diagnosis of human preimplantation embryoviability · pdf filemolecular karyotyping and of...
TRANSCRIPT

...........................................................................................................................
Diagnosis of human preimplantationembryo viabilityDavid K. Gardner1,*, Marcos Meseguer2, Carmen Rubio3,and Nathan R. Treff4
1School of BioSciences, University of Melbourne, Parkville, VIC 3010, Australia 2Laboratorio Fiv, IVI Valencia, Plaza Policıa Local, 3, Valencia46015, Spain 3IGENOMIX and Fundacion Instituto Valenciano de Infertilidad (FIVI)/INCLIVA, Valencia, Spain 4Reproductive Medicine Associatesof New Jersey, Basking Ridge, NJ 07960, USA
*Correspondence address. E-mail: [email protected]
Submitted on June 15, 2014; resubmitted on November 2, 2014; accepted on December 8, 2014
table of contents
† Introduction† Methods† Morphokinetics/time-lapse imaging† Comprehensive chromosome screening
CGH arraysSNP arrays, qPCR and NGS
† Analysis of physiology and the search for biomarkersProteomicsMetabolomicsTargeted metabolomicsGlucose utilizationAmino acid utilization
† Laboratory on a chip† Conclusions
background: Transfer of more than a single embryo in an IVF cycle comes with the finite possibility of a multiple gestation. Even a twinpregnancy confers significant risk to both mother and babies. The move to single-embryo transfer for all patients will be greatly facilitated bythe ability to quantify embryo viability. Developments in time-lapse incubation systems have provided new insights into the developmental kineticsof the human preimplantation embryo. Advances in molecular methods of chromosomal analysis have created platforms for highly effectivescreening of biopsied embryos, while noninvasive analysis of embryo physiology reveals more about the embryo than can be determined bymorphology alone.
methods: Recent developments in time-lapse microscopy, molecular karyotyping and in proteomics and metabolomics have been assessedand presented here in a descriptive review.
results and conclusions: New algorithms are being created for embryo selection based on their developmental kinetics in culture,and the impact of factors such as patient etiology and treatment are being clarified. Potential links between morphokinetic data and embryo kar-yotype are being elucidated. The introduction of new molecular methods of determining embryo chromosomal complement is proving to beaccurate and reproducible, with the future trending toward CGH arrays or next generation sequencing as a rapid and reliable means of analysis,that should be suitable for each IVF clinic to adopt. A relationship between embryo metabolism and viability is established and is now being con-sidered together with morphokinetic data to create more robust algorithms for embryo selection. Microfluidic devices have the capacity and po-tential to be used in human IVF clinics for the routine diagnosis of embryo biomarkers.
Key words: chromosome / metabolism / selection / time lapse / embryo viability
& The Author 2015. Published by Oxford University Press on behalf of the European Society of Human Reproduction and Embryology. All rights reserved.For Permissions, please email: [email protected]
Human Reproduction Update, Vol.21, No.6 pp. 727–747, 2015
Advanced Access publication on January 6, 2015 doi:10.1093/humupd/dmu064

IntroductionThe goal of assisted reproduction in the human is the delivery of a healthysingleton child through the transfer of an individual euploid embryo. Withmajor advances in embryo culture and cryopreservation over the past 15years, resulting in significant increases in embryo implantation rates and incumulative pregnancy rates per retrieval, we have witnessed a significantreduction in the numbers of embryos being transferred, with several coun-tries undertaking routine single-embryo transfer (SET) and confining high-order multiple gestations to the archives of human IVF. However, unlessdiagnosis of embryo viability is applied for embryo selection for transfer,we condemn patients to successive cycles of retrievals, cryopreservationand replacements. Our objective should be to assist couples to attain ahealthy family sooner than later, not sooner or later.
A greater understanding of embryo developmental kinetics and physi-ology will facilitate the identification of the most viable embryo for trans-fer. However, even if we can identify those embryos within a cohort mostable to implant, one cannot guarantee their ploidy, nor in cases of familialgenetic diseases, can we select the unaffected conceptus. Rapid advancesin genetic screening technologies are now improving our ability to screenembryonic cells, to ensure that transfer of a healthy euploid embryobecomes routine. Of significance, relationships between the kinetics ofembryo development, proteomics and metabolism with the chromo-somal status of the embryo are emerging. Consequently, in thoseclinics or countries where detailed genetic analysis is not feasible, thereis at least the potential to decrease the possibility of transferring an aneu-ploid embryo through a more detailed analysis of embryonic develop-mental history.
Mastenbroek et al. (2011a, b) fashioned a challenging argument that‘the path of embryo selection is turning into a dead-end in the questfor optimal IVF success rates’. Furthermore, it was proposed that‘Embryo selection techniques will not only be unable to improve livebirth rates but also they could even lower the success rate of IVF’, andthat in the ‘perfect world’ the best that embryo selection can do isshorten the time to pregnancy. Even if the only reason to performembryo selection is to decrease the time it takes for a couple to conceive,then that in itself is a very good reason. Patients do not consider a reduc-tion in the time to achieve a pregnancy as nonconsequential. Further-more, reducing the number of transfers a patient must undergo alsocarries with it a reduction in financial and emotional pain for thepatient. Every time a patient undergoes a transfer and subsequent preg-nancy test, they experience a dramatic rollercoaster of emotions thatcreates an unparalleled toll on their ‘infertility’ psychology. Routine IVFis already believed to be associated with a 17% drop out rate (Verberget al., 2008). However, without the ability to screen the embryos ofpatients for genetic and chromosomal abnormalities, patients willendure either countless cycles of miscarriage, or the challenges asso-ciated with a subsequent amniocentesis. Here we discuss how develop-ments in the fields of time-lapse microscopy (mophokinetic analysis),genetic diagnosis, and in proteomics and metabolism could facilitatethe identification and transfer of an individual viable euploid embryo,thereby reducing the time to a healthy pregnancy for our patients.
MethodsThe available evidence regarding the clinical use of time-lapse microscopy,molecular karyotyping and of proteomics and metabolomics has been
assessed and presented here. We consider the use and implementation ofnew technologies and their potential impact on facilitating SET and theirability to increase the take-home baby rate. No systematic review hasbeen carried out.
Morphokinetics/time-lapseimagingMorphological evaluation has been the method of embryo selection forover 30 years and remains the primary approach of embryo assessmentduring IVF cycles. However, this evaluation method poses limitations,not only arising from the subjectivity of the embryologist, but alsobecause of the evaluation system itself, which views embryo develop-ment statically. Current embryo evaluation is typically based on the mor-phological appearance of embryos at just a few discrete time points,which limits the frequency of observations made thereby concealingwhat happens during the intervals between observations (Cruz et al.,2012). In addition, it is important to bear in mind that the status of anembryo (and thus its grading) can change markedly within just a fewhours (Mio and Maeda, 2008; Meseguer et al., 2011). These limitationscan be overcome with time-lapse systems by which one cannot onlyanalyze embryo morphology but also provide information about thedynamic changes during the preimplantation period. The key advantagesof time-lapse technology include the provision of an improved culture en-vironment based on embryo evaluation without removal from the incu-bator, and the objective and accurate information obtained, not onlyquantitatively, but also qualitatively. Time-lapse observations have alsobeen useful in defining new or poorly described concepts of embryologysuch as the duration of the first three cell cycles (Lemmen et al., 2008; Mioand Maeda, 2008; Meseguer et al., 2011) and furthermore, redefiningtraditionally used morphology parameters used for embryo selectionsuch as pronuclear score (Azzarello et al., 2012; Aguilar et al., 2014).
Here the most relevant studies performed on time-lapse in the last fewyears are considered; Lemmen et al. (2008) reported that embryosresulting in pregnancies displayed not only a significantly higher cleavagesynchrony but also a higher synchrony in nuclear appearance at thetwo-blastomere stage compared with nonimplanting embryos. Wonget al. (2010) suggested that embryos that reached the blastocyst stagecould be predicted by the timing of early developmental stages: firstcytokinesis 0–33 min, time interval between first and second mitosesof 7.8–14.3 h, and time interval between second and third mitoses of0–5.8 h. Finally, it was observed that in embryos which did not form ablastocyst, fragmentation was rarely reversed, whereas it could be inthose reaching the blastocyst stage. Using morphokinetics it has beenpossible to demonstrate associations between various cleavage stagekinetic parameters and the ability of the embryos to reach the blastocyststage (Cruz et al., 2012). One of the largest data sets on transferred ICSI-generated embryos analyzed by time-lapse was presented by Mesegueret al. (2011), where results obtained were used to design a hierarchicalclassification for the selection of embryos with the highest implantationpotential. The timing of the cleavage to 5 cells (t5) had the best correl-ation with implantation success. The algorithm started with a morpho-logical screening of the embryos to discard nonviable embryos. Thenext step was to exclude embryos fulfilling at least one of the followingexclusion criteria: (i) uneven blastomere size after the first cleavage; (ii)abrupt division from one to three cells or (iii) multinucleation at the
728 Gardner et al.

4-cell stage. After evaluation for these criteria, the remaining levels of thehierarchy were defined according to the exact timings of some of the vari-ables (t5, the duration of the transition from a 2-cell to a 4-cell embryos(s2) and the duration of the second cell cycle (cc2)). Using this algorithmand logistic regression to analyze the predictive abilities of time lapse, acomparison using a receiver operating characteristic curve (ROC) wasmade with standard morphology which gave an area under the curve(AUC) of 0.64, whereas a logistic regression on the time-lapse selectiongave an AUC of 0.72. The higher AUC supports the possibility ofimproved embryo selection using time-lapse. In the largest retrospectivestudy on pregnancy outcomes compared with conventional incubationthere was a relative 20% improvement in pregnancy rates in the time-lapse system (Meseguer et al., 2012). These results have since been con-firmed in a prospective RCT (Rubio et al., 2014). Data reflect improve-ments of the clinical outcome as a consequence of both improvedculture conditions and a better embryo selection process by using a pre-defined morphokinetic algorithm.
Once methods for embryo selection by using time-lapse were estab-lished, issues arose of whether there could be universal application ofthese algorithms, or whether different culture conditions may affecttheir efficacy and restrict their use only to those IVF units where themodels were generated. Basile et al. (2013) compared different culturemedia, by a RCT on sibling oocytes, and observed identical timings andproportion of optimal embryos for the two media analyzed. However,in contrast, Ciray et al. (2012) reported different timings with differentculture media using a similar experimental design. Furthermore,oxygen has been shown to have profound effects on mammalian preim-plantation embryo developmental kinetics (Wale and Gardner, 2010;Kirkegaard et al., 2013a). Consequently, it is important that bothmedia type and oxygen concentration be reported in clinical studies, asboth can affect the timing of key developmental events.
Chamayou and colleagues defined two sets of morphokinetic para-meters (MKP). Firstly, they established optimal ranges for some para-meters which were considered predictive of the embryo’s ability tobecome a viable blastocyst on Day 5 (D5; time of pronuclei formation;t1, time of 2,4, 7 and 8 cell cleavage, t(n); t2, t4, t7, t8) (Chamayouet al., 2006). They then suggested that cc3 was the MKP significantly asso-ciated with implantation and a viable pregnancy (t5-t3; cc3). With theseresults they suggested D3 embryo transfer for those embryos with MKPoutside optimal ranges. Analysis of initial events of development revealedthat PN breakdown occurred significantly later in embryos resulting inlive birth and never earlier than 20 h 45 min (Azzarello et al., 2012),while timings at which second polar body extrusion, pronuclear fadingand length of S-phase occurred were linked successfully to embryo im-plantation (Aguilar et al., 2014).
Potential inter- and intra-observer variability may impact time-lapsemarker interpretation, similar to what has been found with manualembryo morphology grading (Chen et al., 2013). Of note, a prospectivestudy has recently demonstrated the ability of a noninvasive, computer-automated test to improve the prediction of blastocyst formation by D3and provide clinical value to embryologists for D3 selection without anymanual analysis (Conaghan et al., 2013). The test, termed Eeva (earlyembryo viability assessment), is facilitated by dark-field microscopyand cell-tracking software algorithms (based on P2 (t3-t2) and P3(t4-t3) as relevant parameters categorizing embryos into groups witheither high or low likelihood of forming ‘usable blastocysts’). AddingEeva to traditional D3 morphology significantly improved the ability of
experienced embryologists to predict usable blastocyst outcomes andreduced the variability among embryologists (Conaghan et al., 2013).Eeva significantly improved the specificity (84.7 versus 52.1%; P ,
0.0001) and the positive predictive value (54.7 versus 34.5%; P ,
0.0001) of usable blastocyst predictions compared with morphologyevaluation alone (Conaghan et al., 2013). Subsequently, the transferabil-ity of this model in four different clinics has been demonstrated retro-spectively (Kirkegaard et al., 2014). Even though results obtainedshowed a 30% higher implantation rate between the entire cohort andembryos categorized as usable, 50.6% of the embryos that resulted inpregnancy were categorized as nonusable according to the algorithm.Consequently, these data highlight the importance of increasing implant-ation rates with the application of a model, but with the need to bear inmind the importance of a low rejection rate of viable embryos. A furtherstudy has since confirmed cases where poor conventional morphologicalscore and/or suboptimal morphokinetic embryos have resulted in livebirths, highlighting the risk of discarding so-called ‘viable embryos’(Stecher et al., 2014).
The transfer of blastocysts yields higher implantation rates than trans-fers at the cleavage stage, but this outcome must be balanced against thepossible disadvantages of longer culture such as the risk of canceledcycles and concerns over the possible epigenetic effects of prolongedin vitro culture (Kirkegaard et al., 2012a). An analysis of blastocyst transfercombined with time-lapse analysis found that human embryo cleavagerates are suggestive of their ability to develop to the blastocyst stageand to implant (Dal Canto et al., 2012). It was concluded that cleavagefrom the 2- to 8-cell stage occurs progressively earlier in embryos withthe ability to develop to blastocysts and to implant. In this line of research,Kirkegaard et al. (2013b) suggested that high-quality blastocysts could bepredicted within the first 48 h of culture by a short duration of the firstcytokinesis, duration of the 3-cell stage and absence of direct cleavageto 3 cells. The duration of the 2-cell stage either in Kirkegaard et al.(2012a, b) or the study by Cruz et al. (2012) is not found to differbetween high- and low-quality embryos, in contrast to the findings byWong et al. (2010). Further studies may reveal the correlation of key de-velopmental parameters with subsequent blastocyst score. However, acaveat here is that human embryos can have the same embryo grade andyet differ with respect to their proteome and metabolism (discussedfurther on), and that transfer of the cleavage stage embryo to theuterus is asynchronous, which results in compromised fetal developmentin animal models (Barnes, 2000; Walker et al., 2015).
There are several studies that have correlated time-lapse with prob-ability of selecting chromosomally normal embryos (Montag, 2013;Swain, 2013). Davies et al. (2012) observed that embryos withcomplex aneuploidies presented delays on the first two cleavages aswell as prolonged transitions between 2 and 4 cells. They also observedirregular divisions and an asynchronous PN disappearance in abnormalembryos. Chavez’s group observed precise timings in all euploidembryos to the 4-cell stage, whereas only 30% of aneuploid embryospresented parameter values within normal timing windows (Chavezet al., 2012). Campbell and colleagues went on to develop a model to cat-egorize the riskof aneuploidy using time-lapse. They found no differencesin the early stages, but defined as parameters relevant to ploidy the initi-ation of blastulation and the timing of the formation of a full blastocyst(Campbell et al., 2013a, b). Similarly, it has been reported that there isa significant correlation between early times to cavitation, from thefirst cytokinesis and from 5 cells, and reduced prevalence of aneuploidy
Diagnosis of human embryo viability 729

(Hong et al., 2013). Basile and colleagues studied the differences in thecleavage time between chromosomally normal and abnormal embryosin order to elaborate an algorithm to increase the probability of noninva-sively selecting chromosomally normal embryos. A different kinetic be-havior was observed, being able to identify, using a logistic regressionanalysis, t5-t2 and cc3 (t5-t3) as the most relevant parameters relatedto normal chromosomal content (Basile et al., 2014). However, in aretrospective analysis Kramer et al. (2014) have been unable to segregateaneuploid from euploid embryos using MKP. Consequently, selection ofembryos through time-lapse technology should not be considered as areplacement for preimplantation genetic screening (PGS). As reportedby Basile et al. a ROC analysis to determine the predictive propertiesof morphokinetics with respect to chromosomal normality gave anAUC value of 0.634 (95% confidence interval, 0.581–0.687), a valuewhich is below what we should expect from a diagnostic tool that isable to forecast an abnormal embryo (with an abnormal chromosomecontent) (Basile et al., 2014). However, it does represent an excellent se-lection tool for good prognosis patients who are not indicated for PGS, orfor patients who are indicated but who for any legal, social, or economicreasons do not wish to, or cannot, have PGS performed. A summary ofthe papers published in time-lapse research are summarized in Table Iand the graphic representation of the morphokinetic models developedby diverse groups and using different outcomes are shown in Figure 1.
In conclusion, we have tried to convey the issues around the use oftime-lapse technology as a clinical tool. We are conscious that standard-ization in embryo annotation and nomenclature is necessary in order toallow a more accurate interpretation of the data available, and that theexisting literature does not yet provide any certainty on the improvementin live birth rates permitted by time-lapse monitoring (TLM). However,we should emphasize that studies do exist which are based on largesample sizes and include several centers from different countries(Rubio et al., 2012), and that recently a RCT has been published whichstrongly supports time-lapse as a strategy for embryo selection (Rubioet al., 2014). Finally, we wish to convey that conventional morphology as-sessment only allows moderate prediction of the embryo’s implantationability and suffers from relatively limited specificity and sensitivity. In add-ition, it is prone to inter- and intra-observer variations (Baxter Benduset al., 2006) and thus can never represent a truly objective quantifiablemarker.
Comprehensive chromosomescreeningDespite the identical objective of determining the molecular karyotype ofan embryo, not all methods of comprehensive chromosome screening(CCS) are the same (Treff, 2012). In fact, the specific methods used, per-formance characteristics, and level of supportive evidence of each can bequite specific (Treff and Scott, 2012). For example, there are manymethods of DNA amplification, which likely represents the most criticalelement in the success of CCS. The most common method, wholegenome amplification (WGA), can itself be performed using anynumber of commercially available kits, such as RepliG (QIAgen), Geno-miPhi (GE Healthcare), GenomePlex (Sigma Aldrich), PicoPlex (RubiconGenomics) or MALBAC (Yikon Genomics). There are many factorsinvolved in the consideration of which WGA method to use. Forexample, when the objective is accurate genotyping a multiple
displacement amplification (MDA)-based approach (i.e. RepliG) maybe more accurate than PCR-based methods (i.e. GenomePlex) (Treffet al., 2011a). Consistent with this observation, methods such as karyo-mapping (Handyside et al., 2010) and parental support (Johnson et al.,2010a, b) have used MDA-based WGA as they rely upon genotype infor-mation to make diagnoses. In contrast, PCR-based WGA methods havedemonstrated superior performance for copy number based analysessuch as with array comparative genomic hybridization (CGH) (Harperand Harton, 2010), quantitative single nucleotide polymorphism (SNP)array methods (Treff et al., 2010a; Konings et al., 2012), and next-generation sequencing (NGS) (Baslan et al., 2012; Lu et al., 2012; Kohnet al., 2013; Voet et al., 2013; Yin et al., 2013; Zhang et al., 2013). An al-ternative to WGA involves targeted multiplex PCR, a method that is lessexpensive and more rapid than WGA, and that has been applied to quan-titative real-time (q)PCR (Treff et al., 2012a) and NGS-based CCS (Treffet al., 2013a, b). Ultimately, it is difficult to evaluate which method ofinitial amplification from the embryo is optimum, as cross-comparisonstudies have proven to be virtually impossible due to significant commer-cial conflict of interest.
In addition to the wide variety of available methods for amplification,there are also many platforms for evaluating the amplified DNA. Herethe focus is on CGH and SNP arrays, qPCR, and NGS-based CCS. Inall cases, platforms of analysis were developed for applications that donot require extensive pre-amplification of DNA and instead weredesigned for large quantities of starting material [i.e. as in cancer(Nannya et al., 2005), genome-wide association studies (Glessner andHakonarson, 2011) and prenatal diagnosis (Rajcan-Separovic, 2012;Shaffer et al., 2012)]. As a result, performance when starting from onlya single or few cells is an important component to consider before appli-cation to embryo CCS. Despite the obvious benefit of the positivecontrol, few studies exist that have rigorously evaluated performanceon cells with known aneuploidies. Some argue that even cell lines arepoor controls as they may possess significant levels of mosaicism.However, there are many stable cell lines with little to no evidence of mo-saicism by conventional testing methodologies, which provide an ad-equate positive control to evaluate the performance of new testingmethods. Examples of the use of blinded analysis of cell lines exist forthe development of SNP arrays (Treff et al., 2010a), qPCR (Treff et al.,2012a) and targeted NGS (Treff et al., 2013b).
CGH arraysAneuploidies are extremely common in the early human embryo(Harper et al., 1995; Munne and Cohen, 1998). Trisomic and monosom-ic embryos account for at least 10% of human pregnancies and, forwomen nearing the end of their reproductive lifespan the incidencecan exceed 50% (Nagaoka et al., 2012). Age-related defects result inhigher aneuploidy rates in offspring and an increase in spontaneous abor-tions, thereby reducing ongoing implantation rates (Boue et al., 1985).Aneuploidy may also be a contributing factor in other infertile popula-tions. For example, an abnormal embryonic karyotype was found tobe the most frequent cause of recurrent miscarriage (RM) (Sugiura-Ogasawara et al., 2012). Recurrent implantation failure (RIF) remains aclinical challenge and embryonic aneuploidy has been implicated asone of the leading embryonic causes (Margalioth et al., 2006). In malefactor (MF) infertility, an increase in sperm chromosomal abnormalitiesdue to impairment of the meiotic process was described (Rubio et al.,
730 Gardner et al.

.............................................................................................................................................................................................
Table I Summary of studies published from 2010 that have used time-lapse research on human embryos in a clinical setting.
Study Aim
Aguilar et al. (2014) Correlate timings of fertilization events with embryo implantation
Aparicio et al. (2013) Time-lapse review
Athayde Wirka et al. (2014) Characterize atypical dynamic embryo phenotypes identified by time-lapse, and determine their association with embryo development
Azzarello et al. (2012) Study PN development in embryos after ICSI
Basile et al. (2012) Time-lapse review
Basile et al. (2013) Evaluate the effect of two types of culture media on early markers of embryo development
Basile et al. (2014) Elaborate an algorithm to increase the probability of noninvasively selecting a chromosomally normal embryo
Bellver et al. (2013) Assess morphokinetic evaluation of embryos derived from obese women
Campbell et al. (2013a) Develop a model to categorize the risk of aneuploidy in embryos based on morphokinetics
Campbell et al. (2013b) Evaluate the effectiveness of the previously established, morphokinetic-based aneuploidy risk classification model
Chamayou et al. (2013) Determine morphokinetic parameters predictive of embryo development and, competence in producing a clinical pregnancy afterD5 transfer
Chavez et al. (2012) Analyze the chronology of early mitotic events in 4-cell embryos
Chen et al. (2013) Time-lapse review
Ciray et al. (2012) Comparison of different culture media on morphokinetics
Conaghan et al. (2013) Assess a computer-automated platform for time-lapse image analysis and blastocyst prediction
Cruz et al. (2012) Analyze associations between embryo division kinetics and ability to reach blastocyst stage
Cruz et al. (2012) Demonstrate that time-lapse incubation conditions are comparable to conventional incubator
Cruz et al. (2013) Determine if IVF or ICSI techniques influence embryo morphokinetics
Dal Canto et al. (2012) Analyze cleavage timings in relation to blastocyst potential and implantation
Ergin et al. (2014) To compare the detection rate of multinucleation with the time-lapse system and conventional control timing proposed by EuropeanSociety of Human Reproduction and Embryology (ESHRE) consensus and evaluate its impact on pregnancy rates
Freour et al. (2013) Evaluate early embryo morphokinetic parameters according to female smoking status
Hashimoto et al. (2012) Assess the development kinetics of embryos and their ability to develop to blastocysts
Herrero et al. (2013a) Time-lapse review
Herrero et al. (2013b) Updated timings based on morphokinetic studies developed by Meseguer et al. and implementation of morphokinetic selectioncriteria
Iwata et al. (2014) Analyze the timing of initiation of compaction in human embryos
Joergensen et al. (2014) Analyze the cleavage patterns in dipronuclear (2PN) and tripronuclear (3PN) embryos in relation to fertilization method
Kaser and Racowsky (2014) Time-lapse review
Kirkegaard et al. (2012b) Evaluate the effect of blastomere biopsy on early embryonic development using time-lapse
Kirkegaard et al. (2013a) Evaluate the effect of culture oxygen concentrations on embryo development using time-lapse
Kirkegaard et al. (2013b) Apply Wong’s model to a large set of transferred embryos to test this hypothesis and the correlation between published timeintervals and clinical outcome
Kirkegaard et al. (2013b) Test proposed time-lapse parameters during the first 48 h and to analyze if timing during the entire culture period differed betweenembryos destined to implant and to fail
Knez et al. (2013) Elucidate if the presence of sperm vacuoles impacts on early embryo developmental dynamics
Liu et al. (2014) To investigate the prevalence and potential causes of reverse cleavage by human early cleavage embryos and its associations withembryonic development and implantation after transfer
Machtinger and Racowsky(2013)
Time-lapse review
Meseguer et al. (2011) Generate and evaluate an embryo selection tool based on morphokinetics
Meseguer et al. (2012) Compare pregnancy outcomes of treatments in an incubator with time-lapse versus tissue culture chamber
Meseguer et al. (2012) Describe current efforts made to standardize different steps of ART such as time-lapse
Munoz et al. (2012) Evaluate if the type and dose of gonadotrophin influences embryo morphokinetics, and if the estradiol and progesteroneconcentrations in serum has an impact on embryo
Munoz et al. (2013) Evaluate if type of GnRH analog used during controlled ovarian stimulation influences early embryo developmental kinetics
Montag (2013) Current embryo selection strategies and their clinical usefulness, particularly to enhance successful single-embryo transfer
Montag (2013) Review of studies that attempt to correlate timings with embryonic aneuploidy
Continued
Diagnosis of human embryo viability 731

2001; Rodrigo et al., 2010). PGS by fluorescence in situ hybridization(FISH) for a limited number of chromosomes was widely applied foralmost two decades. However, this technique lacked sensitivity andwas far from comprehensive with regards to the coverage of chromo-some complement. Consequently, therewas aclear need for a techniquecapable of CCS, which could also produce reliable and faithful results in ashort period of time. Among other technologies CGH arrays have beenidentified as a robust and accessible diagnostic approach to assess24-chromosome aneuploidy, and consequently IVF programs aremoving toward CCS using CGH arrays (Simpson, 2012; Handyside,2013). This platform was first validated by reanalyzing the sameembryos with FISH, confirming the high efficiency of the platform for an-euploidy detection with only 2.9% of embryos having no results, and theerror rate with discrepancies when compared with FISH was 1.9%(Gutierrez-Mateo et al., 2011). Furthermore, CGH arrays platformwere validated in single cells from embryos previously diagnosed as ab-normal by FISH, with high concordance rates (Mir et al., 2013). Therewere no differences in efficiency and accuracy when comparing cleavageand blastocyst stage biopsies with whole embryo CGH arrayanalysis (Miret al., 2013). Additionally, CGH arrays can be applied to identify unba-lanced embryos in carriers of Robertsonian and reciprocal transloca-tions, with a resolution as high as 6 Mb for trophectoderm biopsies(Alfarawati et al., 2011). Therefore, CCS with CGH arrays can be accur-ately applied at different embryo biopsy stages, and currently, blastocystbiopsy is the most common approach, with some groups also describingsuccessful results with D3 biopsies. For D3 biopsies, euploid embryosare commonly transferred at blastocyst stage, in the same cycle asretrieval. For blastocyst biopsies in most of the centers, blastocysts arebiopsied and vitrified for transfer in a subsequent cycle. For bothapproaches, DNA amplification from a single cell or 4–5 cells is per-formed, followed by DNA labeling with Cy3 and Cy5 fluorophoresand cohybridization onto the arrays for 4–12 h (Fig. 2). Fluorescence in-tensity is detected using a laser scanner and specific software is used fordata processing (Fig. 2).
In cleavage stage biopsies, a retrospective studyof 2858 cycles showedthe clinical benefits of embryo selection based on CGH array analysis inpoor reproductive prognosis couples. The most frequent indication wasadvanced maternal age (AMA), followed by RIF, MF and couples with a
previous trisomic pregnancy. The percentage of informative embryoswas 97.9%, with only 2.1% of samples noninformative mostly due tocell loss or damage during manipulation, resulting in defective amplifica-tion. Aneuploidy rates were 67.2% in women ,38 years and 86.3% inwomen ≥38 years. Differences in the percentage of chromosomal ab-normalities could be attributed to the significant increase in embryoswith complex aneuploidies with AMA (9.6 versus 23.5%, P , 0.05);with no differences in the percentages of embryos with chaotic pattern(15.1 versus 16.7%) or in embryos with segmental aneuploidies (6.9versus 5.1%), in women ,38 years and ≥38 years, respectively. Meanpregnancy rate per transfer and implantation rate in women ,38years were 59.0% and 49.0%, respectively. In women ≥38 years, preg-nancy rates per transfer and implantation rates stand as high as 51.1%and 46.2%, respectively. Miscarriage rates were also comparable in the,38 years and ≥38 years age groups (11.9 versus 9.9%). The increasein chromosomal abnormalities related to female age was reflected in ahigher percentage of cycles without embryo transfer in women ≥38years, showing 62% of cycles with all embryos chromosomally abnormal.However, when a euploid embryo was transferred, outcome was notaffected by maternal age (Rubio et al., unpublished). Another retrospect-ive case–control study comparing the outcome in poor prognosispatients who underwent cleavage stage biopsies with fresh embryotransfer cycles and similar patients undergoing regular IVF cyclesshowed double implantation and ongoing pregnancy rate with a decreasein multiple pregnancy and miscarriage rates in CCS cases (Keltz et al.,2013).
At the blastocyst stage, Wells et al. (2009) found that the probability ofan individual analyzed by CGH arrays of generating a pregnancy was66.7% compared with 27.9% without CGH array testing. A multicenterretrospective study described an increase in the incidence of aneuploidembryos, which correlated with increased maternal age, observingsimilar implantation and ongoing pregnancy rates per transfer afterCCS in patients up to 42 years of age, after which these rates dramaticallydeclined (Harton et al., 2013). Another retrospective study of 106blastocyst biopsy cycles showed an increase in aneuploidy rates withAMA that was associated with the higher percentage of complex abnor-malities in embryos from AMA patients (Rubio et al., unpublished). Thepercentages of chaotic embryos and embryos with segmental
.............................................................................................................................................................................................
Table I Continued
Study Aim
Nakahara et al. (2010) Evaluate the safety of time-lapse observations using a built-in microscope incubator
Pribenszky et al. (2010) Evaluate the correlation between the time of cleavage stages, occurrence of fragmentation and development to the blastocyst.
Rubio et al. (2012) Analyze implantation rate of embryos with cleavage from 2 to 3 cells in ,5 h
Serdarogullari et al. (2014) Study gender-specific embryo development kinetics between human male and female embryos
Stecher et al. (2014) Cases of time-lapse imaging, demonstrating that embryos with poor scores between D1 and D3, can result in the birth of healthybabies
Sundvall et al. (2013) To assess the variability of time-lapse annotations
Swain (2013) Review of studies that attempt to correlate timings with embryonic aneuploidy
Wong et al. (2010) Prediction of embryo potential to reach blastocyst stage using three dynamic imaging parameters
Wong et al. (2013) Time-lapse review
PN, pronuclei; ART, assisted reproductive technology.
732 Gardner et al.

Figure 1 Graphical representation of the published algorithm that, using morphokinetics, suggests a method for embryo selection based on implantationor chromosomal euploidy as the final outcome. Abbreviations are related with timings in hours: t, timing of cleavage from ICSI until the number of cellsconsidered 2,3,4 etc.; cc, cell cycle duration; s, synchrony of the cell cycle. In the figure, the calculations of each of the variables used in the algorithmsare described graphically.
Diagnosis of human embryo viability 733

aneuploidies were similar in both groups. There was a decrease in thepercentage of abnormal embryos compared with cleavage stageembryos that could be mostly attributed to the arrest of embryos withchaotic pattern and complex abnormalities before reaching blastocyststage. Focusing on clinical results, pregnancy rates per transfer and im-plantation rates in women ,38 years were similar to those obtainedin cleavage stage biopsies, at 54.3% and 42.4%, respectively (Rubioet al., unpublished). More recently, PGS by CGH array with singleeuploid blastocyst transfer appears to be a successful strategy for patientswith multiple failed IVF attempts (Greco et al., 2014).
Moreover, CCS using CGH arrays has been also applied in good prog-nosis patients with a high potential to increase overall pregnancy rates inIVF programs and to decrease multiple pregnancies when SET is per-formed. The first RCT comparing SET at blastocyst stage with andwithout CGH arrays in good prognosis patients showed an aneuploidyrate of 44.9% among biopsied blastocysts, with a significantly higher clin-ical pregnancy rate in the CCS group (70.9 versus 45.8%, P ¼ 0.017).There were no twin pregnancies. This study revealed the limitations ofSET when conventional morphology was used alone, even in patientswithout an increased risk for aneuploidy, as the CGH arrays groupimplanted with greater efficiency and yielded a lower miscarriage ratethan those selected without CGH arrays (Yang et al., 2012).
In summary, the incorporation of CGH arrays appears to result in anincrease in pregnancy and implantation rates, showing that aneuploidiesfor any of the 24 chromosomes can occur in preimplantation humanembryos, and therefore can impair embryo viability and implantation.Female age is the factor with a higher impact on the incidence of
aneuploidies, showing that aneuploidies are the main cause of reproduct-ive failure with advancing female age. CCS with CGH arrays is a robustand highly effective approach for the assessment of embryo viability,either on D3 embryos or at blastocyst stage.
SNP arrays, qPCR and NGSSNP arrays allow for both genotyping and copy number predictions atthousands of positions in the genome. Some SNP array-based CCSmethods rely upon the genotypes alone (Handyside et al., 2010;Johnson et al., 2010a,b), whereas others involve quantitative analysis ofcopy number assignments (Treff et al., 2010a; Konings et al., 2012; vanUum et al., 2012; Tan et al., 2013). Typically, the signals at each positionare compared between the embryo biopsy and those obtained fromknown normal samples in order to identify possible imbalances in theembryo (Fig. 3). One advantage of SNP arrays is the ability to characterizecopy number neutral events, such as recombination sites (Handysideet al., 2010), uniparental disomy (Gueye et al., 2014), parental origin ofaneuploidy (Handyside et al., 2010; Rabinowitz et al., 2012) and balancedtranslocations (Treff et al., 2011b). However, like array CGH, disadvan-tages include the time to obtain a result and the expense of the proced-ure, both of which have been overcome by the development ofqPCR-based CCS (Treff et al., 2012a).
One reason qPCR is faster and cheaper is the elimination of the needfor WGA. Instead, a multiplex PCR reaction is performed to preamplify96 copy number neutral positions in the genome, four per chromosome.Each of the 96 positions are then quantified in individual reactions using
Figure 2 Work-flow and different types of chromosomal abnormalities detected by comparative genome hybridization arrays.
734 Gardner et al.

TaqMan primers and fluorescent target sequence specific probes on a384 well plate, followed by normalization to previous data from knownnormal samples (Fig. 3). The process can be completed in 4 h, requiresthe least hands on time, is easily automatable, involves relatively lowtech equipment and is one-third the cost of array based methods.Another advantage is the ease by which additional primers can be incor-porated in order to characterize single gene disorders (Treff et al.,2013a), small duplications and deletions (Treff et al., 2011b), mitochon-drial disorders (Treff et al., 2012b) or unbalanced translocations (Treffet al., 2013c).
Perhaps, the most ideal combination of throughput, cost and capabilityis afforded by NGS-based CCS. While the approach can involve WGA(Baslan et al., 2012; Lu et al., 2012; Yin et al., 2013; Zhang et al., 2013;Fiorentino et al., 2014), it is unclear what if any necessity there is todetect imbalances below the resolution of the whole chromosome, asthere is still considerable debate regarding the prevalence of de novodele-tions and duplications at the preimplantation stage of development. Thisis particularly true given that a large percentage of ‘dup/dels’ observed in
the preimplantation embryo are likely artifacts of the procedures used toidentify them (Van der Aa et al., 2013; Voet et al., 2013). Alternatively,targeted amplification of specific loci in the genome can also be usedprior to NGS-based CCS and may provide the most cost effective meth-odology (Treff et al., 2013c). In either case, the amplified DNA under-goes massively parallel sequencing in order to count the number ofsequence ‘reads’ which align to each of the 24 chromosomes. Theread counts are then normalized to data from known normal samplesin order to define the copy numbers present in the embryo biopsy(Fig. 3). Reduced cost per embryo is provided by the opportunity toperform molecular barcoding, which allows multiple samples to besequenced in parallel in the same reaction and then segregated backout to the original sample using standard bioinformatics methods(Knapp et al., 2012).
For all methods, mosaicism is of course an important variable toconsider, particularly when evaluating consistency of diagnoses frommultiple samples from the same embryo (Taylor et al., 2014). The mis-guided interpretation of discrepant results as true mosaicism instead of
Figure 3 Representation of three contemporary methods of comprehensive chromosome screening (CCS). Quantitative single nucleotide polymorph-ism (SNP) arrays can involvewhole genome amplification (WGA), followed by labeling and hybridization to an array of synthetic oligonucleotides represent-ing thousands of SNPs in the genome. Embryonic DNA profiles arecompared with knownnormal samples in order to characterize relative quantities of eachchromosomeand makea CCS diagnosis. A mupltiplex PCR reaction for four loci perchromosome arepreamplified in a quantitative real-time (q)PCR-basedCCS approach. Individual loci are then quantified in a 384-well plate format using TaqMan fluorescent probes. Normalization to known normal samples isthen performed in order to obtain an embryo CCS diagnosis. Finally, next-generation sequencing (NGS) can involve either WGA or targeted multiplex PCRfollowed by sequencing and determine relative read counts on each chromosome (Chr) for an embryo CCS diagnosis against known normal samples.
Diagnosis of human embryo viability 735

technical artifact has largely overestimated the true frequency of preim-plantation embryonic mosaicism, particularly when estimates weremade using FISH (Northrop et al., 2010; Treff et al., 2010b). As aresult of the well-established technical and clinical limitations of FISH(Mastenbroek et al., 2011a, b), its use to confirm findings of a newmethod of CCS should not be considered a sufficient level of validation(Gutierrez-Mateo et al., 2011; Mir et al., 2013). One way in which toprevent true mosaicism from impacting the evaluation of the technicalconsistency of a new method of CCS is to evaluate rebiopsies ofembryos that had already been given two consistent diagnoses fromtwo separate biopsies using an alternative CCS methodology (Treffet al., 2012a). A number of recent studies have investigated the frequencyand distribution of mosaicism within the blastocyst using contemporarymethods of CCS. For example, Capalbo et al. (2013a, b) found a fre-quency of 15.7% mosaicism in 70 blastocysts and no evidence of prefer-ential segregation of aneuploidy in the inner cell mass or trophectoderm.Johnson et al. (2010a, b) observed 20% mosaicism within 10 blastocystsoriginally diagnosed as aneuploid and again no evidence of preferentialsegregation to either lineage. Fragouli et al. (2011) found 32.4% mosai-cism among 19 blastocysts, and Northrop et al. (2010) found 24% mosai-cism among 50 blastocysts, both also demonstrating a lack of preferentialsegregation.
Not only is it important to develop more rigorous control for and in-cidence estimates of mosaicism but also it is necessary to consider thesensitivity to detection when CCS is performed on multicell trophecto-derm biopsies. This question has been evaluated by preparing artificialmixtures of two cell lines, one aneuploid and one euploid, to determinewhat level of aneuploidy is necessary to allow detection. One studyinvolved the use of array CGH and indicated a sensitivity to detectionat 50% aneuploid:euploid ratios (Mamas et al., 2012). Another studyinvolved the use of SNP arrays and found that 40% aneuploid:euploidratios were sufficient for detection (Northrop et al., 2010). A variety ofNGS-based CCS methods under development are likely to involvesimilar studies as it is anticipated that this technology will furtherimprove sensitivity to detection of mosaic biopsies.
While mosaicism represents an important variable to considerduring preclinical development, it also represents an important chal-lenge to the clinical application of CCS. By definition, it is impossiblefor any CCS method to accurately diagnose a mosaic embryo sincethe outcome of CCS is to diagnose the embryo as either normal or an-euploid. In a mosaic embryo, there may in fact be both normal and an-euploid cells. The question of how critical mosiacism is to the predictivevalue of CCS can only be adequately addressed through the study ofclinical outcomes of embryos diagnosed after they have already beenselected for transfer using standard morphology without regard tothe CCS diagnosis (Scott et al., 2012), and only then is it possible todetermine whether the CCS diagnosis would have made the correctprediction for the actual clinical outcomes. This is particularly criticalto define the false-positive rate of aneuploidy diagnosis, as the standardRCT is incapable of defining it. The risk of using a method that has notundergone such an evaluation is discarding embryos with true repro-ductive potential. This risk becomes more relevant when applied topatients of AMA or with a single gene disorder indication, wherealready less genetically normal embryos will be available for transfer.In the only reported ‘nonselection’ study to date, the predictive valueof an aneuploid diagnosis for a negative clinical outcome was 96%(using a specific SNP array based CCS approach) demonstrating
minimal impact of mosaicism and suggesting the safety of applyingthis approach to the selection of embryos for transfer (Scott et al.,2012).
Debate also remains regarding the optimum timing of embryo biopsy,not only for the safety of the procedure, but also for the predictive valueof CCS for the ultimate clinical outcome. For example, evidence suggeststhat blastomere biopsy reduces the reproductive potential of theembryo (Scott et al., 2013a) and may be one of the most genetically un-stable stages of embryonic development (Vanneste et al., 2009; Fragouliet al., 2013). This has now led to a growth in either polar body or troph-ectoderm based CCS. Proponents of polar body screening often citeeliminating the impact of mosaicism, that maternal meiosis is theprimary origin of aneuploidy, a reduced impact of biopsy, and govern-ment regulations as the primary factors favoring a polar body approach(Montag et al., 2009; Geraedts et al., 2010). In contrast, proponents oftrophectoderm screening suggest the opportunity to detect all originsof aneuploidy, the demonstrated safety of the procedure, multipleRCTs demonstrating clinical efficacy, and reduced cost to the laboratoryand the patient as favorable factors (Kokkali et al., 2007; Forman et al.,2012; Capalbo et al., 2013a, b). Some even maintain that blastomerebiopsy remains a viable approach for patients that fail to produce blasto-cysts making it difficult for a consensus to be reached at the present time(Wilton, 2007; Rubio et al., 2013).
Preclinical accuracy and safety of the procedure represent importantcomponents of new CCS technologies. Retrospective studies may alsobe useful but only as a preliminary analysis to justify the endeavor of anappropriately designed RCT. There are now at least four reportedCCS RCTs across three laboratories and three different CCS technolo-gies, all of which involved testing at the blastocyst stage of development(Schoolcraftet al., 2012; Yanget al., 2012; Forman et al., 2013; Scott et al.,2013b). In all cases, a clear clinical benefit was demonstrated, validatingthe strategy of aneuploidy screening to improve the efficiency of IVF.Although other studies using polar bodies or blastomeres are forth-coming, the success with, and emphasis on, trophectoderm biopsymay mark the beginning of a paradigm shift to this stage of analysis.The impact of CCS on IVF outcomes has been assessed throughseveral RCTs, as listed in Table II.
Despite rapid developments made in increasing the sensitivity andaccuracy of the technologies involved in CCS, together with thegrowing number of trials showing their efficacy, there are still thosewho question the validity of this approach (Gleicher et al., 2014; Mas-tenbroek and Repping, 2014). Future studies will undoubtedlyconfirm which patient populations may or may not benefit fromCCS-based embryo selection.
What is clear is that not all euploid blastocysts result in a heathly new-born. Many fail to implant and progress, indicating that additional factorsare important to the reproductive potential of the embryo. Determiningthe role of the embryonic ‘omics’ or noninvasive predictions obtainedby using the media, cumulus cells or time-lapse imaging, in identifyingbiomarkers is the next challenge. However, the lessons learned fromthe development and application of new methods of CCS in IVF shouldatminimumprovideaframeworkforappropriatevalidationofnewembryoselection technologies prior to routine utilization (Scott and Treff, 2010).Furthermore, appropriately designed studies to develop new biomarkersof reproductive potential should include control over the presence of an-euploidy within cases and controls, as this is one of the most well provenfactors influencing the reproductive potential of the embryo.
736 Gardner et al.

..........................................................................................................................................................................................................................................................
Table II RCTs using comprehensive chromosome screening.
Authors Female age(years)
Intervention Eligibility No. ofcycles
% Abnormalembryos
Ongoing PR/cycle ordelivery rates
MR
Yang et al.,Mol Cytogenet 2012
,35 SET after blastocyst biopsy versusblastocyst transfer(Array CGH)
Young good prognosis, IVF patients,first cycle, no prior miscarriage
55 PGS48 control
44.9% 69.1 PGS versus41.7 (P ¼ 0.0009)
2.6 PGS versus9.1(NS)
Forman et al.,Fertil Steril 2013NCT01408433
,43 SET after blastocyst biopsy versusDET of unscreened blastocysts(qPCR)
All indications≥2 blastocyst for biopsy
89 PGS86 control
31% 60.7 PGS versus 65.1(NS)
11.5 PGS versus20.0(NS)
Scott et al.,2013a, bNCT01219283
21–42 Blastocyst biopsy versus blastocysttransfer
All indications≤1 failed IVF
72 PGS83 control
28.6% 84.7 PGS versus 67.5(P ¼ 0.01)
–
Schoolcraft et al.,ASRM 2012
.35 Fresh blastocyst transfer versusfrozen blastocyst biopsy(SNP microarray)
AMA 47 PGS41 control
– 74.5 PGS versus 53.7(P , 0.05)
–
Rubio et al.,ESHRE 2014NCT01571076
38–41 D3 biopsy with blastocyst transferversus blastocyst transfer(Array CGH)
AMA,2 miscarriages,2 IVF failures
75 PGS86 control
77.9% 42.7 PGS versus 25.6(P ¼ 0.0294)
3.3 PGS versus43.6(P , 0.0001)
ESHRE Study for OocyteEuploidy (ESTEEM)NCT01532284
36–41 Polar body biopsy(Array CGH)
AMARecruiting
Yilun Siu and Shangai Ji AiGenetics & IVF InstituteNCT02223221
18–35 Blastocyst biopsy versus blastocysttransfer(Array CGH)
RPL ≥3 miscarriagesRecruiting
RubioIVINCT01571076
,38 D3 biopsy with blastocyst transferversus blastocyst transfer(Array CGH)
Severe male factor,2 million sperm/mlRecruiting
MunneReprogeneticsNCT01946945
22–42 Blastocyst biopsy versus blastocysttransfer(NGS)
All indicationsRecruiting
ScottRMANJNCT02032264
18–42 DET blastocyst biopsy (NGS) ≤1 prior failed IVFRecruiting
CGH, comparative genomic hybridization; SNP, single nucleotide polymorphism; SET, single-embryo transfer; qPCR, quantitative PCR; NGS, next-generation sequencing; PGS, preimplantation genetic screening; AMA, advanced maternal age; RPL,recurrent pregnancy loss; MR, miscarriage rate.
Diagnosis
ofhuman
embryo
viability737

Analysis of physiology and thesearch for biomarkersViability, defined as the ability of an embryo to implant and give rise to ahealthy baby, varies even among euploid embryos, with factors otherthan chromosome complement reflecting the inherent developmentalpotential of the embryo. Hence the term ‘biomarker’ refers to those pro-teins and metabolites that are associated, either positively or negatively,with pregnancy outcome. Morphology has been used extensively forover three decades now, and through the development of elegantscoring systems for the pronucleate oocyte (Scott, 2003), cleavage(Van Royen et al., 1999) and blastocyst stage (Gardner et al., 2000)embryos, we already have useful metrics on which to base our decisionsregarding which embryo has the highest developmental potential, andwith the advent of robust and effective time-lapse systems, furtherdata are emerging to create/augment selection algorithms. Usingembryo-grading systems has clearly assisted in improving transferoutcome, but assessment of morphology alone is not absolute anddoes not necessarily reflect the physiological status of the embryo. Forexample, it has been established for many years that embryos of thesame alphanumeric grade, from the same patient, can differ enormouslyin their proteomes (Katz-Jaffe et al., 2006a, b) and metabolic activity(Gardner et al., 2001). Clearly, it is time to consider how such analysescan be incorporated into the embryo selection process.
ProteomicsThe proteome is estimated to consistof overa million proteins, and beingderived from the transcriptome, is responsible for cell function. Whereasthe analysis of the proteome requires cellular extraction, it is feasible toanalyse a large number of proteins and peptides secreted by an individualhuman embryo, the secretome, noninvasively by sampling the surround-ing culture medium. Such analyses have become possible with the adventof novel mass spectrometry platforms, such as surface-enhanced laserdesorption/ionization time-of-flight mass spectrometry (SELDI-TOFMS) (Katz-Jaffe et al., 2005; Katz-Jaffe et al., 2006a, b), and electrosprayMS (Beardsley et al., 2010) capable of screening spent embryo culturemedia for biomarkers. Using SELDI-TOF, Katz-Jaffe et al. (2006a, b)were able to identify an 8.5-kDa protein whose abundance increasedonly in the secretome of developing blastocysts, indicating a potential re-lationship between this protein and developmental capability. Proteinisolation and identification by tandem MS with peptide sequencingrevealed the best candidate for this 8.5-kDa protein biomarker to be ubi-quitin. Ubiquitin has been implicated in the implantation process duringmammalian development through the control of the activities and turn-over of key signaling molecules. Other approaches for the analysis of thesecretome have included the use of protein microarrays containing 120antibody targets (Dominguez et al., 2008). Similarly, a targeted proteo-mics platform, Luminex, has provided great insight into the proteomeof the fluids of the female reproductive tract (Hannan et al., 2011).Could such proteomic analyses be of value in the diagnosis of embryoploidy, given that the proteome reflects the transcriptome? A pioneeringstudy by McReynolds et al. (2011) has revealed that the presence oflipocalin-1 is increased in the secretome of aneuploid human blastocysts,data obtained through MS and subsequently confirmed through enzyme-linked immunosorbent assay. Whether it is possible or indeed desirableto analyze the karyotype of an embryo through its secretome remains to
be resolved. What is evident however, is that differences in the prote-ome exist due to difference in gene expression between male andfemale preimplantation embryos (Epstein et al., 1978; Kobayashi et al.,2006; Bermejo-Alvarez et al., 2011), which will reflect differences inphysiology (Gardner et al., 2010, 2011).
It is evident that proteomics is currently providing a unique insight intoembryo (and endometrial) physiology and signaling (Hannan et al., 2011;Binderet al., 2014). Despite of its promise however, the analysis of the em-bryonic secretome has yet to be adopted clinically. A key determinant forthis is the sheer cost of MS platforms and that further basic research isrequired to further identify peptides/proteins that reflect embryo viabilityand ploidy. Once these biomarkers have been identified, then adopting atargetedproteomicapproachbecomes feasible.Withrapiddevelopmentsin the field ofmicrofluidics, it is fair toassumethat itwill become possible toperform quantitative and targeted proteomics within an IVF laboratory.
MetabolomicsDefined as the systematic study of the unique chemical footprints thatspecific cellular processes leave behind, metabolomics is able to charac-terize typically thousands of metabolites (Brison et al., 2007; Singh andSinclair, 2007; Botros et al., 2008). Depending upon the platform tech-nology used, it is feasible to both identify and quantify metabolites ofinterest. Quantitative platforms include RAMAN (Seli et al., 2007) and1H nuclear magnetic resonance (Seli et al., 2008), but the limitation ofsensitivities of these technologies means that although they are powerfulresearch tools in human reproductive biology (Gook et al., 2014),working with individual embryos is not feasible. More sensitive is NearInfrared (NIR) spectroscopy, capable of providing detailed spectra ofspent media from a single embryo (Seli et al., 2011). However, althoughsuch technologies have been used to create algorithms associated withembryo viability, NIR cannot identify nor quantify specific nutrients, al-though this latter characteristic should be of relatively small concern ifthis approach to embryo selection can be shown to provide statisticalincreases in implantation rates. Studies using NIR initially created a rela-tive ‘embryo viability score’,which was intended to reflect embryo devel-opmental potential. It was subsequently determined that this score didindeed correlate to both positive and negative implantation outcomes.Interestingly, when human embryos of similar morphology were exam-ined using the same NIR spectral profile their viability scores varied re-markably in relation to morphology, indicating that the metabolome ofembryos was not necessarily reflective of embryo score (Vergouwet al., 2008). These data are consistent with those of Katz-Jaffe et al.(2006a, b) who determined that the proteome of individual human blas-tocystsof the same grade differed, confirming thatembryo morphology isnot completely linked to its physiology. To date, however, the algorithmscreated using NIR have not been proven in prospective trials (Ahlstromet al., 2011; Hardarson et al., 2012; Vergouw et al., 2012). One of themain issues encountered with the NIR system was that the thresholdof signal distinguishing between a viable and nonviable embryo was sus-ceptible to signal noise. NIR spectroscopy systems and the algorithmsgenerated from them can create models that inadvertently conceal pro-blems in a particular platform. Consequently, a method established andcross-validated on a larger scale can be problematic as the variation canlie within the technical platform itself. Hence, if these problems can bedealt with successfully, it is feasible that NIR could still have a role toplay in embryo selection.
738 Gardner et al.

More recently, two other MS approaches have been piloted to analyzespent human embryo culture media; electrospray ionization mass spec-trometry (ESI-MS) (Cortezzi et al., 2013) and direct injection (DI)-MS(Sheedy et al., 2014). Both represent a fast analytical approach, requiringminimal sample preparation, and the ability to produce large amounts ofdata from culture media obtained from a single embryo. Such approachesmay yet serve as valuable tools in the identification of novel biomarkers.
Targeted metabolomicsAn approach that has been used for over three decades for the advance-ment of assisted reproductive techniques (ART) is referred to here as‘Targeted Metabolomics’, where one or several known metabolitesused by the embryo can be quantified noninvasively. Examples of thisinclude glucose and one of its metabolites lactate, and amino acids to-gether with ammonium. However, unlike genetic diagnosis, which deter-mines whether a specific gene is normal, or whether the correctchromosome complement is present, an accurate diagnosis of embryoviability is confounded by the fact that the proteome/secretome andconsequently physiology and metabolome can be directly affected bythe conditions used to sustain embryo development in the laboratory.Furthermore, there are growing data on differences between male andfemale embryos during the preimplantation period. Therefore, if anembryo is analyzed under one set of conditions, it is plausible that asthe surrounding conditions change, so too do the parameters that aremeasured. Factors affecting the metabolic profile of the humanembryo include: embryonic stage of development, sex of the embryo,medium composition (i.e. differences in nutrient concentrations,albumin source), frequency of medium renewal, rate of ammonium accu-mulation and oxygen concentration. Perhaps, one of the most extensive-ly studied factors that regulate embryonic function is oxygen, having beenshown to affect gene expression, the proteome and metabolic activity ofthe mammalian embryo. In a series of studies, Wale and Gardner (2012)revealed that atmospheric oxygen exhibited a bi-phasic effect on aminoacid utilization by the mouse embryo, enhancing turnover during thecleavage stages but reducing their turnover post-compaction. Further-more, atmospheric oxygen was found to inhibit glucose uptake by theblastocyst (Wale and Gardner, 2012). Strikingly, when atmosphericoxygen was used in the presence of ammonium in the medium, thepattern of amino acid metabolism was dramatically altered (Wale andGardner, 2013). Consequently, biomarker levels are affected understress, a finding that warrants further investigation prior to the routine ap-plication of such technologies, and certainly no single overarching hy-pothesis of embryo metabolism can be applied to all stages and allculture conditions (Gardner and Wale, 2013).
To date, only a handful of studies have been performed on carbohy-drate utilization and subsequent viability of the human embryo. In a retro-spective analysis Conaghan et al. (1993) observed an inverse relationshipbetween pyruvate uptake by 2- to 8-cell embryos cultured in the pres-ence of serum and 20% oxygen and subsequent pregnancy. In a studyon human morulae and blastocysts with different degrees of expansion,no conclusive data were generated on the ability of nutrient consumptionor utilization to predict pregnancy outcome (Jones et al., 2001).However, in both studies the medium used to assess embryo metabol-ism was a simple one, lacking lactate, amino acids and vitamins. Conse-quently, embryos analyzed under such conditions could be expectedto be experiencing considerable metabolic stress.
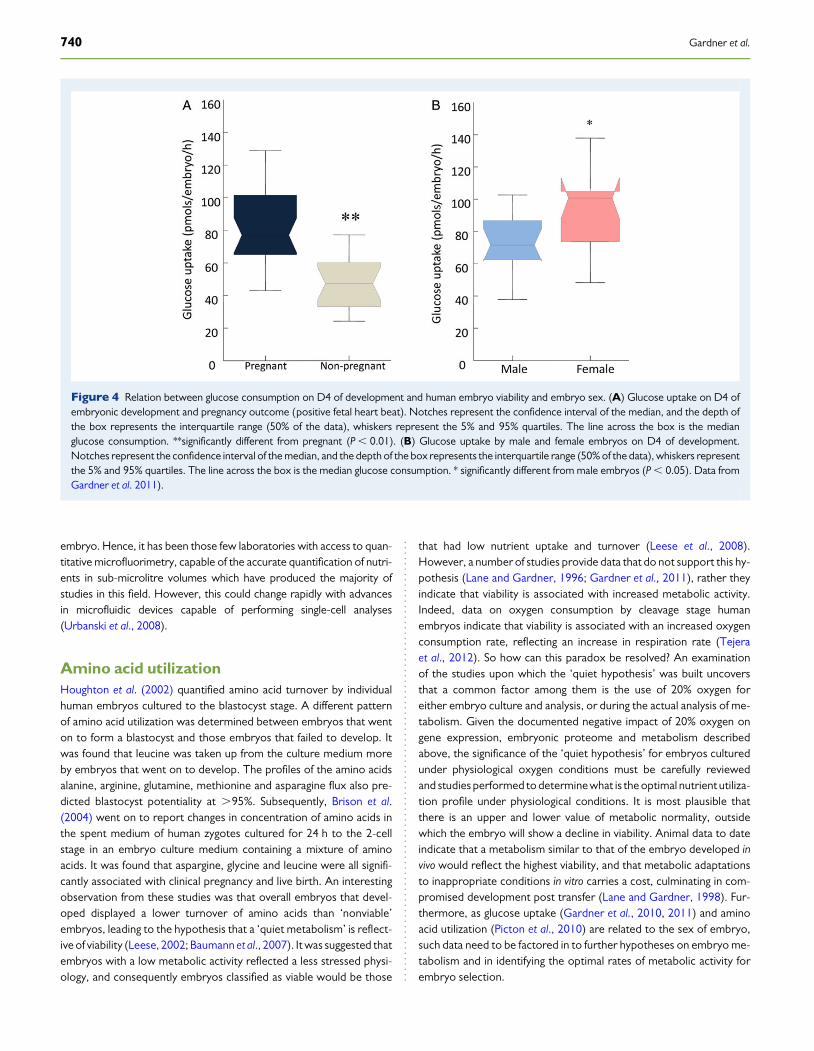
Glucose utilizationA relationship between metabolic activity and embryo development andsubsequent viability has been established over several decades. As earlyas 1970, Menke and McLaren (1970) observed that mouse blastocysts,cultured in a basic medium (lacking amino acids) lost their ability tooxidize glucose. This initial observation was followed by several studiesthat elucidated that changes in embryo metabolism were associatedwith loss of developmental capacity in vitro. In 1980, Renard et al. (1980)determined that D10 cattle blastocysts which consumed glucose at arate greater than 5 mg/h developed better in culture and in vivo aftertransfer than those blastocysts with a glucose uptake below this value. In1987, using noninvasive microfluorescence, Gardner and Leese (1987)went on to measure glucose uptake by individual D4 mouse blastocystsprior to transfer to recipient females. Those embryos that went to termhad a significantly higher glucose uptake in culture than those embryosthat failed to develop after transfer. Interestingly, if glucose uptake islinked with lactate production, to create an indirect measure of glucoseflux through the glycolytic pathway, it creates a second, and potentiallygreater reflective measure of normalcy. In the mouse model, thisapproach has been used to show that even a transient (6 h) exposureto environmental stress at the blastocyst stage, in this case providedby a medium lacking amino acids and vitamins, results in significantelevations in glycolytic activity culminating in a significant reduction inimplantation and fetal development (weight) post transfer (Lane andGardner, 1998). Encouragingly, the levels of glycolysis at the blastocyststage have been used successfully to select prospectively viable mouseblastocysts for transfer by Lane and Gardner (1996). Morphologicallyidentical mouse blastocysts with equivalent diameters were identifiedusing metabolic criteria, as ‘viable’ prior to transfer and had a fetaldevelopment of 80%. In contrast, those embryos that exhibited an ab-normal metabolic profile (compared with in vivo developed controls),developed at a rate of only 6%. Analysis of the relationship betweenhuman embryo nutrition and development in vitro was undertaken byGardner et al. (2001), who determined that glucose consumption onD4 by human embryos was twice as high in those embryos that wenton to form blastocysts. Subsequently, Gardner et al. (2011) went on toconfirm a positive relationship between glucose uptake and humanembryo viability on D4 and D5 of development (Fig. 4). Furthermore,the data generated indicate differences in nutrient utilization betweenhuman male and female embryos, a phenomenon previously documen-ted in other mammalian species (Tiffin et al., 1991; Gardner et al., 2010).Of further interest was the observation by Gardner et al. (2011) that therelationship between glucose uptake by the blastocyst and pregnancyrate was not tied to the alphanumeric score of the embryo (Gardneret al., 2011). In other words, pregnancy was associated with thebiomarker more than with morphology, a factor considered in theabove sections on morphokinetics and proteomics.
Given the evidence of a relationship between glucose metabolism andblastocyst viability, the question is raised—why has glucose uptake notbeen adopted? At first, it would appear straight forward to quantifythe levels of glucose in embryo culture media, given that glucose is mea-sured routinely in biological fluids in so many areas of medicine. Whereasit is feasible to measure glucose concentration in blood and urine, theissue with regards to culture medium is one of accuracy and sensitivity.Standard approaches lack the ability to accurately measure the small dif-ferences in nutrient concentration that can be attributed to an individual
Diagnosis of human embryo viability 739
embryo. Hence, it has been those few laboratories with access to quan-titative microfluorimetry, capable of the accurate quantification of nutri-ents in sub-microlitre volumes which have produced the majority ofstudies in this field. However, this could change rapidly with advancesin microfluidic devices capable of performing single-cell analyses(Urbanski et al., 2008).
Amino acid utilizationHoughton et al. (2002) quantified amino acid turnover by individualhuman embryos cultured to the blastocyst stage. A different patternof amino acid utilization was determined between embryos that wenton to form a blastocyst and those embryos that failed to develop. Itwas found that leucine was taken up from the culture medium moreby embryos that went on to develop. The profiles of the amino acidsalanine, arginine, glutamine, methionine and asparagine flux also pre-dicted blastocyst potentiality at .95%. Subsequently, Brison et al.(2004) went on to report changes in concentration of amino acids inthe spent medium of human zygotes cultured for 24 h to the 2-cellstage in an embryo culture medium containing a mixture of aminoacids. It was found that aspargine, glycine and leucine were all signifi-cantly associated with clinical pregnancy and live birth. An interestingobservation from these studies was that overall embryos that devel-oped displayed a lower turnover of amino acids than ‘nonviable’embryos, leading to the hypothesis that a ‘quiet metabolism’ is reflect-ive of viability (Leese, 2002; Baumann et al., 2007). It was suggested thatembryos with a low metabolic activity reflected a less stressed physi-ology, and consequently embryos classified as viable would be those
that had low nutrient uptake and turnover (Leese et al., 2008).However, a number of studies provide data that do not support this hy-pothesis (Lane and Gardner, 1996; Gardner et al., 2011), rather theyindicate that viability is associated with increased metabolic activity.Indeed, data on oxygen consumption by cleavage stage humanembryos indicate that viability is associated with an increased oxygenconsumption rate, reflecting an increase in respiration rate (Tejeraet al., 2012). So how can this paradox be resolved? An examinationof the studies upon which the ‘quiet hypothesis’ was built uncoversthat a common factor among them is the use of 20% oxygen foreither embryo culture and analysis, or during the actual analysis of me-tabolism. Given the documented negative impact of 20% oxygen ongene expression, embryonic proteome and metabolism describedabove, the significance of the ‘quiet hypothesis’ for embryos culturedunder physiological oxygen conditions must be carefully reviewedand studies performed to determine what is the optimal nutrient utiliza-tion profile under physiological conditions. It is most plausible thatthere is an upper and lower value of metabolic normality, outsidewhich the embryo will show a decline in viability. Animal data to dateindicate that a metabolism similar to that of the embryo developed invivo would reflect the highest viability, and that metabolic adaptationsto inappropriate conditions in vitro carries a cost, culminating in com-promised development post transfer (Lane and Gardner, 1998). Fur-thermore, as glucose uptake (Gardner et al., 2010, 2011) and aminoacid utilization (Picton et al., 2010) are related to the sex of embryo,such data need to be factored in to further hypotheses on embryo me-tabolism and in identifying the optimal rates of metabolic activity forembryo selection.
Figure 4 Relation between glucose consumption on D4 of development and human embryo viability and embryo sex. (A) Glucose uptake on D4 ofembryonic development and pregnancy outcome (positive fetal heart beat). Notches represent the confidence interval of the median, and the depth ofthe box represents the interquartile range (50% of the data), whiskers represent the 5% and 95% quartiles. The line across the box is the medianglucose consumption. **significantly different from pregnant (P , 0.01). (B) Glucose uptake by male and female embryos on D4 of development.Notches represent the confidence interval of the median, and the depth of the box represents the interquartile range (50% of the data), whiskers representthe 5% and 95% quartiles. The line across the box is the median glucose consumption. * significantly different from male embryos (P , 0.05). Data fromGardner et al. 2011).
740 Gardner et al.

Figure 5 Concept microfluidic device for biomechanical analysis of a preimplantation embryo. A blastomere is first biopsied from a cleavage stageembryo (A). The blastomere is then loaded into the microfluidic device and driven through a channel constriction (B). This causes morphological com-pression (C), from which some basic biomechanical properties can be calculated from curvatures of the rear (a), side (b) and front (d) aspects of theplasma membrane. The same blastomere can then be moved to an actuation zone and lysed (D and E), with subcellular fragments divided by simplechannel branching to other regions for parallel molecular analysis on-chip, or to other off-board platforms (F). From Thouas et al. (2013) (with permissionfrom Springer).
Figure 6 Schematic of an embryo culture system for perfusion culture and analysis of biomarkers. Culture media are continuously passed over theembryo(s). The composition of the culture media can be changed according to the specific requirements of each stage of embryonic development.Toxins, such as ammonium, are not able to build up and impair embryo development, while more labile components of the culture system are not dena-tured. Samples of culture media can be removed for biomarker analysis. Adapted from Gardner (1994).
Diagnosis of human embryo viability 741

When morphokinetics meets metabolismA recent study has considered the relationship between embryo mor-phokinetics and metabolic activity. In the first study of its kind, mouseembryos underwent both time-lapse and metabolic analyses. Embryoshad their rate of development quantified together with the utilizationof glucose and amino acids of the resultant blastocysts. It was determinedthat embryos assigned to the fastest quartile gave rise to blastocysts witha higher glucose uptake and lower glycolytic activity (reflective ofincreased viability) than their counterparts which developed from theslowest quartile (Lee et al., 2015). Furthermore, differences in aminoacid utilization were also detectable, with those blastocysts developingfrom fast embryos exhibiting higher aspartate uptake, which is of signifi-cance for the regulation of the malate-aspartate shuttle in the blastocyst(Mitchell et al., 2009; Gardner and Wale, 2013). Of interest, within eachquartile embryos exhibited a range of metabolic activities, supporting themove to use multiple parameters in diagnosing embryo viability. Thesedata indicate that morphokinetics and targeted metabolomics are com-plementary approaches for the assessment of embryo development andviability.
Laboratory on a chipThe human embryo represents limited biological material for quan-titative analysis, which makes it very attractive for analysis throughmicrofluidic devices, which are capable of moving, mixing andaccurate analysis of sub-microlitre volumes. The potential of such‘Laboratory-on-a-Chip’ devices in reproductive biology has been con-sidered for over a decade, and to date microfluidic devices exist,capable of performing several of the tasks associated with humanART including oocyte denudation (Beebe et al., 2002), sperm sorting(Schuster et al., 2003), IVF and embryo culture (Swain et al., 2013),and analysis of biomarkers (Urbanski et al., 2008; Heo et al., 2012). Al-though currently a single platform to undertake all tasks does not exist,it is envisaged that in the near future such devices will be employed clin-ically for specific tasks. Furthermore, such devices could be used forboth genetic and biomarker analysis (see Fig. 5), given that microfluidicdevices are commercially available for qPCR and gene arrays. As it ispossible to visualize the embryo within such devices, it is envisagedthat morphokinetic data will be collected concomitantly, and that the‘IVF-Lab-on-a-Chip’ will reside within a time-lapse incubation system(Fig. 6).
ConclusionsEdwards et al. (1984) observed that faster cleaving embryos gave rise tomore viable pregnancies, and in an animal model, it was determined thatthere existed a relationship between glucose uptake by blastocysts andsubsequent development post-transfer (Renard et al., 1980). Withdevelopments in time-lapse technologies, together with advances inculture systems, we are now confirming, and advancing, such observa-tions made three decades earlier. Furthermore, together with rapiddevelopments and implementation of new molecular means of CCS,we are creating new paradigms for the way human IVF and embryo diag-nosis are being performed. Additionally, we are on the verge of under-standing what relationships exist between embryo morphokinetics,karyotype, gene expression and physiology, which will help to formulate
comprehensive and robust algorithms for embryo selection (as well asthe development of more refined culture systems). Microfluidicdevices for embryo culture and analysis may yet prove to be a practicalmeans for the integration and application of these new technologies.Combined with developments in vitrification technologies for embryocryopreservation (Vajta, 2013), there is now no longer a need to transferthe embryo back to the uterus in same cycle that the oocytes were col-lected, thereby avoiding a uterus whose receptivity has been compro-mised by exposure to exogenous gonadotrophins during ovarianstimulation (Gardner, 2008; Evans et al., 2012, 2014). Rather, followingtime-lapse monitoring, biopsy for karyotyping and analysis of spentculture media for biomarkers, one could rank each euploid embryo fortransfer in a natural cycle on the appropriate day of receptivity for thepatient (Ruiz-Alonso et al., 2013), thereby significantly reducing thetime to conception.
Authors’ rolesAll four authors contributed equally to the inception and writing of thismanuscript.
FundingNo funding was received for the preparation of this work.
Conflict of interestNone of the authors have any conflict of interest.
ReferencesAguilar J, Motato Y, Escriba MJ, Ojeda M, Munoz E, Meseguer M. The human first cell
cycle: impact on implantation. Reprod Biomed Online 2014;28:475–484.Ahlstrom A, Wikland M, Rogberg L, Barnett JS, Tucker M, Hardarson T.
Cross-validation and predictive value of near-infrared spectroscopy algorithms forday-5 blastocyst transfer. Reprod Biomed Online 2011;22:477–484.
Alfarawati S, Fragouli E, Colls P, Wells D. First births after preimplantation geneticdiagnosis of structural chromosome abnormalities using comparative genomichybridization and microarray analysis. Hum Reprod 2011;26:1560–1574.
Aparicio B, Cruz M, Meseguer M. Is morphokinetic analysis the answer? Reprod BiomedOnline 2013;27:654–663.
Athayde Wirka K, Chen AA, Conaghan J, Ivani K, Gvakharia M, Behr B, Suraj V, Tan L,Shen S. Atypical embryo phenotypes identified by time-lapse microscopy: highprevalence and association with embryo development. Fertil Steril 2014;10:1637–1648.
Azzarello A, Hoest T, Mikkelsen AL. The impact of pronuclei morphology anddynamicity on live birth outcome after time-lapse culture. Hum Reprod 2012;27:2649–2657.
Barnes FL. The effects of early uterine environment on the subsequent development ofembryo and fetus. Theriogenology 2000;53:649–658.
Basile N, Meseguer M. Time-lapse technology: evaluation of embryo quality and newmarkers for embryo selection. Expert Rev Obstet Gynecol 2012;7:175–190.
Basile N, Morbeck D, Garcia-Velasco J, Bronet F, Meseguer M. Type of culture mediadoes not affect embryo kinetics: a time-lapse analysis of sibling oocytes. Hum Reprod2013;28:634–641.
Basile N, Nogales Mdel C, Bronet F, Florensa M, Riqueiros M, Rodrigo L,Garcia-Velasco J, Meseguer M. Increasing the probability of selectingchromosomally normal embryos by time-lapse morphokinetics analysis. Fertil Steril2014;101:699–704.
Baslan T, Kendall J, Rodgers L, Cox H, Riggs M, Stepansky A, Troge J, Ravi K, Esposito D,Lakshmi B et al. Genome-wide copy number analysis of single cells. Nat Protoc 2012;7:1024–1041.
742 Gardner et al.

Baumann CG, Morris DG, Sreenan JM, Leese HJ. The quiet embryo hypothesis:molecular characteristics favoring viability. Mol Reprod Dev 2007;74:1345–1353.
Baxter Bendus AE, Mayer JF, Shipley SK, Catherino WH. Interobserver andintraobserver variation in day 3 embryo grading. Fertil Steril 2006;86:1608–1615.
Beardsley AJ, Li Y, O’Neill C. Characterization of a diverse secretome generated by themouse preimplantation embryo in vitro. Reprod Biol Endocrinol 2010;8:71.
Beebe D, Wheeler M, Zeringue H, Walters E, Raty S. Microfluidic technology forassisted reproduction. Theriogenology 2002;57:125–135.
Bellver J, Mifsud A, Grau N, Privitera L, Meseguer M. Similar morphokinetic patterns inembryos derived from obese and normoweight infertile women: a time-lapse study.Hum Reprod 2013;28:794–800.
Bermejo-Alvarez P, Rizos D, Lonergan P, Gutierrez-Adan A. Transcriptional sexualdimorphism during preimplantation embryo development and its consequencesfor developmental competence and adult health and disease. Reproduction 2011;141:563–570.
Binder NK, Evans J, Gardner DK, Salamonsen LA, Hannan NJ. Endometrial signalsimprove embryo outcome: functional role of VEGF isoforms on embryodevelopment and implantation. Hum Reprod 2014;29:2278–2286.
Botros L, Sakkas D, Seli E. Metabolomics and its application for non-invasive embryoassessment in IVF. Mol Hum Reprod 2008;14:679–690.
Boue A, Boue J, Gropp A. Cytogenetics of pregnancy wastage. Adv Hum Gen 1985;14:1–57.
Brison DR, Houghton FD, Falconer D, Roberts SA, Hawkhead J, Humpherson PG,Lieberman BA, Leese HJ. Identification of viable embryos in IVF by non-invasivemeasurement of amino acid turnover. Hum Reprod 2004;19:2319–2324.
Brison DR, Hollywood K, Arnesen R, Goodacre R. Predicting human embryo viability:the road to non-invasive analysis of the secretome using metabolic footprinting.Reprod Biomed Online 2007;15:296–302.
Campbell A, Fishel S, Bowman N, Duffy S, Sedler M, Hickman CF. Modelling a riskclassification of aneuploidy in human embryos using non-invasive morphokinetics.Reprod Biomed Online 2013a;26:477–485.
Campbell A, Fishel S, Bowman N, Duffy S, Sedler M, Thornton S. Retrospective analysisof outcomes after IVF using an aneuploidy risk model derived from time-lapseimaging without PGS. Reprod Biomed Online 2013b;27:140–146.
Capalbo A, Bono S, Spizzichino L, Biricik A, Baldi M, Colamaria S, Ubaldi FM, Rienzi L,Fiorentino F. Sequential comprehensive chromosome analysis on polar bodies,blastomeres and trophoblast: insights into female meiotic errors andchromosomal segregation in the preimplantation window of embryodevelopment. Hum Reprod 2013a;28:509–518.
Capalbo A, Wright G, Elliott T, Ubaldi FM, Rienzi L, Nagy ZP. FISH reanalysis of innercell mass and trophectoderm samples of previouslyarray-CGH screened blastocystsshows high accuracy of diagnosis and no major diagnostic impact of mosaicism at theblastocyst stage. Hum Reprod 2013b;28:2298–2307.
Chamayou S, Ragolia C, Alecci C, Storaci G, Maglia E, Russo E, Guglielmino A. Meioticspindle presence and oocyte morphology do not predict clinical ICSI outcomes: astudy of 967 transferred embryos. Reprod Biomed Online 2006;13:661–667.
Chamayou S, Patrizio P, Storaci G, Tomaselli V, Alecci C, Ragolia C, Crescenzo C,Guglielmino A. The use of morphokinetic parameters to select all embryos withfull capacity to implant. J Assist Reprod Genet 2013;30:703–710.
Chavez SL, Loewke KE, Han J, Moussavi F, Colls P, Munne S, Behr B, Reijo Pera RA.Dynamic blastomere behaviour reflects human embryo ploidy by the four-cellstage. Nat Commun 2012;3:1251.
Chen AA, Tan L, Suraj V, Reijo Pera R, Shen S. Biomarkers identified with time-lapseimaging: discovery, validation, and practical application. Fertil Steril 2013;99:1035–1043.
Ciray HN, Aksoy T, Goktas C, Ozturk B, Bahceci M. Time-lapse evaluation of humanembryo development in single versus sequential culture media--a sibling oocytestudy. J Assist Reprod Genet 2012;29:891–900.
Conaghan J, Hardy K, Handyside AH, Winston RM, Leese HJ. Selection criteria forhuman embryo transfer: a comparison of pyruvate uptake and morphology.J Assist Reprod Genet 1993;10:21–30.
Conaghan J, Chen AA, Willman SP, Ivani K, Chenette PE, Boostanfar R, Baker VL,Adamson GD, Abusief ME, Gvakharia M et al. Improving embryo selectionusing a computer-automated time-lapse image analysis test plus day 3morphology: results from a prospective multicenter trial. Fertil Steril 2013;100:412–419.
Cortezzi SS, Cabral EC, Trevisan MG, Ferreira CR, Setti AS, Braga DP, Figueira Rde C,Iaconelli A Jr, Eberlin MN, Borges E Jr. Prediction of embryo implantation potential by
mass spectrometry fingerprinting of the culture medium. Reproduction 2013;145:453–462.
Cruz M, Garrido N, Herrero J, Perez-Cano I, Munoz M, Meseguer M. Timing of celldivision in human cleavage-stage embryos is linked with blastocyst formation andquality. Reprod Biomed Online 2012;25:371–381.
Cruz M, Garrido N, Gadea B, Munoz M, Perez-Cano I, Meseguer M. Oocyteinsemination techniques are related to alterations of embryo developmentaltiming in an oocyte donation model. Reprod Biomed Online 2013;27:367–375.
Dal Canto M, Coticchio G, Mignini Renzini M, De Ponti E, Novara PV, Brambillasca F,Comi R, Fadini R. Cleavage kinetics analysis of human embryos predictsdevelopment to blastocyst and implantation. Reprod Biomed Online 2012;25:474–480.
Davies S, Christopikou D, Tsorva E, Karagianni A, Handyside AH, Mastrominas M.Delayed cleavage division and a prolonged transition between 2- and 4-cell stagesin embryos identified as aneuploidy at the 8-cell stage by array-CGH. Hum Reprod2012;27:ii84–ii86.
Dominguez F, Gadea B, Esteban FJ, Horcajadas JA, Pellicer A, Simon C. Comparativeprotein-profile analysis of implanted versus non-implanted human blastocysts.Hum Reprod 2008;23:1993–2000.
Edwards RG, Fishel SB, Cohen J, Fehilly CB, Purdy JM, Slater JM, Steptoe PC,Webster JM. Factors influencing the success of in vitro fertilization for alleviatinghuman infertility. J In Vitro Fert Embryo Transf 1984;1:3–23.
Epstein CJ, Smith S, Travis B, Tucker G. Both X chromosomes function beforevisible X-chromosome inactivation in female mouse embryos. Nature 1978;274:500–503.
Ergin EG, Caliskan E, Yalcinkaya E, Oztel Z, Cokelez K, Ozay A, Ozornek HM.Frequency of embryo multinucleation detected by time-lapse system and itsimpact on pregnancy outcome. Fertil Steril 2014;102:1029–1033.
Evans J, Hannan NJ, Hincks C, Rombauts LJF, Salamonsen LA. Defectove soil for a fertileseed? Altered endometrial development is detrimental to pregnancy success. PLoSOne 2012;7:e53098.
Evans J, Hannan NJ, Edgell TA, Vollenhoven BJ, Lutjen PJ, Osianlis T, Salamonsen LA,Rombauts LJF. Fresh versus frozen embryo transfer: backing clinical decisions withscientific and clinical evidence. Hum Reprod Update 2014;20:800–821.
Fiorentino F, Biricik A, Bono S, Spizzichino L, Cotroneo E, Cottone G, Kokocinski F,Michel CE. Development and validation of a next-generation sequencing-basedprotocol for 24-chromosome aneuploidy screening of embryos. Fertil Steril 2014;101:1375–1382 e1372.
Forman EJ, Hong KH, Treff NR, Scott RT. Comprehensive chromosome screening andembryo selection: moving toward single euploid blastocyst transfer. Semin ReprodMed 2012;30:236–242.
Forman EJ, Hong KH, Ferry KM, Tao X, Taylor D, Levy B, Treff NR, Scott RT Jr. In vitrofertilization with single euploid blastocyst transfer: a randomized controlled trial.Fertil Steril 2013;100:100–107 e101.
Fragouli E, Alfarawati S, Daphnis DD, Goodall NN, Mania A, Griffiths T, Gordon A,Wells D. Cytogenetic analysis of human blastocysts with the use of FISH, CGHand aCGH: scientific data and technical evaluation. Hum Reprod 2011;26:480–490.
Fragouli E, Alfarawati S, Spath K, Jaroudi S, Sarasa J, Enciso M, Wells D. The origin andimpact of embryonic aneuploidy. Hum Genet 2013;132:1001–1013.
Freour T, Dessolle L, Lammers J, Lattes S, Barriere P. Comparison of embryomorphokinetics after in vitro fertilization-intracytoplasmic sperm injection insmoking and nonsmoking women. Fertil Steril 2013;99:1944–1950.
Gardner DK. Mammalian embryo culture in the absence of serum or somatic cellsupport. Cell Biol Int 1994;18:1163–1179.
Gardner DK. Blastocyst transfer. In: Rizk BRMB, Garcia-Velasco JA, Sallam HN,Makrigiannakis A (eds). Infertility and Assisted Reproduction. Cambridge: CambridgeUniversity Press, 2008, 531–539.
Gardner DK, Leese HJ. Assessment of embryo viability prior to transfer by thenoninvasive measurement of glucose uptake. J Exp Zool 1987;242:103–105.
Gardner DK, Wale PL. Analysis of metabolism to select viable human embryos fortransfer. Fertil Steril 2013;99:1062–1072.
Gardner DK, Lane M, Stevens J, Schlenker T, Schoolcraft WB. Blastocyst score affectsimplantation and pregnancy outcome: towards a single blastocyst transfer. Fertil Steril2000;73:1155–1158.
Gardner DK, Lane M, Stevens J, Schoolcraft WB. Noninvasive assessment of humanembryo nutrient consumption as a measure of developmental potential. FertilSteril 2001;76:1175–1180.
Diagnosis of human embryo viability 743

Gardner DK, Larman MG, Thouas GA. Sex-related physiology of the preimplantationembryo. Mol Hum Reprod 2010;16:539–547.
Gardner DK, Wale PL, Collins R, Lane M. Glucose consumption of singlepost-compaction human embryos is predictive of embryo sex and live birthoutcome. Hum Reprod 2011;26:1981–1986.
Geraedts J, Collins J, Gianaroli L, Goossens V, Handyside A, Harper J, Montag M,Repping S, Schmutzler A. What next for preimplantation genetic screening? Apolar body approach! Hum Reprod 2010;25:575–577.
Gleicher N,KushnirVA,BaradDH.Preimplantation genetic screening (PGS) still in searchof a clinical application: a systematic review. Reprod Biol Endocrinol 2014;12:22.
Glessner JT, Hakonarson H. Genome-wide association: from confounded to confident.Neuroscientist 2011;17:174–184.
Gook DA, Edgar DH, Lewis K, Sheedy JR, Gardner DK. Impact of oxygen concentrationon adult murine pre-antral follicle development in vitro and the correspondingmetabolic profile. Mol Hum Reprod 2014;20:31–41.
Greco E, Bono S, Ruberti A, Lobascio AM, Greco P, Biricik A, Spizzichino L, Greco A,Tesarik J, Minasi MG et al. Comparative genomic hybridization selection ofblastocysts for repeated implantation failure treatment: a pilot study. BioMed ResInt 2014;2014:457913.
Gueye NA, Devkota B, Taylor D, Pfundt R, Scott RT Jr, Treff NR. Uniparental disomy inthe human blastocyst is exceedingly rare. Fertil Steril 2014;101:232–236.
Gutierrez-Mateo C, Colls P, Sanchez-Garcia J, Escudero T, Prates R, Ketterson K,Wells D, Munne S. Validation of microarray comparative genomic hybridizationfor comprehensive chromosome analysis of embryos. Fertil Steril 2011;95:953–958.
Handyside AH. 24-chromosome copy number analysis: a comparison of availabletechnologies. Fertil Steril 2013;100:595–602.
Handyside AH, Harton GL, Mariani B, Thornhill AR, Affara NA, Shaw MA, Griffin DK.Karyomapping: a universal method for genome wide analysis of genetic diseasebased on mapping crossovers between parental haplotypes. J Med Gen 2010;47:651–658.
Hannan NJ, Paiva P, Meehan KL, Rombauts LJ, Gardner DK, Salamonsen LA. Analysis offertility-related soluble mediators in human uterine fluid identifies VEGF as a keyregulator of embryo implantation. Endocrinology 2011;152:4948–4956.
Hardarson T, Ahlstrom A, Rogberg L, Botros L, Hillensjo T, Westlander G, Sakkas D,Wikland M. Non-invasive metabolomic profiling of Day 2 and 5 embryo culturemedium: a prospective randomized trial. Hum Reprod 2012;27:89–96.
Harper JC, Harton G. The use of arrays in preimplantation genetic diagnosis andscreening. Fertil Steril 2010;94:1173–1177.
Harper JC, Coonen E, Handyside AH, Winston RM, Hopman AH, Delhanty JD.Mosaicism of autosomes and sex chromosomes in morphologically normal,monospermic preimplantation human embryos. Prenat Diagn 1995;15:41–49.
Harton GL, Munne S, Surrey M, Grifo J, Kaplan B, McCulloh DH, Griffin DK, Wells D,Group PGDP. Diminished effect of maternal age on implantation afterpreimplantation genetic diagnosis with array comparative genomic hybridization.Fertil Steril 2013;100:1695–1703.
Hashimoto S, Kato N, Saeki K, Morimoto Y. Selection of high-potential embryos byculture in poly(dimethylsiloxane) microwells and time-lapse imaging. Fertil Steril2012;97:332–337.
Heo YS, Cabrera LM, Bormann CL, Smith GD, Takayama S. Real time culture andanalysis of embryo metabolism using a microfluidic device with deformation basedactuation. Lab Chip 2012;12:2240–2246.
Herrero J, Meseguer M. Selection of high potential embryos using time-lapse imaging:the era of morphokinetics. Fertil Steril 2013a;99:1030–1034.
Herrero J, Tejera A, Albert C, Vidal C, de los Santos MJ, Meseguer M. A time to lookback: analysis of morphokinetic characteristics of human embryo development.Fertil Steril 2013b;100:1602–1609.
Hong K, Forman E, Prodoehl A, Upham K, Treff N, Scott R. Early times to cavitation areassociated with a reduced prevalence of aneuploidy in embryos cultured to theblastocyst stage: a prospective blinded morphokinetic study. Fertil Steril 2013;100:S382.
Houghton FD, Hawkhead JA, Humpherson PG, Hogg JE, Balen AH, Rutherford AJ,Leese HJ. Non-invasive amino acid turnover predicts human embryo developmentalcapacity. Hum Reprod 2002;17:999–1005.
Iwata K, Yumoto K, Sugishima M, Mizoguchi C, Kai Y, Iba Y, Mio Y. Analysis ofcompaction initiation in human embryos by using time-lapse cinematography. JAssist Reprod Genet 2014;31:421–426.
Joergensen MW, Agerholm I, Hindkjaer J, Bolund L, Sunde L, Ingerslev HJ, Kirkegaard K.Altered cleavage patterns in human tripronuclear embryos and their association tofertilization method: a time-lapse study. J Assist Reprod Genet 2014;31:435–442.
Johnson DS, Cinnioglu C, Ross R, Filby A, Gemelos G, Hill M, Ryan A, Smotrich D,Rabinowitz M, Murray MJ. Comprehensive analysis of karyotypic mosaicismbetween trophectoderm and inner cell mass. Mol Hum Reprod 2010a;16:944–949.
Johnson DS, Gemelos G, Baner J, Ryan A, Cinnioglu C, Banjevic M, Ross R, Alper M,Barrett B, Frederick J et al. Preclinical validation of a microarray method for fullmolecular karyotyping of blastomeres in a 24-h protocol. Hum Reprod 2010b;25:1066–1075.
Jones GM, Trounson AO, Vella PJ, Thouas GA, Lolatgis N, Wood C. Glucosemetabolism of human morula and blastocyst-stage embryos and its relationship toviability after transfer. Reprod Biomed Online 2001;3:124–132.
Kaser DJ, Racowsky C. Clinical outcomes following selection of human preimplantationembryos with time-lapse monitoring: a systematic review. Hum Reprod Update 2014;20:617–631.
Katz-Jaffe MG, Linck DW, Schoolcraft WB, Gardner DK. A proteomic analysis ofmammalian preimplantationembryonicdevelopment.Reproduction 2005;130:899–905.
Katz-Jaffe MG, Gardner DK, Schoolcraft WB. Proteomic analysis of individual humanembryos to identify novel biomarkers of development and viability. Fertil Steril2006a;85:101–107.
Katz-Jaffe MG, Schoolcraft WB, Gardner DK. Analysis of protein expression(secretome) by human and mouse preimplantation embryos. Fertil Steril 2006b;86:678–685.
Keltz MD, Vega M, Sirota I, Lederman M, Moshier EL, Gonzales E, Stein D.Preimplantation genetic screening (PGS) with Comparative genomic hybridization(CGH) following day 3 single cell blastomere biopsy markedly improves IVFoutcomes while lowering multiple pregnancies and miscarriages. J Assist ReprodGenet 2013;30:1333–1339.
Kirkegaard K, Agerholm IE, Ingerslev HJ. Time-lapse monitoring as a tool for clinicalembryo assessment. Hum Reprod 2012a;27:1277–1285.
Kirkegaard K, Hindkjaer JJ, Ingerslve HJ . Human embryonic development afterblastomere removal: a time-lapse analaysis. Hum Reprod 2012b;27:97–105.
Kirkegaard K, Hindkjaer JJ, Ingerslev HJ. Effect of oxygen concentration on humanembryo development evaluated by time-lapse monitoring. Fertil Steril 2013a;99:738–744 e734.
Kirkegaard K, Kesmodel US, Hindkjaer JJ, Ingerslev HJ. Time-lapse parameters aspredictors of blastocyst development and pregnancy outcome in embryos fromgood prognosis patients: a prospective cohort study. Hum Reprod 2013b;28:2643–2651.
Kirkegaard K, Campbell A, Agerholm I, Bentin-Ley U, Gabrielsen A, Kirk J, Sayed S,Ingerslev HJ. Limitations of a time-lapse blastocyst prediction model: a largemulticentre outcome analysis. Reprod Biomed Online 2014;29:156–158.
Knapp M, Stiller M, Meyer M. Generating barcoded libraries for multiplexhigh-throughput sequencing. Methods Mol Biol 2012;840:155–170.
Knez K, Tomazevic T, Vrtacnik-Bokal E, Virant-Klun I. Developmental dynamics ofIMSI-derived embryos: a time-lapse prospective study. Reprod Biomed Online2013;27:161–171.
Kobayashi S, Isotani A, Mise N, Yamamoto M, Fujihara Y, Kaseda K, Nakanishi T,Ikawa M, Hamada H, Abe K et al. Comparison of gene expression in male andfemale mouse blastocysts revealed imprinting of the X-linked gene, Rhox5/Pem,at preimplantation stages. Curr Biol 2006;16:166–172.
Kohn AB, Moroz TP, Barnes JP, Netherton M, Moroz LL. Single-cell semiconductorsequencing. Methods Mol Biol 2013;1048:247–284.
Kokkali G, Traeger-Synodinos J, Vrettou C, Stavrou D, Jones GM, Cram DS, Makrakis E,Trounson AO, Kanavakis E, Pantos K. Blastocyst biopsy versus cleavage stage biopsyand blastocyst transfer for preimplantation genetic diagnosis of beta-thalassaemia: apilot study. Hum Reprod 2007;22:1443–1449.
Konings P, Vanneste E, Jackmaert S, Ampe M, Verbeke G, Moreau Y, Vermeesch JR,Voet T. Microarray analysis of copy number variation in single cells. Nat Protoc2012;7:281–310.
Kramer YG, Kofinas JD, Melzer K, Noyes N, McCaffrey C, Buldo-Licciardi J,McCulloh DH, Grifo JA. Assessing morphokinetic parameters via time lapsemicroscopy (TLM) to predict euploidy: are aneuploidy risk classification modelsuniversal? J Assist Reprod Genet 2014;31:1231–1242.
Lane M, Gardner DK. Selection of viable mouse blastocysts prior to transfer using ametabolic criterion. Hum Reprod 1996;11:1975–1978.
744 Gardner et al.

Lane M, Gardner DK. Amino acids and vitamins prevent culture-induced metabolicperturbations and associated loss of viability of mouse blastocysts. Hum Reprod1998;13:991–997.
Lee YSL, Thouas GA, Gardner DK. Developmental kinetics of cleavage stage mouseembryos are related to their subsequent carbohydrate and amino acid utilisationat the blastocyst stage. Hum Reprod 2015;30:543–552.
Leese HJ. Quietplease, do not disturb: a hypothesis of embryometabolism and viability.Bioessays 2002;24:845–849.
Leese HJ, Baumann CG, Brison DR, McEvoy TG, Sturmey RG. Metabolism of the viablemammalian embryo: quietness revisited. Mol Hum Reprod 2008;14:667–672.
Lemmen JG, Agerholm I, Ziebe S. Kinetic markers of human embryo quality usingtime-lapse recordings of IVF/ICSI-fertilized oocytes. Reprod Biomed Online 2008;17:385–391.
Liu Y, Chapple V, Roberts P, Matson P. Prevalence, consequence, and significance ofreverse cleavage by human embryos viewed with the use of the Embryoscopetime-lapse video system. Fertil Steril. 2014;102:1295–1300.
Lu S, Zong C, Fan W, Yang M, Li J, Chapman AR, Zhu P, Hu X, Xu L, Yan L et al. Probingmeiotic recombination and aneuploidy of single sperm cells by whole-genomesequencing. Science 2012;338:1627–1630.
Machtinger R, Racowsky C. Morphological systems of human embryo assessment andclinical evidence. Reprod Biomed Online 2013;26:210–221.
Mamas T, Gordon A, Brown A, Harper J, Sengupta S. Detection of aneuploidy by arraycomparative genomic hybridization using cell lines to mimic a mosaic trophectodermbiopsy. Fertil Steril 2012;97:943–947.
Margalioth EJ, Ben-Chetrit A, Gal M, Eldar-Geva T. Investigation and treatment ofrepeated implantation failure following IVF-ET. Hum Reprod 2006;21:3036–3043.
Mastenbroek S, Repping S. Preimplantation genetic screening: back to the future. HumReprod 2014;29:1846–1850.
Mastenbroek S, Twisk M, van der Veen F, Repping S. Preimplantation genetic screening: asystematic reviewand meta-analysisofRCTs.HumReprodUpdate 2011a;17:454–466.
Mastenbroek S, van der Veen F, Aflatoonian A, Shapiro B, Bossuyt P, Repping S. Embryoselection in IVF. Hum Reprod 2011b;26:964–966.
McReynolds S, Vanderlinden L, Stevens J, Hansen K, Schoolcraft WB, Katz-Jaffe MG.Lipocalin-1: a potential marker for noninvasive aneuploidy screening. Fertil Steril2011;95:2631–2633.
Menke TM, McLaren A. Mouse blastocysts grown in vivo and in vitro: carbon dioxideproduction and trophoblast outgrowth. J Reprod Fertil 1970;23:117–127.
Meseguer M, Herrero J, Tejera A, Hilligsoe KM, Ramsing NB, Remohi J. The use ofmorphokinetics as a predictor of embryo implantation. Hum Reprod 2011;26:2658–2671.
Meseguer M, Rubio I, Cruz M, Basile N, Marcos J, Requena A. Embryo incubation andselection in a time-lapse monitoring system improves pregnancyoutcome comparedwith a standard incubator: a retrospective cohort study. Fertil Steril 2012;98:1481–1489 e1410.
Mio Y, Maeda K. Time-lapse cinematography of dynamic changes occurring duringin vitro development of human embryos. Am J Obstet Gynecol 2008;199:660e661–665.
Mir P, Rodrigo L, Mercader A, Buendia P, Mateu E, Milan-Sanchez M, Peinado V,Pellicer A, Remohi J, Simon C et al. False positive rate of an arrayCGH platformfor single-cell preimplantation genetic screening and subsequent clinicalapplication on day-3. J Assist Reprod Genet 2013;30:143–149.
Mitchell M, Cashman KS, Gardner DK, Thompson JG, Lane M. Disruption ofmitochondrial malate-aspartate shuttle activity in mouse blastocysts impairsviability and fetal growth. Biol Reprod 2009;80:295–301.
Montag M. Morphokinetics and embryo aneuploidy: has time come or not yet? ReprodBiomed Online 2013;26:528–530.
Montag M, van der Ven K, Rosing B, van der Ven H. Polar Body Biopsy: a viablealternative to preimplantation genetic diagnosis and screening. Reprod BiomedOnline 2009;18:6–11.
Munne S, Cohen J. Chromosome abnormalities in human embryos. Hum Reprod Update1998;4:842–855.
Munoz M, Cruz M, Humaidan P, Garrido N, Perez-Cano I, Meseguer M. Dose ofrecombinant FSH and oestradiol concentration on day of HCG affect embryodevelopment kinetics. Reprod Biomed Online 2012;25:382–389.
Munoz M, Cruz M, Humaidan P, Garrido N, Perez-Cano I, Meseguer M. The type ofGnRH analogue used during controlled ovarian stimulation influences earlyembryo developmental kinetics: a time-lapse study. Eur J Obstet Gynecol Reprod2013;168:167–172.
Nagaoka SI, Hassold TJ, Hunt PA. Human aneuploidy: mechanisms and new insightsinto an age-old problem. Nat Rev Genet 2012;13:493–504.
Nakahara T, Iwase A, Goto M, Harata T, Suzuki M, Ienaga M, Kobayashi H, Takikawa S,Manabe S, Kikkawa F, Ando H. Evaluation of the safety of time-lapse observations forhuman embryos. J Assist Reprod Genet 2010;27:93–96.
Nannya Y, Sanada M, Nakazaki K, Hosoya N, Wang L, Hangaishi A, Kurokawa M,Chiba S, Bailey DK, Kennedy GC et al. A robust algorithm for copy numberdetection using high-density oligonucleotide single nucleotide polymorphismgenotyping arrays. Cancer Res 2005;65:6071–6079.
Northrop LE, Treff NR, Levy B, Scott RT Jr. SNP microarray-based 24 chromosomeaneuploidy screening demonstrates that cleavage-stage FISH poorly predictsaneuploidy in embryos that develop to morphologically normal blastocysts. MolHum Reprod 2010;16:590–600.
Picton HM, Elder K, Houghton FD, Hawkhead JA, Rutherford AJ, Hogg JE, Leese HJ,Harris SE. Association between amino acid turnover and chromosome aneuploidyduring human preimplantation embryo development in vitro. Mol Hum Reprod2010;16:557–569.
Pribenszky C, Matyas S, Kovacs P, Losonczi E, Zadori J, Vajta G. Pregnancy achieved bytransfer of a single blastocyst selected by time-lapse monitoring. Reprod BiomedOnline 2010;21:533–536.
Rabinowitz M, Ryan A, Gemelos G, Hill M, Baner J, Cinnioglu C, Banjevic M, Potter D,Petrov DA, Demko Z. Origins and rates of aneuploidy in human blastomeres. FertilSteril 2012;97:395–401.
Rajcan-Separovic E. Chromosome microarrays in human reproduction. Hum ReprodUpdate 2012;18:555–567.
Renard JP, Philippon A, Menezo Y. In-vitro uptake of glucose by bovine blastocysts.J Reprod Fertil 1980;58:161–164.
RodrigoL,PeinadoV,MateuE,Remohi J,PellicerA,SimonC,Gil-SalomM,RubioC. Impactof different patterns of sperm chromosomal abnormalities on the chromosomalconstitution of preimplantation embryos. Fertil Steril 2010;94:1380–1386.
Rubio C, Gil-Salom M, Simon C, Vidal F, Rodrigo L, Minguez Y, Remohi J, Pellicer A.Incidence of sperm chromosomal abnormalities in a risk population: relationshipwith sperm quality and ICSI outcome. Hum Reprod 2001;16:2084–2092.
Rubio I, Kuhlmann R, Agerholm I, Kirk J, Herrero J, Escriba MJ, Bellver J, Meseguer M.Limited implantation success of direct-cleaved human zygotes: a time-lapse study.Fertil Steril 2012;98:1458–1463.
Rubio C, Bellver J, Rodrigo L, Bosch E, Mercader A, Vidal C, De Los Santos MJ, Giles J,Labarta E, Domingo J et al. Preimplantation genetic screening using fluorescence insitu hybridization in patients with repetitive implantation failure and advancedmaternal age: two randomized trials. Fertil Steril 2013;99:1400–1407.
Rubio I, Galan A, Larreategui Z, Ayerdi F, Bellver J, Herrero J, Meseguer M. Clinicalvalidation of embryo culture and selection by morphokinetic analysis: arandomized, controlled trial of the EmbryoScope. Fertil Steril 2014;102:1287–1294.
Ruiz-Alonso M, Blesa D, Diaz-Gimeno P, Gomez E, Fernandez-Sanchez M, Carranza F,Carrera J, Vilella F, Pellicer A, Simon C. The endometrial receptivity array fordiagnosis and personalized embryo transfer as a treatment for patients withrepeated implantation failure. Fertil Steril 2013;100:818–824.
Schoolcraft WB, Surrey E, Minjarez D, Gustofson RL, Scott RT Jr, Katz-Jaffe MG.Comprehensive chromosome screening (CCS) with vitrification results inimproved clinical outcome in women .35 years: a randomized control trial. FertilSteril 2012;98:S1.
Schuster TG, Cho B, Keller LM, Takayama S, Smith GD. Isolation of motile spermatozoafrom semen samples using microfluidics. Reprod Biomed Online 2003;7:75–81.
Scott L. Pronuclear scoring as a predictor of embryo development. Reprod BiomedOnline 2003;6:201–214.
Scott RT Jr, Treff NR. Assessing the reproductive competence of individual embryos:a proposal for the validation of new “-omics” technologies. Fertil Steril 2010;94:791–794.
Scott RT Jr, Ferry K, Su J, Tao X, Scott K, Treff NR. Comprehensive chromosomescreening is highly predictive of the reproductive potential of human embryos: aprospective, blinded, nonselection study. Fertil Steril 2012;97:870–875.
Scott RT Jr, Upham KM, Forman EJ, Zhao T, Treff NR. Cleavage-stage biopsy significantlyimpairs human embryonic implantation potential while blastocyst biopsy does not: arandomized and paired clinical trial. Fertil Steril 2013a;100:624–630.
Scott RT Jr, Upham KM, Forman EJ, Hong KH, Scott KL, Taylor D, Tao X, Treff NR.Blastocyst biopsy with comprehensive chromosome screening and fresh embryotransfer significantly increases in vitro fertilization implantation and delivery rates:a randomized controlled trial. Fertil Steril 2013b;100:697–703.
Diagnosis of human embryo viability 745

Seli E, Sakkas D, Scott R, Kwok SC, Rosendahl SM, Burns DH. Noninvasivemetabolomic profiling of embryo culture media using Raman and near-infraredspectroscopy correlates with reproductive potential of embryos in womenundergoing in vitro fertilization. Fertil Steril 2007;88:1350–1357.
Seli E, Botros L, Sakkas D, Burns DH. Noninvasive metabolomic profiling of embryoculture media using proton nuclear magnetic resonance correlates withreproductive potential of embryos in women undergoing in vitro fertilization. FertilSteril 2008;90:2183–2189.
Seli E, Bruce C, Botros L, Henson M, Roos P, Judge K, Hardarson T, Ahlstrom A,Harrison P, Henman M et al. Receiver operating characteristic (ROC) analysis ofday 5 morphology grading and metabolomic Viability Score on predictingimplantation outcome. J Assist Reprod Genet 2011;28:137–144.
Serdarogullari M, Findikli N, Goktas C, Sahin O, Ulug U, Yagmur E, Bahceci M.Comparison of gender-specific human embryo development characteristics bytime-lapse technology. Reprod Biomed Online 2014;29:193–199.
Shaffer LG, Dabell MP, Fisher AJ, Coppinger J, Bandholz AM, Ellison JW, Ravnan JB,Torchia BS, Ballif BC, Rosenfeld JA. Experience with microarray-based comparativegenomic hybridization for prenatal diagnosis in over 5000 pregnancies. Prenat Diagn2012;32:976–985.
Sheedy JR, Yoshida Y, Gardner DK. Direct injection mass spectrometry reveals uniquemetabolite profiles from spent human embryo culture media due to albumin sourceand pregnancy. Fertil Steril 2014;102:e212–e213.
Simpson JL. Preimplantation genetic diagnosis to improve pregnancy outcomes insubfertility. Best Pract Res Clin Obstet Gynaecol 2012;26:805–815.
Singh R, Sinclair KD. Metabolomics: approaches to assessing oocyte and embryoquality. Theriogenology 2007;68(Suppl. 1):S56–S62.
Stecher A, Vanderzwalmen P, Zintz M, Wirleitner B, Schuff M, Spitzer D, Zech NH.Transfer of blastocysts with deviant morphological and morphokinetic parametersat early stages of in-vitro development: a case series. Reprod Biomed Online 2014;28:424–435.
Sundvall L, Ingerslev HJ, Breth Knudsen U, Kirkegaard K. Inter-, intra-observervariability of time-lapse annotations. Hum Reprod 2013;28:3215–3221.
Sugiura-Ogasawara M, Ozaki Y, Katano K, Suzumori N, Kitaori T, Mizutani E. Abnormalembryonic karyotype is the most frequent cause of recurrent miscarriage. HumReprod 2012;27:2297–2303.
Swain JE. Could time-lapse embryo imaging reduce the need for biopsy and PGS? J AssistReprod Genet 2013;30:1081–1090.
Swain JE, Lai D, Takayama S, Smith GD. Thinking big by thinking small: application ofmicrofluidic technology to improve ART. Lab Chip 2013;13:1213–1224.
Tan YQ, Tan K, Zhang SP, Gong F, Cheng DH, Xiong B, Lu CF, Tang XC, Luo KL, Lin Get al. Single-nucleotide polymorphism microarray-based preimplantation geneticdiagnosis is likely to improve the clinical outcome for translocation carriers. HumReprod 2013;28:2581–2592.
Taylor TH, Gitlin SA, Patrick JL, Crain JL, Wilson JM, Griffin DK. The origin,mechanisms, incidence and clinical consequences of chromosomal mosaicism inhumans. Hum Reprod Update 2014;20:571–581.
Tejera A, Herrero J, Viloria T, Romero JL, Gamiz P, Meseguer M. Time-dependent O(2)consumption patterns determined optimal time ranges for selecting viable humanembryos. Fertil Steril 2012;98:849–857 e843.
Thouas G, Potter DL, Gardner DK. Microfluidic devices for the analysis of gamete andembryo physiology. In: Gardner DK, Seli E, Wells D, Sakkas D (eds), Human Gametesand Preimplantation Embryos: Assessment and Diagnosis. New York: Springer, 2013,281–299.
Tiffin GJ, Rieger D, Betteridge KJ, Yadav BR, King WA. Glucose and glutaminemetabolism in pre-attachment cattle embryos in relation to sex and stage ofdevelopment. J Reprod Fertil 1991;93:125–132.
Treff NR. Genome-wide analysis of human preimplantation aneuploidy. Semin ReprodMed 2012;30:283–288.
Treff NR, Scott RT Jr. Methods for comprehensive chromosome screening of oocytesand embryos: capabilities, limitations, and evidence of validity. J Assist Reprod Genet2012;29:381–390.
Treff NR, Su J, Tao X, Levy B, Scott RT Jr. Accurate single cell 24 chromosomeaneuploidy screening using whole genome amplification and single nucleotidepolymorphism microarrays. Fertil Steril 2010a;94:2017–2021.
Treff NR, Levy B, Su J, Northrop LE, Tao X, Scott RT Jr. SNP microarray-based 24chromosome aneuploidy screening is significantly more consistent than FISH. MolHum Reprod 2010b;16:583–589.
Treff NR, Su J, Tao X, Northrop LE, Scott RT Jr. Single-cell whole-genome amplificationtechnique impacts the accuracy of SNP microarray-based genotyping and copynumber analyses. Mol Hum Reprod 2011a;17:335–343.
Treff NR, Tao X, Schillings WJ, Bergh PA, Scott RT Jr, Levy B. Use of single nucleotidepolymorphism microarrays to distinguish between balanced and normal chromosomesin embryos from a translocation carrier. Fertil Steril 2011b;96:e58–e65.
Treff NR, Tao X, Ferry KM, Su J, Taylor D, Scott RT Jr. Development and validation of anaccurate quantitative real-time polymerase chain reaction-based assay for humanblastocyst comprehensive chromosomal aneuploidy screening. Fertil Steril 2012a;97:819–824.
Treff NR, Campos J, Tao X, Levy B, Ferry KM, Scott RT Jr. Blastocyst preimplantationgenetic diagnosis (PGD) of a mitochondrial DNA disorder. Fertil Steril 2012b;98:1236–1240.
Treff NR, Fedick A, Tao X, Devkota B, Taylor D, Scott RT Jr. Evaluation of targetednext-generation sequencing-based preimplantation genetic diagnosis ofmonogenic disease. Fertil Steril 2013a;99:1377–1384.
Treff NR, Forman EJ, Scott RT Jr. Next-generation sequencing for preimplantationgenetic diagnosis. Fertil Steril 2013b;99:e17–e18.
Treff NR, Forman EJ, Katz-Jaffe MG, Schoolcraft WB, Levy B, Scott RT Jr. Incidentalidentification of balanced translocation carrier patients through comprehensivechromosome screening of IVF-derived blastocysts. J Assist Reprod Genet 2013c;30:787–791.
Urbanski JP, Johnson MT, Craig DD, Potter DL, Gardner DK, Thorsen T. Noninvasivemetabolic profiling using microfluidics for analysis of single preimplantation embryos.Anal Chem 2008;80:6500–6507.
Vajta G. Vitrification in human and domestic animal embryology: work in progress.Reprod Fertil Dev 2013;25:719–727.
Van der Aa N, Cheng J, Mateiu L, Esteki MZ, Kumar P, Dimitriadou E, Vanneste E,Moreau Y, Vermeesch JR, Voet T. Genome-wide copy number profiling of singlecells in S-phase reveals DNA-replication domains. Nucleic Acids Res 2013;41:e66.
Van Royen E, Mangelschots K, De Neubourg D, Valkenburg M, Van de Meerssche M,Ryckaert G, Eestermans W, Gerris J. Characterization of a top quality embryo, a steptowards single-embryo transfer. Hum Reprod 1999;14:2345–2349.
van Uum CM, Stevens SJ, Dreesen JC, Drusedau M, Smeets HJ, Hollanders-Crombach B, Die-Smulders CE, Geraedts JP, Engelen JJ, Coonen E. SNP array-based copy number and genotype analyses for preimplantation genetic diagnosisof human unbalanced translocations. Eur J Hum Genet 2012;20:938–944.
Vanneste E, Voet T, Le Caignec C, Ampe M, Konings P, Melotte C, Debrock S,Amyere M, Vikkula M, Schuit F et al. Chromosome instability is common in humancleavage-stage embryos. Nat Med 2009;15:577–583.
Verberg MF, Eijkemans MJ, Heijnen EM, Broekmans FJ, de Klerk C, Fauser BC,Macklon NS. Why do couples drop-out from IVF treatment? A prospectivecohort study. Hum Reprod 2008;23:2050–2055.
Vergouw CG, Botros LL, Roos P, Lens JW, Schats R, Hompes PG, Burns DH,Lambalk CB. Metabolomic profiling by near-infrared spectroscopy as a tool toassess embryo viability: a novel, non-invasive method for embryo selection. HumReprod 2008;23:1499–1504.
Vergouw CG, Kieslinger DC, Kostelijk EH, Botros LL, Schats R, Hompes PG, Sakkas D,Lambalk CB. Day 3 embryo selection by metabolomic profiling of culture mediumwith near-infrared spectroscopy as an adjunct to morphology: a randomizedcontrolled trial. Hum Reprod 2012;27:2304–2311.
Voet T, Kumar P, Van Loo P, Cooke SL, Marshall J, Lin ML, Zamani Esteki M,Van der Aa N, Mateiu L, McBride DJ et al. Single-cell paired-end genomesequencing reveals structural variation per cell cycle. Nucleic Acids Res 2013;41:6119–6138.
Wale PL, Gardner DK. Time-lapse analysis of mouse embryo development in oxygengradients. Reprod Biomed Online 2010;21:402–410.
Wale PL, Gardner DK. Oxygen regulates amino acid turnover and carbohydrate uptakeduring the preimplantation period of mouse embryo development. Biol Reprod 2012;87:24.
Wale PL, Gardner DK. Oxygen affects the ability of mouse blastocysts to regulateammonium. Biol Reprod 2013;89:75.
Walker KJ, Green MP, Gardner DK. Spatial asynchronous transfer of cleavage stagemouse embryos to the uterus compromises fetal development. Mol Reprod Dev2015;82:80.
Wells D, Fragouli E, Alfarawaty S et al. Increased embryo implantation and high birthrates following comprehensive chromosomal screening of in vitro fertilized
746 Gardner et al.

embryos. 9th Annual International Conference on Preimplantation Genetics, Miami,Florida, April 23–25. Reprod Biomed Online, 2009;18(Suppl):S10
Wilton LJ. In vitro fertilization with preimplantation genetic screening. N Engl J Med2007;357:1770.
Wong CC, Loewke KE, Bossert NL, Behr B, De Jonge CJ, Baer TM, Reijo Pera RA.Non-invasive imaging of human embryos before embryonic genome activationpredicts development to the blastocyst stage. Nat Biotechnol 2010;28:1115–1121.
Wong C, Chen AA, Behr B, Shen S. Time-lapse microscopy and image analysis in basicand clinical embryo development research. Reprod Biomed Online 2013;26:120–129.
Yang Z, Liu J, Collins GS, Salem SA, Liu X, Lyle SS, Peck AC, Sills ES, Salem RD. Selectionof single blastocysts for fresh transfer via standard morphology assessment alone andwith array CGH for good prognosis IVF patients: results from a randomized pilotstudy. Mol Cytogenet 2012;5:24.
Yin X, Tan K, Vajta G, Jiang H, Tan Y, Zhang C, Chen F, Chen S, Zhang C, Pan X et al.Massively parallel sequencing for chromosomal abnormality testing introphectoderm cells of human blastocysts. Biol Reprod 2013;88:69.
Zhang C, Zhang C, Chen S, Yin X, Pan X, Lin G, Tan Y, Tan K, Xu Z, Hu P et al. A singlecell level based method for copy number variation analysis by low coverage massivelyparallel sequencing. PLoS One 2013;8:e54236.
Diagnosis of human embryo viability 747