by vuillemin & bulard,^ trabaud & oustric,^ corral & al ... · 438 ecosystems and...
TRANSCRIPT

Inhibition mechanisms and successional
processes: a case study of Cist us albidus L. in
Provence
C. Robles, T. Dutoit, G. Benin
Laboratoire de Biosystematique et Ecologie Mediterraneenne,
Case 421 Bis, FSTSt Jerome, 13397 Marseille Cedex 20, France
Email: Thierry.Dutoit@bioeco. u-3mrs.fr
Abstract
Cistus are Mediterranean shrubs which are obligate seeders. Their germination is stimulated byfire. After this perturbation, these form pure dense stands which persist for about fifteen years,this period corresponds to the life time of the individuals. So far, shrubland decay was alwaysawarded to the non population renewal, due to the low germination rates observed without heatstimulation.
For one of these species: Cistus albidas L., we have researched to ascertain if anautotoxic phenomenon could also act on the disappearance. We have realised two types ofbioassays on C. albidas seeds: one with C. albidas leachates and the other with essential oils.We have studied the germination rate, seedling viability and growth. No significant effect wasobserved between C. albidas leachates and the same study area rains control. On the otherhand, essential oils of C. albidas leaves affect the seed germination, seedling viability andgrowth. As, otherwise, we could have shown the existence of essential oil in the litter. Duringlitter degradation, oils are likely to be released into the soil and their autotoxic properties canbe exerted . The station decay would not only result from the existence of a low germinationrate without heat stimulation but also a teratogen and lethal action of oils which would act onrare individuals.
1 Introduction
Plants of the genus Cistus (Cistacea) occupy a very important place in theMediterranean vegetation. According to Juhren/ these shrubs colonisedegraded areas and form one of the first shrub stage of successional dynamic.They are obligate seeders and their germination is stimulated by fire, as shown
Transactions on Ecology and the Environment vol 16, © 1997 WIT Press, www.witpress.com, ISSN 1743-3541

438 Ecosystems and Sustainable Development
by Vuillemin & Bulard, Trabaud & Oustric, Corral & al/ Roy & Sonie, .After this perturbation, they colonise large areas forming pure dense standswhich persist for 10 or 15 years. The stands decline at the age of about 10-15years old which corresponds to the life time of the individuals mentioned byother authors, e.g. Juhren/ Roy & Sonie. The stand decay is due to the deathof the individuals and to the absence of renewal. This latter point is explained bythe little germination rates without fire stimulation and inhibitory action of thelight quality (red/far red ratio under Cistus canopies) as shown by Roy &Sonie . However, although this germination is low, in some cases it may not beinconsiderable as suggested by Vuillemin & Bulard, and Trabaud & Oustric.
Therefore, we have researched another cause which could add to otheralready known negative factors. Among the different causes which can explainsuccessional dynamic, allelopathic processes may play an important part. Theymay be responsible for the fast disappearance of a vegetation stage by releasesof toxic compounds which inhibit seeds germination and the seedlings growth.These allelochemicals may be provided by others plants, which will form thenext vegetation stage, or by themselves (autotoxicity). The autotoxicityphenomenon is relatively frequent as noted by Rice. Consequently, theautotoxic hypothesis may explain the difficulties of regeneration encountered byCistus.
To illustrate this study, C. albidus has been chosen for its extendedassessment in Provence. It may grow on both siliceous and calcareous soil.Moreover, its chemical composition has been poorly studied and no mention ofits essential oil has been reported by literature. To estimate the existence and theimpact of the C. albidus autotoxicity processes, bioassays have been realisedwith two types of materials : natural foliar leachates collected in field andessential oils extracted in laboratory. We have studied the secondary effects ofallelopathic processes i.e. their actions on the seed germination rates, seedlingviability and growth.
The purpose of this study is to test the toxic potentiality of C albidus oilsand natural foliar leachates on its own seed germination and seedling growthand to discuss on implications of the results in the successional dynamicprocesses.
2 - Material and methods
Six natural areas have been chosen for sampling different materials : leaves,foliar natural leachates, natural rainfall and seeds. They are localized on bothtypes of substrates with varied ecological conditions. The seeds were collectedeach month from august to november 1995 in each study area. They were keptin paper bags at room temperature sheltered from light. To obtain a highgermination percentage and after several tests according to the literature data,seeds are heat pretreated in dry heater at 80°C for 10 minutes, the same day ofthe experiment. Damaged seeds are eliminated. The germination bioassays havebeen made in 9 cm diameter Petri dishes lined with a sheet of filter paper(Whatmann n°4). 20 intact C. albidus seeds are laid in each Petri dishes. 2ml of
Transactions on Ecology and the Environment vol 16, © 1997 WIT Press, www.witpress.com, ISSN 1743-3541

Ecosystems and Sustainable Development 439
treatment are added initially. Seven Petri dishes are realised for each treatmentof each experimentation. The germination bioassays are realised underphotoperiod conditions for 30 days with a counting every two days. As soon asa seed is germinated, it is removed to another Petri dish. They are regularlywatered with the corresponding treatment for the tests with natural leachatesand distilled water for the oils tests. 14 days after removal, seedling radicle andhypocotyl are measured. Seedlings with necrosis, depigmentation or othersymptoms are not taken into account in the measures. All the seedlingsqualitative observations were listed.
2.1 - Natural foliar leachates
The natural foliar leachates were collected in four sites : Cal, Ca2(calcareous sites), Sil and Si2 (siliceous sites). For each substrate, one of thetwo sites is a C. albidus pure dense stand (Cal and Si2). The foliar leachatesare collected in 10 cm diameter pluviometers placed at random below Cistusplants. They are covered with a net to avoid insect or leaf introduction. Neareach site, several identical pluviometers localised in open area allow to collectnatural rainfall (field control). The pluviometers were placed in april andSeptember 1996. Every time, the foliar leachates and field control were collectedafter several days of rain. The collected samples were filtered (micropore, 2j m)and maintained in freezer. The two samples of a same site were further onmixed up in order to get a sufficient volume to realise the experiment. The foliarleachates were tested on the seeds of the corresponding site. Seeds wereregularly watered with their treatment. Two control were realised, one withdistilled water (laboratory control) and one with natural rainfall (field control).After germination, seedlings were watered with the corresponding treatment.
2.2 - Essential oils
Essential oils are obtained by hydrodistillation for 5 hours of the leavesharvested every 5 weeks for 14 months in each study area. They are kept infreezer. However, because of the very low yields, oils corresponding to thedifferent samples of the different sites of a substrate had to be mixed. So, twotypes of oils are tested : siliceous and calcareous oils, on a mixture of seedsproceeding of all sites for each substrates. Four concentrations are tested : 10 ,10" , 10"*, 10"* (v/v) and a control. Oils are diluted in hexane, and the control isrealised with pure hexane. After solvent evaporation, seeds are deposed andwatered with the water volume corresponding to dilution. No more oil will beadded, and the seeds and the seedlings will be watered with only distilled water.
2.3 - Analysis of data
The variation among treatments is tested with Mann-Whitney's U test forgermination rate and with Student's t test for the seedling growth. Statistical
Transactions on Ecology and the Environment vol 16, © 1997 WIT Press, www.witpress.com, ISSN 1743-3541
440 Ecosystems and Sustainable Development
analysis are performed with Statview software. The threshold for significantdifferences between treatments is taken at p < 0,01.
3 Results
3.1 - The foliar leachates
The mean germination rate of each site and each treatment is listed in table 1.
Germination
c%)
nonviableseedlings-
total seedlingsratio
Study AreaCalCa2SilSi2CalCa2SilSi2
TreatmentLC60,0%52,0%82,0%66,5%0,120,140,060,09
F.C.65,0%49,3%83,0%75,5%0,270,420,080,11
F.L.75,59459,5%85,0%77,0%0,180,290,050,07
Table 1 : Mean germination percentage of C. albidus seeds and number ofnonviable seedlings-total number of seedlings ratio in presence of foliarleachates of C. albidus (F.L.), laboratory control (L.C.) or field control (F.L.).
Germination rate is always higher with the foliar leachates then with the fieldcontrol (except for Ca2) and then with laboratory control. So, the foliarleachates seem to favour seed germination but the rates of treatments arerelatively similar. The Mann-Whitney's U test indicates no statisticallysignificant differences. Among seeds germinated, a part of them showalterations (necrosis, no development, monsters...) and some others died beforethe 14* day. These seedlings (damaged + dead) are considered like nonviablebecause of their great degree of alteration. Table 1 presents the number ofnonviable/total number seedlings ratio. The field control seeds ratio is alwayshigher than the two other treatments ratio. For the calcareous sites, the seedswatered with the foliar leachates show a higher ratio compared to laboratorycontrol whereas the ratio is similar for the siliceous areas. The number of viableseedlings is generally higher for seeds watered with foliar leachates. However,the difference between treatment are lower.
The mean length of radicle, hypocotyl and total is listed in Table 2. Seedlingsgrowth bioassays show, whatever the study area, the distilled water seedlinglength is more important than the length of the two other treatments. However,the difference is only statistically significant for the site Si2. The foliar leachatesseedling total length is generally superior to the seedling total length of the fieldcontrol (except for Ca2), but a Student's t test reveals no significant differencesbetween the lengths.
Transactions on Ecology and the Environment vol 16, © 1997 WIT Press, www.witpress.com, ISSN 1743-3541

Ecosystems and Sustainable Development 441
s
t
u
d
y
A
r
e
a
Cal
Ca2
Sil
Si2
LC
F.C.FL.LC
F.C.
FL
LC
F.C.
F.L.
LC
F.C.F.L.
Radicle length(cm)
1,8 ±0,50
1,6 ±0,48
1,7 ±0,47
1,6 ±0,49
1,5 ±0,41
1,4 ±0,46
2,4 ± 0,65
2,0 ±0,56
2,0 ±0,61
2,3 ±0,52
1,9 ±0,49
1,8 ±0,43
Hypocotyllength (cm)
1,6 ±0,27
1,6 ±0,31
1,6 ±0,34
1,6 ±0,38
1,7 ±0,33
1,6 ±0,29
1,9 ±0,29
2,0 ± 0,29
2,0 ± 0,29
1,9 ±0,29
1,7 ±0,29
1,9 ±0,33
Total length(cm)
3,4 ±0,71
3,2 ± 0,683,4 ± 0,67
3,3 ±0,78
3,3 ±0,65
3,1 ±0,61
4,2 ± 0,84
4,0 ± 0,73
4,0 ± 0,82
4,2 ±0,71
3,6 ± 0,693,7 ±0,67
nb of measuredseedlings
73
64
84
60
39
59
105
106
113
85
9297
Table 2 : Mean and +/- S.E. (standard error) of hypocotyl length, radicle lengthand total length of C. albidus seedlings watered with natural foliar leachates(F.L.), laboratory control (L.C.) or field control (F.L.).
3.2 - Essential oil bioassays
C 104 103 10-2 10-1
-*- Mangpninabon%
-*- Ratio
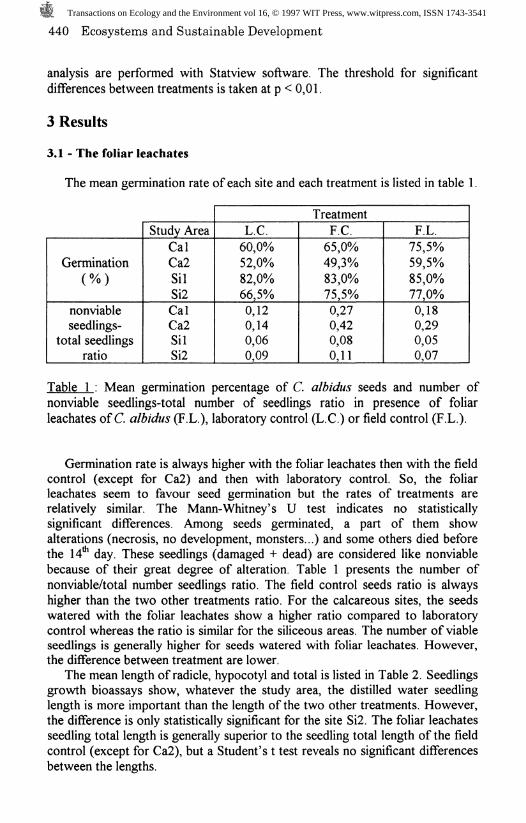
Figure 1 : Mean percentage germination of C. albidus seeds exposed to variousoil concentrations and C : control ( calcareous, -— siliceous).
Transactions on Ecology and the Environment vol 16, © 1997 WIT Press, www.witpress.com, ISSN 1743-3541
442 Ecosystems and Sustainable Development
Figure 1 shows the mean germination percentage per treatment. Germination isstimulated by enhanced oil concentrations until 10" for the calcareous oils and10" for siliceous oils (figure 1). However, the stimulating effect is onlysignificant for 10" and control for siliceous oil. On the other hand, thegermination is severely inhibited for the two oils at 10"*. A Mann-Whitney's Utest show a statistically significant difference between this concentration and theothers or the control. Like foliar leachates bioassays, some seedlings died beforethe 14* day or are damaged (necrosis, monster, depigmentation). Thenonviable/total seedlings ratio enhanced strongly from 1(T* to 10"* where itreaches almost 100% (Figure 1). On the other hand, it's lower at 10 comparedto control. So, the low concentrations seem to stimulate germination and favoura «normal» seedling development whereas higher concentrations inhibitgermination and/or have lethal or teratogen effects on the seedlings.
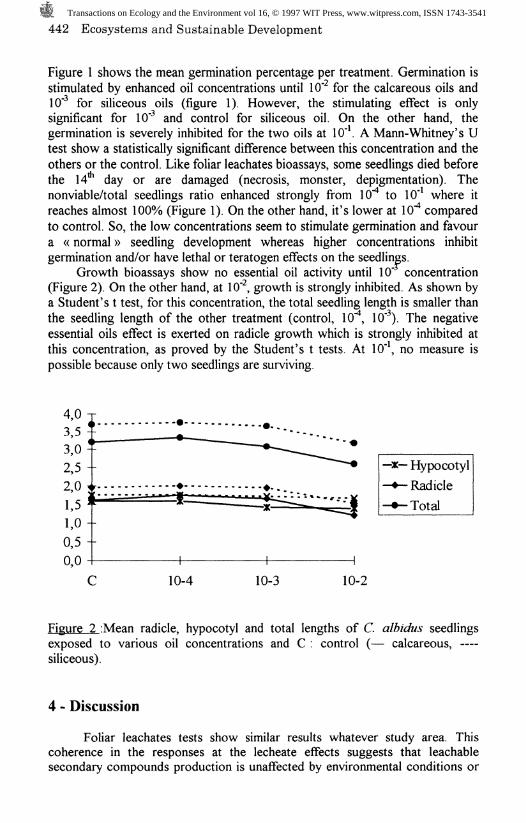
Growth bioassays show no essential oil activity until 10" concentration(Figure 2). On the other hand, at 10" , growth is strongly inhibited. As shown bya Student's t test, for this concentration, the total seedling length is smaller thanthe seedling length of the other treatment (control, 10 , 10" ). The negativeessential oils effect is exerted on radicle growth which is strongly inhibited atthis concentration, as proved by the Student's t tests. At 10"*, no measure ispossible because only two seedlings are surviving.
10-4 10-3 10-2
Figure 2 iMean radicle, hypocotyl and total lengths of C. albidus seedlingsexposed to various oil concentrations and C : control (— calcareous, —siliceous).
4 - Discussion
Foliar leachates tests show similar results whatever study area. Thiscoherence in the responses at the lecheate effects suggests that leachablesecondary compounds production is unaffected by environmental conditions or
Transactions on Ecology and the Environment vol 16, © 1997 WIT Press, www.witpress.com, ISSN 1743-3541

Ecosystems and Sustainable Development 443
too slightly to lead to a difference in their effects. Although osmotic potentialsas shown by Wardle^ may distort the bioassay interpretatios, they have notbeen taking into account in this study because of the field control which allowsa correction compared with the laboratory control. The germination rate resultssuggest that natural foliar leachates tend to enhance germination and seedlingssurvival. However, the differences between the foliar leachates germination andthe other germination rates are lower and above all no statistically significant.We must conclude that foliar leachates have no effects on germination rate andseedlings viability, whatever the ecological condition. Generally, aqueousextracts or leachates are less toxic than organic extracts as shown by Kumari &al. Particularly, Molina & aP show that natural aerial leachates are not aconsistent source of allelochemicals. Moreover, in our study, the samplings arecollected after several days of rain. So, the foliar toxins were diluted which hadaffected their potentialities. For these reasons, the absence of any foliarleachates effect on the germination rate and the seedling survival doesn't provethat leachable allelopathic compounds in C. albidus leaves do not exist.Moreover, Tukey^ has shown that some other climatic events may be moreeffective in leaching allelochemical compounds than rain water.
Seedling growth bioassays show that distilled water seedling length is alwayshigher than that of other treatments. This is likely the result of osmotic effects.Wardle^ has proved that growth measurements are likely to lead to error ifosmotic potentials are not taken into account. No significant difference isobserved between the field control seedling length and the foliar leachateslength. According to Rutherford & Powrie,* foliar leachates usually stimulategrowth by their nutrient supply. Perhaps, no effect is observed in ourexperiment because the growing time (14 days) is too short. They developthemselves with cotyledon stores, so nutrient supply has no effect. The otherhypothesis is that the nutrient leaching on C. albidus leaves is insignificant.
Essential oil bioassays have proved oil negative effects at highconcentrations on all the three measured parameters : germination percentage,seedling viability and growth. C. albidus oils are almost exclusively composedof sesquiterpens (unpublished data). Previous works like Fischer & al's orKomai & aFs^ had shown allelopathic activities of these compounds. In ourstudy, seed germination rate is severely inhibited by oil high concentrations.Moreover, these essential oils have lethal and teratogen action which can beobserved on seedlings. As only an initial quantities of oil is supplied to the seedsand as the seedlings do not receive any oils, we can conclude that essential oilaction must be exerted very early on the embryo (before and/or duringgermination). Embryo potentialities remain affected whatever ulterior conditionsand even if it had no more contact with essential oils. The results suggest thatthese oils have deleterious irreversible effects on seed becoming.
Moreover, these oils inhibit seedlings growth. Radicle elongation is severelydepressed. In a poor nutrient environment, it may be an additional obstacle forseedling establishment. It will be more difficult for the seedlings to take upnecessary nutrients for their growth and development. Moreover, according toRutherford & Powerie, under conditions of environmental stress like
Transactions on Ecology and the Environment vol 16, © 1997 WIT Press, www.witpress.com, ISSN 1743-3541

444 Ecosystems and Sustainable Development
diminished water availability or frost, root-shoot ratio may be increased and thisincrease represents an adjustment factor. On the contrary, the decreasing of theratio observed in our experimentation may cause the seedlings to become moresensitive. This reduced capacity to resist water stress or frost stress leadprobably to a great seedlings mortality. These results agree with observationsmade by numerous authors e. g. Clemente & al/ Trabaud & Oustric ... Theyhave observed important germination during spring and autumn followed by ahigh death rate caused by water or frost stress. This important death rate wouldnot be only stress consequence but also its combined action to a lower seedlingresistence due to a vegetal autotoxic action.
So, essential oils exhibit autotoxic properties. They limit germination rate.Among the germinated seeds, some of them can't survive because theirdevelopment is not normal. Surviving seedlings exhibit radicle length muchsmaller than control radicle length. Thus this multiplies their sensitivity to waterstress which brings on a higher death rate. Existence of autotoxicityphenomenon has been already described for other plants by numerous authors e.g. : Kumai & Kohli/ Vokou & Margaris/* Pelissier,™ Tarayre & al." Theycould allow population regulation and/or save a vital area for the individuals.Yet, our experimentation does not prove the existence of such phenomenon infield. However, further experiments have revealed the presence of theseessential oils in C. albidus leaves litter (unpublished data). During litterdegradation, the oils are probably released in the soil and may exert theirautotoxic effects either on new fallen seeds or on seed soil bank. Steamdistillation of soil below Cistus plants have sometimes shown essential oilexistence (unpublished data). Various studies e. g. : Rice," Molina & al/Fischer & al have shown that litter could be an important carrier forallelochemicals. The fire would then play a double part: stimulating germinationrates and deleting compounds that would not have been leached away.
Further experiences are necessary to test the existence of autotoxic action infield. However, these results show that Cistus albidus autotoxicity phenomenonexists. This fact may strongly limit seedling appearance and growth. The aboveprocess adds to other negative factors already known : absence of heat stimulusand inhibitory red/far red ratio under shrub canopies as shown by Vuillemin &Bulard, Trabaud & Oustric, Roy & Sonie. Thus, the fast stand decay wouldresult of two complementary phenomenon : a low germination rate belowcanopy and without fire, and the existence of an autotoxicity phenomenonwhich strongly reduces germination and has injurious effects on seedlingsviability and stress resistance.
5 - Conclusion
C. albidus essential oils have shown autotoxic potentialities at highconcentrations. Their effects act on the three parameters taken into account inthis study : germination rate, seedling viability and growth. By their inhibitoryactions on germination and by their lethal and teratogen effects, oils may limitstrongly the appearance of new individuals able to develop normally. Radicle
Transactions on Ecology and the Environment vol 16, © 1997 WIT Press, www.witpress.com, ISSN 1743-3541

Ecosystems and Sustainable Development 445
growth inhibition is ascertained in surviving seedlings. This lower radiclegrowth can be a limiting factor for seedling growing in poor nutrientenvironments. These seedlings are likely very sensitive to climatic stress like thedry hot season. This radicle elongation inhibition may provoke a high seedlingdeath rate.
These allelochemicals do not seem to be released by natural foliar leachates.Bioassays realised with them show no significant effect on the measuredparameters. On the other hand, essential oils have been found in leaf litter and insome cases in soil below Cistus plants. The above results suggest that a Cistusalbidus autotoxicity phenomenon could occur in natural conditions. Thecompounds released by litter during its degradation can exert deleteriousactivity against new fallen seeds or even bank soil seeds. The stand decay andespecially the absence of new individuals appearance would not only result fromnegative physical factors but also autotoxic compound actions.
References
1. Clemente, A.S., Rego, F.C., Correia, O.A. Demographic patterns andproductivity of post-fire regeneration in Portuguese Mediterranean maquis,Int. J. WildlandFire, 1996, 6, 1, 5-12.
2. Corral, R., Pita, J.M., Perez-Garcia F. Some aspects of seed germination infour species of CistusL SeedSci. & Technol, 1990, 18, 321-325.
3. Fischer, N.H., Williamson, G.B., Weidenhamer, J.D., Richardson, DR Insearch of allelopathy in the Florida scrub : The role of terpenoids, J. Chem.EcoL, 1994, 20, 6, 1355-1380.
4. Fischer, N.H., Weidenhamer, JD , Bradow, J.M. Inhibition and promotionof germination by several sesquiterpenes, J. Chem., EcoL, 1989, 15, 6,1785-1793.
5. Juhren, M.C. Ecological observations on Cistus in the Mediterraneanvegetation. - Forest science, 1966, 12, 4, 415-426.
6. Komai, K, Tang, C.S., Nishimoto, R.K. chemotypes ofCyperus rotundusin Pacific rim and basin distribution and inhibitory activities of theiressential oils, J. Chem. EcoL, 1991, 17, 1, 1-8.
7. Kumari, A., Kohli, R.K. Autotoxicity of ragweed parthenium (Partheniumhysterophorus\ Weed Science, 1987, 35, 629-632.
Transactions on Ecology and the Environment vol 16, © 1997 WIT Press, www.witpress.com, ISSN 1743-3541

446 Ecosystems and Sustainable Development
8. Kumari, A., Kohli, R.K., Saxene, D.B. Allelopathic effects of Partheniumhysterophorns L leachates and extracts on Brassica campestris L., Annalsof Biology, 1985, 1, 2, 189-196.
9. Molina, A., Reigosa, M.J., Caballeira, A Release of allelochemical agentsfrom litter, throughfall and topsoil in plantations of Eucalyptus globuluslabill in Spain, J. Chem. Ecol., 1991, 17, 1, 147-160.
10. Pelissier, F. Allelopathic inhibition of spruce germination, Acta (Ecologica,1993, 14,2,211-218.
11. Rice, EL Allelopathy, Second Edition , Academic Press, London, 1984.
12. Roy, J, Sonie, L Germination and population dynamics of Cistus speciesin relation to fire, J. of Applied Ecology, 1992, 29, 647-655.
13. Rutherford, M.C., Powrie, L.W. Allelochemic control of biomass allocationin interacting shrub species, J. Chem. Ecol, 1993, 19, 5, 893-906.
14. Tarayre, M, Thompson, J.D., Escarre, J, Lynhart, Y.B. Intraspecificvariation in the inhibitory effects of Thymus vulgaris (Labiatae)monoterpenes en seed germination, Oecologia, 1995, 101, 110-118.
15. Trabaud, L, Oustric, J. Heat requirements for seed germination of threeCistus species in the garrigue of southern France, Flora, 1989, 183, 321-325.
16. Trabaud, L., Oustric, J. Comparaison des strategies de regeneration apresincendie chez deux especes de cistes, Revue Ecol. (terre et vie), 1989, 44,3-13.
17. Tukey, H.B. The leaching of substances from plants, Ann. Rev. PlantPhysioL, 1970, 21, 305-324.
18. Vokou, D, Margaris, N.S. Autoallelopathy of Thymus capitatus, Acta(Ecologica, (Ecol. Plant., 1986, 7 (21), 2, 157-163.
19. Vuillemin, J, Bulard, C. Ecophysiologie de la germination de Cistusalbidus L. et Cistus monspeliensis L , Naturalia Monspeliensa Ser. Bot.,1981,46, 1-11.
20. Wardle, DA, Nicholson, K.S., Ahmed, M. Comparison of osmotic andallelopathic effects of grass leaf extracts on grass seed germination andradicle elongation, Plant and Soil, 1992, 140, 315-319.
Transactions on Ecology and the Environment vol 16, © 1997 WIT Press, www.witpress.com, ISSN 1743-3541