arabidopsis phosphoribosylanthranilate isomerase: molecular
TRANSCRIPT

The Plant Cell, Vol. 7, 447-461, April 1995 O 1995 American Society of Plant Physiologists
Arabidopsis Phosphoribosylanthranilate Isomerase: Molecular Genetic Analysis of Triplicate Tryptophan Pathway Genes
Jiayang Li,'?' Jianmin Zhao,' Alan B. Rose,' Renate Schmidt,b and Robert L. Last a Boyce Thompson lnstitute for Plant Research and Section of Genetics and Development, Cornell University, Tower Road, Ithaca, New York 14853-1801
Department of Molecular Genetics, Cambridge Laboratory, John lnnes Centre, Norwich, NR4 7UJ, United Kingdom
Phosphoribosylanthranilate isomerase (PAI) catalyzes the third step of the tryptophan biosynthetic pathway. Arabidopsis PAI cDNAs were cloned from a cDNA expression library by complementation of an Escherichia co l i trpC- PAI deficiency mutation. Genomic DNA blot hybridization analysis detected three nonallelic genes encoding PAI in the Arabidopsis ge- nome. DNA sequence analysis of cDNA and genomic clones indicated that the PA17 and PA12 genes are virtually identical with only a single conservative amino acid difference throughout the deduced coding region as well as extensive conser- vation of introns and flanking regions. PA13 shows less identity (90%) with PAI7 and PA12. All three PAI polypeptides possess an N-terminal putative plastid target sequence, suggesting that these enzymes all function in plastids. The PAI7 gene is flanked by nearly identical direct repeats of -350 nucleotides. Our results indicate that, in contrast to most microor- ganisms, the Arabidopsis PAI protein i s not fused with indole-3-glycerolphosphate synthase, which catalyzes the next step in the pathway. Yeast artificial chromosome hybridization studies indicated that the PA12 gene is tightly linked to the anthranilate synthase a subunit 1 (ASAI) gene on chromosome 5. PAI1 was mapped to the top of chromosome 1 using recombinant inbred lines, and PA13 is loosely linked to PAH. cDNA restriction mapping and sequencing and RNA gel blot hybridization analysis indicated that all three genes are transcribed in wild-type plants. The expression of an- tisense PAl7 RNA significantly reduced the immunologically observable PAI protein and enzyme activity in transgenic plants. The plants expressing antisense RNA also showed two phsnotypes consistent with a block early in the pathway: blue fluorescence under UV light and resistance to the anthranilate analog 6-methylanthranilate. The extreme nucleotide conservation between the unlinked PA17 and PA12 loci suggests that this gene family is actively evolving.
INTRODUCTION
The tryptophan biosynthetic pathway in higher plants not only produces an essential amino acid but also provides precur- sors for synthesis of a variety of secondary metabolites, including the hormone indole-3-acetic acid (Wright et al., 1991; Normanly et al., 1993), antimicrobial phytoalexins (Tsuji et al., 1993), alkaloids (Cordell, 1974), and glucosinolates (Haughn et al., 1991). Although we have asophisticated understanding of this amino acid biosynthetic pathway in microorganisms, work on the genetics and biochemistry of tryptophan biosyn- thesis in higher plants has only recently begun to receive close scrutiny (reviewed in Last, 1993; Rose and Last, 1994). As shown in Figure 1, mutants of Arabidopsis are now available at four steps in the tryptophan pathway. These rnutants have distinct phenotypic characteristics: trpl phosphoribosylan- thranilate (PR-anthranilate) transferase mutants (defective in the PATl gene) are blue fluorescent under UV light due to
Current address: lnstitute of Genetics, Academica Sinica, Beijing 100101. P.R. China. * To whom correspondence should be addressed.
accumulation of anthranilate compounds (Last and Fink, 1988; Rose et al., 1992); trp4 anthranilate synthase p subunit (ASB) mutants have been identified as suppressors of the blue- fluorescence mutation trpl-100 (Niyogi et al., 1993); trp3 tryp- tophan synthase mutants have been identified as resistant to 5methylanthranilate (Last and Fink, 1988; E.R. Radwanski and R.L. Last, unpublished results); and trp2 mutants are resis- tant to 5-fluoroindole (Last et al., 1991; Barczak et al., 1995).
Because the enzyme plays an obligatory role in indolic sec- ondary metabolism, it is desirable to obtain mutants with reduced phosphori bosylanthranilate isomerase (PAI) activity, which catalyzes the conversion of 5-phosphoribosylanthranilate to l-(O-carboxyphenylamino)-l-deoxyribulose-5-phosphate (CDRP; Figure 1). However, no such mutants have been reported, despite analysis of hundreds of thousands of muta- genized Arabidopsis Columbia (COLO) or Landsberg erecta (Ler) seedlings for the blue-fluorescence phenotype associated with accumulation of anthranilate compounds or resistance to toxic anthranilate analogs. We reasoned that failure to ob- tain such mutants is most likely due to a flaw in the isolation

448 The Plant Cell
strategies employed or the expression of functionally redun- dant isoenzymes.
Genetic redundancy is extremely common in plants, includ- ing the relatively small genome of Arabidopsis (McGrath et al., 1993), and the existence of multiple functional genes might have thwarted the identification of PAI-deficient mutants of Arabidopsis. In fact, gene duplication is a common feature of aromatic amino acid biosynthetic enzymes in flowering plants. For example, of the seven aromatic amino acid biosynthetic pathway enzymes for which genes were previously cloned from Arabidopsis, five were demonstrated to be encoded by two or more nonallelic genes: 3-deoxy-~-arabino-heptulosonate Fphosphate synthase (Keith et al., 1991), 5-enolpyruvylshiki- mate-3-phosphate synthase (Klee et al., 1987), anthranilate synthase a subunit (ASA; Niyogi and f ink, 1992), AS6 (Niyogi et al., 1993), and tryptophan synthase l3 subunit (Berlyn et al., 1989; Last et al., 1991). In some cases, it appears that the in- dividual members of the gene families may serve distinct physiological functions. For example, although the 3-deoxy- D-arabíno-heptulosonate Fphosphate synthase and ASA en- zymes are encoded by duplicate genes in Arabidopsis, only one gene for each enzyme is induced in response to wound- ing and pathogen attack (Keith et al., 1991; Niyogi and Fink, 1992). Furthermore, the duplicate genes encoding ASA (Niyogi and Fink, 1992) and the tryptophan synthase p subunit (Last et al., 1991; Pruitt and Last, 1993) are differentially expressed in an organ-specific or developmentally regulated manner.
It is essential to obtain mutants blocked at all stages of the tryptophan biosynthetic pathway. Such a series of defective plants would allow the precise biochemical identification of the branch points leading to important “secondary” products. Mu- tants blocked at different steps in the pathway may accumulate different secondary products, some of which may confer in- teresting phenotypes or be useful for dissection of the pathways of aromatic secondary metabolism. Another advantage to ob- taining a complete set of pathway mutants is that it will allow a rigorous assessment of the regulated and rate-limiting cata- lytic steps. Because it may be impossible to obtain Mendelian mutants in all steps of the pathway (especially those encoded by multigene families), we asked whether it would be possi- ble to reduce the activity of an enzyme occurring early in the pathway, despite the existence of functional triplicate genes.
In this study, we report the isolation of cDNA and genomic clones encoding Arabidopsis PAI. There are three genetically unlinked genes (PAI7 to fAI3) for this enzyme, and they en- code unusually highly related enzymes. Despite the genetic redundancy, expression of antisense PAI7 RNA in transgenic plants resulted in dramatically reduced levels of immunologi- cally detectable PAI protein and enzyme activity. These antisense plants have phenotypes consistent with a reduced ability to convert PR-anthranilate to CDRP: they are blue fluo- rescent under UV light and resistant to 6-methylanthranilate. These results indicate that antisense gene expression should complement the classic genetic analysis of the tryptophan pathway.
RESULTS
lsolation of Arabidopsis PAI cDNAs by Suppression of an Escherichia coli trpC- Mutation
Functional complementation of microbial mutations has been used to isolate plant or animal cDNA or genomic clones for a variety of metabolic enzymes (for examples, see Henikoff et al., 1981; Elledge et al., 1991; Rose et al., 1992; Niyogi et al., 1993; Senecoff and Meagher, 1993). To obtain clones for Arabidopsis PAI, we selected for cDNAs that would comple- ment the E. coli trpC9830 mutation, which causes tryptophan auxotrophy due to a PAI activity defect (Yanofsky et al., 1971; Schechtman and Yanofsky, 1983), and 18 were chosen for further characterization (pJL19 to pJL36; see Table 1). Char- acterization of the plasmids from these colonies demonstrated that each contained a 1.0- to 1.5-kb insert with very similar re- striction maps; this is consistent with the isolation of cDNAs from either a single PAI gene or highly related genes. DNA sequence analysis confirmed that these cDNA inserts encode PAI polypeptides with similarity to the known microbial PAIS and that the cDNAs represent two distinct but remarkably simi- lar enzymes (see following discussion).
Analysis of Arabidopsis PAI Genomic Clones
The results of DNA sequence analysis of the cDNA inserts suggested the existence of at least two similar genes encod- ing Arabidopsis PAI. The results of genomic DNA gel blot hybridization experiments, such as that shown in figure 2A, suggested that there were three such genes. For example, prob- ing of genomic DNA digested with restriction enzymes EcoRI, Sacl, or Xbal, which do not cut within the 18 cDNA clones ana- lyzed, each yielded sets of three hybridizing bands. In contrast, the PAI7 cDNA hybridized with six Xhol fragments, consistent with the observation that there is a single Xhol restriction site in each class of Arabidopsis cDNA clone identified.
The hypothesis that there are three detectabie genes en- coding PAI was confirmed by probing Arabidopsis bacteriophage h genomic DNA libraries. Restriction enzyme mapping of the resultant clones indicated that approximately equal numbers of clones corresponded to the two classes of cDNAs (PAI7 and PAI2), whereas only one clone for the third locus (PAI3) was found. Detailed restriction maps were constructed for a single representative of each class of PAI genomic DNAs, as shown in Figure 28. Comparison of these data with the genomic DNA gel blot hybridization patterns (Figure 2A) confirmed that there are three highly related genes encoding PAI isoenzymes in Arabidopsis.
Restriction enzyme mapping and DNA sequence analysis of fAI7, PAI2, and PA/3 genomic DNA sequences and com- parison with PAI7 and PAI2 cDNA sequences revealed that all three are quite similar and that PAI7 and PAI2 are almost

Triplicate Arabidopsis Tryptophan Genes 449
Table 1. Plasmids Used in This Study
Designation PAI Vector Description
pJL24 pJL26 pJL28 pJL29 pJL33 pJL58
pJL59
pJL60; 61
pJL62; 63
pJL94
PAI2 phYES PAll plYES PAI7 plYES PA12 phYES PAI2 phYES PAI2 KS+”
PAl l KS+
PAll S K + d
PAll SK+
SK +
pJL118; 120 PAI2 SK+
pJL178 PAI3 SK+
cDNA ( - 48 to 1751)a cDNA ( + 31 4 to 1747)a cDNA ( - 94 to 1 732)a,b cDNA (-77 to 1732)a cDNA ( + 624 to 1631)” Subclone of the 11 5 k b Sal1
fragment from the genomic library in EMBLB
fragment from the genomic library in EMBL3
fragment from pJL59, both orientations
Subclone of the 2.2-kb EcoRV fragment from pJL59, both orientations
Subclone of the direct repeat sequence flanking fA17 excisable with Xhol
Subclone of the 3.6-kb Xbal fragment from pJL58, both orientations
Subclone of the 3.0-kb EcoRl fragment from the genomic library in XGEM-11
Subclone of the 8.5-kb Sal1
Subclone of the 2.0-kb Sacl
pJL193; 194 PA/3 KS+; SK+ Subclone of 5.1-kb Sall-EcoRI fragment from the genomic library in EMBL3
pJL197 a-PAI7 pB1121 Antisense PAll construct containing the Hindlll-Sacl fragment from pJL189
ing the N-terminal truncated PAI3 polypeptide
pJL257 PAI3 SK + Kpnl deletion of pJL194 encod-
a Corresponding nucleotide position in Figure 3.
tached to the 3’ end of PAI7 cDNA in pJL28. Approximately 0.4 kb of unknown Arabidopsis DNA fragment is at-
pBluescript II KS + . pBluescript II SK+.
identical. As shown in Figure 3, there are five exons (protein coding sequences indicated in boldface) and four introns in each PAI gene, with the four introns ranging from 90 to 350 bp. Remarkably, PA17 and PAI2 are nearly identical not only within the protein coding regions but also within regions that are generally less highly conserved; this includes the introns and sequences that are 364 bp upstream of the presumed AUG initiator codons and 470 bp downstream of the inferred stop codons; in total, the region spanning -364 to +1972 nucleo- tides shows 99% nucleotide identity, as indicated in Figure 3. In contrast, the region of high conservation of PA13with PA17 or PA12 extends from 63 bp upstream of the initiator codons through 56 bp downstream of the stop codons (from -82 to
+1556, as shown in Figure 3). The DNA sequence identity between this 1618-nucleotide PA13 sequence and the corre- sponding PA17 or PA12 sequence is 90%. The genomic DNA sequences are available from GenBank under accession num- bers U18970 (PAH), U18968 (PA12), and U18969 (PA13).
Direct Repeats Flank the PAll Gene
Analysis of the PAll genomic DNA sequence revealed an un- expected feature of the organization of this locus: PA17 is located between two 99% identical direct repeat elements of >353 nucleotides flanking the coding region. (The two direct repeat elements correspond to nucleotides 303 to 656 and 3433 to 3785 under GenBank sequence accession number U18970.) The existence of these repeats was verified by genomic DNA gel blot hybridization analysis using a cloned repeat element as a probe, which is shown in Figure 4. Digestion with Hhal, which cleaves once within each element, yielded the expected strongly hybridizing fragment of 3.1 kb and two weaker junc- tion bands. Digestion with Xhol resulted in two expected bands of 0.35 and 6 kb, each corresponding to one of the elements (Figure 4). No other copies of this repeat sequence were de- tected by genomic DNA analysis under these hybridization conditions.
Comparison of the PAl1, PA12, and PAl3 Coding Regions
A comparison of the amino acid sequence of the predicted Arabidopsis and microbial PAI proteins is shown in Figure 5. For each PAI gene, protein translation was assumed to start at the first AUG codon of the longest PAI cDNA (pJL169). The three genes are capable of encoding proteins of 275 amino acid residues, with calculated molecular masses of 29.6 kD. The PAI1 and PAI2 polypeptides differ only by a single con- servative change of Glu to Asp at amino acid 136. The more divergent PAI3 has 18 amino acid residue differences com- pared with PAll and 19 compared with PA12. The inferred plant PAI protein contains severa1 conserved motifs among the microbial proteins. They include two domains of absolutely con- served amino acids (VGVF and VQLHG) as well as three other conserved domains (GGSG, LAGGI, and GIDVSSGI), as indi- cated by the boxed areas in Figure 5. Overall, Arabidopsis PAI protein shares 20 to 27% amino acid identity with the various microbial proteins.
As previously observed for other Arabidopsis enzymes of aromatic amino acid biosynthesis (Rose and Last, 1994), the three Arabidopsis PAI proteins have N-terminal extensions not present in the corresponding microbial proteins (Figure 5). Three lines of evidence suggest that these are plastid transit peptides. First, they are rich in positively.charged and hydrox- ylated amino acids, both characteristic of plant transit sequences (von Heijne et al., 1991). In the first 65 amino acid
450 The Plant Cell
Erythrose-4-P + Phosphoenolpyruvate
I DAMPS (2)
3-Deoxy-D-arab/'no-heptulosonate-7-PIIt
ShikimateI EPSPS(2)
5-&rapyruvylshikimate-3-P
Anthranilate-p'-glucoside
Prephenate
Arogenate
/ \Tyrosine Phenylalanine
Anlhranilatetrpl 1 PAT (1)
5-PhosphorlbosylanthranllateI PAI (3)
1-(O-Carboxyphenylamlno)-1-deoxyrlbulo«e-S-P1 InGPS
Indole-3-glycerolphosphate
trp3 1 TSA
Indole
"P2 I TSB (2)
Tryptophan
Figure 1. The Aromatic Amino Acid Biosynthetic Pathway inArabidopsis.
Selected enzymes are shown with the gene copy number per haploidgenome indicated in parentheses. The mutants of the tryptophan bio-synthetic pathway (from chorismate to tryptophan) are given at thecorresponding steps (trpl to trp4). DAHPS, 3-deoxy-D-arab/no-heptulo-sonate 7-phosphate synthase; EPSPS, 5-eno/pyruvylshikimate-3-phosphate synthase; CM, chorismate mutase; ASA, anthranilatesynthase a subunit; ASB, anthranilate synthase P subunit; PAT, phos-phoribosylanthranilate transferase; PAI, phosphoribosylanthranilateisomerase; InGPS, indole-3-glycerolphosphate synthase; TSA, tryp-tophan synthase a subunit; TSB, tryptophan synthase (5 subunit. Thedashed line indicates that multiple steps are involved. P, phosphate.
residues, RAM and PAI2 each have 15 hydroxylated amino acids(Ser or Thr) and 12 positively charged residues (Arg, His, orLys), whereas PAI3 has 17 hydroxylated amino acids and 13positively charged residues. Second, a truncated PAI2 cDNA(pJL33) lacking the first 65 amino acid residues is able to com-plement the E. co// trpC9830 mutation, indicating that these65 amino acids are not required for producing an active PAIenzyme. Third, the in vitro-synthesized PAI2 precursor proteinis imported into purified spinach chloroplasts and processedinto a smaller polypeptide (Zhao and Last, 1995), with the cleav-age site shown by the arrow in Figure 5.
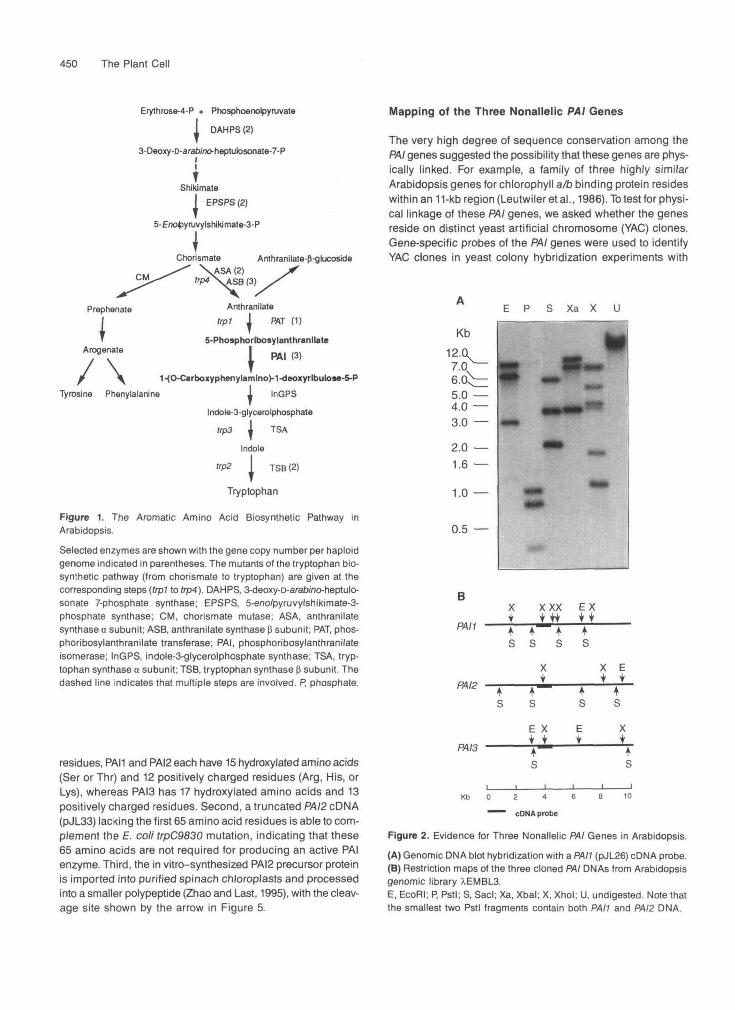
Mapping of the Three Nonallelic PAI Genes
The very high degree of sequence conservation among thePAI genes suggested the possibility that these genes are phys-ically linked. For example, a family of three highly similarArabidopsis genes for chlorophyll a/b binding protein resideswithin an 11-kb region (Leutwiler et al., 1986). To test for physi-cal linkage of these PAI genes, we asked whether the genesreside on distinct yeast artificial chromosome (YAC) clones.Gene-specific probes of the PAI genes were used to identifyYAC clones in yeast colony hybridization experiments with
A
Kb2.07'°\6.0^5.0 —4.0 —3.0 —
2.0 —1.6 —
1.0 —
0.5 —
B
E P S Xa X U
U .*P « **^p *̂ tMMift
•»mm
«•» ***
X X XX E X1 III II
PA" 1 1 1 1S S S S
X X E1 I I
M I I I IS S S S
E X E XI I I 1
i _Kb 0
1 1s s
1 1 1 12 4 6 8 1(
^~~ cDNA probe
Figure 2. Evidence for Three Nonallelic PAI Genes in Arabidopsis.
(A) Genomic DNA blot hybridization with a PAH (DJL26) cDNA probe.(B) Restriction maps of the three cloned PAI DNAs from Arabidopsisgenomic library XEMBL3.E, EcoRI; P, Pstl; S, Sacl; Xa, Xbal; X, Xhol; U, undigested. Note thatthe smallest two Pstl fragments contain both PAH and PAI2 DNA.

PAI 1 PAI2 PAI3
PAIl PAI2 PAI3
PAIl PAI2 PAI3
PAIl PAI2 PAI3
PAIl PAI2 PAI3
PAIl PAI2 PAI3
PAIl PAI2 PAI3
GTG-AITITGCn;CTATCAAT-Cn;C~AG~TGAWCA~ATATTATA~'M'-------TGC~CATG~A~ACTAATAGGAATCWG
A. .T.
TAAGAWAGTTGATGAA TTATITITCCTC ....-c........
.................................................................................................... ........... TTGCTn:.CAA..T... C. ..T.. T
.................
PAIl AGCUVLTATCCACTQRTCTCCA~TCCA~T-~TCT~T~C~CT-TC~TAACTTTTCTTGACTTTTAGTTTTTCTTCAGATT PAI2 . . . . . . . . . . . . . . . . . . A . . C . . . . . . . . . . . . . . . . . G . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . pA13 .......... T ..........cc..... C C . A . . . . . . . G . . . . . . . . . . . . . . . . ~ . . . . . . . . . . T . . . . . . ....G.....-.G....A..G..... PAIl W A C T A G A C C T T ' C T G T G T G T G ~ A T T C W ~ ~ C G G C T A A T T A C ~ ~ ~ C A ~ A ~ ~ ~ C T A C ~ A ~ T C C A ~ ~ PAI2 ........................................................................... ......................... PAI3 ........--A.T..........G................TAT...G.....T.......C.........G....T........A..CT...........
PAIl MTCOAACOOTGTCTTTTTC~GAGTCGGGTAT~TCACUVLCA~~TTGA~T~AGTGTTTCCMCACA~G~TGTA~TCCTM~ATGAT- PAI2 .................................................................................................... p ~ 1 3 ....... A ............A..T...........................................G.... .......................... c .
PAIl ~ T Q R ~ ~ T ~ ~ ~ T Q R T ~ C T C ~ C ~ ~ T ~ ~ T W T C T A C A T T C T T A T C G A T T T T T A T T T T G C W T C A C C ~ G
p~13 ....................c............................................................................... PAI2 ........C........................................................................G........ . . . . . . . . . .
PAIl T G A T T T T C T C A A C T T T A T T A A C G G T T G A T A G T T A T G G T T T C T T A C A ~ - ~ M ~ ~ ~ G ~ ~ ~ T C ~ T ~ ~ ~ PAI2 .................................................................................................... p~13 ........ . . . G . . . . . . . . . . . . . . . . . G . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . . .. A.. . . .
-301
-201
-101
-1
100
200
300
400
500
600
700
800
900
1000
PAIl AGTCATATATGTCCTTMTOCAAATCA~T~~~TCTTGMTGMGTTCC~MGA~ATTGTCATCTT~TGATTGGATTCTTGT~ATAGT~A 1100 PAI2 .................................................................................................... p ~ 1 3 ......................... 0 ......... .....c......~........................c........................... PAIl PAI2 PAI3
ACOOCYrOOOAOGTACCAAG~CAAC~AA~TT-------~~ATTGTAATGTAC~CTTAGAA~~AAGCAAWACTCTAGTACAT- 1200 G ................ T ...T........ A.G ......CGCACTG..T............T.....C..G...CG.G...CA.............C... G
...................................................................................................
PAIl A A A A C C A m G W T T T C A C G A G G T G A ~ G ~ T A W A A G A G T G T G A T A G G T T C A A T C A T ~ C A A T ~ C T G C A G ~ ~ ~ ~ ~ T 1300 PAI2 .................................................................................................... ............................... .T. .... A.A ...................
CTTA~TGOOOGAATCMTCCMC~TGTTTCAGM~TCTTTCTATCCTTCMCCTGAT~ 1400 PAI2 .................................................................................................... PAI3 . . . . . . . A . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
PAIl MTTGATGTTAGTAGCOGTATTT~OOTACAGACOOTATCCAGMOGATMGTCTMGATM~TCCTTTATMCT~AGTTC~TCTGTACACTACT~ 1500
p ~ 1 3 ............c.. T ........... c.T......~....................... ... T ..... c..............~............... PAIl T G G C A A G C A A T A T A A A C C A C G G T A A T T T A T C T T G T A A C T A T A G ~ A ~ A ~ ~ C C T G ~ m C ~ ~ ~ ~ W A ' M ' ~ G ~ ~ ~ T A A C T G 1600
PAI2 .................................................................................................... - - ........................ ............ ................................ .G ......................
PAIl GCTGATAATAATAATAATGCCCTTmTTTcATcAGTAGTAGTAGA--~AAA~~GCG~TTGWACCGTAAT~TGTCG~~W~GAA 1700
PAIl CAAGATCCTXTATG-AATGTATTCTACTGATCATATA'M'ATl'TCATTTGAWG ACAGGmTA
PAI3 AGC. ...........
ACTGCAGTATGTCCTCATGT'M 1900 ........................ PAIl TGWCATXAGTTFEGAWTGCCWATA GCATTGCTIAAAm--TGATAATAGGT 2000 PAI2 ............................. AT.C.CT...TGA..CT.CCC.G..A..
Figure 3. DNA Sequence Alignment of PAI1, PAl2, and PA13.
The inferred protein coding sequences present in the cDNA clones are shown in boldface. A dot represents an identical base, and a dash indicates a gap. The assumed start codon, defined as position 1, is the first ATG found in the correct open reading frame of the longest cDNA clone, and boldface type represents the protein coding regions common to the three genes. Underlined triplet regions indicate locations of the presumed translational initiator and terminator codons. The genomic clone sequences are available from GenBank under the following accession numbers: PAII, U18970; PAl2, U18968; PA13, U18969.

452 The Plant Cell
« oI XKb
12.(k7-°\ IT6.0 vl—,5.0 —4.0 —3.0 — •*-
2.0 —1.6 —
1.0 —
0.5 —
3128 bp
PAH \ I
Figure 4. PAH Is Flanked by Two Nearly Identical Direct RepeatSequences.Arabidopsis DMA (20 ng per lane) was digested with Hhal or Xhol,and the 0.35-kb direct repeat sequence, which is diagrammed below,was used as a probe during DMA gel blot hybridization. Hhal diges-tion produced an expected 3.1-kb strongly hybridizing band and twoweaker junction bands. Digestion with Xhol resulted in two expectedstrong bands during DMA gel blot hybridization with the direct repeatsequence; the weaker band may have resulted from partial digestion.
three YAC libraries (Ecker, 1990; Ward and Jen, 1990; Grilland Somerville, 1991).
A comparison of the coordinates of the YAC clones contain-ing the PAI gene sequences with coordinates of YAC clonesthat previously had been mapped to chromosomes 4 and 5of Arabidopsis (R. Schmidt, J. West, K. Love, and C. Dean,unpublished results) showed that five (EW5A4, EW7E12,EW9F12, EW16C1, and yUP3E1) of the six YAC clones con-taining the PAI2 gene sequences also contained sequencescorresponding to ASA gene 1 (ASA1), which was previouslymapped onto chromosome 5 (Niyogi and Fink, 1992; Jarviset al., 1994). This result, together with the YAC mapping results,places PAI2 on chromosome 5 in the immediate vicinity of ASA1,as shown in Figure 6. It appears that these two genes are within110 kb of each other because YAC clones EW5A4 and EW9F12are 110 and 125 kb in length, and each contains sequencescorresponding to both PAI2 and ASA1.
The PAH gene has been placed on the top of chromosome1 by restriction fragment length polymorphism (RFLP) mappingof recombinant inbred lines (Lister and Dean 1993). A RFLP
for restriction enzyme Fokl was identified between Ler andCol-0, and this polymorphism was examined in 55 recombinantinbred lines. Linkage analysis placed the PAH polymorphism~3 centimorgans (cM) from marker m488. Screening of YAClibraries with a PAH probe identified YAC clones that alsocontain the previously described duplicate cDNA clone 171(McGrath et al., 1993). YAC clones EW13E7 and EW20B12 are125 and 135 kb in length, and both clones hybridized to PAHas well as cDNA 171, suggesting that these sequences areseparated by <125 kb. This result, together with the recom-binant inbred mapping, places one of the loci correspondingto cDNA 171 close to PAH on chromosome 1 (Figure 6). ThePAI3 gene was mapped at ~50 cM on chromosome 1, betweenmarkers nga280 and nga248 (J. Bender and G. Fink, personalcommunication). These results indicate that the three PAIgenes are not tightly linked in the Arabidopsis genome.
PAI Is Not Fused to lndole-3-GlycerolphosphateSynthase
PAI is commonly synthesized as a fusion protein with one ortwo other enzymes of the tryptophan pathway in both fungiand bacteria. For example, it is fused with indole-3-glycerol-phosphate synthase to form a bifunctional enzyme in E. coli(Christie and Platt, 1980) and other bacteria (Horowitz et al.,1983; Crawford et al., 1991). These two activities are fused intoa trifunctional protein containing the ASB in Neurospora crassa(Schechtman and Yanofsky, 1983) and other fungi (Sanchezetal., 1986;Choietal., 1988; Kosetal., 1988; Adams and Royer,1990). In contrast, the Arabidopsis PAIs appear to be synthe-sized as monofunctional proteins. For example, analysis of 1.5kb upstream and 0.5 kb downstream of the PAI genomic DNAsequences revealed no homology with Arabidopsis indole-3-glycerolphosphate synthase or any other tryptophan biosyn-thetic pathway enzyme sequence. In agreement with thesequence results, an anti-PAl antibody cross-reacts with a 29-kDPAI polypeptide, which is the size predicted for PAI enzyme(see following discussion; see also Zhao and Last, 1995). Fur-thermore, cDNAs for Arabidopsis indole-3-glycerolphosphatesynthase are not homologous to the PAI genes (Li et al., 1995).
Expression of PAH, PAI2, and PAI3
Although the high degree of sequence conservation suggestedthat all three genes are expressed, it was not clear whetherthe PAI3 gene was transcribed, because no PAIS cDNA cloneswere identified. Therefore, we directly tested this hypothesisby RNA gel blot hybridization. As shown in Figure 7A, a tran-script of ~1.3 kb was observed when a PAH cDNA fragment(pJL48), which is expected to hybridize to products of all threegenes, was used as a probe. A gene-specific oligonucleotidehybridization probe (JL2) provided a specific assay for the invitro-synthesized PAI3 transcript, whereas oligonucleotide JL1hybridized to PAH (and presumably also to the nearly identical

Triplicate Arabidopsis Tryptophan Genes 453
24 .o 26.9 ' 39.9
50
68.3
113.3
Athl Ath2 Ath3 Sce Eco
Athl Ath2 Ath3 Sce Eco
-
- '
-
+ I lntron 1 I lntron 2 MSTGISTDLHVHFGALNFSKTYKSGLSNRTVSFSRVGYAQNRKLSCSVSNTENVAPKDDERGKDRPLVKMCGITSARDAAMAVEAGADFIGMIIWPHSKR 100 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ......S...L.PR.......S.......K.. ................................
N.V- .L.RGQ..KA.YD.. -1YG.L.FVAT.P.
200 ................................ ..E......V..... G.
QPKEDV.ALVNDYGID . . . . . E.-------WQEYQEFLGLPVIKRLVFPKDCNI..SAASQKPHS H.1ADWDKAKVLS.V EEQLYIDTL-E.LPAH.AIWKALS.G---------ETL-AREFQ
S
T.DPVI.RK..TL-KAYKNSSGT.KY CVNVEQ.QEV---------MA.A.LQ
SRNG--WL .i-j. AGGI PTNVSEALSILQ 292
.PESLHF . . . . .E . .GD. .RLNG GQTLGNV.. . . . AD.CV. .AQT-G
Figure 5. Comparison of Arabidopsis PAII, PA12, and PAI3 Amino Acid Sequences.
The entire inferred amino acid sequence of PAI1 (Athl) is presented using the single-letter code. Amino acid residues identical to the PAII se- quence are represented by dots. The intron positions are shown by vertical lines. The gaps are indicated with dashes. The arrow shows the cleavage site of the chloroplast transit sequences of the Arabidopsis PAI2 protein. The phylogenetically conserved amino acid domains are indi- cated with boxes. Sce, Saccharomyces cerevisiae (Tschumper and Carbon, 1980); Eco, E. coli (Horowitz et al., 1983).
fA12) RNA (Figure 78). The gene-specific JL2 probe identified a transcript from total Arabidopsis leaf RNA (Figure 7A), indi- cating that fA13 is expressed in the leaf.
Consequences of Reduced PAI Expression: Antisense PAl7 Transgenic Plants
The observation that the Arabidopsis Col-O ecotype possesses three functional genes for the PAI enzyme may explain the inability to identify mutants defective for this enzyme, despite the recovery of numerous mutations affecting the previous en- zyme in the pathway, PR-anthranilate transferase (Last and Fink, 1988; Rose et al., 1992; A.B. Rose and R.L. Last, un- published data). It is desirable to obtain plants with reduced PAI activity because, in contrast to tryptophan synthase- deficient mutants (Wright et al., 1991; Normanly et al., 1993), mutants altered in earlier steps of the pathway are expected to be defective in the synthesis of indolic secondary metabo- lites. Consistent with this hypothesis, frp7-7 auxotrophic mutants display a syndrome of morphological defects, consistent with an auxin defect (Last and Fink, 1988). These consideraiions suggested the use of antisense RNA expression to reduce the synthesis of PAI protein. We reasoned that the high degree of nucleotide identity shared among the three genes made it likely that antisense expression of fA17 cDNA would also reduce the expression of the nearly identical fA12 gene and perhaps would affect all three genes. Transgenic plants con- taining the fAl7 antisense cDNA expressed from a cauliflower mosaic virus 35s promoter or the vector control were regen- erated from Agrobacterium-infected Arabidopsis roots on medium containing tryptophan (to support growth of auxo- trophic plants). These primary regenerants were allowed to self-pollinate to produce T2 seed. The kanamycin resistance
1
PAI1 I1 71 m488
g3786
PAI3
GAP-B
ADH
14.8
20.1 25.5
61.5
81 .o
- PAI2/ASA1
- trpl/PATl - tt4
- tt3
- trpWSB1
Figure 6. Map Locations of the Three PAI Genes.
The map distance between PAI7 and m488 was calculated from the 55 recombinant inbred lines using Mapmaker version 2 as described in Methods. PAI7 and PAI2 are located on the map based on the in- tegrated geneticlRFLP map of the Arabidopsis genome (Hauge et al., 1993). The approximate map position of PAI3 was determined by J. Bender (personal communication). The numbers to the left of the chro- mosomes indicate map positions in centimorgans.

454 The Plant Cell
segregation ratio of the T2 plants indicated that four of the 12antisense transgenic lines obtained appeared to have a sin-gle expressed kanamycin resistance locus, and the remainingtransgenic lines had functional inserts at multiple loci (J. Li,data not shown). A true-breeding T3 antisense transgenic line(197-5) and vector control line (199-1) were chosen for detailedcharacterization.
Phenotypes of the PAH Antisense RNA Mutants
Although the growth rate and morphology of the 197-5 anti-sense transgenic plants were identical to those of the vectorcontrol in the presence or absence of tryptophan (Figure 8Band data not shown), this line manifested two whole-plantphenotypes that are consistent with a reduced ability to convertPR-anthranilate to CORP. First, as shown in Figure 8A, the197-5 antisense transgenic plants were resistant to 6-methyl-
Probe =!
anthranilate. Anthranilate analogs have previously been shownto have herbicidal activity against a variety of plant species(Thomas, 1984). The toxicity of at least a subset of these her-bicides appears to be a result of the conversion to tryptophananalogs because Arabidopsis trpl, trp2, and trp3 mutants,which are blocked in three different steps of the tryptophanbiosynthetic pathway (Figure 1), are resistant to 5-methyl-anthranilate (Last and Fink, 1988; Rose et al., 1992). Theseantisense plants produced green true leaves, whereas the wild-type or vector control plants produced chlorotic cotyledons anddid not make true leaves on 6-methylanthranilate-containingmedium. This result is consistent with reduced activity of atleast one enzyme in the tryptophan biosynthetic pathway.
Second, 197-5 plants are blue fluorescent under UV illumin-ation (Figure 8B), suggesting that these plants accumulateanthranilate derivatives. This hypothesis was tested by sub-jecting extracts from 197-5 leaves to reverse phase HPLCanalysis. The antisense plants accumulated a UV-absorptivecompound that cochromatographs with anthranilate (3-glucosidefrom trp1-100 extracts (J. Li, data not shown). To characterizefurther the presumptive anthranilate-containing compoundfrom 197-5, it was purified by HPLC and treated with almondp-glucosidase (Last and Fink, 1988), and the products wereassayed by HPLC. The resultant UV-absorptive compound co-eluted with the authentic anthranilic acid standard (J. Li, datanot shown); this is consistent with the hypothesis that the majorblue-fluorescent compound accumulated in 197-5 is anthrani-late p-glucoside. The accumulation of anthranilate p-glucoside,rather than an N-ribosylated compound, is consistent with previ-ous results indicating that such anthranilate glycosylaminesare chemically unstable in vivo and in extracts (Doy et al., 1961).
B
Probe
£2 — — — 2RNA g: g; g| g: g
Figure 7. RNA Gel Blot Hybridization Analysis.(A) Hybridization of total RNA (20 ng per lane) isolated from rosetteleaves. Probes used in the hybridization are as indicated above thegel. The nonspecific probe for all three PAI genes is a 0.7-kb Pstl PAHcDNA fragment. JL1 and JL2 are synthesized oligonucleotide probesspecific for PAH and PA/2 (JL1) or PA/3 (JL2) mRNAs.(B) RNA blot hybridization of in vitro-transcribed RNA with oligonu-cleotide probes JL1 and JL2 demonstrates that the probes are transcriptspecific.
Biochemical and Molecular Characterization ofAntisense PAI RNA Mutants
The finding that antisense PAH transgenic plants accumulatedanthranilate derivatives and are resistant to 6-methylanthrani-late suggests that there is a significant reduction of PAI enzymeactivity in these plants. As shown in Table 2, assays of PAIenzyme activity in leaf tissue demonstrated that 197-5 plantsproduced ~15% of the PAI activity found in the wild-type, vec-tor control, or trpl-100 positive control plants. In contrast, theactivity of the previous enzyme in the pathway (PR-anthranilatetransferase) was similar in the wild-type and antisense 197-5-and vector 199-1-transformed plants and reduced in the con-trol trpl-100 plants. Consistent with the results of enzymeassays, immunoblot analysis showed that leaves of the an-tisense plants accumulated ^10% of control levels of PAIcross-reactive material (Figure 9A and Table 2). That PR-anthranilate transferase, ASA, and tryptophan synthase a andP subunits accumulated to wild-type levels (Figures 9B to 9Dand Table 2) demonstrates that the antisense transgene is spe-cifically influencing PAI protein. The close agreement betweenresidual enzyme activity (~15%) and protein accumulation(~10°/o) suggests that the polyclonal antiserum raised against

197-5 Col-0
199-1 197-5 Col-0 trp1-100Figure 8. Phenotypes of Antisense PAH Transgenic Plants.
(A) A segregating population from antisense line 197-5 (left) and wild-type Col-0 (right) were grown on PNS agar plates containing 300 \iM 6-methyl-anthranilate and 25 uM L-tryptophan for 3 weeks.(B) Plants of the vector control (199-1). antisense line (197-5). wild-type Col-0, and blue-fluorescent mutant trpl-100 were germinated and grownon PNS agar plate for 10 days and then transplanted into soil for another 10 days of growth. The photograph was taken using a blue-light WrattenGelatin Filter No. 47 (Kodak) under UV light.

456 The Plant Cell
Table 2. Biochemical Characterization of Antisense Plants
Assay3
PAI proteinPAI activityPAT activityBlue-fluorescent compounds
197-5
1014
105670
trpl-100
102101<5860
Col-0999085
118
" Values are given as percentages of the vector control transgenicplant 199-1.
the PAI2 protein reacts with all three isoenzymes present inleaf tissue.
to the case for many bacteria and fungi in which PAI, indole-3-glycerolphosphate synthase, and ASA are often synthesizedas a multifunctional polypeptide (Mutter et al., 1986). This ob-servation extends previous observations indicating that plantPR-anthranilate transferase, ASA, and the tryptophan synthaseP subunit are encoded as monofunctional enzymes (Berlynet al., 1989; Niyogi and Fink, 1992; Rose et al., 1992). Unlikemicroorganisms, the plant tryptophan pathway apparently con-sists of monofunctional enzymes.
Duplication of expressed sequences is a common featurein plants, including the relatively small genome of Arabidop-sis. For example, McGrath et al. (1993) estimated that 17% of90 independent randomly selected cDNAs hybridized to non-tandemly duplicated loci in the Arabidopsis genome. Althoughthe amount of nucleotide conservation varies widely among
DISCUSSION
We have characterized three Arabidopsis genes encoding PAI,the third enzyme of the tryptophan biosynthetic pathway (Fig-ure 1). The cDNA clones were isolated by complementation ofan £ co// trpC9830 mutation with an Arabidopsis cDNA ex-pression library in XYES (Elledge et al., 1991). Comparison ofthe sequences of these cDNAs indicates that they producenearly identical isoforms, encoded by the genes PAH and PAI2.Genomic DNA gel blot hybridization with these cDNAs revealedthat there is an additional PAI gene that was not identified bycDNA analysis (Figure 2).
The predicted amino acid sequences of the three PAI genesare homologous to those from bacteria and fungi, as shownin the boxed areas in Figure 5. Strikingly, the predicted PAHand PAI2 proteins are virtually identical, with only a single con-servative amino acid change from Glu to Asp in a total of 275amino acids. In contrast, the PAI3 protein has 18 or 19 aminoacid residue differences compared with PAH and PAI2. Noneof these changes are located in the five highly conserved do-mains observed in the protein sequence alignment (Figure 5).
Two lines of evidence indicate that all three PAI genes aretranscribed in Arabidopsis leaf tissue. First, approximatelyequal numbers of PAH and PAI2 cDNAs were isolated byfunctional complementation in £ co//. Second, RNA blot hybrid-ization analysis of total Arabidopsis leaf RNA with gene-specificoligonucleotide probes showed that the PAI3 mRNA signal wasapproximately equal to that of PAH and PAI2 combined (Fig-ure 7A). However, the question of whether the three PAI genesare subject to spatial, developmental, or environmental regu-lation awaits further investigation.
YAC hybridization and genetic mapping results are consis-tent with the notion that the three PAI genes are nonallelic andunlinked. PAH mapped close to m488 at the top of chromo-some 1, PAI3 mapped at ~50 cM on chromosome 1, and PAI2was linked to ASA? on chromosome 5 (Figure 6). However, PAI2and ASA1 are still physically separated from each other anddo not form a fused protein. This result is in striking contrast
§o>cn -o g.
ITmm» 4 PAI
B ASA
PAT
Figure 9. Immunoblot Analysis of Tryptophan Pathway Proteins.The sources of the extracts are indicated at top. Equal amounts of to-tal protein from 197-5,199-1, wild-type Col-0, and trp1-100 were appliedto SDS-polyacrylamide gels.(A) Membrane probed with anti-PAl antibody.(B) Membrane probed with anti-ASA antibody.(C) Membrane probed with anti-PR-anthranilate transferase antibody.(D) Membrane probed with anti-tryptophan synthase a and anti-trypto-phan synthase (3 antibodies.

Triplicate Arabidopsis Tryptophan Genes 457
members of well-characterized plant gene families, in published examples, nonallelic genes that are absolutely conserved or nearly identical tend to be present in tandem repeats. For ex- ample, two genes for tobacco sesquiterpene cyclase (€AS7 and €AS2), which are separated by 6 kb, are absolutely identical, including 388 nucleotides upstream and 40 nucleotides down- stream of the transcription start site (Facchini and Chappell, 1992). Similarly, there are six genes for the ice plant ribulose bisphosphate carboxylase/oxygenase small subunit family, five of which are closely linked (DeRocher et al., 1993). Two genes of this tandem cluster (rbcS-4 and rbcS-5) are absolutely conserved at the nucleotide leve1 throughout the transcribed region. Thus, it is remarkable that the PAI7 and PAI2 loci are 99% identical throughout -2350 nucleotides, including introns and flanking nontranscribed regions, despite the fact that they mapped to separate chromosomes.
The finding that the PAI7 and PA/2 genes are virtually iden- tical, including hundreds of base pairs of flanking and intronic sequences (Figure 3), is consistent with an actively evolving gene family. One possibility is that these genes were created by a relatively recent gene duplication event. An alternative hypothesis is that there may be an active recombinational mechanism maintaining the homogeneity of these two unlinked sequences. There is evidence for gene conversion between nonallelic genes influencing the rate of nucleotide divergence in plants (see Meagher et al., 1989). The presence of two di- rect repeat sequences flanking PAI7 (Figure 4) may have a role in maintaining the high degree of sequence similarity. For ex- ample, it is possible that this repeat was created by a recent transpositional or recombinational event. The existence of such highly conserved genes should provide a system for future studies of gene duplication and sequence evolution. If this gene family is as evolutionarily dynamic as our data suggest, it should also provide opportunities for phylogenetic studies of populations of Arabidopsis and related species.
Previous attempts to identify mutants lacking PAI activity failed, presumably due to the presence of three expressed genes in the COLO ecotype. In this study, the antisense RNA approach was successfully employed to obtain PAI-deficient plants, despite the existence of a multigene family. This is an especially important observation because future studies of aro- matic secondary metabolism will benefit from the availability of plants blocked at each step of indole ring biogenesis. The observation that enzyme activity is decreased in the antisense plants confirms that these PAI genes, which were identified by functional complementation, encode Arabidopsis PAI. The phenotype of these antisense PAI7 plants resembles that of leaky trp7 mutants, which are defective in PR-anthranilate transferase (Rose et al., 1992), the enzyme catalyzing the preceding biosynthetic reaction (Figure 1).
It is surprising that no tryptophan-requiring auxotrophs were recovered among the antisense transgenic plants. One pos- sible explanation is that tryptophan auxotrophic calli grew significantly more slowly than the prototrophs, despite the pres- ente of 50 pM tryptophan in the tissue culture media. In this
scenario, the auxotrophs would not be identified as healthy kanamycin-resistant calli or shoots. Such slow growth in tissue culture has been observed for trp7-7 (A.B. Rose, unpublished data). Alternatively, examination of a larger collection of trans- genic lines may be necessary to identify antisense plants that possess sufficiently low PAI activity to require tryptophan for growth. A third possible explanation is that expression of the antisense PAI7 differentially decreases expression of the three genes. Analysis of a larger collection of transgenic lines should allow us to test these hypotheses.
METHODS
Plant Culture
The Arabidopsis thaliana Columbia (Col-0) wild-type ecotype was grown on sterile agar media or Redi-Earth (Grace-Sierra, Milpitas, CA) un- der continuous illumination (80 to 120 pmol m+ sec-l) at 22% as described previously (Last and Fink, 1988). lnhibitor resistance was tested on sterile PNS agar medium (Haughn and Somerville, 1986) containing 300 pM 6-methylanthranilate and 25 pM L-tryptophan. The '
plants were scored 3 weeks postimbibition. Transgenic T3 plants used for phosphoribosylanthranilate (PR-anthranilate) transferase (PAT) or phosphoribosylanthranilate isomerase (PAI) enzyme activity assays were grown on soil-less mix for 2 weeks, after transferring from agar PNS medium containing 50 pg/mL kanamycin.
lsolation of Arabidopsis PAI cDNAs
Descriptions of the plasmids generated in this study are summarized in Table 1. Arabidopsis PAI cDNAs were isolated based on the ability to complement an Escherichia coli trpC- mutation according to the method described by Elledge et al. (1991). E. coli trpC9830, defective in PAI (Yanofsky et al., 1971; Schechtman and Yanofsky, 1983), was lysogenized with 1KC (which encodes the Cre recombinase neces- sary for conversion of 1YES to the plasmid form) to produce strain RLB809. This Trp- strain (1.6 x 1Ol0 cells) was infected with 3.6 x 108 plaque-forming units of an Arabidopsis cDNA expression library in 1YES and grown on the tryptophan-deficient medium of Vogel and Bonner (1956), which contains E salts with 1% dextrose and 2.5% casein hydrolysate (salt- and vitamin-free grade; ICN Biochemicals, Cleve- land, OH) plus 50 pg/mL ampicillin. Eighteen colonies were picked from 111 positive colonies, and the pYES plasmids (derived by Cre- mediated site-specific recombination of 1YES) isolated from them were designated pJL19 through pJL36. The five cDNA inserts of different lengths from pJL24, pJL26, pJL28, pJL29, and pJL33 were subcloned in both orientations into pBluescript I1 KS+ or SK+ (Stratagene), yield- ing pJL166 and pJL167, pJL48 and pJL49, pJL168 and pJL169, pJL170 and pJL171, and pJL50 and pJL51, respectively.
Screening an Arabidopsis Genomic Library
The Pstl fragment containing the 700-bp PAI7 cDNA from pJL48 was used as a hybridization probe to screen an Arabidopsis (Col-O eco- type) genomic library (Clontech, Palo Alto, CA) in lEMBL3 by plaque

458 The Plant Cell
hybridization. A total of 51 positive clones were obtained from the screen- ing of 250,000 pfaques. The representative PAI7, PAI2, and PAI3 genomic DNAs shown in Figure 2 were subcloned into pBluescript II KS+ or SK+. For sequencing purposes, two partia1 PAI7 fragments were fur- ther subcloned in both orientations into pBluescript II SK+: a 2.0-kb Sacl fragment in pJL60 and pJL61, and 2.2 kb of EcoRV in pJL62 and pJL63. For sequencing PAI2 and PAI3, a 3.6-kb Xbal fragment and a 5.1-kb Sall-EcoRI fragment were further subcloned in both orientations into pBluescript I I KS+ or SK+ to create pJL118, pJL120, pJL193, and pJL194 (Table 1).
DNA and RNA Manipulations
Bacteriophage X DNA was prepared according to the method described by Santos (1991) or the Magic Lambda Prep DNA purification system (Promega). Plant DNA and total RNA were extracted from the 2- to 4-week-old rosettes as previously described (Cone, 1989; Nagy et al., 1989). DNA and RNA manipulations and analyses were performed using published protocols (Ausubel et al., 1993). DNA probes longer than 300 bp were radioactively labeled using the random hexamer label- ing method (Feinberg and Vogelstein, 1983). Genomic DNA gel blots were hybridized at 65OC for 18 hr in Church buffer (Church and Gilbert, 1984) and washed twice at 65% for 15 min with 0.1 x SSC (1 x SSC is 0.15 M NaCI, 0.015 M sodium citrate), 0.1% SDS.
The oligonucleotides JL1 (5’-GGATCTTGTTCTTGAGAGCCCAAC- GACATTATTACGGTG-3’) and JL2 (5’-GGGCCAAAAGCAGCTGCCTCT- CGCTCGTCTmGTGGACTAGG-31 were used as gene-specific hybrid- ization probes. JL1 is complementary to bases 1670 to 1708 of the PAI7 and PAI2 sequences, and JL2 is complementary to bases 1684 to 1731 of PA/3 (see Figure 3). The oligonucleotides were 3’end labeled with terminal deoxynucleotidyl transferase, and hybridizations using oligo- nucleotide probes were performed for 16 to 24 hr at 45% as described by Pruitt and Last (1993). The filter hybridized with oligonucleotide JL1 was washed twice at 45OC for 10 min each in 6 x SSC, 0.1% SDS, followed by two 15-min washes at 45OC in 6 x SSC, 0.1% SDS. The filter hybridized with oligonucleotide JL2 was washed in the same way as JL1 except at 65OC. Gene-specific hybridization was confirmed using in vitro-transcribed sense RNAs from PAI2 (pJL167) and PAI3 (pJL257) as templates.
Genomic DNA and cDNA sequencing was performed using the mul- tiwell Microtitre Plate DNA sequencing system (Amersham, Arlington Heights, IL) on overlapping unidirectional deletion series prepared as described by Rose et al. (1992). The GenBank and EMBL protein data bases were searched using the FASTA algorithm (Pearson and Lipman, 1988). Multiple sequence alignment of DNAor protein was performed using the PILEUP program of the University of Wisconsin, Madison, Genetics Computer Group package, and the pairwise identity was de- termined by the Clustal software package (Higgins et al., 1992).
PAI Mapping
The PAI7 gene was mapped using 55 recombinant inbred lines gener- ated from the cross between the Col-O and Landsberg erecta (Ler) ecotypes of Arabidopsis (Lister and Dean, 1993). A genomic DNA fragment of 1347 bp containing a Fokl restriction fragment length poly- morphism (RFLP) was amplified by polymerase chain reaction as described previously (Konieczny and Ausubel, 1993) using the primers JL3 (5’-GTAGAGGATTGAGCTTAAGGCAAG-3? and JL2381 (5‘-AGC- ACGCGAACCATTACCATGAAG-3’) at an annealing temperature of 56%. JL3 is a fAl7-specific sequence (-396 to -373), and JL2381 is complementary to bases 948 to 971 of the PAI7 or ?AI2 sequence
(see Figure 3). Fokl digestion of amplified products (1347 bp) from the Ler or COLO parenta1 ecotypes yielded 979- or 882-bp products, respec- tively. The linkage of PAI7 and the RFLP markers was determined using Mapmaker (Lander et al., 1987), using data obtained from ‘An Arabidop- sis thaliana Database” (AAtDB). The map positions indicated in Figure 6 are from the integrated geneticIRFLP map (Hauge et al., 1993). The relative positions of PAI1 and M488 are reversed from the order indi- cated in Figure 6 on the latest Lister and Dean recombinant inbred line map.
Three yeast artificial chromosome (YAC) libraries (Ecker, 1990; Ward and Jen, 1990; Grill and Somerville, 1991) were screened with gene- specific single-copy hybridization probes located adjacent to the PAI structural genes: the 0.35-kb Xhol fragment containing the directly flank- ing repeated sequence (Figure 4) from pJL94 for PAI7, the 1.0- or 6.5-kb Sall-Xbal fragments from pJL58 for PAI2, and a 0.55-kb Clal fragment from pJL178 for PAl3. Yeast colony filters were prepared as outlined by Coulson et al. (1988) with the modifications described in Schmidt et al. (1992). Probe labeling, hybridization, and washing conditions were performed according to Schmidt et al. (1992). Genomic DNA of yeast colonies was prepared as described by Schmidt et al. (1994). The analysis of YAC clones containing PAI gene sequences was per- formed using DNA gel blots of yeast genomic DNA digested with EcoRl and BamHI. lntact chromosomal DNA of YAC clones was prepared and subsequently separated by pulsed-field gel electrophoresis to ana- lyze the size of YACs (Schmidt et al., 1994). The PAl7 probe detected 12 YAC clones (EG8A12, EG8Bl1, EG22C6, EWllF9, EW13E7, EW18C3, EW20B12, yUP5H2, yUP9D5, yUPlOB6, yUPlOD1, and yUP22C7); PAI2- and PAIS-specific sequences were identified in six (EW5A4, EW7E12, EW8B9, EW9F12, EWl6C1, and yUP3El) and five (EG2F12, EG89F12, EW9E7, yUP6A5, and yUP24B3) YAC clones, respectively.
Antibody Production, lmmunoblot Analysis, and Enzyme Assays
Production of rabbit anti-PAI2 antisera, protein gel electrophoresis, im- munoblotting, and Phosphorlmager (Molecular Dynamics, Sunnyvale, CA) radioactivity quantification were performed as described by Zhao and Last (1995). The activities of PAI and PAT enzymes were assayed according to published methods (Last and Fink, 1988). Protein con- centration was measured with a protein dye binding assay kit (Bio-Rad), using BSA as the standard.
Analysis and ldentification of Anthranilate P-Glucoside by HPLC
Rosette-stage leaf tissue (100 mg) was ground to a fine suspension in 200 pL of 100% (v/v) methanol in asmall ground-glass homogenizer. The extracts were clarified by centrifugation at 14,0009 for 3 min. The pellet was reextracted with 200 pL of distilled water and centrifuged as given previously. Fifty microliters of the combined supernatant was subjected to C18 reverse phase chromatography by a slight modification of a published procedure (Li et al., 1993). A linear gradient of acetoni- trile ran from 2 to 7% for 5 min and was constant at 7% for 15 min. A linear gradient of 7 to 22% was then applied for 25 min and was followed by a linear gradient of 22 to 100% for 10 min. Elution was monitored at 330 nm. For identification of the anthranilate P-glucoside, 25 g of rosette leaf tissue from trp7-700 or JL197-5 plants was extracted with 50 mL of 100% methanol and reextracted with 50 mL of distilled water. The supernatants were concentrated to 0.5 to 1.0 mL under re- duced pressure. The concentrated crude extract was clarified at 14,0009 for 3 min, and chromatography was performed as described previously.

Triplicate Arabidopsis Tryptophan Genes 459
The fractions with the peak at retention time of 10.5 min were collected, dried under reduced pressure, and dissolved in 400 pL of distilled wa- ter. This purified sample (20 pL) was digested with 5 units of almond B-glucosidase (Sigma) at 37OC for 2 hr in 200 pL of 50 mM acetate, pH 5.0. After digestion, a 50-vL sample was analyzed by HPLC with or without the authentic anthranilate standard (Sigma).
Construction of Arabidopsis PAI Antisense Transgenic Plants
An antisense construct was produced that expresses nearly the en- tire PAl7 cDNA coding sequence from the cauliflower mosaic virus 35s promoter. A 0.8-kb PAl7 cDNA fragment from pJL169 (corresponding to the exonic sequences from -83 to +I475 in Figure 3) was created by complete EcoRl digestion, followed by partia1 digestion with Pstl, and this fragment was rendered blunt ended with the Klenow frag- ment of DNA polymerase I. To express this sequence in the antisense orientation, it was cloned behind the 35s promoter present on the pB1121 (Clontech)-derived plasmid pAR140 (pAR140 contains the appropriate EcoRI-Hindlll fragment from pB1121 cloned into EcoRIIHindlll-cleaved pBluescript II KS+). pAR140 was digested with Smal, and the blunt- ended PAl7 fragment was ligated to create pJL189. The 1.8-kb Hindlll- Sacl fragment from pJL189 that contains the 35s-PAI1 cDNA fusion construct was then recloned into the larger Hindlll-Sacl fragment of pB1121 to construct the binary transformation plasmid pJL197, which was introduced into Agrobacterium fumefaciens LBA4404 by electroporation.
Arabidopsis root transformation was performed according to the methods described previously (Valvekens et al., 1988; Rose et al., 1992), with the following modifications. Roots were obtained by germination and 10-day growth of seedlings in a 500-mL Ehrlenmeyer flask con- taining 100 mL of half-strength liquid Murashige-Skoog salts (Sigma), pH 5.8, and 1% sucrose with agitation at 100 to 150 rpm under con- tinuous rmm illumination (10 pmol m-2 sec-l). Kanamycin was omitted from root-inducing medium plates to facilitate root induction, and PNS medium replaced glucose medium for enhanced root growth prior to transfer to soil-less mix. The kanamycin resistance of the transgenic plant progeny was tested on PNS medium containing 50 pg/mL kanamy- cin. All sterile media were supplemented with 50 pM L-tryptophan.
ACKNOWLEDGMENTS
We thank John Mulligan for the LYES Arabidopsis cDNA library and IKC lysogen, Charles Yanofsky for the trpC E. coli strain, and Judy Bender and Gerald Fink for sharing unpublished mapping data. This research was supported by grants from the Biotechnology Research Development Corporation (Peoria, IL) and the National lnstitutes of Health (Grant No. GM43134) and by a National Science Foundation Presidential Young lnvestigator Award (No. DMB-9058134) to R.L.L.
Received for January 13, 1995; accepted February 10, 1995.
REFERENCES
Adams, R.R., and Royer, T. (1990). Complete genomic sequence en- coding trpC from Aspergillus niger var. awamori. Nucl. Acids Res. 18, 4931.
Ausubel, F.M., Brent, R., Kingston, R.E., Moore, D.D., Seidman, J.G., Smith, J.A., and Struhl, K., eds (1993). Current Protocols in Molecular Biology. (New York: Wiley).
Barczak, A.B., Zhao, J., Pruitt, K.D., and Last, R.L. (1995). BFluoroindole resistance identifies tryptophan synthase fl subunit mutants in Arabidopsis thaliana. Genetics 139, in press.
Berlyn, M.B., Last, R.L., and Fink, G.R. (1989). A gene encoding the tryptophan synthase p subunit of Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 86, 4604-4608.
Choi, H.T., Revuelta, J.L., Sadhu, C., and Jayaram, M. (1988). Struc- tural organization of the TRP7 gene of Phycomyces blakesleeanus: lmplications for evolutionary gene fusion in fungi. Gene 71,85-95.
Christie, G.E., and Platt, T. (1980). Gene structure in the tryptophan operon of €scherichia coli. J. MOI. Biol. 142, 519-530.
Church, G.M., and Gilbert, W. (1984). Genomic sequencing. Proc. Natl. Acad. Sci. USA 81, 1991-1995.
Cone, K. (1989). Yet another plant DNA prep. Maize Genet. Newslett. 63, 68.
Cordell, G.A. (1974). The biosynthesis of indole alkaloids. Lloydia 37,
Coulson, A., Waterston, R., Kiff, J., Sulston, J., and Kohara, Y. (1988). Genome linking with yeast artificial chromosomes. Nature 335, 184-186.
Crawford, I.P., Han, C.Y., and Silverman, M. (1991). Sequence and features of the tryptophan operon of Vibfioparahmolyticus. DNA Seq.
DeRocher, E.J., Quigley, F., Macke, R., and Bohnert, H.J. (1993). The six genes of the Rubisco small subunit multigene family from Mesembryanfhemum cptallinum, a facultative CAM plant. MOI. Gen. Genet. 239, 450-462.
Doy, C.H., Rivera, A., and Srlnlvasan, P.R. (1961). Evidence for the enzymatic synthesis of K(5’-phosphoribosyl)anthranilic acid, a new intermediate in tryptophan biosynthesis. Biochem. Biophys. Res. Commun. 4, 83-88.
Ecker, J.R. (1990). PFGE and YAC analysis of the Arabidopsis genome. Methods 1, 186-194.
Elledge, S.J., Mulligan, J.T., Ramer, S.W., Spottswood, M., and Davis, R.W. (1991). LYYES: A multifunctional cDNA expression vec- tor for the isolation of genes by complementation of yeast and Escherichia coli mutations. Proc. Natl. Acad. Sci. USA 88,1731-1735.
Facchini, P.J., and Chappell, J. (1992). Gene family for an elicitor- induced sesquiterpene cyclase in tobacco. Proc. Natl. Acad. Sci. USA 89, 11088-11092.
Feinberg, AR, and Vogelstein, B. (1983). A technique for radiolabeling DNA endonuclease restriction fragments to high specific activity. Ana1 Eiochem. 132, 6-13.
Grill, E., and Somerville, C. (1991). Construction and characteriza- tion of a yeast artificial chromosome library of Arabidopsis which is suitable for chromosome walking. MOI. Gen. Genet. 226,484-490.
Hauge, B.M., Hanley, S.M., Cartinhour, S., Cherry, J.M., Goodman, H.M., Koornneef, M., Stam, P., Chang, C., Kempin, S., Medrano, L., and Meyerowitz, E.M. (1993). An integrated geneticlRFLP map of the Arabidopsis thaliana genome. Plant J. 3, 745-754.
Haughn, G.W., and Somerville, C. (1986). Sulfonylurea-resistant mu- tants of Arabidopsis thaliana. MOI. Gen. Genet. 204, 430-434.
Haughn, G.W., Davin, L., Giblin, M., and Underhill, E.W. (1991). Bio- chemical genetics of plant secondary metabolites in Arabidopsis thaliana: The glucosinolates. Plant Physiol. 97, 217-226.
Henikoff, S., Tatchell, K., Hall, B.D., and Nasmyth, K.A. (1981).
219-298.
1, 189-196,

460 The Plant Cell
lsolation of a gene from Drosophila by complementation in yeast. Nature 289, 33-37.
Higgins, D.G., Bleasby, A.J., and Fuchs, R. (1992). Clustal V Im- proved software for multiple sequence alignment. Comput. Appl. Biosci. 8, 189-191.
Horowitz, H., Van Andell, J., and Platt, T. (1983). Nucleotide se- quence of the trpD and trpC genes of Salmonella typhimurium. J. MOI. Biol. 169, 775-797.
Hiitter, R., Niederberger, P., and DeMoss, J.A. (1986). Tryptophan biosynthetic genes in eukaryotic microorganisms. Annu. Rev. Microbiol. 40, 55-77.
Jarvis, P., Lister, C., Szabo, V., and Dean, C. (1994). lntegration of CAPs markers into the RFLP map generated using recombinant in- bred lines of Arabidopsis thaliana. Plant MOI. Biol. 24, 685-687.
Keith, B., Dong, X., Ausubel, F.M., and Fink, G.R. (1991). Differen- tia1 induction of 3-deoxy-~-arabino-heptulosonate Fphosphate synthase genes in Arabidopsis thaliana by wounding and pathogenic attack. Proc. Natl. Acad. Sci. USA 88, 8821-8825.
Klee, H.J., Muskopf, Y.M., and Gasser, C.S. (1987). Cloning of an Arabidopsis thaliana gene encoding 5-enolpyruvylshikimate-3- phosphate synthase: Sequence analysis and manipulation to ob- tain glyphosate-tolerant plants. MOI. Gen. Genet. 210, 437-442.
Konieczny, A., and Ausubel, F.M. (1993). A procedure for mapping Arabidopsis mutations using co-dominant ecotype-specific PCR- based markers. Plant J. 4, 403-410.
Kos, T., Kuijvenhoven, A., Hessing, H.G.M., Pouwels, P.H., and van den Hondel, C.A.M.J.J. (1988). Nucleotide sequence of the Asper- gillus niger trpC gene: Structural relationship with analogous genes of other organisms. Curr. Genet. 13, 137-144.
Lander, E.S., Green, P., Abrahamson, J., Barlow, A., Daly, M.J., Lincoln, S.E., and Newberg, L. (1987). Mapmaker: An interactive computer package for constructing primary genetic linkage maps of experimental and natural populations. Genomics 1, 174-181.
Last, R.L. (1993). The genetics of nitrogen assimilation and amino acid biosynthesis in flowering plants: Progress and prospects. In lnternational Review of Cytology, Vol. 143, K.W. Jeon, M. Friedlander, and J. Jarvik, eds (San Diego: Academic Press), pp. 297-330.
Last, R.L., and Fink, G.R. (1988). Tryptophan-requiring mutants of the plant Arabidopsis thaliana. Science 240, 305-310.
Last, R.L., Bissinger, P.H., Mahoney, D.J., Radwanski, E.R., and Fink, G.R. (1991). Tryptophan mutants in Arabidopsis: The conse- quences of duplicated tryptophan synthase p genes. Plant Cell 3, 345-358.
Leutwiler L.S., Meyerowitz, E.M., and Tobin, E.M. (1986). Structure and expression of three light-harvesting chlorophyll a/b-binding pro- tein genes in Arabidopsis thaliana. Nucl. Acids Res. 14,4051-4064.
Li, J., Ou-Lee, T.-M., Raba, R., Amundson, R.G., and Last, R.L. (1993). Arabidopsis flavonoid mutants are hypersensitive to UV-B irradiation. Plant Cell 5, 171-179.
Li, J., Chen, S., Zhu, L., and Last, R.L. (1995). lsolation of cDNAs encoding the tryptophan pathway enzyme indole-3-glycerol phos- phate synthase from Arabidopsis thaliana. Plant Physiol., in press.
Lister, C., and Dean, C. (1993). Recombinant inbred lines for map- ping RFLP and phenotypic markers in Arabidopsis thaliana. Plant
McGrath, J.M., Jansco, M.M., and Pichersky, E. (1993). Duplicate sequences with a similarity to expressed genes in the genome of Arabidopsis thaliana. Theor. Appl. Genet. 86, 880-888.
J. 4, 745-750.
Meagher, R.B., Berry-Lowe, S., and Rice, K. (1989). Molecular evo- lution of the small subunit of ribulose bisphosphate carboxylase: Nucleotide substitution and gene conversion. Genetics 123,845-863.
Nagy, F., Kay, S.A., and Chua, N.-H. (1989). Analysis of gene expres- sion in transgenic plants. In Plant Molecular Biology Manual, S.B. Gelvin and R.A. Schilperoort, eds (Dordrecht, The Netherlands: Kluwer), pp. B4:l-29.
Niyogi, K.K., and Fink, G.R. (1992). Two anthranilate synthase genes in Arabidopsis: Defense-related regulation of the tryptophan path- way. Plant Cell 4, 721-733.
Niyogi, K.K., Last, R.L., Fink, G.R., and Keith, B. (1993). Suppres- sors of trpl fluorescence identify a new Arabiodpsis gene, TRP4, encoding the anthranilate synthase p subunit. Plant Cell5,1011-1027.
Normanly, J., Cohen, J.D., and Fink, G.R. (1993). Arabidopsis thaliana auxotrophs reveal a tryptophan-independent biosynthetic pathway for indole-Sacetic acid. Proc. Natl. Acad. Sci. USA 90,10355-10359.
Peanon, W.R., and Lipman, D.J. (1988). lmproved tools for biologi- cal sequence comparison. Proc. Natl. Acad. Sci. USA 85,2444-2448.
Pruitt, K.P., and Last, R.L. (1993). Expression patterns of duplicate tryptophan synthase p genes in Arabidopsis thaliana. Plant Phys- iol. 102, 1019-1026.
Rose, A.B., and Last, R.L. (1994). Molecular genetics of amino acid, nucledide and vitamin biosynthesis. In Arabidopsis, E.M. Meyerowitz and C.R. Somerville, eds(Cold Spring Harbor, NY Cold Spring Har- bor Press), pp. 835-879.
Rose, A.B., Casselman, A.L., and Last, R.L. (1992). A phosphoribo- sylanthranilate transferase gene is defective in blue fluorescent Arabidopsis thaliana tryptophan mutants. Plant Physiol. 100,582-592.
SBnchz, F., Tourino, A., Traseira, S., Perez-Aranda, A., Rubio, V., and Penalva, M.A. (1986). Molecular cloning and characterization of the trpC gene from fenicillium chrysogenum. MOI. Gen. Genet.
Santos, M.A. (1991). An improved method for the small scale prepa- ration of bacteriophage DNA based on phage precipitation by zinc chloride. Nucl. Acids Res. 19, 5442.
Schechtman, M.G., and Yanofsky, C. (1983). Structure of the trifunc- tional trpl gene from Neurospora crassa and its aberrant expression in Escherichia coli. J. MOI. Appl. Genet. 2, 83-99.
Schmidt, R., Cnops, G., Bancroft, I., and Dean, C. (1992). Construc- tion of an overlapping YAC library of the Arabidopsis thaliana genome. Aust. J. Plant Physiol. 19, 341-351.
Schmidt, R., Putterill, J., West, J., Cnops, G., Robson, F., Coupland, G., and Dean, C. (1994). Analysis of clones carrying repeated DNA sequences in two YAC libraries of Arabidopsis thaliana. Plant J. 5, 735-744.
Senecoff, J.F., and Meagher, R.B. (1993). lsolating the Arabidopsis thaliana genes for de novo purine synthesis by complementation of Escherichia coli mutants, I: 5'-PhosphoribosyC5-aminoimidazole synthetase. Plant Physiol. 102, 387-399.
Thomas, G.J. (1984). Herbicidal activity of 6-methylanthranilic acid and analogs. J. Agric. Food Chem. 32, 747-749.
Tschumper, G., and Carbon, J. (1980). Sequence of a yeast DNA frag- ment containing a chromosomal replicator and the TRP7 gene. Gene
Tsuji, J., Zook, M., Somerville, S.C., Last, R.L., and Hammerschmidt, R. (1993). Evidence that tryptophan is not a direct biosynthetic in- termediate of camalexin in Arabidopsis thaliana. Physiol. MOI. Plant Pathol. 43, 221-229.
205, 248-252.
10, 157-166.

Jriplicate Arabidopsis Tryptophan Genes 461
Valvekens, D., Van Montagu, M., and Van Lijsebettens, M. (1988). Agrobacterium tumefaciens-mediated transformation of Arabidop- sis thaliana root explants by using kanamycin selection. Proc. Natl. Acad. Sci. USA 85, 5536-5540.
Vogel, H.J., and Bonner, D.M. (1956). Acetylornithinase of fscher- chia colipartial purification and some properties. J. Biol. Chem. 218,
von Heijne, G., Hirai, T., Klosgen, R.-B., Steppuhn, J., Bruce, E., Keegstra, K., and Herrmann, R. (1991). CHLPEP-A database of chloroplast transit peptides. Plant MOI. Biol. Rep. 9, 104-126.
Ward, E.R., and Jen, G.C. (1990). lsolation of single-copy-sequence clones from a yeast artificial chromosome library of randomly-sheared Arabidopsis thaliana DNA. Plant MOI. Biol. 14, 561-568.
97-106.
Wright, A.D., Sampson, M.B., Neuffer, M.G., Michalcruk, L., Slovin, J.P., and Cohen, J.D. (1991). Indole-3-acetic acid biosynthesis in the mutant maize orangepericarp, a tryptophan auxotroph. Science 254, 998-1000,
Yanofsky, C., Horn, V., Bonner, M., and Staslowski, S. (1971). Polar- ity and enzyme functions in mutants of the first three genes of the tryptophan operon of fscherichia coli. Genetics 69, 409-433.
Zhao, J., and Last, R.L. (1995). lmmunological characterization and chloroplast localization of the tryptophan biosynthetic enzymes of the flowering plant Arabidopsis thaliana. J. Biol. Chem. 270, 6081-6087.