herawati sudoyo - understanding the complexity of genes ... · understanding the complexity of...
TRANSCRIPT

UNDERSTANDING THE COMPLEXITY OF GENES AND DISEASES IN INDONESIAN ARCHIPELAGO
Herawati SudoyoEijkman Institute for Molecular Biology
Jakarta, IndonesiaHealth systems complexity: Bridging Physiome with Health Care through
Computational Modelling Symposium, NITH-NTU, Singapore, 26 January 2015

GENNEKA TUNGGAL IKAUnity in diversity

Indonesia and Infectious Diseases – a Great Challenge to Mitigate Biorisk
§ Problems with emerging and reemerging infectious diseases
§ Most caused mainly by environmental, ecological or demographic factors spread by travel and trade – Indonesia is a maritime country with 17.504 islands, 700 languages, 33 provinces, 230 million population
Problems with people movementRecognize the need to develop, strengthen and maintained the capacity to detect, report and respond to public health events

WHAT ARE WE FACING?
§Indonesia – A very diverse populations –vast genome diversity – disease management complex §Indonesia - A rapidly developing country with serious challenges in infectious (emerging and re-emerging) and zoonotic diseases§New, re-emerging or drug-resistant infections whose incidence in humans has increased within the past two decades or whose incidence threatens to increase in the near future

Indonesia – A rapidly developing country with serious problems in Infectious diseaseMalaria:
15 million cases and 42,000 deaths/year (2005) - highest case number and fatality rate in the world; increasing drug resistant parasites
Tuberculosis:ranked third in TB burden following India and China - TB is third major causes of mortalityEstimation: 269 TB cases/100,000
Dengue: Most important viralborne disease2004: 78,690 cases (CFR- 1.2%)2007: 123,174 cases,1,251 deaths
Hepatitis B:10% of population are carriersModerate-to-high endemic (WHO)

Serotype Legend
4 13 2
42%
30%
8%
20%41%
32%
19%
8%
Makassar 2007-2008N = 111
Merauke 2001
Palembang 1998
Bandung 2002
Yogyakarta 1996
Jayapura 1994Jakarta 2004
Corwin 2001; Suwandono 2006; Porter 2005; Graham 1999; Richards 1997; Sukri 2003
DENV SEROTYPE DISTRIBUTION – SHOWED DIVERSITY
Four antigenically distinct serotypes: infection with one serotype does not provide protection to the other three
Management of Disease is Not Simple, Need a Strong Disease Surveillance
0
50000
100000
150000
200000
250000
1985 1986 1987 1988 1989 1990 1991 1992 1993 1994 1995 1996 1997 1998 1999 2000 2001 2002 2003 2004 2005 2006
C A
S E
Indonesia South-east Asia Region

SCIENCE & TECHNOLOGY IN HEALTH SECURITY –PREPAREDNESS FOR PANDEMIC
RISK ASSESSMENTS§ Molecular Epidemiology:
• Cluster of viral isolates will indicate the presence of new strain
• Surveillance - tracing sources of infection
§ Characteristics of Virus• Alteration of interaction with host
receptors - pandemic need changing in specificity of viral type receptor into human-type
• Change of virulence • Drug resistance

ALL ABOUT COLLABORATION
• Interdisciplinary collaboration between genetics, language, culture and medicine
Genetics
Linguistic
Anthropology Archeology
Medicine
Mathematic

THE AUSTRONESIAN DIASPORA
WMP
WMPCMP
PAPUA
OCEANIC
OCEANIC
SHWNG
§ One of the world largest language group. § Covers a very wide area, from Madagascar Island as the
western most to the Easter Island of Polynesia

EIJKMAN INSTITUTE and THE POPULATION STRUCTURE of the INDONESIAN ARCHIPELAGO
Eijk
man
Inst
itute
• More than 6,000 samples collected• More than 3745 individuals from 35 ethnic
populations examined for mtDNA• 2000 genetic samples from 25 isolated populations of
13 islands for Y STR • Most extensive database on mtDNA sequence
polymorphisms in Indonesian archipelago• Pan Asia SNP initiative – mapping 50K Asian
population using DNA microarray – 234 Indonesian samples
• 200 male samples of Javanese and Batak origin for Rapidly Mutating YSTR
• Forensic population database – STR marker 21 loci, Y filer

§ MtDNA and Y chr diversity of 2740 individuals, 70 communities, 12 islands§ Indonesian genetic diversity
is a direct outcome of complex history of immigration, transitory migrants and populationsTumonggor et al. J Hum Genet. 2013: 58

SETTLEMENT of ISLAND SOUTHEAST ASIATHE FIRST WAVE: FROM AFRICA TO THE EAST
~50,000 ya
Mellars P (2006). Science
The first stage of Indonesian prehistory represent the archipelago’s initial settlement as part of Africa dispersal ~ 50kya
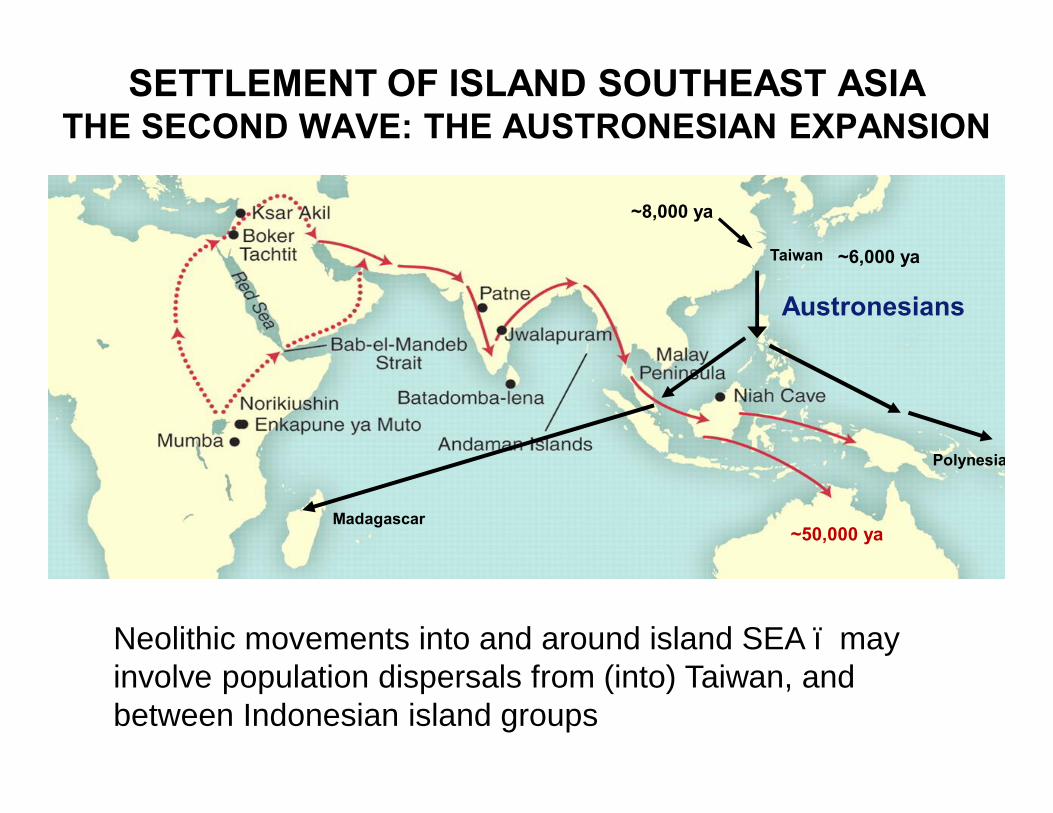
SETTLEMENT OF ISLAND SOUTHEAST ASIATHE SECOND WAVE: THE AUSTRONESIAN EXPANSION
~6,000 ya
Austronesians
~50,000 ya
Taiwan
Madagascar
Polynesia
~8,000 ya
Neolithic movements into and around island SEA – may involve population dispersals from (into) Taiwan, and between Indonesian island groups
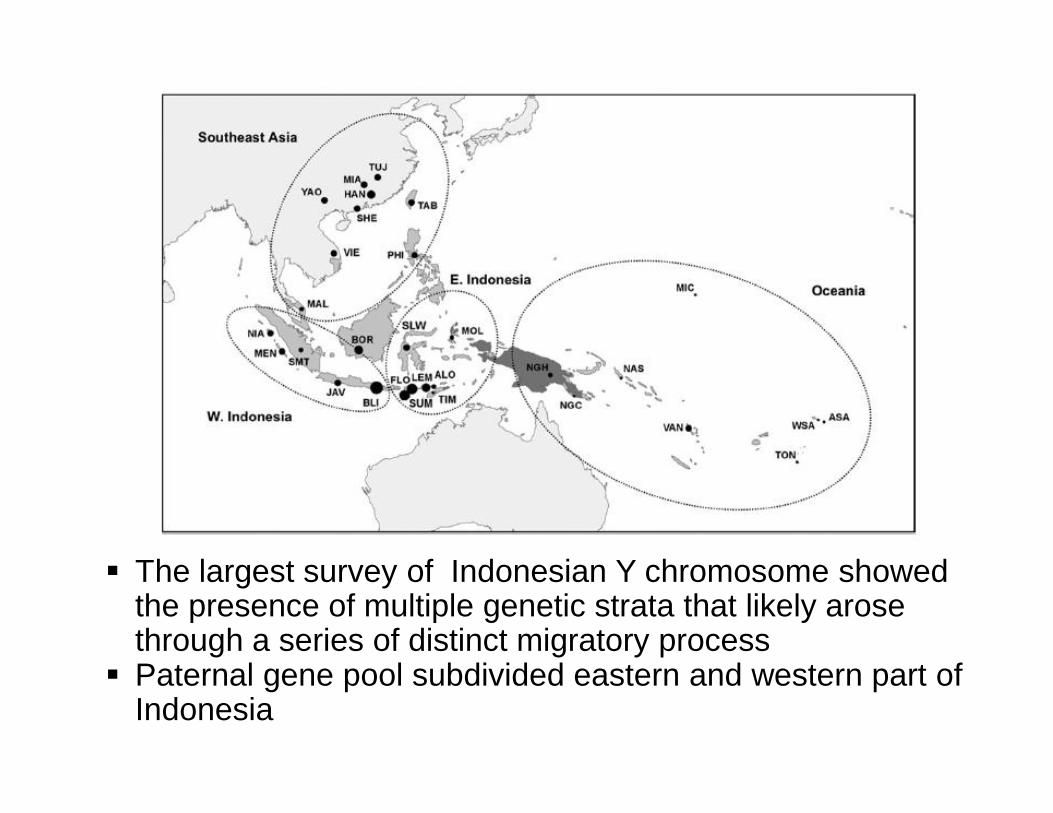
§ The largest survey of Indonesian Y chromosome showed the presence of multiple genetic strata that likely arose through a series of distinct migratory process § Paternal gene pool subdivided eastern and western part of
Indonesia
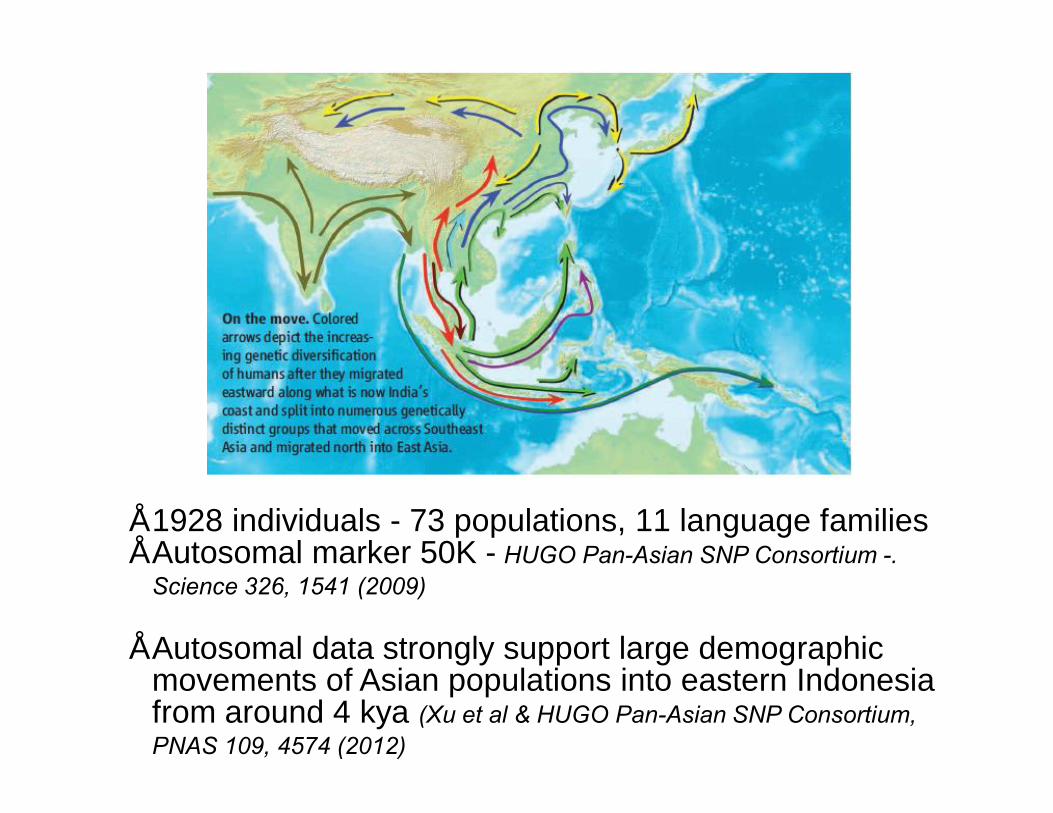
• 1928 individuals - 73 populations, 11 language families • Autosomal marker 50K - HUGO Pan-Asian SNP Consortium -.
Science 326, 1541 (2009)
• Autosomal data strongly support large demographic movements of Asian populations into eastern Indonesia from around 4 kya (Xu et al & HUGO Pan-Asian SNP Consortium, PNAS 109, 4574 (2012)

East Papuan and Alorese
Sumba, Flores, Lembata
Malay,Sumatra, Borneo, Java (Mix with Austroasiatic)

VARIATION OF GENOTYPIC PATTERN OF DISEASES
HEPATITIS AND OVALOCYTOSIS AS A MODEL

• Variation of ethnics in Indonesia: 1. The peopling of the Indonesian archipelago
using mitochondrial-DNA, Y and autosomal chromosome
2. Anthropological and linguistic study -Consistent with cultural and linguistic characteristics
• Variation of susceptibility and resistance to disease (malaria, TB and others)
• Variation of genotypic and phenotypic pattern of diseases (infectious and non-infectious, including hepatitis B/C, dengue,red blood cell disorders )

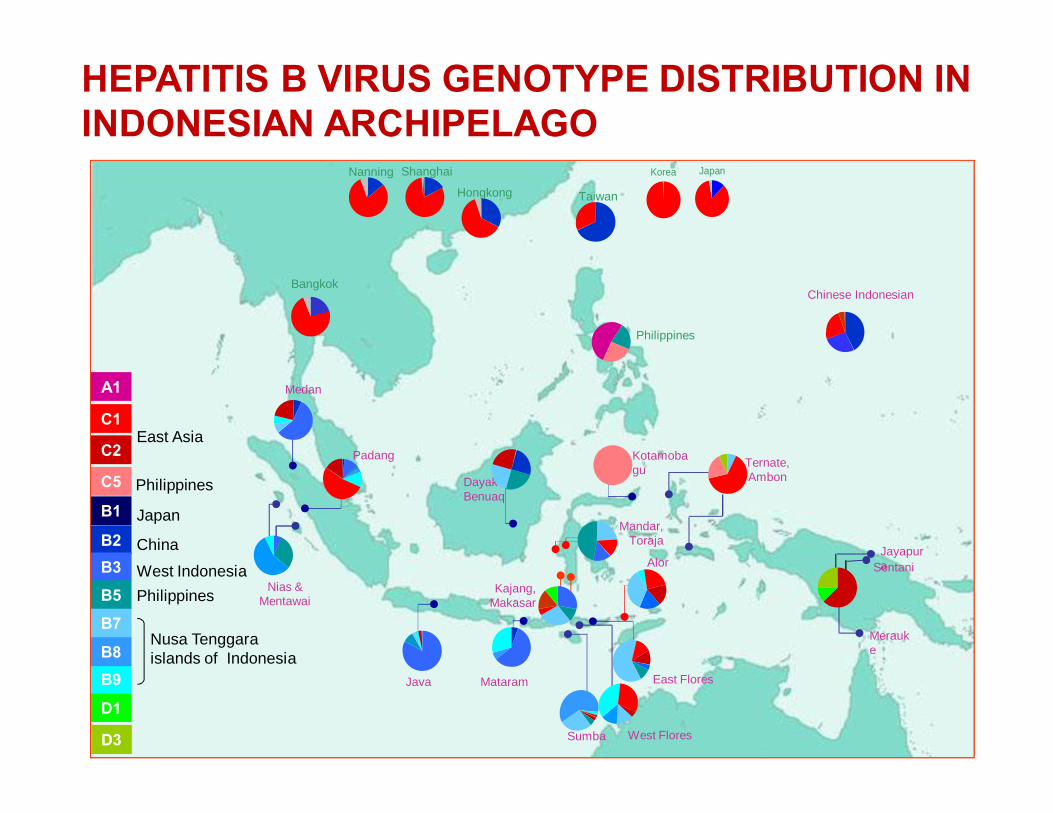
HEPATITIS B VIRUS GENETIC HETEROGENEITY
§ Ten HBV genotypes - A to J have been identified worldwide
§ Shows different geographical distribution, viral characteristics and possibly clinical outcomes and response to treatment, , provide historical information pattern of the local population
Miyakawa & Mizokami, Intervirology 2003;46:329-338
A: North-west Europe, North America, Central Africa; B: Southeast Asia; C: Far east; D: Mediteranean, Middle East, India; E: Sub-Saharan Africa; F: American natives, Polynesian; G.USA samples, H: Mexico

• Indonesian Archipelago:Genotypes B and C are predominant, with sub-genotype B3/adw unique to Indonesia
• Latin America:Genotypes F and H are indigenous in this continent, genotypes A and D might be a mere reflection of a past European migration, and genotypes B and C could represent a consequence of a recent Asian migration.
JapanKorea
Kajang, Makasar
Java
Padang
Nias & Mentawai
Dayak Benuaq
Sumba
Alor
East Flores
Chinese IndonesianBangkok
Mandar, Toraja
West Flores
Kotamobagu
TaiwanHongkong
Nanning Shanghai
JayapuraSentani
Merauke
Mataram
Philippines
Medan
Ternate, Ambon
East Asia
China
Japan
West IndonesiaPhilippines
Nusa Tenggara islands of Indonesia
C1
C2
B2B3B5B7
B9B8
D1
B1
C5
A1
Philippines
D3
HEPATITIS B VIRUS GENOTYPE DISTRIBUTION IN INDONESIAN ARCHIPELAGO

AY226578M57663A1AB246335A1AB241115A1AF297621A1
X70185A2AB222707A2AB126580A2
AB246337A2 DQ463787B6DQ463790B6
DQ463791B6DQ463792B6DQ463788B6
DQ463793B6DQ463794B6
DQ463789B6D23678B1AB073850B1AB073848B1D00329B1
DQ993700B2EU139543B2
DQ993708B2EIH21AB22061B2DQ993711B2EIH45AB2
AP011084B2 AP011089B7AB493833
AB493835AB493836
AB493832EIHB134B3AB033555B3
AP011085B3AB033554B3
D00331B32059B3
1839B3M54923B3
LBY01B7LBY041B7Alr049B7
AP011090B7LBY009B7
AP011091B7FLT32B7
AP011092B7KD048B7
CBL027B7LAR070B7
AP011088B7FLT020B7
AB493830AB493828
AB493829AB493827
AB493831.KDI35B8KDI43B8
KDI04B8KD35B8
AB219426B5AB219429B5
AB219427B5AB219428B5
AP011087B5AP011086B5
PAN09B9PAN011B9
Alo36B9PAN01B9PAN037B9
AP011096B8AB493834
AP011093B8AP011095B8
AP011094B8AY033073B4
AY033072B4AB100695B4
AB073835B4AB112066C1AB112348C1
AB112471C1AB074756C1
EIHB006C1AP011097C1SLK-126
AB033553C2AB113879C2
AB202071C2AP011098C2
X01587C2PhLC14C5
PhLC03C5AB241112C5
AP011101C5AP011099C5AP011100C5
PhCH24C5M154AP011105C7AP011104C7
AP011107C7AP011106C7
AP011108CJ022
STN013AB493837AB493847AB493838AB493840
AB493839AP011102C6AB105172
AB493844AB493843
AB493842AB493841
AP011103C6M007X75656C3
X75665C3AB048705C4
AB048704C4AF280817D1
AY161157D1AY161150D1
M73D1AF151735D1
AB078032D2AB078033D2
AB090268D2AB090269D2AY090452D3
AB493848.AB493845
AB493846DQ315776D3
DQ315777D3AB048702D4
AB048703D4AB048701D4AB033559D4
AB032431EAB205129E
AB106564EX75657E
AB064311GAB064312GEF634480G
AB056513GAB086397F1
AF223963F1AF223962F2AF223965F2
AB166850F2AB214516F2
AY090455F1AB179747H
AB205010HAB266536HAB059660H
100
100
100
100
100100
100
100
100
99
100
100100
100
100
100
100
100
100
100
100
100
100
100
100
100
92
100
92
100
92
100
100
68
100
100
92
92
96
92
100
100100
92
100
92
100
100
97
100
100
100
100
100
100
0.02
100
HF
HF2
F1GG
ED
D1
D2
D3D4
E
C3C4
C6
C7
C5
C2
C1
B4
B8
B5
B9
B7
B3
B2
B1
B6
A1A2
C
B
AInuit populations in Arctic
Japanese, east Asia
Chinese, Asia mainland
Indonesian populations, Southeast Asia
Indonesian populations, Southeast Asia
Indonesian populations, Southeast AsiaPhilippino and Indonesian populations, Southeast Asia
Indonesian populations, Southeast Asia
Vietnamese, Southeast Asia East Asia, China mainland and Southeast Asia
East Asia, China mainland and Southeast Asia
Philippino, East Asia
Indonesian populations, Southeast Asia
Indonesian populations, Southeast Asia
Polynesia, New Caledonia, Pacific region Aborigine populations, Southern Australia
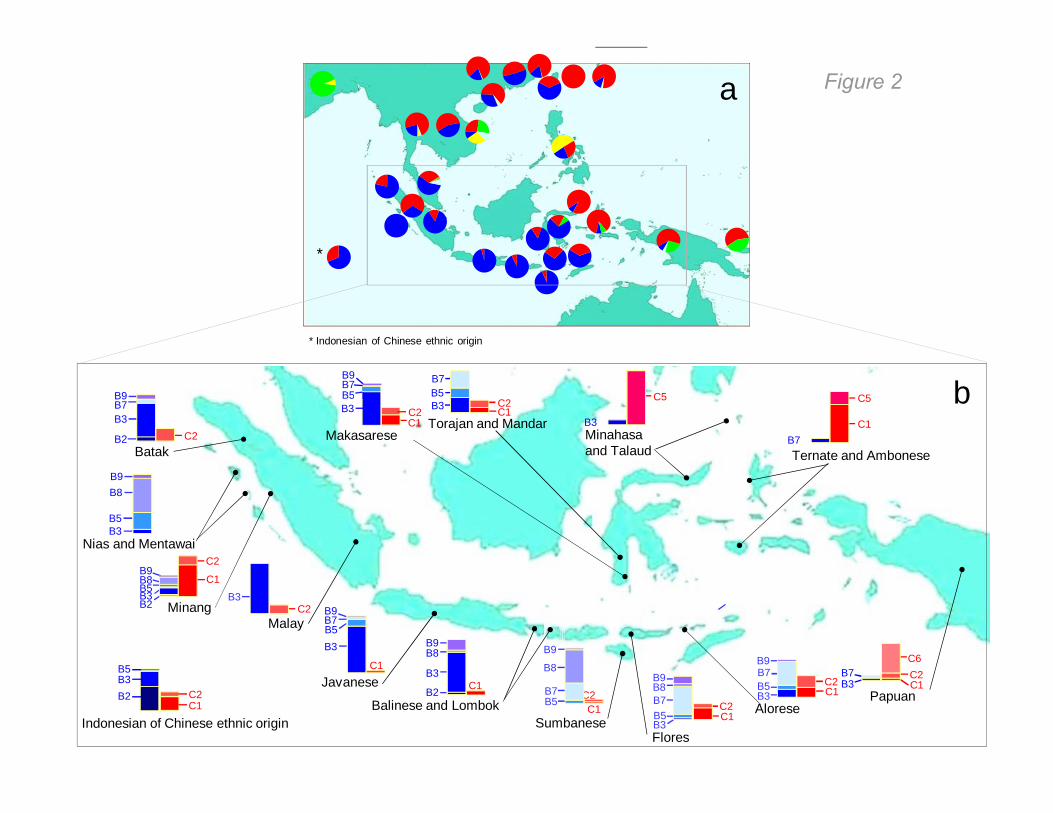
Figure 2
Balinese and LombokSumbanese
Flores
Alorese
Minahasa and Talaud Ternate and Ambonese
C1
C5
BatakB2
B3B7B9
C2
Malay C2
B3B9
C1B2
B3
B8B9
C1C2
B3
B7B5
B8B9
B3B5B7B9
C2C1 Papuan
C1C2C6
B3B7
Indonesian of Chinese ethnic origin
B2B3B5
C1C2
B3B5
B8B9
Nias and Mentawai
Minang
B3
C5
B7
bMakasarese
C1C2
B2B3B5B8B9
C1
C2
Javanese
B5B3
B7
C1
* Indonesian of Chinese ethnic origin
*
a
B7B5
B8
B9
C1C2
Torajan and Mandar B3
B7
C1C2
B5B3B5B7B9

Javanese(Suriname)
Javanese(Holland)
Javanese (Java)
HBV/B3 (adw)
HBV genotype is maintained in Javanese ethnic population separated for centuries

100
AB540583
AF223957C1
AP011097
AF223960AB112471
AB074756
AF068756
AB112348AB112066
AY247031
AF533983
X01587
D23681
AB033553
AB113879
AP011098
AB202071
PhCH24AB241112
PhLC03
PhLC14 AP011100
AP011101
AP011099AP011106
AP011107
AP011104
AP011105
EU670263AP011108
GQ358157
AB493837
GQ358155
GQ358156
AB493840 AB493838AB493842
AB493844
AP011103
AB493841
AB493839
AP011102
X75656
X75665
AB048705
AB048704
AB266536
6
3
4
97
8
5
21
10
H
EAST ASIA AND SEA
Papua-Pacific
North Australia
TWO TYPES HBV/C SUBGENOTYPES: THE ASIAN AND PAPUA-PACIFIC(study on HBV isolates from the Asia-Pacific Region)

• Subgenotypes of HBV/B in Japan (Sugauchi et al):– Bj
– B2 (Ba: prevalent in China)
Maintained Viral Characteristics
In Chinese ethnic population*
Clinical Implications?
DNA Sequence:
*Have been living in Indonesia for > 3
generations
HBV genotype is maintained in Javaneseethnic population separated for centuries
• Subgenotypes of HBV/B in Indonesia:– B3 (adw)– B5 (adw)– B7 (ayw)– B2 (Chinese
Indonesian)

‘a’ determinant (124-147)
309 regular blood donors (2005) from Medan (North Sumatra) and Solo (Central Java) (HBsAg, anti-HCV and anti-HIV neg)• anti-HBc (+) : 134 (43.4%)• HBV DNA (+): 25 (8.09%)• Variants were detected in 7 samples: T123A (1), M133L (1),
T143M (7)
Thedja MD et al. 2010. Occult hepatitis B in blood donors in Indonesia: alter antigenicity of the hepatitis B virus surface protein. Hepatology Int.4, 608
CLINICAL AND PUBLIC HEALTH SIGNIFICANCE:Detection failure of HepB virus in blood donors


• Malaria - caused by Plasmodium parasites- is responsible for high mortality mortality. An estimated 300-500 million cases each year result in more than 1 million deaths
• Available preventive methods are not sufficient• Humans have a number of genetic adaptations
that act to combat Plasmodium
• Understanding the mechanism of malaria interactions with the erythrocytes membrane will provide opportunities for new methods of disease prevention and treatment

Rank Country Mortality Rate
49 INDONESIA 3.1

MALARIA AS A SELECTIVE AGENT IN HUMANS
WHO 2005
SAO, HbE,Gerbich negativity,
α-thalassemia (many), G6PD deficiency (many)
Duffy negativity,G6PD deficiency (A-),
HbS, HbC

COMMON THEMES AMONG MALARIA-RESISTANCE ALLELES EXAMINED to DATE
(G6PD A-, HBC, HBS, DUFFY NEGATIVITY, HBE)
1) Recent origins (<<30,000 years)2) Strong selection coefficients3) Compatible with “Malarial Eve” Hypothesis

• An uncommon variant of hereditary elliptocytosis• Caused by heterozygosity for a 27 bp del in the gene
encoding the erythrocyte membrane protein band 3 – the solute carrier family 4 (SLC4A1) protein on chromosome 17
• SAO erythrocytes are rigid because mutation enhances the tightness of association between band 3, ankyrin and the spectrin lattice
• The condition confers highly specific protection against cerebral malaria
SOUTHEAST ASIA OVALOCYTOSIS

§ Erythrocyte membrane contain variety of proteins –(i) peripheral proteins of bands 1, 2, 4.1, 4.2, 5 and 6, and (ii) integral proteins of bands 3, 7 and PAS 1 - 4.
§ Mutation in SLC4A1 gene enhances the tightness of association between band 3, ankyrin and spectrin lattice

SAO PROTECTS FROM MALARIA-RELATED MORTALITY.
• Strong correlation between SAO and malaria prevalence (Mgone et al. 1996).
• Case-control studies suggest strong (complete?) protection from cerebral malaria (Genton et al. 1995, Allen et al. 1999).

SAO is A BALANCED POLYMORPHISM§ Heterozygote has greatly enhanced fitness
in malarial environments.Heterozygotes have no negative clinical manifestations.
§ SAO homozygote is inviableWhen SAO is common, it can demonstrably increase the miscarriage rate (Liu et al. 1994)
§ SAO as a health burdenOne in for of the pregnancies in these couples will be lost due to SAO inviability.With random mating 12.25% (=0.35x0.35) of couples will both be heterozygous carriers of SAO.

SAO HAS A RESTRICTED GEOGRAPHIC DISTRIBUTION.
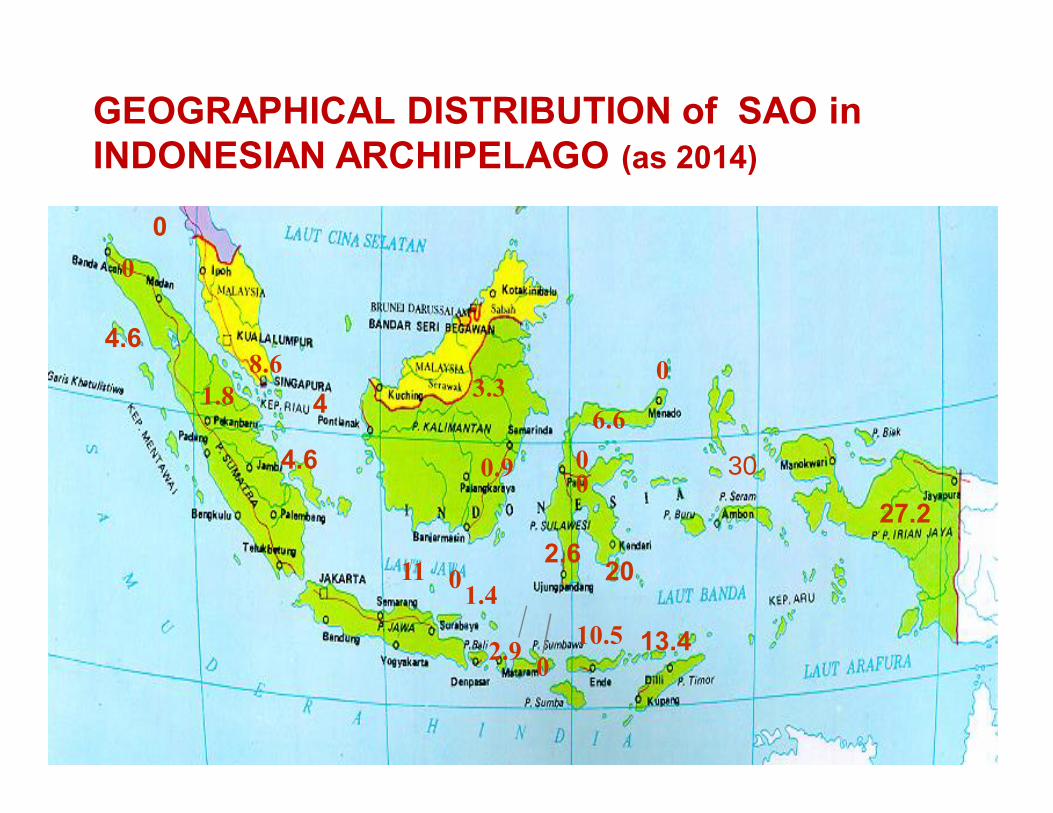
GEOGRAPHICAL DISTRIBUTION of SAO in INDONESIAN ARCHIPELAGO (as 2014)
0
8.6
0
11 01.4
2.9 010.5 13.4
27.20
2.6
0
06.6
1.8
4.6
4 3.3
0.9
20
30
4.6

Eijkman Institute
1.4
8.6
1.8
3.3
4.6
0.9
6.6
10.5
2.9
27
13.4
0
0
0
0
0
0
0
0
0 10 20 30
0 . 1
A f r i c a
A l o r
I r i a n
B a t a k
J a v a
M a k a s s a r
S a s a k
B u g i s
M i n a h a s a
S u m b a
S u m b a w a
T o r a j a
K a i l i
B a n j a r
P a l e m b a n g
T e n g g e r
D a y a k
M i n a n g
M a l a y ( P ’ b a r u )
B a l i
FREQUENCY OF SAO (%)
FREQUENCY OF SAO VARIES SIGNIFICANTLY IN DIFFERENT POPULATIONS (not revised)

DISTRIBUTION OF PAPUAN GENETIC CONTRIBUTION AND SAO PREVALENCE
Papuan Genetic Contribution (%)0 5 10 15 20 25 30
SAO
Pre
vala
nce
(%)
0
10
20
30
40
50
§ SAO is not found in non-endemic area for malaria; but not all endemic area shown SAO
•SAO is found mainly in two clades: a. Malay-related (lowprevalence) and b. Papuan-related (high prevalence)

SAO likely evolved after expansion of P. falciparum into Southeast Asia.
P. falciparum emerges as health threat ~10,000 years before present.
Models of Coevolution between Plasmodium and Humans May Need to Accommodate the Existence of Ancient Malaria-resistance Alleles

FUTURE STUDY§ Bringing human and medical genetics and
computational biology together – to answer the big question on how culture shaped the human genome§ The answer is in your hands
Genetics
Linguistic
Anthropology Archeology
Medicine
Mathematic

Acknowledgment
Steve Lansing, Complex Institute, NTUMurray Cox and Elsa Guillot, Massey University, New ZealandTatyana Karafet, University of ArizonaThe HUGO Pan-Asian SNP Consortium
EIJKMAN INSTITUTE
David Mulyono,Meta Dewi TedjaDin Syafruddin
Helen SuryadiSafarina MalikGludhug PurnomoWindy JoanmawatiAlida HarahapDewi Megawati

Thank you and greetings from Indonesia