complete x fromhumanchorionic villi dna - pnas fileproc. natl. acad. sci. usa vol. 83, pp....
TRANSCRIPT

Proc. Natl. Acad. Sci. USAVol. 83, pp. 2182-2186, April 1986Genetics
Complete reactivation of X chromosomes from human chorionic villiwith a switch to early DNA replication
(dosage compensation/DNA methylation/somatic cell hybrids)
BARBARA R. MIGEON, MALGORZATA SCHMIDT, JOYCE AXELMAN, AND CATHERINE RUTA CULLEN
Department of Pediatrics, The Johns Hopkins University School of Medicine, Baltimore, MD 21205
Communicated by John W. Littlefield, November 22, 1985
ABSTRACT Mammalian sex-dosage compensation is me-diated by maintaining activity of only one X chromosome. Theasynchronous DNA synthesis characterizing the silent human Xchromosome is thought to be reversible only during ontogenyof oocytes. We have previously shown that the glucose-6-phosphate dehydrogenase (G6PD) locus (G6PD) on the al-locyclic X chromosome in chorionic villi is partially expressed.We now show that in hybrids derived from a clone of chorionicvilli cells (heterozygous for G6PD A) and mouse A9 cells, theloci for G6PD, hypoxanthine phosphoribosyltransferase(HPRT) and phosphoglycerate kinase are expressed on bothhuman X chromosomes; the human X chromosomes carryingeither G6PD A or B replicate synchronously with each otherand with murine chromosomes. The X chromosome with G6PDA was identified as the original late-replicating X, becausemethylation in the body of the HPRT gene on this chromosomeremained characteristic of the inactive X chromosome. Theseresults indicate that X-chromosome inactivation is completelyreversible in cells of trophoblast origin; induction of fulltranscriptional activity is accompanied by acquisition ofisocyclic replication, showing an intimate relationship betweenthese processes. The molecular events responsible for thisreversal may be similar to those occurring during maturationof oocytes. Chorionic vilii and derivative hybrids provide invitro models for exploring early events that program the singleactive X chromosome.
In mammals, the sex difference in dosage ofX chromosomesis compensated by silencing all but one X chromosome infemale somatic cells (1). Dosage compensation is not estab-lished simultaneously in all tissues (2) but is programmedalong with other tissue-specific functions. At the time oftissue differentiation, only one X chromosome achievespotential transcriptional activity; the others become inactive,condense at interphase, and replicate asynchronously [laterthan the autosomes and the active X chromosome (3)]. Theinactivity of the allocyclic X chromosome is rigorouslymaintained (4), and spontaneous reversal of the process-i.e., reactivating the entire chromosome and reversing al-locyclic replication-has not been observed in somatic cells.The spontaneous reactivation events that have been noted (5,6) affect some loci but not others, and the chromosomeremains late-replicating (5). On the other hand, reactivationof the entire X chromosome is believed to occur regularlyduring the ontogeny of oocytes. Primordial germ cells in theyolk sac of female rodents have a condensed X chromosome(7), and there is evidence from mouse embryos that glucose-6-phosphate dehydrogenase (G6PD) genes on both X chro-mosomes are expressed shortly after germ cells am've at thegonadal ridges, whereas only one locus was active previously(8).
Although recent observations provide insights into main-tenance ofX chromosome dosage compensation (9-11), littleis known about initiation of the process. Determining theinitial steps in X chromosome differentiation-those thatprogram only a single X chromosome-has been difficultbecause the relevant stages are not readily accessible toexperimental manipulation. We recently reported that theG6PD locus on allocyclic X chromosomes is variably ex-pressed in chorionic villi of the human placenta, based onobservations of cloned diploid cells (12); while some cellsexpress only a single allele, others express the two, but onemore prominently than the other. In either case, the relevantchromosome remains late-replicating. We now report that thepartially active X chromosome from this extraembryonictissue oftrophoblastic origin begins to replicate synchronous-ly and appears to undergo complete reactivation whenintroduced into the milieu of the interspecies hybrid cell.
STRATEGYThe discovery that G6PD alleles on late-replicating X chro-mosomes in chorionic villi were at least partially expressedled us to look for other "active" genes on the "inactive"chromosome. To segregate the relevant X chromosome fromits homologue, we used mouse-human hybrids. The chori-onic villi used for fusion were heterozygous for the commonelectrophoretic variants of G6PD so that each human Xchromosome was marked with a unique form of the enzyme(G6PD A and G6PD B). Because these cells originated froma clone, the population was homogeneous with respect towhich of the two chromosomes was the active one (13). Inthis clone (NB1B-6), the predominant enzyme was G6PD B,whereas G6PD A was the minor variant, and the ABheterodimer was of intermediate abundance (Fig. 1, arrow).The parental mouse cells lacked hypoxanthine phosphori-bosyltransferase (HPRT), so that only hybrids retaining ahuman X chromosome with an active HPRT gene couldsurvive in hypoxanthine/arminopterin/thymidine (HAT) se-lective medium (14). Therefore, the G6PD phenotype of thesehybrids would reveal ifthe HPRTlocus on one or both humanX chromosomes was functional. If the HPRT locus on thelate-replicating X chromosome were active, then hybridswith this chromosome could be examined for activity at otherloci on the chromosome.
MATERIALS AND METHODSCells and Tissues. Cultures were established from the
chorionic villi of a full-term placenta heterozygous for G6PDA. These cells were cloned (5) by plating 10 cells per 60-mmtissue culture dish, and one of the clones (NB1B-6) was usedas the human parent for hybrids. Cells from NB1B-6 had 46
Abbreviations: G6PD, glucose-6-phosphate dehydrogenase; HAT,hypoxanthine/aminopterin/thymidine; HPRT, hypoxanthine phos-phoribosyltransferase; PGK, phosphoglycerate kinase.
2182
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.

Proc. Natl. Acad. Sci. USA 83 (1986) 2183
A
1 2 3 4 5 6 7 8 9 10
FIG. 1. Cellulose acetate electrophoresis, showing the G6PDphenotype of the human chorionic villi clone (NB1B-6) (lane 2,arrow) and ofNBlB-6-A9 hybrids. Controls were mixtures of humanG6PD A and G6PD B (lanes 1, 5, and 8) and mouse A9 (lane 4).Hybrids expressing humanG6PD B and G6PD B-mouse heterodimer(lanes 3 and 9), a hybrid with human G6PD A and G6PD A-mouseheterodimer (lane 7), and hybrids expressing G6PD AB, includingG6PD AB, G6PD A-mouse, and G6PD B-mouse heterodimers Oanes6 and 10) are shown. Note that the G6PD B-mouse heterodimermigrates close to G6PD A. The intensity of the lowest band (mouseenzyme) in lane 10 was reduced during the photographic process toshow the human G6PD A-mouse heterodimer.
normal chromosomes including allocyclic as well as early-replicating X (see below).Hybrids were obtained from-fusion of NB1B-6 with A9
mouse fibroblasts by selection in HAT medium (5).Chromosome Studies. To determine the kinetics of chro-
mosome replication, cultures of chorionic villi were placed inmedium containing BrdUrd (10 ,g/ml) for one cell cycle,which was replaced with medium containing thymidine (25,ug/ml) during the final 6-8 hr before harvest (15). Hybridcells were maintained in HAT medium for these studies,
except that BrdUrd was substituted for thymidine for one cellcycle, and the medium was replaced by HAT mediumenriched with thymidine (25 ug/ml) during the final 6-8 hrbefore harvest.Enzyme Studies. Cell extracts prepared as described (12)
were analyzed for G6PD (12), phosphoglycerate kinase(PGK) (16), and HPRT (17) by cellulose acetate gel electro-phoresis.DNA Studies. DNA was purified from chorionic villi,
cultures of chorionic villi, and hybrid cells as described (9).For methylation studies, DNA was digested with Hpa II/BamHI or with Hpa II/Pst I, blotted, transferred to nitro-cellulose, and hybridized with nick-translated HPRT probepPB1.7 (18) according to standard methods as described (9).
RESULTS
Characteristics of Hybrid Clones. Enzyme phenotype. Hy-brid clones, picked -3 wk after fusion, were assayed for Xchromosome-linked enzymes as soon as enough cells wereavailable (2nd-3rd subculture). The G6PD phenotype washeterogenous (Fig. 1; Table 1); eight hybrids expressed G6PDA, seven expressed G6PD B, and four had both G6PD A andB and the AB heterodimer. As hybrids were selected in HATmedium, all had human HPRT (Fig. 2 Left). Therefore, theheterogeneous G6PD phenotype of these hybrids suggestedthat both human HPRT genes (one coupled with G6PDA andthe other with G6PD B) were functional. This interpretationis supported by the following evidence. The presence of ABheterodimers in four hybrids showed that both G6PD alleleswere in the same cell. The two human X chromosomes fromhybrid 70C, which expressed both human G6PD allelessegregated randomly, as expected ifboth chromosomes were
Table 1. Characteristics of human-mouse hybrid clones
Human enzymes expressedG6PD No. of human chromosomes Percent of cells with
No. of cells with iso- or allocyclic XClone A B AB* PGK HPRT Total X Translocations intact human X Isocyclic Allocyclic
30A + + + 0-1 0-1 2-350 + + + 0-1 0-1 0-170A + + + 5-9 0-1 1100B + + + 1-8 1 2-4102 + + + 0-1 0 1110A 4sc + + + 1 100 tOO110D 3sc + + -1+ 1
7sc +120C 3sc + + ND 5-11 1-2 20 100 010 + - + 0 0 0-130B + + ND 0-2 0-1 0-140 + + + 9-10 1-2 0-160 + + + 2-3 1 170B + + + 2-4 1 180 + + + 6-11 1 0-190 + + + 3-5 1 1-2 29 100 070C 4sc + + + + + 8-11 1-2 1-2 10 100 0
7sc + + + 40 100 (20)t 09sc +
100A + + + + + 1-5 1-2 1120A 4sc + + + + + 4 1-2 50 100 (20)t 0
6sc + + + 20 100 0120B 2sc + + + + + 3-12 1-2
Ssc + + + 100 100 (40)t 0lOsc +
sc, Subculture; ND, not determined.*AB heterodimer.tNumbers in parentheses represent percent with 2 isocyclic X chromosomes.
Genetics: Migeon et al.
Proc. Natl. Acad. Sci. USA 83 (1986)
An::n,|w-w^ A.
-By
.s*:X
1 2 3 4 5 6 1 2 3 4
FIG. 2. Cellulose acetate electrophoresis, showing enzyme phe-notype ofNB1B-6-A9 hybrid clones. (Left) Autoradiograph showingHPRT. Lanes: 1, hybrid Qlone 40 with G6PD B; 2, clone 80 withG6PD B; 3, human/mouse mix; 4, clone 30A with G6PD A; 5, clonelOOB with G6PD B; 6, mouse A9. (Right) Autoradiograph showingPGK. Lanes: 1, Human control; 2, hybrid 90 with G6PD B; 3, hybrid102A with G6PD A; 4, human/mouse mix.
functional: four subclones had only G6PD A, six had onlyG6PD B, and five continued to express both alleles. Further-more, when hybrids containing a single human X chromo-some (11OA expressing G6PD A and 70B with G6PD B) wereplated into medium containing 6-thioguanine, the 6-thiogua-nine-resistant cells in each case lost human G6PD as well asHPRT. This showed that G6PD and HPRT were syntenic,and that each G6PD allele segregated with an active HPRTgene.
Further analysis ofthese hybrids revealed that not only theHPRT and G6PD loci but also the PGK locus was functionalon both human X chromosomes. Except for clone 10 thatlacked an intact X chromosome, all hybrids had human PGK,irrespective of the human G6PD allele expressed (Fig. 2Right).DNA replication. We suspected that G6PD A was the
product of the allocyclic X chromosome, as the parent clonehad less G6PD A than G6PD B (Fig. 1). However, X-linkedenzymes were similarly expressed (based on relative inten-sity of electrophoretic bands) in hybrids with G6PD A orG6PD B (see Fig. 2), so that we no longer could distinguishone X chromosome from the other by the level of enzymeactivity. To determine if the chromosome with G6PD A was,in fact, the allocyclic X chromosome, we examined thereplication of the X chromosomes in these hybrids. By meansof terminal-thymidine or BrdUrd labeling (15), we observed
a
0 _*_ ::
~.o 4.#-
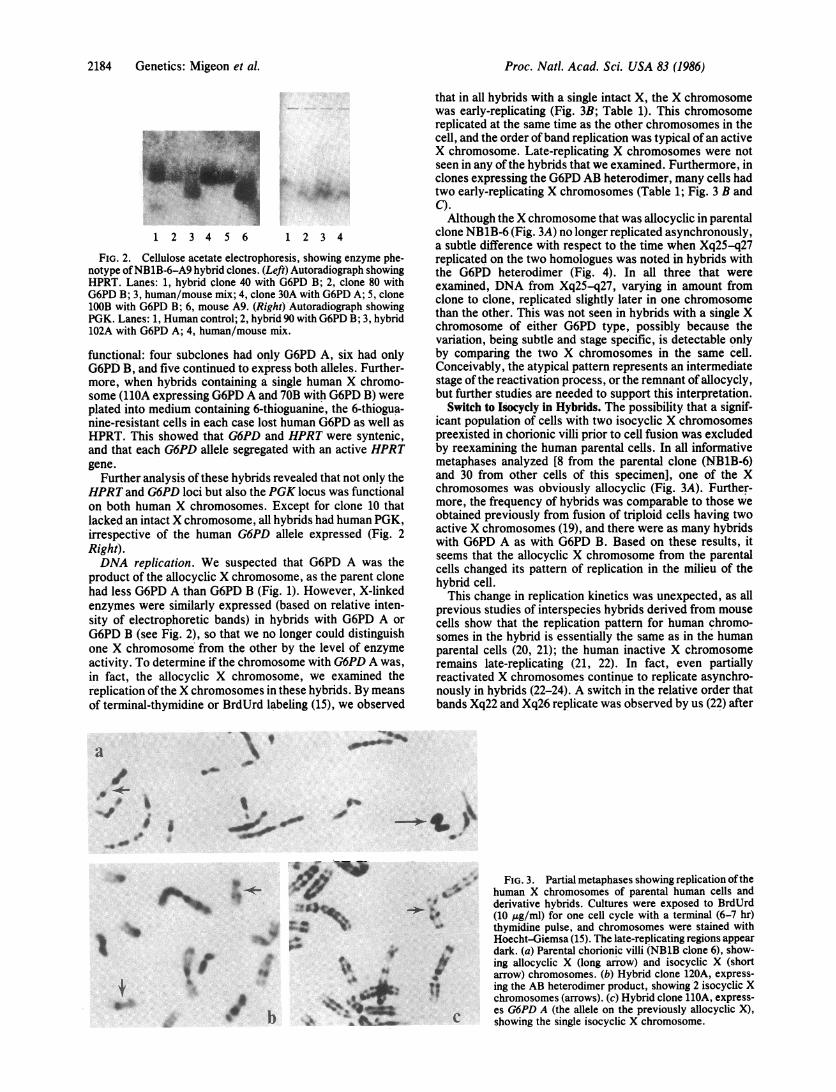
that in all hybrids with a single intact X, the X chromosomewas early-replicating (Fig. 3B; Table 1). This chromosomereplicated at the same time as the other chromosomes in thecell, and the order ofband replication was typical of an activeX chromosome. Late-replicating X chromosomes were notseen in any of the hybrids that we examined. Furthermore, inclones expressing the G6PD AB heterodimer, many cells hadtwo early-replicating X chromosomes (Table 1; Fig. 3 B andC).Although the X chromosome that was allocyclic in parental
clone NB1B-6 (Fig. 3A) no longer replicated asynchronously,a subtle difference with respect to the time when Xq25-q27replicated on the two homologues was noted in hybrids withthe G6PD heterodimer (Fig. 4). In all three that wereexamined, DNA from Xq25-q27, varying in amount fromclone to clone, replicated slightly later in one chromosomethan the other. This was not seen in hybrids with a single Xchromosome of either G6PD type, possibly because thevariation, being subtle and stage specific, is detectable onlyby comparing the two X chromosomes in the same cell.Conceivably, the atypical pattern represents an intermediatestage of the reactivation process, or the remnant of allocycly,but further studies are needed to support this interpretation.
Switch to Isocycly in Hybrids. The possibility that a signif-icant population of cells with two isocyclic X chromosomespreexisted in chorionic villi prior to cell fusion was excludedby reexamining the human parental cells. In all informativemetaphases analyzed [8 from the parental clone (NB1B-6)and 30 from other cells of this specimen], one of the Xchromosomes was obviously allocyclic (Fig. 3A). Further-more, the frequency of hybrids was comparable to those weobtained previously from fusion of triploid cells having twoactive X chromosomes (19), and there were as many hybridswith G6PD A as with G6PD B. Based on these results, itseems that the allocyclic X chromosome from the parentalcells changed its pattern of replication in the milieu of thehybrid cell.
This change in replication kinetics was unexpected, as allprevious studies of interspecies hybrids derived from mousecells show that the replication pattern for human chromo-somes in the hybrid is essentially the same as in the humanparental cells (20, 21); the human inactive X chromosomeremains late-replicating (21, 22). In fact, even partiallyreactivated X chromosomes continue to replicate asynchro-nously in hybrids (22-24). A switch in the relative order thatbands Xq22 and Xq26 replicate was observed by us (22) after
. '
ii
JO.30 .#
*:0
.....tq #
:*^~~ ~ ~ .;" H
.I
+'..
0.d .#P
Stew*.~~~~~~~~~~~~1..I@ots, IS~j. b
FIG. 3. Partial metaphases showing replication oftheY+ human X chromosomes of parental human cells and
derivative hybrids. Cultures were exposed to BrdUrd(10 Ag/ml) for one cell cycle with a terminal (6-7 hr)thymidine pulse, and chromosomes were stained withHoecht-Giemsa (15). The late-replicating regions appeardark. (a) Parental chorionic villi (NB1B clone 6), show-ing allocyclic X (long arrow) and isocyclic X (shortarrow) chromosomes. (b) Hybrid clone 120A, express-ing the AB heterodimer product, showing 2 isocyclic Xchromosomes (arrows). (c) Hybrid clone 11OA, express-es G6PD A (the allele on the previously allocyclic X),
C showing the single isocyclic X chromosome.
2184 Genetics: Migeon et al.
001W
I
0 4L40.
...0
Proc. Natl. Acad. Sci. USA 83 (1986) 2185
I t i
1 2 3 4 5 6 7
Im
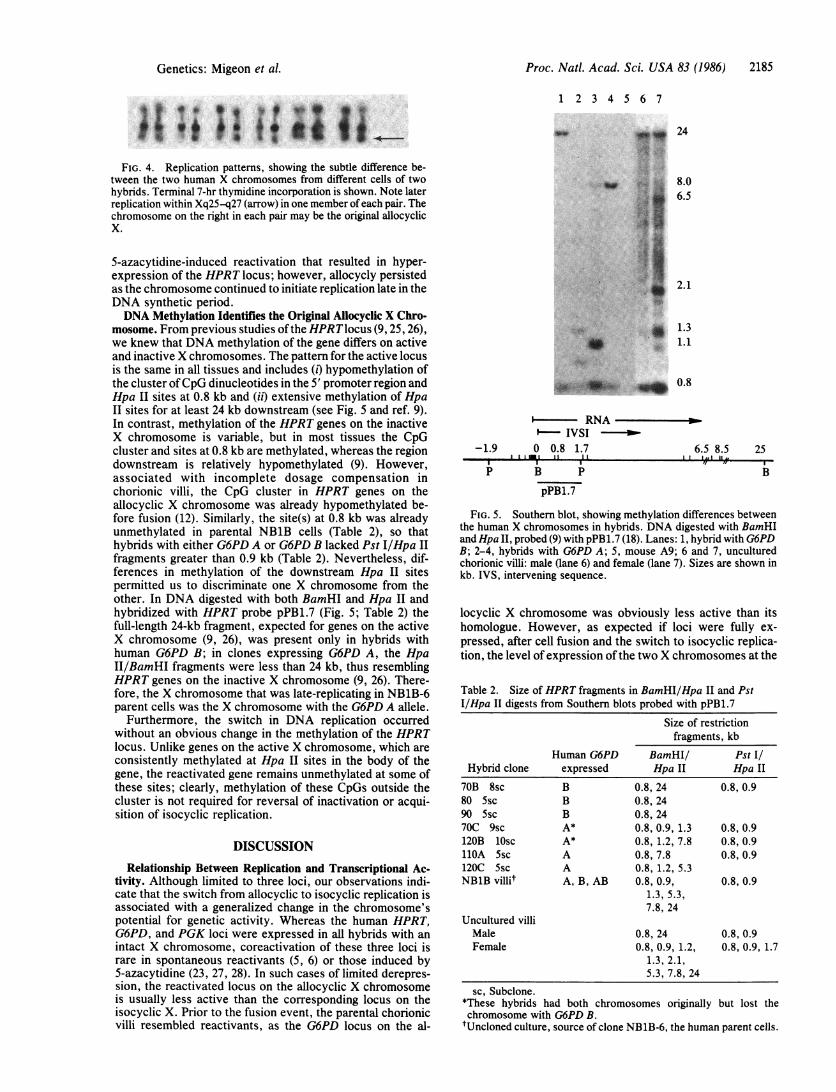
FIG. 4. Replication patterns, showing the subtle difference be-tween the two human X chromosomes from different cells of twohybrids. Terminal 7-hr thymidine incorporation is shown. Note laterreplication within Xq25S-q27 (arrow) in one member of each pair. Thechromosome on the right in each pair may be the original allocyclicX.
5-azacytidine-induced reactivation that resulted in hyper-expression of the HPRT locus; however, allocycly persistedas the chromosome continued to initiate replication late in theDNA synthetic period.DNA Methylation Identifies the Original Allocyclic X Chro-
mosome. From previous studies ofthe HPRTlocus (9, 25, 26),we knew that DNA methylation of the gene differs on activeand inactive X chromosomes. The pattern for the active locusis the same in all tissues and includes (i) hypomethylation ofthe cluster ofCpG dinucleotides in the 5' promoter region andHpa II sites at 0.8 kb and (ii) extensive methylation of HpaII sites for at least 24 kb downstream (see Fig. 5 and ref. 9).In contrast, methylation of the HPRT genes on the inactiveX chromosome is variable, but in most tissues the CpGcluster and sites at 0.8 kb are methylated, whereas the regiondownstream is relatively hypomethylated (9). However,associated with incomplete dosage compensation inchorionic villi, the CpG cluster in HPRT genes on theallocyclic X chromosome was already hypomethylated be-fore fusion (12). Similarly, the site(s) at 0.8 kb was alreadyunmethylated in parental NB1B cells (Table 2), so thathybrids with either G6PD A or G6PD B lacked Pst I/Hpa IIfragments greater than 0.9 kb (Table 2). Nevertheless, dif-ferences in methylation of the downstream Hpa II sitespermitted us to discriminate one X chromosome from theother. In DNA digested with both BamHI and Hpa II andhybridized with HPRT probe pPB1.7 (Fig. 5; Table 2) thefull-length 24-kb fragment, expected for genes on the activeX chromosome (9, 26), was present only in hybrids withhuman G6PD B; in clones expressing G6PD A, the HpaII/BamHI fragments were less than 24 kb, thus resemblingHPRT genes on the inactive X chromosome (9, 26). There-fore, the X chromosome that was late-replicating in NB1B-6parent cells was the X chromosome with the G6PD A allele.
Furthermore, the switch in DNA replication occurredwithout an obvious change in the methylation of the HPRTlocus. Unlike genes on the active X chromosome, which areconsistently methylated at Hpa II sites in the body of thegene, the reactivated gene remains unmethylated at some ofthese sites; clearly, methylation of these CpGs outside thecluster is not required for reversal of inactivation or acqui-sition of isocyclic replication.
DISCUSSIONRelationship Between Replication and Transcriptional Ac-
tivity. Although limited to three loci, our observations indi-cate that the switch from allocyclic to isocyclic replication isassociated with a generalized change in the chromosome'spotential for genetic activity. Whereas the human HPRT,G6PD, and PGK loci were expressed in all hybrids with anintact X chromosome, coreactivation of these three loci israre in spontaneous reactivants (5, 6) or those induced by5-azacytidine (23, 27, 28). In such cases of limited derepres-sion, the reactivated locus on the allocyclic X chromosomeis usually less active than the corresponding locus on theisocyclic X. Prior to the fusion event, the parental chorionicvilli resembled reactivants, as the G6PD locus on the al-
24
8.06.5
2.1
ww'. 1.3
*i~: 1. 1..0
II *_-.~ 0.8
-1.9
p
i- RNAp- IVSI0 0.8 1.7 6..
II IttI 1 IIt L
B PpPB1.7
.5 8.5 25,.., ,,I.
FIG. 5. Southern blot, showing methylation differences betweenthe human X chromosomes in hybrids. DNA digested with BamHIand Hpa II, probed (9) with pPB1.7 (18). Lanes: 1, hybrid with G6PDB; 2-4, hybrids with G6PD A; 5, mouse A9; 6 and 7, unculturedchorionic villi: male (lane 6) and female (lane 7). Sizes are shown inkb. IVS, intervening sequence.
locyclic X chromosome was obviously less active than itshomologue. However, as expected if loci were fully ex-pressed, after cell fusion and the switch to isocyclic replica-tion, the level of expression ofthe two X chromosomes at the
Table 2. Size of HPRT fragments in BamHI/Hpa II and PstI/Hpa II digests from Southern blots probed with pPB1.7
Size of restrictionfragments, kb
Human G6PD BamHI/ Pst I/Hybrid clone expressed Hpa II Hpa II
70B 8sc B 0.8, 24 0.8, 0.980 Ssc B 0.8, 2490 5sc B 0.8, 2470C 9sc A* 0.8, 0.9, 1.3 0.8, 0.9120B lOsc A* 0.8, 1.2, 7.8 0.8, 0.9110A Ssc A 0.8, 7.8 0.8, 0.9120C Ssc A 0.8, 1.2, 5.3NB1B villit A, B, AB 0.8, 0.9, 0.8, 0.9
1.3, 5.3,7.8, 24
Uncultured villiMale 0.8, 24 0.8, 0.9Female 0.8, 0.9, 1.2, 0.8, 0.9, 1.7
1.3, 2.1,5.3, 7.8, 24
sc, Subclone.*These hybrids had both chromosomes originally but lost thechromosome with G6PD B.tUncloned culture, source of clone NB1B-6, the human parent cells.
Genetics: Migeon et al.
';

Proc. Natl. Acad. Sci. USA 83 (1986)
three loci we examined did not obviously differ. That induc-tion of full activity is associated with a switch to earlierreplication supports the hypothesis of a relationship betweenthe potential for transcriptional activity and the kinetics ofchromosome replication (reviewed in ref. 29). Localizedreactivation affecting some loci but not others may beaccompanied by changes in the order in which specificregions ofthe chromosome replicate (22) but does not reverseallocycly. To reverse asynchronous replication of the entirechromosome seems to require more extensive changes orperhaps another mechanism-e.g., derepression of a specificregion ofthe chromosome. Determining the time that specificloci replicate before and after reactivation should help toclarify the relationship between replication and gene activity.
Reversibility of Inactivation Characterizes Early Differenti-ating Tissues. The only other evidence consistent with com-plete reversal of X chromosome inactivation comes fromstudies of intraspecies mouse hybrids derived from malignantembryonal carcinoma cells (30). These tumor cells inducereactivation of a previously silent PGK locus and isocyclicreplication of the inactive X chromosome in fetal thymocytesand bone marrow cells, presumably because the tumororiginates from embryonic cells that have not yet undergonedosage compensation (31). Surprisingly, these teratocarci-noma stem cells do not induce reactivation when fused withother differentiated somatic cells (32, 33) or culturedheteroploid cells (34). On the other hand, the potential forisocyclic replication and full transcriptional activity seems tobe a consistent feature of cultured cells from chorionic villi,as dosage compensation is reversible in each of the threechorionic villi specimens that we have fused with A9 mousefibroblasts (unpublished observations). Unlike teratocarci-noma stem cells, chorionic villi are well differentiated, butdifferentiation of this extrembryonic tissue occurs earlierthan that of tissues from the embryo proper (2). Clearly, thereare tissue differences in the maintenance of X dosage com-pensation; the inactive state in germ cells and chorionic villiis locked in less firmly than in other tissues, where derepres-sion events occur rarely, and are piecemeal, rather thanaffecting the chromosome as a whole.
Tissue differences in stability of dosage compensationcould result from factors that are tissue specific or uniquelypresent (or absent) at specific stages of development, induc-ing (or inhibiting) reversal of the process. Such factors maybe more (or less) prevalent in ovarian tissue, where completereversal occurs regularly, than in chorionic villi, where geneson the inactive X chromosome are incompletely and variablyexpressed. Whereas the milieu of the chorionic villus is notsufficient to induce complete reversal, that provided by thehybrid cell can assure that reactivation occurs, not as a rareevent, but with regularity. Whether the induction of synchro-nous replication by fusion with A9 represents an extension ofthe events initiated in the chorionic villus or results fromnovel modifications contributed by the mouse genome re-mains to be seen. Clearly the hybrid cell environment is notsufficient in itself because fusion of these mouse cells withother human tissues does not reverse inactivation but onlyinduces infrequent and spotty reactivation. Demethylation ofspecific loci, along with the prolonged survival bestowed onthe human genome by hybridization, may play a role in
localized reactivation events; however, other explanationsmust be invoked for the frequent (and perhaps obligatory)reversal that takes place in hybrid cells of trophoblast origin.In any event, studies ofthe chorionic villi, derivative hybrids,and ovary should provide the means to identify the relevantfactors and their cellular localization.
We gratefully acknowledge G. Russell Evans III for assistancewith the DNA methylation studies and Dr. David C. Kaslow for hiscritical reading of this manuscript. This work was supported byNational Institutes of Health Grant HD05465. M.S. was a Fellow ofthe Epply Foundation.
1. Lyon, M. F. (1972) Biol. Rev. Cambridge Philos. Soc. 42, 1-35.2. Monk, M. & Harper, M. I. (1979) Nature (London) 281, 311-313.3. German, J. (1962) Trans. N. Y. Acad. Sci. 24, 395-407.4. Migeon, B. R. (1972) Nature (London) 239, 87-89.5. Migeon, B. R., Wolf, S. F., Mareni, C. & Axelman, J. (1982) Cell 29,
595-600.6. Kahan, B. & DeMars, R. (1975) Proc. Nati. Acad. Sci. USA 72,
1510-1514.7. Ohno, S. (1963) in Second International Conference on Congenital
Malformations, ed. Fishbein, M. (National Foundation, New York), pp.36-40.
8. Kratzer, P. G. & Chapman, V. M. (1981) Proc. Natl. Acad. Sci. USA78, 3093-3097.
9. Wolf, S. F., Jolly, D. J., Lunnen, K. D., Friedmann, T. & Migeon,B. R. (1984) Proc. Natl. Acad. Sci. USA 81, 2806-2810.
10. Wolf, S. F., Dintzis, S., Toniolo, D., Persico, G., Lunnen, K.,Axelman, J. & Migeon, B. R. (1984) Nucleic Acids Res. 12, 9333-9348.
11. Wolf, S. F. & Migeon, B. R. (1985) Nature (London) 314, 467-469.12. Migeon, B. R., Wolf, S. F., Axelman, J., Kaslow, D. C. & Schmidt, M.
(1985) Proc. NatI. Acad. Sci. USA 82, 3390-3394.13. Davidson, R. G., Nitowski, H. M. & Childs, B. (1963) Proc. Natl. Acad.
Sci. USA 50, 481-483.14. Littlefield, J. W. (1966) Exp. Cell Res. 41, 190-196.15. Schmidt, M., Stolzmann, W. M. & Baranovskaya, L. J. (1982)
Chromosoma 85, 405-412.16. Pai, G. S., Sprenkle, J. A., Do, T. T., Mareni, C. & Migeon, B. R.
(1980) Proc. Natl. Acad. Sci. USA 77, 2810-2813.17. Der Kaloustian, V. M., Byrne, R., Young, W. J. & Childs, B. (1969)
Biochem. Genet. 3, 299-302.18. Jolly, D. J., Esty, A. C., Bernard, H. U. & Friedmann, T. (1982) Proc.
Natl. Acad. Sci. USA 79, 5038-5041.19. Migeon, B. R., Sprenkle, J. A. & Do, T. T. (1979) Cell 18, 637-641.20. Farber, R. A. & Davidson, R. L. (1979) Somatic Cell Genet. 5, 519-528.21. Willard, H. F. in Cytogenetics of the Mammalian X Chromosome, Part
A, ed. Sandberg, A. (Liss, New York) p. 427.22. Schmidt, M., Wolf, S. F. & Migeon, B. R. (1985) Exp. Cell Res. 158,
301-310.23. Mohandas, T. T., Sparkes, R. S., Bishop, D. F., Desnick, R. J. &
Shapiro, L. J. (1984) Am. J. Hum. Genet. 36, 916-925.24. Jones, P. A., Taylor, S. M., Mohandas, T. & Shapiro, L. J. (1982) Proc.
Natl. Acad. Sci. USA 79, 1215-1219.25. Yen, P. H., Patel, P., Chinault, A. C., Mohandas, T. & Shapiro, L. J.
(1984) Proc. Natl. Acad. Sci. USA 81, 1759-1763.26. Vogelstein, B., Fearon, E., Hamilton, S. R. & Feinberg, A. P. (1985)
Science 227, 642-645.27. Hors Cayla, M. C., Heuertz, S. & Frezal, J. (1983) Somatic Cell Genet.
9, 645-657.28. Lester, S. C., Korn, N. J. & DeMars, R. (1982) Somatic Cell Genet. 8,
265-284.29. Goldman, M. A., Holmquist, G. P., Gray, M. C., Caston, L. A. & Nag,
A. (1984) Science 224, 686-692.30. Takagi, N., Yoshida, M. A., Sugawara, 0. & Sasaki, M. (1983) Cell 34,
1053-1062.31. Martin, G. R., Epstein, C. J., Travis, B., Tucker, G., Yatziv, S.,
Martin, D. W., Jr., Clift, S. & Cohen, S. (1978) Nature (London) 271,329-333.
32. Von Kap-Herr, C. & Mukherjee, B. B. (1977) Exp. Cell Res. 104,369-376.
33. Graves, J. A. M. & Young, G. J. (1982) Exp. Cell Res. 141, 87-97.34. McBurney, M. W. & Strutt, B. J. (1980) Cell 21, 357-364.
2186 Genetics: Migeon et al.