head shape variation in response to diet in triatoma ...hugoabenitez.com/pdf/lunardietal2017.pdf ·...
TRANSCRIPT

R
H(v
RWa
Bb
a
ARRAAC
KMSDV
1
pa1(p(tal
mcp
h0
Zoologischer Anzeiger 267 (2017) 187–193
Contents lists available at ScienceDirect
Zoologischer Anzeiger
jou rn al hom epage : w ww.elsev ier .com/ locate / j cz
esearch paper
ead shape variation in response to diet in Triatoma williamiHemiptera, Reduviidae: Triatominae), a possible Chagas diseaseector of legal Amazônia
osaline Rocha Lunardia,∗, Hugo A. Benítezb, Thaís Peres Câmaraa, Letícia Pinho Gomesa,agner Welber Arrais-Silvaa
Laboratório de Parasitos e Vetores, Campus Universitário do Araguaia, Universidade Federal de Mato Grosso, 78600-000, Barra do Garc as, Mato Grosso,razilDepartamento de Recursos Ambientales, Facultad de Ciencias Agronómicas, Universidad de Tarapacá, Arica, Chile
r t i c l e i n f o
rticle history:eceived 11 November 2016eceived in revised form 3 April 2017ccepted 4 April 2017vailable online 5 April 2017orresponding Editor: Sven Bradler.
eywords:orphometrics
a b s t r a c t
Triatoma williami is naturally infected by Trypanosoma cruzi, the ethiological agent of Chagas disease,the most significant cause of morbidity and mortality in South and Central America. There is a lack ofdata demonstrating the bionomic aspects, the vectorial competence or the natural ecotope and the wildhosts of T. williami, although this species may be considered secondary vector because it maintains theirwild condition and shows synanthropic potential, colonizing the peridomiciles and frequently invadingthe household. The synanthropy represents a secondary adaptation by sylvatic species in response toenvironmental changes, and this adaptability to human dwellings depends on the plasticity of triatomine.This study describes for the first time the shape plasticity of T. williami in response to diet (blood). Two
hapeiet influenceector ecology
groups of triatomines were exposed to a sole blood meal source, mammalian and bird, to assess the effectof blood meal source on head morphology. The variation was analyzed using geometric morphometrictools and showed that T. williami has higher morphological variation in shape associated with bloodsource. This results represent an important representation of the shape adaptation of T. williami and acontribution to the knowledge of morphometrics variation of insect vector of diseases.
© 2017 Elsevier GmbH. All rights reserved.
. Introduction
Morphological adaptation has been a core element in com-ared biology, especially in the study of organismal diversificationnd evolutionary innovation (Pigliucci et al., 2006; West-Eberhard,989; West-Eberhard, 2005). It is well known that all organismsincluding the sessile animals) exhibit at least some degree oflasticity, that is, their function is affected by external conditionsFoster, 1979; Mokady et al., 1999). Despite the obvious adap-ive significance of plasticity, it is clearly not without limits andpparent constraints. Organisms are not infinitely plastic but ratherimited in the range of environments they can respond to.
Triatomines are well known as a high plastic species, where
orphological analyses have been used to understand the pro-ess of plasticity and infer about the changes of this morphometrichenomena (Dujardin et al., 2009; Hernández et al., 2011; Nattero
∗ Corresponding author.E-mail address: [email protected] (R.R. Lunardi).
ttp://dx.doi.org/10.1016/j.jcz.2017.04.001044-5231/© 2017 Elsevier GmbH. All rights reserved.
et al., 2013). In the last few years’ studies about phenotypic plas-ticity using morphometric have been analysing the genetic effectof this pattern. Nattero et al. (2013) found that mostly the effect ofvariation of these species are related to quality of blood ingestedduring nymph instars and adult stages. In the last decades, evolu-tionary biologists have combined accurate mathematical tools thatreduce error in determining the morphological changes and effectson species (Bookstein, 1991; Rohlf and Marcus, 1993; Wagner,1984). One of the methods developed from this process is theshape analysis based on statistical multivariate techniques andnovel visualisation methods. Geometric morphometrics (GMM) is acoordinate-based method, so that their primary data are 2D or 3DCartesian coordinates of anatomically distinguishable landmarks(i.e. discrete anatomical points that are homologous among all theindividuals under analysis) (Adams et al., 2004; Adams et al., 2013;Bookstein, 1998; Rohlf and Marcus, 1993; Rohlf and Slice, 1990).
In the present study we will analyse the influence of the diet atthe head shape variation in one of the species of the subfamilyTriatominae.
1 her An
2tam2osdfrtaTttfc
si1dvomngiWfle
ettvtchtpea
qt
2
2
biGpstimgta
88 R.R. Lunardi et al. / Zoologisc
This subfamily includes 18 genera and 141 species (Jurberg et al.,010). These insects are obligatory hematophagous and are poten-ial vectors of Trypanosoma cruzi (Chagas, 1909), that is the etiologicgent of Chagas Disease or American trypanosomiasis, one of theost important parasitic diseases in Latin America (Soares et al.,
014). Although triatomines are primarily sylvatic, some speciesccasionally are attracted to the interior of the dwellings and a fewpecies, such as Triatoma infestans and Rhodnius prolixus, are mainlyomestic (Depickère et al., 2012; Noireau et al., 2005). The speciesound in sylvatic habitats generally play a limited epidemiologicalole compared to domestic species, but they may act as synan-hropic vectors of T. cruzi and as a reinfestation source of dwellingsfter insecticide spraying (Marti et al., 2014; Noireau et al., 2000).hese species are considered secondary vectors because they main-ain their wild condition, they are generally autochthonous andhey show synanthropic potential, colonizing the peridomiciles andrequently invading the household where they sometimes establisholonies (Noireau et al., 2005; Rodríguez et al., 2013).
The synanthropy represents a secondary adaptation by sylvaticpecies in response to environmental changes, and this adaptabil-ty depends on the plasticity of triatomines (Forattini and Service,989). In general, all triatomines are considered plastic insects thatevelop rapid morphological changes in response to environmentalariability. The organisms that have this flexibility in the expressionf a character have a potential advantage for dealing with environ-ental heterogeneity (Ernande and Dieckmann, 2004). This ability
amed phenotypic plasticity, is defined as the capacity of a singleenotype to exhibit variable phenotypes in response to variationn the environment (Fordyce, 2006; Whitman and Agrawal, 2009;
hitman and Ananthakrishnan, 2009), and is considered essentialor understanding the development and maintenance of morpho-ogical variation (Nattero et al., 2013a; Pigliucci, 2005; Pigliuccit al., 2006).
A better understanding of different aspects in adaptation andvolution of Triatominae populations may contribute to the selec-ion of appropriate vector control strategies in endemic areas. Inhis way, the quantitative study of phenotypic traits and nutritionalariables could be informative about the population structure andhe relative mobility of vectors (Hernández et al., 2011). In thisontext, morphological plasticity in response to blood source mayave important ecological consequence and may predict the synan-hopic success of species. Blood source would influence on differenthenotypic dimensions in ontogenetic trajectory, and representnvironments that are different enough to cause significant vari-tions in head shape and size of triatomines (Nattero et al., 2013a).
Therefore, the principal aims of this study was investigate anduantify the phenotypic shape variation of T. williami triggered bywo different blood sources, in mammals and birds.
. Materials and methods
.1. Data collection
Adult specimens of T. williami were spontaneously collectedetween May and August 2012 in the outside of a military build-
ng located in a natural reserve, in the municipality of Barra doarc as, Mato Grosso. The nymphs of the first generation of wildarental were randomly selected, forming two groups with 100pecimens each. The groups were fed at a determined source fromhe first nymphal stage and therefore there was no selection accord-ng to sex (macroscopically visible only in adults). The accumulated
ortality was 57% in the group fed in mammals and 84% in theroup fed on birds, until the N5 stage. The insects were identifiedhrough their external morphological characters, according to Lentnd Wygodzinsky (1979). They were maintained in a controlled
zeiger 267 (2017) 187–193
environment at a temperature of 27 ± 1 ◦C and relative humidityof 70% ± 10, fed weekly on quail (Coturnix coturnix). The bird group(AV) was fed exclusively on quail (average feeding time 32.9 min)and the mammal group (MAM) exclusively on mice (average feed-ing time 50.7 min) (Mus musculus).
2.2. Morphometric analysis
Eight anatomical landmarks (based on their anatomical traits)were digitized in the head of T. williami, in dorsal and profile view inorder to have the complete head variation in a 2D view (Fig. 1), usingthe software program tpsDIG V2.17 (Rohlf, 2013), for which coor-dinates were generated from all the landmarks and the head shapeinformation was extracted using a full Procrustes fit. Procrustessuperimposition is a procedure that removes the information ofsize, position and orientation in order to obtain shape variables(Rohlf and Slice, 1990).
For the dorsal view, the symmetry was taken into account, andthe symmetric component of shape was calculated from the aver-ages of original and reflected copies (Klingenberg et al., 2002). Inorder to avoid any inaccuracy in the sampling and landmarkingprocess it is critically important to perform a procedure to testthe measurement error (ME), which was conducted comparing theoriginal dataset with a control of repeated measures comparingthe values of the MS of the individual values with the error by aProcustes ANOVA (e.g. Klingenberg and McIntyre, 1998).
In order to quantify the shape variation related with the shapedimensions, a principal component analysis (PCA) was carried out,and to evaluate the differences between sexual dimorphism andtheir differences in diets a canonical variate analysis (CVA) wasperformed Both analysed traits were evaluated by a two wayANOVA in order to find a significant influence in shape. The resultswere reported as Procrustes distances and the respective P val-ues for these distances, after permutation tests (10000 runs), werereported. All the above-mentioned analyses were performed usingMorphoJ v1.05d (Klingenberg, 2011).
2.3. Size and centroid size comparison
A proxy of size was used in order to get the maximum of infor-mation from the morphological plasticity effect of the blood mealdifferences. The proxy of size was the centroid size (Bookstein,1986, 1989; Mosimann, 1970). The centroid size (CS) defined as thesquare root of the sum of squared distances of a set of landmarksfrom their centroid (Dryden and Mardia, 1998). The level of statis-tic differentiation between shape and centroid size was assessedby computing a two way ANOVA using two factors, sex and foodsource. The results were reported as sums of squares (SS) and meansquares (MS), which are dimensionless (Arnqvist and Martensson,1998; Cardini and Elton, 2007).
3. Results
The Procrustes ANOVA for assessing the measurement error ofboth views (dorsal and profile) showed that the MS for individ-ual variation exceeded the measurement error (Table 1). The PCAshows that the first few PC’s accumulated most shape variationin very few dimensions for both views. In the head dorsal view,the first three PC’s account for 80.3% (PC1: 50.3%, PC2: 21.1%, PC3:8.9%) and for the head profile view the first two PC’s account for62.3% (PC1: 27.4%, PC2: 21.4%, PC3: 13.4%) providing a reasonableapproximation of the total amount of variation.
The average head shape shows that blood feed affects the mor-phology and they are also clearly differentiated by sex. In theindividuals (male and female average morphology) that were feed-ing with the mammal blood, the variation at the dorsal view was

R.R. Lunardi et al. / Zoologischer Anzeiger 267 (2017) 187–193 189
Fig. 1. Representation of the 8 morphological landmarks identified in two head views of Triatoma williami. A: Dorsal View, B: Profile View.
Table 1Measurement error procrustes analysis of variance for both centroid size and head shape of Triatoma williami.Sums of squares (SS) and mean squares (MS) are in units ofProcrustes distances (dimensionless).
Dorsal View
Centroid size
Effect SS MS df F P Pillai tr. P(param)
Individual 2185488.05 40472.001 54 5050.79 <0.0001 n.a n.aError 1 440.714998 8.013 55
ShapeEffectIndividual 0.03976375 6.14E-05 648 40.82 <0.0001 11.09 <0.0001Error 1 0.00099226 1.5034E-06 660
Profile ViewCentroid size
Effect SS MS df F P Pillai tr. P(param)Individual 1101127.227 22471.9 49 1821.82 <0.0001 n.a n.aError 1 616.745492 12.33491 50
Shape
pamamOn
aiapns
aGCTvtoe
EffectIndividual 0.05579351 9.49E-05 588
Error 1 0.00067368 1.1228E-06 600
rincipally narrowing in the landmarks 1 and 8 (Apex of gena) and symmetric left broadening of the central landmaks of the head,aking the morphology a bit narrow between the tip of the head
nd the intersection of the eye (Fig. 2A). For the profile view, theorphology shows a broadening of the landmarks 4 and 5 withcelo anterior and the intersection of the head base and neck, andarrowing in the landmark 1 and 8 (Fig. 2B).
For the individuals feeding the bird blood, the dorsal shape show vector movement of the landmarks 1 and 8 showing a broaden-ng pattern and global expansion of to the central landmarks giving
thinner morphology. For the profile shape the landmarks dis-lacement were principally from the central landmarks. Where aarrowed variation was noticeable of the landmark 5 on the inter-ection of head base and neck.
The grouped CVA showed a strong differentiation between dietnd sex that were quantified comparing the Procustes distances.raphically this variation can also be observed using the first twoVs are aligned with the major axes of variation among groups.herefore, they account for the maximum amount of among-groupariance relative to within-group variance (Fig. 3a and b). In order
o confirm the significance of the diet and sex variation the ANOVAf the two investigated positions, showed highly significant differ-nces for shape between sex and diet, nevertheless, the centroid84.51 <0.0001 11.61 <0.0001
size only shows differences between the sex and not clear differ-ences on the diet (Table 2).
4. Discussion
This study has used geometric morphometrics to analyse thephenotypic plasticity of the T. williami as a consequence of the bloodsource diet in mammals and birds. This tool was found to be usefulfor exploring and evaluating the levels of morphological variationand sexual shape dimorphism, confirming a significant variationdue to the blood sources in mammals and birds.
Our results demonstrate that the diets on two food sources, withcomplete different nutritional background (mammal and birds)induced phenotypic plasticity. Findings confirm previous studies inT. infestans suggesting that morphological expression on the pheno-type is a consequence of developmental allocation to tissue growththat maintains growth and development of head (Nattero et al.,2013a; Nattero et al., 2013b). The individuals that are feeding onthe mammals blood shown a wider and shorter morphology effect
of the narrowed landmarks 1 and 8 in contrast to the individualsfeeding on bird blood, where in accordance with previous studies ofNattero et al. (2013a), the effect of the bird diet on the morphologygenerates a thinner head shape (Fig. 2).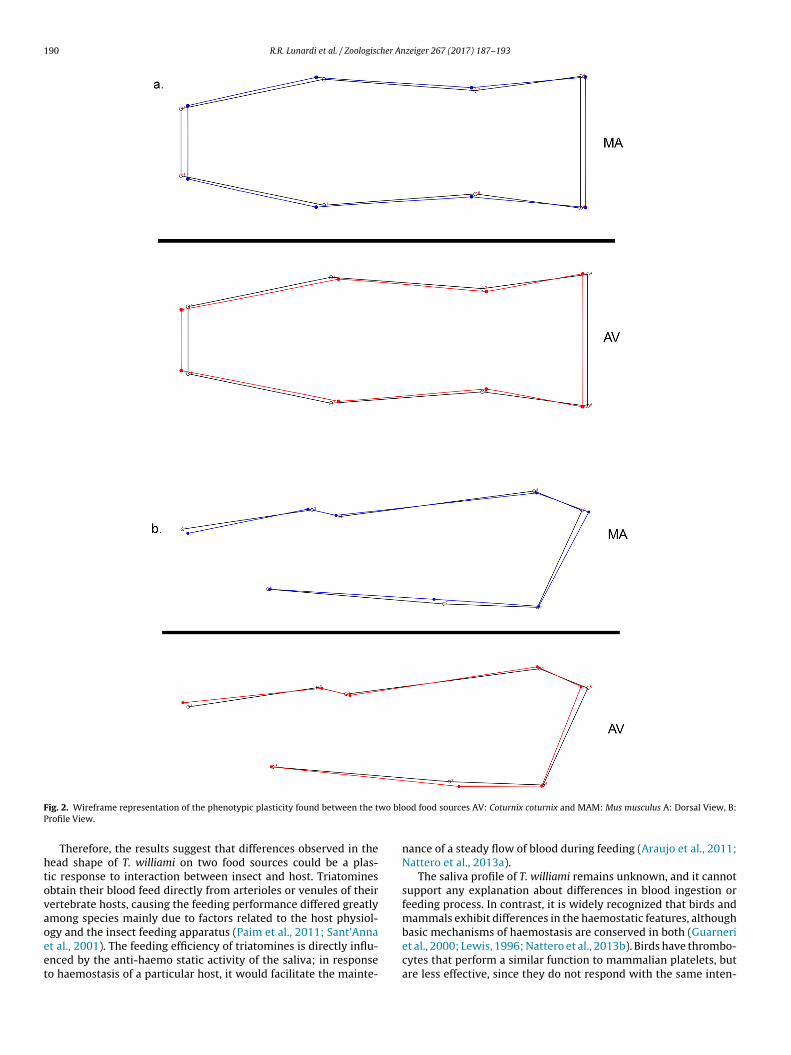
190 R.R. Lunardi et al. / Zoologischer Anzeiger 267 (2017) 187–193
F wo bloP
htovaoeet
ig. 2. Wireframe representation of the phenotypic plasticity found between the trofile View.
Therefore, the results suggest that differences observed in theead shape of T. williami on two food sources could be a plas-ic response to interaction between insect and host. Triatominesbtain their blood feed directly from arterioles or venules of theirertebrate hosts, causing the feeding performance differed greatlymong species mainly due to factors related to the host physiol-
gy and the insect feeding apparatus (Paim et al., 2011; Sant’Annat al., 2001). The feeding efficiency of triatomines is directly influ-nced by the anti-haemo static activity of the saliva; in responseo haemostasis of a particular host, it would facilitate the mainte-od food sources AV: Coturnix coturnix and MAM: Mus musculus A: Dorsal View, B:
nance of a steady flow of blood during feeding (Araujo et al., 2011;Nattero et al., 2013a).
The saliva profile of T. williami remains unknown, and it cannotsupport any explanation about differences in blood ingestion orfeeding process. In contrast, it is widely recognized that birds andmammals exhibit differences in the haemostatic features, although
basic mechanisms of haemostasis are conserved in both (Guarneriet al., 2000; Lewis, 1996; Nattero et al., 2013b). Birds have thrombo-cytes that perform a similar function to mammalian platelets, butare less effective, since they do not respond with the same inten-
R.R. Lunardi et al. / Zoologischer Anzeiger 267 (2017) 187–193 191
Fig. 3. Canonical variate analysis of the head shape comparing the sexual shape dimorphism and blood food sources shape for the different views A: Dorsal View, B: ProfileView. The wireframe represents the average head shape by sex and blood food source.
Table 2Procrustes ANOVA for both centroid size and shape of Triatomawilliami. Sums of squares (SS) and mean squares (MS)are in units of Procrustes distances (dimensionless).
Dorsal View
Centroid size
Effect SS MS df F P
Sex 468461.3279 468461.3279 1 39.45 <0.0001Blood Source 3337.56206 3337.56206 1 0.28 0.5983Individual 617544.1754 11875.84953 52 n.a n.a
ShapeEffect SS MS df F PSex 0.00147508 0.000245846 6 5.31 <0.0001Blood Source 0.00127951 0.000213252 6 4.61 0.0002Individual 0.00317193 9.7899E-06 324 n.a n.a
Profile ViewCentroid size
Effect SS MS df F PSex 271858.7489 271858.7489 1 47.44 <0.0001Blood Source 8160.938121 8160.938121 1 1.42 0.2387Individual 269339.2275 5730.621863 47 n.a n.a
ShapeEffect SS MS df F P
7
05
05
sel
Sex 0.00306804 0.0002556Blood Source 0.00098303 8.19188E-Individual 0.02428738 4.30627E-
ity to platelet aggregation inducers (Araujo et al., 2009; Araujot al., 2011; Martinez-Abadias et al., 2011). Birds also appear toack some coagulation factors, particularly in the intrinsic coag-
12 5.94 <0.000112 1.9 0.0316564 n.a n.a
ulation system (Lewis, 1996). As a consequence of differences inthe haemostatic mechanisms and blood viscosity, higher in mam-mals compared to birds (Baskurt, 2007), the sucking process of

1 her An
twt
mimhporaDtsbsdipiefbt
waVrGnsoa2
mncia2cwfieT
pocc
A
dCS
R
A
92 R.R. Lunardi et al. / Zoologisc
riatomines may require more mechanical effort in mammals thatas principally reflected on a wider head, the morphology adapted
o more “complicated type of food” (Fig. 2).The differences in blood features presented by birds and mam-
als may affect the ingestion of blood by triatomines. This bloodntake capacity of the insect is promoted by the cibarial pump, the
usculature of which occupies practically the full length of theead (Bennet-Clark, 1963). This powerful pump creates negativeressure between the opening of the food canal and the lumenf the alimentary compartment; therefore, blood viscosity, size ofed blood cells and their capacity to deform can be directly associ-ted with the muscular effort of the cibarial pump (Kingsolver andaniel, 1995). Considering that cibarial pump muscles nearly fill
he head capsule, that viscosity of the diet interferes with the blooducking and that the haemostasis mechanism is different betweenirds and mammals. The phenotypic plasticity observed in the headhape of T. williami could be due to an adaptation to its hosts, withifferent development of the cibarial pump muscles. This possibil-
ty was shown in T. infestans, that had greater development of theump muscles, when fed on mammal compared to birds, result-
ng in higher head shapes changes (Nattero et al., 2013a; Natterot al., 2013b). In addition to the factors stated above, several otheractors may also affect insect feeding, especially on live hosts, thelood diet varies on the same host depending on the blood flow ofhe cannulated vessel (Araujo et al., 2009).
The sexual dimorphism presented in head shape and size of T.illiami (Fig. 3), with females greater than males, occur frequently
mong triatomine species (Gaspe et al., 2012; Nattero et al., 2013a;argas et al., 2006). In many species, shape and size variationareelated with nutritional quality of blood (Feliciangeli et al., 2007;uarneri et al., 2000). It is worth mentioning that T. williami didot show variations in the head centroid size related to the bloodource, unlike the observations made in T. infestans feeding on birdsr mammals, whose head was greater in feeding on mammals,nd the body was greater in those feeding on birds (Nattero et al.,013a).
The lack of association between these sizes would indicateore variation in the insect feeding behavior or in the host blood
utritional value, whereas size is often considered a more labileharacter than shape (Dujardin et al., 2014). Size variation is morenfluenced by environmental factors, whereas shape variation has
stronger genetic component (Klingenberg, 2002; Klingenberg,010; Klingenberg et al., 2010; Klingenberg and Leamy, 2001). Inontrast, T. williami head centroid size was more stable than shape,hen exposed to different blood diet sources. Despite the shapets, the idea of a genetic trait, however, also varies according to thenvironmental conditions, as observed here in T. williami and also. dimidiate (e.g. Dujardin et al., 2009).
We showed for the first time that T. williami has higher mor-hological plasticity in shape associated with blood source, in spitef the presence of stasis in size. However, there still remains thehallenge to test whether plasticity confers a fitness advantage toulminate in domiciliation.
cknowledgements
This work was supported by Fundac ão de Amparo à Pesquisao Estado de Mato Grosso, Conselho Nacional do Desenvolvimentoientífico and Coordenac ão de Aperfeic oamento de Pessoal de Níveluperior.
eferences
dams, D.C., Rohlf, F.J., Slice, D.E., 2004. Geometric morphometrics: ten years ofprogress following the ‘revolution’. Ital. J. Zool. 71, 5–16.
zeiger 267 (2017) 187–193
Adams, D.C., Rohlf, F.J., Slice, D.E., 2013. A field comes of age: geometricmorphometrics in the 21 st century. Hystrix-Ital. J. Mammal. 24, 7–14.
Araujo, R.N., Pereira, M.H., Soares, A.C., Pereira, I.D., Diotaiuti, L., Gontijo, N.F.,Lehane, M.J., Guarneri, A.A., 2009. Effect of intestinal erythrocyte agglutinationon the feeding performance of Triatoma brasiliensis (Hemiptera: Reduviidae). J.Insect Physiol. 55, 862–868.
Araujo, R.N., Soares, A.C., Gontijo, A.F., Guarneri, A.A., Pereira, M.H., Gontijo, N.F.,2011. Electromyogram of the Cibarial Pump and the Feeding Process inHematophagous Hemiptera. INTECH Open Access Publisher.
Arnqvist, G., Martensson, T., 1998. Measurement error in geometricmorphometrics: empirical strategies to assess and reduce its impact onmeasures of shape. Acta Zool. Acad. Sci. Hungaricae 44, 73–96.
Baskurt, O.K., 2007. Handbook of Hemorheology and Hemodynamics. IOS press.Bennet-Clark, H., 1963. Negative pressures produced in the pharyngeal pump of
the blood-sucking bug, Rhodnius prolixus. J. Exp. Biol. 40, 223–229.Bookstein, F.L., 1986. Size and shape spaces for landmark data in two dimensions.
Stat. Sci., 181–222.Bookstein, F.L., 1989. Size and shape: a comment on semantics. Syst. Biol. 38,
173–180.Bookstein, F.L., 1991. Morphometric Tools for Landmark Data: Geometry and
Biology. Cambridge University Press, Cambridge.Bookstein, F.L., 1998. A hundred years of morphometrics. Acta Zool. Acad. Sci.
Hungaricae 44, 7–59.Cardini, A., Elton, S., 2007. Sample size and sampling error in geometric
morphometric studies of size and shape. Zoomorphology 126, 121–134.Depickère, S., Buitrago, R., Sinani, E., Baune, M., Monje, M., Lopez, R., Waleckx, E.,
Chavez, T., Brenière, S.F., 2012. Susceptibility and resistance to deltamethrin ofwild and domestic populations of Triatoma infestans (Reduviidae: triatominae)in Bolivia: new discoveries. Memórias do Instituto Oswaldo Cruz 107,1042–1047.
Dryden, I., Mardia, K., 1998. Statistical Shape Analysis. John Wiley and Son,Chichester.
Dujardin, J.-P., Costa, J., Bustamante, D., Jaramillo, N., Catalá, S., 2009. Decipheringmorphology in Triatominae: the evolutionary signals. Acta Trop. 110, 101–111.
Dujardin, J.-P., Kaba, D., Solano, P., Dupraz, M., McCoy, K., Jaramillo-O, N., 2014.Outline-based morphometrics, an overlooked method in arthropod studies?Infection. Gen. Evol. 28, 704–714.
Ernande, B., Dieckmann, U., 2004. The evolution of phenotypic plasticity inspatially structured environments: implications of intraspecific competition,plasticity costs and environmental characteristics. J. Evol. Biol. 17, 613–628.
Feliciangeli, M.D., Sanchez-Martin, M., Marrero, R., Davies, C., Dujardin, J.-P., 2007.Morphometric evidence for a possible role of Rhodnius prolixus from palm treesin house re-infestation in the State of Barinas (Venezuela). Acta Trop. 101,169–177.
Forattini, O.P., Service, M., 1989. Chagas’ disease and human behavior. Demogr.Vector-borne Dis., 107–120.
Fordyce, J.A., 2006. The evolutionary consequences of ecological interactionsmediated through phenotypic plasticity. J. Exp. Biol. 209, 2377–2383.
Foster, A.B., 1979. Phenotypic plasticity in the reef corals Montastraea annularis(Ellis & solander) and Siderastrea siderea (Ellis & solander). J. Exp. Mar. Biol.Ecol. 39, 25–54.
Gaspe, M., Schachter-Broide, J., Gurevitz, J., Kitron, U., Gürtler, R., Dujardin, J.-P.,2012. Microgeographic spatial structuring of Triatoma infestans (Hemiptera:reduviidae) populations using wing geometric morphometry in the ArgentineChaco. J. Med. Entomol. 49, 504–514.
Guarneri, A.A., Diotaiuti, L., Gontijo, N.F., Gontijo, A.F., Pereira, M.H., 2000.Comparison of feeding behaviour of Triatoma infestans, Triatoma brasiliensisand Triatoma pseudomaculata in different hosts by electronic monitoring of thecibarial pump. J. Insect Physiol. 46, 1121–1127.
Hernández, M.L., Abrahan, L.B., Dujardin, J.P., Gorla, D.E., Catalá, S.S., 2011.Phenotypic variability and population structure of peridomestic Triatomainfestans in rural areas of the arid Chaco (western Argentina): spatial influenceof macro-and microhabitats. Vector-Borne Zoonotic Dis. 11, 503–513.
Kingsolver, J., Daniel, T., 1995. Mechanics of Food Handling by Fluid-feedingInsects, Regulatory Mechanisms in Insect Feeding. Springer, pp. 32–73.
Klingenberg, C.P., Leamy, L.J., 2001. Quantitative genetics of geometric shape in themouse mandible. Evolution 55, 2342–2352.
Klingenberg, C.P., McIntyre, G.S., 1998. Geometric morphometrics ofdevelopmental instability: analyzing patterns of fluctuating asymmetry withprocrustes methods. Evolution 52, 1363–1375.
Klingenberg, C.P., Barluenga, M., Meyer, A., 2002. Shape analysis of symmetricstructures: quantifying variation among individuals and asymmetry. Evolution56, 1909–1920.
Klingenberg, C.P., Debat, V., Roff, D.A., 2010. Quantitative genetics of shape incricket wings: developmental integration in a functional structure. Evolution64, 2935–2951.
Klingenberg, C.P., 2002. Morphometrics and the role of the phenotype in studies ofthe evolution of developmental mechanisms. Gene 287, 3–10.
Klingenberg, C.P., 2010. Evolution and development of shape: integratingquantitative approaches. Nat. Rev. Genet. 11, 623–635.
Klingenberg, C.P., 2011. MorphoJ: an integrated software package for geometric
morphometrics. Mol. Ecol. Resour. 11, 353–357.Lewis, J.H., 1996. Comparative Hemostasis in Vertebrates. Springer, New York.Marti, G., Echeverria, M., Waleckx, E., Susevich, M., Balsalobre, A., Gorla, D., 2014.
Triatominae in furnariid nests of the Argentine Gran Chaco. J. Vector Ecol. 39,66–71.

her An
M
M
M
N
N
N
N
P
P
P
R
differences. Proc. Natl. Acad. Sci. 102, 6543–6549.Whitman, D.W., Agrawal, A.A., 2009. What is phenotypic plasticity and why is it
R.R. Lunardi et al. / Zoologisc
artinez-Abadias, N., Heuze, Y., Wang, Y., Jabs, E.W., Aldridge, K., Richtsmeier, J.T.,2011. FGF/FGFR signaling coordinates skull development by modulatingmagnitude of morphological integration: evidence from apert syndromemouse models. PLoS One 6.
okady, O., Loya, Y., Achituv, Y., Geffen, E., Graur, D., Rozenblatt, S., Brickner, I.,1999. Speciation versus phenotypic plasticity in coral inhabiting barnacles:Darwin’s observations in an ecological context. J. Mol. Evol. 49, 367–375.
osimann, J.E., 1970. Size allometry – size and shape variables withcharacterizations of lognormal and generalized gamma distributions. J. Am.Stat. Assoc. 65, 930–945.
attero, J., Malerba, R., Rodríguez, C.S., Crocco, L., 2013a. Phenotypic plasticity inresponse to food source in Triatoma infestans (Klug 1834) (Hemiptera,Reduviidae: Triatominae). Infect. Genet. Evol. 19, 38–44.
attero, J., Rodríguez, C.S., Crocco, L., 2013b. Effects of blood meal source on foodresource use and reproduction in Triatoma patagonica Del Ponte (Hemiptera,Reduviidae). J. Vector Ecol. 38, 127–133.
oireau, F., Flores, R., Gutierrez, T., Abad-Franch, F., Flores, E., Vargas, F., 2000.Natural ecotopes of Triatoma infestans dark morph and other sylvatictriatomines in the Bolivian Chaco. Trans. R. Soc. Trop. Med. Hyg. 94, 23–27.
oireau, F., Carbajal-de-La-Fuente, A.L., Lopes, C.M., Diotaiuti, L., 2005. Someconsiderations about the ecology of Triatominae. Anais da Academia Brasileirade Ciências 77, 431–436.
aim, R.M., Araújo, R.N., Soares, A.C., Lemos, L.C.D., Tanaka, A.S., Gontijo, N.F.,Lehane, M.J., Pereira, M.H., 2011. Influence of the intestinal anticoagulant inthe feeding performance of triatomine bugs (Hemiptera; Reduviidae). Int. J.Parasitol. 41, 765–773.
igliucci, M., Murren, C.J., Schlichting, C.D., 2006. Phenotypic plasticity and
evolution by genetic assimilation. J. Exp. Biol. 209, 2362–2367.igliucci, M., 2005. Evolution of phenotypic plasticity: where are we going now?Trends Ecol. Evol. 20, 481–486.
odríguez, C.S., Crocco, L., Altamirano, A., Catalá, S., 2013. Changes related togender, geographic population and habitat in the antennal phenotype of
zeiger 267 (2017) 187–193 193
Triatoma patagonica Del Ponte, 1929 (Hemiptera: reduviidae). Acta Trop. 125,143–149.
Rohlf, F.J., Marcus, L.F., 1993. A revolution in morphometrics. Trends Ecol. Evol. 8,129–132.
Rohlf, F.J., Slice, D., 1990. Extensions of the Procustes methods for the optimalsuperimposition of landmarks. Syst. Zool. 39, 40–59.
Rohlf, F.J., 2013. TPSdig, v. 2.17. State University at Stony Brook., NY.Sant’Anna, M.c.R.V., Diotaiuti, L., de Figueiredo Gontijo, A., de Figueiredo Gontijo,
N., Pereira, M.H., 2001. Feeding behaviour of morphologically similar Rhodniusspecies: influence of mechanical characteristics and salivary function. J. InsectPhysiol. 47, 1459–1465.
Soares, A.C., Araújo, R.N., Carvalho-Tavares, J., de Figueiredo Gontijo, N., Pereira,M.H., 2014. Intravital microscopy and image analysis of Rhodnius prolixus(Hemiptera: reduviidae) hematophagy: the challenge of blood intake frommouse skin. Parasitol. Int. 63, 229–236.
Vargas, E., Espitia, C., Patino, C., Pinto, N., Aguilera, G., Jaramillo, C., Bargues, M.D.,Guhl, F., 2006. Genetic structure of Triatoma venosa (Hemiptera: reduviidae):molecular and morphometric evidence. Memórias do Instituto Oswaldo Cruz101, 39–45.
Wagner, G.P., 1984. On the eigenvalue distribution of genetic and phenotypicdispersion matrices – evidence for a nonrandom organization of quantitativecharacter variation. J. Math. Biol. 21, 77–95.
West-Eberhard, M.J., 1989. Phenotypic plasticity and the origins of diversity. Annu.Rev. Ecol. Syst., 249–278.
West-Eberhard, M.J., 2005. Developmental plasticity and the origin of species
important. Phenotypic Plast. Insects 10, 1–63.Whitman, D.W., Ananthakrishnan, T.N., 2009. Phenotypic Plasticity of Insects:
Mechanisms and Consequences. Science Publishers, Inc.