editorial - universidad autónoma del estado de...
TRANSCRIPT



EDITORIAL
En julio de 2010 se tomó la determinación de editar la revista CIENCIAS AGRICOLAS INFORMA con una
periodicidad semestral. El primer número, el 19: 3, presentó siete artículos. Posteriormente, en los números siguientes
se incluyeron cinco contribuciones científicas; es decir, actualmente se publican 10 artículos científicos por año. Se ha
conseguido mantener por tres años esta periodicidad; de 2011 a la fecha, de los 30 documentos publicados, 26 han sido
de autores nacionales y cuatro de autores extranjeros, dos de Cuba, particularmente de la Universidad de la Juventud
“Jesús Montané Oropesa” y dos de España, de las Universidades de Málaga y Politécnica de Catalunya. Durante este
lapso, la Universidad Autónoma del Estado de México ha firmado 30% de los documentos publicados, siguiéndolo en
orden descendente, el Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (inifap), 17%; Colegio de
Postgraduados y Benemérita Universidad Autónoma de Puebla, 10%; Universidad Autónoma del Estado de Morelos y
Universidad de la Cañada, 7%. Con un menor porcentaje de publicaciones están la Universidad Nacional Autónoma de
México y la Universidad Juárez Autónoma de Tabasco. La revista CIENCIAS AGRICOLAS INFORMA ha logrado
una mayor difusión, y su meta siguiente es ser incluida en bases de datos académicas. Nuevamente, agradecemos a los
autores su apoyo para lograr los objetivos planteados y los nuevos que deseamos alcanzar a la brevedad.
.
M. en fit. arteMio BalBuena Melgarejo
Director de la Facultad de Ciencias Agrícolas
Universidad Autónoma del Estado de México

UNIVERSIDAD AUTÓNOMA DEL ESTADO DE MÉXICO
Dr. en D. Jorge Olvera García Rector
Dr. en Ed. Alfredo Barrera Baca Secretario de Docencia
Dra. en Est. Lat. Ángeles Ma. del Rosario Pérez Bernal Secretaria de Investigación y Estudios Avanzados
M. en D. José Benjamín Bernal Suárez Secretario de Rectoría
M. en E. P. y D. Ivett Tinoco García Secretaria de Difusión Cultural
M. en C. I. Ricardo Joya Cepeda Secretario de Extensión y Vinculación
M. en E. Javier González Martínez Secretario de Administración
Dr. en C. Pol. Manuel Hernández Luna Secretario de Planeación y Desarrollo Institucional
M. en A. Ed. Yolanda E. Ballesteros Sentíes Secretaria de Cooperación Internacional
Dr. en D. Hiram Raúl Piña Libien Abogado General
L. en Com. Juan Portilla Estrada Director General de Comunicación Universitaria
Lic. Jorge Bernaldez García Secretario Técnico de la Rectoría
M. en A. Emilio Tovar Pérez Director General de Centros Universitarios y Unidades Académicas Profesionales
M. en A. Ignacio Gutiérrez Padilla Contralor Universitario
FACULTAD DE CIENCIAS AGRÍCOLAS
M. en Fit. Artemio Balbuena Melgarejo Director
M. A. Antonio Díaz Víquez Subdirector Académico
C. P. Román Apolinar Padilla Subdirector Administrativo
Dr. José Francisco Ramírez Dávila Coordinador del Centro de Investigación y Estudios Avanzados en Fitomejoramiento
Ing. Eduardo Enrique Lovera Coordinador de Difusión Cultural y Extensión
EQUIPO EDITORIAL
Correctores de redacción y estilo
Thomas H. Norman Mondragón María del Carmen Corona Rodríguez Carlos Gustavo Martínez Rueda
Correctora de idioma inglés
L.L.I. María Esther Calzado Fierros
Apoyo editorial y responsable de página web Eder Torres Miranda
Diseño y formato
Programa Editorial de la uaeM
Fotografía de portada
Cortesía Mtro. José Luis Hernández De la Cruz
COMITÉ EDITORIAL
Editor principal Omar Franco Mora
Comité Editorial Internacional
Jorge M. Fonseca Arizona University, United States of America
Daniel Valero Garrido Universidad Miguel Hernández, España
Edilberto Pozo Velázquez Universidad Central “Marta Abreu”, Cuba
Juan Miguel Pérez Universidad Nacional de Agricultura, Honduras
Comité Editorial Nacional
Jesús Jasso Mata Colegio de Postgraduados, México
Juan Guillermo Cruz Castillo Universidad Autónoma Chapingo, México
Héctor González Rosas Colegio de Postgraduados, México
Humberto Vaquera Huerta Colegio de Postgraduados, México
Marcos Pérez Sato Benemérita Universidad Autónoma de Puebla, México

CIENCIAS AGRICOLAS INFORMA
Año 22, No. 2, julio-diciembre 2013, es una publicación semestral editada por la Universidad Autónoma del Estado de México, a través de la Facultad
de Ciencias Agrícolas, Campus Universitario “El Cerrillo”, El Cerrillo Piedras Blancas, Toluca, México. Km 12.5, C.P. 50200, tel. y fax: (722) 296-55-18,
296-55-29 y 296-55-31 ext. 148, [email protected]. Editor responsable: Dr. Omar Franco Mora. Reserva de Derechos al Uso Exclusivo No. 04-
2006-102710130900-102, ISSN 1870-7378, ambos otorgados por el Instituto Nacional del Derecho de Autor, Licitud de Título y Contenido No. 15510,
otorgado por la Comisión Calificadora de Publicaciones y Revistas Ilustradas de la Secretaría de Gobernación. Impresa por CigoMe S. A. de C. V.
Este número se terminó de imprimir el 15 de diciembre de 2013 con un tiraje de 200 ejemplares.
Cada autor es responsable del contenido de su texto. Se autoriza la reproducción total o parcial, siempre y cuando se cite el crédito literario de la
fuente. Esta revista no responde por artículos no solicitados.
SumARIO
69
83
89
97
107
Línea de investigación Genética Vegetal y Fisiología
Evaluación del rendimiento de grano y de los componentes del rendimiento en trigo harineroEvaluation of grain yield and its components in soft wheat
Angélica Torres-Ramírez, Sanjaya Rajaram-Devi, Andrés González-Huerta, Artemio Balbuena-Melgarejo
Línea de investigación Recursos Naturales y Protección Ambiental
Aceite de ricino (Ricinus communis L.) con aplicaciones en comunicaciones ópticasCastor oil plant (Ricinus communis L.) with applications in optical communications
Ernesto Díaz-López, Israel Jesús Orlando-Guerrero, Jesús Manuel Campos-Pastelín, Irma Brena-Hernández, Juan Manuel Loeza-Corte
Línea de investigación Sanidad Vegetal
Tácticas para el control del tizón gomoso del tallo en el cultivo de sandía Integration of tactics for the control of the gummy stem blight in the cultivation of watermelon
Jesús Pérez-González, Benedicto Martínez-Coca, Salvador Guadarrama-Valentín, Sonia Estrada-Terra, Danay Infante-Martínez, Yanisia Duarte-Leal, Claudio Esquivel-Álvarez
Línea de investigación Biotecnología
Caracterización molecular de 20 clones de Hevea brasiliensis del jardín de propagación del inifap en TabascoMolecular characterizacion in 20 clones of Hevea brasiliensis in inifap propagation gardens at Tabasco state
José Luis Hernández-De la Cruz, Julia María Lesher-Gordillo, Armando Romo-López, José Miguel Hernández-Cruz
Caracterización molecular de cuatro variedades de Gerbera jamesonii Bolus, mediante microsatélites anclados y rapdsMolecular characterization of four varieties of Gerbera jamesonii Bolus, through anchored microsatellites and rapds
Amaury Martín Arzate-Fernández, José Luis Piña-Escutia, Luis Miguel Vázquez-García, Adolfo Carrillo-Velázquez


69
Evaluación del rendimiento de grano y de los componentes del rendimiento en trigo harinero
Evaluation of grain yield and its components in soft wheat
Angélica Torres-Ramírez,1* Sanjaya Rajaram-Devi,2 Andrés González-Huerta,1 Artemio Balbuena-Melgarejo1
CIENCIAS AGRICOLAS INFORMA 201322(2): 69-82
Recibido: 27 de febrero de 2013Aceptado: 2 de octubre de 2013
REsumEn
El presente trabajo se desarrolló en condiciones de temporal en el ciclo agrícola primavera-verano de 2009 en San Miguel Chapultepec, Estado de México. El objetivo principal fue analizar el rendimiento de grano y los componentes del rendimiento en 15 genotipos de trigo harinero (Triticum aestivum L.). Los genotipos de trigo de mayor producción de grano fueron los identificados como 1, 5 y 11 (casi 4 750 kg ha-1). Los tres genotipos más resistentes a roya amarilla fueron 5, 8 y 12 (3,3, 5,0 y 5,0%, respectivamente). Los genotipos más resistentes a acame fueron 6, 12 y 14 (0%). El aumento de rendimiento de grano en el material genético fue atribuido significativamente a un incremento en la altura de la planta, peso de 1 000 granos y peso hectolítrico del grano, pero hubo genotipos precoces con menor número de espiguillas por espiga y granos por espiga que tuvieron alto rendimiento en grano.
Palabras clave: Análisis de componentes principales, trigos sobresalientes, Triticum aestivum, Valles Altos, Valle de Toluca.
abstRact
Present research was developed under unirrigated conditions in the agricultural cycle spring-summer 2009 in San Miguel Chapultepec, State of Mexico. The main objective was to analyze the grain yield and grain yield components in 15 bread wheat (Triticum aestivum L. ) genotypes. The genotypes with greater grain yield production were those identified as 1, 5 and 11 (nearly 4 750 kg ha-1). The three genotypes more resistant to yellow rust were 5, 8 and 12 (3.3, 5.0 and 5.0%, respectively). The genotypes more resistant to lodging were 6, 12 and 14 (0.0%). The increase in grain yield in the genetic material was significantly attributed to an increase in plant height, 1 000 grain weight and hectolitric weight of grain, but there was early genotypes with a smaller number of spikelets per spike and grain per spike that had high grain yield.
Key words: High Valleys, outstanding wheat genotypes, principal components analysis, Toluca Valley, Triticum aestivum.
1 Facultad de Ciencias Agrícolas, Universidad Autónoma del Estado de México, México. 2 Centro Internacional de Investigaciones Agrícolas en Zonas Áridas, Siria.
*Autora para correspondencia: [email protected]
70
Torres et al., 2013. Rendimiento en trigo
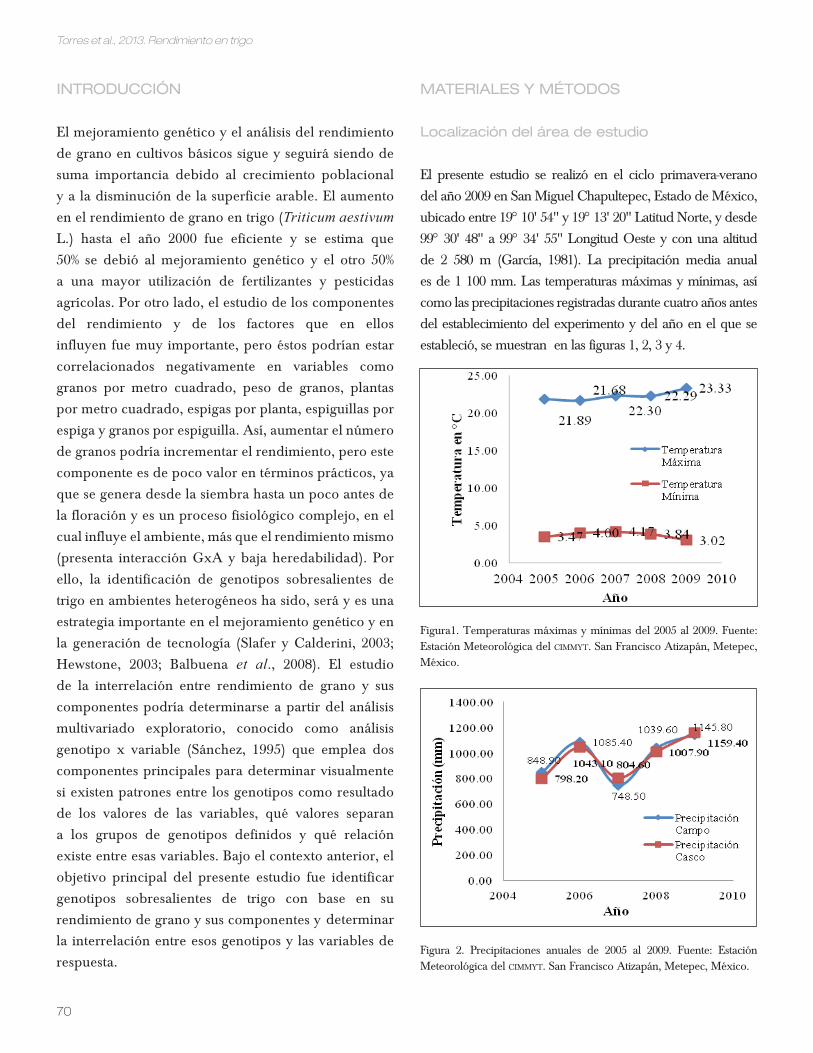
Figura1. Temperaturas máximas y mínimas del 2005 al 2009. Fuente: Estación Meteorológica del CiMMyt. San Francisco Atizapán, Metepec, México.
Figura 2. Precipitaciones anuales de 2005 al 2009. Fuente: Estación Meteorológica del CiMMyt. San Francisco Atizapán, Metepec, México.
IntRoduccIón
El mejoramiento genético y el análisis del rendimiento
de grano en cultivos básicos sigue y seguirá siendo de
suma importancia debido al crecimiento poblacional
y a la disminución de la superficie arable. El aumento
en el rendimiento de grano en trigo (Triticum aestivum
L.) hasta el año 2000 fue eficiente y se estima que
50% se debió al mejoramiento genético y el otro 50%
a una mayor utilización de fertilizantes y pesticidas
agrícolas. Por otro lado, el estudio de los componentes
del rendimiento y de los factores que en ellos
influyen fue muy importante, pero éstos podrían estar
correlacionados negativamente en variables como
granos por metro cuadrado, peso de granos, plantas
por metro cuadrado, espigas por planta, espiguillas por
espiga y granos por espiguilla. Así, aumentar el número
de granos podría incrementar el rendimiento, pero este
componente es de poco valor en términos prácticos, ya
que se genera desde la siembra hasta un poco antes de
la floración y es un proceso fisiológico complejo, en el
cual influye el ambiente, más que el rendimiento mismo
(presenta interacción GxA y baja heredabilidad). Por
ello, la identificación de genotipos sobresalientes de
trigo en ambientes heterogéneos ha sido, será y es una
estrategia importante en el mejoramiento genético y en
la generación de tecnología (Slafer y Calderini, 2003;
Hewstone, 2003; Balbuena et al., 2008). El estudio
de la interrelación entre rendimiento de grano y sus
componentes podría determinarse a partir del análisis
multivariado exploratorio, conocido como análisis
genotipo x variable (Sánchez, 1995) que emplea dos
componentes principales para determinar visualmente
si existen patrones entre los genotipos como resultado
de los valores de las variables, qué valores separan
a los grupos de genotipos definidos y qué relación
existe entre esas variables. Bajo el contexto anterior, el
objetivo principal del presente estudio fue identificar
genotipos sobresalientes de trigo con base en su
rendimiento de grano y sus componentes y determinar
la interrelación entre esos genotipos y las variables de
respuesta.
matERIalEs y métodos
localización del área de estudio
El presente estudio se realizó en el ciclo primavera-verano
del año 2009 en San Miguel Chapultepec, Estado de México,
ubicado entre 19° 10‘ 54‘‘ y 19° 13‘ 20‘‘ Latitud Norte, y desde
99° 30‘ 48‘‘ a 99° 34‘ 55‘‘ Longitud Oeste y con una altitud
de 2 580 m (García, 1981). La precipitación media anual
es de 1 100 mm. Las temperaturas máximas y mínimas, así
como las precipitaciones registradas durante cuatro años antes
del establecimiento del experimento y del año en el que se
estableció, se muestran en las figuras 1, 2, 3 y 4.

71
CienCias agRiColas infoRma 22(2): 69-82. Julio-Diciembre 2013
Figura 3. Temperaturas mensuales máximas y mínimas del año 2009. Fuente: Estación Meteorológica del CiMMyt. San Francisco Atizapán, Metepec, México.
Figura 4. Precipitación mensual en milímetros del año 2009. Fuente: Estación Meteorológica del CiMMyt. San Francisco Atizapán, Metepec, México.
material genético
Se utilizaron 14 genotipos de trigo harinero provenientes
del Programa de Mejoramiento Genético de la Empresa
Resource Seed Mexicana, S. A. de R. L. de C. V. y un
genotipo comercial empleado como testigo (cv. Tollocan)
(Cuadro 1).
Cuadro 1. Genotipos de trigo harinero evaluados en San Miguel Chapultepec, México, durante el ciclo primavera-verano, 2009.
No. Origen Cruza Pedigree
1 México TOLLOCAN F2005 CV. Tollocan F2005
2 Argentina PSN/BOW//SERI/3/MILAN/4/ATTILA ARG-0T
3 India SW90.1057/3/KAUZ*2/YACO//KAUZ/4/S91.12331
INDIA-0CJ
4 India BL2064//SW89.5124*2/FASAN/3/TILHI INDIA-0CJ
5 México ALD/CEP75630//CEP75234/PT7219/3/BUC/BJY/4/CBRD/5/TNMU/PF85487
CMSS93Y0363IT-6Y-3AL-010M-10Y-0M-0LPY-2SJ-0Y¨05T-05CJ-0T
6 U.S.A. 04W40483 USA-1T-02CJ-0T
7 U.S.A. 04W40677PPO USA-5T-02CJ-0T
8 U.S.A. 05W90706 USA-4T-02CJ-0T
9 U.S.A. 06W30565 USA-1T-02CJ-0T
10 U.S.A. 06W30565 USA-3T-02CJ-0T
11 U.S.A. 06W30565 USA-4T-02CJ-0T
12 U.S.A. 06W30596 USA-3T-02CJ-0T
13 U.S.A. 05W90911 USA-08CJ-0T
14 U.S.A. 07W50636 USA-08CJ-0T
15 U.S.A. 07W50744 USA-08CJ-0T

72
Torres et al., 2013. Rendimiento en trigo
diseño experimental y tamaño de la parcela
El material genético fue evaluado en campo bajo un
diseño experimental de bloques completos al azar con
tres repeticiones. La unidad experimental consistió de dos
surcos a doble hilera, de 3 m de longitud y 0,18 m entre
hileras; la separación entre parcelas fue de 0,76 m. En
el experimento se sembraron bordos de protección. La
parcela útil fueron los dos surcos a doble hilera (área de
4,56 m2).
desarrollo del trabajo experimental
La fertilización se realizó en dos etapas: en la primera
se aplicaron 80-75-60 de NPK utilizando como fuentes
urea (N, 46%), superfosfato de calcio triple (P2O, 46%) y
cloruro de potasio (K2O, 60%), los cuales se aplicaron en
el último rastreo, sobre los residuos de rastrojo del ciclo
pasado para acelerar el proceso de descomposición de la
materia orgánica e inmediatamente se preparó el suelo.
La segunda fertilización se efectuó antes de la segunda
escarda, adicionando 80 unidades de nitrógeno/ha (urea)
y 60 unidades de potasio/ha (cloruro de potasio). Estos
tratamientos se definieron con base en el análisis de
suelo realizado antes de la siembra.
La preparación del suelo consistió de tres pasos
de rastra efectuados 60, 30 y 10 días previos al
establecimiento del experimento. La siembra se realizó
mecánicamente en donde se depositaron 68 g de semilla
de cada genotipo. Las labores culturales del experimento
consistieron en dos escardas: cuando la planta tuvo cinco
hojas y durante la etapa de amacollamiento; ambas
labores dependieron de la disponibilidad de humedad
en el suelo y cuyo propósito fue aporcar las plantas y
controlar parcialmente la maleza.
La maleza de hoja ancha se controló con Tifensulfuron
(20 g ha-1 en 200 L de agua) y Metsulfuron Metil (6 g
ha-1 en 200 L de agua) cuando las plántulas tenían 3 cm
de altura. La maleza de hoja angosta se controló con
Clodinafop-propargil; la aplicación se hizo ocho días
antes del amacollamiento. Por último, se asperjaron
20 ml de Sencor y 100 ml de Paraquat por cada 15
L de agua en la etapa de espigamiento; se utilizó
una boquilla 8004 y una campana para evitar daños
al trigo.
La cosecha se realizó manualmente cuando el material
genético alcanzó la madurez fisiológica, y consistió en
recoger todas las espigas que se produjeron en las parcelas
útiles, las cuales se trillaron mecánicamente. Los granos
fueron colocados en bolsas que permitieron su aireación
y secado a un contenido de humedad de 14%.
Variables de respuesta
El acame de tallo se determinó cuando las plantas
alcanzaron la madurez fisiológica, y presentaron
inclinación de 30° o más. Los días a espigamiento fueron
los días transcurridos desde la siembra hasta que más de
60% de las plantas espigaron. Los días a floración fueron
los días transcurridos desde la siembra hasta que más de
60% de las plantas presentaron antesis. Los días a madurez
fisiológica fueron los días transcurridos desde la siembra
hasta que más de 60% de la población tuvo un color verde
alimonado. Se determinó el área de la hoja bandera en
centímetros cuadrados; registrando en antesis con la
fórmula: longitud de la hoja x ancho de la hoja x 0,75.
El área de la segunda hoja (en centímetros cuadrados), es
decir, la penúltima hoja de la planta se registró en antesis
con la misma fórmula. La longitud del primero y segundo
entrenudo (expresado en centímetros), se registró al medir
el largo del primero y segundo entrenudo de la planta en
madurez fisiológica.
Se determinó la incidencia de roya amarilla (Puccinia
striformis) en madurez fisiológica y se expresó en
porcentaje (Zadoks et al. 1974). La altura de planta se
midió con una regla y se registró en centímetros, desde
el nivel del suelo hasta la punta de la espiga. El número
de tallos fueron los existentes en un metro lineal de la
parcela útil; mientras que el número de espigas fueron las
verdaderas localizadas en un metro lineal de la parcela
útil. Para obtener el rendimiento de grano se pesó el
grano de cada una de las parcelas experimentales útiles

73
CienCias agRiColas infoRma 22(2): 69-82. Julio-Diciembre 2013
y se extrapoló a kilogramos por hectárea. Para el peso
hectolítrico del grano se empleó una báscula digital y
se pesó el grano limpio proveniente de cada parcela
experimental útil y se obtuvo su peso en relación con
el volumen de un litro. De cada parcela se pesaron 100
granos y su peso se extrapoló a 1 000 granos; mientras
que para obtener los granos por espiga, se empleó una
muestra aleatoria de 10 espigas de cada parcela útil
desgranadas individualmente y se contabilizó el número
de granos. Para la longitud de la espiga se muestrearon
10 espigas por cada parcela y se midió su longitud en
centímetros. El peso de grano por espiga se registró en
gramos en 10 espigas tomadas al azar en cada parcela.
Finalmente, el número de espiguillas por espiga se
determinó de una muestra de 10 espigas provenientes de
cada parcela, se contabilizaron las espiguillas por espiga
y se registró su media.
análisis estadístico
Los datos se analizaron con el Sistema para Análisis
Estadístico (Statistical Analysis System (sas) versión
para Windows). Las pruebas de f y de comparación de
medias entre genotipos de trigo se hicieron a un nivel de
significancia del 0,01 (Tukey); de igual manera se realizó
un análisis de correlación lineal simple y de componentes
principales (Martínez, 1988; Sánchez, 1995).
REsultados y dIscusIón
análisis de varianza para las variables
fisiológicas y morfológicas
En el Cuadro 2 se puede observar que los efectos
entre genotipos fueron significativos para la mayoría
de las variables registradas, excepto para roya amarilla
(ra) y acame (pa). Las diferencias entre repeticiones,
asociadas a la heterogeneidad del suelo donde se
estableció el ensayo, sólo fueron significativas en ra,
altura de planta (ap) y área de la segunda hoja (ash).
Los coeficientes de variación fueron aceptables para
siete variables (valores de 1,1 a 10,6%), pero no para
roya y acame; estos resultados podrían explicarse por
el hecho de que estas dos últimas no se distribuyen
uniformemente en todas las parcelas que conforman el
experimento y específicamente, en las que recibieron
el mismo tratamiento.
comparación de medias para las variables
fisiológicas y morfológicas
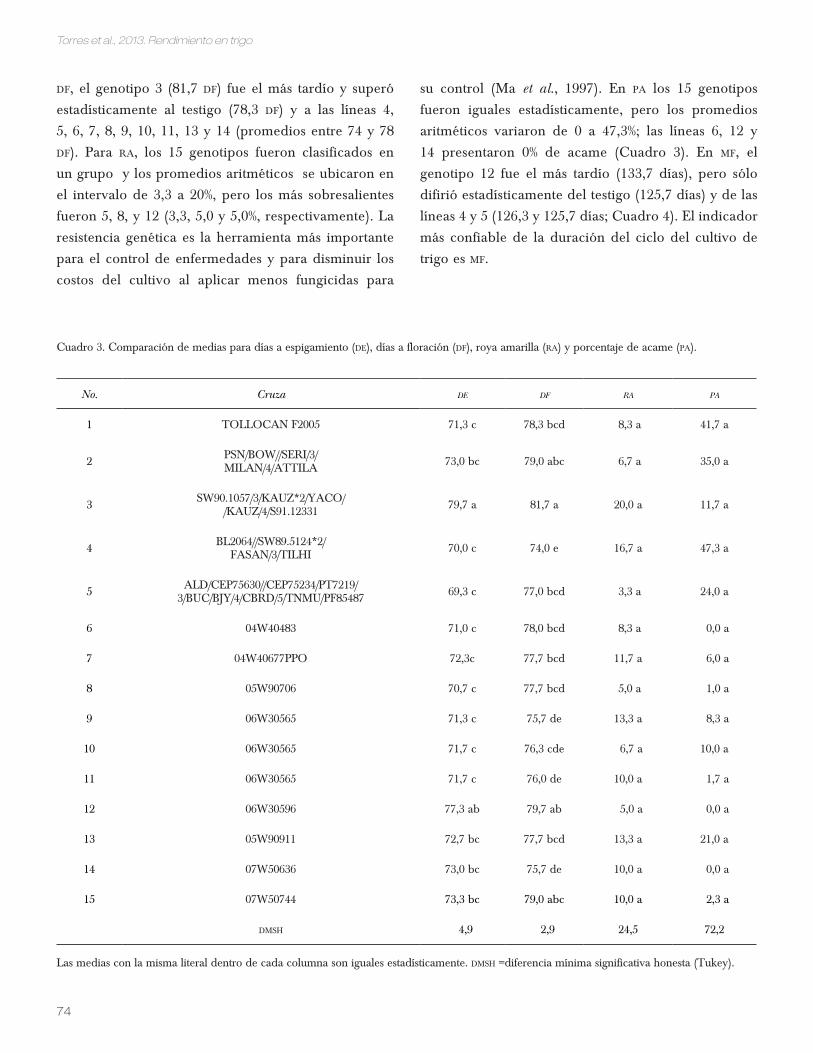
En de, los genotipos 3 y 12 (79,7 y 77,3 días) superaron
estadísticamente al testigo (71,3 días), pero las líneas
identificadas como 2, 4, 5, 7, 8, 9, 10, 13, 14 y 15 fueron
tan precoces o más que éste (entre 69,3 y 73,3 de). Para
Cuadro 2. Análisis de varianza, media general y coeficiente de variación para días a espigamiento (de), días a floración (df), roya amarilla (ra), porcentaje de acame (pa), madurez fisiológica (Mf), altura de la planta (ap), área de la hoja bandera (ahB), área de la segunda hoja (ash) y longitud de entrenudos (len).
F.V. G.L. de df ra pa mf ap ahb ash len
Genotipos 11,7** 15,4** 1,3 ns 2,0 NS 4,2** 12,0** 4,3* 5,4** 20,8**
Repeticiones 2 0,3 NS 0,2 NS 3,6* 1,4 NS 0,21 NS 9,41** 2,64 NS 10,8** 3.55 NS
Media 72,6 77,6 9,9 13,3 130,1 79,2 21,0 21,9 51,3
C.V. 1,9 1,1 69,3 151,3 1,6 3,5 10,6 8,2 3,8
* Significativo al 0,05; ** altamente significativo al 0,01; ns no significativo.
74
Torres et al., 2013. Rendimiento en trigo
df, el genotipo 3 (81,7 df) fue el más tardío y superó
estadísticamente al testigo (78,3 df) y a las líneas 4,
5, 6, 7, 8, 9, 10, 11, 13 y 14 (promedios entre 74 y 78
df). Para ra, los 15 genotipos fueron clasificados en
un grupo y los promedios aritméticos se ubicaron en
el intervalo de 3,3 a 20%, pero los más sobresalientes
fueron 5, 8, y 12 (3,3, 5,0 y 5,0%, respectivamente). La
resistencia genética es la herramienta más importante
para el control de enfermedades y para disminuir los
costos del cultivo al aplicar menos fungicidas para
su control (Ma et al., 1997). En pa los 15 genotipos
fueron iguales estadísticamente, pero los promedios
aritméticos variaron de 0 a 47,3%; las líneas 6, 12 y
14 presentaron 0% de acame (Cuadro 3). En Mf, el
genotipo 12 fue el más tardío (133,7 días), pero sólo
difirió estadísticamente del testigo (125,7 días) y de las
líneas 4 y 5 (126,3 y 125,7 días; Cuadro 4). El indicador
más confiable de la duración del ciclo del cultivo de
trigo es Mf.
Cuadro 3. Comparación de medias para días a espigamiento (de), días a floración (df), roya amarilla (ra) y porcentaje de acame (pa).
No. Cruza de df ra pa
1 TOLLOCAN F2005 71,3 c 78,3 bcd 8,3 a 41,7 a
2 PSN/BOW//SERI/3/MILAN/4/ATTILA 73,0 bc 79,0 abc 6,7 a 35,0 a
3 SW90.1057/3/KAUZ*2/YACO//KAUZ/4/S91.12331 79,7 a 81,7 a 20,0 a 11,7 a
4 BL2064//SW89.5124*2/FASAN/3/TILHI 70,0 c 74,0 e 16,7 a 47,3 a
5 ALD/CEP75630//CEP75234/PT7219/3/BUC/BJY/4/CBRD/5/TNMU/PF85487 69,3 c 77,0 bcd 3,3 a 24,0 a
6 04W40483 71,0 c 78,0 bcd 8,3 a 0,0 a
7 04W40677PPO 72,3c 77,7 bcd 11,7 a 6,0 a
8 05W90706 70,7 c 77,7 bcd 5,0 a 1,0 a
9 06W30565 71,3 c 75,7 de 13,3 a 8,3 a
10 06W30565 71,7 c 76,3 cde 6,7 a 10,0 a
11 06W30565 71,7 c 76,0 de 10,0 a 1,7 a
12 06W30596 77,3 ab 79,7 ab 5,0 a 0,0 a
13 05W90911 72,7 bc 77,7 bcd 13,3 a 21,0 a
14 07W50636 73,0 bc 75,7 de 10,0 a 0,0 a
15 07W50744 73,3 bc 79,0 abc 10,0 a 2,3 a
dMsh 4,9 2,9 24,5 72,2
Las medias con la misma literal dentro de cada columna son iguales estadísticamente. dMsh =diferencia mínima significativa honesta (Tukey).

75
CienCias agRiColas infoRma 22(2): 69-82. Julio-Diciembre 2013
La mayor ap se registró en la variedad testigo y su promedio
aritmético (88,1 cm) fue diferente estadísticamente
del registrado en las líneas 6, 9, 10, 11 y 14 (medias
de 72,7, 74,2, 74,8, 76,0 y 69,4 cm, respectivamente;
Cuadro 4). El testigo también presentó un pa del 41,7%
pero su producción de grano fue de 4 740 kg ha-1. El
otro genotipo superior, el identificado como 11, tuvo
menos altura de planta (76,0 cm) y menor pa (1,7%) y su
producción de grano (rg) fue igual a la del testigo. Para
el ahB, los genotipos más sobresalientes fueron 7, 5, 12
y 13 (25,1, 24,7, 23,5 y 23,2 cm2, respectivamente); este
grupo de cuatro genotipos fue estadísticamente igual y
sólo difirió significativamente del genotipo 10 (15,1 cm2;
Cuadro 4. Comparación de medias entre genotipos para madurez fisiológica (Mf días), altura de la planta en centímetros (ap), área de la hoja bandera en centímetros cuadrados (ahB), área de la segunda hoja en centímetros cuadrados (ash) y longitud de entrenudos en cm (len).
No. Cruza mf ap ahb ash len
1 TOLLOCAN F2005 125 b 88,1 a 21,2 ab 26,8 a 59,1 a
2 PSN/BOW//SERI/3/MILAN/4/ATTILA 131 ab 88,0 ab 20,8 ab 21,7 abc 59,7 a
3 SW90.1057/3/KAUZ*2/YACO//KAUZ/4/S91.12331
132 ab 81,0 abc 19,7 ab 22,1 abc 46,1 de
4 BL2064//SW89.5124*2/FASAN/3/TILHI
126 b 78,8 abcd 18,9 ab 19,6 bc 50,7 bcd
5 ALD/CEP75630//CEP75234/PT7219/3/BUC/BJY/4/CBRD/5/TNMU/PF85487
125 b 87,1 ab 24,7 a 25,1 ab 58,6 a
6 04W40483 130 ab 72,7 cd 21,7 ab 22,8 abc 48,0 cde
7 04W40677PPO 130 ab 79,7 abc 25,1 a 24,4 ab 50,8 bcd
8 05W90706 131 ab 79,1 abcd 21,4 ab 20,1 bc 53,5 abc
9 06W30565 129 ab 74,2 cd 18,9 ab 19,7 bc 46,5 de
10 06W30565 130 ab 74,8 cd 15,1 b 17,3 c 49,1 cde
11 06W30565 129 ab 76,0 cd 18,3 ab 20,9 abc 49,3 cde
12 06W30596 133 a 80,0 abc 23,5 a 21,6 abc 50,3 bcd
13 05W90911 131 ab 78,3 bcd 23,2 a 21,9 abc 56,3 ab
14 07W50636 131 ab 69,4 d 22,9 ab 22,9 abc 42,6 e
15 07W50744 132 ab 80,1 abc 19,6 ab 21,0 abc 49,0 cde
dMsh 7,3 9,8 7,9 6,4 6,9
Las medias con la misma literal dentro de cada columna son iguales estadísticamente. dMsh= diferencia mínima significativa honesta (Tukey).
Cuadro 4). Los rendimientos de grano de los cuatro
genotipos fueron de 4 740, 3 720 y 3 519 kg ha-1. En ash
el genotipo 1 tuvo el valor más alto (26,8 cm2) y superó
estadísticamente a 4, 8, 9 y 10 con 19,6, 20,1, 19,7 y 17,3
cm2 (Cuadro 4). El genotipo 1 (testigo) también produjo el
mayor rg. La len en los genotipos 1, 2 y 5 (59,1, 59,7 y 58,6
cm, respectivamente) fue igual estadísticamente pero éstos
difirieron significativamente de 9 líneas (medias de 42,6
a 50,8 cm; Cuadro 4). len y ap están muy relacionados,
por lo que ambas variables son indicadores importantes
del porte de planta; los genotipos de mayor len también
serían los de mayor altura de planta.

76
Torres et al., 2013. Rendimiento en trigo
Cuadro 5. Análisis de varianza, media general y coeficiente de variación para número de tallos por metro lineal (nt), número de espigas por metro lineal (ne), rendimiento de grano por hectárea (rg), peso hectolítrico del grano (ph), peso de mil granos (pMg), granos por espiga (ge), longitud de la espiga (le), espiguillas por espiga (ee) y peso de grano por espiga (pge).
F. V. G.L. nt ne rg ph pmg ge le ee pge
Genotipos 14 8,8** 3,53** 1,8 ns 7,3** 11,4** 15,7** 18,1** 11,3** 2,9**
Repeticiones 2 0,05 ns 0,9 ns 3,7 ns 0,8 ns 2,8 ns 0,3 ns 3,0 ns 0,9 ns 0,6 ns
Media 890,2 766,2 4163,9 73,8 38,4 59,0 9,8 19,2 2,4
C.V. 13,2 16,9 14,4 1,4 6,2 4,1 2,9 3,3 11,4
*Significativo al 0,05; **significativo al 0,01; NS, no significativo.
comparación de medias para las variables
del rendimiento de grano
Para nt, el genotipo 9 tuvo el mayor promedio
(1 364 tallos) y superó al testigo (783 tallos) y a 10 líneas
(entre 716 y 920 tallos; Cuadro 6). Los genotipos con
valores de nt entre 1 126 y 1 364 tuvieron rg entre
4 232 y 4 743, pero hubo genotipos con medias entre
751 y 920 que también produjeron rg entre 4 089 y
4 740 kg ha-1, por lo que se infiere que el incremento
o la disminución de la densidad de siembra en este
material genético también aumentaría la producción de
grano, pero estará en función del genotipo que se quiera
sembrar. Las medias para ne se clasificaron en tres grupos.
El genotipo más sobresaliente fue el 9 (1 079 espigas),
pero éste sólo se diferenció estadísticamente de la línea
7 (581 espigas). El testigo (genotipo 1) tuvo 724 espigas
(Cuadro 6). Aun cuando el incremento en el ne está
relacionado significativamente con un aumento en
nt, ambas variables podrían no contribuir a mejorar
el potencial productivo del trigo, debido a que habrá
genotipos que no respondan favorablemente a una mayor
o menor densidad de siembra.
Para el rg estadísticamente no hubo diferencias entre
genotipos, pero los de mayor rendimiento fueron 11, 5 y 1,
con 4 743,8, 4 740,1 y 4 740 kg ha-1 (Cuadro 6). El testigo
fue sobresaliente en ap, ahB, ash, len, ne, ph, pMg y pge;
el genotipo 11 sobresalió por su precocidad, resistencia al
acame, menor altura de planta, mayor número de tallos y
de espigas por metro lineal y peso hectolítrico del grano.
El genotipo 5 se distinguió por su precocidad, resistencia
a roya amarilla, mayor altura de planta, mayor longitud de
entrenudos, mayor área de la hoja bandera y de la segunda
hoja, mayor número de espigas por metro lineal, mayores
pesos hectolítrico y de 1 000 granos.
De acuerdo con la clasificación de producción de
Villaseñor y Espitia (2000), 10 genotipos (11, 5, 1, 4,
10, 2, 8, 9, 6 y 14, con 4 743,8, 4 740,1, 4 740, 4 535,4,
4 502,5, 4 429,5, 4 246,7, 4 232,1, 4 129,8 y 4 089,6 kg
ha-1, respectivamente) se encuentran en la categoría
“favorable”, la cual contempla una producción superior a
4 000 kg ha-1, dentro de ellos se encuentra el testigo. Los
cinco genotipos restantes (3, 12, 15, 13 y 7, con 3 877,5,
3 720,4, 3 661,9, 3 519,4 y 3 325,7 kg ha-1, respectivamente)
se encuentran en la categoría “regular”, que va de 2 000 a
4 000 kg ha-1. En la clasificación de Rodríguez et al. (2005)
sólo tres genotipos (11, 5 y 1, con 4 743,8, 4 740,1 y 4 740
kg ha-1, respectivamente) se agruparon en “favorable”, con
producciones de 4 715 a 6 173 kg ha-1, y los 12 genotipos
restantes (4, 10, 2, 8, 9, 6, 14, 3, 12, 15, 13 y 7, con 4 535,4,
análisis de varianza para las variables
del rendimiento de grano
El rendimiento de grano (rg) entre genotipos no fue
significativo (P > 0,05), pero en el resto de las variables
evaluadas los efectos fueron altamente significativos
(P < 0,01). Entre repeticiones tampoco hubo efectos
significativos (P > 0,05) (Cuadro 5).

77
CienCias agRiColas infoRma 22(2): 69-82. Julio-Diciembre 2013
4 502,5, 4 429,5, 4 246,7, 4 232,1, 4 129,8, 4 089,6, 3 877,5,
3 720,4, 3 661,9, 3 519,4 y 3 325,7 kg ha-1, respectivamente)
se encuentran en la categoría “regular 2” (de 3 134 a 4 338
kg ha-1). Estas clasificaciones de producción demuestran
que los genotipos evaluados tienen un buen potencial
de rendimiento para la región considerada en el presente
estudio.
Slafer et al. (2002) comentaron que el rendimiento
de grano depende principalmente del peso promedio
del grano y del número de granos por metro cuadrado,
pero ambas variables están correlacionadas negativa y
significativamente. También granos por metro cuadrado
está determinada por granos por espiga. Espigas por metro
cuadrado también depende de plantas por metro cuadrado
y de espigas por planta, mientras que granos por espiga
está determinada por espiguillas por espiga y por granos
por espiguilla. Estos son los componentes del rendimiento
más importantes que deben favorecerse a través de un
programa de mejoramiento o por medio de la aplicación
de tecnología para incrementar la producción en trigo.
Los genotipos con los valores de ph más altos fueron
2 y 11, con 76,2 y 75,5 g L-1, respectivamente, pero
ambos fueron iguales estadísticamente que el testigo.
Los dos genotipos más sobresalientes sólo difirieron
estadísticamente de las líneas 3, 7 y 12 (70,93, 70,63 y
71,50 g L-1) (Cuadro 6). Los genotipos con altos ph (2,
11 y 1) también produjeron rendimientos de grano de
4 740, 4 743 y 4 429 kg ha-1, mientras que los de menores
promedios en ph también rindieron 3 877, 3 325 y 3 720 kg
ha-1, por lo que se infiere que el peso hectolítrico del grano
puede ser un indicador confiable para la identificación de
genotipos sobresalientes.
Cuadro 6. Comparación de medias entre genotipos para número de tallos por metro lineal (nt), número de espiguillas por espigas (ne), rendimiento de grano en kilogramos por hectárea (rg) y peso hectolítrico del grano en gramos por litro (ph).
No. Cruza nt ne rg ph
1 TOLLOCAN F2005 783,3 cd 724,4 ab 4740 a 74,77 abc
2 PSN/BOW//SERI/3/MILAN/4/ATTILA 853,4 bcd 772,2 ab 4429 a 76,17 a
3 SW90.1057/3/KAUZ*2/YACO//KAUZ/4/S91.12331
673,3 d 630,0 ab 3877 a 70,93 cd
4 BL2064//SW89.5124*2/FASAN/3/TILHI 751,1 cd 692,2 ab 4535 a 73,67 abcd
5 ALD/CEP75630//CEP75234/PT7219/3/BUC/BJY/4/CBRD/5/TNMU/PF85487
768,9 cd 688,9 ab 4740 a 74,77 abc
6 04W40483 792,2 bcd 686,7 ab 4129 a 74,73 abc
7 04W40677PPO 857,8 bcd 581,1 b 3325 a 70,63 d
8 05W90706 947,8 abcd 814,4 ab 4246 a 74,97 ab
9 06W30565 1364,5 a 1078,9 a 4232 a 72,43 abcd
10 06W30565 1126,2 abc 950,0 ab 4502 a 75,43 ab
11 06W30565 1208,9 ab 998,9 ab 4743 a 75,50 a
12 06W30596 716,7 d 691,1 ab 3720 a 71,50 bcd
13 05W90911 766,6 cd 677,8 ab 3519 a 74,27 abcd
14 07W50636 920,0 bcd 792,2 ab 4089 a 73,37 abcd
15 07W50744 786,7 cd 714,4 ab 3661 a 73,33 abcd
dMsh 419,6 463,9 2148 3,9
Las medias con la misma literal dentro de cada columna son iguales estadísticamente. dMsh=diferencia mínima significativa honesta.

78
Torres et al., 2013. Rendimiento en trigo
Cuadro 7. Comparación de medias entre genotipos para peso de mil granos en gramos (pMg), granos por espiga (ge), longitud de la espiga en centímetros (le), espiguillas por espiga (ee) y peso de granos por espiga en gramos (pge).
No. Cruza pmg ge le ee pge
1 TOLLOCAN F2005 41 abc 56 cde 9,2 de 18,7 bcde 2,5 a
2 PSN/BOW//SERI/3/MILAN/4/ATTILA 41 abc 55 cde 11,2 a 18,3 cde 2,7 a
3 SW90.1057/3/KAUZ*2/YACO//KAUZ/4/S91.12331 39 abcd 60 bcde 9,9 bcde 21,3 a 2,7 a
4 BL2064//SW89.5124*2/FASAN/3/TILHI 47 a 52 e 9,2 de 17,3 e 2,6 a
5 ALD/CEP75630//CEP75234/PT7219/3/BUC /BJY/4/CBRD/5/TNMU/PF85487
39 abcd 55 cde 9,1 de 17,0 e 2,2 a
6 04W40483 45 ab 54 de 10,4 abc 18,0 de 2,7 a
7 04W40677PPO 31 d 56 cde 8,9 e 19,0 bcde 2,2 a
8 05W90706 39 abcd 70 a 10,1 bcd 20,0 abcd 2,8 a
9 06W30565 31 d 55 cde 9,7 cde 19,0 bcde 1,8 a
10 06W30565 36 bcd 57 bcde 9,2 de 18,7 bcde 2,2 a
11 06W30565 34 dc 56 cde 9,4 de 19,0 bcde 2,1 a
12 06W30596 34 dc 66 ab 10,8 ab 20,7 ab 2,3 a
13 05W90911 41 abc 63 abc 9,8 bcde
19,7 abcd 2,3 a
14 07W50636 37 bcd 62 abcd 10,6 abc 20,3 abc 2,4 a
15 07W50744 35 cd 69 a 9,9 bcd 20,3 abc 2,4 a
dMsh 8,5 8,6 1,0 2,3 0,9
Las medias con la misma literal dentro de cada columna son iguales estadísticamente. dMsh= diferencia mínima significativa honesta (Tukey).
El genotipo con mayor valor para pMg fue 4 con
47,5 g; éste se diferenció estadísticamente de los
genotipos 7, 9, 10, 11, 12, 14 y 15 (31,7, 31,7, 36,7, 34,2,
37,5 y 35,8 g, respectivamente) (Cuadro 7). Aun cuando
la línea 4 obtuvo alto rendimiento de grano (4 535 kg
ha-1) y alto pMg, en las líneas 7, 9, 10, 11, 12, 14 y 15,
estadísticamente inferiores en pMg, se registraron rg
tanto altos como bajos, por lo que se deduce que pMg
podría no ser considerada como un indicador confiable
para identificar genotipos de mayor rendimiento. Los
genotipos 8 y 15 tuvieron mayor número de ge (69,6 y
68,8) y ambos superaron estadísticamente al testigo y a
otros nueve materiales genéticos evaluados (52 granos)
(Cuadro 7).
Los genotipos 8 y 15 produjeron 4 246 y 3 661 kg ha-1.
El hecho de que los genotipos de mayor producción
de grano, 1, 5 y 11 con 4 740, 4 740 y 4 743 kg ha-1,
no sean los de mayor número de ge, sugiere que este
componente del rendimiento no debe ser considerado
como un indicador confiable para la identificación
de genotipos con mayor rg. De acuerdo con Slafer
et al. (2002), el rg depende principalmente de granos
por metro cuadrado y del peso promedio del grano;
granos por metro cuadrado tiene como componentes
principales a granos por espiga y espigas por m2, por
lo que quizás esta última variable podría ser de mayor
utilidad en el mejoramiento genético de este cereal.

79
CienCias agRiColas infoRma 22(2): 69-82. Julio-Diciembre 2013
El genotipo con mayor le fue el 2 (11,2 cm) y éste
superó al testigo (9,2 cm) y a otras 10 líneas (medias
entre 8,9 y 10,1 cm) (Cuadro 7). En teoría se esperaría
que un genotipo con mayor longitud de la espiga
también tuviera mayor rendimiento de grano, pero
esto no siempre es así debido a que, como Slafer et al. (2002) lo señalaron, el rg depende fundamentalmente
del número de granos por metro cuadrado y del peso
de grano promedio. El genotipo de mayor longitud de
espiga, identificado como 2, podría tener un menor
número de granos por espiga, pero mayor peso
promedio que un genotipo de menor longitud de espiga,
como el testigo, pero con granos de mayor peso
promedio.
Para la variable ee, el genotipo 3 tuvo el mayor valor
(21.3 ee) y superó estadísticamente al testigo (18,7 ee) y a
otras 9 líneas (medias entre 17 y 19 ee) (Cuadro 7). Esta
variable no es un indicador confiable para incrementar
el rg en trigo, debido a que el mejor genotipo (3) en ee
no tuvo el mayor rg. Slafer et al. (2002) indicaron que ee
y granos por espiguilla son los principales componentes
del número de granos por espiga, pero ee podría tener
poco valor práctico en la selección de mejores genotipos,
especialmente si ésta presenta correlación negativa y
significativa con rg, como se observó en el presente
estudio.
Hernández (1984) observó que el rg y ap se
correlacionaron negativa y significativamente, pero que
ee, le y ge tuvieron asociación positiva y significativa
con rg. Se detectó poca variabilidad fenotípica en pge.
Las diferencias entre los 15 genotipos, estadísticamente
no significativas, variaron de 1,8 a 2,8 g. Los genotipos
más sobresalientes fueron 8, 2, 3 y 6 (Cuadro 7).
Estos resultados sugieren que existe poca variabilidad
genética para pge en el germoplasma evaluado, pero
también podrían estar relacionados con lo expuesto
por Slafer et al. (1996), quienes sugirieron que en los
cultivares modernos hubo mayor número de granos
por metro cuadrado y menor peso de grano en los
cultivares antiguos, por lo que el avance en la mejora
genética del rendimiento se atribuye principalmente
a un incremento en la primera variable, pero que el
incremento en el potencial de rendimiento de grano
a través de un mayor número de granos por metro
cuadrado podría ser impedido o retardado por un
efecto compensatorio de la reducción del peso de
grano. Otra explicación sería que el incremento en el
número de granos por metro cuadrado se explicaría
por un mayor número de granos por espiga, pero de
menor peso.
análisis de correlación y de componentes
principales
El rendimiento de grano (rg) se correlacionó positiva
y significativamente con altura de la planta (ap) y peso
hectolítrico del grano (ph; r = 0,39** y r = 0,53**) y
negativa y significativamente con días a espigamiento
(de), madurez fisiológica (Mf) y espiguillas por espiga
(ee); (valores de r de -0,38*, -0,36*, -0,38* y 0,38*;
Cuadro 8, Figura 1). Estos resultados concuerdan con
los observados por Hsu y Watson (1971) y por Lupton
et al. (1974), quienes demostraron que el número
de espigas, granos por espiga y peso de granos por
espiga fueron los componentes del rendimiento más
importantes. En este mismo contexto, Slafer y Calderini
(2003) y Hewstone (2003) señalaron que el rg podría
incrementarse al favorecer la expresión fenotípica de
un mayor número de plantas por metro cuadrado,
de granos por espiguilla, de espigas por planta, de
espiguillas por espiga, del peso de grano promedio y
de granos por metro cuadrado, pero las correlaciones
observadas dependen de la variabilidad genética en las
variables evaluadas, del ambiente y de la interacción
genotipo x ambiente, por lo que estos resultados son
relativos.

80
Torres et al., 2013. Rendimiento en trigo
Cua
dro
8. A
nális
is de
cor
rela
ción
par
a dí
as a
esp
igam
ient
o (d
e) ,
días
a fl
orac
ión
(df)
, roy
a am
arill
a (r
a),
porc
enta
je d
e ac
ame
(pa),
mad
urez
fisio
lógi
ca (M
f), a
ltura
de
la p
lant
a (a
p), n
úmer
o de
tallo
s ( n
t),
núm
ero
de e
spig
as (n
e), r
endi
mie
nto
de g
rano
(rg),
peso
hec
tolít
rico
del g
rano
(ph),
peso
de
mil
gran
os (p
Mg),
gran
os p
or e
spig
a (g
e), l
ongi
tud
de la
esp
iga
(le)
, esp
igui
llas
por
espi
ga ( e
e), p
eso
de g
rano
por
esp
iga
(pg
e), á
rea
de la
hoj
a ba
nder
a (a
hB),
áre
a de
la s
egun
da h
oja
(ash
) y lo
ngitu
d de
ent
renu
dos
(len
).
de
df
ra
pam
fa
pn
tn
er
gph
pmg
ge
le
eepg
ea
hb
ash
len
de
0,71
**0,
17n
s- 0
,22n
s0,
56**
0 n
s-0
,33*
-0,2
5ns
-0,3
8*-0
,52*
*-0
,2n
s0,
36*
0,35
*0,
67**
0,04
ns
-0,0
3ns
-0,0
9ns
-0,3
2*
df
-0,0
1ns
-0,1
4ns
0,4*
*0,
4**
-0,4
6**
-0,3
5*-0
,24n
s-0
,23n
s-0
,03n
s0,
36*
0,36
*0,
51**
0,27
ns
0,13
ns
0,19
ns
0,13
ns
ra
0,09
ns
0,05
ns
-0,1
2ns
-0,0
3ns
-0,0
3ns
-0,2
1ns
-0,1
8ns
0ns
-0,1
5ns
-0,1
6ns
0,12
ns
0ns
-0,1
6ns
-0,1
2ns
-0,2
7ns
pa-0
,37*
0,41
**-0
,24n
s-0
,17n
s0,
16n
s0n
s0,
36*
-0,3
*-0
,16n
s-0
,3*
0,24
ns
-0,1
ns
0,2n
s0,
46**
Mf
-0,2
9ns
0,08
ns
0,07
ns
-0,3
6*-0
,39*
*-0
,34*
0,53
**0,
49**
0,64
**0,
05n
s0,
01n
s-0
,22n
s-0
,35*
ap
-0,4
**-0
,22n
s0,
35*
0,2n
s0,
24n
s-0
,07n
s-0
,06n
s-0
,18n
s0,
24n
s0,
21n
s0,
44**
0,78
**
nt
0,87
**0,
15n
s0,
17n
s-0
,46*
*-0
,18n
s-0
,17n
s-0
,07n
s-0
,38*
*-0
,29n
s-0
,29n
s-0
,31*
ne
0,25
ns
0,28
ns
0,29
ns
-0,0
9ns
0ns
-0,0
3ns
-0,1
9ns
-0,3
*-0
,2n
s-0
,22*
rg
0,53
**0,
39**
-0,3
8*-0
,1n
s-0
,38*
0,1n
s-0
,19n
s0,
15n
s0,
29n
s
ph0,
43**
-0,1
9ns
0,09
ns
-0,4
2**
0,21
ns
-0,2
2ns
-0,0
7ns
0,44
**
pMg
-0,2
3ns
0,12
ns
-0,3
6*0,
48**
-0,0
4ns
0,12
ns
0,35
*
ge
0,38
**0,
71**
0,17
ns
0,12
ns
-0,0
9ns
-0,1
2ns
le0,
39**
0,33
*0,
11n
s0n
s-0
,07n
s
ee
0,13
ns
0,03
ns
-0,0
7ns
-0,4
5**
pge
0,06
ns
0,16
ns
0,17
ns
ah
B0,
68**
0,2n
s
ash
0,36
*
len
* Si
gnifi
cativ
o al
0,0
5; *
* al
tam
ente
sig
nific
ativ
o al
0,0
1; n
s no
sig
nific
ativ
o.

81
CienCias agRiColas infoRma 22(2): 69-82. Julio-Diciembre 2013
Figura 1. Análisis de componentes principales para 15 genotipos de trigo, representados con números, y 18 variables agronómicas, escritas con letras mayúsculas.
En el Cuadro 8 y en la Figura 1 también se puede observar
que de se correlacionó positiva y significativamente con
días a floración (df), madurez fisiológica (Mf), granos por
espiga (ge), longitud de entrenudos (len) y espiguillas
por espiga (ee) (valores r de 0,71, 0,56, 0,36, 0,35 y
0,67, respectivamente). de también se asoció negativa
y significativamente con número de tallos (nt), peso
hectolítrico del grano (ph) y longitud de la espiga (le)
(Valores r de -0,33, -0,52 y -0,32, respectivamente). Otras
correlaciones positivas y negativas significativas también
se pueden detectar en el Cuadro 8 y éstas pueden
confirmarse en la gráfica del biplot de la Figura 1.
El genotipo 11 (06w30565) y el testigo superaron a los
demás al presentar el mayor rendimiento y tener mejores
características agronómicas (menos acame, más tallos
por metro lineal, más espigas por metro lineal y mayor
peso hectolítrico del grano), que permiten asegurar un
mayor rendimiento y disminuir riesgos de pérdidas en la
producción de grano.
conclusIonEs
El genotipo 11 (4 743,8 kg ha-1) y el testigo tuvieron
la misma producción de grano y ambos superaron
numéricamente las líneas 7 y 13 (3 326 y 3 519 kg
ha-1, respectivamente), con casi 1 400 y 1200 kg ha-1. Los
genotipos precoces tuvieron mayor rendimiento, como el
genotipo 11 (06w30565) con 130 días a madurez fisiológica
(Mf). El genotipo más tardío fue el 12 (06w30596) con 134
días a Mf y rendimiento de 3 720,4 kg ha-1. El genotipo
11 (06w30565) presentó un porcentaje de acame de 1,6,
y altura de 0,76 cm; el testigo (cv. Tollocan) presentó
41,7% de acame y altura de 88,1 cm. La capacidad de
amacollamiento en el genotipo 11 (06w30565) fue mejor
(1 209 tallos y 999 espigas por metro lineal); en el testigo
(cv. Tollocan) se registraron 783 tallos y 724 espigas. El
peso hectolítrico del grano en el genotipo 11 (06w30565)
fue de 75,5 g L-1 y el del testigo (cv. Tollocan) fue de 74,77
g L-1. Los tres genotipos más resistentes a roya amarilla

82
Torres et al., 2013. Rendimiento en trigo
fueron 5, 8 y 12 (3,3, 5,0 y 5,0%, respectivamente). Los
genotipos más resistentes al acame fueron 6, 12 y 14 (0%).
Los genotipos con mayor longitud de la espiga fueron 2,
12 y 14 (11,2, 10,8 y 10,6 cm). El material con mayor peso
específico del grano fue 2, 10 y 11 (76,17, 75,43 y 75,50
g L-1). El aumento en el rendimiento de grano en los 15
genotipos de trigo fue atribuido significativamente a un
aumento en su altura de planta, peso de 1 000 granos y
peso hectolítrico del grano; la disminución en su potencial
productivo se asoció significativamente a su precocidad y
a un menor número de espiguillas por espiga y de granos
por espiga.
REFEREncIas bIblIoGRÁFIcas
1. Balbuena, B. A., A. González H., E. Rosales R., A. Domínguez L., O. Franco M. y D. J. Pérez L. 2008. Identificación de genotipos sobresalientes de trigo en el Valle de Toluca, México. Agric. Téc. Méx. 34: 257-261.
2. García, E. 1981. Modificaciones al sistema de clasificación climática de Köppen. Larios. México, 252 p.
3. Hernández, S. A. 1984. Antecedentes del mejoramiento genético del trigo en México. Germen. Año 2, núm. 2. Texcoco, México. 5 p.
4. Hsu, P. and P. D. Walton. 1971. Relationships between yield and its components and structures above flag leaf node in spring wheat. Crop Sci. 11: 190-193.
5. Hewstone, M. C. 2003. Rediseño de componentes de rendimiento y su interacción con el manejo. En: Mohan K. M., M. Díaz A., M. Castro (eds.). Seminario Internacional sobre Estrategias y Metodologías Utilizadas en el Mejoramiento de Trigo: un Enfoque Multidisciplinario. CiMMyt-inia. La Estanzuela, Uruguay. pp. 25-35.
6. Lupton, F. G. H., R. H. Oliver and P. Ruckenbauer. 1974. An analysis of the factors determining yield in crosses between semi-dwarf and taller wheat varieties. J. Agric. Sci. Camb. 82: 483-496.
7. Ma, H., R. P. Singh and A. Mujeeb-Kazi. 1997. Resistance to stripe rust in durum wheats, A-genome diploids, and their amphiploids. Euphytica. 94: 279-286.
8. Martínez, G. A. 1988. Diseños experimentales. Métodos y elementos de teoría. Trillas. México. 756 p.
9. Rodríguez-Pérez, J. E., J. Sahagún-Castellanos, H. E. Villa Señor-Mir, J. D. Molina-Galán y A. Martínez-Garza. 2005. La interacción genotipo por ambiente en la caracterización de áreas temporaleras de producción de trigo. Agrociencia. 39: 51-64.
10. Sánchez, G. J. J. 1995. El análisis biplot en clasificación. Rev. Fito-tecnia Mex. 18: 188-203.
11. Slafer, G. A. 1996. Differences in physic development rate amongst wheat cultivars independent of responses to photoperiod and vernalization. A viewpoint of the intrinsic earliness hypoth-esis. J. Agron. Crop Sci. 126: 403-419.
12. Slafer, G. A. y D. F. Calderini. 2003. Herramientas fisiológicas para el mejoramiento del rendimiento de trigo. En: M. Mohan K., M. Díaz A., M. Castro (eds.). Seminario Internacional sobre Estrategias y Metodologías Utilizadas en el Mejoramiento de Trigo: un Enfoque Multidisciplinario.CiMMyt-inia. La Estanzuela, Uruguay. pp. 13-24.
13. Slafer, G. A., R. Savin, D. F. Calderini y D. J. Miralles. 2002. Bases fisiológicas para el manejo y mejoramiento de cebada. En: Actas del III Congreso Latinoamericano de Cebada. inia. Colonia, Uruguay. pp. 54-63.
14. Villaseñor, M. H. E. y E. Espitia R. 2000. Características de las áreas productivas de trigo de temporal: problemática y condiciones de producción. En: Villaseñor, M. H. E. y E. Espitia R. (eds.). El trigo de temporal en México. Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias. Secretaría de Agricultura y Ganadería. México. pp: 85–98.
15. Zadoks, J., C. Ghang and C. F. Konzak. (1974). A decimal code for the growth stages of cereals. Weed Res. 14: 415-421.

83
Aceite de ricino (Ricinus communis L.) con aplicaciones en comunicaciones ópticas
oil from castor oil plant (Ricinus communis l.) with applications in optical communications
Ernesto Díaz-López,1* Israel Jesús Orlando-Guerrero,1 Jesús Manuel Campos-Pastelín,1
Irma Brena-Hernández,1 Juan Manuel Loeza-Corte1
CIENCIAS AGRICOLAS INFORMA 201322(2): 83-87
Recibido: 9 de agosto de 2013Aceptado: 1 de diciembre de 2013
REsumEn
En la Cañada Oaxaqueña se colectaron semillas de ricino (Ricinus communis L.) de tres accesiones, de las cuales se obtuvo aceite para fabricar una fibra óptica monomodo, y se determinaron posibles aplicaciones en comunicaciones. Con el núcleo líquido de aceite se evaluó la pérdida y atenuación de luz respecto a la longitud de fibra, cuyo aislante fue vidrio crown con un índice de refracción 1,52 y utilizando como fuente luz blanca y un láser rojo He-Ne. Los resultados indican que la pérdida, así como la atenuación, son mayores a una longitud de 0,80 m en la fibra, lo que de alguna forma limita su uso en comunicaciones, pero puede tener otros usos como en endoscopía con aplicaciones médicas.
Palabras clave: Atenuación, Cañada Oaxaqueña, fibra monomodo, ricino.
abstRact
In the Cañada Oaxaqueña, the castor oil plant seed (Ricinus communis L.) from three accessions were collected, Seed-oil obtained was used to fabricate a single-mode fiber, and its potential applications in communications was identified. With the liquid core of oil it was evaluated and compared the loss and light attenuation in respect to the fiber length, that fiber was insulated with crown glass, having a refractive index of 1.52 and a red He-Ne laser was uses as a source of white light. The results indicate that loss and attenuation, are higher at a length of 0.80 m in the fiber, which really limits its use in communications, but might have other uses such as in medical endoscopy applications.
Key words: Attenuation, Cañada Oaxaqueña, castor oil plant, single-mode fiber.
1 Universidad de la Cañada, México.
*Autor para correspondencia: [email protected].

84
Díaz-lópez et al., 2013. Ricino en comunicaciones ópticas
IntRoduccIón
El petróleo ha sido por muchos años la materia principal
para sintetizar polímeros, los cuales tiene diferentes
aplicaciones como: plásticos, hule espuma e incluso
materiales que pueden transmitir ondas luminosas
que tienen uso en comunicaciones, tal es el caso de las
fibras ópticas (Meneses et al., 2007). Actualmente, los
suministros de este recurso no renovable han disminuido
considerablemente, lo que obliga al ser humano a buscar
nuevas fuentes de energía y materiales que puedan ser
útiles en la fabricación de polímeros (Stevens, 2002).
Una alternativa a esta problemática puede ser el uso de
materiales de origen vegetal que crecen en ecosistemas
áridos y secos como la Cañada Oaxaqueña, tal es el caso
del ricino (Ricinus communis L.) multidendricaule (plantas
que presentan múltiples ramificaciones) que tiene su origen
en el sur del continente africano, y que fue introducido en
América por los portugueses, principalmente en América
del Sur de donde fue llevado a México por los españoles
(Acevedo et al., 1997). Esta planta fue utilizada por muchos
años para la extracción de ácidos grasos de sus semillas,
de las cuales se extrae un aceite secante de alta calidad.
Con el incremento en la explotación del petróleo, después
de la segunda guerra mundial, este cultivo fue marginado
hasta quedar casi en el olvido (Acevedo y Lima, 2001).
Se reconoce que el uso indiscriminado de hidrocarburos
ha provocado un incremento en la temperatura mundial,
fenómeno conocido como “calentamiento global”
ocasionado por los gases de efecto invernadero que
impiden la disipación del calor del planeta hacia el
espacio, por ello surge la necesidad de encontrar nuevos
recursos que funcionen como alternativas y sustitutos de
los productos obtenidos del petróleo crudo, y que sean
amigables con el ambiente (Laine, 2009), además de tener
uso en las telecomunicaciones. Una posible solución a
esto, puede ser la utilización de aceites vegetales como el
del ricino, que posee algunas características de interés para
las comunicaciones como la transmitancia de fotones por
medio de fibras líquidas (Meier y Metzgerb, 2007; Cruz et al., 2012). A este respecto, se han realizado investigaciones
sobre las propiedades químicas de ácidos grasos y sus
propiedades físicas; Cano et al. (2008) mencionan que las
propiedades físicas del aceite de Monthostachys dependen
principalmente de la composición química, lo que trae
como consecuencia que sus propiedades ópticas sean
diferentes a las de otras especies del género. Fernando
et al. (2012) comentan, por otro lado, que los aceites
vegetales poseen altos índices dieléctricos en comparación
con los aceites provenientes de derivados del petróleo,
y concluyen que aquéllos pueden tener aplicaciones en
electrónica, electricidad y óptica, debido a sus propiedades
dieléctricas. Otros investigadores como Franco (2005)
mencionan también que por su bajo índice de refracción,
algunos aceites poseen propiedades importantes para
refractar la luz y por ello se utilizan en microscopía
como el aceite de inmersión con un bajo índice de
refracción (1,517), el cual es empleado en microscopía
de fluorescencia para identificar bacterias con la lente
objetivo de 100X. Basados en estas investigaciones, el
presente trabajo pretende argumentar a favor del uso
de estos metabolitos y sus posibles aplicaciones en la
fabricación de equipos, como las fibras ópticas.
matERIalEs y métodos
El presente estudio se llevó a cabo en la Cañada Oaxaqueña
ubicada a 18o 06‘ de L.N. y 98o 06‘ L.W. y a 880 msnm,
bajo un clima Bs1e´g que corresponde a un clima seco,
con una temperatura media anual que oscila entre 18 oC ≤
X ≤ 27 oC. La precipitación es de junio a septiembre con
un total de 550 mm, oscilación de la temperatura mayor
a 14 oC y el mes más cálido se presenta antes del solsticio
de verano, que para la región ocurre en abril (García,
2005). El material genético (semillas) fue colectado de tres
accesiones localizadas en la comunidad de San Antonio
Nanahuatipam, las cuales se encontraban en los límites de
plantaciones de caña de azúcar (Saccharum officinarum
L.), ubicadas a 18o 07‘ L.N. y 98o 05‘ L.W. para la accesión
uno, 18o 08‘ L.N. y 98o 04‘ L.W. para la accesión dos y
18o10‘ L.N. y 98o 03‘ L.W. para la tercera; estas
localizaciones se realizaron con ayuda de un gps (Garmín,
modelo eTrex Legend HCX).

85
CienCias agRiColas infoRma 22(2): 83-87. Julio-Diciembre 2013
Las plantas madre (accesiones) fueron identificadas con
las claves especializadas para la familia Euphorbiaceae,
de las cuales se colectaron los frutos (cápsulas), que
contienen aproximadamente cuatro semillas, éstas se
transportaron en bolsas de papel previamente identificadas
por accesión. De cada accesión se extrajo el aceite por
el método de prensado, utilizando una prensa Cropland
modelo CLB-300 con motor de 7,5 HP trifásico. Una vez
extraído el aceite en bruto, éste fue separado con ayuda
de un rota-vapor marca EG Technic modelo 9200 y un
solvente (hexano), con el objeto de separar y obtener un
mayor grado de pureza. A continuación se procedió a
almacenarlo en recipientes de vidrio color ámbar, para
posteriormente realizar la fibra óptica líquida; el llenado
de ésta se llevó a cabo aplicando presión hidrostática
constante con ayuda de una bomba de vacío para evitar la
formación de burbujas, el aislante fue un capilar de vidrio
crown cuyo índice de refracción fue de 1,52 y su diámetro
interior de 0,006 m, el largo del tubo fue de un metro y el
núcleo de aceite de ricino presentó un índice de refracción
de 1,57, el cual se determinó utilizando un refractómetro
portátil marca Atago. La luz blanca de un led y de un
láser de Helio-Neón fue acoplado a la fibra óptica líquida
utilizando un adaptador SX marca Edmund. Una vez
lleno el capilar, se realizaron las pruebas (mediciones) de
pérdidas de ancho de banda vs. atenuación y atenuación
vs. longitud efectiva de fibra, para fines de caracterización
de la fibra óptica. En la primera medición se obtuvieron
gráficas de ancho de banda en un rango de 300-800 nm de
longitud de onda contra pérdida espectral (dB m), y en la
segunda se obtuvieron gráficas de atenuación para 632,8
nm contra longitud efectiva de la fibra (m). La primera
medida fue realizada utilizando un analizador de espectro
óptico con una resolución nominal de 5 nm. La pérdida
de la fibra en función del ancho de banda se calculó
utilizando la relación (Saleh y Teich, 1991):
Donde L es la longitud de la fibra, I1 es la intensidad
de entrada de la fibra y I2 es la intensidad de salida
de la fibra. La atenuación (dB/m) presente en la fibra
fue medida con la técnica “cutback metod” o método
reducido, la cual consiste en comparar la potencia de
entrada y salida para segmentos de la fibra óptica. Esto
se llevó a cabo con un láser de He-Ne cuya longitud de
onda central es de 632,8 nm, las potencias de entrada y
salida fueron obtenidas con un medidor de potencia con
segmentos de fibra de 0,05 m. La atenuación sufrida por
la fibra óptica para cada segmento se calculó utilizando
la relación (Saleh y Teich, 1991):
Donde I0 es la intensidad de entrada de la fibra y I1
es la intensidad de salida de la fibra para cada
segmento.
REsultados y dIscusIón
La Figura 1 muestra algunas características respecto a la
pérdida espectral sufrida para diferentes longitudes de
onda, en ella se puede observar que la fibra óptica líquida
sufre pérdidas menores a 1,5 dB m-1 para longitudes de
onda que van de 500 a 800 nm, es decir, cerca de la región
infrarroja. De este modo se afirma que la fibra puede
transmitirse de forma adecuada dentro de esta región.
Asimismo, en esta misma figura se puede observar que
la fibra óptica líquida sufre pérdidas significativas para
longitudes de onda de 300 a 500 nm, donde las pérdidas
abarcan un rango de 7 a 1,5 dB m-1, lo que hace que la
fibra no sea viable para transmitir en esas longitudes de
onda. Estos resultados coinciden con los reportados por
Urrutia et al. (2006), Broeng et al. (1999), Gambling et al. (1972), Stone (1972a) y Stone (1972b), quienes mencionan
que las pérdidas en dB en una fibra óptica tienen que ver
con el material del cual está elaborado el núcleo, así se
trate de fibras de cristal fotónico (fCf) o fibras líquidas, y
especifican que el rango de pérdida oscila entre los 300 a
450 nm.
En la Figura 2 se muestra la atenuación contra longitud
efectiva de fibra, y se puede apreciar que los datos obtenidos
se ajustaron a un modelo lineal, con un coeficiente de

86
Díaz-lópez et al., 2013. Ricino en comunicaciones ópticas
determinación alto de 0,99, lo que indica que 99% de la
atenuación en la fibra monomodo se debe a la longitud de la
fibra. Respecto a la pendiente, ésta indica que por cada metro
lineal de fibra, se tuvo una atenuación en la misma de 1,9 dB,
demostrando así que el núcleo de ricino puede transmitir luz
blanca y fotones emitidos por un láser rojo. Los resultados
coinciden con los datos experimentales de Gutiérrez et al. (2009) y Wang et al. (1998), quienes aseveran que
el ensanchamiento espectral de la atenuación en una
fibra se debe a la inestabilidad modulacional y al auto-
desplazamiento de la frecuencia, y esto a su vez depende de
la longitud de fibra y de la potencia de bombeo de la fuente.
De acuerdo con lo encontrado en la presente investigación,
la fibra monomodo con núcleo de aceite de ricino tendría
pocas aplicaciones en comunicaciones debido a la pérdida
y atenuación, ya que en este tipo se utilizan fibras de
longitudes grandes. No obstante, la fibra monomodo con
núcleo de aceite de ricino podría tener aplicaciones como
Figura 1. Pérdida espectral contra longitud de onda en una fibra líquida monomodo con núcleo de aceite de ricino (Ricinus communis L.), en la Cañada Oaxaqueña. Primavera 2013.
Figura 2. Atenuación contra tamaño efectivo de fibra con núcleo de aceite de ricino (Ricinus communis L.), en la Cañada Oaxaqueña. Primavera 2013.

87
CienCias agRiColas infoRma 22(2): 83-87. Julio-Diciembre 2013
fuente de iluminación y tener uso en la endoscopía con
aplicaciones médicas. A este respecto, se sugiere que en
trabajos de esta índole se estudien diferentes materiales
que sirvan como aislante al núcleo líquido de ricino y
se analice su comportamiento en cuanto a la pérdida y
atenuación.
conclusIonEs
Las mayores pérdidas, así como los valores de
atenuación en la fibra líquida monomodo con
núcleo de aceite de ricino, se presentaron en longitudes de
0,80 m, por ello se infiere que este tipo de fibras presenta
grandes limitaciones para ser utilizadas en comunicaciones,
pero pueden funcionar como fuentes luminosas que
podrían usarse en medicina con uso en endoscopía. A
este respecto, los ecosistemas con climas secos como la
Cañada Oaxaqueña, pueden ser una alternativa para
producir materiales aplicables en la industria e incluso
en la medicina, al sembrar plantas como el ricino
que prosperan en ambientes donde otras especies no
lo harían y obtener así aceite útil en aspectos como se
mencionaron anteriormente. De manera general, el
ricino puede ser una fuente importante para fabricar
fibras líquidas monomodo que podrían emplearse como
fuentes luminosas.
aGRadEcImIEntos
Los autores del presente estudio agradecen el apoyo
económico recibido del Programa de Mejoramiento del
Profesorado (proMep) para el proyecto con clave idCa 11749.
REFEREncIas bIblIoGRÁFIcas
1. Acevedo, D. M. P. y E. F. Lima. 2000. O agronegocio da mamona no Brasil. Embrapa informacao tecnológica. Brasilia, Brasil. 350 p.
2. Acevedo, D. M. P, E. F. Lima, F. A. S. Batista, N. E. Beltrao, J. J. Soares, R. M. Vieria y J. A. Moreira. 1997. Recomendacoes técnicas para o cultivo da mamona (Ricinus communis L.) no
nordeste do Brasil. Circular Técnica 25. eMBrapa-Cnpa. Campina Grande, Brasil. 39 p.
3. Akahashi, H. 1985. Optical transmission loss of liquid-core silica fibers in theinfrared region. Optical Comm. 53: 164-168.
4. Broeng, D., D. Mogilevstev, S. E. Barkou and A. Bjarklev. 1999. Photonic crystal fibers: A new class of optical waveguides. Optical Fiber Tech. 5: 305-330.
5. Cano, C., P. Bonilla, M. Roque y J. Ruiz. 2008. Actividad antimicótica in vitro y metabolitos del aceite esencial de las hojas de Minthostachys Millis (Muña). Rev. Peru. Med. Exp. Salud Pública. 25: 298-301.
6. Cruz, A. K., G. A. Sáenz, S. J. Montañez, A. C. Noé y L. E. Flores. 2012. Aceites vegetales: una fuente renovable y económica para obtener plásticos. Rev. Cient. Univ. Aut. Coahuila. 4: 1-8.
7. Fernando, N. D., R. H. Cadavid y E. D. Fernando. 2012. Aplicación del aceite dieléctrico de origen vegetal en transformadores eléctricos. Ingeniería y Universidad. 16: 201-223.
8. Franco, C. Y. 2005. Reseña de la microscopía de fluorescencia y su aplicación en el diagnóstico de bacterias fitopatogénicas. Fito-sanidad. 9: 65-68.
9. Gambling, W. A. 1972. Dispersion in low-loss liquid-core optical fibres. Electr. Lett. 8: 2-3.
10. García, E. 2005. Modificaciones al sistema de clasificación climática de Köppen, para adaptarlo a las condiciones de la República Mexicana. unaM. México. 217 p.
11. Gutiérrez, J. G., T. M. Vargas, S. C. Romero, F. O. Hernández, E. A. Kuzin, A. M. Estudillo, C. R. Grajales, L. R. Rojas, y Z. F. Gutiérrez. 2009. Influencia de la inestabilidad modulacional en la generación de un espectro continuo en fibras ópticas con pulsos de nanosegundos. Rev. Mex. Física. 55: 359-366.
12. Laine, J. 2009. Ciento cincuenta años de combustión de hidrocarburos fósiles: las alternativas emergentes. Ingeniería y Ciencia. 5: 11-31.
13. Meier, M. A. R. y J. O. Metzgerb. 2007. Plant oil renewable re-sources as green alternatives in polymer science. Chem. Soc. Rev. 36: 1788-1802.
14. Meneses, J., C. M. Corrales y M. Valencia. 2007. Síntesis y caracterización de un polímero biodegradable a partir del almidón de yuca. Rev. Esc. Ing. Antioquia. 8: 57-67.
15. Saleh, B. E. A. and M. C. Teich. 1991. Fundamentals of photonics. John Wiley & Sons. New York, USA. 966 p.
16. Stevens, E. S. 2002. Green plastics: an introduction to de new science of biodegradable plastics. Princeton University Press. New Jersey, USA. 238 p.
17. Stone, J. 1972a. Optical transmission in liquid-core quartz fibers. Appl. Physics Lett. 20: 239-240.
18. Stone, J. 1972b. Optical transmission loss in liquid-core hollow fibers. ieee J. Select Topics Quantum Electron. March: 386–388.
19. Urrutia, L. M., D. González, L. C. Lomer, P. B. Quintela, A. García, y H. M. López. 2006. Sensibilidad de fibras de cristal fotónico con sus parámetros estructurales: su aplicación en sensores. En: XXI Simposium Nacional de la Unión Científica Internacional de Radio. Oviedo, España (Cd).
20. Wang, W. 1998. Spectrophotometry with liquid-core optical fibre in aqueous solution phase in the ultraviolet region. Anal. Chem. Acta. 375: 261–267.


89
Tácticas para el control del tizón gomoso del tallo en el cultivo de sandía
Integration of tactics for the control of the gummy stem blight in the cultivation of watermelon
Jesús Pérez-González,1* Benedicto Martínez-Coca,2 Salvador Guadarrama-Valentín,3 Sonia Estrada-Terra,1 Danay
Infante-Martínez,2 Yanisia Duarte-Leal,2 Claudio Esquivel-Álvarez3
CIENCIAS AGRICOLAS INFORMA 201322(2): 89-95
Recibido: 10 de abril de 2013Aceptado: 14 de junio de 2013
REsumEn
En la Isla de la Juventud (Cuba) el tizón gomoso del tallo de la sandía, causado por Didymella bryoniae, se presenta anualmente en 31,68% de las hectáreas dedicadas al cultivo. Por ello se propuso como objetivo evaluar la eficacia de la integración de tácticas culturales, biológicas y químicas para el manejo del tizón gomoso del tallo en sandía. Los resultados reflejaron que la azoxistrobina, tiofanato de metilo y clorotalonilo tienen una acción residual de hasta 18 días en época de primavera y de invierno. La integración de las tácticas, como siembra en áreas con cuatro años de barbecho herbáceo, inoculaciones de la cepa A- 34 de Trichoderma harzianum Rifai al suelo, semillas y follaje, y la aplicación de azoxistrobina, disminuyó la intensidad de la enfermedad e incrementó los rendimientos en sandía en ambas épocas.
Palabras clave: Barbecho, fungicidas, manejo de gomosis, químicos, Trichoderma.
abstRact
One of the diseases influencing watermelon (Citrullus lanatus (Thunb.) Matsum and Nakai) yield decrement, is the gummy stem blight caused by Didymella bryoniae (Fuckel) Rehm. In the Isle of Youth, the illness is presented annually in 31.68% of the hectares sowed with this crop. Thus, objective of present research was to evaluate the effectiveness of four fungicides and the integration of cultural, biological and chemical management, for their control. The results reflected that azoxistrobina, methyl tiofanato and clorotalonilo have a residual action up to 18 days in spring and winter. Mixing techniques such as plantation during four years of herbaceous fallow, with inoculations to the soil, seeds and applications to the foliage with the strain A - 34 of Trichoderma harzianum Rifai, and the azoxistrobina application, diminished the intensity of the illness and it increased the yields in watermelon in both times.
Key words: Chemical, fallow, fungicides, handling of gummy stem blight, Trichoderma.
1 Universidad Isla de la Juventud “Jesús Montané Oropesa”, Cuba. ²2 Centro Nacional de Sanidad Agropecuaria, Cuba. 3 Facultad de Ciencias Agrícolas, Universidad Autónoma del Estado de México, México.
*Autor para correspondencia: [email protected]

90
Pérez et al., 2013. Tizón gomoso del tallo
IntRoduccIón
Las pérdidas causadas en el cultivo de sandía por el
tizón gomoso difieren por países y por época. En Estados
Unidos, la enfermedad ha provocado pérdidas desde
15% de la producción (Keinath et al., 1997) hasta 43%
(Keinath y Duthie, 1998), fundamentalmente en época de
primavera. En Brasil, Figueiredo et al. (1966) observaron
que la severidad de la enfermedad en las frutas varió de 19
a 51% en función de la época de siembra. Su incidencia en
la India aumentó de 13% en 1999 a 21% en 2003 (Sudisha
et al., 2004). En la Isla de la Juventud, Cuba, el tizón
gomoso del tallo (Didymella bryoniae (Fuckel) Rehm (syn.
Mycosphaerella melonis (Pass) W. F. Chiu y J. C. Walker) se
presenta anualmente en 31,68% del área sembrada
de sandía (Citrullus lanatus (Thunb.) Matsum
and Nakai).
Los síntomas del tizón gomoso del tallo aparecen en
hojas, tallos y frutos. Estos se extienden desde el centro de
la planta de sandía hacia los extremos o guías. Las manchas
al inicio se muestran como pequeñas áreas cloróticas o
puntos que crecen rápidamente, frecuentemente aparecen
por el borde de las hojas y entre las nerviaciones. En las
manchas foliares se distingue un halo pequeño de color
amarillo con exudado gomoso. Al envejecer éstas, se
observan en el centro fructificaciones de color negro,
que en las hojas jóvenes generalmente se corresponden
con picnidios producidos en la fase anamórfica Phoma cucurbitacearum (Fr.:(Fr) Sacc.) y en las viejas con
pseudotecios de la fase teleomórfica Didymella bryoniae (Fuckel) Rehm (Pérez et al., 2012a).
Las lesiones en el tallo primario comienzan por la base,
sobre todo en el primer metro de longitud del mismo,
pero se pueden encontrar incluso en los secundarios. Al
inicio son de forma elipsoidal y de color parduzco pálido.
Posteriormente, la mancha se alarga y se hace visible una
hendidura en su centro con exudado gomoso, con presencia
de cuerpos de fructificación, los picnidios parcialmente
inmersos en el tejido y los pseudotecios sobre el mismo
(Pérez et al., 2012a).
A pesar de que internacionalmente se han investigado
diferentes tácticas de control de la enfermedad, la química
es la más recomendada (Santos et al., 2005; Keinath y
DuBose, 2009). También, esta es la más utilizada en la
Isla de la Juventud, a pesar del alto costo económico y
ecológico. Son muchos los fungicidas recomendados
para el control del tizón gomoso del tallo. Sikora
(1994) recomendó el clorotalonilo, mancozeb, maneb y
tiofanato metilo. Keinath (2001) combinó clorotalonilo
con benomil y mancozeb. Santos et al. (2005), en Brasil,
indicaron mancozeb y difenoconazol, tiofanato metilo y
clorotalonilo, mancozeb, trifloxistrobina, propiconazol,
oxicloruro de cobre para el control en melón. Keinath y
DuBose (2009) recomendaron en sandía, switch alternado
con clorotalonilo y cobre. A pesar de emplearse una gama
de productos químicos para combatir la enfermedad, aún
el control no es eficaz.
Otras medidas recomendadas para el control de la
enfermedad son: rotación de cultivos hasta cuatro años
(Pérez et al., 2012b); utilización de suelos sueltos, arenosos,
con buen drenaje para la siembra de cucurbitáceas; evitar
riego por aspersión; no provocar daños mecánicos; control
de posibles insectos vectores, uso de variedades comerciales
con aceptables resistencia, manejo poscosecha (Ferguson et al., 2009), así como la aplicación al suelo, semillas y al follaje
de Trichoderma harzianum Rifai (Martínez et al., 2012).
Se ha investigado la epifitiología y control del tizón
gomoso del tallo, pero para su manejo sólo se ha realizado
una sumatoria de los resultados obtenidos en diferentes
países. Por tal motivo el objetivo del presente trabajo fue
integrar tácticas culturales, biológicas y químicas, para el
manejo del tizón gomoso del tallo en la Isla de la Juventud.
matERIalEs y métodos
Los experimentos se realizaron en la Isla de la Juventud,
en época de primavera y de invierno, en suelos alíticos
de baja actividad arcillosa amarillento típico (Hernández
et al., 1999). Las atenciones culturales se realizaron según Minag (1998). Los productos químicos utilizados en los
experimentos fueron seleccionados a partir de la estrategia
recomendada por la Estación de Protección de Plantas de
la Isla de la Juventud y la literatura especializada para

91
CienCias agRiColas infoRma 22(2): 89-95. Julio-Diciembre 2013
el control del tizón gomoso del tallo (Keinath, 2001;
Santos et al., 2005). Las aspersiones de los productos
fitosanitarios se realizaron con una mochila Matabi de 16
L de capacidad, con boquilla cónica. Las evaluaciones se
realizaron diariamente hasta la aparición de las primeras
manchas, y posteriormente cada siete días, de las manchas
activas en todas las guías y hojas. Se utilizó la escala de
cuatro grados (Pérez et al., 2003).
Se utilizaron diseños de bloques al azar con cinco
repeticiones por tratamiento y 10 plantas como unidad
experimental; las parcelas experimentales tenían un área
de 168 m2 y el marco de siembra fue de 2,80 x 1,0 m. Las
cosechas se realizaron en función de la época de siembra.
La intensidad de la enfermedad se determinó por la fórmula
de McKiney (1923): y los datos se transformaron con la
expresión arcos p+1. Los rendimientos se determinaron
por área evaluativa en cada parcela. Los mismos fueron
sometidos a un análisis de varianza de clasificación simple
y las medias se compararon según la Dócima de Rangos
Múltiples de Duncan (Lerch, 1997).
duración de eficacia de algunos fungicidas
en el control del tizón gomoso del tallo
en sandía
Los experimentos se realizaron en los años 2004 y 2005.
Las semillas de sandía (0,5 kg) se inocularon con D. bryoniae (105 conidios/mL-1), por el método de imbibición
durante 20 min. Después de la siembra, el cultivo fue
monitoreado hasta que la enfermedad en las parcelas
alcanzó una intensidad homogénea. Posteriormente, se
aplicaron cuatro fungicidas en parcelas independientes:
1. clorotalonilo (Bravo SC 72 a 1 kg i.a. ha-1), 2. tiofanato
metilo (Topsin- M PH 70 a 0,3% i.a. ha-1); 3. azoxistrobina
(Amistar SC 25 100 g i.a. ha-1); 4. mancozeb (Mancozeb PH
80 a 2,4 kg i.a. ha-1); 5. testigo (tratamiento sin aplicación). Las
aplicaciones en los tratamientos comenzaron a los 25 días
de sembrado el cultivo. Se realizaron cuatro evaluaciones
a partir del quinto o séptimo día de aplicación, hasta que
el último de los tratamientos indicara señal de una nueva
aplicación del control químico.
Integración de tácticas para el manejo
del tizón gomoso del tallo
En áreas sembradas con sandía en los meses de mayo
y noviembre de los años 2007 y 2009, manejadas con
las diferentes tácticas, se evaluó la intensidad de la
enfermedad y los rendimientos. Las tácticas evaluadas
fueron: Siembra en un suelo con cuatro años de barbecho
herbáceo, inoculado con T. harzianum (cepa A-34) (109
conidios por hoyo) tres días antes de la siembra, así como
inoculación de las semillas y al follaje (tres aplicaciones),
cuando comenzó la emisión de las primeras guías,
floración y fructificación (concentración de 108 conidios/
mL-1 y dosis de 1011 conidios/ha-1). Se realizó aplicación
de azoxistrobina (Amistar SC 25 100 g i.a. ha-1) en época
de primavera y en época de invierno de azoxistrobina y
mancozeb (Mancozeb PH 80 a 2,4 kg i.a. ha-1) cuando la
enfermedad alcanzó el umbral económico. Para evitar el
efecto de la azoxistrobina y mancozeb sobre T. harzianum,
se realizaron las aplicaciones 10 días antes o después del
antagonista (Morera, 2009).
En el tratamiento testigo, se procedió como recomienda
el Minag (2006): dos años el suelo en barbecho herbáceo
y aplicaciones con cobre (Cuproflow SC 37 a 2 kg i.a.
ha-1), alternando con mancozeb (Mancozeb 2,4 kg i.a.
ha-1) cuando el cultivo alcanzó 5% de intensidad de la
enfermedad.
REsultados y dIscusIón
duración de la eficacia de algunos fungicidas en
el control del tizón gomoso del tallo en sandía
En el Cuadro 1 se observa en el año 2004, que a los cinco
días de realizadas las aplicaciones de los fungicidas en época
de primavera no se detectaron diferencias significativas
entre los tratamientos con azoxistrobina, tiofanato metilo
y clorotalonilo, pero estos difirieron del mancozeb, que lo
hizo a su vez con el testigo. Sin embargo, en el año 2005
azoxistrobina y tiofanato metilo tuvieron diferencias con los
restantes fungicidas.

92
Pérez et al., 2013. Tizón gomoso del tallo
El tratamiento donde se aplicó azoxistrobina mostró
mayor eficacia, pero sin diferencias significativas con
tiofanato metilo en primavera para ambos años. No
obstante, cuando se aplicó azoxistrobina, la señal
para una nueva aplicación de control químico fue
más tardía que con el resto de los fungicidas. Estos
resultados tienen correspondencia con los notificados
por Barlett et al. (2002), con azoxistrobina para el
control de D. bryoniae y con los de Hopkins (2002),
que señaló que las aplicaciones de fungicidas
convencionales como azoxistrobina y clorotalonilo
disminuyeron en este mismo orden los porcentajes de
incidencia del tizón gomoso del tallo en el cultivo de
melón. Concuerdan además, con los de Rizzo et al. (2003), quienes demostraron en Brasil la efectividad
de las aplicaciones de azoxistrobina, alternando con
clorotalonilo y acibenzolar-S-metilo en el control de D. bryoniae en condiciones controladas.
En la campaña de invierno el ciclo de desarrollo
de la sandía es más largo, por lo que las plantas
están expuestas más tiempo al ataque del patógeno,
Cuadro 1. Acción residual de fungicidas frente al tizón gomoso del tallo en época de primavera.
Tratamientos
Intensidad de la enfermedad (Medias transformadas)
2004 2005
5días
12 días
18días
23días
5días
12 días
18días
23días
Azoxistrobina 0,88 a 1,18 a 1,76 a 1,95a 0,88a 1,28a 1,84a 1,99a
Tiofanato metilo 0,88 a 1,28 a 1,81ab 2,03ab 0,88a 1,43a 1,86ab 2,03a
Clorotalonilo 0,88 a 1,58 b 1,88b 2,07b 1,08b 1,69b 1,96bc 2,11b
Mancozeb 1,48 b 1,84 c 1,99c 2,09b 1,58c 1,97c 2,05c 2,13b
Testigo 1,73 c 1,91 c 2,01c 2,09b 1,83d 1,91c 2,07c 2,16b
CV 22,91 20,51 6,48 3,85 23,28 17,84 6,61 4,06
ESx 0,077 0,066 0,024 0,016 0,08 0,059 0,026 0,017
Medias con letras diferentes en la columna difieren significativamente (p< 0,05).
pero los bajos porcentajes de humedad relativa
propician la disminución de la incidencia y severidad
de esta enfermedad. Sin embargo, el aumento de la
temperatura en época de invierno ha inducido a un
incremento considerable del índice de intensidad de
la enfermedad en los últimos 15 años, por lo que fue
necesario la evaluación de fungicidas químicos en esta
época.
En invierno los productos de mayor eficacia son:
azoxistrobina y tiofanato metilo (Cuadro 2), con
azoxistrobina el índice de intensidad para una nueva
aplicación del control químico se alcanzó a los 24
días en el año 2004. Clorotalonilo no tiene diferencias
significativas con tiofanato metilo, pero presentó índice
de intensidad para la aplicación del control químico
a los 19 días aproximadamente. Bharath et al. (2005)
obtuvo buenos resultados en la disminución de la
severidad de la enfermedad con el empleo de tiofanato
metilo. Por otro lado, los resultados con mancozeb
tienen la misma tendencia que los de primavera.
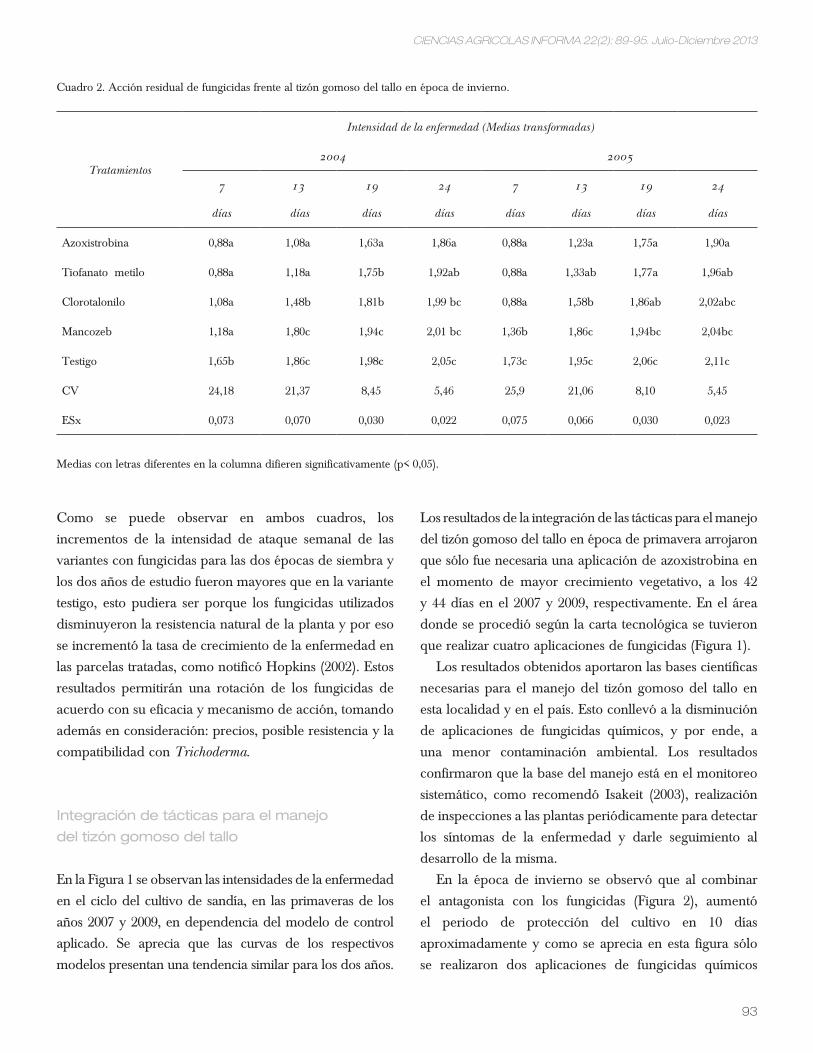
93
CienCias agRiColas infoRma 22(2): 89-95. Julio-Diciembre 2013
Como se puede observar en ambos cuadros, los
incrementos de la intensidad de ataque semanal de las
variantes con fungicidas para las dos épocas de siembra y
los dos años de estudio fueron mayores que en la variante
testigo, esto pudiera ser porque los fungicidas utilizados
disminuyeron la resistencia natural de la planta y por eso
se incrementó la tasa de crecimiento de la enfermedad en
las parcelas tratadas, como notificó Hopkins (2002). Estos
resultados permitirán una rotación de los fungicidas de
acuerdo con su eficacia y mecanismo de acción, tomando
además en consideración: precios, posible resistencia y la
compatibilidad con Trichoderma.
Integración de tácticas para el manejo
del tizón gomoso del tallo
En la Figura 1 se observan las intensidades de la enfermedad
en el ciclo del cultivo de sandía, en las primaveras de los
años 2007 y 2009, en dependencia del modelo de control
aplicado. Se aprecia que las curvas de los respectivos
modelos presentan una tendencia similar para los dos años.
Los resultados de la integración de las tácticas para el manejo
del tizón gomoso del tallo en época de primavera arrojaron
que sólo fue necesaria una aplicación de azoxistrobina en
el momento de mayor crecimiento vegetativo, a los 42
y 44 días en el 2007 y 2009, respectivamente. En el área
donde se procedió según la carta tecnológica se tuvieron
que realizar cuatro aplicaciones de fungicidas (Figura 1).
Los resultados obtenidos aportaron las bases científicas
necesarias para el manejo del tizón gomoso del tallo en
esta localidad y en el país. Esto conllevó a la disminución
de aplicaciones de fungicidas químicos, y por ende, a
una menor contaminación ambiental. Los resultados
confirmaron que la base del manejo está en el monitoreo
sistemático, como recomendó Isakeit (2003), realización
de inspecciones a las plantas periódicamente para detectar
los síntomas de la enfermedad y darle seguimiento al
desarrollo de la misma.
En la época de invierno se observó que al combinar
el antagonista con los fungicidas (Figura 2), aumentó
el periodo de protección del cultivo en 10 días
aproximadamente y como se aprecia en esta figura sólo
se realizaron dos aplicaciones de fungicidas químicos
Tratamientos
Intensidad de la enfermedad (Medias transformadas)
2004 2005
7
días
13
días
19
días
24
días
7
días
13
días
19
días
24
días
Azoxistrobina 0,88a 1,08a 1,63a 1,86a 0,88a 1,23a 1,75a 1,90a
Tiofanato metilo 0,88a 1,18a 1,75b 1,92ab 0,88a 1,33ab 1,77a 1,96ab
Clorotalonilo 1,08a 1,48b 1,81b 1,99 bc 0,88a 1,58b 1,86ab 2,02abc
Mancozeb 1,18a 1,80c 1,94c 2,01 bc 1,36b 1,86c 1,94bc 2,04bc
Testigo 1,65b 1,86c 1,98c 2,05c 1,73c 1,95c 2,06c 2,11c
CV 24,18 21,37 8,45 5,46 25,9 21,06 8,10 5,45
ESx 0,073 0,070 0,030 0,022 0,075 0,066 0,030 0,023
Cuadro 2. Acción residual de fungicidas frente al tizón gomoso del tallo en época de invierno.
Medias con letras diferentes en la columna difieren significativamente (p< 0,05).

94
Pérez et al., 2013. Tizón gomoso del tallo
(azoxistrobina y mancozeb). En el tratamiento sugerido por la carta tecnológica se aplicaron fungicidas en cinco ocasiones
(cobre alternado con mancozeb). Al igual que en primavera de los años 2007 y 2009, las curvas de intensidad de la
enfermedad en época de invierno mantienen una tendencia similar para cada modelo de control.
Como se aprecia en ambas figuras, los picos de infección disminuyen, sobre todo hasta los 50 días, edad del cultivo, lo
que pudiera estar relacionado con la alta tasa de crecimiento diario de la planta de sandía, con un promedio de 4,6 cm.
Eso implica que cuando se aplique una medida de control el agente causal tiende a frenar su avance, pero el cultivo sigue
creciendo, por lo que en la próxima evaluación a los siete días disminuye el porcentaje de área foliar afectada y con ello
el pico de la curva en el tiempo.
Cuando se integraron las tácticas: suelo con cuatro años en barbecho, inoculación al suelo, semillas y aplicación
al follaje con Trichoderma, el tratamiento con azoxistrobina en época de primavera y en invierno con azoxistrobina y
mancozeb, los rendimientos difirieron significativamente del tratamiento regido por la carta tecnológica (Cuadro 3). La
táctica en que se inocula Trichoderma incrementó los rendimientos de primavera en 2,35 t ha-1 y en 3,11 t ha-1 en época
de invierno, con respecto al tratamiento que siguió la carta tecnológica descrita para la Isla de la Juventud. Esto puede
ser por efecto de la compensación de daños o por un posible efecto estimulante que pudo ejercer Trichoderma (Inbar
et al., 1994; Martínez et al., 2013).
Figura 1. Comparación de modelos de control tizón gomoso del tallo en sandía durante la época de primavera en el año 2007 y 2009.
Figura 2. Comparación de modelos de control tizón gomoso del tallo en sandía durante la época de invierno en el año 2007 y 2009.
Cuadro 3. Influencia de las tácticas de manejo del tizón gomoso del tallo sobre el rendimiento de la sandía.
Modelos de control
Rendimientos ( t ha-1)
Primavera Invierno
2007 2009 2007 2009
Integración de tácticas 17,14 a 16,78 a 23,11 a 22,55 a
Carta tecnológica 14,90 b 14,33 b 20,05 b 19,39 b
CV 7,42 8,32 7,50 7,7
ESx 0,37 0,40 0,51 0,5
Medias con letras diferentes en la columna difieren significativamente (p< 0,05).

95
CienCias agRiColas infoRma 22(2): 89-95. Julio-Diciembre 2013
conclusIonEs
La mayor eficacia en el control del tizón gomoso del
tallo en sandía se obtuvo con azoxistrobina, tiofanato
de metilo y clorotalonilo, con acción residual de hasta
18 días en ambas estaciones. Cuando se integraron las
tácticas: siembras realizadas en suelo con cuatro años de
barbecho herbáceo, con inoculaciones al suelo, semillas y
aplicaciones al follaje con la cepa A- 34 de T. harzianum,
y la aplicación de azoxistrobina, disminuyó la intensidad
de la enfermedad e incrementó los rendimientos en sandía
en época de primavera en 2,35 t ha-1 y en 3,11 t ha-1 en
invierno.
REFEREncIas bIblIoGRÁFIcas
1. Barlett, D., J. Clogh, J. Godwin, A. Hall and M. Hamer. 2002. The strobilurin fungicides. Rev. Pest Manag. Sci. 58: 649-662.
2. Bharath, B. G., S. Lokesh, V. R. Rai, H. S. Prakash, B. Yashovarma and H. S. Shetty. 2005. Role of foliar spray in the infection and management of fungal diseases of watermelon [Citrullus lanatus (Thunb.) Matsum and Nakai]. Rev. World J. Agric. Sci. 1: 105-108.
3. Ferguson, G., R. Cerkauskas and S. Khosla. 2009. Gummy stem blight of greenhouse cucumber. Ministry of Agriculture, Food and Rural Affairs. Ontario, Canada (consultado en www.ontario.ca/omafra, fecha de consulta 29 de septiembre 2010).
4. Figueiredo, M. B., R. M. G. Cardoso and J. Abrahao. 1966. Perdas causadas pelo fungo Mycosphaerella melonis (Pass.) Chiu y J. C. Walker, em aboboreira de moita. O Biológico. 32: 203-205.
5. Hernández, A., J. Pérez, M. D. Bosch y L. Rivero. 1999. Nueva versión de la clasificación genética de suelos de Cuba. Inst. Sue-los, agrinfor. Cuba. 64 p.
6. Hopkins, D. L. 2002. Control of gummy stem blight of watermelon with plant defense activators combined with fungicides. Proc. Fla. State Hort. Soc. 115: 183-186.
7. Inbar, J., M. Abramsky, D. Cohen and I. Chet. 1994. Plant growth enhancement and disease control by Trichoderma harzianum in vegetable seedlings grown under commercial conditions. Europ. J. Plant Pathol. 100: 337-346.
8. Isakeit, T. 2003. Resistance of the gummy stem blight fungus to strobilurin fungicides: A potential problem for watermelon production in Texas. Veg. Product. 13: 12.
9. Keinath, A. P. 2001. Effect of fungicide applications scheduled to control gummy stem blight on yield and quality of watermelon fruit. Rev. Plant Dis. 85: 53-58.
10. Keinath, A. P. and I. A. Duthie. 1998. Yield and quality reductions in watermelon due to anthracnose, gummy stem blight and black rot. Recent Res. Develop. Plant Pathol. Research Signpost. 2: 77-90.
11. Keinath, A. P. and V. B. DuBose. 2009. Evaluation of triazoles and other fungicides for control of gummy stem blight on watermelon. Rev. Plant Dis. Manag. Reports. 3: 34.
12. Keinath, A. P., W. H. May and V. B. Dubose. 1997. Comparison of eight fungicide intervals to control gummy stem blight on wa-termelon. Fungicide & Nematicide Tests. 52: 195.
13. Lerch, G. 1977. La experimentación agrícola en las ciencias biológicas y agrícolas. Científico-Técnico. Cuba. 452 p.
14. Martínez, B. C., J. G. Pérez, D. Infante, Y. Duarte y M. Moreno. 2012. Evaluación in vitro de aislamientos de Trichoderma spp. frente a Didymella bryoniae (Fuckel) Rehm. Rev. Protección Veg. 27: 3.
15. Martínez, B. C., D. Infante y Y. Reyes. 2013. Trichoderma spp. y su papel en el control de plagas en los cultivos. Rev. Protección Veg. 28: 1.
16. McKiney, H. H. 1923. Influence of soil, temperature and moisture on infection of wheat seedlings by Helmintosporium sativum: Rev. J. Agric. Res. 26: 195-217.
17. Ministerio de la Agricultura (Minag). 1998. Carta tecnológica del cultivo de melón. Grupo territorial de hortaliza. Isla de la Juventud, Cuba. 12 p.
18. Ministerio de la Agricultura (Minag). 2006. Carta tecnológica del cultivo de melón. Delegación Territorial de la Agricultura (Ministerio de la Agricultura). Isla de la Juventud, Cuba. 8 p.
19. Morera, J. 2009. TricoFung. Producto biológico a base de Trichoderma sp. Dossier informativo (consultado en www.morera.com, fecha de consulta 2 de febrero de 2009).
20. Pérez, J. G., B. C. Martínez, E. A. Alfaro y A. Cervantes. 2003. Elaboración de una escala para la evaluación del tizón gomoso del tallo en el cultivo de sandía, bajo condiciones de la Isla de la Juventud, Cuba. En: VIII Encuentro de Agricultura Orgánica y Sostenible. Isla de la Juventud, Cuba. 7 p.
21. Pérez, J. G., B. C. Martínez, B. C. Covas y H. García. 2012a. Sintomatología e identificación del agente causal del tizón gomoso del tallo en sandía (Citrullus lanatus (Thunb.) Matsum and Nakai) en la Isla de la Juventud. Rev. Protección Veg. 27: 3.
22. Pérez, J. G., B. C. Martínez, S. V. Guadarrama, A. Ll. Cervantes y C. A. Esquivel. 2012b. Tácticas agronómicas para la regulación del tizón gomoso del tallo [Didymella bryoniae (Fuckel) Rehm] en el cultivo de sandía [Citrullus lanatus (Thunb.) Matsum and Nakai]. Ciencias Agrícolas Informa. 21: 106-114.
23. Rizzo, A. A. N., M. R. Ferreira and L. T. Braz. 2003. Ação de acibenzolar-S-methyl (BTH) isolado e em combinação com fungicidas no controle do cancro da haste em melão rendilhado. Rev. Hort. Bras. 21: 238-240.
24. Santos, G. R., A. C. Cafe´-Filho and L. M. F. Saboya. 2005. Controle químico do crestamento gomoso do caule em melancia. Rev. Fitopatol. Bras. 30: 155-163.
25. Sikora, E. J. 1994. Gummy stem blight of cucurbits. Plant Disease. ANR-871. Oct. (consultado en http: \\www.desaveal.ual.es\sifa\ayudas\tmelon.htm, fecha de consulta 31 de mayo 2004).
26. Sudisha, J., K. T. Vasanth, S. R. Niranjana and H. S. Shekar. 2004. First report of gummy stem blight caused by Didymella bryoniae on muskmelon (Cucumis melo) in India. New Dis. Rep. (7 y 9).


97
Caracterización molecular de 20 clones de Hevea brasiliensis del jardín de propagación del inifap en Tabasco
molecular characterizacion in 20 clones of Hevea brasiliensis in inifap propagation gardens at tabasco state
José Luis Hernández-De la Cruz,1 Julia María Lesher-Gordillo,1* Armando Romo-López,1 José Miguel Hernández-Cruz2
CIENCIAS AGRICOLAS INFORMA 201322(2): 97-105
Recibido 15 de febrero de 2013. Aceptado 30 de diciembre de 2013
REsumEn
Hevea brasiliensis o árbol de hule es cultivado en los países tropicales y ecuatoriales, es la fuente principal para la producción de hule natural, el cual tiene más de 4 000 usos lo que lo hace muy importante en la industria. En este trabajo se propuso como objetivos, conocer la diversidad genética entre 20 tipos de clones de H. brasiliensis, propagados en el jardín del Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias en Tabasco, y determinar el polimorfismo presente entre estos individuos. La variabilidad genética entre los genotipos se estimó con la distancia genética Dice y upgMa. Los cuatro microsatélites empleados detectaron un total de 55 bandas polimórficas. Los resultados obtenidos del aMova mostraron que la mayor diversidad genética se encontró entre clones (82%), con un P = 0,01, mientras que la variación intra clones fue de 18% de la variación total, indicando una baja variabilidad genética.
Palabras clave: Diversidad genética, Hevea brasiliensis, microsatélites, polimorfismo.
abstRact
Hevea brasiliensis or rubber tree, cultivated in ecuatorial and tropical countries, is the primary plant used in natural rubber production, it has more tan 4 000 uses; thus is very important for diverses industries. The aim of this research was to determine the genetic diversity in 20 inbred lines of H. brasiliensis, this lines are grow in multiplication gardens in Tabasco State. Also we get the polymorphism among this lines. Genetic variability among genotypes was estimated with 4 select polymorphic SSRs, by way of Dice Genetic distance and upgMa clustering. We observed 55 polymorphic bands. In the aMova analysis the highest genetic diversity among lines was 82% and significative P = 0,01. The variability intralines was 18%, suggesting a low genetic variability.
Key words: Genetic diversity, Hevea brasiliensis, microsatellites, polymorphism.
1 Universidad Juárez Autónoma de Tabasco, México.2 Campo Experimental-Huimanguillo, Tabasco, México.
*Autor para correspondencia: [email protected]

98
Hernández et al., 2013. Caracterización de Hevea
IntRoduccIón
El árbol del hule es una planta perenne que pertenece
al género Hevea de la familia Euphorbiaceae. El
género Hevea está compuesto por diez especies, todas
originarias de las cuencas bajas del Río Amazonas en
Brasil (Schultes, 1990). Su domesticación comenzó en
el siglo XIX y se empezó a distribuir en otros países
en 1876, por Wickman (Omokhafe y Álika, 2003). La
colección de Wickman que llegó al sureste de Asia
y África, fue el principal germoplasma que se usó
para el mejoramiento genético de Hevea en forma de
clon. H. brasiliensis es la especie más importante en
la producción de látex a nivel comercial. El caucho
natural es altamente valorado por sus propiedades
físico-químicas, ya que no tiene sustituto sintético
comparable debido a su elasticidad y resistencia a altas
temperaturas (Lynen, 1969).
El árbol de hule se propaga vegetativamente en
forma comercial por injerto (clon). Para la producción
masiva de clones de hule se requiere contar con jardines
de propagación, como fuente para la obtención del
material clonal. La producción de clones, con fines
agrícolas, enfrenta el desafío de implementar sistemas
que le permitan garantizar la calidad de sus productos.
La mejora genética ha permitido disponer de clones
con características productivas adecuadas a cada área
geográfica. Caracterizar molecularmente a los clones
de Hevea que existen en el jardín de propagación
del Instituto Nacional de Investigaciones Forestales,
Agrícolas y Pecuarias (inifap) permitirá constatar si
corresponden a un mismo genotipo y también para la
contribución a la certificación del material comercial
de H. brasiliensis de mayor interés para la entidad. De
los 35 clones de H. brasiliensis que se propagan en el
jardín, sólo se seleccionaron 20 considerados los más
importantes para la producción del látex en Tabasco.
Los avances en biología molecular han permitido
obtener mejores estimaciones de la diversidad
genética, cuya importancia radica en el desarrollo
de estrategias de conservación y mejoramiento
genético en especies de interés comercial. Las
técnicas empleadas en biología molecular tales como
marcadores moleculares han permitido conocer,
caracterizar y estimar la diversidad genética existente
(inter e intrapoblacional). Los marcadores moleculares
ssr generados en Hevea han sido reportados en
estudios de análisis de diversidad genética de H. brasiliensis en materiales cultivados y/o silvestres (Saha
et al., 2005, Saha et al., 2007; Nakkanong et al., 2008;
Gouvêa et al., 2010), así como en la identificación de
loci genéticos implicados en la expresión de rasgos
agronómicos (Lespinasse et al., 2000, Le Guen et al., 2007). Asimismo, el uso de los microsatélites ha
permitido la construcción de bibliotecas genómicas
para los clones de hule GT1 y RRII105 (Roy et al. 2004). La aplicación de los microsatélites, por lo tanto,
ofrece una nueva herramienta de apoyo al programa
de mejoramiento, certificación y conservación genética
de H. brasiliensis.Los objetivos de este trabajo fueron establecer la
identidad de los tipos de clones de Hevea con los que
se cuenta en el jardín de propagación del estado de
Tabasco, y determinar el polimorfismo presente entre las
poblaciones de los mismos.
matERIalEs y métodos
Se realizó el muestreo en el jardín de propagación de
clones de hule en Tabasco, que se encuentra ubicado en
la Ranchería el Chichonal 1ra sección, localizado en el
municipio de Jalapa (17o 47‘ 43‘‘ L.N. y al 92o 40‘ 49‘‘ L.W.).
Se colectaron un total de 512 muestras correspondientes a
los 20 tipos de clones (Cuadro 1).

99
CienCias agRiColas infoRma 22(2): 97-105. Julio-Diciembre 2013
Cuadro 1. Clon, número de plantas muestreadas y país de origen de los clones de H. brasiliensis colectados en el jardín de propagación del Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (inifap) Tabasco.
Clon Número de plantas muestreadas País de origen
1. PB330 28 Malasia
2. PB260 34 Malasia
3. PB312 22 Malasia
4. RRICC100 32 Sri Lanka
5. PB340 32 Malasia
6. RRIM600 21 Malasia
7. BPM24 23 Indonesia
8. RRIM901 22 Malasia
9. RRIM712 38 Malasia
10. RRIM728 22 Malasia
11. IRCA109 36 Costa de Marfil
12. PB5/51 20 Malasia
13. RRIM921 24 Malasia
14. IAN710 28 Brasil
15. IAN873 24 Brasil
16. IAN754 24 Brasil
17. PB235 20 Malasia
18. IRCA209 20 Costa de Marfil
19. RRIM710 20 Malasia
20. RRIM711 22 Malasia
TOTAL 512
Para la extracción del dna genómico hoja, se empleó el
protocolo descrito por Doyle y Doyle, (1990), modificado
por Bekesiova et al. (1999). Para la amplificación del
dna genómico se utilizaron cuatro iniciadores de tipo
microsatélite específicos para H. brasiliensis (hmac4,
hmac5, hmct1 y hmct5) (sigMa) diseñados por Saha et al. (2005) (Cuadro 2).

100
Hernández et al., 2013. Caracterización de Hevea
Cuadro 2. Nombre y secuencia de los iniciadores utilizados para la amplificación del dna de H. brasiliensis reportados por Saha et al. (2005).
Iniciadores Secuencia de los iniciadores ( 5´ 3´ )
hmac4 5´ GTTTTCCTCCGCAGACTCAG 3´ 5´ ATCCACCAAATAAGGCATGA 3´
hmac5 5´ TCGGTTGGTTTACCATGACA 3´ 5´ ACATCACATGAGTGTATCTCTGATCTC 3´
hmct1 5´ AACCAGAAGGGTGTCATGCT 3´ 5´ GGAATCCCATGACAATCCAC 3´
hmct5 5´ ATGTATGTGTGCGCAGGAAG 3´ 5´ CTGTAGTCATGGCAGCAGGA 3´
Las reacciones de amplificación se llevaron a cabo
con 20 μL de volumen final el cual contenía 11,55 μL
de agua desionizada estéril, 1 μL de dna genómico
(aproximadamente 20 ng), 2,3 μL de MgCl₂, 2,0 μL
de buffer de reacción al 10X, 1 μL de cada uno de los
iniciadores sentido y antisentido, 1 μL de dNTPs y 0,15
μL de polimerasa AmpliTaq Gold. El programa de pCr
para llevar a cabo la amplificación fue: desnaturalización
con un ciclo inicial de 95 °C por 5 min, seguido por 7
ciclos a 94 °C por 30 s, 63 °C por 1 min y 72 °C por 1 min.
Posteriormente estas temperaturas fueron seguidas por 23
ciclos a 94 °C por 30 s, 56 °C por 1 min, 72 °C por 1 min,
y una temperatura final de elongación a 72 °C por 10 min.
El dna fue amplificado en un equipo MyCycler (Biorad).
El producto de pCr fue sometido a electroforesis en gel de
agarosa 2,0%. Los geles fueron fotografiados, tomando las
bandas intensas como presentes [1] o como ausentes [0]
en cada genotipo con el programa UVIgeltec, creando así
una matriz de valores binarios que se analizó a través de
la aplicación del software NTSys 2,0. Posteriormente los
datos se ejecutaron con el archivo NTS. Con el objetivo
de construir un dendrograma y determinar el grado de
similitud que hay entre el material genético analizado se
utilizó el índice de Dice (Nei y Li, 1979). Para conocer el
porcentaje de polimorfismo se empleó la siguiente fórmula
P= npj/ntotal (Donde P= proporción de loci polimórficos,
npj= número de loci polimórfico y ntotal= número total de
loci), y para estimar la variación molecular entre clones
se utilizó el método de aMova con el programa de
GenAlex 6.41.
REsultados y dIscusIón
Los cuatro iniciadores evaluados en el jardín de
propagación de clones de H. brasiliensis, mostraron
polimorfismo. En conjunto generaron un total de 55
regiones polimórficas (Cuadro 3).
Cuadro 3. Porcentaje de polimorfismo obtenido en los 20 tipos de clones de H. brasiliensis y tamaño de bandas observadas por cada iniciador de microsatélites.
Iniciador Bandas polimórficas
Bandas monomórficas
Porcentaje de polimorfismo
Tamaño de la banda
hmac4 2 0 100 280-300 pb
hmac5 19 2 90.4 246-300 pb
hmct1 18 1 94.7 180-260 pb
hmct5 16 2 88.8 160-227 pb
Con este trabajo y los resultados reportados por
Hernández (2009) y Saha et al. (2005), los cuatro
iniciadores que fueron utilizados para identificar los
clones de H. brasiliensis, demostraron ser eficientes
para conocer los genotipos específicos de los clones. Estos resultados permitieron corroborar que entre
mayor sea el número de individuos analizados sobre
sus genotipos (Cuadro 4), se identificará un alto nivel de
polimorfismo en una población de una misma especie
(Koskinen et al. 2004; Nybon, 2004). Sin embargo,
cuando las posibilidades de hacer este tipo de estudios
están limitadas por los recursos, es muy importante
poder seleccionar los iniciadores más eficientes para la
determinación de la variabilidad genética.

101
CienCias agRiColas infoRma 22(2): 97-105. Julio-Diciembre 2013
Cuadro 4. Estudios citados, número de plantas analizadas en sus genotipos y número de bandas polimórficas reportados con los cuatro iniciadores.
Estudios citados Número de plantas analizadas
Total de bandas polimórficas
Saha et al. (2005) 27 19
Hernández (2009) 154 45
Los resultados obtenidos del aMova señalan que
la mayor diversidad genética encontrada fue entre
clones (82%), con P = 0,01. La variación dentro
(intra clones) sólo cuantificó 18% de la variación
total. El haber encontrado mayor porcentaje de
variación genética entre clones (Cuadro 5), puede ser
resultado del origen geográfico del que proceden los
clones evaluados en nuestro estudio (Cuadro 1). Sin
embargo, estos resultados difieren de lo reportado por
Lam et al. (2009), quienes analizaron 59 accesiones
procedentes de 13 distritos de los estados de Mato
Groso, Rondonia y Acre localizadas en el país de
Brasil, y encontraron que la variación interpoblacional
representó sólo 14,1% de la varianza genética total,
mientras que la variación intrapoblacional representó
85,9%. Los autores argumentan que puede haber un
cierto flujo de genes entre los distritos, posiblemente
debido al cruzamiento de la especie como un modo
de reproducción y dispersión de semillas por una
red de ríos en la cuenca del Amazonas y esto explica
porqué las accesiones silvestres son más polimórficas
que los clones cultivados. Estos resultados coinciden
con lo reportado por Lekawipat et al. (2003), quienes
evaluaron 12 marcadores de microsatélites en 108
accesiones de Hevea (40 cultivados y 68 silvestres),
siendo en los cultivados donde se detectó el menor
polimorfismo.
Cuadro 5. Resultados del aMova para la variación intra clonal de H. brasiliensis.
Fuentes de variación
Gl Suma de Cuadrados
CuadradosMedios
Var. Est.
% Total
Entre clones 19 527,168 27,746 1,077 82%
Intra clones 492 118,211 0,240 0,240 18%
Total 511 645,379 1,075 100%
Gl= Grados de libertad, Var. Est.= Varianza estimada, p = 0,01
En el dendrograma obtenido (Figura 1) se observaron
nueve agrupaciones; los trece genotipos de Malasia
(PB235, PB330, PB312, RRIM711, RRIM901, PB5/51,
PB260, RRIM710, RRIM728, RRIM600, PB340, RRIM712
e RRIM921) se distribuyeron en seis grupos (1, 2, 3,
4, 6, 7). Los genotipos de Costa de Marfil (IRCA109 e
IRCA209) estuvieron en los grupos 4 y 5. Por otra parte,
los genotipos de Brasil (IAN710, IAN873 e IAN 754)
fueron distribuidos en tres grupos (6, 7 y 9), mientras que
el genotipo de Indonesia (BPM24) y el genotipo de Sri
Lanka (RRICC100) se distribuyeron en el grupo 1 y 8. Los
clones IAN754 del grupo 9 se excluyó de los ocho grupos,
lo que significa que es baja la información genética que
comparten con los otros 19 tipos de clones.
En el dendrograma se observó un total de 24 individuos
que son diferentes en su estructura genética dentro de
todas las 20 poblaciones de clones de H. brasiliensis. Esto
significa que la variación genética de H. brasiliensis en los
jardines de propagación de Tabasco es alta, aunque todos
proceden de un mismo germoplasma (línea Wickman),
debido a que se ha enriquecido la constitución genética
de esta especie mediante cruzas con clones mejorados
que proceden de África, Indonesia, Sri Lanka y Brasil;
haciendo a estas poblaciones con mayor variabilidad. Esta
variación tiene ventajas ya que hace que las plantaciones
de H. brasiliensis sean más resistentes a las plagas,
condiciones ambientales y enfermedades.

102
Hernández et al., 2013. Caracterización de Hevea
Las muestras analizadas de los clones IRCA209, al
analizar el dendrograma se observa que se distribuyen
en dos grupos y uno de los cuales está más cercano a
RIMM728 con una distancia genética de 0,44 y el otro
grupo más cercano a RIMM600 con una diferencia
genética de 0,57 por lo que se considera que un
grupo de las muestras de IRCA209 no corresponden
a este clon. Del grupo 5 se observaron las diferencias
genéticas dentro del grupo de los clones IRCA109 en
donde se distribuyen en tres grupos: los IRCA109**
e IRCA109***, con una diferencia genética de 0,70.
También se presentó una separación marcada entre
ambos grupos con los grupos IRCA109* que osciló
entre 0,47 de distancia genética. En las muestras
IAN710 se distribuyeron en dos grupos: en el grupo
6 se observa los IAN710*- RIMM921 con un índice
de diferencia genética de 0,45. En el grupo 7 se puede
observar que muestras del clon IAN710** e RIM712
tienen una distancia genética de 0,33. Asimismo un
grupo de muestras de IAN710 no corresponden al clon
indicado.
Los clones IRCA109, IAN710 e IRCA209 presentaron
dentro de su misma población diferencias en sus
genotipos y por lo tanto no presentan una uniformidad
en su información genética (Figura 2 y 3). Este estudio
refuerza lo encontrado por Medina et al. (2007),
quienes emplearon la técnica de aflp para determinar
el nivel de polimorfismo que existe dentro de la
población de clones IAN710 y IAN873, y encontraron
que entre árboles de un mismo clon (IAN873) existió
un porcentaje del 15,1, mientras que en los árboles del
clon IAN710 se encontró un porcentaje del 36,7. Los
resultados han permitido corroborar que la variación
observada entre árboles no es consecuencia de la
técnica, sino de un material plantado en los jardines
clonales, el cual no es homogéneo.
Figura 1. Dendrograma realizado a partir de las distancias genéticas de los 20 tipos de clones de hule utilizando el índice de Nei.
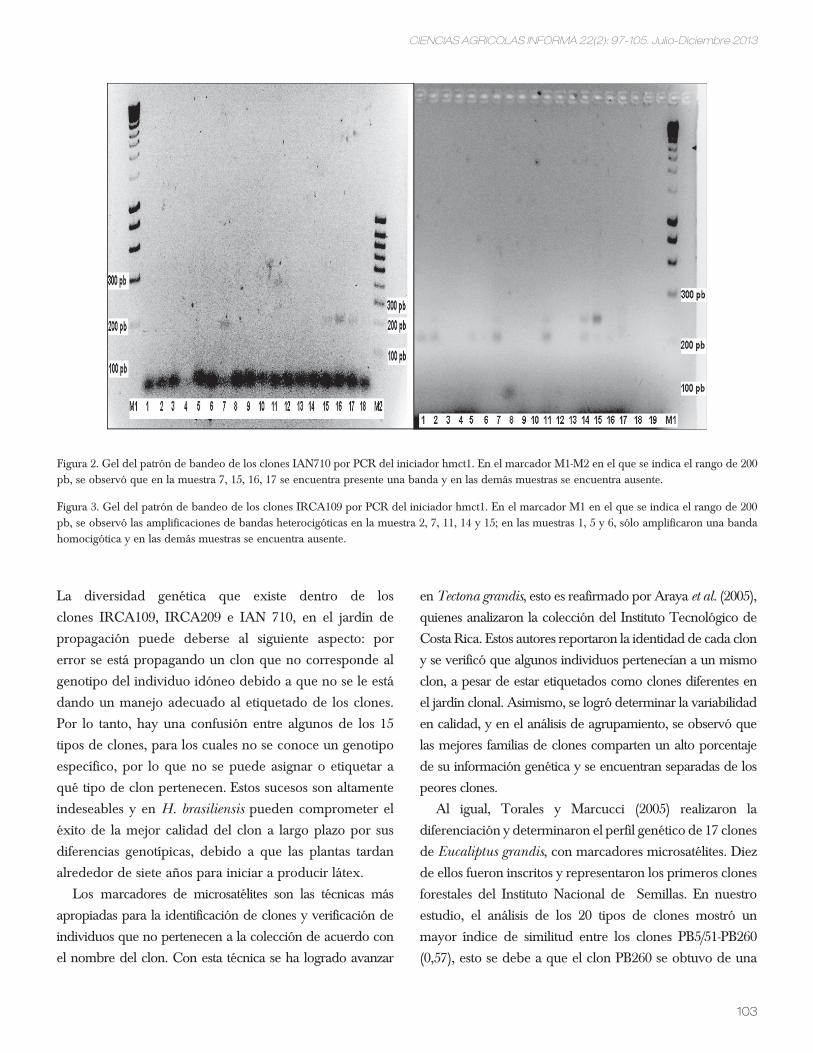
103
CienCias agRiColas infoRma 22(2): 97-105. Julio-Diciembre 2013
La diversidad genética que existe dentro de los
clones IRCA109, IRCA209 e IAN 710, en el jardín de
propagación puede deberse al siguiente aspecto: por
error se está propagando un clon que no corresponde al
genotipo del individuo idóneo debido a que no se le está
dando un manejo adecuado al etiquetado de los clones.
Por lo tanto, hay una confusión entre algunos de los 15
tipos de clones, para los cuales no se conoce un genotipo
específico, por lo que no se puede asignar o etiquetar a
qué tipo de clon pertenecen. Estos sucesos son altamente
indeseables y en H. brasiliensis pueden comprometer el
éxito de la mejor calidad del clon a largo plazo por sus
diferencias genotípicas, debido a que las plantas tardan
alrededor de siete años para iniciar a producir látex.
Los marcadores de microsatélites son las técnicas más
apropiadas para la identificación de clones y verificación de
individuos que no pertenecen a la colección de acuerdo con
el nombre del clon. Con esta técnica se ha logrado avanzar
en Tectona grandis, esto es reafirmado por Araya et al. (2005),
quienes analizaron la colección del Instituto Tecnológico de
Costa Rica. Estos autores reportaron la identidad de cada clon
y se verificó que algunos individuos pertenecían a un mismo
clon, a pesar de estar etiquetados como clones diferentes en
el jardín clonal. Asimismo, se logró determinar la variabilidad
en calidad, y en el análisis de agrupamiento, se observó que
las mejores familias de clones comparten un alto porcentaje
de su información genética y se encuentran separadas de los
peores clones.
Al igual, Torales y Marcucci (2005) realizaron la
diferenciación y determinaron el perfil genético de 17 clones
de Eucaliptus grandis, con marcadores microsatélites. Diez
de ellos fueron inscritos y representaron los primeros clones
forestales del Instituto Nacional de Semillas. En nuestro
estudio, el análisis de los 20 tipos de clones mostró un
mayor índice de similitud entre los clones PB5/51-PB260
(0,57), esto se debe a que el clon PB260 se obtuvo de una
Figura 2. Gel del patrón de bandeo de los clones IAN710 por PCR del iniciador hmct1. En el marcador M1-M2 en el que se indica el rango de 200 pb, se observó que en la muestra 7, 15, 16, 17 se encuentra presente una banda y en las demás muestras se encuentra ausente.
Figura 3. Gel del patrón de bandeo de los clones IRCA109 por PCR del iniciador hmct1. En el marcador M1 en el que se indica el rango de 200 pb, se observó las amplificaciones de bandas heterocigóticas en la muestra 2, 7, 11, 14 y 15; en las muestras 1, 5 y 6, sólo amplificaron una banda homocigótica y en las demás muestras se encuentra ausente.

104
Hernández et al., 2013. Caracterización de Hevea
cruza entre el clon PB5/51 x PB49, asimismo, los clones
IRCA209** (GT1 x RRIM601)-RRIM600 (Tjir x PB81),
que mostraron un índice de 0,57, son individuos que se
obtuvieron mediante cruzas de clones que proceden de
Malasia, mientras que IRCA109**-IRCA109***, con índice
de 0,70, son clones que proceden del mismo progenitor con
una diferencia genética baja. Nuestros resultados coindicen
con lo reportado por Nakkanong et al. (2008) y Gouvêa
et al. (2010), quienes mostraron una baja similitud y alta
diversidad genética entre clones de H. brasiliensis usando
marcadores de tipo microsatélites.
Los tipos de clones que presentaron un genotipo
homogéneo en el dendrograma fueron BPM24, PB235,
PB330, PB312, RRIM711, RRIM901, PB5/51, PB260,
RRIM710, RRIM728, RRIM600, PB340, IAN873,
RRIM712, RRIM921, RRICC100 e IAN754. La importancia
de conservar la variabilidad de los diferentes tipos de
clones, se debe a que no todos los tipos de clones van a
interaccionar en un ambiente de una misma forma y en
otro se comporten de manera diferente. Mantener el jardín
policlonal atenúa este riesgo, por esta razón es indispensable
conservar los diferentes tipos de clones aunque no sean
los más productivos en cada sitio, si no se está seguro del
comportamiento del clon.
conclusIonEs
Los marcadores de microsatélites permitieron identificar
las diferencias entre los 20 tipos de clones de H. brasiliensis evaluados. Los cuatro microsatélites detectaron un
total de 55 bandas polimórficas entre los clones. En los
clones IRCA109, IAN710 e IRCA209 la población es
heterogénea debido a que se identificaron individuos con
genotipos diferentes.
aGRadEcImIEntos
El financiamiento del presente estudio fue obtenido
gracias a Fundación Produce Tabasco. Para la realización
del trabajo de “Caracterización molecular de los clones
del árbol de hule [Hevea brasiliensis (Muell.) Arg.] de los
jardines de propagación del estado de Tabasco”.
REFEREncIas bIblIoGRÁFIcas
1. Araya, E., O. Murillo, G. Aguilar y O. Rocha. 2005. Uso de marcadores genéticos en silvicultura clonal. Rev. For. Kur. 6: 1-14.
2. Bekesiova, I., J. P. Nap and L. Mlynarova. 1999. Isolation of high quality dna and rna from leaves of the carnivorous plant Drosera rotundifolia. Plant Mol. Biol. Rep. 17: 269-277.
3. Doyle, J. J. and J. L. Doyle. 1990. Isolation of plant dna from fresh tissue. Focus 12: 13-15.
4. Gouvêa, L., L. Benchimol, A. F. Chioratto, M. I. Zucchi and P. S. Gonçalves. 2010. Genetic divergence of rubber tree estimated by multivariate techniques and microsatellite markers. Genet. Mol. Biol. 2: 308-318.
5. Hernández, D. J. L. 2009. Utilización de la técnica de microsatélites para la determinación de la variabilidad genética en las plantaciones de H. brasiliensis (Muell.) Arg. del Estado de Tabasco. Tesis de licenciatura. Universidad Juárez Autónoma de Tabasco. Villahermosa, Tabasco. 50 p.
6. Koskinen, M. T., H. Hirnoven, P. A. Landry and C. Primmer. 2004. The benefits of increasing the number of microsatellites used in genetic population studies: an empirical perspectiva. Hereditas. 141: 61-67.
7. Lam, V. L., T. Thanh, V. T. Quynh and L. Mau. 2009. Genetic diversity of Hevea IRRDBQ´81 collection assessed by rapd markers. Mol. Biotech. 42: 292-298.
8. Le Guen V. D., C. R. Garcia, S. R. Matto, F. Doare´, D. Lespi-nasse and M. Seguin. 2007. Bypassing of a polygenic Microcyclus ulei resistance in rubber tree, analyzed by qtl detection. New Phytol. 173: 335-345.
9. Lekawipat N., K. Teerawatanasuk, M. Rodier-Goud, M. Seguin, A.Vanavichit, T. Toojinda, S.Tragoonrung. 2003. Genetic diversity analysis of wild germplasm and cultivated clones of Hevea brasiliensis Muell. Arg. by using microsatellite markers. J. Rubb. Res. 6: 36-47.
10. Lespinasse. D., M. Rodier-Goud, L. Grivet, A. Leconte, Legnate H, M. Seguin. 2000. A saturated genetic linkage map of rubber tree (Hevea spp.) based on rflp, aflp, microsatellite, and isozyme markers. Theor. Appl. Genet. 100: 127-138.
11. Lynen, F. 1969. Biochemical problems of rubber synthesis. Rubb. Res. Inst. Malaysia. 21: 389–406.
12. Medina C., I. García., M. Caro y F. A. Aristizábal. 2007. Análisis aflp de variación somaclonal en embriones somáticos de Hevea brasiliensis. Rev. Col. Cienc. Quim. Farm. 36: 70-80.
13. Nakkanong K., C. Nualsri and S. Sdoodee. 2008. Analysis of genetic diversity in early introduced clones of rubber tree (Hevea brasiliensis) using rapd and microsatellite markers. Songklanakarin J. Sci. Technol. 30: 553-560.
14. Nei, M. and W. H. Li. 1979. Mathematical model for studying genetic variation in terms of restriction endonucleasas. Proceedings of the National Academy of Sciencies. New York, USA. 76: 5269-5273.

105
CienCias agRiColas infoRma 22(2): 97-105. Julio-Diciembre 2013
15. Nybon, H. 2004. Comparison of different nuclear dna markers for estimating intraspecific genetic diversity in plants. Mol. Ecol. 13: 1143-1155.
16. Omokhafe, K. O. and J. E. Álika. 2003. Phenetic relationship of rubber tree clones. Biol. Plant. 46: 217-222.
17. Roy, B. C., M. A. Nazeer and T. Saha. 2004. Identification of simple sequence repeats in rubber (Hevea brasiliensis). Cur. Sci. 87: 807-811.
18. Saha, T., C. B. Roy, M. Ravindran, K. Binin and M. A. Nazeer. 2007. Allelic diversity revealed through ssr polymorphisms at the locus encoding HMG-CoA reductase in rubber (Hevea brasiliensis). Sil. Gen. 2: 58-65.
19. Saha, T., C. Bindu and M. A. Nazeer. 2005. Microsatellite variability and its use in the characterization of cultivated clones of Hevea brasiliensis. Plant Breed. 124: 86-92.
20. Schultes, R. E. 1990. A brief taxonomic view of the genus Hevea. In: MrrdB. Monograph, Kuala Lumpur, Malaysia. 57 p.
21. Torales, S. y S. Marcucci. 2005. Identificación genética de clones utilizando microsatélites en Eucalyptus grandis. Rev. Idia. 21: 191-194 (consultado en http://www.biblioteca.org.ar/libros/210588.pdf, fecha de consulta en 28 de septiembre del 2010).


107
Caracterización molecular de cuatro variedades de gerbera jamesonii Bolus, mediante microsatélites anclados y rapds
molecular characterization of four varieties of Gerbera jamesonii bolus, through anchored microsatellites and rapds
Amaury Martín Arzate-Fernández,1* José Luis Piña-Escutia,1 Luis Miguel Vázquez-García,2
Adolfo Carrillo-Velázquez1
CIENCIAS AGRICOLAS INFORMA 201322(2): 107-115
Recibido: 2 de mayo de 2013 Aceptado: 18 de septiembre de 2013
REsumEn
El manejo no autorizado de cultivares puede generar confusión debido a los diferentes nombres que reciben, además de ocasionar pérdidas al obtentor vegetal. En el presente trabajo se caracterizaron molecularmente cuatro variedades de Gerbera jamesonii (Blanca, Bicolor, Blanca-Verdosa y Ave María), mediante microsatélites anclados (assr), y de polimorfismo de dna al azar (rapds), usando tejido de hoja y de lígula. Los marcadores assr evaluados en hoja generaron 23 bandas y 73% de polimorfismo, mientras que con el tejido de lígula se generaron 18 bandas y 88% de polimorfismo. En contraste, los marcadores rapds evaluados en hoja, generaron 22 bandas y 63% de polimorfismo, mientras que con el tejido de lígula se generaron 25 bandas y 80% de polimorfismo. Se concluyó que con el uso de ambos marcadores, independientemente del tejido usado, fue posible caracterizar a las cuatro variedades de gerbera y diferenciarlas genéticamente entre sí.
Palabras clave: assr, caracterización molecular, gerbera, polimorfismo, rapds.
abstRact
The unauthorized management of cultivars can cause confusion due to the variation in the names that they received, it also causes some money losses to the plant breeder. In this work four varieties of Gerbera jamessonii (Blanca, Bicolor, Blanca-Verdosa y Ave María) were molecularly characterized through anchored microsatellites (assr), and the polymorphism of dna at random (rapds), using leaf and ligula tissues. The assr markers evaluated in leaf produced 23 bands and 73% of polymorphism, whereas with the ligula tissue 18 bands and 88% of polymorphism were generated. In contrast, the rapds markers evaluated in leaf produced 22 bands and 63% of polymorphism, whereas with the ligula tissue 25 bands and 80% of polymorphism were generated. In conclusion, with both markers, regardless the used tissue, it was possible to characterize the four varieties of gerbera and differentiate them genetically.
Key words: assr, gerbera, molecular characterization, polymorphism, rapds.
1 Facultad de Ciencias Agrícolas, Universidad Autónoma del Estado de México, México. 2 Centro Universitario, Universidad Autónoma del Estado de México, Tenancingo, México.*Autor para correspondencia: [email protected].

108
arzate fernández et al., 2013. Caracterización de gerbera
IntRoduccIón
El género Gerbera pertenece a la familia Asteraceae y
comprende entre 40 y 50 especies, siendo uno de los
géneros más demandados en el mercado de flores (Ozkinis
y Lisiencka, 1990). En México, la producción de Gerbera jamesonii se concentra en el Estado de México donde
es considerada una planta altamente demandada como
flor de corte. Esto ha generado nuevas variedades por
mejoramiento genético, entre las que destacan: Blanca,
Bicolor, Blanca-Verdosa y Ave María, las cuales se han
diversificado entre productores de diferentes estados
de la República Mexicana. Sin embargo, el manejo
no autorizado de variedades puede causar pérdidas
al obtentor vegetal, generando controversias legales.
En este sentido, se considera que las variedades antes
mencionadas pueden tener una constitución genética
similar y que los distintos colores de sus lígulas pueden
ser sólo consecuencia del ambiente donde se cultivan.
Los marcadores morfológicos o fenotípicos han sido
utilizados tradicionalmente para distinguir variedades
de especies vegetales estrechamente relacionados (Piña-
Escutia et al., 2009). Sin embargo, tienen el inconveniente
de que algunas características pueden variar por el efecto
del ambiente. Por su parte, los marcadores de dna son
fenotípicamente neutros, termoestables, presentan mayor
polimorfismo y pueden ser evaluados en cualquier etapa
de desarrollo de la planta (Welsh y McClelland, 1999).
Los marcadores más utilizados por su rapidez, economía
y su capacidad de polimorfismo son los assr (Secuencias
Simples Repetidas Ancladas) y los rapds (Polimorfismo
de dna Amplificado al Azar) (Alzofeifa-Delgado, 2006).
Estos marcadores han sido utilizados para determinar
la huella genética tanto de especies silvestres como de
cultivadas, entre las que destacan: Jacaranda mimosifolia
(Escandón et al., 2005a), Tigridia pavonia (Piña-Escutia
et al., 2009), Piperia yadoni (George et al., 2009),
Chrysanthemum spp. (Miñano et al., 2009), Rosa spp.
(Mohapatra y Rout, 2006), Gerbera spp. (Da Mata et al., 2009; Kwon et al., 2001), Passiflora sp. (Pérez-Almeida et al., 2009), Heliconia spp. (Marouelli et al., 2010), entre
otros.
El objetivo del presente trabajo fue caracterizar cuatro
variedades de G. jamesonii Bolus, para lograr diferenciarlas
genéticamente entre sí, mediante el uso de marcadores
moleculares tipo assr y rapds.
matERIalEs y métodos
material vegetal
Se usaron cuatro variedades de G. jamesonii Bolus:
Blanca, Bicolor, Blanca-Verdosa y Ave María. Dichos
materiales colectados provenían del banco de recursos
fitogenéticos del Centro Universitario Tenancingo de la
uaeM. El estudio se realizó en el Laboratorio de Biología
Molecular Vegetal de la Facultad de Ciencias Agrícolas
de la Universidad Autónoma del Estado de México.
Extracción de dna
Para la extracción del dna genómico, se utilizaron
aproximadamente 100 mg de tejido fresco de hoja y de
lígula, con el método CtaB (Zhou et al., 1999). El dna fue
resuspendido en 50 μL de agua desionizada esterilizada y
almacenado a -20 oC hasta su uso.
amplificación del dna por pcr
Se utilizaron cinco iniciadores de tipo microsatélites
anclados (assr) y cinco de polimorfismo de dna al
azar (rapds) (Cuadro 1). La reacción en cadena de la
polimerasa (pCr) se efectuó en un volumen final de 10 μL
que contenía 7,2 μL de agua desionizada esterilizada, 1
μL de amortiguador 10X PCR con amonio (15 mM), 0,5
μL de MgCl2 (25 mM), 1 μL de dNTPs (10 mM) (aplied
BiosysteMs®), 0,1 μL de Taq dna polimerasa (MerCury
reagents®), 0,1 μL de iniciador (20 μM) (invitrogen®) y
0,1 μL de dna genómico (10 ng μL-1). Las condiciones de
amplificación para los iniciadores assr fueron las reportadas
por Piña-Escutia et al. (2009), y para los iniciadores rapds

109
CienCias agRiColas infoRma 22(2): 107-115. Julio-Diciembre 2013
fueron las reportadas por Debener y Mattiesch (1998).
La amplificación del dna se realizó en un termociclador
de gradiente Mastercycler (eppendorf®) modelo Hamburg
22331. Los productos amplificados fueron separados por
electroforesis en un gel de agarosa Tipo II (sigMa®) a 1%,
al cual se adicionaron 3 μL de bromuro de etidio (sigMa®),
las bandas amplificadas se observaron y fotografiaron en un
transiluminador (uvp®) modelo MP20.
caracterización molecular
Los patrones de bandeo generados con los assr y los rapd
se utilizaron para construir una matriz binaria de datos
(MBd) donde se consideró la presencia de una banda
como “1” y la ausencia de la misma, como “0”, para este
análisis sólo se contemplaron las bandas nítidas. La MBd
fue usada para la caracterización molecular de las cuatro
variedades de G. jamesonii evaluadas.
Para los dos tipos de marcadores, el porcentaje de
polimorfismo (%p), y el número total de bandas (Bt)
fueron los parámetros calculados con el programa
popgene versión 1.32 (Yeh y Boyle, 1999). En ambos
casos, para determinar las distancias genéticas y la
forma de agrupación de las variedades analizadas, se
usó el método upgMa basado en la matriz de distancia
genética (DG) de Nei (1972), a partir de la cual se
construyeron los dendrogramas correspondientes para
cada uno de los análisis.
Cuadro 1. Nombre y secuencia de los iniciadores utilizados en la caracterización molecular de cuatro variedades de G. jamesonii.
Tipo de marcador Nombre del iniciador Secuencia 5´ 3´ Número de bases
**assr
3´ -assr02 (CT)7 ATC 17
3´ -assr15 (CT)7 ATG 17
3´ -assr20 (CT)7 GCA 17
3´ -assr29 (CT)7 GTA 17
3´ -assr35 (CT)7 TGA 17
*rapds
Y24 AACCGCGCTC 10
Y29 TTCGGGCCGT 10
Y37 GTCAAGCGCG 10
Y38 TAACCGCGCC 10
Y41 GCGTCCTGGG 10
*Yamagishi (1995), **Yamagishi et al. (2002).

110
arzate fernández et al., 2013. Caracterización de gerbera
REsultados y dIscusIón
marcadores assr
Tejido de hoja. Los iniciadores assr utilizados en el presente trabajo mostraron un grado de polimorfismo desde 0% con
el iniciador assr29 hasta 100% con el assr02. Se generó un total de 23 bandas, 73% de polimorfismo y un promedio de
4,6 bandas por iniciador (Cuadro 2).
Cuadro 2. Caracterización molecular de cuatro variedades de G. jamesonii, mediante cinco iniciadores assr, usando tejido de hoja y de lígula. Bandas totales (Bt), bandas monomórficas (BM), bandas polimórficas (Bp), porcentaje de polimorfismo (%p).
IniciadorHoja Lígula
bt bm bp % p bt bm bp % p
3´-assr02 5 0 5 100 4 0 4 100
3´-assr15 7 1 6 85 6 1 5 83
3´-assr20 5 2 3 60 5 0 5 100
3´-assr29 1 1 0 0 1 1 0 0
3´-assr35 5 2 3 60 2 0 2 100
TOTAL (promedio) 23 6 17 (73) 18 2 16 (88)
En la Figura 1 se observan los productos de amplificación
y el tamaño aproximado de los fragmentos (pb), obtenidos
con el iniciador assr15, tanto en el tejido de hoja como
en el de lígula, mostrando los patrones de bandeo que
permitieron caracterizar a cada una de las variedades
evaluadas.
En el dendrograma generado por los iniciadores
assr, usando tejido de hoja (Figura 2a), se muestran las
distancias genéticas. Se observó la formación de un solo
grupo. Sin embargo, las variedades que mostraron mayor
similitud genética fueron Blanca y Blanca-Verdosa con
una DG = 0,11, mientras que las más distantes fueron Ave
María y Bicolor con un valor de DG = 0,91.
Tejido de lígula. El uso de los iniciadores assr permitió
generar un total de 18 bandas, 88% de polimorfismo y
un promedio de 3,6 bandas por iniciador. De manera
específica, con el iniciador assr29 no se encontró
polimorfismo, mientras que con los iniciadores assr02,
assr20 y assr35 se observó 100% (Cuadro 2). En el
dendrograma (Figura 2b) se observó nuevamente la
formación de un solo grupo. Las variedades menos
emparentadas genéticamente fueron Blanca y Bicolor,
presentando una DG = 0,50; mientras que las variedades
con mayor similitud genética fueron Blanca-Verdosa
y Ave María, con una DG = 0,12. De acuerdo con
los resultados obtenidos con los iniciadores assr, se
pudo observar un mayor porcentaje de polimorfismo
usando el tejido de lígula (88%), en comparación con
el de hoja (73%) (Cuadro 2). Sin embargo, con ambos
tejidos fue posible la identificación de las cuatro
variedades evaluadas, permitiendo obtener su perfil
molecular. Estos resultados ilustran la factibilidad de
utilizar estos marcadores en el análisis genético de
cultivares estrechamente relacionados. La eficacia de
los assr también ha sido reportada en estudios de otras
especies ornamentales como: Nierembergia lineariefolia

111
CienCias agRiColas infoRma 22(2): 107-115. Julio-Diciembre 2013
Figura 1. Productos amplificados con el iniciador assr15, utilizado en la caracterización molecular de cuatro variedades de G. jamesonii, usando tejido de hoja y de lígula. BCa: Blanca, BiC: Bicolor, B-v: Blanca- Verdosa, aM: Ave María.
Figura 2. Dendrogramas generados a partir de las distancias genéticas obtenidas con cinco iniciadores assr, usando tejido de hoja (a) y tejido de lígula (b) en cuatro variedades de G. jamesonii.

112
arzate fernández et al., 2013. Caracterización de gerbera
(Escandón et al., 2005b), Tacca chantrieri (Zhang et al., 2006) y en Sprekelia formosissima (L.) (Bautista-Puga et al., 2011), entre otros. Pradeep et al. (2002) y Hu et al. (2003) señalan que la efectividad de los iniciadores
assr posiblemente se debe a la secuencia motivo y la de
su ancla, donde una secuencia motivo Ct presenta un
mayor polimorfismo con respecto a los que tienen una
at y son los más abundantes y altamente polimórficos
en el genoma de las plantas, aunque dificultan la
amplificación de bandas específicas; sugiriendo que se
debe a la semicomplementariedad del iniciador en la
etapa de alineación de la pCr (Fang y Rose, 1997).
La secuencia del ancla consiste de uno, dos o tres
nucleótidos los cuales se anclan al final del extremo
3´ o 5´ de un microsatélite (assr), asegurando el
reconocimiento del iniciador con el dna genómico, con
la finalidad de incrementar la distinción de fragmentos
polimórficos (Pradeep et al., 2002; Yamagishi et al., 2002). En el presente estudio, el nivel de distinción
entre las cuatro variedades también dependió del tipo
de tejido y de la secuencia del ancla del iniciador. Así,
usando el tejido de hoja, únicamente con el iniciador
3´-assr02, cuya secuencia es atC, fue posible obtener
100% de polimorfismo. De igual manera, usando el tejido
IniciadorHoja Lígula
bt bm bp % p bt bm bp % p
Y24 2 2 0 0 2 2 0 0
Y29 5 1 4 80 5 0 5 100
Y37 3 3 0 0 3 2 1 33
Y38 6 0 6 100 8 0 8 100
Y41 6 2 4 66 7 1 6 85
TOTAL (promedio) 22 8 14 (63) 25 5 20 (80)
de lígula, se observó 100% de polimorfismo no sólo con el
mismo iniciador 3´-assr02, sino también con el iniciador
3´-assr20, cuya secuencia es gCa y el 3´-assr35, cuya
secuencia es tga (Cuadro 2). Esto sugiere que la técnica
assr puede ser un sistema altamente informativo, rápido,
confiable para la identificación de cultivares, como lo
señala Tapia et al. (2005a).
marcadores rapds
Tejido de hoja. Los iniciadores rapds presentaron un
intervalo de polimorfismo de 0% con Y24 e Y37, hasta 100%
con Y38 (Cuadro 3). Con estos iniciadores se generó un
total de 22 bandas, 63% de polimorfismo, y un promedio
de 3,6 bandas por iniciador.
En la Figura 3 se observan los patrones de bandeo
generados con el iniciador Y38, utilizando tejido de hoja
y de lígula, permitiendo una caracterización molecular
eficaz con ambos tejidos evaluados, y con cada una de las
cuatro variedades de G. jamesonii analizadas.
En el dendrograma generado con el tejido de hoja
(Figura 4a), se observó la formación de un solo grupo.
Cuadro 3. Caracterización molecular de cuatro variedades de G. jamesonii, mediante cinco iniciadores rapds, usando tejido de hoja y de lígula. Bandas totales (Bt), bandas monomórficas (BM), bandas polimórficas (Bp), porcentaje de polimorfismo (%p).
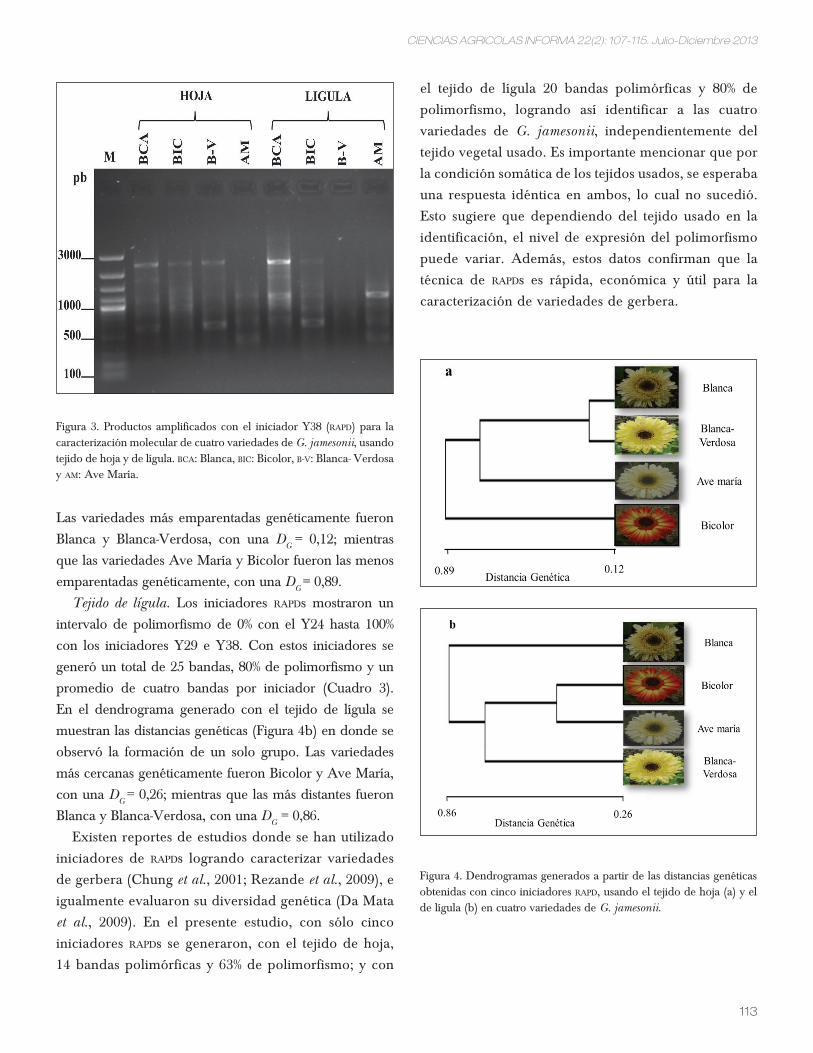
113
CienCias agRiColas infoRma 22(2): 107-115. Julio-Diciembre 2013
Las variedades más emparentadas genéticamente fueron
Blanca y Blanca-Verdosa, con una DG = 0,12; mientras
que las variedades Ave María y Bicolor fueron las menos
emparentadas genéticamente, con una DG = 0,89.
Tejido de lígula. Los iniciadores rapds mostraron un
intervalo de polimorfismo de 0% con el Y24 hasta 100%
con los iniciadores Y29 e Y38. Con estos iniciadores se
generó un total de 25 bandas, 80% de polimorfismo y un
promedio de cuatro bandas por iniciador (Cuadro 3).
En el dendrograma generado con el tejido de lígula se
muestran las distancias genéticas (Figura 4b) en donde se
observó la formación de un solo grupo. Las variedades
más cercanas genéticamente fueron Bicolor y Ave María,
con una DG = 0,26; mientras que las más distantes fueron
Blanca y Blanca-Verdosa, con una DG = 0,86.
Existen reportes de estudios donde se han utilizado
iniciadores de rapds logrando caracterizar variedades
de gerbera (Chung et al., 2001; Rezande et al., 2009), e
igualmente evaluaron su diversidad genética (Da Mata
et al., 2009). En el presente estudio, con sólo cinco
iniciadores rapds se generaron, con el tejido de hoja,
14 bandas polimórficas y 63% de polimorfismo; y con
Figura 3. Productos amplificados con el iniciador Y38 (rapd) para la caracterización molecular de cuatro variedades de G. jamesonii, usando tejido de hoja y de lígula. BCa: Blanca, BiC: Bicolor, B-v: Blanca- Verdosa y aM: Ave María.
el tejido de lígula 20 bandas polimórficas y 80% de
polimorfismo, logrando así identificar a las cuatro
variedades de G. jamesonii, independientemente del
tejido vegetal usado. Es importante mencionar que por
la condición somática de los tejidos usados, se esperaba
una respuesta idéntica en ambos, lo cual no sucedió.
Esto sugiere que dependiendo del tejido usado en la
identificación, el nivel de expresión del polimorfismo
puede variar. Además, estos datos confirman que la
técnica de rapds es rápida, económica y útil para la
caracterización de variedades de gerbera.
Figura 4. Dendrogramas generados a partir de las distancias genéticas obtenidas con cinco iniciadores rapd, usando el tejido de hoja (a) y el de lígula (b) en cuatro variedades de G. jamesonii.

114
arzate fernández et al., 2013. Caracterización de gerbera
conclusIonEs
Con los marcadores assr y rapds, usando tejido de hoja
y lígula, se generaron patrones de bandeo diferentes,
permitiendo caracterizar a las cuatro variedades de gerbera
evaluadas, lo que reveló que todas son genéticamente
diferentes. La eficacia de los assr en la generación de
polimorfismo en ambos tejidos fue mayor que la obtenida
con los rapds.
REFEREncIas bIblIoGRÁFIcas
1. Alzofeifa-Delgado, A. 2006. Uso de los marcadores moleculares en plantas; aplicaciones en frutales del trópico. Agron. Mesoam. 17: 221-242.
2. Bautista-Puga, M. D., L. M. Vázquez, G., H. Leszczynska, B. M. W. Borys y A. M. Arzate-Fernández. 2011. Caracterización del lirio azteca, mediante marcadores morfológicos y moleculares. Agrociencia. 45: 413-422.
3. Chung Y. M., Kim H. A., Kim K. Y., Park S. W., Yi Y. B., Lee J. H. and Chang O. K. 2001. Morphological characteristics and genetic variation of gerbera (Gerbera hybrida hort.). J. Plant Biotechnol. 3: 145–149.
4. Da Mata, L. T., M. I. Segeren, A. Seregen, F. and C. A. Colombo. 2009. Genetic divergence among accessions evaluated by rapd. Sci. Hort. 121: 92-96.
5. Debener, T. and L. Mattiesch. 1998. Effective pairwise combina-tion of long primers for rapds analyses in rose. Plant Breed. 117: 147-151.
6. Escandón A. S., M. Pérez-De la Torre, A. Acevedo, P. Marcucci and I. Miyajima. 2005a. Anchored issr as molecular marker to characterize accessions of Jacaranda mimosifolia L. Don. Acta Hort. 683: 121-127.
7. Escandón, A., M. Pérez-De la Torre, M. S. Soto y N. Zelener N. 2005b. Identificación de clones selectos de Nierembergia linariaefolia mediante microsatélites anclados. Rev. Invest. Agropec. 34: 5-17.
8. Fang, D. Q. and M. L. Roose. 1997. Identification of closely re-lated citrus cultivars with inter-simple sequence repeat markers. Theor. Appl. Genet. 95: 408-417.
9. George, S., J. Sharma and V. L. Yadon. 2009. Genetic diversity of the endangered and narrow endemic Piperia yadonii (Orchidaceae) assessed with issr polymorphisms. Am. J. Bot. 96: 2022-2030.
10. Hu, J., M. Nakatani, L. A. Garcia, T. Kuranouchi and T. Fujimura. 2003. Genetic analysis of sweetpotato and wild relatives using inter-simple sequence repeats (issrs). Breed. Sci. 53: 297-304.
11. Marouelli, P. L., P. W. Inglis, M. A. Ferreira and G. S. C. Busto. 2010. Genetic relationships among Heliconia (Heliconiaceae) spe-cies based on rapd markers. Gen. Mol. Res. 9: 1377-1387.
12. Miñano, S. H., M. E. González B. and C. Martin. 2009. Molecular characterization and analysis of somaclonal variation in chrysan-themum cultivars using rapd markers. Sci. Hort. 122: 238-243.
13. Mohapatra, A. and G. R. Rout. 2006. Optimization of primer screening for evaluation of genetic relationship in rose cultivars. Biol. Plant. 50: 295-299.
14. Nei, M. 1972. Original measure of genetic identity and genetic distance genetic. Am. Naturalist. 106: 283-292.
15. Oszkinis, K. and A. Lisiencka. 1990. Gerbera. edaMex. México, México. 14 p.
16. Pérez-Almeida, I., S. Vásquez G., D. Pérez., O. De la Rosa y E. Salazar. 2009. Huella genética de genotipos silvestres y comerciales de Passiflora spp. Utilizando patrones rapd. Bioagro. 21: 203-208.
17. Piña-Escutia, J. L., C. Vences C., M. G. Gutiérrez M., L. M. Vázquez G. y A. M. Arzate-Fernández. 2009. Caracterización morfológica y molecular de nueve variedades botánicas de Tigrida pavonia (L.F) DC. Agrociencia. 44: 147-148.
18. Pradeep, R. M., N. Sarla, and E. A. Siddiq. 2002. Inter simple sequence repeat (issr) polymorphism and its application in plant breeding. Euphytica. 128: 9-17.
19. Rezende, S. K. R., L. Vilela P., R. Paiva, A. Chalfun J., P. Pereira T. and T. E. Mesetto. 2009. Genetic divergence among cultivars of gerbera using rapd markers. Ciência Rural. 39: 2435-2440.
20. Tapia, C. E., A. H. Guillén y M. A. Gutiérrez. 2005a. Caracterización genética de materiales de piña (Ananas spp.) mediante rapd e issr. Rev. Fitotec. Mex. 28: 187-194.
21. Welsh, J. and M. Mc Clelland. 1999. Genomic fingerprinting using arbitrarily primed pCr and a matrix of pairwise combina-tions of primers. Nucleic Acids Res. 19: 5275-5279.
22. Yamagishi, M. 1995. Detection of section specific amplificated polymorphic dna (rapd) markers in Lilium. Theor. App. Gen. 91: 830-835.
23. Yamagishi, M., H. Abe., M. Nakano and A. Nakatsuka. 2002. PCR-based molecular markers in Asiatic hybrid lily. Sci. Hort. 96: 225-234.
24. Zhang, L., L. Jun, Q., L. Taol H., J. Chen and L. Zhu, D. 2006. Genetic diversity and geographic differentiation in Tacca chantri-eri (Taccaceae): an autonomous selfing plant with floral display. Ann. Bot. 98: 449-457.
25. Zhou, Z., M. Miwa and T. Hogestsu. 1999. Analysis of genetic structure of a Suillus grevillei population in a Larix caempfleri stand by polymorphism of inter-simple sequence repeat issr. New Phytol. 144: 55-63.

115
LINEAmIENTOS PARA AuTORES Y DICTAmINADORES DE LA REVISTA “CIENCIAS AGRICOLAS INFORmA”
Enero de 2014
A) Aspectos generales del texto
• La revista CIENCIAS AGRICOLAS INFORMA (CAI) es una publicación semestral editada por la Facultad de Ciencias
Agrícolas de la Universidad Autónoma del Estado de México. Los trabajos publicados corresponden a las CIENCIAS
AGROPECUARIAS Y LOS RECURSOS NATURALES, incluyendo aspectos educativos y de comercialización.
La revista tiene las siguientes secciones: Genética vegetal y fisiología, Recursos naturales y protección ambiental,
Sanidad vegetal, Biotecnología, Manejo de suelo y mecanización agrícola, Agroindustrias, Producción pecuaria,
Administración y economía agrícola y Educación en ciencias agropecuarias. Los artículos pueden estar escritos
en español o inglés; Cai no tiene el servicio de traducción, por lo que en caso de escritos en idioma inglés deberá
presentarse una carta de la empresa en donde se realizó la revisión gramatical.
• Se recibirán artículos producto de una investigación original, ensayos, notas bibliográficas, o revisiones de libros
recién editados, que no hayan sido publicados en otras revistas. Las propuestas de artículos de una investigación
tendrán una extensión no mayor a 25 cuartillas, mientras que las notas breves, ensayos, revisiones bibliográficas y de
libros tendrán una extensión hasta de diez cuartillas. El trabajo deberá enviarse por correo electrónico a cieagrinfo@
gmail.com
B) Formato
• El escrito deberá estar en letra Times New Roman, tamaño 12 puntos, a doble espacio y con número de
renglón (continuo en todo el documento) en el lado izquierdo de la página y tendrá márgenes de 2.5 cm por
los cuatro lados.
• Las unidades y fórmulas deberán escribirse siguiendo las normas del Sistema Internacional de Unidades. Los números
sin abreviación de unidad deberán escribirse con letra del cero al nueve, del 10 en adelante con número arábigo. La
separación de decimales será con coma y no con punto. Ejemplo: 0,01 y no 0.01.
• El trabajo escrito llevará una página de presentación donde se escribirán los datos generales del trabajo
como título, nombre completo de los autores (nombre y apellidos), lugar de adscripción, dirección postal,
teléfono y correo electrónico donde pueda recibir mensajes o intercambiar información. Indicar al autor
responsable.
• El título del artículo, tanto en español como en inglés se escribirá en mayúsculas y minúsculas alineado a la izquierda,
así como el nombre de los autores.

lineamientos para autores y dictaminadores de la revista CienCias agRiColas infoRma
116
• Los encabezados de segundo orden, i.e., INTRODUCCIÓN, RESUMEN, ABSTRACT, MATERIALES Y
MÉTODOS, etc., se escribirán en mayúsculas. Los encabezados de tercer orden se escribirán en minúsculas,
excepto la letra inicial, p.e. Análisis estadístico. En caso de existir encabezados de cuarto orden, se escribirán
en minúsculas, excepto la letra inicial, y en cursivas: Extracción de pectina.
• Las partes a considerar en el artículo son: TÍTULO, TÍTULO EN INGLÉS, RESUMEN, ABSTRACT,
INTRODUCCIÓN, MATERIALES Y MÉTODOS, RESULTADOS Y DISCUSIÓN, CONCLUSIONES Y
REFERENCIAS BIBLIOGRÁFICAS.
• TÍTULO. Debe ser preciso y resaltar el aspecto más importante del trabajo. Además de ser breve, no exceder
de 15 palabras. Escrito en mayúsculas y minúsculas, los nombres científicos irán en cursivas. Inmediatamente
después del título en español se incluirá su traducción correcta al inglés.
• RESUMEN Y ABSTRACT con una extensión no mayor a 150 palabras.
• PALABRAS CLAVE. Después del RESUMEN o ABSTRACT incluir, en orden alfabético, tres o cuatro palabras
clave o key words, según sea el caso y de preferencia no aparezcan en el título del trabajo.
• INTRODUCCIÓN. Definir el problema de estudio, antecedentes que contextualicen el problema
(con las respectivas referencias bibliográficas que apoyen este apartado), objetivos e hipótesis de trabajo.
• MATERIALES Y MÉTODOS. Consistirá de una breve descripción del lugar y condiciones en donde se
realizó la investigación. También se mencionarán los materiales, equipo, metodologías y procedimientos
utilizados congruentes con los objetivos. Asimismo, se deberán indicar las variables de estudio involucradas,
modelo estadístico utilizado y los análisis de estudio implicados. Sólo en caso de que se utilice una
metodología innovadora, ésta deberá ser descrita con mayor detalle, así como los autores que respaldan
la metodología en cuestión.
Caracterización molecular de cuatro variedades de gerbera jamesonii Bolus, mediante microsatélites anclados y rapds
molecular characterization of four varieties of Gerbera jamesonii bolus, through anchored microsatellites and rapds
Amaury Martín Arzate-Fernández,1* José Luis Piña-Escutia,1 Luis Miguel Vázquez-García,2
Adolfo Carrillo-Velázquez1

117
lineamientos para autores y dictaminadores de la revista CienCias agRiColas infoRma
• RESULTADOS Y DISCUSIÓN. Presentación e interpretación escrita en forma ordenada, clara, objetiva
e imparcial del fenómeno observado, sin repetición de la información de cuadros y figuras. Comparación
de los resultados del trabajo en relación con lo publicado por otros autores, así como el señalamiento de
posibles causas de las respuestas observadas del fenómeno de estudio, evitando caer en especulaciones de
cualquier tipo. Preferentemente deberán presentarse los resultados en cuadros y figuras apegándose a los
lineamientos que más adelante se describen.
• CONCLUSIONES. Deberán ser de manera categórica, breve y precisa enunciando las aportaciones
concretas al conocimiento de acuerdo con los objetivos planteados y apoyados en los resultados obtenidos
en el trabajo.
• REFERENCIAS BIBLIOGRÁFICAS. Todas las publicaciones citadas en el artículo deberán enlistarse
alfabéticamente y numeradas. Evitar consultas de tesis o memorias de congresos sin arbitraje o en corto.
Las referencias bibliográficas deberán incluirse en estricto orden alfabético con el apellido paterno del
primer autor.
1. Alzofeifa-Delgado, A. 2006. Uso de los marcadores moleculares en plantas; aplicaciones en frutales del trópico. Agron. Mesoam. 17: 221-242.
2. Bautista-Puga, M. D., L. M. Vázquez, G., H. Leszczynska, B. M. W. Borys y A. M. Arzate-Fernández. 2011. Caracterización del lirio azteca, mediante marcadores morfológicos y moleculares. Agrociencia. 45: 413-422.
3. Chung Y. M., Kim H. A., Kim K. Y., Park S. W., Yi Y. B., Lee J. H. and Chang O. K. 2001. Morphological characteristics and genetic variation of gerbera (Gerbera hybrida hort.). J. Plant Biotechnol. 3: 145–149.
4. Da Mata, L. T., M. I. Segeren, A. Seregen, F. and C. A. Colombo. 2009. Genetic divergence among accessions evaluated by rapd. Sci. Hort. 121: 92-96.
5. Debener, T. and L. Mattiesch. 1998. Effective pairwise combination of long primers for rapds analyses in rose. Plant Breed. 117: 147-151.
6. Escandón A. S., M. Pérez-De la Torre, A. Acevedo, P. Marcucci and I. Miyajima. 2005a. Anchored issr as molecular marker to characterize accessions of Jacaranda mimosifolia L. Don. Acta Hort. 683: 121-127.
7. Escandón, A., M. Pérez-De la Torre, M. S. Soto y N. Zelener N. 2005b. Identificación de clones selectos de Nierembergia linariaefolia mediante microsatélites anclados. Rev. Invest. Agropec. 34: 5-17.
8. Fang, D. Q. and M. L. Roose. 1997. Identification of closely related citrus cultivars with inter-simple sequence repeat mark-ers. Theor. Appl. Genet. 95: 408-417.
9. George, S., J. Sharma and V. L. Yadon. 2009. Genetic diversity of the endangered and narrow endemic Piperia yadonii (Orchidaceae) assessed with issr polymorphisms. Am. J. Bot. 96: 2022-2030.
10. Hu, J., M. Nakatani, L. A. Garcia, T. Kuranouchi and T. Fujimura. 2003. Genetic analysis of sweetpotato and wild relatives using inter-simple sequence repeats (issrs). Breed. Sci. 53: 297-304.
11. Marouelli, P. L., P. W. Inglis, M. A. Ferreira and G. S. C. Busto. 2010. Genetic relationships among Heliconia (Heliconia-ceae) species based on rapd markers. Gen. Mol. Res. 9: 1377-1387.
12. Miñano, S. H., M. E. González B. and C. Martin. 2009. Molecular characterization and analysis of somaclonal variation in chrysanthemum cultivars using rapd markers. Sci. Hort. 122: 238-243.
13. Mohapatra, A. and G. R. Rout. 2006. Optimization of primer screening for evaluation of genetic relationship in rose cul-tivars. Biol. Plant. 50: 295-299.
14. Nei, M. 1972. Original measure of genetic identity and genetic distance genetic. Am. Naturalist. 106: 283-292.15. Oszkinis, K. and A. Lisiencka. 1990. Gerbera. edaMex. México, México. 14 p.16. Pérez-Almeida, I., S. Vásquez G., D. Pérez., O. De la Rosa y E. Salazar. 2009. Huella genética de genotipos
silvestres y comerciales de Passiflora spp. Utilizando patrones rapd. Bioagro. 21: 203-208.

lineamientos para autores y dictaminadores de la revista CienCias agRiColas infoRma
118
A continuación se citan algunas formas para las referencias bibliográficas:
En español:
Libro con varios autores
Satorre, E. H., A. R. L. Beneche, G. A. Slafer, E. B. de la Fuente, D. J. Miralles, M. E. Otegui y R. Savin. 2003.
Producción de granos. Bases funcionales para su manejo. Facultad de Agronomía. Universidad de Buenos Aires. Buenos
Aires, Argentina. 783 p.
Libro con dos autores
Domínguez, P. J. y A. Castañeda V. 2002. Guía técnica para la producción de chirimoya en el Estado de México.
Fundación Salvador Sánchez Colín-CiCtaMex. Coatepec Harinas, México. 30 p.
Libro con un autor
Cadahia, L. C. 2006. Fertirrigación. Cultivos hortícolas y ornamentales. Mundi-Prensa. Madrid, España. 55 p.
Capítulo de libro
Loyola, V. M. y J. R. López. 1985. El cultivo de tejidos vegetales para la producción de sustancias naturales. En:
M. L. Robert y V. M. Loyola (comp.) El cultivo de tejidos vegetales en México. Centro de Investigación Científica
de Yucatán, A.C., Consejo Nacional de Ciencia y Tecnología. México D. F. pp. 125-132.
Sección de libro
Ayala, F. J. y J. A. Kiger, Jr. 1984. Genética moderna. Omega. Madrid, España. pp. 183-299.
Autoría institucional
Centro Internacional de Mejoramiento de Maíz y Trigo (CiMMyt). 1985. Desarrollo, mantenimiento y multiplicación de
semilla de variedades de maíz de polinización libre. El Batán, Texcoco, México. 11 p.
Artículo en revista
González, H. A., J. Sahagún C., L. M. Vázquez G., J. E. Rodríguez P., D. J. Pérez L., A. Domínguez L., O. Franco M.
y A. Balbuena M. 2009. Identificación de variedades de maíz sobresalientes considerando el modelo aMMi y los índices
Eskridge. Agric. Tec. Méx. 35: 189-200.
En inglés:
Libro
Valero, D. and M. Serrano. 2010. Postharvest biology and technology for preserving fruit quality. CrC. Boca Raton, USA.
287 p.

119
lineamientos para autores y dictaminadores de la revista CienCias agRiColas infoRma
Capítulo de libro
Miller, W. 1993. Lilium longiflorum. In: De Hertogh, A. and M. Le Nard (eds.). The physiology of flower and bulbs.
Elsevier. Amsterdam, Netherlands. p. 391-422.
Artículo en revista
Chamani, E., A. Khalighi, C. D. Joyce, E. D. Irving, A. Z. Zamani, Y. Mostofi and M. Kafi. 2005. Ethylene and anti-
ethylene treatment effects on cut ‘First Red’ rose. J. Applied Hort. 7: 3-7.
Becker, H. B. and J. León. 1988. Stability analysis in plant breeding. Plant Breed. 101: 1-23.
Cockerham, C. C. 1961. Implications of genetic variances in a hybrid breeding program. Crop Sci. 1: 47-52.
Fox, P. N., B. Skovmand, B. K. Thompson, H. J. Braun and R. Cormier. 1990. Yield and adaptation of hexaploid spring
triticale. Euphytica. 47: 57-64.
Maddonni, G., M. E. Otegui, B. Andrieu, M. Chelle and J. J. Casal. 2002. Maize leaves turn away from neighbors. Plant
Physiol. 130: 1181-1189.
Fuentes de información electrónica en línea:
Mercy A. O., N. S. Lang, F. W. Ewers and S. A. Owens. 2006. Xylem vessel anatomy of sweet cherries grafted onto dwarfing
and nondwarfing rootstocks. J. Amer. Soc. Hort. Sci. 131: 577-585 (consultado en http://www.electronic.com/JournalEZ/
toc.cfm?code=0420001&lssueSelector=13105&CFID=2188994&CFTOKEN=16427495F6F3-439D-A4BF7749EOFBD4AF,
fecha de consulta 09 de noviembre de 2006).
Valenzuela, V. H., T. Herrera, M. I. Gaso, E. Pérez-Silva y E. Quintero. 2004. Acumulación de radiactividad en hongos
y su relación con roedores en el bosque del Centro Nuclear de México. Rev. Int. Contaminación Amb. 20: 141-
146 (consultado en http://redalyc.uaemex.mx/redalyc/src/inicio/ArtPdfRed.jsp?iCve=3702401&iCveNum=1699, fecha de
consulta 09 de noviembre de 2006).
El hecho de no respetar el formato de referencias bibliográficas derivará en la devolución inmediata de la propuesta sin
iniciar el proceso de revisión.
C) Para los autores
• El proceso de publicación en la revista Cai iniciará con una revisión de la propuesta por parte del editor principal.
Si la decisión es que el artículo es de interés de Cai, el autor responsable recibirá una clave de identificación del
manuscrito, para su posterior seguimiento. El artículo será enviado a dos pares académicos de reconocido prestigio
en el área del conocimiento correspondiente para su evaluación anónima; modalidad denominada “doble ciego”.
El autor responsable recibirá, en un plazo no mayor de dos meses, la comunicación de los comentarios de los
revisores a su artículo. El resultado podrá ser: ACEPTADO EN SU FORMA ACTUAL, ACEPTADO CON
CORRECCIONES MENORES, CONDICIONADO A CORRECCIONES SUSTANTIVAS, RECHAZADO.
Una vez que el autor responsable reciba las sugerencias de los árbitros, tendrá tres semanas para remitir la versión
corregida de su trabajo. Cai se reserva el derecho de rechazar los trabajos cuyo autor responsable no cumpla el
plazo sugerido.

lineamientos para autores y dictaminadores de la revista CienCias agRiColas infoRma
120
Cuando en un plazo mayor a dos meses, los dictaminadores no hayan emitido su respuesta, el editor principal
comunicará al autor responsable si desea cambiar, por única ocasión, de evaluadores. En caso de un nuevo retraso
en la evaluación, se comunicará al autor principal del rechazo del documento.
D) Para los revisores
• Los revisores tendrán un plazo máximo de 30 días hábiles para remitir el dictamen correspondiente mediante un
oficio dirigido a la Coordinación Editorial de la Revista y el dictamen por separado sin firma ni nombre. En el oficio
se debe incluir si el escrito es aceptado en su estado actual, aceptado con correcciones menores, condicionado y
sujeto a modificaciones que mejoren su presentación o si es rechazado con la debida argumentación.
E) Anexos
Presentación de cuadros
• Los cuadros deben presentarse numerados en forma sucesiva (por ejemplo: Cuadro 1, 2, 3, ..., n). Su colocación será
inmediatamente después de haber sido citado. Los cuadros deben presentarse en el formato de “tabla” de Word
únicamente (no se aceptarán cuadros hechos con tabulaciones).
• El título de los cuadros deberá colocarse en la parte superior de éstos y escribirse en letra mayúscula inicial. El mismo
procedimiento se seguirá para los encabezados de las columnas o hileras en caso de tratarse de cuadros de doble
entrada.
• En los cuadros solamente se aceptarán tres líneas principales en forma horizontal, sin líneas verticales. Los números
deberán alinearse por el punto. Se sugiere un tamaño de línea de 1.5 puntos para las líneas principales del cuadro y
de 0.5 puntos para la línea que divide los títulos del cuadro y los datos del cuadro.
Ejemplos de cuadros
Cuadro 1. Análisis de varianza, media general y coeficiente de variación para días a espigamiento (de), días a floración (df), roya amarilla (ra), porcentaje de acame (pa), madurez fisiológica (Mf), altura de la planta (ap), área de la hoja bandera (ahB), área de la segunda hoja (ash) y longitud de entrenudos (len).
F.V. G.L. de df ra pa mf ap ahb ash len
Genotipos 11,7** 15,4** 1,3 ns 2,0 NS 4,2** 12,0** 4,3* 5,4** 20,8**
Repeticiones 2 0,3 NS 0,2 NS 3,6* 1,4 NS 0,21 NS 9,41** 2,64 NS 10,8** 3.55 NS
Media 72,6 77,6 9,9 13,3 130,1 79,2 21,0 21,9 51,3
C.V. 1,9 1,1 69,3 151,3 1,6 3,5 10,6 8,2 3,8
* Significativo al 0,05; ** altamente significativo al 0,01; ns no significativo.

121
lineamientos para autores y dictaminadores de la revista CienCias agRiColas infoRma
Presentación de figuras
• Se consideran como figuras las fotografías, grabados, gráficas, dibujos, mapas, planos de localización y esquemas
que den idea del fenómeno estudiado. Las figuras no deben ser repetición de los cuadros o del texto y también se
numeran consecutivamente: Figura 1, 2, 3,...,n.
• Las figuras se colocarán inmediatamente después de haber sido referidas. El título de figura deberá colocarse al pie
de ésta con letra mayúscula inicial. La figura debe explicarse por sí misma para evitar repeticiones en el texto.
• Sólo podrán presentarse figuras en blanco y negro.
• Las gráficas deberán enviarse en Excell o Sigmaplot, indicando la versión utilizada. Se recomienda no presentar
gráficas con efectos en tercera dimensión a menos que sea estrictamente necesario o la naturaleza de la gráfica así
lo requiera.
Cuadro 2. Acción residual de fungicidas frente al tizón gomoso del tallo en época de primavera.
Tratamientos
Intensidad de la enfermedad (Medias transformadas)
2004 2005
5días
12 días
18días
23días
5días
12 días
18días
23días
Azoxistrobina 0,88 a 1,18 a 1,76 a 1,95a 0,88a 1,28a 1,84a 1,99a
Tiofanato metilo 0,88 a 1,28 a 1,81ab 2,03ab 0,88a 1,43a 1,86ab 2,03a
Clorotalonilo 0,88 a 1,58 b 1,88b 2,07b 1,08b 1,69b 1,96bc 2,11b
Mancozeb 1,48 b 1,84 c 1,99c 2,09b 1,58c 1,97c 2,05c 2,13b
Testigo 1,73 c 1,91 c 2,01c 2,09b 1,83d 1,91c 2,07c 2,16b
CV 22,91 20,51 6,48 3,85 23,28 17,84 6,61 4,06
ESx 0,077 0,066 0,024 0,016 0,08 0,059 0,026 0,017
Medias con letras diferentes en la columna difieren significativamente (p< 0,05).

lineamientos para autores y dictaminadores de la revista CienCias agRiColas infoRma
122
LTCL VTCL AVENA
Núme
ro de
tallo
s (m2
)
0
100
200
300
400
500
LTCL VTCL AVENA
Peso
fresco
de fo
rraje (
t ha-1
)
0
10
20
30
40
50
LTCL VTCL AVENA
Peso
seco
de fo
rraje (
t ha-1
)
0
2
4
6
8
10
LTCL VTCL AVENA
Peso
seco
de ta
llos (
g)
0
5
10
15
20
25
(a) (b)
(c) (d)
Ejemplos de figuras
Figura 1. Comportamiento promedio de cinco líneas de triticale (ltCl), dos variedades de triticale (vtCl) y una variedad de avena para peso fresco de forraje (a) y peso seco de forraje (b). Las líneas verticales de cada barra indica el error estándar de la media de cada grupo.
Figura 2. Producción de materia seca como una función de los días de crecimiento de la planta en etapa de antesis.
Núm
ero
de ta
llos
(m2 )


Ciencias Agrícolas Informa No. 22(2) es una
revista publicada por la Facultad de Ciencias
Agrícolas, se terminó de imprimir en el mes de
diciembre de 2013, en Editorial CigoMe s.a. de C.v. La edición consta de 200 ejemplares.