deletion of the phosphoglucose isomerase structural gene makes growth and sporulation glucose...
TRANSCRIPT

M ol Gen Genet (1986) 204:310-316
© Springer-Verlag 1986
Deletion of the phosphoglucose isomerase structural gene makes growth and sporulation glucose dependent in Saccharomyces cerevisiae AndrOs Aguilera Institut fiir Mikrobiologie, Technische Hochschule Darmstadt, Schnittspahnstrasse 10, D-6100 Darmstadt, Federal Republic of Germany
Summary. The structural gene PGH coding for phosphoglu- cose isomerase was replaced by the LEU2 gene in the ge- nome of Saccharomyces cerevisiae. Plasmids carrying the LEU2 gene between genomic regions flanking the PGH gene were constructed and used to transform a PGH/pgil diploid strain. Stable transformants lacking the PGI1 allele were isolated. Southern analysis of their meiotic products showed that haploid strains with a deletion of 1.6 kb within the 2.2 kb PGI1 coding region were viable. Thus, the PGH gene is not essential in yeasts. However, unlike pgil mutants with residual phosphoglucose isomerase activity, no growth was detected in the pgilA haploid strains when fructose was supplied as sole carbon source. The wild-type growth rate could be restored by adding 0.1% glucose to the medi- um. Furthermore, pgil mutants with residual enzymatic ac- tivity grew very slowly on fructose-supplemented media containing up to 2% glucose. Strains carrying the deletion allele, however, failed to grow at glucose concentrations higher than 0.5%. Also the pgilA strains did not grow in glucose as sole carbon source. On the other hand pgilA/ pgi13 diploid strains did not sporulate on the usual acetate medium. This defect could be alleviated by the addition of 0.05% glucose to the sporulation medium. Under these conditions the pgilA mutants sporulated with an efficiency of 25% compared with the wild type. These results suggest that (a) the phosphoglucose isomerase reaction is the only step catalysing the interconversion of glucose-6-P and fruc- tose-6-P, (b) glucose-6-P is essential in yeasts, and (c) the oxidation of glucose-6-P through the glucose-6-P dehy- drogenase reaction is not sufficient to support growth in yeasts.
Key words: Gene replacement - PGI1 deletion - Glucose-6- P - Glucose dependence - Saccharomyces cerevisiae
Introduction
Glucose-6-P and fructose-6-P are interconverted in yeasts and bacteria by the phosphoglucose isomerase reaction (for reviews see Fraenkel and Vinopal 1973; Fraenkel 1982). In yeasts the enzyme is a dimer composed of two identical subunits (Kempe et al. ~974) coded by the gene PGII (Mai- tra and Lobo 1977). The PGH gene has been isolated re- cently and molecular analysis has revealed that there is only one copy per haploid genome (Aguilera and Zimmermann
1986). No evidence has been found for the existence of isoenzymes for phosphoglucose isomerase. Several muta- tions of the PGH gene have been isolated (Maitra 1971; Clifton et al. 1978; Herrera and Pascual 1978; Ciriacy and Breitenbach 1979). All of them show residual phosphoglu- cose isomerase activity.
Glucose-6-P is required for the synthesis of inositol (Eisenberg 1978) necessary for membrane phospholipids (Henry 1982), and for the synthesis of glucans of the cell wall (Ballou 1982). It is also required as substrate for the hexose monophosphate oxidation pathway to generate NADPH (Fraenkel 1982). All these facts suggest that the phosphoglucose isomerase reaction is an essential metabolic step in yeasts, However, in Escherichia coli phosphoglucose isomerase and glucose itself seem to be dispensable because an amber mutant and a phage Mu insertion, presumably leading to complete loss of enzymatic activity, could grow on medium with glucose, mannose or glycerol as sole car- bon sources (Vinopal et al. 1975).
To elucidate the role of phosphoglucose isomerase and glucose-6-P in yeast cells the PGI1 flanking regions were used to construct deletions in which more than 70% of the PGH coding sequence was substituted by the yeast LEU2 sequence. Haploids carrying this deletion had no de- tectable enzymatic activity but were still able to grow under certain conditions. The importance of glucose-6-P in the mitotic as well as in the meiotic cycle of Saccharomyces cerevisiae is discussed.
Materials and methods
Strains. Diploid strain DD1 (MAT~/MATa leu2-3, II2/leu2-3, 112 trpl/trpl ura3-52/ura3-52 PGI1/pgil) was used as recipient for transformation. Strains UTL-7A (MAT~ leu2-3, 112 trpl ura3-52), DMA4 (MATa pgil leu2-3, 112 trpl ura3-52), 83L (MATa spoil ura3 cam cyh2 ade2 his7 hom3) and 84L (MAT~ spoll ura3 canl cyh2 ade2 his7 hom3 tyrl) were used as sources for genomic DNA as well as for constructing isogenic diploids. Strain RR1 of Escherichia cob was used for the preparation of plasmids.
Media and growth conditions. YEP media containing 2% peptone (Difco, Detroit, MI, USA), 1% yeast extract (Ox- oid, Basingstoke, England) supplemented with 2% glucose (YEPD) or 2% fructose (YEPF) as carbon sources, were

311
used as rich media. YEP itself contains trace amounts of glucose. The synthetic medium was based on 1.7 g/1 Difco yeast nitrogen base without amino acids and ammonium sulphate, supplemented with 5 g/1 (NIt4)2SO4, amino acids and bases as required and 2% glucose (SD) or 2% fructose (SF) as carbon sources. Media were solidifed with 15 g/1 agar (Oxoid). Liquid cultures were grown on a shaker at 28 ° C. Classical genetical analysis was performed following published procedures (Sherman et al. 1979).
Preparation of crude extracts and enzyme assays. Crude ex- tracts were prepared as described by Ciriacy and Breiten- bach (1979) using ballotini glass beads (Sigma, St. Louis, MO, USA) for breaking the cells. Phosphoglucose isomer- ase activity was assayed as previously described (Aguilera and Zimmermann 1986) in 50 mM imidazole buffer, pH 7.0, containing 10 mM Mg + +
Nystatin enrichment of pgi- strains. This was basically per- formed as described by Snow (1966). Cells were grown in fructose-supplemented media up to early stationary phase and kept overnight in 1.6% yeast nitrogen base without amino acids and bases. The cell titre was adjusted to 10 v cells/ml in a synthetic glucose medium supplemented with tryptophan and uracil. After 3-4 h nystatin was added at a final concentration of 10 gg/ml and cells were incubated for 90 rain at 28 ° C. Cells were washed three times and permited to grow in YEPF for 2 h, before plating on YEPF media.
Transformation experiments. E. coli was transformed as de- scribed by Cohen et al. (1972), and yeast cells according to Beggs (1978). Sphaeroplasts were prepared using zymo- lyase 20T (Miles GmbH, Frankfurt, FRG) at a final con- centration of 50 gg/ml. Tryptophan and leucine were omit- ted fi'om the transformation media to select for plasmid markers.
Nucleic acid preparations. Yeast chromosomal DNA was prepared according to Ciriacy and Williamson (1981). Plas- mid DNA was prepared from yeast transformants accord- ing to Nasmyth and Reed (1980) and from E. coli according to Bolivar and Backman (1979) or Birnboim (1983) for small- or large-scale preparations, respectively.
Preparation of DNA fragments and subcloning. DNA restric- tion fragments were electrophoresed in 0.6% agarose gels with 89 mM Tris, 2.5 mM EDTA, 19 mM acetic acid, pH 8.3. Bands of interest were cut out of the gel, equilibrated with 0.3 M sodium acetate, 1 mM EDTA, pH 7.0. DNA fragments were isolated from the gel by the optimized freeze-squeeze method of Tautz and Renz (1983). This DNA was either radioactively labelled and used for hybrid- ization or ligated into vector DNA according to Mertz and Davis (1972), using T4 DNA ligase (Boehringer, Mann- heim, FRG).
DNA electrophoresis and hybridization. DNA electrophore- sis was performed in 0.83% agarose, 89 M Tris, 112 mM H3BO3, 2.5 mM EDTA, pH 8.3. DNA was transferred to nitrocellulose filters as described by Wahl et al. (1979). DNA used as, radioactive probe was labelled by nick trans- lation using c~3sS-dATP (New England Nuclear, Boston, MA, USA) and DNA polymerase I (Bethesda Research
Laboratories, Gaithersburg, MD, USA) as described by Rigby et al. (1977). DNA-DNA hybridizations were per- formed in 0.6 M NaC1, 0.5 M sodium phosphate, pH 6.2, 3 mM EDTA, 1% sarcosyl and 0.1 mg/ml sonicated herring sperm DNA at 65 ° C for 48 h.
Results
Substitution of the chromosomal PGI1 gene by the LEU2 gene
Cloned DNA fragments containing the PGI1 gene (Agui- lera and Zimmermann 1986) were used to delete most of the PGI1 coding sequence at its chromosomal locus in order to characterize the phenotype of a true null mutation. The pgil mutants isolated so far by classical genetic procedures have residual phosphoglucose isomerase activity.
For this purpose plasmids pC and pPGI1-4 carrying the PGH gene and its flanking regions (Aguilera and Zim- mermann 1986) were digested with restriction endonucle- ases BamHI and J(hoI, and with SalI-BglII or SalI-XhoI, respectively. The 8.4 kb BamHI-XhoI fragment carrying the TRP1 gene and 3.6 kb of the flanking region at one side of the PGH gene were isolated from plasmid pC. The 2.16 kb SalI-35hoI fragment carrying the LEU2 marker, and the 6.5 kb SalI-BglII fragment containing the 2 ~m replica- tion origin and 3.0 kb of flanking region from the opposite side of PGH were purified from plasmid pPGI1-4. The three isolated fragments were mixed and ligated with T4 DNA ligase. Plasmids p25 and p34 carrying the three ex- pected fragments were identified in E. coli RR1 transfor- mants (Fig. 1). Both plasmids contain the LEU2 marker inserted between the flanking regions of the PGH gene but in different orientations. The plasmids also carry two repli- cation origins (ARSI and the 2 ~m replicon). Because the 2 ~tm replication origin leads to instability of the chromo- somes upon integration (Falco et al. 1982) it was removed from the plasmids. Partial digestion of both plasmids with Kpnl was performed and fragments lacking the 2.7 kb KpnI fragment carrying the 2 gm origin were religated. Plasmids p25d and p34d were finally prepared from amp r tet s trans- formants of E. coli (Fig. 1).
Diploid strain DDI homozygous for leu2 and trpl and heterozygous for PGI1 was transformed with plasmids p25, p34, p25d and p34d. The rationale was to replace the PGH allele in the chromosome with the LEU2 marker in the plasmids. This requires a double recombination event at both PGI1 flanking regions (one-step; Rothstein 1983) or integration of the plasmids at the PGI1 locus as a primary event and subsequent removal of the PGI1 allele by single recombination of appropriate flanking regions (two-step process). Therefore unstable LEU+TRP + transformants that were PGI +, due to the PGH allele in one of the chro- mosomes were used to select for strains with the stable phenotype LEU+trp-pg i - . They should carry the LEU2 gene instead of the PGI1 allele at the PGI1 chromosomal locus.
Because there is no way of selecting such a phenotype the method of nystatin enrichment (Snow 1966) was used. After nystatin enrichment cells were plated on YEP fructose and replica plated on YEP glucose. Colonies not growing on the latter medium were selected and the LEU, TRP and URA phenotypes were established. From a total of 32,787 colonies screened, 582 pgi- strains were selected,

312
..ARSI • /~-~=~qL TRPI
or~ARSI ~ ~ K s ~ ~mp- ,~ TRP~ a s
'x ho, oo.,
/sotv'Bgm X / SaLI/XhoI ×
I LEU2 S AR~ ARSl
X [ - ~ //"" 'K Tg DNA Ligase
PGI__11\ LEU2 .LEU2 S
,,KprlI pQrtiol ~- T~ DNA Ligose
x LEO2
Fig. 1. Construction of plasmids carrying the LEU2 gene inserted between both flanking regions of the PGH gene. The yeast DNA sequences ARS1, TRP1, LEU2 and the 2 gm replication origin are shown as closed boxes. The PGI1 coding region is indicated by stippled boxes and its flanking regions by broad open boxes. Single lines represent pBR322 sequences and the thin open boxes represent flanking sequences from the TRP1 or LEU2 genes. Arrows indicate the restriction targets used for subsequent subcloning experiments and dots represent the ligation points of different fragments used to construct the plasmids. Only the very important restriction sites are symbolized. These are as follows: B, BglII ; Ba, BamHI; K, KpnI; S, SalI; X, XhoI. See text for further details
Table 1. Genetic linkage between the phenotypes LEU + and small colony size in the stable transformants D-25d2, D-25d5 and D- 34d2
Strain Complete Number of complete tetrads tetrads with segregation
2 ss LEU ÷ :2 WT leu- others
D-25d2 9 9 0 D25-d5 7 7 0 D-34d2 12 12 0
WT, wild-type colony size; ss, small colony size
most of them arising from mitotic recombination. Only 12 stable LEU+trp-ura - strains were recovered, three from transformants with plasmid p34d, nine with p25d and none with plasmids p25 and p34 containing the 2 gm replication origin.
Tetrad analysis of these strains revealed a 4:0 segrega- tion on YEP fructose medium for the pgi- phenotype, but a 2:2 segregation for colony size (two colonies always ap- pearing very small). All segregants leading to small-size col- onies were LEU + and all of the segregants with normal-size colonies were leu- (Table l). This result is what would be expected for a substitution of the PGII allele by the LEU2 gene in the diploid strain, if P G H were a non-essential gene.
Southern blot analysis was performed to show at the molecular level that the P G H gene had been substituted. Chromosomal DNA was prepared from each segregant of
one complete tetrad of diploids D25d5 and D34d2, from the haploid strains, DMA4, UTL-7A and from the diploid strain DDI. The radioactive probes used were: the 2.16 kb SaII-J(hoI fragment of plasmid pPGI1M containing the LEU2 gene, the 1.55 kb HindIII-BglII fragment of plasmid pC carrying part of the P G H gene that is thought to be deleted in the mutants, and plasmid pBd carrying the TRP1 and P G H markers (Aguilera and Zimmermann 1986). Fig- ure 2 shows that the PGH band is missing in two spores of each tetrad (the 15 kb BamHI and the 2.05 kb EcoRI band). Then, a new BamHI fragment of 15 kb appears be- sides the wild-type 9 kb fragment corresponding to the LEU2 marker. When DNA was digested with EcoRI which cuts in the LEU2 gene, two new bands of 3.45 and 1.7 kb (spores from D34d) and of 3.85 and 1.3 kb (spores from D25d) appeared in addition to the two original LEU2 frag- ments (3 and 7 kb). These new bands correspond to the two EcoRI LEU2-carrying fragments present in plasmids p34d and p25d, respectively. The TRP1 band appears as a 1.45 kb EcoRI fragment as in the wild-type strains. These results proved that PGI1 had been successfully replaced by LEU2 in the mutants. The additional weakly labelled bands appearing in the Southern blots correspond to the repetitive 5 sequence contained in probes carrying the LEU2 gene and/or to 2 gm contaminant sequences from the plas- mids from which the probe carrying part of the P G H gene was isolated.
Finally the enzymatic activity was measured in the se- gregants of the tetrads. In the spores carrying the pgi lA the phosphoglucose isomerase activity was below the level of detection (< 0.5 mU/mg protein; Table 2).

313
Fig. 2. Southern blot analysis of one complete tetrad (a-d) of stable transformants D-25d5 and D-34d2, the diploid DD1 (PGI1/pgiI) (1), and the haploid strains DMA4 (pgil) (2) and UTL-TA (PGI1) (3). Chromosomal DNA was digested with endonucleases BamHI (left) or EcoRI (right). The DNA sequences used as radioactive probes were: a 2.2 kb SalI-XhoI fragment containing the LEU2 gene (LEU*), a 1.55 kb HindIII-BglII fragment containing part of the PGH gene (PGI*), both isolated from plasmid pPGII-4, and plasmid pBd (Aguilera and Zimmermann 1986) which contains the TRP1 maker and part of the PGH gene (PGI*/TRP*)
T a b l e 2. Phosphoglucose isomerase enzymatic activity of the wild- type strain UTL-7A, the pgi- mutant DMA4 and different spores derived from diploid stable transformants D-25d5 and D-34d2
Strain Genotype mU/mg protein Relative activity"
UTL-7A PGI1 1,499 1.0 DMA4 pgil 29 0,02 25d5-Ia pgil 28 0,02 25d5-Ib pgilA n.d. b - 34d2-Ia pgil 25 0.02 34d2-Ic pgilA n.d. b -
" Wild-type activity taken as 1.0 b Not detectable in 0.5 mg of total protein
Growth and sporulation are glucose-dependent processes
Haplo id strains carrying pgilA failed to grow on synthetic media with fructose as sole carbon source. Growth on a synthetic fructose medium supplemented with increasing amounts of glucose was studied in the wild type, UTL-7A, a pgil mutant (strain D M A 4 ) with 1 % - 2 % of the wild-type
activity and the pgilA mutants described above. Wild- type growth was not affected by increasing glucose concentra- tions (Fig. 3). The pgil mutant grows slower than the wild type at various glucose concentrat ion (Fig. 3A) but is able to reach a s tat ionary phase cell concentrat ion similar to the wild type (Fig. 3 B). However the pgiiA mutant , which is not able to grow in a synthetic fructose medium, grew when small amounts of glucose were added. The growth is p ropor t iona l to the glucose concentrat ion in the range below 0.1% but when the glucose concentrat ion exceeds 0.1% the growth rate is reduced and no growth occurs above 0.5% (Fig. 3). On the other hand no growth of the pgilA mutant could be detected at any glucose concen- t rat ion from 0.01% to 2% w/v when used as sole carbon source, whereas the amount of growth of the wild-type strain was concentrat ion dependent (Fig. 4).
Growth curves of the pgilA mutan t show that the addi- t ion of 0.1% glucose to synthetic fructose medium restored the wild-type growth rate (Fig. 5) but growth was dramat i - cally reduced when 2% glucose was added. The same result was obtained when 0.5%, 1.0% and 4% glucose was sup- plied. On the Other hand, experiments performed with Dur-
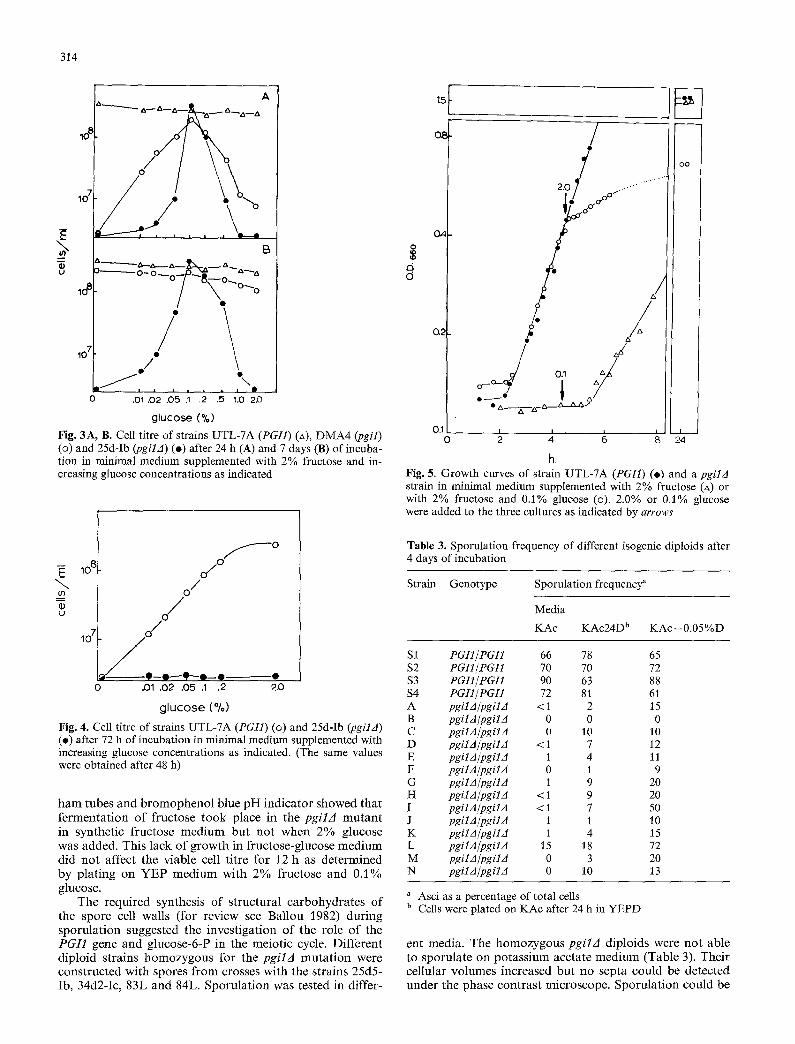
314
,o 8
g~
,J
107
\
A
B
u / ~ 0 ~ . 0 . . ~ °
/ 1 / , / | | , , , , , •
0 .01.02 .05 .1 .2 .5 1.0 2.0
glucose (%)
Fig. 3A, B. Cell titre of strains UTL-7A (PGI1) (zx), DMA4 (pgil) (o) and 25d-Ib (pgilA) (o) after 24 h (A) and 7 days (B) of incuba- tion in minimal medium supplemented with 2% fructose and in- creasing glucose concentrations as indicated
if)
0~ t.)
S o
lO 8 ° 7"0
o /
0 .01 .02 .05 .1 .2 2.0
g l u c o s e (%)
Fig. 4. Cell titre of strains UTL-7A (PGI1) (o) and 25d-Ib (pgilA) (e) after 72 h of incubation in minimal medium supplemented with increasing glucose concentrations as indicated. (The same values were obtained after 48 h)
ham tubes and bromophenol blue pH indicator showed that fermentation of fructose took place in the pgiIA mutan t in synthetic fructose medium but not when 2% glucose was added. This lack of growth in fructose-glucose medium did not affect the viable cell titre for 12 h as determined by plating on YEP medium with 2% fructose and 0.1% glucose.
The required synthesis of structural carbohydrates of the spore cell walls (for review see Ballou 1982) during sporulation suggested the investigation of the role of the P G H gene and glucose-6-P in the meiotic cycle. Different diploid strains homozygous for the pgi lA mutat ion were constructed with spores from crosses with the strains 25d5- Ib, 34d2-Ic, 83L and 84L. Sporulation was tested in differ-
0
ci d
0.1 L 0
!
o/ .............. 2. , - . "" clo.O - c / O f
/ /
0.1 ~ / o_o- I j
• ~ Z x - ~ j
2 4 6 8 24
h. Fig. 5. Growth curves of strain UTL-7A (PGI1) (o) and a pgilA strain in minimal medium supplemented with 2% fructose (zx) or with 2% fructose and 0.1% glucose (o). 2.0% or 0.1% glucose were added to the three cultures as indicated by arrows
Table 3. Sporulation frequency of different isogenic diploids after 4 days of incubation
Strain Genotype Sporulation frequency a
Media
KAc KAc24D b KAc + 0.05 % D
S1 PGI1/PGI1 66 78 65 $2 PGH/PGH 70 70 72 $3 PGH/PGI1 90 63 88 $4 PGI1/PGH 72 81 61 A pgilA/pgilA < 1 2 15 B pgilA/pgilA 0 0 0 C pgilA/pgilA 0 10 10 D pgilA/pgilA < 1 7 12 E pgilA/pgil3 1 4 i l F pgilA/pgilA 0 1 9 G pgilA/pgilA 1 9 20 H pgilA/pgilA < 1 9 20 I pgilA/pgilA < 1 7 50 J pgilA/pgilA I t 10 K pgilA/pgiIzJ 1 4 15 L pgilA/pgilA 15 18 72 M pgilA/pgilA 0 3 20 N pgilA/pgil3 0 10 13
" Asci as a percentage of total cells b Cells were plated on KAc after 24 h in YEPD
ent media. The homozygous pgi lA diploids were not able to sporulate on potassium acetate medium (Table 3). Their cellular volumes increased but no septa could be detected under the phase contrast microscope. Sporulation could be

partially restored by adding 0.05% glucose to the medium or by allowing the cells to accumulate glucose-6-P in YEP glucose before plating on sporulation media (Table 3).
Discussion
Strains with a deleted PGI1 gene have two remarkable phe- notypes: they require small amounts o f glucose to grow but high glucose concentrations in the medium completely inhibit growth (Fig. 5).
In contrast to mutants with residual phosphoglucose isomerase activity (Maitra 1971; Ciriacy and Breitenbach 1979) the pgilA mutants cannot grow on fructose as the sole carbon source, although fructose can still be fermented to CO2. Growth is only possible when fructose is supplied along with low concentrations of glucose. Glucose becomes a limiting factor for growth in strains with undetectable phosphoglucose isomerase activity (Fig. 3). Therefore in S. cerevisiae as well as in E. coli (Vinopal et al. 1975) glucose is an essential component. The latter conclusion also ex- tends to sporulation. Diploid pgilA/pgilA strains were not able to sporulate unless small amounts of glucose were sup- plied (Table 3).
These facts could be explained because cell wall glucans (Ballou 1982) and the inositol of membrane phospholipids (Henry 1982) which are both synthesized during cell growth, require glucose-6-P as a precursor, and because N A D P H in yeast is generated from glucose-6-P through direct oxidation (see Fraenkel 1982). Sporulation also re- quires the synthesis of glucans for spore cell wall formation (Kane and Roth 1974). Moreover, mutants requiring glu- cosamine are defective in sporulation (Whelan and Ballou 1975). Unless glucose-6-P derived from glucose was pro- vided for a pgilA mutant, mitosis and meiosis but not fruc- tose fermentation were blocked. This result clearly shows that phosphoglucose isomerase is the only enzyme catalys- ing the interconversion of fructose-6-P and glucose-6-P in yeasts.
As already observed with other gylcolytic mutants and also with pgi mutants (Ciriacy and Breitenbach 1979), high glucose concentrations in the medium inhibit the growth of the pgilA mutants. This inhibition is much more dramatic than in a mutant with around 1% of the wild-type phosphoglucose isomerase activity (Fig. 3). The cultures are not able to duplicate their biomass and, therefore, to com- plete a cell cycle (Fig. 5). The role that glucose-6-P accumu- lation (Maitra 1971), ATP depletion (Ciriacy and Breiten- bach 1979) or the competition of both fructose and glucose molecules for both hexokinases play in this inhibition is difficult to clarify. But the complete inability to grow on high glucose concentrations and the immediate reduction of the growth rate after adding glucose support the sugges- tion of Ciriacy and Breitenbach (1979) that ATP depletion is the primary effect leading to growth cessation on glucose.
Finally, the pgilA mutants are unable to grow at any glucose concentration from 0.01% to 2% when supplied as sole carbon source (Fig. 4). This result demonstrates that in S. cerevisiae the direct oxidation of glucose-6-P is not able to support growth, in contrast to E. coli mutants with- out phosphoglucose isomerase activity (Vinopal et al. J 975) which are still able to grow on glucose. Therefore in S. cerevisiae the, oxidative metabolism of glucose-6-P is not sufficient to support growth. The oxidation of glucose-6-P and its catabolism through glycolysis via the pentose-P
315
pathway, which could produce fructose-6-P and glyceralde- hyde-3-P through the transketolase and transaldolase reac- tions (see Fraenkel 1982), seems to be insufficient to supply all the components required in metabolism.
Acknowledgements. I wish to thank Prof. F.K. Zimmermann for many fruitful discussions and careful reading of the manuscript, and I. Aguilera for typing the manuscript. This investigation was supported by a grant of the Deutsche Forschungsgemeinschaft to F.K. Zimmermann. A.A. is a "Fundaci6n Juan March" (Spain) Postdoctoral Fellow.
References
Aguilera A, Zimmermann FK (1986) Isolation and molecular anal- ysis of the phosphoglucose isomerase structural gene of Sac- charomyces cerevisiae. Mol Gen Genet 202:83-89
Ballou CE (1982) Yeast cell wall and cell surface. In: Strathern NJ, Jones EW, Broach JR (eds) The molecular biology of the yeast Saccharomyces. Metabolism and gene expression. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York, pp 335-360
Beggs JD (1978) Transformation of yeast by a replicating hybrid plasmid. Nature 275:104-108
Bolivar F, Backman K (1979) Plasmids of Escherichia coli as clon- ing vectors. Methods Enzymol 68 : 245-267
Birnboim HC (1983) A rapid alkaline extraction method for the isolation of plasmid DNA. Methods Enzyrnol 100:243-255
Ciriacy M, Breitenbach I (1979) Physiological effects of seven dif- ferent blocks in glycolysis in Saccharomyces cerevisiae. J Bacter- iol 139:152-160
Ciriacy M, Williamson VM (1981) Analysis of mutations affecting Ty-mediated gene expression in Saccharomyces cerevisiae. Mol Gen Genet 182:159-163
Clifton D, Weinstock SB, Fraenkel DG (1978) Glycolysis mutants in Saccharomyces cerevisiae. Genetics 88:1-11
Cohen SN, Chang ACY, Hsu L (1972) Nonchromosomal antibiotic resistance bacteria: genetic transformation of Escherichia coli by R-factor DNA. Proc Natl Acad Sci USA 69:2110 2114
Eisenberg F (1978) Intermediates in the myo-inositol-l-phosphate synthase reaction. In: Wells W, Eisenberg F (eds) Cyclitols and phosphoinositides. Academic Press, New York pp 269-275
Falco SC, Yuyang L, Broach JR, Botstein D (1982) Genetic prop- erties of chromosomally integrated 2 ~ plasmid DNA in yeast. Cell 29 : 573-584
Fraenkel DG (1982) Carbohydrate metabolism. In: Strathern NJ, Jones EW, Broach JR (eds) The molecular biology of the yeast Saccharomyces. Metabolism and gene expression. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York, pp 1-37
Fraenkel DG, Vinopal RT (1973) Carbohydrate metabolism in bacteria. Annu Rev Microbiol 27:6%100
Henry SA (1982) Membrane lipids of yeast: Biochemical and genet- ic studies. In: Strathern N J, Jones EW, Broach JR (eds) The molecular biology of the yeast Saccharomyces. Metabolism and gene expression. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York, pp 101 158
Herrera LS, Pascual (1978) Genetical and biochemical studies of glucosephosphate isomerase deficient mutants in Saccharo- myces cerevisiae. J Gen Microbiol 108:305-310
Kane SM, Roth R (1974) Carbohydrate metabolism during asco- spore development in yeast. J Bacteriol 118:8-14
Kempe TD, Gee DM, Hathaway GM, Noltmann EA (1974) Sub- unit and peptide compositions of yeast phosphoglucose isomer- ase isoenzymes. J Biol Chem 249:4625-4633
Maitra PK (1971) Glucose and fructose metabolism in a phospho- glucoisomeraseless mutant of Saccharomyces cerevisiae. J Bac- teriol 107:75%769
Maitra PK, Lobo Z (1977) Genetic studies with a phosphoglucose isomerase mutant of Saccharomyces cerevisiae. Mol Gen Genet 156: 55-60

316
Mertz JE, Davis RW (1972) Cleavage of DNA by R1 restriction endonuclease generates cohesive ends. Proc Natl Acad Sci USA 69: 3370-3374
Nasmyth KA, Reed SI (1980) Isolation of genes by complementa- tion in yeast: Molecular cloning of a cell cycle gene. Proc Natl Acad Sci USA 77:2119-2123
Rigby PWJ, Dieckmann M, Rhodes C, Berg P (1977) Labelling deoxyribonucleic acid to high specific activity by nick transla- tion with DNA polymerase I. J Mol Biol 113:237-251
Rothstein RJ (1983) One-step gene disruption in yeast. Methods Enzymol 101 : 202-211
Sherman F, Fink GR, Lawrence CW (1979) Methods in yeast genetics. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York
Snow R (1966) An enrichment method for auxotrophic yeast mu- tants using the antibiotic nystatin. Nature 211:206-207
Tautz D, Renz M (1983) An optimized freeze-squeeze method for
the recovery of DNA fragments from agarose gels. Anal Bio- chem 132:14-19
Vinopal RT, Hillman JD, Schulman H, Reznikoff WS, Fraenkel DG (1975) New phosphoglucose isomerase mutants of Eseherichia coli. J Bacteriol 122:1172-1174
Wahl GN, Stern M, Stark GR (1979) Efficient transfer of large DNA fragments from agarose gels to diazobenzyl-oxymethyl- paper and rapid hybridization by using dextran sulfate. Proc Natl Acad Sci USA 76:3683-3687
Whelan WL, Ballou CE (1975) Sporulation in D-glucosamine auxo- trophs of Saccharomyces cerevisiae: Meiosis with defective as- cospore wall formation. J Bacteriol 124:1545-1557
Communicated by C.P. Hollenberg
Received February 13 /April 15, 1986