functional analysis of genes involved in the regulation of
TRANSCRIPT

Functional analysis of genes involved in the regulation of developmentof reproductive organs in rice (Oryza sativa)Chen, Y.
CitationChen, Y. (2011, December 20). Functional analysis of genes involved in the regulation ofdevelopment of reproductive organs in rice (Oryza sativa). Retrieved fromhttps://hdl.handle.net/1887/18262 Version: Corrected Publisher’s Version
License: Licence agreement concerning inclusion of doctoral thesis in theInstitutional Repository of the University of Leiden
Downloaded from: https://hdl.handle.net/1887/18262 Note: To cite this publication please use the final published version (if applicable).

Chapter 3
Functions of OsJAR1 in Floret Opening and Anther
Dehiscence in Rice (Oryza sativa)
Yuguo Xiao*1, 2, Yi Chen*1, Jeroen Heijmans1, Angela Hoogenboom1, Tatsiana Charnikhova3, Patrick P.J. Mulder4, Adamantia Agalou1, Zhen Zhu2, Mei
Wang1, 5, Ludovico Dreni6, Martin M. Kater6, Corinne Michel7, Jean-Benoit Morel7, Harro Bouwmeester3, and Pieter B.F. Ouwerkerk1
1 Department of Molecular and Developmental Genetics, Institute Biology Leiden, Leiden University, Sylvius Laboratory, Sylviusweg 72, PO Box 9505, 2300 RA Leiden, The Netherlands 2 State Key Laboratory of Plant Genomics, Institute of Genetics and Developmental Biology, Chinese Academy of Sciences, No.1 West Beichen Road, Chaoyang District, Beijing, 100101 China 3 Plant Research International BV, Droevendaalsesteeg 1 6708 PB, PO Box 16, 6700 AA, Wageningen, The Netherlands 4 RIKILT– Institute of Food Safety, Bornsesteeg 45, 6708 PD, PO Box 230 6700 AE, Wageningen, The Netherlands
5 SU BioMedicine-TNO, Utrechtseweg 48, 3704 HE Zeist, PO Box 360, 3700 AJ Zeist, The Netherlands 6 Dipartimento di Scienze Biomolecolari e Biotecnologie, Universita` degli Studi di Milano, Via Celoria 26, 20133 Milan, Italy 7 UMR BGPI INRA/CIRAD/SupAgro, Campus International de Baillarguet, TA A 54 / K34398 Montpellier, France
* These authors contribute equally to the work


Chapter 3 Functions of OsJAR1 in floret opening and anther dehiscence
79
Abstract In literature two rice JASMONATE RESISTANT 1 (osjar1) lines were described that were mutated by the endogenous retrotransposon Tos17. The gene disrupted in the two independent lines, named osjar1-2 and osjar1-3 respectively, is predicted to encode a putative auxin-responsive GRETCHEN HAGEN 3 (GH3) family protein named OsJAR1. In this chapter, we present a detailed characterization of the osjar1 phenotype and present a model for the biological functions of OsJAR1. In the osjar1 mutants, the florets fail to close again after opening because the withering of the lodicules is delayed by several days whereas this normally happens within hours. The reduced fertility of osjar1 plants is caused by a delay in anther dehiscence. Normally, OsJAR1 mRNA is present in floral and vegetative organs but is down-regulated in osjar1-2 and the length is shorter in osjar1-3 due to the Tos17 insertions. In contrast to the wild type, floret opening in osjar1-2 and osjar1-3 cannot be induced by exogenous application of methyl jasmonate (MeJA) and root growth is not inhibited by MeJA. Overexpression of OsJAR1 under control of the rice actin promoter rescued the osjar1-2 phenotype and restored floret opening in response to MeJA. osjar1-3 is more sensitive to rice blast disease and overexpression of OsJAR1 in wild type rice plants increases resistance to blast disease, indicating a role of OsJAR1 in defence against pathogens. The OsJAR1 protein specifically conjugates alanine, cysteine, leucine, isoleucine, methionine, phenylalanine, tryptophan and valine to JA in in vitro assays and JA-Ile levels were reduced in osjar1-2 plants. We conclude that OsJAR1 is a JA-amino acid synthetase that is required to convert JA into JA-Ile for optimal flower opening, anther dehiscence, seed development and resistance to blast disease.
Introduction
Anthesis is the period during which a flower is fully open and functional and is obviously an essential step in the production of progeny. Events that take place during anthesis include flower opening and closure, anther dehiscence, release of pollen grains and fertilization. In general, flower opening and closure is caused by the movement of flower organs. In tulip and taraxacum, for example, petal movement due to unequal growth on both sides of the petals results in the opening of flower (Tanaka et al., 1988; Van Doorn and Van Meeteren, 2003). In rice, the opening and closure of the florets result from the separation of lemma from palea caused by the expansion and subsequent atrophy or withering of the lodicules (Matsuo and Hoshikawa, 1993; Qin et al., 2005). External factors such as light (Kaihara and Takimoto, 1980, 1981; Bieleski et al., 2000) and temperature (Kaihara and Takimoto, 1981; Ichimura and Suto, 1998) were described to have effects on flower opening and closure in several plant species. The plant hormones ethylene and MeJA were also shown to have critical functions in regulation of flower opening and closure. Ethylene promotes or inhibits flower opening depending on the species. For instance, in cut roses, flower opening is promoted by exogenous ethylene application (Ma et al., 2005). MeJA induces floret opening in rice and the numbers of induced opening florets are correlated with MeJA concentration in the wild type (Zeng et al., 1999). Thus far, no gene directly related to regulation of flower opening and closure has been identified.
Anther dehiscence and pollen grain release is another important step during anthesis. Several genes involved in anther dehiscence have been identified and characterized in several plant species.

Functions of OsJAR1 in floret opening and anther dehiscence
80
These genes include, AOS/DDE2-2 (Park et al., 2002; Von Malek et al., 2002), AtMYB26 (Steiner-Lange et al., 2003), COI1 (Xie et al., 1998), DAD1 (Ishiguro et al., 2001), NST1/NST2 (Mitsuda et al., 2005), OPR3/DDE1 (Sanders et al., 2000; Stintzi and Browse, 2000) and RPK2 (Mizuno et al., 2007) from Arabidopsis and NEC1 (Ge et al., 2001) and PhETR2 (Wang and Kumar, 2007) from petunia. Loss-of-function mutants of these genes show obvious anther developmental defects in timing of anther dehiscence, filament elongation or pollen viability. AOS/DDE2-2, COI1, DAD1 and OPR3/DDE1 encode proteins involved in the jasmonic acid (JA) pathway, which indicates that this plant hormone may be a critical regulator of anther dehiscence in plants. In rice, the Myb domain protein AID1 (Zhu et al., 2004) was shown to be involved in anther development. The aid1 mutation gives rise to partial completion of anther indehiscence and this phenotype could not be complemented by exogenous application of MeJA, suggesting that AID1 is not involved in MeJA-mediated anther dehiscence in rice.
Riemann et al. (2008) described two Tos17 mutants named osjar1 and osjar2 respectively after JASMONATE RESISTANT 1. The gene disrupted in two independent lines osjar1-2 and osjar1-3 is predicted to encode a putative auxin-responsive GRETCHEN HAGEN 3 (GH3) family protein which is closely related to JAR1 of Nicotiana glutinosa (NgJAR1) and JAR1 of Arabidopsis (AtJAR1). However the relation between the osjar1 phenotype and the biological function of OsJAR1 remained unclear. Simultaneously we identified the same osjar1 mutant in a population of T-DNA and transposon mutants in genes predominantly expressed in developing flowers and grains. The osjar1 lines showed a very obvious phenotype of open-staying flowers and elongated seeds. In this chapter we report on a detailed characterization of the osjar1 phenotype and propose a biological function of OsJAR1. In wild type plants, OsJAR1 mRNA is present in both floral and vegetative organs, but it is down-regulated in osjar1-2 whereas its length is shorter in osjar1-3. Analysis of an OsJAR1 promoter-β-glucuronidase construct revealed that the OsJAR1 promoter is predominantly active in the vasculature of floral and vegetative tissues. We confirmed localization of OsJAR1 expression in anthers using RNA in situ hybridization. Floret opening in osjar1-2 and osjar1-3 cannot be induced by exogenous application of MeJA which demonstrated that MeJA-induced floret opening pathway is defective in osjar1-2 and osjar1-3. Overexpression of OsJAR1 in the background of osjar1-2 rescued the osjar1-2 phenotype. osjar1-2 mutants show reduced root growth inhibition by MeJA which suggested that OsJAR1 is probably also involved in other aspects of MeJA-mediated plant growth and developmental processes. In addition, we observed that osjar1-3 has reduced resistance to the blast fungus M. oryzae and overexpression of OsJAR1 in control rice plants enhanced resistance towards blast disease, confirming a link between JA and disease resistance. Thin-layer chromatography (TLC) and liquid chromatography-tandem mass spectrometry (LC-MS/MS) analysis of products produced in vitro by recombinant OsJAR1 protein demonstrated that this GH3 protein is able to form conjugates between JA and alanine, cysteine, isoleucine, leucine, methionine, phenylalanine, tryptophan and valine but not with the twelve other amino acids. In osjar1-2 we observed a significantly decreased level of JA-Ile in both JA treated and untreated leaves which confirm the function of OsJAR1 in conjugation of JA to isoleucine. We propose that this step and JA-Ile plays a crucial role in timing of flower opening and closure, anther dehiscence and disease resistance.

Chapter 3
81
Results Phenotypical characterization of osjar1 Normally in rice, anthesis only happens during a specific period of the day and lasts for 40 to 90 min depending on the cultivar. In our greenhouse settings, florets of rice Nipponbare rice plants open at noon and keep opened for approximately 90 minutes. During anthesis, florets open and the filaments of the stamen elongate, so that the anthers are positioned outside the florets; simultaneously anther dehiscence and pollination are taking place. At the end of anthesis, florets are closing again (Figure 1A-1E). The opening and closure of the florets is driven by the expansion and subsequently atrophy of the lodicules. The rice floret has two lodicules just below the base of its stamina and on the lemma side. The lodicules swell just before the anthesis, pushing the lemma away resulting in the opening of the florets. At the end of anthesis, closure of rice florets is due to a quick shrinking of lodicules due to withering (Figure 1K-1M) (Matsuo and Hoshikawa, 1993). In a population of Tos17
Figure 1. Comparison of the phenology of florets from wild-type and osjar1-2.plants. A-E. Time course of floret anthesis in wild type plants. Photos were taken just before anthesis or 15, 30, 60 and 90 minutes after the floret started to open, respectively. F-J. Time course of floret anthesis in osjar1-2. Photos were taken just before anthesis or 2 hours, one day, three days and five days after the floret started to open, respectively. K-M. Close-up of the lodicules of the wild-type were monitored just before floret opening (K), during floret opening (L) and just the end of florets opening (M). P-R. Close-up of lodicules of osjar1-2 were monitored just before floret opening (P), during floret opening (Q) and three days after florets started to open (R), respectively. N. Comparison of spikelet in the wild type and osjar1-2. From left to right, mature osjar1-2 spikelet, osjar1-2 spikelet at the filling stage and mature wild type spikelet. O. Comparison of wild type grains and osjar1-2 grains. From left to right, osjar1-2 grains at the filling stage, mature osjar1-2 grains, wild type grains at the filling stage and mature wild type grains. S. Wild type mature grains. T. osjar1-2 mature grains. lo, lodicules. Bars (A-J, N, O, S, T) = 1 mm; Bars (K-M, P-R) =0.5 mm.

Functions of OsJAR1 in floret opening and anther dehiscence
82
Figure 2. Comparison of anthers from wild type and osjar1-2 plants. A and B. Floret organs of wild type (A) and osjar1 (B) just before floret opening. C. Stamen of the wild-type (a) and the osjar1-2 mutant (b and c) just before floret opening. D. Stamen of the wild-type (d and e) and the osjar1-2 mutant (f and g) during floret opening. E. Wild-type anther at the pollen mature stage before anther dehiscence. F. Dehisced wild type anther. G. osjar1-2 anther at the pollen mature stage before floret opening. H and I. osjar1-2 anther during floret opening. J-N. Close-up of the region including the connective of anther in E-I, respectively. en, endothelium; ca, cavity; ccc, circular cell cluster; se, septum; ep, epidermis; P, pollen; V, vascular bundle. Bars (A-B) = 1 mm; Bars (C-D) = 100 μm; Bars (E-I) = 50 μm; Bars (J-N) = 25 μm. mutants, two different osjar1 mutants were isolated which were characterized by an obvious open staying phenotype of the flower (Riemann et al., 2008). In a detailed study we observed ourselves in total five major differences between control plants and osjar1. First, the opening of the osjar1 florets happens at random, which means also during the night instead of in a certain period of the day as normally occurs with control flowers. Second, osjar1 florets failed to close after the opening (Figure 1G-1J). Third, 87.2% of the anthers in osjar1 are not dehiscing (Figure 1G-1J, 2D). Fourth, senescence of the floral organs of osjar1 is delayed compared to control plants. For instance, in osjar1 florets, the lodicules are still big enough to keep the floret open even three days after the floret starts open (Figure 1R) whereas wild type florets already close after 90 minutes (Figure 1M). Fifth, osjar1 plants are less fertile and have abnormal seed morphology (Figure 1N, O, S and T). Only 7.5-14.5% of osjar1 florets have seeds set and the vast majority of florets are sterile. The osjar1 mutation is in cultivar Nipponbare, which is a Japonica type and has thick and round seeds (Figure 1O and 1S). In contrast, osjar1 grains are narrower and longer (Figure 1O and 1T). More than 99% of the grains from osjar1 homozygous plants show this phenotype. In addition, grains from osjar1 plants show wrinkled, curved and over-dried endosperm and the withered stigma is still kept attached to the mature grain (Figure 1N and 1O). Compared to the wild type, the osjar1 anther is shorter and a little crooked (Figure 2 A-C). Normally, the anther wall consists of four cell layers

Chapter 3
83
from outside to inside, epidermis and endothecium, middle layer and tapetum (Figure 2E, 2J). Adjacent to the circular cell cluster there is a dehiscent cavity present, which is a single layer of specialized epidermal cells connecting each other of the pair of loculi and is the final breakage site of anther dehiscence (Figure 2J) (Zhu et al., 2004). The endothelium close to the cavity and circular cell cluster shows obvious well-developed U-shaped cell wall thickenings, which is considered to be important for anther dehiscence (Figure 2J) (Matsui et al., 1999). During floret opening, the pollen increase to swell resulting in increased pressure and simultaneously cells in the anther wall lose water. As a result, the anther wall gets twisted and the circular cell cluster splits resulting in the rupture of anther loculi and pollen release (Figure 2D, 2F and 2K) (Matsui et al., 1999). Cross sections of osjar1-2 anther show no obvious difference except that the cell shape of several cells during the stage just before floret opening and anther dehiscence is different. That is, the epidermis cells, endothelium cells and septum cells in osjar1 anther are more irregular than normally (Figure 2E, 2G, 2J and 2L). In osjar1, 87.2% of the osjar1-2 anthers are not or only partly dehisced after floret opening (Figure 2D, 2H and 2I). In the non-dehisced anthers, the circular cell cluster was still kept intact and not splitted although the anther wall was shrunken considerably (Figure 2H, 2M). The osjar1 phenotype co-segregates with a Tos17 insertion in the OsJAR1 gene and correlates with the occurrence of abnormal transcripts in mutants osjar1-2 and osjar1-3 The osjar1 phenotype was found in populations derived from Tos17 insertion lines NC2728 (osjar1-2) and ND0051 (osjar1-3), which originated from the Hirochika Laboratory (Miyao et al., 2003). Three out of 19 NC2728 plants and four out of 16 ND0051 plants displayed the osjar1 phenotype. All progeny of three osjar1-2 plants (NC2728-7, 14, 19) exhibited an identical phenotype. The phenotypic segregation in line NC2728 was according to a ratio of three wild type versus one mutant plants. The same result was obtained in line osjar1-3 which demonstrates that the osjar1 phenotype is very likely caused by a recessive mutation. Database searches at NCBI (http://www.ncbi.nlm.nih.gov) retrieved 16 FSTs (flanking sequence tags) from mutant lines NC2728 and ND0051. By BLASTing these with the rice genomic database in TIGR (http://rice.plantbiology.msu.edu/), only FSTs T06540T and ND0051_0_403_1A from lines NC2728 and ND0051 respectively, were found to be associated with locus LOC_Os05g50890. This locus is predicted to contain five exons and encodes a GH3 family protein (Jain et al., 2006) named OsJAR1 (Riemann et al., 2008). The Tos17 alleles in mutant lines NC2728 and ND0051 were found to be inserted into exon 3 and 4 of OsJAR1, respectively (Figure 3A). Because these two mutants show the same phenotype, we hypothesized that it is caused by Tos17 insertions in this locus leading to disruption of gene function. This hypothesis was strongly supported by Southern blot hybridization of XbaI-digested genomic DNA from wild type plants and plants from the two mutant populations (lines NC2728 and ND0051) with OsJAR1 specific probes. The wild type plants were predicted to contain a hybridizing restriction fragment of 2.5 kb and 1.2 kb. Because there is an XbaI site in the Tos17 retrotransposon, homozygous Tos17 insertion plants were predicted to contain a fragment of 5 kb in mutant line NC2728 and a 2.2 kb fragment in ND0051 including a partial Tos17 fragment which is in agreement with the Southern blot hybridization results (Figure 3D to F). Because only homozygous Tos17 insertion plants (Tos17/Tos17) in lines NC2728 and ND0051 show the osjar1 phenotype, this result confirms that the osjar1 phenotype is co-segregating with the Tos17 insertion in OsJAR1. This was confirmed by comparing the phenotype and genotype of the first

Functions of OsJAR1 in floret opening and anther dehiscence
84
Figure 3. Association of the osjar1 phenotype with Tos17 insertions in OsJAR1. A. Schematic structure of OsJAR1 and positions of Tos17. OsJAR1 contains five exons (black bars) and four introns (white bars). ATG and TGA indicate start and stop codons respectively. Triangles indicate the positions of Tos17 insertions in mutant line NC2728, NE8577 and ND0051 respectively. Black arrows indicate the orientations of the Tos17 insertions. The lines below the OsJAR1 gene indicate the positions of the probes used in Southern and northern blot analyses (P1, P2 and N1, N2, respectively). Xb, XbaI. B and C. Positions and orientations of Tos17 in osjar1-2 (B) and osjar1-3 (C). Letters in grey indicatedthe repeated sequence caused by Tos17 insertion, black letters indicate nucleotides from exon 4 and exon 5 of ogm1 and ogm2, respectively. D, E and F. Co-segregation of Tos17 in OsJAR1 with the mutant phenotype of osjar1. As a result of Tos17 insertions, 5 kb, 2.2 kb and 2.5 kb polymorphic bands appear in mutant lines NC2728, ND0051 and NE8544, respectively, whereas the wild-type OsJAR1 gene showed a 2.5 kb, 1.2 kb and 1.2 kb band in segregatns of mutant line NC2728, ND0051 and NE8544, respectively. The 5 kb and 2.2 kb polymorphic bands in mutant lines NC2728 and ND0051 were fully associated with the osjar1 phenotype (O). W, OsJAR1 in a wild type plant. H, heterozygous OsJAR1/osjar1 plant. P, wild-type Nipponbare control. G. OsJAR1 mRNA expression in wild type and osjar1 mutants. Northern blot hybridized with probe N2 and N1 which were indicated in Figure 3A. Blots were hybridized with 17S rRNA to check for the total RNA loading. L leaves; S stems; R, roots; P, panicles. generation of heterozygous Tos17 insertion plants in both mutant lines. The progeny of heterozygous Tos17 insertion plants segregated at a 1:2:1 ratio of homozygous (Tos17/Tos17), heterozygous (Tos17/+) and Tos17 null (+/+) and only Tos17/Tos17 plants display the osjar1 phenotype. The Tos17 insertion positions in osjar1-2 and osjar1-3 were confirmed by sequencing the FSTs of the Tos17 insertions in osjar1-2 and osjar1-3 genomic DNA with two primers, one is OsJAR1-specific and adjacent to the annotated Tos17 insertion site, and the other is Tos17 specific. The sequencing results on the FSTs show that the Tos17 retrotransposons in the osjar1-2 and osjar1-3 mutants were inserted after amino acid 354 and 498 of the OsJAR1 ORF, respectively. FST sequences obtained from osjar1-2 and osjar1-3 were submitted to NCBI with Genbank Accession codes FI494725-FI494726 and FI494727-FI494728, respectively. The insertions in osjar1-2 and osjar1-3 caused a duplication

Chapter 3
85
of five nucleotides OsJAR1 target sequence adjacent to the border of Tos17 (Figure 3B-3C). To check the expression of the OsJAR1 gene in osjar1-2, osjar1-3 and control plants, northern
blot analysis was performed using two OsJAR1-specific probes named N1 and N2 (Figure 3A). With probe N1, a 2.4 kb transcript was detected in different tissues including root, shoot, leaf and young panicle in the wild type, but similar sized transcripts were not detected in osjar1-2 and osjar1-3 (Figure 3G). For osjar1-2, we only observed a faint and bigger transcript, which probably is caused by the presence of Tos17 (4.1 kb) in the transcript. For osjar1-3, we detected a transcript smaller than in the wild type. With probe N2, we could only detect an OsJAR1 mRNA signal in the wild type (Figure 3G), thus in osjar1-3 the Tos17 insertion causes premature termination of transcription of OsJAR1 mRNA. To summarize, we conclude that transcription of the OsJAR1 is disrupted in osjar1-2 and osjar1-3 and that no active OsJAR1 protein can be formed. Rescue of osjar1 phenotype by overexpression of OsJAR1 To confirm that the osjar1 phenotype was indeed due to Tos17 insertions in OsJAR1, we set up a complementation test by overexpressing a wild type copy of the OsJAR1 cDNA under control of the rice Act1 promoter (construct ProActin::OsJAR1) in stably transformed rice plants. When osjar1-2 was transformed with construct ProActin::OsJAR1, the anther indehiscence phenotype was rescued in the resulting plants. Three ProActin::OsJAR1 transgenic lines with a single T-DNA insertion were selected and they showed a normal floret opening and closure phenotype. Normal seed set was produced as well (Figure 4B). Northern blot analysis of the three single T-DNA transgenic lines showed expression of wild type OsJAR1 mRNA (Figure 4C). Because the transgenic lines were
Figure 4. Rescue of the osjar1-2 phenotype by transgenic overexpression of OsJAR1.A. Effect of MeJA on the induction of floret opening of Actin::OsJAR1 transgenic lines (□ and ×) and wild type (△and ◇). Each data point represents the mean (with S.D.) of ten replicates. Concentrations of MeJA were 0 mM (□ and △) and 0.4 mM (◇ and ×). B. Morphology of grains (from left to right) from wild type, osjar1-2 and Actin::OsJAR1 plants, respectively. C. Northern blot analysis of OsJAR1 in osjar1-2, wild type and Actin::OsJAR1 transgenic lines. WT, the wild type; A173, A61 and A143 represent three independent single T-DNA insertion lines with construct ProActin::OsJAR1. Hybridization with 17S rRNA was to check the loading of total RNA.

Functions of OsJAR1 in floret opening and anther dehiscence
86
made via a tissue culture-based protocol that in principle can activate Tos17 elements, Southern blot analysis was performed to verify any Tos17 activity from OsJAR1. However, the Tos17 element was still inserted at the same position in the transgenic osjar1-2/ProActin::OsJAR1 lines tested (data not shown). OsJAR1 mRNA is expressed in floral and vegetative organs The temporal and spatial expression pattern of the OsJAR1 gene was analyzed in further detail using an OsJAR1 promoter-β-glucuronidase (ProOsJAR1::GUS) fusion construct. For this, a 4.5 kb promoter fragment was translationally coupled to the GUS reporter gene and introduced in Nipponbare rice via Agrobacterium-mediated transformation. In total, 25 independent transgenic plants were made expressing construct ProOsJAR1::GUS. GUS reporter gene activity was detected in leaf blade, sheath, root, floret and stem of mature plants (Figure 5A-5D). In the fibrous root system, ProOsJAR1::GUS was predominantly expressed in young adventitious root rather than older adventitious roots (Figure 5A). Strongest GUS activity was found on the base of new lateral roots and no GUS activity occurred at the tip of the root (Figure 5A). Analysis of cross section of ProOsJAR1::GUS main root showed that ProOsJAR1::GUS is more abundant in vascular cylinder than in cortex cells (exodermis, sclerenchymatous layer cells, cortical parenchyma cells and endodermis) and epidermis (Figure 5O). In vascular cylinder, no GUS activity was detected in protoxylem and metaxylem (mxI and mxII); strong GUS activity was detected in pericycle, parenchyma cells and primary phloem; primary phloem has the strongest ProOsJAR1::GUS expression (Figure 5P). ProOsJAR1::GUS show similar expression patterns in mature leaf blade, leaf sheath and stem. That is, strongest expression was detected in the vascular bundle of these three tissues (Figure 5E-5L). In addition to the vascular bundle, weak GUS activity was detected in mesophyll cells and bulliform cell, but not in the sclerenchymatous layer cells in mature leaf blade (Figure 5F). In mature leaf sheath and stem, GUS activity was detected in the parenchyma cells between two vascular bundles (Figure 5H, 5J, and 5K). In the vascular bundle of these three tissues, GUS activity was detected in bundle sheath, protoxylem parenchyma cells, metaxylem parenchyma cells, sieve-tube cells, companion cells and metaphloem parenchyma cells as well (Figure 5L-5N). Companion cells and metaphloem parenchyma cells has the strongest ProOsJAR1::GUS expression. No GUS activity was detected in the late metaxylem (mxII) and weak GUS activity was detected in early metaxylem (mxI) (Figure 5M-5N).
The ProOsJAR1::GUS construct showed a dynamic expression pattern in floret during development. In the early developmental stage (one week before floret opening), GUS activity was detected in glumes only, but not in stamen, pistil and lodicules (Figure 6A). When florets get older (2~4 days before anther dehiscence), GUS activity was detected in the lodicules, the upper part of ovary and the connective of anthers adjacent to the filament (Figure 6B). Up to one or two days before floret opening till the end of floret opening, obvious GUS activity was detected in all parts of the connective tissue of the anthers (Figure 6C, 6D and 6E). Furthermore, the part of filament close to the anther showed ProOsJAR1::GUS expression during anther dehiscence (Figure 6D) and GUS activity was found in the whole filament at the end of anther dehiscence (Figure 6E). The expression of ProOsJAR1::GUS construct in stamen becomes more obvious by comparing anthers from different developmental stages together (Figure 6O). By checking cross sections of anthers from different developmental stages, we realized that the majority of anthers in the early or middle

Chapter 3
87
Figure 5. Histochemical localization of ProOsJAR1::GUS in root, leaf sheath and stem of transgenic rice plants. A. Adventitious roots. B. Internodes. C. Segment of leaf blade. D. Segment of leaf sheath. E. Transverse section through the region of main vein of a mature leaf blade. F. Transverse section through the region of lateral large vascular bundle of a mature leaf blade. G. Close-up of main vascular bundle in (E). H. Transverse section through the first internode showing the internode wall. I. Close-up of large vascular bundle in (H). J. Transverse section through a leaf sheath. K. Transverse section through the midrib region of a leaf sheath. L. Close-up of the large vascular bundle in (K). M, N. Details of the large vascular bundle in (L). O. Transverse section through a primary root 2 mm from the tip. P. Details of central vascular cylinder in (O). bf, bulliform cell; bs, bundle sheath; cc, companion cells; cpc, cortical parenchyma cells ,ed, endodermis; ep, epidermis; ex, exodermis; mc, mesophyll cells; mpp, metaphloem parenchyma cells; mvb, main vascular bundle; mxI, early metaxylem; mxII, late metaxylem; mxp, metaxylem parenchyma cells; pc, pericycle; pp, primary phloem; pxl, protoxylem lacuna; pxp, protoxylem parenchyma cells; px, protoxylem; sc, sieve-tube cells; sl, sclerenchymatous layer cells; vb, vascular bundle; vc, vascular cylinder; vpc, parenchyma cells in vascular bundle. Bar (A) = 0.2 mm; Bars (B-D) = 1 mm; Bars (E, K) = 100 μm; Bars (F, H, Q) = 50 μm; Bars (G, I, L) = 25 μm; Bar (J) = 250 μm; Bars (M, N, P) = 15 μm. microspore stage had no ProOsJAR1::GUS expression (Figure 6F), but there is still a minority of anthers in the early or middle microspore stage showing weak GUS activity in the vascular bundle cells (VBC) of the anther connective tissue (Figure 6K). Obvious GUS activity was found in the

Functions of OsJAR1 in floret opening and anther dehiscence
88
Figure 6. Localization of ProOsJAR1::GUS expression in florets of transgenic rice plants. A-E. Floret and stamen at one week before floret opening (A), the stage of 2~4 days before anther dehiscence (B), the early stage of anther dehiscence (C), the middle stage of anther dehiscence (D) and the end of anther dehiscence (E). F-K. Transverse sections of anther in different developmental stages, which is microspore stage (F, K), the stage of 2~4 days before anther dehiscence (G), the stage of 1~2 days before anther dehiscence (H), the early stage of anther dehiscence (I) and the end of anther dehiscence (J). L Details of the vascular bundle in the stage just before anther dehiscence. M-N. In situ hybridization of OsJAR1 transcript in the transverse sections across the region of the connective tissue of the anther obtained from a wild type floret just before dehiscence. (M) Antisense probe. (N) Sense probe. O. Stamen in different developmental stages. an, anther; ft, filament; pa, palea; lo, lodicules; V, vascular bundle. Bars (A-E) = 1 mm; Bars (F-J) = 50 μm; Bars (K-O) = 25 μm. VBC from the late microspore stage onwards (Figure 6G). Compared to the late microspore stage, a more intense GUS activity was detected in the VBC during the pollen mature stage (Figure 6H). During the early stage of floret opening, GUS activity was detected not only in the VBC, but also in the septum (Figure 6I). Till the end of anther dehiscence, GUS activity was detected in the vascular bundle of the anther connective tissue, septum and anther wall as well (Figure 6J). Strong expression of construct ProOsJAR1::GUS in the VBC of the anther connective tissue is consistent with the results obtained from in situ hybridization of cross sections of anthers with an anti-sense OsJAR1 RNA probe (Figure 6M). A sense probe did not show any positive signal in the VBC of the anther connective tissue (Figure 6N).
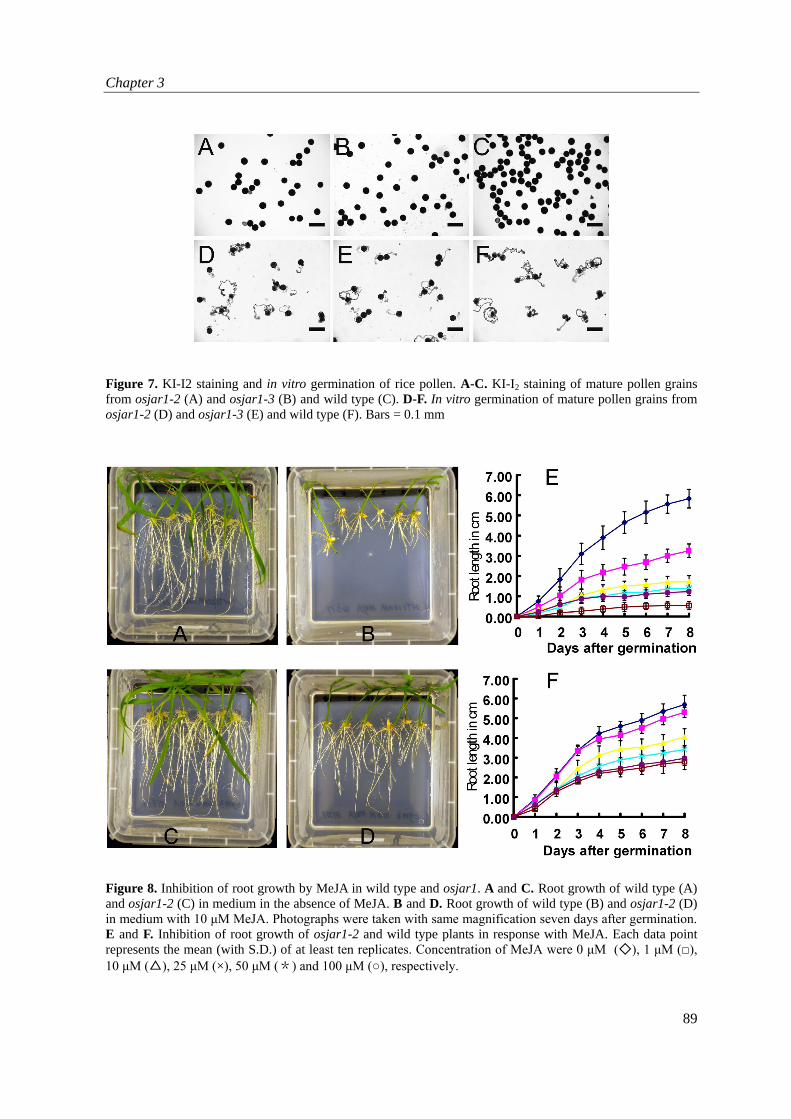
osjar1 has normal pollen viability and pistil perceptiveness Plants with the osjar1-2 (30 plants tested) and osjar1-3 (10 plants tested) mutations showed 9.1% fertility (ranging from 7.5% to 14.5%) whereas wild type plants showed 98% fertility. Fertility was measured as the ratio between filled grains and total number of florets. In order to find out which defects in flower organs resulted in the observed lower fertility of osjar1, crossing assays and pollen viability tests were performed. After crossing of 25 osjar1-2 pistils with mature pollen from control Nipponbare plants, 25 seeds were obtained. All the progeny of the crossed lines showed no
Chapter 3
89
Figure 7. KI-I2 staining and in vitro germination of rice pollen. A-C. KI-I2 staining of mature pollen grains from osjar1-2 (A) and osjar1-3 (B) and wild type (C). D-F. In vitro germination of mature pollen grains from osjar1-2 (D) and osjar1-3 (E) and wild type (F). Bars = 0.1 mm
Figure 8. Inhibition of root growth by MeJA in wild type and osjar1. A and C. Root growth of wild type (A) and osjar1-2 (C) in medium in the absence of MeJA. B and D. Root growth of wild type (B) and osjar1-2 (D) in medium with 10 μM MeJA. Photographs were taken with same magnification seven days after germination. E and F. Inhibition of root growth of osjar1-2 and wild type plants in response with MeJA. Each data point represents the mean (with S.D.) of at least ten replicates. Concentration of MeJA were 0 μM (◇), 1 μM (□), 10 μM (△), 25 μM (×), 50 μM (*) and 100 μM (○), respectively.

Functions of OsJAR1 in floret opening and anther dehiscence
90
differences in phenotype compared to control plants, indicating that the stigma of osjar1-2 is able to accept pollen for fertilization. Furthermore, in vitro pollen germination and iodine stainings were carried out to check viability of mature osjar1-2 pollen. Results show that 95.6% (478/500) of wild type pollen and 93% (465/500) pollen from osjar1-2 were stained with iodine (Figure 7A-7C). In the pollen germination assay, 90.8% (590/650) and 86.8% (564/650) of the pollen from control and osjar1-2 plants, respectively, germinated two hours after incubation in a pollen germination medium (Figure 7D-7E). Essentially, the same results were obtained for mutant osjar1-3, which suggested that pollen of osjar1-2 and osjar1-3 are able to pollinate the stigma and that osjar1 mutants have a normal, functioning and receptive pistil. osjar1-2 shows reduced root growth inhibition on MeJA It has been reported that MeJA inhibits root growth in Arabidopsis (Staswick et al., 1992). In contrast to wild type, the Arabidopsis jar1-1 loss-of-function mutant exhibits moderate insensitivity to MeJA in a root growth inhibition assay (Staswick et al., 1992). We examined whether root growth of rice is sensitive too to MeJA by germinating rice seeds on media containing various concentrations of MeJA. As shown in Figure 8A-8D, seven days after germination, control seedlings showed 45% and 70% inhibition of primary root growth on medium with 1 μM and 10 μM MeJA, respectively. On higher concentrations MeJA (25, 50 or 100 μM), a more serious inhibition of primary root growth occurred (Figure 8E). In contrast to the obvious inhibition of root growth of control plants, osjar1-2 roots exhibited considerable MeJA-resistance and showed only 7% and 30% growth inhibition on media with 1 μM and 10 μM MeJA, respectively. Addition of 100 μM MeJA resulted in 50% root growth inhibition in osjar1-2 seedlings (Figure 8F). osjar1 mutants show reduced resistance to the blast fungus Magnaporthe oryzae JA is implicated in disease resistance mechanisms in plants (Berger, 2002; O'Donnell et al., 2003; Jones and Dangl, 2006; Qiu et al., 2007) and several defects in disease resistance were found in the Arabidopsis jar1 mutant (Staswick et al., 1998; Ferrari et al., 2003; Traw et al., 2003). We investigated the possible role of the OsJAR1 gene in resistance to the fungus M. oryzae which is the causative agent of rice blast disease. When inoculated with the avirulent strain CL367, mutants osjar1-2 and osjar1-3 showed larger necrotic lesions than wild type plants (Figure 9A). Consistently, ProActin::OsJAR1 lines overexpressing the OsJAR1 gene in cultivar Nipponbare showed reduced susceptibility to the virulent isolate FR13 (Figure 9B) which normally is pathogenic to Nipponbare rice. This suggests that the OsJAR1 gene is required for blast fungus resistance. We further investigated regulation of OsJAR1 by following the expression of the ProOsJAR1::GUS construct in infected plants. Surprisingly, GUS activity decreased around the lesions triggered by the avirulent isolate CL367 (Figure 9C). In virulent-infected plants (isolate FR13), GUS activity was also reduced with the notable exception of the immediate surroundings of the lesions (Figure 9C). The results show that OsJAR1 is tightly controlled by infection with M. oryzae with contrasting effects depending on the distance to the infection site. MeJA and JA-mediated induction of floret opening in osjar1 is impaired It has been reported that the opening of florets in rice can be induced by different concentrations of MeJA (Zeng et al., 1999). We checked if opening of florets from osjar1-2 were also inducible by

Chapter 3
91
Figure 9. OsJAR1 is a positive regulator of resistance towards rice blast disease. A. The insertion mutant osjar1-3 shows reduced resistance (enhanced necrosis) to the avirulent strain M. oryzae CL367. Similar results were obtained with osjar1-2. B. The over-expression line ProActin::OsJAR1 shows reduced susceptibility (grayish lesions) to the virulent strain M. oryzae FR13. Similar results were obtained with five independent transgenic lines. C. Expression of construct ProOsJAR1::GUS is regulated by infection with M. oryzae. (a) Non-infected plants only treated with gelatin used for resuspending M. oryzae spores. GUS activity is detected in the entire leaf blade. (b) Plants inoculated with the avirulent strain M. oryzae CL367. The brown spots are typical for a resistance response. GUS activity is reduced around the necrotic lesions. (c) Plants inoculated with the virulent strain M. oryzae FR13. The non-stained around the infection sites are surrounded by intense GUS staining represent typical susceptible lesions after infection with M. oryzae. In contrast, GUS activity is reduced in tissues distant from the infection sites. All experiments were repeated three times on four independent transgenic lines.
Figure 10. Effect of MeJA on induction of floret opening in wild type and osjar1-2. A. Control plants. B. osjar1-2 mutant. Each data point represents the mean (with S.D.) of at least ten replicates. Panicles were treated with 0, 0.04, 0.4 and 4 mM MeJA, respectively. Concentration of MeJA were 0 μM (◇), 1 μM (□), 10 μM (△), 25 μM (×), 50 μM (*) and 100 μM (○), respectively.

Functions of OsJAR1 in floret opening and anther dehiscence
92
MeJA. Prior to treatments with MeJA, any already flowering florets were removed. This experiment was taken three hours in advance of the predicted opening time of florets from control Nipponbare plants. Indeed, floret opening in control plants was inducible by MeJA. The first opening of florets on control plants occurred in 17 to 25 minutes after MeJA treatment, depending on the different concentrations of MeJA. A higher concentration of MeJA was inducing more floret opening per panicle (Figure 10A). Surprisingly, florets from osjar1-2 plants were still closed, even 90 minutes after treatment in various tested MeJA concentrations (Figure 10B). Essentially, similar results were obtained with osjar1-3. In summary, the results show that the MeJA signaling pathway involved in floret opening is defective in the osjar1 mutants. Furthermore, by treating transgenic ProActin::OsJAR1 lines with MeJA during anthesis, responsiveness of floret opening and closure by MeJA was fully restored, which confirmed that the MeJA-mediated pathway for floret opening was rescued by overexpression of the wild type OsJAR1 cDNA in osjar1-2 (Figure 4A). We also used JA to treat osjar1-2 and wild type flowers. Similar results were observed in which the induction of floret opening in wild type was impaired in osjar1 too (data not shown). Rescue of the phenotype of osjar1-2 by exogenous JA treatment One of the main characteristics of the phenotype of osjar1 is that the lodicules keep on swelling even several days after opening of the florets started. We checked if JA treatment could rescue the phenotype of the osjar1 lodicule. Ten flowers from two osjar1-2 plants at 1 DAF were treated with JA and ten other osjar1-2 flowers were treated with DMSO as control treatment. The treatment was started at 9 am and no obvious change was observed throughout the daytime till 9 pm. However, all JA-treated lodicules were found withered (Figure 11B) and the flowers closed 24 hours after treatment, whereas the control osjar1-2 lodicules did not (Figure 11C) and florets stayed opened. OsJAR1 is responsive to wounding and exogenous JA/MeJA Riemann et al. (2008) described that the expression of OsJAR1 to responsive to exogenous application of MeJA within 45 minutes. Since osjar1 exhibits insensitivity towards MeJA treatment in different assays with a variety of timeframes of treatments, we further investigated the temporal responsiveness of OsJAR1 expression by exogenous JA and MeJA application. Northern blot analysis was used to monitor OsJAR1 expression in segments of wild type seedling leaves cut into two centimeter pieces and incubated with JA and MeJA respectively. DMSO and water were used as controls. Initially, we checked the effects on OsJAR1 expression within two hours of incubation. Accumulation of OsJAR1 mRNA started within half an hour in all treatments including the control, which indicates that wounding also triggers OsJAR1 expression. However, MeJA application extra induced OsJAR1 expression at 0.5 hr (Figure 12). For JA, no obvious differences compared to the controls were discovered. To further study the temporal regulation of OsJAR1, we analyzed the expression profile at time points ranging from 3 to 24 hr after starting the experiment. Extra inductions were both found with JA and MeJA treatment at 3hr. After that, the mRNA level OsJAR1 of all treatments decreased at 6 hr to the original level but increased again to a comparatively higher level than zero from 12 to 24 hr. Thus, the expression may be controlled by the circadian rhythm too.

Chapter 3
93
Figure 11. Rescue of the osjar1 flower phenotype by JA treatment. Ten lodicules of osjar1-2 flowers at 1 DAF were treated with JA by pipetting 3 µl of 5mM JA onto the lodicules very carefully. Ten osjar1-2 flowers were treated with 4% DMSO as controls. A. osjar1-2 flower two days after flowering start. B. osjar1-2 flower 24 hours after treated with JA. C. osjar1-2 flower 24 hours after treated with 4% DMSO.
Figure 12. OsJAR1 is responsive to wound and application with JA/MeJA. Northern blot analysis of leaf segments from three-week-old seedlings after treatment with 100 μM JA, 0.4 mM MeJA, 1% DMSO and water controls in a time-dependent manner under continuous light. Total RNA was extracted at the time points (in hours) indicated above the lanes. The lane indicated with 0 indicates the start of the experiment.
OsJAR1 encodes an enzyme catalyzing the formation of JA-amino acid conjugates OsJAR1 is predicted to encode a protein of 581 amino acids that belongs to the plant-specific GH3 family. Database searches and phylogenetic analysis showed that OsJAR1 is the most closest to NaJAR4 and AtJAR1 from tobacco and Arabidopsis respectively (Wang et al., 2007). Alignment of the deduced OsJAR1 amino acid sequence showed that it has ~61% identity (79% similarity) and ~67% identity (83% similarity) with AtJAR1 and NaJAR4 respectively, which implicated that OsJAR1 might have similar functions like AtJAR1 and NaJAR4. Both AtJAR1 and NaJAR4 encode a JA-Ile synthetase capable of conjugating JA to isoleucine (Staswick and Tiryaki, 2004; Kang et al., 2006). To study if OsJAR1 protein has this function too, recombinant protein was expressed as a glutathione S-transferase (GST) fusion in E. coli. After purification, the putative activity of the recombinant GST-OsJAR1 protein was assayed in independent enzymatic reactions with all 20 different L-amino acids. Reactions without GST-OsJAR1 protein were set up as negative controls. After analysis by thin-layer chromatography (TLC), eight amino acids were found conjugated to (±)-JA in presence of GST-OsJAR1. Figure 13 shows that the putative conjugates for JA-Ala, JA-Cys,

Functions of OsJAR1 in floret opening and anther dehiscence
94
Figure 13. Conjugation of eight different amino acids with JA by recombinant GST-OsJAR1 protein. Reactions with JA and each amino acid (three letter abbreviations) were analyzed by TLC and stained for JA with vanillin dye. (+) and (-) represent with and without recombinant protein respectively.
Figure 14. Variation of in vitro synthesized JA-AA conjugates and measurement of JA-AA content in leaves from wild-type and osjar1-2 plants. A. LC-MS of enzymatic synthesized JA-AA. B and C UPLC-MS measurement of JA-AA content in flag leaves from wild type (B) and osjar1-2 (C) plants, five hours after 10mM JA treatment. D and E UPLC-MS measurement of JA-AA content in flag leaves from wild-type (D) and osjar1-2 (E).

Chapter 3
95
JA-Ile, JA-Leu, JA-Met, JA-Phe, JA-Trp and JA-Val all have reduced mobility compared to JA. There are also obvious differences in intensity of the JA spots when either of the eight conjugates were formed, which indicates the reduction of free JA because of the conversion into conjugated forms.
Six of the in vitro conjugation products of JA with amino acids (Ile, Leu, Val, Met, Phe, Trp) were further analysed using ultra performance liquid chromatography coupled to tandem mass spectrometry (UPLC-MS/MS). For each conjugate, five or six peaks of variable intensity were observed in the corresponding chromatograms (Figure 14A). MS/MS fragmentation of the individual peaks produced very similar fragmentation spectra, strongly suggesting that the peaks belong to (diastereo) isomeric compounds. So far the chromatographic separation of jasmonate-N-amino acid isomers has only been described for chiral HPLC columns and has not been possible with classical C18 reversed phase columns (Kramell et al., 1999). With UPLC a much better separation of closely eluting compounds is achieved. Without comparison with authentic standards it is very difficult to assign the correct chemical structure to each of the isomers. The synthetic (±) JA that was used is a mixture of a number of (stereo) isomers. According to the UPLC-MS/MS chromatogram, the main constituent at 4.85 min is a mixture of the (3R,7R) and (3S,7S)-2-Z-pentenyl stereo isomers (approximately 89% according to LC-MS/MS analysis). The (3R,7S) and (3S,7R)-2-Z-pentenyl stereo-isomers (9%) elute at 4.95 min, while the minor peak (2%) at 5.00 min could be a mixture of the (3R,7R) and (3S,7S)-2-E-pentenyl stereo isomers (Holbrook et al., 1997). The relative concentration of the JA-AA isomers produced will be a function of the starting concentration and of the substrate specificity of the recombinant GST-OsJAR1 protein towards each of the JA stereo-isomers. For all JA-AA conjugates isomer 3 (assigned on the basis of elution order in the chromatograms) is the major metabolite formed. It is likely that this is the (3R, 7R)-jasmonate-N-amino acid isomer (Vick and Zimmerman, 1984; Mueller and Brodschelm, 1994; Holbrook et al., 1997). Reduced levels of JA-AA conjugates in leaves from osjar1-2 plants Based on the observation that recombinant OsJAR1 protein is able to catalyze synthesis of eight different JA-AA conjugates, leaves from wild type and osjar1-2 plants were analyzed for JA-AA content using UPLC-MS/MS. In the first experiment we analyzed leaf materials treated with 10mM JA in order to confirm if this treatment would affect levels of JA-AA conjugates. In the MRM chromatograms of extracts collected from wild type leaves treated with (±) JA, four or five isomeric jasmonate conjugates were detected that correspond to Ile, Leu, Val, Met and Phe, conjugation of JA to Trp could not be detected (Figure 14B). There are clear differences between extracts obtained from wild type and osjar1-2 (Figure 14B and C). Isomer 3 of JA-Ile and JA-Leu dominate in the wild type extract. Significant amounts of JA-Val isomer 3 and isomer 1 of JA-Leu and JA-Val are present as well, while isomer 4 is absent for all AAs. However, in the same leaf tissue from osjar1-2 the overall concentration of jasmonate-N-amino acid conjugates is significantly lower than in wild type (aproximately 5-fold). This is mainly due to the fact that the reduced concentration of isomer 3 of JA-Ile, JA-Leu and JA-Val in osjar1-2 compared to wild type (25-fold reduction of JA-Ile and 10-fold reduction of JA-Leu and JA-Val). The other isomers are present more or less in the same quantities as in wild type, with isomer 1 of JA-Leu and JA-Val as predominant conjugates. In the chromatograms of extracts obtained from untreated wild type and osjar1-2 leaves, the same isomers

Functions of OsJAR1 in floret opening and anther dehiscence
96
of JA-Ile, JA-Leu, JA-Val and JA-Phe conjugates were detected as in the JA-treated leaf extracts, but in concentrations more than 100-fold reduced (Figure 14D and E). Thus, JA treatment induces accumulation of JA-AA conjugates in rice flag leaves. The distribution of isomers and relative abundance of JA-AAs in untreated wild type leaves is very similar to that of JA-treated wild type. In both samples, isomer 3 of JA-Ile and JA-Leu are the most abundant JA-conjugates. The conjugate profile for the extract from osjar1-2 bears a closer resemblance to that of wild type rather than that of JA-treated osjar1-2. The high preference for isomer 1 JA-AA conjugates in leaves from osjar1-2, treated with JA, is not found in untreated osjar1-2. JA-Ile and JA-Val isomer 1 together with JA-Ile isomer 5 are most abundant in extracts from osjar1-2 leaves. On average the levels of JA-AA conjugates are three times lower in osjar1-2 extracts. Discussion JA is a growth regulator and has been shown to control a wide range of processes in plants, ranging from responses to biotic and abiotic stresses to various developmental and environmental response processes (Ellis and Turner, 2001; Berger, 2002; Mei et al., 2006; Acosta et al., 2009). However, the molecular mechanisms involved in these processes are yet poorly understood. Previously, the OsJAR1 gene was reported to be regulated by exogenous MeJA and two different photoreceptors, phytochrome A (phyA) and phytochrome B (phyB). The corresponding mutants, osjar1-2 and osjar1-3 are characterized by a very obvious floret phenotype with open husks and also includes low-fertility and malformed seeds (Riemann et al., 2008). In this Chapter, we studied the phenotype of osjar1 and the molecular and physiological actions of OsJAR1 in more detail and show that this protein has a biochemical function as an enzyme conjugating amino acids to JA resulting in active hormones which is confirmed by the finding that JA conjugates are lower in osjar1-2 plants. The lowered fertility was shown to be caused by a strongly reduced anther dehiscence and not be any obvious defect of the pistil since these can be pollinated normally. The osjar1 pollen itself could also be ruled out as the cause of reduced fertility since they were shown to be perfectly viable in pollen assays. We tested for disease susceptibility of plants misexpressing OsJAR1 and also showed a role for this gene in resistance towards the rice blast fungus Magnaporthe oryzae. Finally, we postulate that the lowered levels of the JA conjugates cause the open husk phenotype of the two osjar1 alleles studied here. The genetic basis of the phenotype of osjar1-2 and osjar1-3 In rice, opening and closure of the florets results from swelling of lodicules which pushes away the palea from the lemma and subsequent withering, respectively (Matsuo and Hoshikawa, 1993). Normally, rice florets close again in less than 90 minutes after opening because the lodicules are withering quickly after swelling. However, in both Tos17 alleles for osjar1 that we studied, the lodicules remained swollen even five days after floret opening started. Consequently, movement of the palea and lemma and thus, closure of the flower are impaired. Other phenotypical aspects of osjar1 are that the seeds are narrower and more elongated and that the style remains attached. The fertility is much lower than in wild type Nipponbare plants grown under the same conditions. This phenotype is observed in both alleles of osjar1 which both segregate as a recessive allele. On northern blots we observed abnormal osjar1 transcripts for both alleles. For osjar1-2 in line NC2728,

Chapter 3
97
expression was much reduced and the length of the band correlated with the insertion of the 4.5 kb Tos17 retrotransposon in exon 3. For osjar1-3 in line ND0051, we observed that the osjar1 mRNA was shorter which may be caused by premature termination of transcription or incorrect splicing. We also identified a third Tos17 allele in line NE8577, but this line has no observable phenotype. This Tos17 insert is in the fourth intron and usually such Tos17 inserts are spliced out together with the intron without having too many effects on transcription. Strong support for the hypothesis that the two mutant lines with the osjar1 phenotype are caused by Tos17 insertions in OsJAR1 and thereby impairing the function of the gene product is by the experiment in which we retransformed osjar1-2 with an actin promoter-driven OsJAR1 overexpression construct. Five independent transgenic lines were verified for overexpression of OsJAR1 and showed complementation of the osjar1 phenotype. The seeds of these transgenic plants could not be distinguished from those from wild type Nipponbare plants and the induction of opening of the florets by MeJA is restored as well without any other observable effects on the phenotype Because Tos17 is activated by tissue culture we confirmed by Southern blotting that Tos17 is still present in all the transgenic lines. OsJAR1 has functions as a JA-AA synthetase OsJAR1 was confirmed to encode a JA-AA synthetase by TLC and LC-MS/MS assays. For this, recombinant OsJAR1 protein was purified from E. coli and was shown to catalyze the formation of JA-Phe, JA-Ile, JA-Leu, JA-Met, JA-Trp, JA-Val, JA-Cys and JA-Ala in in vitro reactions analyzed with TLC assays. All eight amino acids that were conjugated by OsJAR1 to JA belong to the so-called non-polar group (Alberts et al., 2008), indicating OsJAR1 has a certain substrate preference. The identity of JA-Ile, isolated from TLC plates, was further confirmed by LC-MS/MS analysis. Furthermore, UPLC-MS/MS analysis was used to determine JA-AA levels in wild type and osjar1 plants. In total, five conjugates were confirmed that corresponded to Ile, Leu, Val, Met and Phe of which JA-Leu and JA-Ile are the dominant forms. Unlike the jar1 mutants in Arabidopsis which only show an obvious decrease of JA-Ile (Staswick and Tiryaki, 2004), in osjar1-2 plants we observed significant reduction in levels of five out of six measured JA-AA conjugates and in particular the level of JA-Ile is 25-fold lower. Also JA-Leu and JA-Val levels are strongly affected with a ten-fold decrease. It is therefore very likely that the low levels of JA-Ile and JA-Leu or JA-Val are responsible for the observed phenotype of the open-staying flowers and delayed anther dehiscence. There are a number of other studies that implicate a function of JA in these processes. The Arabidopsis mutants fad3-2 fad7-2 fad8 (McConn and Browse, 1996) and opr3 (Stintzi and Browse, 2000) have defects in JA biosynthesis and all have lowered levels of JA. In these mutants, flower organs initially develop normally, but like with osjar1-2 and osjar1-3, anther locules do not dehisce at the right time of flower opening. A major difference is that the filaments do not elongate normally and pollen grains are inviable whereas these are unaffected in osjar1-2 and osjar1-3. An experiment involving treatment of leaves from osjar1-2 and wild type plants with JA further verified the role of OsJAR1 as a JA-AA synthetase in vivo. UPLC-MS/MS results showed that this treatment resulted in a strong accumulation of JA-AA conjugates and that in wild type leaves, isomer 3 of JA-Ile was 25-fold higher than in osjar1-2 and also JA-Leu and JA-Val were 10-fold higher. Although the accumulation of JA-AA conjugates was much less in osjar1-2, the level of JA-AA conjugates also increased considerably compared to the untreated leaves, indicating there are also other, redundant JA-AA synthetase expressed and active. The induction of the level of isomer 1 JA-Leu

Functions of OsJAR1 in floret opening and anther dehiscence
98
and JA-Val in osjar1-2 and wild type was not so different, which further demonstrated the activity of other JA-AA synthetases. Taken together, from the results we can conclude that OsJAR1 has substrate preference and is mainly responsible for the conjugation of isomer 3 JA-AA molecules. In the experiment of flower opening induced by JA treatment, the induction was found impaired in osjar1-2 and osjar1-3. Because the most apparent difference in the level of JA-AA conjugates between wild type and osjar1-2 was found in isomer 3 JA-Ile and JA-Leu, we speculate that these two are the most essential conjugates involved in flower opening and closure and anther dehiscence. Treatment of osjar1-2 lodicules with exogenous JA can rescue the lodicule phenotype in osjar1-2 yet needs a comparatively long period (24 hours), indicating that other JA-AA isomers or even other JA-related pathways may also be involved in flower opening but it could also be that it is difficult for JA to get into the lodicule. Function of JA-AA conjugates in plants Experiments in Arabidopsis show that JA-Ile is an important signaling molecule in the COI1-mediated JA-response, and acts by promoting the physical interaction between COI1 and JAZ1 proteins and initiates SCFCOI1 dependent proteasome degradation of JAZ repressor proteins. In turn, this results in release of the transcription factor MYC2 (and probably other proteins) from the MYC2-JAZ complex, and subsequent activation of transcriptional activation of JA response target genes (Farmer, 2007; Thines et al., 2007). Since the JA-Ile content in leaves of osjar1 plants is much lower, it is very well likely that this will have effects on downstream genes involved in lodicule swelling and withering, anther dehiscence and seed-shape. Effects on gene expression also has been shown for the Arabidopsis opr3 mutant where transcriptome studies of the stamen (Mandaokar et al., 2006) showed groups of genes that are specifically induced or repressed by JA, amongst which are many transcription factors and transporters. Recently, Staswick (2009) reported that JA-Trp functions as an endogenous auxin inhibitor in Arabidopsis and that exogenous JA-Trp application caused agravitropic root growth in seedlings. The biological functions of other JA-AA conjugates are still unclear. It is known that the homeostasis of an active hormone is rather important for normal plant development and defence responses and that the homeostasis of active auxin is maintained by IAA-AA hydrolases and IAA-amido synthetases (Staswick et al., 2005; Schuller and Ludwig-Muller, 2006). It is very well possible that JA-AA conjugates especially JA-Ile and JA-Trp and associated biosynthesis genes including OsJAR1 and others have an important function in the homeostasis of active jasmonates in planta. If so, there are probably yet unknown JA-AA hydrolases acting together with the OsJAR1 protein in the conversion of active jasmonates into inactive JA-AA conjugates, resulting in the homeostasis of active jasmonates in plants. The lower fertility of osjar1 mutants is caused by a reduction of anther dehiscence Although osjar1 mutant has a 7.5-14.5% lower seedset because of flower sterility, it showed normal pistil receptiveness in crossing experiments. Furthermore, KI-I2 stainings in in vitro pollen germination assays showed that nothing is wrong with viability of the pollen themselves. However, we found that in fact anther dehiscence is much reduced in osjar1 plants. While normally anthers are dehiscing and withering after pollen release, most of the osjar1 anthers remain closed or only partially opened although they seem to develop normally and are completely and normally filled with pollen grains. Dehiscence of the anthers is the result of a multistep process that starts with

Chapter 3
99
swelling of the pollen as the driving force for the final rupture of the septum, the splitting of the stomium and the locule wall (Matsui et al., 1999). However, the force exerted by the swollen pollen only is not sufficient for opening of the theca. At the stomium, small groups of cells are actively dissociated from each other thereby contributing to anther opening and pollen release (Keijzer, 1987). Enzymes, such as polygalacturonases are likely to be involved in this process (Ogawa et al., 2009). Another important process during anther development and dehiscence is PCD (Programmed Cell Death). In tomato it has been shown that cells of the septum, connective tissue, middle layer and endothecium and epidermal cells surrounding the stomium show features consistent with progression through PCD (Senatore et al., 2009). Studies with Arabidopsis mutants have shown that the different steps in anther dehiscence are not necessarily linked, as mutants have been described that lack pollen and still display anther dehiscence (Aarts et al., 1993, 1997). We showed that anther tissue and pollen of osjar1 consist of the same cell layers and structures as wild type anthers and the locules are completely filled with trapped but viable pollen, as the crossing experiments show. One obvious morphologic difference on the cellular level, however, is the more irregular cell shape in septum, anther wall and around the vascular bundle and we hypothesize that this somehow may impair dehiscence. On the other hand, we cannot exclude that JA-Ile synthesized by OsJAR1 is involved in regulation of enzymes involved in dissociation of stomium cells or in PCD. Light signaling and JA regulate OsJAR1 and floret opening in rice Anthesis in rice is confined to a specific period of the day and is thus tightly regulated. On the contrary, osjar1 florets open randomly even at night and show loss of timing control of anthesis. The homolog of OsJAR1 in Arabidopsis, AtJAR1/Fin219 has been reported to be involved in phyA-mediated FR light signaling and to interact with a glutathione S-transferase, FIP1, to regulate cell elongation and flowering in response to light (Chen et al., 2007). The loss-of-function mutant fin219 exhibited a long hypocotyl phenotype under continuous far-red (cFR) and is slightly delayed in flowering (Hsieh et al., 2000). Microarray studies with rar-red light revealed that FIN219 may act downstream of FHY1, FAR and FHY3 and controls the expression of smaller sets of genes (Wang et al., 2002). Riemann et al. (2008) reported that phytochrome A (phyA) and phytochrome B (phyB) act redundantly to induce the accumulation of OsJAR1 mRNA by red light. Our data shows that OsJAR1 has a dynamic expression profile in florets during floret development and anther dehiscence. The loss-of-function mutant osjar1 exhibits the open husk phenotype and a defect in anther dehiscence, which suggests that dynamic expression of OsJAR1 mRNA in florets and perhaps other tissues, is important for normal anther dehiscence, timing of floret opening and closure and grain development. Taking all data together, we hypothesize that both light and JA are important regulators of flower opening and development in rice and that OsJAR1 plays a significant role in the light induced and JA-mediated regulation of rice anthesis, floret opening/closure and development. A role for OsJAR1 in disease resistance JA-Ile is also studied as a signal molecule in regulation of plant defence such as the responses to herbivore and wounding (Suza and Staswick, 2008; Wang et al., 2008). The most obvious ortholog for OsJAR1 in Arabidopsis, AtJAR1 has been reported to be involved in protection against a variety of stress factors (Staswick et al., 1992, 2002) and NcJAR4 from tobacco has been reported to be connected to MeJA-mediated anti-nutritive defense against the insect Manduca sexta (Kang et al.,

Functions of OsJAR1 in floret opening and anther dehiscence
100
2006). Our studies showed that osjar1 mutants have increased sensitivity to the blast fungus M. oryzae. In agreement with this, when OsJAR1 was overexpressed in transgenic rice lines using the constitutive rice Actin1 promoter, we found that sensitivity was decreased and in fact plants become more resistant than the wild type and osjar1-2. Using the ProOsJAR1::GUS line we show the local effect of M. oryzae infection on OsJAR1 expression. Directly around the infection site, expression strongly increased but elsewhere in the leaf expression decreased. Our results show that OsJAR1 too, like its homologs in Arabidopsis and tobacco, is involved in defence response and resistance. Considering the difference in levels of JA-Ile, JA-Leu and JA-Val between wild type rice and osjar1-2 it is very likely, that these molecules are active in regulation of disease resistance in rice to M. oryzae and possibly other diseases. Another homolog of OsJAR1, GH3-8, has also been implicated in disease resistance (Ding et al., 2008). However, GH3-8 belongs to a different group II set of proteins in a phylogeny analysis (Jain et al., 2006), whereas OsJAR1, AtJAR1 and NcJAR4 belong to group I. In general, GH3 group II proteins encode IAA-amino acid synthetases that conjugate amino acids to IAA and regulate plant growth and development (Staswick et al., 2005) by preventing free IAA accumulation. GH3-8 activates disease resistance in a pathway independent of JA and salicylic acid, thus although the homology with AtJAR1 and OsJAR1 is apparent both groups of genes seem to function in different pathways. Conclusions OsJAR1 is the first gene reported to be related to MeJA-signaling regulating floret opening and anther dehiscence in rice. In this Chapter, we presented a detailed morphological description of osjar1 and used genetic and biochemical data to propose a function of OsJAR1. But there are still several questions remaining. Such as, how does loss-of-function of OsJAR1 cause osjar1 phenotype on the molecular level? Which are the target genes of the pathway controlled by JA-Ile? Why do lodicules in osjar1 mutant keep swelling instead of drooping quickly during anthesis? How does light and JA signaling regulate coordinately rice flowering and anther dehiscence? Further genetic and physiological studies on the osjar1 mutant would provide useful information to elucidate the roles of light, MeJA signaling and OsJAR1 in floret development and plant development and allow us to develop a better mechanistic understanding of JA signaling in plants.
Materials and methods Plant materials and growth conditions Rice cultivars and mutant lines used in this study were Japonica rice (O. sativa L. cv. Nipponbare) and the three mutant lines NC2728, ND0051 and NE8577 containing a Tos17 insertion in OsJAR1. Seeds were surface sterilized and immersed in water for two days in the darkness at 28°C, and then sowed into soil in a greenhouse (at 28°C, 85% relative humidity and a 12-h light/dark period). Plant tissues for nucleic acid extraction collected from seedlings, panicles, leaves, shoot and roots were immediately frozen in liquid nitrogen and stored at –80°C until further processing. JA and MeJA treatments JA (racemic mixture) and MeJA were purchased from Sigma-Aldrich (The Netherlands) and dissolved in DMSO as stocks of 0.4 M, and diluted with water to the concentrations described in the

Chapter 3
101
figure legends. Vector constructions In order to make an OsJAR1 promoter-GUS reporter (ProOsJAR1::GUS) construct, a 5.1 kb OsJAR1 promoter-leader fragment (TIGR Release 5, coordinates on chromosome 5 are 29114475-29119558, LOC_Os05g50890) was obtained by PCR using Phusion polymerase (Bioke, the Netherlands) on wild type genomic DNA from cultivar Nipponbare using the primers OsJAR1Pup-BamHI, 5’-GGATCCAACCGGTACTAACGACGCCTC-3’ and OsJAR1Plow-NcoI, 5’-CCATGGTCATTGTAATGGTAGAATCCTG-3’. The resulting PCR product was cloned into pCR2.1-TOPO (Invitrogen) to form construct pCR2.1-OsJAR1P that was subsequently sequenced (Baseclear, Leiden, The Netherlands). Next, a 4.5 kb OsJAR1 promoter-leader fragment was obtained from pCR2.1-OsJAR1P by EcoRI and NcoI and translationally fused to the start codon of the gusA reporter gene in binary vector pCAMBIA-1391Z (Genbank Accession AF234312), to form construct ProOsJAR1::GUS.
To construct the Actin1 promoter-OsJAR1 fusion construct (ProAct1::OsJAR1) in a binary vector, a 1.5 kb promoter fragment including the entire OsJAR1 coding sequence (LOC_Os05g50890) was obtained by RT-PCR on total mRNA isolated from Nipponbare seedlings with primers OsJAR1Rup-NcoI, 5’-CCATGGCAATGACGATCTGCAGCTGTGAGG-3’ and OsJAR1Rlow-HindIII, 5’-AAGCTTGTTTTCTTCCGGTGGAGACTACAG-3’. The resulting PCR product was cloned into pCR4Blunt-TOPO (Invitrogen) to form construct pCRB4-OsJAR1T that was subsequently sequenced (Baseclear, Leiden, The Netherlands). Next, the 1.5 kb fragment was rescued from pCRB4-OsJAR1T construct by HindIII and NcoI partial digestion. The rice Actin1 promoter was released by XhoI and NcoI from a pSP72 (Promega) based vector ACT-GFP (P.B.F.Ouwerkerk, unpublished materials) which is based on (McElroy et al., 1990). Both fragments were inserted between HindIII and XhoI sites of binary vector pC1300intC (Genbank Accession AF294978), to obtain binary vector ProAct1::OsJAR1.
For construct GST-OsJAR1, the 1.5 kb fragment including the entire OsJAR1 coding sequence was rescued from pCRB4-OsJAR1T construct by HindIII and NcoI partial digestion and inserted between HindIII and NcoI of GST expression vector pGEX-KG (Guan and Dixon, 1991), resulting in construct GST-OsJAR1. Rice transformation Transformation of rice (O. sativa L. cv. Nipponbare) was performed as previously described by (Scarpella et al., 2000), except that Agrobacterium tumefaciens strain LBA 4404 was used instead of LBA 1119. Plantlets were maintained in tissue culture on half-strength Murashige-Skooge (MS) medium, 10 g/l sucrose medium before transfer to the greenhouse. Crossing experiments The crossing assay was performed using mature pollen of wild-type Nipponbare plants to fertilize pistils during floret opening of wild-type and osjar1-2 plants. To avoid contamination of self-fertilization of osjar1-2, anthers on the osjar1-2 mutant were removed before manual crossing.

Functions of OsJAR1 in floret opening and anther dehiscence
102
Iodine-potassium iodide staining assay and in vitro pollen germination Mature pollen was stained with 1% iodine-potassium iodide solution for two minutes and was observed with a microscope (Leica) using bright-field illumination. Wild type pollen grains were directly collected on a glass slide by shaking dehiscing anthers. For indehiscent anthers of the osjar1-2 mutant, pollen grains were manually squeezed onto a glass slide. Ten spikelets of wild type and osjar1-2 plants were checked.
In vitro pollen germination was carried out according to the method of Han et al. (2007). Pollen grains from dehisced anthers from Nipponbare and osjar1-2 plants (opened manually) were placed on glass slides at 28°C for 2 h in pollen germination medium consisting of 1 mM CaCl2, 1 mM KCl, 0.8 mM MgSO4, 1.6 mM H3BO3, 30 mM CaSO4, 0.03% casein, 0.3% 2-(N-morpholino) ethanesulfonic acid, 10% sucrose and 12.5% polyethylene glycol 4,000. Humidity was maintained above 90%. Observations were with a Zeiss Axioplan 2 microscope using the bright-field option. Opening of florets induced by MeJA Floret opening assays were carried out according to the method of Zeng et al. (1999). Panicles having florets close to anthesis, in which several florets had flowered before the experimental date, were selected for the tests. Experiments were conducted at 8 am. Intact panicles were carefully immersed in various concentrations of MeJA or control solution for two minutes. For each panicle, the time lag between treatment and the occurrence of the first opening floret was carefully monitored, and the numbers of opening florets were recorded at fixed intervals. During the experiments, shaking or any form of contact stimuli was carefully avoided. MeJA was dissolved in 95% ethanol and then diluted to the desired concentrations. The control was water with the same amount of ethanol as in the MeJA solutions. Root growth inhibition assays Dehusked seeds from Nipponbare and osjar1-2 plants were soaked into water for one day at dark and then sowed on medium with various concentrations of MeJA. Seeds were grown in tissue culture at 28°C and a light/dark cycle of 12 h/12 h and 80% humidity. Measurement of root length was carried out at the same moment of the day and lasted for eight subsequent days. Fungal infection of rice plants Plants were spray-inoculated when three weeks old with spore suspensions of 50,000 spores/ml as described in Vergne et al. (2007). Disease symptoms or GUS activity of infected leaves were monitored at seven days-post inoculation. Southern and Northern blot analyses For Southern blot analysis, rice genomic DNA was isolated from leaf samples collected from young developing greenhouse grown plants in 96 tube-racks (Qiagen) and dry-ground using a Mixer Mill MM300 (Retsch, Germany) with 4 mm stainless steel beads. Per sample 5 µg DNA was digested with XbaI, fractionated on a 0.8% agarose gel run in TAE and transferred onto Hybond N+ membranes (Amersham) under alkaline conditions. Hybridizations were performed with 32P-labeled OsJAR1-specific probes at 65°C in hybridization mixture (10% dextran sulphate, 1 M NaCl, 1% SDS, 100 µg/ml of denatured salmon sperm DNA). The membranes were washed once in 2x SSC

Chapter 3
103
and 1% SDS at 65°C for 30 min and once in 1x SSC and 0.1% SDS at 65°C for 30 min. For northern blot analysis, total RNA from various tissues was isolated and 20 μg per sample was electrophorized in formaldehyde agarose gels (Memelink et al., 1994) transferred to Hybond-N+ membrane. Hybridizations were performed at 65°C overnight. Next, membranes were washed twice in 0.1 × SSPE, 0.5% SDS at 42°C for 30 min before exposition to X-Ray films. Detection of GUS and GFP expression, cytological techniques and microscopy Histochemical detection of GUS activity, cytological techniques and microscopy were performed as described earlier (Scarpella et al., 2000). Samples were viewed using a Leica MZ12 stereo microscope or Zeiss Axioplan 2 and images were acquired with a Sony 3CCD Digital Photo Camera DKC-5000. RNA in situ hybridyzation Digoxygenin-labled anti-sense and sense RNA probes were prepared by in vitro transcription according to the instruction provided with the DIG RNA Labeling Kit (SP6/T7; Roche). The OsJAR1 cDNA used was amplified on the template of plasmid pCRB4-OGM1T with following primers: 5’-TTGTGAACCCATCAAACAGCA-3’ (forward) and 5’-TAATACGACTCACTATAGGGACTTTGCAGCACTTGTTACGGG-3’ (reverse, with T7 promoter); 5’-TAATACGACTCACTATAGGGTTGTGAACCCATCAAACAGCA-3’ (forward, with T7 promoter) and 5’-ACTTTGCAGCACTTGTTACGGG-3’ (reverse). Young florets of wild type rice (Nipponbare) were fixed in 3.7% FAA (5% acetic acid, 50% ethanol, 3.7% formaldehyde in water), dehydrated through an ethanol series and embedded in paraffin. Paraffin blocks were cut into 7 µm sections and mounted on POLYSINETM slides (Menzel GmbH & Co KG). Pretreatment of the sections was done as described before (Ferrandiz and Sessions, 2008). Subsequently, the sections were hybridized as described previously (Cañas et al., 1994) with modified hybridization temperature at 50°C. Formamide washing and immunological detection were performed as described (Kouchi and Hata, 1993) with minor modifications. The slides were washed twice with 2xSSC, 50% formamide at 50°C for 45 minutes after hybridization and washed twice with 1xSSC, 50% formamide at 55°C for 60 minutes after RNAse A treatment. After washing the sections were incubate with 1:1000 diluted alkaline phosphatase-conjugated antidigoxigenin antibody at 4°C overnight and then incubated at 30°C about 5 hours for color reaction. Slides were mounted with a coverslip and observed with Zeiss Axioplan 2 upright microscope. Photographs were made on a Sony 3CCD Digital Photo Camera DKC-5000. Expression and purification of the recombinant protein In order to extract recombinant protein expressed from construct GST-OsJAR1, 5 ml of overnight cultures of BL21 (DE3, pLys; Novagen) carrying pGEX plasmids were used to inoculate 500 ml LB medium containing 200 µg/ml carbenicillin and 25 µg /ml chloroamphenicol and grown at 37°C to an optical density at 600 nm of 0.5. Next, protein synthesis was induced by the addition of solid IPTG to final concentration of 1 mM and cultures were grown for 4 hours at 29°C. The harvested cells were suspended in 20 ml PBS and frozen in liquid nitrogen. After thawing on ice, the bacteria were lysed by sonication (eight times 10 s burst; 5 s pause between bursts), centrifuged (at 18,000

Functions of OsJAR1 in floret opening and anther dehiscence
104
rpm for 30 min at 4°C) and the supernatant was filtered through a 0.45 µm membrane. Protein purification was performed using Poly-Prep Chromatography columns (Biorad 731-1550) containing 0.5 ml settled Glutathion-Sepharose 4B beads (Amersham Biosciences). Columns were first washed two times with 10 ml PBS before bacterial extract was passed through. After binding, columns were washing with 10 ml PBS and bound proteins were eluted in 2.5 ml (10 x 0.25 ml) glutathion elution buffer (100 mM glutathione, 500 mM Tris-HCL pH 8.0). Eluted protein was concentrated using Microcone centrifugal filter devices (Millipore) according to manufacturer’s protocol and the protein content was determined by the method of Bradford. The GST-OsJAR1 fusion protein was stored at -80°C in 10% glycerol. In vitro synthesis of Jasmonate-amino acid conjugates The synthesis of JA-AA conjugates was based on the method described previously (Staswick and Tiryaki, 2004) with minor modifications. Reactions included 100 mM Tris-HCl (pH8.6), 6 mM MgCl2, 6 mM ATP, 2 mM DTT, 2 mM JA, 3 mM amino acid and 1 µg GST-OsJAR1 protein. Reactions were incubated at 25°C for two hours and run on TLC Silica gel 60 F254 sheets (Merck kGaA, Germany) with 35% chloroform, 55% ethyl acetate and 10% formic acid. The plates were stained with 6% vanillin and 1% sulfuric acid (in ethanol) and baked at 110°C until spots appeared. The products of the enzyme reactions were eluted from TLC-Silica gel (~ 1 ml) with 5 ml CHCl3/EtOAc (1:1) and 5 ml MeOH/FA (99:1). Of each fraction 1 ml was evaporated to dryness under a stream of nitrogen, dissolved in 0.5 ml of MeCN/H2O/FA (25:75:0.1), filtered and stored at -20oC. The chemical identity of the synthesized conjugates was verified by Time-of-Flight (ToF) mass spectrometry using a QTOF Ultima V4.00.00 mass spectrometer (Waters-Micromass, UK). The accurate masses obtained and the calculated elemental compositions correlated very well with the expected chemical structures of the JA-AA conjugates. Sample preparation for LC-MS/MS analysis Fresh flag leaves were harvested from six individual rice plants (cultivar Nipponbare or osjar1-2) frozen in liquid nitrogen, grinded, and either directly used for extractions or stored at -80°C. Similarly, leaves from six Nipponbare and osjar1-2 plants that had been sprayed with 10 mM JA were harvested five hours after treatment for LC-MS/MS analysis. Each experiment was done in triplicates. The grinded fresh rice leaves (approximately 500 mg) from each plant were extracted with 2 ml of cold ethyl acetate in a 10 ml glass tube. The tubes were vortexed and sonicated for 10 min in a Branson 3510 ultrasonic bath (Branson Ultrasonics, USA). Samples were centrifuged for 10 min at 1,350 g in a MSE Mistral 2000 centrifuge (Mistral Instruments, UK), and the organic phase was carefully transferred to 10 ml glass tubes. The pellet was re-extracted with 2 ml of ethyl acetate. The ethyl acetate fractions were combined and dried under a flow of nitrogen and the residue was dissolved in 250 µl of MeCN/H2O/FA (25:75:0.1). Before LC-MS/MS analysis, samples were filtered through Minisart SRP4 0.45 µm filters (Sartorius, Germany). Analysis of JA-AA conjugates using UPLC-MS/MS Analysis was performed using a Waters Micromass Quattro Premier XE tandem mass spectrometer (Waters, USA) equipped with an ESI source and coupled to an Acquity UPLC system (Waters, USA). Chromatographic separation was achieved on an Acquity UPLC BEH C18 column (150 × 2.1 mm,

Chapter 3
105
1.7 µm) (Waters, USA), applying a MeCN/H2O gradient containing 0.1% FA. The gradient started at 0% MeCN for 1 min, raised to 35% MeCN in 1 min, followed by an 8 min gradient to 40% MeCN, followed by a 1 min gradient to 90% MeCN, which was then maintained for 1 min and followed by a 0.2 min gradient back to the starting conditions. The column was equilibrated at this solvent composition for 2.8 min. Total run time was 15 min. The column was operated at 50ºC with a flow-rate of 0.4 ml min-1 and sample injection volume was 20 μl. The mass spectrometer was operated in positive and negative electrospray ionization (ESI) mode. The nebuliser and desolvation gas flows were 50 and 800 l h-1, respectively. The capillary voltage was set at 2.7 kV, the cone voltage at 20 V for ESI+ and 30 V for ESI-, the source temperature at 120ºC and the desolvation gas temperature at 450ºC. Fragmentation was performed by collision induced dissociation with argon at 3.6 × 10-3 mbar.
Multiple reaction monitoring (MRM) was used for analysis of JA and JA-AA conjugates. The MRM transitions were set according to the full-scan and MS/MS spectra obtained for synthetic JA and the enzymatically synthesized JA-AA (JA-Ile, JA-Leu, JA-Val, JA-Phe, JA-Met, JA-Trp) conjugates. Full scan and MS/MS spectra were recorded in ESI+ and ESI-. The collision energy was optimized for each individual compound for at least two fragment ions in positive mode and one in negative mode. The route of fragmentation was similar for all analyzed JA-AA conjugates. In ESI- the most intense fragment resulted from the loss of the amino acid moiety as has been observed before (Tamogami and Kodama, 1998; Wang et al., 2007). Under ESI+ conditions a larger number of fragment ions were produced the most abundant fragment ions were the [M+H]+-46 ion, formed by loss of formic acid from the amino acid and a fragment with m/z 151, [C10H15O]+ (2-pentenyl-pentan-3-one) originating from the JA moiety. For JA-Ile and JA-Leu also a fragment with m/z 86 [C5H12N]+ derived from the AA moiety was obtained. The MRM method operated in ESI+ mode is more specific for the identification of JA-AA conjugates then in ESI-. Moreover, a better response was obtained for the product ions in ESI+ mode. Therefore the data obtained in ESI+ were used for the quantification of JA-AA conjugates. MassLynx 4.1 software (Waters, USA) was used for data acquisition and analysis. Treatment of lodicules with JA Ten flowers from two osjar1-2 independent plants one day after flowering start were treated with 5 mM JA by pipetting with 3 µl of 5 mM JA onto the lodicules very carefully. To avoid the latent mechanical influences, ten osjar1-2 flowers were treated with 4% DMSO as controls. The treatment was started at 9 am. Accession Numbers Sequence data from this article can be found in the EMBL/GenBank databases with accession codes FI494725-FI494728. FI494725 and FI494726 are Tos17 FSTs from osjar1-2 and FI494727 and FI494728 are from osjar1-3, respectively. Acknowledgements Tos17 lines used in this work were developed by Drs. Akio Miyao and Hirohiko Hirochika (Miyao et al., 2003) and were provided by the Rice Genome Resource Center (Tsukuba, Japan). We are grateful to Gerda Lamers for her technical support in microscopy. We thank Ward de Winter for tissue culture media and Elly Schrijnemakers for plant caretaking. This research was supported by

Functions of OsJAR1 in floret opening and anther dehiscence
106
the Programme for Strategic Scientific Alliances (04-PSA-BD-04) between China and the Netherlands of the Royal Netherlands Academy of Arts and Sciences (KNAW) for AA, MW, PBFO, YX, ZZ and the KNAW China Exchange Programme (04CDP022, 06CDO033, 07CDP005, 08CDP042) for YX, YC, MW, ZZ and PBFO. We acknowledge financial support from The Netherlands Organisation for Scientific Research (NWO; VICI-grant) to HJB, TC and PPJM. References Aarts, M.G., Hodge, R., Kalantidis, K., Florack, D., Wilson, Z.A., Mulligan, B.J., Stiekema, W.J., Scott,
R., and Pereira, A. (1997). The Arabidopsis MALE STERILITY 2 protein shares similarity with reductases in elongation/condensation complexes. Plant J. 12, 615-623.
Aarts, M.G.M., Dirkse, W.G., Stiekema, W.J., and Pereira, A. (1993). Transposon tagging of a male sterility gene in Arabidopsis. Nature 363, 715-717.
Acosta, I.F., Laparra, H., Romero, S.P., Schmelz, E., Hamberg, M., Mottinger, J.P., Moreno, M.A., and Dellaporta, S.L. (2009). tasselseed1 is a lipoxygenase affecting jasmonic acid signaling in sex determination of maize. Science 323, 262-265.
Alberts, B., Wilson, J.H., and Hunt, T. (2008). Molecular biology of the cell. (New York: Garland Science). Berger, S. (2002). Jasmonate-related mutants of Arabidopsis as tools for studying stress signaling. Planta 214,
497-504. Bieleski, R., Elgar, J., Heyes, J., and Woolf, A. (2000). Flower opening in Asiatic Lily is a rapid process
controlled by dark-light cycling. Ann. Bot. (Lond) 86, 1169-1174. Cañas, L.A., Busscher, M., Angenent, G.C., Beltran, J.P., and Van Tunen, A.J. (1994). Nuclear
localization of the petunia MADS box protein FBP1 in petal and stamen cells. Plant J. 6, 597-604. Chen, I.C., Huang, I.C., Liu, M.J., Wang, Z.G., Chung, S.S., and Hsieh, H.L. (2007). Glutathione S-
transferase interacting with far-red insensitive 219 is involved in phytochrome A-mediated signaling in Arabidopsis. Plant Physiol. 143, 1189-1202.
Ding, X., Cao, Y., Huang, L., Zhao, J., Xu, C., Li, X., and Wang, S. (2008). Activation of the indole-3-acetic acid-amido synthetase GH3-8 suppresses expansin expression and promotes salicylate- and jasmonate-independent basal immunity in rice. Plant Cell 20, 228-240.
Ellis, C., and Turner, J.G. (2001). The Arabidopsis mutant cev1 has constitutively active jasmonate and ethylene signal pathways and enhanced resistance to pathogens. Plant Cell 13, 1025-1033.
Farmer, E.E. (2007). Plant biology: jasmonate perception machines. Nature 448, 659-660. Ferrandiz, C., and Sessions, A. (2008). Nonradioactive In Situ Hybridization of RNA Probes to Sections of
Plant Tissues, pp. pdb.prot4943.abstract Ferrari, S., Plotnikova, J.M., De Lorenzo, G., and Ausubel, F.M. (2003). Arabidopsis local resistance to
Botrytis cinerea involves salicylic acid and camalexin and requires EDS4 and PAD2, but not SID2, EDS5 or PAD4. Plant J. 35, 193-205.
Ge, Y.X., Angenent, G.C., Dahlhaus, E., Franken, J., Peters, J., Wullems, G.J., and Creemers-Molenaar, J. (2001). Partial silencing of the NEC1 gene results in early opening of anthers in Petunia hybrida. Mol.Genet.Genomics 265, 414-423.
Guan, K., and Dixon, J.E. (1991). Eukaryotic proteins expressed in Escherichia coli: An improved thrombin cleavage and purification procedure of fusion proteins with glutathione S-transferase. Anal. Biochem. 192, 262-267.
Han, M.J., Jung, K.H., Yi, G., Lee, D.Y., and An, G. (2007). Rice immature pollen 1 (RIP1) is a regulator of late pollen development. Plant Cell Physiol. 48, 204.
Holbrook, L., Tung, P., Ward, K., Reid, D.M., Abrams, S., Lamb, N., Quail, J.W., and Moloney, M.M. (1997). Importance of the Chiral Centers of Jasmonic Acid in the Responses of Plants (Activities and Antagonism between Natural and Synthetic Analogs). Plant Physiol. 114, 419-428.
Hsieh, H.L., Okamoto, H., Wang, M., Ang, L.H., Matsui, M., Goodman, H., and Deng, X.W. (2000). FIN219, an auxin-regulated gene, defines a link between phytochrome A and the downstream regulator COP1 in light control of Arabidopsis development. Genes Dev. 14, 1958-1970.
Ichimura, K.A.Z.U., and Suto, K.E.N.I. (1998). Environmental Factors Controlling Flower Opening and Closing in aPortulacaHybrid. Ann. Bot. (Lond) 82, 67-70.
Ishiguro, S., Kawai-Oda, A., Ueda, J., Nishida, I., and Okada, K. (2001). The DEFECTIVE IN ANTHER

Chapter 3
107
DEHISCIENCE gene encodes a novel phospholipase A1 catalyzing the initial step of jasmonic acid biosynthesis, which synchronizes pollen maturation, anther dehiscence, and flower opening in Arabidopsis. Plant Cell 13, 2191-2209.
Jain, M., Kaur, N., Tyagi, A.K., and Khurana, J.P. (2006). The auxin-responsive GH3 gene family in rice (Oryza sativa). Funct. Integr. Genomics 6, 36-46.
Jones, J.D., and Dangl, J.L. (2006). The plant immune system. Nature 444, 323-329. Kaihara, S., and Takimoto, A. (1980). Studies on the light controlling the time of flower-opening in Pharbitis
nil. Plant Cell Physiol. 21, 21-26. Kaihara, S., and Takimoto, A. (1981). Effects of Light and Temperature on Flower-opening of Pharbitis nil.
Plant Cell Physiol. 22, 215-221. Kang, J.H., Wang, L., Giri, A., and Baldwin, I.T. (2006). Silencing threonine deaminase and JAR4 in
Nicotiana attenuata impairs jasmonic acid-isoleucine-mediated defenses against Manduca sexta. Plant Cell 18, 3303-3320.
Keijzer, C.J. (1987). The processes of anther dehiscence and pollen dispersal. I. The opening mechanism of longitudinally dehiscing dnthers New Phytol. 105, 487-498.
Kouchi, H., and Hata, S. (1993). Isolation and characterization of novel nodulin cDNAs representing genes expressed at early stages of soybean nodule development. Mol. Gen. Genet. 238, 106-119.
Kramell, R., Porzel, A., Miersch, O., and Schneider, G. (1999). Analysis of synthetic isoleucine conjugates of cucurbic acid isomers by liquid chromatography. Phytochem. Anal. 10, 82-87.
Ma, N., Cai, L., Lu, W., Tan, H., and Gao, J. (2005). Exogenous ethylene influences flower opening of cut roses (Rosa hybrida) by regulating the genes encoding ethylene biosynthesis enzymes. Sci.China C. Life Sci. 48, 434-444.
Mandaokar, A., Thines, B., Shin, B., Lange, B.M., Choi, G., Koo, Y.J., Yoo, Y.J., Choi, Y.D., and Browse, J. (2006). Transcriptional regulators of stamen development in Arabidopsis identified by transcriptional profiling. Plant J. 46, 984-1008.
Matsui, T., Omasa, K., and Horie, T. (1999). Mechanism of anther dehiscence in rice (Oryza sativa L.). Ann. Bot. (Lond) 84, 501-506.
Matsuo, T., and Hoshikawa, K. (1993). Science of the rice plant (Tokyo: Rural Culture Association), pp. 341-345.
McConn, M., and Browse, J. (1996). The critical requirement for linolenic acid is pollen development, not photosynthesis, in an Arabidopsis mutant. Plant Cell 8, 403-416.
McElroy, D., Rothenberg, M., and Wu, R. (1990). Structural characterization of a rice actin gene. Plant Mol. Biol. 14, 163-171.
Mei, C., Qi, M., Sheng, G., and Yang, Y. (2006). Inducible overexpression of a rice allene oxide synthase gene increases the endogenous jasmonic acid level, PR gene expression, and host resistance to fungal infection. Mol. Plant Microbe Interact. 19, 1127-1137.
Memelink, J., Swords, K.M.M., Staehelin, L.A., and Hoge, J.H.C. (1994). Southern, Northern and Western blot analysis. In Plant Molecular Biology Manual, Gelvin SB and S. RA, eds (Dordrecht: Kluwer Academic Publishers), pp. F1-F23.
Mitsuda, N., Seki, M., Shinozaki, K., and Ohme-Takagi, M. (2005). The NAC transcription factors NST1 and NST2 of Arabidopsis regulate secondary wall thickenings and are required for anther dehiscence. Plant Cell 17, 2993-3006.
Miyao, A., Tanaka, K., Murata, K., Sawaki, H., Takeda, S., Abe, K., Shinozuka, Y., Onosato, K., and Hirochika, H. (2003). Target Site Specificity of the Tos17 Retrotransposon Shows a Preference for Insertion within Genes and against Insertion in Retrotransposon-Rich Regions of the Genome. Plant Cell 15, 1771-1780.
Mizuno, S., Osakabe, Y., Maruyama, K., Ito, T., Osakabe, K., Sato, T., Shinozaki, K., and Yamaguchi-Shinozaki, K. (2007). Receptor-like protein kinase 2 (RPK 2) is a novel factor controlling anther development in Arabidopsis thaliana. Plant J. 50, 751-766.
Mueller, M.J., and Brodschelm, W. (1994). Quantification of jasmonic acid by capillary gas chromatography-negative chemical ionization-mass spectrometry. Anal. Biochem. 218, 425-435.
O'Donnell, P.J., Schmelz, E., Block, A., Miersch, O., Wasternack, C., Jones, J.B., and Klee, H.J. (2003). Multiple hormones act sequentially to mediate a susceptible tomato pathogen defense response. Plant Physiol. 133, 1181-1189.
Ogawa, M., Kay, P., Wilson, S., and Swain, S.M. (2009). ARABIDOPSIS DEHISCENCE ZONE POLYGALACTURONASE1 (ADPG1), ADPG2, and QUARTET2 are polygalacturonases required

Functions of OsJAR1 in floret opening and anther dehiscence
108
for cell separation during reproductive development in Arabidopsis. Plant Cell 21, 216-233. Park, J.H., Halitschke, R., Kim, H.B., Baldwin, I.T., Feldmann, K.A., and Feyereisen, R. (2002). A
knock-out mutation in allene oxide synthase results in male sterility and defective wound signal transduction in Arabidopsis due to a block in jasmonic acid biosynthesis. Plant J. 31, 1-12.
Qin, Y., Yang, J., and Zhao, J. (2005). Calcium changes and the response to methyl jasmonate in rice lodicules during anthesis. Protoplasma 225, 103-112.
Qiu, D., Xiao, J., Ding, X., Xiong, M., Cai, M., Cao, Y., Li, X., Xu, C., and Wang, S. (2007). OsWRKY13 mediates rice disease resistance by regulating defense-related genes in salicylate- and jasmonate-dependent signaling. Mol. Plant Microbe Interact. 20, 492-499.
Riemann, M., Riemann, M., and Takano, M. (2008). Rice JASMONATE RESISTANT 1 is involved in phytochrome and jasmonate signalling. Plant, Cell Env. 31, 783-792.
Sanders, P.M., Lee, P.Y., Biesgen, C., Boone, J.D., Beals, T.P., Weiler, E.W., and Goldberg, R.B. (2000). The arabidopsis DELAYED DEHISCENCE1 gene encodes an enzyme in the jasmonic acid synthesis pathway. Plant Cell 12, 1041-1061.
Scarpella, E., Rueb, S., Boot, K.J., Hoge, J.H.C., and Meijer, A.H. (2000). A role for the rice homeobox gene Oshox1 in provascular cell fate commitment. Development 127, 3655-3669.
Schuller, A., and Ludwig-Muller, J. (2006). A family of auxin conjugate hydrolases from Brassica rapa: characterization and expression during clubroot disease. New Phytol. 171, 145-157.
Senatore, A., Trobacher, C.P., and Greenwood, J.S. (2009). Ricinosomes Predict Programmed Cell Death Leading to Anther Dehiscence in Tomato. Plant Physiol. 149, 775-790.
Staswick, P.E. (2009). The Tryptophan Conjugates of Jasmonic And Indole-3-Acetic Acids Are Endogenous Auxin Inhibitors. Plant Physiol. 150, 1310-21.
Staswick, P.E., and Tiryaki, I. (2004). The oxylipin signal jasmonic acid is activated by an enzyme that conjugates it to isoleucine in Arabidopsis. Plant Cell 16, 2117-2127.
Staswick, P.E., Su, W., and Howell, S.H. (1992). Methyl jasmonate inhibition of root growth and induction of a leaf protein are decreased in an Arabidopsis thaliana mutant. Proc.Natl.Acad. Sci.USA 89, 6837-6840.
Staswick, P.E., Yuen, G.Y., and Lehman, C.C. (1998). Jasmonate signaling mutants of Arabidopsis are susceptible to the soil fungus Pythium irregulare. Plant J. 15, 747-754.
Staswick, P.E., Tiryaki, I., and Rowe, M.L. (2002). Jasmonate response locus JAR1 and several related Arabidopsis genes encode enzymes of the firefly luciferase superfamily that show activity on jasmonic, salicylic, and indole-3-acetic acids in an assay for adenylation. Plant Cell 14, 1405-1415.
Staswick, P.E., Serban, B., Rowe, M., Tiryaki, I., Maldonado, M.T., Maldonado, M.C., and Suza, W. (2005). Characterization of an Arabidopsis enzyme family that conjugates amino acids to indole-3-acetic acid. Plant Cell 17, 616-627.
Steiner-Lange, S., Unte, U.S., Eckstein, L., Yang, C., Wilson, Z.A., Schmelzer, E., Dekker, K., and Saedler, H. (2003). Disruption of Arabidopsis thaliana MYB26 results in male sterility due to non-dehiscent anthers. Plant J. 34, 519-528.
Stintzi, A., and Browse, J. (2000). The Arabidopsis male-sterile mutant, opr3, lacks the 12-oxophytodienoic acid reductase required for jasmonate synthesis. Proc. Natl. Acad. Sci. USA 97, 10625-10630.
Suza, W.P., and Staswick, P.E. (2008). The role of JAR1 in Jasmonoyl-L: -isoleucine production during Arabidopsis wound response. Planta 227, 1221-1232.
Tamogami, S., and Kodama, O. (1998). Quantification of amino acid conjugates of jasmonic acid in rice leaves by high-performance liquid chromatography-turboionspray tandem mass spectrometry. J. Chromatogr. 822, 310-315.
Tanaka, O., Tanaka, Y., and Wada, H. (1988). Photonastic and thermonastic opening of capitulum in dandelion,Taraxacum officinale and Taraxacum japonicum. J. Plant Res. 101, 103-110.
Thines, B., Katsir, L., Melotto, M., Niu, Y., Mandaokar, A., Liu, G., Nomura, K., He, S.Y., Howe, G.A., and Browse, J. (2007). JAZ repressor proteins are targets of the SCF(COI1) complex during jasmonate signalling. Nature 448, 661-665.
Traw, M.B., Kim, J., Enright, S., Cipollini, D.F., and Bergelson, J. (2003). Negative cross-talk between salicylate- and jasmonate-mediated pathways in the Wassilewskija ecotype of Arabidopsis thaliana. Mol. Ecology 12, 1125-1135.
Van Doorn, W.G., and Van Meeteren, U. (2003). Flower opening and closure: a review. J. Exp. Bot. 54, 1801-1812.
Vergne, E., Ballini, E., Marques, S., Mammar, B.S., Droc, G., Gaillard, S., Bourot, S., DeRose, R.,

Chapter 3
109
Tharreau, D., Notteghem, J.L., Lebrun, M.H., and Morel, J.B. (2007). Early and specific gene expression triggered by rice resistance gene Pi33 in response to infection by ACE1 avirulent blast fungus. New Phytol. 174, 159-171.
Vick, B.A., and Zimmerman, D.C. (1984). Biosynthesis of jasmonic acid by several plant species. Plant Physiol. 75, 458-461.
Von Malek, M.B., Van der Graaf, G.E., Schneitz, K., and Keller, B. (2002). The Arabidopsis male-sterile mutant dde2-2 is defective in the ALLENE OXIDE SYNTHASE gene encoding one of the key enzymes of the jasmonic acid biosynthesis pathway. Planta 216, 187-192.
Wang, H., Ma, L., Habashi, J., Li, J., Zhao, H., and Deng, X.W. (2002). Analysis of far-red light-regulated genome expression profiles of phytochrome A pathway mutants in Arabidopsis. Plant J. 32, 723-733.
Wang, L., Halitschke, R., Kang, J.H., Berg, A., Harnisch, F., and Baldwin, I.T. (2007). Independently silencing two JAR family members impairs levels of trypsin proteinase inhibitors but not nicotine. Planta 226, 159-167.
Wang, Y., and Kumar, P.P. (2007). Characterization of two ethylene receptors PhERS1 and PhETR2 from petunia: PhETR2 regulates timing of anther dehiscence. J. Exp. Bot. 58, 533-544.
Wang, Z., Cao, G., Wang, X., Miao, J., Liu, X., Chen, Z., Qu, L.J., and Gu, H. (2008). Identification and characterization of COI1-dependent transcription factor genes involved in JA-mediated response to wounding in Arabidopsis plants. Plant Cell Reports 27, 125-135.
Xie, D.X., Feys, B.F., James, S., Nieto-Rostro, M., and Turner, J.G. (1998). COI1: an Arabidopsis gene required for jasmonate-regulated defense and fertility. Science 280, 1091-1094.
Zeng, X., Zhou, X., Zhang, W., Murofushi, N., Kitahara, T., and Kamuro, Y. (1999). Opening of Rice Floret in Rapid Response to Methyl Jasmonate. J. Plant Growth Reg.. 18, 153-158.
Zhu, Q.H., Ramm, K., Shivakkumar, R., Dennis, E.S., and Upadhyaya, N.M. (2004). The ANTHER INDEHISCENCE1 gene encoding a single MYB domain protein is involved in anther development in rice. Plant Physiol. 135, 1514-1525.
