovu lat ions per ewe compared to
1 . 7 5 ovu l a t ions per ewe ) , and they were depressed to a greater degree by
oes trogenic luce rne ( 2 9 percent compared to 2 2 percen t ) . Th i s grea ter
sens i t ivi ty of hi ghly f e cund ewes to the coumes tans was also repor t ed
1n ear lier t r i a l s ( Scales and Mo s s , 1 9 7 6 ; S cales e t a l . , 1 9 7 7 ) .
This breed var iation 1n sens i t ivi ty to coume stans wou ld appear to
co rrespond to ob s erved breed d i f f erences 1n sens i t iv i t y to exogenous
s teroid oes trogen s . Th e admini s t rat ion of exog enous s te roi d oes trogens
to cycl ing ewe s increases the dur a t i on o f oes trus (Land , 1 9 7 0 ) and br eed s
of higher fecund i ty d i sp l ay a greater re sponse than d o the less pr o l i f i c
breeds (Land e t a l . , 1 9 7 2 ) . Ewes of high f ecund i ty are however , l e s s
sens i t ive t o exogenous oes tro gen- indu ced L H secret ions ( Land e t al . , 1 9 7 6 ) .
An apparent contradic tion to the higher sens i t iv i ty to coumes tans 1 n the
depr essio n of ovu lat ion r a te , is the repo r t of Land e t aL ( 1 9 7 6 ) of a l ower
sens i t iv i ty to the exogenous oes tradiol- induced depr e s s i on of ovu l a t ion
rate .
From the data co l l e c ted in Exper iment I I , i t is c l early evident
tha t it i s the inges tion o f c oumes tans in the l a t ter ha l f o f the
oes trous cyc le that is invo lved in ovu l a t ion r a te depre s s io n . The
gra z i ng of oes trogenic lucerne early inthe oes trous cyc l e had no e f f e c t
o n ovu l a t i on r a tes provided that the ewe s gra zed non-oes trogenic p as ture

in the l a t ter hal f o f the cyc le . The se f ind ings are con s i s tent wi th
the c la im tha t the number o f f o l l i c l e s whic h are dest ined to ovula t e ,
is d e t e rmined within the three days p r ior to oestrus (Land , 1 9 7 3 ;
Findlay and Cumming , 1 9 7 7 ; Bherer e t al . , 1 97 7 ) .
6 1 .
Hemica s t r a t ion o f ewes p rior to day 1 4 o f the oest rnus cycle , l eads
to a trans ient inc r ease in FSH levels coinc id ing with fal l ing o e s tradiol
levels , and increas ed foll icular growth in the r ema ining ovary over
the f ina l three days o f the cycle ( Ramire z and Sawyer , 1 9 74 ; Findl ay
end Cumming , 1 9 7 7 ) . In exper imen t s where the feeding o f lupin gra in
to ewes inc reased ovula t ion ra t e s , inc r ea ses in FSH l evels were a l so
reported ( Br i en e t a l . , 1 9 7 6) . This st rong . al though ind irec t ,
corr ela t ion b e tween high FSH l evels and incr eased ovulat ion rat e s ,
promp t s t he s ugges t ion tha t t he converse si tua t ion occurs in ewe s
graz ing lucerne . FSH l evels in ewe s inges t ing coumes tans may b e
depressed due to t h e inten s i f ica t ion of t h e negat ive-f eedback inf luence
of oest rogens on gonado trophin relea s e .
(E) FOLLICLE POPULATION S
I t h a s previously been shown t ha t breeds w i t h a high inher ent
ovula t ion ra t e , ovula t e a higher proport ion o f the Graaf ian follic les
tha t develop (Brad ford e t a l . , 1 9 7 1 ; Turnbull e t a l . , 1 9 7 7b ) . Th is
chara c t e ris t ic i s eviden t in the fol l ic l e populat ion data of Experimen t
I . When a l l corpora lutea and unru p tured follicles with a diamet er
o f gre a t er than 3 . 5mm a r e included in an e s t ima t e o f the l a rge f o l l ic l e
popul a t ion , t r end s between t h e number o f fo l l ic l es and ovul a t ion rate
are apparen t . Ewe s o f the Border Lec i s t er x Romney subgroup had
f ewer f o l l i c l e s per ovula t ion t han d id ewes f rom the Romney subgroup .
The l ow numbers o f ewes involved p robab ly p recludes the s e d i f f e r ences
f rom b eing s ta t i s t ically signnican t .

6 2 .
O e s t rogenic l ucerne incr ea s ed the numb er o f fol l i c l e s pre sent per
fo l l ic l e ovu lat ing in the ewes of h igh fecund i t y , a t r end which para l l eled
the correlation b e tween h igh fecund i t y and ovul a t ion ra t e d epress ion .
In Experimen t I I , a l l surface fol l icles o f a d iameter greater than
2mm were incor pora t ed in es t ima tes o f fol l ic l e popul a t ions , and s imilar
trends were shown . The inges t ion of o e s t ro genic lucerne in t he l a t ter
hal f o f the oest rous cyc l e increased the number of fol l ic l e s present
per ovul ation . Expo sure to coumes tan s in the ear ly s tages o f the cyc l e
d id no t a f f ec t f o l l ic l e number s . To tal fol l icle popula t ions , recorded
after s l ic ing t he ovaries , showed t rend s s imilar to tho s e s hown by
su rface f o l l i c l e popul a t ions .
I t would app ear tha t i f the coume s tans do suppress the ma tura t ion
of f o l l icles , then this suppression does no t restric t inc r eases in
fol l ic l e size . Al though fol l ic l e s devel op to a comparab l e s iz e in
ovar ies o f sheep graz ing lucerne , there i s some phy s io logic a l res tric t ion
whic h al lows only the mo st advanc e d fol l i c l es to develop the po t en t ia l
t o ovula te . Al t hough there is no direct ev idence , a d e f i c iency o f FSH
at this s tage o f foll icular developmen t would appear to b e a pos s ib l e
cau s e o f ovula t ion ra te depr e s s ion . Th e hemicas trat ion exp er imen ts
of Ramirez and Sawyer ( 1 9 74 ) and F ind lay and Cumming ( 1 9 7 7 ) give s trong
ind i rec t evidence of the requi rement for FSH . Fur thermore , r epor t s o f
the proper t ies o f exogenous oestro gens i n a l t er ing p i tui tary respons ive
nes s , could ac coun t for suppre s s io n of FSH l evels by the coume s tans .
The a dm in i s t r a t ion o f o e s tradiol t o ewes a f ew hours ( 4 -6) p r ior to
Gn-RH , depres ses the gonado t rophin response (Pellet ier and S igno re t , 1 9 7 0 ;
Liber tum e t a l . , 1 9 74 ) . I t wou l d al so appear tha t FSH i s much more
sens i t ive to this inhib i tory e f f ec t o f o e s t ro gen than i s LH ( S a l amons en
et a l . , 1 9 7 3 ; Baird an d Scaramuz z i , 1 9 7 6 ; F indlay and Cumming , 1 9 7 7 ) .

6 3 .
I t ha s been established , tha t in sheep tho se follicl es des t ined
to ovula t e would have a d iame ter of grea t er than O . Smm by day 6-7
of the o e s t rous cycle ( Turnbul l e t al . , 1 9 7 7a) . As the ingest ion o f
o e s tr ogenic lucerne i n the f i r s t hal f o f the cycle ( up to day 9 )
does no t inf luence ovu l a t ion ra t e s , t he ' in i t ia t ion ' o f follicular
growth is apparen tly unimpor tant in ovula tion ra te d epression . I t has
been sugge s t ed that large follicles re l ease a subs tanc e that suppresses
follicle ini t ia t ion (Peters et al . , 1 9 7 3 ) . As a consequence o f
lowered ovula t ion ra te , ewes graz ing lucerne will have a grea ter number
o f large f o l l icles presen t early in the next oes trous cyc l e . However ,
even if there is a depression in the number s of follicles ' in i t ia t ed '
i t is unl ikely that this woul d be a factor limi t ing the det ermina t ion
o f ovul a t ion ra tes .
There were no signi f i cant d i f f e r enc e s be tween groups in mean
ovar ian weights nor in the number of follicles present per gram o f
ovar ian t i ssue . Al t hough t hese are l imi t ed observa t ions , this would
sugge s t that the coumes tans do not hav e any maj o r e f fec t s on ovar ian
me tabolism tha t re sul t in weight inc r eases .
( F ) PROGESTERONE SECRETION
Proges t erone may inf luence foll icular growth ind ep end en t l y of i t s
e f f ec t s v i a the hypo t halomo-pitui tary axis . Fo llic ular growth is mor e
rapid i n ova ries that conta in a corpus lut eum ( Dufour e t al . , 1 9 7 1 b ) .
Inj ec t ion o f proge s t e rone into one ovary o f anoestrous ewes , toge ther
with sys t emic adminis t r a t ion o f PMS G , resul t s in mor e rapid f o l l icular
growth and in a grea t e r n umb er o f f o l l icles and corpora lutea in t he
inj ec t ed t han in the con t rol ovary ( Harned and Casiday 1 9 7 1 ) . Thi s
t r ea tmen t does not a l t er oes t rogen secre t ion ( Rexroad and Casida , 1 9 7 7 ) .

Smi th and Robinson ( 1 9 6 9 ) found the d iame t er o f a co rpus lut eum
( an e s t ima t e o f i t s ma s s ) to be h ighly correlated to proges t erone
conc en t r a t ion in the ovar ian vein blood pla sma ( an e s t ima te o f luteal
f unc t ion ) . The inges t ion o f coumes tan s did not inf luence l u t eal
f unc t ion as e s t ima ted by corpora lutea size .
(G) COUMESTAN S AND THE REPRODUCTIVE TRACT
64 .
The ing e s t ion o f coumes tans by overiec tomi zed ewes resul t s in the
expre s s ion o f the ' c lass ical ' o e s t rogenic character i s t i c s in the
r eproduc t ive trac t . The administra t ion o f 7 . 5 to 2 5mg of coumes trol
per day , induces maxima l s t imu l a t ion of u t e r ine growth (Braden e t a l . ,
1 9 6 7 ; Shut t e t al . , 1 9 6 9 ; Kel l y et al . , 1 9 76b) . The coumes tans
also induce corn i f ication o f r ep roduc tive trac t t issue in ovariec tomized
ewes , s imilar to tha t occurring at oes trus in in tac t ewes ( Braden et al . ,
1 9 6 7 ) . S imilarly , the ing e s t ion of o e s t rogenic luc erne causes the
secret ion of c ervical mucus , comparable to l evels found at oes trus ,
in ova r ie c tomized (Kelly , 1 9 7 2 ; Kelly e t al . , 1 9 7 6b ) or in in tact ano e s trous
ewe s (Kel ly et al . , 1 9 7 6a) . Synthe t ic coumes tans have s imilar e f f e c t s
( Shu t t e t al . , 1 9 6 9 ) .
In Exper imen t II reproduc t ive trac t s were examined thr ee days
po s t -oes trus , when t ho se o e s t ro g enic changes which occur at oe strus would
s t il l have b een evident . Ther e were no s ignif icant d if f er ences b e tween
ewes gra z ing pa s ture and ewe s grazing oes tro genic l uc e rne , in the
measurement of those oestro gen ic traits examined - i . e . u t er ine weigh t s
and e p i t hel ial c e l l he igh t s o f r eproduc t ive trac t t i s su e . Apparen t l y
t h e levels of en dogenous o e s t ro gens p resent a t o es t ru s masked t h e e f f ec t s
o f the inges t ed coumestan s .
I t ha s b een d emon s t ra t ed t ha t t he coumes tans compete with
o e s tradiol f o r oes trogen-receptors ( Shemesh � al . , 1 97 2 ) . I t wou l d

appear tha t , hav ing bound to oes tro gen r e c ep tor s , the coumes tan
rec eptor complex is capab le o f fully exp r e s s ing tho se oestro genic
changes induc ed by the oes trad iol -r eceptor complex , a t l ea s t within
65 .
the reproduct ive trac t . Newsome and K i t t s ( 1 9 7 7 ) recen t ly found tha t
there were r educed l evel s o f o es trad iol-receptor bind ing in ewes
graz ing oes trogen ic lucerne , al though o e s trogenic parame ters of the
r eproduc t ive trac t are unc hanged .
(H) CON CLUS IONS
The l evels of coumes tan intake by ewes grazing luc erne in these
exper imen t s were suf f ic i en t to induce a l l o f i t s o e s trogen ic e f f ec t s .
These oestro genic changes - inc rea s ed u t erine weigh t , inc reased c ervical
mucus produc t ion and corn i f ic a t ion o f r eproduc t ive trac t t i s sue - would
be r ead ily expr e s sed in ovar iec tomized ewes subj ec t ed to s imilar l evels
of coume s tan s . However , in in tac t cyc l i ng ewes these parame t er s d id
no t d i f f er from tho s e shown by ewe s graz ing non-oe s t rogen ic ryegra s s /
clover pa s tur e . The s ingl e chara c t er is tic o f the r eproduc t ive sy s t em
wh ic h was cons i s t en t ly a f f ec t ed by oestrogen ic luc ern e , was a r educ t ion
in the numbers of eggs ovula ted . Al tho ugh not pa r t of th is s tudy ,
observa t ions show tha t the eggs wh ich a r e produced , are capable of
normal f er t il i t y .
I t ha s b een shown tha t ovula t ion ra te i s propo r t iona l to t he
dura t ion o f o e s t rus (Land , 1 9 7 0 ; Hannahan and Qui rk e , 1 9 7 5 ) , whic h
in turn can b e influenced by exog enous o es t rogens ( S caramu z z i e t al . ,
1 9 7 1 a ; Land e t a l . , 1 97 2 ) . The admin i s tra t ion o f exogenous s t ero id
oes trogen s inc r ea s e s the l ength of the o estrus pe riod . However ,
a s ingle admin i s tra t ion o f exogenous o e s trogen is mo s t e f f ec t ive, a s
ewes show r efrac toriness to r ep ea t ed do s es ( S caramu z z i e t al . , 1 9 7 2 ) .

6 6 .
Mor l ey e t al . , ( 1 963) found tha t daily adminis tration of s t ilboes trol
to cyc l ing ewes decrea sed ovulat ion ra tes , a respon s e which pa ra l l e l s
tha t o f the coumes tans . I t may be tha t a s ingl e , appropria t ely- t imed
dose of coumest an s will inc r ea se the dura t ion of oest rus and inc r ea s e
ovu l a t ion ra t e s 1 a s d o e s a si ngle dose o f st ero id o e s t ro gens .
Although Newsome and Ki t t s ( 1 9 7 7 ) fo und tha t t he inge s t ion o f
coume s tans r educ ed the l evel s o f oes trad io l , oestrone and oes trio l
which bound to recep tors , they did no t measure to tal f ree endogenous
o es trogen l evels . Scaramu z z i ( 1 9 7 5 ) and Fa irc lough e t al . , ( 1 9 7 6 )
found tha t the dep r e s s ion o f endog enous o e s t rogen levels , b y the
administrat ion of an t i-sera , p r evented t he occurrence o f behav ioural
o es t r u s . The coume s tans d o not sign if icantly inc r ea s e t he inc id enc e
of ' s il en t ' hea t s . I t is a l s o unl ikely tha t the coumes tans are
involved in the expression of behavioural oestrus and ther efore ma sk
lowered endogenous oes trogen l evel s , as immunizat ion aga in s t phyto
oes tro gens does no t prevent o es trus ( Cox and Wilson , 1 9 7 6 ) .
The demonst ra tion in Exp er imen t I I , tha t the ing es t ion o f o e s t ro genic
luc erne la t e in the oes trous c yc l e depresses ovulat ion rates , wherea s
ing e s t ion early i n the cyc le ha s no effec t , is ana logous t o t h e e f f ec t s
o f hemica s tration before or a f t er day 1 4 of the cyc le . Un ilateral
ova r i ec tomy before day 1 4 results in a c ompensa tory increase in f o l l icular
growth in t he rema in ing ovary , and cons equen t ly , ovulat ion rates are
unc hanged f rom t ho se o f intac t ewes ( Ramir e z and Sawyer , 1 9 74 ; Findlay
and Gumming , 1 9 7 7 ) . Ther e i s an inc r ea se in FSH l evel s fol lowing
hemicas tra t ion and t he sub s equent fall in endogenous o e s t ro gen l evel s
(Findlay and Gumming , 1 9 7 7 ) . A s imilar relat ionship b e tween c hanges
in FSH level s and foll icular developmen t could be involved in the

6 7 .
coumestan-induced depres s ion o f ovula t ion ra t es . The ing e s t ion o f
o e s trogen ic lucerne may d epre s s F S H leve l s and consequen tly , inhibit the
la t t er s tages of foll icle ma turat ion , so d ecr eas ing the numb er o f
Graa f ian f o l l ic l e s l ikely to ovula t e . Smi th e t al . , ( unpub l i shed data)
recen t ly found t ha t t he administra t ion of Pregnan t Mares S er um
Gonado trophin (PMSG) to ewe s gra zing o e s t rogenic l uc erne , woul d overcome
the coumes t an-induced depression in ovula t ion rates .
A depr e s s ion o f FSH secret ion by the coumes tans could a r i s e f rom
ei ther the direc t ac t ion of the coumes tans on t he p i t u itary , in a manner
s imilar to the n ega t ive-fe edback ac t ion of the endo g enous o e s t ro gens , o r
from a coumes tan-induced inc r ease in endo genous oest rogen l eve l s .
Al though N ewsome and Ki t t s ( 1 9 7 7 ) showed tha t the coumes tans d epressed
the level s o f receptor-bound endo genous oestrogens , they did no t mea sure
leve ls o f ' free ' endo genous o e s t ro gens . The coumes tans may e f f ec t ively
inc r ease pla sma oestrogen level s a c t ing a t the hypo thalamo-p i tu i tary
leve l , by reduc ing the amount o f endo genous oestro gens bo und to
reproduc t ive tract oestro gen recep tors over the per i-oestrus period .
I t ha s been d emonstra ted t ha t the coumes tans compete with endogenous
oes trogens for oes trogen recep tors ( S heme sh et al . , 1 9 7 2 ) .
Al t erna t ive ly , t he coumes tans may l ower pla sma endo genous oes trogen
levels and a f f ec t the p i t u i tary r esponse to Gn -RH . A s t rong correlat ion
be tween t he oes trad iol : progest erone r a t io and the pitui tary r esponse
to Gn-RH ha s been r epor ted ( Chamley e t al . , 1 9 7 4a ; 1 9 74b ; P e l l e t ier ,
1 9 7 6 ) . Coumes tans may a l t er gonad otrophin secret ion pa t t erns by
lowering the oes trad iol : progesterone ra t io and c onsequently , the
response to Gn-RH .
F indlay and Cumming ( 1 9 7 7 ) sugges t ed that FSH secret ion i s much

more s en s i t ive to the nega t ive-f eedback a c t ion o f oes trogen t han is
LH . Such variat ion in s en s i t ivity ha s been d emonstra t ed with the
admin i s t rat ion o f exogenous o e s t rogens ( S alamonsen et a l . , 1 9 7 3 ;
Ba ird and Sca ramuz zi , 1 9 7 6 ) . I f there is such a wid e var ia t ion in
6 8 .
re sponse to coumes tans , a f a l l in FSH l eve l s o f suf f ic i en t magni tude to
in f luence ovu l a t ion ra tes may have occ urred , while the changes in LH
secretion were not large enough to be d e t ec ted by the proc edures used
in this tr ial .
The inge s t ion of coumes tans depresses ovula t ion rates but sel dom
preven t s ovul a t ion f rom occurr ing . How could a depression in FSH
l evel s accoun t for this ? It ha s been shown tha t the pr esence of FSH
l eads to an inc rease in the numb er of FSH r ec ep tors ; tha t FSH and
oest rad io l in concert inc rease LH receptor l eve l s , and tha t LH inc r ea s e s
L H receptor l evels (Richards a n d Midgley , 1 9 7 6 ) . I t is feasible t ha t
t he depress ion of FSH (and the subs equent inc r ease i n LH : FSH ratio )
af ter expo sure t o coumes tans , will r esul t in a depr e s s ion i n FSH
rec ep tor l evels and an inc r ea s e in LH receptor l evel s wi thin d eveloping
fol l ic l es . Presumably , a s FSH l eve l s are l imi ting , the ava ilab l e
FSH wi l l be taken u p preferen t ially b y the fol l ic l e ( o r f o l l ic l e s )
a t t h e highe s t s tage of developmen t , wh ich wil l possess the l a rge s t
numb er of F S H receptor s . Thi s wil l in turn l ead t o an incr ea s e in FSH
receptor l evels and con s equen t l y , to greater FSH up take . This
' ca scading ' e f f ec t wi l l r e su l t in the largest fol l ic l e (or f o l l ic l e s )
developin g to the stage o f po t ential ovul a t ion , a t t b e exp ense o f o ther
Graaf ian f o l l ic l e s .
The inc r ea s ed suscep t ib i l ity o f high fecundity b r eeds to the
coume s tan s may be related to FSH sec r e t io n . Al t hough there is no

69 .
ev idenc e to show tha t ewes o f high fecund i t y have h igh pla sma FSH
l evel s , i t has been shown t ha t s uch ewes are less s ens i t ive to the
nega tive-f eedback ac t ions of exogenous oes trogens ( Land et al . , 1 9 7 2 ;
1 9 7 6 ; Land , 1 9 7 6) . I f these ewes are al so less s ens i t ive to
endogenous oes trogen s , they would presumably have h igher FSH level s ,
a lower LH : FSH ra t io , and a s a cons equence o f these pla sma gonado t ro ph in
l evels , would develop mor e follicles with t he potential to ovula t e .
The mor e follicles with this po t en t ial to ovula te tha t are produced in
normal ewe s , the grea t er i s the pos sible inf luence of the coume s tans .
Dir e c t ev idence in sup por t o f this po s tula t ed a c t ion o f the coumes t an s
awa i t s f u r t her exper imen ta t ion .
POS S IBLE FUTURE TRIALS
Fur t her wo rk needs to be don e to determine the extent to which
FSH is impl ica t ed in the depression o f ovula t ion r a t e s , in ewes grazing
oes trogenic lucerne .
I f a depression in FSH secret ion does occur in ewes graz ing
oes trogenic luc ern e , and FSH is r espons ible for fo l l icular hyp e r t rophy
a f t er hemicas tra tion , t hen if hemica s t rated ewes a r e subj ec t ed to
coumes tan s , fol l icular hype r trophy will be impa ired . Trials involving
hemicas t r a t ion and o e s t ro genic luc erne t rea tmen t s at d i f fer en t t imes
in t he o e s trous cyc l e could d e t ermine thi s . The admini s t ra t ion o f
PMSG to h emicastrated , or intac t ewes , a t d i f f eren t t imes d ur ing the
cyc l e may add fur t her s up po r t to the claim that FSH i s involved in
d e t ermin ing ovula t ion r a t e s .
Fur t her as says o f pla sma gonado trophin l evels in ewes grazing
oestrogenic lucerne , are necessary . These s hould involve inten s ive
blood sampl ing regimes which would enable accura t e quant i ta t ive

7 0 .
secr e t ion r a t e s t o b e calculated . Inves tiga t ions into the mechanisms
which inf luence gonadotrophin s ec r e t io n , i f plasma gonado t rophin levels
are al tered by the coumes tans , could inc lude the e s t ima t ion of plasma
endogenous o e s t rogen l evels and the r e sponses to exo genous Gn- RH
of ewes gra z ing o e s t rogenic luc erne .
PRACTICAL APPLICATIONS OF THIS INVESTIGAT ION
In t ho se area s o f New Zealand wh ich exper ience dry summers , lucerne
often outproduces pas ture in l a t e summer and autumn . Luc erne may be
the only availabl e forage prior to ma t ing , when it is impor tant to
ma inta in breeding ewes a t high livewe igh t s .
The resul t s ob tained in these t r ials sugges t tha t o e s t ro gen ic
lucerne may b e util ized to achieve op t imum pre-ma t ing ewe l iv ewe ight s ,
wi thou t any de trimen tal e f f e c t s on reproduc t ive performanc e i f ewes are
removed from the luc erne eigh t days p r ior to ma t ing . In fac t removal
from lucerne as late as three days pr ior to the ons e t of o e s t rus may be
all that is required to res tore no rmal f ecund i ty . In prac t ical terms ,
i t could b e recommended to remove ewe s from oes trogenic luc erne
immedia t e ly pr ior to ma t ing , as only a small percen t age of ewes would
exhib i t oes trus w i thin this three day perio d . Al terna tively , i t could
be sugge s t ed to r emove ewes a week p r ior t o ma ting .
Fur t her work involving much larger numbers o f ewes , needs t o be
conduc t ed to verify t ha t removal f rom lucerne at mid-cyc l e r estores
normal ovulat ion rates , and henc e lambing rat e s . S imila r l y , the
e f f ec t s of longer t erm gra zing of oes trogenic lucerne prior t o r emoval
( such as f o r 1 -2 mont hs pre-ma t ing) , mus t also be inves t iga t e d before
these f indings are s t rongly advo c a t e d .

B I B L I 0 G R A P H Y

B I B L I 0 G R A P H Y
Ad ams , N . R . ( 1 9 7 3 ) . Relat ionship be tween u te r ine cys t s and ovar � an
ac tiv i ty in We s tern Au s tr a l ian ewe s . Aus t . Ve t . J . 4 9 : 1 7 6 .
7 1 .
Adams , N . R . ( 19 7 6 a ) . Cervical mucus change s in infer t i le ewe s previously
expo sed to oes tro genic sub terranean c lover . Re s . Ve t . Sc i . 2 1 : 5 9 -6 3 .
Adams , N . R . ( 19 7 6b ) . Al tered ovar ian func t io n in ewe s af ter pro longed
expo sure to p l ant oe s trogens . Theriogeno logy . �: 6 2 5 .
Adams , N . R . ( 1 9 7 7 ) . C ervical mucus and reproduc tive e f f i c iency �n ewe s
a f ter expo sure t o oe s trogenic pas ture s . Aus t . J . Agr ic . Re s . 2 8 :
4 8 1-489 .
Adams , N . R . ( 19 7 8 ) . Presis ten t l y a l tered sen s i t ivity to o e s t r ogens � n
ewes af ter pro longed expo sure t o p l ant oes trogens . Ab s t . 6 th As ia
and Ocean ia Cong . Endocr ino l pp . 7 1 - 7 2 .
Ad ler , J . H . and We i t zkin-Ne iman , G . ( 1 9 70) . Deme thy l a t i on o f methyl-0-
coumes trol by rumen micro-organi sms �n v i tro . Refuah . Ve t . 2 7 : 5 1 -55 .
Al exand er , G . and Ro s s i ter , R . C . ( 1 9 5 2 ) . The effects o f f er t i l i zer
tr ea tmen t s on the oes trogenic po tency o f Tr i fo l ium s ub ter raneum .
Aus t . J . Agr i c . Re s . l= 24-2 8 .
Al len , D . M . and Lamming , G . E . ( 1 9 6 1 ) . Nu t r i t i on and reproduc t ion �n
the ewe . J . Agr i c . S c i . (Camb . ) 5 6 : 6 9 -7 9 .
Al len, E . and Da i s y , E . A . ( 1 9 2 3 ) . An ovar ian hormone : pre l iminary repor t
on i t s l oc a l i z a t i o n , extrac t ion and par t ial pur i f i c a t ion and a c t ion on
te s t anima l s . J . Amer . Med . As s n . 8 1 : 8 1 9-8 2 1 .
A l l i son , A . J . ( 19 6 8 ) . The inf luence o f l ive we ight on ovu l a t ion rate in
the ewe . Pro c . N . Z . Soc . Anim . Prod . 2 8 : 1 15 - 1 1 9 .
A l l i s on , A . J . ( 19 7 2 ) . Produc tion o f cerv i c a l mucus i n ewe s treated w i th
exogenous proge s t agen and oe s trogen . Aus t . J . Agr i c . Re s . 2 3 : 4 7 3-48 1 .

Ander son, J . N . , Peck , E . J . , and C lark , J . H . ( 1 9 7 5 ) . Es trogen- induced
uterine re s ponses and growth : Re lat i onship to recep tor e s trogen
b i nd ing by u ter ine nuc l e i . Endocr ino l . 9 6 : 1 60- 1 6 7 .
Arms trong , D . T . , Gr inwi ch , D . L . , Moon , Y . S . and Zamecni k , J . ( 1 9 7 4 ) .
Inhib i t ion of ovu l a t i on in r abb i t s by intraf o l l icular inj e c tion o f
i ndome thac in and pros tagland in F . ant i s era . L i f e S c i . 1 4 : 1 29 - 1 40 .
Bahr , J . , Kao , L . and Nalbandov , A . V . ( 1 9 7 4 ) . The r o le o f the ca te
cho lamines and nerves in ovu l a t i on . B io l . Repr od . 1 0 : 2 7 3-290 .
Baird , D . T . , Baker , T . G . , McNa t t y , K . P . and Neal , P . ( 1 9 7 5 ) . Re l a t i onship
b e tween the secre tion of the corpus luteum and the l ength o f the
f o l l i cular pha se o f the ovar ian cyc l e . J . Repr od . Fe r t . 45 :
6 1 1 -6 1 9 .
Baird , D . T . and Scarmuz z i , R . J . ( 1 9 7 6 ) . Changes in the s e c r e t ion o f
ovar ian s teroids and p i tui t ary L H i n the per i-ovu l atory per iod i n
the ewe ; the e f f e c t of proges terone . J . Endocr ino l . 70 : 2 3 7 - 2 4 5 .
Ba ird , D . T . , Swans ton , I . and Scaramu z zi , R . J . ( 1 9 7 6 ) . Pu l sa t i l e r e l ea s e
7 2 .
o f LH and secret ion o f ovar ian s tero ids i n sheep dur ing the l u tea l phase
o f the oe s trous cyc l e . Endocrino l . 9 8 : 1 490- 1 49 6 .
Bar c ikowski , B . , Car l so n , J . C . , Wi l son , L . and McCr acken , J . A . ( 1 9 7 4 ) .
The ef f e c t s of endogenous and exogenous oes tradio l - 1 7 8 on the r e l ease
o f pro s taglandin F 2cr from the ovine u teru s . Endocrino l . 9 5 : 1 340- 1 349 .
Barre l l , G . K . ( 19 7 6 ) . Studies o f neuroendocrine mechani sms inf luenc i ng
seasona l var iat ion s in semen produ c t ion and plasma hormone leve l s i n
rams . Ph . D . The s i s . Mas s ey Univer s i ty .
Barre t t , J . F . , George , J . M . and Lamond , D . R . ( 19 6 5 ) . Reprodu c t ive
performance o f Mer ino ewe s grazing red c lover (Tr i f o l ium pratense L )
improved p a s ture or na tive pas ture . Aus t . J . Agr i c . Re s . 1 6 : 1 89 - 200 .
Barre t t , J . F . , Mou l e , G . R . , Braden , A . W . H . and Harr i s , A . N . A . ( 19 6 1 ) .
Cys t i c g l andu l ar hyper p l a s ia o f the endome tr ium i n ewe s in New South
Wa l e s and Queensland . Aus t . Ve t . J . 3 7 : 1 4 .

7 3 .
B a t terham , T . J . , Har t , N . K . , Lamb e r t on , J . A . and Braden , A . W . H . ( 19 6 5 ) .
Me t ab o l i sm o f oestrogenic i s o f lavone s i n sheep . Natur e . 206 : 509 .
Bat terham , T . J . , Shut t , D . A . , Har t , N . K . , Braden , A . W . H . and Tweeda l e , H . J .
( 19 7 1 ) . Me t ab o l i sm of intrarumina l l y adminis t e red 4- 1 4c-formonone t i n
and 4- 1 4c-b i ochanin A i n sheep . Au s t . J . Agr ic . Re s . �: 1 3 1- 1 3 8 .
Beck , A . B . and Knox , J . R . ( 1 9 7 1 ) . The acyla ted i s o f lavone g l yco s i d e s
f rom sub terranean c lover and r e d c l over . Au s t . J . Chem . 24 :
1 50 9 - 1 5 1 8 .
Beck , T . W. and Reeve s , J . J . ( 1 9 7 3 ) . Serum LH i n ewe s trea ted wi th
var ious dosages of 1 7 8- oes trad io l at three s tages of the anoes trous
sea son . J . Anim . Sci . 36 : 5 6 6-5 70 .
Benne t , D . ( 1 9 6 6 ) . The e f f e c t o f s t i lboe s tr o l d ipropriona te and leguminous
pas tures on the mammary secre t ion o f ewes . Aus t . Ve t . J . 4 2 :
3 64-3 68 .
Benne t t , D . , Mor l e y , F . W . H . and Axe l s en , A . ( 19 6 7a) . Bioas say re sponse
of ewe s to l egume sward s : 1 . Uter ine we i ght r e s pons e ; var iab i l i ty ,
c a l ibra ti on and pred ic t ion . Au s t . J . Agr i c . Re s . �: 485-49 4 .
Benne t t , D . , Mor l ey , F . W . H . and Axe l s en , A . ( 1 9 6 7b ) . Bioas say r e sponses
o f ewes to legume sward s : 2 . Uter ine we i ght res u l t s f r om sward s .
Aus t . J . Agr i c . Re s . � : 4 9 5 -504 .
Benne t ts , H . W . , Underwood , E . J . and Shier , F . L . ( 1 9 4 6 ) . A spec i f ic
breed ing prob l em o f sheep on sub terranean c lover pas tures i n We s tern
Aus tralia . Aus t . Vet . J . 2 2 : 2 - 1 1 .
Bhere r , J . , Ma t ton , P . and Dufour , . J . ( 1 9 7 7 ) . Fate o f the two large s t
f o l l ic l e s i n the ewe a f ter inj e c t ion o f gonado trophins a t two s tages
o f the oe s trous cyc le . Pro c . Soc . Exp . B i o l . Med . 1 5 4 : 4 1 2-4 1 4 .

74 .
Bicko f f , E . M . , Boo t h , A . N . , Lyman , R . L . , Livingston , A . L . , Thompson , C . R .
and De Ed s , F . ( 1 9 5 7 ) . Coume s t r o l , a new oestrogen i so l a t ed f rom
f orage crops . Sc ience . 1 2 6 : 9 6 9 - 9 7 0 .
Bicko f f , E . M . , Boo t h , A . N . , L iving s ton e , A . L . and Hendr ickson , A . P . ( 1 960a ) .
Ob s erva t ions o f the e f f ec t s o f drying on oes trogenic a c t ivity o f
a l f a l f a samp l e s o f varying ma tur i ty . J . Anim . S c i . 1 9 : 7 4 5 -7 5 3 .
Bicko f f , E . M . , Boo th , A . N . , Living s ton e , A . L . , Hendr ickson , A . P . and
Lyman , R . L . ( 1 9 59 ) .
and dried forage .
Det ermina t ion o f es trogenic a c t ivity in fresh
J . Anim . S c i . 1 8 1 000- 1009 .
Bicko f f , E . M . , Boo th , A . N . , Lyman , R . L . , L iving s ton , A . L . , Thomp son , C . R .
and Kohl er , G . O . ( 1 9 5 8 ) . I s o l a t ion o f a new est rogen f rom Ladino
c l over . �ric . Food Chem . 6 : 536-5 3 9 .
Bicko f f , E . M . , Liv ings tone , A . L . , Boo th , A . N . , Thomp son , C . R . , Hol lowe l l ,
E . A . and Beinhar t , E . G . ( 1 9 60b ) . Some variat ion in e s t rogenic
a c t iv i ty in f resh and dr ied whi t e c lover c lones and the Lad ino var i e ty .
J . Anim . S c i . 1 9 : 1 1 4 3 - 1 1 4 9 .
Bicko f f , E . M . , Livings tone , A . L . , Hendrickson , A . P . and Boo th , A . N . ( 1 9 6 2 ) .
Rel a t ive potenc ies o f several o e s t rogen-l ike compounds found in
forages . J . Agr ic . Food Chem . 1 0 : 4 1 0 -4 1 2 .
B icko f f , E . M . , Liv ings tone , A . L . and Guggo l z , J . ( 1 9 65 ) . Isol a t ion o f
pheno l ic compounds f rom Lad ino c l over . J . Agr ic . Food Chem . 1 3
Bicko f f , E . M . , Liv ings ton , A . L . and W i t t , S . C . ( 1 964) . I s o l a t ion o f
coumes trol and o ther phenol ic s f r om a l fal fa b y coun t ercur r en t d i s t -
r ibut ion . J . Pharm . Sc i . 5 3 : 1 4 9 6- 1 4 9 9 .
1 5 1 - 1 55 .
Bindon , B . M . , Chang , T . S . and New ton-Turner , H . ( 1 9 7 1 ) . Ovar ian r es ponse
to gonado troph ins by Mer ino ewes selec t ed for f ecund i t y . Aus t . J .
Agr ic . Re s . 22 : 809-820 .

B j e r s ing , L . , Hay , M . F . , Kann , G . , Moo r , R . M . , Na f to l in , F . ,
Scaramu z z i , R . J . , Sho r t , R . V . and Youngla i , E . V . ( 1 9 7 2 ) .
7 5 .
Changes in gonado trophin s , ovar ian s t eroids and follicular morphol o gy
in sheep at o es trus . J . Endoc r inol . 52 : 4 6 5-4 7 9 .
Bognadove , E . M . ( 1 964) . The role o f the brain in the regula t ion o f
p i t u i tary gonado t rophin secre t ion . Vit . and Harm . 2 2 : 206-260 .
Bol t , D . J . and Hawk , H . W . ( 1 9 7 1 ) . C . L . regres s io n induc ed by E 1 E 2 8
and DES in ewe s . J . An im . S c i . 33 : 24 9 .
Bol t , D . J . , Kel l y , H . E . and Hawk , H . W . ( 1 9 7 1 ) . Rel ease o f LH by
o e s t ra diol in cycl ing ewes . B io l . Reprod . 4 : 3 5 -4 0 .
Bradbury , R . B . and Wh i t e , D . E . ( 1 9 5 1 ) . The chemistry o f sub t erranean
clover . 1 . The isolat ion o f formononetin and gen i s t e in .
J . Chem . Soc . 1 9 5 1 344 7 - 344 9 .
Bradbury , R . B . and Whi t e , D . E . ( 1 9 54 ) . Es trogen s and rela ted sub s tance s
i n p lan ts . Vit . and Harm . 1 2 : 207-2 33 .
Braden , A . W . H . , Har t , N . K . and Lamber ton , J . A . ( 1 96 7 ) . The o e s t rogenic
ac t iv i ty and me tabol i sm o f certain i s o f l avone s in sheep .
Aus t . J . Agr ic . Res . � : 335-34 9 .
Bradford , G . E . , Qui rke , J . F . and Har t , R. ( 1 9 7 1 ) . Na tural and induced
ovula t ion rate o f Finn ish Landrace and o ther breeds o f sheep .
Anim . Prod . 1 3 : 6 2 7 -635 .
Brand , A . and De Jong , W . H . R . ( 1 9 7 3 ) . Qualita t iv e and quan t i t a t ive
mic romorpholo gical inve s t iga t ions o f the ter t iary foll i c l e popul a t ion
dur ing the oestrous cyc l e in sheep . J . Reprod . Fer t . 3 3 : 4 3 1 -4 3 9 .
Br ien , F . D . , Bax t e r , R . W . , Findlay , J . K . and Gumming , I . A . ( 1 9 7 6 ) .
E f f e c t o f l up in grain supplementation on ovu l a t ion rate and pla sma
FSH concentrat ion in mai den and ma ture Mer ino ewes .
S o c . Anim . Prod . 1 1 : 2 3 7 -240 .
Pro c . Aus t .

7 6 .
But enand t , A . and Jacob i , H . ( 1 9 3 3 ) . Uber die Dar s tell un g e ines
kry s ta l l i s i er t en pfla z l ichen Tokokinins ( Thelykinins) und s eine
Iden t i f i z ierung mit d em « -Fo l l i kelhormon .
Z . Phys io l . Chem . 2 1 8 : 1 04 - 1 1 2 .
Butl er , G . W . , S t eemer s , M . A . and Wong , E . ( 1 9 6 7 ) . The e f f ec t o f
I no trogen , phosphorus and pota s s ium supply on the isoflavone con t en t
o f l eaves o f red c lover (Trifol ium pra t ense) . N . Z . J . Agr ic . Res .
1 0 : 3 1 2 -3 1 5 .
Capony , F . and Ro chefor t , H . ( 1 97 7 ) . I n v i t ro and In vivo in t erac t ions o f
( 3H ) d ime thyl s t ilbo es trol with the es trogen recep t o r . Mo l . and Cel l .
Endocr inol . � : 4 7 -64 .
Carl son , J . C . , Barc ikowski , B . , Cargill , V . and McCracken , J . A . ( 1 9 7 4 ) .
The blockade o f LH release by indometha c in .
Metab . 3 9 : 399-402 .
J . Cl in . Endocrinol .
Carson, R . S . , Findlay , J . K . and Bur ger , H . G . ( 1 97 8 ) . Gonado trophin
receptors wi thin the ovine ovar ian foll icl e . Ab s t s . 6th Asia and
Oc ean ia Cong. Endoc rinol . 1 9 7 - 1 98 .
Cayen , M . N . and Common , R . H . ( 1 96 5 ) . An inves tiga t ion o f ur inary
conver s ion produc t s of coume s t rol in the fowl . B iochem. B iophys .
Ac ta . 1 00 : 567-5 7 3 .
Chaml ey , W . A . , F indlay , J . K . , Cumming , I . A . , Buckma s t er , J . M . and Godin g ,
J . R . ( 1 9 74 a ) . Effect o f pr egnancy on LH response to synthetic
gonado t roph in-r eleasing hormone in the ewe . Endocrinol . 94 : 2 9 1-2 9 3 .
Chamley , W .A . , F indlay , J . K . , Jona s , H . , Cumming , I . A . and God ing , J . R .
( 1 9 7 4 b ) . E f f e c t o f pregnancy on the FSH respons e to synthetic
gonado t rophin-r eleasing hormone in ewe s . J . Reprod . Fer t . 3 7 :
1 09- 1 1 2 .

Chamley , W . A . , Cumming , I . A . and Goding , J . R . ( 1 9 74 c ) . S tudi es on
oestro gen b inding to u t erine cytosol r ec ep tor s prepared f rom
norma l and c lover-infer t il e ewes . J . Reprod . Fer t . 3 6 : 4 9 1 .
7 7 .
Chang , M . C . ( 1 96 6 ) . Transport o f eggs from the Fallopian tub e to the
uterus as a func t ion of oestrogen . Na ture 2 1 2 : 1 048- 1 04 9 .
Chan g , T . S . ( 1 963) . Geno type-pas ture int erac t ion in the f er t i l i ty o f
Romney Marsh ewe s . Proc . N . Z . Soc . Anim . Prod . 2 3 : 1 00- 1 06 .
Channing , C . P . ( 1 9 7 0 ) . Inf luences of t he In vivo and In vitro hormonal
environment upon lut ein iza t ion in t i ssue cul t ur e .
Ha rm . Res . 2 6 : 589 -6 2 2 .
Rec en t Prog.
Cheng , E . , S tory , C . D . , Payn e , L . C . , Yoder , L . and Burroughs , W . J .
( 1 9 5 3 ) . De t ec t ion o f o es t rogenic subs tanc es in alfalfa and c lover
hays f ed to fatten ing l ambs . J . Anim . S c i . 1 2 507 -5 14 .
Cheng, E . , Yoder , L . , S tory , C . D . and Burroughs , W . J . ( 19 54 ) . Oes trogenic
ac t iv i ty of some iso f 1avone deriva t i es . Sc ience 1 2 0 : 5 7 5-5 7 6 .
Cidlowsk i , J . A . and Mul doon , T . G . ( 1 9 7 6 ) . Dis s imilar e f f ec t s o f ant i
e s tro gens upon e s tro gen r ecep tors i n respons ive t i s sues o f mal e
and f emale rats . Bio l . Reprod . 12 38 1-389 .
Cidlowski , J . A . and Mul doon , T . G . ( 1 9 7 8 ) . The dynamics o f int racellular
es trogen rec ep tor regul at ions as influenced by 1 7 � -es tradiol .
Biol . Reprod . � : 234-24 6 .
Clar k , J . H . , Paszko , Z and P eck , E . J . ( 1 9 7 7 ) . Nuc l ear b inding and
retention of the r ec ep tor-o e s trogen complex : Relat ion o f the
agon is t ic and antafJ)nis t ic prcpert ies of estrio l . Endoc r ino l .
1 0 0 : 9 1 -96 .
Clark, N . T . ( 1 9 6 5 ) . A comparison o f the r eproduc t ive perfo rmance o f
ewes grazing Dwal ganup or Bacchus Marsh sub terranean c lover o r
a mixture o f lucerne a n d Burr medic before a n d during mat ing.
Aus t . J . Exptal . Agri c . 5 : 106- 1 09 .

C l os e , R . C . ( 1 9 6 7 ) . Di sease of lucerne 1 n New Zealand . From ' The
Lucerne Crop ' . ed . R . H . M . Langer . Pub l i shed : A . H . & A . W . Ree d .
Wg tn . N . Z .
C o l eman , E . A . , S tab , M . and Ke im , W . F . ( 1 9 6 5 ) . Uterotrophic ac t iv i ty
in Lotus corni culatu s L . Crop S c i . 5 : 2 7 6- 2 7 7 .
Coop , I . E . ( 1 9 6 6 a ) . Effec t o f f lushi ng o n r eproduc t ive performance of
ewes . J . Agr i c . Sci . �: 305-3 2 3 .
Coop , I . E . ( l9 6 6b ) . The re spons e o f ewes to f lus hing . Wor l d Rev .
Anim . Prod . 2 ( 4 ) : 6 9 - 7 4 .
7 8 .
Coop , I . E . ( 1 9 7 7 ) . Depress ion o f l amb ing percentage f r om ma t i ng o n lucerne .
Proc . N . Z . Soc . Anim . Prod . 3 7 : 1 4 9 - 1 5 1 .
Coop , I . E . and C l ark , V . R . ( 1 960) . The reproduc tive perfo rmanc e o f
ewe s mated on lucerne . N . Z . J . Agr i c . Re s . l: 9 2 2-9 3 3 .
Cos t e l l o , C . H . and Lynn , E . V . ( 19 50 ) . An oe s trogenic compound f r om
l i quor ice roo t extrac t . J . Amer . Pharm . 39 : 1 7 7 - 1 8 1 .
Cox, R . I . and Braden , A . W . H . ( 1 9 7 4 ) . The me tab o l i sm and phys i o l o g i cal
effects o f phy to-oes trogens in l ives tock . Proc . Aus t . Soc . Anim .
Prod . 10 : 1 2 2- 1 29 .
Cox , R . I . , Mat tner , P . E . and Thorbur n , G . D . ( 1 9 7 1 ) . Changes i n ovar 1an
secre t ion of oe s trad io l - 1 7 8 around oes trus in the sheep . J .
Endocr ino l . 49 : 345-346 .
Cox , R . I . , Thorburn G . D . , Curr i e , W . B . and Re s ta l l , B . J . ( 1 9 7 4 ) .
Interr e l a t ionshi ps be tween oes trogens and pros tag l and in F
concentr a t i ons i n the u tero-ovarian venous plasma o f the ewe .
J . Reprod . Fer t . 36 : 448-449 .
Cox , R . I . and Wi l son , P . A . ( 19 7 6 ) . E f f e c t s of immuni z i ng sheep to
s teroid oe s t ro gens and phyto-oe s trogen s . J . Reprod . Fer t . 46 : 5 2 4 .
Cumming , I . A . ( 19 7 7 ) . Rel a t ionships i n the sheep o f ovu l a t ion r a t e wi th
l iveweigh t , br eed , season and nut r i t i o n . Aus t . J . Expt a l . Agr i c .
and Anirn . Hu sh . 1 7 : 234-24 1 .

/ ':J , Cumming , I . A . , Buckmaster , J . M . , Blockey , M. A . , God ing , J . R . ,
Winf ield e . G . and Baxter , R . W . ( 1 97 3 ) . Constancy o f interval
b etween LH release and ovulation in the ewe . Biol . Rep rod . 2_
Cumming , I. A . and McDonal d , M . F . ( 1 9 6 7 ) . The production o f ova by
New Zealand Romney ewes following ho rmonal s t imul a t ion .
N . Z . J . Agr ic . Res . lQ : 226-2 36 .
2 4 -29 .
Curnow , D . H . , Robinson , T . J . and Underwood , E . J . ( 1 94 8 ) . Oestrogenic
ac t ion of extrac ts of subterranean c lover (!. sub t erranean �- var .
Dwalganup) . Aus t . J . Exptal B iol . Med . S c i . 26 : 1 7 1 - 1 80 .
Curnow , D .H . and Ro ssiter , R . C . ( 1 9 5 5 ) . The occurrence o f genistein
in subt erranean clover (!. sub t erranean �) and o ther Trifol ium
specie s . Aus t . J . Exptal B iol . Med . Sci . 33 : 24 3 � 248 .
Da t tagup ta , P . C . , Khastgir , H . N . and S engup ta , P . ( 1 960) . Inves t igations
on the st ruc ture of psora l idin . Chem . and Ind . 1 9 60a : 4 8 -4 9 .
Davies , H . L . and Dud z inski , M . L . ( 1 9 6 5 ) . Fur ther s tudies on o e s t rogenic
a c t ivity in strains of sub terranean c lover (Trifol ium sub terranean �)
in so uth-we s t ern Aus tral ia . Aus t . J . Agr ic . Res . � : 9 3 7 -9 5 0 .
Davies , H . L . , Ro s s it er , R . C . and Mal l er , R . ( 1 9 7 0 ) . The effects o f
d i f ferent cul tivars of sub t erranean clover (!. sub terranean L )
on sheep reproduc tion in the south-west of Wes tern Aus tral ia .
Aus t . J . Agric . Res . ll : 3 5 9 -3 69 .
Diekman , M . A . and Malvern , P . V . ( 1 97 3) .
oes trad iol on LH pa t terns in ewes .
Effect o f ovariectomy and
J . Anim . S c i . 37 : 562-56 7 .
Dorhn , M . Faur e , W . , Pol l , H . and Blo tevogel , W . ( 1 9 2 6 ) . - ci t ed by
K. Kal lela , Thesis , Hels inki , 1 964 . Med . Kl in . (Munich) 2 2 : 1 4 1 7 .
Ducker , M . J . and Boyd , J . S . ( 1 9 7 7 ) . The effect o f body size and body
condit ion on the ovulat ion rate o f ewes . Anim . Pro d . 2 4 : 3 7 7 - 38 5 .
Dufour , J . ( 1 9 7 6 ) . Carac terist iques des foll icul es ovariens lors de
l ' ovula tion s imple ou double chez le brebis e t developpment de
l eurs corps j aunes . Inf orma t ions Veterinaires 18 : 1 6- 1 7 .

Dufour , J . , Gin ther , O . J . and Cas ida , L . E . ( 1 9 7 Ja) . _ Compensatory
hypertrophy a f t er unila t eral ovariectomy and the des truc t ion o f
8 0 .
f o l l ic l es i n t he anoes t rous ewe . Proc . Soc . Exptal B io l . Med . 1 38
1 068- 1 07 2 .
Dufour , J . , Gin ther , O . J . and Cas ida , L . E . ( 1 9 7 1b ) . Corpus lut eum
a c t ion on ovar ian fo l l icular developmen t after des t ruct ion o f
mac ro scop ically v i s ib l e follicles in ewe s .
Med . 1 38 : 4 7 5-4 7 8 .
Proc . Soc . Exptal . B io l .
Dufo ur , J . and Ma t ton , P . ( 1 9 7 7 ) . Iden t i f ica t ion o f ovar ian f o l l i c l e s
a t es trus and developmen t o f the ir ensu ing corpora lutea in s ingle
and mul t ip l e ovula t ing ewes on two feed ing regimes .
S c i . 5 7 : 64 7 -652 .
Can . J . Anim .
Ea s t , J . , Underwood , E . J . and Bennet t s , H . W . ( 19 4 9 ) . Oes t ro genic
effects o f sub ter ranean clover ( T sub terranean � var . Dwalganup)
Pro tec t ive ac t ion o f androgens in the cas tra t e mal e .
B iol . Med . Sc i . 2 7 : 1 0 5 - 1 1 3 .
Aus t . J . Exptal .
Eas twoo d , K . C . and McDona ld , M . F . ( 1 97 5 ) . Reproduc t ive d i f f erences in
two-year -o l d Romney and Border Le i c e s ter cro ss Romney ewes a f t�r
gonad o trophin treatmen t . N . Z . J . Agr i c . Re s . 1 8 : 1 09 - 1 1 7 .
Ech t ernkamp , S . E . and La s t er , D . B . ( 1 9 7 6 ) . P lasma LH conc en t r a tions for
pr e-puberal , po s t -pub eral , anoest rous and cyc l ing ewes o f varying
fecund i t y .
Ed ey , T . N . ( 1 9 6 8 ) .
J . Anim . S c i . 4 2 : 444-4 4 7 .
Bodyweight and ovu l a t ion ra t e in sheep .
Soc . An im . Pro d . 7 : 1 8 8 - 1 9 1 .
Proc . Aus t .
Edwards , R . G . ( 1 9 7 4 ) . Fol l icular f luid . J . Reprod . Fer t . 12 : 1 89-2 1 9 .
Espe y , L . L . ( 1 97 4 ) . Ovar ian prot eolyt ic enzymes and ovula t ion .
B io l . Reprod . 1 0 : 2 1 6-2 3 5 .

8 1 .
Espey , L . L . and Coon s , P . J . ( 1 9 7 6 ) . Fac to r s which inf luence ovula tory
d egradat ion o f rabbit ovar ian follicl es . Biol . Reprod . 1 4 : 2 33-24 5 .
Fairclough , R . J . , Smith , J . F . and Peter son , A . J . ( 1 9 7 6 ) . Pas s ive
immunizat ion against o es t radio l - 1 7 8 and i t s e f f ec t on luteolys i s ,
oestrus and ovu la t ion in t he ewe . J . Reprod . Fer t . 48 : 1 6 9 - 1 7 1 .
Farka s , G . L . and Kir aly , Z . ( 1 9 6 2 ) . Ro le o f pheno l ic compounds in the
physiology of p lan t d i s ea s e s and d i s eas e resis tanc e .
Phytopa th . Z . 4 4 1 0 5 - 109 .
Fellner , 0 . 0 . ( 1 9 2 6 ) . - c i t ed by K . Kallela , Thes is , H e l s ink i , 1 9 64 .
Med . Kl in . (Mun ich) � : 1886- 1 888 .
Ferguson , E . R . and Ka t z enell enbogen , B . S . ( 1 9 7 7 ) . A compara t ive s tudy
o f an t i- es trogen a c t ion : Temporal pat terns o f antagonism o f
e s tro gen- s t imulated uterine growth and e f f ec t s o f e s t ro gen receptor
levels . Endocr inol . 1 00 1 2 4 2 - 1 2 5 1 .
Findlay , J . K . and Gumming , I . A . ( 1 9 7 6a ) . F SH in the ewe : E f fec t s of
sea son , l ivewei ght and plane o f nut r i t ion on pl asma FSH and ovula t ion
ra t e . Biol . Reprod . 1 5 : 3 3 5-34 2 .
Findlay , J . K . and Gumming , I . A . ( 1 9 7 6b ) . Increases in ovula t ion r a t e
i n sheep fol lowing admin i s t ra t ion o f a n LH-RH analogue .
1 5 1 1 5- 1 1 7 .
Biol . Reprod .
F indlay , J . K . and Gumming , I . A . ( 1 9 7 7 ) . The e f fec t o f un ila teral
ovar iectomy on p lasma gonadotrophin l evel s , es t rus and ovulat ion
rate in sheep . B iol . Reprod . l2 1 78 - 1 8 3 .
F indlay , J . K . , Gumming , I . A . , Chamley , W . A . , Buckma s t e r , J . M . , Go ding, J . R .
and Hearnshaw , H . ( 1 9 7 3) . The release o f LH by o es t radio l - 1 7 8 and
a synthe t ic Go-RH in ewes a f fected with Clover Disease . J . Reprod .
Fer t . 3 2 : 34 1 .

F i r t h , J . H . • , Salamon , S . and Maxwel l , W . M . C . ( 1 9 7 7 ) . Observations on
oes trus wi thou t ovulation in sheep . Pro c . Aus t . Soc . B iol . Reprod .
9 : 58 .
Fle t cher , I . C . ( 19 7 1 ) . E f f e c t s o f nu tr i t ion, livewe i gh t and season on
the inc idence o f twin ovu lation in Sou th Aus tralian s tr ong wool
Merino ewe s . Au s t . J . Agric . Res . 2 2 : 3 2 1-330 .
8 2 .
Fle t cher , I . C . and Lindsay , D . R . ( 1 9 7 l a) . Effect o f oe s tr ogen on oes trus
b ehavi our and its var i a t i on wi th season in the ewe . J . Endocr inol 50 :
6 85-69 6 .
Fle tcher , I . C . and Lindsay , D . R . ( 1 9 7 lb ) . E f f e c t o f rams on the dura t i on
o f oes trus behaviour � n ewes . J . Reprod . Fer t . 25 : 253-259 .
Ford , S . P . , We ems , C . W . , P i t t s , R . E . , Pex ton , J . E . , But cher , R . L . and
Inskeep , E . K . ( 1 9 7 5 ) . E f f ects o f o e s trad iol- 1 7 8 and proge s t er one
on pros tag l and in F in she ep uteri and uterine venous plasma .
J . Anim . Sc i . 4 1 : 1407- 1 4 1 3 .
Foster , J . P . and Cri ghton , D . B . ( 1 9 7 4 ) . Lu teiniz ing Hormone (LH) re lease
af ter single inj e c t ions of synthe t i c LH-releas ing hormone (LH-RH) in
the ewe a t three d i f f erent reproduc t ive s tages and compari sons w i th
natural LH release a t oe s tru s . Theriogeno logy. l: 8 7 - 1 00 .
Franc i s , C . M . and Mil l inton , A . J . ( 1 9 6 4 ) . Var ietal var i a t i on in the
i sof lavone con tent of sub terr anean c lover : I t s e s t imat ion by a
microtechnique . Au s t . J . Agr ic . Re s . �: 5 5 7 - 5 6 4 .
Franc i s , C . M . and Mi ll ing ton , A . J . ( 1 9 6 5 a ) . We ther b ioas say of annual
p a s ture legumes : IV . The oes trogenic a c t iv i t y of annual med ic
p a s tur e s . Aus t . J . Agr i c . Re s . 1 6 : 9 2 7 -9 3 5 .
Franc i s , C . M . and Mi l l ing ton , A . J . ( 19 6 5 b ) . Isof lavone mu tati ons � n
sub terranean clover : I . The i r p rodu c t ion, charac t er i s t i c s and
i nher i tance . Aus t . J . Agric . Re s . 1 6 : 5 6 5 -5 7 3 .
Franc i s , C . M. and Mi l l ington , A . J . ( 19 6 5c ) . We ther b ioas say o f annual
pas tur e legumes : I l l . The oes trogenic potency o f dry sub te r r anean
clover pas tures , and of leaf b lade and p e t iole i n the green s tate .
Aus t . J . Agr ic . Res . 1 6 : 23-30 .

8 3 .
Franc i s , C . M . , Mi l l ing ton , A . J . and Ba i l ey , E . T . ( 1 9 6 7 ) . The d i s t r ib u t ion
o f oes trogenic i s o f l avone s in the genus Tr i fo l ium . Aus t . J . Agr i c .
Re s . 1 8 : 4 7 -54 .
Gibbons , R . A . and Mat tner , P . ( 1 9 6 6 ) . S ome aspe c t s o f the chemi s try of
cerv i c a l mucu s . Int . J . Fer t . 1 1 : 3 6 6-3 7 2 .
Cod ing , J . R . , B l o ckey , M .A . , Brown , J . M . , Ca t t , K . J . and Cumming , I . A .
( 1 9 70 ) . The role o f oes trogen � n the control o f the o e s trous cyc le
�n th e ewe . J . Reprod . Fe r t . �: 3 6 8-369 .
Cod ing , J . R . , Cat t , K . J . , Brown , J . M . , Ka l tenb ach , C . C . , Cumming , I . A .
and Mo l e , B . J . ( 1 969 ) . R . I . A . f o r ovine LH : Secr e t ion o f LH
hormone dur i ng oes trus and f o l l owing oes trogen admini s t r a t i on �n the
sheep . Endo crino l . 85 : 1 3 3 - 1 4 2 .
Gor ski , J . and Cannon , F . ( 19 7 6 ) . Current mode l s o f s teroid hormone
act ion ; a c r i t i que . Ann . Rev . Phy s io l . 3 8 : 4 2 5 -450 .
Gri sebach , H . and Bar z , W . ( 1 9 6 3 ) . The b i o genesis o f coumes trol �n
lucerne . Z i e t schr i f t Fur Na tur fors chung . 18B 46 6-4 70 .
Guggo l z , J . , L iv ings tone , A . L . and B i cko f f , E . M . ( 19 6 1 ) . D e t e c t ion o f
d iad zein , f ormonone tin , gen i s tein and b iochanin A in forage s .
J . Agr i c . Food Chem . �: 330-3 3 2 .
Hamada , Y . , Brons on , R . A . , Wr igh t , K . H . and Wal lach , E . E . ( 1 9 7 7 ) .
Ovu l a t ion in the perfused r abb i t ovary ; the inf luence o f
pro s tagland ins and pro s tag l and i n i nh ib itor s . B i o l . Reprod . 1 7 : 5 8-6 3 .
Hammond , J . ( 19 4 5 ) . Induced ovu l a t ion and heat in anoes trus sheep .
J . Endocr ino l . 4 : 1 6 9 - 1 8 0 .
Hanrahan , J . P . and Qu i rke , J . F . ( 1 9 7 5 ) . Repeatib i l i ty o f the dura t i on o f
oe s trus and breed d i f f erences � n the re l a t ionships b e tween dur a t ion
o f oe s trus and ovu l a t ion rate in s heep . J . Reprod . Fer t . �: 29-36 .
Hanson , C . H . , Loper , G . M . , Koh ler , G . O . , B i cko f f , E . M . and T ay l o r , K . N . ( 1 9 6 5 )
Var i a t ion i n coume s tr o l content o f a l f a l f a a s r e l ated t o l ocat ion ,
var i e t y , cu t t ing , year , s tage o f grow th and d i s ea s e . U . S . D . A . Tech •
Bu l l . 1 3 3 3 .

84 .
Harned , M . A . and Casida , L . E . ( 1 97 1 ) . Local response to proges t erone in
the ewe ovary . J . Anim . S c i . 3 3 : 1 1 5 8 .
Hawk , H . W . ( 1 97 4 ) . Oes trogenic e f f e c t s on uterine mo t il i ty in ewes .
J . Anim . Sci . 39 : 2 09- 2 1 0 .
Hawk , H . W . ( 1 9 7 5 ) . Enhancement by exogenous oes tradiol o f u t er ine
mo t ility in oes trous ewes . J . An im . S c i . 4 1 : 5 7 2 -5 7 7 .
Hawk , H . W . and Bol t , D . J . ( 1 9 70) . Luteolytic effect o f oes t radiol - 1 7 S
when adminis tered af t er mid-cyc l e i n the ewe . Biol . Repro d . 2
2 7 5- 2 7 8 .
Hawk , H . W . and Cooper , B . S . ( 1 97 5 ) . Improvement o f sperm t ranspo r t by
the admin is tra t ion o f oestradiol to oes t rous ewe s . J . Anim . S c i . 4 1
1 4 00- 1 4 0 6 .
Hawk , H . W . and Cooper , B . S . ( 1 9 7 6 ) . Ovum f e r t ilizat ion and embryo
survival in ewes treated with o es tradiol immed iately prior to ma t ing .
J . Anim . Sc i . 4 2 6 7 7-680 .
Hay , M . F . and Moor , R . M . ( 1 9 7 5a ) . Di s t r ibut ion of �5-3 S-hydroxy s t eroid
dehydrogena se a c t ivity in the Graaf ian follicle o f the sheep .
J . Repr o d . Fer t . � : 3 1 3- 32 2 .
Hay , M . F . and Moor , R . M . ( 1 9 7 5b ) . Funct ional and s t ruc tural rela t ionships
in the Graaf ian follicle popul a t ion of the sheep ovary .
Fer t . 4 5 : 5 8 3-59 3 .
J . Reprod .
Heape , W . ( 1 8 9 9 ) . Abortions , barr enness and fertility in sheep .
J . Roy. Agric . Soc . 60 : 2 1 7 -2 4 8 .
Hearnshaw , H . , Brown , J . M . , Cumming , I . A . , Goding , J . R. and Na irn , M .
( 1 9 7 2 ) . Endocrino logical and histopa thological aspects o f t he
in f e r t i l ity in the ewe caused by o e s trogenic c lover . J . Reprod .
Fer t . 2 8 : 1 60- 1 6 1 .

8 5 .
Hearnshaw , H . , Cumming , I . A . and God ing , J . R . ( 1 9 7 7 ) . E f f e c t o f inge s t ion
o f oestrogenic c love r on Lut e in i z ing Hormone release in the
ovar i ec tom ized ewe . Aus t . J . B io l . S c i . 3 0 : 2 1 7 - 2 2 3 .
Hohlweg , W . ( 1 9 34 ) . Veranderungen des hypohys en-vorderl appens und
des ovar iums nach Behand l ingen mit g ro s sen-dosen van fol l ikel ho rmon .
Kl in . Wchnshr 1 3 : 92-9 5 .
Ho l s t , P . J . and Braden , A . W . H . ( 1 97 2 ) .
Aus t . J . B io l . Sc i . 2 5 : 1 6 7 - 1 7 3 .
Ovum transport in the ewe .
Hoo l ey , R . D . , Baxter , R . W . , Chaml ey , W . A . , Cumming , I . A . , Jonas , H . A .
and Find lay , J . K . ( 1 9 74 ) . FSH and LH respons e to gonado troph in-
relea s ing ho rmone during t he ovine oes trous cyc l e and f o ll owing
progesterone administrat ion . Endocrinol . 95 : 9 3 7 -94 2 .
Howland , B . E . , Akbar , A . M . and S tormshak , F . ( 1 9 7 1 ) . S erum LH l evels
and luteal weight in ewe s fol lowing a s ingle inj e c t ion o f e s t rad io l .
B io l . Reprod . 2 : 2 5-2 9 .
Howland , B . E . and Palmer , W . M . ( 1 9 73 ) . Es trogen-induced c hanges in
serum l u t einiz ing hormone leve l s in ovariectomized ewes . Can . J . Anim .
S c i . 5 2 4 1 3-4 14 .
Hs euh , A . J . W . , Peck , E . J . , and Clark , J . H . ( 1 9 7 6 ) . Cont ro l o f u t er ine
e s t rogen r ec ep tor l evel s by pro gest erone . Endocrinol 9 8 : 4 38 -4 4 4 .
Hul e t , C . V . , P r i c e , D . A . and Foo te , W . C . ( 1 9 6 8 ) . E f f e c t s o f var ia t ion
in l ight , mont h o f year and nut rient intake on reproduc t ive phenomena
in ewes during the breed ing season . J . Anim . S c i . 2 7 : 684 -690
Hu t chinson , J . S . M . and Rober t son , H . A . ( 1 9 6 6 ) . The growth o f the fol l ic l e
and corpus l u teum in t h e ovary o f t h e sheep . Res . Vet . S c i . , 7 : 1 7 -2 4 .
Ingham , J . L . ( 1 97 2 ) . Phy toal exins and o ther natural produ c t s a s
fac tors i n p l an t d isease resis t anc e . The Bo tan ical Rev . 3 8 34 3-424 .

Jagu s ch , K . T . , Smi th , J . F . and Ke l l y , R . W . ( 19 7 7 ) . E f f e c t s o f f ee d i ng
luc erne dur ing ma ting on the fer t i l i ty o f ewes . Proc . N . Z .
Nu t r i t ion Soc . 1 9 7 7 : 2 6 -28 .
Jensen , E . V . and De Sombr e , E . R . ( 1 9 7 3 ) . Es t rogen-r eceptor in terac t ion
S c i ence 1 8 2 : 1 2 6-1 3 2 .
Jonas , H . A . , Salamons en , L . A . , Burger , H . A . , Chamley , W . A . , Cumming ,
I . A ., F ind l ay , J . K . and God ing , J . R . ( 1 9 7 3 ) . Re l ease of FSH af ter
adminis tra t ion of Gn-RH or oes trad i o l to the anoes trous ewe .
Endocrino l . 9 2 : 862-86 5 .
Jones , E . E . and Nalbandov , A . V . ( 1 9 7 2 ) . Ef fec t s of intraf o l l icular
inj e c t ion of gonado trophins on ovu l a t i on or l u t e i ni z a t ion o f ovarian
f o l l i c l e s . B io l . Reprod . I: 8 7 -9 3 .
86 .
Jones , H . E . H . and Pope , G . S . ( 1 9 6 1 ) . A me thod for the i s o l a t ion o f
miroes t r o l f rom Peuraria mir i f ica . J . Endocr inol 2 2 : 303- 3 1 2 .
Ka l le l a , K . ( 1 9 6 4 ) . The incidence of plant oes trogens in Finni sh pas ture
and fodder p l ant s wi th special reference to the i r po s s ib l e e f f e c t s
in cases of s ter i l i ty i n ruminants . Thes is . C o l lege o f Veter inary
Medi c i ne , He l s ink i , F inl and .
Ka l tenbach , C . C . and Davies , H . L . ( 1 9 70 ) . Fer t i l i za t ion, sperm t r anspor t
and ear ly embryonic loss in ewe s gra zed on cu l t ivar s o f sub terranean
c l over . ( T . Sub terranean) Au s t . J . Agr i c . Re s . �: 107 - 1 1 5 .
Ka t zene l l enbogen, B . S . , Fergu s on , E . R . and Lan , N . C . ( 19 7 7 ) . Fundamental
d i f f erences in the a c t ion o f e s t rogens and ant i - e s trogens on the
u terus : Compar i son b e tween compound s wi th s imi l ar dur a t ion o f a c t i on .
Endocr i no l . 100 : 1 25 2 - 1 259 .
Ke l ly , R . W . ( 1 9 7 2 ) . The oes t rogenic ac t iv i ty of c oumes tans �n ovar i e c t omi zed
ewe s . J . Reprod . Fer t . �: 1 5 9 .
Ke l l y , R . W . ( 1 9 7 6 ) . Mod i f ied extract ion technique f o r d e te rmina t ion of
amoun t s of coumes tans � n lucerne - pers comm . c i ted b y McCu t cheon ;
19 7 6 . Invermay Agr ic . Re s . Centre , Mo sgiel , N . Z .

8 7 .
Kelly , R . W . , Adams , N . R . and Lindsay , D . R . ( 1 9 7 6a) . · E f f ec t of
coumes tans on r ep roduc t ion in t he ewe . Aus t . J . Agric . Res .
27 : 2 5 3 -2 5 9 .
Kelly , R . W . , Al l i so n , A . J . and Shirley , D . K . ( 1 976b) . Int era c t ions
be tween phyto-oe stro gens and s t eroids in the cervical mucu s and
uterine we ight responses in ewes . Aus t . J . Agr ic . Res . 12 : 1 0 rl07
Kil leen , I . D . ( 1 9 6 7 ) . The effec ts of bodyweight and l evel o f nut r i t ion
before , during and a f ter j o ining on ewe f ecun d i ty .
Exptal Agr ic . Anim . Husb . 2 : 1 2 6 - 1 36 .
Aus t . J .
Kohl er , G . O . and B icko f f , E . M. ( 1 96 1 ) . Coumes trol and o ther b iologically
a c t ive compounds in forages . Feed s tu f f s 3 3 : 4 6 .
Ko l igian , K . B . and S to rmshak , F . ( 1 9 7 7 a ) . Proge s t erone inhib i t ion o f
es t rogen recep tors r epleni shmen t i n ovine endome trium. B iol .
Reprod . l2 : 4 1 2-4 1 6 .
Ko ligian , K . B . and S to rmshak , F . ( 1 9 7 7b ) . Nuclear and cytoplasmic
es trogen r ecep tor s in ovine endome tr ium during the estrous cyc l e .
Endocrino l . 1 0 1 : 5 24 - 5 3 3 .
Krarup , T . , P eder son , T . and Faber , M . ( 1 9 6 9 ) . Regula t ion o f oocyte
growth in the mou s e ovary . Na ture , 224 : 1 8 7 - 1 8 8 .
Kuehl , F . A . , Humes , J . L . , Tarno f f , J . , Cir illo , V . J . and Ham , E . A . ( 1 9 7 0 ) .
Pro s taglandin receptor s i t e s ; evidence for an essent ial role in
the ac t ion of LH . S c ienc e . 1 69 8 83-886 .
Lamming , G . E . ( 1 9 7 8 ) . Personal communica t ion . Phys io logy Dept . ,
School o f Agr ic . No t t ingham Un ive r s i ty .
Lan , N . C . and Ka t z enellenbo gen , B . S . ( 1 9 7 6 ) . Temporal r e l a t ionships
be tween hormone r eceptor b inding and b iological r e spons e s in the
u t erus : S tudies with shc r t and long a c t ing d e r ivat ives of e s t r iol .
Endocrino l . 98 220-22 7 .

88 .
Land , R . B . ( 1 9 7 0) . A r el a t ionsh ip b e tween the dur a tion o f o e s t ru s ,
ovul a t ion rate and l i t t er size o f sheep . J . Reprod . Fer t . 2 3
4 9 -53 .
Land , R . B . ( 1 9 7 3 ) . Ovula t ion rate in F inn-Do rse t sheep fol lowing
un i l a t eral ova r i ec tomy or chlorproma z ine t rea tmen t , at d i f feren t
s tages o f the o e s t rous cyc l e . J . Repro d . Fert . 3 3 9 9 - 1 05 .
Land , R . B . ( 1 9 7 6 ) . The s ens i t ivity o f the ovulat ion rate o f F inn ish
Landrace and S co t t i sh Blackface ewe s to exogenous o e s t rogens .
J . Reprod . Fer t . 4 8 2 1 7 -2 1 8 .
Land , R . B . , Pel l e t ier , J . , Thimon ier , J . and Maul eon , P . ( 1 9 7 3 ) . A
quan t ita t ive s tudy o f gene t ic di fferences in the inc idence o f
o e s t rus , ovula t ion and pla sma LH concentrat ion i n sheep .
J . Endo crinol 58 305 -3 1 7 .
Land , R . B . , Thomp son , R . and Baird , D . T . ( 1 9 7 2 ) . The dura t ion o f
o e s t rus in ovar iec tomized Finn ish Landrace and S c o t t i sh B lackface
ewes followin g proge s terone and oestrogen t r eatmen t . J . Reprod .
Fer t . 30 : 3 9 -44 .
Land , R . B . , Wheeler , A . G . and Carr , W . R . ( 1 9 7 6 ) . S eas onal var i a t ion in
t he oestrogen-induced LH di scharge o f ovariectomized Finn i sh
Landrace and S co t t i s h B lackface ewes . Ann . B io l . Anim . B io ch .
B iophys . 1 6 : 5 2 1 -5 2 8 .
Lewi s , P . E . and Warren , J . E . ( 1 9 7 5 ) . Indome tha c in inhibi t s luteo l y s is
in t he ewe . J . Anim . S c i . 4 1 : 365-366 .
Libertum , D . , Or ia s , R . and McCann , S . M . ( 1 9 7 4 ) . B iphasic e f f ec t o f
o e s t rogen o n t he sen s i t ivity o f the p i t u i tary t o LH-RH Endoc r ino l .
94 : 1 094- 1 1 00 .

8 9 .
Lightfoo t , R . J . , Croker , K . P . and Neil , H . G . ( 1 967 ) . Failure o f sperm
t ranspo r t in rela t ion to ewe infertil ity following prolonged grazing
on o e s t rogenic pa s tur e s . Aus t . J . Agric . Re s . � : 7 55 -7 6 5 .
Lightfoo t , R . J . , Mar shall , T . and Croker , K . P . ( 1 9 7 6 ) . E f f e c t s o f
rate and dura t ion o f lupin grain supplementat ion on ovu l a t ion and
f er t il it y of Merino ewes . Proc . Aus t . Soc . Anim . Prod . 1 1 5 . p .
Lightfoo t , R . J. , Smi t h , J . F . , Cumming , I . A . , Marsha l l , T . , Wra t h , R . H .
and Hearnshaw , H . ( 1 9 7 4 ) . Infe r t i l i ty in ewes c aused by p rolonged
grazing of oes trogenic pastures : O e s trus , f er t il iza t ion and
c ervical mucus . Aus t . J . B iol . S c i . 2 7 : 409-4 1 4 .
Lightfoo t , R . J . and Wra th, R . H . ( 1 9 7 4 ) . The mechanism o f t empo rary
infer t il i t y in ewes graz ing oes trogenic subterranean clover prior
to and du ring j o ining . Proc . Aus t . Soc . Anim . Prod . 1 0 1 30- 1 34 .
Lindner , H . R . ( 1 96 7 ) . S tudy of the f a t e o f phyto-o e s t ro gens in sheep
by det ermina t ion o f isoflavones and coumestrol in the plasma and
adipo s e t i s su e . Aus t . J . Agric . Re s . 1 8 : 305- 3 3 3 .
Lind say , D . R . , Cognie , Y . , Pellet ier , J and S ignoret , J . P . ( 1 9 7 5 ) .
Infl uence o f the presence of rams on the timing o f ovulat ion and
di scharge of LH in ewe s . Phys io l . and B ehav . 1 5 : 4 2 3-4 2 6 .
Lind say , D . R. and Francis , C . M . ( 1 96 8 ) . Cervical mucus measuremen t in
ovariec tomiz ed ewes as a b ioassay of syn thetic and phyto-oes tro gens .
Aus t . J . Agr ic . Res . 1 9 : 1 06 9 - 1 07 6 .
Lipner , H . and Greep , R . O . ( 1 97 1 ) . Inhibi t ion o f s teroid s yn th e s is a t
various s i t es i n the b io synt he t i c pa thway in rela t ion to induced
ovulat ion . Endocrino l . 88 : 6 0 2 -6 1 7 .
L iv ing s tone , A . L . , B ic ko f f , E . M . , Guggo l z , J . and Thomps on , C . R . ( 19 6 1 ) .
Quan t i ta t ive determina t ion o f coume s t ro l in fresh and d r ied lucerne .
J . Agr ic . Food Chem . 9 1 3 5-1 3 7 .

9 0 .
Loewe , S . , Lange , F . and Spoh r , E . ( 19 2 7 ) - c i ted b y K . Kal l e l a , The s i s ,
He l s ink i , 1964 . B iochem . Z . 1 80 : 1 .
Loper G . M . ( 1 9 6 8 ) . E f f e c t o f aph id inf e s tat ion on the coumes trol
content of a l f a l f a var i e t i es d i f f er ing in aph id r e s i s tance .
Crop Sc i . �: 104-106 .
Loper , G . M . and Han son , C . H . ( 1 96 4 ) . Inf luence of c ontro l l ed
environmental fac tor s and two fo l iar pa thogens on coume s t r o l
in a l f a l f a . Crop Sc i . �: 4 80-4 8 2 .
Loper , G . M . , Hanson , C . H . and Graham , J . H . ( 1 96 7 ) . Coume s trol content
o f a l f a lfa as e f f e c ted b y s e l ec ti on for res i s tance to f o l iar
d i seases . Crop S c i . I: 1 8 9 - 1 9 2 .
Luine , V . N . and McEwen , B . S . ( 1 9 7 7 ) . E f f e c t s o f an es trogen an tagon i s t
o n enzyme activi t i es and (3
H ) es trad i o l nuclear b ind i ng 1 n the
u teru s , pitui tary and b rain . Endocr ino l . 100 : 903-9 1 0 .
Lunenf i e l d , B . , Kraiem , Z . and Eshko l , A . ( 19 7 5 ) . The func t ion o f the
growing f o l l i c l e . J . Reprod . Fer t . 45 : 567-5 7 4 .
Lyman , R . L . , Bickof f , E . M . , Boo th , A . N . and Livings tone , A . L . ( 1 9 5 9 ) .
Detec tion of coume s t r o l i n Leguminous plant s . Arch . Biochem .
Biophys . 80 : 6 1-6 7 .
McCut cheo n , S . N . ( 1 9 7 6 ) . A s tudy o f the reproduc t ive performance o f
Bo rder Le ices ter X Romney ewe s ma ted o n oe s trogenic lucerne .
B . Agr i c . Sci ( Hons . ) D i s ser tation, Ma s sey Univers i ty .
McDona ld , M . F . and Chang , T . S . ( 1 96 6 ) . Var i a t ion i n ovar 1an a c t iv i ty o f
Romney Mar sh ewe s . Proc . N . Z . Soc . Anim . Prod . 26 : 98-106 .
Mc lnne s , P . and Smi th , M . D . ( 19 6 6 ) . The e f f e c t s of nu tri tion b e fore
ma t ing on the performance of Mer ino ewe s . Aus t . J . Expt a l Agr i c .
and Anim . Husb . 6 : 45 5-45 9 .
McLean , J . W . ( 1 9 6 7 ) . Luc e r ne and phy to-oes trogens . From ' The Lucerne
Crop ' Ed . R . M. Langer . Pub l i shed A . H . & A . W . Reed . Wg tn . N . Z .

9 1 .
Mal l arnpa t i , R . S . and Casida , L . E . ( 19 70 ) . Ovar ian compensatory
hyper trophy f o l lowing uni l ateral ovar i e c t omy dur ing the breed ing
season in the ewe . B io l . Repr od . l= 4 3-46 .
Mar sh , J . M . ( 1 9 7 1 ) . The e f f e c t s o f pro s tagland ins on the adenyl
cyc lase o f the bovine corpus l u teum . Ann . New York Acad . S c i .
1 80 : 4 1 6-425 .
Ma t tner , P . R . ( 1 9 6 6 ) . Forma tion and re ten t ion of the sperma ta zoan
r e servo � r �n the cervix of the ruminan t . Nature 2 1 2 : 1 4 7 9 - 1 4 80 .
Ma t t on , P . , Bherer , J . and Duf our , J . J . ( 1 9 7 7 ) . Morpho logy and
re spons ivene s s of the two large s t ovar ian fo l l i c l e s in anoes trous
ewes . Can . J . Anim . Sci . 5 7 : 459-464 .
Mi che l i , R . A . , Boo th , A . N . , Livings tone , A . L . and B i ckof f , E . M . ( 1 9 6 2 ) .
Coume s tro l , p l ant pheno l i c s and s ynth e t i c oe s t ro gens ; a corr e l a t ion
o f s truc ture and a c t iv i ty . J . Med . Pharm . Chem . 5 : 3 2 1- 3 2 5 .
Mi l l er , B . G . , Murphy , L . and S tone , G . M . ( 1 9 7 7b ) . Hormone receptor
l eve l s and hormone , RNA and pro t e i n metab o l i sm in the geni t a l tract
dur ing the oes trous cyc l e in the ewe . J . Endocr ino l . 7 3 : 9 1- 9 8 .
Mi l ler , W . L . , Knight , M . M . , Gr imek , H . J . and Gorsk i , J . ( 1 9 7 7 a ) .
Es tro gen regu l a t i on o f Fol l i c l e S t imu l a t ing Hormone in c e l l cul tures
of sheep p i tu i tar ies . Endocrino l . lOO : 1306- 1 3 1 6 .
Mi l l i ngton , A . J . , Franc i s , C M . and McKeown , N . R . ( 1 964a) . We ther
b ioas say o f annual pas ture l egume s . I . Oes trogeni c a c t iv i ty � n
Med i cago tr ibu l o ide s desr . var . Cyprus r e lative t o f our s tr ains o f
(Tr i f o l ium sub t e rranean L . ) Au s t . J . Agr ic . Re s . 1 5 : 5 20-5 26 .
Mi l l ing ton , A . J . , Franc i s , C . M . and McKeown , N . R . ( 19 6 4b ) . Wether
b ioassay o f annual pa s ture l egume s . I I . The o e s t r ogenic a c t iv i ty
o f nine s trains of (Tr i f o l ium sub terranean L . ) Aus t . J . Agr ic . Re s .
1 5 : 5 2 7 -5 3 6 .
Moo r , R . M . ( 19 7 3 ) . Oes trogen produc t i on by i ndividual f o l l ic l e s
expl an ted f r om ovar ies o f sheep . J . Reprod . Fer t . 3 2 : 5 45-54 8 .

9 2 .
Moor , R . M . ( 19 7 4 ) . The ovar ian f o l l i c l e o f the sheep ; i nhib i t ion o f
o e s trogen secr e t ion o f l u te ini z ing hormone . J . Endocrino l . 6 1 :
455-463 .
Moor, R . M . ( 1 9 7 7 ) . S i tes o f s teroid produc t i on 1n ov i ne Graaf i an
f o l l i c l e s in cul ture . J . Endocr ino l . 7 3 : 143-15 0 .
Moor , R . M . , Hay , M. F . and Caldwe l l , B . V . ( 19 7 1 ) . The sheep f o l l ic l e ;
r e l a t ion be tween the s i tes o f s tero id d ehydrogenase ac t iv i ty ,
gonadotrophic s t imu l a tion and s teroid produc t i on . J . Reprod .
Fer t . 2 7 : 484-485 .
Moore , N . W . , Barre t , S . , Brown , J . B . , Schind l er , I . , Smi th , M . A . and
Smyth, B . ( 196 9 ) . Oes trogen and proges terone content of ovar ian
vein b l ood o f the ewe during the oes trous cycl e . J . Endocr ino l .
44 : 55-6 2 .
Moor e , N . W. and Rob inson , T . J . ( 1 95 7 ) . The behaviour a l and vag ina l
response of the spayed ewe to oe s t rogen inj ec ted a t var ious t ime s
r e l a t ive to the inj e c tion o f proges terone . J . E .ocrino l . 1 5 :
360-36 5 .
Mor l e y , F . H . W . , Axe l s en , A . and Benne t t , D . ( 1 963) . E f f ec t s o f o e s tro gen
on fer t i l i ty in ewe s . Na ture . 199 : 403-404 .
Mor l e y , F . H . W . , Axe l s en , A . and Benne t t , D . ( 1964) . E f f e c t s o f graz i ng
red c l over (Tr if o l ium pr a t ense) dur ing the j o i ning sea son on ewe
f e r t i l i ty . Proc . Au s t . Soc . Anim . Prod . 5 : 5 8-6 1 .
Mor ley , F . H . W . , Axe l s en , A . and Benne t t , D . ( 19 6 6 ) . Recovery o f
f er t i l i ty af ter graz ing o n oes trogenic r ed c l over . Aus t . Ve t . J .
4 2 : 204-20 6 .
Mor l e y , F . H . W . , Benne t t , D . , B r aden , A . W . H . , Turnbul l , K . E . and
Axe l s en , A . ( 19 68 ) . Comparison o f mice , gu inea-p igs and sheep
as te s t an ima l s for b ioas s ay o f o e s t rogenic pas ture l e gume s .
Proc . N . Z . Soc . Anim . Prod . 28 : 1 1- 2 1 .

Morr i s , I . D . ( 19 76 ) . Change s i n brain , p i tu i tary and u te r ine
cytop l asmic oes trogen receptors induc ed b y oe s tradiol-1 7 8
in the ovariec tomi zed ra t s . J . Endo c r ino l . 7 1 : 343-349 .
Moule , G . R . , Braden , A . W . H . and Lamond , D . R . ( 1 96 3 ) . The s ignif i c ance
of o e s trogens in pas ture p l ants 1 n r e lation to anima l produc t ion .
Anim . Brdg . Ab s t s . 3 1 : 1 3 9 - 1 5 7 .
Murphy , L . , S tone , G . M . , Mil ler , B . G . , and Moo re , N . W . ( 19 7 7 ) .
Oes trad i o l and proge s t erone : Solub l e recep tor l ev e l s and
me tabo l i sm in the u terus of ovar iec tomi zed ewe s . Aus t . J . B i o l .
Sc i . 30 : 2 2 5 - 2 2 8 .
Newsome , F . E . and Ki t t s , W . D . ( 19 7 7 ) . E f f e c t s of a l f a l fa consump t ion
on oes trogen l eve l s 1n ewe s . Can . J . Anim. S c i . 5 7 : 5 3 1 - 5 5 5 .
9 3 .
N i l s son , A . ( 1 9 6 1 ) . On the In v i tro me tabo l i sm of the p l ant o e s trogen
bi ochanin A 1n rumen f luid . Ark . Kem i 1 7 : 305 -3 10 .
Ni l s son, A . ( 19 6 3 ) . 0-deme thy l a t ion of b i ochanin A and some o ther
i s o f l avones and me thylated o e s trogens by microsomal l iver enzyme s .
Ark . Kemi . 2 1 : 9 7 - 1 2 1 .
N i l s son, A . , H i l l , J . A . and Dav i e s , H . L . ( 1 9 6 7 ) . An In v i tr o s tudy
of formonone tin and b io chanin A me tab o l i sm in r umen f lu i d f r om
sheep . B iochim . B iophys . Ac ta . 1 4 8 : 9 2-98 .
Ni swender , G . D . , Reicher t , L . E . , Midg l e y , A . R . and Nalbandov , A . V .
( 1969 ) . Rad i o immuno a s s ay for bovine and ovine l u t e i ni s ing
hormone . Endocrino l . 84 : 1 1 66-1 1 7 3 .
Ob s t , J . M . and Seamark , R . F . ( 1 9 70 ) , P l a sma proges te rone concentrati ons
dur ing the r eproduc t iv e cyc l e o f ewe s grazing Yar l oop c l ov e r .
J , Reprod , Fer t . 2 1 : 545-547 .

O ' Ma l l ey , B . W . and Schrader , W . T . ( 1 9 7 6 ) . The recep tors o f s teroid
hormone s . S c i . Amer . 234 ( 2 ) : 3 2-43 .
O ' Shea , J . D . ( 1 9 7 1 ) . Smo o th-musc le-l ike c e l l s 1n the thec a ex terna
of ovar ian f o l l i c l es in the sheep . J . Reprod . Fer t . 24 : 2 8 3-285 .
O ' Shea , J . D . and Phi l l ips , R . E . ( 1 9 7 4 ) . Contrac t i l i ty o f ovar 1 an
fo l l i c l e s from sheep In v i tro . J . Reprod . Fer t . �: 4 5 7 .
Owman , C . , Sj ober g , N . O . , Svens son , K . G . and Wa l l es , B . ( 19 7 5 ) .
Au tonomic nerv e s med i a t i ng contrac t i l i ty in the -human Gr aaf i an
fo l l i c l e . J . Reprod . Fer t . 45 : 5 3 3-556 .
Pant , H . C . ( 1 9 7 7 ) . E f f e c t of oes trad io l infu s ion on p l a sma g onado
trophins and ovar ian ac t iv i ty in proge s terone-pr imed and unpr imed
anoes trous ewe s . J . Endocrino l . 7 5 : 2 2 7 -233 .
Pe l l e t i er , J . ( 1 9 7 6 ) . Inf luence of L H-RH on LH and FSH r e l ea se
in domes t i c annima l s . Ann . B io l . An im . B iochm . B i ophys . 16 :
2 1 3-234 .
Pe l l e t i er , J . and S i gnore t , J . P . ( 1 9 7 0 ) . pp . 215-2 2 3 . from
' L ' inhib i t ion de l ' ovu l a t i on ' Ed . Ma s son e t Cie .
Peter s , H . , Byskov , A . G . , Lintern-Moore , S . and Faber , M . ( 1 9 7 3 ) .
Fo l l i c le grow th ini t i a t ion in the immature mous e ovar y ;
extra-ovar i an o r intra-ovarian ovar i an contro l . J . Reprod .
F er t . 3 5 : 6 1 9-620 .
Peter s , H . , Byskov , A . G . , Hime l s te i n-Braw , R . and Faber , M . ( 19 7 5 ) .
Fo l l i cu l ar grow th ; the b a s i c event i n the mouse and human ovary .
J . Reprod . Fer t . 45 : 559-5 6 6 .
9 4 .

9 5 .
Phelps , C . P . , Coy , D . H . , S chal ly , A . V . and Sawy er , C . H : ( 1 9 7 7 ) .
Blockade o f LH release and ovul a t ion in the rabb i t with inhib i tory
ana logues o f LH-RH . Endoc r ino l . 100 : 1 5 2 6 - 1 53 2 .
Phi , L . T . , Moon , Y . S . and Arms t rong , D . T . ( 1 9 7 7 ) . E f f e c t s o f ·sy s t emic
and intrafoll i cular inj ec t ions o f LH , prostagland ins and indome tha c in
on the luteiniza t ion o f rabb i t Graaf ian fol l i c les .
1 3 54 3-5 52 .
Pro s tagland ins
Pieter s e , P . J . S . and And rews , F . N . ( 1 9 56 ) . The o e s t rogenic a c t ivity
o f al falfa and o ther f ee d s t uf f s . J . Anim . S c i . 1 5 2 5-36 .
P iper , E . L . and Foo t e , W . C . ( 1 9 6 8 ) . Ovul a t ion and corpus lut eum
maintenance in ewes t rea ted with 1 7 8-oes t radiol . J . Reprod . Fert .
1 6 : 2 5 3- 25 9 .
Pop e , G . S . ( 1 9 54 ) . The impor tance o f pa s t ure plant o e s t ro gens in the
r eproduc t ion and lac t a t ion o f graz ing an imals . Da iry S c i . Ab s t s .
1 6 : 333-3 56 .
Pop e , G . S . and Wright , H . G . ( 1 9 54 ) . Oes t rogenic isof lavones in red
c lover and sub t erranean c lover . Chem . Ind . 1 9 54 : 1 0 1 9 - 1 0 20 .
Radford , H . M . and Wallac e , A . L . C . ( 1 9 7 4 ) . C . N . S . blockade o f the
of the oestradiol - s t imul a t ed rel ease of LH in the ewe . J . Endo c r inol .
6 0 : 247-25 2 .
Radford , H . M. , Wa llac e , A . L . and Whea tley , I . S . ( 1 9 7 0 ) . LH relea s e ,
ovul a t ion and oes trus fol lowing the trea tment o f ano e s trous ewes
with ovar ian s t ero id s . J . Reprod . Fert . 2 1 : 3 7 1 - 3 7 2 .
Rad for d , H . M . , Whea tley , I . S . and Walla c e , A . L . C . ( 1 9 6 9 ) . The e f f e c t s
o f o e s tradiol b en zoa t e and progesterone on secret ion o f l u t e in i z ing
hormone in the ova r i e c tomized ewe . J . Endoc r inol . 4 4 : 1 3 5 - 1 3 6 .

9 6 .
Ramirez , V . D . and Sawyer , C . H . ( 1 9 7 4 ) . A sex d i f fer ence �n the r a t
p i tu i t ary FSH response to uni l a teral gonadec tomy as r eveal ed i n
p l a sma rad i o immunoas says . Endocr ino l . 9 4 : 4 7 5-4 8 2 .
Reardon , T . F . and Rob inson , T . J . ( 1 9 6 1 ) . Seasona l var i a t ion � n the
reac t iv i t y to oes trogen in the ovariec tomi zed ewe .
Au s t . J . Agr ic . Re s . 1 2 : 3 20-326 .
Reeve s , J . J . , Ar imura , A . and Scha l l y , A . V . ( 19 7 l a) . P i tu i tary
r e sponsivene s s to pur i f ied luteini z ing hormone-r e l e a s i ng hormone
(LH-RH ) at various s t ages o f the oes trous cyc l e in sheep . J . Anim .
Sci . 3 2 : 1 2 3 -126 .
Reeves , J . J . , Arimura , A . and Scha l ly , A . V . ( 1 9 7 lb ) . Changes in
p i tu i tary r e spons ivene s s to LH-RH in anoes trous ewe s pre-trea ted
wi th oes tradiol benzoa t e . B i o l . Reprod . �: 88-9 2 .
Reeves , J . J . , O ' Donne l l , D . A . and Denor s c i a , F . ( 1 9 7 2 ) . E f f e c t of
ov ar iectomy on serum lute ini z ing hormone (LH) concen tration s in the
ano e s t rous ewe . J . Anim . Sc i . 3 5 : 7 3 - 7 8 .
Reeve s , J . J . , Beck , T . W . and Ne t t , T . M . ( 1 9 7 4 ) . Serum F SH �n the
anoe s trous ewe trea ted wi th 1 7 6-oes trad i o l . J . Anim . S c i . 38 :
3 7 4 -3 7 7 .
Re s ta l l , B . J . ( 1 9 6 6 ) . The f a l lop ian tub e o f the sheep . I I . The
inf luence of proge s t erone and oes trogen on the secre tary a c t iv i t ie s
o f t h e f a l l op ian tub e . Aus t . J . B io l . Sci . 1 9 : 1 8 7 -19 7 .
Rexroad , C . E . and Barb , C . R . ( 1 9 7 7 ) . Cerv ical mucus in o e s t r u s ewe s
af ter trea tment wi th oes trog en , proge s tagens and u t er ine dev i c e s .
J . Anim . S c i . 44 : 102-105 .
Rexro ad , C . E . , and C as ida , L :E . ( 19 7 7 ) . E f f e c t o f i nj e c t ion o f
proges terone into one ovary of PMSG- trea ted ano e s t r ou s ewe on
f o l l i c l e growth and ovar i an oes trad i o l - 1 7 6 . J. Anim. S c i . 44 : 84-8 8 .

9 7 .
Richa rd s , J . S . and Midgley , A . R . ( 1 97 6 ) . Pro t ein hormone a c t ion : A
key to unders tand ing ovarian foll icular and luteal c el l development .
B iol . Reprod . li : 8 2 -94 .
Robj son , G . A . , But cher , R . W . and Sutherland , E . W . ( 1 9 7 1 ) .
' Cycl i c AMP ' Academic Press , New York .
f rom
Rodbard , D . ( 1 9 68 ) . Mec han ic s of ovu l a t ion . J . C l in . Endo c r inol .
Metab . 2 8 : 849 -86 1 .
Rond el l , P . ( 1 9 74 ) . Rol e of s t eroid synthes i s in t he pro c e s s o f
ovulat ion . B iol . Reprod . lQ : 1 9 9 - 2 1 5 .
Ro s s i t er , R . C . ( 1 9 6 9 ) . Phys io logical and ecological s tud ies on the
oest rogenic isoflavones in sub t er ranean c lover . V I I . E f f e c t
1 04 3 - 1 05 1 . of nitrogen supp l y . Aus t . J . Agric . Res . 20
Ro s s i ter , R. C . ( 1 9 7 0 ) . Fac to r s effec t ing the oestrogen content o f
sub terranean c l over pa stur es . Au s t . Ve t . J . 4 6 : 1 4 1 - 144 .
Ro s s i t er , R . C . and Ba rrow , N . J . ( 1 9 7 2 ) . Phy siological and ecological
studies on the o e s trogen ic isoflavones in sub t erranean c lover .
IX. Ef f e c t s of sulphur supply . Aus t . J . Agric . Res . � : 4 1 1 -4 1 8 .
Ros s i ter , R . C . and Beck, A . B . ( 1 9 66a) . Phys io logical and ecological
stud ies on the o e s t rogenic isoflavone content o f sub terranean
clover . I . E f fec t s of t empera tur e . Aus t . J . Agric . Res . 1 7
29-37 .
Ro s s iter , R . C . and Beck, A . B . ( 1 9 66b) . Phy s iological and eco logical
stud ies on the o es trogenic isoflavone s in sub t erran ean c lover .
I I . E f f e c t s o f phospha t e supply . Aus t . J . Agr ic . Res . !l 4 4 7 -4 5 6 .
Ruh , T . S . and Ruh , M . F . ( 1 97 4 ) . The e f f e c t of ant i-o es trogens on the
nuc l ear b ind ing of t he oes trogen recep t o r . S tero ids 24 2 09-224 .

Salamonsen , L . A . , Jonas , H . A . , Burger , H . G . , Buckrnas ter , J . M . ,
Chamley , W . A . , Gumming , I . A . , F i nd l a y , J . K . and Cod ing , J . R .
( 1 9 7 3 ) . A heterol ogou s rad i o immuno a s s ay for Fol l ic l e S t imu l a t ing
Hormone ; app l i cation to measurement o f FSH in the ov ine o e s t r ous
cyc l e and in s evera l o ther spec1es inc l ud ing man . Endocrino l .
9 3 : 6 10-6 1 8 .
Sanger , V . L . and Be l l , M . S . ( 1 9 59 ) . Es trogenic activ i t y 1 n green
f orage crops and i t s e f f e c t s on breed ing ewes . J . Arner . V e t .
Assn . 1 3 4 : 23 7-239 .
Scal es , G . H . and Mo ss , R . A . ( 1 9 7 6 ) . Ma ting ewe s on luc erne .
N . Z . J . Agr i c . 1 3 2 : 2 1 -2 2 .
9 8 .
Scal es , G . H . , Mo s s , R . A . and Ke l l y , R . W . ( 1 9 7 7 ) . Reprodu c t ive performance
of ewe s ma ted on luce rne . Proc . N . Z . Soc . Anim . Prod . 3 7 :
1 5 2 - 1 5 7 .
Scaramu z zi , R . J . ( 1 9 7 5 ) . Inhib i t ion of oe s tru s b ehav i our in ewe s by
pas s ive immunizat ion agains t oestrad iol- 1 7 8 . J . Reprod . Fer t .
42 : 1 4 5 - 1 48 .
Scararnuzzi , R . J . , Lind s ay , D . R . and She l ton , J . N . ( 1 9 7 l a ) . The e f f e c t
of oes trad i o l ben zoate o n the dura t i on o f oe s trus behaviour i n the
ovar i e c tom i z ed ewe . J . Endocr ino l 5 0 : 3 45-346 .
Scararnuzzi , R . J . , Lind s ay , D . R . and Shel t on , J . N . ( 19 7 2 ) . E f f e c t o f
repea t ed oes trogen adminis tra tion on oes trus b ehav iour i n ovari-
ec tom i zed ewes . J. Endocrino l . 5 2 : 269-2 78 .
Scararnuzzi , R . J . , Ti l l so n , S . A . , Thorneycrof t , I . H . and Caldwe l l , B . V .
( 19 7 lb ) . Ac tion of exogenous proges terone and e s trogen on b ehavioural
e s tru s and luteiniz ing hormone l eve l s in the ovar iec tomized ewe .
Endoc r ino l . 8 8 : 1 1 8 4- 1 1 89 .

9 9 .
Schinckel , P . G . ( 1 94 8 ) . Infer t i l i ty in ewe s graz ing sub terranean
clover pas tures . Obs erva t ions on breeding behaviour f o l l owing
tran s f e r to ' sound ' coun try . Aust . Vet . J . 2 4 : 2 89-2 94 .
Schwar t z , N . B . ( 1 974) . The rol e o f FSH and LH and o f their ant ibod i e s
o n f o l l i c l e growth and o n ovul a t ion . Biol . Reprod . lQ : 2 36-2 7 2 .
S eamark , R. F . ( 1 9 78) . Intraovarian factors in ovar ian f unc t ion .
Proc . 6 th Asia and Oceania Cong. Endo c r ino l : pp . 1 65 - 1 7 1 .
S eamark , R. F . , Moor , R . M . and Mc in tosh, J . E . A . ( 1 9 7 4 ) . S t ero id hormone
produc t ion by sheep ovar ian fo l l ic l es cul tured In vitro .
J . Re£!o d . Fer t . 4 1 1 4 3- 1 58 .
Shemesh , M . , Lindner , H . R . and Ayalon , N . ( 1 9 72 ) . Af f inity o f rabbit
uterine oes tradiol receptor for phyto-oes trogens and its use in a
compe t i t ive pro tein b inding R. I . A . for plasma coumes t ro l .
J . Reprod . Fer t . 2 9 : 1 -9 .
Shut t , D . A . , Axelsen , A . and Lindner , H . R . ( 1 96 7 ) . Free and conj uga t ed
iso flavones in the pla sma o f sheep fol lowing inges tion o f o e s t rogenic
clover . Aus t . J . Agric . Res . 1 8 647-655 .
Shut t , D . A . and Braden , A . W . H . ( 1 9 68 ) . The sign i f i cance o f equal in
relation to the oestrogenic responses in sheep inge s t ing c l over
with a high formononeton c ontent . Aus t . J . Agr i c . Res . 1 9 : 54 5-555 .
Shu t t , D . A . , Braden , A . W . H . and Lindner , H . R . ( 1 9 6 9 ) . Plasma coumes t ro l
l evels i n sheep following admin i s t r a t ion of s ynthetic coumes trol
o r inge s t ion of medic hay (Med icago l i t toral i s ) .
20 : 6 5 - 69 .
Aus t . J . Agr ic . Res .
Shu t t , D . A . , Wes ton , R . H . and Hogan , J . P . ( 1 9 70) . Quan t i t a t ive aspec t s
o f phyto-oes trogen metabol ism in sheep f ed on sub terranean c lover
( Trifol ium sub terranean ) cul t ivar Clare or red clover ( Tr if o l ium
·pr aten s e ) . Aus t , J . Agr i c . Res . 2 1 : 7 1 3-7 22 .

1 00 .
Skar zynski , B . ( 1 9 3 3 ) . An o e s t ro genic sub s tance from p lant ma ter ial .
Nature 1 3 1 : 7 6 6 .
Smea ton , T . C . and Rober t son , H . A . ( 1 97 1 ) . S tudies on the growth and
a t resia of Graaf ian follicles in the ovary of the sheep .
J . Reprod . Fer t . 2 5 : 2 4 3-2 5 2 .
Smith, J . F . ( 1 97 1 ) . S tudies on ovine inf e r t ility in agricul tural
reg ions of We st ern Aus tralia : Cervical mucus product ion by f e r t i l e
and inf ert il e ewe s . Aus t . J . Agr ic . Res . � 5 1 3- 5 1 9 .
Smi th , J . F . ( l 9 7 5 ) . S tud ies on ovine inf e r t ility in agricul t ural
reg ions of We s t ern Aus tral ia : Plasma proges t erone l evel s of f e r t i l e
and infer t il e ewes . Aus t . J . Biol . S c i . 2 8 : 85-88 .
Smith , J . F . , Jagusch , K . T . and B runswick , L . ( 1 9 7 8 ) . unpub l i shed data .
Ruakura Agric . Re s . Centre . Hamil ton New Zeal and .
Smith , J . F . and Rob inson , T . J . ( 1 9 6 9 ) . Lu t eal func t ion in Merino
ewe s and the e f f e c t of exo genous proge s t agens . J . Endo crino l . 44
7 9 -8 9 .
Sned ecor , G . W . and Co chrane , W . G . ( 1 9 6 7 ) . f rom ' S ta t i s t ical me thods '
Iowa S t a t e Unive r s i ty Pres s .
Squire s , V . R . ( 1 9 6 6 ) . S easonal inc idence of o e s t ro genic a c t iv i t y in
i r rigated pa s ture as a s s es sed b y we ther t ea t l ength incr ea s e .
Proc . Aus t . Soc . Anim . Prod . 6 : 1 1 7 - 1 2 2 .
S t einberger , E . and Chowdhu ry , M . ( 1 9 7 7 ) . The e f f ec ts o f t e s t o s t erone
propriona t e and es tradiol b en zoa t e on the In v i tro syn thesis
of FSH . B io l . Reprod . � 4 03-4 08 .
S tob , M . , Davis , R . L . and Andrews , F . N . ( 1 9 57 ) . S t rain d i f f er ences
in the oest rogenic i ty of alfal fa . J . Anim . S c i . 1 6 : 8 5 0-8 5 3 .
S to rmshak , F . , Kelly , H . E . and Hawk , H . W . ( 1 9 6 9 ) . Suppr e s s ion o f ovine
l u t eal forma t ion b y o e s t radio l - 1 7 8 . J . Anim . S c i . 2 9 : 4 7 6 -4 7 8 .

1 0 1 .
S tuthman , D . D . , B ickof f , E . M . , Davis , R . L . and S t ob , M . ( 1 9 6 7 ) .
Coumes trol d i f f er ences in Medicago sat iva L . free o f fol iar d i s ease
s ymp toms . Crop Sc i . � : 3 3 3 - 3 34 .
Sundaram , S . K . and S t o b , M . ( 1 96 7 ) . Ef fec t of uni l a teral ova r i ec tomy
on r eproduc t ion and induced ovulat ion in ewes .
3 7 4 -3 7 6 .
J . Anim . Sc i . 2 6
Sutherland , R . L . and Mes ter , J . ( 1 9 7 6 ) . E f f ec t of o e s t radiol and
progest erone on the concen t ra t ion o f oestrogen receptor s it e s in
the c hic ken oviduc t . J . Endo crino l . ll : 55p .
Thimonier , J . , Terqu i , M . and P el l e t ier , J . ( 1 9 7 4 ) . Plasma e s trad iol
and p roges terone l eve l s and pituitary re spons iveness to LH-RH
in t he ewe . J . S t eroid Biochem 5 : 388 .
To j o , S . , Fukun i shi , H . , Tsuchihashi , T . , Shimura , T . , Mikami , K . and
Kaku , H . ( 1 9 7 5 ) . E l e c t r ical ac t iv i ty recorded f rom ova r ian t issue
in per fused human ut ero- tuba-ovar ian un it . J . Repro d . Fer t . 44
5 8 7 - 5 8 9 .
Trounson , A . O . , Chamley , W . A . , Kennedy , J . P . and Tas s el l , R . ( 1 9 74 ) .
P r imordial follicle numbers in ovaries and levels o f LH and FSH
in p i tui taries and p lasma of lamb s selec t ed for and aga inst
mul t iple bir ths . Aus t . J . Biol . Sc i . 2 7 : 2 9 3-2 9 9 .
Tsa f r ir i , A . , Lindner , H . A . , Zor , U . and Lamprech t , S . A . ( 1 9 7 2 ) .
Phys iol ogical role o f pros taglandins in the induc t ion o f ovulat ion .
P r o s t agland ins l : 1 - 1 0 .
Turnbul l , K . E . , Braden , A . W . H . and George , J . M. ( 1 96 6 ) . Fer t il i za t ion
and early emb ryonic losses in ewes tha t had grazed o e s t rogen ic
pas tur e for 6 year s . Aus t . J . Agric . Res . 1 7 : 907-9 1 7 .

Turnbu l l , K . E . , Braden , A . H . H . and Ma t tner , P . E . ( 1 9 7 7 a ) , The
pat tern of f o l l i cular growth and a tr e sia in the ov ine ovary .
Aus t . J . Biol . Sc i . 30 : 229-24 1 .
Turnbu l l , K . E . , Land , R . B . and Scaramu z z i , R . J . ( 1 9 7 7 b ) . Pat terns o f
growth o f Graaf ian f o l l i c les i n sheep and the i r r e l a t ionship to
ovu l a t ion r a t e . Proc . Au s t . Soc . Reprod . B io l . 9 : 4 5 .
Turnbu l l , K . E . , Shut t , D . A . and Brad e n , A . W . H . ( 19 6 7 ) . Increase
in chlor ine content of cerv ical mucu s a s a simp le test for
impend i ng ovulat ion and for oes trogenic s t imu la t ion �n ewe s .
Au s t . J . Exptal. Anim . Husb . 7 : 3 1 4 -3 1 7 .
Underwood , E . J . and Sh ie r , F . L . ( 1 9 5 1 ) . The permanence o f the
oes trogenic e f f e c t s o f sub te rranean clover graz ing on the ewe .
Au s t . Ve t . J . 2 7 : 63-6 7 .
Wa llace , L . R . ( 1 9 6 1 ) . Inf luence of l ivewe i ght and cond i t ion on ewe
f er t i l i ty . Proc . Ruakura Farmer s Conf . p . l4 .
Wa lles , B . , Edv ins son , L . , Owman , C . , S j oberg , N . O . and Sporrong , B .
( 1 9 7 6 ) . Chol inerg i c nerves and r e ceptors med i a t ing contract ion
of the Gr aaf ian f o l l i c le . Biol . Reprod , 15 : 5 6 5-5 7 2 .
102 .
Wa l les , B . , Falck , B . , Owman , C . and Sj oberg , N . ( 1 9 7 7 ) . Characteri zat ion
of au tonomic recep tor s in the smoo th mu scula ture of human Graa f i an
f o l l ic l e . B io l . Reprod , �: 4 23-43 1 .
We i s s , T . J . and Arms trong , D . J . ( 1 9 7 7 ) . Adenylate cyc l a s e a c tivi ty �n
sheep ovar ian f o l l i cu lar t i s su e ; developmental change s in hormone
re spons e . Proc . Aus t . Soc . Reprod . Biol . �: 46 .
We lchen , R . and Du l l aar t , J . ( 1 9 7 6 ) . Admini s tr a t i on o f ant i-serum
agains t ovine FSH or ovi ne LH at pro-Qe s tru s in the r a t ; e f f e c t s
on f o l l i cular deve lopment dur ing the oncoming cyc l e . J . Endocr ino l .
7 0 : 301-306 .

103 .
Wheeler , A . G . , Baird , D . T . , Land , R . B . and Sc aramu z z i , R . J . ( 1 9 7 7 a) .
Seasonal var i a t ion in oe s trus , the se cre t ion o f o e s tr ogen and
proge s terone and LH leve l s the ewe . J . Reprod . Fer t . 5 1 : 4 2 7 -4 3 2 .
Whee ler , A . G . , Baird , D . T . , Land , R . B . and Scaramu z z i , R . J . ( 1 9 7 7b ) .
Gene t i c variat ion in the secre t ion o f oes trogen in the ewe .
J . Endo cr inol . 7 5 : 337-3 3 8 .
Wi se , P . M . and Payne , A . H . ( 1 9 7 5 ) . The o e s t r ogen cytosol recep tor �n
fema l e ov ine p i tu i tary . Mo le c . C e l l . Endocr i nol . 3 : 385-395 .
Wong , E . ( 1 9 6 2 ) . D e t e c t ion and e s t ima tion of oes trogenic con s t i tuents
�n red c l over . J . Sci . Food Agr i c . !l: 304 .
Wong , E . and F lux , D . S . ( 19 6 2 ) . The oes trogenic a c t iv i ty o f red c lover
i so f l avones and some of their degrad a t ion produ c t s . J . Endo crino l .
24 : 34 1-348 .
Wong , E . , Flux , D . S . and Latch , G . C . M . ( 1 9 7 1 ) . The o e s t rogen i c ac tivity
of whi t e clove r . N . Z . J . Agr i c . Re s . 14 : 6 39-645 .
Wong , E . and Latch , G . C . M . ( 1 9 7 1 ) . E f f ect of fungal d i seases on pheno l i c
con tents of whi t e clover . N . Z . J . Agr i c . Re s . �: 6 3 3-638 .
Wong , M . S . F . and Cox , R . I . ( 1 9 7 1 ) . Evidence for phy to-o e s tr ogens as
su lpho-c onj uga t e s in sheep p l asma . Proc . 4 th Asia and O ceania
Cong. Endoc r i nol . Ab s t s . P . l 2 2 .
Wong , M . S . F . and Cox, R . I . ( l9 7 2 ) . Mi croanalysis o f phyto-oe s trogens .
J . Reprod . Fer t . 28 : 1 58 - 1 5 9 .
Wra th , R . H . and L i gh t f oo t , R . J . ( 1 9 7 6 ) . A s tudy o f reprodu c t ive was tage
among commerc ia l sheep f l ocks grazing oes trogenic pas tures �n
south We s tern Aus tralia . Pro c . Au s t . Soc . Anim . Prod . 1 1 : 2 25-23 2 .
Yang , N . S . T . , Mar s h , J . M. and Le Mai r e , W . J . ( 1 9 7 4 ) . Pos t ovul atory
change s in the concentr a t ion o f pros tagland ins i n r abb i t Gra f f ian
f o l l i c le s . Pros taglandins �: 3 7-44 .
Yu thas a s trako sol , P . , Howland , B . E . , Simarak s , s . and Palme r , W . M . ( 19 7 4 ) .
Oe s trogen-induced LH release in proges terone-trea ted ovar i e c tomized
ewe s . Can . J . Anim . S c i . 54 : 5 6 5 -5 7 2 .