biotransformations leading to toxic metabolites: …€¦ · 676 chapter 33 biotransformations...
TRANSCRIPT

674Copyright © 2008, Elsevier Ltd
All rights reserved.Wermuth’s The Practice of Medicinal Chemistry
Chapter 33
I . HISTORICAL BACKGROUND
As drugs are usually foreign chemicals, history of concern for the biotransformations of drugs leading to toxic metabolites formation is intrinsically linked to the history of xenobiotic metabolism studies. The International Society for the Study of Xenobiotics (ISSX) website ( http://www.issx.org ) presents an overview of the fi eld history where some key fi gures may be pointed out.
One is probably Richard Tecwyn Williams who intro-duced the Phase I and II classifi cation of xenobiotics metab-olism reactions. Although his emblematic book 1 was called “ Detoxication mechanisms, ” he estimated that, in some cases, metabolism may increase toxicity. He also considered that this “ bioactivation ” may occur during the Phase II reac-tions (usually considered as detoxication reactions), and not only that of Phase I (functionalization reactions).
Quite at the same time, Bernard Brodie studied the antimalarial atabrine (quinacrine) metabolism in order to
avoid the toxic side effects of the drug. He also developed some new analytical methodologies, necessary for meta-bolic studies. Then he put together a group of researchers (including Julius Axelrod, James Gillette and many others) in this fi eld, and they published many studies of great impor-tance related to drug metabolism, most famous probably concerning acetaminophen. Among these works, these scientists developed the covalent binding theory concept, which provides an explanation for the toxic side effects of drugs. Following the work of James and Elizabeth Miller on covalent binding of polycyclic aromatic hydrocarbon electrophilic metabolites on DNA in the 1940s, Brodie et al. suggested that in vivo bioactivation may lead to the formation of electrophilic entities, which are capable of linking with biological macromolecules, thus inducing dis-turbances in cellular functions.
The discovery of mixed function oxidases during the 1950s and the characterization of cytochrome P450 by Omura and Sato 2 were a “ revolution ” in the fi eld of
I. HISTORICAL BACKGROUND II. INTRODUCTION III. REACTIONS INVOLVED
IN THE BIOACTIVATION PROCESS
A. Oxidation B. Oxidative stress C. Reduction D. Substitutions: hydrolysis and
conjugation
E. Eliminations F. Further biotransformations
leading to the ultimate toxicant
IV. EXAMPLES OF METABOLIC CONVERSIONS LEADING TO TOXIC METABOLITES
A. Acetaminophen B. Tienilic acid
C. Halothane D. Valproic acid E. Troglitazone V. CONCLUSION REFERENCES
“ La matière demeure et la forme se perd. ” “ The matter remains and the form is lost. ”
Pierre de Ronsard
Biotransformations Leading to Toxic Metabolites: Chemical Aspect Anne-Christine Macherey and Patrick M. Dansette
Ch33-P374194.indd 674Ch33-P374194.indd 674 5/30/2008 6:35:15 PM5/30/2008 6:35:15 PM

675
xenobiotic metabolism. Remmer discovered that cyto-chrome P450 may be induced by phenobarbital, and Conney characterized the induction with 3-methyl cholanthrene. These works 3 were of great importance for mechanistic studies of drug metabolism. A new step was done in 1999 with the fi rst crystallization of a mammalian cytochrome P450 by Johnson, which provided new perspectives in safer drug design.
Induction of cytochrome P450 synthesis suggests that xenobiotics may exert an effect on the genome: the use of genomics and proteomics represents a new challenge for predictive toxicology in drug design.
II . INTRODUCTION
Toxicity is the result of the more or less harmful action of chemicals on a living organism. Toxicology, the study of toxicity, is situated at the border of chemistry, biology, and in some cases, physics. Molecular toxicology tries to eluci-date the mechanisms by which chemicals exert their toxic effects. Because many foreign chemicals enter the body in inert but unexcretable forms, biotransformations are an important aspect of the fate of xenobiotics. 4,5 In the case of drugs, metabolic conversions may be required for thera-peutic effect ( “ prodrugs ” ; see Chapter 36 for a detailed dis-cussion of prodrugs). In other cases, metabolism results in a loss of the biological activity. Sometimes, biotransforma-tions produce toxic metabolites. The last process is called toxifi cation or bioactivation. It should be emphasized that the general principles of pharmacology embrace the occur-rence of toxic events: although biotransformation processes are often referred to as detoxifi cation, the metabolic products are, in a number of cases, more toxic than the parent com-pounds. For drugs, whether biotransformations lead to the formation of toxic metabolites or to variations in therapeu-tic effects depends on intrinsic (such as the genetic poly-morphism of some metabolism pathways) and extrinsic (such as the dose, the route or the duration) factors. The biochemical conversions are usually of an enzymatic nature and yield reactive intermediates, which may be implicated in the toxicity as far as the fi nal metabolites. The primary events, which constitute the beginning of the toxic effect may result, after metabolism, from an inhibition of a spe-cifi c (and in most cases enzymatic) cellular function, an alkylating attack or an oxidative stress.
With regard to the toxicity arising from metabolites ( “ indirect toxicity ” ), three cases may be distinguished ( Figure 33.1 ):
A. Biotransformation begins with the transient formation of a reactive intermediate, whose lifetime is long enough to allow an attack on cellular components. This occurs when a reactive intermediate (generally radicals or electrophiles such as a carbonium ion) is formed and reacts rapidly with cellular macromolecules (such as unsaturated lipids,
proteins, nucleic acids…), thus leading to their degrada-tion and fi nally to cellular necrosis.
B. The fi rst step of the metabolic process yields a primary metabolite, which can, in some cases, accumulate in the cell and react with cellular components before being transformed.
C. The fi nal metabolites, when in excess, may accumulate and react with cellular macromolecules.
Usually, metabolic conversions are divided into two major types of reactions (see chapter 32 for a detailed dis-cussion of metabolic biotransformations). Phase I reactions, or functionalization reactions, involve the introduction of a polar functionality such as a hydroxyl group into the xeno-biotic structure. During Phase II reactions, this group is subsequently coupled (or conjugated) with an endogenous cofactor, which contains a functional group that is usually ionized at physiological pH. This ionic functional group facilitates active excretion into the urinary and/or hepato-biliary system. The elimination by transport mechanism is sometimes also called “ Phase III. ”
Because bioactivation is mainly an activation of xeno-biotics to electrophilic forms, which are entities capable of reacting irreversibly with tissue nucleophiles, biotrans-formations leading to toxic metabolites are in most cases Phase I reactions. But Phase II reactions may also give rise to toxic phenomena, for example, when conjugation pro-duces a toxic metabolite, or when it is responsible for a specifi c target organ toxicity by acting as a delivery form to particular sites in the body where it is hydrolyzed and exerts a localized effect. Also, the fi nal toxic metabolite may be formed by combinations of several Phase I and Phase II reactions. Because of the increasing understanding of drug metabolizing enzymes, some authors 6 claim that Williams “ Phases I and II ” classifi cation is now inaccurate and even misleading. Pointing out the fact that Williams only introduced the classifi cation at the end of his book and did not use it in his monograph, they consider it would be now wiser to avoid using any special category.
FIGURE 33.1 Indirect toxicity.
II. Introduction
Ch33-P374194.indd 675Ch33-P374194.indd 675 5/30/2008 6:35:15 PM5/30/2008 6:35:15 PM

CHAPTER 33 Biotransformations Leading to Toxic Metabolites: Chemical Aspect676
III . REACTIONS INVOLVED IN THE BIOACTIVATION PROCESSES
During the biotransformations affecting xenobiotics, fi ve major kinds of chemical reactions may occur: oxidations (by far the most important), reductions, hydrolysis, substi-tutions, and eliminations. As Phase I and II reactions are part of this classifi cation, each class of reactions can give rise to toxic metabolites.
A . Oxidation
Several enzymatic systems are involved during the oxida-tive transformations of xenobiotics. Whether substances act upon one enzyme rather than another depends not only on its specifi c function, but also on the electromolecular environment. The most important is the microsomal drug metabolizing system known as cytochrome P450 (CYP) monooxygenase, which is localized mainly in the liver and is involved in most biological oxidations of xenobiot-ics. 7–9 Those include C- , N- and S- oxidations, N- , O- and S- dealkylation, deaminations, and certain dehalogenations. Under anaerobic conditions, it can also catalyze reductive reactions. The CYP monooxygenase system is a multien-zymatic complex constituted by the CYP hemoprotein, the fl avoprotein enzyme NADPH CYP reductase, and the unsaturated phospholipid phosphatidylcholine. The iso-forms involved in xenobiotic metabolism are membrane bound enzymes situated in the endoplasmic reticulum. After
cell lysis for in vitro studies, they are found in the micro-somal fraction. There are numerous isoforms (more than 6,000 known in all species). Thus a nomenclature based on their sequence similarity has been designed, and they are classifi ed in families and subfamilies: for instance CYP3A4 is the major human CYP, CYP is for cytochrome P450, 3 for the number of the family (more than 40% sequence identity), A for the letter of the subfamily (more than 55% sequence identity) and 4 the number in the subfamily. The human genome shows 57 complete CYP sequences plus a number of pseudo-genes. The CYPs involved in xenobiotic metabolism 9 (about 15) belong to families 1 to 4. The cata-lytic mechanism of CYP involves a formal (FeO) 3 � com-plex formed by the elimination of H 2 O from the iron site after two electrons have been added ( Figure 33.2 ).
Another oxidative enzyme is the FAD-containing monooxygenase, which is capable of oxidizing nucleophilic nitrogen, sulfur, and organophosphorus compounds. The fl avoprotein binds NADPH, oxygen and then the substrate. The oxidized metabolite is released, followed by NADP. Alcohol dehydrogenase and aldehyde dehydrogenase cata-lyze the oxidation of a variety of alcohols and aldehydes into aldehydes and acids, respectively, in the liver. Xanthine oxidase oxidizes several purine derivatives such as theo-phylline. Monoamine oxidase (MAO) and diamine oxidase convert amines into alkyl or aryl aldehydes by oxidation of the amine to an imine followed by subsequent hydrolysis. Peroxidases are oxidative enzymes, which couple the reduc-tion of hydrogen peroxide and lipid hydroperoxides to the oxidation of other substrates. This co-oxidation is responsi-ble for the production of reactive electrophiles from aromatic amines (e.g. the highly carcinogenic benzidine), phenols, hydroquinones, polycyclic aromatic hydrocarbons, etc.
The oxidation reactions can be described in terms of a rather common chemistry that involves the abstraction of either a hydrogen atom or a nonbonded (or π ) electron by the iron-oxo porphyrin complex ( Figure 33.3 ). The high-valent complex electronic confi guration is unknown, but is usually written as Fe V —— O.
The one-electron oxidation yields transient radicals ( Figure 33.4 ), which are transformed into more stable forms.
These radicals can incorporate an oxygen atom by abstraction of a hydroxyl group from the CYP iron-oxo spe-cies. This yields an oxidized derivative that may be some-times more toxic than the parent compound or susceptible to further metabolic conversions. Free radicals may also
FIGURE 33.2 Catalytic cycle of cytochrome P450 (CYP) monooxygenase.
FIGURE 33.3 CYP oxidation process.
Ch33-P374194.indd 676Ch33-P374194.indd 676 5/30/2008 6:35:16 PM5/30/2008 6:35:16 PM

677
bind to the site of their formation, thus leading to inhibition or inactivation of the enzyme. When the radical is not effi -ciently controlled by the iron, it may leave the active site. The subsequent released radical is able to produce damage to unsaturated fatty acids, thus leading to lipid peroxidation and destruction of the cellular structure. Another mode of the radical stabilization is a second one-electron oxidation, which consists of the loss of another electron. The fate of free radicals is now extensively studied because of their great capacities for forming covalent bonds with cellular macromolecules. 10–12
1 . C -H bond oxidations
These oxidations, which are usually catalyzed by CYP monooxygenases, produce hydroxylated derivatives. 13 When the C -H bond is located in the α position to a heteroatom (such as O, S, N, halogen), the α hydroxylated derivative obtained is usually unstable and may be further oxidized or cleaved ( Figure 33.5 ).
The antibiotic chloramphenicol is oxidized by CYP monooxygenase to chloramphenicol oxamyl chloride formed by the oxidation of the dichloromethyl moiety of chloramphenicol followed by elimination of hydrochloric acid 14 ( Figure 33.6 ). The reactive metabolite reacts with the � -amino group of a lysine residue in CYP 15 and inhibits the enzymatic reaction progressively with time. This type of inhibition is a time-dependent inhibition or a mechanism-based inhibition or inactivation, and the substrate involved historically has been called a suicide substrate because the enzymatic reaction yields a reactive metabolite, which destroys the enzyme. 16
In the case of chloroform, the unstable trichlorometha-nol loses hydrochloric acid and forms phosgene, which is very reactive ( Figure 33.7 ). 17
Tertiary amines containing at least one hydrogen on the α carbon may either be N- oxidized (leading to an N- oxide in the case of tertiary amines), or C- oxidized, thus leading to a carbinolamine. The latter, usually being unstable, splits into a secondary amine and an aldehyde moiety ( Figure 33.8 ). Several electron transfer mechanisms have been proposed. 7–9
During the oxidation of nitrosamines, the hydroxylated derivative formed cleaves spontaneously into highly reactive metabolites capable of alkylating nucleophilic sites in the cellular components.
FIGURE 33.4 One-electron oxidation.
FIGURE 33.5 C -H bond oxidation in the α -position to a heteroatom.
FIGURE 33.6 Metabolic activation of chloramphenicol.
FIGURE 33.7 Oxidation of chloroform.
III. Reactions Involved in the Bioactivation Process
Ch33-P374194.indd 677Ch33-P374194.indd 677 5/30/2008 6:35:18 PM5/30/2008 6:35:18 PM

CHAPTER 33 Biotransformations Leading to Toxic Metabolites: Chemical Aspect678
2 . Unsaturated bond oxidations
Double bonds are oxidized by CYP monooxygenases into epoxides, which are generally very reactive. Epoxides are considered responsible for the toxicity of the unsaturated compounds.
The hepatocarcinogenicity of afl atoxin B1 (AFB1) is known to be due to the epoxide (AFB1-oxides) formed, which binds directly with the N-7 atom of a guanine molecule in DNA ( Figure 33.9 ). 18
Aromatic chemicals are metabolized into unstable arene-oxides, which, as epoxides, are comparable to potentially equivalent electrophilic carbocations. These metabolites react easily with thiol groups derived from proteins, leading, for example, to hepatotoxicity. Bromobenzene seems to tar-get a large group of functionally diverse hepatic proteins, as demonstrated recently in a proteomic analysis. 19 The chem-ical is oxidized ( Figure 33.10 ) into a 3,4-epoxide, which
does not exhibit mutagenic or carcinogenic activity, but reacts nonenzymatically with liver proteins and produces hepatic necrosis. 20 The isomeric 2,3-epoxide rearranges very quickly to 2-bromophenol and is less toxic. A sec-ondary CYP-catalyzed oxidation to hydroquinone and benzoquinone also can occur. In this alternative pathway, conjugation with glutathione can lead to the formation of products, which may elicit their toxicity elsewhere than the liver and especially in the kidney. 20
3 . N -oxidations
Tertiary amines are transformed into N- oxides (generally less toxic), but primary and secondary amines are oxidized into hydroxylated derivatives (hydroxylamines). This oxi-dation is responsible for the hepatotoxicity and mutagenic-ity of acetamino-2-fl uorene ( Figure 33.11 ). 21
Nitrenium ions may occur during bioactivation of aro-matic amines and amides, which are usually N- oxidized into N- hydroxylated derivatives. By sulfation or esterifi ca-tion followed by elimination of the newly formed leaving group, the latter may be transformed into highly reactive nitrenium ions. In the case of aromatic nitrenium ions they are in equilibrium with their tautomeric aromatic carboca-tions, which react with cellular nucleophilic macromolecules (nucleic acids, etc.).
4 . Heteroatom oxidations
Heteroatoms such as nitrogen or sulfur are oxidized at their nonbonded peripheric electrons as described for thiophene ( Figure 33.12a ). 22 Thiophene is oxidized to thiophene sulfoxide, which is unstable and dimerizes spontaneously to thiophene S- oxide dimers through a Diels–Alder reac-tion. 23,24 They also react with nucleophiles like the thiol group of glutathione or proteins, giving glutathione or pro-tein adducts. In addition, thiophenes are oxidized to unsta-ble thiophene epoxides, which rearrange spontaneously to thiolenones as found recently for 2- and 3-phenylthiophenes ( Figure 33.12b ). In fact, there is a competition between S- oxidation (sulfoxide pathway) and double bond oxidation (epoxide pathway). In the presence of glutathione, adducts formed from both reactive intermediates have been found, in addition to thiophene S- oxide dimers and the thiolenones tautomers of hydroxythiophenes. 25,26
Halogenated aromatic compounds may also be oxidized by CYP monooxygenases, yielding hypervalent halogen-ated compounds.
B . Oxidative stress
Oxidative stress has been defi ned as a disturbance in the pro-oxidant–antioxidant balance in favor of the pro-oxidant state resulting from alterations in the redox state of the cell. The stepwise reduction of oxygen into superoxide anion,
FIGURE 33.8 Oxidation of a tertiary amine.
FIGURE 33.9 Oxidation of afl atoxin B 1 .
Ch33-P374194.indd 678Ch33-P374194.indd 678 5/30/2008 6:35:21 PM5/30/2008 6:35:21 PM

679
FIGURE 33.10 Metabolism of bromobenzene.
FIGURE 33.11 N -oxidation of acetamino-2-fl uorene.
III. Reactions Involved in the Bioactivation Process
Ch33-P374194.indd 679Ch33-P374194.indd 679 5/30/2008 6:35:23 PM5/30/2008 6:35:23 PM

CHAPTER 33 Biotransformations Leading to Toxic Metabolites: Chemical Aspect680
hydrogen peroxide, hydroxyl radical, and fi nally water, which accounts for about 5% of the normal oxygen reduc-tion (versus 95% by means of the mitochondrial electron transport chain), may be increased by the redox cycling of some xenobiotics such as quinones or nitro-aromatic derivatives. These compounds are susceptible to one-electron reduction, which yields radical structures that may be back-oxidized to the parent compound. During this reoxidation, oxygen is reduced into superoxide anion. The oxygen reduction products are highly reactive entities that attack all the cellular components, especially when their normal degradation systems (superoxide dismutase, glutathione peroxidase, catalase) are overburdened. The polyunsaturated lipids are especially sensitive to these attacks because they are susceptible to a membrane-degrading peroxidation.
C . Reduction
Reductive biotransformations of several compounds such as polyhalogenated, keto, nitro and azo derivatives, are
catalyzed by a variety of enzymes that differ according to the substrates and the species. The liver CYP-dependent drug metabolizing system is capable of reducing N- oxide, nitro and azo bonds, whereas the cytosolic nitrobenzene reductase activity is mainly due to CYP reductase, which transforms nitrobenzene into its hydroxylamino derivative. NADPH cytochrome c reductase is also able to catalyze the reduction of nitro compounds. These metabolic conversions may also be brought about by gastrointestinal anaerobic bacteria.
Reductive processes that occur during the metabolism of xenobiotics involve either one-electron reduction or a two-electron transfer.
Ionic reduction using a hydride occurs in vivo during the reduction catalyzed by NADH or NADPH enzymes, whereas one-electron reduction releases a radical structure, which may contribute to the toxic effect. Figure 33.13 illustrates the biotransformations affecting the anthracycline antitumor drug daunomycin. 27 Recent studies suggest that nitric oxide synthases may contribute to the cardiotoxicity, probably because of their structural similarities with CYP reductase. 28
FIGURE 33.12 (a) oxidation of thiophene, (b) oxidation of 2-phenylthiophene.
Ch33-P374194.indd 680Ch33-P374194.indd 680 5/30/2008 6:35:25 PM5/30/2008 6:35:25 PM

681
1 . Reduction of polyhalogenated compounds
Some polyhalogenated compounds, such as CCl4, BrCCl3 and halothane (CF3-CHBrCl), when in the presence of the reduced form of CYP, may undergo one-electron reduc-tion 29,30 ( Figure 33.14 ), which leads to a radical that may be transformed by different pathways.
The radical formed may add directly on the unsaturated lipid bonds or initiate an unsaturated lipid peroxidation or undergo another one-electron reduction. The last reaction yields a carbene that can form a complex with the iron of the reduced form of CYP. Reduction of polyhalogenated compounds gives rise to several reactive intermediates, such as radicals, carbenes and peroxides, whose participa-tion in the toxic effect varies greatly. 13
2 . Reduction of nitro compounds
The different steps of the biotransformations that produce a primary amine from an aromatic nitro compound involve a nitro radical-anion, a nitroso derivative, a nitroxyl radical, a hydroxylamine and then the primary amine ( Figure 33.15 ).
Each of these different intermediates may contribute to the toxicity. Hydroxylamines are often responsible for methemoglobinemia,31 whereas mutagenic and carcinogenic activity may be due to the combination of nitro radical-anion, nitroso derivatives or esterifi ed hydroxylamine (such as sulfate derivatives) with cellular macromolecules.
Carcinogenicity may also be the result of the oxidative stress subsequent to the formation of oxygen–reduction prod-ucts (superoxide anion, hydrogen peroxide, hydroxyl radical)
FIGURE 33.13 Biotransformations of daunorubicin.
III. Reactions Involved in the Bioactivation Process
Ch33-P374194.indd 681Ch33-P374194.indd 681 5/30/2008 6:35:27 PM5/30/2008 6:35:27 PM

CHAPTER 33 Biotransformations Leading to Toxic Metabolites: Chemical Aspect682
during redox cycling of the nitro radical-anion, which restores the parent nitro compound.
3 . Reduction of azo compounds
Azo compounds are susceptible to reduction, fi rst to hydrazo intermediates, which are reductively cleaved into the appro-priate amines. It has been proposed 32 that the fi rst step, as with nitro compounds, is the formation of an azo-anion radical.
D . Substitutions: hydrolysis and conjugation
Among substitution reactions, ester and amide hydroly-sis are common, and often operate during detoxifi cation processes. Both specifi c enzymatic and chemical hydroly-sis may occur. Acid-catalyzed reactions may occur in the stomach and the kidney, whereas base-catalyzed reactions may be assisted by the alkaline pH of the intestine.
FIGURE 33.14 Reduction of polyhalogenated compounds.
FIGURE 33.15 Reductive biotransformation of nitro arene compounds.
Ch33-P374194.indd 682Ch33-P374194.indd 682 5/30/2008 6:35:28 PM5/30/2008 6:35:28 PM

683
Phase II, or conjugation reactions, are also substitution reactions, which proceed by means of an endogenous and generally activated electrophile. In mammals, fi ve major conjugation reactions of xenobiotics exist and are mediated by transferase enzymes. Acid compounds, through their acyl-CoA ester, may also be conjugated with amino acids such as glycine, glutamine, and taurine. The specifi city for the endogenous agent is high, but the specifi city for the xenobiotic is broader.
To a great extent, conjugation produces excretable and nontoxic metabolites and thus is referred to as detoxifi cation, but exceptions exist in each class of conjugation reaction. A more-in-depth discussion of Phase II metabolism can be found in Chapter 32.
1 . Glucuronic acid conjugation
This substitution involves the transfer of a glucuronic acid from uridine diphosphate glucuronic acid (UDPGA) to a functional group in the xenobiotic substrate. The group may be a hydroxyl, carboxylic acid, amino or sulfur functional group. Glucuronides are never directly impli-cated in toxicity but are sometimes responsible for target-organ toxicity. Aromatic amines may be converted in the liver into hydroxylamine O- glucuronides, which are excreted in the urine and broken down in the bladder (if its pH is acidic) to liberate the proximate hydroxylamine carcinogen.
2 . Sulfation
Sulfate conjugation gives a polar and ionized conjugate by means of the esterifi cation of a hydroxyl group with sulfate ion (transferred from 3 � -phosphoadenosine-5 � -phosphosul-fate or PAPS). The reaction is catalyzed by a hydrosoluble sulfotransferase. Sulfation sometimes gives rise to reactive intermediates that may undergo further reactions to yield electrophilic metabolites. In the case of 2-acetaminofl u-orene, the O- sulfate moiety is a facile leaving group, and this cleavage produces nitrenium ions, which act as alkylating agents for DNA ( Figure 33.11 ).
3 . Acetylation
Acetylation is a very common metabolic reaction, which occurs with amino, hydroxyl or sulfhydryl groups. The acetyl group is transferred from acetyl-Coenzyme A, and the reaction is cat-alyzed by acetyltransferases. An important aspect of this kind of substitution is the genetic polymorphism of one acetyltrans-ferase in humans, who are divided into fast and slow acetyla-tors. In a few cases, the conjugates are further metabolized to toxic compounds, as is seen with isoniazid. Some evidence exists that acetylation of the antitubercular isoniazid leads to enhanced hepatotoxicity of the drug. 33,34 Acetylation followed by hydrolysis and CYP-dependent oxidation yields free acetyl
radicals 35 or acylium cations, which may acetylate the nucle-ophilic macromolecule functions ( Figure 33.16 ).
4 . Glutathione conjugation
Substitution reactions of xenobiotics with glutathione are the most important and contribute effi ciently to detoxifi cation. Nevertheless, in some cases, such as vicinal dihalogenated compounds, glutathione conjugation produces monosub-stituted derivatives, which may cyclize into a highly elec-trophilic episulfonium ion ( Figure 33.17 ). 36
5 . Methylation
Methylation is rarely of quantitative importance in the metabolism of xenobiotics. The methyl group is transferred from the nucleotide S- adenosyl- l -methionine (SAM) by means of a methyltransferase. The functional groups that undergo methylation include primary, secondary and tertiary amines, pyridines, phenols, catechols, thiophenols. The aza-heterocycle pyridine is metabolized to the N- methylpyridin-ium ion, which is more toxic than pyridine itself 37 ( Figure 33.18 ). The binding properties of the ionized metabolite are disturbed by the loss of its hydrophobic feature, resulting from the polarity inversion.
E . Eliminations
Eliminations of hydrogen and a halogen occur sometimes during the metabolism of halogenated xenobiotics and lead to an alkene. The double bond may be oxidized into an epoxide by means of oxidative enzyme systems as dis-cussed above. Dehydrogenation, dehydrochlorination and dechlorination are (with oxidation) the different metabolic pathways of the γ -isomer of the insecticide hexachloro-cyclohexane (lindane). 38
F . Further biotransformations leading to the ultimate toxicant
Other reactions must be mentioned beside the major reac-tions described above. These reactions may be responsible for the transformation of a toxic metabolite into the ulti-mate toxicant. 39 Rearrangements and cyclizations are exam-ples of reactions involved in these processes. In the case of the solvent hexane ( Figure 33.19 ), the toxic metabolite, 2, 5-hexanedione, is formed by four successive oxidations of the molecule. The condensation of the γ -dicetone with the lysyl amino group of a neurofi lament protein is followed by a Paal–Knorr cyclization reaction. This is the initial proc-ess that explains the hexane-induced neurotoxicity. 40 A fur-ther auto-oxidation of the N- pyrrolyl derivatives leads to the cross-linking of the axonal intermediate fi lament proteins and the subsequent occurrence of peripheral neurotoxicity. 41
III. Reactions Involved in the Bioactivation Process
Ch33-P374194.indd 683Ch33-P374194.indd 683 5/30/2008 6:35:29 PM5/30/2008 6:35:29 PM

CHAPTER 33 Biotransformations Leading to Toxic Metabolites: Chemical Aspect684
FIGURE 33.16 Bioactivation of isoniazid.
FIGURE 33.17 Bioactivation to episulfonium ion.
FIGURE 33.18 Bioactivation of pyridine.
Analogous pyrrolyl derivatives are also found as furan metabolites. Furans are oxidized by CYP to reac-tive furan-epoxides, which rearrange to ene-dial or ene-keto-aldehyde metabolites ( Figure 33.20 ). 26,42,43 After
reaction with thiols and amines like lysine, they form stable pyrrolic derivatives. This fi rst depletes the cell of glutathione then creates cross-links in proteins and toxicity.
Ch33-P374194.indd 684Ch33-P374194.indd 684 5/30/2008 6:35:29 PM5/30/2008 6:35:29 PM

685
IV . EXAMPLES OF METABOLIC CONVERSIONS LEADING TO TOXIC METABOLITES
The formation of toxic metabolites and/or intermediates during the metabolism of drugs may occur by a considerable variety of pathways that are mediated by several enzyme systems. The following fi ve examples do not represent an exhaustive list of the bioactivation processes, but are samples of original, signifi cant and/or well-known drugs whose biotransforma-tions lead to toxic compounds by the main types of reactions discussed above. Two of them (acetaminophen and tienilic acid) are CYP-mediated oxidations. Halothane acts through both oxidative and reductive biotransformations. Valproic acid is toxic through its elimination product. The toxicity of troglitazone seems to involve two distinct metabolic path-ways, leading to both alkylating and oxidative stresses.
A . Acetaminophen
The analgesic acetaminophen (4-hydroxyacetanilide, para-cetamol) exhibits hepatotoxicity when administered in very high doses (approximately 250 mg/kg in rat and about 13 g for a 75 kg human ). 44 The metabolite responsible is known to be the N- acetyl- p- benzoquinone imine (NAPQI) ( Figure 33.21 ). 45
The formation of NAPQI may proceed via CYP2E1, 46 but also via peroxidases such as prostaglandin hydroperoxidase.
The most commonly described mechanism proposes that metabolic activation occurs through N- oxidation of acetami-nophen to N- hydroxyacetaminophen followed by dehydra-tion to NAPQI ( Figure 33.22 ). 47
However, it seems that N- hydroxyacetaminophen is not a major intermediate in the oxidation of acetaminophen. The formation of NAPQI probably proceeds by two successive one-electron oxidations 48 ( Figure 33.23 ).
During the fi rst step, a one-electron oxidation yields a phenoxy radical (Ar-O • ). 49 The presence of the radical was supported by fast fl ow ESR spectroscopy in the presence of horseradish peroxidase. In the second one-electron oxidation, the phenoxy radical is oxidized to NAPQI. As described in Figure 33.21 , the highly electrophilic NAPQI may easily react with glutathione or protein thiol groups according to a Michael-type addition. The attack of liver protein thiol
FIGURE 33.19 Bioactivation of hexane.
FIGURE 33.20 Bioactivation of furans.
IV. Examples of Metabolic Conversions Leading to Toxic Metabolites
Ch33-P374194.indd 685Ch33-P374194.indd 685 5/30/2008 6:35:31 PM5/30/2008 6:35:31 PM

CHAPTER 33 Biotransformations Leading to Toxic Metabolites: Chemical Aspect686
groups and the subsequent adduct formation is frequently mentioned in the mechanism of acetaminophen hepatotoxicity. In mice, a number of proteins were identifi ed such as glyc-eraldehyde-3-phosphate dehydrogenase, 50 calreticulin and the thiol: protein disulfi de reductases Q1 and Q5 51 and this number is increasing with the advances of proteomics. 52
Another hypothesis for the mechanism of toxicity is supported by the oxidative potency of NAPQI, but still suffers from lack of evidence. 53 NAPQI is a good oxidant
for thiols of cellular components and pyridine nucleotides. Moreover, it may undergo a redox cycling with formation of superoxide anion by means of an oxygen one-electron reduction ( Figure 33.24 ).
The stepwise reduction of oxygen produces hydrogen peroxide, and fi nally, a hydroxyl radical, which is a strong oxidant implicated in cellular oxidative stress. This oxidative stress causes glutathione depletion, a disruption of the cellular calcium regulation and modifi cations of cellular proteins, thus
FIGURE 33.21 Biotransformation pathway of acetaminophen.
FIGURE 33.22 Oxidation of acetaminophen according to the “ N -hydroxyacetaminophen pathway. ”
Ch33-P374194.indd 686Ch33-P374194.indd 686 5/30/2008 6:35:33 PM5/30/2008 6:35:33 PM

687
immunoallergic hepatitis was associated with the appearance of circulating antireticulum antibodies called anti-LKM2 antibodies, which are directed toward a liver endoplasmic reticulum protein. 58,59 From these observations, the mecha-nism of the immunotoxicity associated with the prolonged use of tienilic acid was elucidated by the Mansuy group. 60–62
Tienilic acid is oxidized in the liver by CYP monooxy-genase to 5-hydroxytienilic acid, which is the major urinary metabolite (about 50% in human). In humans, the bioacti-vation of tienilic acid depends on CYP2C9. This isoform is one of the major forms of CYP in the human liver. This oxidation occurs through an electrophilic intermediate capable of alkylating very specifi cally the CYP 61,63,64 leading to its inactivation. This mechanism-based inactivation is also observed with many xenobiotics such as alkenes with terminal unsaturation, alkynes, strained cycloalkylamines, 4-alkyldihydropyridines, benzodioxoles, and some tertiary amines. 16,65 The irreversible binding of the compound with CYP leads to an immune response and to generation of antibodies against both the modifi ed protein and its native form. In fact, the autoantibodies anti-LKM2 present in hepatitis patients recognize CYP2C9 both as native pro-tein and as modifi ed protein. In addition, patient sera con-tain antibodies to tienilic acid-modifi ed proteins. It has also been demonstrated in a rat model that tienilic acid modifi ed CYP2C9 is exported to the plasma membrane of hepato-cytes 66 and has been shown that tienilic acid treated rabbit hepatocytes, when fi rst incubated with anti-LKM2, were lysed by human NK cells. 67 Thus, it is hypothesized that appearance of tienilic acid bound proteins on the hepato-cyte surface triggers their cytolysis.
In the case of tienilic acid, the electrophilic reactive spe-cies is unknown. This is either a thiophene sulfoxide, as has been demonstrated for its 3-isomer, 68 or a thiophene epoxide ( Figure 33.25 ). In both cases the electrophilic character of the intermediate is enhanced by the presence of an activat-ing 2-keto group. In any event, this electrophilic species reacts with the enzyme CYP2C9 where it is produced and inactivates it effi ciently (one inactivation event every 13 turnover). 61 The covalent binding of tienilic acid to CYP2C9 has been directly observed by mass spectrometry. 69 This reaction occurs in all patients with active CYP2C9 using this drug; however, very few produce anti-LKM2 and have hepatitis, which suggests some specifi city in their immune response.
C . Halothane
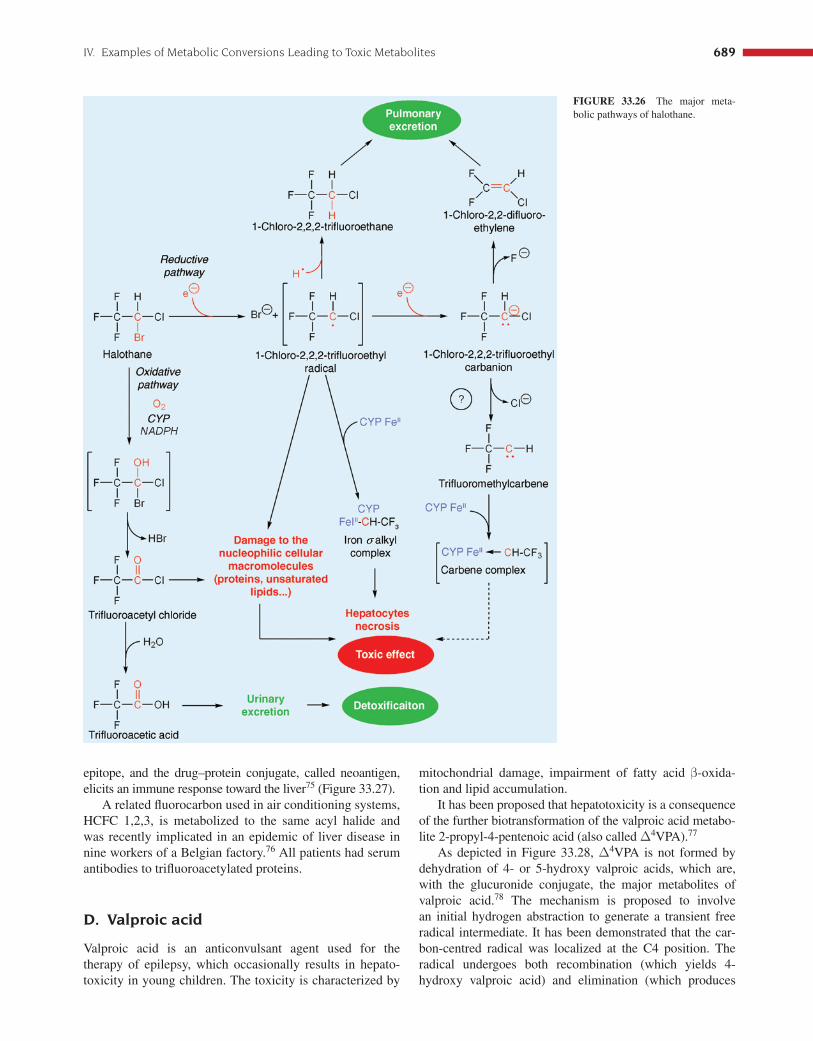
Halothane is a widely used anesthetic drug that occa-sionally results in severe hepatitis. About 60–80% of the dose is eliminated in unmetabolized form during the 24 h following administration to patients. This compound is metabolized in the presence of CYP monooxygenase CYP2E1 according to the two main pathways 13 depicted in Figure 33.26 .
FIGURE 33.23 Oxidation of acetaminophen by means of the phenoxy radical.
FIGURE 33.24 Redox cycling of N -acetyl- p -benzoquinone imine.
IV. Examples of Metabolic Conversions Leading to Toxic Metabolites
leading to cell death. Some biochemical parameters related to necrotic and apoptotic processes are affected in acetami-nophen-exposed PC12 cells transfected with CYP2E1. 54,55
It therefore appears that both covalent (e.g. alkylation) and noncovalent (e.g. oxidative stress) interactions play a major role in the pathogenesis of acute lethal cell injury caused by NAPQI. 56 At present, it is not possible to identify which of these two interactions is the critical event in initiating aceta-minophen hepatotoxicity, even if some authors suggest that the characteristic features of oxidative stress are more likely the consequences of damage mediated by protein adduction. 57
B . Tienilic acid
Tienilic acid is a uricosuric diuretic drug that may cause immunoallergic hepatitis in 1 in 10,000 patients, a side effect that resulted in its withdrawal from the market. The
Ch33-P374194.indd 687Ch33-P374194.indd 687 5/30/2008 6:35:34 PM5/30/2008 6:35:34 PM

CHAPTER 33 Biotransformations Leading to Toxic Metabolites: Chemical Aspect688
The major biotransformation pathway involves an oxidative step with introduction of an oxygen atom and subsequent formation of halohydrin. The unstable halohy-drin loses hydrobromic acid to yield trifl uoroacetyl chloride, which in turn is hydrolyzed to trifl uoroacetic acid. This fi nal metabolite is found in the urine. 70
In conditions of low levels of oxygen, a reductive path-way (10%) is enhanced and yields a free radical interme-diate characterized as 1-chloro-2,2,2-trifl uoroethyl radical. Another one-electron reduction produces the 1-chloro-2,2,2-trifl uoroethyl carbanion, which may undergo two possible kinds of eliminations.
One is the abstraction of a fl uoride ion according to a E1cB elimination, which yields 1-chloro-2,2-difl uoroethylene. This metabolite is eliminated by exhalation. Early studies suggested that a second elimination process might be an α -elimination of a chloride ion, which produces trifl uoromethylcarbene, 71 but this was later reconsidered. 72 It was hypothesized that a
carbene complex with the FeII in the active site might lead to inactivation of the CYP, but this inactivation is now thought to be due to the formation of an iron- σ -alkyl complex derived from the 1-chloro-2,2,2-trifl uoroethyl radical.
The initially formed 1-chloro-2,2,2-trifl uoroethyl radical may also cause a radical attack of polyunsaturated lipids, which produces 1-chloro-2,2,2-trifl uoroethane. This mecha-nism is similar to the pathway described with the trichlo-romethyl radical formed during the one-electron reduction of carbon tetrachloride ( Figure 33.14 ). The trichloromethyl radical may initiate a peroxidation of unsaturated lipids from the membrane with subsequent liberation of chloroform.
Several studies have demonstrated that halothane hepato-toxicity is mainly due to an immune reaction toward modifi ed proteins of the liver. In fact, these proteins are trifl uoro-acetylated on their � -NH2-lysyl residue by the trifl uoroacetyl chloride formed during the oxidative metabolism of halothane. 73,74 The product of the reaction can act as a foreign
FIGURE 33.25 Tienilic acid bio-transformation to reactive intermediates and stable metabolites.
Ch33-P374194.indd 688Ch33-P374194.indd 688 5/30/2008 6:35:35 PM5/30/2008 6:35:35 PM
689
epitope, and the drug–protein conjugate, called neoantigen, elicits an immune response toward the liver 75 ( Figure 33.27 ).
A related fl uorocarbon used in air conditioning systems, HCFC 1,2,3, is metabolized to the same acyl halide and was recently implicated in an epidemic of liver disease in nine workers of a Belgian factory. 76 All patients had serum antibodies to trifl uoroacetylated proteins.
D . Valproic acid
Valproic acid is an anticonvulsant agent used for the therapy of epilepsy, which occasionally results in hepato-toxicity in young children. The toxicity is characterized by
mitochondrial damage, impairment of fatty acid β -oxida-tion and lipid accumulation.
It has been proposed that hepatotoxicity is a consequence of the further biotransformation of the valproic acid metabo-lite 2-propyl-4-pentenoic acid (also called Δ 4 VPA). 77
As depicted in Figure 33.28 , Δ 4 VPA is not formed by dehydration of 4- or 5-hydroxy valproic acids, which are, with the glucuronide conjugate, the major metabolites of valproic acid. 78 The mechanism is proposed to involve an initial hydrogen abstraction to generate a transient free radical intermediate. It has been demonstrated that the car-bon-centred radical was localized at the C4 position. The radical undergoes both recombination (which yields 4-hydroxy valproic acid) and elimination (which produces
FIGURE 33.26 The major meta-bolic pathways of halothane.
IV. Examples of Metabolic Conversions Leading to Toxic Metabolites
Ch33-P374194.indd 689Ch33-P374194.indd 689 5/30/2008 6:35:36 PM5/30/2008 6:35:36 PM

CHAPTER 33 Biotransformations Leading to Toxic Metabolites: Chemical Aspect690
the unsaturated derivative Δ 4VPA). The formation of these metabolites is catalyzed in rat by CYP4B1 79 and in human by CYP2C9. 9 Δ4VPA is a hepatotoxic and strong teratogenic compound in animal models. In addition to that metabolic pathway, valproic acid undergoes biotransforma-tion leading to (E) Δ 2 VPA, which is devoid of embryotoxic effect in rodents. 80
Further biotransformations of Δ 4VPA involve both the liver microsomal CYP enzymes and the fatty acid β -oxidation pathway ( Figure 33.29 ). The mixed-function-oxidase system metabolizes the unsaturated metabolite to a γ -butyrolactone 81 derivative through a chemically reactive entity that is a mechanism-based inhibitor of CYP. The alkylation of the prosthetic heme by means of the radical occurs prior to formation of the epoxide. 82 Thus, the epox-ide is not involved in the CYP inhibition.
The β -oxidation cycle activates Δ 4 VPA to its Coenzyme A derivative and, through sequential steps of β -oxidation, yields the Coenzyme A ester of 3-oxo-2-propyl-4-pentenoic acid. 83 This fi nal metabolite is believed to be a
reactive electrophilic species that alkylates 3-ketoacyl-CoA thiolase (the terminal enzyme of β -oxidation) by means of a Michael-type addition through nucleophilic attack at the olefi nic terminus. 84 Oxidative stress may also be impli-cated, at least in part, in valproic acid hepatotoxicity, as suggested by experimental data on the effect of the drug on reactive oxygen species. 85
E . Troglitazone
Troglitazone (( � )-5-[4-(6-hydroxy-2,5,7,8-tetramethyl-chroman-2-ylmethoxy) benzyl]-2,4-thiazolidinedione) is an oral insulin sensitizer belonging to the thiazolidinedione class of compounds used for the treatment of type II diabetes. Its withdrawal from the US market was the consequence of the recent occurrence of hepatic failure leading sometimes to death.
It was fi rst demonstrated that troglitazone is metabo-lized mainly to sulfate and glucuronide conjugates. 86 Also
FIGURE 33.27 Biotransformation of halothane to trifl uoroacetyl chloride and the subsequent binding to protein.
FIGURE 33.28 Bioactivation of valproic acid to Δ 4 VPA.
Ch33-P374194.indd 690Ch33-P374194.indd 690 5/30/2008 6:35:37 PM5/30/2008 6:35:37 PM

691
troglitazone is an inducer of CYP3A. 87 The mechanism of toxicity is still unclear, but seems to proceed according to two distinct pathways. This is supported by the demonstra-tion that incubation of troglitazone with P450 isoforms in the presence of glutathione give rise to at least fi ve GSH conjugates. 88,89 Identifi cation of these adducts provided evidence for the two pathways described in Figures 33.30 and 33.31 .
As described in Figure 33.30 , oxidative cleavage of the thiazolidinedione ring probably generates highly electrophilic α -ketoisocyanate and sulfenic acid intermediates. This CYP 3A mediated oxidation would afford a reactive sulfoxide inter-mediate, which undergoes a spontaneous ring opening.
The second pathway ( Figure 33.31 ) consists of a CYP3A-mediated 90 one-electron oxidation of the phenolic hydroxyl group leading to an unstable hemiacetal, which opens spon-taneously to form the quinone metabolite. This undergoes thiazolidinedione ring oxidation according to the pathway shown in Figure 33.30 . Alternatively, a CYP-mediated hydrogen abstraction may occur on the phenoxy radical, leading to an o -quinone methide derivative.
It is now well established that troglitazone undergoes several metabolic transformation mediated by CYP3A4, leading to numerous electrophilic species. 91 Thus toxicity
FIGURE 33.29 Bioactivation of Δ 4 VPA.
FIGURE 33.30 Oxidation of the thiazolidinedione ring of troglitazone.
IV. Examples of Metabolic Conversions Leading to Toxic Metabolites
Ch33-P374194.indd 691Ch33-P374194.indd 691 5/30/2008 6:35:38 PM5/30/2008 6:35:38 PM

CHAPTER 33 Biotransformations Leading to Toxic Metabolites: Chemical Aspect692
acts probably both by covalent binding to hepatic proteins and oxidative stress through a redox cycling process. The implication of the thiazolidinedione moiety is less likely since the more recent drugs of this series seems devoid of toxicity. Recent studies using mitochondrial manganese superoxide dismutase partially defi cient mice also suggested that genetic defi ciencies may be, at least partially, responsi-ble for the liver failure in troglitazone-treated patients. 92,93
V . CONCLUSION
In the foregoing discussion, it has been emphasized that almost all metabolic reactions are capable of producing
reactive metabolites. This bioactivation yields toxic com-pounds that may act directly or indirectly 56 ( Figure 33.32 ). The emergence of toxicity may be the outcome of the interactions of metabolites or reactive intermediates with biological targets such as cellular macromolecules. Some compounds exhibit their toxicity by inducing the genera-tion of reactive oxygen species, thus producing alterations in the redox state of the cell. Often, covalent bonds are formed during a phenomenon that may be referred to as “ alkylating stress. ” Bioactivation of drugs followed by drug protein adduction is then considered as a key sequence in the occurrence of toxic side effects. 94 As the precise dam-ages of adducts on cellular functions are not fully under-stood, the formation of electrophilic metabolites is to be
FIGURE 33.31 Oxidation of the chromane ring of troglitazone.
Ch33-P374194.indd 692Ch33-P374194.indd 692 5/30/2008 6:35:39 PM5/30/2008 6:35:39 PM

693
avoided in drug design. Proteome profi ling (proteomics) may help to identify and compare proteins implicated in alkylating stress due to drugs, but this fi eld remains to be developed and methods are to be validated. The specifi c inhibition of an enzyme by its own substrate is a peculiar feature of alkylating stress. Determination and monitor-ing of drug protein adducts have important implications in drug development, for example, in identifying CYP3A4 inactivation, since this CYP isoform is responsible for the metabolism of about 50% of the therapeutic drugs. 95 Thus medicinal chemists have set a threshold of acceptable covalent binding when developing a new drug. For exam-ple, this value for covalent binding levels to liver proteins was less than 50 pmol-equiv/mg protein under standard conditions at Merck 96 and can be subject to discussion on a case-to-case basis. This target represents about 1/20th of the level of binding for model hepatotoxins. Often the mol-ecule can be modifi ed to decrease this type of unwanted reaction without losing too much pharmacological activity. 96–98
Such a variety of mechanisms makes it diffi cult to point at molecular functions susceptible to produce toxic effects through bioactivation. However, some major toxo-phoric groups may be highlighted ( Table 33.1 ). They may be implicated in acute or chronic toxicity. These patterns must be of particular concern in drug design. A number of recent papers on these matters have been published on how to avoid those toxic events in drug design. 99–101
Generally, the formation of toxic metabolites is not the only pathway of biotransformation, and the overall metab-olism is constituted toward detoxication and bioactivation processes. The toxic metabolites are themselves often fur-ther detoxifi ed. The duality between a benefi cial detoxica-tion phenomenon (metabolism, drug resistance) and the occurrence of a toxic effect represents the cost for adapt-ability of metabolic enzymes to the diversity of xenobiot-ics. For those interested, a recent review applies the above chemistry to predict drug safety. 102
REFERENCES
1. Williams , R. Detoxication Mechanisms , 2nd Edition . John Wiley & Sons : New York , 1959 , p. 796 .
2. Omura , T. , Sato , R. The carbon monoxide-binding pigment of liver microsomes. I. evidence for its hemoprotein nature . J. Biol. Chem. 1964 , 239 , 2370 – 2378 .
3. Snyder , R. Microsomal enzyme induction . Toxicol. Sci. 2000 , 55 , 233 – 234 .
FIGURE 33.32 Alkylating and oxidative stresses.
V. Conclusion
TABLE 33.1 Some Major Toxophoric Groups and Their Bioactivation Mechanisms
Toxophoric group Bioactivation mechanism
Azocompounds
Nitrenium ions, tautomeric carbonium ions
Acetamides
Aromatic/heterocyclic amines
Nitro compounds
Nitroaromatic compounds
Radical formation/oxidative stress
Bromoarenes Arene oxide formation
Ethinyl Ketene formation/heme destruction
Furanes Furane epoxide and ene-dial formation
Pyrroles Pyrrole oxide
Nitrogen mustard Aziridium ions
Nitroso compounds Diazonium ions/heme adduct/radical formation Hydrazines
Nitrosamines Carbenium ions/DNA alkylation
Polyhalogenated compounds
Radical and carbene formation/episulfonium with GSH
Quinone Semiquinone radical formation/oxidative stress/thiol trapping
Thioamides Thiourea formation
Thiophene Thiophene sulfoxide or thiophene epoxide formation
Vinyl Epoxidation/heme destruction
Ch33-P374194.indd 693Ch33-P374194.indd 693 5/30/2008 6:35:40 PM5/30/2008 6:35:40 PM

CHAPTER 33 Biotransformations Leading to Toxic Metabolites: Chemical Aspect694
4. Sipes , G. , Gandolfi , A. J. Biotransformation of toxicants . In Casarett and Doull’s Toxicology: The Basic Science of Poisons ( Amdur , M. O. , Doull , J. , Klaassen , C. D. , Eds) , 4th Edition . Pergamon Press : New York , 1991 , pp. 88 – 126 .
5. Alvares , A. P. , Pratt , W. B. Pathways of drug metabolism . In Principles of Drug Action: The Basis of Pharmacology ( Pratt , W. B. , Taylor , P. , Eds) , 3rd Edition . Churchill Livingstone : New York , 1990 , pp. 227 – 300 .
6. Josephy , P. D. , Guengerich , F. P. , Miners , J. O. “ Phase I and phase II ” drug metabolism: terminology that we should phase out? Drug Metab. Rev. 2005 , 37 , 575 .
7. Guengerich , F. P. Common and uncommon cytochrome P450 reac-tions related to metabolism and chemical toxicity . Chem. Res. Toxicol. 2001 , 14 , 611 – 650 .
8. Guengerich , F. P. Cytochrome P450 oxidations in the generation of reactive electrophiles: epoxidation and related reactions . Arch. Biochem. Biophys. 2003 , 409 , 59 – 71 .
9. Isin , E. M. , Guengerich , F. P. Complex reactions catalyzed by cyto-chrome P450 enzymes . Biochim. Biophys. Acta 2007 , 1770 , 314 – 329 .
10. Mason , R. P. , Chignell , C. F. Free radicals in pharmacology and toxi-cology – selected topics . Pharmacol. Rev. 1981 , 33 , 189 – 211 .
11. Aust , S. D. , Chignell , C. F. , Bray , T. M. , Kalyanaraman , B. , Mason , R. P. Free radicals in toxicology . Toxicol. Appl. Pharmacol. 1993 , 120 , 168 – 178 .
12. Singal , P. K. , Petkau , A. , Gerrard , J. M. , Hrushovetz , S. , Foerster , J. Free radicals in health and disease . Mol. Cell. Biochem. 1988 , 84 , 121 – 122 .
13. Anders , M. W. , Pohl , L. R. Halogenated alkanes . In Bioactivation of Foreign Compounds ( Anders , M. W. , Ed.) , Academic Press : Orlando, FL , 1985 , pp. 284 – 315 .
14. Pohl , L. R. , Nelson , S. D. , Krishna , G. Investigation of the mechanism of the metabolic activation of chloramphenicol by rat liver micro-somes. Identifi cation of a new metabolite . Biochem. Pharmacol. 1978 , 27 , 491 – 496 .
15. Halpert , J. R. , Miller , N. E. , Gorsky , L. D. On the mechanism of the inactivation of the major phenobarbital-inducible isozyme of rat liver cytochrome P-450 by chloramphenicol . J. Biol. Chem. 1985 , 260 , 8397 – 8403 .
16. Fontana , E. , Dansette , P. M. , Poli , S. M. Cytochrome P450 enzymes mechanism based inhibitors: common sub-structures and reactivity . Curr. Drug Metab. 2005 , 6 , 413 – 454 .
17. Cresteil , T. , Beaune , P. , Leroux , J. P. , Lange , M. , Mansuy , D. Biotransformation of chloroform by rat and human liver microsomes; in vitro effect on some enzyme activities and mechanism of irreversible binding to macromolecules . Chem. Biol. Interact. 1979 , 24 , 153 – 165 .
18. Benasutti , M. , Ejadi , S. , Whitlow , M. D. , Loechler , E. L. Mapping the binding site of afl atoxin B1 in DNA: systematic analysis of the reactivity of afl atoxin B1 with guanines in different DNA sequences . Biochemistry 1988 , 27 , 472 – 481 .
19. Koen , Y. M. , Gogichaeva , N. V. , Alterman , M. A. , Hanzlik , R. P. A proteomic analysis of bromobenzene reactive metabolite targets in rat liver cytosol in vivo . Chem. Res. Toxicol. 2007 , 20 , 511 – 519 .
20. Rietjens , I. M. , den Besten , C. , Hanzlik , R. P. , van Bladeren , P. J. Cytochrome P450-catalyzed oxidation of halobenzene derivatives . Chem. Res. Toxicol. 1997 , 10 , 629 – 635 .
21. Verna , L. , Whysner , J. , Williams , G. M. 2-Acetylaminofl uorene mech-anistic data and risk assessment: DNA reactivity, enhanced cell prolif-eration and tumor initiation . Pharmacol. Ther. 1996 , 71 , 83 – 105 .
22. Dansette , P. M. , Thang , D. C. , el Amri , H. , Mansuy , D. Evidence for thiophene- S -oxide as a primary reactive metabolite of thiophene in vivo : formation of a dihydrothiophene sulfoxide mercapturic acid . Biochem. Biophys. Res. Commun. 1992 , 186 , 1624 – 1630 .
23. Treiber , A. , Dansette , P. M. , ElAmri , H. , Girault , J. P. , Ginderow , D. , Mornon , J. P. , Mansuy , D. Chemical and biological oxidation of thio-phene: preparation and complete characterization of thiophene S -oxide dimers and evidence for thiophene S -oxide as an intermediate in thiophene metabolism in vivo and in vitro . J. Am. Chem. Soc. 1997 , 119 , 1565 – 1571 .
24. Treiber , A. , Dansette , P. M. , Mansuy , D. Mechanism of the aromatic hydroxylation of thiophene by acid-catalyzed peracid oxidation . J. Org. Chem. 2002 , 67 , 7261 – 7266 .
25. Dansette , P. M. , Bertho , G. , Mansuy , D. First evidence that cytochrome P450 may catalyze both S -oxidation and epoxidation of thiophene derivatives . Biochem. Biophys. Res. Commun. 2005 , 338 , 450 – 455 .
26. Dalvie , D. K. , Kalgutkar , A. S. , Khojasteh-Bakht , S. C. , Obach , R. S. , O’Donnell , J. P. Biotransformation reactions of fi ve-membered aro-matic heterocyclic rings . Chem. Res. Toxicol. 2002 , 15 , 269 – 299 .
27. Gaudiano , G. , Koch , T. H. Redox chemistry of anthracycline antitu-mor drugs and use of captodative radicals as tools for its elucidation and control . Chem. Res. Toxicol. 1991 , 4 , 2 – 16 .
28. Garner , A. P. , Paine , M. J. , Rodriguez-Crespo , I. , Chinje , E. C. , Ortiz De Montellano , P. , Stratford , I. J. , Tew , D. G. , Wolf , C. R. Nitric oxide synthases catalyze the activation of redox cycling and bioreductive anticancer agents . Cancer Res. 1999 , 59 , 1929 – 1934 .
29. Butler , T. C. Reduction of carbon tetrachloride in vivo and reduction of carbon tetrachloride and chloroform in vitro by tissues and tissue constituents . J. Pharmacol. Exp. Ther. 1961 , 134 , 311 – 319 .
30. Mico , B. A. , Branchfl ower , R. V. , Pohl , L. R. Formation of elec-trophilic chlorine from carbon tetrachloride – involvement of cyto-chrome P-450 . Biochem. Pharmacol. 1983 , 32 , 2357 – 2359 .
31. Gross , P. Biologic activity of hydroxylamine: a review . Crit. Rev. Toxicol. 1985 , 14 , 87 – 99 .
32. Mason , R. P. Redox cycling of radical anion metabolites of toxic chemicals and drugs and the Marcus theory of electron transfer . Environ. Health Perspect. 1990 , 87 , 237 – 243 .
33. Grant , D. M. , Hughes , N. C. , Janezic , S. A. , Goodfellow , G. H. , Chen , H. J. , Gaedigk , A. , Yu , V. L. , Grewal , R. Human acetyltrans-ferase polymorphisms . Mutat. Res. 1997 , 376 , 61 – 70 .
34. Timbrell , J. A. , Mitchell , J. R. , Snodgrass , W. R. , Nelson , S. D. Isoniazid hepatoxicity: the relationship between covalent binding and metabolism in vivo . J. Pharmacol. Exp. Ther. 1980 , 213 , 364 – 369 .
35. Sinha , B. K. Activation of hydrazine derivatives to free radicals in the perfused rat liver: a spin-trapping study . Biochim. Biophys. Acta 1987 , 924 , 261 – 269 .
36. Weber , G. L. , Steenwyk , R. C. , Nelson , S. D. , Pearson , P. G. Identifi cation of N -acetylcysteine conjugates of 1,2-dibromo-3-chloropropane: evidence for cytochrome P450 and glutathione medi-ated bioactivation pathways . Chem. Res. Toxicol. 1995 , 8 , 560 – 573 .
37. D’Souza , J. , Caldwell , J. , Smith , R. L. Species variations in the N -methylation and quaternization of [ 14 C]pyridine . Xenobiotica 1980 , 10 , 151 – 157 .
38. Videla , L. A. , Barros , S. B. , Junqueira , V. B. Lindane-induced liver oxidative stress . Free Radic. Biol. Med. 1990 , 9 , 169 – 179 .
39. Miller , E. C. , Miller , J. A. Mechanisms of chemical carcinogenesis . Cancer 1981 , 47 , 1055 – 1064 .
40. DeCaprio , A. P. , Strominger , N. L. , Weber , P. Neurotoxicity and pro-tein binding of 2,5-hexanedione in the hen . Toxicol. Appl. Pharmacol. 1983 , 68 , 297 – 307 .
41. Genter St Clair , M. B. , Amarnath , V. , Moody , M. A. , Anthony , D. C. , Anderson , C. W. , Graham , D. G. Pyrrole oxidation and protein cross-linking as necessary steps in the development of gamma-diketone neuropathy . Chem. Res. Toxicol. 1988 , 1 , 179 – 185 .
42. Chen , L. J. , Hecht , S. S. , Peterson , L. A. Characterization of amino acid and glutathione adducts of cis -2-butene-1,4-dial, a reactive metabolite of furan . Chem. Res. Toxicol. 1997 , 10 , 866 – 874 .
43. Baer , B. R. , Rettie , A. E. , Henne , K. R. Bioactivation of 4-ipomeanol by CYP4B1: adduct characterization and evidence for an enedial intermediate . Chem. Res. Toxicol. 2005 , 18 , 855 – 864 .
44. Thomas , S. H. Paracetamol (acetaminophen) poisoning . Pharmacol. Ther. 1993 , 60 , 91 – 120 .
45. Dahlin , D. C. , Miwa , G. T. , Lu , A. Y. , Nelson , S. D. N -acetyl- p -ben-zoquinone imine: a cytochrome P-450-mediated oxidation product of acetaminophen . Proc. Natl. Acad. Sci. USA 1984 , 81 , 1327 – 1331 .
46. Chen , W. , Koenigs , L. L. , Thompson , S. J. , Peter , R. M. , Rettie , A. E. , Trager , W. F. , Nelson , S. D. Oxidation of acetaminophen to its toxic
Ch33-P374194.indd 694Ch33-P374194.indd 694 5/30/2008 6:35:40 PM5/30/2008 6:35:40 PM

695
quinone imine and nontoxic catechol metabolites by baculovirus-expressed and purifi ed human cytochromes P450 2E1 and 2A6 . Chem. Res. Toxicol. 1998 , 11 , 295 – 301 .
47. Mitchell , J. R. , Jollow , D. J. , Gillette , J. R. , Brodie , B. B. Drug metabo-lism as a cause of drug toxicity . Drug Metab. Dispos. 1973 , 1 , 418 – 423 .
48. Ramakrishna Rao , D. N. , Fischer , V. , Mason , R. P. Glutathione and ascorbate reduction of the acetaminophen radical formed by per-oxidase. Detection of the glutathione disulfi de radical anion and the ascorbyl radical . J. Biol. Chem. 1990 , 265 , 844 – 847 .
49. Fischer , V. , West , P. R. , Harman , L. S. , Mason , R. P. Free-radical metabolites of acetaminophen and a dimethylated derivative . Environ. Health Perspect. 1985 , 64 , 127 – 137 .
50. Dietze , E. C. , Schafer , A. , Omichinski , J. G. , Nelson , S. D. Inactivation of glyceraldehyde-3-phosphate dehydrogenase by a reactive metabo-lite of acetaminophen and mass spectral characterization of an arylated active site peptide . Chem. Res. Toxicol. 1997 , 10 , 1097 – 1103 .
51. Zhou , L. , McKenzie , B. A. , Eccleston , E. D. , Jr , Srivastava , S. P. , Chen , N. , Erickson , R. R. , Holtzman , J. L. The covalent binding of [ 14 C]acetaminophen to mouse hepatic microsomal proteins: the spe-cifi c binding to calreticulin and the two forms of the thiol:protein disulfi de oxidoreductases . Chem. Res. Toxicol. 1996 , 9 , 1176 – 1182 .
52. Welch , K. D. , Reilly , T. P. , Bourdi , M. , Hays , T. , Pise-Masison , C. A. , Radonovich , M. F. , Brady , J. N. , Dix , D. J. , Pohl , L. R. Genomic identifi cation of potential risk factors during acetaminophen-induced liver disease in susceptible and resistant strains of mice . Chem. Res. Toxicol. 2006 , 19 , 223 – 233 .
53. Rosen , G. M. , Singletary , W. V. , Jr , Rauckman , E. J. , Killenberg , P. G. Acetaminophen hepatotoxicity. An alternative mechanism . Biochem. Pharmacol. 1983 , 32 , 2053 – 2059 .
54. Holownia , A. , Mapoles , J. , Menez , J. F. , Braszko , J. J. Acetaminophen metabolism and cytotoxicity in PC12 cells transfected with cyto-chrome P4502E1 . J. Mol. Med. 1997 , 75 , 522 – 527 .
55. Dai , Y. , Cederbaum , A. I. Cytotoxicity of acetaminophen in human cytochrome P4502E1-transfected HepG2 cells . J. Pharmacol. Exp. Ther. 1955 , 273 , 1497 – 1505 .
56. Nelson , S. D. , Pearson , P. G. Covalent and noncovalent interactions in acute lethal cell injury caused by chemicals . Annu. Rev. Pharmacol. Toxicol. 1990 , 30 , 169 – 195 .
57. Josephy , P. D. The molecular toxicology of acetaminophen . Drug Metab. Rev. 2005 , 37 , 581 .
58. Homberg , J. C. , Andre , C. , Abuaf , N. A new anti-liver-kidney micro-some antibody (anti-LKM2) in tienilic acid-induced hepatitis . Clin. Exp. Immunol. 1984 , 55 , 561 – 570 .
59. Dansette , P. M. , Bonierbale , E. , Minoletti , C. , Beaune , P. H. , Pessayre , D. , Mansuy , D. Drug-induced immunotoxicity . Eur. J. Drug Metab. Pharmacokinet. 1998 , 23 , 443 – 451 .
60. Beaune , P. , Dansette , P. M. , Mansuy , D. , Kiffel , L. , Finck , M. , Amar , C. , Leroux , J. P. , Homberg , J. C. Human anti-endoplasmic reticulum autoantibodies appearing in a drug-induced hepatitis are directed against a human liver cytochrome P-450 that hydroxylates the drug . Proc. Natl. Acad. Sci. USA 1987 , 84 , 551 – 555 .
61. Lopez-Garcia , M. P. , Dansette , P. M. , Mansuy , D. Thiophene deriva-tives as new mechanism-based inhibitors of cytochromes P-450: inac-tivation of yeast-expressed human liver cytochrome P-450 2C9 by tienilic acid . Biochemistry 1994 , 33 , 166 – 175 .
62. Lecoeur , S. , Bonierbale , E. , Challine , D. , Gautier , J. C. , Valadon , P. , Dansette , P. M. , Catinot , R. , Ballet , F. , Mansuy , D. , Beaune , P. H. Specifi city of in vitro covalent binding of tienilic acid metabolites to human liver microsomes in relationship to the type of hepatotoxicity: comparison with two directly hepatotoxic drugs . Chem. Res. Toxicol. 1994 , 7 , 434 – 442 .
63. Mansuy , D. Molecular structure and hepatotoxicity: compared data about two closely related thiophene compounds . J. Hepatol. 1997 , 26 ( Suppl 2 ) , 22 – 25 .
64. Lopez Garcia , M. P. , Dansette , P. M. , Valadon , P. , Amar , C. , Beaune , P. H. , Guengerich , F. P. , Mansuy , D. Human-liver cytochromes P-450 expressed in yeast as tools for reactive-metabolite formation studies.
Oxidative activation of tienilic acid by cytochromes P-450 2C9 and 2C10 . Eur. J. Biochem. 1993 , 213 , 223 – 232 .
65. Masubuchi , Y. , Horie , T. Toxicological signifi cance of mechanism-based inactivation of cytochrome p450 enzymes by drugs . Crit. Rev. Toxicol. 2007 , 37 , 389 – 412 .
66. Robin , M. A. , Maratrat , M. , Le Roy , M. , Le Breton , F. P. , Bonierbale , E. , Dansette , P. , Ballet , F. , Mansuy , D. , Pessayre , D. Antigenic targets in tienilic acid hepatitis. Both cytochrome P450 2C11 and 2C11-tienilic acid adducts are transported to the plasma membrane of rat hepatocytes and recognized by human sera . J. Clin. Invest. 1996 , 98 , 1471 – 1480 .
67. Neuberger , J. , Williams , R. Immune mechanisms in tienilic acid asso-ciated hepatotoxicity . Gut 1989 , 30 , 515 – 519 .
68. Valadon , P. , Dansette , P. M. , Girault , J. P. , Amar , C. , Mansuy , D. Thiophene sulfoxides as reactive metabolites: formation upon microsomal oxidation of a 3-aroylthiophene and fate in the presence of nucleophiles in vitro and in vivo . Chem. Res. Toxicol. 1996 , 9 , 1403 – 1413 .
69. Koenigs , L. L. , Peter , R. M. , Hunter , A. P. , Haining , R. L. , Rettie , A. E. , Friedberg , T. , Pritchard , M. P. , Shou , M. , Rushmore , T. H. , Trager , W. F. Electrospray ionization mass spectrometric analysis of intact cyto-chrome P450: identifi cation of tienilic acid adducts to P450 2C9 . Biochemistry 1999 , 38 , 2312 – 2319 .
70. Harris , J. W. , Pohl , L. R. , Martin , J. L. , Anders , M. W. Tissue acyla-tion by the chlorofl uorocarbon substitute 2,2-dichloro-1,1,1-trifl uoro-ethane . Proc. Natl. Acad. Sci. USA 1991 , 88 , 1407 – 1410 .
71. Mansuy , D. , Nastainczyk , W. , Ullrich , V. The mechanism of haloth-ane binding to microsomal cytochrome P450 . Naunyn Schmiedebergs Arch. Pharmacol. 1974 , 285 , 315 – 324 .
72. Ahr , H. J. , King , L. J. , Nastainczyk , W. , Ullrich , V. The mechanism of reductive dehalogenation of halothane by liver cytochrome P450 . Biochem. Pharmacol. 1982 , 31 , 383 – 390 .
73. Pohl , L. R. An immunochemical approach of identifying and char-acterizing protein targets of toxic reactive metabolites . Chem. Res. Toxicol. 1993 , 6 , 786 – 793 .
74. Kenna , J. G. , Neuberger , J. , Williams , R. Evidence for expression in human liver of halothane-induced neoantigens recognized by antibod-ies in sera from patients with halothane hepatitis . Hepatology 1988 , 8 , 1635 – 1641 .
75. Pohl , L. R. , Kenna , J. G. , Satoh , H. , Christ , D. , Martin , J. L. Neoantigens associated with halothane hepatitis . Drug Metab. Rev. 1989 , 20 , 203 – 217 .
76. Hoet , P. , Graf , M. L. , Bourdi , M. , Pohl , L. R. , Duray , P. H. , Chen , W. , Peter , R. M. , Nelson , S. D. , Verlinden , N. , Lison , D. Epidemic of liver disease caused by hydrochlorofl uorocarbons used as ozone-sparing substitutes of chlorofl uorocarbons . Lancet 1997 , 350 , 556 – 559 .
77. Baillie , T. A. Metabolic activation of valproic acid and drug-medi-ated hepatotoxicity. Role of the terminal olefi n, 2- n -propyl-4-pente-noic acid . Chem. Res. Toxicol. 1988 , 1 , 195 – 199 .
78. Rettie , A. E. , Rettenmeier , A. W. , Howald , W. N. , Baillie , T. A. Cytochrome P-450-catalyzed formation of delta 4-VPA, a toxic metabolite of valproic acid . Science 1987 , 235 , 890 – 893 .
79. Rettie , A. E. , Sheffels , P. R. , Korzekwa , K. R. , Gonzalez , F. J. , Philpot , R. M. , Baillie , T. A. CYP4 isozyme specifi city and the rela-tionship between omega-hydroxylation and terminal desaturation of valproic acid . Biochemistry 1995 , 34 , 7889 – 7895 .
80. Kassahun , K. , Baillie , T. A. Cytochrome P-450-mediated dehydro-genation of 2-n-propyl-2(E)-pentenoic acid, a pharmacologically active metabolite of valproic acid in rat liver microsomal prepara-tions . Drug Metab. Dispos. 1993 , 21 , 242 – 248 .
81. Prickett , K. S. , Baillie , T. A. Metabolism of unsaturated derivatives of valproic acid in rat liver microsomes and destruction of cyto-chrome P-450 . Drug Metab. Dispos. 1986 , 14 , 221 – 229 .
82. Ortiz de Montellano , P. R. , Yost , G. S. , Mico , B. A. , Dinizo , S. E. , Correia , M. A. , Kumbara , H. Destruction of cytochrome P-450 by 2-isopropyl-4-pentenamide and methyl 2-isopropyl-4-pentenoate: mass spectrometric characterization of prosthetic heme adducts and
References
Ch33-P374194.indd 695Ch33-P374194.indd 695 5/30/2008 6:35:41 PM5/30/2008 6:35:41 PM

CHAPTER 33 Biotransformations Leading to Toxic Metabolites: Chemical Aspect696
nonparticipation of epoxide metabolites . Arch. Biochem. Biophys. 1979 , 197 , 524 – 533 .
83. Rettenmeier , A. W. , Gordon , W. P. , Prickett , K. S. , Levy , R. H. , Baillie , T. A. Biotransformation and pharmacokinetics in the rhesus monkey of 2-n-propyl-4-pentenoic acid, a toxic metabolite of valp-roic acid . Drug Metab. Dispos. 1986 , 14 , 454 – 464 .
84. Rettenmeier , A. W. , Prickett , K. S. , Gordon , W. P. , Bjorge , S. M. , Chang , S. L. , Levy , R. H. , Baillie , T. A. Studies on the biotransfor-mation in the perfused rat liver of 2-n-propyl-4-pentenoic acid, a metabolite of the antiepileptic drug valproic acid. Evidence for the formation of chemically reactive intermediates . Drug Metab. Dispos. 1985 , 13 , 81 – 96 .
85. Chang , T. K. , Abbott , F. S. Oxidative stress as a mechanism of valproic acid-associated hepatotoxicity . Drug Metab. Rev. 2006 , 38 , 627 – 639 .
86. Kawai , K. , Kawasaki-Tokui , Y. , Odaka , T. , Tsuruta , F. , Kazui , M. , Iwabuchi , H. , Nakamura , T. , Kinoshita , T. , Ikeda , T. , Yoshioka , T. , Komai , T. , Nakamura , K. Disposition and metabolism of the new oral antidiabetic drug troglitazone in rats mice and dogs . Arzneimittelforschung 1997 , 47 , 356 – 368 .
87. Ramachandran , V. , Kostrubsky , V. E. , Komoroski , B. J. , Zhang , S. , Dorko , K. , Esplen , J. E. , Strom , S. C. , Venkataramanan , R. Troglitazone increases cytochrome P-450 3 A protein and activity in primary cultures of human hepatocytes . Drug Metab. Dispos. 1999 , 27 , 1194 – 1199 .
88. Kassahun , K. , Pearson , P. G. , Tang , W. , McIntosh , I. , Leung , K. , Elmore , C. , Dean , D. , Wang , R. , Doss , G. , Baillie , T. A. Studies on the metabolism of troglitazone to reactive intermediates in vitro and in vivo . Evidence for novel biotransformation pathways involv-ing quinone methide formation and thiazolidinedione ring scission . Chem. Res. Toxicol. 2001 , 14 , 62 – 70 .
89. Prabhu , S. , Fackett , A. , Lloyd , S. , McClellan , H. A. , Terrell , C. M. , Silber , P. M. , Li , A. P. Identifi cation of glutathione conjugates of troglitazone in human hepatocytes . Chem. Biol. Interact. 2002 , 142 , 83 – 97 .
90. Yamazaki , H. , Shibata , A. , Suzuki , M. , Nakajima , M. , Shimada , N. , Guengerich , F. P. , Yokoi , T. Oxidation of troglitazone to a qui-none-type metabolite catalyzed by cytochrome P-450 2C8 and P-450 3A4 in human liver microsomes . Drug Metab. Dispos. 1999 , 27 , 1260 – 1266 .
91. Smith , M. T. Mechanisms of troglitazone hepatotoxicity . Chem. Res. Toxicol. 2003 , 16 , 679 – 687 .
92. Jaeschke , H. Troglitazone hepatotoxicity: Are we getting closer to understanding idiosyncratic liver injury? Toxicol. Sci. 2007 , 97 , 1 – 3 .
93. Ong , M. M. , Latchoumycandane , C. , Boelsterli , U. A. Troglitazone-induced hepatic necrosis in an animal model of silent genetic mito-chondrial abnormalities . Toxicol. Sci. 2007 , 97 , 205 – 213 .
94. Zhou , S. , Chan , E. , Duan , W. , Huang , M. , Chen , Y. Z. Drug bioac-tivation, covalent binding to target proteins and toxicity relevance . Drug Metab. Rev. 2005 , 37 , 41 – 213 .
95. Yang , X. X. , Hu , Z. P. , Chan , S. Y. , Zhou , S. F. Monitoring drug-pro-tein interaction . Clin. Chim. Acta. 2006 , 365 , 9 – 29 .
96. Evans , D. C. , Watt , A. P. , Nicoll-Griffi th , D. A. , Baillie , T. A. Drug-protein adducts: an industry perspective on minimizing the potential for drug bioactivation in drug discovery and development . Chem. Res. Toxicol. 2004 , 17 , 3 – 16 .
97. Tang , C. , Subramanian , R. , Kuo , Y. , Krymgold , S. , Lu , P. , Kuduk , S. D. , Ng , C. , Feng , D. M. , Elmore , C. , Soli , E. , Ho , J. , Bock , M. G. , Baillie , T. A. , Prueksaritanont , T. Bioactivation of 2,3-diaminopy-ridine-containing bradykinin B1 receptor antagonists: irreversible binding to liver microsomal proteins and formation of glutathione conjugates . Chem. Res. Toxicol. 2005 , 18 , 934 – 945 .
98. Doss , G. A. , Baillie , T. A. Addressing metabolic activation as an integral component of drug design . Drug Metab. Rev. 2006 , 38 , 641 – 649 .
99. Baillie , T. A. , Cayen , M. N. , Fouda , H. , Gerson , R. J. , Green , J. D. , Grossman , S. J. , Klunk , L. J. , LeBlanc , B. , Perkins , D. G. , Shipley , L. A. Drug metabolites in safety testing . Toxicol. Appl. Pharmacol. 2002 , 182 , 188 – 196 .
100. Obach , R. S. , Walsky , R. L. , Venkatakrishnan , K. Mechanism-based inactivation of human cytochrome p450 enzymes and the predic-tion of drug–drug interactions . Drug Metab. Dispos. 2007 , 35 , 246 – 255 .
101. Kalgutkar , A. S. , Obach , R. S. , Maurer , T. S. Mechanism-based inactivation of cytochrome P450 enzymes: chemical mechanisms, structure–activity relationships and relationship to clinical drug–drug interactions and idiosyncratic adverse drug reactions . Curr. Drug Metab. 2007 , 8 , 407 – 447 .
102. Guengerich , F. P. , MacDonald , J. S. Applying mechanisms of chemi-cal toxicity to predict drug safety . Chem. Res. Toxicol. 2007 , 20 , 344 – 369 .
Ch33-P374194.indd 696Ch33-P374194.indd 696 5/30/2008 6:35:41 PM5/30/2008 6:35:41 PM