genetic control of autoimmunity: protection from … · genetic control of autoimmunity: protection...
TRANSCRIPT

of June 8, 2018.This information is current as
Congenic StrainBiliary Disease in a Nonobese Diabetic
Autoimmunefrom Diabetes, but Spontaneous Genetic Control of Autoimmunity: Protection
Laurence B. Peterson and William M. RidgwayFenyk-Melody, Daniel B. Rainbow, Linda S. Wicker,
JudithSass, Michael Nalesnik, John A. Todd, Paul A. Lyons, Syuichi Koarada, Yuehong Wu, Noreen Fertig, David A.
http://www.jimmunol.org/content/173/4/2315doi: 10.4049/jimmunol.173.4.2315
2004; 173:2315-2323; ;J Immunol
Referenceshttp://www.jimmunol.org/content/173/4/2315.full#ref-list-1
, 14 of which you can access for free at: cites 40 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2004 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on June 8, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on June 8, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Genetic Control of Autoimmunity: Protection from Diabetes,but Spontaneous Autoimmune Biliary Disease in a NonobeseDiabetic Congenic Strain1
Syuichi Koarada,* Yuehong Wu,* Noreen Fertig,* David A. Sass,* Michael Nalesnik,†
John A. Todd,� Paul A. Lyons,� Judith Fenyk-Melody,¶ Daniel B. Rainbow,� Linda S. Wicker,�
Laurence B. Peterson,ठand William M. Ridgway 2*
At least 20 insulin-dependent diabetes (Idd) loci modify the progression of autoimmune diabetes in the NOD mouse, an animalmodel of human type 1 diabetes. The NOD.c3c4 congenic mouse, which has multiple B6- and B10-derivedIdd-resistant alleles onchromosomes 3 and 4, respectively, is completely protected from autoimmune diabetes. We demonstrate in this study, however,that NOD.c3c4 mice develop a novel spontaneous and fatal autoimmune polycystic biliary tract disease, with lymphocytic peribili-ary infiltrates and autoantibodies. Strains having a subset of theIdd-resistant alleles present in the NOD.c3c4 strain show com-ponent phenotypes of the liver disease: NOD mice with B6 resistance alleles only on chromosome 3 have lymphocytic liverinfiltration without autoantibody formation, while NOD mice with B10 resistance alleles only on chromosome 4 show autoantibodyformation without liver infiltration. The liver disease is transferable to naive NOD.c3c4 recipients using splenocytes from affectedNOD.c3c4 mice, demonstrating an autoimmune etiology. Thus, substitution of non-NOD genetic intervals into the NOD strain canprevent diabetes, but in turn cause an entirely different autoimmune syndrome, a finding consistent with a generalized failure ofself-tolerance in the NOD genetic background. The complex clinical phenotypes in human autoimmune conditions may be sim-ilarly resolved into largely overlapping biochemical pathways that are then modified, potentially by alleles at a few key chromo-somal regions, to produce specific autoimmune syndromes.The Journal of Immunology, 2004, 173: 2315–2323.
S pontaneous murine autoimmune diseases (e.g., diabetes inNOD mice or lupus in New Zealand Black (NZB)3/NewZealand White) mice are characterized by stereotypic clin-
ical phenotypes and genetically fixed (inbred) backgrounds. Hu-man autoimmune diseases, on the contrary, are characterized by acomplex array of overlapping phenotypes occurring in outbred,genetically heterogeneous backgrounds. It has been reported thatrelatives of autoimmune probands show an increased incidence ofboth the target autoimmune disease (e.g., type 1 diabetes (T1D)) aswell as an increased incidence of other autoimmune diseases, suchas rheumatoid arthritis or autoimmune thyroiditis (1, 2). Multiple,clinically disparate autoimmune diseases have also been reported
in the same patient, e.g., autoimmune diabetes coexisting with ei-ther Sjogren’s syndrome, autoimmune thyroiditis, or primary bil-iary cirrhosis (PBC) (3, 4). Metanalysis of multiple linkage studiesdemonstrated that many autoimmune disease-associated loci mapto a limited number of distinct gene clusters, suggesting that phe-notypically diverse diseases may result from a partially overlap-ping set of genes (5, 6). These studies suggest the hypothesis thatdivergent clinical disease phenotypes could arise from similar bio-chemical pathways.
In this study, we present a model system manifesting completelydifferent autoimmune syndromes in NOD and NOD congenicstrains differing at critical insulin-dependent diabetes (Idd) inter-vals. The NOD mouse has been extensively studied as a model ofgenetic control of autoimmunity. NOD mice develop immune-me-diated pancreatic � cell destruction (insulitis), progressing to dia-betes in 70–80% of female mice by 30 wk of age (7). The initialgenome scan, performed over a decade ago, identified many po-tential B10 protective Idd loci (8). Many of these loci cluster inregions of the mouse genome associated with other mouse auto-immune syndromes (5). A series of congenic mice have been pro-duced, using microsatellite-defined non-MHC Idd loci derivedfrom diabetes-resistant strains of mice introgressed onto the NODbackground; these mice show decreased diabetes incidence, andhence validate the initial linkage studies. NOD.c3 and NOD.c4congenic mice have a diabetes frequency of 3 and 5%, respectively(Fig. 1) (9–11). As we predicted, combining NOD.c3 (chromo-some 3, Idd3, Idd10, Idd17, and Idd18) with NOD.c4 (chromo-some 4, Idd9.1, Idd9.2, and Idd9.3) totally eliminated diabetes (9).We show in this study, however, that while a combination of pro-tective c3/c4 loci can effectively prevent diabetes, they interactwith the NOD genome to result in a novel, genetically controlledautoimmune liver disease.
Divisions of *Rheumatology and Immunology and †Pathology, School of Medicine,University of Pittsburgh, Pittsburgh, PA 15261; Departments of ‡Immunology andRheumatology, §Pharmacology, and ¶Comparative Medicine, Merck Research Lab-oratories, Rahway, NJ 07065; and �Wellcome Trust Centre for Molecular Mechanismsin Disease, Cambridge University, Cambridge, United Kingdom
Received for publication March 2, 2004. Accepted for publication June 8, 2004.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 L.S.W., J.A.T., P.A.L., and D.B.R. are supported by grants from the Juvenile Dia-betes Research Foundation and the Wellcome Trust. The availability of NOD con-genic mice through the Taconic Farms Emerging Models Program has been supportedby grants from the Merck Genome Research Institute, National Institute of Allergyand Infectious Diseases, and the Juvenile Diabetes Research Foundation. W.M.R. issupported by National Institutes of Health, National Institute of Diabetes and Diges-tive and Kidney Diseases 60714, and the Juvenile Diabetes Research Foundation.2 Address correspondence and reprint requests to Dr. William M. Ridgway, S725Biomedical Science Tower, 200 Lothrop Street, Pittsburgh, PA 15261. E-mail ad-dress: [email protected] Abbreviations used in this paper: NZB, New Zealand Black; ANA, antinuclear Ab;Idd, insulin-dependent diabetes; IP, immunoprecipitation; PBC, primary biliary cir-rhosis; PDC, pyruvate dehydrogenase complex; SLE, systemic lupus erythematosus;Sm, Smith; snRNP, small nuclear ribonucleoprotein; T1D, type 1 diabetes.
The Journal of Immunology
Copyright © 2004 by The American Association of Immunologists, Inc. 0022-1767/04/$02.00
by guest on June 8, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

Materials and MethodsMice
The development of the NOD.c3c4 strain from the parental strains, NOD.B10Idd9.1/9.2/9.3 and NOD.B6 Idd3/17/10/18, was described previously (9). Thec3c4 strain has �80 cM genetic intervals on chromosomes 4 and 3 intro-gressed from B10 and B6, respectively (Fig. 1). The NOD.B10 Idd9.1/9.2/9.3R28 strain (9), called c4(a) and used as the parental chromosome 4 con-genic strain in this study, has a smaller introgressed region (�60 cM) from theB10 strain as compared with the NOD.B10 Idd9.1/9.2/9.3 parental strain(which is no longer extant). The c3(a) strain was described previously (10) asthe NOD.B6 Idd3R1/Idd10R2 strain. Although the Idd17 resistance gene is notpresent in the c3(a) strain, diabetes resistance of this strain is identical with thatof the NOD.B6 Idd3/17/10/18 parental strain (which is also no longer extant),which was called NOD.B6 (Idd3/Idd10)R1 (10). Please note that subsequent tothe publication of Ref. 10, the Idd10 region was shown to consist of two Iddgenes, Idd10 and Idd18 (11), and therefore the strain designations were up-dated for the descriptions used in the current study. The c3c4 and c3(a) strainswere selectively bred to produce the novel c3(a)c4 strain. The c3c4, c3(a),c4(a), and c3(a)c4 strains are available from Taconic Farms (Germantown,NY) through the Emerging Models Program (lines 1112, 1538, 1104, and1802, respectively).
Histology
Organs were fixed in 10% buffered formalin and processed for paraffinembedding. Tissue sections (5 �m) were stained with H&E and evaluatedmicroscopically for leukocytic infiltration and cyst scores.
Antinuclear Ab (ANA) immunofluorescence
Prepared HEP-2 slides were purchased from Diasorin (Stillwater, MN).Mouse sera were diluted 1/20 in 10 mM PBS, pH 7.3. A total of 25 �l ofthe diluted sera was incubated on the slide for 30 min in a dark, moistchamber and washed in 10 mM PBS in a Coplin jar for 10 min. Fluores-cein-conjugated goat Ab to mouse IgG, IgM, and IgA (Valeant Pharma-ceuticals, Costa Mesa, CA) was diluted 1/100 in PBS. A total of 35 �l ofthe diluted conjugate was placed on each well, and the slide was incubated30 min in a dark, moist chamber and washed in PBS for 10 min. Slideswere viewed and scored at �400 magnification with a dry lens of aLeitz fluorescence microscope equipped with a mercury vapor lamp.(Leica Microsystems, Bannockburn, IL).
Protein immunoprecipitation (IP)
Protein IP assays were set up, as previously described (12). Briefly, rapidlygrowing, undifferentiated K562 cells were grown at 37°C and 5% CO2 inDMEM supplemented with 10% FBS. Cells were collected in 200-ml con-ical tubes, centrifuged at 800 rpm for 8 min, washed once with prewarmedmethionine-free DMEM culture medium, and centrifuged again. The me-dium was removed, and for each sample the following were added: 5 ml ofmethionine-free DMEM supplemented with 3% FBS, 50 �Ci of [35S]me-thionine (ExpreSS; New England Nuclear, Boston, MA), and 1 � 106
K562 cells. The cells were labeled at 37°C overnight (8–16 h), collected,washed, and suspended in 5–10 ml of IP buffer: 10 mM Tris-HCl, pH 8.0,500 mM NaCl, and 0.1% Igepal. They were then sonicated, and the son-icate was centrifuged for 10 min at 4°C at 14,000 rpm to remove debris. A
FIGURE 1. Genetic map of the NOD.c3c4, c3(a), c4(a), and c3(a)c4 strains showing the boundaries of the contributions from chromosomes 3 and 4.Shaded intervals denote B6/B10 genetic origin.
2316 GENETIC CONTROL OF AUTOIMMUNE BILIARY DISEASE
by guest on June 8, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

10-�l serum sample was bound overnight to 2 mg of protein A-SepharoseCL4b beads (Pharmacia), washed three times with IP buffer, and incubated2 h at 4°C with the [35S]methionine-radiolabeled extract from �1 � 106
rapidly dividing K562 cells. The beads were washed three times with IPbuffer, suspended in 2� Laemmli sample buffer, loaded on a standard size8–15% SDS-PAGE gel with a 5% stack, and electrophoresed at 200 V. Thegels were enhanced by soaking for 30 min in 0.5 M sodium salicylate. Thegels were dried and autoradiographed 2–7 days. Apparent m.w. of positivebands were determined by plotting the m.w. of known 14C-labeled stan-dards run on the same gel.
Anti-DNA assays
Anti-DNA assays were set up, as previously described (13), with the fol-lowing modifications. Calf thymus DNA (Sigma-Aldrich, St. Louis, MO)was dissolved in solution (33 mM Na acetate, 50 mM NaCl, 0.03 mMZnCl, pH 4.5) and incubated with S1 nuclease (Invitrogen Life Technol-ogies, Carlsbad, CA) at 0.1 U/�g for 45 min. The reaction was stopped bythe addition of Na2 EDTA (pH 8.0) to a final concentration of 10 mM. Anequal volume of isoamyl alcohol/chloroform (1:24) was added to removeprotein. After centrifugation, the DNA was precipitated with cold 95%ethanol and dissolved in SSC (0.1 M NaCl, 0.014 M Na citrate, pH 8.0).The concentration of DNA was determined by spectrophotometry; the 260/280 ratio was higher than 1.9. ssDNA was prepared by boiling for 3 minwith subsequent rapid cooling on ice. For dsDNA assays, S1-treated DNAwas used. Each well of an ELISA 96-well plate was precoated with 100 �lof protamine sulfate working solution (0.0001%), then coated with 100 �lof 5 �g/ml DNA 1 h at room temperature. Wells were blocked with PBSsupplemented with 1% BSA, pH 7.2. A total of 50 �l of appropriatelydiluted sera was added to each well and incubated for 1 h at room tem-perature, washed, then stained with biotin-conjugated goat anti-mouse IgGor IgM in duplicate or triplicate for 1 h at room temperature. Europium-labeled streptavidin, followed by enhancement solution, was added, andtime-resolved fluorometry was performed on a Wallac 1420 Victor2 flu-orometer (PerkinElmer Wallac, Upsala, Sweden). A pool of B6 control serawas run with each sample, providing a consistent reference standard. Theresults are expressed as a stimulation index � Eu counts (experimental)/Eucounts (control serum).
Transfer studies
We followed the standard protocol used to transfer diabetes into NODrecipients (14). Seven- to 12-wk-old female NOD.c3c4 recipient mice wereirradiated with 700–750 rad. A total of 25 � 106 donor splenocytes sus-pended in PBS was transferred i.v. into the irradiated recipients; irradiatedcontrols received PBS alone. The livers of the donors were examined his-tologically. The recipients were aged 90 days posttransfer, then analyzedfor histological evidence of liver disease.
ResultsSevere biliary tract disease in NOD.c3c4 mice
Predicting that the combination of the NOD.B6 Idd3/17/10/18(chromosome 3, c3) and NOD.B10 Idd9.1/9.2/9.3 (chromosome 4,c4) congenic strains would create an NOD-related strain, whichhas no autoimmune diabetes, we derived a new double congenicstrain, NOD.c3c4 (9) (see genetic map; Fig. 1). As expected,NOD.c3c4 mice were not susceptible to autoimmune diabetes (nomice developed diabetes by 12 mo of age), and almost no insulitiswas observed (9). Unexpectedly, the NOD.c3c4 strain developedan entirely different syndrome, with biliary disease resulting inmassive hepatomegaly (Fig. 2). Liver failure leading to a moribundstate developed in �50% of females and 25% of males at 9–11 moof age. Histological examination showed marked biliary cyst for-mation progressing with age (Fig. 3, A–C), and progressive lym-phocytic infiltrates adjacent to the cyst walls (Fig. 3, G–I). As theliver disease progressed in the NOD.c3c4 mice, biliary obstructionworsened, resulting in ascites formation and a moribund condition.Elevated serum transaminases (aspartate aminotransferase, alanineaminotransferase) accompanied histological evidence of disease(data not shown). Cysts were not found in any other organ. Con-sistent with the observation that none of the NOD.c3c4 mice de-veloped diabetes, the pancreatic islets showed only minimal peri-insulitis (Fig. 3, D–F), and the salivary glands showed �50% of
the lymphocytic infiltration observed in NOD mice (data notshown).
Kinetic studies of disease onset showed that 44% of c3c4 micehad early cyst formation and lymphocytic infiltrate at 8 wk. By 16wk, 71% showed histological disease, while 90% of mice had his-tological evidence of disease over age 30 wk. The histologicalscores for cyst formation and lymphocytic infiltration increased inan approximately linear fashion over time (data not shown); allmice with histological evidence of cyst formation had coexistentperibiliary lymphocytic infiltration histologically.
Component immunophenotypes in donor strains: NOD micecongenic for resistance alleles on chromosome 3 micedevelop lymphocytic liver infiltrates not seen in other NODcongenic mice
Notably, like the NOD strain, neither of the parental strains thathad been bred together to develop the NOD.c3c4 strain, c3(NOD.B6 Idd3/10/17/18) and c4 (NOD.B10 Idd9.1/9.2/9.3),showed any hepatomegaly after aging for over 1 year (n � 100females for each strain). However, we hypothesized that subclin-ical abnormalities potentially related to the liver disease might bepresent in one or both of these strains. Because the c3 and c4
FIGURE 2. A, Massive hepatomegaly in NOD.c3c4 mice (left) com-pared with NOD mice (right). B, Comparison of a liver lobe fromNOD.c3c4 (left) showing massive hepatomegaly compared with the samelobe from NOD (right).
2317The Journal of Immunology
by guest on June 8, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

strains used to develop the NOD.c3c4 strain are not extant, theexamination of chromosome 3 and 4 congenic strains for compo-nent immunophenotypes was performed with related congenicstrains (see Materials and Methods for strain derivation),NOD.c3(a) and NOD.c4(a) (Fig. 1). Upon histological examina-tion, four of four 33-wk-old female NOD.c3(a) mice demonstratedperibiliary lymphocytic aggregations without cyst formation (Fig.4). This abnormality was not present in NOD mice nor inNOD.c4(a) mice.
Component immunophenotypes in donor strains: NOD.c4(a)mice develop antinuclear and anti-Smith autoantibodies not seenin other NOD congenic mice
ANA testing was performed on each strain. Female NOD.c3c4mice began developing positive ANA at �12 wk, with 45% (5 of11) positive at 16 wk and 83% (58 of 70) ANA� at 22 wk andolder (Fig. 5). NOD.c4(a) mice also had a high incidence of ANApositivity, with 12 of 23 female mice ANA� at 20 wk and older.Most (greater than 95%) NOD, B6, and NOD.c3(a) mice wereANA negative (Fig. 5). In c3c4 mice, serum antinuclear autoanti-bodies generally formed after the development of histological dis-ease. However, the correlation was not absolute because occasion-ally c3c4 mice were ANA� while showing no histologicalevidence of biliary disease.
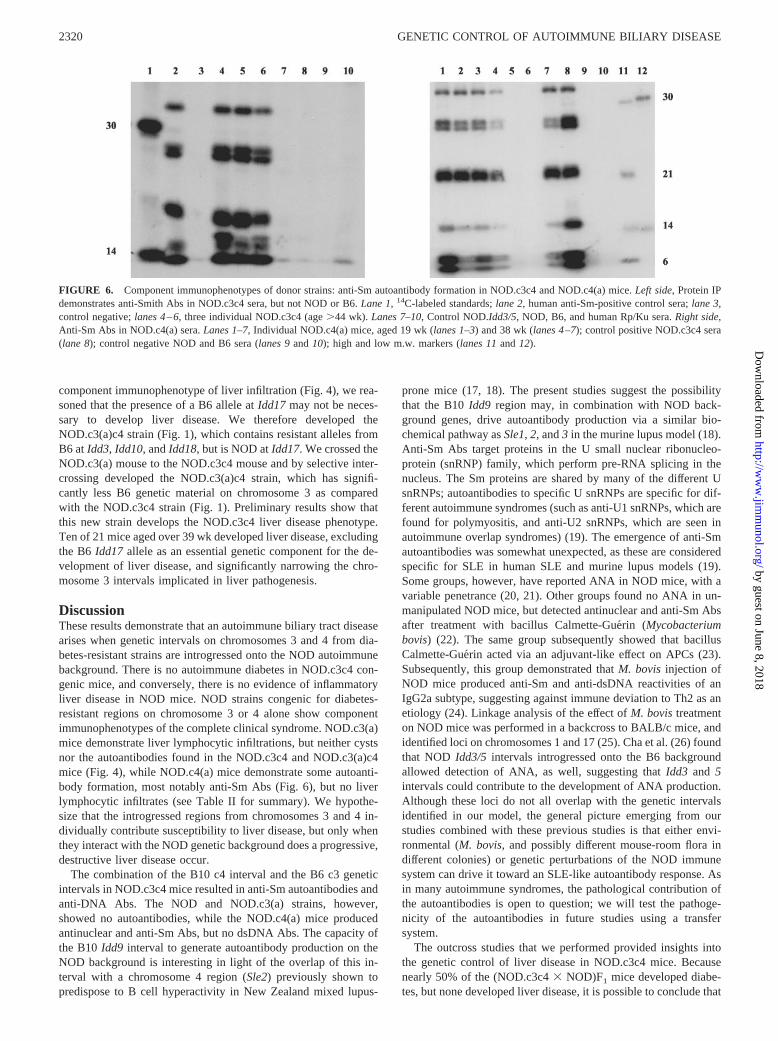
Autoantibody production was further investigated using IP.NOD.c3c4 mice demonstrated anti-Sm autoantibodies on IP,which are considered highly specific for systemic lupus erythem-atosus (SLE) in humans (Fig. 6). At 16 wk, 5 of 8 NOD.c3c4 micetested were positive for anti-Sm Abs by IP. By 35 wk, 17 of 29(58%) NOD.c3c4 mice were anti-Sm�. The specificity for Sm Agwas confirmed by RNA IP (data not shown). NOD, B6, andNOD.c3(a) mice were negative for anti-Sm Abs. NOD.c4(a) mice,however, formed anti-Sm Abs at a similar percentage toNOD.c3c4 mice, with 13 of 25 anti-Sm� at 20 wk or older (Fig. 6).
FIGURE 3. Kinetics of liver and pan-creatic histopathology in NOD.c3c4mice. A–C, Progressive biliary cyst for-mation in 8 (A)-, 16 (B)-, and 40 (C)-wk-old NOD.c3c4 mice. D–F, Minimal pan-creatic islet inflammation in the sameindividual mice. G–I, Progressiveperibiliary tract lymphocytic infiltrationin the same mice. Arrows point into bil-iary cyst cavities. Lymphocytes infiltratethe pericystic space.
FIGURE 4. Component immunophenotypes of donor strains: lympho-cytic infiltrates were found in NOD.c3(a) (left), but not NOD (right) norNOD.c4(a) strains.
2318 GENETIC CONTROL OF AUTOIMMUNE BILIARY DISEASE
by guest on June 8, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

Overall, the �50% penetrance of anti-Sm Abs in the NOD.c3c4and NOD.c4(a) mice is similar to that observed in the autoimmuneMRL/Mp-lpr/lpr mouse (16).
Anti-DNA autoantibodies in NOD.c3c4 mice
We further characterized autoantibody production in the NOD,NOD.c3(a), NOD.c4(a), B6, and NOD.c3c4 strains by analyzinganti-ssDNA and anti-dsDNA Ab levels. The IgM dsDNA titer wassignificantly elevated in NOD.c3c4 mice aged less than 6 mo, butin none of the other strains (Fig. 7A). As the mice aged past 8 mo,only NOD.c3c4 mice demonstrated significant elevations of IgGdsDNA autoantibody compared with the other genetically relatedstrains (Fig. 7A). The increase in titers was statistically equivalentto anti-DNA titers in NZB mice (Fig. 7B). NOD.c3c4 mice olderthan 8 mo also demonstrated increased IgM dsDNA and ssDNA,and IgG ssDNA autoantibody titers (Fig. 7C, and data not shown).Older female NOD.c3c4 mice demonstrated significant increasesin titers of IgM dsDNA, IgG ssDNA, and IgG dsDNA comparedwith male mice (Fig. 7C), suggesting a more severe autoantibodyresponse in female mice.
Splenocytes from diseased NOD.c3c4 mice can transfer liverdisease to naive irradiated NOD.c3c4 recipients
One fundamental test of autoimmune etiology in experimentalmodels is transfer of autoimmunity by lymphocytes from affectedto unaffected mice (14). An autoimmune etiology for NOD diabe-tes was first suggested by the transfer of diabetes with splenocytesfrom diabetic mice (14, 15). We used the NOD splenic transferprotocol to test whether the liver disease could be transferred withimmunocytes or whether the liver inflammation was secondary toa degenerative process intrinsic to biliary/hepatic cells. A total of25 � 106 splenocytes, or PBS alone, was transferred from diseasedolder mice to young, irradiated NOD.c3c4 recipients. Five of sevensplenocyte recipients developed histological evidence of liver dis-ease, compared with one of four PBS recipients (Fig. 8 and TableI). Consistent with results in NOD, irradiation was protective ofdisease in absence of cotransfer of pathogenic splenocytes (14).
Infectious agents are not cofactors for liver disease inNOD.c3c4 mice
To exclude any contribution by an infectious agent, NOD.c3c4mice were rederived by embryo transfer at Taconic Farms. Theliver disease emerged with equal penetrance following rederiva-tion, excluding a pathogenic bacterial/viral/parasitic etiology forour observations.
Diabetes, not liver disease, is dominant in (NOD.c3c4 �NOD)F1 mice
To begin to dissect the relationship of the genetic control of T1Dand the autoimmune liver disease, we tested for liver disease anddiabetes in two F1 crosses. Of 20 (NOD.c3c4 � NOD)F1 miceaged to over 1 year, 9 developed diabetes and none developed liverdisease. Thus, one dose of the chromosome 3 and 4 liver diseasealleles is insufficient to induce liver disease in the context of theNOD background. (NOD.c3c4 � B6.H2g7)F1 mice showed neitherdiabetes nor liver disease. This result demonstrated that the B6/B10 alleles, which are homozygous in this F1 (assuming B6 andB10 mice share disease alleles at both chromosomes), must inter-act with additional NOD genes present in a homozygous state onthe NOD genetic background to generate the NOD.c3c4 liver dis-ease phenotype. These results also demonstrate that a spontaneousdominant mutation is not the cause of liver disease in thec3c4 mice.
Autoantibody studies were performed in the F1 mice. Seven of10 (NOD.c3c4 � NOD)F1 mice were ANA�, and 4 of 5 tested byIP were anti-Sm�. In contrast, 6 of 17 (NOD.c3c4 � B6.H2g7)F1
mice were ANA�, and 0 of 17 were anti-Sm�. This result dem-onstrates that c3c4 loci are dominant for autoantibody productionwhen present on a homozygous NOD background. In contrast,anti-Sm autoantibodies cannot arise when the B6/B10 c3c4 allelesare homozygous on a heterozygous NOD/B6 background (see Ta-ble II).
The presence of B6 Idd17 is not required for the development ofliver disease
To determine whether the same genes that contribute protectionfrom T1D confer liver disease susceptibility or, alternatively, thatthe colocalization is due to linked loci, we have initiated the de-velopment of congenic strains that will continue to be susceptibleto liver disease if the Idd genes themselves are also the liver dis-ease genes. Because the NOD.c3(a) and the no longer extantNOD.c3 strains had the same degree of protections from diabetes(see Materials and Methods) and the NOD.c3(a) mice showed the
FIGURE 5. Component immunophenotypes of donor strains: antinu-clear autoantibody formation in NOD.c3c4 and NOD.c4(a) mice.NOD.c4(a) serum: speckled immunofluorescence antinuclear pattern.NOD.c3c4: homogenous antinuclear immunofluorescence pattern. NOD:negative ANA immunofluorescence.
2319The Journal of Immunology
by guest on June 8, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
component immunophenotype of liver infiltration (Fig. 4), we rea-soned that the presence of a B6 allele at Idd17 may not be neces-sary to develop liver disease. We therefore developed theNOD.c3(a)c4 strain (Fig. 1), which contains resistant alleles fromB6 at Idd3, Idd10, and Idd18, but is NOD at Idd17. We crossed theNOD.c3(a) mouse to the NOD.c3c4 mouse and by selective inter-crossing developed the NOD.c3(a)c4 strain, which has signifi-cantly less B6 genetic material on chromosome 3 as comparedwith the NOD.c3c4 strain (Fig. 1). Preliminary results show thatthis new strain develops the NOD.c3c4 liver disease phenotype.Ten of 21 mice aged over 39 wk developed liver disease, excludingthe B6 Idd17 allele as an essential genetic component for the de-velopment of liver disease, and significantly narrowing the chro-mosome 3 intervals implicated in liver pathogenesis.
DiscussionThese results demonstrate that an autoimmune biliary tract diseasearises when genetic intervals on chromosomes 3 and 4 from dia-betes-resistant strains are introgressed onto the NOD autoimmunebackground. There is no autoimmune diabetes in NOD.c3c4 con-genic mice, and conversely, there is no evidence of inflammatoryliver disease in NOD mice. NOD strains congenic for diabetes-resistant regions on chromosome 3 or 4 alone show componentimmunophenotypes of the complete clinical syndrome. NOD.c3(a)mice demonstrate liver lymphocytic infiltrations, but neither cystsnor the autoantibodies found in the NOD.c3c4 and NOD.c3(a)c4mice (Fig. 4), while NOD.c4(a) mice demonstrate some autoanti-body formation, most notably anti-Sm Abs (Fig. 6), but no liverlymphocytic infiltrates (see Table II for summary). We hypothe-size that the introgressed regions from chromosomes 3 and 4 in-dividually contribute susceptibility to liver disease, but only whenthey interact with the NOD genetic background does a progressive,destructive liver disease occur.
The combination of the B10 c4 interval and the B6 c3 geneticintervals in NOD.c3c4 mice resulted in anti-Sm autoantibodies andanti-DNA Abs. The NOD and NOD.c3(a) strains, however,showed no autoantibodies, while the NOD.c4(a) mice producedantinuclear and anti-Sm Abs, but no dsDNA Abs. The capacity ofthe B10 Idd9 interval to generate autoantibody production on theNOD background is interesting in light of the overlap of this in-terval with a chromosome 4 region (Sle2) previously shown topredispose to B cell hyperactivity in New Zealand mixed lupus-
prone mice (17, 18). The present studies suggest the possibilitythat the B10 Idd9 region may, in combination with NOD back-ground genes, drive autoantibody production via a similar bio-chemical pathway as Sle1, 2, and 3 in the murine lupus model (18).Anti-Sm Abs target proteins in the U small nuclear ribonucleo-protein (snRNP) family, which perform pre-RNA splicing in thenucleus. The Sm proteins are shared by many of the different UsnRNPs; autoantibodies to specific U snRNPs are specific for dif-ferent autoimmune syndromes (such as anti-U1 snRNPs, which arefound for polymyositis, and anti-U2 snRNPs, which are seen inautoimmune overlap syndromes) (19). The emergence of anti-Smautoantibodies was somewhat unexpected, as these are consideredspecific for SLE in human SLE and murine lupus models (19).Some groups, however, have reported ANA in NOD mice, with avariable penetrance (20, 21). Other groups found no ANA in un-manipulated NOD mice, but detected antinuclear and anti-Sm Absafter treatment with bacillus Calmette-Guerin (Mycobacteriumbovis) (22). The same group subsequently showed that bacillusCalmette-Guerin acted via an adjuvant-like effect on APCs (23).Subsequently, this group demonstrated that M. bovis injection ofNOD mice produced anti-Sm and anti-dsDNA reactivities of anIgG2a subtype, suggesting against immune deviation to Th2 as anetiology (24). Linkage analysis of the effect of M. bovis treatmenton NOD mice was performed in a backcross to BALB/c mice, andidentified loci on chromosomes 1 and 17 (25). Cha et al. (26) foundthat NOD Idd3/5 intervals introgressed onto the B6 backgroundallowed detection of ANA, as well, suggesting that Idd3 and 5intervals could contribute to the development of ANA production.Although these loci do not all overlap with the genetic intervalsidentified in our model, the general picture emerging from ourstudies combined with these previous studies is that either envi-ronmental (M. bovis, and possibly different mouse-room flora indifferent colonies) or genetic perturbations of the NOD immunesystem can drive it toward an SLE-like autoantibody response. Asin many autoimmune syndromes, the pathological contribution ofthe autoantibodies is open to question; we will test the pathoge-nicity of the autoantibodies in future studies using a transfersystem.
The outcross studies that we performed provided insights intothe genetic control of liver disease in NOD.c3c4 mice. Becausenearly 50% of the (NOD.c3c4 � NOD)F1 mice developed diabe-tes, but none developed liver disease, it is possible to conclude that
FIGURE 6. Component immunophenotypes of donor strains: anti-Sm autoantibody formation in NOD.c3c4 and NOD.c4(a) mice. Left side, Protein IPdemonstrates anti-Smith Abs in NOD.c3c4 sera, but not NOD or B6. Lane 1, 14C-labeled standards; lane 2, human anti-Sm-positive control sera; lane 3,control negative; lanes 4–6, three individual NOD.c3c4 (age �44 wk). Lanes 7–10, Control NOD.Idd3/5, NOD, B6, and human Rp/Ku sera. Right side,Anti-Sm Abs in NOD.c4(a) sera. Lanes 1–7, Individual NOD.c4(a) mice, aged 19 wk (lanes 1–3) and 38 wk (lanes 4–7); control positive NOD.c3c4 sera(lane 8); control negative NOD and B6 sera (lanes 9 and 10); high and low m.w. markers (lanes 11 and 12).
2320 GENETIC CONTROL OF AUTOIMMUNE BILIARY DISEASE
by guest on June 8, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

the two disease processes, T1D and the biliary autoimmune pro-cess, cannot occur simultaneously, as if the autoimmune responsefocuses on one organ system. However, the observed result ofdiabetes in the F1 mice could be explained by a recessive mode ofinheritance for the liver disease as opposed to a dominant mode forT1D. Regardless, it is intriguing to speculate that the underlyinggenetic pathway controlling these two different organ-specific au-toimmune syndromes may overlap substantially (require the NODgenetic background), but that the choice of target organ is influ-enced by other loci. This genetic result has implications for un-derstanding the pathogenesis of autoimmune syndromes in hu-mans. The occurrence of a diverse clinical set of autoimmunesyndromes in a human kindred could be explained by a sharedautoimmune-prone genetic background modified by additional ge-netic loci that then focus the clinical manifestations of the break-down of tolerance. Similarly, one can hypothesize that autoim-mune syndromes/diseases that appear radically different from aclinical perspective may share genetic/biochemical pathways thatdiffer only at a few points.
The discovery of a spontaneous, autoimmune biliary tract dis-ease in NOD.c3c4 mice raises the question of analogy to any sim-ilar human disease process. PBC has been described at a greaterincidence in relatives of patients with T1D (3, 4). PBC is a humanautoimmune disease characterized by lymphocytic infiltration into
Table I. NOD.c3c4 transfer data
MouseAge at
TransferAge atHarvest Treatment
Days afterTransfer
LiverDisease
C37 7 wk 133 days PBS 91 �C38 7 wk 133 days PBS 91 �C39 7 wk 133 days Spleen 91 �C40 7 wk 133 days Spleen 91 �C41 7 wk 133 days Spleen 91 �T1 11 wk 166 days Spleen 85 �T2 11 wk 166 days PBS 85 �T3 11 wk 166 days Spleen 85 �T4 11 wk 166 days PBS 85 �T15 13 wk 181 days Spleen 87 �T16 13 wk 181 days Spleen 87 �T17 13 wk 181 days PBS 87 �
FIGURE 7. Anti-DNA Ab production by NOD.c3c4 and related NODcongenic mice. A, IgM anti-dsDNA Abs in male and female mice youngerthan 6 mo; at this age, there was no sex difference in Ab levels. NOD.c3c4mice (n � 19) had a significantly increased IgM dsDNA level comparedwith B6 (n � 16), NOD (n � 4), and NOD.c3(a) (n � 7) (p � 0.003, 0.02,and 0.01); the p value compared with NOD.c4(a) mice (n � 7) was 0.09. B,IgG anti-dsDNA in female congenic mice older than 6 mo. NOD.c3c4 mice(n � 11) had a significantly increased level of IgG dsDNA compared with B6(n � 4) NOD.c3(a) (n � 10), NOD.c4(a) (n � 11), NOD (n � 4), and
New Zealand White (n � 6) mice (p � 0.04, 0.002, 0.001, 0.04, and 0.02,respectively), but equivalent levels to NZB (n � 6) (p � 0.95). C, Male(n � 16) vs female (n � 11) NOD.c3c4 anti-DNA responses in mice agedolder than 8 mo. Female mice had significantly higher IgM dsDNA, IgGssDNA, and IgG dsDNA titers than male NOD.c3c4 mice (p � 0.003, 0.01,and 0.0006, respectively).
FIGURE 8. Splenocytes transfer liver disease to naive recipients. PBS(left) or 25 � 106 whole splenocytes (right) from 40-wk-old NOD.c3c4mice were transferred to irradiated 7- to 13-wk-old NOD.c3c4 recipients.Mice were sacrificed after 3 mo, and livers were examined for pathology.Three experiments were performed. See Table I for complete results.
2321The Journal of Immunology
by guest on June 8, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

the biliary epithelium; progressive biliary epithelial damage result-ing in fibrosis, biliary tract obstruction, and ultimately liver failure;and a specific immune response to mitochondrial enzymes (thepyruvate dehydrogenase complex (PDC)) (27). A series of studiesdemonstrated that the anti-PDC Ab is predominantly directed tothe E2 enzyme of the PDC complex (28–31). A second series ofstudies demonstrated T cell epitopes in the E2 complex, and sup-port the hypothesis that PBC is a T cell-mediated disease driven byPDC-reactive autoimmune T cells with a Th1 phenotype (32–36).As we demonstrate in this study, NOD.c3c4 mice develop a spon-taneous biliary tract disease characterized by lymphocytic portaltract infiltrations, biliary obstruction, and eventual mortality due toobstructive liver failure, and the disease is transferable by spleno-cytes, implicating likely T cell involvement. The major differencebetween NOD.c3c4 mice and human PBC is that NOD.c3c4 micedeveloped biliary epithelial hyperplasia (resulting in cysts) ratherthan the biliary epithelial damage with fibrosis seen in humans.Nonetheless, it is possible that a different pathological response ofmurine biliary epithelia to infiltrating lymphocytes could be due togenetically determined differences in tissue injury response. Topursue this issue, future studies will determine B and T cell re-sponses of NOD.c3c4 mice to murine pyruvate dehydrogenase.
The results from the current study, that a resistance region forone disease (T1D) contains genetic elements that confer increasedsusceptibility to another lethal syndrome (biliary disease), com-plement the recent demonstration that a single NOD Idd suscep-tibility allele can contribute to the pathogenesis of two differentautoimmune diseases, T1D and experimental autoimmune enceph-alomyelitis (37). The concept of the NOD genetic background pro-viding a pan-autoimmune diathesis that is focused by specificmodification is consistent with the observation that the B7-2knockout placed on the NOD background develops an autoim-mune demyelinating disease, and that HLA-DQ8 placed on theNOD background produces autoimmune myocarditis (38, 39). Thedevelopment of biliary tract disease, Sjogren’s syndrome (40), thy-roiditis (41), and autoimmune diabetes in a set of genetically sim-ilar NOD and NOD congenic strains serves as a model for humanpedigrees containing multiple autoimmune diseases, including di-abetes and primary biliary cirrhosis. Our results interpreted in thecontext of these other studies demonstrate that a genetically de-termined autoimmune pathway emerges from the NOD back-ground genes, which can be changed to a completely differentautoimmune phenotype by the introduction of different chromo-somal intervals.
References1. Lin, J. P., J. M. Cash, S. Z. Doyle, S. Peden, K. Kanik, C. I. Amos, S. J. Bale,
and R. L. Wilder. 1998. Familial clustering of rheumatoid arthritis with otherautoimmune diseases. Hum. Genet. 103:475.
2. Griffiths, M. M., J. A. Encinas, E. F. Remmers, V. K. Kuchroo, and R. L. Wilder.1999. Mapping autoimmunity genes. Curr. Opin. Immunol. 11:689.
3. Ko, G. T., C. C. Szeto, V. T. Yeung, C. C. Chow, H. Chan, and C. S. Cockram.1996. Autoimmune polyglandular syndrome and primary biliary cirrhosis.Br. J. Clin. Pract. 50:344.
4. Humbert, P., and J. L. Dupond. 1988. Multiple autoimmune syndromes. Ann.Med. Interne (Paris) 139:159.
5. Vyse, T. J., and J. A. Todd. 1996. Genetic analysis of autoimmune disease. Cell85:311.
6. Becker, K. G., R. M. Simon, J. E. Bailey-Wilson, B. Freidlin, W. E. Biddison,H. F. McFarland, and J. M. Trent. 1998. Clustering of non-major histocompati-bility complex susceptibility candidate loci in human autoimmune diseases. Proc.Natl. Acad. Sci. USA 95:9979.
7. Bach, J. F., and D. Mathis. 1997. The NOD mouse. Res. Immunol. 148:285.8. Todd, J. A., T. J. Aitman, R. J. Cornall, S. Ghosh, J. R. Hall, C. M. Hearne,
A. M. Knight, J. M. Love, M. A. McAleer, J. B. Prins, et al. 1991. Geneticanalysis of autoimmune type 1 diabetes mellitus in mice. Nature 351:542.
9. Lyons, P. A., W. W. Hancock, P. Denny, C. J. Lord, N. J. Hill, N. Armitage,T. Siegmund, J. A. Todd, M. S. Phillips, J. F. Hess, et al. 2000. The NOD Idd9genetic interval influences the pathogenicity of insulitis and contains molecularvariants of Cd30, Tnfr2, and Cd137. Immunity 13:107.
10. Podolin, P. L., P. Denny, C. J. Lord, N. J. Hill, J. A. Todd, L. B. Peterson,L. S. Wicker, and P. A. Lyons. 1997. Congenic mapping of the insulin-dependentdiabetes (Idd) gene, Idd10, localizes two genes mediating the Idd10 effect andeliminates the candidate Fcgr1. J. Immunol. 159:1835.
11. Podolin, P. L., P. Denny, N. Armitage, C. J. Lord, N. J. Hill, E. R. Levy,L. B. Peterson, J. A. Todd, L. S. Wicker, and P. A. Lyons. 1998. Localization oftwo insulin-dependent diabetes (Idd) genes to the Idd10 region on mouse chro-mosome 3. Mamm. Genome 9:283.
12. Falkner, D., J. Wilson, N. Fertig, K. Clawson, T. A. Medsger, Jr., andP. A. Morel. 2000. Studies of HLA-DR and DQ alleles in systemic sclerosispatients with autoantibodies to RNA polymerases and U3-RNP (fibrillarin).J. Rheumatol. 27:1196.
13. Gilkeson, G. S., J. P. Grudier, D. G. Karounos, and D. S. Pisetsky. 1989. Induc-tion of anti-double stranded DNA antibodies in normal mice by immunizationwith bacterial DNA. J. Immunol. 142:1482.
14. Wicker, L. S., B. J. Miller, and Y. Mullen. 1986. Transfer of autoimmune dia-betes mellitus with splenocytes from nonobese diabetic (NOD) mice. Diabetes35:855.
15. Wicker, L. S., J. A. Todd, and L. B. Peterson. 1995. Genetic control of autoim-mune diabetes in the NOD mouse. Annu. Rev. Immunol. 13:179.
16. Eisneberg, R. A., S. Y. Craven, R. W. Warren, and P. L. Cohen. 1987. Stochasticcontrol of anti-Sm autoantibodies in MRL/Mp-lpr/lpr mice. J. Clin. Invest.80:691.
17. Mohan, C., L. Morel, P. Yang, and E. K. Wakeland. 1997. Genetic dissection ofsystemic lupus erythematosis pathogenesis: Sle2 on murine chromosome 4 leadsto B cell hyperactivity. J. Immunol. 159:454.
18. Morel, L., B. P. Croker, K. R. Blenman, C. Mohan, G. Huang, G. Gilkeson, andE. K. Wakeland. 2000. Genetic reconstitution of systemic lupus erythematosisimmunopathology with polycongenic murine strains. Proc. Natl. Acad. Sci. USA97:6670.
19. Craft, J. E. 1997. Anti-snRNP antibodies. In Dubois’ Lupus Erythematosis, 5thEd. D. J. Wallace and B. H. Hahn, eds. Williams and Wilkins, Baltimore, pp.457–470.
20. Arai, T., K. Noguchi, N. Machida, M. Sasaki, Y. Oki, and K. Komeda. 1989.Detection of nuclear protein antigens to antinuclear antibodies in serum of NODmouse. Jikken Dobutsu 38:159.
Table II. Genotypic and phenotypic summary
NOD NOD.c3a NOD.c4a B6.H2G7(NOD.c3c4� NOD)F1
(NOD.c3c4 �B6.H2G7)F1 NOD.c3c4
GenotypeC3 N/N B/B N/N B/B B/N B/B B/BC4 N/N N/N B/B B/B B/N B/B B/BGenome N/N N/N N/N B/Ba N/N N/B N/NAutoimmune phenotypesANA � � ��� � ��� � ����Anti-dsDNA � � � � � � ����Anti-Sm � � �� � �� � ��Pathological phenotypesDiabetes ��� � � � �� � �Pancreatic insulitis ���� � ���� � �� � �Lymphocytic biliary infiltrates � �� � � � � ����
a Except the MHC region including I-Ag7. �, 0–25%; ��, 25–50%; ���, 50–75%; ����, �75%.
2322 GENETIC CONTROL OF AUTOIMMUNE BILIARY DISEASE
by guest on June 8, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

21. Humphreys-Beher, M. G., L. Brinkley, K. R. Purushotham, P. L. Wang,Y. Nakagawa, D. Dusek, M. Kerr, N. Chegini, and E. K. Chan. 1993. Charac-terization of antinuclear autoantibodies present in the serum from nonobese di-abetic (NOD) mice. Clin. Immunol. Immunopathol. 68:350.
22. Baxter, A. G., A. C. Horsfall, D. Healey, P. Ozegbe, S. Day, D. G. Williams, andA. Cooke. 1994. Mycobacteria precipitate an SLE-like syndrome in diabetes-prone NOD mice. Immunology 83:227.
23. Baxter, A. G., D. Healey, and A. Cooke. 1994. Mycobacteria precipitate auto-immune rheumatic disease in NOD mice via an adjuvant-like activity. Scand.J. Immunol. 39:602.
24. Horsfall, A. C., R. Howson, P. Silveira, D. G. Williams, and A. G. Baxter. 1998.Characterization and specificity of B-cell responses in lupus induced by Myco-bacterium bovis in NOD/Lt mice. Immunology 95:8.
25. Jordan, M. A., P. A. Silveira, D. P. Shepherd, C. Chu, S. J. Kinder, J. Chen,L. J. Palmisano, L. D. Poulton, and A. G. Baxter. 2000. Linkage analysis ofsystemic lupus erythematosus induced in diabetes-prone nonobese diabetic miceby Mycobacterium bovis. J. Immunol. 165:1673.
26. Cha, S., H. Nagashima, V. B. Brown, A. B. Peck, and M. G. Humphreys-Beher.2002. Two NOD Idd-associated intervals contribute synergistically to the devel-opment of autoimmune exocrinopathy (Sjogren’s syndrome) on a healthy murinebackground. Arthritis Rheum. 46:1390.
27. Yeaman, S. J., J. A. Kirby, and D. E. Jones. 2000. Autoreactive responses topyruvate dehydrogenase complex in the pathogenesis of primary biliary cirrhosis.Immunol. Rev. 174:238.
28. Van de Water, J., D. Fregeau, P. Davis, A. Ansari, D. Danner, P. Leung,R. Coppel, and M. E. Gershwin. 1988. Autoantibodies of primary biliary cirrhosisrecognize dihydrolipoamide acetyltransferase and inhibit enzyme function. J. Im-munol. 141:2321.
29. Frazer, I. H., I. R. Mackay, T. W. Jordan, S. Whittingham, and S. Marzuki. 1985.Reactivity of anti-mitochondrial autoantibodies in primary biliary cirrhosis: def-inition of two novel mitochondrial polypeptide autoantigens. J. Immunol.135:1739.
30. Van de Water, J., A. Cooper, C. D. Surh, R. Coppel, D. Danner, A. Ansari,R. Dickson, and M. E. Gershwin. 1989. Detection of autoantibodies to recom-binant mitochondrial proteins in patients with primary biliary cirrhosis. N. Engl.J. Med. 320:1377.
31. Gershwin, M. E., I. R. Mackay, A. Sturgess, and R. L. Coppel. 1987. Identifi-cation and specificity of a cDNA encoding the 70 kd mitochondrial antigen rec-ognized in primary biliary cirrhosis. J. Immunol. 138:3525.
32. Krams, S. M., J. Van de Water, R. L. Coppel, C. Esquivel, J. Roberts, A. Ansari,and M. E. Gershwin. 1990. Analysis of hepatic T lymphocyte and immunoglob-ulin deposits in patients with primary biliary cirrhosis. Hepatology 12:306.
33. Van de Water, J., A. A. Ansari, C. D. Surh, R. Coppel, T. Roche, H. Bonkovsky,M. Kaplan, and M. E. Gershwin. 1991. Evidence for the targeting by 2-oxo-dehydrogenase enzymes in the T cell response of primary biliary cirrhosis. J. Im-munol. 146:89.
34. Jones, D. E., J. M. Palmer, O. F. James, S. J. Yeaman, M. F. Bassendine, andA. G. Diamond. 1995. T-cell responses to the components of pyruvate dehydro-genase complex in primary biliary cirrhosis. Hepatology 21:995.
35. Lohr, H., B. Fleischer, G. Gerken, S. J. Yeaman, K. H. Meyer zum Buschenfelde,and M. Manns. 1993. Autoreactive liver-infiltrating T cells in primary biliarycirrhosis recognize inner mitochondrial epitopes and the pyruvate dehydrogenasecomplex. J. Hepatol. 18:322.
36. Palmer, J. M., A. G. Diamond, S. J. Yeaman, M. F. Bassendine, and D. E. Jones.1999. T cell responses to the putative dominant autoepitope in primary biliarycirrhosis (PBC). Clin. Exp. Immunol. 116:133.
37. Encinas, J. A., L. S. Wicker, L. B. Peterson, A. Mukasa, C. Teuscher, R. Sobel,H. L. Weiner, C. E. Seidman, J. G. Seidman, and V. K. Kuchroo. 1999. QTLinfluencing autoimmune diabetes and encephalomyelitis map to a 0.15-cM regioncontaining IL2. Nat. Genet. 21:158.
38. Elliott, J. F., J. Liu, Z.Yuan, N. Bautista-Lopez, S. L. Wallback, K. Suzuki,D. Rayner, P. Nation, M. A. Robertson, G. Liu, and K. M. Kavanagh. 2003.Autoimmune cardiomyopathy and heart block develop spontaneously in HLA-DQ8 transgenic IA� knockout NOD mice. Proc. Natl. Acad. Sci. USA100:13447.
39. Salomon, B., L. Rhee, H. Bour-Jordan, H. Hsin, A. Montag, B. Soliven,J. Arcella, A. M. Girvin, J. Padilla, S. D. Miller, and J. A. Bluestone. 2001.Development of spontaneous autoimmune peripheral polyneuropathy in B7-2-deficient NOD mice. J. Exp. Med. 194:677.
40. Bernard, N. F., F. Ertug, and H. Margolese. 1992. High incidence of thyroiditisand anti-thyroid autoantibodies in NOD mice. Diabetes 41:40.
41. Goillot, E., M. Mutin, and J. L. Touraine. 1991. Sialadenitis in nonobese diabeticmice: transfer into syngeneic healthy neonates by splenic T lymphocytes. Clin.Immunol. Immunopathol. 59:462.
2323The Journal of Immunology
by guest on June 8, 2018http://w
ww
.jimm
unol.org/D
ownloaded from