dna and lipid molecular stratigraphic records of haptophyte
TRANSCRIPT

Earth and Planetary Science Letters 284 (2009) 610–621
Contents lists available at ScienceDirect
Earth and Planetary Science Letters
j ourna l homepage: www.e lsev ie r.com/ locate /eps l
DNA and lipid molecular stratigraphic records of haptophyte succession in the BlackSea during the Holocene
Marco J.L. Coolen a,⁎, James P. Saenz a, Liviu Giosan b, Nan Y. Trowbridge a, Petko Dimitrov c,Dimitar Dimitrov c, Timothy I. Eglinton a
a Marine Chemistry and Geochemistry Department, Woods Hole Oceanographic Institution, Woods Hole, MA 02543, USAb Geology and Geophysics Department, Woods Hole Oceanographic Institution, Woods Hole, MA 02543, USAc Bulgarian Academy of Sciences, Institute of Oceanology, Marine Geology and Archaeology Department, P.O. Box 152, 9000 Varna, Bulgaria
⁎ Corresponding author. Tel.: +1 508 289 2931.E-mail address: [email protected] (M.J.L. Coolen).
0012-821X/$ – see front matter. Published by Elsevierdoi:10.1016/j.epsl.2009.05.029
a b s t r a c t
a r t i c l e i n f oArticle history:Received 15 August 2008Received in revised form 18 May 2009Accepted 22 May 2009Available online 21 June 2009
Editor: M.L. Delaney
Keywords:Black SeaHoloceneancient DNAfossil DNApaleogeneticsalkenonesEmliania huxleyiIsochrysis
Previous studies suggest that the coccolithophorid haptophyte Emiliania huxleyi entered the Black Sea ~3400 yrsago and since thena coccolith oozedefined asUnit I has developed.Unit I sediments contain long-chain alkenonesderived from E. huxleyiwhereas the alkenone distribution of the deeper coccolith-free sapropel (Unit II) is ratherunusual. Alkenone-derived past sea surface temperature (SST) estimates suggest a large difference between UnitII and Unit I, which is likely a result of unusual biological precursors of the alkenones in Unit II. Here, we report ahigh-resolution stratigraphic analysis of ancient haptophyte DNA to establish the Holocene succession ofhaptophytes as sources of the alkenones in the Black Sea. Haptophytes related to brackish Isochrysis spp. were theinitial sources of alkenones, and appeared immediately after the onset of sapropel deposition (~7550 yrs beforepresent [a BP]). As salinity increased, Isochrysis-related haptophyteswere slowly replaced by a complex suite of E.huxleyi strains as sources of alkenones. Our paleogenetic data showed that E. huxleyi colonized the Black Seashortly after the onset of sapropel deposition, ~4000 yrs earlier than previously recognized based on theirpreserved coccoliths. E. huxleyi strains were the most likely source of the previously reported abundant andunusual C36 di-unsaturated “Black Sea alkenone”. Strong haptophyte species and strain-specific effects wereobserved on the level of unsaturation of alkenones which resulted in spurious alkenone-derived SST estimatesbefore 5250 a BP. In contrast, from ~5250 a BP onwards a relatively stable haptophyte assemblage dominated by adifferent suite of E. huxleyi strains yielded robust alkenone-SST values and indicated a gradual cooling from 19 °Cto ~15 °C at the top of the record (~450 a BP).
Published by Elsevier B.V.
1. Introduction
Climate shifts over Eurasia, together with global sea level changes,modulated freshwater and saltwater inputs to the Black Sea during thedeglaciation and early Holocene, leading to major hydrologic changesin the basin (Kaminski et al., 2002; Mudie et al., 2002; Major et al.,2002; Aksu et al., 2002a,b; Kaplin and Selivanov, 2004; Major et al.,2006; Bahr et al., 2006; Mudie et al., 2007; Hiscott et al., 2007; Van derMeer et al., 2008). Various studies have shown that the Black Sea wasoriginally a lake with low salinity waters, underlain by largely oxicsurface sediments (Unit III). Due to the postglacial sea-level rise, aconnection across the shallow sill of the Bosphorus to the Aegean andMediterranean Seawas established no later than 9400 a ago (e.g. Ryanet al., 1997; Aksu et al., 1999). Following the Bosphorus connection, astrong pycnocline developed between the brackish surface water,influenced by the large riverine inflow, and the more saline bottomwater of Mediterranean origin. Due to the stable stratification, anoxia
B.V.
developed below the pycnocline, and from~7500 a ago an organic-richsapropel (Unit II) accumulated in the deepbasin. The coccolithophorid,Emiliania huxleyi (from now on abbreviated as Eh), is believed to haveinvaded the Black Sea ~3400 a ago, and since then a laminatedcoccolith ooze, defined as Unit I, has been deposited (Hay et al., 1991;Arthur and Dean, 1998). Unit I also contains abundant long-chain (C37,C38 and C39) unsaturated methyl and ethyl ketones (alkenones) (Xuet al., 2001) which are characteristic of haptophyte algae including Eh(Volkman et al., 1980; Marlowe et al., 1984; Patterson et al., 1994;Volkman et al.,1995; Rontani et al., 2004; Sawada and Shiraiwa, 2004).These compounds are of great interest to paleoceanographers becauseof the strong empirical relationship between the degree of unsatura-tion in alkenones (U37
K ′ unsaturation parameter) and growth tempera-ture, which forms the basis for their use as a molecular proxy of pastSSTs (e.g. Brassell et al., 1986; Prahl and Wakeham, 1987).
On the other hand, alkenone distributions in Unit II werecharacterized by high concentrations of a novel alkenone (a di-unsaturated ethyl ketone with 36 carbon atoms [C36:2 eK]) and theexclusive presence of C37:2 mK (i.e. no C37:3 mK) (Xu et al., 2001).Alkenone-derived SST estimates suggest a large difference between

611M.J.L. Coolen et al. / Earth and Planetary Science Letters 284 (2009) 610–621
Unit II (average, 19.8 °C) and Unit I (average, 14.6 °C) (Xu et al., 2001).A shift of similar magnitude occurs between the uppermost interval ofUnit II and the first invasion of Eh, implying an unrealistic change inSST on timescales of ~200 yrs. The absence of Eh coccoliths in Unit IIsuggests that other factors must be responsible for the abrupt changesin U37
K ′, and different biological precursors for the C36:2 and C37:2alkenones appear to be a likely possibility. This inference is supportedby the stable carbon isotopic compositions (δ13C) of alkenones foundin Unit I and Unit II, which differ significantly (Xu et al., 2001).
In a previous study we identified and quantified past phytoplanktonmembers based on their ancient DNA preserved in the sediment record(i.e., fossil DNA) using molecular biological tools (i.e. “paleogenetics”)(Coolen et al., 2006).Ehwas the only source of alkenones in theUnit I but,unfortunately, the short multicore did not cover the sapropel includingthe interval with the unusual C36:2 alkenone (Coolen et al., 2006).
In this study, we extended the record of haptophyte successionbased on their preserved 18S ribosomal RNA gene (18S rDNA)signatures (~60 yr resolution) throughout the complete sapropeland the top 20 cm of the lacustrine Unit III in order to probe foradditional sequences of non-coccolithophorid haptophytes, whichcould be the sources of alkenones in Unit II sediments. 18S rDNA is asuitable phylogenetic marker to differentiate between non-cocco-lithophorid and coccolithophorid alkenone producers (Coolen et al.,2004), but Eh and the closely related Gephyrocapsa oceanica haveidentical 18S rDNA sequences (Medlin et al., 1994). To distinguishbetween closely related strains of both species, we targeted the fasterevolving mitochondrial cytochrome oxidase subunit I encoding gene(COI) as a phylogenetic marker (Stothard and Rollinson, 1997; Quanet al., 2004; Gunasekera et al., 2005; Monti et al., 2005; Paine et al.,2008). These paleogenetic studies were made in conjunction withdown-core alkenone compositional analyses so that relationshipsbetween species/strain succession and the lipid biomarker SST proxyrecord could be carefully evaluated.
2. Material and methods
2.1. Sampling
Alkenones and DNA of ancient haptophytes were analyzed fromGiant Gravity Core GGC18, recovered from awater depth of 971m in thewestern basin of the Black Sea (42°46.569″N:28°40.647″E) (Fig. S1)during cruise AK06 on the R/V Akademik (Institute of Oceanology,Bulgarian Academy of Sciences; IOBAS) in September 2006. On board,the corewas split in half and the upper 140 cmwas sectioned into 1 cm-intervals and sub-sampled. This section spans most of Unit I (N.B.: theupper 12 cm, representing the final 450 yrs of deposition of Unit I, wasnot recoveredwith thegravitycorer), the completeUnit II, and theupper20 cm of Unit III. Precautions were taken to prevent (cross) contamina-tion with foreign DNA (Supplementary material). Samples for DNAextraction were stored in liquid nitrogen during the cruise and shippedon dry ice to the Woods Hole Oceanographic Institution (WHOI). Theremaining sampled and archived core halves were shipped refrigeratedto WHOI where the remaining part of each 1 cm-interval was sub-sampled and transferred into pre-combusted glass jars and freeze-driedfor lipid geochemistry, total organic carbon (TOC) content, and 14C AMSdating of selected intervals.
2.2. Elemental composition by X-ray fluorescence (XRF)-scanning
The archived core half was digitally photographed, subjected toX-radiography, and scanned for bulk elemental composition using anITRAX™micro-XRF scanner (Croudace et al., 2005; St-Onge et al., 2007).The XRF measurements were performed using a molybdenum X-raytube at an exposure time of 10 s per measurement and a step-size of200 µm to allow for analysis of bulk chemistry at lamina and intra-lamina scales. The downcore variability in calcium was used herein to
characterize the core lithostratigraphy. In calibration studies with thesame instrument (Bertrand et al., 2007) as well as other XRF scanningstudies in the Black Sea (Kwiecien et al., 2009), itwas found that calciumis largely a direct measure of calcium carbonate content in sediments.
2.3. Age model for GGC18
Thirteen one-cm-intervals were selected for 14C AMS dating of bulkorganic carbon at the National Ocean Sciences Accelerator MassSpectrometry (NOSAMS) Facility at WHOI. Radiocarbon dating ofindividual organic compounds in concert with bulk organic carbon(Eglinton et al.,1997) suggests that pre-aged detrital organic carbon is aminor component of the carbon inHolocene Black Sea sediments. This issupported by a new estimate for the onset of sapropel at ~8000 yrs BP(Bahr et al., 2006), which is significantly earlier than previouslyproposed (Jones and Gagnon, 1994). However, until a better under-standing of the time variability of detrital organic carbon inputs isdeveloped, and for reasons of consistency with previous studies, weapplied a detrital carbon correction of 580 yrs to our radiocarbon dates(Jones and Gagnon, 1994). An age model for the core was thendeveloped by calibrating the corrected radiocarbon dates to calendaryears B.P. (1950) with Calib 5.0.1 (Stuiver and Reimer, 1993) using theINTCAL04 calibration curve (Reimer et al., 2004).
2.4. TOC content, stable carbon isotopic values (δ13C, ‰), and lipidgeochemistry (alkenone analysis)
All quantitative fossil DNA and alkenone data in this study werenormalized to TOC content. The weighted percentage of organic carbonand stable (organic) carbon isotopic valueswas determined on acidifiedsamples according to Eglinton et al. (2002) using a Carlo Erba EA 1108elemental analyzer interfaced via a Finnigan-MAT Conflo-II open splitdevice to a DeltaPlus isotope ratio monitoring mass spectrometer.
Extraction and analysis of alkenones was based on methodsdescribed previously (Xu et al., 2001). Total lipid extracts (TLEs) wereobtained from ~2 g dry weight sediment. The sediment was freeze-dried and extracted by Accelerated Solvent Extraction (ASE) using 9:1dichloromethane/methanol (100 °C, 1000 psi). The TLE was thendried under N2 and saponified using 2 M KOH in 4:1 (v/v) methanol/water (5 mL, 80 °C, 1 h). NaCl-saturated distilled water was added(5 mL), and neutral components, including alkenones, were extractedthree times in 1:4 (v/v) dichloromethane/hexane at basic pH. Theneutral TLE fraction was then loaded onto a glass column containingactivated Si-gel and separated into two fractions by eluting with 3 mLhexane (F1:hydrocarbons) and 15 mL dichloromethane (F2: alke-nones). Prior to GC analysis, 10 µg/µl of C36 n-alkane was added tofraction F2 as an internal standard. Analysis of alkenones wasperformed by GC-FID on a Hewlett Packard 5890 using a CPSil5-CBcolumn (Chrompack, 30 m×0.32 mm I.D.). Samples were injected at40 °C (1 min). The oven was programmed after Xu et al. (2001).Compound identificationwas based on comparison of retention timeswith samples characterized previously (Xu et al., 2001).
2.5. Extraction of total DNA
Total DNAwas extracted from 5 g of wetweight sediment using thePowerMax™ Soil DNA Isolation Kit following the recommendations ofthe manufacturer (Mobio, Carlsbad, CA). See the Supplementarymaterial for precautions to prevent contamination with foreign DNAand controls for contamination. DNA extracts were tested for thepresence of PCR-inhibiting impurities and, if necessary, subjected tofurther purification (Supplementary material).
The concentration of extracted DNA was quantified with thefluorescent dye PicoGreen (Molecular Probes) (Coolen et al., 2006).The genomic DNA of each individual 1 cm-interval served as templatefor the steps outlined in Sections 2.6 and 2.7.

612 M.J.L. Coolen et al. / Earth and Planetary Science Letters 284 (2009) 610–621
2.6. QPCR amplification of fossil genetic haptophyte markers
All reactions were performed in a Realplex qPCR cycler (Eppendorf)and involved initial denaturing, followed by 38 cycles including denatur-ing (30 s at 94 °C), 40 s of primer annealing at temperatures described inTable S1, primer extension (60 s at 72 °C) and imaging of newly formed
Fig. 1. Holocene sediment record GGC18 with (A) calcium content from coccolithophorid alsedimentary DNA per gramdryweight sediment and per gramTOC; (D) total number of preserhaptophyte 18S rDNAandalkenones and the ratiobetweenEhCOI and alkenones as anassessmenplotted against calendar ages. The gray-shaded parts of the figure mark the timing when the v
SYBRgreen labelled double-stranded DNA (80 °C for 20 s). With theexception of the primers (0.2 µMeach) used in this study, PCR ingredientswere similar to those described previously (Coolen et al., 2006).
Partial (498-bp-long) haptophyte 18S rDNA was selectively ampli-fiedusinghaptophyte-specific primers (Coolen et al., 2004).18S rDNAofIsochrysis galbana and Isochrysis-related haptophytes was amplified
gae as revealed by XRF elemental scanning; (B) TOC content and δ13C values; (C) totalved haptophyte 18S rDNA and (E) Eh COI copies as revealed by qPCR; (F) The ratio betweentof the level ofpreservationof both types of haptophytemolecularmarkers. All data pointsarious alkenone-producing haptophytes were present in the Black Sea.

Table 1Major events or observations related to Holocene haptophyte distributions in GGC18.
Note: the calendar ages of the sediment intervals are based on the age model used.The gray-shaded parts of the table mark the timing when the various alkenone-producing haptophytes were present in the Black Sea as outlined in the legend below the table.
Table 2Radiocarbon and calendar age determined from bulk organic matter in selected 1 cm-intervals of GGC18.
613M.J.L. Coolen et al. / Earth and Planetary Science Letters 284 (2009) 610–621
with the general haptophyte primer “Prym429f” (Coolen et al., 2004)and a novel Isochrysis-specific reverse primer (Isochr_742r; Table S1).The latter primerwas designed for this study based on aligned 18S rDNAsequences available through theSilva database (Pruesse et al., 2007) andby using the Probe_Design tool implemented in the phylogenetic treeprogram ARB (Ludwig et al., 2004). Partial (317 bp-long) COI gene of Ehor G. oceanica was selectively amplified using primers EhuxGoce_coiFand EhuxGoce_coiR (Table S1). The latter COI primerswere designed forthis study based on aligned mitochondrial COI sequences of Eh CCMP1742 (AY342361) and G. oceanica (AB000118). To further test thespecificity of the COI primers, 317 bp-long fragments were amplifiedfrom the mitochondrial genome of seventeen Eh and two G. oceanicastrains listed in Table S1.
Between 101 and 108 copies (10-fold dilution series) of 18S rDNA orCOI of Eh CCMP371 were added to reaction mixtures and served asstandards during qPCR to calibrate the copy numbers of fossilhaptophyte DNA in the Black Sea samples. The above qPCR productswere separated based on species/strain-specific variations in thenucleotide positions by Denaturing Gradient Gel Electrophoresis(DGGE) (Muyzer et al., 1993).
2.7. Phylogenetic analysis of sequenced DGGE bands
The polyacrylamide gels (6%, wt/vol) contained a denaturinggradient of 20–50% (with 100% denaturant equaling 7 M urea and 40%formamide) for haptophyte 18S rDNA and 20–70% for COI. Gels wererun for 5 h at 12.5 V cm−1 at 60 °C using a PhorU2 system (Ingeny,Leiden, Netherlands). DGGE bands were excised from the gel andprepared for subsequent cycle sequencing (Coolen et al., 2006) usingthe facilities of Agencourt, Beverly, MA.
The 18S rDNA sequences were analyzed using the ARB phylogenysoftware package (Ludwig et al., 2004). The Fast Aligner V1.03 toolimplemented in ARB was used for automatic alignment of therecovered Black Sea sequences with their closest relatives from theNational Center for Biotechnology Information (NCBI) database(http://www.ncbi.nlm.nih.gov/). Then, the phylogenetic bootstraptrees (1000 replications) were first reconstructed based on 1200 bp-long available sequences of the closest relatives employing theNeighbor-Joining method (Saitou and Nei, 1987). The shorter alignedenvironmental 18S rDNA sequences from this study were inserted

614 M.J.L. Coolen et al. / Earth and Planetary Science Letters 284 (2009) 610–621
afterwards without changing the overall tree topology, employing theParsimony Interactive tool implemented in ARB.
The alignment of nucleotide and protein sequences of the COIrecovered from the Black Sea and CCMP and CCAP cultures wasperformed in ARB using COI sequences of Eh (AY342361) and G.oceanica (AB000118) as references. Then, the phylogenetic trees werereconstructed employing the Neighbor-Joining method. Sequencesobtained in this study have been deposited in the NCBI sequencedatabase under accession numbers GQ245983–GQ246021.
3. Results
3.1. Chronology
The stratigraphy of Black Sea sediments deposited under sulfidicconditions has beenwell described (Calvert et al.,1987; Hay,1988; Hayet al., 1991; Arthur et al., 1994; Jones and Gagnon, 1994; Bahr et al.,2005). XRF scanning revealed that the upper 47 cm of sediment
Fig. 2. Phylogenetic position of haptophyte 18S rDNA sequences retrieved from GGC18 sehaptophyte sequences from GGC18 were determined from the DGGE represented in Fig. S2biosynthesizing haptophytes of the order Isochrysidaleswhereas sequences Hap2–Hap5 are rof alkenones. The time intervals when the various alkenone-producing haptophytes were pmutations per nucleotide. Numbers at nodes give bootstrap values out of 1000 resamplings
represents the calcium-rich, coccolith-bearing laminated sedimentlayers of Unit I (Fig. 1A, Table 1). AMS radiocarbon (14C) dating wasperformed on 13 selected sediment layers spanning the marine Unit Iand II (Table 2). Thefirst occurrence of Eh coccoliths in our core is datedat 2576±63 a BP (Table 1, Fig.1A). The initiation of the organic carbon-rich, Unit II sapropel is dated at 7550 a BP (Table 1). The averagesedimentation rates were 0.21 mm yr−1 (Unit I) and 0.15 mm yr−1
(Unit II). Therefore, each 1 cm interval represents on average ~40 yrs ofdeposition in Unit I, and ~60 yrs of deposition in Unit II.
3.2. TOC, δ13C, and sedimentary DNA content
The TOC-content was on average 3.75±0.49% in the Unit Isediments and on average two times higher in the upper part of theUnit II sapropel (7.12±0.62%). With the exception of the lower TOC-content (4.2%) between 6700 and 6830 a BP, TOC concentrations wereslightly higher in the sapropel deposited prior to 4900 a BP (9.81±1.67%) (Fig. 1B). The TOC-content was significantly lower in the upper
diment layers relative to reference sequences obtained from the NCBI database. The. Holocene Black Sea sequences Hap1 and Hap6 through Hap9 are related to alkenone-elated to haptophytes outside the Isochrysidales cluster, and not likely to be past sourcesresent in the Black Sea are denoted after each sequence. Bar indicates 0.1 fixed pointfor phylogenetic trees calculated by Neighbor Joining/Maximum Parsimony methods.

615M.J.L. Coolen et al. / Earth and Planetary Science Letters 284 (2009) 610–621
part of Unit III (2.41±0.79) (data not shown) and at the Unit III/Unit IItransition (3.0%) at 7550 a BP (Fig. 1B).
The δ13C values were relatively consistent at −24.36±0.45‰ inUnit I and Unit II sediments younger than 4900 a, although therewas aslight trend in more enriched δ13C values towards the recent past(Fig. 1B). Between 5400 and 6300 a BP, the most enriched δ13C valueswere observed (close to −23‰). δ13C values were most depletedtowards the transition to Unit III (i.e. −26.5‰).
The total DNA concentration in the sediment layers depositedbetween 450 and 7570 a BP was on average 10.1±6.2 µg per dryweight sediment [µg (g dw sed)−1] or 196±173 µg per gram TOC [µg(g TOC)−1] (Fig. 1C). DNA contents were more constant in sediments
Fig. 3.Holocene sediment record GGC18 with (A) relative abundance (max 100%) of main alkrelative abundance of other haptophyte species not related to alkenone producers of the ordstrains of Eh as identified based on COI as the phylogenetic marker; (C) concentration of the ugram dry weight sediment and (E) per gram TOC; (F) alkenone-derived (U37
K and U37K ′) SST va
gray-shaded parts of the figure mark the timing when the various alkenone-producing hap
deposited before 3500 a BP [8.7±2.5 µg (g dw sed)−1 or 102±44 µg(g TOC)−1] as compared to the DNA contents in younger sedimentlayers [11.3±7.9 µg (g dw sed)−1 or 278±202 µg (g TOC)−1](Fig. 1C). DNA contents declined sharply at the Unit II/III transitionwith 0.78±0.48 µg (g dw sed)−1 or 30±16 µg (g TOC)−1in the upper20 cm of Unit III sediments (data not shown).
3.3. Markers indicative of Holocene haptophyte algae in the Black Sea
3.3.1. 18S rDNA stratigraphyQPCR analysis revealed that the amount of fossil haptophyte 18S
rDNA increased approximately 10-fold over ~7000 yrs of deposition.
enone producers based on pixel densities of 18S rDNA bands in digitized DGGE gels (theer Isochrysidales is not included); (B) the relative abundance of 11 different Holocenenusual Black Sea alkenone (C36:2 ethyl ketone); (D) concentrations of C37 alkenones perlues calibrated after (Prahl et al., 1988). All data points plotted against calendar ages. Thetophytes were present in the Black Sea.

616 M.J.L. Coolen et al. / Earth and Planetary Science Letters 284 (2009) 610–621
Unit I layers<2000 a BP contained the highest amount of haptophyte18S rDNA [between 1.9·105 and 6.5·106 copies (g TOC)−1]. A highervariability could be observed in the remainder of the core [between1.5·103 and 3·106 copies (g TOC)−1] (Fig. 1D). A comparable depthprofile was observed when the fossil haptophyte 18S rDNA contentwas normalized to dry weight sediment (Fig. 1D). Haptophyte 18SrDNAwas below the detection limit [<150 copies (g dw sed)−1] in thelacustrine Unit III sediments (data not shown).
Separation of haptophyte 18S rDNA by DGGE (Fig. S2) andsubsequent phylogenetic analysis of sequenced DGGE bands resultedin the identification of nine unique environmental sequences (i.e.phylotypes) throughout the Holocene record (Fig. 2). The firsthaptophytes were related to Isochrysis species and appeared at theonset of sapropel deposition at 7550 a BP (Figs. 2, 3A, Table 1).Phylotype Hap8 (Fig. 2) (i.e., DGGE band 8 in Fig. S2) was the mostfrequently occurring Isochrysis-related haptophyte and comprised20–100% of haptophyte 18S rDNA within sediments >4900 a BP(Fig. 3A). Isochrysis-related phylotypes Hap6, 7, and 9 were lessabundant and persisted only for brief intervals during early sapropeldeposition (Fig. S2). All fossil Isochrysis-related phylotypes showedhighest similarity to sequences found in Holocene sediments from apermanently stratified brackish lake on the Antarctic coastline (AceLake, Vestfold Hills) (Coolen et al., 2004) (Fig. 2). Sediment layers<4900 a BP did not contain Isochrysis DNA (Fig. 3A, Table 1). Theabsence of IsochrysisDNA in sediment layers<4900 a BPwas confirmedby qPCR with our Isochrysis-specific primers: 107 copies of Eh 18S rDNAtemplate generated just asmany amplicons aswhen only 102 copies of I.galbana 18S rDNA template were used. Thus, the Isochrysis-specificprimer-set was five orders of magnitude more specific for Isochrysis-than for Eh 18S rDNA (data not shown). Using this highly specific andsensitive PCR approach, no PCR products could be generated fromsamples <4900 a BP after 40 cycles, indicating that Isochrysis 18S rDNAwas indeed absent from those layers (data not shown).
18S rDNA of Eh was almost continuously present following its firstappearance at 6430 a BP (Fig. 3A, Table 1) and Eh was the onlyhaptophyte found in Unit I (Figs. 3A, S2). Our haptophyte-specificprimers also target and amplify 18S rDNA of Chrysotila lamellosa (datanot shown) but this benthic alkenone producer (Rontani et al., 2004)was never detected in the analyzed sediment layers.
In addition, haptophyte sequences were found with no clearaffiliation to alkenone producers of the order Isochrysidales (phylo-
Fig. 4. Phylogenetic tree showing the relationship of partial Eh COI sequences retrieved fromHfrom the NCBI database and our newly sequenced COI genes from fifteen Eh and two G. oc(CCMP), and two Eh strains from the Culture Collection of Algae and Protozoa (CCAP). The Ehindicates 0.05 fixed point mutations per nucleotide. The time intervals when the various Eh
types 2–5, Figs. 2, S2). Since these phylotypes were also not directlyrelated to named species of other haptophytes, they are irrelevant forthis paper and will not be discussed further.
3.3.2. COI stratigraphyTo distinguish between Eh strains and G. oceanica, we designed
primers targeting the mitochondrial COI-gene (Stothard and Roll-inson, 1997; Quan et al., 2004; Gunasekera et al., 2005; Monti et al.,2005; Paine et al., 2008). Nine nucleotide positions were differentwithin the 317 bp-long COI fragment between Eh (AY342361) and G.oceanica (AB000118) whereas the COI sequence of I. galbana(AB000119) was substantially different. Using COI, we could dis-criminate between Eh and G. oceanica but only 4 out of the 17analyzed Eh strains from the CCMP and CCAP culture collections hadunique COI sequences (Figs. 4, S4). Despite this low COI diversityamong cultivated Eh strains from different oceanic settings (Figs. 4,S4), we found 11 unique COI sequences in the fossil record, indicativethat at least eleven unique Eh strains thrived in the Holocene photiczone of the Black Sea (Fig. 4). None of the fossil COI sequences wereclosely related to G. oceanica (Fig. 4).
EhCOI-6 (Figs. 3B, 4, S3 DGGE band 6) represents the first Eh strainto colonize the Black Sea at 7270 a BP (Table 1), which is ~300 yrs afterthe onset of sapropel deposition and after Isochrysis appeared. The useof specific primers targeting Eh COI thus slightly lowered the detectionlimit for Eh DNA since the oldest 18S rDNA of Eh (i.e. with generalhaptophyte primers) was found at 6430 a BP (Table 1). Eh COI wasabsent from the short interval between ~7550 and 7270 a BP whereonly 18S rDNA sequences related to Isochrysis spp. were found(Fig. 3A, B, Table 1).
Before 5250 a BP, EhCOI-6 was the dominant Eh, with short-termsuccessions by EhCOI-11 (~6800 a BP) and −2 (6550–6430 a BP).EhCOI-6 disappeared around ~5250 a BP (Fig. 3B), which is shortlybefore all Isochrysis-related haptophytes disappeared (Fig. 3A). After5250 a BP, EhCOI-1 occurred most frequently and predominated formany centuries. EhCOI-5, −7, −8, and 9 only appeared for brief(~50 yrs) periods, whereas EhCOI-6 shortly re-occurred and coincidedwith EhCOI-5 at 3360±68 a BP (Fig. 3B, Table 1). EhCOI-4 colonizedthe photic zone as early as 3600 a BP, but was abruptly replaced byEhCOI-1 when Unit I started to accumulate. The three strains found inUnit I (EhCOI-1, 2 & 3) appear to have predominated for periods of~300 yrs between 1700 and ~450 a BP; Fig. 3B). Note that EhCOI-2 also
olocene Black Sea sediments of GGC18 (colored boxes) to reference sequences obtainedeanica strains from the Provasoli-Guillard Center for Culture of Marine PhytoplanktonCOI sequences from GGC18 were determined from the DGGE represented in Fig. S3. Barstrains were present in the Black Sea are denoted after each sequence.
617M.J.L. Coolen et al. / Earth and Planetary Science Letters 284 (2009) 610–621
appeared during early sapropel deposition whereas EhCOI-3 wasunique to Unit I sediments.
QPCR revealed that the number of fossil EhCOI copies were ingeneral 1 to 2 orders of magnitude lower than those of totalhaptophyte 18S rDNA. As for haptophyte 18S rDNA, the fossil COIcontent increased by one order of magnitude from its first occurrenceat 7270 a BP towards the most recent analyzed layer (Fig. 1E).
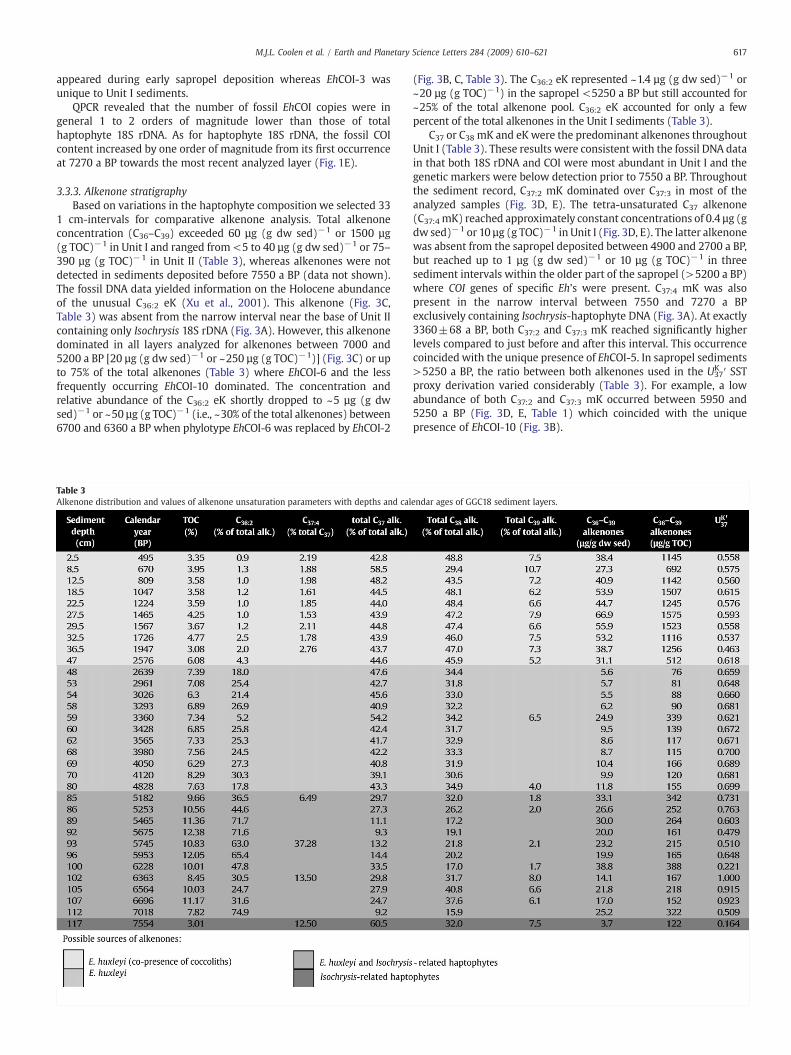
3.3.3. Alkenone stratigraphyBased on variations in the haptophyte composition we selected 33
1 cm-intervals for comparative alkenone analysis. Total alkenoneconcentration (C36–C39) exceeded 60 µg (g dw sed)−1 or 1500 µg(g TOC)−1 in Unit I and ranged from <5 to 40 µg (g dw sed)−1 or 75–390 µg (g TOC)−1 in Unit II (Table 3), whereas alkenones were notdetected in sediments deposited before 7550 a BP (data not shown).The fossil DNA data yielded information on the Holocene abundanceof the unusual C36:2 eK (Xu et al., 2001). This alkenone (Fig. 3C,Table 3) was absent from the narrow interval near the base of Unit IIcontaining only Isochrysis 18S rDNA (Fig. 3A). However, this alkenonedominated in all layers analyzed for alkenones between 7000 and5200 a BP [20 µg (g dw sed)−1 or ~250 µg (g TOC)−1)] (Fig. 3C) or upto 75% of the total alkenones (Table 3) where EhCOI-6 and the lessfrequently occurring EhCOI-10 dominated. The concentration andrelative abundance of the C36:2 eK shortly dropped to ~5 µg (g dwsed)−1 or ~50 µg (g TOC)−1 (i.e., ~30% of the total alkenones) between6700 and 6360 a BP when phylotype EhCOI-6 was replaced by EhCOI-2
Table 3Alkenone distribution and values of alkenone unsaturation parameters with depths and cal
(Fig. 3B, C, Table 3). The C36:2 eK represented ~1.4 µg (g dw sed)−1 or~20 µg (g TOC)−1) in the sapropel <5250 a BP but still accounted for~25% of the total alkenone pool. C36:2 eK accounted for only a fewpercent of the total alkenones in the Unit I sediments (Table 3).
C37 or C38 mK and eK were the predominant alkenones throughoutUnit I (Table 3). These results were consistent with the fossil DNA datain that both 18S rDNA and COI were most abundant in Unit I and thegenetic markers were below detection prior to 7550 a BP. Throughoutthe sediment record, C37:2 mK dominated over C37:3 in most of theanalyzed samples (Fig. 3D, E). The tetra-unsaturated C37 alkenone(C37:4 mK) reached approximately constant concentrations of 0.4 µg (gdw sed)−1 or 10 µg (g TOC)−1 in Unit I (Fig. 3D, E). The latter alkenonewas absent from the sapropel deposited between 4900 and 2700 a BP,but reached up to 1 µg (g dw sed)−1 or 10 µg (g TOC)−1 in threesediment intervals within the older part of the sapropel (>5200 a BP)where COI genes of specific Eh's were present. C37:4 mK was alsopresent in the narrow interval between 7550 and 7270 a BPexclusively containing Isochrysis-haptophyte DNA (Fig. 3A). At exactly3360±68 a BP, both C37:2 and C37:3 mK reached significantly higherlevels compared to just before and after this interval. This occurrencecoincided with the unique presence of EhCOI-5. In sapropel sediments>5250 a BP, the ratio between both alkenones used in the U37
K ′ SSTproxy derivation varied considerably (Table 3). For example, a lowabundance of both C37:2 and C37:3 mK occurred between 5950 and5250 a BP (Fig. 3D, E, Table 1) which coincided with the uniquepresence of EhCOI-10 (Fig. 3B).
endar ages of GGC18 sediment layers.

618 M.J.L. Coolen et al. / Earth and Planetary Science Letters 284 (2009) 610–621
Alkenone-inferred SSTs using the U37K ′ index calibration for Eh of
(Prahl et al., 1988) exhibited dramatic swings between 5 and 30 °C inUnit II sediments >5250 a BP (Fig. 3F). The alkenone SST slowlydecreased over the last few millennia from 19 °C to 15 °C with theexception of a colder interval (~12 °C) at around 1950 a BP. Thissystematic cooling and absence of dramatic fluctuations in alkenone-derived temperatures from ~5250 to ~450 a BP (Fig. 3F) coincidedwith the absence of Isochrysis-related species (Fig. 3A), lower C36:2 eKcontent (Fig. 3C), as well as a different assemblage of Eh strains ascompared to earlier in the record (Fig. 3B). Together, these observa-tions imply strong species/strain effects on alkenone composition.
3.4. Preservation of haptophyte DNA vs. alkenones
The ratio between total haptophyte 18S rDNA [copies (g TOC)−1]and total alkenones [µg (g TOC)−1] was determined from the Unit Iand II sediment layers to be an assessment of the level of preservationof both types of haptophyte molecular markers (Fig. 1F). The totalhaptophyte rDNA/alkenone ratio was fairly constant in the coccolith-bearing Unit I sediments (i.e., 1130±653), but the variability in thisratio was higher (5200±9125) in the coccolith-free sapropeldeposited after 4900 a BP. A decreasing trend of this ratio (435±610) could be observed from the oldest part of the sapropel(>4900 a BP). The EhCOI/alkenone ratio was a factor 5 lower in theUnit I sediments and fluctuated less (190±127 in sediments youngerthan 4900 a) as compared to the 18S rDNA/alkenone ratio. Adeceasing trend in the sapropel older than 4900 yrs was paralleledby the COI/alkenone ratio, although the latter ratio was ~9 timeslower than the 18S rDNA/alkenone ratio.
4. Discussion
4.1. Sediment ages and lithology
The lithology of Quaternary Black Sea sediments at depths below500 m is well known, facilitating identification of the distinctdepositional phases in our Holocene sediment record. The first visibleoccurrence of coccoliths is at 2576±63 a BP which is significantly laterthan estimated by Jones and Gagnon (1994) (cf. 3400 a BP). Althoughspeculative, this difference may be partly attributable to spatialdifferences in the first occurrence of coccolithophorid Eh strains ordifferences in the preservation of coccoliths. In contrast, our calibratedage for the onset of sapropel (Unit II) depositionwas ~7550 a BP, closelymatching the age determined by (Jones and Gagnon,1994) (7540 a BP).
4.2. Preservation of nucleic acids, alkenones, and coccoliths
The deep water anoxia that persisted during most of the Holocene(Sinninghe Damsté et al., 1993) resulted in excellent preservation oforganic matter in Unit I and II (~10 wt.%) (Calvert et al., 1987; Arthurand Dean,1998; Calvert and Karlin,1998; Galimov et al., 2002; and thisstudy). Such conditions, including the presence of hydrogen sulfide(e.g. 400 µM sulfide in the Black Sea today(Jørgensen et al., 2001), hasbeen shown to also provide excellent preservation conditions for fossilDNA in Holocene sediments (Coolen et al., 2004, 2006; D'Andrea et al.,2006; Coolen et al., 2007; Manske et al., 2008) and even late-Pleistocene sediments (Coolen and Overmann, 2007) althoughpreservation of fossil DNA seems not to be restricted to sulfidicenvironments (Bissett et al., 2005; Coolen et al., 2008; Corinaldesi etal., 2008). The total DNA content was high throughout Units I and II,with values typical of marine surface sediments containing freshlydeposited organic matter (Dell'Anno et al., 1999, 2002; Corinaldesi etal., 2005). Especially in Unit II sediments older than 3500 yrs, the totalDNA concentration showed only minor fluctuations, suggesting anexcellent preservation of the DNA. The DNA concentration was morevariable in sediment layers younger than 3500 yrs despite comparable
TOC contents in the sapropel deposited between 2550 and 4900 a BP.This variability in DNA content does not per se reflect small scalevariations in the preservation of the sedimentary DNA, but could alsoreflect differences in the biological origin of the bulk sedimentary DNAsince the total DNA content between organisms varies. During ourprevious study we showed that the ratio between the 18S rDNA copynumbers of haptophytes and total alkenones did not significantlychange throughout Unit I (Coolen et al., 2006). This finding indicatesthat preservation efficiencies of sedimentary nucleic acids in Unit I aresimilar to those of the relatively refractory alkenones.
In the current study, we were able to examine this ratio throughoutboth Units I and II (Fig. 1F). The ~10 fold decrease in this ratio in Unit IIsediments deposited between ~7550 and 4900 a BP suggests somepreferential degradation of haptophyte DNA relative to the alkenones.Since not all haptophyte phylotypes recovered from the sedimentsreflect that of alkenone producers, we also compared the ratio betweenEh COI and total alkenones. Again, this ratio was consistent down to~4900 a-old sediments, but decreased 4-fold in the older Unit IIsediments. However, the predominance of Isochrysis-related hapto-phytes as additional alkenone sources in this deeper interval alsocontributed to a lowering of the Eh COI/alkenone ratio. Overall, our dataimplies that haptophyteDNA iswell preserved in theHoloceneBlack Seasedimentary record.
4.3. Genetic markers for Eh
Recently, a gene encoding a protein with calcium-binding motifs,designated GPA, and involved in the formation of coccoliths in Eh, hasbeen shown to be a potentially powerful genomic marker differentiat-ing between various Eh strains (Schroeder et al., 2005), although G.oceanica cannot be identified based on the GPA gene. Our multipleattempts to amplify fossil GPA of Eh in the sediment record wereunsuccessful, yielding amplicons of the wrong length (data notshown). This could be indicative of the absence of the correct templateDNA in the fossil record.
Our study showed that, in contrast to GPA, COI appeared to be asuitable marker in discriminating between Eh and G. oceanica. Sincemany mitochondria are present in eukaryotic cells, mitochondrial(COI) gene copy numbers likely exceed that of the nuclear GPA gene,increasing the chances of successful amplification after severalthousand years of burial. Based on the partial COI gene, we wereable to follow the succession of at least 11 Eh strains throughout theHolocene in the Black Sea, although an even higher diversity waslikely since wewere unable to resolve all of the tested CCMP and CCAPEh strains. Nevertheless, a relationship between the haptophytesequences in the Holocene Black Sea and environmental change isapparent (see Sections 4.5 and 4.6).
4.4. Haptophyte sources for alkenones in the Holocene Black Sea sediments
The fossil DNA survey shows that the most abundant alkenoneproducers during early sapropel deposition were related or identicalto Isochrysis spp. from the Holocene sediment record of the brackishAce Lake, Antarctica (Coolen et al., 2004). Interestingly, Ace Lakeshares a similar hydrological history with the Black Sea: thepostglacial sea-level rise led to the incursion of marine waters intothe initially freshwater-filled meltwater lake (Bird et al., 1991). Thisinflux then resulted in density stratification and subsequent bottomwater anoxia, leading to the formation of an organic carbon-richsapropel. All alkenones in Ace Lake were derived from Isochrysis-related phylotypes since coccoliths or 18S rDNA of Eh were absent(Coolen et al., 2004). A comparison between the alkenones found inAce Lake and those from the Black Sea thus enables us to differentiateamong alkenones derived from Isochrysis vs. Eh.
For example, C37:4 mK, which is an important alkenone found in I.galbana (Marlowe et al., 1984; Patterson et al., 1994) is also the main

619M.J.L. Coolen et al. / Earth and Planetary Science Letters 284 (2009) 610–621
alkenone found in the Ace Lake sediments (Coolen et al., 2004). In AceLake, the closely related Isochrysis phylotypes were further sources ofC38 and C39 alkenones and occasionally produced the C37:3 mK, but notthe C37:2 mK (Coolen et al., 2004). This is in agreement with thealkenone distribution between 7550 and 7270 a BP (i.e. the presenceof C37:4 mK and the absence of the C37:2 mK) where Isochrysis is thesole source of alkenones.
Cultivars of Eh have also been shown to produce C37:4 mK(Volkman et al., 1980; Marlowe et al., 1984; Prahl et al., 1988) andsince DNA of Isochrysis (or Chrysotila) was absent from Unit I, thisalkenone must have been solely biosynthesized by Eh. A closer look atthe coinciding presence of fossil C37:4 mK and Eh DNA suggests thatthe biosynthesis of this alkenone is a feature that was unique tocertain Holocene Eh strains in the Black Sea. Specifically, C37:4 mKmaxima during early sapropel deposition coincided with the presenceof unique Eh strains (EhCOI-1, 2, and 10). If Isochrysis was the source,this alkenone should have been present in all analyzed layers of thiscore section. Whereas EhCOI-10 was unique to sapropel layers>5250 a BP, EhCOI-1 and −2 reappeared as a putative C37:4 mKproducer in Unit I. EhCOI-1 in the younger part of the sapropel did notproduce C37:4 mK, and might have been a different strain with anidentical sequence to EhCOI in Unit I. The strains that were unique tothe sapropel layers <5250 a BP (EhCOI-4, 5, 7, 8, and 9) were clearlynot C37:4 mK producers. In addition, EhCOI-6 was never a source ofC37:4 mK>5250 a ago and still did not make this alkenone when itreturned for only 50 yrs at ~3360 a BP. It thus seems more likely thatC37:4 mK production in Eh is a strain-specific feature and that thosestrains were selected by the lower salinities prevailing during Unit Ideposition (Van der Meer et al., 2008) and during early sapropeldeposition (Mudie et al., 2002).
We were also able to unravel the source of the unusual alkenone(C36:2 eK) in the Black Sea sapropel. Indirect indications for Eh as thesource of C36:2 eK came from a recent study of Prahl et al. (2006).These authors reported that a culture of E huxleyi strain CCMP 1742,which was used to establish the original U37
K ′-growth temperaturecalibration (Prahl et al., 1988), recently started to produce majoramounts of three new alkenones including a C36:2 eK (Prahl et al.,2006). However, since the positions of the double bonds of the C36:2eK in CCMP 1742 were different (ω15,22-C36 eK) (Prahl et al., 2006)than those reported for the Black Sea sediments (ω15,20-C36 eK) (Xuet al., 2001; Prahl et al., 2006), a biological precursor other than Ehcould not be excluded (Prahl et al., 2006).
Our fossil DNA data demonstrated that Eh strains rather than theIsochrysis-related haptophytes were sources of the C36:2 eK in theBlack Sea, since this compound was also abundant during latesapropel deposition (<4900 a BP), where only DNA of Eh, and notthat of Isochrysis, was found. This alkenone was furthermore absentfrom the narrow interval (7550–7270 cal. yr BP) where we onlydetected DNA of Isochrysis-related phylotypes but not that of Eh. Inaddition, since this alkenone was not reported from Ace Lake(Volkman et al., 1988; Schouten et al., 2001; Coolen et al., 2004) orfrom cultivars of I. galbana (Marlowe et al., 1984; Patterson et al.,1994), we can rule out Isochrysis-related species as sources of the C36:2eK in the Black Sea. Neither Gephyrocapsa nor Chrysotila evercolonized the Black Sea and, therefore, were also not sources.
Since DGGE is biased to identify only sequences that representmore than ~2% of the total pool of PCR-amplified DNA (e.g. Coolen etal., 2007), it is possible that certain haptophyte species weremissed aspossible additional sources of alkenones during this study. In order tosearch for haptophytes that were missed with the PCR/DGGEapproach, we recently explored “next generation” 454 tag pyrose-quencing (e.g Sogin et al., 2006) as a paleogenetic tool. This approachrevealed the near-to-complete eukaryotic diversity in the present-daywater column of the Black Sea and within sediment intervals ofGGC18, which were selected for 454 sequencing based on thehaptophyte community shifts observed by PCR/DGGE during this
study. This approach revealed ~30,000 eukaryotic 18S rDNAsequences per sample. The only additional haptophyte that wasrecovered from most of the analyzed samples using the ultrasensitive454 tag pyrosequencing technology was a species of the haptophytegenus Pavlova (Coolen and Davis, unpublished results). Despite thefact that our PCR/DGGE primers are selective for the 18S rDNA of themajority of haptophytes, 18S rDNA sequences of the genus Pavlova donot completely match with our primers. This explains why Pavlovawas not amplified during PCR and escaped detection. However, Pav-lova is not known to produce alkenones and is rather distantly relatedto the cluster of alkenone-producing haptophytes (Fig. 2).
4.5. Implications for alkenone-based SST reconstructions
SST reconstruction in the Black Sea is difficult because of the lack ofconventional proxies such as foraminiferal Mg/Ca ratios (Elderfieldand Ganssen, 2000). However, the exceptional preservation condi-tions for organic matter under anoxic deposition (no bioturbation)and relatively high sedimentation rates (~15–20 cm kyr−1) suggestthat exquisite paleoclimate records might be retrieved for theHolocene (e.g. spanning the Medieval Warm Period or the Little IceAge) using organic geochemical proxies. So, establishing whetheralkenone paleothermometry is feasible is an important goal.
It is likely that salinity was an important influence on theoccurrence of Isochrysis-related haptophytes and the presence ofunique Eh strains (i.e. EhCOI-6, 10 and 11). Highly variable amounts ofC37 alkenones but mostly low levels of the C37:3 mK were produced byIsochrysis and/or Eh during this early phase of sapropel deposition.Thus, species- and even strain-specific variations in alkenonecompositions would then explain the spurious U37
K ′ values andcorresponding SSTs that oscillated between 5 and 30 °C. The inclusionof C37:4 mK as part of the original U37
K index (Brassell et al., 1986)yielded lower apparent SSTs, but did not reduce the unrealisticallylarge swings in the alkenone SST record. Similarly, applying thealkenone SST calibration for I. galbana (Versteegh et al., 2001) did notmarkedly change the SST estimates (data not shown). This indicatesthat Isochrysis species from the Holocene Black Sea and Ace Lake havea different alkenone composition than I. galbana. Similarly, thealkenone composition of the Eh strains that thrived in the Black Sea>5250 a BP differed from the strains that prevailed in the Black Seaafter that period. Unless cultures of these haptophytes becomeavailable, it will remain impossible to confidently apply alkenoneunsaturation as a proxy for SST in the Black Sea (or in Ace Lake) duringearly sapropel deposition.
In contrast, from ~5250 a BP onwards a relatively stablehaptophyte assemblage dominated by a different suite of Eh strainsyielded robust alkenone-SSTestimates and indicated a gradual coolingfrom 19 °C to ~15 °C at the top of the record (~450 a BP). C36:2 eK stillrepresented 25% of the total alkenones in the younger sapropel layersand, therefore, the presence of this alkenone alone is not an indicatorfor future settings with unreliable alkenenone SST estimates.
A general cooling trend was also observed from alkenoneunsaturation ratios from mid-Holocene to recent Marmara Seasediments (Sperling et al., 2003). Although a direct comparison ofthe Black Sea SST recordwith that from theMarmara Sea is most likelynot straightforward and warrants further investigation, it seems thatfrom ~5250 a BP to the present, the alkenone-based SST reconstruc-tion at our coring site appears robust.
4.6. Possible environmental controls on the biosynthesis of the “Black Seaalkenone”
C36:2eK has also recently been found enriched in glacial depositsfrom the Japan Sea (Fujine et al., 2006). During glacial intervals,connectivity between the Japan Sea and the open Pacific was morerestricted and surface waters may have been considerably fresher

620 M.J.L. Coolen et al. / Earth and Planetary Science Letters 284 (2009) 610–621
owing to enhanced river input (Fujine et al., 2006). It was, therefore,suggested that the presence of this alkenone was salinity related.However, C36:2eK is also present in modern-day surface waters of theDYFAMED station in the Ligurian Sea (NW Mediterranean) (Rontaniet al., 2001) where Eh is a well-recognized component of thephytoplankton community (Ternois et al., 1997). Based on ourpaleogenetic data, we suspect that yet-to-be identified strains of Ehmight be the source of the C36:2 eK at this location. The salinity of thesurface waters in the Ligurian Sea varies between 20 and 35 whereasthe salinity inferred from past dinoflagellate compositions (i.e.dinocysts) at the onset of the deposition of the Black Sea's sapropel,was around 7 (Mudie et al., 2001, 2007; Hiscott et al., 2007). If Ehstrains were indeed the source of the C36:2 eK in both the HoloceneBlack Sea and the modern-day Ligurian Sea, salinity may not be thekey trigger for Eh to biosynthesize the Black Sea alkenone. Further-more, the calibration benchmark strain of Eh (CCMP 1742) was shownto produce increased levels of a similar C36 eK when the cultivar wasincubated in the dark and under nutrient-limited conditions (Prahlet al., 2006). Perhaps, therefore, nutrient availability was the trigger,possibly as a consequence of highly stratified surface waters in theBlack Sea during sapropel deposition (Sinninghe Damsté et al., 1993;Huang et al., 2000). Further studies will be required to address thisquestion.
5. Conclusions
Our combined lipid biomarker and fossil DNA analyses revealedthat haptophytes related to brackish Isochrysis spp. were the initialsources of alkenones in the Black Sea, and appeared immediately afterthe onset of sapropel deposition (~7550 a BP). Isochrysis-relatedhaptophytes were slowly replaced by a complex suite of E. huxleyi (Eh)strains as sources of alkenones. Paleogenetics showed that Eh's werepresent during sapropel deposition, ~4000 yrs earlier than previouslyrecognized, based on their preserved coccoliths. The unusual C36:2 eK“Black Sea alkenone”wasmost abundant between 7270 and 5250 a BPand coincided with the presence of three Eh strains which wereunique for that part of the core, suggesting that they were biologicalprecursors of C36:2 eK. In addition, we found that Isochrysis as well ascertain E. huxleyi strains were capable of producing the C37:4 mK atcertain intervals during sapropel and Unit I deposition.
Strong haptophyte species- and strain-specific effects wereobserved on the level of unsaturation of alkenones which resultedin spurious alkenone-derived SST estimates before 5250 a BP. Incontrast, from ~5250 a BP onwards, a relatively stable haptophyteassemblage dominated by a different suite of Eh strains yielded robustalkenone-SST estimates and indicated a gradual cooling from 19 °C to~15 °C at the top of the record (~450 a BP).
Acknowledgments
Wewould like to thank Al Gagnon, Daniel Montluçon, Drs. CorneliaWuchter and Angela Dickens (WHOI) as well as the R/V Akademikcrew and IOBAS researchers, and in particular Delcho Solakov, for theirextensive organizational and participatory help with the cruise, andCarl G. Johnson (WHOI) for the extensive TOC measurements. Inaddition, we would like to thank two anonymous and dedicatedreviewers for their suggestions to improve the manuscript. We aregrateful for the financial support from US National Science Foundationgrant OCE 0602423, as well as funding from WHOI's Access to the Seaprogram, and a grant from the Andrew W. Mellon FoundationEndowed Fund for Innovative Research.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.epsl.2009.05.029.
References
Aksu, A.E., Abrajano, T., Mudie, P.J., Yasar, D., 1999. Organic geochemical andpalynological evidence for terrigenous origin of the organic matter in Aegean Seasapropel S1. Mar. Geol. 153, 303–318.
Aksu, A.E., Hiscott, R.N., Kaminski, M.A., Mudie, P.J., Gillespie, H., Abrajano, T., Yasar, D.,2002a. Last glacial–Holocene paleoceanography of the Black Sea and Marmara Sea:stable isotopic foraminiferal and coccolith evidence. Mar. Geol. 190, 119–149.
Aksu, A.E., Mudie, P.J., Rochon, A., Kaminski, M.A., Abrajano, T., Yasar, D., 2002b.Persistent Holocene outflow from the Black Sea to the Eastern Mediterraneancontradicts Noah's Flood hypothesis. GSA Today 12, 4–10.
Arthur, M.A., Dean, W.E., 1998. Organic-matter production and preservation andevolution of anoxia in the Holocene Black Sea. Paleoceanography 13, 395–411.
Arthur, M.A., Dean, W.A., Neff, E.D., Hay, B.J., King, J., Jones, G., 1994. Varve calibrationrecords of carbonate and organic carbon accumulation over the last 2000 years inthe Black Sea. Glob. Biogeochem. Cycles 8, 195–217.
Bahr, A., Lamy, F., Arz, H., Kuhlmann, H., Wefer, G., 2005. Late glacial to Holocene climateand sedimentation history in the NW Black Sea. Mar. Geol. 214, 309–322.
Bahr, A., Arz, H.W., Lamy, F., Wefer, G., 2006. Late glacial to Holocene paleoenviron-mental evolution of the Black Sea, reconstructed with stable oxygen isotope recordsobtained on ostracod shells. Earth Planet. Sci. Lett. 241, 863–875.
Bertrand, S., Hughen, K., Sepúlveda, J.A., Pantoja, S., Lange, C., 2007. Late Holoceneclimate variability of Northern Patagonia reconstructed by a multi-proxy analysisfjord sediments (44–47° S). Geophysical Research Abstracts. InEuropean Geos-ciences Union (EGU), Vienna, p. 01568.
Bird, M.I., Chivas, A.R., Radnell, C.J., Burton, H.R., 1991. Sedimentological and stable-isotope evolution of lakes in the Vestfold Hills, Antarctica. Palaeogeogr. Palaeocli-matol. Palaeoecol. 84, 109–130.
Bissett, A., Gibson, J.A.E., Jarman, S.N., Swadling, K.M., Cromer, L., 2005. Isolation,amplification, and identification of ancient copepod DNA from lake sediments.Limnol. Oceanogr. Methods 3, 533–542.
Brassell, S.C., Eglinton, G., Marlowe, I.T., Pflaumann, U., Sarnthein, M., 1986. Molecularstratigraphy: a new tool for climatic assessment. Nature 320, 129–133.
Calvert, S.E., Karlin, R.E., 1998. Organic carbon accumulation in the Holocene sapropel ofthe Black Sea. Geology 26 (2), 107–110 1998. Geology 26, 107–110.
Calvert, S.E., Vogel, J.S., Southon, J.R., 1987. Carbon accumulation rates and the origin ofthe holocene sapropel in the Black Sea. Geology 15, 918–921.
Coolen, M.J.L., Overmann, J., 2007. 217,000-year-old DNA sequences of green sulfurbacteria inMediterranean sapropels and their implications for the reconstruction ofthe paleoenvironment. Environ. Microbiol. 9, 238–249.
Coolen, M.J.L., Muyzer, G., Rijpstra, W.I.C., Schouten, S., Volkman, J.K., Sinninghe Damsté,J.S., 2004. Combined DNA and lipid analyses of sediments reveal changes inHolocene haptophyte and diatom populations in an Antarctic lake. Earth Planet. Sci.Lett. 223, 225–239.
Coolen, M.J.L., Boere, A., Abbas, B., Baas, M., Wakeham, S.G., Sinninghe Damsté, J.S., 2006.Fossil DNA derived from alkenone-biosynthesizing haptophytes and other algae inHolocene sediment from the Black Sea. Paleoceanography 21, PA1005. doi:10.1029/2005PA001188.
Coolen, M.J.L., Volkman, J.K., Abbas, B., Muyzer, G., Schouten, S., Sinninghe Damsté, J.S.,2007. Identification of organic matter sources in sulfidic late Holocene Antarcticfjord sediments from fossil rDNA sequence analysis. Paleoceanography 22, PA2211.doi:10.1029/2006PA001309.
Coolen, M.J.L., Talbot, H.M., Abbas, B.A., Ward, C., Schouten, S., Volkman, J.K., SinningheDamsté, J.S., 2008. Sources for sedimentary bacteriohopanepolyols as revealed by16S rDNA stratigraphy. Environ. Microbiol. 10, 1783–1803.
Corinaldesi, C., Danovaro, R., Dell'Anno, A., 2005. Simultaneous recovery of extracellularand intracellular DNA suitable for molecular studies from marine sediments. Appl.Environ. Microbiol. 71, 46–50.
Corinaldesi, C., Beolchini, F., Dell'Anno, A., 2008. Damage and degradation rates ofextracellular DNA in marine sediments: implications for the preservation of genesequences. Mol. Ecol. 17, 3939–3951.
Croudace, I.W., Rindby, A., Rothwell, G.R., 2005. ITRAX: description and evaluation of anew X-ray core scanner. In: Rothwell, G.R. (Ed.), New Techniques in Sediment CoreAnalysis. InGeological Society, London, pp. 51–63.
D'Andrea, W.J., Lage, M., Martiny, J.B.H., Laatsch, A.D., Amaral-Zettler, L.A., Sogin, M.L.,Huang, Y., 2006. Alkenone producers inferred from well-preserved 18S rDNA inGreenland lake sediments. J. Geophys. Res. 111, G0313. doi:10.1029/2005JG000121.
Dell'Anno, A., Fabiano, M., Mei, M.L., Danovaro, R., 1999. Pelagic-benthic coupling ofnucleic acids in an abyssal location of the northeastern Atlantic ocean. Appl.Environ. Microbiol. 65, 4451–4457.
Dell'Anno, A., Bompadre, S., Danovaro, R., 2002. Quantification, base composition, andfate of extracellular DNA in marine sediments. Limnol. Oceanogr. 47, 899–905.
Eglinton, T.I., Benitez-Nelson, B.C., Pearson, A., McNichol, A.P., Bauer, J.E., Druffel, E.R.M.,1997. Variability in radiocarbon ages of individual organic compounds frommarinesediments. Science 277, 796–799.
Eglinton, T.I., Eglinton, G., Dupont, L., Sholkovitz, E.R., Montlucon, D., Reddy, C.M., 2002.Composition, age, and provenance of organic matter in NW African dust over theAtlantic Ocean. Geochem. Geophys. Geosyst. 3 2001GC000269.
Elderfield, H., Ganssen, G., 2000. Past temperature and δ 18O of surface ocean watersinferred from foraminiferal Mg/Ca ratios. Nature 405, 442–445.
Fujine, K., Yamamoto, M., Tada, R., Kido, Y., 2006. A salinity-related occurrence of a novelalkenone and alkenoate in Late Pleistocene sediments from the Japan Sea. Org.Geochem. 37, 1074–1084.
Galimov, E.M., Kodina, L.A., Zhiltsova, L.I., Tokarev, V.G., Vlasova, L.N., Bogacheva, M.P.,Korobeinik, G.S., Vaisman, T.I., 2002. Organic carbon geochemistry in the north-western Black Sea–Danube River system. Estuar. Coast. Shelf Sci. 54, 631–641.

621M.J.L. Coolen et al. / Earth and Planetary Science Letters 284 (2009) 610–621
Gunasekera, R.M., Patil, J.G., McEnnulty, F.R., Bax, N.J., 2005. Specific amplification of mt-COI gene of the invasive gastropod Maoricolpus roseus in planktonic samplesreveals a free-living larval life-history stage. Mar. Freshw. Res. 56, 901–912.
Hay, B.J., 1988. Sediment accumulation in the central western Black Sea over the past5100 years. Paleoceanography 3, 491–508.
Hay, B.J., Arthur, M.A., Dean, W.A., Neff, E.D., Honjo, S., 1991. Sediment deposition in theLate Holocene abyssal Black Sea with climatic and chronological implications.Deep-Sea Res. 38, 1211–1235.
Hiscott, R.N., Aksu, A.E., Mudie, P.J., Marret, F., Abrajano, T., Kaminski, M.A., Evans, J.,Cakiroglu, A.I., Yasar, D., 2007. A gradual drowning of the southwestern Black Seashelf: evidence for a progressive rather than abrupt Holocene reconnectionwith theeasternMediterranean Sea through theMarmara Sea Gateway. Quat. Int.167,19–34.
Huang, Y., Freeman, K.H., Wilkin, R.T., Arthur, M.A., Jones, A.D., 2000. Black Seachemocline oscillations during the Holocene: molecular and isotopic studies ofmarginal sediments. Org. Geochem. 31, 1525–1531.
Jones, G.A., Gagnon, A.R., 1994. Radiocarbon chronology of Black Sea sediments. Deep-Sea Res., Part 1, Oceanogr. Res. Pap. 41, 531–557.
Jørgensen, B.B., Weber, A., Zopfi, J., 2001. Sulfate reduction and anaerobic methaneoxidation in black Sea sediments. Deep-Sea Res. I 48, 2097–2120.
Kaminski, M.A., Aksu, A., Box, M., Hiscott, R.N., Filipescu, S., Al-Salameen, M., 2002. LateGlacial to Holocene benthic foraminifera in the Marmara Sea: implications for BlackSea–Mediterranean Sea connections following the last deglaciation. Mar. Geol. 190,165–202.
Kaplin, P.A., Selivanov, A.O., 2004. Late Glacial and Holocene sea level changes in semi-enclosed seas of North Eurasia: examples from the contrasting Black and WhiteSeas. Palaeogeogr. Palaeoclimatol. Palaeoecol. 209, 19–36.
Kwiecien, O., Arz, H.W., Lamy, F., Plessen, B., Bahr, A., Haug, G.H., 2009. North Atlanticcontrol on precipitation pattern in the eastern Mediterranean/Black Sea regionduring the last glacial. Quat. Res. 71, 375–384.
Ludwig, W., Strunk, O., Westram, R., Richter, L., Meier, H., Yadhukumar, Buchner, A., Lai,T., Steppi, S., Jobb, G., Förster, W., Brettske, I., Gerber, S., Ginhart, A.W., Gross, O.,Grumann, S., Hermann, S., Jost, R., König, A., Liss, T., Lüßmann, R., May, M., Nonhoff,B., Reichel, B., Strehlow, R., Stamatakis, A., Stuckmann, N., Vilbig, A., Lenke, M.,Ludwig, T., Bode, A., Schleifer, K.-H., 2004. ARB: a software environment forsequence data. Nucleic Acids Res. 32, 1363–1371.
Major, C., Ryan,W., Lericolais, G., Hajdas, I., 2002. Constraints on Black Sea outflow to theSea of Marmara during the last glacial–interglacial transition. Mar. Geol. 190, 19–34.
Major, C.O., Goldstein, S.L., Ryan, W.B.F., Lericolais, G., Piotrowski, A.M., Hajdas, I., 2006.The co-evolution of Black Sea level and composition through the last deglaciationand its paleoclimatic significance. Quat. Sci. Rev. 25, 2031–2047.
Manske, A.K., Henssge, U., Glaeser, J., Overmann, J., 2008. Subfossil 16S rRNA genesequences of green sulfur bacteria in the Black Sea and their implications for pastphotic zone anoxia. Appl. Environ. Microbiol. 74, 624–632.
Marlowe, I.T., Green, J.C., Neal, A.C., Brassell, S.C., Eglinton, G., Course, P.A., 1984. Long-Chain (n-C37–C39) alkenones in the Prymnesiophyceae. Distribution of alkenonesand other lipids and their taxonomic significance. Br. Phycol. J. 19, 203–216.
Medlin, L., Barker, G.L.A., Baumann, M., Hayes, P.K., Lange, M., 1994. Molecular biologyand systematics. In: Green, J.C., Leadbeater, B.S.C. (Eds.), The Haptophyte Algae.InClarenden Press, Oxford, pp. 393–411.
Monti, M.M., Nappo, A.G., Giorgini, M., 2005. Molecular characterization of closelyrelated species in the parasitic genus Encarsia (Hymenoptera:Aphelinidae) basedon the mitochondrial cytochrome oxidase subunit I gene. Bull. Entomol. Res. 95,401–408.
Mudie, P.J., Aksu, A.E., Yasar, D., 2001. Late Quaternary dinoflagellate cysts from theBlack, Marmara and Aegean seas: variations in assemblages, morphology andpaleosalinity. Mar. Micropaleontol. 43, 155–178.
Mudie, P.J., Rochon, A., Aksu, A.E., Gillespie, H., 2002. Dinoflagellate cysts, freshwateralgae and fungal spores as salinity indicators in Late Quaternary cores fromMarmara and Black seas. Mar. Geol. 190, 203–231.
Mudie, P.J., Marret, F., Aksu, A.E., Hiscott, R.N., Gillespie, H., 2007. Palynological evidencefor climatic change, anthropogenic activity and outflow of Black Sea water duringlate Pleistocene and Holocene: centennial- to decadal-scale records from the Blackand Marmara Seas. Quat. Int. 167, 73–90.
Muyzer, G., De Waal, E.C., Uitterlinden, A.G., 1993. Profiling of complex microbialpopulations by denaturing gradient gel electrophoresis analysis of polymerasechain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microbiol. 59,695–700.
Paine, M.A., McDowell, J.R., Graves, J.E., 2008. Specific identification using COI sequenceanalysis of scombrid larvae collected off the Kona coast of Hawaii Island. Ichthyol.Res. 55, 7–16.
Patterson, G.W., Tsitsa-Tsardis, E., Wikfors, G.H., Gladu, P.K., Chitwood, D.J., Harrison, D.,1994. Sterols and alkenones of Isochrysis. Phytochemistry 35, 1233–1236.
Prahl, F.G., Wakeham, S.G., 1987. Calibration of unsaturation patterns in long-chainketone compositions for palaeotemperature assessment. Nature 330, 367–369.
Prahl, F.G., Muehlhausen, L.A., Zahnle, D.B., 1988. Further evaluation of long-chainalkenones as indicators of paleoceanographic conditions. Geochim. Cosmochim.Acta 52, 2303–2310.
Prahl, F.G., Rontani, J.F., Volkman, J.K., Sparrow,M.A., Royer, I.M., 2006. Unusual C-35 andC-36 alkenones in a paleoceanographic benchmark strain of Emiliania huxleyi.Geochim. Cosmochim. Acta 70, 2856–2867.
Pruesse, E., Quast, C., Knittel, K., Fuchs, B.M., Ludwig, W.G., Peplies, J., Glockner, F.O.,2007. SILVA: a comprehensive online resource for quality checked and alignedribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 35,7188–7196.
Quan, J.X., Zhuang, Z.M., Deng, J.Y., Dai, J.X., Zhang, Y.P., 2004. Phylogenetic relationshipsof 12 Penaeoidea shrimp species deduced from mitochondrial DNA sequences.Biochem. Genet. 42, 331–345.
Reimer, P.J., Baillie, M.G.L., Bard, E., Bayliss, A., Beck, J.W., Bertrand, C.J.H., Blackwell, P.G.,Buck, C.E., Burr, G.S., Cutler, K.B., Damon, P.E., Edwards, R.L., Fairbanks, R.G.,Friedrich, M., Guilderson, P., Hogg, A.G., Hughen, K.A., Kromer, B., McCormac, G.,Manning, S., Ramsey, C.B., Reimer, R.W., Remmele, S., Southon, J.R., Stuiver, M.,Talamo, S., Taylor, F.W., Van der Plicht, J., Weyhenmeyer, C.E., 2004. IntCal04terrestrial radiocarbon age calibration, 0–26 cal kyr BP. Radiocarbon 46, 1029–1058.
Rontani, J.-F., Marchand, D., Volkman, J.K., 2001. NaBH4 reduction of alkenones to thecorresponding alkenols: a useful tool for their characterisation in natural samples.Org. Geochem. 32, 1329–1341.
Rontani, J.-F., Beker, B., Volkman, J.K., 2004. Long-chain alkenones and relatedcompounds in the benthic haptophyte Chrysotila lamellosa Anand HAP 17.Phytochemistry 65, 117–126.
Ryan, W.B.F., Pitman III, W.C., Major, C.O., Shimkus, K., Moskalenko, V., Jones, G.A.,Dimitrov, P., Gorür, N., Sakinç, M., Yüce, H., 1997. An abrupt drowning of the BlackSea shelf. Mar. Geol. 138, 119–126.
Saitou, N., Nei, M., 1987. The neighbor-joiningmethod: a newmethod for reconstructingphylogenetic trees. Mol. Biol. Evol. 4, 406–425.
Sawada, K., Shiraiwa, Y., 2004. Alkenone and alkenoic acid compositions of themembrane fractions of Emiliania huxleyi. Phytochemistry 65, 1299–1307.
Schouten, S., Rijpstra, W.I.C., Kok, M., Hopmans, E.C., Summons, R.E., Volkman, J.K.,Sinninghe Damsté, J.S., 2001. Molecular organic tracers of biogeochemical processesin a saline meromictic lake (Ace Lake). Geochim. Cosmochim. Acta 65, 1629–1640.
Schroeder, D.C., Biggi, G.F., Hall, M., Davy, J., Martinez, J.M., Richardson, A.J., Malin, G.,Wilson, W.H., 2005. A genetic marker to separate Emiliania huxleyi (Prymnesio-phyceae) morphotypes. J. Phycol. 41, 874–879.
Sinninghe Damsté, J.S., Wakeham, S.G., Kohnen, M.E.L., Hayes, J.M., De Leeuw, J.W., 1993.A 6000-year sedimentary molecular record of chemocline excursions in the BlackSea. Nature 362, 827–829.
Sogin, M.L., Morrison, H.G., Huber, J.A., Welch, D.M., Huse, S.M., Neal, P.R., Arrieta, J.M.,Herndl, G.J., 2006. Microbial diversity in the deep sea and the underexplored “rarebiosphere”. Proc. Natl. Acad. Sci. 103, 12115–12120.
Sperling, M., Schmiedl, G., Hemleben, C., Emeis, K.C., Erlenkeuser, H., Grootes, P.M.,2003. Black Sea impact on the formation of eastern Mediterranean sapropel S1?Evidence from theMarmara Sea. Palaeogeogr. Palaeoclimatol. Palaeoecol. 190, 9–21.
St-Onge, G., Mulder, T., Francus, P., Long, B., 2007. Continuous physical properties ofcored marine sediments. In: Hillaire-Marcel, C., De Vernal, A. (Eds.), Proxies in LateCenozoic Paleoceanography. InElsevier, Amsterdam, pp. 63–98.
Stothard, J.R., Rollinson, D., 1997. Partial DNA sequences from the mitochondrialcytochrome oxidase subunit I (COI) gene can differentiate the intermediate snailhosts Bulinus globosus and B-nasutus (Gastropoda:Planorbidae). J. Nat. Hist. 31,727–737.
Stuiver, M., Reimer, P.J., 1993. Extended C-14 data-base and revised calib 3.0 C-14 agecalibration program. Radiocarbon 35, 215–230.
Ternois, Y., Sicre, M.A., Boireau, A., Conte, M.H., Eglinton, G., 1997. Evaluation of long-chain alkenones as paleo-temperature indicators in the Mediterranean Sea. Deep-Sea Res., Part 1, Oceanogr. Res. Pap. 44, 271–286.
Van der Meer, M.T.J., Sangiorgi, F., Baas, M., Brinkhuis, H., Sinninghe Damsté, J.S.,Schouten, S., 2008. Molecular isotopic and dinoflagellate evidence for late Holocenefreshening of the Black Sea. Earth Planet. Sci. Lett. 267, 426–434.
Versteegh, G.J.M., Riegman, R., De Leeuw, J.W., Jansen, J.H.F., 2001. U(37)(K′)values forIsochrysis galbana as a function of culture temperature, light intensity and nutrientconcentrations. Org. Geochem. 32, 785–794.
Volkman, J.K., Eglinton, G., Corner, E.D.S., Forsberg, T.E.V., 1980. Long-chain alkenes andalkenones in the marine coccolithophorid Emiliania huxleyi. Phytochemistry 19,2619–2622.
Volkman, J.K., Burton, H.R., Everitt, D.A., Allen, D.I., 1988. Pigment and lipidcompositions of algal and bacterial communities in Ace Lake, Vestfold Hills,Antarctica. Hydrobiologia 165, 41–57.
Volkman, J.K., Barrett, S.M., Blackburn, S.I., Sikes, E.L., 1995. Alkenones in Gephyrocapsaoceanica: implications for studies of paleoclimate. Geochim. Cosmochim. Acta 59,513–520.
Xu, L., Reddy, C.M., Farrington, J.W., Frysinger, G.S., Gaines, R.B., Johnson, C.G., Nelson, R.K.,Eglinton, T.I., 2001. Identification of a novel alkenone in Black Sea sediments. Org.Geochem. 32, 633–645.