dynamics, control and synchronization of some models of neuronal oscillators
TRANSCRIPT

UNIVERSITE DE DSCHANG
************
ECOLE DOCTORALE
************
UNITE DE FORMATION DOCTORALE
SCIENCES FONDAMENTALES ET
TECHNOLOGIES
*************
UNIVERSITY OF DSCHANG
************
POSTGRADUATE SCHOOL
************
DOCTORAL TRAINING UNIT
FUNDAMENTAL SCIENCES AND
TECHNOLOGIES
*************DEPARTEMENT DE PHYSIQUE
DEPARTMENT OF PHYSICS
LABORATOIRE D’ELECTRONIQUE ET DE TRAITEMENT DU SIGNAL
LABORATORY OF ELECTRONICS AND SIGNAL PROCESSING (LETS)
THESIS
Presented for the achievement of the grade ofDoctorat / Ph.D degree in Physics
Option: Electronics
ByMEGAM NGOUONKADI Elie Bertrand
Registration number: 02S099
M. Sc. in Physics, Option: Electronics
Under the supervision of:FOTSIN Hilaire Bertrand
Associate Professor
2014-2015
Dynamics, control and synchronization of some models of neuronal oscillators

Dynamics, control and
synchronization of some models
of neuronal oscillators
Title
2/42

Outline
General Introduction
1. Bifurcation and multistability in the
extended Hindmarsh-Rose neuronal
oscillator2. Phase synchronization of bursting neural
networks
General conclusion and outlook
3/42
3. Robust synchronization of a small
pacemaker neuronal ensemble via
nonlinear controller: electronic circuit
design

General introduction
Figure 1: Electrical phenomena are the main information processing in the brain
General introduction 5/42
Neurosciences are one the main research topics of the century. The human brain is one ofthe most complex systems in science and understanding how it works is as old a question asmankind. The expression "computational neuroscience" reflects the possibility ofgenerating theory of brain function in terms of the information-processing properties ofstructures that make up the nervous system. It implies that we ought to be able to exploitthe conceptual and technical resources of computational research to help find explanationsof how neuronal structures achieve their effects, what functions are executed by neuronalstructures, and the nature of the states represented by the nervous system.
Neuronal activity modeling and network models
(a) (b)
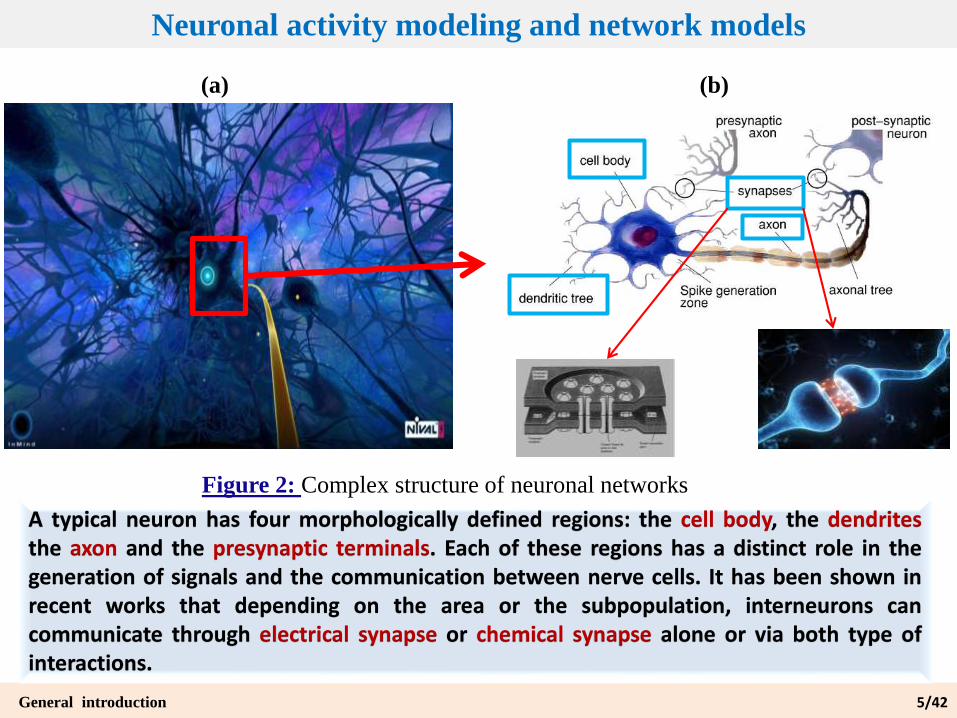
Figure 2: Complex structure of neuronal networks
General introduction 5/42
A typical neuron has four morphologically defined regions: the cell body, the dendritesthe axon and the presynaptic terminals. Each of these regions has a distinct role in thegeneration of signals and the communication between nerve cells. It has been shown inrecent works that depending on the area or the subpopulation, interneurons cancommunicate through electrical synapse or chemical synapse alone or via both type ofinteractions.

Neuronal activity modeling and network models
FitzHugh-Nagumo model: Fitzhugh and Nagumo 1961
Morris-Lecar model: Morris and Lecar 1981
Hindmarsh-Rose model : Hindmarsh J. L. et al. 1982, 1984
Extended Hindmarsh-Rose model : Selverston A. I.et al. 2000, Pinto R. et al. 2000
Why The extended Hindmarsh-Rose (eHR) neuronal model?
Several details of the shape of spiking-bursting activity, can be adjusted with the helpof this extended model.
It can describe the calcium exchange between intracellular warehouse and thecytoplasm, to completely produce the chaotic behavior of the stomatogastric ganglionneurons.
A better adjustment of the behavior of electronics neurons, when connected to itsliving counterpart, is better represented by the fourth order HR model.
General introduction 6/42
The integate and fire model: Lapicque L. 1907
Using the Hodking-Huxley model is biophysically prohibitive, since we can only simulate a
handful of neurons in real time.
In contrast, using the integrate-and-fire model is computationally effective, but the model is
simple and unable to produce the rich spiking and bursting dynamics exhibited by cortical
neurons.

Model’s equations:
Here, a, b, c, d, e, f , g, μ, s, h, v, k, r and l, are constants which express thecurrent and conductance based dynamics.
IDC represents the injected current.
x represents the membrane voltage, and y a fast current. z is a slow current, w isa slow dynamical process.
The parameters μ and v play a very important role in neuron activity. The firstrepresents the ratio of time scales between fast and slow fluxes across theneuron’s membrane and the second controls the speed of variation of the slowcurrent.
2 3
2
- -
- - -
-
-
DCx ay bx cx dz I
y e fx y gw
z z s x h
w v kw r y l
&
&
&
&
(1)
General introduction 7/42

Problems and objectives
1- From a nonlinear dynamical systems point of view, does the eHR neuronaloscillator behavior bring out how neurons respond to stimulus? Does themodel present the multistability mechanism?
2- How the dynamic chemical synapse, particularly the neurotransmittersbinding time constant influences the time delayed interactions of burstingneurons, since one knows that chemical synapses and neurons are dynamicalnonlinear devices? Are there time-delay induced phase-flip transitions to or outof synchrony when the chemical and electrical synapses are takensimultaneously into account?
3- The third goal of this work is to study the synchronized behavior of anexternal neuron and a complex network constituted of the pacemaker groupneurons of the lobster's pyloric CPG.
General introduction 8/42
In 1948, Hodgkin found that, injecting a DC-current of different amplitude in isolatedaxons, results in the production of repetitive spiking and inhibition with differentfrequencies.These observations were investigated a few decades later by Rinzel and Ermentrout. Theyshow that the observed behaviors are due to different bifurcation mechanisms.

Nonlinear Physics formalisms and assumptions
Phase space reconstructionFractal dimensionsThe Lyapunov exponentsSymbolic dynamicsAutocorrelation and cross correlation functionsMutual information
General introduction 9/42
1- The coupled individual dynamicalsystems are all identical2- The same function of the componentsfrom each dynamical system is used tocouple networks’ nodes.3- The synchronization manifold is aninvariant manifold.4- The couplings are linear andnonlinear.5- A modification of the Watts-Strogatznetwork suggested by Monasson Will beconsidered.
→ Assumptions 1 and 3 guarantee theexistence of a unified synchronizationhyperplane.→ Assumption 2, allows us to make thestability diagram specific to the differentchoice of dynamical systems.→ Assumptions 4 and 5, help to choose alarge class of coupling structures and aspecified network model, whichthemselves include many real-worldapplications.
ImplicationsAssumptions
Nonlinear formalisms

1. Bifurcation and multistability in
the Extended Hindmarsh-Rose
Neuronal Oscillator
10/42
E. B. Megam Ngouonkadi et al. , Chaos Solitons and Fractals, (2016) 85
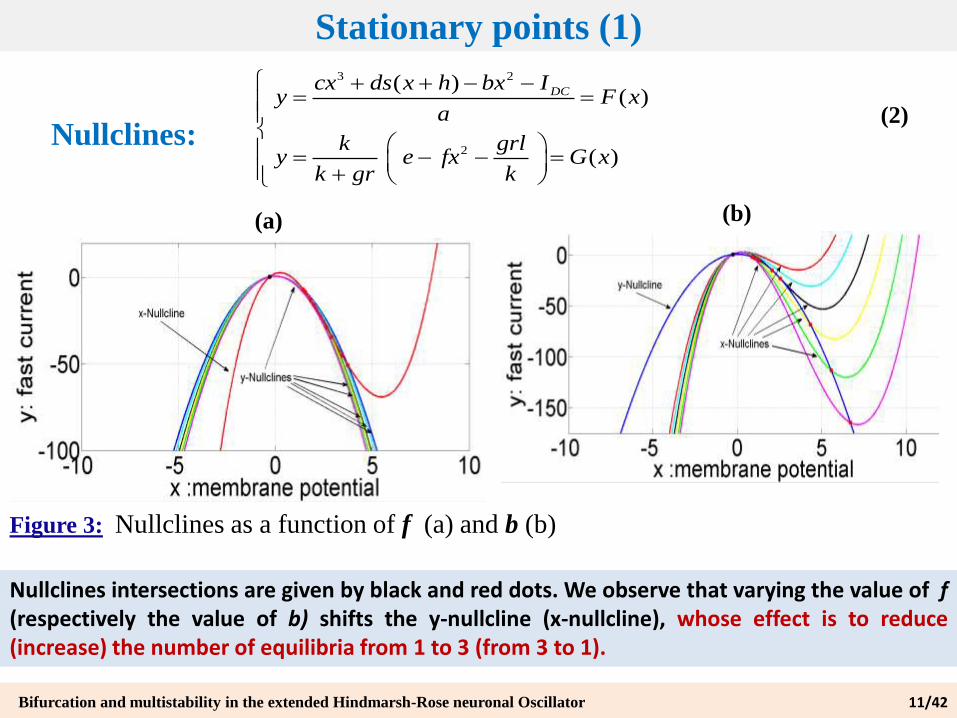
Stationary points (1)
Nullclines:
3 2
2
( )( )
( )
DCcx ds x h bx I
y F xa
k grly e fx G x
k gr k
Figure 3: Nullclines as a function of f (a) and b (b)
(a) (b)
(2)
Bifurcation and multistability in the extended Hindmarsh-Rose neuronal Oscillator 11/42
Nullclines intersections are given by black and red dots. We observe that varying the value of f(respectively the value of b) shifts the y-nullcline (x-nullcline), whose effect is to reduce(increase) the number of equilibria from 1 to 3 (from 3 to 1).
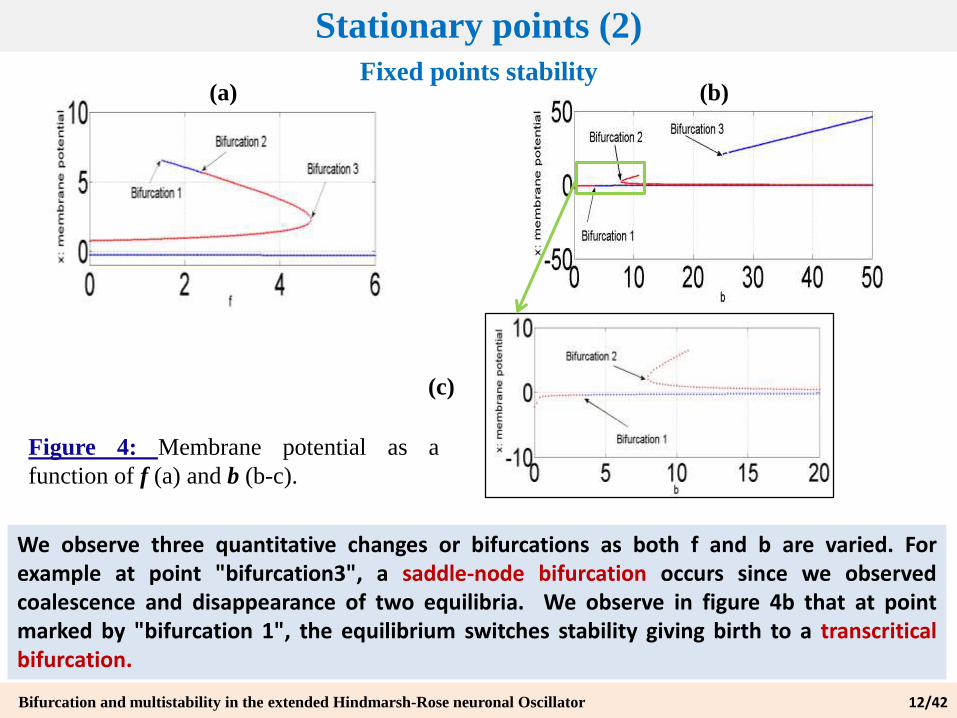
Fixed points stability
Figure 4: Membrane potential as a
function of f (a) and b (b-c).
(a) (b)
(c)
Bifurcation and multistability in the extended Hindmarsh-Rose neuronal Oscillator 12/42
Stationary points (2)
We observe three quantitative changes or bifurcations as both f and b are varied. Forexample at point "bifurcation3", a saddle-node bifurcation occurs since we observedcoalescence and disappearance of two equilibria. We observe in figure 4b that at pointmarked by "bifurcation 1", the equilibrium switches stability giving birth to a transcriticalbifurcation.

Hopf Bifurcation (1)
0
2 1 0
0 0
0 0
e
e
a d
fx gJ
s
vr kv
Characteristic equation : 4 3 2
1 2 3 40a a a a
with:
1
2
3
4
1
2 ( 1 )
2 2
( )
2 ( )
e
e e
e
a kv
a vrg kv kv sd fx a kv
a vrg kv sd kv kvafx sd fax
vrg kv kv
a vrg sd kv sd kv fax kv vrg
(3)
(4)
Bifurcation and multistability in the extended Hindmarsh-Rose neuronal Oscillator 13/42
Jacobian matrix at
equilibrium Se:
(5)
What happens when a pair of complex-conjugate characteristic exponents of an equilibriumstate crosses over the imaginary axis? The drain of stability is directly connected to thedisappearance or the birth of a periodic orbit. This bifurcation represents the most mechanismfor transition from a stationary regime to oscillations and can highlight proper interpretationof numerous physical phenomena.

Hopf Bifurcation (2)
1.0; 3.0; 1.0; 0.99; 1.01; 5.0128;
0.0278; 3.966; 1.605; 0.0009;
0.9573; 3.0; 1.619; 3.024972.
DC
a b c d e f
g s h v
k r l IThe unique equilibrium:
0.7553399395
1.8314834490.
3.3697518000
0.6658835764
e
e
e
e
e
x
yS
z
w
The critical value of the bifurcation parameter μ is:
0.1230628577 c
(6)
(7)
(8)
Parameter values , Hindmasrh et al. 1984; Selverston et al. 2000
Bifurcation and multistability in the extended Hindmarsh-Rose neuronal Oscillator 14/42
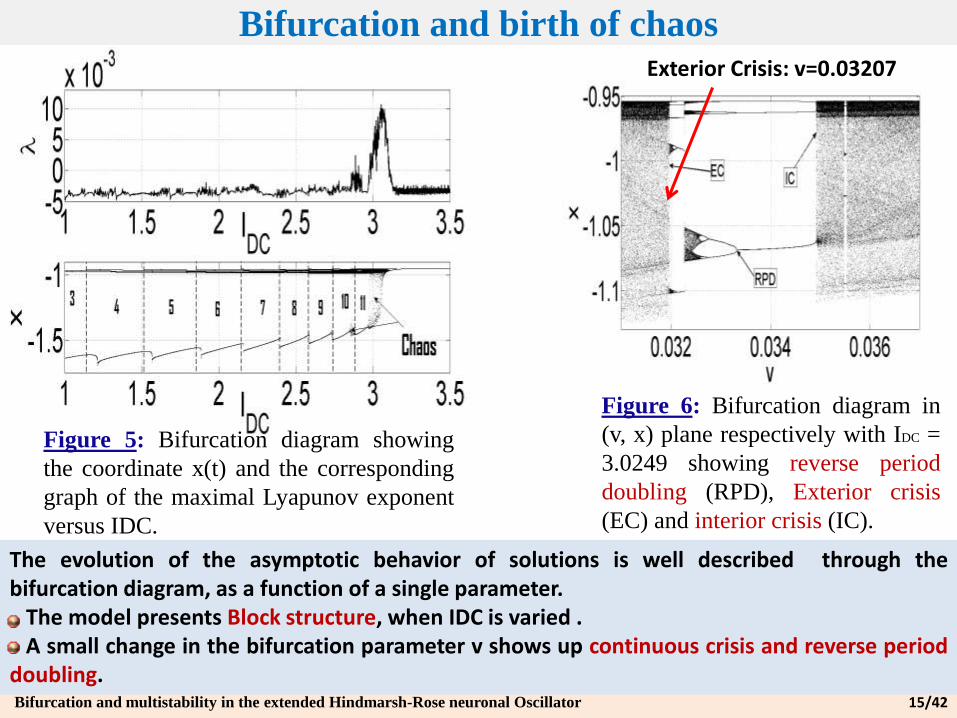
Bifurcation and birth of chaos
Figure 5: Bifurcation diagram showing
the coordinate x(t) and the corresponding
graph of the maximal Lyapunov exponent
versus IDC.
Figure 6: Bifurcation diagram in
(v, x) plane respectively with IDC =
3.0249 showing reverse period
doubling (RPD), Exterior crisis
(EC) and interior crisis (IC).
Bifurcation and multistability in the extended Hindmarsh-Rose neuronal Oscillator 15/42
Exterior Crisis: v=0.03207
The evolution of the asymptotic behavior of solutions is well described through thebifurcation diagram, as a function of a single parameter.
The model presents Block structure, when IDC is varied .A small change in the bifurcation parameter v shows up continuous crisis and reverse period
doubling.
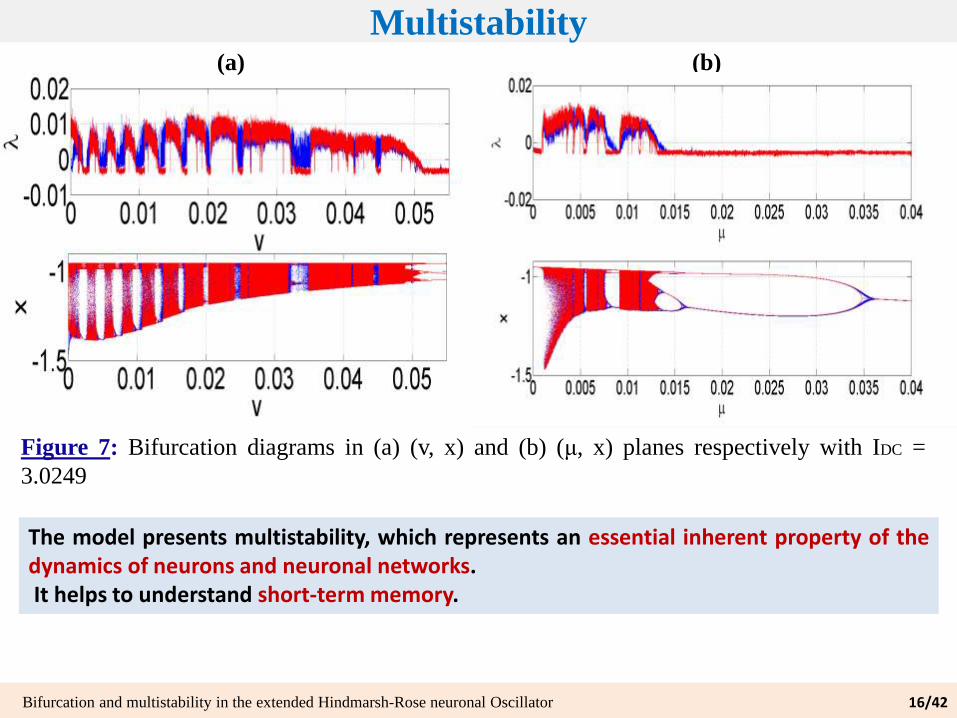
Multistability
Figure 7: Bifurcation diagrams in (a) (v, x) and (b) (μ, x) planes respectively with IDC =
3.0249
(a) (b)
Bifurcation and multistability in the extended Hindmarsh-Rose neuronal Oscillator 16/42
The model presents multistability, which represents an essential inherent property of thedynamics of neurons and neuronal networks.It helps to understand short-term memory.

2. Phase synchronization of
bursting neural networks
17/42
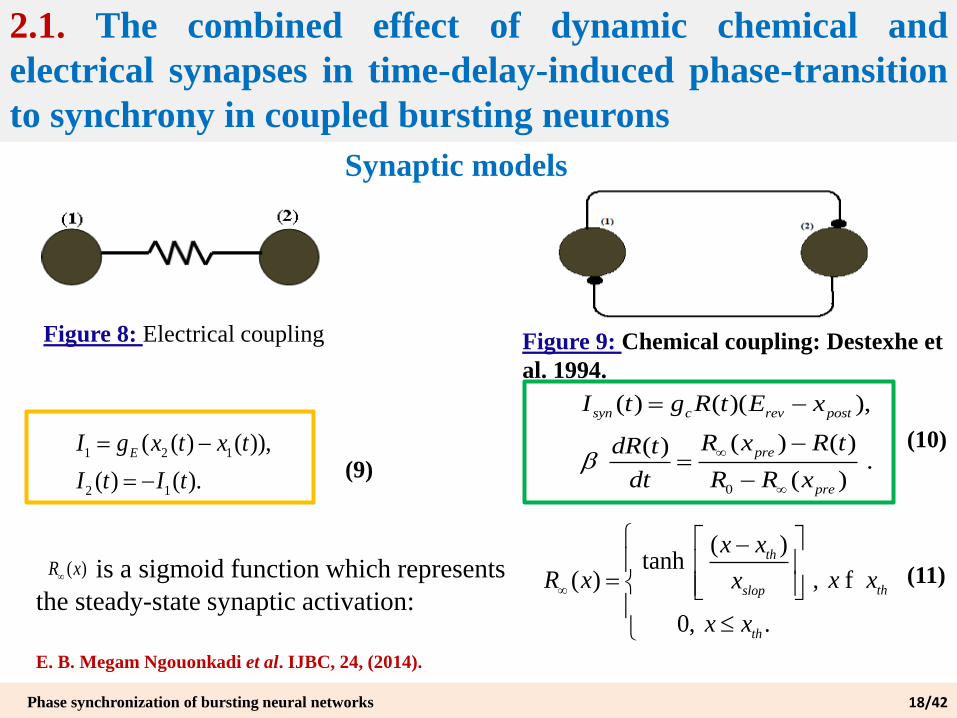
2.1. The combined effect of dynamic chemical and
electrical synapses in time-delay-induced phase-transition
to synchrony in coupled bursting neurons
Synaptic models
Figure 8: Electrical coupling
1 2 1
2 1
( ( ) ( )),
( ) ( ).
EI g x t x t
I t I t
Figure 9: Chemical coupling: Destexhe et
al. 1994.
0
( ) ( )( ),
( ) ( )( ).
( )
syn c rev post
pre
pre
I t g R t E x
R x R tdR t
dt R R x
is a sigmoid function which represents
the steady-state synaptic activation:
( )R x
(9)
(10)
(11)
Phase synchronization of bursting neural networks 18/42
( )tanh
( ) ,
0, .
f
th
thslop
th
x x
R x x xx
x x
E. B. Megam Ngouonkadi et al. IJBC, 24, (2014).

In the case of delayed electrical (linear coupling) and delayed chemical (nonlinear coupling)interactions, the whole system is described by the following equations:
1 2
1, 1,
0
( ) ( , , ) ( )( ).
( ) ( ) ( )where , 1, 2 and .
( )
&N N
i i i E ij i j c ij j rev i
j i j j i j
j j j
j
X F X g G H x x g C R t E x
dR t R x R ti j
dt R R x
1 1( , , ) ( ) ( ) .
i j j iH x x x t x t
From physiological experiments, 1=0 and 2 0
(12)
(13)
Phase synchronization of bursting neural networks 19/42
Network equations
Figure 10: Electrical and chemical coupling
Xi: represents an m-dimensional vector of dynamical variables of neuron i.Fi: is the velocity field.Gij: Electrical connection matrix.Cij: describes the way neurons are chemically coupled.H: represents the electrical coupling between nodes i and j.

Phase:
Phase difference:
1 ( )( ) tan
( )
i
i
i
x tt
x t
%
1 1 2 1 2
1 2
1 2 1 2
( ) ( ) ( ) ( )( ) ( ) tan
( ) ( ) ( ) ( )
x t x t x t x tt t
x t x t x t x t
% %
% %
(15)
(16)
(17)
Phase synchronization of bursting neural networks 20/42
Statistical quantities
The synchronization of slow bursts between two coupled neurons is analyzed based on themethod described by Pinto et al. (2000). It uses the normalized maximal deviation.
The normalized maximal deviation: max
max min
1 1/f f f
N dx x x
1 2( ) ( ) ( ) f f f
dx t x t x t
(14)
Phase-flip transitions to synchronization are analyzed using the instantaneous phase and phase difference of the time series.
Pinto et al. (2000) ; Pikovsky et al. (2003)
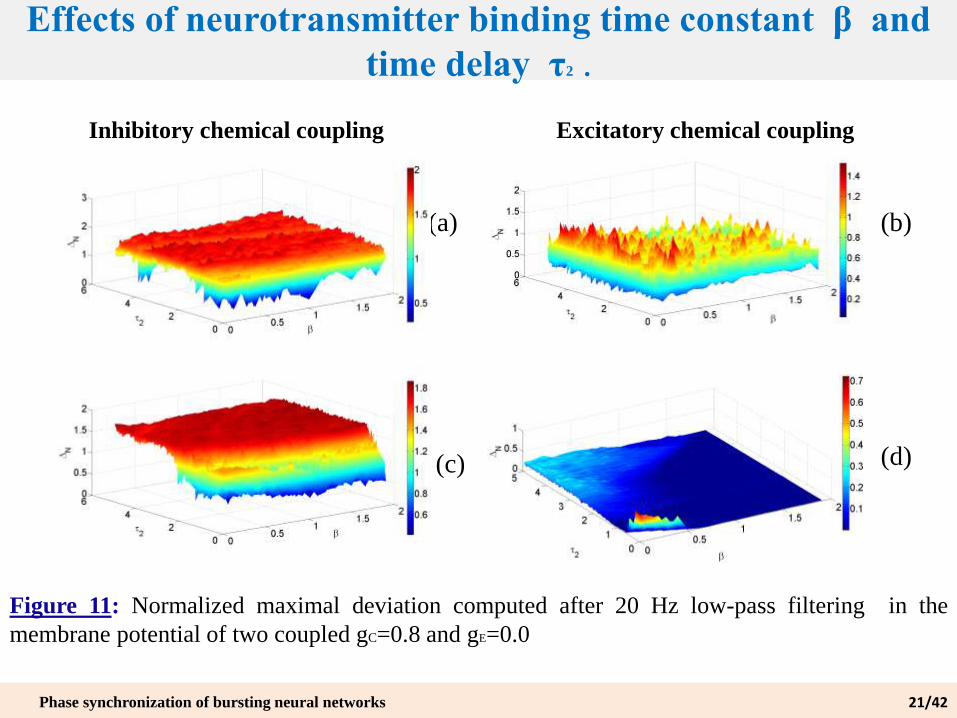
Effects of neurotransmitter binding time constant β and
time delay τ2 .
Figure 11: Normalized maximal deviation computed after 20 Hz low-pass filtering in the
membrane potential of two coupled gC=0.8 and gE=0.0
(a) (b)
(d)(c)
Inhibitory chemical coupling Excitatory chemical coupling
Phase synchronization of bursting neural networks 21/42
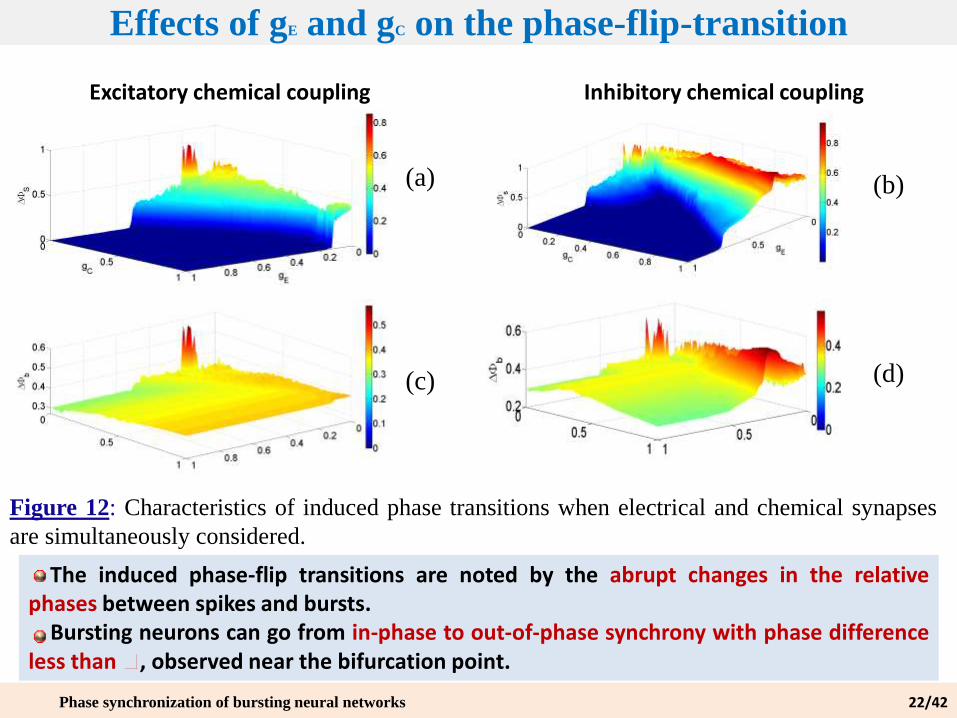
Effects of gE and gC on the phase-flip-transition
Figure 12: Characteristics of induced phase transitions when electrical and chemical synapses
are simultaneously considered.
(a) (b)
(c) (d)
Excitatory chemical coupling Inhibitory chemical coupling
Phase synchronization of bursting neural networks 22/42
The induced phase-flip transitions are noted by the abrupt changes in the relativephases between spikes and bursts.
Bursting neurons can go from in-phase to out-of-phase synchrony with phase differenceless than , observed near the bifurcation point.

int 1ij
c
%
0
( ) ( ) ( ).
( )
ij j ij
j
dR t R x R t
dt R R x
( )( ) ( )( )
syn ij C ij ij rev iI t g R t E x (19)
(18)
(20)
2.2. Phase synchronization of bursting neural networks
with electrical and delayed dynamic chemical couplings
Phase synchronization of bursting neural networks 23/42
E. B. Megam Ngouonkadi et al., European Physical Journal B, 88, (2015).
Many studies have confirmed that, large-scale brain has small-world property as
anatomical networks (Buzsaki G. 2006, Sporns O.et al. 2006).
We use a modification of the Watts-Strogatz network suggested by Monasson R.In this configuration no rewiring takes place but additional long range links are addedrandomly. This allows the network quantities mathematically easier accessible.
Delay is not still constant, but vary with some probabilistic law. It is spatially distributed and its value depends on the distance between neurons.
Int[x] represents the integer part of x.
: Gaussian white noise with zero mean and unitary standard deviation.: the distances fluctuations in realistic neural systems.c%

Small-world network topology
(a) (b)
Figure 13: Example of small-world network topology
N=8 N=16
Order parameter: measure of the spikes synchrony
( )
1 1
1 f
j
T Ni t
t jf
eNT
1
1
( ) 2 , ,j
j ji
j i ij j
i i
t Tt T t T
T T
j = 1,…,N and N is the number of nodes in the network.
ρ turns to unity for complete phase coherence and near zero for weak coherence among thephases of spike trains.
(24)
(23)
Phase synchronization of bursting neural networks 24/42
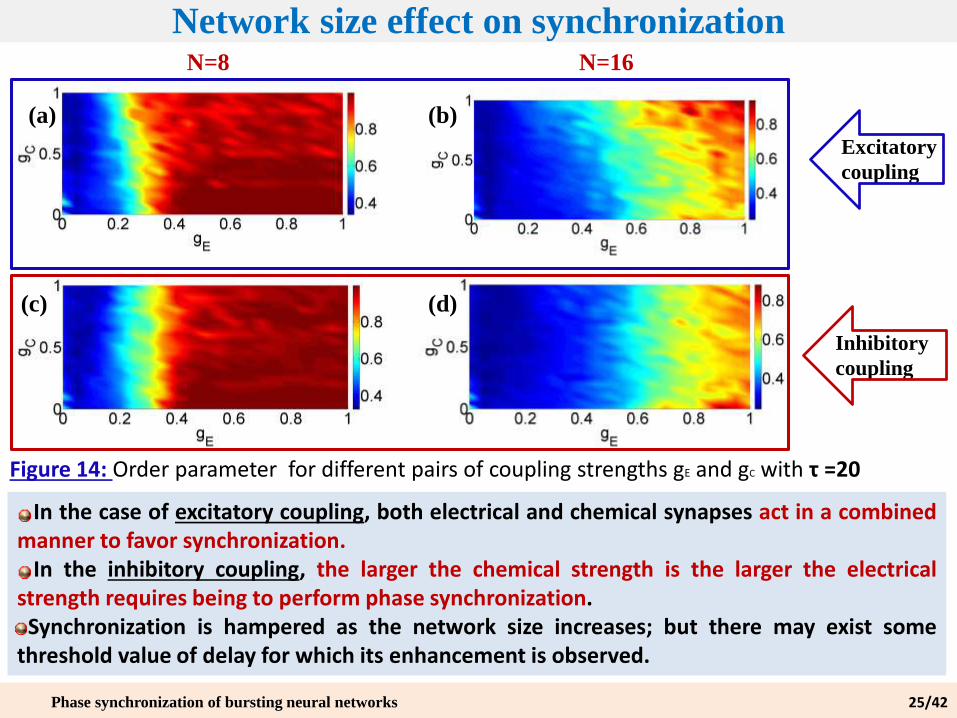
Network size effect on synchronization
Figure 14: Order parameter for different pairs of coupling strengths gE and gC with τ =20
N=8 N=16
Phase synchronization of bursting neural networks 25/42
(a)
(c)
(b)
(d)
Inhibitory
coupling
Excitatory
coupling
In the case of excitatory coupling, both electrical and chemical synapses act in a combinedmanner to favor synchronization.
In the inhibitory coupling, the larger the chemical strength is the larger the electricalstrength requires being to perform phase synchronization.
Synchronization is hampered as the network size increases; but there may exist somethreshold value of delay for which its enhancement is observed.
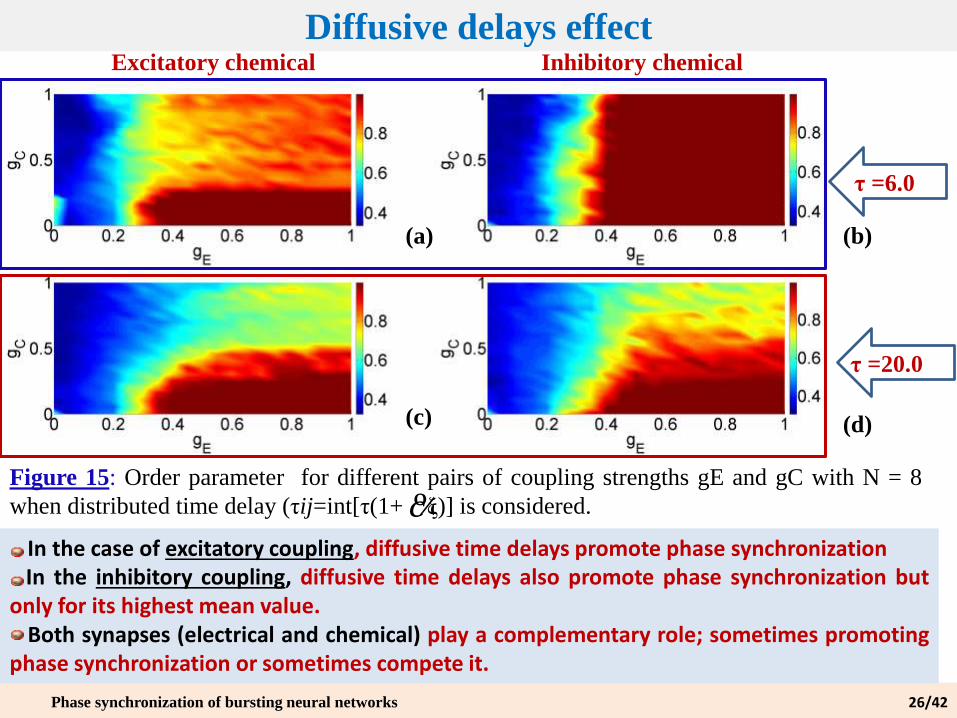
Diffusive delays effect
(a) (b)
(c) (d)
Excitatory chemical Inhibitory chemical
τ =6.0
τ =20.0
Phase synchronization of bursting neural networks 26/42
Figure 15: Order parameter for different pairs of coupling strengths gE and gC with N = 8
when distributed time delay (τij=int[τ(1+ ξ)] is considered.c%
In the case of excitatory coupling, diffusive time delays promote phase synchronizationIn the inhibitory coupling, diffusive time delays also promote phase synchronization but
only for its highest mean value.Both synapses (electrical and chemical) play a complementary role; sometimes promoting
phase synchronization or sometimes compete it.

3. Robust synchronization of
a small pacemaker neuronal
ensemble via nonlinear
controller: electronic circuit
design
27/42
E. B. Megam Ngouonkadi et al., Cognitive Neurodynamics, In revision
Robust synchronization of a small pacemaker neuronal ensemble via nonlinear controller: electronic circuit design 28/42
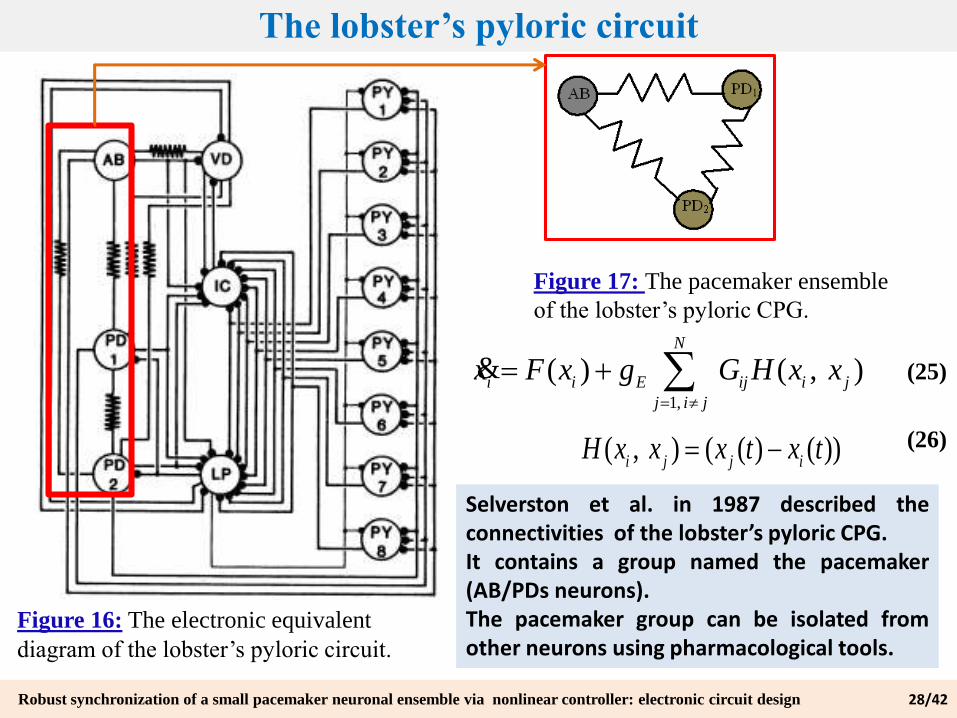
The lobster’s pyloric circuit
Figure 16: The electronic equivalent
diagram of the lobster’s pyloric circuit.
Figure 17: The pacemaker ensemble
of the lobster’s pyloric CPG.
1,
( ) ( , )N
i i E ij i j
j i j
x F x g G H x x
&
( , ) ( ( ) ( ))i j j i
H x x x t x t
(25)
(26)
Selverston et al. in 1987 described theconnectivities of the lobster’s pyloric CPG.It contains a group named the pacemaker(AB/PDs neurons).The pacemaker group can be isolated fromother neurons using pharmacological tools.
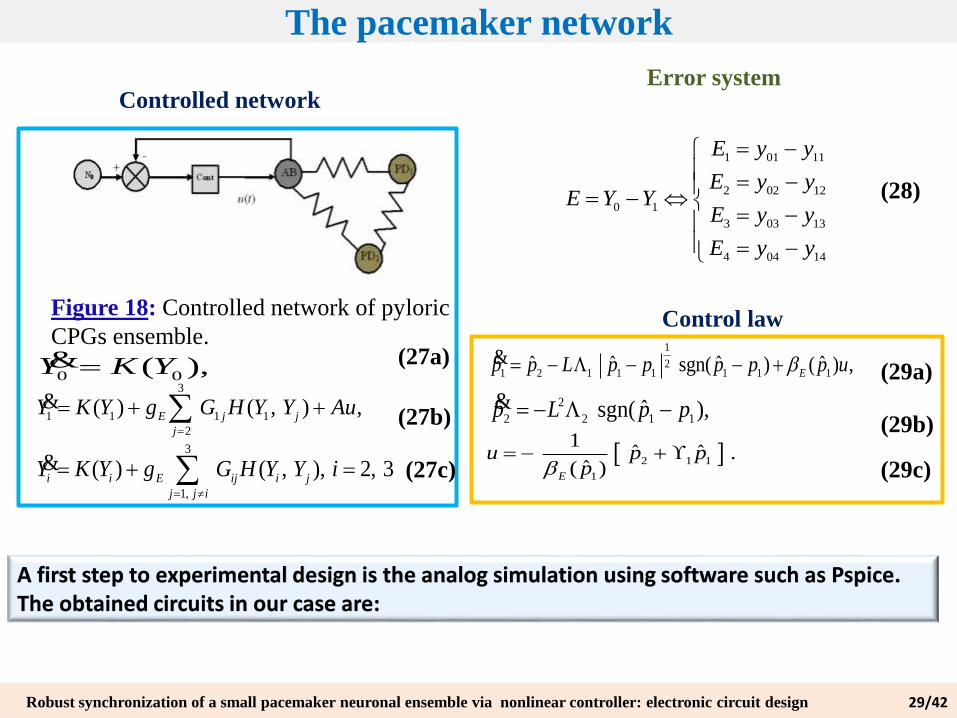
The pacemaker network
1 01 11
2 02 12
0 1
3 03 13
4 04 14
E y y
E y yE Y Y
E y y
E y y
Controlled network
Figure 18: Controlled network of pyloric
CPGs ensemble.
0 0( ),Y K Y&
3
1 1 1 1
2
( ) ( , ) ,E j j
j
Y K Y g G H Y Y Au
&
3
1,
( ) ( , ), 2, 3i i E ij i j
j j i
Y K Y g G H Y Y i
&
Control law1
21 2 1 1 1 1 1 1
ˆ ˆ ˆ ˆ ˆsgn( ) ( ) ,E
p p L p p p p p u &
2
2 2 1 1ˆ ˆsgn( ),p L p p &
2 1 1
1
1ˆ ˆ .
ˆ( )E
u p pp
Error system
(28)
(27a)(29a)
Robust synchronization of a small pacemaker neuronal ensemble via nonlinear controller: electronic circuit design 29/42
(27b)
(27c)
(29b)
(29c)
A first step to experimental design is the analog simulation using software such as Pspice. The obtained circuits in our case are:
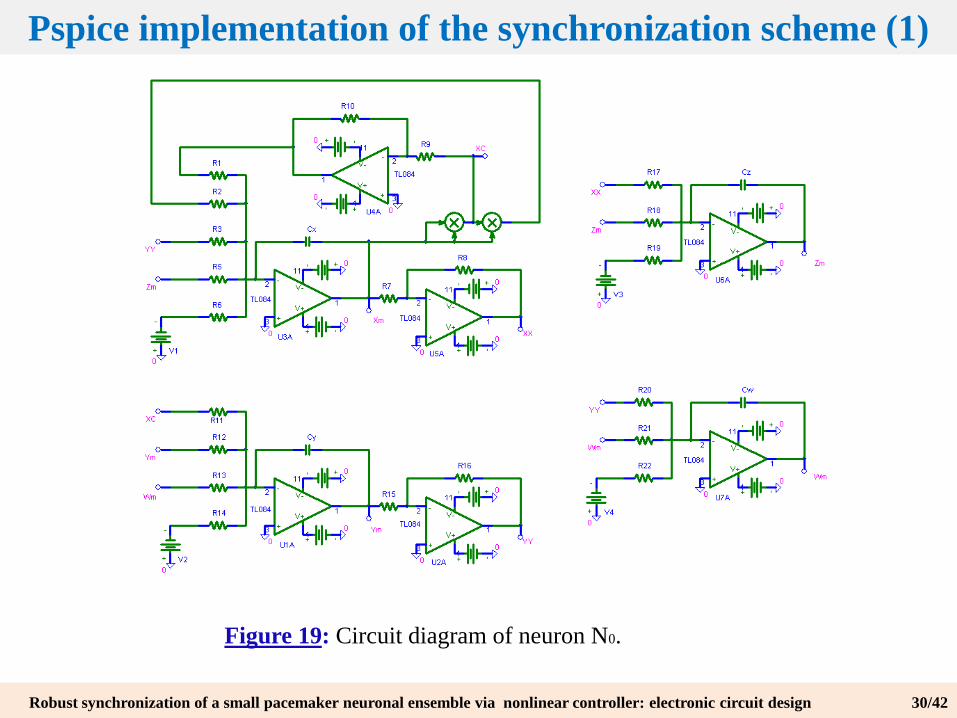
Figure 19: Circuit diagram of neuron N0.
Pspice implementation of the synchronization scheme (1)
Robust synchronization of a small pacemaker neuronal ensemble via nonlinear controller: electronic circuit design 30/42
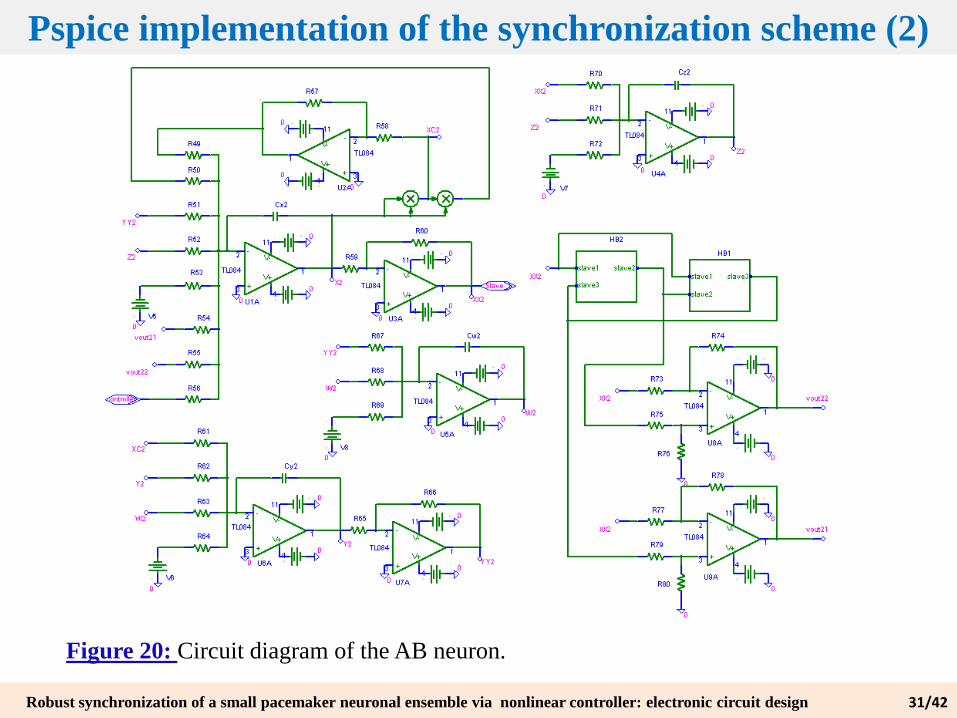
Figure 20: Circuit diagram of the AB neuron.
Pspice implementation of the synchronization scheme (2)
Robust synchronization of a small pacemaker neuronal ensemble via nonlinear controller: electronic circuit design 31/42
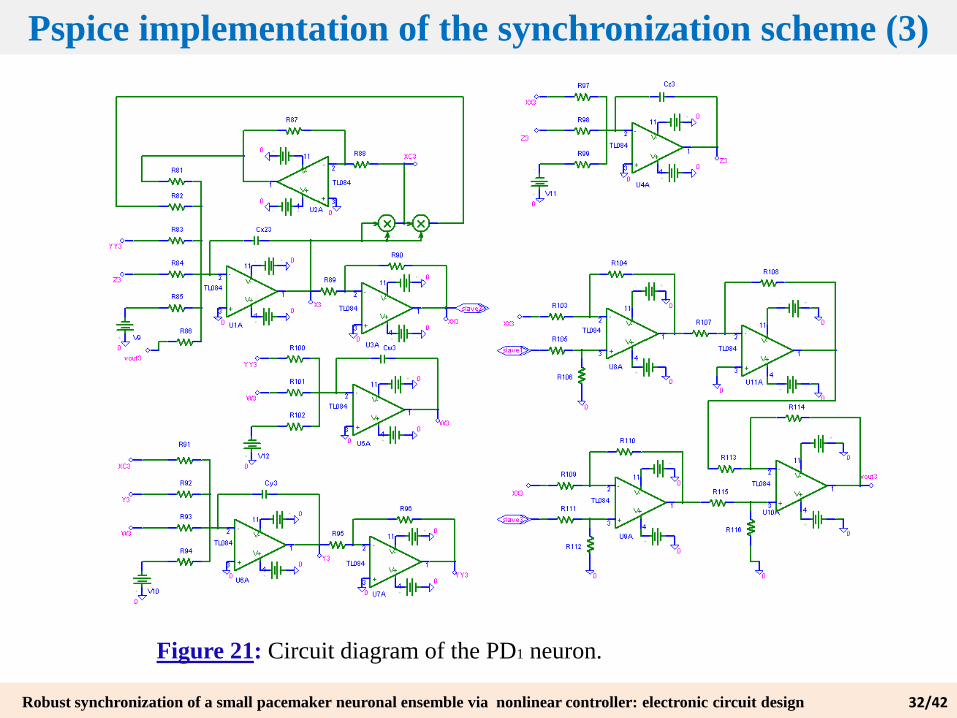
Figure 21: Circuit diagram of the PD1 neuron.
Pspice implementation of the synchronization scheme (3)
Robust synchronization of a small pacemaker neuronal ensemble via nonlinear controller: electronic circuit design 32/42
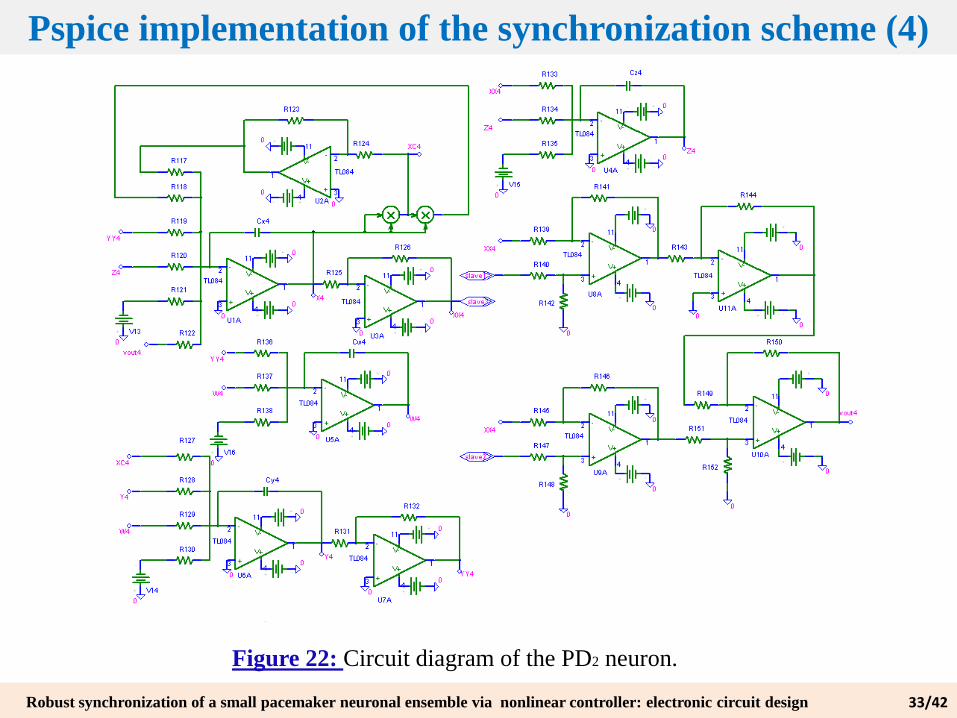
Figure 22: Circuit diagram of the PD2 neuron.
Pspice implementation of the synchronization scheme (4)
Robust synchronization of a small pacemaker neuronal ensemble via nonlinear controller: electronic circuit design 33/42
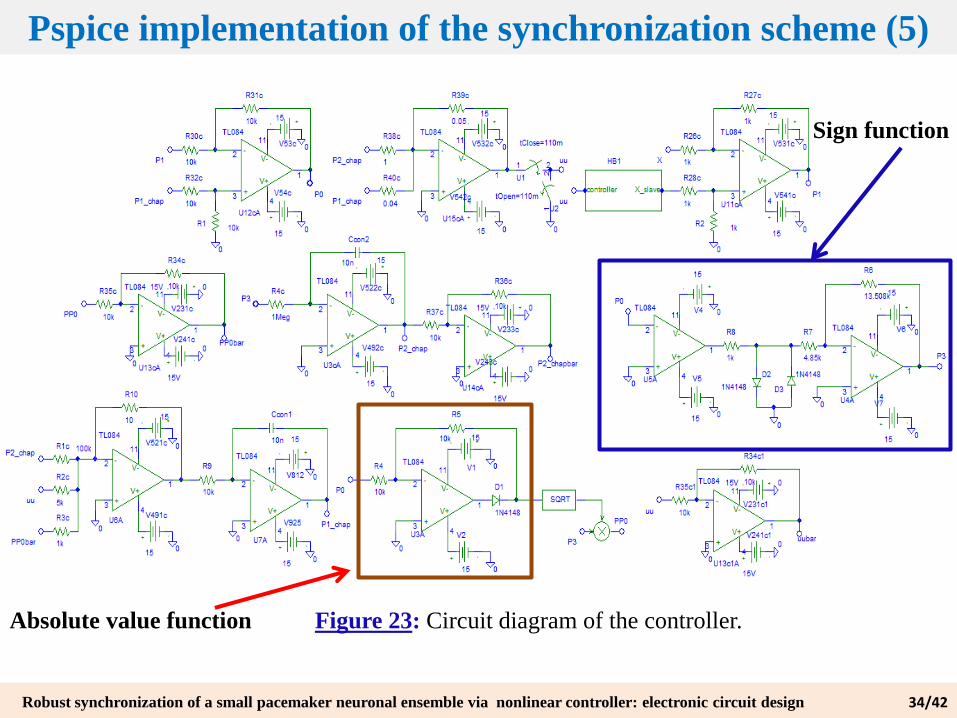
Figure 23: Circuit diagram of the controller.
Pspice implementation of the synchronization scheme (5)
Absolute value function
Sign function
Robust synchronization of a small pacemaker neuronal ensemble via nonlinear controller: electronic circuit design 34/42
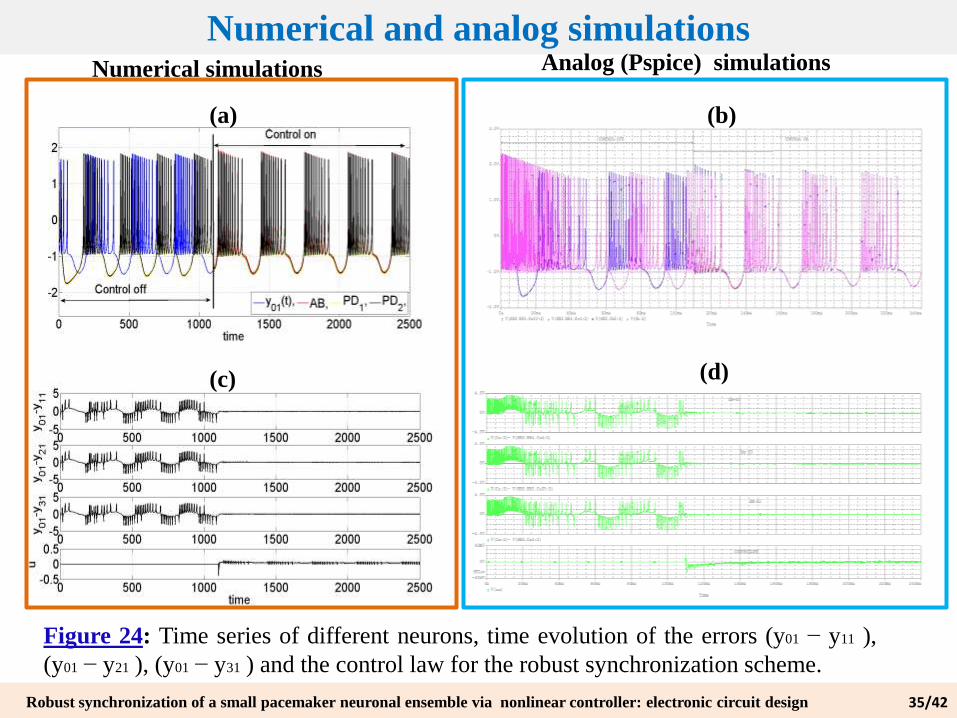
Figure 24: Time series of different neurons, time evolution of the errors (y01 − y11 ),
(y01 − y21 ), (y01 − y31 ) and the control law for the robust synchronization scheme.
(c) (d)
Numerical and analog simulations
(a) (b)
Numerical simulations Analog (Pspice) simulations
Robust synchronization of a small pacemaker neuronal ensemble via nonlinear controller: electronic circuit design 35/42

General conclusion and outlook
2. It is clear that multistability has important implications for information processing anddynamical memory in a neuron. It seems to be a major mechanism of operation in the area ofmotor control, particularly in the operation of multifunctional central pattern generators.These results furnish potentially useful information for enhancing our knowledge on the wayby which the neuronal system works and encodes information.
4. As the synchronization error depends on the control gain and the synchronization of thenetwork on the coupling strength, a compromise exists between both the coupling strengthand the control gain.
3. With the fact that, chemical synapses are responsible of the non-local nature of thesynapses, we have also shown that the distributed time-delays affect the phasesynchronization of the network cells.
1. Possible mechanisms to highlight how a nervous system give rapid response to stimulus aredescribed by the abrupt changes observed in the system’s dynamics named, the Hopfbifurcation and the interior crisis.
Main results
General conclusion and outlook 36/42

Outlook1. An interesting point to consider, is to carry the dynamics of extended HR excitable systemswith delayed coupling.
2. An extension of the master stability function towards more than one delay time is desirable,and when both, electrical and chemical synapses are taken into account.
3. A system of memristive coupled neurons can show information creation and recovery,expressed quantitatively by the information recovery inequality, in distinction to propertiesestablished for passive communication channels. Instead, these aspects of nonlinear activityshould provide an interesting framework for understanding the rich properties of memristorsynapses and realistic neuronal networks.
General conclusion and outlook 37/42
General conclusion and outlook

Personal references (1)Articles coming from the thesis
4. Implementing a Memristive Van Der Pol Oscillator Coupled to a Linear Oscillator:
Synchronization and Application to Secure Communication, E. B. Megam Ngouonkadi, H.
Fotsin, P. Louodop Fotso, Physica Scripta, 89, (2014).
3. The combined effect of dynamic chemical and electrical synapses in time-delay-induced
phase-transition to synchrony in coupled bursting neurons, E. B. Megam Ngouonkadi, H.
Fotsin, P. Louodop Fotso, International Journal of Bifurcation and Chaos, 24, (2014).
2. Phase synchronization of bursting neural networks with electrical and delayed dynamic
chemical couplings, E. B. Megam Ngouonkadi, M. Kabong Nono, V. Kamdoum Tamba and H.
B. Fotsin, European Physical Journal B, 88, (2015).
1. Bifurcation of Periodic Solutions and multistability in the Extended Hindmarsh-Rose
Neuronal Oscillator, E. B. Megam Ngouonkadi, H. B. Fotsin, P. Louodop Fotso, V. Kamdoum
Tamba and Hilda A. Cerdeira, Chaos Solitons and Fractals, (2016) 85.
5. Noise effects on robust synchronization of a small pacemaker neuronal ensemble, via
nonlinear controller: electronic circuit design, E. B. Megam Ngouonkadi, H. B. Fotsin, M.
Kabong Nono and Louodop Fotso Patrick Herve, Cognitive Neurodynamics, In revision
Personal references 38/42

Other articles
1. Emergence of complex dynamical behavior in Improved Colpitts oscillators:
antimonotonicity, chaotic Bubbles, coexisting attractors and transient chaos, V. Kamdoum
Tamba, H. B. Fotsin, J. Kengne, E. B. Megam Ngouonkadi, and P.K. Talla, International
Journal of Dynamics and Control, (2016) 1-12.
2. Finite-time synchronization of tunnel diode based chaotic oscillators, P. Louodop, H. Fotsin,
M. Kountchou, E. B. Megam Ngouonkadi, Hilda A. Cerdeira and S. Bowong, Physical
Review E, (2014) 89.
3. Effective Synchronization of a Class of Chua’s Chaotic systems Using an Exponential
Feedback Coupling, P. Louodop, H. Fotsin, E. B. Megam Ngouonkadi, S. Bowong and Hilda
A. Cerdeira, Journal of Abstract and Applied Analysis, (2013).
4. Dynamics, analysis and implementation of a new multiscroll memristor based chaotic circuit,
N. Henry Alombah, H. B. Fotsin, E. B. Megam Ngouonkadi, Tekou Nguazon, International
Journal of Bifurcation and Chaos, In revision.
5. Dynamics and indirect finitie-time stability of modified relay-coupled chaotic systems, P.
Louodop, E. B. Megam Ngouonkadi, H. Fotsin, S. Bowong and H. A. Cerdeira, Physical
Review E, In revision.
Personal references 39/42
Personal references (2)

Acknowledgements
I thank the University of Dschang for all facilities that they gave us,
the Abdus Salam International Center for Theoretical Physics (ICTP) whichpermit us to present some parts of this work during conferences.
I also thank Professor Hilda Cerdeira for collaboration and moral support,
which resulted in the publication of some works related to this thesis.
I thank all the jury's members who kindly accepted to review and evaluate
this work.
I thank my family for moral and financial supports.
I thank the LETS (Laboratory of Electronics and Signal Processing)
members for their collaborations.
I also thank the public for their kind attentions.
Acknowledgements 40/42

41/42