‘de rol van dendritische cellen in intestinale...
TRANSCRIPT

FACULTEIT GENEESKUNDE EN
GEZONDHEIDSWETENSCHAPPEN
Academiejaar 2012 - 2013
‘De rol van dendritische cellen in intestinale homeostase’
Hanne VAN BEVEREN
Promotor: Prof. Dr. Claude Cuvelier
Scriptie voorgedragen in de 2de Master in het kader van de opleiding
MASTER OF MEDICINE IN DE GENEESKUNDE


Voorwoord
Deze masterproef werd voorgesteld in het kader van de opleiding ‘Master in de Geneeskunde’
aan de UGent.
Vooreerst zou ik mijn promotor, professor C. Cuvelier, willen bedanken voor de
ondersteuning en feedback die hij me bood tijdens het schrijven van deze masterproef. Ik zou
ook graag professor K. Vermaelen willen bedanken om op te treden als commissaris bij de
verdediging van mijn masterproef.
Graag bedank ik mijn ouders bedanken voor de steun die ze me gaven tijdens mijn studies en
het schrijven van mijn masterproef.
Tenslotte wil ik u bedanken, beste lezer, voor de interesse die u toont voor deze masterproef.
Ik hoop dat deze masterproef nuttig voor u kan zijn.

Abstract……………………………………………………….……………………………….1
I. Inleiding…………………………………………………………………………..…………2
II. Methodologie……………………………………………………………………….………4
III. Resultaten………………………………………………………………………………….5
1. Dendritische cellen………………………………………………………………..…5
1.1 Dendritische cellen………………………………………………………....5
1.1.1 Soorten dendritische cellen………………………….……………5
1.1.2 Lokalisatie en migratie………………………………..…...……..6
1.1.3 Maturatie………………………………………………..…..……7
1.1.4 Opname van bacteriën……………………………………..……..8
1.2 Microbioom…………………………………………………………...……9
1.3 Homeostase………………………………………………………….……11
2. Ontstaan van de tolerante dendritische cellen………………………………...…....12
2.1 Conditionering door intestinale epitheliale cellen……………………..….12
2.1.1 Intestinale epitheliale cellen……………………………...……..12
2.1.2 TSLP………………………………………………….…………15
2.1.3 TGF-β………………………………………………………..….16
2.1.4 RA……………………………………………………………….17
2.2 Neurologische beïnvloeding……………………………………………....20
2.3 Apoptotische cellen……………………………………………………….21
2.3.1 Apoptotische dendritische cellen…………………………..……21
2.3.2 Andere apoptotische cellen…………………………………..….23
2.4 IDO…………………………………………………………………….….25
2.5 IL-10………………………………………………………………….…...26
2.6. HLA-G en ILT4…………………………………………………….…….26
2.7 IL-27……………………………………………………………........……27
2.8 PD-1/PD-1L………………………………………………………...….….27
3. Functies van tolerante dendritische cellen……………………………...………….28
3.1 Invloed op T-cellen…………………………………………………….….28
3.1.1 Inductie van Tregs………………………………………….…...28
3.1.1.1 Foxp3+Tregs………………………………………..…28
3.1.1 1.1 Ontstaan……………………………………..28
3.1.1.1.1.1 Natuurlijke Foxp3+Tregs………….28
3.1.1.1.1.2 Geïnduceerde Foxp3+Tregs…….…28

3.1.1.1.2 Kenmerken van Foxp3+Tregs……………….30
3.1.1.2 TR1s…………………………………………...………31
3.1.1.2.1 Ontstaan van Tr1……………………...……..31
3.1.1.2.2 Kenmerken van Tr1s……………………..….33
3.1.1.3 Functies van Tregs…………………………………….34
3.1.2 Suppressie van T-cellen…………………………………………36
3.2 IgA………………………………………………………………….….….37
3.2.1 Ontstaan……………………………………………….….….….37
3.2.2 De rol van IgA in homeostase………………………….……….39
3.3 Gut homing……………………………………………………………..…40
3.4 Co-stimulatoire molecules: PD-1 en CTLA-4……………….……..……..41
3.5 Overzicht: 3 gebieden homeostase…………………………………..……42
3.5.1 Auto-reactieve T-cellen…………………………………………42
3.5.2 Tolerantie………………………………………………….….…45
3.5.3 Indijken van inflammatie…………………………………….….45
4. Invloed van de omgeving………………………………………………………..…46
4.1 IECs…………………………………………………………………...…..46
4.2 Tregs…………………………………………………………….......…….46
4.3 Microbioom……………………………………………………………….46
IV. Discussie………………………………………………………………………..………...48
1. Samenvatting…………………………………………………………………….…48
2. Onderzoek……………………………………………………………………….…51
3. Therapie…………………………………………………………………………….52
V. Referentielijst………………………………………………………………………...……53
1. Wetenschappelijke artikels……………………………………………………...….53
2. Proefschriften…………………………………………………………………..…..58
3. Websites………………………………………………………………………...….58
VI. Bijlagen
1. Lijst met afkortingen
2. Overzicht figuren en tabellen
2.1 Figuren
2.2 Tabellen

1
Abstract
De darm bevindt zich op een kritieke plaats in het lichaam. Het vormt de
eerste plaats van verdediging tegen binnendringende pathogenen. Daarom vinden we in de
darm een geavanceerd immuunsysteem terug -het GALT genoemd- dat onmiddellijk in actie
treedt om deze pathogenen uit te schakelen. De darm komt ook met andere antigenen in
contact nl. antigenen van commensale bacteriën en antigenen afkomstig van voedsel. Dit zijn
onschadelijke antigenen en daar mag het immuunsysteem niet tegen reageren. Gebeurt dit
toch, dan krijgen we te maken met chronische inflammatoire darmziekten en voedsel-
allergieën. Het immuunsysteem moet dus onderdrukt worden, zodat het niet reageert tegen
deze onschadelijke bacteriën. Het moet echter wel steeds paraat staan om onmiddellijk in
actie te treden wanneer het toch in contact komt met schadelijke bacteriën. Dit heet
homeostase: het bewaren van de balans tussen te veel, dan wel te weinig reageren. Daarom
moet voortdurend de beslissing gemaakt worden of het immuunsysteem in actie moet treden
of niet. Dendritische cellen (DCs) spelen in dit proces een belangrijke rol. DCs vormen de
link tussen het innate en het adaptieve immuunsysteem. Ze bevinden zich in de eerste
verdedigingslinie. Ze nemen antigenen op en presenteren die vervolgens aan de cellen van het
adaptieve immuunsysteem zoals T-cellen. De DCs bevinden zich op een belangrijk kruispunt.
Zij bepalen immers welke weg het immuunsysteem opgaat: tolerantie of inflammatie. Dit is
mogelijk doordat DCs het verschil kunnen maken tussen schadelijke antigenen en
onschadelijke antigenen. DCs presenteren de antigenen op een inflammatoire of een tolerante
manier aan het adaptieve immuunsysteem. Op die manier weet het adaptieve systeem welk
antwoord het moet geven. DCs bepalen dus het antwoord dat het adaptieve immuunsysteem
geeft op een bepaald antigen.
DCs bezitten twee belangrijke eigenschappen die ze inschakelen om
homeostase ter hoogte van de darm te realiseren. Ze zijn in staat regulatoire T-cellen (Tregs)
en plasmacellen te induceren die IgA produceren. Hiermee kunnen ze homeostase realiseren
op drie gebieden: uitschakelen van auto-reactieve T-cellen, bewerkstelligen van tolerantie ten
opzichte van onschadelijke antigenen en tijdig indijken van inflammatoire reacties.
In dit proces spelen een aantal cellen een grote rol: DCs, intestinale epitheliale cellen (IECs),
het microbioom en Tregs. Deze cellen staan niet los van elkaar en beïnvloeden elkaar
voortdurend. DCs hebben hierin een centrale rol.

2
I. Inleiding
Vandaag de dag bedraagt de wereldwijde prevalentie van IBD (inflammatory bowel
diseases, hieronder verstaan we de ziekte van Crohn en colitis ulcerosa) 116 à 133 per
100.000, dit betekent dat er wereldwijd 8 tot 9 miljoen personen leven met IBD. Er zijn ook
veel mensen die leiden aan een voedselallergie of voedselintolerantie. Deze pathologieën zijn
te verklaren door een overactiviteit van het immuunsysteem ter hoogte van de darm ten
opzichte van onschadelijke antigenen. Anders gezegd, bepaalde aspecten die zorgen voor
tolerantie van het immuunsysteem ten opzichte van deze onschadelijke antigenen lopen fout
bij deze pathologieën.
Onze darm komt voortdurend in contact met allerlei antigenen. Velen daarvan zijn
onschadelijk en behoeven geen inflammatoire immuunrespons. Dit zijn de commensale
bacteriën in onze darm en antigenen afkomstig van voedsel. Wanneer hier toch tegen
gereageerd wordt, ontstaat een chronische inflammatie. Anderzijds mag ons immuunsysteem
ook niet volledig in toom gehouden worden. Het moet immers nog steeds in staat zijn
onmiddellijk te reageren wanneer het in contact komt met antigenen die wel schadelijk zijn.
Men zou zich nu de volgende vraag kunnen stellen: wat zorgt ervoor dat ons immuunsysteem
in staat is te reageren tegen schadelijke antigenen, maar toch niet tegen onschadelijke
antigenen maw. hoe wordt homeostase ter hoogte van de darm bewerkstelligd? DCs spelen
hierin een belangrijke rol, zij liggen immers in de eerste verdedigingslinie tegen schadelijke
pathogenen die de darm binnendringen.
Tegenwoordig is het onderzoek naar de processen die een rol spelen in homeostase ter hoogte
van de darm in volle opgang. Vooral het onderzoek naar het microbioom en zijn rol in
homeostase kent een enorme toename. In 2011 werd de Nobelprijs voor geneeskunde
toegekend aan professor Ralph Steinman voor zijn ontdekking van de DC en de rol ervan in
het aanpassingsproces van immuniteit. Dit alles bewijst dat we meer en meer het belang
inzien van de rol van DCs in homeostase ter hoogte van de darm.
Het is belangrijk dat dit onderzoek gebeurt, want hoe meer we weten over de fysiologische
processen, hoe beter we kunnen verstaan wat er precies gebeurt wanneer deze processen
mislopen. Als we de fysiologie en fysiopathogenese begrijpen, kunnen we hiermee tevens aan
de slag om nieuwe en betere therapieën te ontwikkelen.

3
In deze masterproef wordt een overzicht gegeven van wat we tot nu toe
weten over de rol van DCs in homeostase ter hoogte van de darm. Deze kennis is echter vrij
fragmentarisch, daarom werd geprobeerd om deze zo goed mogelijk te bundelen en een
algemene lijn aan te brengen in deze informatie. Met deze masterproef proberen we
kapstokken aan te reiken waaraan al deze informatie kan worden opgehangen.
Vele puzzelstukjes zijn reeds gekend. Hier proberen we de puzzel samen te leggen, wetende
dat er nog steeds puzzelstukken ontbreken.

4
II. Methodologie
Opzoeken van literatuur
De informatie voor deze literatuurstudie is afkomstig wetenschappelijke artikels die werden
opgezocht in medische databanken zoals Pubmed, NCBI en Web of Science. Er werd gewerkt
met volgende zoektermen: homeostasis, intestinal dendritic cells. Via de citaties en referenties
van gevonden artikels werden andere artikels opgezocht. Reviews werden gebruikt om een
globaal beeld te krijgen van wat er reeds bestond aan kennis. De verschillende elementen uit
de reviews werden vervolgens verder uitgediept. Er werd ook gezocht op auteur. Hiervoor
werd ook gebruikt gemaakt van de website van Biomedexperts.
Alle artikels zijn verschenen in vooraanstaande wetenschappelijk tijdschriften met een hoge
impact factor.
Er werd gekozen om zoveel mogelijk artikels te gebruiken die gepubliceerd zijn na 2005.
Aangezien het onderzoek rond dit onderwerp in volle opgang is, verschijnen er op
regelmatige basis nog artikels rond dit onderwerp. Er is gepoogd de informatie zo up-to-date
mogelijk te houden.
Er werd een databank via EndNote aangelegd. Er werd ook gebruik gemaakt van Reference
Manager.
De wetenschappelijke artikels werden met een kritische blik beoordeeld. Er werd gezocht naar
tegenstrijdigheden, verbanden en hiaten tussen de artikels.
Schrijven van de masterproef
Er is geopteerd voor een beschrijvende onderzoeksmethode. Het is de bedoeling een overzicht
te bieden van de huidige kennis. Er werd geprobeerd deze kennis in één groot geheel te
brengen en het niet te beschrijven als een aaneenschakeling van losse studieresultaten.
Er werd geprobeerd structuur aan te brengen in het geheel dmv. onderverdeling in
hoofdstukken. Men moet natuurlijk in het achterhoofd houden dat dit eigenlijk vrij artificieel
is, aangezien de processen door elkaar lopen en bepaalde zaken overlappen, maar deze
onderverdeling zal de lezer helpen om het geheel beter te begrijpen en een beter overzicht te
bewaren van het geheel.
Tijdens het schrijven werden de teksten voortdurend geëvalueerd.

5
III. Resultaten
1. Inleiding
1.1 Dendritische cellen
DCs maken deel uit van het innate immuunsysteem. Het zijn antigen-presenterende cellen.
DCs scannen voortdurend de omgeving af opzoek naar pathogenen. Wanneer ze pathogenen
tegenkomen, gaan ze de pathogenen phagocyteren, verwerken en vervolgens de antigenen via
MHC-moleculen op het celoppervlak presenteren aan andere immuuncellen zoals de
T-cellen. Deze worden zo geactiveerd en starten een immuunrespons op. Op die manier
vormen de DCs een link tussen het innate en het adaptieve immuunsysteem.
DCs bevinden zich dus in de eerste verdedigingslinie tegen pathogenen.
1.1.1 Soorten dendritische cellen
DCs ontstaan in het beenmerg. Daar verblijven ze als precursoren tot ze het beenmerg
verlaten via het bloed.
We kunnen twee grote groepen DCs onderscheiden op basis van hun expressie van specifieke
merkers en een verschillend antwoord op stimuli voor maturatie en differentiatie.
We onderscheiden myeloïde en plasmacytoïde DCs. In de darm bevinden zich zowel
myeloïde en plasmacytoïde DCs. Beide soorten spelen een rol in de homeostase in de darm.
Fig. 1. (1) Soorten DCs. Vanuit precursoren in het beenmerg ontstaan twee cellijnen: myeloïde en plasmacytoïde
DCs. Uit de myeloïde cellijn ontstaan interstitiële DCs en Langerhanscellen die zich in de perifere weefsels
bevinden en myeloïde DCs die zich in de bloedbaan bevinden. Uit de plasmacytoïde cellijn ontstaan
plasmacytoïde DCs die zich in de bloedbaan bevinden. Alle DCs kunnen na activering migreren naar de
secundaire lymphe-organen.

6
1.1.2 Lokalisatie en migratie
Het immuunsysteem in de darm bestaat uit het GALT (gut-associated lymphoid tissue) en de
mesenterische lymphnodi (MLNs). Het GALT bestaat uit de lamina propria (LP), de peyerse
platen (PP) en de geïsoleerde lymfefollikels (ILF). Het GALT behoort tot de secundaire
lymphe-organen. DCs bevinden zich in de darm ter hoogte van het GALT en de MLNs. (2)
Boven het GALT bevindt zich het intestinaal epitheel. Het intestinaal epitheel bestaat uit vier
celtypes: enterocyten, entero-endocriene cellen, paneth-cellen en mucusproducerende cellen
(goblet-cellen). Het epitheel in de dunne darm bestaat uit vili en crypten, het epitheel in de
dikke darm bestaat niet uit vili en is plat.
De LP is de bindweefsellaag onder het epitheel. In de LP bevinden zich allerlei immuuncellen
waaronder DCs, T-cellen, B-cellen en IgA producerende plasmacellen.
De PP bevinden zich onder het epitheel. Ze bestaan uit aggregaten van follikels. In de follikels
bevinden zich voornamelijk B-cellen en DCs. Tussen de follikels bevinden zich
interfolliculaire ruimtes die T-cellen en DCs bevatten.
De PP hebben een koepelvormige structuur en puilen zo als het ware uit in de epitheliale laag.
Dit gebied noemen we de subepitheliale dome (SED). Het epitheel boven de SED is speciaal
epitheel nl. het follikel geassocieerd epitheel (FAE). Het bestaat niet uit crypten en vili en het
bevat de M-cellen (Microfold-cellen). Deze cellen spelen een rol in het transport van micro-
organismen naar de DCs in het GALT.
Ter hoogte van de LP vindt vooral opname van antigenen door de DCs plaats, ter hoogte van
de andere delen van het GALT vinden voornamelijk immuunresponsen plaats. (3)
Fig. 2. (96) GALT en intestinaal epitheel.

7
Na hun ontstaan in het beenmerg verlaten de DCs het beenmerg via de bloedbaan. De DCs
moeten nu naar verschillende locaties in het lichaam migreren. Migratie naar de darm gebeurt
onder invloed van de IECs. De IECs secreteren chemokineliganden (CCLs) die de DCs
aantrekken naar de PP waar ze vervolgens antigenen kunnen opnemen. Deze CCLs zijn CCL5
en CCL9. (4) Ze binden met de chemokinereceptor (CCR) CCR1 op de DCs. Wanneer DCs
antigenen hebben opgenomen, presenteren ze vervolgens de antigenen aan andere
immuuncellen zoals T-cellen die zich ter hoogte van de MLNs bevinden. De DCs moeten dus
vanuit de LP naar de MLNs kunnen migreren. Dit gebeurt met behulp van hun
chemokinereceptor CCR7 die bindt met CCL19 en CCL21. (5) In steady state, dwz. wanneer
er geen inflammatie aan de gang is, migreren de DCs voortdurend vanuit de LP naar de
MLNs.
1.1.3 Maturatie
Mature en immature DCs verschillen phenotypisch en functioneel van elkaar.
DCs matureren onder invloed van verschillende signalen afkomstig van pathogenen en
inflammatie/infectie zoals inflammatoire cytokines (TNF-α, IL-1, IFN, urinezuur,…), binding
van CD40 met CD40-ligand, pathogenen via PRRs,… (6)
DCs brengen tijdens de maturatie co-stimulatoire molecules tot expressie op hun celoppervlak
nl. CD86, CD80 en CD83. Deze oppervlaktemolecules maken deel uit van de B7 familie.
Mature DCs seceteren ook andere cytokines dan immature DCs. Mature DCs produceren
pro-inflammatoire cytokines zoals IL-12, TNF-α en IFN-γ.
Immature en mature DCs hebben verschillende functies. Immature DCs zijn gespecialiseerd
in de opname van antigenen, terwijl mature DCs gespecialiseerd zijn in de presentatie van
antigenen.
De maturatiestatus van een DC speelt ook een rol in tolerantie: Immature DCs zijn tolerante
DCs. Om een T-cel te activeren zijn twee signalen tussen de DCs en de T-cel vereist. Het
eerste signaal is de binding tussen MHC van de DC en de TCR van de T-cel. Het tweede
signaal is het co-stimulatoir signaal: oppervlaktemolecules van de B7 familie binden met
CD28 op de T-cellen. Wanneer het tweede signaal niet aanwezig is, worden de T-cellen
anergisch of gaan ze in apoptose. Immature DCs brengen geen co-stimulatoire signalen tot
expressie en vervullen op die manier een tolerante functie. Immature DCs secreteren bepaalde
cytokines die T-cellen laten differentiëren tot Tregs en bekomen tolerantie via de effecten van
Tregs. Sommige tolerantiemechanismen maken gebruik van de tolerante status van immature
DCs: ze verwezenlijken tolerantie door de DCs in een immature status te houden.

8
Tolerante DCs vertonen een specifiek phenotype. Ze zijn resistent tegenover signalen die
maturatie induceren bij DCs, maw. tolerante DCs matureren niet.
Ze vertonen gedaalde antigenpresenterende capaciteiten, ze vertonen een verminderde
productie van molecules die nodig zijn om T-cellen te activeren en een verminderde secretie
van inflammatoire cytokines zoals IL-12, TNF-α, IL-6 en ze brengen minder co-stimulatoire
molecules zoals CD80, CD86 tot expressie. (7)
1.1.4 Opname van bacteriën
DCs fungeren als sentinel cellen: ze komen als eerste in contact met bacteriën in de
darm. Wanneer ze een bacterie hebben gephagocyteerd, zijn ze in staat te bepalen of het gaat
om een pathogene dan wel een commensale bacterie.
Zoals eerder vermeld, bezitten immature DCs een hoge capaciteit tot phagocytose. Tijdens
steady state nemen ze dan ook voortdurend bacteriën op uit hun omgeving. Er zijn
verschillende manieren waarop een DC in contact komt met bacteriën om die dan vervolgens
op te nemen. (8)
Opname van bacteriën gebeurt voornamelijk ter hoogte van de PP. (9)
1. Transport via M-cellen
Dit is de traditionele route waarlangs bacteriën worden opgenomen.
M-cellen bevinden zich in het FAE. M-cellen transporteren de antigenen van het lumen naar
de SED.
2. Via apoptotische epitheliale cellen
DCs kunnen apoptotische IECs phagocyteren. Op die manier nemen de DCs bacteriën op via
indirecte wijze. Een alternatief mechanisme is de opname van antigen-exosomen afkomstig
van IECs. DCs die zich in de subepitheliale follikels bevinden zijn in staat om antigenen
rechtstreeks van apoptotische IECs op te nemen.
3. Rechtstreeks contact
Tijdens inflammatie ontstaan er discontinuïteiten in de epitheliale cellaag. De bacteriën
kunnen dan doorheen deze openingen het GALT bereiken. Op die manier komen de DCs
rechtstreeks in contact met de bacteriën.
4. Foetale Fc receptor
Een recent ontdekt mechanisme van opname van bacteriën is opname dmv. de foetale
Fc receptor. De foetale Fc receptor vinden we terug in zowel foetale als volwassen weefsels.
De foetale Fc receptoren nemen bacteriën op vanuit het lumen en transporteren de bacteriën
doorheen de epitheliale cellaag naar het GALT.

9
5. Intestinale cellen
Ook intestinale cellen zijn betrokken in de opname van bacteriën door de DCs. IECs laten
exosomen en antigenen los die vervolgens worden opgenomen door de DCs.
6. Dendrieten
Een recent ontdekt en tevens zeer intrigerend mechanisme is de opname van bacteriën door
DCs via dendritische uitstulpingen. (10)
Tussen de IECs bevinden zich tight junctions. De DCs zijn in staat deze tight junctions te
openen. Doorheen de openingen sturen de DCs vervolgens hun dendrieten die in staat zijn om
antigenen op te nemen. Tijdens dit proces blijft de integriteit van de epitheliale barrière steeds
bewaard, doordat de DCs eiwitten produceren gelijkaardig aan de eiwitten waaruit de tight
junctions zijn opgebouwd (nl. occludines en claudines). Na dit proces blijft het epitheel dus
onbeschadigd achter.
Verschillende factoren hebben een invloed op de opname van antigenen door DCs.
(11) Ten eerste secreteren de IECs chemokines die DCs aantrekken naar de PP alwaar ze
antigenen kunnen opnemen. Deze chemokines zijn CCL5 en CCL9. (4)
Ten tweede zorgt de herkenning van micro-organismen door TLRs ervoor dat de
antigenopname en antigenpresentatie door de DCs bevorderd wordt.
De werking van de M-cellen wordt verstrekt door IgA.
Hoe herkennen de DCs nu bacteriën? DCs herkennen bacteriën met behulp van
bepaalde receptoren nl. PRR (patern recognition receptor). Er bestaan verschikkende soorten
PRRs. Bij de DC vinden we drie categorieën PRRs terug: Toll-like receptoren (TLR),
nucleotide oligomerization receptoren (NLR) en C-type lectin receptors. Deze PRRs
herkennen specifieke onderdelen van de micro-organismen nl. microbe-associated molecular
patterns (MAMP). (8)
1.2 Microbioom
Het microbioom is het geheel van bacteriën in het maag-darmstelsel.
Het gaat om commensale bacteriën dwz. bacteriën die in een gastheer leven zonder die schade
te berokkenen. We kunnen de darm beschouwen als een ecosysteem waarin de mens en het
microbioom samenleven. Dit is een mooi voorbeeld van mutualisme: een samenlevingsvorm
van verschillende soorten waar beiden een voordeel van ondervinden.
Het microbioom bevat zo’n 1000-tal verschillende soorten bacteriën, in totaal zijn er 3 triljoen
bacteriën. (Ter vergelijking: dit komt overeen met 10 bacteriën voor 1 menselijke cel)
Het microbioom ontstaat in de eerste dagen na onze geboorte. De samenstelling wordt
beïnvloed door onze voeding, bepaalde pathologieën en medicatie (12).

10
Het microbioom heeft verschillende functies in de darm. (13)
Het speelt het een rol in de vertering. Bacteriën breken namelijk stoffen af die de mens niet
zelf kan verteren. Bacteriën produceren een aantal stoffen waaronder vitamine K. Het
microbioom heeft ook een anti-pathogeen effect. Het heeft een ruimte-innemend effect,
waardoor pathogene bacteriën geen kans krijgen die plaats te bezetten. Het microbioom
concurreert met pathogene bacteriën voor metabolieten, waardoor de pathogene bacteriën niet
genoeg metabolieten hebben en daardoor niet kunnen overleven. Het microbioom produceert
anti-bacteriële peptiden. Het speelt ook een actieve rol in immuunprocessen. Het heeft
namelijk een invloed op IECs en DCs. De mens is dus afhankelijk van het microbioom.
Anderzijds stelt het ons voortdurend voor een uitdaging om er niet tegen te reageren. Daarom
zijn er in het immuunsysteem mechanismen ingebouwd die ervoor zorgen dat we tolerantie
vertonen ten opzichte van het microbioom. Een belangrijke rol hierin is weggelegd voor de
DCs.
De laatste jaren neemt de interesse voor het microbioom in grote mate toe. Er wordt
momenteel veel onderzoek gevoerd naar het microbioom. Omwille van deze onderzoeken zijn
er reeds een aantal zeer interessante ontdekkingen gedaan.
Ten eerste is er de ontdekking van enterotypes. We kunnen het microbioom onderverdelen in
drie enterotypes. Deze enterotypes bevatten dezelfde soorten bacteriën, maar verschillen in de
soort bacteriën die dominant zijn. Men weet nog niet zeker of ons enterotype stabiel is dan
wel veranderlijk en wat dan ons type beïnvloed. (14) De volgende stap in dit onderzoek is te
bepalen of het soort enterotype een risico inhoudt voor bepaalde pathologieën.
Ten tweede is men te weten gekomen dat het microbioom een rol speelt in de fysiopathologie
van verschillende pathologieën nl. obesitas, diabetes (15) en inflammatoire darmziekten (16).
Ook qua therapie is men tot een aantal ontdekkingen gekomen. Zo heeft men ontdekt dat
patiënten met een therapieresistente darminfectie met E.coli zijn gebaat met een transplantatie
van hun microbioom. Dit gebeurt door de toediening van feces aan de patiënt afkomstig van
gezonde donoren. (17)
Tot slot kan men besluiten dat al deze onderzoeken ons één belangrijk feit vertellen nl. dat het
microbioom een grote rol speelt in de gezondheid van ons lichaam. Het is belangrijk dat men
dit beseft en meer en meer rekening houdt met deze bacteriën. Men zou het microbioom
kunnen beschouwen als een tweede genoom in ons lichaam, naast het genoom van onze
lichaamscellen. Men zou verder kunnen gaan in dit denkproces en zich de vraag stellen of we
onze levensstijl kunnen aanpassen aan ons enterotype om zo pathologieën te voorkomen. Men
zou het kunnen vergelijken het met predictieve geneeskunde op basis van je genoom. (95)

11
1.3 Homeostase
Homeostase is het vermogen om het interne milieu constant te houden. Wanneer we
homeostase toepassen op immuniteit betekent dit dat het immuunsysteem ervoor moet zorgen
dat het interne milieu niet beschadigd wordt door een onaangepaste immuunrespons. Het
immuunsysteem moet een balans vinden tussen te weinig of te veel reageren.
Homeostase uit zich in drie gebieden. Homeostase moet auto-immuniteit voorkomen, ervoor
zorgen dat een immuunrespons tijdig wordt ingedijkt, zodat deze niet escaleert en leidt tot
schade en ten slotte moet homeostase ervoor zorgen dat het lichaam tolerantie vertoont
tegenover commensale bacteriën en voedingsantigenen, maar toch steeds paraat staat om snel
en adequaat te reageren wanneer het in contact komt met pathogene antigenen. Vooral dit
laatste is typerend en van groot belang in de darm. De darm komt immers voortdurend in
contact met commensale bacteriën van het microbioom en met voedingsantigenen. DCs
spelen een voorname rol in deze homeostase. Hierbij is het van groot belang dat DCs in de
darm een onderscheid kunnen maken tussen allo-antigenen en auto-antigenen, commensale en
pathogene bacteriën en voedingsantigenen. Om homeostase binnen deze drie gebieden te
vrijwaren, bezitten de DCs twee grote eigenschappen: ze kunnen Tregs induceren en de
productie van IgA bevorderen.

12
2. Ontstaan van de tolerante dendritische cellen
2.1 Conditionering door intestinale epitheliale cellen
2.1.1 Intestinale epitheliale cellen
Vroeger werd het epitheel louter beschouwd als een fysische barrière, nu beseft men
meer dan ooit dat het epitheel een actieve, cruciale rol speelt in homeostase. De (tolerante)
functie van DCs is niet zozeer een intrinsieke eigenschap verbonden aan een bepaalde type
van DCs. Het is afhankelijk van een phenotype dat verworven wordt onder invloed van
bepaalde factoren uit de omgeving. De DCs liggen in nauw contact met de IECs, waardoor de
IECs makkelijk een invloed kunnen uitoefenen op de DCs. De IECs vormen inderdaad een
bron van bepaalde factoren die het phenotype van de DCs kunnen bepalen. Deze factoren
leiden niet tot maturatie in de DCs, maar conditioneert hen tot een bepaald phenotype. Het
phenotype van de DCs uit zich in de molecules die ze tot expressie brengen op hun
celoppervlak en in de molecules die ze gaan produceren en secreteren. IECs conditioneren dus
de DCs, zodat deze differentiëren naar een tolerant phenotype. IECs produceren voortdurend
allerlei mediatoren waaronder TLSP (thymic stromal lymphopoietin), TGF-β (transforming
growth factor bèta) en RA (retinoic acid). (18). Deze productie neemt nog eens toe wanneer
IECs worden gestimuleerd door commensale bacteriën. (19-21)
De soort factor waarmee de DC geconditioneerd wordt, bepaalt welke functie de DC zal
uitoefenen. Dit komt doordat de factor het phenotype van de DC bepaalt en het phenotype
bepaalt dan weer de functie. Het samenspel tussen IECs, het microbioom en DCs is dus van
cruciaal belang voor het behoud van homeostase in de darm.
Ongedifferentieerde DCs bereiken de darm via het bloed. Via de binding tussen de
E-cadherine ligand op de IECs en de overeenkomstige receptoren op de DCs worden de DCs aan het
intestinaal epitheel gebonden, waarna de IECs hun invloed kunnen uitoefenen op de DCs.
IECs zijn de sensoren van de omgeving. Ze komen immers als eerste in contact met de
bacteriën. Het is dan ook belangrijk dat zij het verschil kunnen maken tussen pathogeen en
commensaal. Wanneer het gaat om een pathogeen, geven ze het signaal aan de DCs om een
inflammatoire respons te geven. Wanneer het gaat om een commensale bacterie,
conditioneren ze de DCs om een tolerant phenotype aan te nemen. Via dit mechanisme
kunnen de DCs in een tolerante staat gehouden worden en wanneer het nodig is, onmiddellijk
naar een inflammatoire staat worden gebracht.

13
IECs herkennen bacteriën met behulp van bepaalde receptoren nl. PRRs. Bij de IECs vinden
we alleen TLRs en NLRs terug. Vooral de TLRs zijn van belang in het behoud van de
homeostase. Aan het oppervlak is er een dus voortdurende interactie tussen de commensale
bacteriën en de IECs via de PRRs. (22) Er zijn elf soorten TLRs te onderscheiden. Elke TLR
herkent verschillende soorten micro-organismen, bevindt zich op verschillende cellen en
bevindt zich op verschillende plaatsten bij de cel.
Tabel 1. (22) Karakteristieken van de verschillende TLRs.

14
De verschillende TLRs functioneren via verschillende intracellulaire pathways.
Fig. 3. (23) Overzicht van de verschillende intracellulaire pathways van de TLRs.
De vraag is nu hoe de IECs het onderscheid kunnen maken tussen schadelijke en
onschadelijke bacteriën. We kunnen drie strategieën onderscheiden. (12, 13) De expressie,
distributie en cellulaire lokalisatie van de TLRs speelt een rol in het onderscheid tussen
pathogeen en commensaal (24) NLRs zijn voornamelijk intracellulair gelokaliseerd en TLRs
zijn voornamelijk gelokaliseerd ter hoogte van de celmembraan. Meer specifiek zouden
TLR2, TLR4 en TLR5 zich voornamelijk intracellulair en op de basolaterale celmembraan
bevinden.

15
Dit betekent dat enkel bacteriën die door de cellaag geraken in staat zijn TLR2-4-5 te
activeren en dat enkel bacteriën die intracellulair geraken NLRs en TLR2-4-5 kunnen
activeren.
Een kleine hoeveelheid TLR2 en TLR4 op de apicale membraan is verantwoordelijk voor het
ontwikkelen van tolerantie voor endotoxine. Dit wil zeggen dat de TLRs niet meer
geactiveerd worden bij daaropvolgende blootstelling aan endotoxines bv. LPS
(lipopolysaccharide).
Over het derde mechanisme bestaat nog geen zekerheid. Het zou kunnen dat er om een
inflammatoire respons te veroorzaken twee signalen gelijktijdig vereist zijn nl activering van
de TLR op de apicale membraan en activering van de intracellulaire TLR.
Het is bewezen dat een zeker niveau van signalisatie vereist is om homeostase te
behouden. Toch mogen de IECs niet voortdurend reageren op het microbioom. Verschillende
mechanismen werden ingebouwd om dit te verkomen.
Enerzijds speelt de polarisatie van de IEC een cruciale rol. Recent is aangetoond dat de
activering van TLR9 via de apicale dan wel de basolaterale membraan leidt tot verschillende
uitkomsten. Activatie van TLR9 via de apicale membraan leidt tot een homeostatische
respons, terwijl de activering van TLR9 via de basolaterale membraan leidt tot een
pro-inflammatoire respons via NF-κB (nuclear factor kappa-light-chain-enhancer of activated
B-cells). (25)
Anderzijds bestaan er inhibitoren van TLR bv. IRAK-M, TOLLIP, SIGIR, A20 en PPAR-γ
(peroxisome proliferator-activated receptor-gamma). Zij zorgen ervoor dat chronische
imflammatie of vernietigende signalen van TLR niet optreden. (12)
2.1.2 TSLP
Een eerste factor die IECs gebruiken om DCs te conditioneren is TLSP. (18) TSLP
wordt gesecreteerd door de IECs oa. in de thymus en darm. (26) De receptor van TSLP is een
heterodimere receptor die bestaat uit een complex van TSLP-receptor en IL-7-receptor.
Binding van TSLP op zijn respectievelijke receptor leidt tot de phosphorylatie en zo ook de
activering van de transcriptiefactor STAT5. Dit leidt vervolgens tot de transcriptie van een
aantal genen die een rol spelen in de differentiatie van de DCs. (26)
TSLP heeft een invloed op de DCs, waardoor deze specifieke functies kunnen
uitoefenen: induceren van Tregs en IgA producerende plasmacellen en het elimineren van
auto-reactieve T-cellen. (13, 26)
TSLP speelt een rol in zowel centrale als perifere tolerantie. TSLP conditioneert DCs, zodat
deze een tolerant phenotype verkrijgen en T-cellen laten differentiëren naar Tregs.

16
TSLP dat geproduceerd wordt door de epitheliale cellen in de thymus speelt samen met DCs
een rol in centrale tolerantie. (26) TSLP dat geproduceerd wordt door de IECs speelt een rol
in perifere tolerantie.
DCs geconditioneerd met TSLP vertonen een specifiek phenotype: gedaalde secretie van
IL-12 en TNF-α, gestegen secretie van IL-6 en IL-10. DCs die geconditioneerd zijn door
TSLP verliezen dus hun vermogen om IL-12 te produceren en zo ook om Th1 (T-helpercel1)
te activeren. In plaats daarvan activeren ze Th2 (T-helpercel2) door hun gestegen productie
van IL-10. Zelfs wanneer deze geconditioneerde DCs gestimuleerd worden door bacteriën
zijn ze niet in staat Th1 te activeren, dit komt doordat ze geen IL-12 meer kunnen produceren.
Geconditioneerde DCs kunnen geen IL-12 meer produceren, omdat hun productie van IL-12
geblokkeerd is door TSLP. Dit is concentratie-afhankelijk: de inhibitie van de productie van
IL-12 door TSLP gebeurt enkel wanneer de concentratie van TSLP zich binnen bepaalde
grenzen bevindt. DCs kunnen Th1 activeren wanneer ze nog niet in contact gekomen zijn met
IECs of wanneer de concentratie van TSLP zich buiten de grenzen bevindt waarin het IL-12
blokkeert. Bacteriën maken hier handig gebruik van. Sommige bacteriën veroorzaken een
toename van TSLP wanneer ze gekoppeld zijn aan de basolaterale membraan van IECs. Op
die manier komt de concentratie van TSLP buiten de grenzen en kunnen DCs alsnog
differentiatie naar Th1 bewerkstelligen.
TSLP speelt een rol in het ontstaan van IgA producerende plasmacellen doordat het de
vrijstelling van BAFF (B-cell activating factor) en APRIL (a proliferation-inducing ligand)
door de IECs bevordert. (27)
2.1.3 TGF-β
IECs secreteren TGF-β. TGF-β conditioneert DCs tot het CD103+-phenotype.
(20, 27) CD103+DCs ontstaan in de darm. CD103+DCs vertonen een tolerant phenotype.
CD103 is een dus marker voor tolerantie bij DCs. De tolerante status van CD103+DCs
kenmerkt zich door een specifiek patroon van cytokine secretie: gedaalde productie van IL-12
en IL-6 en gestegen productie van IL-10, TGF-β en RA. TGF-β zorgt dus via de inductie van
CD103 voor de tolerante status bij de DCs.
DCs die geconditioneerd zijn met TGF-β afkomstig van IECs produceren vervolgens
zelf TGF-β. TGF-β wordt geproduceerd als een inactief complex en moet geactiveerd worden.
Dit gebeurt door ανβ8 (een integrine). (28) TGF-β is latent aanwezig in zowel CD103-DCs als
CD103+DCs, maar CD103+DCs bezitten grotere hoeveelheden actief TGF-β.

17
Dit is niet te wijten aan een hogere productie van latent TGF-β, maar aan het feit dat
CD103+DCs een verhoogd vermogen hebben om hun latent TGF-β te activeren. (8, 28, 29)
Dit verhoogd vermogen om TGF-β te activeren is te wijten aan het feit dat CD103+DCs een
verhoogde hoeveelheid ανβ8 bezitten. (28)
CD103+DCs bezitten ook grote hoeveelheden tgfb2 (transforming growth factor, bèta 2),
plat (tissue plasminogen activator) en ltbp3 (latent TGF-β binding protein 3). (5, 30) Dit zijn
enzymes die betrokken zijn in omzetting van TGF-β van de latente, inactieve vorm naar de
actieve vorm. CD103-DCs produceren slechts lage hoeveelheden tgfb2, plat, ltbp3 en ανβ8,
waardoor ze niet goed in staat zijn latent TGF-β om te zetten in actief. Bovenstaande feiten
verklaren waarom CD103-DCs geen Foxp3+Tregs (Forkhead box p3 Tregs) kunnen
induceren.
Deze productie van TGF-β heeft twee functies. Ten eerste zou het zelf geproduceerd TGF-β
op een autocriene manier werken en zo kunnen de DCs hun eigen activering onderdrukken.
(13, 31) Ten tweede is deze productie vereist voor het uitvoeren van hun functies.
CD103+DCs hebben vier grote functies. Ze zijn in staat Tregs te induceren op een
TGF-β en RA dependente wijze. Ze zorgen er ook voor dat de T-cellen migreren vanuit de
MLNs naar de darm. Hun tolerante status zorgt er ook voor dat ze de inflammatoire responsen
van Th1 en Th17 (T-helpercel17) onderdrukken. Ze spelen een rol in de inductie van IgA
producerende plasmacellen.
2.1.4 RA
Verscheidene factoren uit het milieu bevorderen de productie van RA in de DCs.
Dit doen ze oa. door de transcriptie van aldh1a2 te induceren, het gen dat codeert voor
RADLH (retinal dehydrogenase, een sleutelenzyme dat betrokken is in de metabolisatie van
vitamine A tot RA). Hierdoor zijn de DC nu zelf in staat de metabolisatie van vitamine A tot
RA uit te voeren. (32)
Enkele van deze signalen worden hier opgesomd. RA afkomstig van de epitheliale cellen.
ERK (extracellular signal related kinase). Micribiota of endogene TLR agonisten binden op
TLR1/TLR2. Dit leidt tot intracellulaire signalisatie via MyD88 (myeloid differentiation
primary response gene 88) en JNK-pathway, waardoor RALDH2 geïnduceerd wordt. (33)
Opname van Vitamine A uit de voeding in de dunne darm zorgt voor een constante
hoeveelheid vitamine A in de dunne darm. Epitheliale cellen zetten het vitamine A om tot RA
en bieden dit vervolgens aan aan de DCs. De lever stelt voortdurend hoge hoeveelheden
retinol vrij in de gal.
18
Deze gal induceert via de RA receptor op de DCs de capaciteit om retinol te metaboliseren en
zo RA te kunnen produceren in de DCs. (34) Epitheliale cellen zetten het vitamine A om tot
RA en bieden dit vervolgens aan aan de DCs. IL-4 en GM-CSF (granulocyte-macrophage
colony stimulating factor) induceren op synergistische wijze de productie van RA in DCs.
(35) β –catenine en PPAR-γ (peroxisome proliferator-activated receptor)
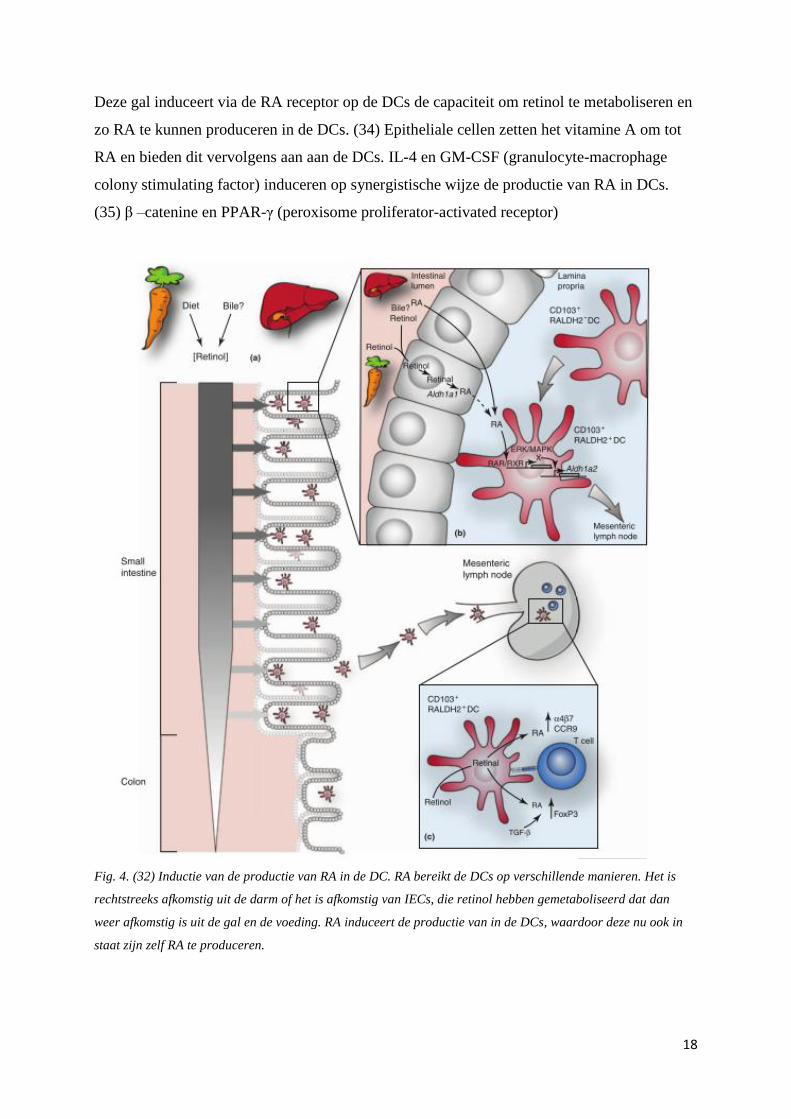
Fig. 4. (32) Inductie van de productie van RA in de DC. RA bereikt de DCs op verschillende manieren. Het is
rechtstreeks afkomstig uit de darm of het is afkomstig van IECs, die retinol hebben gemetaboliseerd dat dan
weer afkomstig is uit de gal en de voeding. RA induceert de productie van in de DCs, waardoor deze nu ook in
staat zijn zelf RA te produceren.

19
De metabolisatie van vitamine A tot RA gebeurt in twee stappen. Retinol wordt
omgezet tot retinal door alcohol dehydrogenase. Retinal wordt vervolgens geoxideerd tot RA
door RALDH. Er bestaan twee iso-vormen van RALDH: RALDH1 en RALDH2. RALDH2 is
de voornaamste iso-vorm die we terugvinden in DCs. RALDH wordt gecodeerd door het
aldh1a1 en aldh1a2. (32)
Fig. 5. (36) Metabolisatie van vitamine A tot RA. Retinol wordt omgezet tot retinal door alcohol dehydrogenase.
Retinal wordt vervolgens geoxideerd tot RA door RALDH.
Net zoals TGF-β conditioneert RA de DCs tot het CD103+-phenotype. (20, 27) TGF-β
en RA zorgen beiden via de inductie van CD103 voor de tolerante status bij de DCs. Het is
vooral RA die zorgt voor de inductie van CD103. TGF-β en RA kunnen elk apart de expressie
van CD103 induceren, maar dan enkel wanneer ze in zeer hoge concentraties aanwezig zijn.
In fysiologische concentraties zijn ze beiden vereist om dit effect te bereiken.
Net zoals DCs die geconditioneerd zijn met TGF-β zelf TGF-β gaan produceren, gaan ook
DCs die geconditioneerd zijn met RA vervolgens zelf RA produceren. Tijdens hun
conditionering met RA wordt de expressie van aldh1a2 geïnduceerd. Dit heeft dezelfde
functies als bij TGF-β. Ten eerste zou het zelf geproduceerd RA op een autocriene manier
werken en zo kunnen de DCs hun eigen activering onderdrukken. (13, 31) Ten tweede is deze
productie is vereist voor het uitvoeren van hun functies.
De mogelijkheid van DCs om RA te produceren speelt een belangrijke rol in veel van
hun functies: de inductie van Foxp3+Tregs, de migratie van T-cellen vanuit de MLNs naar de
darm, de inductie van IgA producerende plasmacellen, de suppressie van Th17.

20
2.2 Neurologische beïnvloeding
De darm en de hersenen staan met elkaar in contact via de HPA-as en het autonoom
zenuwstelsel. Op die manier kunnen ze steeds met elkaar communiceren. Het intestinaal
autonoom zenuwstelsel bevat zowel orthosympathische en parasympatische zenuwvezels.
Deze bevinden zich in plexussen in het darmweefsel nl. de plexus van Meisner en de plexus
van Auerbach. We vinden ook gliale cellen terug in de submucosa van het darmweefsel.
De N. Vagus maakt ook deel uit van de bezenuwing van de darm. In het darmweefsel vinden
we vele verschillende neurotransmitters terug waaronder acetylcholine, adrenaline, VIP
(vasoactive intestinal peptide) en PACAP (pituitary adenylate cyclase-activating polypeptide).
Allemaal hebben ze een invloed op de DCs.
Immature DCs behandeld met adrenaline worden gerekruteerd naar de MLNs. Daar
vergemakkelijkt adrenaline de antigenpresentatie door de DCs. Dit leidt tot een betere
inductie van tolerantie. (21)
VIP wordt vrijgesteld door de zenuwen, maar wordt ook gesecreteerd door bepaalde
immunologische cellen onder invloed van pathogene antigenen en tijdens inflammatoire
condities. VIP fungeert dan als een anti-inflammatoire factor. De effecten van VIP/PACAP op
DCs zijn verschillend naargelang er inflammatie aanwezig is of niet. (37)
VIP en PACAP binden op dezelfde receptoren: VPAC1 en VPAC2. VPAC1 is veel meer
aanwezig en is de belangrijkste receptor van de twee. (37) Ze behoren beiden tot de familie
van de G-gekoppelde proteïnes.
VIP/PACAP is ook in staat de DCs te conditioneren (38, 39) Ze hebben een invloed op
de maturatie van DCs, de expressie van co-stimulatoire molecules en de migratie van DCs.
Deze effecten verschillen naargelang het stadium van maturatie waarin de DC zich bevindt op
het moment dat ze geconditioneerd wordt door VIP/PACAP: de effecten van VIP/PACAP
zijn dus verschillend voor immature en mature DCs. (21, 37)
Immature DCs die geconditioneerd met VIP en PACAP nemen een tolerogeen phenotype aan:
ze produceren lage hoeveelheden pro-inflammatoire cytokines zoals TNF, IFN-γ, IL-12 en
IL-6, daarentegen produceren ze hoge hoeveelheden IL-10. Ze zijn niet meer in staat volledig
te matureren na stimulatie met inflammatoire stimuli. Wanneer ze geactiveerd worden met
LPS, brengen ze geen co-stimulatoire molecules zoals CD80, CD86, CD83 en CD40 meer tot
expressie. (38-40)

21
Het moleculair mechanisme verloopt als volgt: VIP/PACAP bindt op VPAC1adenylaat
cyclasecAMPPKAinhibitie van de activatering van NF-κB door de inhibitie van de
translocatie naar de kerninhibitie van de transcriptie van de genen die instaan voor de
productie van co-stimulatoire molecules en pro-inflammatoire cytokinesgeen productie van
co-stimulatoire molecules en pro-inflammatoire cytokines. (40) Op die manier verliezen ze
hun capaciteit om de proliferatie van T-cellen en differentiatie naar Th2-type T-cellen te
stimuleren.
DCs geconditioneerd met VIP/PACAP vertonen een verhoogde capaciteit voor endocytose en
dus een verhoogde antigen-capterende capaciteit. (21, 38)
VIP/PACAP heeft een effect op de migratie van mature DCs (21, 41). VIP/PACAP inhibeert
het tot expressie brengen van CCR7 en stimuleert het tot expressie brengen van CCR1. (41)
Dit gebeurt op niveau van het mRNA. Hierdoor kunnen deze niet migreren naar de MLNs,
zodat ze geen T-cellen kunnen activeren. Op deze manier wordt een inflammatoire respons
voorkomen. Ze hebben dit effect niet op immature DCs.
2.3 Apoptotische cellen
2.3.1 Apoptotische dendritische cellen
Apoptotische DCs spelen ook een rol in het bewerkstelligen van homeostase. (42)
DCs hebben een beperkte levensduur. De levenscyclus van een DC loopt van differentiatie
vanuit een progenitor cel tot apoptose na het presenteren van een antigen. In steady state
condities is er een snelle graad van turnover. Tijdens de maturatie van DCs zorgen bepaalde
genen ervoor dat de DCs nu een maximale immuunrespons kunnen bewerkstelligen. Tijdens
de maturatie worden er eveneens genen geactiveerd die een rol spelen in de apoptose van de
DC. Doordat DCs in apoptose gaan nadat ze antigenen gepresenteerd hebben, wordt de
omvang van de immuunrespons beperkt. Dit gebeurt met behulp van twee systemen.
Enerzijds doordat wanneer de DCs in apoptose gaan ze geen antigenen meer kunnen
presenteren en dus ook geen immuuncellen meer kunnen activeren. Anderzijds doordat
apoptotische DCs kunnen opgenomen worden door andere DCs en dit leidt tot tolerantie
doordat deze DCs het ontstaan van regulatoire T-cellen bewerkstelligen.
A. Welke pathways spelen een rol in de regulering van apoptose bij dendritische cellen?
Verschillende pathways zijn verantwoordelijk voor de regulering van apoptose van de DCs.
Zij kunnen anti-apoptotisch zijn via het activeren van anti-apoptotische genen of
pro-apopotitisch via het activeren van pro-apoptotische genen. Dit gebeurt zowel via
T-cel-dependente als T-cel-independente mechanismen. T-cel-dependente mechanismen zijn:
CD154 (CD40L)-CD40, TRANCE-RANK/OPG en FasL (CD95L)-Fas (CD95).

22
De T-cel-independente mechanismen zijn: LPS-CD14, TRAIL-DR4/DR5, NUR77 familie,
HLA-DR, CCR7-CCL19/CCL21, Leptine-CD295, IFN Type I-IFN Type I Receptor en
amyloïde peptiden-GPCR/FPRL1.
Men kan twee soorten pathways onderscheiden: een extrinsieke en een intrinsieke pathway. In
de extrinsieke pathway bindt een apoptose-inducerend ligand aan een receptor, hierdoor wordt
een apoptose-inducerend signalisatiecomplex gevormd, dat caspase 8 en 10 activeert. In de
intrinsieke pathway komen de signalen vanuit de cel. Deze signalen induceren een verhoogde
permeabiliteit van de buitenste membraan van de mitochondria. Normaal wordt de
permeabiliteit van de mitochandria onderdrukt door Bcl-2 door de pro-apoptotische proteïnes
Bax en Bak, die de membraan depolariseren, te inhiberen. Bcl-2 is in hoge mate aanwezig in
immature DCs, maar vermindert naarmate de DCs matureerd.
Fig. 6. (42) Inductie van apoptose in de DCs. De activering van verschillende receptoren induceert apopotose in
de DCs. Dit gebeurt via de tussenkomst van T-cellen of zonder de tussenkomst van T-cellen. De activering van
deze receptoren leidt via verschillende intracellulaire mechanismen tot apoptose.

23
B. Welke signalen triggeren de apoptose van DCs?
Verschillende stimuli kunnen apoptose bij DCs induceren. Dit leidt tot immuunsuppressie. De
verschillende stimuli gebruiken dan ook de inductie van apopotose bij DCs als een
mechanisme om een gedaalde immuniteit te bekomen.
Volgende stimuli kunnen leiden tot de inductie van apoptose: virussen, parasieten en
bacteriën; pathologische omstandigheden zoals borstkanker, sepsis en trauma;
glucocorticoïden, tumoren, UV-straling en Foxp3+Tregs.
C. Opname van apoptotische DCs door levende DCs leidt tot tolerantie
Apoptotische DCs zijn in staat tot de inductie van een tolerant phenotype in DCs.
Onder invloed van de binding met apoptotische DCs gaan de DCs meer TGF-β produceren.
Dit gebeurt door twee mechanismen. In omstandigheden zonder stimulatie is er altijd een
basale hoeveelheid TGF-β aanwezig in de DC. Dit wordt onderdrukt door LPS. Na opname
van apoptotische DCs wordt de DC ongevoelig voor signalen van LPS. Op die manier wordt
het TGF-β niet meer onderdrukt door LPS. Een ander mechanisme verloopt via de
mTOR-pathway. Binding van een nog niet gekende ligand afkomstig van de apoptotische
DCs met een nog niet gekende receptor op de DC leidt tot de activering van mTOR. mTOR
activeert vervolgens eIF4E dat in staat voor de transcriptie van TGF-β mRNA. Zoals eerder
vermeld, bevindt het gevormde TGF-β zich nog in de inactieve vorm en moet het nog
geactiveerd worden door ανβ8. De ligand hiervoor, afkomstig van apoptotische DCs, is nog
niet gekend. De tolerante DCs zijn vervolgens via hun secretie van TGF-β in staat
Foxp3+Tregs te induceren. Tevens vermindert hun vermogen om T-cellen tot Th17-cellen te
laten differentiëren. Dit komt vermoedelijk door hun gedaalde productie van IL-6. (42, 43)
Opname van apoptotische DCs door DCs zou ook kunnen leiden tot de activering van
PD-L1 (programmed death ligand 1) en PD-L2 (programmed death ligand 2). PD-L1 en
PD-L2 binden vervolgens met de PD-1 (programmed death-1) receptor op T- en B-cellen. Op
die manier wordt de immunologische capaciteit van de T- en B-cellen onderdrukt. (42)
De opname van apopototische DCs door DCs leidt dus tot tolerantie via twee mechanismen.
2.3.2 Andere apoptotische cellen
DCs worden beïnvloed door de omgeving waarin ze zich bevinden. Zo worden ze ook
beïnvloed door naburige celdood. DCs zijn in staat allerlei soorten apoptotische cellen op te
nemen. Dit om de homeostase in de omgeving te bewaren. Maar ze doen zelfs meer: ze
kunnen ook actief immunosuppressie bewerkstelligen. (44) Opname van apoptotische cellen
voorkomt auto-immuniteit door tolerantie tegenover auto-antigenen te bewerkstelligen.

24
Apoptotische cellen zijn immers een bron van auto-antigenen. Opname van deze apoptotische
cellen leidt tot tolerantie tegenover deze auto-antigenen doordat de DCs deze vervolgens op
een tolerante wijze te presenteren aan het adaptive immuunsysteem. (45)
Wanneer DCs apoptotische cellen opnemen, dan leidt dit ook tot de inductie van een
tolerant phenotype in de DCs. De DCs krijgen zo een immuunregulerende functie bij opname
van apoptotische cellen. (44)
Bij de inductie van een tolerant phenotype zijn volgende receptoren betrokken: CD36 receptor
en de integrines ανβ3en ανβ5. (32) Maw. apoptotische cellen veranderen dus het phenotype
en de functie van DCs.
Blootstelling van DCs aan apoptotische T-cellen leidt tot IFN-γ gemediëerde inductie
van IDO (indoleamine 2,3-dioxygenase) (47). IDO onderdrukt vervolgens de proliferatie van
T-cellen. (45)
Fig. 7. (42) Inductie van een tolerant phenotype in DCs via de opname van apoptotische DCs. Apoptotische
cellen waaronder apoptotische DCs hebben een invloed op DCs via de activering van receptoren. Dit leidt tot de
inductie van een verhoogde hoeveelheid TGF-β dat vervolgens een rol zal spelen in de functies van DCs
waaronder de inductie van Foxp3+Tregs.

25
2.4 IDO
DCs produceren steeds een kleine basale hoeveelheid IDO, maar verschillende
signalen zijn in staat de productie van IDO op te drijven: activering van de aryl hydrocarbon-
receptor (AhR), TLR9, TGF-β, apoptotische cellen, CTLA-4 (cytotoxic T-lymphocyte
antigen-4) van Tregs. (48) De fysiologische ligand die de Ahr op de DCs bindt, is nog niet
gekend. Binding van CpG oligodeoxynucleotides op TLR9 leidt tot de stijging van de
productie van IDO. Dit leidt tevens tot het tot expressie brengen van enkele
oppervlaktemolecules die vereist zijn in het induceren van Foxp3+Tregs. (49) Blootstelling
van DCs aan apoptotische T-cellen leidt tot IFN-γ gemediërde inductie van IDO (47).
Autocrien TGF-β zorgt ervoor dat de DCs hun tolerant phenotype dat geïnduceerd
werd door IDO langdurig behouden. TGF-β induceert en onderhoudt de productie van IDO in
DCs. Dit gebeurt via de atypische NF-κB -pathway. De PI3K/Akt-pathway (PI3K activiteit en
Akt posphorylatie) of ook wel mTor-pathway genoemd, is de tussenschakel die TGF-β met de
NF-κB-pathway verbindt. Het moleculair mechanische verloopt als volgt:
TGF-βactivering van de PI3K/Akt-pathwayphosporylatie van IDO-ITIM
(=immunoreceptor tyrosine-based inhibitory motifs) door kinase FYN +verhoogde productie
van SHP-1 en SHP-2 (=tyrosine phospatases)vorming van SHP-1/SHP-2-IDO-
complexenactivering van de phosphorylatie capaciteit van SHP-1 en SHP-2.
Activering van SHP-1 en SHP-2 leidt vervolgens tot volgende gebeurtenissen:
activering van de NF-κB -pathwayproductie van IDO.
IRAK1-pathwayproductie van IFN-α productie van IDO.
De onderhouden productie van IDO zorgt ook voor de onderhouden productie van
TGF-β via de metabolisatie van trypthophan tot 3-HAA.
Via eenn rol in het onderhouden van de productie van TGF-β speelt IDO dus ook een rol in
het onderhouden van de eigen productie en dus zo voor het onderhouden van het tolerante
phenotype.
Hoe zorgt IDO er nu voor dat de DCs een tolerant phenotype aannemen?
IDO is een enzyme dat de eerste stap in de metabolisatie van trypthophan tot kynurenine
kataboliseert. Kynurenine wordt vervolgens verder gemetaboliseerd tot 3-HAA. 3-HAA
veroorzaakt suppressie van de productie van IL-6 en stimuleert de productie van TGF-β. (50)
Zoals eerder vermeld zorgt TGF-β voor het onderhouden van de productie van IDO en het
tolerant phenotype. Het speelt ook een rol hun functie.

26
IDO en TGF-β werken dus samen om een tolerant phenotype in de DCs te induceren en te
onderhouden, doordat ze elkaars productie onderhouden: TGF-β zorgt voor de verhoogde
productie van IDO. IDO zorgt dan oa. voor de productie van TGF-β. TGF-β werkt in op
autocriene wijze op de DCs en onderhoudt de productie van IDO en dus zo ook de eigen
productie.
DCs die IDO produceren spelen een rol in de inductie van Foxp3+Tregs en in de
suppressie van T-cellen. (204) (51)
2.5 IL-10
DCs gemodificeerd met IL-10 zijn tolerant, doordat IL-10 ervoor zorgt dat ze hun
immature status behouden. IL-10 induceert ook de expressie van CD276 (B7-H3) en VTCN1
(B7-H4). (52)
Deze DCs spelen een rol in de inductie van Tr1s. (53, 54)
2.6. HLA-G en ILT4
De binding van HLA-G (Human leukocyte antigen-G) en ILT4 (Ig-like transcript 4) is
ook in staat een tolerant phenotype in de cel te induceren.(55-58) Dit gebeurt door ervoor te
zorgen dat de DCs hun immature status behouden door een stop in de maturatie van DCs te
bewerkstelligen.
Hoe zorgt de activering van ILT-4 door HLA-G nu voor inhibitie van maturatie in
DCs? (58, 59) Binding van ILT4 met HLA-G leidt tot de phosphorylatie van ILT4. Hierdoor
worden de phosphatasen SHP-1 en SHP-2 gerekruteerd. SHP-1/2 activeert vervolgens NF-κB
dat de productie van IL-6 induceert. De activering van NF-κB door SHP-1/2 is afhankelijk
van MAP-kinase. (58) IL-6 is een inflammatoir cytokine, maar wanneer de productie van IL-6
niet op de gebruikelijke wijze geïnduceerd werd – zoals hier het geval is – dan gedraagt IL-6
zich als een anti-inflammatoir cytokine. (59) IL-6 induceert de activering van STAT3. STAT3
activeert cathepsin S, dat een stop veroorzaakt in de pathway die zorgt voor de samenstelling
en transport van MHC class II molecules naar het celoppervlak. Daardoor is er een minder
efficiënte expressie van MHC molecules en de MHC molecules vertonen ook structurele
afwijkingen. Daardoor worden er kan de DC minder goed antigenen presenteren. (58)

27
Fig. 8. (59) Inductie van tolerantie in DCs door HLA-G en ILT4.
ILT4+HLA-GSHP-1/2NF-κBIL-6STAT3cathepsin Sinhibitie MHCimmature, tolerante DCs.
Deze DCs spelen een rol in de inductie van Tr1s.
2.7 IL-27
DCs ontvangen bepaalde signalen die de productie van IL-27 in de cel induceren. (46)
Het gaat om signalen afkomstig van TLR3, TLR4, TLR9 ,commensale gram negatieve
bacteriën (maar niet gram positieve), IFN-α, IFN-β, IFN-γ, invaderende pathogenen, LPS,
CD40 en inflammatoire cytokines (60, 61)
Deze DCs produceren en secreteren vervolgens zelf IL-27.
IL-27 zorgt voor de onderdrukking van de productie van IL-6, IL-23 en IL-1 β in de DC. (50)
Deze DCs spelen een rol in de inductie van Tr1s, de suppressie van T-cellen en de
inductie van IgA producerende plasmacellen.
2.8 PD-1/PD-1L
Soluble PD-1 (sPD-1) is in staat DCs te conditioneren.(62). Dit gebeurt vermoedelijk
via ‘reverse signaling’, dwz. dat een ligand gaat fungeren als receptor. PD-L1 op de DCs
fungeert hier dus als receptor van sPD-1. Op die manier zorgt sPD-1 voor een tolerant
phenotype. Dit PD-1 kan afkomstig zijn van Tregs. De DCs gaan vervolgens IL-10
produceren.
DCs die geconditioneerd zijn via de activering van hun PD-1L zijn in staat IL-10 te
produceren. Deze DCs spelen een rol in de inductie van Foxp3+Tregs en Tr1.

28
3. Functies van tolerante dendritische cellen
3.1 Invloed op T-cellen
3.1.1 Inductie van Tregs
3.1.1.1 Foxp3+Tregs
Foxp3+Tregs spelen een belangrijke rol in het behoud van tolerantie en het voorkomen van
auto-immuniteit. Ze worden gekenmerkt door de expressie van Foxp3+ op hun celoppervlak
Ze ontstaan uit naïeve CD4+T-cellen en in mindere mate uit CD8+T-cellen die onder invloed
van bepaalde factoren Foxp3+ tot expressie brengen. We onderscheiden twee soorten
Foxp3+Tregs: de natuurlijk voorkomende Foxp3+Tregs die ontstaan in de thymus en de
geïnduceerde Foxp3+Tregs die ontstaan in de periferie zoals de darm.
3.1.1 1.1 Ontstaan
3.1.1.1.1.1 Natuurlijke Foxp3+Tregs
De natuurlijke Foxp3+Tregs ontstaan in de thymus onder invloed van DCs die
geconditioneerd zijn met TSLP afkomstig van de epitheliale cellen in de thymus.TSLP stelt
de DCs in staat Foxp3+Tregs te induceren door de expressie van Foxp3+ bij de naïeve
T-cellen te induceren.
De hoeveelheid natuurlijke Foxp3+Tregs kan later verder uitgebreid worden door mature
DCs. (63) Deze uitbreiding is afhankelijk van CD80/CD86 en IL-2 dat een essentiële rol
speelt in de overleving van Tregs. (64)
3.1.1.1.1.2 Geïnduceerde Foxp3+Tregs
Geïnduceerde Foxp3+Tregs ontstaan in de periferie onder invloed van CD103+DCs
die zijn ontstaan onder invloed van de conditionering met TGF-β, RA, IDO en PD-1.
Deze DCs induceren vervolgens Foxp3+Tregs met behulp van TGF-β, RA, IDO en PD-1L.
A. De inductie van Foxp3+Tregs door TGF-β en RA
CD103+DCs produceren TGF-β en RA die de expressie van Foxp3+ in naïeve T-cellen
induceren. (5, 30, 65, 66) Deze T-cellen worden vervolgens Foxp3+Tregs. Dit gebeurt ter
hoogte van de MLNs. (5)
TGF-β en RA afkomstig van CD103+DCs bindt op hun overeenkomstige receptoren op de
T-cellen, waardoor deze Foxp3 tot expressie brengen. Deze molecule speelt een rol in hun
ontwikkeling: het zet een genetische programma in gang dat nodig is voor hun differentiatie.
Het precieze mechanisme hoe de invloed van TGF-β en RA leidt tot de expressie van Foxp3
is niet gekend.

29
CD103+DCs induceren tevens de RA-receptor in de nieuw gevormde Foxp3+Tregs, wat hen
gevoelig maakt voor RA dat ook een rol speelt in andere processen. (66)
TGF-β en RA zijn beiden vereist en werken samen in een synergistisch verband. (65)
DCs werken ook samen met B-cellen om Foxp3+Tregs induceren (64). Wanneer ze samen
werken, verhogen ze elkaars capaciteit om Foxp3+Tregs te induceren. Het gaat dus ook om
een synergistische samenwerking.
B. De inductie van Foxp3+Tregs door IDO
CD103+DCs produceren IDO. IDO zorgt via twee molecules nl. trypthophan en
kynurenine voor de inductie tolerantie.
Ten eerste leidt IDO tot een depletie van trypthophan in de DC. Dit leidt tot de inductie van
de receptoren ILT3 en ILT4 die een rol spelen in de inductie van Foxp3+Tregs. (50) Hoe dat
precies gebeurt, is nog niet gekend.
Ten tweede leidt IDO tot de productie van kynurenine, een metaboliet van tryptophan. Het
bindt op de AhR op de T-cellen en induceert zo een tolerant phenotype. (67)
TGF-β speelt hier ook een rol in: het vergemakkelijkt de binding tussen kynurenine en AhR.
TGF-β zorgt eveneens voor een verhoogde presentatie van AhR op de celmembraan van
T-cellen.
De inductie van Foxp3+Tregs door DCs die IDO produceren vereist cel-celcontact.
Volgende molecules op de celmembraan van de DCs zijn vereist voor dit cel-celcontact:
CD80, CD86 en HLA-DR. (49, 68)
In het lichaam is een beschermmechanisme ingebouwd om toch nog Foxp3+Tregs te
kunnen induceren, wanneer de productie van IDO geblokkeerd is. (68) Naast het endogene
kynurenine geproduceerd met behulp van IDO, is er ook exogeen kynurenine. Dit kan dan
ingeschakeld worden om de functie van het endogeen kynurenine over te nemen.
C. De inductie van Foxp3+Tregs door PD-1L
Binding tussen PD-1L op de DCs met PD-1 op de naïeve T-cellen leidt in de aanwezigheid
van TGF-β tot de differentiatie van de naïeve T-cel naar een Foxp3+Tregs. (69) De
Foxp3+Tregs bezitten ook de PD-1L. Zo leidt binding tussen PD-L1 op de Foxp3+Tregs en
PD-1 op de naïeve T-cellen ook tot de differentiatie van de naïeve T-cel naar Foxp3+Tregs.
(69) Foxp3+Tregs kunnen dus zelf zorgen voor een uitbreiding van hun aantal. De
PD-1/PD-1L pathway fungeert ook als een hulp voor Foxp3+Tregs. Foxp3+Tregs die
gestimuleerd worden door PD-L1 zijn beter in staat om Foxp3 tot expressie te brengen en
vertonen een betere suppressieve capaciteit. Dat is voornamelijk zo wanneer de hoeveelheid
Foxp3+Tregs laag is ten opzichte van de hoeveelheid effector T-cellen. (69)
30
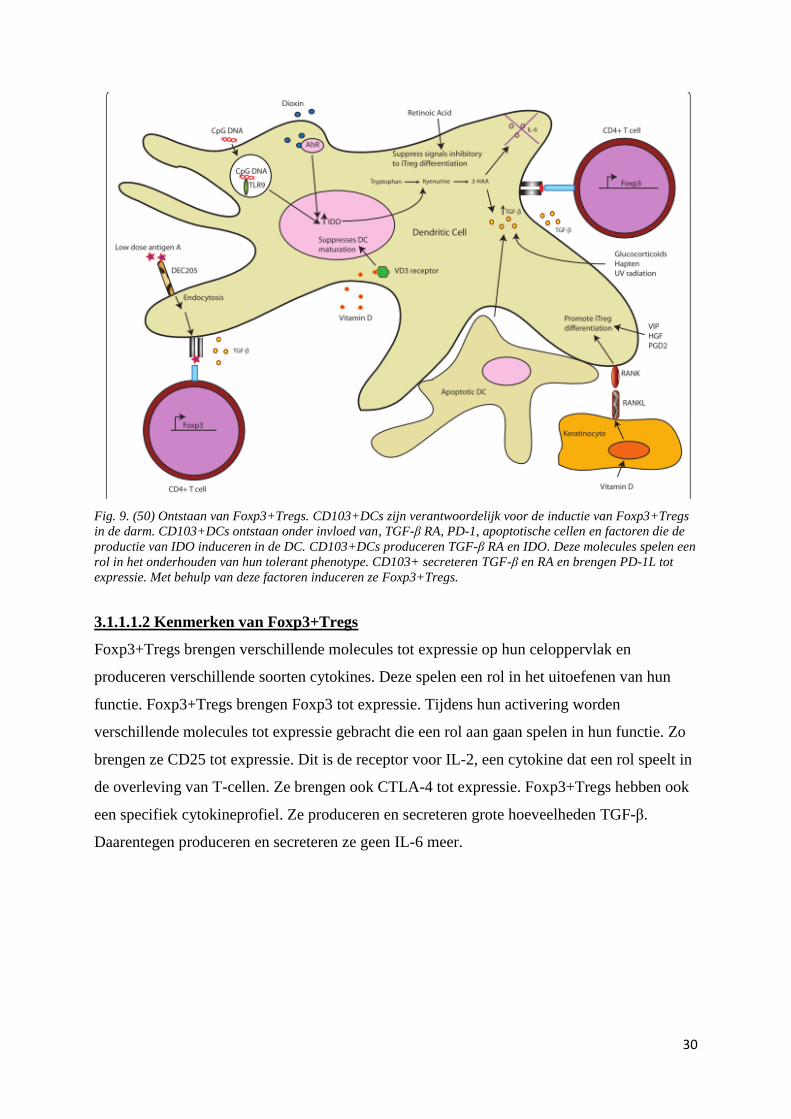
Fig. 9. (50) Ontstaan van Foxp3+Tregs. CD103+DCs zijn verantwoordelijk voor de inductie van Foxp3+Tregs
in de darm. CD103+DCs ontstaan onder invloed van, TGF-β RA, PD-1, apoptotische cellen en factoren die de
productie van IDO induceren in de DC. CD103+DCs produceren TGF-β RA en IDO. Deze molecules spelen een
rol in het onderhouden van hun tolerant phenotype. CD103+ secreteren TGF-β en RA en brengen PD-1L tot
expressie. Met behulp van deze factoren induceren ze Foxp3+Tregs.
3.1.1.1.2 Kenmerken van Foxp3+Tregs
Foxp3+Tregs brengen verschillende molecules tot expressie op hun celoppervlak en
produceren verschillende soorten cytokines. Deze spelen een rol in het uitoefenen van hun
functie. Foxp3+Tregs brengen Foxp3 tot expressie. Tijdens hun activering worden
verschillende molecules tot expressie gebracht die een rol aan gaan spelen in hun functie. Zo
brengen ze CD25 tot expressie. Dit is de receptor voor IL-2, een cytokine dat een rol speelt in
de overleving van T-cellen. Ze brengen ook CTLA-4 tot expressie. Foxp3+Tregs hebben ook
een specifiek cytokineprofiel. Ze produceren en secreteren grote hoeveelheden TGF-β.
Daarentegen produceren en secreteren ze geen IL-6 meer.

31
3.1.1.2 TR1s
3.1.1.2.1 Ontstaan van Tr1
Tr1s (type 1 regulatoire T-cellen) worden geïnduceerd door DCs die IL-27, IL-10 en TGF-β
secreteren en ICOS-ligand (inducible T-cell costimulator-ligand) tot expressie brengen. De
productie van IL-27 wordt geïnduceerd door allerlei signalen waaronder IFN-γ. IL-27 zorgt
vervolgens voor de onderdrukking van de productie van IL-6, IL-23 en IL-1 β in de DC. De
DC zal ook dit IL-27 secreteren. Verschillende factoren zorgen voor de inductie van de
productie van IL-10 in de DC nl. IL-10, VIP en de activering van PD-1. De DCs zullen ook
het IL-10 dat ze produceren, gaan secreteren.
A. Inductie van Tr1s door IL-27
De productie van IL-27 in de DCs heeft een invloed op de inductie van Tr1.
DCs die geconditioneerd werden met signalen die de productie van IL-27 in de cel induceren,
produceren dus IL-27. Zij secreteren vervolgens hun IL-27 en dat induceert verschillende
effecten in de T-cel, waaronder de productie van IL-10 en IL-21. Deze T-cellen differentiëren
zo naar Tr1s. (46) IL-27 heeft volgende effecten op de T-cel:
Secretie van IL-27 door de DCinductie van de transcriptiefactoren c-Maf+AhR in
CD4+T celactivering van de promotors van IL-10 nl. STAT1 en STAT3
inductie van de productie van IL-10Tr1-inductie (70-72)
secretie van IL-27 door de DC inductie van de transcriptiefactoren c-Maf+AhR in
CD4+T celactivering van de promotor van IL-21 IL-21 productieTr1-expansie (70-72)
Secretie van IL-27 door DCinductie van Ahr. (70)
Secretie van IL-27 door DC inductie van ICOSL op de celmembraan van de T-cel. (71, 72)
Secretie van IL-27 door DCinductie van het tot expressie brengen van de IL-21 receptor.
(72)
Binding van IL-27 op de IL-27 receptor leidt tot de phosphorylatie van STAT1 en
STAT3. STAT1 en STAT3 zijn promotors van de IL-10 transcriptie en brengen zo de
productie van IL-10 op gang. (60, 73) De functie van deze DCs om Tr1 te induceren is dus
afhankelijk van STAT1 en STAT3. (226) De functie van deze DCs om Th17 te onderdrukken
daarentegen is enkel afhankelijk van STAT1. (73)
IL-21 werkt op autocriene wijze op de T-cellen en zorgt zo voor de amplificatie van
Tr1. (71, 72) Dit gebeurt door het versterken van de effecten van IL-27 op de T-cel. (70)
Signalisatie van ICOS zorgt voor het onderhouden van de productie van IL-10 en IL-21 door
de T-cellen.

32
Dit gebeurt door het onderhouden van het tot expressie brengen van c-Maf (71, 72) en Ahr.
(70) IL-21 en ICOS zorgen er dus voor dat Tr1s hun phenotype behouden, onafhankelijk van
IL-27.
Fig. 10. (71) Effecten van IL-27. IL-27 induceert de productie van IL-21 en IL-10 in de T-cel, waardoor deze
differentiëren naar Tr1s. IL-27 leidt tevens tot de verhoogde expressie van receptoren voor IL-21 en ICOSL.
IL-21 werkt op autocriene wijze en onderdhoudt zo de productie van IL-21 en IL-10.
IL-27 werkt samen met TGF-β om de inductie van Tr1s te verbeteren. TGF-β helpt het
effect van IL-27 te vergroten. (53, 72) IL-27 en TGF-β hebben het volgende effect op naïeve
T cellen: (53)
T cellen die alleen door IL-27 worden geactiveerd produceren IL-10, een beetje IFN-γ, IL-5
en IL-13. T-cellen die daarentegen geactiveerd werden door IL-27 én TGF-β produceren meer
IL-10, minder Il-4, IL-5, IL-13.
IL-27 induceert IL-10, TGF-β zorgt voor onderhoud van IL-10.
De inductie van Ahr is groter wanneer de DCs geactiveerd wordt door IL-27 én TGF-β.
(70)
B. Inductie van Tr1s door IL-10
DCs geconditioneerd met IL-10 behouden hun immature phenotype en produceren
grote hoeveelheden IL-10. Herhaaldelijke stimulatie van T-cellen met immature DCs leidt tot
de inductie van een Tr1 phenotype in de T-cellen. Dit mechanisme is afhankelijk van IL-10.
(54) De inductie van Tr1 door DCs die IL-10 secreteren is ook afhankelijk van HLA-G en
ILT4. (55)
33
Autocriene productie van IL-10 door DCs zorgt voor de verhoogde expressie van ILT4 en
HLA-G op DCs en is ook vereist voor de verhoogde expressie van HLA-G op de CD4+T
cellen. (55, 57) Dit verklaart de rol van IL-10 in de inductie van Tr1 door DCs.
C. Inductie van Tr1s door ICOSL
IL-27 staat in voor de inductie van ICOS op de T-cel.
De activering van ICOS op de T-cel door ICOSL op de DCs leidt tot de inductie van de
productie van IL-21 en IL-10 via activering van c-Maf. (72, 74)
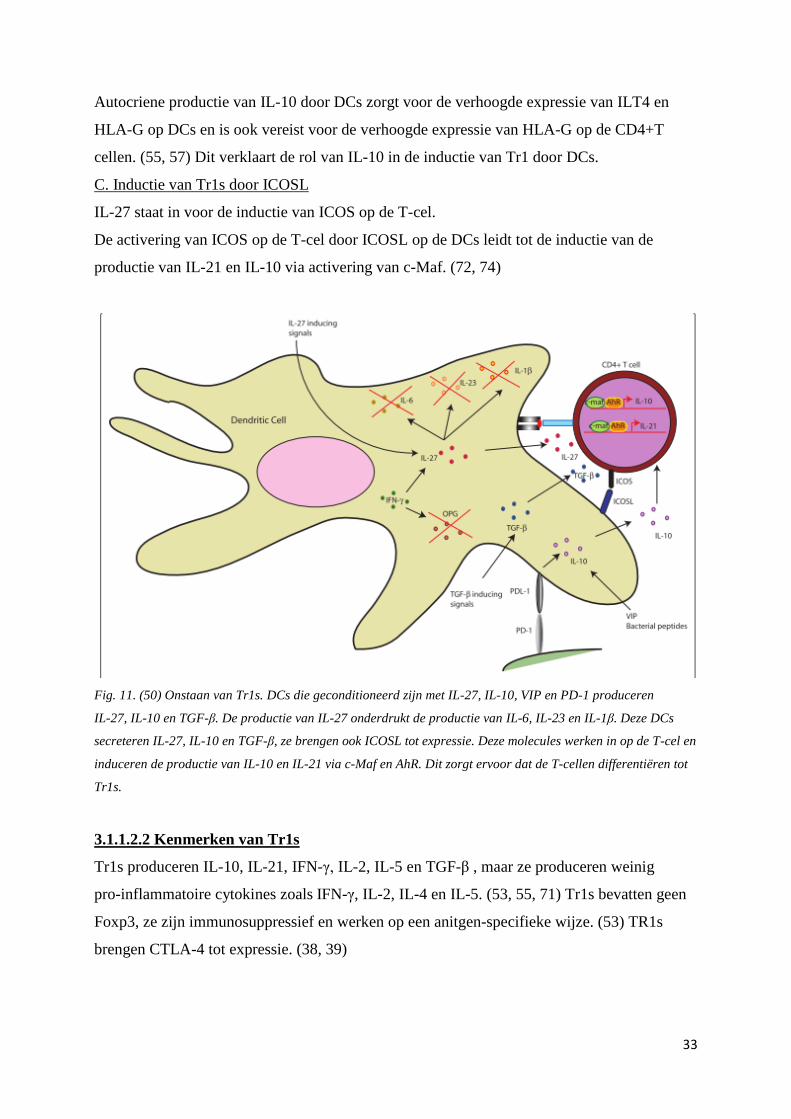
Fig. 11. (50) Onstaan van Tr1s. DCs die geconditioneerd zijn met IL-27, IL-10, VIP en PD-1 produceren
IL-27, IL-10 en TGF-β. De productie van IL-27 onderdrukt de productie van IL-6, IL-23 en IL-1β. Deze DCs
secreteren IL-27, IL-10 en TGF-β, ze brengen ook ICOSL tot expressie. Deze molecules werken in op de T-cel en
induceren de productie van IL-10 en IL-21 via c-Maf en AhR. Dit zorgt ervoor dat de T-cellen differentiëren tot
Tr1s.
3.1.1.2.2 Kenmerken van Tr1s
Tr1s produceren IL-10, IL-21, IFN-γ, IL-2, IL-5 en TGF-β , maar ze produceren weinig
pro-inflammatoire cytokines zoals IFN-γ, IL-2, IL-4 en IL-5. (53, 55, 71) Tr1s bevatten geen
Foxp3, ze zijn immunosuppressief en werken op een anitgen-specifieke wijze. (53) TR1s
brengen CTLA-4 tot expressie. (38, 39)

34
3.1.1.3 Functies van Tregs
A. Invloed op T-cellen
De precieze functie van Tregs is nog niet goed gekend. In het algemeen bezitten de
Tregs een negatief immuunmodulerende rol. Die rol oefenen ze uit op verschillende soorten
cellen waaronder T-cellen, DCs, natural killer cellen (NK-cellen) en antigen-presenterende
cellen (APCs). Bepaalde cytokines zoals IL-10, TGF-β en IL-2 spelen een belangrijke rol in
de functie van Tregs. Deze cytokines hebben een invloed op de celcyclus en zijn in staat
apoptose te induceren in T-cellen. Tevens hebben deze cytokines een invloed op de
co-stimulatie en de maturatie van DCs. De natuurlijke Foxp3+Tregs, de geïnduceerde
Foxp3+Tregs en de Tr1s produceren verschillende cytokines wat zich uit in verschillende
functies. Natuurlijke Foxp3+Tregs produceren voornamelijk TGF-β en granzyme-B,
Foxp3+Tregs voornamelijk TGF-β en Tr1s voornamelijk IL-10.
De inductie van Foxp3 in T-cellen door DCs stimuleert het tot expressie brengen van
de IL-2 receptor in de Foxp3+Tregs. Zo zijn de Foxp3+Tregs gevoeliger voor IL-2.
IL-2 speelt een belangrijke rol in overleving van Tregs. De verhoogde gevoeligheid zorgt
tevens voor depletie van IL-2 in de omgeving waar effector T-cellen zich bevinden. Dit tekort
aan IL-2 leidt in de effector T-cellen tot een apoptose en een verminderde proliferatie (8, 68).
Andere studies concluderen dat Foxp3+Tregs ook in staat zijn rechtstreeks de productie van
IL-2 in effector T-cellen te inhiberen. (75)
Via de productie van IL-10 en TGF-β onderdrukken de Tregs de T-cel responsen via
IL-10 en TGF-β dependente mechanismen. (54) Deze mechanismen zijn niet goed gekend.
Via de productie van IL-10 en TGF-β en zijn de Tregs in staat de proliferatie van Th1 te
inhiberen. Dit doen ze door de productie van IL-2 in de Th1 te inhiberen. (38, 39) In mindere
mate zijn ze ook in staat de proliferatie van Th2 te inhiberen. (38)
Tregs reduceren eveneens de productie van cytokines bij Th1, ze hebben daarentegen geen
invloed op het cytokineprofiel van de Th2. (38) Tregs maken de T-cellen hypo-responsief
voor secundaire stimulatie met allo-antigenen. (68) Natuurlijke Tregs veroorzaken cytolyse
van NK-cells en CD8+T-cells dmv. hun secretie van granzyme B en porferine. (75)

35
B. Invloed op DCs
In steady state vindt er een voortdurende interactie plaats tussen DCs en Tregs. Het is
dus niet alleen zo dat DCs een invloed hebben op de Tregs, de Tregs hebben ook een invloed
op de DCs. Enerzijds induceren DCs de Tregs, anderzijds hebben de Tregs een invloed op het
phenotype en de functie van de DCs. Tregs zorgen ervoor dat DCs een immunosuppressief
phenotype aannemen. Op die manier zijn de Tregs in staat hun aantal uit te breiden. (46)
Tregs kunnen migreren naar de MLNs. Daar komen ze in contact met DCs.
Tregs zijn in staat het contact tussen DCs en T-cellen te blokkeren, waardoor de
T-cellen niet meer geactiveerd kunnen worden door DCs en dus geen immuunresponsen
kunnen uitvoeren. (76)
Tregs kunnen tevens het phenotype van DCs beïnvloeden. Tregs produceren IL-10 en TGF-β.
Deze molecules onderdrukken de maturatie van DCs en bezorgen de DCs op die manier een
tolerant phenotype. (46) Dit kan ook door rechtstreeks contact tussen de Tregs en de DCs via
de binding tussen PD-1 op de Tregs en PD-1L op de DCs. (75) Dit tolerant phenotype
kenmerkt zich als volgt: productie van IL-10, gestegen expressie van B7-H4 en B7-H3 op hun
celoppervlak en gedaalde expressie van co-stimulatoire molecules op hun celoppervlak. De
expressie van B7-H4 en B7-H3 gebeurt onder invloed van IL-10. (52) Via hun productie van
IL-10 en B7-H4 en B7-H3 op hun celoppervlak kunnen DCs dan de activiteit van de T-cellen
blokkeren of de T-cellen anergisch maken. We zien dus dat de Tregs via dit mechanisme ook
een onrechtstreekse invloed hebben op T-cellen.
Foxp3+Tregs zijn in staat IL-27 te induceren in DCs. (53, 71) Ze zijn ook in staat apoptose te
induceren bij de DCs. (42) Tregs kunnen ook de productie van IDO induceren bij de DCs.
(48)

36
Fig. 12. (46) Beïnvloeding van de DCs door Tregs. Tregs blokkeren het contact tussen de DCs en de T-cellen,
waardoor deze niet kunnen geactiveerd worden. Tregs beïnvloeden het phenotype van de DCs, waardoor deze
tolerant worden en de T-cellen inhiberen.
C. IgA
Via TGF-β vormen de Foxp3+Tregs IgM B-cellen om naar IgA-B-cellen. (77) Op die manier
ontstaat er productie van IgA op een T-cel dependente wijze.
In het hoofdstuk over IgA gaan we hier uitgebreider op in.
D. Inflammatie indijken
CTLA-4 speelt naast TGF-β ook een belangrijke rol in de functie van natuurlijke
Foxp3+Tregs. (78) CTLA-4 inhibeert de T-cel activiteit doordat het een hogere affiniteit heeft
voor CD28 dan CD86 en CD80, die een rol spelen in inflammatoire responsen. Op die manier
wordt de inflammatoire respons tijdig gestopt.
3.1.2 Suppressie van T-cellen
DCs die IL-27 produceren, inhiberen de differentiatie van T-cellen naar Th17. (79) Dit
komt doordat IL-27 de productie van IL-6, IL-23 en IL-1 β in de DC onderdrukt. (50, 72) Op
die manier kan de DC geen T-cellen meer laten differentiëren tot Th17. Deze DCs inhiberen
tevens de productie van bepaalde cytokines in de T-cel bv. IL-17 dat instaan voor de functie
en het behoud van Th17.
DCs die IL-27 secreteren, inhiberen de ook differentiatie van T-cellen naar Th1 en Th2. (61)
IL-27 reduceert de expressie van transcriptiefactoren: T-bet voor Th1, GATA-3 voor Th2 en
RORC voor Th17. (79) IL-27 heeft dus een effect op vele verschillende T cellen. Dit is te
verklaren door het feit dat IL-27 een effect heeft op T-cel proliferatie via de suppressie van de
productie van IL-2 dat van belang is bij T-cel proliferatie. (61)

37
DCs die TGF-β en RA produceren, onderdrukken Th1 en Th17. Dit komt doordat deze
DCs geen IL-6 meer produceren. Het is vooral RA die ervoor zorgt dat ze de inductie van
Th17 inhiberen.
IDO onderdrukt de T-cellen op twee manieren. Ten eerste leidt IDO tot depletie van
tryptophan in de omgeving waar de T-cellen zich bevinden. Dit veroorzaakt bij de T-cellen
een stop in de celcyclus tijdens de G1-fase. Ten tweede zorgt 3-HAA ervoor dat de productie
van IL-6 in de DCs wordt geïnhibeerd. Op die manier kan de DC geen T-cellen meer laten
differentiëren tot Th17.
3.2 IgA
3.2.1 Ontstaan
A. T-cel dependent
Het ontstaan van IgA producerende plasmacellen onder invloed van T-cellen vindt
plaats ter hoogte van de PP. (80) DCs zorgen via de invloed van verschillende cytokines voor
de differentiatie van T-cellen tot verschillende Tregs.
Deze Tregs produceren vervolgens ook cytokines die IgM-B-cellen laten omschakelen
tot IgA-B-cellen die vervolgens differentiëren tot IgA producerende plasmacellen. (80, 77)
Zoals reeds blijkt uit vorige hoofdstukken worden DCs geconditioneerd door factoren
uit de omgeving nl. TSLP, TGF-β, RA en IL-10, waardoor ze differentiëren naar een tolerant
phenotype dat Tregs induceert. Dit doen ze met behulp van de secretie van bepaalde
cytokines. TGF-β samen met RA induceert Foxp3+Treg, IL-10 en IL-27 samen met TGF-β
induceert Tr1. De Tregs produceren vervolgens allerlei cytokines die van belang zijn in het
ontstaan van IgA-B-cellen nl. TGF-β, IL-4, en IL10. TGF-β activeert samen met de CD40-
ligand IgM-B-cellen en laat hen omschakelen naar IgA-B-cellen. (77) IL-4 en IL-10 zijn
verantwoordelijk voor de expansie van de IgA-B-cellen en de differentiatie naar IgA
producerende plasmacellen. IgA producerende plasmacellen produceren naast IgA ook enkele
cytokines nl. IL-6, IL-10 en TGF-β. Deze cytokines vergroten de inductie van Tregs en de
IgA producerende plasmacellen dragen dus op die manier ook bij tot hun eigen uitbreiding.
(80) Het IL-6 werkt ook op een autocriene manier in op de IgA producerende plasmacellen,
hetgeen verder bijdraagt tot de IgA productie. (81)

38
Fig. 13. (80) Ontstaan van IgA producerende plasmacellen op T-cel afhankelijke wijze. DCs produceren allerlei
cytokines die de differentiatie van Tregs bevorderen. Deze Tregs produceren op hun beurt ook cytokines die een
rol spelen in het onstaan van IgA producerende plasmacellen. IgA producerende plasmacellen produceren naast
IgA ook cytokines die een rol spelen in de verdere uitbreiding van Tregs en IgA. Zo houden ze hun eigen
productie in stand.
B. T-cel independent
Het ontstaan van de IgA producerende plasmacellen zonder tussenkomst van T-cellen
vindt plaats ter hoogte van de LP en in de MLN. (80)
Vijf molecules -RA, APRIL, BAFF, TGF-β en NO- binden op hun receptor op de B-cel,
waardoor de B-cel differentieert naar een IgA producerende plasmacel. IECs en DCs zijn
verantwoordelijk voor de productie van deze vijf molecules.
TGF-β wordt geproduceerd door IECs en DCs en bindt op de TGF-β receptor. iNOS
(inducible nitrogen oxide) wordt geproduceerd door de DCs onder invloed van de binding van
microbiota op TLRs. iNOS is belangrijk om een goede TGF-β signalisatie te hebben in de
B-cel. RA wordt geproduceerd door de IECs en de DCs. RA bindt op de RA receptor op de
B-cel. RA induceert ook op een indirecte manier IgA producerende plasmacellen, doordat het
de secretie van TGF-β en iNOS verhoogd. APRIL wordt geproduceerd door de IECs onder
invloed van signalen afkomstig van hun TLRs. (82) APRIL en BAFF worden ook
geproduceerd door DCs onder invloed van TSLP (82) dat op zijn beurt wordt geproduceerd
door IECs onder invloed van signalen afkomstig van hun TLRs.

39
iNOS is ook van belang in de productie van APRIL en BAFF door de DCs. APRIL en BAFF
binden op de TACI-receptor op de B-cellen.
De activering van de TACI-receptor leidt tot de verhoogde productie van het enzym AID
(activation-induced cytidine deaminase). VIP speelt ook een rol in de inductie van IgA
producerende B-cellen.
Fig. 14. (80) Ontstaan van IgA producerende plasmacellen op een T-cel onafhankelijke wijze. In het proces dat
leidt tot het ontstaan van IgA producerende plasmacellen op een T-cel onafhankelijke wijze kunnen we drie
hoofdrolspelers onderscheiden: IECs, DCs en het microbioom. Deze produceren gezamenlijk vijf molecules: RA,
APRIL, BAFF, TGF-β en NO. Deze molecules zorgen voor het onstaan van IgA producerende plasmacellen.
3.2.2 De rol van IgA in homeostase
De belangrijkste functie van IgA is het beschermen van het lichaam tegen het binnendringen
van allerlei ziekteverwekkers. IgA staat in voor immuunexclusie, dwz. dat IgA ervoor zorgt
dat pathogene bacteriën niet kunnen binden met het epitheel. IgA zorgt er ook voor dat het
microbioom en de immuuncellen beter contact kunnen maken met elkaar. (83) IgA staat in
voor de opsionisatie van antigenen, zodat deze beter kunnen opgenomen worden door
M-cellen. (80, 83). IgA zorgt ervoor dat bacteriën minder gemakkelijk vanuit het lumen van
de darm naar de mucosa kunnen gaan. (84) IgA voorkomt de overgroei van commensale
bacteriën.

40
3.3 Gut homing
Na hun inductie door de DCs in de MLNs moeten de Tregs naar de darm kunnen
migreren om daar hun functie uit te oefenen. CD103+DCs induceren twee
oppervlaktemolecules bij de Tregs, waardoor deze naar de darm kunnen migreren. Deze
molecules zijn het integrine α4β7 en de chemokinereceptor CCR9. (85)
De ligand voor α4β7 is MadCAM1 (mucosal addressin cell adhesion molecule-1) dat
tot expressie wordt gebracht op de postcapillaire endotheliale cellen in de dunne en dikke
darm. De ligand voor CCR9 is CCL25 dat tot expressie wordt gebracht op endotheliale cellen
van de venules in de dunne darm.
Cellen die α4β7 en CCR9 bezitten, kunnen enkel migreren naar de darm. De plaats
waar de Tregs naartoe migreren is ook afhankelijk van de lymphenodus waarin ze zijn
geïnduceerd. Tregs die geactiveerd zijn in MLNs kunnen dus enkel migreren naar intestinaal
weefsel. Op deze manier bereikt men dus weefselspecifieke migratie. (85, 86) Eenmaal
gemigreerd naar de intestinale mucosa, zijn er andere factoren vereist om de T-cellen daar ter
plaatse te houden. Vermoedelijke factoren zijn CCR2 en CCR6. (85)
De inductie van α4β7 en CCR9 op T-cellen door CD103+DCs is afhankelijk van RA.
(36) RA bindt op de RA-receptor op de Tregs. De stimulatie van de RA-receptor lijkt alleen
effectief te zijn wanneer de Tregs tegelijkertijd ook gestimuleerd worden via de TCR en
CD28. (36) De inductie van α4β7en CCR9 bij B-cellen vereist naast RA ook IL-5 en/of IL-6.
(87)
Stromale cellen in MLNs kunnen de DCs ondersteunen in de inductie van α4β7 en
CCR9 op T-cellen.(88) Stromale cellen zijn immers een grote bron van RALDH en ze
produceren dan ook grote hoeveelheden RA.
Fig. 15. (36) Migratie van Tregs naar de darm. Tregs die geïnduceerd werden in de MLNs migreren onder
invloed van α4β7en CCR9 naar de darm. Inductie van α4β7en CCR9 gebeurt door de DCs met behulp van RA.
Ter hoogte van de darm bindt α4β7met MadCAM1 en CCR9 met CCL25.

41
3.4 Co-stimulatoire molecules: PD-1 en CTLA-4
In dit hoofdstuk bekijken we hoe oppervlaktemolecules een rol spelen in homeostase
via hun invloed op effector T-cellen nl. de controle van T-cel activering en hun rol in perifere
tolerantie nl. inductie van Tregs. (69, 75)
PD-1 en CTLA-4 vinden we terug op geactiveerde lymfocyten zoals T-cellen. PD-1 en
CTLA-4 worden verhoogd tot expressie gebracht wanneer T-cellen co-stimulatoire signalen
ontvangen via CD28, maw. tijdens hun activering. CD28 bereidt zo als het ware de T-cel voor
op latere stimulatie van PD-1 en/of CTLA4. (75)
PD-1 en CTLA-4 hebben twee liganden nl. PD-L1 en PD-L2. Beide liganden vinden we terug
bij DCs. (89) PD-2L vinden we zelfs uitsluitend terug bij DCs. PD-1L is standaard aanwezig
op de DCs en de aanmaak wordt gestimuleerd bij activering door IFN-α, IFN-β, IFN-γ via de
JAK/STAT en de MAPK pathways (69). PD-2L is enkel aanwezig op de DCs na inductie.
(90)
PD-L1 en PD-L2 zijn betrokken in zowel de Treg intrinsieke als extrinsieke vorm van
perifere tolerantie. (69) Bij de Treg extrinsieke vorm is er geen tussenkomst van Tregs en
oefent de DCs haar invloed rechtstreeks uit op de T-cellen Bij de Treg intrinsieke manier is er
een tussenkomst van Tregs. De functies van PD-L1 en PD-L2 overlappen elkaar.(91) Hoewel
PD-1 en CTLA-4 gestimuleerd worden door dezelfde liganden, zijn hun functies toch licht
verschillend. Dit is gedeeltelijk te verklaren door het feit dat beide receptoren verschillend tot
expressie worden gebracht in tijd en plaats. (75) CTLA-4 wordt vooral tot expressie gebracht
door de T-cel wanneer die zich in de MLNs bevindt. CTLA-4 zal dus eerder een rol spelen in
de vroege fase. PD-1 wordt vooral tot expressie gebracht door de T-cellen, wanneer die zich
in de weefsels bevinden. PD-1 zal dus eerder een rol spelen in de latere fase. Anders gezegd:
CTLA-4 speelt vooral een rol in de inductie van tolerantie, PD-1 speelt vooral een rol in het
behoud van tolerantie.
A. T-cel extrinsiek mechanisme
De stimulatie van PD-1 en CTLA-4 leidt tot inhibitie van de T-cel activiteit
(verminderde cytokine productie, verminderde proliferatie) of induceert de apoptose van de
T-cel. (62) PD-L1 en PD-L2 zijn zo in staat auto-reactieve T–cellen te verwijderen door
apoptose te induceren in deze cellen. (75) Dit gebeurt op twee manieren:
Via de Fas-FasL pathway en onrechtstreeks via de onderdrukking van het CD28 signaal.
Co-stimulatie met CD28 verschaft immers overlevingssignalen aan de cel. Dit gebeurt via
competitie tussen CTLA-4 en CD80/CD86. CTLA-4 heeft een hogere affiniteit voor CD28
dan CD80/CD86.

42
PD-1 en CTLA-4 kunnen het TCR signaal beëindigen (75). Binding van PD-L1 en
PD-L2 met PD-1 en CTLA-4 leidt tot de phosphorylatie van PD-1 en CTLA-4 ter hoogte van
SHP1/2. Dit leidt tot de dephosphorylatie van secundaire signaalmolecules, waardoor de
intracellulaire signalisatie van TCR beëindigd wordt.
Immature DCs kunnen tolerantie induceren in CD8+T-cellen via een T-cel extrinsiek
mechanisme. Zowel PD-1 als CTLA-4 zijn hiervoor vereist, wanneer één van beide afwezig
is, veroorzaakt dit immuniteit ipv. tolerantie. (92) De T-cellen die gestimuleerd worden via dit
mechanisme worden hypo-responsief tegenover de antigenen die de DCs presenteerden samen
met PD-1 en CTLA-4. Dit noemen we anergie.
PD-L1 en PD-L2 zijn niet zozeer in staat T-cel activering zelf te inhiberen. Ze zijn wel
in staat de productie van IL-2 en IFN-γ te onderdrukken tijdens de activering van CD4+T-
cellen. (89) en de proliferatie van T-cellen te inhiberen (91).
PD-L1 en PD-L2 kunnen het signaal tussen CD80/CD86 op de DC met CD28 op de T-
cel ongedaan maken wanneer de T-cel slechts zwak wordt gestimuleerd door antigenen. (91)
Dit mechanisme wordt bevestigd door het feit dat de hoeveelheid PD-1 op de T-cel groter is
bij zwakke antigen-stimulatie.
B. T-cel intrinsiek mechanisme
Zoals beschreven werd in vorige hoofdstukken zijn DCs in staat Foxp3+Tregs en Tr1s te
induceren met behulp van PD-1L. Deze Tregs zorgen voor homeostase.
3.5 Overzicht: 3 gebieden homeostase
In de vorige hoofdstukken werden de functies van DCs besproken. In dit hoofdstuk wordt
gekeken hoe deze functies leiden tot homeostase binnen de drie gebieden nl. elimineren van
auto-reactieve T-cellen, induceren van tolerantie ten opzicht van. onschadelijke antigenen en
indijken van inflammatie.
3.5.1 Auto-reactieve T-cellen
Auto-reactieve T-cellen zijn T-cellen die auto-antigenen herkennen met hun receptor.
Zij kunnen zo leiden tot auto-immuniteit doordat ze een immuurespons kunnen opstarten
tegen auto-antigenen. Dit moet dus vermeden worden. Daarom heeft het immuunsysteem
twee mechanismen ingebouwd nl. centrale en perifere tolerantie.
Centrale tolerantie vindt plaats in de thymus en perifere tolerantie vindt plaats in de perifere
weefsels waaronder de darm. DCs spelen een rol in beide mechanismen.
De thymus is de plaats waar nieuwe T-cellen ontstaan. Deze nieuwe T-cellen hebben een
nieuw samengestelde T-cel receptor (TCR) waarmee het de antigenen, die aangeboden
worden door de DCs via MHC, kan binden.

43
De TCR is antigen-specifiek dwz. dat elke TCR slechts een soort antigen kan binden. Onder
deze nieuwgevormde TCRs kunnen zich TCRs bevinden die auto-antigenen binden. T-cellen
die zo’n TCR bezitten zijn dus auto-reactief. Het is van belang dat deze auto-reactieve
T-cellen vernietigd worden voor ze de thymus kunnen verlaten en een auto-immune respons
kunnen veroorzaken in het lichaam. Dit gebeurt dmv. twee selectieprocessen: positieve en
negatieve selectie. DCs spelen een rol in beide selectieprocessen. (26). Bij de positieve
selectie krijgen T-cellen met een TCR die in staat is te binden met MHC, het signaal om
verder te matureren. T-cellen die binden met MHCI worden CD8+T-cellen en T-cellen die
binden met MHCII worden CD4+T-cellen. T-cellen met een TCR die niet bindt met MHC
krijgen het een signaal om in apoptose te gaan. Bij negatieve selectie worden T-cellen met een
TCR die een hoge affiniteit vertoont voor auto-antigenen geëlimineerd. T-cellen met een TCR
die een lage affiniteit vertoont voor auto-antigenen krijgen het signaal om te matureren en te
migreren naar de perifere weefsels. Het is gebleken dat niet elke T-cel met een TCR die hoge
affiniteit vertoont voor auto-antigenen wordt geëlimineerd. De T-cellen die niet worden
geëlimineerd differentiëren tot Tregs. (26)
Dit lijkt een paradox te zijn: enerzijds elimineren de DCs T-cellen met een TCR die een hoge
affiniteit vertonen, anderzijds selecteren de DCs deze T-cellen uit om te laten differentiëren
tot Tregs. Dit is te verklaren door het feit dat deze verschillende functies door verschillende
types van DCs worden uitgevoerd. (26) TSLP dat gesecreteerd wordt door de epitheliale
cellen in de thymus heeft een invloed op de DCs: DCs die beïnvloed werden door TSLP zijn
in staat Tregs te induceren. T-cellen met een TCR die een hoge affiniteit vertoont voor
auto-antigenen die binden met deze DCs krijgen geen negatief signaal, maar een tweede
positief signaal, waardoor ze niet geëlimineerd worden, maar differentiëren naar Tregs en de
thymus verlaten. Dit zijn de natuurlijke Foxp3+Tregs.

44
Fig. 16. (26) Centrale tolerantie in de thymus. T-cellen ondergaan eerst positieve selectie: T-cellen met een TCR
die in staat is te binden met MHC krijgen het signaal om verder te matureren. Vervolgens ondergaan de T-cellen
negatieve selectie: T-cellen met een TCR die een hoge affiniteit vertoont voor auto-antigenen krijgen het signaal
om in apoptose te gaan. Wanneer echter deze T-cellen geconfronteerd worden met DCs die geconditioneerd
werden met TSLP krijgen ze het signaal te differentiëren naar Tregs. T-cellen met een TCR die een lage affiniteit
vertoont voor auto-antigenen krijgen het signaal verder te matureren en de thymus te verlaten. Dit zijn de naïeve
T-cellen.
Helaas zijn niet alle mogelijke auto-antigenen aanwezig in de thymus. Dit betekent dat
T-cellen met een TCR die een hoge affiniteit vertoont voor auto-antigenen die niet aanwezig
zijn in de thymus dus niet uitgeselecteerd worden. Deze kunnen de thymus verlaten en de
periferie bereiken. Daarom zijn er in de periferie ook mechanismen ingebouwd om de
activiteit van deze auto-reactieve T-cellen te voorkomen. Dit is het zogenaamde mechanisme
van perifere tolerantie. DCs kunnen met de auto-antigenen in contact komen via de opname
van apoptotische DCs die auto-antigenen bevatten. Deze opname leidt tot de inhibitie van
maturatie in de DC en de inductie van een tolerant phenotype.
Deze immature DCs brengen geen co-stimulatoire signalen tot expressie. De opname van
apoptotische DCs leidt tot de inductie van TGF-β, IDO en PD-1L in de DC. De DCs migreren
vervolgens naar de MLNs. Daar bieden ze de auto-antigenen aan aan de aanwezige T-cellen
zonder co-stimulatoir signaal. Ter hoogte van de MLNs hebben deze DCs twee effecten. Ten
eerste stimuleren ze de aanwezige auto-reactieve T-cellen met behulp van
PD-1L, waardoor deze anergisch worden of in apoptose gaan. Ten tweede induceren ze
tolerantie in de aanwezige T-cellen met behulp van hun productie van TGF-β en IDO. Deze
worden zo Foxp3+Tregs. Deze Tregs verlaten de MLNs en migreren naar de darm waar ze
vervolgens de auto-reactieve T-cellen blokkeren.

45
Fig. 17. (46) Perifere tolerantie. Via de opname van apoptotische DCs die auto-antigenen bevatten, kunnen de
DCs in contact komen met auto-antigenen. Deze opname blokkeert de maturatie in de DCs, waardoor deze een
tolerante functie gaan vervullen. Deze opname induceert ook de productie van TGF-β, IDO en PD-1L in de DCs.
De DCs migreren vervolgens naar de MLNs. Daar induceren ze anergie in de T-cellen en apoptose in de
T-cellen via PD-1L. Via hun productie van TGF-β en IDO induceren ze Foxp3+Tregs. De Tregs verlaten
vervolgens de MLNs en migreren naar de darm waar ze auto-reactieve T-cellen kunnen blokkeren.
3.5.2 Tolerantie
Het is belangrijk dat het immuunsysteem tolerantie vertoont tegenover commensale
bacteriën en voedingsantigenen. Ook hier is de functie van DCs van cruciaal. DCs induceren
via vele mechanismen verschillende soorten Tregs. Deze Tregs induceren vervolgens anergie
in T-cellen. De DCs kunnen ook via rechtstreeks contact met de T-cellen anergie induceren.
Dit gebeurt via PD-1L.
3.5.3 Indijken van inflammatie
Ten slotte is het van belang dat een inflammatoire respons geen al te grote proporties
aanneemt en tijdig wordt gestopt voor het kan leiden tot schade. De inflammatoire respons
moet dus tijdig worden ingedijkt.
CTLA-4 inhibeert de T-cel activiteit doordat het een hogere affiniteit heeft voor CD28
dan CD86 en CD80, die een rol spelen in inflammatoire responsen. Op die manier kan de
inflammatoire respons tijdig gestopt worden.
De opgedreven inductie van IDO door IFN-γ gebeurt enkel in inflammatoire omstandigheden.
Dit mechanisme dient om hyper-inflammatie te vermijden.

46
4. Invloed van de omgeving
Doorheen de voorbije hoofdstukken werd duidelijk dat DCs niet de enige cellen zijn die een
rol vervullen in het behoud van homeostase. In de verwezelijking van homeostase kunnen we
verschillende hoofdrolspelers onderscheiden. Zij staan voortdurend met elkaar in contact en
oefenen voortdurend een invloed uit op elkaar. Het gaat om de DCs, de Tregs, het (intestinaal)
epitheel en het microbioom.
4.1 IECs
De taak van IECs bestaat vooral uit het onderscheid maken tussen schadelijke en
onschadelijke antigenen om zo de DCs op gepaste wijze te conditioneren.
4.2 Tregs
Tregs zijn in staat het contact tussen DCs en T-cellen te blokkeren, waardoor de
T-cellen niet meer geactiveerd kunnen worden door DCs en dus geen immuunresponsen
kunnen uitvoeren. (76) Tregs kunnen tevens het phenotype van DCs beïnvloeden.
Foxp3+Tregs zijn in staat IL-27 te induceren in DCs. (53, 71) Ze zijn ook in staat apoptose te
induceren bij de DCs. (42) Tregs kunnen ook de productie van IDO induceren bij de DCs.
(48)
4.3 Microbioom
Het microbioom speelt ook een rol in homeostase. Dit gebeurt door de beïnvloeding
van IECs en DCs. Een zekere activering van de TLRs door het microbioom is vereist om
homeostase te behouden.
Het microbioom speelt een rol in verschillende processen.
De activering van sommige TLRs door het microbioom leidt tot de productie van
anti-microbiële factoren. (12, 22) Dit verloopt via MyD88 dependente pathways. Op die
manier worden er defensines en anti-microbiële factoren geïnduceerd. Deze factoren
inhiberen de groei van bacteriën, zowel pathogene als commensale, in de crypten van de
dunne darm. Eén van de anti-microbiële factoren zijn de β-defensines. Daarnaast hebben zij
ook nog andere functies. β-defensines hebben een chemo-attractante invloed op immature
DCs door hun interactie met CCR6. β-defensines-3 hebben een invloed op de functie van DCs
doordat ze binden met TLR1/2 op de DCs.
TLRs hebben ook een invloed op de opname van antigenen door DCs. (11)
Ten eerste secreteren de IECs uit het FAE chemokines die DCs aantrekken naar de PP. Ten
tweede zorgt de herkenning van micro-organismen door TLRs ervoor dat de antigenopname
en antigenpresentatie door de DCs bevorderd wordt.

47
Zoals eerder vermeld kunnen de IECs het phenotype van de DCs beïnvloeden door de
secretie van bepaalde cytokines die een invloed hebben op de DCs bv. TSLP, TGF-β en RA.
Het microbioom beïnvloedt dit proces. (22, 93) Het microbioom heeft dus een onrechtstreekse
invloed op de DCs via het induceren van de productie en secretie van bepaalde factoren in
IECs die de DCs conditioneren. IECs produceren IL-8, TSLP en TGF-β onder invloed van
zowel gram+ als gram- commensale bacteriën. (19) De productie van IL-8 en TSLP is het
hoogst wanneer IECs werden gestimuleerd door gram- bacteriën. De grootte van het effect
van TSLP en TGF-β is afhankelijk van het soort commensale bacterie waarmee de DCs in
contact is gekomen. (19)
Interactie tussen IECs en het microbioom verhindert dat een inflammatoire respons
optreedt die normaal optreedt wanneer IECs in contact komen met bacteriën. (93)

48
IV. Discussie
1. Samenvatting
Het immuunsysteem ter hoogte van de darm komt voortdurend in contact met
antigenen afkomstig van commensale bacteriën en voedsel. Het is van belang dat het
immuunsysteem niet reageert op deze antigenen en dus geen inflammatoire respons induceert
tegen deze antigenen maw. het immuunsysteem moet tolerant zijn ten opzichte van deze
antigenen. Anderzijds mag het immuunsysteem niet volledig onderdrukt zijn, het moet
immers nog steeds in staat zijn te reageren, wanneer het in contact komt met antigenen die
wel pathogeen zijn. Daarom moet er een evenwicht gevonden worden in de mate waarop het
immuunsysteem reageert. Dit heet homeostase. Homeostase uit zich in verschillende
gebieden: homeostase moet auto-immuniteit voorkomen, ervoor zorgen dat een
immuunrespons tijdig wordt ingedijkt, zodat deze niet escaleert en leidt tot schade en tenslotte
moet homeostase ervoor zorgen dat het lichaam tolerantie vertoont tegenover commensale
bacteriën en voedingsantigenen, maar toch steeds paraat staat om snel en adequaat te reageren
wanneer het om pathogene antigenen gaat.
In dit proces spelen een aantal cellen een grote rol: DCs, IECs, het microbioom en
Tregs. DCs spelen een centrale rol en treden op als coördinator in het proces. Deze cellen
staan niet los van elkaar, maar beïnvloeden elkaar voortdurend. Ze communiceren met elkaar
via rechtstreeks contact of via de secretie van cytokines. Een aantal molecules spelen een
voorname rol in de functies van deze cellen. Het gaat om IL-10, TGF-β en RA.
DCs bezitten twee belangrijke eigenschappen die ze gebruiken om homeostase ter
hoogte van de darm te realiseren. Zo zijn ze in staat Tregs en IgA producerende plasmacellen
te induceren. Hiermee kunnen ze homeostase realiseren op drie gebieden: het uitschakelen
van auto-reactieve T-cellen, het bewerkstelligen van tolerantie ten opzichte van onschadelijke
antigenen en het tijdig indijken van inflammatoire reacties.
Een algemeen werkingsmechanisme dat regelmatig terugkeert is het volgende: Een
cytokine uit de omgeving bv. afkomstig van IECs heeft een invloed op een DC: het induceert
namelijk een tolerant phenotype in de DC. Deze tolerante DC produceert vervolgens zelf
bepaalde cytokines die een invloed hebben op naïeve T-cellen: het cytokine afkomstig van de
DC converteert de naïeve T-cel naar een Treg. Deze Treg produceert vervolgens zelf bepaalde
cytokines die een invloed hebben op effector T-cellen: het cytokine onderdrukt de effector
T-cel.

49
Volgende figuren geven een overzicht van bepaalde DCs. Ze vertellen ons hoe de DCs
geconditioneerd worden, welke molecules ze vervolgens produceren en secreteren en welke
functie ze uitvoeren.
Apoptotische cellen TGF-β /RA IDO stimulatoren PD-1
TGF-β/RA TGF-β/RA
CD103+DCs IDO geen IL-6
PD-1L
Foxp3+Tregs suppressie van T-cellen IgA Gut-homing
Fig. 18. Overzicht DCs. CD103+DCs worden geconditioneerd door apoptotische cellen, TGF-β, RA, signalen
die de productie van IDO induceren in de cel en PD-1. Deze DCs produceren vervolgens TGF-β, RA en IDO.
Deze productie stelt hen in staat tot de secretie van TGF-β en RA en het tot expressie brengen van PD-1L. Ze
onderdrukken tevens de secretie van IL-6. Deze DCs kunnen zo Foxp3+Tregs induceren, T-cellen onderdrukken,
IgA producerende plasmacellen induceren en cellen laten migreren naar de darm.

50
IL-10 IL-27 stimulatoren VIP PD-1
IL-27
IL-10
DCs ICOSL
TGF-β
TR1s suppressie IgA
van T-cellen
Fig. 19. Overzicht DCs. DCs die geconditioneerd worden door IL-10, signalen die de productie van IL-27
stimuleren, VIP en PD-1 produceren en secreteren IL-27, IL-10 en TGF-β en brengen ICOSL tot expressie op
hun membraan. Via de secretie van deze molecules zijn ze in staat TR1s te induceren, T-cellen te onderdrukken
en IgA producerende plasmacellen te induceren.

51
TSLP
DCs
Auto-reactieve Foxp3+Tregs IgA T-cellen elimineren
Fig. 20. Overzicht DCs. DCs die geconditioneerd werden met TSLP zijn in staat auto-reactieve T-cellen te
elimineren, Foxp3+Tregs te induceren en IgA producerende plasmacellen te induceren.
2. Onderzoek
Het onderzoek naar immunologie en homeostase in de darm staat momenteel nog in de
kinderschoenen. Wetenschappers zijn de laatste jaren meer en meer overtuigd van het belang
van kennis over immunologie ter hoogte van de darm en meer bepaald welke invloed een
defecte immuunregulatie speelt in allerlei inflammatoire aandoeningen. Hierdoor kent het
onderzoek een enorme boost, waardoor het aantal artikels rond dit onderwerp nog elke dag
toeneemt.
Hoewel we momenteel regelmatig nieuwe zaken ontdekken, blijven er nog steeds heel
wat hiaten over in de kennis. Vaak kent men het algemene werkingsmechanisme, maar kent
men niet de precieze details van het mechanisme zoals bv. de pathways die een rol spelen.
Nu is het vooral belangrijk om de details te onderzoeken. Er is dus zeker nog ruimte voor
verder onderzoek. Vooral de rol van het microbioom in homeostase vergt nog heel wat
onderzoek. Zo ook de precieze manier hoe Tregs hun immuunregulerende functie uitoefenen.

52
Veel onderzoeken zijn uitgevoerd op murine cellen en niet op humane cellen of
uitgevoerd in vitro en niet in vivo. Daarom moet men voorzichtig zijn bij het extrapoleren van
deze resultaten naar de mens. Er moet dus meer onderzoek komen in vivo en op humane
cellen. Aangezien dit praktisch niet altijd mogelijk is, staan de onderzoekers in de toekomst
voor een uitdaging.
Tussen de verschillende artikels zijn er weinig tegenstrijdigheden te bemerken. Ze
vermelden meestal dezelfde resultaten.
3. Therapie
De reden waarom dit onderzoek naar de processen die een rol spelen bij homeostase
zo belangrijk is, is het feit dat een betere kennis over de fysiologie leidt tot een beter begrip
van de fysiopathogenese en hoe beter we dit begrijpen, hoe beter we nieuwe en verbeterde
therapieën kunnen creëren.
Vanuit de kennis die er momenteel voorhanden is over homeostase kunnen er een
aantal mogelijke therapeutische strategieën voorgesteld worden.
Het induceren van tolerante DCs door het inbrengen van bepaalde molecules is een mogelijke
optie. Dit zou kunnen via een aantal factoren die een rol spelen in de inductie van tolerante
DCs zoals IL-10, TGF-β en TSLP. Hiernaar zijn reeds onderzoek en experimenten
uitgevoerd. Het inbrengen van IL-10 via een lactobacillus bacterie leek veelbelovend bij
murine cellen, maar had niet het verhoopte resultaat bij humane cellen.
Het induceren van tolerante DCs in het labo en deze vervolgens in te brengen is ook een
mogelijkheid. Eén onderzoeksteam is er reeds in geslaagd tolerogene DCs op te wekken in het
labo dmv. VitD3 en Dexomathesone. (94) Deze DCs zijn zeer effectief in vitro. VitD3 en
Dexamethasone houden de DCs in een immature status en zoals eerder vermeld, fungeren
immature DCs als tolerante DCs. Tolerante DCs die geïnduceerd zijn met VitD3 en
Dexomathesone produceren grote hoeveelheden IL-10, een molecule dat een belangrijke rol
speelt in de tolerante functie van DCs. DCs die geïnduceerd zijn met VitD3 zijn ook in staat
naïeve T-cellen te converteren naar Tregs. Ze kunnen ook apoptose induceren in
effector T-cellen. Ze hebben ook een invloed op auto-reactieve cellen: ze veranderen namelijk
het cytokine antwoord van de auto-reactieve T-cellen. Het is nog niet onderzocht of deze DCs
ook effectief functioneren bij de mens.
Het onderzoek naar homeostase heeft reeds vele zaken kunnen ophelderen. Tegelijkertijd
komt men hierbij tot het besef dat men nog vele aspecten niet kent en dat verder onderzoek
noodzakelijk blijft.

53
V. Referentielijst
1. Wetenschappelijke artikels
1. Blanco P, Palucka AK, Pascual V, Banchereau J. Dendritic cells and cytokines in human inflammatory
and autoimmune diseases. Cytokine & growth factor reviews. 2008;19(1):41-52. Epub 2008/02/09.
2. Johansson C, Kelsall BL. Phenotype and function of intestinal dendritic cells. Semin Immunol. 2005
Aug;17(4):284-94.
3. Izcue A, Coombes JL, Powrie F. Regulatory T cells suppress systemic and mucosal immune activation
to control intestinal inflammation. Immunol Rev. 2006 Aug;212:256-71.
4. Zhao X, Sato A, Dela Cruz CS, Linehan M, Luegering A, Kucharzik T, et al. CCL9 is secreted by the
follicle-associated epithelium and recruits dome region Peyer's patch CD11b+ dendritic cells. Journal of
immunology (Baltimore, Md : 1950). 2003 Sep 15;171(6):2797-803. PubMed PMID: 12960300. Epub
2003/09/10. eng.
5. Coombes JL, Siddiqui KR, Arancibia-Carcamo CV, Hall J, Sun CM, Belkaid Y, et al. A functionally
specialized population of mucosal CD103+ DCs induces Foxp3+ regulatory T cells via a TGF-beta and retinoic
acid-dependent mechanism. The Journal of experimental medicine. 2007 Aug 6;204(8):1757-64. PubMed PMID:
17620361. Pubmed Central PMCID: PMC2118683. Epub 2007/07/11. eng.
6. Kelsall BL, Leon F. Involvement of intestinal dendritic cells in oral tolerance, immunity to pathogens,
and inflammatory bowel disease. Immunological reviews. 2005 Aug;206:132-48. PubMed PMID: 16048546.
Epub 2005/07/29. eng.
7. Veldhoen M, Moncrieffe H, Hocking RJ, Atkins CJ, Stockinger B. Modulation of dendritic cell function
by naive and regulatory CD4+ T cells. Journal of immunology (Baltimore, Md : 1950). 2006 May
15;176(10):6202-10. PubMed PMID: 16670330. Epub 2006/05/04. eng.
8. Ng SC, Kamm MA, Stagg AJ, Knight SC. Intestinal dendritic cells: their role in bacterial recognition,
lymphocyte homing, and intestinal inflammation. Inflammatory bowel diseases. 2010 Oct;16(10):1787-807.
PubMed PMID: 20222140. Epub 2010/03/12. eng.
9. Lelouard H, Fallet M, de Bovis B, Méresse S, Gorvel JP. Peyer's patch dendritic cells sample antigens
by extending dendrites through M cell-specific transcellular pores. Gastroenterology. 2012 Mar;142(3):592-
601.e3. doi: 10.1053/j.gastro.2011.11.039. Epub 2011 Dec 8.
10. Rescigno M, Urbano M, Valzasina B, Francolini M, Rotta G, Bonasio R, et al. Dendritic cells express
tight junction proteins and penetrate gut epithelial monolayers to sample bacteria. Nature immunology. 2001
Apr;2(4):361-7. PubMed PMID: 11276208. Epub 2001/03/29. eng.
11. Chieppa M, Rescigno M, Huang AY, Germain RN. Dynamic imaging of dendritic cell extension into
the small bowel lumen in response to epithelial cell TLR engagement. The Journal of experimental medicine.
2006 Dec 25;203(13):2841-52. PubMed PMID: 17145958. Pubmed Central PMCID: PMC2118178. Epub
2006/12/06. eng.
12. Kelly D, Mulder IE. Microbiome and immunological interactions. Nutrition reviews. 2012 Aug;70
Suppl 1:S18-30. PubMed PMID: 22861803. Epub 2012/08/17. eng.
13. Iliev ID, Matteoli G, Rescigno M. The yin and yang of intestinal epithelial cells in controlling dendritic
cell function. The Journal of experimental medicine. 2007 Oct 1;204(10):2253-7. PubMed PMID: 17893197.
Pubmed Central PMCID: PMC2118441. Epub 2007/09/26. eng.
14. Arumugam M, Raes J, Pelletier E, Le Paslier D, Yamada T, Mende DR, et al. Enterotypes of the human
gut microbiome. Nature. 2011 May 12;473(7346):174-80. PubMed PMID: 21508958. Epub 2011/04/22. eng.
15. Qin J, Li Y, Cai Z, Li S, Zhu J, Zhang F, et al. A metagenome-wide association study of gut microbiota
in type 2 diabetes. Nature. 2012 Oct 4;490(7418):55-60. PubMed PMID: 23023125. Epub 2012/10/02. eng.
16. Erickson AR, Cantarel BL, Lamendella R, Darzi Y, Mongodin EF, Pan C, et al. Integrated
metagenomics/metaproteomics reveals human host-microbiota signatures of Crohn's disease. PloS one.
2012;7(11):e49138. PubMed PMID: 23209564. Pubmed Central PMCID: PMC3509130. Epub 2012/12/05. eng.
17. Brandt LJ. American Journal of Gastroenterology Lecture: Intestinal microbiota and the role of fecal
microbiota transplant (FMT) in treatment of C. difficile infection. Am J Gastroenterol. 2013 Feb;108(2):177-85.
doi: 10.1038/ajg.2012.450. Epub 2013 Jan 15.
18. Rimoldi M, Chieppa M, Salucci V, Avogadri F, Sonzogni A, Sampietro GM, et al. Intestinal immune
homeostasis is regulated by the crosstalk between epithelial cells and dendritic cells. Nature immunology. 2005
May;6(5):507-14. PubMed PMID: 15821737. Epub 2005/04/12. eng.

54
19. Zeuthen LH, Fink LN, Frokiaer H. Epithelial cells prime the immune response to an array of gut-
derived commensals towards a tolerogenic phenotype through distinct actions of thymic stromal lymphopoietin
and transforming growth factor-beta. Immunology. 2008 Feb;123(2):197-208. PubMed PMID: 17655740.
Pubmed Central PMCID: PMC2433297. Epub 2007/07/28. eng.
20. Iliev ID, Mileti E, Matteoli G, Chieppa M, Rescigno M. Intestinal epithelial cells promote colitis-
protective regulatory T-cell differentiation through dendritic cell conditioning. Mucosal immunology. 2009
Jul;2(4):340-50. PubMed PMID: 19387433. Epub 2009/04/24. eng.
21. Nijhuis LE, Olivier BJ, de Jonge WJ. Neurogenic regulation of dendritic cells in the intestine.
Biochemical pharmacology. 2010 Dec 15;80(12):2002-8. PubMed PMID: 20615391. Epub 2010/07/10. eng.
22. Wells JM, Rossi O, Meijerink M, van Baarlen P. Epithelial crosstalk at the microbiota-mucosal
interface. Proceedings of the National Academy of Sciences of the United States of America. 2011 Mar 15;108
Suppl 1:4607-14. PubMed PMID: 20826446. Pubmed Central PMCID: PMC3063605. Epub 2010/09/10. eng.
23. Lavelle EC, Murphy C, O'Neill LA, Creagh EM. The role of TLRs, NLRs, and RLRs in mucosal innate
immunity and homeostasis. Mucosal immunology. 2010 Jan;3(1):17-28. PubMed PMID: 19890268. Epub
2009/11/06. eng.
24. Wells JM, Loonen LM, Karczewski JM. The role of innate signaling in the homeostasis of tolerance and
immunity in the intestine. International journal of medical microbiology : IJMM. 2010 Jan;300(1):41-8. PubMed
PMID: 19783476. Epub 2009/09/29. eng.
25. Lee J, Mo JH, Katakura K, Alkalay I, Rucker AN, Liu YT, Lee HK, Shen C, Cojocaru G, Shenouda S,
Kagnoff M, Eckmann L, Ben-Neriah Y, Raz E. Maintenance of colonic homeostasis by distinctive apical TLR9
signalling in intestinal epithelial cells. Nat Cell Biol. 2006 Dec;8(12):1327-36. Epub 2006 Nov 26.
26. Liu YJ, Soumelis V, Watanabe N, Ito T, Wang YH, Malefyt Rde W, et al. TSLP: an epithelial cell
cytokine that regulates T cell differentiation by conditioning dendritic cell maturation. Annual review of
immunology. 2007;25:193-219. PubMed PMID: 17129180. Epub 2006/11/30. eng.
27. Iliev ID, Spadoni I, Mileti E, Matteoli G, Sonzogni A, Sampietro GM, et al. Human intestinal epithelial
cells promote the differentiation of tolerogenic dendritic cells. Gut. 2009 Nov;58(11):1481-9. PubMed PMID:
19570762. Epub 2009/07/03. eng.
28. Paidassi H, Acharya M, Zhang A, Mukhopadhyay S, Kwon M, Chow C, et al. Preferential expression of
integrin alphavbeta8 promotes generation of regulatory T cells by mouse CD103+ dendritic cells.
Gastroenterology. 2011 Nov;141(5):1813-20. PubMed PMID: 21745448. Pubmed Central PMCID:
PMC3202651. Epub 2011/07/13. eng.
29. Worthington JJ, Czajkowska BI, Melton AC, Travis MA. Intestinal dendritic cells specialize to activate
transforming growth factor-beta and induce Foxp3+ regulatory T cells via integrin alphavbeta8.
Gastroenterology. 2011 Nov;141(5):1802-12. PubMed PMID: 21723222. Epub 2011/07/05. eng.
30. Siddiqui KR, Powrie F. CD103+ GALT DCs promote Foxp3+ regulatory T cells. Mucosal
immunology. 2008 Nov;1 Suppl 1:S34-8. PubMed PMID: 19079226. Epub 2008/12/23. eng.
31. del Rio ML, Bernhardt G, Rodriguez-Barbosa JI, Forster R. Development and functional specialization
of CD103+ dendritic cells. Immunological reviews. 2010 Mar;234(1):268-81. PubMed PMID: 20193025. Epub
2010/03/03. eng.
32. Agace WW, Persson EK. How vitamin A metabolizing dendritic cells are generated in the gut mucosa.
Trends in immunology. 2012 Jan;33(1):42-8. PubMed PMID: 22079120. Epub 2011/11/15. eng.
33. Wang S, Villablanca EJ, De Calisto J, Gomes DC, Nguyen DD, Mizoguchi E, et al. MyD88-dependent
TLR1/2 signals educate dendritic cells with gut-specific imprinting properties. Journal of immunology
(Baltimore, Md : 1950). 2011 Jul 1;187(1):141-50. PubMed PMID: 21646294. Pubmed Central PMCID:
PMC3119749. Epub 2011/06/08. eng.
34. Jaensson-Gyllenback E, Kotarsky K, Zapata F, Persson EK, Gundersen TE, Blomhoff R, et al. Bile
retinoids imprint intestinal CD103+ dendritic cells with the ability to generate gut-tropic T cells. Mucosal
immunology. 2011 Jul;4(4):438-47. PubMed PMID: 21289617. Pubmed Central PMCID: PMC3130189. Epub
2011/02/04. eng.
35. Yokota A, Takeuchi H, Maeda N, Ohoka Y, Kato C, Song SY, et al. GM-CSF and IL-4 synergistically
trigger dendritic cells to acquire retinoic acid-producing capacity. International immunology. 2009
Apr;21(4):361-77. PubMed PMID: 19190084. Pubmed Central PMCID: PMC2660862. Epub 2009/02/05. eng.
36. Iwata M. Retinoic acid production by intestinal dendritic cells and its role in T-cell trafficking.
Seminars in immunology. 2009 Feb;21(1):8-13. PubMed PMID: 18849172. Epub 2008/10/14. eng.
37. Delgado M, Reduta A, Sharma V, Ganea D. VIP/PACAP oppositely affects immature and mature
dendritic cell expression of CD80/CD86 and the stimulatory activity for CD4(+) T cells. Journal of leukocyte
biology. 2004 Jun;75(6):1122-30. PubMed PMID: 15020654. Epub 2004/03/17. eng.
38. Delgado M, Gonzalez-Rey E, Ganea D. The neuropeptide vasoactive intestinal peptide generates
tolerogenic dendritic cells. Journal of immunology (Baltimore, Md : 1950). 2005 Dec 1;175(11):7311-24.
PubMed PMID: 16301637. Epub 2005/11/23. eng.

55
39. Gonzalez-Rey E, Chorny A, Fernandez-Martin A, Ganea D, Delgado M. Vasoactive intestinal peptide
generates human tolerogenic dendritic cells that induce CD4 and CD8 regulatory T cells. Blood. 2006 May
1;107(9):3632-8. PubMed PMID: 16397128. Pubmed Central PMCID: PMC1895772. Epub 2006/01/07. eng.
40. Delgado M, Gonzalez-Rey E, Ganea D. Vasoactive intestinal peptide: the dendritic cell --> regulatory T
cell axis. Annals of the New York Academy of Sciences. 2006 Jul;1070:233-8. PubMed PMID: 16888172. Epub
2006/08/05. eng.
41. Weng Y, Sun J, Wu Q, Pan J. Regulatory effects of vasoactive intestinal peptide on the migration of
mature dendritic cells. Journal of neuroimmunology. 2007 Jan;182(1-2):48-54. PubMed PMID: 17081624. Epub
2006/11/04. eng.
42. Kushwah R, Hu J. Dendritic cell apoptosis: regulation of tolerance versus immunity. Journal of
immunology (Baltimore, Md : 1950). 2010 Jul 15;185(2):795-802. PubMed PMID: 20601611. Epub 2010/07/06.
eng.
43. Kushwah R, Wu J, Oliver JR, Jiang G, Zhang J, Siminovitch KA, et al. Uptake of apoptotic DC
converts immature DC into tolerogenic DC that induce differentiation of Foxp3+ Treg. European journal of
immunology. 2010 Apr;40(4):1022-35. PubMed PMID: 20101618. Epub 2010/01/27. eng.
44. Zhong K, Song W, Wang Q, Wang C, Liu X, Chen D, et al. Murine myeloid dendritic cells that
phagocytose apoptotic T cells inhibit the immune response via NO. PloS one. 2012;7(11):e49378. PubMed
PMID: 23166651. Pubmed Central PMCID: PMC3499560. Epub 2012/11/21. eng.
45. Williams CA, Harry RA, McLeod JD. Apoptotic cells induce dendritic cell-mediated suppression via
interferon-gamma-induced IDO. Immunology. 2008 May;124(1):89-101. PubMed PMID: 18067553. Pubmed
Central PMCID: PMC2434374. Epub 2007/12/11. eng.
46. Mahnke K, Johnson TS, Ring S, Enk AH. Tolerogenic dendritic cells and regulatory T cells: a two-way
relationship. Journal of dermatological science. 2007 Jun;46(3):159-67. PubMed PMID: 17428639. Epub
2007/04/13. eng.
47. Ravishankar B, Liu H, Shinde R, Chandler P, Baban B, Tanaka M, et al. Tolerance to apoptotic cells is
regulated by indoleamine 2,3-dioxygenase. Proceedings of the National Academy of Sciences of the United
States of America. 2012 Mar 6;109(10):3909-14. PubMed PMID: 22355111. Pubmed Central PMCID:
PMC3309765. Epub 2012/02/23. eng.
48. Pallotta MT, Orabona C, Volpi C, Vacca C, Belladonna ML, Bianchi R, et al. Indoleamine 2,3-
dioxygenase is a signaling protein in long-term tolerance by dendritic cells. Nature immunology. 2011
Sep;12(9):870-8. PubMed PMID: 21804557. Epub 2011/08/02. eng.
49. Moseman EA, Liang X, Dawson AJ, Panoskaltsis-Mortari A, Krieg AM, Liu YJ, et al. Human
plasmacytoid dendritic cells activated by CpG oligodeoxynucleotides induce the generation of CD4+CD25+
regulatory T cells. Journal of immunology (Baltimore, Md : 1950). 2004 Oct 1;173(7):4433-42. PubMed PMID:
15383574. Epub 2004/09/24. eng.
50. Kushwah R, Hu J. Role of dendritic cells in the induction of regulatory T cells. Cell & bioscience.
2011;1(1):20. PubMed PMID: 21711933. Pubmed Central PMCID: PMC3125210. Epub 2011/06/30. eng.
51. Matteoli G, Mazzini E, Iliev ID, Mileti E, Fallarino F, Puccetti P, et al. Gut CD103+ dendritic cells
express indoleamine 2,3-dioxygenase which influences T regulatory/T effector cell balance and oral tolerance
induction. Gut. 2010 May;59(5):595-604. PubMed PMID: 20427394. Epub 2010/04/30. eng.
52. Kryczek I, Wei S, Zou L, Zhu G, Mottram P, Xu H, et al. Cutting edge: induction of B7-H4 on APCs
through IL-10: novel suppressive mode for regulatory T cells. Journal of immunology (Baltimore, Md : 1950).
2006 Jul 1;177(1):40-4. PubMed PMID: 16785496. Epub 2006/06/21. eng.
53. Awasthi A, Carrier Y, Peron JP, Bettelli E, Kamanaka M, Flavell RA, et al. A dominant function for
interleukin 27 in generating interleukin 10-producing anti-inflammatory T cells. Nature immunology. 2007
Dec;8(12):1380-9. PubMed PMID: 17994022. Epub 2007/11/13. eng.
54. Levings MK, Gregori S, Tresoldi E, Cazzaniga S, Bonini C, Roncarolo MG. Differentiation of Tr1 cells
by immature dendritic cells requires IL-10 but not CD25+CD4+ Tr cells. Blood. 2005 Feb 1;105(3):1162-9.
PubMed PMID: 15479730. Epub 2004/10/14. eng.
55. Gregori S, Tomasoni D, Pacciani V, Scirpoli M, Battaglia M, Magnani CF, et al. Differentiation of type
1 T regulatory cells (Tr1) by tolerogenic DC-10 requires the IL-10-dependent ILT4/HLA-G pathway. Blood.
2010 Aug 12;116(6):935-44. PubMed PMID: 20448110. Epub 2010/05/08. eng.
56. Chui CS, Li D. Role of immunolglobulin-like transcript family receptors and their ligands in suppressor
T-cell-induced dendritic cell tolerization. Human immunology. 2009 Sep;70(9):686-91. PubMed PMID:
19524004. Epub 2009/06/16. eng.
57. Gregori S, Magnani CF, Roncarolo MG. Role of human leukocyte antigen-G in the induction of
adaptive type 1 regulatory T cells. Human immunology. 2009 Dec;70(12):966-9. PubMed PMID: 19664675.
Epub 2009/08/12. eng.

56
58. Wu J, Horuzsko A. Expression and function of immunoglobulin-like transcripts on tolerogenic dendritic
cells. Human immunology. 2009 May;70(5):353-6. PubMed PMID: 19405174. Pubmed Central PMCID:
PMC2775504. Epub 2009/05/01. eng.
59. Liang S, Ristich V, Arase H, Dausset J, Carosella ED, Horuzsko A. Modulation of dendritic cell
differentiation by HLA-G and ILT4 requires the IL-6--STAT3 signaling pathway. Proceedings of the National
Academy of Sciences of the United States of America. 2008 Jun 17;105(24):8357-62. PubMed PMID:
18550825. Pubmed Central PMCID: PMC2448841. Epub 2008/06/14. eng.
60. Iyer SS, Ghaffari AA, Cheng G. Lipopolysaccharide-mediated IL-10 transcriptional regulation requires
sequential induction of type I IFNs and IL-27 in macrophages. Journal of immunology (Baltimore, Md : 1950).
2010 Dec 1;185(11):6599-607. PubMed PMID: 21041726. Epub 2010/11/03. eng.
61. Stumhofer JS, Hunter CA. Advances in understanding the anti-inflammatory properties of IL-27.
Immunology letters. 2008 May 15;117(2):123-30. PubMed PMID: 18378322. Pubmed Central PMCID:
PMC2390685. Epub 2008/04/02. eng.
62. Kuipers H, Muskens F, Willart M, Hijdra D, van Assema FB, Coyle AJ, et al. Contribution of the PD-1
ligands/PD-1 signaling pathway to dendritic cell-mediated CD4+ T cell activation. European journal of
immunology. 2006 Sep;36(9):2472-82. PubMed PMID: 16917960. Epub 2006/08/19. eng.
63. Yamazaki S, Steinman RM. Dendritic cells as controllers of antigen-specific Foxp3+ regulatory T cells.
Journal of dermatological science. 2009 May;54(2):69-75. PubMed PMID: 19286352. Pubmed Central PMCID:
PMC2680740. Epub 2009/03/17. eng.
64. Moore C, Sauma D, Reyes PA, Morales J, Rosemblatt M, Bono MR, et al. Dendritic cells and B cells
cooperate in the generation of CD4(+)CD25(+)FOXP3(+) allogeneic T cells. Transplantation proceedings. 2010
Jan-Feb;42(1):371-5. PubMed PMID: 20172352. Epub 2010/02/23. eng.
65. Sun CM, Hall JA, Blank RB, Bouladoux N, Oukka M, Mora JR, et al. Small intestine lamina propria
dendritic cells promote de novo generation of Foxp3 T reg cells via retinoic acid. The Journal of experimental
medicine. 2007 Aug 6;204(8):1775-85. PubMed PMID: 17620362. Pubmed Central PMCID: PMC2118682.
Epub 2007/07/11. eng.
66. Jaensson E, Uronen-Hansson H, Pabst O, Eksteen B, Tian J, Coombes JL, et al. Small intestinal
CD103+ dendritic cells display unique functional properties that are conserved between mice and humans. The
Journal of experimental medicine. 2008 Sep 1;205(9):2139-49. PubMed PMID: 18710932. Pubmed Central
PMCID: PMC2526207. Epub 2008/08/20. eng.
67. Mezrich JD, Fechner JH, Zhang X, Johnson BP, Burlingham WJ, Bradfield CA. An interaction between
kynurenine and the aryl hydrocarbon receptor can generate regulatory T cells. J Immunol. 2010 Sep
15;185(6):3190-8. doi: 10.4049/jimmunol.0903670. Epub 2010 Aug 18
68. Chen W, Liang X, Peterson AJ, Munn DH, Blazar BR. The indoleamine 2,3-dioxygenase pathway is
essential for human plasmacytoid dendritic cell-induced adaptive T regulatory cell generation. J Immunol. 2008
Oct 15;181(8):5396-404.
69. Francisco LM, Sage PT, Sharpe AH. The PD-1 pathway in tolerance and autoimmunity. Immunological
reviews. 2010 Jul;236:219-42. PubMed PMID: 20636820. Pubmed Central PMCID: PMC2919275. Epub
2010/07/20. eng.
70. Apetoh L, Quintana FJ, Pot C, Joller N, Xiao S, Kumar D, et al. The aryl hydrocarbon receptor interacts
with c-Maf to promote the differentiation of type 1 regulatory T cells induced by IL-27. Nature immunology.
2010 Sep;11(9):854-61. PubMed PMID: 20676095. Pubmed Central PMCID: PMC2940320. Epub 2010/08/03.
eng.
71. Pot C, Apetoh L, Awasthi A, Kuchroo VK. Molecular pathways in the induction of interleukin-27-
driven regulatory type 1 cells. Journal of interferon & cytokine research : the official journal of the International
Society for Interferon and Cytokine Research. 2010 Jun;30(6):381-8. PubMed PMID: 20540648. Pubmed
Central PMCID: PMC2964360. Epub 2010/06/15. eng.
72. Pot C, Jin H, Awasthi A, Liu SM, Lai CY, Madan R, et al. Cutting edge: IL-27 induces the transcription
factor c-Maf, cytokine IL-21, and the costimulatory receptor ICOS that coordinately act together to promote
differentiation of IL-10-producing Tr1 cells. Journal of immunology (Baltimore, Md : 1950). 2009 Jul
15;183(2):797-801. PubMed PMID: 19570826. Pubmed Central PMCID: PMC2768608. Epub 2009/07/03. eng.
73. Stumhofer JS, Silver JS, Laurence A, Porrett PM, Harris TH, Turka LA, et al. Interleukins 27 and 6
induce STAT3-mediated T cell production of interleukin 10. Nature immunology. 2007 Dec;8(12):1363-71.
PubMed PMID: 17994025. Epub 2007/11/13. eng.
74. Bauquet AT, Jin H, Paterson AM, Mitsdoerffer M, Ho IC, Sharpe AH, et al. The costimulatory
molecule ICOS regulates the expression of c-Maf and IL-21 in the development of follicular T helper cells and
TH-17 cells. Nature immunology. 2009 Feb;10(2):167-75. PubMed PMID: 19098919. Pubmed Central PMCID:
PMC2742982. Epub 2008/12/23. eng.

57
75. Fife BT, Bluestone JA. Control of peripheral T-cell tolerance and autoimmunity via the CTLA-4 and
PD-1 pathways. Immunological reviews. 2008 Aug;224:166-82. PubMed PMID: 18759926. Epub 2008/09/02.
eng.
76. Tadokoro CE, Shakhar G, Shen S, Ding Y, Lino AC, Maraver A, et al. Regulatory T cells inhibit stable
contacts between CD4+ T cells and dendritic cells in vivo. The Journal of experimental medicine. 2006 Mar
20;203(3):505-11. PubMed PMID: 16533880. Pubmed Central PMCID: PMC2118249. Epub 2006/03/15. eng.
77. Zhang X, Kedl RM, Xiang J. CD40 ligation converts TGF-beta-secreting tolerogenic CD4-8- dendritic
cells into IL-12-secreting immunogenic ones. Biochemical and biophysical research communications. 2009 Feb
20;379(4):954-8. PubMed PMID: 19135981. Epub 2009/01/13. eng.
78. Cong Y, Wang L, Konrad A, Schoeb T, Elson CO. Curcumin induces the tolerogenic dendritic cell that
promotes differentiation of intestine-protective regulatory T cells. European journal of immunology. 2009
Nov;39(11):3134-46. PubMed PMID: 19839007. Epub 2009/10/20. eng.
79. Murugaiyan G, Mittal A, Lopez-Diego R, Maier LM, Anderson DE, Weiner HL. IL-27 is a key
regulator of IL-10 and IL-17 production by human CD4+ T cells. Journal of immunology (Baltimore, Md :
1950). 2009 Aug 15;183(4):2435-43. PubMed PMID: 19625647. Pubmed Central PMCID: PMC2904948. Epub
2009/07/25. eng.
80. Cerutti A, Rescigno M. The biology of intestinal immunoglobulin A responses. Immunity. 2008
Jun;28(6):740-50. PubMed PMID: 18549797. Pubmed Central PMCID: PMC3057455. Epub 2008/06/14. eng.
81. Mora JR, von Andrian UH. Role of retinoic acid in the imprinting of gut-homing IgA-secreting cells.
Seminars in immunology. 2009 Feb;21(1):28-35. PubMed PMID: 18804386. Pubmed Central PMCID:
PMC2663412. Epub 2008/09/23. eng.
82. Turnbull EL, Yrlid U, Jenkins CD, Macpherson GG. Intestinal dendritic cell subsets: differential effects
of systemic TLR4 stimulation on migratory fate and activation in vivo. Journal of immunology (Baltimore, Md :
1950). 2005 Feb 1;174(3):1374-84. PubMed PMID: 15661895. Epub 2005/01/22. eng.
83. Rol N, Favre L, Benyacoub J, Corthesy B. The role of secretory immunoglobulin A in the natural
sensing of commensal bacteria by mouse Peyer's patch dendritic cells. The Journal of biological chemistry. 2012
Nov 16;287(47):40074-82. PubMed PMID: 23027876. Pubmed Central PMCID: PMC3501041. Epub
2012/10/03. eng.
84. Suzuki K, Fagarasan S. How host-bacterial interactions lead to IgA synthesis in the gut. Trends in
immunology. 2008 Nov;29(11):523-31. PubMed PMID: 18838301. Epub 2008/10/08. eng.
85. Johansson-Lindbom B, Agace WW. Generation of gut-homing T cells and their localization to the small
intestinal mucosa. Immunological reviews. 2007 Feb;215:226-42. PubMed PMID: 17291292. Epub 2007/02/13.
eng.
86. Agace W. Generation of gut-homing T cells and their localization to the small intestinal mucosa.
Immunology letters. 2010 Jan 18;128(1):21-3. PubMed PMID: 19808049. Epub 2009/10/08. eng.
87. Rossetti M, Gregori S, Roncarolo MG. Granulocyte-colony stimulating factor drives the in vitro
differentiation of human dendritic cells that induce anergy in naive T cells. European journal of immunology.
2010 Nov;40(11):3097-106. PubMed PMID: 20957751. Pubmed Central PMCID: PMC2997328. Epub
2010/10/20. eng.
88. Molenaar R, Greuter M, van der Marel AP, Roozendaal R, Martin SF, Edele F, et al. Lymph node
stromal cells support dendritic cell-induced gut-homing of T cells. Journal of immunology (Baltimore, Md :
1950). 2009 Nov 15;183(10):6395-402. PubMed PMID: 19841174. Epub 2009/10/21. eng.
89. Keir ME, Liang SC, Guleria I, Latchman YE, Qipo A, Albacker LA, et al. Tissue expression of PD-L1
mediates peripheral T cell tolerance. The Journal of experimental medicine. 2006 Apr 17;203(4):883-95.
PubMed PMID: 16606670. Pubmed Central PMCID: PMC2118286. Epub 2006/04/12. eng.
90. Keir ME, Francisco LM, Sharpe AH. PD-1 and its ligands in T-cell immunity. Current opinion in
immunology. 2007 Jun;19(3):309-14. PubMed PMID: 17433872. Epub 2007/04/17. eng.
91. Latchman Y, Wood CR, Chernova T, Chaudhary D, Borde M, Chernova I, et al. PD-L2 is a second
ligand for PD-1 and inhibits T cell activation. Nature immunology. 2001 Mar;2(3):261-8. PubMed PMID:
11224527. Epub 2001/02/27. eng.
92. Probst HC, McCoy K, Okazaki T, Honjo T, van den Broek M. Resting dendritic cells induce peripheral
CD8+ T cell tolerance through PD-1 and CTLA-4. Nature immunology. 2005 Mar;6(3):280-6. PubMed PMID:
15685176. Epub 2005/02/03. eng.
93. Rescigno M. Gut commensal flora: tolerance and homeostasis. F1000 biology reports. 2009;1:9.
PubMed PMID: 20948601. Pubmed Central PMCID: PMC2920680. Epub 2009/01/01. eng.
94. Unger WW, Laban S, Kleijwegt FS, van der Slik AR, Roep BO. Induction of Treg by monocyte-derived
DC modulated by vitamin D3 or dexamethasone: differential role for PD-L1. European journal of immunology.
2009 Nov;39(11):3147-59. PubMed PMID: 19688742. Epub 2009/08/19. eng.

58
2. Proefschriften
95. Van Beveren H. Predictieve geneeskunde op basis van genoominformatie: fictie of realiteit? Gent:
UGent; 2010
3. Websites
96. Vakgroep celbiologie en histolgie van de Universiteit van Amsterdam. Celbiologie en histologie.
Internetsite van het AMC 2010-2011. Beschikbaar via:
http://os1.amc.nl/celbiologie/20102011/2.4/dudacol/html/046-6.html. Geraadpleegd op 9 maart 2013.

59

VI. Bijlagen
1. Lijst met afkortingen
AhR: Aryl hydrocarbon-Receptor
AID: activation-induced cytidine deaminase
APRIL: a proliferation-inducing ligand
BAFF: B-cell activating factor
cAMP: Cyclisch adenosinemonofosfaat
CCL: Chemokine Ligand
CCR: Chemokine Receptor
CTLA-4: cytotoxic T-lymphocyte antigen-4
DCs: Dendritische Cellen
FAE: Follikel Geassocieerd Epitheel
Foxp3: Forkhead box p3
GALT: Gut-Associated Lymphoid Tissue
HLA-DR: human leukocyte antigen-DR
HLA-G: human leukocyte antigen-G
ICOS: Inducible T-cell COStimulator
IDO: indoleamine 2,3-dioxygenase
IECs: Intestinale Epitheliale Cellen
IFN: Interferon
Ig: Immunoglobuline
IL: InterLeukine
ILT4: Ig-like transcript 4
iNOS: inducible Nitrogen Oxide
ITIM: immunoreceptor tyrosine-based inhibitory motifs
JAK: Just Another Kinase
JNK-pathway: c-Jun N-terminal kinases
LP: Lamina Propria
LPS: Lipopolysaccharide
ltbp3: latent TGF-β binding protein 3
MadCAM1: mucosal addressin cell adhesion molecule-1
MAMP: Microbe-Associated Molecular Pattern

MAPK: Mitogen-activated protein kinases
M-cellen: Microfold cellen
MHC: Major Histocompatibility Complex
MLN: Mesenterische LympheNodi
MyD88: myeloid differentiation primary response gene 88
NF-κB: nuclear factor kappa-light-chain-enhancer of activated B cells
NLR: Nucleotide Oligomerization Receptor
NO: Nitrogen Oxide
PACAP: Pituitary Adenylate Cyclase-Activating Polypeptide
PD-1/2: programmed death-1/2
PD-1/2L: programmed death-1/2-ligand
PKA: Proteïne Kinase A
plat: tissue plasminogen activator
PP: Peyerse Platen
PPAR-γ: peroxisome proliferator-activated receptor-gamma
PPR: Patern Recognition Receptor
RA: Retinoic Acid
RADLH: Retinaldehyde dehydrogenase
RLR: Retinoic acid inducible gene I-Like Receptor
RORC: RAR-related orphan receptor C
SED: SubEpitheliale Dome
TCR: T-Cel Receptor
TGF-β: Transforming Growth Factor Bèta
tgfb2: transforming growth factor, bèta 2
Th1: T-helpercel1
Th2: T-helpercel2
Th17: T-helpercel17
TNF: Tumor Necrosis Factor
TLR: Toll-Like Receptor
Tregs: Regulatoire T-cellen
TSLP: Thymic Stromal LymphoPoietin
VIP: Vasoactive Intestinal Peptide

2. Overzicht figuren en tabellen
2.1 Figuren
Fig. 1. (1) Soorten DCs.
Fig. 2. (96) GALT en intestinaal epitheel.
Fig. 3. (23) Overzicht van de verschillende intracellulaire pathways van de TLRs.
Fig. 4. (32) Inductie van de productie van RA in de DC.
Fig. 5. (36) De metabolisatie van vitamine A tot RA.
Fig. 6. (42) Inductie van apoptose in de DCs.
Fig. 7. (42) Inductie van een tolerant phenotype in DCs via de opname van apoptotische DCs.
Fig.8. (59) Inductie van tolerantie in DCs door HLA-G en ILT4.
Fig. 9. (50) Ontstaan van Foxp3+Tregs.
Fig. 10. (71) Effecten van IL-27.
Fig. 11. (50) Onstaan van Tr1s.
Fig. 12. (46) Beïnvloeding van de DCs door Tregs.
Fig. 13. (80) Ontstaan van IgA producerende plasmacellen op T-cel afhankelijke wijze.
Fig. 14. (80) Ontstaan van IgA producerende plasmacellen op een T-cel onafhankelijke wijze.
Fig. 15. (36) Migratie van Tregs naar de darm.
Fig. 16. (26) Centrale tolerantie in de thymus.
Fig. 17. (46) Perifere tolerantie.
Fig. 18. Overzicht DCs.
Fig. 19. Overzicht DCs.
Fig. 20. Overzicht DCs.
2.2 Tabellen
Tabel 1. (22) Karakteristieken van de verschillende TLRs.