corpus luteum function and parturition in cattle and pigs in the cow and pig is preceeded by a...
TRANSCRIPT

Arch. Tierz., Dummerstorf 44 (2001) Special Issue, 37-50
Department of Farm Animal Health, University of Utrecht, The Nelherlands
MARCEL A. M. TAVERNE
Corpus Luteum function and parturition in cattle and pigs
Summary At term pregnancy in cattle and pigs progesterone is from luteal origin. Like in other domestic animal species, parturition in the cow and pig is preceeded by a sudden drop in the matemal plasma progesterone concentration. An increased Output of Prostaglandin from the pregnant uterus is most likely responsible for prepartum luteolysis. The withdrawal of progesterone is not immediately followed by activation ofthe myometrium; rather a period of (relative) myometrial quiescence allows for functional changes to take place in the cervix and myometrium, before parturient uterine contractions develop (between 12 and 14 hours, and between 6 and 9 hours before onset of the expulsive stage, in cows and pigs respectively). These will only subsequently cause a widening ofthe cervix. Maintenance of progesterone dominance, by treatment with exogenous progesterone/ progestagens or by induction of a new set of corpora lutea (pigs), at the time that parturition is initiated, will postpone delivery. However, uterine Prostaglandin output (cows) still increases under these conditions (cows) and many of the calves and piglets are bom dead when (prolonged) expulsion finally takes place after relief from progesteron. This indicates that progesterone is not able to counteract completely the parturition promoting signals generated by the pregnant uterus.
Key Words: parturition, corpus luteum, progesterone, induction of parturition, Prostaglandins, cervix, myometrium, cattle, pigs
Zusammenfassung Titel der Arbeit: Corpus luteum-Funktion und Geburt bei Rind und Schwein Zum Graviditätsende sind Corpora lutea die entscheidenden Progesteronproduzenten bei Rind und Schwein. Bei beiden Spezies geht der Geburt, wie bei anderen Haustieren auch, ein plötzlicher Abfall der Progesteronkonzentration voraus. Diese Luteolyse wird höchstwahrscheinlich durch Prostaglandin uterinen Ursprungs induziert. Die Verringerung der Progesteronkonzentration ist nicht sofort von einer Zunahme der Aktivität des Myometriums begleitet. Vielmehr erlaubt eine Phase der relativen Myometriuminaktivität funktionelle Veränderungen in der Zervix und im Myometrium, bevor die Geburtswehen beginnen. Diese treten beim Rind etwa 12 bis 14 h und beim Schwein etwa 6 bis 9 h vor der Austreibungsphase auf. Eine Aufrechterhaltung der Progesterondominanz zum Zeitpunkt des Geburtsbeginns, hervorgerufen durch die Verabreichung von Progesteron/Progestagenen bzw. durch die Induktion neuer Corpora lutea (Schwein) wird die Geburt verzögern. Jedoch kommt es auch unter diesen Bedingungen zu einem Anstieg der Prostaglandinkonzentrationen. Findet nach der Verringerung der Progesteronkonzentrationen dann die Austreibung statt, so steigt der Anteil totgeborener Kälber und Ferkel. Das zeigt, dass Progesteron nicht vollständig in der Lage ist, die von Uterus stammenden, geburtsfbrdernden Signale zu unterdrücken.
Schlüsselwörter: Geburt, Gelbkörper, Progesterone, Geburtsinduktion, Prostaglandinen, Cervix, Myometrium, Rind, Schwein
Introduction There are several reasons to study the endocrinology and physiology of parturition in domestic animals in general, and in cows and pigs in particular. While abortion or premature parturition may be caused by different factors, the underlying endocrine mechanisms of premature pregnancy interruptions can also be

38
quite different but usually they lead to (partial) cessation of progesterone production by luteal tissues. Conversely, prolonged gestation, which mainly occurs in some breeds of cattle (HOLM and SHORT, 1962) but which might also become evident in sows with very small numbers of piglets (RANDALL, 1987), is associated by a failure of or a delay in the demise of the Corpora lutea of pregnancy. Knowledge about the endocrine regulation of pregnancy and parturtion should provide us with tools to induce parturition, either for reasons of farm management or because of clinical indications. The pathophysiological mechanisms of several forms of dystocia, such as incomplete dilatation of the cervix and (presumed) uterine inertia or placental retention, should lead us to the development of more optimal ways to prevent or to treat clinical cases. The key role of progesterone, originating from the corpus luteum, in keeping the uterus closed and the importance of luteal regression to allow release of uterine contents, is elegantly illustrated by a model, the pseudopregnant goat. These animals do have a uterine content but no conceptus. In pseudopregnant goats the corpus luteum of an oestrous cycle persists and after about 30 days of exposure to progesterone, fluid Starts to accumulate in the uterus, sometimes reaching a volume of more than 12 liters. These cases of so-called hydrometra heal themselves when the corpus luteum caeses to produce progesterone. They can also be treated with exogenous Prostaglandin F2a, which causes regression of the persistent CL, relaxation of the cervix and uterine contractions, so that the uterine content is expelled (review by TAVERNE et al., 1995). This contribution wil not pay attention to the mechanisms leading to and involved in (prepartal) luteal regression itself (for which the reader is referred to the review by McCRACKEN et., 1999), but it will rather emphasize on data describing effects of manipulating corpus luteum function and progesterone production during late gestation in cows and pigs. These data will illustrate that luteal regression, leading to withdrawal of a progesterone influence, Starts a sequence of immediate prepartal events in such a way, that a window of time is created allowing physiological preparations for the birth process to occur.
Corpus luteum and progesterone around calving In cows the corpus luteum is the main source of progesterone throughout pregnancy. LH is known to be a major factor in the maintenance of CL-function in cattle (HOFFMANN et al., 1974) and basal concentrations and episodic release of LH were less after day 250 than at earlier stages of gestation (SCHALLENBERGER et al, 1985). There are, however, no indications for a change in LH secretion during the peripartal period (HOFFMANN et al., 1973). Tissue slices ofthe CL immediately post partum were unresponsive to LH, as judged by progesterone production in vitro, whereas tissues from prepartum CL responded to LH (GROSS et al., 1985). There is no evidence to suggest that the prepartum decline of maternal plasma progesterone concentrations, which Starts already several weeks before calving, but shows a more precipitous fall at about 30-40 hours before delivery (POPE et al., 1969; STABENFELDT et al., 1970; EDQVIST et al., 1978), is due to withdrawal of a maternal gonadotrophin support. However, the corpus luteum is not the sole source of progesterone during gestation. After ovariectomy during the third trimester of pregnancy maternal peripheral plasma

39
progesterone concentrations decrease but do not reach basal levels and pregnancy is maintained in some of these animals for a variable period of time (MACDONALD et al., 1953; ESTERGREEN et al., 1967; WENDORFF et al., 1983; LINDELL, 1981). Maternal adrenalectomy caused all ovariectomized pregnant cows to abort or to calve prematurely (WENDORFF et al, 1983), suggesting that the maternal adrenal produces amounts of progesterone sufficient to maintain gestation after day 215. A third and probably more relevant source of progesterone is the placenta: placental cells produce progesterone in vitro (AINSWORTH and RYAN, 1967; SHEMESH et al, 1983, 1984a; REIMERS et al, 1985; SCHULER et al, 1994) and a positive uterine venous-arterial difference of the plasma progesterone concentration was maintained after ovariectomy on day 250, but not on day 270 of gestation (PIMENTEL et al, 1986). Therefore the placenta may only secrete progesterone during a limited period of gestation when it could easily Substitute luteal progesterone after ovariectomy because of its close proximity to the main target tissues, the endo-metrium and myometrium. Rising levels of Prostaglandin F, or the 13-14 dihydro-15-keto metabolite of PGF (PGFM), have been measured both in peripheral blood (EDQVIST et al, 1978, 1981; BOSU et al, 1984) and in utero-venous blood (FAIRCLOUGH et al, 1975a; HUNTER et al, 1977), at the time that the final and rapid phase of progesterone withdrawal before spontaneous calving, corticosteroid-induced calving (FAIRCLOUGH et al, 1981; LINDELL et al, 1977; JANSZEN et al, 1990) or prostaglandin-induced (KOETS et al, 1998) parturition takes place. Prostaglandin F2a
most likely represents the main endogenous luteolytic Stimulus. A systemic rather than a local and ipsilateral, effect of this luteolytic factor is indicated by embryo transfer studies, in which an embryo of a cow developed in the uterine hörn of a Zebu dam contralateral to the side where the single corpus luteum of pregnancy and a Zebu foetus were situated. Pregnancy in the Zebu normally lasts for about 296 days but in this case the cow foetus appeared to initiate the birth process in the Zebu at about 284 days (SUMMERS et al, 1983). While Prostaglandin release from the uterus takes place in the form of just a small number of peaks around luteolysis during the oestrous cycle (BASU and KINDAHL, 1987), no PGFM-pulses were found (with high frequent sampling) during prepartum luteolysis, but plasma levels increases more gradually (KINDAHL, personal communication). When parturition is induced by treating cows with PGF2a (BOSC, 1981) it is because of its luteolytic but not myometrial stimulating action (ZEROBIN et al, 1973; JANSZEN et al, 1993). Uterine contractility in the cow has not been recorded concurrently with measurements of hormonal changes around the spontaneous onset of parturition. RUSSE (1964) postulated that the onset of parturition in the cow is characterized by a fall in uterine tone, after which contractions typical for labour can develop. More recently JANSZEN et al. (1990,1993) described a period of about 8 hours with complete myometrial electrical quiescence which coincided with induced prepartum luteolysis. The cause and functional significance of this temporary uterine inhibition are not known, but it has been postulated that this period allows for a proper biochemical and morphological preparation ofthe cervix and myometrium before the typical labor contractions start to develop (JANSZEN, 1993). Recent data from our lab (KOETS et al, 1998) showed that exactly during this period of myometrial inhibition, maternal aorta plasma levels
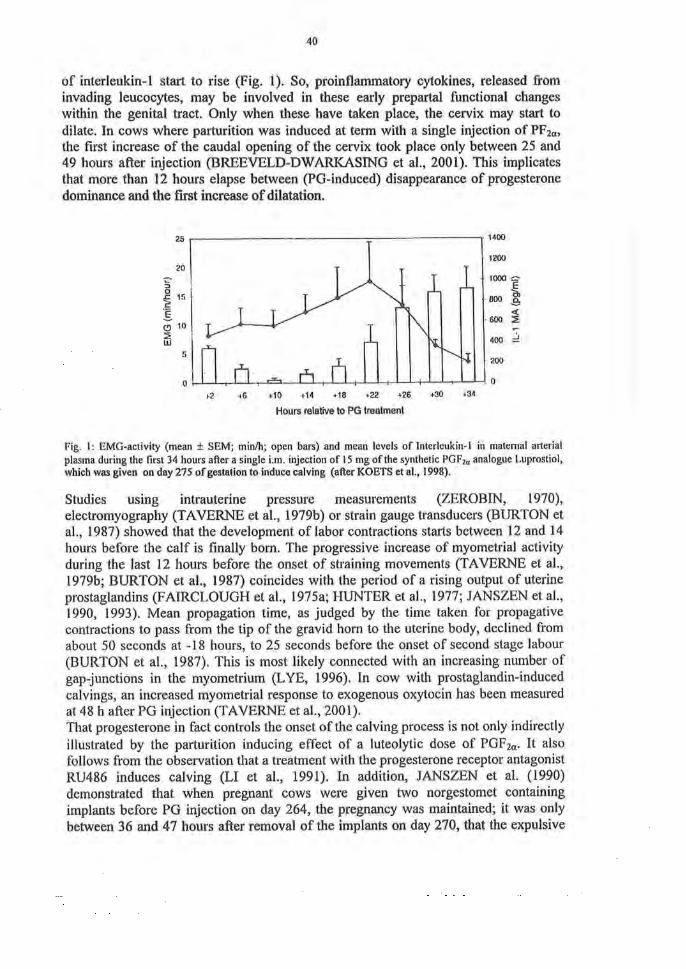
40
of interleukin-1 start to rise (Fig. 1). So, proinflammatory cytokines, released from invading leueocytes, may be involved in these early prepartal functional changes within the genital tract. Only when these have taken place, the cervix may start to dilate. In cows where parturition was induced at term with a single injection of PF2a, the first increase of the caudal opening of the cervix took place only between 25 and 49 hours after injection (BREEVELD-DWARKASING et al, 2001). This implicates that more than 12 hours elapse between (PG-induced) disappearance of progesterone dominance and the first increase of dilatation.
46 +10 +14 +18 +22 +26
Hours relative to PG treatment
Fig. 1: EMG-activity (mean ± SEM; min/h; open bars) and mean levels of Interleukin-1 in matemal arterial plasma during the first 34 hours after a single i.m. injection of 15 mg ofthe synthetic PGF2a analogue Luprostiol, which was given on day 275 of gestation to induce calving (after KOETS et al., 1998).
Studies using intrauterine pressure measurements (ZEROBIN, 1970), electromyography (TAVERNE et al, 1979b) or strain gauge transducers (BURTON et al, 1987) showed that the development of labor contractions Starts between 12 and 14 hours before the calf is finally born. The progressive increase of myometrial activity during the last 12 hours before the onset of straining movements (TAVERNE et al, 1979b; BURTON et al, 1987) eoineides with the period of a rising output of uterine Prostaglandins (FAIRCLOUGH et al, 1975a; HUNTER et al, 1977; JANSZEN et al, 1990, 1993). Mean propagation time, as judged by the time taken for propagative contractions to pass from the tip of the gravid hörn to the uterine body, declined from about 50 seconds at -18 hours, to 25 seconds before the onset of second stage labour (BURTON et al, 1987). This is most likely connected with an increasing number of gap-junetions in the myometrium (LYE, 1996). In cow with prostaglandin-induced calvings, an increased myometrial response to exogenous Oxytocin has been measured at 48 h after PG injection (TAVERNE et al, 2001). That progesterone in fact controls the onset of the calving process is not only indirectly illustrated by the parturition inducing effect of a luteolytic dose of PGF2c(. It also follows from the Observation that a treatment with the progesterone reeeptor antagonist RU486 induces calving (LI et al, 1991). In addition, JANSZEN et al. (1990) demonstrated that when pregnant cows were given two norgestomet containing implants before PG injection on day 264, the pregnancy was maintained; it was only between 36 and 47 hours after removal of the implants on day 270, that the expulsive
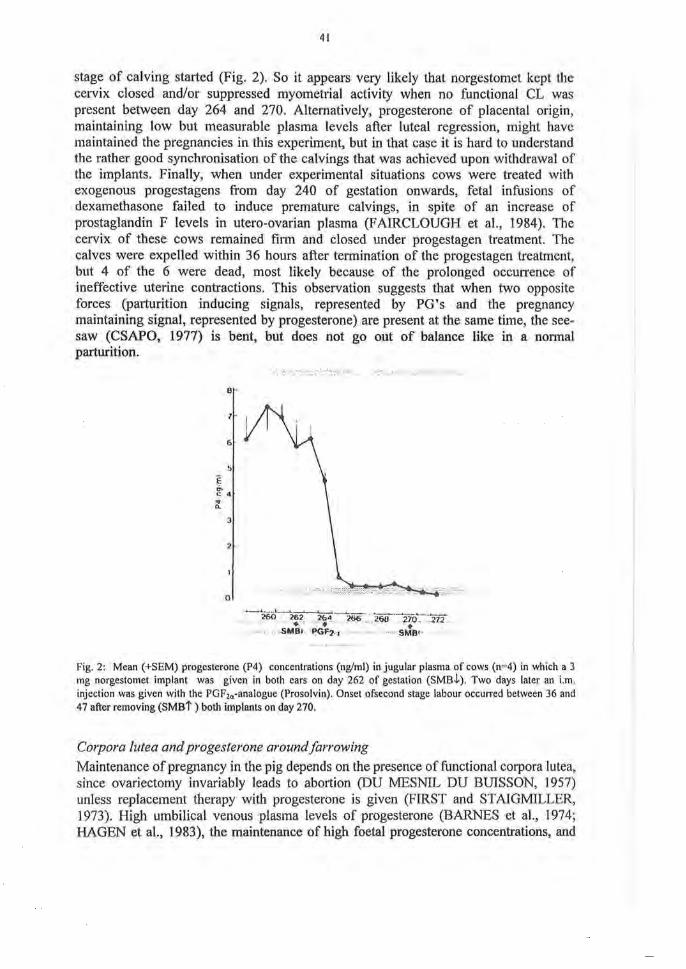
41
stage of calving started (Fig. 2). So it appears very likely that norgestomet kept the cervix closed and/or suppressed myometrial activity when no functional CL was present between day 264 and 270. Altematively, progesterone of placental origin, maintaining low but measurable plasma levels after luteal regression, might have maintained the pregnancies in this experiment, but in that case it is hard to understand the rather good Synchronisation of the calvings that was achieved upon withdrawal of the implants. Finally, when under experimental situations cows were treated with exogenous progestagens from day 240 of gestation onwards, fetal infusions of dexamethasone failed to induce premature calvings, in spite of an increase of Prostaglandin F levels in utero-ovarian plasma (FAIRCLOUGH et al, 1984). The cervix of these cows remained firm and closed under progestagen treatment. The calves were expelled within 36 hours after termination of the progestagen treatment, but 4 of the 6 were dead, most likely because of the prolonged oecurrence of ineffective uterine contractions. This Observation suggests that when two opposite forces (parturition inducing signals, represented by PG's and the pregnancy maintaining signal, represented by progesterone) are present at the same time, the see-saw (CSAPO, 1977) is bent, but does not go out of balance like in a normal parturition.
5 | •: 4
.26° 262 ...264... ,?fi<S.,L:26fl .. ..7X0.. ~2Ü.. SMBl PGFj,, SMB'
Fig. 2: Mean (+SEM) progesterone (P4) concentrations (ng/ml) in jugular plasma of cows (n=4) in which a 3 mg norgestomet implant was given in both ears on day 262 of gestation (SMBl). Two days later an i.m. injection was given with the PGF2<I-analogue (Prosolvin). Onset ofsecond stage labour occurred between 36 and 47 after removing (SMBt) both implants on day 270.
Corpora lutea and progesterone around farrowing Maintenance of pregnancy in the pig depends on the presence of functional corpora lutea, since ovariectomy invariably leads to abortion (DU MESNIL DU BUISSON, 1957) unless replacement therapy with progesterone is given (FIRST and STAIGMILLER, 1973). High umbilical venous plasma levels of progesterone (BARNES et al, 1974; HAGEN et al, 1983), the maintenance of high foetal progesterone concentrations, and

42
the negative arterio-venous differences in umbilical vessels of foetuses in ovariectomized sows (RANDALL and TSANG, 1986) suggest that the porcine placenta is capable of progesterone synthesis, although its functional significance is not yet clear. Hypophyseal luteotrophic support is essential for the continuation of pregnancy because maternal hypophysectomy results in abortion, unless progesterone is given afterwards (DU MESNIL DU BUISSON and DENAMUR, 1969). LH is the main luteotrophic hormone (BRINKLEY and YOUNG, 1974; COOK et al, 1967) but there is very little information on LH secretion and luteal progesterone production during the peripartal period. A decreased sensitivity to LH at 41 to 17 hours before parturition was indicated by the loss of a clear relationship between episodic increases of LH and progesterone in the miniature pig (PARVIZI et al, 1976). Regression of corpora lutea, as judged by declining maternal plasma progesterone concentrations, proceeds spontaneous parturition (ROBERTSON and KING, 1974; ASH and HEAP, 1975; PARZIVI et al, 1976). The decline usually Starts and gradually proceeds during the last few weeks and a more abrupt fall takes place between 24 and 10 hours before expulsion ofthe first piglet (TAVERNE et al, 1979a). Progesterone was found to suppress uterine contractility in the ovariectomized sow (ZEROBIN and SPÖRRI, 1972; TAVERNE, 1982; PORTER and WATTS, 1986). Although prepartal withdrawal of progesterone is not complete during farrowing and fetal expulsion occassionally occurs in the presence of elevated maternal plasma progesterone concentrations (TAVERNE, 1982), it is reasonable to assume that parturition in the pig normally occurs after a release from the progesterone block on the myometrium. The Stimulus for luteolysis partly originates in the uterus, because hysterectomy prolonges the lifespan ofthe corpora lutea (FELDER et al, 1986). It may reach the corpora lutea by way of the peripheral circulation, because transplantation of the ovaries to the body wall does not affect gestation length (MARTIN et al, 1978). However, an alternate route through the mesosalpinx has been proposed for the cycling sow (KOTWICA et al, 1983). There is indirect evidence that Prostaglandins are involved in luteolysis: parturition can easily be induced by a single injection of either PGF2a (DIEHL et al, 1977) or its synthetic analogues (ASH and HEAP, 1973; SILVER et al, 1983), and administration of indomethacin delayed luteolysis and delivery of piglets for several days (NARA and FIRST, 1981; TAVERNE et al, 1982). Direct measurements of Prostaglandins in maternal plasma around parturition have given equivocal results. While some authors, using frequent (KINDAHL et al, 1982) or infrequent (NARA and FIRST, 1981) sampling regimens, reported elevated Prostaglandin F metabolite concentrations coinciding with the fall in maternal plasma progesterone levels, other workers failed to show such a relationship (SILVER et al, 1979; GOONERATNE et al, 1982, 1983). In a study in which hourly bleeding took place during the spontaneous onset of parturition, the initial fall in plasma progesterone concentrations occurred prior to significant increases of PGFM in the uterine artery and PGF2a in uterine venous blood (RANDALL et al, 1986). What mechanisms operate at prepartum luteolysis in the pig is therefore not completely clear. Recent studies on luteolysis in cycling pigs indicate that intraluteal, cytokine-mediated paracrine processes, may be involved as well (WUTTKE et al, 1995). Profiles of plasma Prostaglandin concentrations on the day of parturition clearly suggest that Prostaglandins (mainly PGF2a) are responsible for the early Stimulation of uterine labor-like contractility, at the time that the inhibitory influences on the myometrium are
43
fading out. One of these inhibitory factors might be relaxin. Relaxin is stored in luteal cells during pregnancy (BELT et al, 1971) and is released at minor but physiologically significant quantities during gestation. A more substantial release of relaxin, as reflected by peak concentrations in maternal plasma, occurs during the last days before parturition (SHERWOOD et al, 1975, 1981; TAVERNE et al, 1982; KENDALL et al, 1982; ELDRIDGE et al, 1989). Relaxin release can be initiated by luteolytic (SHERWOOD et al, 1976) and non-luteolytic doses of Prostaglandin F2a (NARA et al, 1982), and can be blocked by treatment with indomethacin (SHERWOOD et al, 1979; TAVERNE et al, 1982) but not with meclofenamic acid (GOONERATNE et al, 1983). However, early peaks of relaxin concentrations in maternal plasma were, like the initial decrease of progesterone, not associated with altered PGFM concentrations in uterine arterial plasma (RANDALL et al, 1986). So it is not yet sure what initiates the prepartum relaxin release; in sows hysterectomized during pregnancy, relaxin also reaches peak values in the circulation around day 114 (ANDERSON et al, 1983).
Cervical softening, as judged by the ability to pass a series of rods of different diameters into the cervical canal, increases gradually from 3 days before parturition (KERTILES and ANDERSON, 1979). The appearance of labor-like uterine contractions clearly follows the decline of progesterone (TAVERNE et al, 1979a), but coincides with rapidly falling relaxin levels and increased output of uterine Prostaglandin F (RANDALL et al, 1986). This temporal relationship of hormonal and uterine events indicates that progesterone, relaxin and Prostaglandins are the key hormones during the onset of porcine parturition. The relaxin surge complets cervical softening and might Substitute for progesterone to inhibit the onset of regulär uterine contractions at an early stage of parturition (PORTER, 1979; PORTER and WATTS, 1986), when luteolysis and progesterone withdrawal have started.
21s
20
t 16
i 12 •
o 8 h
! 4
\ /
h \
TV
-4 -3 -2 -T 0
PG .
_ , , ,1
10 :
16 24 32 40 48 56 64 72 80 -4 .5' h o u r .s 4 days
rM..-:-i:P: ; : , P 6 . - ' -.-!,•
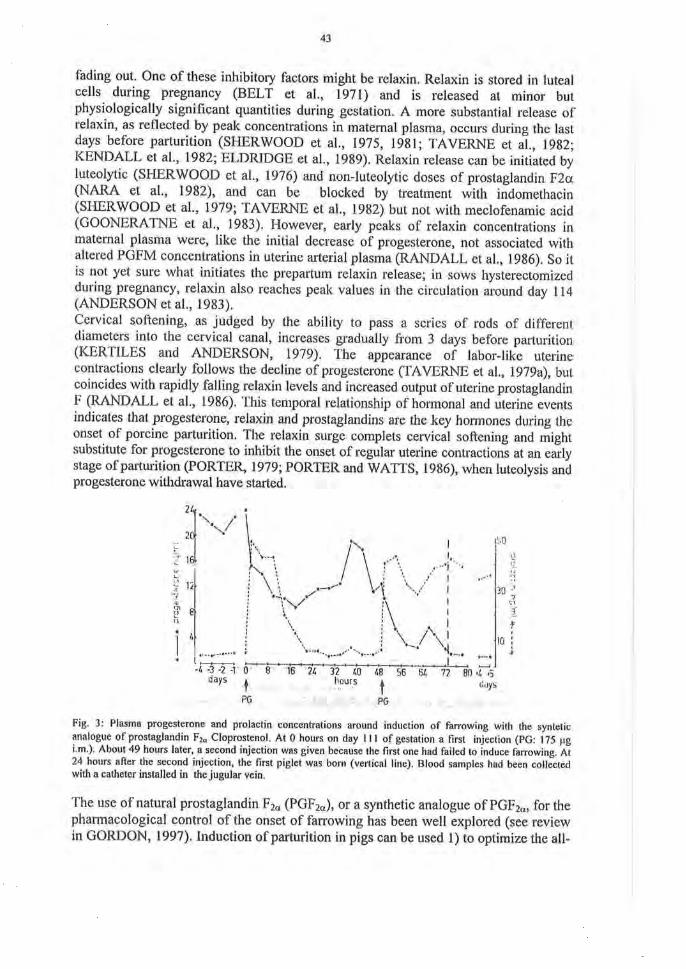
Fig. 3: Plasma progesterone and prolactin concentrations around induction of farrowing with the syntetic analogue of Prostaglandin F2o Cloprostenol. At 0 hours on day 111 of gestation a first injection (PG: 175 pg i.m.). About 49 hours later, a second injection was given because the first one had failed to induce farrowing. At 24 hours after the second injection, the first piglet was bom (vertical line). Blood samples had been collected with a catheter installed in the jugular vein.
The use of natural Prostaglandin F2a (PGF2a), or a synthetic analogue of PGF2a, for the pharmacological control of the onset of farrowing has been well explored (see review in GORDON, 1997). Induction of parturition in pigs can be used 1) to optimize the all-

44
in/all-out use of farrowing house facilities, 2) to facilitate cross fostering of piglets, 3) to obtain piglets from diseased pregnant as soon as possible, so that they can be culled, and 4) to guarantee optimal supervision during and shortly after parturition in order to reduce perinatal piglet losses as much as possible. Although it has been demonstrated that very intensive supervision ofthe farrowing process (following its pharmacological induction) may substantially reduce perinatal mortality under farm conditions
197 T U . daV°9 * " ^ " 4min. 7
1C0ÜJ.V A
197 day 110 D
-#• 1—m ——-«*# 1-• - I
-*•—' w—r~ 1#- h-
HMilL u j ü lyij. Ja iiu,i r ™ * r H W rrF ™ ~Wf,
-MHIk y^t ^ ^ui. L.L
Hiigi i i» r
• * - H l HM- HM • # N * 1 * * ' . . . .
Fig. 4: Myometrial electrical activity in a sow around induction of farrowing with the synthetic analogue of Prostaglandin F ^ Cloprostenol. Two surface electrodes had been implanted on the myometrium over two foetal Chambers ofthe same uterine hörn at day 97 of gestation. A: a spontaneous contracture on day 109 of gestation; B: i.m. injection (arrow) of 175 ug Cloprostenol on day 110 does not induce any emg-activity; C: at 6 hours after injection, when plasma P4 measured 6.7 ng/ml, the uterus is still quiescent; D: at 23 hours after injection, a parturient pattern of emg-activity was recorded and plasma P4 had dropped to 3.4 ng/ml; E: 30 hours after injection and 1 hour before the first piglet was bom; F: during expulsion ofthe 2nd and 3'd piglet (•)

45
(HOLYOAKE et al, 1995), financial investments involved in such supervision might be substantial, so that a farm-based cost-benefit analysis should be undertaken, before such a programme is implemented. Moreover, it has become quite clear that induction of farrowing with PGF2a does not reduce the length of the period over which sows farrowed relative to non-induced sows (HOLYOKE et al, 1995), because a relative small proportion of the sows will not farrow as a result of the injection. The latter seems to be related to failure or incompleteness of luteal regression (Fig. 3 and SILVER et al, 1983). To narrow the window during which sows start to farrow after PG treatment, several combinations of drugs have been used, including injections of Oxytocin at a fixed time after PG-treatment. From a clinical point of view this seems to be a wrong approach, as adverse effects of Oxytocin are to be expected when a sow with a still insufficiently dilated cervix, is already exposed to overstimulation of uterine contractility. Indeed it has been reported that requirements of manual interventions were highest in those treatment schedules (20 IU of Oxytocin at 24 hours after PG-injection) which resulted in the highest synchrony and the most rapid onset of farrowing (DIAL et al, 1987). It should be realized that even the primary injection of natural PGF2a to induce the farrowing process causes a temporary, though significant reduction in fetal oxygenation, most likely because of a diminished uteroplacental bloodflow during an induced prolonged myometrial contracture (RANDALL, 1990). Injection of the synthetic PGF2o analogue cloprostenol, on the other hand, does not have an immediate effect on myometrial activity (Fig. 4), although fetal plasma cortisol levels were found to be significantly higher at two hours after injection (SILVER et al, 1983).
Synchronisation of farrowing by treatment with exogenous progesterone or progestagens should not be recommended. Treatment with exogenous progesterone (NELLOR et al, 1975; TAVERNE et al, 1982) or induction ofa set of new corpora lutea with eCG/hCG at day 103 of gestation (MARTIN et al, 1977) results in prolonged gestation. However, a relative high percentage of stillborn piglets and a prolonged duration of the expulsive stage have been observed when delivery finally occurred after the P4 treatment was stopped or the new corpora lutea had regressed. This suggests that, like in the cow, some kind of subclinical, ineffective parturition process is initiated at about the normal time, although uterine Prostaglandin output and myometrial activity have not been measured under these circumstances in the pig.
References AINSWORTH, L.; RYAN, K.J.:
Steroid hormone transformations by endocrine organs from pregnant mammals: II. Formation and metabolism of progesterone by bovine and sheep placenta in vitro. Endocrinology, 81 (1967), 1349
ANDERSON, L.L.; ADAIR, V.; STROMER, M.H.; McDONALD, W.G.: Relaxin production and release after hysterectomy in the pig. Endocrinology, 113 (1983), 677-686
ASH,R.W.;Heap, R.B.: Oestrogen, progesterone and corticosteroid concentration in peripheral plasma of sows during pregnancy, parturition, lactation and after weaning. J. Endocr., 64 (1975), 141-154
BARNES, R.J.; COMLINE, R.S.; SILVER, M.: Foetal and maternal progesterone concentrations in the sow. J. Endocr., 62 (1974), 419-420
BASU, S.; KINDAHL, H.: Development of a continuous blood collection technique and a detaiied study of Prostaglandin F2a release during luteolysis and early pregnancy in heifers. J. Vet. Med. A., 34 (1987), 487-500
BELT, W.D.; ANDERSON, L.L.; CAVAZOS, L.F.; MELAMPY, R.M.: Cytoplasmic granules and relaxin levels in porcine corpora lutea. Endocrinology, 89 (1971), 1-10

46
BOSC, MJ.: Le velage. Mechanisme et contröle. In: A. Constatin and E. Meissonnier (Editors), L'uterus de la vache. Anatomie, Physiologie, Pathologie. Pub. Soc. Fr. Buiatrie, (1981), 149-168
BOSU, W.T.K.; LIPTRAP, R.M.; LESLIE, K.: Peripartal changes in plasma progesterone and 15-keto-13,14-dihydro-prostaglandin F2a concentrations in Holstein cows with or without retained foetal membranes. Anim. Reprod. Sei., 7 (1984), 497-510
BREEVELD-DWARKASING, V.N.A.; STRUIJK, P.C.; EIJSKOOT, F.; LOTGERING, F.K.; vanDISSEL-EMILIANI, F.M.F.; vanderWEIJDEN, G.C; TAVERNE, M.A.M.: Ultrasonic cervimetry to study the dilatation of the caudal cervix of the cow at parturition. Theriogenology, in press
BRINKLEY, H.J.; Young, E.P.: Effect of ovariectomy on the concentration of plasma LH, FSH and PRL in pig. J. Anim. Sei., 39 (1974), 229
BURTON, M.J.; DZIUK, H.E.; FAHNING, M.L; ZEMJANIS, R.: Myometrial activity during natural and dexamethasone-induced parturition in the cow. Am. J. Vet. Res., 48(1987), 37-44
COOK, B.; KALTENBACH, C.C; NORTON, H.W.; NALBANDOV, A.V.: Synthesis of progesterone in vitro by porcine corpora lutea. Endocr., 81 (1967), 573-584
CSAPO, A.I.: The see-saw theory of parturition. In: The Fetus and Birth, Ciba Foundation Symposium 47 (new series; 1977), 159-195
DIAL, G.D.; ALMOND, G.W.; HILLEY, H.D.; REPASKY, M.S.; HAGAN, J.: Oxytocin precipitation ofprostaglandin-induced farrowing in swine: determination of the optimal dose of Oxytocin and optimal interval between Prostaglandin F2a and Oxytocin, Am. J. Vet. Res., 48 (1987), 966-970
DIEHL, J.R.; BAKER, D.H.; DZIUK, P.J.: Effect of PGF2a on sow and litter Performance during and following parturition. J. Anim. Sei., 44 (1977), 89-94
DU MESNIL du BUISSON, F.; DAUZIER, L.: Influence de I' ovariectomie chez la truie pendant la gestation. CR. Biol., 151 (1957), 311-313
DU MESNIL du BUISSON, F.; DENAMUR, R.: Mechanisme du contröle de la function luteale chez la truie, la brebis et la vache. In: C. Gual (Editor), Progress in Endocrinology, Excerpta Medica Foundation, Amsterdam, The Netherlands, (1969), 927-934
EDQVIST, L.E.; KINDAHL, H.; STABENFELDT, G.: Release of Prostaglandin F2 during the bovine peripartal period. Am. J. Physiol., 16 (1978), 111-120
EDQVIST, L.E.; LINDELL, J.O.; KINDAHL, H.: Prostaglandin F2o release at premature and normal term deliveries in the cow. Acta Vet. Scand., 77 (1981), 267-277
ELDRIDGE-WHITE, R.; EASTER, R.A.; HEATON, D.M.; O'DAY, M.B.; PETERSEN.G.C; SHANKS, R.D.; TARBELL, M.K.; SHERWOOD, O.D.: Hormonal control ofthe cervix in pregnant gilts. I. Changes in the physical properties ofthe cervix correlate temporally with elevated serum levels of estrogen and relaxin. Endocrinology 125 (1989), 2996-2003
ESTERGREEN, V.L.,; FROST, O.L.; GOMES, M.A.; ERTS, R.E.; BULLARD, J.F.: Effect of ovariectomy on pregnancy maintenance and parturition in dairy cows. J. Dairy. Sei., 50 (1967), 1293-1295
FAIRCLOUGH, R.J.; HUNTER, J.T.; WELCH, R.A.S.: Peripheral plasma progesterone and utero-ovarian Prostaglandin F concentrations in the cow around parturition. Prostaglandins, 9 (1975), 901-914
FAIRCLOUGH, R.J.; HUNTER, J.T.; PETERSON, A.J.; WELCH, R.A.S.: Plasma concentration of progesterone, oestrogens and Prostaglandin F in maternal blood and corticosteroids and oestrogens in foetal blood of cows during dexamethasone induced deliveries. Acta Endocr., 96(1981), 401-412
FAIRCLOUGH, R.J.; KALTENBACH, C.C; PETERSON, A.J.; WELCH, R.A.S.; COX, R.I.; WONG, M.S.F.: Failure of exogenous progesteron to block dexamethasone-induced Prostaglandin F release from the uterus of late pregnant cows. Biol. Reprod., 30 (1984), 112-118
FELDER, K.J.; MOLINA, J.R.; BENOIT, A.M.; ANDERSON, L.L.: Precise timing for relaxin and decreased progesterone secretion after hysterectomy in the pig. Endocrinology 119 (1986), 1502-1505
FIRST, N.L.; STAIGMILLER, R.B.:

47
Effects of ovariectomy, dexamethasone and progesterone on the maintenance of pregnancy in swine. J. Anim. Sei., 37 (1973), 1191-1194
GOONERATNE, A.D.; HARTMANN, P.E.; BAKER, J.: Influence of meclofenamic acid on the initation of parturition and lactation in the sow. J. Reprod. Fert., 65(1982), 157-162
GOONERATNE, A.D.; BRYANT-GREENWOOD, G.; WALKER, F.M.; NOTTAGE, HM.; HARTMANN, P.E.: Pre-partum changes in the plasma concentrations of progesterone, relaxine, Prostaglandin F-2 and 13, 14-dihydro-15-keto Prostaglandin F-2 in meclofenamic aeid-treated sows. J. Reprod. Fert., 68 (1983), 33-40
GORDON, [.: Controlled Reproduction in Pigs. CAB Interantional, Oxon, UK (1997)
GROSS, T.S.; WILLIAMS, CS. ; GALINA, CS.; MANSPEAKER, J.; HAALAND, M.A.; RUSSEK, E.: Functional Status ofthe peri-partum bovine corpus luteum. Anim. Reprod. Sei., 8 (1985), 193-201
HAGEN, D.R.; HOUTZ, S.K.; GRIEL Jr., L.C; KAVANAUGH, J.F.: Concentrations of progesterone in arterial and venous plasma of fetal pigs and their dams in late gestation. Biol. Reprod., 29 (1983), 919-923
HOFFMANN, B.; SCHAMS, D.; GIMENEZ, T.; ENDER, J.L.; HERRMAN, Ch.; KARG, H.: Changes of progesterone, total oestrogens, corticosteroids, prolactin and LH in bovine peripheral plasma around parturition with special reference to the effect of exogenous corticoids and a prolactin inhibitor respectively. Acta Endocr., 73 (1973), 385-395
HOFFMANN, B.; SCHAMS, D.; BOPP, R.; ENDER, M.L.; GIMENEZ, T.; KARG, H.: 1974. Luteotrophic factors in the cow: evidence for LH rather than prolactin. J. Reprod. Fert., 40 (1974), 77-85
HOLM, L.W.; SHORT, R.V.: Progesterone in the peripheral blood of guernsey and friesian cows during prolonged gestation. J. Reprod. Fert., 4 (1962), 137-141
HOLYOAKE, P.K.; DIAL, G.D.; TRIGG, T.; KING, V.L.: Reducing pig mortality through supervision during the perinatal period. J. Anim. Sei., 73 (1995), 3543-3551
HUNTER, J.T.; FAIRCLOUGH, R.J.; PETERSON, A.J.; WELCH, R.A.S.: 1977. Foetal and maternal hormonal changes preceding normal bovine parturition. Acta Endocr., 84 (1977), 653-662
JANSZEN, B.P.M.; KNIJN, H.; van der WEIJDEN, G.C; BEVERS, M.M.; DIELEMAN, S.J.; TAVERNE, M.A.M.: Flumethason-induced calving is preceded by a period of myometrial inhibition during luteolysis. Biol. Reprod., 43 (1990a), 466-471
JANSZEN, B.P.M.; BEVERS, M.M.; DIELEMAN, S.J.; vanderWEIJDEN, G.C; TAVERNE, M.A.M: Synchronized calvings after withdrawal of norgestomet implants from cows treated near term with Prostaglandins. Vet.Rec. 127 (1999b), 405-407
JANSZEN, B.P.M.; BEVERS, M.M.; RAVENHORST, M.M.; van der WEIJDEN, G.C; DIELEMAN, S.J.; TAVERNE, M.A.M.: The relation between prostaglandin-induced luteolysis and temporary inhibition of myometrial activity in late pregnant cows with progestagen containing ear implants. J. Reprod. Fert., 97 (1993), 457-461
KENDALL, J.Z.; RICHARDS, J.E.; SHIK, L.N.; FARRIS, T.S.: Plasma relaxin concentrations in the pig during the periparturient period: association with prolactin, estrogen and progesterone concentration. Theriogenology, 17 (1982), 677-687
KERTILES, L.P.; ANDERSON, L.L.: Effect of relaxin on cervical dilatation, parturition and lactation in the pig. Biol. Reprod., 21 (1979), 57-68.
KINDAHL, H.; ALONSO, R.;, CORT, N.; EINARSSON, S.: Release of Prostaglandin F2 during parturition in the sow. Zbl. Vet. Med., 29 (1982), 504 -510
KOETS, A.P.; de SCHWARTZ, N.; TOOTEN, P.; KANKOFER, M.; BROEKHUIJSEN-DAV1ES, J.M.; RUTTEN, V.P.M.G.; van LEENGOED, L.A.M.G.; TAVERNE, M.A.M.; GRUYS, E.: Release of proinflammatory cytokines related to luteolysis and the periparturient acute phase response in prostaglandin-induced parturition in cows. Theriogenology 49 (1998), 797-812
KOTWICA, J.; KRZYMOWSKI, T.; STEFANCZYK, S.; KOZIOROWSKI, M.„ CZAMOCKI, J; RUSZCYK, T.: A new mute of Prostaglandin F2 transfer from the uterus into the ovary in swine. Anim. Reprod. Sei., 5 (1983), 303-309
LI, Y.; PEREZGROVAS, R.; GAZAL, O.S.; SCHABE, C ; ANDERSON, L.L.:

48
Antiprogesterone, RU 486, facilitates parturition in cattle. Endocrinology 129 (1991) 765-770 LINDELL, G.O.; KINDAHL, H; EDQVIST, L.E.:
Prostaglandin release at dexamethasone induced parturition in cows. Acta Vet. Scand 18 (1977) 257-265
LINDELL, G.O.: Studies on induced abortion and parturition and uterine involution in the bovine. Thesis Swedish University of Agric. Sei., (1981) Uppsala, Sweden
LYE, S.J.: Initiation of parturition. Anim. Reprod.Sci., 42 (1996), 495-503
McCRACKEN, J.A.; CUSTER, E.E.; LAMSA, J.C: Luteolysis: a neuroendoerine-mediated event. Physiol. Rev. 79 (1999), 263-323
McDONALD.L.E.; McNUTT, S.H.; NICHOLS, R.E.: On the essentiality ofthe bovine corpus luteum of pregnancy. Am. J. Vet. Res 14 (1953) 539-541
MARTIN, PA.; NORTON, H.W.; DZIUK, P.J.: The effect of corpora lutea induced during pregnancy on the length of gestation in the pig. Biol Reprod., 17(1977), 712-717
MARTIN, P.A.; BEVIER, G.W.; DZIUK, P.J.: The effect of disconnecting the uterus and ovary on the length of gestation in the pig. Biol. Reprod 18 (1978), 428-433
NARA, B.S.; FIRST, N.L.: Effect of indomethacin and Prostaglandin F2 on parturition in swine. J. Anim. Sei., 52 (1981) 1360-1370
NARA B.S.; BALL, G.D.; RUTHERFORD, J.E.; SHERWOOD, OD.; FIRST, N.L.: Release of relaxin by a nonluteolytic dose of Prostaglandin F2 in pregnant sheep. Biol. Reprod 27-(1982), 1190-1195
NELLOR, J.E.; DANIELS, R.W.; HOEFER, J.A.; WILDT, D.E.; DUKELOW, W.R.: Influence of induced delayed parturition on foetal survival in pigs. Theriogenology, 4 (1975), 23-31
PARVIZI, N.; ELSAESSER, F.; SMIDT.D.; ELLENDORFF, F.: Plasma luteinizing hormone and progesterone in the adult female pig during the oestrous cycle, late
pregnancy and lactation, and after ovariectomy and pentobarbitone treatment. J. Endocr., 69 (1976) 193-203
PIMENTEL, M.; PIMENTEL, CA.; WESTON, P.G.; HIXON, J.E.; WAGNER, W.C: Progesterone secretion by the bovine fetoplacental unit and responsiveness of corpora lutea to steroidogenic Stimuli at two stages of gestation. Am. J. Res., 47 (1986), 1967-1971
POPE, G.S.; GUPTA, S.K.; MURRO, J.B.: Progesterone levels in the systemic plasma of pregnant, cycling and ovariectomized cows. J. Reprod. Fert., 20 (1969), 369-381
PORTER, D.G.: The myometrium and the relaxin enigma. Anim. Reprod. Sei., 2 (1979), 77-96
PORTER, D.G.; Watts, A.D.: Relaxin and progesterone are myometrial inhibitors in the ovariectomized non-pregnant mini-pig. J. Reprod. Fert., 76 (1986), 205-213
RANDALL, G.C.B.: Influence of fetal numbers and hypophysectomy on the length of gestation in the pig. Can. J. Anim. Sei., 67 (1987), 1125-1127
RANDALL, G.C.B.: Induction of parturition in pigs: short term effetcs of Prostaglandin F2a on chronically catheterised fetuses at term. The Veterinary Record, 126 (1990) 61-63
RANDALL, G.C.B.; TSANG, B.K.: Influence of the fetal pituitary-adrenal axis on fetal and maternal progesterone and unconjugated oestrogen concentrations in the pig. J. Reprod. Fert., 78 (1986), 65-72
RANDALL, G.C.B.; TAVERNE, M.A.M.; CHALLIS, J.R.G.; KENDALL, J.Z.; TSANG, B.K.: Interrelationships between endocrine changes in peripheral and uterine-venous blood an uterine activity at parturition in the pig. Anim. Reprod. Sei., 11 (1986), 283-294
REIMERS, T.J.; ULLMANN, M.B.; HANSEL, W.: Progesterone and Prostanoid production by bovine binucleate trophoblastic cells. Biol. Reprod., 33 (1985), 1227-1236
ROBERTSON, H.A.; King, G.J.: Plasma concentrations of progesterone, oestrone, oestradiol-17beta and of oestrone sulphate in the pig at implantation, during pregnancy and at parturition. J. Reprod. Fert., 40 (1974), 133-141
RUSSE, M.: Der Geburtsablauf beim Rind. Arch. Exp. Veterinärmed., 19 (1964), 763-870

49
SCHALLENBERGER, E.; RAMPP, J.; WALTERS, D.L.: Gonadotrophins and ovarian Steroids in cattle. II. Pulsatile changes of concentrations in the jugular vein throughout pregnancy. Acta Endocrinologica, 108 (1985), 322-330
SCHULER, G ; HÄRTUNG, F.; HOFFMANN, B.: Investigations on the use of C-21 Steroids as precursors for placental oestrogen synthesis in the cow. Exp. Clin. Endocrinol., 102 (1994), 169-174
SHEMESH, M.; HANSEL, W.; STRAUSS III, J.F.: Bovine placentomes contain factors which decrease progesterone secretion. Biol. Reprod., 29 (1983), 856-862
SHEMESH, M.; HANSEL, W.; STRAUSS III, J.F.: Calcium-dependent, cyclic nucleotide-independent steroidogenesis in the bovine placenta. Proc. Natl. Acad. Sei. USA, 81 (1984), 6403-6407
SHERWOOD, O.D.; CHANG, C.C; BEVIER, G.W.; DZIUK, P.J.: Radioimmunoassay of plasma relaxin levels throughout pregnancy and at parturition in the pig.
Endocrinology, 97 (1975), 834-837 SHERWOOD, O.D.; CHANG, C.C; BEVIER, G.W.; DIEHL, J.R.; DZIUK, P.J.:
Relaxin concentrations in pig plasma following the administration of Prostaglandin F2 during täte pregnancy. Endocrinology, 98 (1976), 875-879
SHERWOOD, O.D.; NARA, B.S.; CRNEKOVIC, V.E.; FIRST, N.L.: Relaxin concentration in pig plasma after the administration of indomethacin and Prostaglandin F2 during late pregnancy. Endocrinology, 104 (1979), 1716-1721
SHERWOOD, O.D.; NARA, B.S.; WELK, F.A.; FIRST, N.L.; RUTHERFORD, J.E.: Relaxin levels in the maternal plasma ofpigs before, during and after parturition and before, during and after suckling. Biol. Reprod., 25 (1981), 65-71
SILVER, M.; BARNES, R.J.; COMLINE, R.S.; FOWDEN, A.L.; CLOVER, L.; MITCHELL, M.D.: Prostaglandins in the foetal pig and pre-partum endocrine changes in mother and foetus. Anim. Reprod. Sei., 2 (1979), 305-322
SILVER, M.; BARNES, R.J.; FOWDEN, A.L.: Fetal and maternal endocrine changes during the induction of parturition with the PGF analoque, cloprostenol, in chronically catheterized sows and fetuses. J. Devel. Physiol, 5 (1983), 307-321
STABENFELDT, GH.; OSBURN, B.J.; EWING, L.L.: Peripheral plasma progesterone levels in the cow during pregnancy and parturition. Am. J. Physiol., 218 (1970), 571-575
SUMMERS, A.M.; SHELTON, J.N.; EDWARDS, J.: The production of mixed species Bos taurus-Bos indicus twin calves. Anim. Reprod. Sei., 6 (1983), 79-89
TAVERNE, M.A.M.; NAAKTGEBOREN, C ; ELSAESSER, F.; FORSLING, M.L.; van der WEIJDEN, G.C; ELLENDORFF, F.; SMIDT, D.:
Myometrial electrical activity and plasma concentrations of progesterone, estrogen and Oxytocin during late pregnancy and parturition in the miniature pig. Biol. Reprod., 21 (1979a), 1125-1134
TAVERNE, M.A.M.; van der WEIJDEN.G.C; FONTIJNE, P.: Preliminary observations on myometrial electrical activity before, during and after parturition in the cow. In: B. Hoffmann, f.L. Mason and J. Schmidt (Editors): Calving Problems and Early Viability of the Calf, Martinus Nijhoff, The Hague, The Netherlands (1979b), 297-311
TAVERNE, M.A.M.: Myometrial activity during pregnancy and parturition in the pig. In: D.J.A. Cole and G.R. Foxcroft (Editors), Control of Pig Production, Butterworth Scientific, (1982), 419-436
TAVERNE, M.A.M.; BEVERS, M.M.; BRADSHAW, J.M.C; DIELEMAN, S.J.; WILLEMSE, A.H.; PORTER, D.G.: Plasma concentrations of prolactin, progesterone, relaxin and oestradiol-I7beta in sows treated with
progesterone, bromoeriptine or indomethacin during late pregnancy. J. Reprod. Fert., 65 (1982), 85-96 TAVERNE, M.A.M.; HESSELINK, J.W.; BEVERS, M.M.; van OORD, H.A.; KORNALIJNSLIJPER, J.E.:
Aetiology and endocrinology of pseudopregnancy in the goat. Reprod. Dornest. Anim., 30 (1995), 228-230
TAVERNE, M.A.M.; de SCHWARTZ, N.C.M.; KANKOFER, M.; BEVERS, M.M.; van OORD.H.A.; SCHAMS, D.; GUTJAHR, St.; van der WEIJDEN, G.C: Uterine responses to exogenous Oxytocin before and after prepartum luteolysis in the cow. Reprod. Dornest. Anim., (2001) in press
WENDORFF, G.L.; LAWYER, M.S.; First, N.L.: Role ofthe adrenals in the maintenance of pregnancy in cows. J. Reprod. Fert., 68 (1983), 281-287
WUTTKE, W.; JARRY, H.; PITZEL, L ; KNOKE, 1.; SPIESZ, H.; THEILING, K.: Some aspects about luteolysis in the porcine corpus luteum. Reprod.Dom. Anim., 30 (1995), 208-210

50
ZEROBIN, K.: Die Uterusbewegungen bei Kühen während der Geburt und der Nachgeburtsphase. Schweizer Arch. Tierheilk., 112 (1970), 544-560
ZEROBIN, K; SPÖRRI, H.: Mobility ofthe bovine and porcine uterus and fallopian tube. Adv. Vet. Sei. Comp. Med., 16 (1972), 303-354
ZEROBIN, K.; JÖCHLE, W.; STEINGRUBER, CH.: Termination of pregnancy with Prostaglandins (PGE2 and PGF2) in cattle. Prostaglandins, 4 (1973), 891-901
Aulhor's address
Prof. MARCEL A.M. TAVERNE, PhD. Department of Farm Animal Health University of Utrecht, 3508 TD Utrecht, The Netherlands
E-Mail: [email protected]