clin pathol: neutrophil elastase and g and in bone a...
TRANSCRIPT

MJClin Pathol: Mol Pathol 1995;48:M28-M34
Neutrophil elastase and cathepsin G proteinand messenger RNA expression in bone marrow
from a patient with Chediak-Higashi syndrome
D Burnett, C J Ward, R A Stockley, R G Dalton, A J Cant, S Hoare, J Crocker
LungImmunobiochemicalResearch Laboratory,The General Hospital,BirminghamD BurnettD J WardA R Stockley
Department ofPathology,Cheltenham GeneralHospitalR G Dalton
Children'sDepartment,Newcasde GeneralHospital, Newcastleupon TyneA J CantS Hoare
Department ofHistopathology,BirminghamHeartlands TrustHospitalJ Crocker
Correspondence to:Dr D Bumett,Liver and Hepatobiliary UnitResearch Laboratories,Clinical Research Block,Queen Elizabeth Hospital,Edgbaston, Birmingham B 152TH.
Accepted for publication8 November 1994
AbstractAims-To determine whether neutrophilelastase and cathepsin G are expressed, attranscriptional or translational levels, inthe bone marrow from a patient withChediak-Higashi syndrome.Methods-Blood neutrophils were isolatedfrom three patients with Chediak-Higashidisease and bone marrow was collectedfrom one. Cell lysates were analysed forneutrophil elastase and cathepsin G ac-tivity by enzyme linked immunosorbentassay and western immunoblotting.Northern blotting was used to detect mes-senger RNA (mRNA) for cathepsin G, el-astase and ,-actin in bone marrowextracts, and immunohistochemistry wasused to localise the enzymes in marrowmyeloid cells.Results-Elastase and cathepsin G werenot detected in blood neutrophils from thepatients with Chediak-Higashi disease,but were present in bone marrow cells,although immunohistochemistry showedthey were not within cytoplasmic granules.The concentrations of elastase and ca-thepsin G in Chediak-Higashi bone mar-row were about 25 and 15%, respectively,of those in normal marrow. Quantitativescanning of northern blots showed thatelastase and cathepsin GmRNA, correctedfor ,-actin mRNA, were expressed equallyin normal marrow.Conclusions-Transcription of elastaseand cathepsin G mRNA in promyelocytesof patients with Chediak-Higashi diseaseis normal, but the protein products aredeficient in these cells and absent in ma-ture neutrophils. This suggests that thetranslated proteins are not packaged intoazurophil granules but are degaded or se-creted from the cells.( Clin Pathol: Mol Pathol 1995;48:M28-M34)
Keywords: Chediak-Higashi syndrome, elastase, ca-thepsin G.
Chediak-Higashi syndrome is an autosomalrecessive disorder characterised by abnormalcytoplasmic granules in a variety of cells.' Amajor sequela of Chediak-Higashi syndromeis recurrent infection, which is thought to resultfrom inefficient microbial killing by leucocytes,despite normal phagocytic activity and a normalrespiratory burst.2 This defect therefore appearsto be associated with a deficiency in killing ofbacteria by microbicidal proteins in phago-
lysosomes. Blood neutrophils of patients withChediak-Higashi syndrome are deficient in theazurophil granule enzymes cathepsin G andelastase. Myeloperoxidase and the defensins,however, are present in normal or slightly re-duced amounts,3 4 myeloperoxidase havingbeen localised to the granules of the cells.5
Cathepsin G and elastase are also deficientin blood neutrophils of the Chediak-Higashianimal model, the beige mouse.6 The bonemarrow of the beige mouse, however, containsabout half the elastase and cathepsin G contentof normal mouse marrow, within neutrophilprecursors but not mature cells.7 These resultssuggest that in this animal model the cathepsinG and elastase genes are transcribed and pro-tein translation does occur, but the enzymesdo not survive to the stage of mature cells. Itis not known whether a similar situation existsin human Chediak-Higashi disease, or indeedwhether transcription of these proteins is nor-mal. The purpose of the present study wasto analyse the messenger RNA (mRNA) andprotein content of cathepsin G and elastase inthe bone marrow of patients with Chediak-Higashi syndrome.
MethodsThree patients with Chediak-Higashi syn-drome were studied. Diagnosis was based ona history of recurrent infection, depigmentedareas of skin, hair or retina, and characteristicgiant granules in leucocytes. None had evid-ence of neuropathy.
Patient JF is a 25 year old man who wasdiagnosed when aged 13 years. He has sufferedfrom recurrent gingivitis, eventually requiringextraction of all his teeth, and multiple skininfections which have been treated with anti-biotics and occasional surgical drainage. He hashad no major chest or urinary tract infections.
Patient GH is a 10 year old boy, diagnosedwhen five years old. He has had a history offrequent respiratory tract and throat infections,oral ulceration, paronychia, and skin bruisingfollowing minor trauma. Relatively minor ab-rasions to the skin were delayed in healing andoften became septic.
Patient AE is a four year old girl with ahistory of recurrent infections of the urinarytract, respiratory tract and throat from the ageof six weeks. She has recently suffered frommouth ulcers and paronychia.
Blood neutrophils were purified as describedby Jepsen and Skottun.' Venous blood wascollected, from the three patients and from a
M28
on 4 July 2018 by guest. Protected by copyright.
http://mp.bm
j.com/
Clin M
ol Pathol: first published as 10.1136/m
p.48.1.M28 on 1 F
ebruary 1995. Dow
nloaded from

Chediak-Higashi azurophil proteinases
healthy volunteer, into lithium heparin anti-coagulant and each blood sample was dilutedwith an equal volume of 0 15 mol/l NaCl so-lution. The diluted blood (5 ml) was layeredonto two layers of Percoll (Pharmacia AB,Uppsala, Sweden); the upper layer was 2 ml1-075 g/ml Percoll, the lower 3 ml 1-096 g/mlPercoll. The tubes were centrifuged at 2000 x gfor 20 minutes. The neutrophils were harvestedfrom the interface between the Percoll layers,washed three times in TRIS buffered RPMI1640 medium (Flow Laboratories, Rick-mansworth, UK), counted and resuspended inRPMI 1640 medium. The polymorphonuclearleucocytes (PMN) were >96% pure and >98%viable (assessed by exclusion of trypan blue).The neutrophil preparations from each subjectwere separated into two samples, centrifugedand the cell pellets retained. One sample fromeach subject (for enzyme activity assays) waslysed by the addition of 0 5 M Tris/HCl, pH7-8, containing 1-0M NaCl and 0-1% (v/v)Triton X-100 and sonicated (6 x 20 W) usinga Lucas Dawe Ultrasonics Soniprobe. The sec-ond sample (for western immunoblotting andenzyme linked immunosorbent assay (ELISA))was lysed by sonication with 0 5 M Tris/HCl,pH 7-8 containing 1 0M NaCl and 0 1% (v/v)Triton X-100, after the addition of proteinaseinhibitors; 25 5 ,ul 100 mM diisopropyl fluoro-phosphate dissolved in dimethyl sulphoxide,0 1 ml 0 1 M 1,10 phenanthroline in methanoland 0-1 ml 0-1 M iodoacetic acid in water. Theneutrophil lysates were stored at - 70°C untilanalysed.Bone marrow (5 ml) was aspirated from the
superior iliac spine of one of the patients (JF)after obtaining informed consent. After re-moval of a sample for routine haematologicalanalysis, the remainder was dispensed into30 ml RPMI 1640 culture medium, containing10% (v/v) fetal calf serum and 2000 unitsof heparin anticoagulant. A portion of bonemarrow was also retained from a haema-tologically normal bone marrow donor. Themarrow samples were centrifuged at 200 x gfor 20 minutes through 5 ml 1 096 g/ml Percollto remove erythrocytes and erythroid pre-cursors. The cells remaining above the Percolllayer were collected, washed in RPMI 1640medium and cytospin preparations made fordifferential cell counts and immuno-histochemistry. The remainder of the cells werecounted and lysates were obtained for westernblotting and enzyme assays as for blood neu-trophils. Preparations of mRNA were also ob-tained from bone marrow for northern blotting.The mRNA was prepared from the bone mar-row using a Promega PolyATract System 1000kit (Promega, Southampton, UK) and finallysolubilised and stored as described by Chom-czynski.9Three complementary DNA (cDNA) probes
were used. The elastase probe'0 was a 516 basepair partial cDNA (supplied by Dr G Salvesen,University of Georgia, Athens, USA) sub-cloned into the EcoRl site of pBluescriptKS(±+/-). The cathepsin G cDNA" (suppliedby Dr T Ley, Jewish Hospital, St Louis, USA)was 1-0 kilobase subcloned into the EcoRl site
of Bluescript. A [-actin probe'2 (also suppliedby Dr T Ley) was a 1 9 kilobase cDNA in theBamHl site of Puc9.Northem blotting was performed essentially
as described by Maniatis et al.'3 The mRNApreparations (about 10,ug) from normal andChediak-Higashi bone marrow samples, sus-pended in denaturing buffer (MOPS con-taining formamide and formaldehyde), weresubjected to electrophoresis in 1% (w/v) aga-rose gels in 0-2 M MOPS buffer (pH 7-0)containing 2-2M formaldehyde, 50mM so-dium acetate and 1 mM EDTA. Following elec-trophoresis, the RNA samples were transferredby capillary blotting to Genescreen TM (NENResearch, Boston, USA). The Genescreen wasprehybridised for 12 hours at 42°C before theaddition ofthe cathepsinG probe or the elastaseprobe (106 ng), radiolabelled with ["P] dCTPto 109 dpm/mg DNA using a commercial ran-dom prime labelling kit (Boehringer Mann-heim, Lewes, UK). Hybridisation took placefor 18 hours at 42°C in the presence of 50%formamide. The screens were then washed fourtimes with 2 x standard sodium citrate (SSC)at 20°C and twice with 0-2 x SSC at 65°C(1 x SSC = 0 15MNaCl,0-015Msodiumcitrate, pH 7*0). The screens were auto-radiographed at -70°C and that which hadbeen probed with the elastase cDNA wasscanned with LKB Wallach 1205 [ plate flatbed counter in the scintillation mode. Whenthe radioactivity of the elastase probed screenhad decayed to undetectable levels, it was re-probed with radiolabelled [3-actin cDNA, auto-radiographed and scanned again. The ratiosof radioactivity caused by hybridisation ofelastase mRNA and P-actin mRNA in normaland Chediak-Higashi bone marrow sampleswere calculated from the scans.
Neutrophil and bone marrow lysates weresubjected to sodium dodecyl sulphate poly-acrylamide gel electrophoresis (SDS-PAGE),as described by Laemmli,'4 on an LKB "Mid-get" system (Pharmacia LKB Biotechnology,Milton Keynes, UK) using a 5% poly-acrylamide stacking gel and 12-5% running gel.Each sample loaded contained lysate from 104cells. Following electrophoresis, the proteinswere transferred to nitrocellulose paper(Hybond-C; Amersham International) by elec-troblotting. The nitrocellulose sheets were in-cubated with either sheep antineutrophilelastase'5 or sheep anticathepsin G,'6 followedby donkey antisheep IgG labelled with horse-radish peroxidase (Binding Site, Birmingham,UK). Binding of peroxidase labelled antibodywas visualised using 3-amino-9-ethylcarbazoleas the substrate.
Cathepsin G was measured in blood neu-trophil and bone marrow lysates using a directcapture ELISA. Nunc Immuno 96 well plateswere loaded with 0-2 ml sheep antihumancathepsin G, 20 jtl/ml in phosphate bufferedsaline (PBS) (pH 7 2) and incubated overnightat 4°C. The wells were washed three times withwashing buffer (PBS containing 0-1% Tween-20 and 1% (v/v) Haemaccel (Hoechst)) beforethe addition of 0-2 ml of each cell lysate dilutedin washing buffer with 0 5M NaCl. Reference
M29
on 4 July 2018 by guest. Protected by copyright.
http://mp.bm
j.com/
Clin M
ol Pathol: first published as 10.1136/m
p.48.1.M28 on 1 F
ebruary 1995. Dow
nloaded from
Burnett, Ward, Stockley, Dalton, Cant, Hoare, et al
standards consisted of dilutions of cathepsinG, purified from an empyema as described byMartodam et al. 7 The plates were incubated fortwo hours at room temperature before washing,followed by the addition of a horseradish per-oxidase conjugated sheep anticathepsin G. Af-ter a further two hours of incubation, the wellswere washed and substrate added; 0-2 ml/well1 mg/ml o-phenylenediamine dihydrochloride(Sigma) in 0 1 M citrate/phosphate buffer (pH5 0) with 1 ,ul/ml hydrogen peroxide. The col-our reaction was stopped with 50 gil 0-5 Mcitric acid/well and the absorbance measuredwith a Dynatech MR 5000 plate reader at490 nm. Cathepsin G concentrations in the celllysates were calculated by interpolation fromthe reference values obtained with pure en-zyme. The lower limit of detection was3 ng/ml.
Neutrophil elastase in cell lysates was meas-ured using an indirect competition ELISA.Nunc Immuno 96 well plates were loaded with0-2 ml elastase, 1 ,ug/ml in PBS, pH 7-2, with0-5 M NaCl and incubated overnight at 4°C.The cell lysates and dilutions of elastase, puri-fied from an empyema,17 were each mixed withequal volumes of 25 jg/ml sheep antielastase,which were also incubated overnight at 4°C.The ELISA plates were washed and the lysatesand reference mixtures (0.2 ml/well) added andincubated for two hours at room temperature.After further washing, the plate wells wereloaded with 0-2 ml horseradish peroxidase con-jugated donkey antisheep IgG (Binding Site)and incubated for two hours at room tem-perature, before the addition of substrate so-lution and measurement of the absorbance at490 nm, as described earlier. The elastase con-
centrations in the cell lysates were calculatedby interpolation from the inhibition bindingreference line obtained with pure elastase, thelower limit of detection being 3 ng/ml. Resultsfor both enzymes were expressed as pg protein/myeloid cell.
Cathepsin G and elastase activities in bloodneutrophil and bone marrow lysates were meas-ured in microtitre plate wells using fluorimetricsubstrates, Succ-Ala-Ala-Pro-Phe-paranitro-anilide and Succ-Ala-Ala-Ala-paranitroanilide,respectively (Bachem Feinchemikalien, Bu-bendorf, Switzerland). Samples (40 p1) of thecell lystates and dilutions of calibration stand-ards ofpure cathepsin G or elastase were mixedwith 140 jil substrate buffer (0 05 mol/l TRIS/HCI buffer containing 0 5 mol/l NaCl and 1%(v/v) Triton X-1 00 and 1 mg/ml of the ap-propriate substrate). The plates were incubatedat room temperature until colour change wassufficient and the absorbance of each well wasmeasured at 410 nm with a DynatechMR 5000plate reader. Cathepsin G and elastase con-centrations in cell lysates were calculated byinterpolation from the calibration curves ob-tained with pure enzymes. The lower limits ofdetection for cathepsin G and elastase activitywere 190 and 150 ng/ml, respectively. Resultswere expressed as activity representing pg en-zyme/myeloid cell.The cytocentrifuge preparations from the
bone marrow samples were stained immuno-histochemically for cathepsin G and elastase asdescribed previously.'5 16.
ResultsNORTHERN BLOTTINGAutoradiographs of northern blots obtained
CATH46
ch.
TA 8-.E
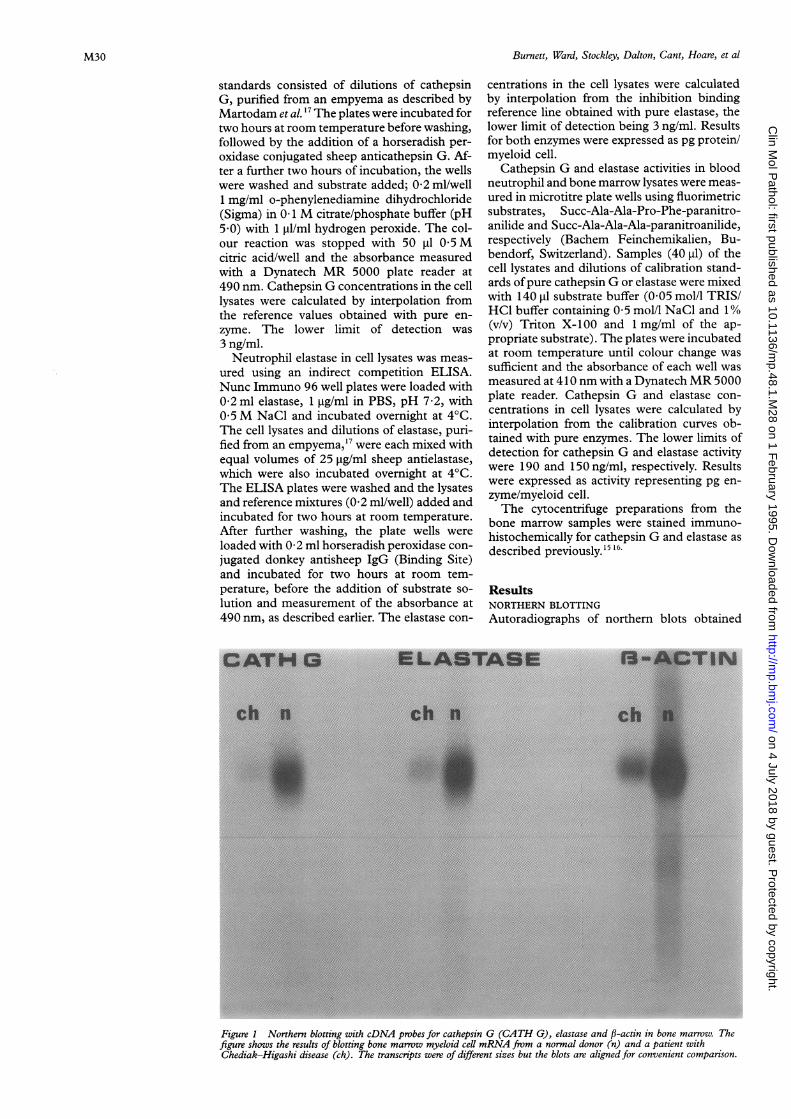
Figure 1 Northern blotting with cDNA probes for cathepsin G (CATH G), elastase and f,-actin in bone marrow. Thefigure shows the results of blotting bone marrow myeloid cell mRNA from a normal donor (n) and a patient withChediak-Higashi disease (ch). The transcripts were of different sizes but the blots are aligned for convenient comparison.
TIN.1.:
M30
...
ISO
....
................. ....................... ...........
...........
......... ........ .........................................................
on 4 July 2018 by guest. Protected by copyright.
http://mp.bm
j.com/
Clin M
ol Pathol: first published as 10.1136/m
p.48.1.M28 on 1 F
ebruary 1995. Dow
nloaded from
Chediak-Higashi azurophil proteinases
1 2 3 1 2 3
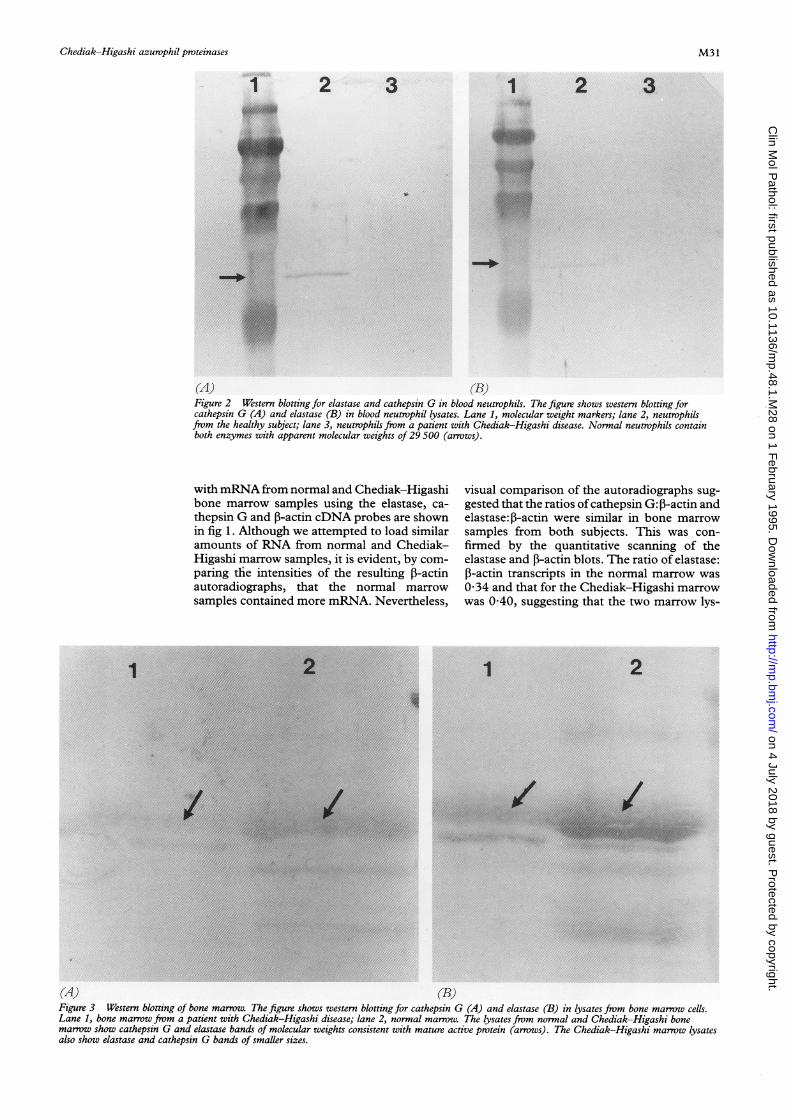
(A) (B)Figure 2 Western blotting for elastase and cathepsin G in blood neutrophils. The figure shows western blotting forcathepsin G (A) and elastase (B) in blood neutrophil lysates. Lane 1, molecular weight markers; lane 2, neutrophilsfrom the healthy subject; lane 3, neutrophils from a patient with Chediak-Higashi disease. Normal neutrophils containboth enzymes with apparent molecular weights of 29 500 (arrows).
withmRNA from normal and Chediak-Higashibone marrow samples using the elastase, ca-thepsin G and P-actin cDNA probes are shownin fig 1. Although we attempted to load similaramounts of RNA from normal and Chediak-Higashi marrow samples, it is evident, by com-paring tie intensities of the resulting f-actinautoradiographs, that the normal marrowsamples contained more mRNA. Nevertheless,
visual comparison of the autoradiographs sug-gested that the ratios ofcathepsin G: P-actin andelastase: P-actin were similar in bone marrowsamples from both subjects. This was con-firmed by the quantitative scanning of theelastase and P-actin blots. The ratio of elastase:j-actin transcripts in the normal marrow was0-34 and that for the Chediak-Higashi marrowwas 0 40, suggesting that the two marrow lys-
;: 2
.:::.:~~ ~ ~ ~ ~ ~ ~~~~~~~i
1
O/
2
./
(A) (B)
Figure 3 Western blotting of bone marrow. The figure shows western blotting for cathepsin G (A) and elastase (B) in lysates from bone marrow cells.Lane 1, bone marrow from a patient with Chediak-Higashi disease; lane 2, normal marrow. The lysates from normal and Chediak-Higashi bonemarrow show cathepsin G and elastase bands of molecular weights consistent with mature active protein (arrows). The Chediak-Higashi marrow lysatesalso show elastase and cathepsin G bands of smaller sizes.
1
M31
:mi
on 4 July 2018 by guest. Protected by copyright.
http://mp.bm
j.com/
Clin M
ol Pathol: first published as 10.1136/m
p.48.1.M28 on 1 F
ebruary 1995. Dow
nloaded from

Burnett, Ward, Stockley, Dalton, Cant, Hoare, et al
.
Al'* ...;.' .,:t...... .
, ..........
... .....
(13),
.:..:. ......'
v.....
(( (1))Figure 4 Immunohistochemical staining of bone marrow promyeloid cells for cathepsin G and elastase. (A) CathepsinG staining of normal marrw. (B) Elastase staining of normal marrow. (C) Cathepsin G staining of Chediak-Higashimarrow. (D) Elastase staining of Chediak-Higashi marrow. Both enzymes are evident within granules of normalmarrow cells, but do not stain within the granules of Chediak-Higashi cells.
ates contained similar amounts of the elastasemRNA transcript relative to that of f-actin.
WESTERN IMMUNOBLOTTING
Cathepsin G and elastase were not detected inthe lysates from blood neutrophils isolated fromany of the three Chediak-Higashi patients,whereas both proteins (molecular weight about29 500) were evident in the normal neutrophils(fig 2). Figure 3 shows the results of westernblotting using lysates from the bone marrow
samples. Both normal and Chediak-Higashimarrow lysates contained cathepsin G andelastase protein.
CATHEPSIN G AND ELASTASE CONCENTRATIONS
IN BLOOD AND MARROW MYELOCYTES
The cathepsin G and elastase concentrationsin Chediak-Higashi blood neutrophil lysates,measured using ELISA or enzyme activity, werebelow the limits of detection of the assays.
Cathepsin G and elastase were detected,using ELISA, in the bone marrow from theChediak-Higashi patient. Cathepsin G was
measured at a concentration of0X23 pg/myeloidcell, compared with a concentration in normalmarrow of 0-97 pg/cell. Elastase was also pres-ent in the Chediak-Higashi marrow at a con-
centration of 0-37 pg/myeloid cell, comparedwith the normal marrow value of 2 3 pg/cell.Thus, Chediak-Higashi marrow myeloid cellscontained 24 and 16% of the cathepsin Gand elastase proteins, respectively, found in thenormal marrow. Cathepsin G activity in themarrow cells represented 0 09 pg/cell inChediak-Higashi marrow and 0-93 pg/cell innormal marrow. The elastase activity inChediak-Higashi marrow was 0 15 pg/cell andwas 1-2 pg/cell in normal marrow. Thus,cathepsin G and elastase activity in Chediak-Higashi marrow cells represented 9-8 and15-8%, respectively, of that in normal marrow
cells.
l, 1,)
M32
on 4 July 2018 by guest. Protected by copyright.
http://mp.bm
j.com/
Clin M
ol Pathol: first published as 10.1136/m
p.48.1.M28 on 1 F
ebruary 1995. Dow
nloaded from

Chediak-Higashi azurophil proteinases
IMMUNOHISTOCHEMISTRY OF CYTOSPINPREPARATIONSFigure 4 shows the results of immuno-histochemical staining of bone marrow, fromthe Chediak-Higashi patient and a normaldonor, for cathepsin G. In the normal marrow
cathepsin G could be seen within the granulesof the cells, whereas the Chediak-Higashi pro-
myelocyte granules showed no staining.
DiscussionIn the present study we have confirmed theobservation of Ganz et al' that elastase andcathepsin G are undetectable in mature bloodneutrophils from patients with Chediak-Higashi disease. The fundamental cause ofneu-trophil cathepsin G and elastase deficiency inChediak-Higashi syndrome has not, however,been established. Elastase, coded for on chro-mosome 19pter'8 and cathepsin G, on chro-mosome 1lq14 2,1" are both transcribed,synthesised and packaged within the pro-
myelocyte during myeloid differentiation in thebone marrow,"' 1920 when the azurophil granulesare formed21; transcription and synthesis cease
before the cells become mature neutrophils.Both enzymes are translated as pre-pro-pro-
teins and thence processed via enzymaticallyinactive pro-proteins to the mature active en-
zymes.22 Activation from the pro-enzyme ap-
pears to involve the proteolytic removal of an
acidic N terminal dipeptide, Ser-Glu in thecase of elastase and Gly-Glu for cathepsin G,and C terminal extensions of 20 and 11 aminoacids in elastase and cathepsin G, respectively.The N terminal sequences of the mature pro-teins share considerable homology, suggestingcommon dipeptidyl peptidases are responsiblefor processing the pro-proteins.22 The C ter-minal extensions of the pro-proteins are nothomologous. The mechanisms determiningpackaging to the azurophil granules are notknown. The deficiency of these enzymes inChediak-Higashi syndrome could potentiallyoccur at any of these several stages of pro-cessing. The identification of the mature en-
zymes within bone marrow promyelocytes ofthe beige mouse suggested, however, that tran-scription of the genes for these enzymes andtranslation of the proteins do occur. Thus, thedeficiency would be a consequence of post-translational events.
In the present study we have shown that inbone marrow from a patient with Chediak-Higashi syndrome the levels of the mRNAtranscripts for cathepsin G and elastase, whencorrected for ,B-actin mRNA levels, were similarto those of normal marrow. Thus, if 1-actintranscription is normal in Chediak-Higashimyeloid cells the results would suggest thattranscription both of cathepsin G and elastaseis also normal.The bone marrow myeloid cells of the
patients with Chediak-Higashi syndrome con-
tained mature, active elastase and cathepsin G.As in the beige mouse, the concentrations ofthese enzymes were lower than those ofmarrowfrom a normal subject and undetectable inmature blood neutrophils. The Chediak-
Higashi marrow protein concentrations wereonly about one fifth of those seen in normalmarrow. This is less than the value for the beigemouse marrow, which was reported to be about60% of normal.6Enzyme activity measurements in marrow
samples were slightly lower than the proteinconcentrations estimated using ELISA. Thisdifference may represent a technical dis-crepancy. Alternatively, it is possible that someinactive, unprocessed, pro-elastase and pro-cathepsin G were present within the marrowcells, although there was no evidence for thison the western blots. The western blots ofelastase and cathepsin G from Chediak-Higashi bone marrow samples showed, in ad-dition to molecular weight bands consistentwith the mature proteins, smaller forms whichwere not observed in lysates of normal marrowor of blood neutrophils. These small formscould represent differences in glycosylation.Alternatively, they might represent proteolysedforms although proteinase inhibitors wereadded to the marrow and neutrophils beforelysis to prevent proteolytic modification of pro-teins during the processing of samples for blot-ting. The presence of the "smaller" form ofelastase and cathepsin G in Chediak-Higashimarrow suggests, therefore, that some proteo-lytic degradation may have occurred beforesampling-that is, in vivo. This has relevanceto the possible fate of these proteins inChediak-Higashi marrow, but further studieswould be necessary to establish the reasons forthis observation. Nevertheless, the presence ofactive elastase and cathepsin G in the myeloidcells of the Chediak-Higashi marrow showsthat these proteins had been synthesised andprocessed to mature, active enzymes. This in-dicates that a deficiency in proteinases involvedin processing of the pro-enzymes is not re-sponsible for the absence of elastase and cath-epsin G in Chediak-Higashi neutrophils.Takeuchi and Swank23 characterised in-
hibitors of elastase and cathepsin G, whichformed covalent complexes with the enzymes,in mature neutrophils of beige mice; these in-hibitors were absent from normal mouse cellsand beige mouse pro-myelocytes. It was sug-gested that the absence of elastase and ca-thepsin G activities in beige mouse neutrophilswas because of the presence of these inhibitors.This would not, however, explain the absenceof immunoreactive elastase and cathepsin G inmature neutrophils, either as native proteins orcomplexed with putative inhibitors. In ad-dition, the presence of elastase and cathepsinG within marrow myeloid cells, but not ap-parently within the cells' granules, suggeststhat these enzymes are absent from matureneutrophils in Chediak-Higashi syndrome be-cause of a failure of transport to, or packagingwithin, the primary granules of the marrowpromyelocytes.
Granule defects are a common feature ofseveral cell types in Chediak-Higashi syn-drome, including cells which do not expresscathepsin G or elastase.1 This suggests that afundamental defect in granule structure may
M33
on 4 July 2018 by guest. Protected by copyright.
http://mp.bm
j.com/
Clin M
ol Pathol: first published as 10.1136/m
p.48.1.M28 on 1 F
ebruary 1995. Dow
nloaded from

Burnett, Ward, Stockley, Dalton, Cant, Hoare, et al
prevent the packaging of some, but not all,neutrophil azurophil proteins. Presumablyelastase and cathepsin G, but not myelo-peroxidase or the defensins, share commonrequirements for packaging. Little is known,however, about the mechanisms responsible forintracellular transport of azurophil proteins. Itis likely that failure to package cathepsin G orelastase successfully may result in these proteinsbeing degraded intracellularly, resulting in theirabsence from mature neutrophils, followingcessation of synthesis. This would explain thesmaller molecular weight forms of these en-
zymes observed on the western blots from theChediak-Higashi bone marrow. Should thishypothesis be correct, the detection of cath-epsin G and elastase activity within Chediak-Higashi marrow cells would suggest that post-translational processing of these enzymes takesplace before the proteins enter the azurophilgranules. The site of processing events has, todate, been unclear.22
In conclusion, our results suggest that theabsence of elastase and cathepsin G inChediak-Highashi blood neutrophils resultsfrom a primary defect in azurophil granulestructure which precludes the successful pack-aging of these enzymes, but permits that ofother azurophil granule proteins. Further stud-ies are indicated to identify the fundamentaldefect in Chediak-Higashi disease and to es-
tablish whether failure of phagolysosome form-ation or the deficiencies of specific granuleproteins are responsible for defective bacterialkilling by leucocytes in this disease.
Supported by grants from the British Lung Foundation, theBirmingham Hospitals Endowment Fund and the General Hos-pital Bicentenary Fund.
1 Windorst DB, Pagett GA. The Chediak-Higashi syndromeand the homologous trait in animals. _7 Invest Dernmatol1 968;60:529-37.
2 Root RK, Rosenthal AS, Balestra DJ. Abnormal bactericidal,metabolic, and lysosomal functions of Chediak-Higashisyndrome leukocytes. .7 Clin Invest 1972;51:649-65.
3 Vassali JD, Pipemo-Granelli A, Griscelli C, Reich E. Specificprotease deficiency in polymorphonuclear leukocytes ofChediak-Higashi syndrome and beige mice. Exp Med
1 978;147: 1285-90.4 Ganz T, Metcalf JA, Gallin JI, Boxer LA, Lehrer RI. Mi-
crobicidal/cytotoxic proteins of neutrophils are deficient intwo disorders: Chediak-Higashi syndrome and "specific"granule deficiency. . Clin Invest 1988;82:522-6.
5 Gilbert CS, Parmley RT, Rice WG, Kinkade JM. Hetero-geneity of peroxidase-positive granules in normal humanand Chediak-Higashi neutrophils. 7 Histochemn Cvtochen1993;41:837-49.
6 Takeuchi K, Wood H, Swank RT. Lysosomal elastase andcathepsin G in beige mice. Neutrophils of beige (Chediak-Higashi) mice selectivelv lack lvsosomal elastase and ca-thepsin G. Y Exp Med 1986;163:665-77.
7 Takeuchi KH, McGarry MP, Swank RT. Elastase and ca-thepsin G activities are present in immature bone marrowneutrophils and absent in late marrow and circulatingneutrophils of beige (Chediak-Higashi) mice. 7 Exp Mcd1987;166: 1362-76.
8 Jepsen LV, Skottun TA. A rapid one-step method for theisolation of human granulocytes from whole blood. ScandY7 Clin Lab Inivest 1982;42:235-8.
9 Chomczynski P. Solubilization in formamide protects RNAfrom degradation. Nrucleic Acids Res 1992;14:3791-2.
10 Farley D, Salvesen G, Travis J. Molecular cloning ofhumanneutrophil elastase. Biol Chenm Hoppe-Secler 1988;369(Suppl) :3-7.
11 Hohn PA, Popescu NC, Hanson RD, Salvesen G, Lev TJ.Genomic organisation and chromosomal localization ofthe human cathepsin G gene. 7 Biol Chemii 1989;264:13412-19.
12 Ponte P, Ng S-Y, Engel J, Gunning P, Kedes L. Evolutionaryconservation of the untranslated regions of actin mRNAs:DNA sequence of a human beta-actin cDNA. NucleicAcids Res 1984;12:1687-96.
13 Maniatis T, Fritsch EF, Sambrook J. Molecular cloning.A laboratory 7mianual. Cold Spring Harbor: Cold SpringHarbor Press, 1982.
14 Laemmli UK. Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature 1970;227:680-5.
15 Crocker J, Jenkins R, Burnett D. Immunohistochemicaldemonstration of leucocvte elastase in human tissues. 7Clin Pathol 1984;37:1114-18.
16 Crocker J, Jenkins R, Burnett D. Immunohistochemicallocalisation of cathepsin G in human tissues. AnmY SuirgPathol 1985;9:338-43.
17 Martodam RR, Baugh RJ, Twumasi DY, Liener IE. A rapidprocedure for the large scale purification of elastase andcathepsin G from human sputum. Prep Biochenm 1979;9:15-31.
18 Zimmer M, Medcalf RL, Fink TM, Mattmann C, LichterP, Jenne DE. Three human elastase-like genes coordinatelyexpressed in the myelomonocyte lineage are organized asa single genetic locus on 19pter. Proc Natl Acad Sci USA1992;89:8215-19.
19 Heusel JW, Scarpati EM, Jenkins NA, Gilbert DJ, CopelandNG, Shapiro SD, et al. Molecular cloning, chromosomallocation and tissue-specific expression of the murine ca-thepsin G gene. Blood 1993;81:1614-23.
20 Fouret P, duBois RM, Bernaudin JF, Takahashi H, FerransVJ, Crvstal RG. Expression of the neutrophil elastase geneduring human bone marrow cell differentiation. Y ExpMed 1989;160:833-45.
21 Bainton DF, Ullyot JL, Farquhar MG. The developmentof neutrophilic polymorphonuclear leucocytes in humanbone marrow: origin and content of azurophil and specificgranules. .T Exp Med 197 1;134:907-34.
22 Salvesen G, Enghild JJ. An unusual specificity in the ac-tivation of neutrophil serine proteinase zymogens. Bio-chemistry 1990;29:5304-8.
23 Takeuchi KH, Swank RT. Inhibitors of elastase and ca-thepsin G in Chediak-Higashi (beige) neutrophils. Y7 ClinInvest 1989;254:7431-6.
M34
on 4 July 2018 by guest. Protected by copyright.
http://mp.bm
j.com/
Clin M
ol Pathol: first published as 10.1136/m
p.48.1.M28 on 1 F
ebruary 1995. Dow
nloaded from