dna x rna - physik.uni-muenchen.de · editing zu handeln. insgesamt - bei unabhängigem gebrauch...
TRANSCRIPT

RNA InteractionsMotivation
Often in Systems Biology, the concentrations of RNA is measured to infer the concentra-tions of the expressed proteins. While this is possible in a highly parallel way (-> Affyme-trix, Realtime PCR). However this approach makes several crucial assumptions such as:- Protein decay is not dependent on protein- No regulation or splicing at the RNA level happens.
To understand the latter caveat, we will give an introduction to possible interactions at the RNA level.
DNA
RNA
Protein
X

Recapitulation: Neural Guidance
Neurale Zelladhäsionsmoleküle der Immunoglobulin-Superfamilie bei Verte-braten und Invertebraten. Die Familienzugehörigkeit wird durch die soge-nannten Immunoglobulin-Domänen des extrazellulären Bereichs festgelegt (durch die Loopstruktur und S S im Schema angedeutet). Dies sind Abschnitte von 40-70 AS, die durch Disulfidbrücken in einer b-Faltblattstruktur zusam-mengehalten werden. Die rechteckigen Boxen stehen für eine zweite Art von wiederholten Domänen, über die einige der gezeigten Moleküle verfügen. Diese Sequenzelemente von etwa 100 AS Länge zeigen Homologie zu der Fibronec-tin-Typ-III-Domäne.
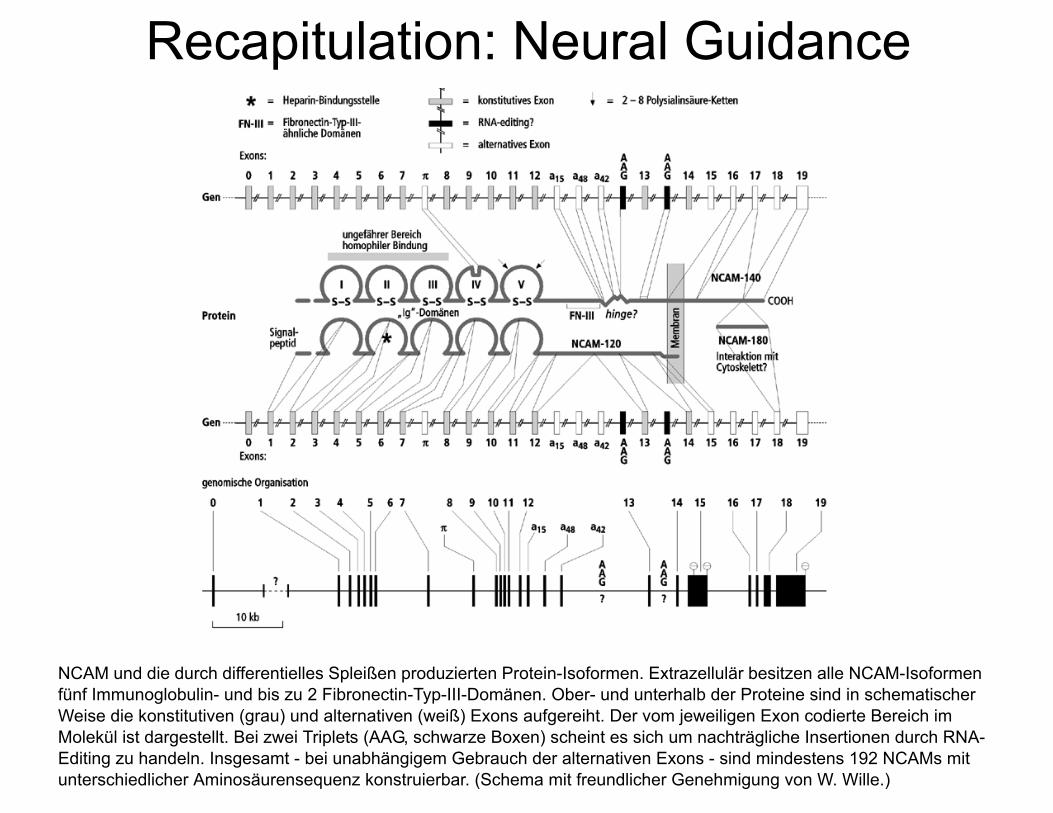
NCAM und die durch differentielles Spleißen produzierten Protein-Isoformen. Extrazellulär besitzen alle NCAM-Isoformen fünf Immunoglobulin- und bis zu 2 Fibronectin-Typ-III-Domänen. Ober- und unterhalb der Proteine sind in schematischer Weise die konstitutiven (grau) und alternativen (weiß) Exons aufgereiht. Der vom jeweiligen Exon codierte Bereich im Molekül ist dargestellt. Bei zwei Triplets (AAG, schwarze Boxen) scheint es sich um nachträgliche Insertionen durch RNA-Editing zu handeln. Insgesamt - bei unabhängigem Gebrauch der alternativen Exons - sind mindestens 192 NCAMs mit unterschiedlicher Aminosäurensequenz konstruierbar. (Schema mit freundlicher Genehmigung von W. Wille.)
Recapitulation: Neural Guidance

SplicingInternal, non-coding, segments of the primary transcript (introns) are removed, i.e. spliced out by the spliceosome. The exons remain and are joined together to make the mature mRNA. The spliceosome is a ribonucleoprotein complex. Each intron is flanked by consen-sus splice donor (GU) and splice acceptor (AG) sites at the 5' and 3' end of the intron, respectively.
Eukaryotic genes contain introns

SplicingHow does splicing work? The spliceosome removes the intron in a chemically interesting way: A specific adenosine residue within the intron attacks the 5’ phosphate end of the intron with its 2’OH residue (RNA!) thus forming a branched (lariat) RNA structure. Then the free 3’ OH of exon I attacks the 5’ end of exon II thus replacing the 3’ end of the intron. The splicing is catalyzed by small nuclear ribonucleoprotein particles (snRNPs), which con-tain both RNAs (U1, U2, ... to U6) and proteins. Some of the snRNAs are complementary to exon-intron boundaries.

R-Looping Experiments
DNA created by reverse transcrip-tion of mature RNA hybridizes with RNA by forming loops of the spliced introns.
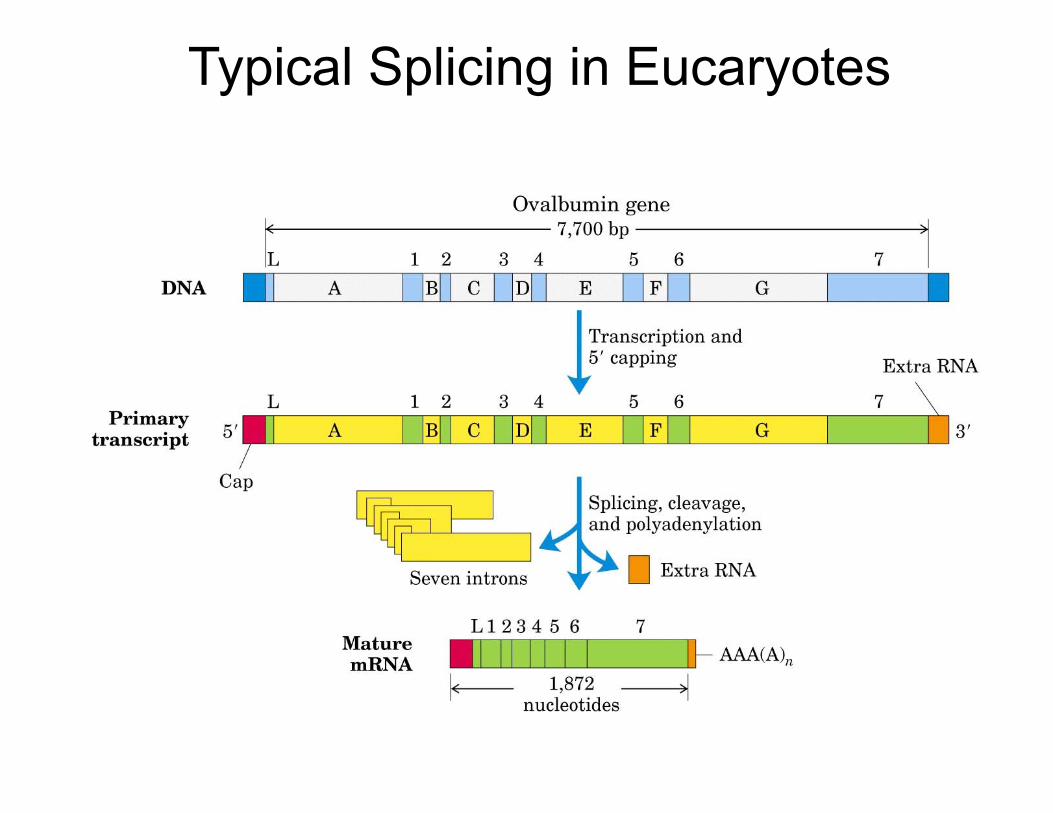
Typical Splicing in Eucaryotes

Definition of IntronSpecial Sequences define the start and the end of the intron.

Group I introns, for example in the rRNA of the ciliate Tetrahymena, are somewhat more primitive than the group II introns. These introns use a free GTP as a cofactor. The GTP is bound by the intron and used as a 'key' to unlock the 5' end of the intron from exon I. Then the 3' OH of exon I can take the place of the 3' OH of the intron to excise the intron entirely and thus fuse exon I to exon II. Tom Cech shared the Nobel prize with Sid Altmann for this discovery of a catalytic activity of RNA.
Self-Splicing Introns

Self-Splicing IntronsThe process requires complex secondary structure of RNA.

Alternative SplicingDepending on the state of a cell, special exons can also be spliced with the flanking introns, leading to shortened proteins depending on cofactors:

RNA processing occurs by a variety of mechanisms to convert a primary transcript into a final function RNA product
Eukaryotic pre-mRNAs are capped, polyadenylated, and spliced to yield one or more mature mRNAs before transport to the cytoplasm. These processes are coupled in the nucleus so that only properly processed mRNAs are exported to the cytoplasm.
The role of introns is still controversial but the favored hypothe-sis is that they arose early in evolution and allowed recombina-tion between mini-genes. They have been almost eliminated in bacteria and many lower eukaryotes perhaps because these
Conclusion Splicing