chromosomal aberrations: formation, identification and distribution · chromosomal aberrations:...
TRANSCRIPT

Mutation Research 504 (2002) 17–36
Chromosomal aberrations: formation, identificationand distribution
G. Obea,∗, P. Pfeiffera, J.R.K. Savageb, C. Johannesa, W. Goedeckea, P. Jeppesenc,A.T. Natarajand, W. Mart́ınez-Lópeze,f , G.A. Follee, M.E. Dretse
a University of Essen, Department of Genetics, 45227 Essen, Germanyb Radiation and Genome Stability Unit, Medical Research Council, Harwell, Didcot, Oxfordshire OX11 0RD, UK
c Medical Research Council, Human Genetics Unit, Edinburgh EH4 2XU, UKd Deptartment of Radiation Genetics & Chemical Mutagenesis, Leiden University Medical Center,
Wassenaarseweg 72, 2333 AL Leiden, The Netherlandse Instituto de Investigaciones Biológicas, Clemente Estable, Avda. Italia 3318, 11600 Montevideo, Uruguay
f Faculty of Sciences, University of the Republic Montevideo, Montevideo, Uruguay
Received 8 November 2001; received in revised form 12 December 2001; accepted 17 January 2002
Abstract
Chromosomal aberrations (CA) are the microscopically visible part of a wide spectrum of DNA changes generated bydifferent repair mechanisms of DNA double strand breaks (DSB). The method of fluorescence in situ hybridisation (FISH)has uncovered unexpected complexities of CA and this will lead to changes in our thinking about the origin of CA. Theinter- and intrachromosomal distribution of breakpoints is generally not random. CA breakpoints occur preferentially inactive chromatin. Deviations from expected interchromosomal distributions of breakpoints may result from the arrangementof chromosomes in the interphase nucleus and/or from different sensitivities of chromosomes with respect to the formationof CA. Telomeres and interstitial telomere repeat like sequences play an important role in the formation of CA. Subtelomericregions are hot spots for the formation of symmetrical exchanges between homologous chromatids and cryptic aberrations inthese regions are associated with human congenital abnormalities. © 2002 Elsevier Science B.V. All rights reserved.
“Chromosomal studies have long promised tobecome a kind of visible chemistry”[1].
1. Introduction
Chromosomal aberrations (CA) are one of theimportant biological consequences of human expo-sure to ionizing radiation and other genotoxic agents.In epidemiological studies, it has been shown that
∗ Corresponding author.E-mail address: [email protected] (G. Obe).
people with elevated frequencies of CA in their pe-ripheral blood lymphocytes have a significantly el-evated risk of developing cancer[2–6]. Many typesof cancers are associated with specific types of CAwhich are etiologic for the cancer in question[7].Dose–response curves (DRC) for CA are differentdepending on the inducing agent, the cell cycle stageexposed and the type of CA analyzed. From DRCfor CA induced by radiation of low linear energytransfer (LET) first interpretations of the origin of CAin a mechanistic sense were deduced[8]. DRC fordicentric chromosomes and reciprocal translocationsinduced by radiation in G0 lymphocytes in vivo and
0027-5107/02/$ – see front matter © 2002 Elsevier Science B.V. All rights reserved.PII: S0027-5107(02)00076-3

18 G. Obe et al. / Mutation Research 504 (2002) 17–36
in vitro are similar and are therefore used in biolog-ical dosimetry[9–12]. The frequency of inborn CAin human population is quite high and it is of utmostimportance to understand how CA originate and howthey are transmitted to progenies[13,14]. Testingagents for their ability to induce CA has a firm placein screening strategies for mutagenic/carcinogenicagents[15,16]. CA are a small fraction of a hugeamount of changes in chromosomal DNA and reflectan enormous plasticity of the genome which has farreaching consequences for evolution[17].
In the present study, the current status of the mech-anisms of induction of CA, their identification andclassification as well as their distribution among thegenome is summarized and discussed.
2. DNA damage and repair lead tochromosomal aberrations
With the exception of polytene chromosomes, eu-karyotic chromosomes are uninemic. They containone continuous DNA molecule in the presyntheticphase of the cell cycle which is replicated during theS-phase. These DNA molecules are extremely longwhen compared to metaphase chromosomes, or to therespective fibrillar structures in interphase chromatin[18,19]. According to calculations made by DuPraw[18], human chromosome 1 contains a DNA moleculeof 7.5 cm length which in interphase is packed in afibrillar structure of 1350�m. In metaphase, chromo-some 1 is about 10�m long. These packaging prob-lems are mainly solved by proteins of various types[20,21]. Due to their enormous dimensions, DNAmolecules in chromosomes are permanent targets ofchemical or physical damage of diverse origin. Onemicroscopically visible result of such damage are CA.
CA are induced by agents that damage chromo-somal DNA [22–24]. Experimental analyses haveshown that DNA double strand breaks (DSB) arethe principal lesions in the process of CA formation[25–27]. DSB arise spontaneously at quite significantfrequencies through a variety of cellular processes,and can be directly induced by ionizing radiation, cer-tain antibiotics, or endonucleases[27,28]. Sources ofspontaneously induced DSB are DNA replication andDNA excision repair by accumulating single strandbreaks (for reviews see[29,30]), transposition[31],
VDJ-recombination[32–34], antibody class switch-ing [35], mitotic recombination[36–38]and oxidativedamage[39].
UV-radiation and the majority of chemical muta-gens are not able to induce DSB directly but leadto other lesions in chromosomal DNA which duringrepair, or DNA synthesis, may give rise to DSB andeventually to CA.
If left unrepaired, DSB may lead to broken chromo-somes. In view of the enormous amount of DNA in achromosome and its very complex protein-associatedpacking, it is likely that many isolated DSB will be sosplinted with proteins that they probably never appearas breaks (severances) in metaphase chromosomes.Unrepaired DSB but also CA (that arise by misrepairof DSB, see below) may be cell lethal at interphaseor mitosis, respectively. If repaired improperly, DSBmay lead to mutations, chromosome rearrangements,and oncogenic transformation.
In eukaryotic cells, DSB are repaired by at leastthree different mechanisms: (i) homologous recom-bination repair (HRR), a highly accurate process thatusually precisely restores the original sequence atthe break; (ii) single-strand annealing (SSA), whichleads to the formation of mainly interstitial deletions;(iii) nonhomologous DNA end joining (NHEJ) whichjoins two broken ends directly and usually generatessmall scale alterations (base pair substitutions, inser-tions and deletions) at the break site. Although NHEJis considered the major pathway of DSB repair in ver-tebrate cells, evidence has emerged recently that ver-tebrate cells are also quite proficient at HRR[40–44].This is consistent with the increasing number of ver-tebrate genes (e.g. XRCC2 and XRCC3) homologousto the members of theSaccaromyces cerevisiae Rad52gene group responsible for HRR in yeast. These re-sults are in contrast to the current dogma that onlyyeast but not vertebrate cells are capable of repairingDSB efficiently by HRR and change our picture ofDSB repair in vertebrate cells[45–49].
HRR and SSA involve the members of the Rad52gene group and strictly require regions of extensivesequence homology. NHEJ depends on the productsof the genes XRCC4-7 and can dispense with se-quence homology (for a more detailed overview ofthe DSB repair processes and the proteins involvedrefer to some recent reviews[28,46–48,50]). The es-sential requirement of HRR for sequence homology

G. Obe et al. / Mutation Research 504 (2002) 17–36 19
is reflected by the fact that it occurs preferentiallybetween sister chromatids in cells undergoing mitoticcell cycles or between homologous chromosomes incells undergoing meiotic cell cycles where it maylead to gene conversion. In addition, HRR can alsooccur between homologous DNA sequences in differ-ent chromosomes (ectopic HRR) which may lead toexchange type CA such as dicentrics and transloca-tions (Fig. 1, upper panel). HRR is usually initiatedby one single DSB to generate both correct intra-chromosomal repair products, and incorrect exchange
Fig. 1. Examples for the possible formation of CA by different mechanisms.
type CA (the two DNA ends of a single DSB interactwith the two strands of a homologous unbroken DNAduplex). DSB between two direct repeat sequencescan be repaired by SSA which leads to the deletion ofone repeat unit and the intervening sequence (Fig. 1,lower panel). NHEJ rejoins DSB in the absence ofextended sequence homology and is, therefore, a uni-versally applicable pathway for the repair of DSB oc-curring within chromosome regions without sequencehomology. SSA and NHEJ need a single DSB togenerate intrachromosomal repair products (the two

20 G. Obe et al. / Mutation Research 504 (2002) 17–36
DNA ends of a single DSB interact with each other).In contrast to the restitution generated by HRR, therepair products usually contain small scale changesNHEJ and deletions SSA. To generate exchange typeCA, both NHEJ and SSA require at least two initialDSB (the four ends of two DSB interact cross-wisewith each other). The repair pathways mentioned be-fore compete actively for the repair of a DSB. Theregulation of these pathways (what circumstancesdetermine which pathway is used) is still enigmatic,but appears to occur at different levels during devel-opment and cell cycle progression[51]. Likewise, itis not known as to what extent a given DSB repairprocess contributes to the formation of a certain typeof CA.
It is important to note that, in principle, each of themechanisms shown inFig. 1is able to form any possi-ble exchange type CA. It is also important to note thatthe number of initial DSB required to induce an ex-change type CA depends on the mechanism involved.A single DSB is sufficient for homologous recombina-tion (HR) while at least two initial DSB are requiredfor NHEJ and SSA. The schemes and interpretationsshown in Fig. 1 are confined to CA formed with amaximum of two DSB. More than two DSB can beinvolved in the formation of CA by the DSB-repairmechanisms described, and it is imaginable that thesedifferent mechanisms can participate in the formationof complex aberrations (seeFig. 2 andFig. 3e and f).HR, the most accurate process, depends on the Rad52and Rad51 proteins and occurs usually between ex-tended regions of sequence homology located on sisterchromatids in cells undergoing mitotic cell cycles oron homologous chromatids in cells undergoing mei-otic cell cycles. If, however, HR occurs within regionsof sequence homology (e.g. repeats, pseudogenes)located on different chromosomes (ectopic HR), ex-change type CA like reciprocal translocations (Fig. 1(1)) or dicentrics and the corresponding fused acen-tric fragments (Fig. 1 (2)) can arise. In the examplesgiven, a single initial DSB is created within a repeatsequence (red box) on the green chromosome. Thesubsequent steps are shown in the double-strand-breakrepair (DSBR) model on the left side (DNA duplexesrepresented by two lines with arrowheads pointing in3′-direction; centromeres by vertical bars). The result-ing DSB-ends are 5′–3′exonucleolytically resected toproduce long 3′-single-stranded tails that invade the
blue chromosome at a site of a homologous DNAsequence (red hatched box). The 3′-ends of the in-vading strands serve as primers for semi-conservativerepair synthesis (black dotted arrows) of several kilo-bases length so that one newly synthesized strand ispresent in each of the donor and recipient. The re-sulting joint molecule contains a heteroduplex regionbordered by two Holliday junctions that are endonu-cleolytically resolved to yield (as shown inFig. 1 (1))reciprocal crossover or non-crossover products (notshown). Depending on the original orientation of thesequence repeat (arrows above red boxes) in the blueand green chromosomes, reciprocal translocations ordicentrics can arise. NHEJ requires two DSB to yieldan exchange type CA but is independent of sequencehomology. In the non-reciprocal example shown inFig. 1 (3), the green centromere-containing fragmentfuses with the blue centromere-containing fragmentto yield a dicentric while the two acentric fragmentsdo not fuse so that breaks arise. For this, the originalDSB-ends have to be enzymatically modified (fill-inDNA synthesis, exonucleolytic trimming) to form aligatable structure. These steps are enhanced by theKu70/Ku80 heterodimer complex. It is important tonote that the reciprocal event (dicentric plus fusedacentric fragment) or the formation of a reciprocaltranslocation is possible as well. InFig. 1 (4), a singleDSB occurs between two direct repeat sequences onthe green chromosome and is processed by the Rad52-and homology-dependent SSA mechanism: extensive5′–3′-exonucleolytic resection of the DSB-ends yieldslong 3′-tails that anneal at regions of sequence homol-ogy. Unpaired ends are nucleolytically removed andgaps filled. The resulting intrachromosomal deletioncomprises one copy of the repeat and the interveningsequence. The formation of exchange-type CA bySSA (not shown) would require two initial DSB (asshown for NHEJ) which must occur within or nearregions of sequence homology on two chromosomes.
In most cases, at least as long as the number of ini-tial breaks remains small, DSB are likely to be repairedcorrectly or lead to small scale DNA alterations in therange of a few base pairs or kilobases which can beanalyzed only by restriction mapping or sequencing.In some cases, DSB may lead to large scale alterationsvisible as CA under the light microscope. Therefore,CA are not special phenomena resulting from spe-cific cellular activities, but are just the microscopically
G. Obe et al. / Mutation Research 504 (2002) 17–36 21
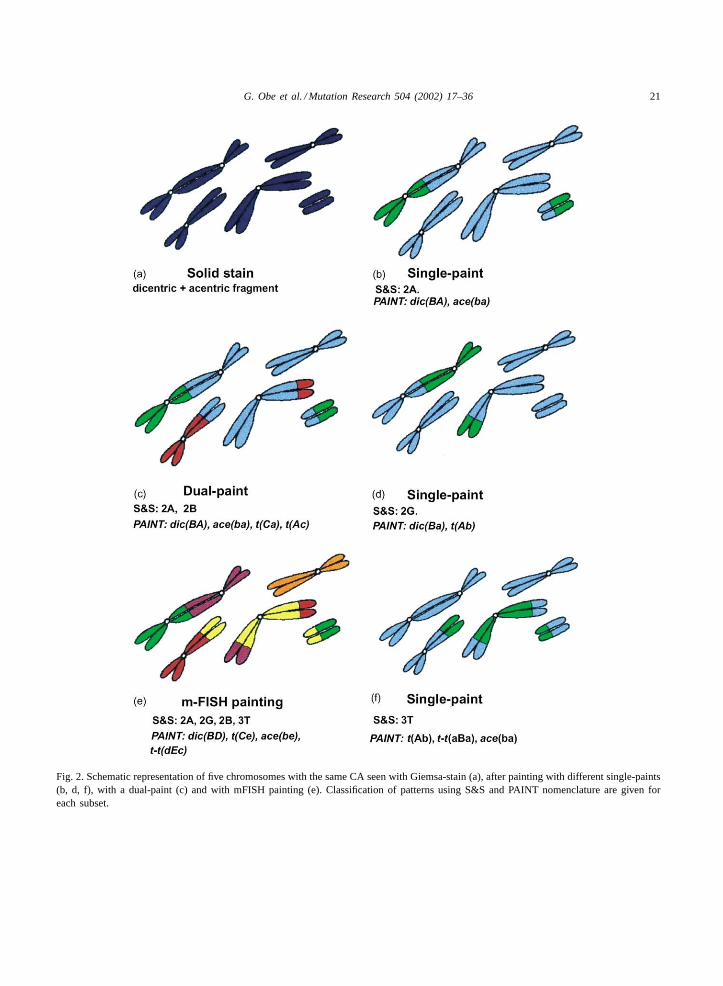
Fig. 2. Schematic representation of five chromosomes with the same CA seen with Giemsa-stain (a), after painting with different single-paints(b, d, f), with a dual-paint (c) and with mFISH painting (e). Classification of patterns using S&S and PAINT nomenclature are given foreach subset.
22 G. Obe et al. / Mutation Research 504 (2002) 17–36
Fig. 3.

G. Obe et al. / Mutation Research 504 (2002) 17–36 23
visible part of a wide spectrum of products generatedby different DSB repair mechanisms.
Microscopical analyses of CA depend on stainingof chromosomes. Especially, the methodology of fluo-rescence in situ hybridisation (FISH) uncovered unex-pected complexities of CA and this will be discussedin Section 3.
3. The impact of FISH-painting on the scoring andinterpretation of chromosome-type aberrations
For many decades, studies of CA, and the theo-ries derived from them, have been based primarilyupon solid-stained microscopical preparations. Onlyasymmetrical forms of CA which give rise to acentricfragments [52] are readily visible in solid-stainedpreparations, and it seems that the dicentric shownin Fig. 2ais directly related to the acentric fragment,and has arisen from a simple, reciprocal, pairwiseexchange following irradiation in G1 (a cyclical ex-change of order 2, or “c2”[53,54]. Solid-stainedchromosome-type aberrations almost always lead tothis conclusion, so that classification, interpretation,and foundational quantitative theory are all basedupon the assumption of an almost exclusive domi-nance of c2 exchanges.
Studies of chromatid-type aberrations indicate thatmany exchanges are “complex”, involving multiplebreaks and/or chromosomes[52]. Such events were
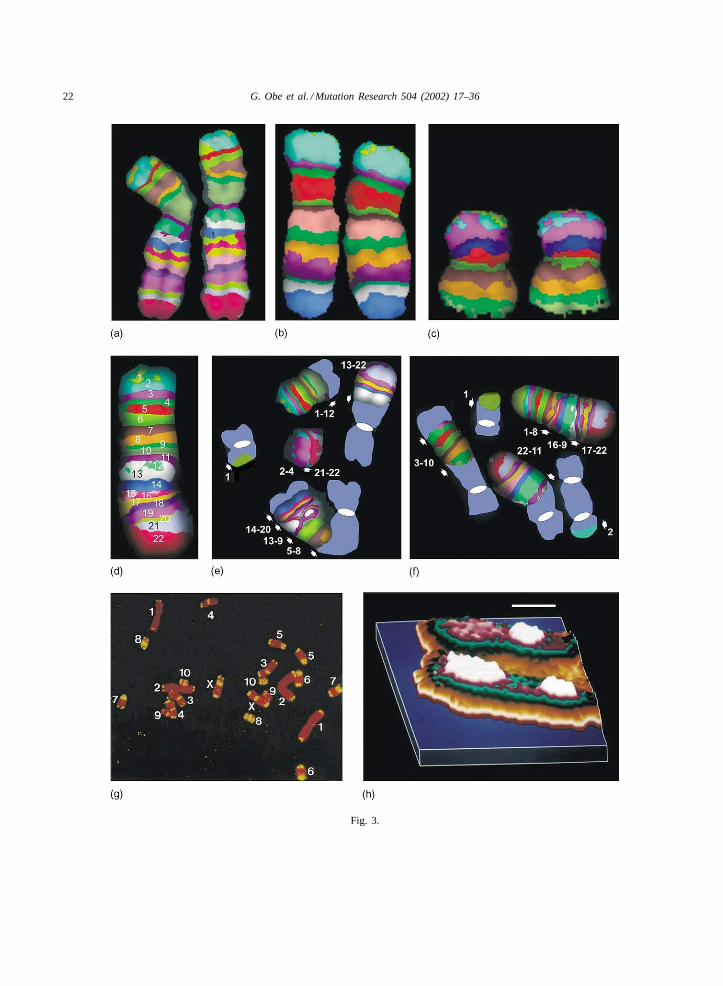
�Fig. 3. (a–c) Human chromosomes 2 (a), 6 (b), and 16 (c) showing pseudocolor bands following high resolution multi-color bandingfluorescence in situ hybridization (images kindly provided by Dr. Ilse Chudoba, MetaSystems, Germany); (d) human chromosome 5 with22 pseudocolor bands indicated by numbers, the centromere corresponds to band 7; (e–f) damaged metaphases following treatment ofhuman G1 lymphocytes with 30 unitsAluI, breakpoints are indicated by arrows. The numbers correspond to the bands shown in (d).DAPI-stained chromosomes are shown in blue with centromeres as white ovals; (e) the banded chromosome in the upper right corneris involved in a reciprocal translocation. The other banded chromosome was apparently broken into six pieces. The p-terminal piece (1)fused with a DAPI-stained chromosome, two pieces (2–4; 21–22) form an acentric fragment. The complex dicentric chromosome containsthree fragments of chromosome 5 with an inverted segment in the middle (14–20; 13–9; 5–8); (f) One chromosome 5 has a paracentricinversion (16–9). The other chromosome 5 apparently broke into four pieces resulting in two exchanges (1 and 22–11) one dicentricchromosome with an inserted fragment of chromosome 5 (3–10) and another dicentric chromosome with a small piece of chromosome 5(2) at one end; (g) metaphase of Chinese hamster primary fibroblast. Telomeres are labeled with a pantelomeric probe (yellow), counterstain is propidium iodide (red). Apart from the telomeres at the ends of the chromosomes, interstitial telomeric sequences occur in mostchromosomes. Chromosome pairs are marked with numbers 1–10, the X-chromosomes are marked with the letter X; (h) example of aminute subtelomeric exchange (t-SCE) detected in a CHO chromosome illustrated as a pseudo-third-dimension graphic image. Only thesubtelomeric region of a chromosome is shown. A Zeiss MSP65 microphotometer associated to a DEC Micro VAX 3300, a TEK 4211graphic terminal and application software were used for scanning Giemsa stained T-banded subtelomeric segments. Density measurementswere sorted using an interval limit of 5 absorbance units per class. Data obtained were interpolated assigning false colors to each thresholdlimit and amplified linearly to generate graphic images (for further technical details see[189]). This procedure allowed the detection ofsmall variations of the distribution of chromatin densities. White color denotes the highest densities detected by the system. Bar= 1.0�m.
deemed to be rare for chromosome-type changes, andfew are seen even with the application of chromosome-banding techniques. However, in retrospect, earlywork from Drosophila [55–57] and from clinicalfindings [13,14,58]should have made us think morecarefully.
When fluorescence in situ hybridization methodsusing chromosome-specific probes (“FISH-painting”)[59–64]began to be applied to induced chromosome-type aberrations, the picture changed and the olderaberration scoring systems[52] proved to be inade-quate.The positional arrangement of postulated initialbreaks within chromosomes can be used to classifypotential complex exchange configurations into fam-ilies by the CAB system (CAB: number of chromo-somes, number of arms, number of breaks)[65,66].Random rejoining interactions between the break-endsproduce many different exchange configurations fromeach family, each of which can generate a number ofdifferent single-paint patterns. Classification of thesepatterns forms the basis for the S&S nomenclaturesystem that can be used for scoring[65,67–69].
The alternative scoring nomenclature system,“PAINT” [69,70] treats each painted element in iso-lation, assigning the rejoining events to one of fivecategories, and also provides a very versatile schemefor descriptive mapping of each element. Descriptionsof patterns by both systems are used inFig. 2.
In the case of complex configurations (defined asthree or more breaks in two or more chromosomes),

24 G. Obe et al. / Mutation Research 504 (2002) 17–36
a variety of different single-paint patterns can arisefrom the same configuration, depending upon the posi-tion occupied by the painted participant. And becausethere are a limited number of possible patterns, differ-ent configurations can generate the same single-paintpattern[65,68,70].
Therefore, when only one, or a few chromosomesare painted, false inferences can be drawn about thedegree of complexity (or otherwise) of the underly-ing exchange. Conversely, it is seldom possible from asingle-paint pattern, to assign the CAB family of ori-gin. Solid-staining inFig. 2ashows a simple two-break(c2) asymmetrical interchange, “dicentric+ acentricfragment”.
If with a single-chromosome FISH protocol, oneof the dicentric participants is painted (Fig. 2b), thepattern (S&S 2A) is precisely that expected for a c2dicentric, and, taken alone, confirms our inference.Multi-color protocols have been developed, two tothree homologue pairs being painted in distinctivecolors, often with the addition of a pan-centromericprobe. These protocols increase the number of aber-rations recovered and can enhance the accuracy ofinterpretation[64,71–82]. Fig. 2cshows a result froma dual-paint protocol, and now, by chance, a sym-metrical interchange between two of the other chro-mosomes (a “reciprocal translocation”, S&S 2B) isdetected suggesting again a simple c2 exchange. Themajority of symmetrical events are not seen with solidstaining, yet these are the types that will be transmit-ted to future cell generations, and are therefore mostlikely to be of genetical significance. It is this effi-ciency to detect symmetrical, transmitted events thatis making FISH so useful in the fields of long-termretrospective dosimetry[83–95], and of clinical andcancer cytogenetics[96–99].
If the initial single-paint had colored the other par-ticipant in the dicentric (Fig. 2d) we find that its termi-nal segment has gone to another centric chromosome,and not to the fragment. Formally, such an event re-quires the interaction of at least three breaks (a c3exchange), so this signal (S&S 2G) indicates that theunderlying exchange configuration is “complex”. The2A pattern ofFig. 2b has given us a false signal; itis a pseudosimple pattern[66,82]. In critical quanti-tative work concerning aberration mechanistics, it isnecessary to correct for suchpseudosimple patterns[69,100].
Anomalous single-paint patterns of many kinds arerelatively common at radiation doses used in cytoge-netic experiments, increasing in number and varietyas dose or LET rises[66,76,100–111]. Because suchanomalous patterns are not seen with solid stain-ing, it is obvious that for many years we have beenunder-estimating the cytogenetic damage that radia-tion causes, and of course this will have had an effectupon our theoretical ideas about aberration formation.
These data show that the underlying exchange con-figuration is “simple” or “complex”, not the pattern.The pattern is only a signal that alerts us (or fails toalert us) to the status of the exchange from which itwas generated.
Relatively recently, it has become possible withthe aid of differential fluorochrome mixes and so-phisticated computer software to paint (pseudocolor)distinctively all the chromosomes in the karyotype[112–117]. So, for the first time, we can actually viewthe exchange configuration which is generating thesingle-paint patterns. This removes much (but not all)of the ambiguity of interpretation which arises fromsingle- and multi-paint protocols. It has, however, in-troduced a number of other problems, not least theclassification of actual configurations, which we havenot faced before[53].
Fig. 2e shows a “multiplex-FISH” (m-FISH) pic-ture of our chromosome group, and it can be seen atonce that the situation is even more complicated thandeduced from previous paintings. Using S&S nomen-clature the configuration comprises a group of foursingle-paint patterns, 2A/2B/2G/3T. Simplistically,this suggests that it originates from the interactionof five breaks in four chromosomes, probably com-plex family CAB 4/5/5. Of the 544 possible rejoininginteractions (restitution excluded) amongst the 10break-ends, just 24 will give this collection of patterns.However, 12 of these involve all 10 ends in a single ex-change cycle (an irreducible c5). The other 12 can beresolved, visually, into two cycles, a c2 and a c3. Suchreducible complex patterns are termed “sequential ex-change complexes”[53,82,118]. Whilst these consti-tute genuine random possibilities from a c5 exchange,they could also have arisen from two space-separated,independent exchanges, but there is no way to de-cide which is the actual origin. The relevance ofthe distinction lies in the fact that in the irreduciblec5 situation, the red/yellow 2B is apseudosimple,

G. Obe et al. / Mutation Research 504 (2002) 17–36 25
but in the c2+ c3 case, it is a truly simple (c2) recip-rocal translocation. These formation-mode problemsare an inherent feature of all configurations derivedfrom complex-generating families≥C/(A/4).
Interestingly, had the single-paint used happened tocorrespond to the yellow chromosome (Fig. 2f) theS&S 3T pattern would have signaled that a minimumof five breaks was involved in the exchange, and thatthe underlying configuration belongs to eitherCABfamily 3/5/5, 4/5/5, or even higher. Comparison ofpanels (b), (d) and (f) emphasizes the limitations andcare needed in interpreting patterns when only a fewchromosomes are painted[68].
FISH-painting has thrown up a further anomalynot encountered in solid-staining studies, namelythe “one-way” exchange (also called “non-reciprocaltranslocations”, “incomplete exchanges”, “terminaltranslocations”[70]. These patterns are “incomplete”in the sense that certain expected segments appear tobe missing. With single-paint protocols, there are sixfrequently recurring patterns (I → VI [100]) plus afew rarer types. At reasonable radiation dose levels,one-way exchanges constitute typically∼20–30% oftotal exchange patterns, and currently, this frequencyappears to be independent of dose or radiation quality.
Initially, it was thought that these represented ex-changes where some segments had failed to join up(i.e. “incomplete” in the accepted cytogenetic defi-nition [52]), but careful work using telomere probeshas shown that the majority arise from complete ex-changes where a terminal segment is so small that itdoes not register as a distinct visible signal[119–128].As inferred from earlier studies[52] incompleteness israre for chromosome-type aberrations (2–6%). This isin sharp contrast to chromatid-type exchanges whereincompleteness of exchanges is between 30 and 50%.
The frequency and degree of chromosome involve-ment in complex exchanges has surprised everyone,and raised a host of problems regarding mechanismsof formation. How and where within the nucleus doso many chromosome regions and lesions come to-gether? The situation is exacerbated by the findingsthat the nucleus has a very compartmentalized archi-tecture, chromosome arms occupying very localized“domains” [52,129–137]. So chromosome “painting”has not only become a useful tool, but has given freshimpetus to re-examine the foundations of our think-ing about the origin of CA. This section shows that
if we paint DNA molecules participating in CA withdifferent colors we may get different types of CA.
FISH studies discussed so far cannot be used toanalyze paracentric inversions, transpositions withinthe same chromosome, longitudinal orientation ofinserted segments and homologue/homologue events.G-banding would allow to do this to a certain extent,but the analysis is difficult and needs great experi-ence. The methodology of m-BAND in which singlechromosomes are covered with pseudocolored trans-verse bands is well suited to recognize such CA types[138,139]. Fig. 3a–f shows examples of differentm-banded human chromosomes and of chromosome5 involved in complex aberrations induced with therestriction endonucleaseAluI.
Not only frequencies and types of CA but alsointer- and intrachromosomal distributions of break-points have been intensively studied. This aspect isdiscussed inSection 4.
4. Intra- and interchromosomal distribution ofbreakpoints
The intrachromosomal distribution of CA break-points in plant and animal (including human) cellsis not always random[140–156]. Results of intra-chromosomal localization of “breakpoints” are diffi-cult to interpret.
Firstly, we are only able to observe and score aselection of the original number, the residue that hasbeen “fixed” in visible aberrations. It is by no meanscertain that these represent an unbiased sample withrespect to positioning. As a constant reminder ofthis fact, it has been proposed that aberration derivedbreakpoints be termed “selected observed residualbreakpoints” (SORB)[157].
Secondly, metaphase chromosomes are highly com-pacted and coiled structures (Section 1) and the con-densation process, which is often differential along thechromosome, may alter the observed position relativeto that in the extended interphase chromatin, wherethe aberration was originally formed.
Thirdly, with respect tochromosome-type (but notchromatid-type) aberrations observed in G-bandedchromosomes, there is an inherent pattern recognitionartifact (the “three-band uncertainty”) which biasesplacement of the SORB to a pale band[158]. This,

26 G. Obe et al. / Mutation Research 504 (2002) 17–36
coupled with the variable efficiency with which dif-ferent kinds of aberrations can be detected withindifferent banded regions[159] introduces further biasinto SORB location.
Fourthly, primary lesions are handled by differentrepair pathways which rarely give rise to CA and bythis influence the location of SORB.
The discussion shows that SORB are at best afar-away echo of the location of primary lesions.
Our own analyses showed that in G-banded Chinesehamster ovary (CHO) cells about 75% of SORB inchromosome and chromatid aberrations induced by therestriction endonucleasesAluI and BamHI as well asof chromatid type aberrations induced by DNaseI oc-curred in Giemsa-light bands. Breakpoint clusters pro-duced by these endonucleases showed co-localizationin specific G-light bands of CHO chromosomes, theheterochromatic long arm of the X chromosome wasnearly devoid of breakpoints[160–162]. Exposure ofmetaphase spreads to these enzymes led to G-bandpatterns after Giemsa staining, but no endonucleasedigestion was evident in the heterochromatin of theX chromosome[162]. It is possible that SORB posi-tions reflect a better access of the enzymes to nucleasesensitive sites in G-light bands which are preferen-tially digested by the enzymes even in fixed metaphasepreparations. SORB induced by neutrons and�-raysin CHO chromosomes were also not randomly dis-tributed, more than 60% occurred in G-light bandsand major breakpoint clusters co-localized with thoseproduced by endonucleases[163]. Following exposureto ionizing radiation energy deposition per unit vol-ume should be random in the nucleus. However, itcan be expected that less DSB are induced in highlycondensed as compared to loose chromatin, becausethe DNA in the former will be more protected fromradiation-induced radicals. This could be one explana-tion for the higher density of radiation-induced SORBin G-light, as compared to G-dark bands.
Of the various properties relating chromatin struc-ture and chromosome organization to the distributionof SORB the visually most striking is the close corre-spondence with sites of hyperacetylation observed inCHO chromosomes[164].
There is an intimate relationship between histoneacetylation status of chromatin and its transcriptionalactivity [165,166]. Acetylation of histones by histoneacetyltransferases (HATs) is in dynamic equilibrium
with deacetylation, mediated by histone deacetylases(HDACs) [167] and many protein modifiers of tran-scription have been shown to possess either HAT orHDAC components (for a review see[168]). SeveralDNA methylation-dependent transcriptional repres-sors appear to be multi-subunit complexes that, inaddition to binding specifically to methylated CpGsequences, also contain HDACs[169–172].
Transcriptionally active chromatin has a moreaccessible structure than transcriptionally inactivechromatin, as indicated by its increased sensitivity tonuclease digestion[162,173]. Current ideas on chro-matin remodeling during transcriptional activation en-visage conversion of the highly ordered and compact30 nm fiber, thought to be formed from a solenoidalarrangement of six to seven nucleosomes per heli-cal turn and including histone H1 linkers[174], toan open ‘beads-on-a-string’ 10 nm fiber, lacking H1,that allows progression of RNA polymerase duringtranscription.
During interphase, transcribed and potentially tran-scribable (poised) chromatin (henceforth collectivelyreferred to as active chromatin) have both a more openconformation and a higher level of histone acetyla-tion than transcriptionally silent, or ‘heterochromatic’chromatin. When antibodies recognizing acetylatedisoforms of histone H4 were used to investigate thenuclear distribution of acetylated chromatin by im-munofluorescence in situ[175,176], it was found, per-haps surprisingly, that hyperacetylated H4 was presentin transcriptionally inactive metaphase chromosomes.The distribution of acetylation is not uniform, butreflects almost exactly the distribution of activelytranscribed DNA during interphase[176–178]. Theproduction of G-band patterns in metaphase chro-mosomes exposed to endonucleases also shows thatcharacteristics of interphase chromatin can be tracedto metaphase chromosomes[162].
Principally, the same problems discussed with re-spect to the intrachromosomal distribution of SORB,pertain to their interchromosomal distribution. It hasbeen shown repeatedly that the interchromosomaldistributions of SORB induced by low LET radi-ation in presynthetic human lymphocytes in vitroare not random[179–181]. One reason for devia-tions from randomness may be non-random arrange-ments of chromosomes in the interphase nucleus[129,134,182,183].

G. Obe et al. / Mutation Research 504 (2002) 17–36 27
The interchromosomal distribution of SORB ofchromatid-type aberrations induced by low LET ra-diation in S-phase CHO cells showed interestingdeviations from randomness. In the majority of chro-mosomes the distribution of SORB was as expectedaccording to chromosome length for simple breaks orfor relative corrected lengths in exchanges calculatedaccording to Savage and Papworth[157,184]. Chro-mosomes 2 and 8 were more often involved in exchan-ges than expected from randomness and Z3 showedmore chromatid breaks and interchanges[185].
Probably, chromosomes have different pheno-types with respect to their characteristics to formradiation-induced CA. Chromosomes 2 and 8 mayhave an “exchange” phenotype, chromosome Z3 a“break” and “exchange” phenotype[185].
In conclusion, it can be stated that deviations fromrandom intra- and interchromosomal distributions ofSORB reflect the complex interplay of primary lesionsin chromosomal DNA, structure and intranuclear ar-rangement of chromosomes at the time of exposure toa chromosome damaging agent and different types ofDSB repair activities. Telomeres, interstitial telomererepeat like sequences and subtelomeric regions seemalso to play important roles in the formation of CAand this will be discussed inSection 5.
5. Subtelomeric segments and telomeres
The incubation of chromosome preparations in abuffered salt solution at high temperature followed byGiemsa staining produces darkly stained subtelomericsegments (T-bands) with the rest of the chromosomelightly stained[186]. Electron microscopic analysesshowed that T-banded sections of human chromo-somes are intricate fibrous structures[187]. T-bandsare a subset of GC-rich R-bands highly resistant toheat denaturation and to digestion with the restrictionendonucleaseMseI [188].
Scanning microphotometer systems and applica-tion software were used to generate computer graphicimages allowing to obtain quantitative data on thestructure of nuclei and chromosomes at the light mi-croscopical level[189,190]. We used this system tostudy T-banded chromosome segments and detecteddifferential distributions of high-density chromatin inthe chromatids of these regions[191,192].
In some chromosomes high density areas in sisterchromatids were arranged like sister chromatid ex-changes and these were calledt-SCE (Fig. 3h) [193].The density patterns and the minutet-SCE may resultfrom characteristic DNA-protein complexes in T-bandareas of the chromosomes[194]. These findings fitto the idea that subtelomeric regions of chromosomesseem to be hot spots for the formation of symmetricalexchanges (SCE) which are not visible in the normallight microscope[195].
Cryptic aberrations in subtelomeric chromosomalregions have been shown to be associated with severalhuman congenital abnormalities[196–199].
Since eukaryotic chromosomes are linear, theyhave two ends with special structures called telomereswhich guarantee DNA synthesis at the ends of chro-mosomal DNA[200–205]and seem to play an im-portant role in the formation of CA. The mammaliantelomere is composed of a TG-rich double strandedregion with a long 3′-extension of (T2AG3)n repeats.According to proposed models, the 3′ tail displacesparts of the double stranded TG-rich sequence andforms a so called t-loop. Telomeres are stabilized byseveral telomere specific proteins including telomererepeat binding factors TRF1 and TRF2[206,207].Disruption of this structure, for example by over-expression of a dominant negative TRF2 protein,leads to DNA damage and to end-to-end fusions ofchromosomes[208,209].
In contrast to these t-loop-guarded telomere ends,ends created by DSB are recombinogenic and aretherefore used as substrates for DSB repair mecha-nisms (seeSection 1). In addition, when DSB occurat interstitial telomeric repeat arrays, they can bestabilized by de novo synthesis of telomeric repeats[210–213]. It is supposed that the formation of newtelomeric sequences protects DSB from nucleolyticdegradation in a similar way to that which naturaltelomeres do[214]. The rather common “one-wayexchanges” in which telomeric regions are involvedare discussed inSection 3.
Apart from telomere specific proteins, DNA repairproteins are also associated with telomeres, and theseprovide the basis for a further mechanism, by whichtelomeres may interfere with DSB repair activities.In yeast, DNA repair proteins stored in telomeres aretranslocated to sites of DSB due to DNA damage[215,216]. In mammalian cells, localization of DNA

28 G. Obe et al. / Mutation Research 504 (2002) 17–36
repair proteins on telomeres has been shown as welland it appears that the intact telomere structure is re-quired to maintain chromosomal integrity by DSB re-pair [217].
Thus, apart from guaranteeing DNA synthesis atthe ends of chromosomal DNA, telomeres may havea dual role in (i) providing a stable chromosome endand (ii) saving the integrity of damaged chromosomalDNA by supplying stored DNA repair proteins.
In addition to the telomeric repeats at the ends ofthe chromosomes, rodent cells contain large intersti-tial telomere repeat-like sequences which have beenshown to be involved in chromosomal rearrangements[218–222]. Except for chromosomes 1 and 2, Chi-nese hamster chromosomes have interstitial telomericblocks near the centromeric regions (Fig. 3g). Theseregions tend to associate together in the interphase nu-cleus forming a chromocenter like organization. Thepresence of repetitive sequences and the proximityof such regions from different chromosomes promoteexchange aberration formation following treatmentwith ionizing radiation, short wave UV and restrictionendonucleases[223–226]. In addition, these types ofaberrations occur spontaneously during culture, lead-ing to amplification of telomeric repeats as well as tothe origin of marker chromosomes[226]. It has alsobeen demonstrated that the telomerase activity is up-regulated both in vitro (Chinese hamster cells, humantumor cell lines)[227,228] and in vivo [201]. Thiseffect should have consequences for repair of DNAdamage as well as formation of aberrations in theseregions.
6. Concluding remarks
In conclusion it can be said that CA are microscop-ically visible changes in the single DNA molecules ofchromosomes and chromatids and therefore can be re-garded as “a kind of visible chemistry” in the sense ofDarlington’s statement given at the beginning of thisreview.
References
[1] C.D. Darlington, The problem of chromosome breakage: anintroduction, Heredity 6 (1953) V–VIII.
[2] S. Bonassi, A. Abbondandolo, L. Camurri, L. Dal Pra, M.De Ferrari, F. Degrassi, A. Forni, L. Lamberti, C. Lando,P. Padovani, Are chromosome aberrations in circulatinglymphocytes predictive of future cancer onset in humans?Preliminary results of an Italian cohort study, Cancer Genet.Cytogenet. 79 (1995) 133–135.
[3] S. Bonassi, L. Hagmar, U. Stromberg, A.H. Montagud,H. Tinnerberg, A. Forni, P. Heikkila, S. Wanders, P.Wilhardt, I.-L. Hansteen, L.E. Knudson, H. Norppa,Chromosomal aberrations in lymphocytes predict humancancer independently of exposure to carcinogens, CancerRes. 60 (2000) 1619–1625.
[4] L. Hagmar, A. Brogger, I.L. Hansteen, S. Heim, B. Hogstedt,L. Knudsen, B. Lambert, K. Linnainmaa, F. Mitelman, I.Nordenson, Cancer risk in humans predicted by increasedlevels of chromosomal aberrations in lymphocytes: nordicstudy group on the health risk of chromosome damage,Cancer Res. 54 (1994) 2919–2922.
[5] L. Hagmar, S. Bonassi, U. Stromberg, A. Brogger,L.E. Knudsen, H. Norppa, C. Reuterwall, Chromosomalaberrations in lymphocytes predict human cancer: a reportfrom the European Study Group on Cytogenetic Biomarkersand Health (ESCH), Cancer Res. 58 (1998) 4117–4121.
[6] L. Hagmar, S. Bonassi, U. Stromberg, Z. Mikoczy, C.Lando, I.L. Hansteen, A.H. Montagud, L. Knudsen, H.Norppa, C. Reuterwall, H. Tinnerberg, A. Brogger, A.Forni, B. Hogstedt, B. Lambert, F. Mitelman, I. Nordenson,S. Salomaa, S. Skerfving, Cancer predictive value ofcytogenetic markers used in occupational health surveillanceprograms: a report from an ongoing study by the EuropeanStudy Group on Cytogenetic Biomarkers and Health, Mutat.Res. 405 (1998) 171–178.
[7] F. Mitelman, F. Mertens, B. Johansson, A breakpoint map ofrecurrent chromosomal rearrangements in human neoplasia,Nat. Genet. 15 (1997) 417–474.
[8] J.R. Savage, Insight into sites, Mutat. Res. 366 (1996) 81–95.
[9] M. Durante, S. Bonassi, K. George, F.A. Cucinotta, Riskestimation based on chromosomal aberrations induced byradiation, Radiat. Res. 156 (2001) 2622–2627.
[10] IAEA. Biological Dosimetry: Chromosomal AberrationAnalysis for Dose Assessment, Vol. 260, InternationalAtomic Energy Agency, Vienna, 1986.
[11] M. Bauchinger, Retrospective dose reconstruction of humanradiation exposure by FISH/chromosome painting, Mutat.Res. 404 (1998) 89–96.
[12] A.T. Natarajan, G. Obe, Biological dosimetry of absorbedradiation dose based on the frequencies of chromosomalaberrations in human lymphocytes, in: C. Baumstark-Khan,S. Kozubek, G. Horneck (Eds.), Fundamentals for theAssessment of Risks from Environmental Radiation, KluwerAcademic Publishers, Dordrecht, 1999, pp. 179–186.
[13] R.J.M. Gardner, G.R. Sutherland, Chromosome Abnor-malities and Genetic Counseling, Oxford University Press,New York, 1996.
[14] O.J. Miller, E. Therman, Human Chromosomes, Springer,New York, 2001.

G. Obe et al. / Mutation Research 504 (2002) 17–36 29
[15] M. Ishidate, K.F. Miura, T. Sofuni, Chromosome aberrationassays in genetic toxicology testing in vitro, Mutat. Res.404 (1998) 167–172.
[16] D. Kirkland, Chromosome aberration testing in genetictoxicology-past, present and future, Mutat. Res. 404 (1998)173–185.
[17] L.H. Caporale (Ed.), Molecular Strategies in BiologicalEvolution, Ann. NY Acad. Sci. 870, 1999.
[18] E.J. DuPraw, DNA and Chromosomes, Holt, Rinehart andWinston, New York, 1970.
[19] R. Kavenoff, L.C. Klotz, B.H. Zimm, On the nature ofchromosome-sized DNA molecules, in: Proceedings of theCold Spring Harbour Symposium on Quantitative Biology,Vol. XXXVIII, 1974, pp. 1–8.
[20] K.E. van Holde, Chromatin, Springer, New York, 1989.[21] A. Wolffe, Chromatin, Structure and Function, Academic
Press, New York, 1998.[22] A.T. Natarajan, Molecular aspects of the origin of
chromosome structural changes, Biol. Zbl. 95 (1976) 139–156.
[23] J.J. Roberts, The repair of DNA modified by cytotoxic,mutagenic, and carcinogenic chemicals, in: J.T. Lett, H.Adler (Eds.), Advances in Radiation Biology, AcademicPress, New York, 1978, pp. 211–442.
[24] B. Singer, D. Grunberger, Molecular Biology of Mutagensand Carcinogens, Plenum Press, New York, 1983.
[25] P. Bryant, The signal model: a possible explanation forthe conversion of DNA double-strand breaks into chromatidbreaks, Int. J. Radiat. Biol. 73 (1998) 243–251.
[26] A.T. Natarajan, G. Obe, Molecular mechanisms involvedin the production of chromosomal aberrations. Part I.Utilization of neurospora endonuclease for the study ofaberration production in G2 stage of the cell cycle, Mutat.Res. 52 (1978) 137–149.
[27] G. Obe, C. Johannes, D. Schulte-Frohlinde, DNA double-strand breaks induced by sparsely ionizing radiation andendonucleases as critical lesions for cell death, chromosomalaberrations, mutations and oncogenic transformation,Mutagenesis 7 (1992) 3–12.
[28] P. Pfeiffer, W. Goedecke, G. Obe, Mechanisms of DNAdouble-strand break repair and their potential to inducechromosomal aberrations, Mutagenesis 15 (2000) 289–302.
[29] K.W. Caldecott, Mammalian DNA single-strand breakrepair: an X-ra(y)ted affair, Bioessays 23 (2001) 447–455.
[30] G.L. Dianov, P. O’Neill, D.T. Goodhead, Securinggenome stability by orchestrating DNA repair: removal ofradiation-induced clustered lesions in DNA, Bioessays 23(2001) 745–749.
[31] R. Kooistra, A. Pastink, J.B. Zonneveld, P.H. Lohman, J.C.Eeken, The Drosophila melanogaster DmRAD54 gene playsa crucial role in double-strand break repair after P-elementexcision and acts synergistically with Ku70 in the repair ofX-ray damage, Mol. Cell. Biol. 19 (1999) 6269–6275.
[32] A. Errami, V. Smider, W.K. Rathmell, D.M. He, E.A.Hendrickson, M.Z. Zdzienicka, G. Chu, Ku86 defines thegenetic defect and restores X-ray resistance and V(D)Jrecombination to complementation group 5 hamster cellmutants, Mol. Cell. Biol. 16 (1996) 1519–1526.
[33] U. Grawunder, D. Zimmer, P. Kulesza, M.R. Lieber,Requirement for an interaction of XRCC4 with DNAligase IV for wild-type V(D)J recombination and DNAdouble-strand break repair in vivo, J. Biol. Chem. 273 (1998)24708–24714.
[34] L.J. Kienker, E.K. Shin, K. Meek, Both V(D)J recombinationand radioresistance require DNA-PK kinase activity, thoughminimal levels suffice for V(D)J recombination, Nucl. AcidsRes. 28 (2000) 2752–2761.
[35] J.P. Manis, Y. Gu, R. Lansford, E. Sonoda, R. Ferrini, L.Davidson, K. Rajewsky, F.W. Alt, Ku70 is required for lateB cell development and immunoglobulin heavy chain classswitching, J. Exp. Med. 187 (1998) 2081–2089.
[36] A. Bergerat, B. de Massy, D. Gadelle, P.C. Varoutas, A.Nicolas, P. Forterre, An atypical topoisomerase II fromArchaea with implications for meiotic recombination, Nature386 (1997) 414–417.
[37] S. Keeney, N. Kleckner, Covalent protein–DNA complexesat the 5′ strand termini of meiosis-specific double-strandbreaks in yeast, Proc. Natl. Acad. Sci. U.S.A. 92 (1995)11274–11278.
[38] S. Keeney, C.N. Giroux, N. Kleckner, Meiosis-specific DNAdouble-strand breaks are catalyzed by Spo11, a member ofa widely conserved protein family, Cell 88 (1997) 375–384.
[39] T.G. Newcomb, L.A. Loeb, Oxidative DNA damage andmutagenesis, in: J.A. Nickoloff, M.F. Hoekstra (Eds.), DNADamage and Repair, Humana Press, Totowa, New Jersey,1998, pp. 65–84.
[40] R.D. Johnson, N. Liu, M. Jasin, Mammalian XRCC2promotes the repair of DNA double-strand breaks byhomologous recombination, Nature 401 (1999) 397–399.
[41] F. Liang, M. Han, P.J. Romanienko, M. Jasin, Homology-directed repair is a major double-strand break repair pathwayin mammalian cells, Proc. Natl. Acad. Sci. U.S.A. 95 (1998)5172–5177.
[42] M.E. Moynahan, J.W. Chiu, B.H. Koller, M. Jasin, Brca1controls homology-directed DNA repair, Mol. Cell 4 (1999)511–518.
[43] A.J. Pierce, R.D. Johnson, L.H. Thompson, M. Jasin,XRCC3 promotes homology-directed repair of DNA damagein mammalian cells, Genes Dev. 13 (1999) 2633–2638.
[44] M. Takata, M.S. Sasaki, E. Sonoda, C. Morrison, M.Hashimoto, H. Utsumi, Y. Yamaguchi-Iwai, A. Shinohara,S. Takeda, Homologous recombination and non-homologousend-joining pathways of DNA double-strand break repairhave overlapping roles in the maintenance of chromosomalintegrity in vertebrate cells, EMBO J. 17 (1998) 5497–5508.
[45] J.E. Haber, DNA recombination: the replication connection,Trends Biochem. Sci. 24 (1999) 271–275.
[46] J.E. Haber, Recombination: a frank view of exchanges andvice versa, Curr. Opin. Cell Biol. 12 (2000) 286–292.
[47] J.H. Hoeijmakers, Genome maintenance mechanisms forpreventing cancer, Nature 411 (2001) 366–374.
[48] K.K. Khanna, S.P. Jackson, DNA double-strand breaks:signaling, repair and the cancer connection, Nat. Genet. 27(2001) 247–253.

30 G. Obe et al. / Mutation Research 504 (2002) 17–36
[49] D.C. van Gent, J.H. Hoeijmakers, R. Kanaar, Chromosomalstability and the DNA double-stranded break connection,Nat. Rev. Genet. 2 (2001) 196–206.
[50] P. Karran, DNA double strand break repair in mammaliancells, Curr. Opin. Genet. Dev. 10 (2000) 144–150.
[51] J.E. Haber, Partners and pathways repairing a double-strandbreak, Trends Genet. 16 (2000) 259–264.
[52] J.R. Savage, Classification and relationships of inducedchromosomal structual changes, J. Med. Genet. 13 (1976)103–122.
[53] M.N. Cornforth, Analyzing radiation-induced complexchromosome rearrangements by combinatorial painting,Radiat. Res. 155 (2001) 643–659.
[54] R.K. Sachs, A.M. Chen, P.J. Simpson, L.R. Hlatky, P.Hahnfeldt, J.R. Savage, Clustering of radiation-producedbreaks along chromosomes: modelling the effects onchromosome aberrations, Int. J. Radiat. Biol. 75 (1999) 657–672.
[55] H. Bauer, Roentgenausloesung von Chromosomenmuta-tionen bei Drosophila melanogaster. Part I. Bruchhaeufigkeit:Verteilung und Rekombination nach Speicheldruesenun-tersuchung, Chromosoma 1 (1939) 343–389.
[56] H. Bauer, M. Demerec, B.P. Kaufmann, X-ray inducedchromosomal alterations in Drosophila melanogaster,Genetics 23 (1938) 610–630.
[57] B.P. Kaufmann, A complex induced rearrangement ofDrosophila chromosomes and its bearing on the problem ofchromosome recombination, Proc. Natl. Acad. Sci. U.S.A.29 (1943) 8–12.
[58] D.S. Borgaonkar, R. Shafer, N. Reisor, Repository of HumanChromosomal Variants and Anomalies. An InternationalRegistry of Abnormal Karyotypes, Medical Center ofDelaware, Newark, USA, 1987.
[59] T. Cremer, S. Popp, P. Emmerich, P. Lichter, C. Cremer,Rapid metaphase and interphase detection of radiation-induced chromosome aberrations in human lymphocytes bychromosomal suppression in situ hybridization, Cytometry11 (1990) 110–118.
[60] J.W. Gray, J.N. Lucas, D. Pinkel, A. Awa, Structuralchromosome analysis by whole chromosome painting forassessment of radiation-induced genetic damage, J. Radiat.Res. (Tokyo) 33 (Suppl.) (1992) 80–86.
[61] J.W. Gray, D. Pinkel, J.M. Brown, Fluorescence in situhybridization in cancer and radiation biology, Radiat. Res.137 (1994) 275–289.
[62] S.S. Habeebu, D.H. Spathas, M.A. Ferguson-Smith,Non-radioactive in situ hybridization of DNA probes tochromosomes and nuclei. A comparison of techniques, Mol.Biol. Med. 7 (1990) 423–435.
[63] R.R. Swiger, J.D. Tucker, Fluorescence in situ hybridization:a brief review, Environ. Mol. Mutagen. 27 (1996) 245–254.
[64] H.U. Weier, J.N. Lucas, M. Poggensee, R. Segraves,D. Pinkel, J.W. Gray, Two-color hybridization with highcomplexity chromosome-specific probes and a degene-rate alpha satellite probe DNA allows unambiguousdiscrimination between symmetrical and asymmetricaltranslocations, Chromosoma 100 (1991) 371–376.
[65] J.R. Savage, P.J. Simpson, FISH painting patterns resultingfrom complex exchanges, Mutat. Res. 312 (1994) 51–60.
[66] P.J. Simpson, J.R. Savage, Estimating the true frequencyof X-ray-induced complex chromosome exchanges usingfluorescence in situ hybridization, Int. J. Radiat. Biol. 67(1995) 37–45.
[67] S. Knehr, H. Zitzelsberger, M. Bauchinger, FISH-basedanalysis of radiation-induced chromosomal aberrations usingdifferent nomenclature systems, Int. J. Radiat. Biol. 73(1998) 135–141.
[68] J.R. Savage, P. Simpson, On the scoring of FISH-paintedchromosome-type exchange aberrations, Mutat. Res. 307(1994) 345–353.
[69] J.R. Savage, J.D. Tucker, Nomenclature systems forFISH-painted chromosome aberrations, Mutat. Res. 366(1996) 153–161.
[70] J.D. Tucker, W.F. Morgan, A.A. Awa, M. Bauchinger, D.Blakey, M.N. Cornforth, L.G. Littlefield, A.T. Natarajan,C. Shasserre, A proposed system for scoring structuralaberrations detected by chromosome painting, Cytogenet.Cell Genet. 68 (1995) 211–221.
[71] M. Bauchinger, E. Schmid, H. Zitzelsberger, H. Braselmann,U. Nahrstedt, Radiation-induced chromosome aberrationsanalysed by two-colour fluorescence in situ hybridizationwith composite whole chromosome-specific DNA probesand a pancentromeric DNA probe, Int. J. Radiat. Biol. 64(1993) 179–184.
[72] J.W. Breneman, R.R. Swiger, M.J. Ramsey, J.L. Minkler,J.G. Eveleth, R.A. Langlois, J.D. Tucker, The developmentof painting probes for dual-color and multiple chromosomeanalysis in the mouse, Cytogenet. Cell Genet. 68 (1995)197–202.
[73] E. Gebhart, S. Neubauer, G. Schmitt, S. Birkenhake, J.Dunst, Use of a three-color chromosome in situ suppressiontechnique for the detection of past radiation exposure, Radiat.Res. 145 (1996) 47–52.
[74] K.L. Johnson, J.D. Tucker, J. Nath, Frequency, distributionand clonality of chromosome damage in human lymphocytesby multi-color FISH, Mutagenesis 13 (1998) 217–227.
[75] S. Knehr, H. Zitzelsberger, H. Braselmann, M.Bauchinger, Analysis for DNA-proportional distribution ofradiation-induced chromosome aberrations in various triplecombinations of human chromosomes using fluorescence insitu hybridization, Int. J. Radiat. Biol. 65 (1994) 683–690.
[76] S. Knehr, R. Huber, H. Braselmann, H. Schraube, M.Bauchinger, Multicolour FISH painting for the analysis ofchromosomal aberrations induced by 220 kV X-rays andfission neutrons, Int. J. Radiat. Biol. 75 (1999) 407–418.
[77] J.N. Lucas, T. Tenjin, T. Straume, D. Pinkel, D. Moore,M. Litt, J.W. Gray, Rapid human chromosome aberrationanalysis using fluorescence in situ hybridization, Int. J.Radiat. Biol. 56 (1989) 35–44.
[78] J.N. Lucas, M. Poggensee, T. Straume, Translocationsbetween two specific human chromosomes detected bythree-color chromosome painting, Cytogenet. Cell Genet. 62(1993) 11–12.

G. Obe et al. / Mutation Research 504 (2002) 17–36 31
[79] J.N. Lucas, R.K. Sachs, Using three-color chromosomepainting to test chromosome aberration models, Proc. Natl.Acad. Sci. U.S.A. 90 (1993) 1484–1487.
[80] E. Schmid, H. Schraube, M. Bauchinger, Radiation-inducedchromosome aberrations analyzed by fluorescence in situhybridization with a triple combination of composite wholechromosome-specific probes, Int. J. Radiat. Biol. 62 (1992)673–678.
[81] P.J. Simpson, J.R. Savage, Detecting hidden exchangeevents within X-ray-induced aberrations using multicolourchromosome paints, Chromosome Res. 3 (1995) 69–72.
[82] P.J. Simpson, D.G. Papworth, J.R. Savage, X-ray-inducedsimple, pseudosimple and complex exchanges involving twodistinctly painted chromosomes, Int. J. Radiat. Biol. 75(1999) 11–18.
[83] M. Bauchinger, K. Salassidis, H. Braselmann, A. Vozilova,S. Pressl, G. Stephan, G. Snigiryova, V.P. Kozheurov, A.Akleyev, FISH-based analysis of stable translocations in aTecha River population, Int. J. Radiat. Biol. 73 (1998) 605–612.
[84] J.L. Fernandez, A. Campos, V. Goyanes, C. Losada, C.Veiras, A.A. Edwards, X-ray biological dosimetry performedby selective painting of human chromosomes 1 and 2, Int.J. Radiat. Biol. 67 (1995) 295–302.
[85] P. Finnon, D.C. Lloyd, A.A. Edwards, Fluorescence insitu hybridization detection of chromosomal aberrations inhuman lymphocytes: applicability to biological dosimetry,Int. J. Radiat. Biol. 68 (1995) 429–435.
[86] L.G. Littlefield, A.F. McFee, S.I. Salomaa, J.D. Tucker,P.D. Inskip, A.M. Sayer, C. Lindholm, S. Makinen, R.Mustonen, K. Sorensen, M. Tekkel, T. Veidebaum, A.Auvinen, J.D. Boice, Do recorded doses overestimate truedoses received by Chernobyl cleanup workers? Results ofcytogenetic analyses of Estonian workers by fluorescence insitu hybridization, Radiat. Res. 150 (1998) 237–249.
[87] J.N. Lucas, A. Awa, T. Straume, M. Poggensee, Y. Kodama,M. Nakano, K. Ohtaki, H.U. Weier, D. Pinkel, J. Gray,Rapid translocation frequency analysis in humans decadesafter exposure to ionizing radiation, Int. J. Radiat. Biol. 62(1992) 53–63.
[88] A.T. Natarajan, S.J. Santos, F. Darroudi, V. Hadjidikova,S. Vermeulen, S. Chatterjee, M. Berg, M. Grigorova, E.T.Sakamoto-Hojo, F. Granath, A.T. Ramalho, M.P. Curado,137Cesium-induced chromosome aberrations analyzed byfluorescence in situ hybridization: eight years follow upof the Goiania radiation accident victims, Mutat. Res. 400(1998) 299–312.
[89] D. Pinkel, J. Landegent, C. Collins, J. Fuscoe, R. Segraves,J. Lucas, J. Gray, Fluorescence in situ hybridization withhuman chromosome-specific libraries: detection of trisomy21 and translocations of chromosome 4, Proc. Natl. Acad.Sci. U.S.A. 85 (1988) 9138–9142.
[90] A.T. Ramalho, A.C.H. Nascimento, A.T. Natarajan, Doseassessments by cytogenetic analysis in Goiania radiationaccident, Radiat. Protection Dosimetry 25 (1988) 97–100.
[91] K. Salassidis, V. Georgiadou-Schumacher, H. Braselmann,P. Muller, R.U. Peter, M. Bauchinger, Chromosome painting
in highly irradiated Chernobyl victims: a follow-up studyto evaluate the stability of symmetrical translocations andthe influence of clonal aberrations for retrospective doseestimation, Int. J. Radiat. Biol. 68 (1995) 257–262.
[92] G. Snigiryova, H. Braselmann, K. Salassidis, V.Shevchenko, M. Bauchinger, Retrospective biodosimetry ofChernobyl clean-up workers using chromosome painting andconventional chromosome analysis, Int. J. Radiat. Biol. 71(1997) 119–127.
[93] G. Stephan, S. Pressl, Chromosome aberrations in humanlymphocytes analysed by fluorescence in situ hybridizationafter in vitro irradiation, and in radiation workers, 11 yearsafter an accidental radiation exposure, Int. J. Radiat. Biol.71 (1997) 293–299.
[94] J.D. Tucker, E.J. Tawn, D. Holdsworth, S. Morris, R.Langlois, M.J. Ramsey, P. Kato, J.D. Boice, R.E. Tarone,R.H. Jensen, Biological dosimetry of radiation workers atthe Sellafield nuclear facility, Radiat. Res. 148 (1997) 216–226.
[95] R. Zaire, C.S. Griffin, P.J. Simpson, D.G. Papworth,J.R. Savage, S. Armstrong, M.A. Hulten, Analysis oflymphocytes from uranium mineworkers in Namibiafor chromosomal damage using fluorescence in situhybridization (FISH), Mutat. Res. 371 (1996) 109–113.
[96] A.S. Goldman, Z. Fomina, P.A. Knights, C.J. Hill, A.P.Walker, M.A. Hulten, Analysis of the primary sex ratio, sexchromosome aneuploidy and diploidy in human sperm usingdual-colour fluorescence in situ hybridisation, Eur. J. Hum.Genet. 1 (1993) 325–334.
[97] S.J. Knight, S.W. Horsley, R. Regan, N.M. Lawrie, E.J.Maher, D.L. Cardy, J. Flint, L. Kearney, Developmentand clinical application of an innovative fluorescence insitu hybridization technique which detects submicroscopicrearrangements involving telomeres, Eur. J. Hum. Genet. 5(1997) 1–8.
[98] R.H. Martin, E. Spriggs, A.W. Rademaker, Multicolorfluorescence in situ hybridization analysis of aneuploidy anddiploidy frequencies in 225,846 sperm from 10 normal men,Biol. Reprod. 54 (1996) 394–398.
[99] M.J. Ramsey, D.H. Moore, J.F. Briner, D.A. Lee, L. Olsen,J.R. Senft, J.D. Tucker, The effects of age and lifestylefactors on the accumulation of cytogenetic damage asmeasured by chromosome painting, Mutat. Res. 338 (1995)95–106.
[100] P.J. Simpson, J.R. Savage, Dose–response curves for simpleand complex chromosome aberrations induced by X-raysand detected using fluorescence in situ hybridization, Int. J.Radiat. Biol. 69 (1996) 429–436.
[101] A.A. Edwards, P.A. Hone, J.E. Moquet, D.C. Lloyd, Simplechromosome exchanges are not linear with dose, Int. J.Radiat. Biol. 75 (1999) 1113–1117.
[102] P. Finnon, J.E. Moquet, A.A. Edwards, D.C. Lloyd, The60Co gamma ray dose–response for chromosomal aberrationsin human lymphocytes analysed by FISH; applicability tobiological dosimetry, Int. J. Radiat. Biol. 75 (1999) 1215–1222.
[103] C.S. Griffin, S.J. Marsden, D.L. Stevens, P. Simpson, J.R.Savage, Frequencies of complex chromosome exchange

32 G. Obe et al. / Mutation Research 504 (2002) 17–36
aberrations induced by238Pu alpha-particles and detected byfluorescence in situ hybridization using single chromosome-specific probes, Int. J. Radiat. Biol. 67 (1995) 431–439.
[104] C.S. Griffin, D.L. Stevens, J.R. Savage, Ultrasoft 1.5keV aluminum K X rays are efficient producers ofcomplex chromosome exchange aberrations as revealed byfluorescence in situ hybridization, Radiat. Res. 146 (1996)144–150.
[105] C.S. Griffin, M.A. Hill, D.G. Papworth, K.M. Townsend,J.R. Savage, D.T. Goodhead, Effectiveness of 0.28 keVcarbon K ultrasoft X-rays at producing simple and complexchromosome exchanges in human fibroblasts in vitrodetected using FISH, Int. J. Radiat. Biol. 73 (1998) 591–598.
[106] M. Grigorova, R. Brand, Y. Xiao, A.T. Natarajan, Frequen-cies and types of exchange aberrations induced by X-raysand neutrons in Chinese hamster splenocytes detected byFISH using chromosome-specific DNA libraries, Int. J.Radiat. Biol. 74 (1998) 297–314.
[107] M.P. Hande, A.T. Natarajan, Induction and persistenceof cytogenetic damage in mouse splenocytes followingwhole-body X-irradiation analysed by fluorescence in situhybridization. Part IV. Dose response, Int. J. Radiat. Biol.74 (1998) 441–448.
[108] C. Lindholm, S. Luomahaara, A. Koivistoinen, T. Ilus, A.A.Edwards, S. Salomaa, Comparison of dose–response curvesfor chromosomal aberrations established by chromosomepainting and conventional analysis, Int. J. Radiat. Biol. 74(1998) 27–34.
[109] J.N. Lucas, F. Hill, C. Burk, T. Fester, T. Straume, Dose–response curve for chromosome translocations measured inhuman lymphocytes exposed to 60Co�-rays, Health Phys.68 (1995) 761–765.
[110] J.N. Lucas, C.S. Hill, C.E. Burk, A.D. Lewis, A.K. Lucas,A.M. Chen, F.C. Sailes, T. Straume, Dose–response curve forchromosome translocations induced by low dose rate137Cs�-rays, Radiat. Protection Dosimetry 71 (1997) 279–282.
[111] P.J. Simpson, J.R. Savage, Identification of X-ray-inducedcomplex chromosome exchanges using fluorescence in situhybridization: a comparison at two doses, Int. J. Radiat.Biol. 66 (1994) 629–632.
[112] R.M. Anderson, S.J. Marsden, E.G. Wright, M.A. Kadhim,D.T. Goodhead, C.S. Griffin, Complex chromosomeaberrations in peripheral blood lymphocytes as a potentialbiomarker of exposure to high-LET alpha-particles, Int. J.Radiat. Biol. 76 (2000) 31–42.
[113] R. Eils, S. Uhrig, K. Saracoglu, K. Satzler, A. Bolzer,I. Petersen, J. Chassery, M. Ganser, M.R. Speicher, Anoptimized, fully automated system for fast and accurateidentification of chromosomal rearrangements by multiplex-FISH (M-FISH), Cytogenet. Cell Genet. 82 (1998) 160–171.
[114] P. Fuchs, M. Molls, Rapid detection of radiation-inducedchromosomal aberrations in lymphocytes and hematopoieticprogenitor cells by M-FISH, Mutat. Res. 452 (2000) 73–81.
[115] B.D. Loucas, M.N. Cornforth, Complex chromosomeexchanges induced by�-rays in human lymphocytes: anM-FISH study, Radiat. Res. 155 (2001) 660–671.
[116] E. Schrock, M.S. du, T. Veldman, B. Schoell, J. Wienberg,M.A. Ferguson-Smith, Y. Ning, D.H. Ledbetter, I. Bar-Am,
D. Soenksen, Y. Garini, T. Ried, Multicolor spectralkaryotyping of human chromosomes, Science 273 (1996)494–497.
[117] M.R. Speicher, S.G. Ballard, D.C. Ward, Karyotypinghuman chromosomes by combinatorial multi-fluor FISH,Nat. Genet. 12 (1996) 368–375.
[118] A.A. Edwards, J.R. Savage, Is there a simple answer to theorigin of complex chromosome exchanges? Int. J. Radiat.Biol. 75 (1999) 19–22.
[119] J.J. Boei, A.T. Natarajan, Combined use of chromosomepainting and telomere detection to analyse radiation-inducedchromosomal aberrations in mouse splenocytes, Int. J.Radiat. Biol. 73 (1998) 125–133.
[120] J.J. Boei, S. Vermeulen, J. Fomina, A.T. Natarajan, Detectionof incomplete exchanges and interstitial fragments inX-irradiated human lymphocytes using a telomeric PNAprobe, Int. J. Radiat. Biol. 73 (1998) 599–603.
[121] J.M. Brown, J.W. Evans, M.S. Kovacs, Mechanism ofchromosome exchange formation in human fibroblasts:insights from chromosome painting, Environ. Mol. Mutagen.22 (1993) 218–224.
[122] J.M. Brown, M.S. Kovacs, Visualization of nonreciprocalchromosome exchanges in irradiated human fibroblasts byfluorescence in situ hybridization, Radiat. Res. 136 (1993)71–76.
[123] W. Deng, J.N. Lucas, Combined FISH with pan-telomericPNA and whole chromosome-specific DNA probes to detectcomplete and incomplete chromosomal exchanges in humanlymphocytes, Int. J. Radiat. Biol. 75 (1999) 1107–1112.
[124] Y. Kodama, M. Nakano, K. Ohtaki, R. Delongchamp,A.A. Awa, N. Nakamura, Estimation of minimal size oftranslocated chromosome segments detectable by fluore-scence in situ hybridization, Int. J. Radiat. Biol. 71 (1997)35–39.
[125] J.N. Lucas, Cytogenetic signature for ionizing radiation, Int.J. Radiat. Biol. 73 (1998) 15–20.
[126] M. Nakano, E. Nakashima, D.J. Pawel, Y. Kodama, A. Awa,Frequency of reciprocal translocations and dicentrics inducedin human blood lymphocytes by X-irradiation as determinedby fluorescence in situ hybridization, Int. J. Radiat. Biol. 64(1993) 565–569.
[127] H. Wu, K. George, T.C. Yang, Estimate of true incompleteexchanges using fluorescence in situ hybridization withtelomere probes, Int. J. Radiat. Biol. 73 (1998) 521–527.
[128] H. Wu, K. George, T.C. Yang, Estimate of the frequency oftrue incomplete exchanges in human lymphocytes exposedto 1 GeV/u Fe ions in vitro, Int. J. Radiat. Biol. 75 (1999)593–599.
[129] C. Cremer, C. Muenkel, M. Granzow, A. Jauch, S. Dietzel,R. Eils, X.-Y. Guan, P.S. Meltzer, J.M. Trent, J. Langowski,T. Cremer, Nuclear architecture and the induction ofchromosomal aberrations, Mutat. Res. 366 (1996) 97–116.
[130] C. Cremer, P. Edelmann, H. Bornfleth, G. Kreth, H. Muench,H. Luz, M. Hausmann, Principles of spectral precisiondistance confocal microscopy for the analysis of molecularnuclear structure, in: B. Jaehne, H. Haussecker, P. Geissler(Eds.), Handbook of Computer Vision and Applications,Academic Press, San Diego, 1999, pp. 839–857.

G. Obe et al. / Mutation Research 504 (2002) 17–36 33
[131] T. Cremer, G. Kreth, H. Koester, R.H.A. Fink, R.Heintzmann, C. Cremer, I. Solovei, D. Zink, C. Cremer,Chromosome territories, interchromatin domain compart-ment, and nuclear matrix: an integrated view of thefunctional nuclear architecture, Crit. Rev. Eukaryot. GeneExpr. 12 (2000) 179–212.
[132] T. Cremer, C. Cremer, Chromosome territories, nucleararchitecture and gene regulation in mammalian cells, Nat.Rev. Genet. 2 (2001) 292–301.
[133] R. Heintzmann, G. Kreth, C. Cremer, Reconstruction of axialtomographic high resolution data from confocal fluorescencemicroscopy: a method for improving 3D FISH images, Anal.Cell. Pathol. 20 (2000) 7–15.
[134] G. Kreth, C. Muenkel, J. Langowski, T. Cremer, C. Cremer,Chromatin structure and chromosome aberrations: modelingof damage induced by isotropic and localized irradiation,Mutat.Res. 404 (1998) 77–88.
[135] J.R. Savage, Interchange and intra-nuclear architecture,Environ. Mol. Mutagen. 22 (1993) 234–244.
[136] J.R. Savage, Cancer. Proximity matters, Science 290 (2000)62–63.
[137] D. Zink, H. Bornfleth, A. Visser, C. Cremer, T. Cremer,Organization of early and late replicating DNA in humanchromosome territories, Exp. Cell Res. 247 (1999) 176–188.
[138] I. Chudoba, A. Plesch, T. Loerch, J. Lemke, U. Claussen,G. Senger, High resolution multicolor-banding: a newtechnique for refined FISH analysis of human chromosomes,Cytogenet. Cell Genet. 84 (1999) 156–160.
[139] C. Johannes, I. Chudoba, G. Obe, Analysis of X-ray-inducedaberrations in human chromosome 5 using high-resolutionmulticolour banding FISH (mBAND), Chromosome Res. 7(1999) 625–633.
[140] L. Barrios, R. Miro, M.R. Caballin, C. Fuster, F. Guedea,A. Subias, J. Egozcue, Cytogenetic effects of radiotherapy.Breakpoint distribution in induced chromosome aberrations,Cancer Genet. Cytogenet. 41 (1989) 61–70.
[141] M.E. Drets, G.A. Folle, W. Martinez-Lopez, R. Bonomi,J.E. Duarte, B.H. Mechoso, J. Larranaga, Quantitativelocalization of chromatid breaks induced by Alu I in the longarms of chromosome number 1 of Chinese hamster ovary(CHO) cells by microphotometric scanning, in: G. Obe,A.T. Natarajan (Eds.), Chromosomal Aberrations. Origin andSignificance, Springer, Heidelberg, 1994, pp. 169–183.
[142] M. Holmberg, J. Jonasson, Preferential location of X-rayinduced chromosome breakage in the R-bands of humanchromosomes, Hereditas 74 (1973) 57–67.
[143] G.P. Holmquist, Chromosome bands, their chromatin flavors,and their functional features, Am. J. Hum. Genet. 51 (1992)17–37.
[144] A. Kiuru, A. Lindholm, A. Auvinen, S. Salomaa,Localization of radiation-induced chromosomal breakpointsalong human chromosome 1 using a combination ofG-banding and FISH, Int. J. Radiat. Biol. 76 (2000) 667–672.
[145] G. Kuenzel, K.I. Gescheff, I. Schubert, Different chromo-somal distribution patterns of radiation-induced interchangebreakpoints in barley: first post-treatment mitosis versusviable offspring, Genome 44 (2001) 128–132.
[146] W. Mart́ınez-López, R. Bonomi, G.A. Folle, M.E. Drets,Microphotometric scanning of chromatid gaps and breaksinduced byAluI and BamHI in Chinese hamster ovary cells,Braz. J. Genet. 19 (1996) 577–582.
[147] J. McLeish, The action of maleic hydrozide in Vicia,Heredity 6 (1953) 125–147.
[148] Y. Nakagome, T. Matsubara, H. Fujita, Distribution of breakpoints in human structural rearrangements, Am. J. Hum.Genet. 35 (1983) 288–300.
[149] B. Porfirio, B. Tedeschi, P. Vernole, D. Caporossi, B.Nicoletti, The distribution of MspI-induced breaks in humanlymphocyte chromosomes and its relationship to commonfragile sites, Mutat. Res. 213 (1989) 117–124.
[150] S.H. Revell, Chromosome breakage by X-rays andradionimetic substances in Vicia, Heredity 6 (1953) 107–124.
[151] I. Schubert, R. Rieger, On the expressivity of aberrationhot spots after treatment with mutagens showing delayed ornon-delayed effects, Mutat. Res. 44 (1977) 337–344.
[152] I. Schubert, C. Huebner, R. Rieger, A. Michaelis, Effectsof chromosome repatterning inVicia faba L. Part VI.Translocation clustering in the NOR as affected by itschromosomal position, Biol. Zent. bl. 103 (1984) 529–542.
[153] P. Slijepcevic, A.T. Natarajan, Distribution of radiation-induced G1 exchange and terminal deletion breakpoints inChinese hamster chromosomes as detected by G banding,Int. J. Radiat. Biol. 66 (1994) 747–755.
[154] P. Slijepcevic, A.T. Natarajan, Distribution of X-ray-indu-ced G2 chromatid damage among Chinese hamsterchromosomes: influence of chromatin conformation, Mutat.Res. 323 (1994) 113–119.
[155] B. Tedeschi, B. Porfirio, D. Caporossi, P. Vernole, B.Nicoletti, Structural chromosomal rearrangements inHpaII-treated human lymphocytes, Mutat. Res. 248 (1991) 115–121.
[156] J.D. Tucker, J.R. Senft, Analysis of naturally occurring andradiation-induced breakpoint locations in human chromo-somes 1, 2 and 4, Radiat. Res. 140 (1994) 31–36.
[157] J.R. Savage, Testing the participation of chromosomesin structural aberrations, in: R.C. Sobti, G. Obe (Eds.),Eukaryotic Chromsomes: Structure and Functional Aspects,Narosa Press, New Delhi, 1991, pp. 111–125.
[158] J.R. Savage, Assignment of aberration breakpoints in bandedchromosomes, Nature 270 (1977) 513–514.
[159] J.R. Savage, Application of chromosome banding techniquesto the study of primary chromosome structural changes, J.Med. Genet. 14 (1977) 362–370.
[160] G.A. Folle, G. Obe, Localization of chromosome breakpointsinduced byAluI and BamHI in Chinese hamster ovary cellstreated in the G1 phase of the cell cycle, Int. J. Radiat. Biol.68 (1995) 437–445.
[161] G.A. Folle, G. Obe, Intrachromosomal localization ofbreakpoints induced by the restriction endonucleasesAluIandBamHI in Chinese hamster ovary cells treated in S phaseof the cell cycle, Int. J. Radiat. Biol. 69 (1996) 447–457.
[162] G.A. Folle, E. Boccardo, G. Obe, Localization ofchromosome breakpoints induced by DNase I in Chinese

34 G. Obe et al. / Mutation Research 504 (2002) 17–36
hamster ovary (CHO) cells, Chromosoma 106 (1997) 391–399.
[163] W. Mart́ınez-López, E.M. Boccardo, G.A. Folle, V.Porro, G. Obe, Intrachromosomal localization of aberrationbreakpoints induced by neutrons and gamma rays in Chinesehamster ovary cells, Radiat. Res. 150 (1998) 585–592.
[164] W. Mart́ınez-López, G.A. Folle, G. Obe, P. Jeppesen,Chromosome regions enriched in hyperacetylated histone H4are preferred sites for endonuclease- and radiation-inducedbreakpoints, Chromosome Res. 9 (2001) 69–75.
[165] T.R. Hebbes, A.W. Thorne, C. Crane-Robinson, A directlink between core histone acetylation and transcriptionallyactive chromatin, EMBO J. 7 (1988) 1395–1402.
[166] T.R. Hebbes, A.W. Thorne, A.L. Clayton, C. Crane-Robinson, Histone acetylation and globin gene switching,Nucl. Acids Res. 20 (1992) 1017–1022.
[167] B.M. Turner, Histone acetylation and control of gene ex-pression, J. Cell Sci. 99 (1991) 13–20.
[168] A.P. Wolffe, J.J. Hayes, Chromatin disruption and modi-fication, Nucl. Acids. Res. 27 (1999) 711–720.
[169] P.L. Jones, G.J. Veenstra, P.A. Wade, D. Vermaak, S.U. Kass,N. Landsberger, J. Strouboulis, A.P. Wolffe, MethylatedDNA and MeCP2 recruit histone deacetylase to represstranscription, Nat. Genet. 19 (1998) 187–191.
[170] X. Nan, H.H. Ng, C.A. Johnson, C.D. Laherty, B.M. Turner,R.N. Eisenman, A. Bird, Transcriptional repression bythe methyl-CpG-binding protein MeCP2 involves a histonedeacetylase complex, Nature 393 (1998) 386–389.
[171] H.H. Ng, Y. Zhang, B. Hendrich, C.A. Johnson, B.M. Turner,H. Erdjument-Bromage, P. Tempst, D. Reinberg, A. Bird,MBD2 is a transcriptional repressor belonging to the MeCP1histone deacetylase complex, Nat. Genet. 23 (1999) 58–61.
[172] H.H. Ng, P. Jeppesen, A. Bird, Active repression ofmethylated genes by the chromosomal protein MBD1, Mol.Cell Biol. 20 (2000) 1394–1406.
[173] T.R. Hebbes, A.L. Clayton, A.W. Thorne, C. Crane-Robinson, Core histone hyperacetylation co-maps withgeneralized DNase I sensitivity in the chicken beta-globinchromosomal domain, EMBO J. 13 (1994) 1823–1830.
[174] F. Thoma, T. Koller, A. Klug, Involvement of histone H1 inthe organization of the nucleosome and of the salt-dependentsuperstructures of chromatin, J. Cell Biol. 83 (1979) 403–427.
[175] P. Jeppesen, A. Mitchell, B. Turner, P. Perry, Antibodiesto defined histone epitopes reveal variations in chromatinconformation and underacetylation of centric heterochro-matin in human metaphase chromosomes, Chromosoma 101(1992) 322–332.
[176] P. Jeppesen, B.M. Turner, The inactive X chromosome infemale mammals is distinguished by a lack of histone H4acetylation, a cytogenetic marker for gene expression, Cell74 (1993) 281–289.
[177] P. Jeppesen, Histone acetylation: a possible mechanism forthe inheritance of cell memory at mitosis, Bioessays 19(1997) 67–74.
[178] P. Jeppesen, Immunofluorescence in cytogenetic analysis:method and applications, Gen. Mol. Biol. 23 (2000) 1107–1114.
[179] S. Cigarran, L. Barrios, J.F. Barquinero, M.R. Caballin,M. Ribas, J. Egozcue, Relationship between the DNAcontent of human chromosomes and their involvementin radiation-induced structural aberrations, analysed bypainting, Int. J. Radiat. Biol. 74 (1998) 449–455.
[180] K.L. Johnson, D.J. Brenner, J. Nath, J.D. Tucker, C.R.Geard, Radiation-induced breakpoint misjoining in humanchromosomes: random or non-random? Int. J. Radiat. Biol.75 (1999) 131–141.
[181] A. Wojcik, C. Streffer, Comparison of radiation-inducedaberration frequencies in chromosmes 1 and 2 of two humandonors, Int. J. Radiat. Biol. 74 (1998) 573–581.
[182] G. Obe, Die Kombinationshaeufigkeiten menschlicherLeukozytenchromosomen in chemisch induzierten Translo-kationen, Chromosoma 27 (1969) 321–326.
[183] F. Vogel, T.M. Schroeder, The internal order of the interphasenucleus, Hum. Genet. 25 (1974) 265–297.
[184] J.R. Savage, D.G. Papworth, Frequency and distributionstudies of asymmetrical versus symmetrical chromosomeaberrations, Mutat. Res. 95 (1982) 7–18.
[185] W. Mart́ınez-López, V. Porro, G.A. Folle, L. Mendez-Acuna,J.R. Savage, G. Obe, Interchromosomal distribution ofgamma ray-induced chromatid aberrations in Chinesehamster ovary (CHO) cells, Gen. Mol. Biol. 23 (2000) 1071–1076.
[186] B. Dutrillaux, Nouveau système de marquage chromo-somique: les bandes T, Chromosoma 41 (1973) 395–402.
[187] T.D. Allen, E.M. Jack, C.J. Harrison, The three dimensionalstructure of human metaphase chromosomes determinedby scanning electron microscopy, in: K.W. Adolph (Ed.),Chromosomes and Chromatin, CRC Press, Boca Raton, FL,1988, pp. 51–72.
[188] P. Ludena, C. Sentis, F. De Cabo, M. Velazquez, J.Fernandez-Piqueras, Visualization of R-bands in humanmetaphase chromosomes by the restriction endonucleaseMseI, Cytogenet. Cell Genet. 57 (1991) 82–86.
[189] M.E. Drets, G.A. Drets, P.J. Queirolo, F.J. Monteverde,Computer graphics as a tool in cytogenetic research andeducation, Comput. Appl. Biosci. 11 (1995) 463–468.
[190] M.E. Drets, F.J. Monteverde, Automated cytogenetics withmodern computerized scanning microscope photometersystems, in: G. Obe, A. Basler (Eds.), Cytogenetics. Basicand Applied Aspects, Springer, Berlin, 1987, pp. 48–64.
[191] M.E. Drets, G.A. Folle, F.J. Monteverde, Quantitativedetection of chromosome structures by computerized micro-photometric scanning, in: G. Obe, A.T. Natarajan (Eds.),Chromosomal Aberrations Basic and Applied Aspects,Springer, Heidelberg, 1989, pp. 1–2.
[192] M.E. Drets, M. Mendizabal, The underlying structure ofthe subtelomeric region detected by microphotometricalscanning and chromosome graphic image analysis, Mutat.Res. 404 (1998) 13–16.
[193] M.E. Drets, G. Obe, F.J. Monteverde, G.A. Folle, I.I.Medina, M.G. De Galvez, J.E. Duarte, B.H. Mechoso,Computerized graphic and light microscope analyses ofT-banded chromosome segments of Chinese hamster ovarycells and human lymphocytes, Biol. Zent. bl. 111 (1992)204–214.

G. Obe et al. / Mutation Research 504 (2002) 17–36 35
[194] M.E. Drets, Insights into the structure of the subtelomericchromosome segments, Gen. Mol. Biol. 23 (2000) 1087–1093.
[195] M.N. Cornforth, R.L. Eberle, Termini of human chromo-somes display elevated rates of mitotic recombination,Mutagenesis 16 (2001) 85–89.
[196] E. Holinski-Feder, E. Reyniers, S. Uhrig, A. Golla, J.Wauters, P. Kroisel, P. Bossuyt, I. Rost, K. Jedele,H. Zierler, S. Schwab, D. Wildenauer, M.R. Speicher,P.J. Willems, T. Meitinger, R.F. Kooy, Familial mentalretardation syndrome ATR-16 due to an inherited crypticsubtelomeric translocation, t(3;16)(q29;p13.3), Am. J. Hum.Genet. 66 (2000) 16–25.
[197] J. Brown, S.W. Horsley, C. Jung, K. Saracoglu, B. Janssen,M. Brough, M. Daschner, B. Beedgen, G. Kerkhoffs, R. Eils,P.C. Harris, A. Jauch, L. Kearney, Identification of a subtlet(16;19)(p13.3;p13.3) in an infant with multiple congenitalabnormalities using a 12-colour multiplex FISH telomereassay, M-TEL, Eur. J. Hum. Genet. 8 (2000) 903–910.
[198] F. Speleman, B. Callens, K. Logghe, N. Van Roy,S.W. Horsley, A. Jauch, M.R. Verschraegen-Spae, J.G.Leroy, Subtelomeric familial translocation t(2;7)(q37;q35)leading to partial trisomy 7q35-->qter: molecular cytogeneticanalysis and clinical phenotype in two generations, Am. J.Med. Genet. 93 (2000) 349–354.
[199] P. Warburton, S. Mohammed, C.M. Ogilvie, Detection ofsubmicroscopic subtelomeric chromosome translocations: anew case study, Am. J. Med. Genet. 91 (2000) 51–55.
[200] E.H. Blackburn, The end of the (DNA) line, Nat. Struct.Biol. 7 (2000) 847–850.
[201] M.P. Hande, P.M. Lansdorp, A.T. Natarajan, Inductionof telomerase activity by in vivo X-irradiation of mousesplenocytes and its possible role in chromosome healing,Mutat. Res. 404 (1998) 205–214.
[202] D. Kipling, The Telomere, Oxford University Press, Oxford,1995.
[203] P. Slijepcevic, Telomeres and mechanisms of Robertsonianfusion, Chromosoma 107 (1998) 136–140.
[204] P. Slijepcevic, P. Bryant, Chromosome healing, telomerecapture and mechanisms of radiation-induced chromosomebreakage, Int. J. Radiat. Biol. 73 (1998) 1–13.
[205] P. Slijepcevic, A.T. Natarajan, P. Bryant, Telomeres andradiation-induced chromosome breakage, Mutagenesis 13(1998) 45–49.
[206] M.E. Luderus, B. van Steensel, L. Chong, O.C. Sibon, F.F.Cremers, T. de Lange, Structure, subnuclear distribution,and nuclear matrix association of the mammalian telomericcomplex, J. Cell Biol. 135 (1996) 867–881.
[207] A. Smogorzewska, B. van Steensel, A. Bianchi, S. Oelmann,M.R. Schaefer, G. Schnapp, T. de Lange, Control of humantelomere length by TRF1 and TRF2, Mol. Cell Biol. 20(2000) 1659–1668.
[208] J. Karlseder, D. Broccoli, Y. Dai, S. Hardy, T. de Lange,p53- and ATM-dependent apoptosis induced by telomereslacking TRF2, Science 283 (1999) 1321–1325.
[209] B. van Steensel, A. Smogorzewska, T. de Lange, TRF2protects human telomeres from end-to-end fusions, Cell 92(1998) 401–413.
[210] M. Chamankhah, W. Xiao, Formation of the yeastMre11–Rad50–Xrs2 complex is correlated with DNA repairand telomere maintenance, Nucl. Acids Res. 27 (1999)2072–2079.
[211] P.S. Meltzer, X.Y. Guan, J.M. Trent, Telomere capturestabilizes chromosome breakage, Nat. Genet. 4 (1993) 252–255.
[212] C.N. Sprung, G.E. Reynolds, M. Jasin, J.P. Murnane,Chromosome healing in mouse embryonic stem cells, Proc.Natl. Acad. Sci. U.S.A. 96 (1999) 6781–6786.
[213] P. Slijepcevic, Y. Xiao, A.T. Natarajan, Chromosome-specifictelomeric associations in Chinese hamster embryonic cells,Genes Chromosomes Cancer 28 (2000) 98–105.
[214] S.M. Bailey, J. Meyne, D.J. Chen, A. Kurimasa, G.C. Li,B.E. Lehnert, E.H. Goodwin, DNA double-strand breakrepair proteins are required to cap the ends of mammalianchromosomes, Proc. Natl. Acad. Sci. U.S.A. 96 (1999)14899–14904.
[215] S.G. Martin, T. Laroche, N. Suka, M. Grunstein, S.M.Gasser, Relocalization of telomeric Ku and SIR proteins inresponse to DNA strand breaks in yeast, Cell 97 (1999)621–633.
[216] K.D. Mills, D.A. Sinclair, L. Guarente, MEC1-dependentredistribution of the Sir3 silencing protein from telomeresto DNA double-strand breaks, Cell 97 (1999) 609–620.
[217] K.K. Wong, S. Chang, S.R. Weiler, S. Ganesan, J. Chaudhuri,C. Zhu, S.E. Artandi, K.L. Rudolph, G.J. Gottlieb, L. Chin,F.W. Alt, R.A. DePinho, Telomere dysfunction impairs DNArepair and enhances sensitivity to ionizing radiation, Nat.Genet. 26 (2000) 85–88.
[218] L. Alvarez, J.W. Evans, R. Wilks, J.N. Lucas, J.M.Brown, A.J. Giaccia, Chromosomal radiosensitivity at intra-chromosomal telomeric sites, Genes Chromosomes Cancer8 (1993) 8–14.
[219] J.P. Day, B.A. Marder, W.F. Morgan, Telomeres and theirpossible role in chromosome stabilization, Environ. Mol.Mutagen. 22 (1993) 245–249.
[220] J.P. Day, C.L. Limoli, W.F. Morgan, Recombinationinvolving interstitial telomere repeat-like sequencespromotes chromosomal instability in Chinese hamster cells,Carcinogenesis 19 (1998) 259–265.
[221] J.L. Fernandez, J. Gosalvez, V. Goyanes, High frequencyof mutagen-induced chromatid exchanges at interstitialtelomere-like DNA sequence blocks of Chinese hamstercells, Chromosome Res. 3 (1995) 281–284.
[222] J. Meyne, R.J. Baker, H.H. Hobart, T.C. Hsu, O.A. Ryder,O.G. Ward, J.E. Wiley, D.H. Wurster-Hill, T.L. Yates,R.K. Moyzis, Distribution of non-telomeric sites of the(TTAGGG)n telomeric sequence in vertebrate chromosomes,Chromosoma 99 (1990) 3–10.
[223] L. Alvarez, J.W. Evans, R. Wilks, J.N. Lucas, J.M.Brown, A.J. Giaccia, Chromosomal radiosensitivity at intra-chromosomal telomeric sites, Genes Chromosomes Cancer8 (1993) 8–14.
[224] A.S. Balajee, H.J. Oh, A.T. Natarajan, Analysis of restrictionenzyme-induced chromosome aberrations in the interstitialtelomeric repeat sequences of CHO and CHE cells by FISH,Mutat. Res. 307 (1994) 307–313.

36 G. Obe et al. / Mutation Research 504 (2002) 17–36
[225] A.S. Balajee, I. Dominguez, V.A. Bohr, A.T. Natarajan,Immunofluorescent analysis of the organization of telomericDNA sequences and their involvement in chromosomal aber-rations in hamster cells, Mutat. Res. 372 (1996) 163–172.
[226] P. Slijepcevic, Y. Xiao, I. Dominguez, A.T. Natarajan, Spon-taneous and radiation-induced chromosomal breakage at in-terstitial telomeric sites, Chromosoma 104 (1996) 596–604.
[227] M.P. Hande, A.S. Balajee, A.T. Natarajan, Induction oftelomerase activity by UV-irradiation in Chinese hamstercells, Oncogene 15 (1997) 1747–1752.
[228] H.J. Oh, M.P. Hande, P.M. Lansdorp, A.T. Natarajan,Induction of telomerase activity and chromosomal aberra-tions in human tumour cell lines following X-irradiation,Mutat. Res. 401 (1998) 121–131.