huang, s., eronen, j., janis, c., saarinen, j., silvestro ... · 115 collections (average across...
TRANSCRIPT

Huang, S., Eronen, J., Janis, C., Saarinen, J., Silvestro, D., & Fritz, S. (2017).Mammal body size evolution in North America and Europe over 20 Myr:similar trends generated by different processes. Proceedings of the RoyalSociety B: Biological Sciences, 284(1849), [20162361].https://doi.org/10.1098/rspb.2016.2361
Peer reviewed version
Link to published version (if available):10.1098/rspb.2016.2361
Link to publication record in Explore Bristol ResearchPDF-document
This is the author accepted manuscript (AAM). The final published version (version of record) is available onlinevia the Royal Society at http://rspb.royalsocietypublishing.org/content/284/1849/20162361]. Please refer to anyapplicable terms of use of the publisher.
University of Bristol - Explore Bristol ResearchGeneral rights
This document is made available in accordance with publisher policies. Please cite only the publishedversion using the reference above. Full terms of use are available:http://www.bristol.ac.uk/pure/about/ebr-terms

Mammal body size evolution in North America and Europe over 20 million years: 1
Similar trends generated by different processes 2
3
Shan Huang1,*, Jussi T. Eronen2, 3, Christine M. Janis4, Juha J. Saarinen2, 5, Daniele 4
Silvestro6, Susanne A. Fritz1,7 5
6
1Senckenberg Biodiversity & Climate Research Centre (BiK-F), Frankfurt am Main, 7
Germany; 8
2Department of Geosciences and Geography, University of Helsinki, Helsinki, Finland; 9
3BIOS Research Unit, Helsinki, Finland; 10
4Department of Ecology and Evolutionary Biology, Brown University, Providence, USA; 11
5Natural History Museum, London, UK; 12
6Department of Biological and Environmental Sciences, University of Gothenburg, 13
Gothenburg, Sweden; 14
7Institute of Ecology, Evolution and Diversity, Goethe University, Frankfurt am Main, 15
Germany. 16
17
*e-mail address: [email protected] 18
19
20

ABSTRACT 21
Because body size interacts with many fundamental biological properties of a species, body 22
size evolution can be an essential component of the generation and maintenance of 23
biodiversity. Here we investigate how body size evolution can be linked to the clade-specific 24
diversification dynamics in different geographic regions. We analyze an extensive body size 25
dataset of Neogene large herbivores (covering ~50% of the 970 species in the orders 26
Artiodactyla and Perissodactyla) in Europe and North America in a Bayesian framework. We 27
reconstruct the temporal patterns of body size in each order on each continent independently, 28
and find significant increases of minimum size in three of the continental assemblages (except 29
European perissodactyls), suggesting an active selection for larger bodies. Assessment of 30
trait-correlated birth-death models indicates that the common trend of body size increase is 31
generated by different processes in different clades and regions. Larger-bodied artiodactyl 32
species on both continents tend to have higher origination rates, and both clades in North 33
America show strong links between large bodies and low extinction rate. Collectively, our 34
results suggest a strong role of species selection and perhaps of higher-taxon sorting in 35
driving body size evolution, and highlight the value of investigating evolutionary processes in 36
a biogeographic context. 37
38
Keywords: species body mass, diversification, biogeography, species selection, Cope’s rule, 39
higher-taxon sorting. 40
41
42

Body size has been considered one of the most important traits of an organism, a species or 43
higher taxon, because it co-varies with a wide range of morphological, physiological, and 44
ecological traits [1-3, 4 and references therein]. Therefore, the study of broad-scale spatial and 45
temporal patterns of body size, central to ecology and evolutionary biology, can shed light 46
onto underlying processes shaping biodiversity patterns [5, 6], for which the rich body of 47
literature on body size biology provides a solid basis. Here, we present a cross-continent 48
investigation of the temporal dynamics of species body size in relation to the means of 49
generation and maintenance of biodiversity over 20 million years (myr). 50
Large-scale assessments comparing a range of taxa often found a variety of 51
trajectories of body size evolution, including directional changes, stasis, and range expansion 52
to contain both larger and smaller sizes through time [4, 7-11]. Particularly in the geologic 53
past, many taxa evolved to have larger bodies, a pattern referred to broadly as Cope's rule, 54
often coinciding with a cooling climate [12-15]. The current and projected climate change and 55
human impact are thus anticipated to cause higher stress for large-bodied species [16-18] and 56
lead to general dwarfism of the present-day fauna [19]. However, a spatially-explicit 57
framework has been suggested essential for sensible prediction [20, 21], and the variation in 58
patterns calls for mechanistic explanations and a deeper understanding of the 59
macroevolutionary processes behind body size change through time. 60
Several different macroevolutionary processes can generate an increasing trend of 61
body size through time. If body size is generally irrelevant to diversification processes of 62
lineages, the temporal pattern of body size should mimic a diffusive process, perhaps 63
asymmetrically bounded by a limit on the minimum size for a viable animal body (a passive 64
evolutionary process) [4, 22-24]. This passive process gives rise to an increasing maximum, 65
and if given enough time, an increasing median body size without raising the minimum. In 66
contrast, more active evolutionary processes like species selection (general correlation of 67
body size with species origination and/or extinction rates), higher-taxon sorting (turnover of 68

higher taxa with different body sizes), and a within-lineage tendency of smaller ancestors 69
giving rise to larger descendants (Cope’s rule in a strict sense) should affect the minimum 70
body size of a clade and diversification processes at relevant scales [4, 22-25]. Assessments of 71
the relationship between body size and diversification processes cannot rely on molecular 72
phylogenies alone, because separating changes in speciation and extinction is difficult without 73
including the extinct lineages [26, 27]. The fossil record serves as a direct window to the long 74
history of biodiversity, and can thus provide an excellent opportunity for investigating 75
evolutionary processes [e.g. 12, 13, 28-30]. Specifically in Cenozoic mammals, Raia et al. 76
attributed apparent size increases in several clades to frequent expansions into new niches 77
favoring large bodies, with the finding of generally later appearances of larger-bodied clades 78
in the fossil record [12]. Within closely related species pairs, however, large-bodied species 79
appeared earlier in the fossil record than their small-bodied relatives in the same geographic 80
region, rejecting the hypothesis of within-lineage evolution towards larger sizes but also 81
raising the question of potential sampling bias against small bodies in older fossils at a fine 82
scale. The context-related heterogeneity shown in their findings invites further investigation 83
of clade-specific dynamics in a biogeographic context. 84
In this study, we investigate whether clade-specific body size variation can be 85
associated with differential origination and/or extinction rates, and can thus be directly linked 86
to the clade-specific diversification dynamics in different geographic regions. We focus on 87
the two extant orders of ungulates, Artiodactyla and Perissodactyla, which have an excellent 88
fossil record of their high diversity throughout the Neogene (around 23–2 million years ago; 89
Ma), allowing a cross-clade, cross-region comparison of evolutionary processes. To derive 90
robust results from the fossil record, we analyze our paleontological data in a Bayesian 91
framework that accounts for the heterogeneous preservation potential. We analyze the two 92
orders separately in two major continental faunas, Europe and North America (thus four 93
groups of continental assemblages in total). We expect an apparent increase of body size in all 94

four of our focal groups with a backdrop of climate cooling during the Neogene [12, 13, 31, 95
32], and we further distinguish whether this trend is generated by a bounded diffusive process 96
or by an active evolutionary process due to selection for large-bodied species. To explain the 97
active process suggested in some groups, we further investigate whether large bodies were 98
selected through elevated origination rate and/or reduced extinction rate (i.e. species 99
selection), whether family/subfamily replacements could have contributed to substantial body 100
size changes (i.e. higher-taxon sorting), and/or whether there is a within-genus tendency of 101
larger species appearing later than smaller species (i.e. strict Cope’s rule). 102
103
Materials and methods 104
We combined data of 6432 fossil occurrences for 970 ungulate species (Orders: Artiodactyla 105
and Perissodactyla) from two sources for Europe (the NOW database [33]) and North 106
America [34, 35] respectively (see figure S1 for sample locations and [36] for a standardized 107
and taxonomically unified dataset), throughout the last ~21 myr, excluding the last ~2 myr 108
(after Gelasian). The geologic ages (reported in Ma) of all fossil data are based on the 109
continental time units for each continent: the Mammal Neogene (MN) units for European 110
occurrences (as followed by the NOW database for their European data [37]), and the North 111
American Land Mammal Ages (NALMAs) for the North American occurrences [38]. We 112
complemented the body mass data available from the initial data sources (NOW and [34, 35]) 113
with additional estimates from the published literature and measurements from museum 114
collections (average across specimen when multiple were measured). Specimen-specific 115
dating was usually not possible for considering temporal intra-specific variation. Our final 116
dataset contains species-level average body mass for 483 of the included species (~50%) (data 117
used in our analyses available at: [Senckenberg data archive address available once the 118
manuscript is accepted]). Despite the common belief that fossil data tend to be more complete 119
close to the Recent, we did not find such bias in our data (see figure S2 and statistics in the 120

figure caption). In addition, an increase of body size towards Recent is the opposite 121
expectation of a biased fossil record – small-bodied animals are more likely to be 122
underrepresented in older, poorly sampled fossil collections but more abundant in the younger 123
record [39, 40]. To assess potential sampling bias against small-bodied species within the two 124
focal orders, we also included preservation rate in our models and tested for a correlation with 125
body size (see below). 126
We analyzed the fossil data in a Bayesian framework to simultaneously estimate the 127
origination and extinction times of each species, the preservation rate (the average expected 128
number of fossil records per species per myr), and parameters of a birth-death model 129
describing the diversification dynamics of an entire clade including correlation of body mass 130
with origination and extinction rates (expected number of origination/extinction events per 131
lineage per myr) using the Python program PyRate [41, 42]. For each order in each 132
continental fauna, we generated 99 replicate datasets to sample random occurrences within 133
the geological time bins [following the procedure in 41, 43], and with each of these 99×4 134
datasets, we ran 10 million iterations (excluding the additional 50,000 burn-in iterations), 135
using Birth-Death Markov Chain Monte Carlo to sample the parameters from their posterior 136
distribution for a covar birth-deadth model of the correlation of body size (!) with the 137
preservation rate (#), origination rate ($) and extinction rate (%; see more details of the 138
methods in ESM). Because we are interested in the first-order processes generating the 139
temporal trend of body size in a biogeographic context, we do not distinguish in-situ 140
origination and extinction from range dynamics (i.e. invasion from outside the focal region 141
and regional extinction due to emigration). However, the long duration and coarse temporal 142
resolution examined here probably decreases impacts of short-term range dynamics. 143
To compare body size patterns with higher taxa replacement, we reconstructed 144
temporal patterns of relative diversities in families (in Artiodactyla) and subfamilies (in 145
Perissodactyla) using the origination and extinction times estimated from the PyRate analyses. 146

To test for within-genus tendency of body size increase from older to younger species, we fit 147
linear mixed-effect models to the relationship between species body size and their order of 148
origination (fixed effect), with genus assignment as a random effect. We compared three 149
models for each group (Artiodactyla and Perissodactyla in Europe and North America) based 150
on AIC: one model with genus effect on the intercepts, one on the slopes, and one on both. 151
Without further evidence from a phylogenetic analysis, a positive correlation between body 152
size and order of origination within a genus can be considered partial support for the tendency 153
of smaller ancestors giving rise to larger descendants. 154
The PyRate analyses were conducted in Python 2.7 (available at 155
http://www.python.org). All additional analyses and data visualizations were conducted in R 156
3.2.2 [44], with the package CODA [45] for summarizing MCMC iteration logs and the 157
packages lme4 [46] and lmerTest [47] for testing and interpreting linear models with mixed 158
effects. 159
160
Results 161
We summarized the body size distribution of species in each geologic time bin (using the 162
continent-specific geologic time scales, see Methods) based on the estimated species 163
origination and extinction times (summarized in: [Senckenberg data archive address available 164
once the manuscript is accepted]) for each order in Europe and North America respectively 165
(thus four groups in total), as well as in a combination of the two regions (figure 1 and S3; 166
table 1). In North America, both Artiodactyla and Perissodactyla increased significantly in 167
minimum body size through time, although only artiodactyls showed a significant increase of 168
median size. However, comparisons of artiodactyl family and perissodactyl subfamily 169
diversities show no consistent replacement of smaller-bodied clades by larger ones (figure 2). 170
In the Neogene of Europe, only artiodactyls increased significantly in minimum, 171
median and maximum body size (table 1), which coincided with apparent declines of several 172

small-bodied families (e.g. Tragulidae, Moschidae, and Cainotheriidae), diversity increase of 173
the larger-bodied Bovidae, and arrivals of even larger hippos and camels (figure 2). European 174
perissodactyls only expanded their range of body size to include a larger maximum over time 175
(table 1). Compared with the other groups, European perissodactyl species already had 176
relatively large bodies owing to the large diversity of rhinos and chalicotheres (figure 2) at the 177
beginning of the Neogene (figure 1 and S3), and as a group, remained large throughout 178
Neogene despite the rises and falls seen in different subclades (figure 2); the lack of 179
significant trend in body size is also apparent at the family level (figure S4). When we 180
combined the two regional faunas, our results suggest significant increases in both minimum 181
and maximum body size, masking the variation between regional patterns (table 1). 182
Although similar trends of increasing body size are shared between the continental 183
faunas, we found different relationships between body size and diversification dynamics 184
among the groups we examined. In the North American fauna, larger body sizes were 185
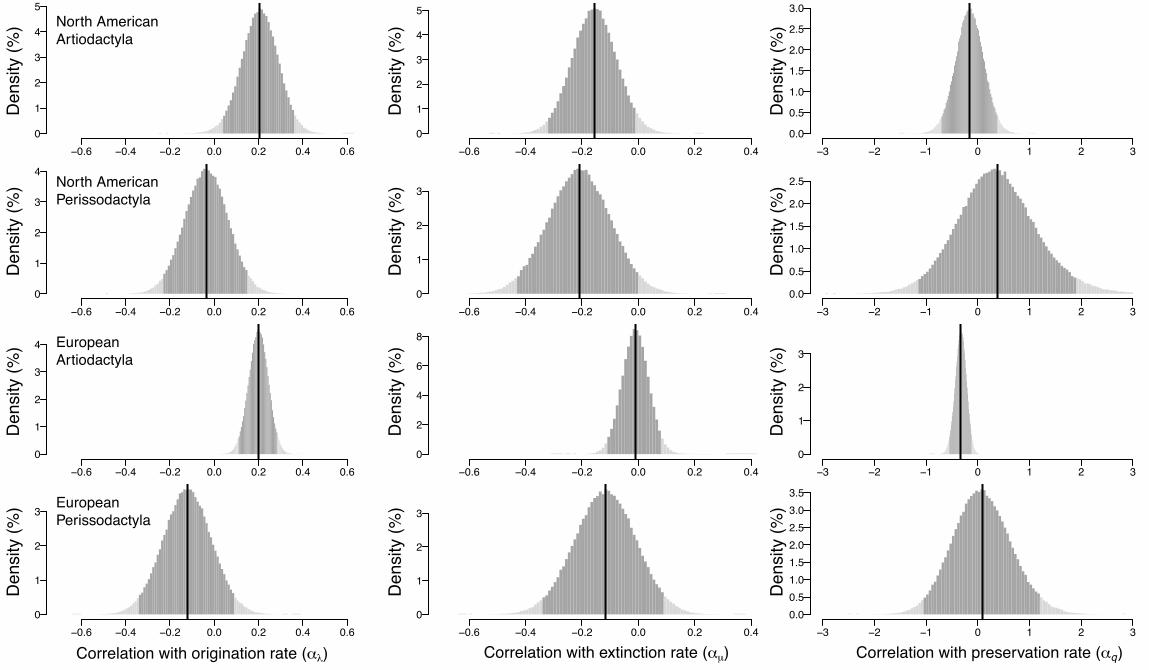
associated with significantly lower extinction rates in both Artiodactyla (mean !% = -0.16) 186
and Perissodactyla (mean !% = -0.21, although the credible interval’s upper bound falls 187
slightly above 0 at 0.005), and with significantly higher origination rates in Artiodactyla 188
(mean !$ = 0.21; see the posterior distributions in figure 3 and effects illustrated in figure 4). 189
Neither of the groups showed a correlation between body size and preservation rate. Among 190
European artiodactyl species, larger bodies are also strongly associated with higher 191
origination rates (mean !$ = 0.20), as well as lower preservation rates (mean !# = -0.33), an 192
unexpected pattern opposite from the commonly assumed sampling bias against small-bodied 193
animals (see Discussion). 194
For all four groups we examined for within-genus relationship between species body 195
size and origination order, the model including a random effect of genus on the intercept was 196
always the best model, although the differences in AIC were marginal (table S1). Based on 197

the best models, we found no correlation between species body size and order of origination 198
within a genus in any of the groups (p ³ 0.20; table S1). 199
200
Discussion 201
We find significant increases in minimum, median, and/or maximum body size in all four 202
focal groups (artiodactyls and perissodactyls in North America and Europe), but this common 203
trend seems to have been generated by different processes in different groups and regions. 204
Three groups (except European perissodactyls) show significant increases in minimum size 205
through time, so their overall body size trends are generated by selection for large bodies, 206
rather than a bounded diffusive process. Our finding of significant correlations of body size 207
with speciation rate (positively) in Artiodactyla on both continents, but with extinction rate 208
(negatively) in both orders in the North American fauna, further supports the idea of a driven 209
evolution towards larger body sizes through species selection, and highlights phylogenetic as 210
well as regional differences in evolutionary processes. The turnover of higher taxa, especially 211
the temporal variation in relative diversities of families in European Artiodactyla, also plays a 212
role in driving the body size patterns. The fourth group, the European perissodactyls, 213
maintained their large sizes throughout the Neogene, except for a temporary drop of their 214
median size with the rise of the Equinae. Notably, the maximum body size in this group 215
increased through time (with the addition of several large rhino species), implying a diffusion-216
like process bounded by selection against smaller sizes. We found no evidence of consistent 217
within-genus evolutionary trends towards larger bodies, thus no support for the strict-sense 218
Cope’s rule to be the dominant process of body size evolution. 219
Further sampling effort for fossil data is unlikely to alter the patterns, as our analyses 220
did not find larger bodies associated with higher preservation rates in any of our groups. In 221
only one group, the European artiodactyls, we found larger bodies to be associated with lower 222
preservation rates, a trend opposite from the expectation of large bodies to be overrepresented 223

in the fossil record [39, 40, 48, 49]. Larger bodies also coincide with lower population density 224
in the extant artiodactyl species that are classified as Least Concern in terms of extinction risk 225
by the IUCN Red List (Spearman’s r = -0.24, p =0.043, n = 93 species, data from [20]; see 226
also [50]), which might have reduced the chance of entering the fossil record [48, 51]. 227
Capturing this trend is probably a signal of extensive sampling for the European fauna. The 228
trend of body size increase towards the present in this group can only change if additional 229
large species had been under-sampled so far and were added into the beginning of our focal 230
time period, i.e. Early-Mid Miocene (by ~12 myr ago). Such addition is highly unlikely given 231
the already higher coverage of body size data for the European species in our dataset (57% 232
artiodactyls and 58% perissodactyls, compared with 38% and 42% respectively in North 233
America). Nevertheless, adding large species to the Early-Mid Miocene cannot change the 234
pattern of increasing minimum body size in European Artiodactyla, and thus cannot reject the 235
hypothesis that larger bodies were preferentially generated in this group. 236
The link between larger bodies and higher origination rates in mammals has been 237
suggested by previous studies examining multiple clades collectively based on fossil body 238
size [12] or based on inferred body size from extant relatives [52], but we only find this link 239
in Artiodactyla. In addition, we also find that both of our focal clades tend to have lower 240
extinction rates for larger-sized species in North America, contradicting previous results [12, 241
52]. In other words, while baseline extinction rates tend to increase towards the Recent (Fig. 242
S5), the elevation of extinction rate affected small-sized species more strongly than large 243
ones. Previous studies hypothesized that adopting new life styles, and thus entering new niche 244
space, allowed some large-bodied lineages to rapidly diversify, implying a role of higher-245
taxon sorting [12, 15, 53, 54]. Our results indicate that this process might be specific to 246
artiodactyls, which show consistently higher origination rates of large-bodied species on both 247
continents. Comparisons of artiodactyl family diversities also show expanding of some large-248
bodied families (e.g. Hippopotamidae and Camelidae) and the thriving of Bovidae in Europe, 249

but little obvious change in North America except for the expansion of Camelidae in the 250
Pliocene (figure 2). However, the niche-opening hypothesis cannot explain the removal of 251
small-bodied species (indicated by the increase in minimum body size and declines of small-252
bodied families such as Tragulidae, Moschidae, and Cainotheriidae in Europe), especially in 253
North America where the lower extinction rate in large-bodied artiodactyls and perissodactyls 254
is more likely to be related to survivorship in face of reportedly less productive habitats since 255
Late Miocene [55, 56] within the continent’s biogeographic setup (see below). Large bodies 256
are evidently associated with (a) capability of tracking suitable conditions due to longer 257
dispersal distance [2] and larger geographic ranges [57, 58], and (b) relaxation of 258
physiological constraints due to larger gut size (especially the hindgut in the case of 259
perissodactyls) for digesting larger quantities of low-nutrient plant materials [59] and larger 260
volume-to-surface ratio for preserving energy [2] and thus improving fasting endurance [60, 261
61], all of which can contribute to resisting extinction. 262
Variation exists not only between the two clades, but also between the two focal 263
regions we examine, suggesting different mechanisms generated the apparently similar trend 264
of body size increase shared by regions. Compared with the European fauna, the North 265
American fauna shows stronger signals of extinction rate in driving body size evolution of 266
both clades we examine. One important biogeographic distinction between the two continents 267
is that, before the initial rise of the Isthmus of Panama at ~6 Ma [62], North America lacked a 268
southern pathway for frequent migration out of the region and later return. The tapering shape 269
of the North American continent also contributes to restricting dispersal towards lower 270
latitudes while the climate transitioned to a colder state. With changes in physical conditions 271
as well as biotic interactions, North American species might be more prone to selective 272
extinction than European species, where more dynamic ranges are possible with access to 273
surrounding regions. Spatial dynamics can thus be another key process shaping the 274
evolutionary history of body size and macroevolutionary patterns in general [30, 55, 63, 64]; 275

comparisons of its role with those of in-situ origination and extinction in further investigation 276
will be valuable. Further, metabolic theory predicts that large bodies are favored in colder 277
climates in various environmental scenarios [13, 14, 19, 54, 65], so that an increase of body 278
size in both temperate regions is not surprising along a generally cooling trajectory of the 279
global climate during the Neogene [32, 66, 67]. However, there is substantial heterogeneity of 280
the climatic conditions across space at all times [56, 68-70], and the resulting 281
appearance/disappearance of habitats (e.g. mid-latitude savannas) can affect faunal 282
composition greatly. It is also possible that body size was not the only trait that was selected 283
for but was entangled in a composite morphological and functional evolution, especially traits 284
tightly related to interaction with the ever-changing environments, and we propose a future 285
direction of assessing how traits interact to play different roles in driving diversification 286
dynamics in different environments. 287
As discussed above, higher-taxon sorting might also have contributed to shaping the 288
body size trends, but the patterns shown here are more complex than clear. In particular, 289
North American perissodactyls show increases in minimum and (marginally) maximum body 290
size but not in the median, suggesting a change in the shape of their size distribution through 291
time rather than a systematic tendency towards larger bodies (a pattern more likely for 292
artiodactyls, shown in figure 1 and S3). The replacement (in terms of relative diversities) of, 293
for example, the generally smaller-bodied subfamily Anchitheriinae by the larger Equinae, 294
might contribute to shaping the patterns in this group, but Anchitheriinae as a clade also 295
appears to have increased in body size through time to an extent that Late Miocene 296
anchitheres were larger than the contemporaneous equines in North America (figure S4). 297
Further, the increase in body size of the smaller-bodied subfamilies (Equinae, Anchitheriinae, 298
and the family Tapiridae) might have contributed to the general increase in minimum body 299
size of North American Perissodactyla throughout the record. There are also other taxa 300
turnover patterns that complicate the picture, such as the early disappearance of the large-301

bodied subfamily Schizotheriinae from the region by Mid-Miocene and the persistence of the 302
generally small-bodied Equinae (figure 2 & S4). The variety of temporal trajectories for 303
family body size shown in figure S4 suggest that within an order, the families might have 304
undergone different evolutionary processes, but species selection might still be an important 305
mechanism in family body size evolution given the general negative correlation between body 306
size and extinction rates, and the lack of evidence for a strict Cope’s rule. Further 307
distinguishing (and comparing) the roles of cross-clade species selection and higher-taxon 308
sorting in shaping the larger-scale body size evolutionary pattern is difficult with current data 309
(the coverage is less satisfactory when analyzed at the family or subfamily level), but a 310
fruitful next step will be rigorous analyses using fossil phylogenies to investigate the 311
phylogenetic distributions of origination and extinction events in relation to phenotypic 312
evolution in similar biogeographical contexts. 313
314
Conclusion 315
A general trend of increasing body size through time is frequently found and discussed in 316
mammalian studies, although not without exceptions [4, 12, 13]. Results from this study 317
suggest species selection and potentially higher-taxon sorting are the most likely driving 318
mechanism for active evolution of larger body size. Passive evolution, a diffusive process 319
bounded by the minimum requirement, is also supported in one group, the European 320
Perissodactyla. The variety of patterns and processes we have identified in the Neogene 321
ungulates from Europe and North America, and others found in different study systems [e.g. 7, 322
10, 14], points to the importance of scale for studying macroevolutionary dynamics [71]. 323
More importantly, similar macroevolutionary trends shared among clades in different regions 324
might actually be generated by different processes – even one single trait (i.e. body size, 325
though admittedly a composite one) can associate with the origination and extinction rates 326
differently across clades and regions, perhaps even differently at different levels of the 327

taxonomic hierarchy [63, 71]. As evolution takes place within the biogeographic setup, 328
apparent patterns are products of the interaction between the focal clade and the regional 329
environment, including the regional topography, climatic conditions, and other biotic 330
components such as vegetation and infectious disease in the system [55, 72, 73]. Therefore, 331
both the clade’s biology and regional environment must be considered to fully understand the 332
evolution of body size, and in general, macroevolutionary patterns [74-76]. 333
334
Data accessibility 335
The data with the average body mass, estimated origination time and estimated extinction 336
time for each species are available at [Senckenberg data archive address available once the 337
manuscript is accepted]. 338
339
Authors’ contributions 340
S.H., J.T.E. and S.A.F. designed the study; S.H., J.J.S. and C.M.J. collected data; S.H., D.S. 341
and S.A.F. performed analyses; S.H. wrote the first draft of the manuscript; all authors 342
contributed to interpretation of data and revisions. 343
344
Competing interests 345
We declare we have no competing interests. 346
347
Funding 348
S.H. is supported by Alexander von Humboldt Foundation through a postdoc fellowship. 349
J.T.E. was supported by Kone Foundation during this work. J.S. was funded by Oskar 350
Huttunen Foundation, Helsinki, Finland (year 2015) and by Jenny and Antti Wihuri Fund, 351
Helsinki, Finland (year 2016). D.S. received funding from the Swedish Research Council 352

(2015-04748). S.F. is funded by the DFG (German Research Foundation grant FR 3246/2-1). 353
This article is a contribution to the integrative Climate Change Biology (iCCB) programme. 354
355
Acknowledgement 356
We thank C. Weiland at the Senckenberg Data Center for technical assistance with the cluster 357
server, the Böhning-Gaese working group at Senckenberg BiK-F for discussion at various 358
stages of the project, and collection accesses (and assistance by personnel in parentheses) at 359
Natural History Museum of Vienna, Austria (U. Göhlich); Natural History Museum, London, 360
UK (A. Currant and P. Brewer); Torquay Museum, Torquay, UK (B. Chandler and C. Jones); 361
British Geological Survey, Nottingham, UK (L. Neep and P. Shepherd); Sedgwich Museum 362
(M. Riley) and University Museum of Zoology (M. Lowe), Cambridge, UK; Staatliches 363
Museum für Naturkunde, Stuttgart, Germany (R. Ziegler); Staatliches Museum für 364
Naturkunde, Karlsruhe, Germany (D. Schreiber); Research Station of Quaternary 365
Palaeontology of the Senckenberg Institute, Weimar, Germany (R.-D. Kahlke); National 366
Museums of Kenya, Nairobi, Kenya (M. Muungu), and Field Natural History Museum (W. F. 367
Simpson). We also thank F. Smith and two anonymous reviewers for constructive comments. 368
369
References: 370
1. Calder W.A. 1984 Size, function, and life history. Cambridge, MA, Harvard 371
University Press. 372
2. Peters R.H. 1983 The ecological implications of body size. Cambridge, UK, 373
Cambridge University Press. 374
3. Blanckenhorn W.U. 2000 The evolution of body size: What keeps organisms small? Q 375
Rev Biol 75(4), 385-407. 376
4. Smith F.A., Payne J.L., Heim N.A., Balk M.A., Finnegan S., Kowalewski M., Lyons 377
S.K., McClain C.R., McShea D.W., Novack-Gottshall P.M., et al. 2016 Body size evolution 378

across the Geozoic. Annu Rev Earth Plannet Sci 44(1), 523-553. (doi:10.1146/annurev-earth-379
060115-012147). 380
5. Huang S., Stephens P.R., Gittleman J.L. 2012 Traits, trees and taxa: global dimensions 381
of biodiversity in mammals. Proc R Soc B 279, 4997-5003. (doi:10.1098/rspb.2012.1981). 382
6. Slater G.J. 2013 Phylogenetic evidence for a shift in the mode of mammalian body 383
size evolution at the Cretaceous-Palaeogene boundary. Methods in Ecology and Evolution 384
4(8), 734-744. (doi:10.1111/2041-210X.12084). 385
7. Jablonski D. 1997 Body-size evolution in Cretaceous molluscs and the status of 386
Cope's rule. Nature 385(6613), 250-252. 387
8. Clapham M.E., Karr J.A. 2012 Environmental and biotic controls on the evolutionary 388
history of insect body size. Proc Natl Acad Sci USA 109(27), 10927-10930. 389
(doi:10.1073/pnas.1204026109). 390
9. Hunt G. 2007 The relative importance of directional change, random walks, and stasis 391
in the evolution of fossil lineages. Proc Natl Acad Sci USA 104(47), 18404-18408. 392
(doi:10.1073/pnas.0704088104). 393
10. Novack-Gottshall P.M., Lanier M.A. 2008 Scale-dependence of Cope's rule in body 394
size evolution of Paleozoic brachiopods. Proc Natl Acad Sci USA 105(14), 5430-5434. 395
(doi:10.1073/pnas.0709645105). 396
11. Gardner J.L., Peters A., Kearney M.R., Joseph L., Heinsohn R. 2011 Declining body 397
size: a third universal response to warming? Trends Ecol Evol 26(6), 285-291. 398
(doi:10.1016/j.tree.2011.03.005). 399
12. Raia P., Carotenuto F., Passaro F., Fulgione D., Fortelius M. 2012 Ecological 400
specialization in fossil mammals explains Cope’s rule. Am Nat 179(3), 328-337. 401
(doi:10.1086/664081). 402
13. Alroy J. 1998 Cope's rule and the dynamics of body mass evolution in North 403
American fossil mammals. Science 280(5364), 731-734. (doi:10.1126/science.280.5364.731). 404

14. Hunt G., Roy K. 2006 Climate change, body size evolution, and Cope's Rule in deep-405
sea ostracodes. Proc Natl Acad Sci USA 103(5), 1347-1352. (doi:10.1073/pnas.0510550103). 406
15. Payne J.L., Boyer A.G., Brown J.H., Finnegan S., Kowalewski M., Krause R.A., 407
Lyons S.K., McClain C.R., McShea D.W., Novack-Gottshall P.M., et al. 2009 Two-phase 408
increase in the maximum size of life over 3.5 billion years reflects biological innovation and 409
environmental opportunity. Proc Natl Acad Sci USA 106(1), 24-27. 410
(doi:10.1073/pnas.0806314106). 411
16. McCain C.M., King S.R.B. 2014 Body size and activity times mediate mammalian 412
responses to climate change. Global Change Biol 20(6), 1760-1769. (doi:10.1111/gcb.12499). 413
17. Olden J.D., Hogan Z.S., Zanden M.J.V. 2007 Small fish, big fish, red fish, blue fish: 414
size-biased extinction risk of the world's freshwater and marine fishes. Global Ecol Biogeogr 415
16(6), 694-701. (doi:10.1111/j.1466-8238.2007.00337.x). 416
18. Dirzo R., Young H.S., Galetti M., Ceballos G., Isaac N.J.B., Collen B. 2014 417
Defaunation in the Anthropocene. Science 345(6195), 401-406. 418
(doi:10.1126/science.1251817). 419
19. Sheridan J.A., Bickford D. 2011 Shrinking body size as an ecological response to 420
climate change. Nature Climate Change 1(8), 401-406. (doi:10.1038/nclimate1259). 421
20. Fritz S.A., Bininda-Emonds O.R.P., Purvis A. 2009 Geographical variation in 422
predictors of mammalian extinction risk: big is bad, but only in the tropics. Ecol Lett 12(6), 423
538-549. 424
21. Turvey S.T., Fritz S.A. 2011 The ghosts of mammals past: biological and 425
geographical patterns of global mammalian extinction across the Holocene. Philos Trans R 426
Soc B 366(1577), 2564-2576. (doi:10.1098/rstb.2011.0020). 427
22. McShea D.W. 1994 Mechanisms of large-scale evolutionary trends. Evolution 48(6), 428
1747-1763. (doi:10.2307/2410505). 429

23. McShea D.W. 2000 Trends, tools, and terminology. Paleobiology 26(3), 330-333. 430
(doi:10.1666/0094-8373(2000)026<0330:TTAT>2.0.CO;2). 431
24. Gould S.J. 1988 Trends as changes in variance: A new slant on progress and 432
directionality in evolution. J Paleontol 62(3), 319-329. 433
25. Jablonski D. 2008 Species selection: Theory and data. Annual Review of Ecology, 434
Evolution, and Systematics 39(1), 501-524. 435
(doi:doi:10.1146/annurev.ecolsys.39.110707.173510). 436
26. Rabosky D.L. 2010 Extinction rates should not be estimated from molecular 437
phylogenies. Evolution 64(6), 1816-1824. (doi:10.1111/j.1558-5646.2009.00926.x). 438
27. Quental T.B., Marshall C.R. 2010 Diversity dynamics: molecular phylogenies need 439
the fossil record. Trends Ecol Evol 25(8), 434-441. (doi:10.1016/j.tree.2010.05.002). 440
28. Smith J.T., Roy K. 2006 Selectivity during background extinction: Plio-Pleistocene 441
scallops in California. Paleobiology 32(3), 408-416. (doi:10.1666/05078.1). 442
29. Finarelli J. 2007 Mechanisms behind active trends in body size evolution of the 443
Canidae (Carnivora: Mammalia). Am Nat 170(6), 876-885. (doi:10.1086/522846). 444
30. Fritz S.A., Schnitzler J., Eronen J.T., Hof C., Böhning-Gaese K., Graham C.H. 2013 445
Diversity in time and space: wanted dead and alive. Trends Ecol Evol 28(9), 509-516. 446
(doi:10.1016/j.tree.2013.05.004). 447
31. Hansen J., Sato M., Russell G., Kharecha P. 2013 Climate sensitivity, sea level and 448
atmospheric carbon dioxide. Philos Trans R Soc A 371(2001). (doi:10.1098/rsta.2012.0294). 449
32. Zachos J., Pagani M., Sloan L., Thomas E., Billups K. 2001 Trends, rhythms, and 450
aberrations in global climate 65 Ma to present. Science 292(5517), 686-693. 451
(doi:10.1126/science.1059412). 452
33. Fortelius M. (coordinator) 2015 New and Old Worlds Database of Fossil Mammals 453
(NOW). University of Helsinki. Date of access: November 1st, 2015. 454

34. Janis C.M., Gunnell G.F., Uhen M.D. 2008 Evolution of Tertiary Mammals of North 455
America: Volume 2, Small Mammals, Xenarthrans, and Marine Mammals. (Cambridge, UK, 456
Cambridge University Press. 457
35. Janis C.M., Scott K.M., Jacos L.L. 1998 Evolution of Tertiary Mammals of North 458
America: Volume 1, Terrestrial Carnivores, Ungulates, and Ungulate Like Mammals. 459
(Cambridge, UK, Cambridge University Press. 460
36. Fritz S.A., Eronen J.T., Schnitzler J., Hof C., Janis C.M., Mulch A., Böhning-Gaese 461
K., Graham C.H. 2016 Twenty-million-year relationship between mammalian diversity and 462
primary productivity. Proc Natl Acad Sci USA. (doi:10.1073/pnas.1602145113). 463
37. Steininger F.F. 1999 Chronostratigraphy, geochronology and biochronology of the 464
Miocene “European Land Mammal Mega-Zones (ELMMZ)” and the Miocene “Mammal-465
Zones (MN-Zones)”. In The Miocene Land Mammals of Europe (eds. Rössner G.E., Heissig 466
K.), pp. 9-24. München, Germany, Friedrich Pfeil. 467
38. Woodburne M.O. (eds) 2004 Late Cretaceous and Cenozoic Mammals of North 468
America. New York, USA, Columbia University Press. 469
39. Plotnick R.E., Smith F.A., Lyons S.K. 2016 The fossil record of the sixth extinction. 470
Ecol Lett 19(5), 546-553. (doi:10.1111/ele.12589). 471
40. Behrensmeyer A.K., Chapman R.E. 1993 Models and simulations of taphonomic 472
time-averaging in terrestrial vertebrate assemblage. In Taphonomic Approaches to Time 473
Resolution in Fossil Assemblages: Short Courses in Paleontology No 6 (eds. Kidwell S., 474
Behrensmeyer A.K.), pp. 125-149, Paleontological Societ Knoxville, Tennessee. 475
41. Silvestro D., Salamin N., Schnitzler J. 2014 PyRate: a new program to estimate 476
speciation and extinction rates from incomplete fossil data. Methods in Ecology and Evolution 477
5(10), 1126-1131. (doi:10.1111/2041-210X.12263). 478

42. Silvestro D., Schnitzler J., Liow L.H., Antonelli A., Salamin N. 2014 Bayesian 479
estimation of speciation and extinction from incomplete fossil occurrence data. Syst Biol. 480
(doi:10.1093/sysbio/syu006). 481
43. Silvestro D., Antonelli A., Salamin N., Quental T.B. 2015 The role of clade 482
competition in the diversification of North American canids. Proc Natl Acad Sci USA 483
112(28), 8684-8689. (doi:10.1073/pnas.1502803112). 484
44. R Development Core Team 2015 R: A language and environment for statistical 485
computing (3.2.2) 486
45. Plummer M., Best N., Cowles K., Vines K. 2006 CODA: Convergence diagnosis and 487
output analysis for MCMC. R News 6, 7-11. 488
46. Bates D., Mächler M., Bolker B., Walker S. 2015 Fitting linear mixed-effects models 489
using lme4. Journal of Statistical Software 67(1), 48. (doi:10.18637/jss.v067.i01). 490
47. Kuznetsova A., Brockhoff P., Christensen H. 2016 lmerTest: tests for random and 491
fixed effects for linear mixed effect models (lmer objects of lme4 package), v. 2.0–25 (R 492
package version 2.0-32) 493
48. Turvey S.T., Blackburn T.M. 2011 Determinants of species abundance in the 494
Quaternary vertebrate fossil record. Paleobiology 37(3), 537-546. (doi:10.1666/09075.1). 495
49. Miller J.H., Behrensmeyer A.K., Du A., Lyons S.K., Patterson D., Tóth A., Villaseñor 496
A., Kanga E., Reed D. 2014 Ecological fidelity of functional traits based on species presence-497
absence in a modern mammalian bone assemblage (Amboseli, Kenya). Paleobiology 40(4), 498
560-583. (doi:10.1666/13062). 499
50. Damuth J. 1981 Population density and body size in mammals. Nature 290(5808), 500
699-700. 501
51. Western D., Behrensmeyer A.K. 2009 Bone assemblages track animal community 502
structure over 40 years in an African savanna ecosystem. Science 324(5930), 1061-1064. 503
(doi:10.1126/science.1171155). 504

52. Liow L.H., Fortelius M., Bingham E., Lintulaakso K., Mannila H., Flynn L., Stenseth 505
N.C. 2008 Higher origination and extinction rates in larger mammals. Proc Natl Acad Sci 506
USA 105(16), 6097-6102. (doi:10.1073/pnas.0709763105). 507
53. Sibly R.M., Brown J.H. 2007 Effects of body size and lifestyle on evolution of 508
mammal life histories. Proc Natl Acad Sci USA 104(45), 17707-17712. 509
(doi:10.1073/pnas.0707725104). 510
54. Smith F.A., Boyer A.G., Brown J.H., Costa D.P., Dayan T., Ernest S.K.M., Evans 511
A.R., Fortelius M., Gittleman J.L., Hamilton M.J., et al. 2010 The evolution of maximum 512
body size of terrestrial mammals. Science 330(6008), 1216-1219. 513
(doi:10.1126/science.1194830). 514
55. Janis C.M. 1993 Tertiary mammal evolution in the context of changing climates, 515
vegetation, and tectonic events. Annu Rev Ecol Syst 24, 467-500. 516
56. Pound M.J., Haywood A.M., Salzmann U., Riding J.B. 2012 Global vegetation 517
dynamics and latitudinal temperature gradients during the Mid to Late Miocene (15.97–5.33 518
Ma). Earth-Sci Rev 112(1–2), 1-22. (doi:10.1016/j.earscirev.2012.02.005). 519
57. Pyron M. 1999 Relationships between geographical range size, body size, local 520
abundance, and habitat breadth in North American suckers and sunfishes. J Biogeogr 26(3), 521
549-558. (doi:10.1046/j.1365-2699.1999.00303.x). 522
58. Stanley S.M. 1986 Population size, extinction, and speciation: The fission effect in 523
Neogene Bivalvia. Paleobiology 12(1), 89-110. 524
59. Clauss M., Frey R., Kiefer B., Lechner-Doll M., Loehlein W., Polster C., Rössner 525
G.E., Streich W.J. 2003 The maximum attainable body size of herbivorous mammals: 526
morphophysiological constraints on foregut, and adaptations of hindgut fermenters. 527
Oecologia 136(1), 14-27. (doi:10.1007/s00442-003-1254-z). 528
60. Millar J.S., Hickling G.J. 1990 Fasting Endurance and the Evolution of Mammalian 529
Body Size. Funct Ecol 4(1), 5-12. (doi:10.2307/2389646). 530

61. Lindstedt S.L., Boyce M.S. 1985 Seasonality, fasting endurance, and body size in 531
mammals. Am Nat 125(6), 873-878. 532
62. Bacon C.D., Silvestro D., Jaramillo C., Smith B.T., Chakrabarty P., Antonelli A. 2015 533
Biological evidence supports an early and complex emergence of the Isthmus of Panama. 534
Proc Natl Acad Sci USA 112(19), 6110-6115. (doi:10.1073/pnas.1423853112). 535
63. Allen C.R., Garmestani A.S., Havlicek T.D., Marquet P.A., Peterson G.D., Restrepo 536
C., Stow C.A., Weeks B.E. 2006 Patterns in body mass distributions: sifting among 537
alternative hypotheses. Ecol Lett 9(5), 630-643. (doi:10.1111/j.1461-0248.2006.00902.x). 538
64. Jablonski D. 2008 Extinction and the spatial dynamics of biodiversity. Proc Natl Acad 539
Sci USA 105(Supplement 1), 11528-11535. (doi:10.1073/pnas.0801919105). 540
65. Secord R., Bloch J.I., Chester S.G.B., Boyer D.M., Wood A.R., Wing S.L., Kraus 541
M.J., McInerney F.A., Krigbaum J. 2012 Evolution of the earliest horses driven by climate 542
change in the Paleocene-Eocene Thermal Maximum. Science 335(6071), 959-962. 543
(doi:10.1126/science.1213859). 544
66. Zhang Y.G., Pagani M., Liu Z. 2014 A 12-million-year temperature history of the 545
tropical Pacific Ocean. Science 344(6179), 84-87. (doi:10.1126/science.1246172). 546
67. Zhang Y.G., Pagani M., Liu Z., Bohaty S.M., DeConto R. 2013 A 40-million-year 547
history of atmospheric CO2. Philos Trans R Soc A 371(2001). (doi:10.1098/rsta.2013.0096). 548
68. Böhme M., Winklhofer M., Ilg A. 2011 Miocene precipitation in Europe: Temporal 549
trends and spatial gradients. Palaeogeogr, Palaeoclimatol, Palaeoecol 304(3–4), 212-218. 550
(doi:10.1016/j.palaeo.2010.09.028). 551
69. Herold N., Huber M., Müller R.D., Seton M. 2012 Modeling the Miocene climatic 552
optimum: Ocean circulation. Paleoceanography 27(1), PA1209. 553
(doi:10.1029/2010PA002041). 554

70. Dowsett H.J., Robinson M.M., Stoll D.K., Foley K.M., Johnson A.L.A., Williams M., 555
Riesselman C.R. 2013 The PRISM (Pliocene palaeoclimate) reconstruction: time for a 556
paradigm shift. Philos Trans R Soc A 371(2001). (doi:10.1098/rsta.2012.0524). 557
71. Jablonski D. 2007 Scale and hierarchy in macroevolution. Palaeontology 50(1), 87-558
109. (doi:10.1111/j.1475-4983.2006.00615.x). 559
72. Hoffmann A.A., Hercus M.J. 2000 Environmental stress as an evolutionary force. 560
Bioscience 50(3), 217-226. (doi:10.1641/0006-3568(2000)050[0217:esaaef]2.3.co;2). 561
73. Ricklefs R.E. 2015 Intrinsic dynamics of the regional community. Ecol Lett 18(6), 562
497-503. (doi:10.1111/ele.12431). 563
74. Condamine F.L., Rolland J., Morlon H. 2013 Macroevolutionary perspectives to 564
environmental change. Ecol Lett 16(s1), 72-85. (doi:10.1111/ele.12062). 565
75. Smith F.A., Lyons S.K. 2011 How big should a mammal be? A macroecological look 566
at mammalian body size over space and time. Philos Trans R Soc B 366(1576), 2364-2378. 567
(doi:10.1098/rstb.2011.0067). 568
76. Cooper N., Purvis A. 2010 Body size evolution in mammals: complexity in tempo and 569
mode. Am Nat 175(6), 727-738. (doi:10.1086/652466). 570
77. Gradstein F.M., Ogg G., Schmitz M. 2012 The Geologic Time Scale 2012. Oxford, 571
UK, Elsevier. 572
573
574

Table and figure legends
Table 1 Temporal trends of body size of Artiodactyla and Perissodactyla in Europe and North
America. The relationship between body size and time to Recent is tested using Spearman’s
rank-order correlation test (n = 14 time bins for all cases); a negative correlation indicates an
increase of body size towards Recent. The combined data set is based on the European
geologic time scale, but results are qualitatively consistent when using the North American
time scale. The number of species with body size data (N) is given for each order on each
continent. Results are qualitatively consistent with Bonferroni correction of the p-values
except for the maximum size in European Artiodactyla (adjusted-p = 0.09).
Body size variables
Artiodactyla Perissodactyla All
combined Europe
(N = 223)
North America (N = 101)
Combined Europe (N = 86)
North America (N = 72)
Combined
Minimum r = -0.973 p < 0.001
r = -0.982 p < 0.001
r = -0.935 p < 0.001
r = -0.103 p = 0.727
r = -0.837 p < 0.001
r = -0.837 p < 0.001
r = -0.935 p < 0.001
Median r = -0.998 p < 0.001
r = -0.844 p < 0.001
r = -0.987 p < 0.001
r = -0.330 p = 0.250
r = -0.198 p = 0.497
r = -0.068 p = 0.816
r = -0.952 p < 0.001
Maximum r = -0.550 p = 0.042
r = -0.987 p < 0.001
r = -0.987 p < 0.001
r = -0.827 p < 0.001
r = -0.485 p = 0.079
r = -0.955 p < 0.001
r = -0.965 p < 0.001
Figure 1 Temporal trends of body size of Artiodactyla and Perissodactyla in Neogene Europe
(left) and North America (right), based on their species’ origination and extinction times
estimated in PyRate analyses (excluding species without body size data). Lines indicate
median size in each time bin and color shades represent the 25%-75% quantiles for between-
clade comparisons. See statistical results of the trends in Table 1 and full range of body size
through time in Figure S3. Geologic epochs (Early-, Mid-, and Late-Miocene, and Pliocene)
following the time scale by Gradstein et al [77] are indicated for reference.

Figure 2 Temporal patterns of relative diversity (proportion of species) in artiodactyl families
(upper) and perissodactyl subfamilies (lower) in relation to the median body size of their
species, ordered (for both the polygons and the legends) as increasing in size from the bottom
(i.e. larger-bodied families are towards the top). Family (and subfamily) duration are based on
their species’ origination and extinction times estimated in PyRate analyses. Color similarity
loosely illustrates the commonly assumed phylogenetic relationship among the families and
subfamilies (see caption for Figure S4 for taxonomic details), with the same color scheme
applied in Europe [left] and North America [right]. Families and subfamilies without any
species body size data are indicated with an asterisk (*), and the white space represents
species without sufficient information for family assignment (position not indicative of their
body size). See Figure S4 for the temporal trend of family or subfamily median body size.
Figure 3 Posterior distributions of the correlation parameter estimates (Covar model)
between body size and three rates: origination rate (!$, left), extinction rate (!%, middle) and
preservation rate (!#, right) in Artiodactyla and Perissodactyla in Europe and North America
respectively. Solid lines indicate the mean estimate and color-filled bars represent the 95%
credible intervals.
Figure 4 The differential origination (white) and extinction rates (grey) in relation to
variation in body size in Artiodactyla and Perissodactyla in Europe and North America
respectively, illustrated using the highest rates during the Neogene (based on the mean
temporal trajectory of all posterior rates from the PyRate analysis, see figure S5) as the
baseline rates ($0 and %0). The baseline rates were then corrected for lineages sized at the
10% and 90% quantiles of all body sizes (b10 and b90, in Kg) in each group, based on the
posterior distribution of the correlative parameters (shown in figure 3) from the Covar model.
Therefore, the bars representing the corrected rates show that artiodactyls of different sizes

are expected to have different origination rates ($10 and $90) in on both continents (the upper
row) and different extinction rates (%10 and %90) in both clades in North America (the left
column). Origination rate of larger-sized artiodactyls could be twice as high as small-sized
ones in Europe.

20 15 10 5Geologic Time (Ma)
PlioLate−MioMid−MioEarly−MioNorth America
Artiodactyla Perissodactyla
20 15 10 5Geologic Time (Ma)
PlioLate−MioMid−MioEarly−Mio
Europe4
6
8
10
log 2
Bod
y m
ass
(Kg)
4
6
8
10

15 10 50.0
0.2
0.4
0.6
0.8
1.0
Geologic Time (Ma)
Prop
ortio
nal d
ivers
ityN. American Artiodactyla
Entelodontidae*Hypertragulidae*Gelocidae*LeptomerycidaeMoschidaeAntilocapridaeCervidaeBovidaeMerycoidodontidaeTayassuidaeDromomerycidaeProtoceratidaeCamelidaeAnthracotheriidae
Increasing median body size
15 10 50.0
0.2
0.4
0.6
0.8
1.0
Geologic Time (Ma)
Prop
ortio
nal d
ivers
ity
European Artiodactyla
Hoplitomerycidae*Haplobunodontidae*CainotheriidaeAndegamerycidaeMoschidaePalaeochoeridaeSanitheriidaeTragulidaeCervidaeBovidaeSuidaeCamelidaePalaeomerycidaeAnthracotheriidaeGiraffidaeHippopotamidae
Increasing median body size
15 10 50.0
0.2
0.4
0.6
0.8
1.0
Geologic Time (Ma)
Prop
ortio
nal d
ivers
ity
N. American Perissodactyla
AnchitheriinaeTapiridaeEquinaeMenoceratinaeDiceratheriinaeAceratheriinaeSchizotheriinaeTeleoceratinae
Increasing median body size
15 10 50.0
0.2
0.4
0.6
0.8
1.0
Geologic Time (Ma)
Prop
ortio
nal d
ivers
ity
European Perissodactyla
AnchitheriinaeTapiridaeEquinaeMenoceratinaeAceratheriinaeChalicotheriinaeSchizotheriinaeTeleoceratinaeRhinocerotinaeElasmotheriinae
Increasing median body size
Den
sity
(%)
−0.6 −0.4 −0.2 0.0 0.2 0.4 0.60
1
2
3
4
5North AmericanArtiodactyla
Den
sity
(%)
−0.6 −0.4 −0.2 0.0 0.2 0.40
1
2
3
4
5
Den
sity
(%)
−3 −2 −1 0 1 2 30.0
0.5
1.0
1.5
2.0
2.5
3.0D
ensi
ty (%
)
−0.6 −0.4 −0.2 0.0 0.2 0.4 0.60
1
2
3
4North AmericanPerissodactyla
Den
sity
(%)
−0.6 −0.4 −0.2 0.0 0.2 0.40
1
2
3
Den
sity
(%)
−3 −2 −1 0 1 2 30.0
0.5
1.0
1.5
2.0
2.5
Den
sity
(%)
−0.6 −0.4 −0.2 0.0 0.2 0.4 0.60
1
2
3
4 EuropeanArtiodactyla
Den
sity
(%)
−0.6 −0.4 −0.2 0.0 0.2 0.40
2
4
6
8
Den
sity
(%)
−3 −2 −1 0 1 2 30
1
2
3
Den
sity
(%)
−0.6 −0.4 −0.2 0.0 0.2 0.4 0.60
1
2
3EuropeanPerissodactyla
Den
sity
(%)
−0.6 −0.4 −0.2 0.0 0.2 0.40
1
2
3
Den
sity
(%)
−3 −2 −1 0 1 2 30.00.51.01.52.02.53.03.5
Correlation with origination rate (αλ) Correlation with extinction rate (αµ) Correlation with preservation rate (αq)

●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
λ10 λ90 µ10 µ900
1
2
3
Size
cor
rect
ed ra
tes
North American Artiodactylaλ0 = 0.48µ0 = 0.83b10 = 16b90 = 441µ = mm
Origination (λ)Extinction (µ)
λ10 λ90 µ10 µ90
0.5
1.0
1.5
2.0
Size
cor
rect
ed ra
tes
European Artiodactylaλ0 = 0.35µ0 = 0.58b10 = 14b90 = 584µ = mm
●●●
●
●●
●●
●
●
●
●
●
●●●●●
●
●
●●●
●
●●●●
●●
●
●
●
●●
●
●
●
●
●
●
●●●
●●
●●●
●
●
●
●
●
●●●●
●
●●●
●●●
●●●●
●
●●●
●
●
●
●●●
●
●
●
●
●
●●
●
●●●●
●●●●
●●●●●
●●●
●
●
●●●●●
●
●
●
●●
●
●●●●●●●●●
●
●
●
●●●●
●
●●●
●●
●
●
●
●●●
●
●
●
●
●
●●●
●●
●●
●●●
●
●●●●
●●
●●
●
●●●
●
●
●●●
●
●●●
●
●
●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●
●●●●
●
●
●●●●●●
●
●
●
●●
●
●
●
●
●●●●●●
●
●●●●●●
●
●
●●
●
●●
●●
●●●
●
●
●
●
●●●●●
●
●●
●
●●●●●
●●●●
●
●●●
●●
●
●
●●
●●●
●●
●
●●●●
●
●
●●●
●
●
●●●
●●
●
●●
●●
●●
●●
●
●●
●●●
●●●●
●
●
●●
●
●●●
●
●●
●●
●●
●●●
●●●●
●
●
●●
●
●
●●●●●●
●
●
●
●●
●●
●
●
●
●●
●●●
●
●
●
●●
●●●
●
●
●
●●●
●
●●
●
●●
●
●●●●●●
●
●●●●●●●
●●●
●●●●●
●●●
●●
●●●●●●●
●●
●
●
●●
●●●●●
●
●●
●●
●
●●●
●
●●
●●●●
●
●●
●●●●●
●
●●
●
●
●
●●●
●
●
●●●●●
●●
●
●●
●●
●●●●●●
●●●●
●●
●●
●
●●●●
●
●
●
●
●
●
●
●●●
●
●
●
●
●●
●
●
●
●
●●
●
●
●●
●●●●
●●●
●●
●
●
●●●
●●●
●●●
●●
●
●
●
●
●●
●
●●●●
●
●●●●
●
●
●
●●
●
●●●●●
●
●●●
●
●●●●
●
●●●●●●
●●●●
●●
●
●
●
●●
●●●
●
●
●●
●●
●●
●
●
●
●
●
●●
●
●●
●
●●●
●
●●
●●
●●●●●
●●●
●
●
●
●
●
●
●
●●
●
●
●
●●●●●●
●●●
●●●
●
●●●
●
●
●
●
●
●
●
●
●●●
●
●●●
●
●
●●
●
●
●●
●
●●
●
●
●●
●
●
●
●●●
●
●
●●●
●
●●●
●●
●●
●●
●
●●
●
●●●
●●●
●
●●
●●●
●●
●
●
●●
●●●●
●
●
●●
●
●
●●●
●
●●●
●
●
●
●●●
●
●●
●●
●●●
●
●●
●
●
●●●●
●
●
●
●
●
●●●
●●●
●
●
●
●
●
●●●●
●●●●
●
●●
●●
●
●●●●
●
●●
●
●
●●
●●●
●
●●●
●
●●
●●●●●
●●
●●●
●
●
●
●●●●
●
●
●
●●●
●●●
●
●
●●●●
●●
●
●●●
●
●●●●●
●
●●●
●●●●
●●
●
●●
●●
●●
●
●
●●
●
●
●●●
●●●
●
●
●●●●●
●
●
●●
●
●
●
●
●●●●●●●●●●●
●●●●
●
●●●
●
●●
●
●●●●●
●●●
●●
●
●●
●●●
●
●●
●
●
●
●●●●●●●●
●
●
●
●●●●
●●●
●
●
●
●
●●●●●●
●
●●
●●
●●●
●
●●
●●●
●●
●●
●
●
●●
●●
●
●●●●
●
●
●●●●●
●
●
●
●
●
●
●●●
●
●
●●
●
●●
●●●
●
●
●●●●
●
●
●●●
●
●●●●●●●●●●●●
●
●●●
●
●●●
●
●
●
●
●●●
●
●
●
●●●
●●
●
●●●●
●
●●
●●●●
●
●●
●
●●
●●●●
●
●
●
●
●
●●
●
●●
●●●●
●●●●
●
●●●
●
●●●
●●
●
●
●●●
●●
●
●
●●●
●●
●●
●
●
●
●●●
●
●●●
●
●●
●
●
●●●
●●●●●●●●
●●
●●●
●
●●
●●
●●
●●
●
●●●
●
●●●●
●●●
●●
●
●
●
●
●
●
●
●●
●●
●●●
●
●●
●●
●●●
●●●●
●
●
●
●●●●●●●●
●●●
●
●
●
●●
●
●●●
●
●●
●
●●●●
●●
●●●
●
●
●●●
●
●●
●
●
●
●
●
●●●
●●
●●●
●
●
●●
●●●●●●●
●
●
●
●●
●●●
●●
●
●●●
●
●
●
●●
●
●●
●●●
●●
●●●●●●
●
●
●
●
●
●●●
●●
●
●●
●●●●●
●
●●
●●
●
●
●
●●●
●
●
●
●
●
●
●●●
●
●●
●●●●
●
●●●●●
●
●●
●●●
●●●●
●●
●●
●
●●
●●●
●●
●●●
●
●
●●●●
●●●
●
●●
●●
●●
●
●●●
●
●●
●●
●
●
●●
●
●
●
●●●
●
●
●
●●●
●
●●●●●●
●●●
●
●
●●●
●●
●●●
●●●
●●●
●
●●
●
●
●
●
●
●●●●
●●
●
●
●●
●
●●
●
●●●
●●●
●
●
●
●●●
●
●●
●●
●
●●
●
●●●
●
●
●
●●
●
●
●●
●●
●●●
●●
●
●●●
●●
●●
●
●●
●
●●
●
●
●
●●
●
●●●
●
●●
●●
●●●●
●●
●
●●●
●
●
●●●
●●
●●
●●●
●●
●●●
●●
●●●
●●●●
●
●●●
●●●
●●
●
●●
●
●●●●
●●
●
●●
●
●●●
●
●●
●●
●●
●
●●●
●
●
●●●●
●
●
●●
●●
●●
●●●
●●
●
●●
●●●●●
●
●
●
●●
●
●●●●
●
●
●
●
●
●●●
●●
●●
●
●
●●●
●●
●
●●●
●●
●
●●
●●
●
●●
●●
●●
●●
●●●●●●
●
●
●
●●
●●
●●●●●●
●●●
●●●
●●●●
●
●●
●●●●
●
●●
●●●●●●●●
●●
●●●●
●●●
●●
●
●
●●●
●
●
●●●●
●●
●
●
●
●
●
●●
●●
●●
●
●
●
●●
●●●
●
●
●
●
●●
●
●●
●●
●●
●
●●●
●●●●●●
●
●●●●●
●●
●
●●
●●
●●
●●●
●
●●
●
●
●
●
●●●
●●
●
●●
●●●
●
●
●
●●●
●●
●●●●●●●
●
●
●●●●
●●
●●
●
●●●
●
●
●●●●
●●●●
●●●●
●●
●
●●
●
●
●
●
●●
●
●
●●
●●●
●
●●
●
●
●
●
●
●
●●
●●
●
●
●●
●
●
●●●
●●●
●
●●●●●●
●●
●
●
●
●●
●
●
●
●●
●
●
●●
●
●●●●●●●
●●●
●
●●●●●
●●●
●
●●●●●●●●
●
●
●●
●
●●
●
●
●●●
●●●●●●
●●●●
●●●●●
●●
●●
●●●
●
●
●
●●●●●●●●
●
●●
●●●
●
●●
●
●●●
●●●●●●
●●
●●●
●●●
●
●●●●●●●
●●●
●
●●
●
●●●●
●●
●●
●
●●
●
●
●
●●
●●●
●●
●●●●●●
●
●
●
●●●●
●
●
●●
●
●●●●
●
●
●●●
●
●●
●
●
●
●●●
●●
●●
●
●
●●●●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●●
●
●
●
●
●●●
●●
●
●●●
●●●●●●●●●●●●
●●●
●
●
●●●
●
●
●
●
●
●●●●●●
●●
●
●
●●●●
●
●●●●
●
●
●●●●●
●●
●
●●●
●
●
●
●●●
●
●
●
●
●
●●●
●
●●●●●●●●●●
●
●
●
●●●
●●
●
●●
●●●●●
●
●●
●
●●●
●
●●
●●●
●
●●●●
●●●
●
●
●●●
●●●
●
●●
●
●
●●
●
●●
●
●
●●●
●
●●●
●●
●
●●●
●●●●
●●●
●●●
●
●●●●
●●
●●●
●
●
●
●
●
●●●●
●●
●
●●●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●●
●●
●●
●
●
●●●●●●
●●
●
●
●●●
●
●
●
●
●●●●●●
●●
●●●
●
●
●
●●
●
●●
●
●●
●●●
●
●●●
●●
●
●●●●●●●
●
●
●
●
●●●
●●●●●
●●
●
●●
●
●
●
●
●●●
●●
●
●●
●
●●
●●●●●●
●●
●●
●●●
●
●
●●
●
●●
●●●●
●
●
●
●
●
●
●●
●
●
●●
●
●●
●●●●●
●●
●
●●
●
●●●●●●●●●●●●●
●●
●●
●●●
●
●
●
●●●
●
●●●
●
●●
●●●
●●
●
●●
●
●●●
●
●●
●●●●
●●●
●
●
●
●
●
●
●
●●
●●
●●●
●
●
●●●●
●●●●
●
●
●
●
●
●●
●●
●●●
●
●
●
●
●●●●
●
●●
●
●●
●●
●
●
●●
●
●
●
●
●●●
●
●●
●●
●●
●●●●●
●●●
●●●
●
●●●●
●●●●●
●●●
●
●
●●
●●●●●
●●
●
●●
●●
●
●
●
●●
●●
●
●●
●●●●●
●
●
●●
●
●●●●●●
●
●
●●
●
●
●
●●●
●
●
●
●●
●
●
●●●
●
●●
●
●●
●●
●
●
●
●
●●
●●
●●
●●●
●
●●
●●
●
●
●●
●
●●●
●●
●
●
●
●●●●●
●
●●●
●
●●
●
●
●●●●
●●
●
●
●
●●●●
●●●●
●●●
●●
●●●●
●●
●●
●●
●
●●●●
●
●
●●●●
●
●●
●●
●
●●
●
●●●●
●●●
●●●
●
●●
●
●
●●
●
●
●
●●
●●
●
●
●
●
●●●●
●
●
●
●
●●
●
●●
●●●
●●●
●●
●
●
●
●●
●●
●●●●●●●
●
●
●●
●
●
●
●●●
●●●●●
●
●
●
●●●
●
●
●
●
●●
●●
●
●
●●●
●●
●●
●●
●
●
●●●
●●●●●
●
●
●●
●
●●●●●
●
●
●●
●●●
●●●●
●●●
●
●
●
●●
●
●
●●
●●●●●
●●
●
●●●
●●●●
●●
●
●
●
●
●●
●●
●●●
●
●●●
●
●
●
●●●
●
●
●
●●
●
●●●
●
●●
●
●
●●●●●
●●●●
●
●
●●●
●
●
●
●
●
●●●●●●●●●
●●●
●●●
●●●●●●●
●
●●●
●
●●●●●●●●●●●●●●●
●●●
●
●
●
●
●
●
●
●
●
●
●●●●
●
●●●●
●●
●●
●●●●●
●
●
●●
●●
●●
●●
●●
●
●●●●
●●●
●●
●
●●
●●
●●●●●●
●●
●●●
●
●●
●
●
●
●●
●
●
●●●●●●●
●●●●●
●●
●●
●●
●●
●
●●
●
●●
●
●
●
●●
●●●
●
●●●
●●●●●●
●
●
●
●
●●●●
●●●
●●●●
●●●●
●
●
●●
●
●●●●
●●
●●●●
●
●
●
●
●
●
●
●●
●●
●●
●●●●
●●
●
●●
●
●
●●●●●
●●
●●
●
●
●●
●
●●
●
●●●
●
●●
●●
●
●●
●●
●
●●
●
●
●●
●●
●●
●
●●●
●
●●●
●
●
●
●●●
●●●●●●
●●
●●●●
●●
●
●●
●●
●
●
●●
●
●
●●●
●●
●●
●
●●
●●
●●●●
●●
●●●
●●●●
●
●
●
●
●●
●
●
●●●●
●●●
●●●●
●●
●
●
●●
●
●●
●●
●
●●
●
●
●●
●●●
●
●●●
●
●
●●
●●●
●●
●
●●
●
●
●●
●
●
●●●
●
●
●
●●●
●●●
●
●
●
●●●●●●
●
●●
●●●●
●
●
●
●
●●●●
●●●●●●
●
●
●
●●●●
●●
●
●●●●●
●●●●
●
●●
●
●●
●●●●
●●
●●●
●
●●
●●●●
●●
●
●
●●
●●●●
●
●
●
●
●
●●●
●●●●●●●●●●
●
●
●
●●
●
●●
●
●●●
●●●●
●
●●
●●
●
●
●●●●
●
●●
●
●
●
●●
●●●
●
●
●
●
●
●●
●●●
●
●●●●●
●
●●●●
●
●●
●
●●
●
●
●●●●●●●
●
●
●
●●
●
●●●●●
●
●
●●
●●
●●
●
●
●
●●
●●●
●●●●
●●
●
●●●●
●●●
●●●
●
●●
●●
●●
●
●
●●●●●
●●
●
●
●●
●
●
●●
●●●
●
●
●
●
●
●
●
●
●●
●
●●
●●
●●●
●
●●
●●
●●
●●
●●●
●
●●●●
●●●●
●●
●●●
●
●●●●●●●
●●
●
●
●
●
●
●
●●
●●●●●●
●
●●
●
●●●
●
●
●●
●
●
●●
●●●●●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●●●
●●
●
●●
●
●
●
●
●●●●
●●
●●
●●●
●
●
●
●●●
●●●●
●
●●
●●
●●●
●●●●●●●
●
●●●●
●
●
●●●●
●
●
●
●●●●●
●
●
●●
●●
●
●●
●●●
●
●●●
●
●
●●●
●
●
●
●
●
●
●
●●●
●
●
●●●●
●
●●●●●●
●●●●●●
●
●
●
●●
●
●
●
●
●
●●
●
●●●●
●
●
●●●●●●●●●●●●●●
●●●
●
●
●
●●
●
●●
●
●
●
●●
●●●
●
●
●
●●●
●●●
●
●●
●●●●●●●●●
●●
●●●
●
●
●
●●●●
●●
●●
●●●●●●●●
●●●
●●
●
●●●
●
●
●
●
●●●
●
●●●●
●
●
●
●
●
●
●
●
●
●●
●●
●●●
●●●
●
●●●●
●●
●
●
●
●●
●
●
●
●●
●●
●
●
●
●
●
●●
●●●●●
●
●●●
●●●●
●●●
●
●●●●●
●●●
●
●
●
●
●
●
●●
●●●
●
●●●
●
●●
●
●●
●●
●
●●●●●●●●●
●
●
●
●
●●●●●
●
●
●●
●
●●●●
●
●
●
●
●
●●●
●
●●
●●
●
●●
●
●●●
●
●
●
●●●●
●●
●
●●
●●●●
●
●●●
●
●
●
●
●●
●
●●●
●●
●●
●
●
●●●
●●●
●
●
●●●
●●
●●
●●
●
●
●
●●●●
●●
●
●●●
●
●
●
●
●●
●
●●●●
●
●
●
●
●
●
●
●
●●●
●●●
●●
●●
●
●●●●●●●●●●●
●●
●
●
●●
●●●●
●
●
●
●
●●
●●
●●●●
●●●
●●
●
●
●●●●●●
●●
●
●●●●
●●
●
●
●●●●
●●
●●
●●
●●
●
●●
●
●
●
●●●●●
●●●
●
●
●●●●●
●
●
●●
●
●
●●
●
●●
●
●
●●
●
●●●
●●●●●
●
●●
●●●
●●●
●●
●●
●●●●
●●●●
●●●
●●●
●●●●
●
●●
●
●
●●●
●●
●●
●●●
●
●●●
●
●
●●●
●●●
●●
●
●
●
●●●
●
●●●●
●●
●●
●
●
●
●●●
●●●
●
●●●●●●●●●●
●
●
●
●
●●●
●●●
●
●
●●
●
●
●
●●●●●●●
●
●●
●●
●●
●●●
●
●●●●
●
●●●
●
●●
●
●
●●●
●
●●●
●
●●●●
●
●
●
●
●
●
●
●
●●●
●
●●●
●●
●
●●●
●
●
●
●
●
●
●●
●
●●●●●●●●
●
●●
●
●●
●
●●
●
●
●●●●
●
●
●
●●
●●●
●
●
●●●●
●●
●
●●●●
●
●
●●
●●
●●
●●
●
●
●●●
●●
●
●●●●●●
●
●
●
●●●
●●
●●
●●●●●●
●●●●
●
●
●
●●●
●●
●●●
●●
●
●●●
●
●●●●
●
●
●●●
●●
●●●●
●
●
●●●
●●
●●
●●
●
●
●
●
●●
●
●●
●
●●
●
●
●
●
●●
●
●●●●●
●
●●
●●
●●●●
●●
●
●
●
●
●
●
●
●
●
●
●●●
●●●●
●
●
●●●
●
●●●●
●●●●●
●
●●●●●
●●
●
●●●
●
●
●●●●●
●●
●
●●
●
●●
●●
●
●●●
●
●●
●●
●●
●●
●●●●●
●
●●●
●●●●●
●
●●●
●
●●●
●
●●
●●●●●
●
●
●●
●
●
●●●●
●
●
●
●●
●
●
●●
●
●●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●
●●
●●
●●●
●●
●●
●
●
●
●
●
●●●
●
●
●
●●
●●●
●
●●
●
●
●●
●●●●●●
●●●●
●●
●●●
●●
●
●
●●
●
●●
●●
●●
●
●
●
●
●
●
●●
●
●
●●
●●
●
●●●●
●
●●●●
●
●●●●
●●
●
●●
●●
●●
●●●
●●●●●●
●●
●
●●●●●
●●
●
●●
●
●
●●
●●●●
●●●
●
●
●●
●
●●●
●●
●
●●●
●
●●
●●●●●
●
●●●
●
●●●●
●●●●
●
●●●
●●●
●
●
●
●●
●
●
●●●●●●●
●
●●
●●
●
●
●
●
●●
●●
●●
●
●
●●
●
●●●●●●
●●
●
●●
●●
●●
●●
●
●
●●●
●●●●●●●●
●
●●
●●
●●●
●●●●●●●●●●●●●●●●●●●
●
●●
●
●
●●
●●
●●
●
●
●●
●●
●●
●●
●●
●
●●●●●●●
●
●●●●●●●●
●
●
●
●●●●●●
●
●
●●
●
●●
●
●
●●●●
●●
●●●●
●●
●
●●●
●
●●
●
●●●
●
●●
●
●●
●●●●
●●●●●
●●●
●●●●●●●●●●●
●
●●●●
●●●●
●●●
●●●
●
●
●●
●
●●●●●●●
●
●●●
●●
●●
●●●
●
●
●●
●●●●
●●●●●
●
●●
●●
●
●●
●
●●
●
●●●●●
●●
●●●
●●●
●●
●
●
●
●
●
●
●●●●●
●
●●●●●
●●●●●●
●●●●●●●●
●
●●
●
●
●
●●
●
●●●●
●●
●●
●●●●●
●●●●
●●●●●
●●●●●
●
●●●
●
●●●
●
●●●
●●●●●
●
●
●
●
●●
●●●●
●
●
●●●●●●●●●●●●
●●
●
●
●●
●
●●●
●
●
●
●●●●●
●●●●
●●
●
●
●
●
●●●
●
●●●●
●●●
●●
●●●●●
●●
●●●●●●●
●●
●●●
●
●●●
●
●●●●●●●
●
●
●●●●●●
●
●●●●●●●
●●
●
●●●●●
●
●
●●
●●
●●●●
●●●●●●●
●●●
●
●●●●●●●
●
●●●
●
●●
●
●●●●●
●
●●●●●
●
●●
●●●●
●
●
●●
●
●
●
●●●●
●
●●
●
●●
●
●
●
●●
●
●
●●●
●●●●
●●
●●
●●●●●
●
●●●
●●●●
●
●
●●●●
●
●●●●●●
●●●●●
●
●
●●
●
●●●
●
●
●
●●●
●
●
●●
●
●
●
●●●●●●
●
●
●●
●
●●
●
●●●●●●
●
●
●
●
●●●●
●
●
●●●●
●●●●
●
●
●●
●
●●
●●
●
●
●●●
●●●
●●●●●●
●
●●
●
●●
●
●
●
●●
●●●
●
●
●●●●
●
●
●●●
●●●
●
●●
●
●●●●●●●
●●
●
●
●●●
●
●●
●●●
●●
●●●
●
●●
●
●
●●
●
●●●
●●●●●●●●●
●
●●
●●●●●●
●
●
●●
●●
●●●●
●●●
●
●●●●●
●
●●
●●●
●
●
●
●
●●
●●
●●
●
●
●●●●●
●
●●●
●
●●
●●
●●●
●
●
●
●●●
●
●●
●●●●●●
●
●●●
●●●
●●●●
●
●
●●
●●
●●●●
●●
●●●
●●●●●
●
●●
●●
●
●
●●●●●
●
●
●
●
●●
●●
●●
●●
●
●●●
●
●●●●●●●●●●●
●●●●●●●●
●●●
●
●●
●●●●●●●
●●
●
●
●
●
●●
●
●●
●
●
●
●
●
●●
●
●
●
●●
●●
●
●●
●●●●●
●
●●●●●●●●
●
●●●●●●
●●●●●
●
●
●●
●●
●
●
●●
●
●●●
●●●●
●
●
●
●
●
●●
●●●●●●
●●
●
●
●●●●
●
●
●
●
●●●●
●●●●●●●●●●
●
●●
●
●●●●●
●
●●●●
●
●
●
●
●●
●
●
●●●●
●
●
●
●●●●●
●●
●●●●
●
●
●
●●●
●
●●
●●●
●●●●
●
●
●●●●●●●●●●
●
●
●
●●
●●
●●●
●●●●
●●
●●●●●●●●●
●
●●●●●
●
●●
●●●●●●
●●
●●
●
●
●
●
●●
●
●
●●●●●
●●●●
●●●●●
●●
●●
●
●
●
●
●
●●
●
●
●●●●●
●
●
●
●
●●●●●
●●
●●
●●●
●●●
●
●
●
●●
●●●●
●
●
●●
●
●●●●
●
●●●
●
●●●●
●
●●
●
●
●●●
●●●●●●●
●
●
●●●●●●
●●
●●
●●
●
●
●●
●
●
●
●●●
●●●●●●
●
●
●
●
●
●
●
●●●●
●●●●
●
●●
●●
●
●
●
●●
●●●●●
●●●
●
●
●
●●●●●●
●●
●●●
●
●●●
●●●
●
●●●
●●
●
●
●●●
●●●●
●
●
●●●●●●
●●
●●●
●●●
●●●
●●●●
●●●●●●
●
●
●
●
●
●
●●
●●●●
●●
●
●
●●●●
●
●●●●●●●
●●●●●●
●●●●
●
●●●
●●
●
●
●
●●
●●●●
●
●
●
●●
●●
●●
●
●●
●
●
●●●
●●●●●●
●●●
●
●
●
●
●
●●●●
●●●●●●●
●●
●●●●●●
●
●●
●●●
●●●●●●
●
●
●
●
●
●
●●●
●●●
●●
●●
●●●
●
●●
●
●
●●●
●●●●
●
●●
●
●●
●
●●●●●
●●
●
●●●●●
●●●
●●
●
●●
●
●●
●
●
●●
●
●
●●●●●●
●●
●
●
●●●
●●●
●
●
●●
●
●
●
●
●
●●●●●●●●●
●●
●
●●
●●●●●●●●●
●
●●
●●●
●
●●●
●●
●●●●●●
●
●
●
●
●●●
●●●
●
●
●●
●●
●●●
●●
●
●●●●
●
●●●
●
●
●
●
●●
●
●
●●
●
●●●●●
●
●●●●
●
●●●●●●
●
●●●
●
●●
●●●●●●
●
●●●●●●●●●
●
●
●
●●●●
●●●●
●
●●●●
●
●●●●
●
●
●
●
●●●●●
●
●
●●
●●●●●●
●●
●●
●
●●●●
●●●●
●●
●
●
●●●
●
●●●●
●●
●●●
●
●
●●●
●●
●●●●
●●●
●
●●●●●●●●●●●
●●●●
●
●
●
●
●
●●
●●
●●●
●
●
●
●
●●●●
●
●●●●●●●●●●●
●●●
●●
●●●●
●
●●●●●●
●
●●●●●
●●●
●●
●●●
●●
●
●
●●
●
●
●
●
●●●
●
●●●
●
●
●
●
●
●
●●●●
●
●●●●
●●●●
●●●●●
●●●●
●
●●●
●●●
●●
●●
●
●●
●
●●●●
●●●
●
●
●●●
●●●●
●●
●●
●●●
●●●
●
●●●●●●●●●
●●
●●●
●
●●
●
●●●●
●
●
●
●●
●
●
●
●●●●●
●
●
●
●●
●●
●
●
●●●●●
●
●●●●
●
●●●●●●●●
●●●●
●
●●●
●
●●●
●
●
●●●
●●
●
●●
●
●●●
●
●●●●●
●
●●●●●●●●
●
●
●●●●
●●●
●●
●●
●●
●
●
●●
●
●
●●
●●●
●
●●●
●
●
●●
●
●●●
●●●
●
●
●●
●●
●
●●
●
●●
●
●●●●●
●●●
●
●●
●●●●●
●●●
●●
●●●
●●●●●
●
●
●
●
●
●●●
●●
●
●●●●
●●
●●●●●●
●●
●
●
●●●
●●●
●●
●●
●
●
●●●●
●●●
●
●●
●●●●
●●
●●
●
●●●●●●●
●
●
●●●
●●●
●●●●
●●
●●●●●
●
●
●●●
●
●●
●
●●
●●●●
●●
●
●
●●
●
●
●
●●
●
●
●
●●●
●●●●
●●●
●
●
●●●●
●
●●●●●●
●●●●●
●●●
●
●
●●●●●●●
●
●
●
●●
●
●●
●●
●●●
●●●
●●
●
●
●
●●●●●●
●●
●●●
●
●●●●●●●
●●
●●●
●
●
●●●●
●●
●
●●
●●●●
●●●●●
●
●
●
●
●
●
●
●●
●
●
●●●
●●
●
●●●●
●
●
●
●●
●
●●●
●●●
●●●
●
●
●
●
●
●●●
●
●
●●●
●
●●●●●
●
●●●
●●
●●
●●
●
●
●
●
●
●
●●●
●●
●●●
●●
●
●●●
●
●
●●●●●●●●●●●●●
●
●
●●
●
●●●●●●●
●
●●
●●
●
●●●●
●●
●
●●●●●●●●
●
●●●●●
●●●●●
●●
●●●●
●●
●
●
●●
●●
●
●●
●
●●
●●●
●
●●
●
●●●●●●●●
●●
●
●●●
●
●
●
●
●
●●
●
●●●
●●
●
●
●
●
●●●●●
●●●
●●
●●●●●●●●●●●
●
●
●
●●
●
●●
●●●●
●●●●●●
●
●●
●●●●●
●●●
●●●
●●●
●
●●
●●
●
●●●
●●●
●
●●
●●●●
●●●
●●●
●
●●
●●
●●
●●●●
●
●
●●
●●●●
●●
●
●●
●●
●
●
●●●
●●●●
●
●●●
●
●
●●
●●●
●
●●●
●
●●
●●●●●●●●●
●
●●
●
●●
●●●●
●●●
●●●
●●●
●●●●●●●●●
●
●
●●
●●●
●
●●
●
●●
●
●
●●●●●
●
●●
●
●
●●
●●
●●●
●
●
●
●
●
●●
●●
●●
●
●●●
●
●●
●
●
●
●
●●●●
●
●
●●
●
●●●
●
●
●●●●
●●
●
●●●●
●●●●●
●●
●●●●●
●
●
●
●
●
●●●
●
●●
●●
●
●
●●
●●
●●
●●
●
●
●
●●
●●
●●●●●
●●
●●
●
●
●●
●
●●●●●●●
●
●●●
●●●
●
●●●
●
●●
●●●●●
●●●●●●●●●●●
●●
●●
●●
●●
●●●●
●
●
●●
●
●
●●
●●
●
●●●
●●
●
●
●●●
●●●●
●
●
●●●
●
●●●●●
●●●●
●●●●●●●●
●●
●
●
●●
●
●
●
●
●●●
●
●
●
●
●
●
●
●●●●
●●●
●
●
●●●●
●●●
●●●
●
●
●
●
●●●
●●●
●●●
●
●
●
●●
●
●
●
●
●●
●●
●
●
●
●
●
●●●●●●
●●●●●
●
●●
●●
●●
●●●
●●
●
●
●
●●
●●●
●●●
●
●●
●
●
●●
●●
●●●●
●●●
●●●●●●
●●●●
●●●●●
●●●●●
●
●
●●
●●●●●
●●
●
●●●
●
●
●
●
●●
●
●
●●●
●
●
●●
●●●●
●
●●●●●●
●●●
●
●●
●
●●●
●
●
●
●●
●●
●●●
●
●●
●●
●●●
●
●
●●●
●●●●
●
●●
●
●●●●●●●
●
●●
●
●●
●●●●
●
●
●●
●
●
●●●●
●
●
●
●●●
●●
●
●●
●●●
●
●●●●
●●
●●●
●●●●
●●
●
●●●●
●
●●●●●●●●●●●●●●
●●●
●
●
●
●●●●●
●
●●●●●●●●●●●
●
●●
●●
●●●●●●●●
●
●●●●
●●
●●●
●●●●
●●●●●
●●●
●
●●
●●
●●
●
●
●
●
●●●●●●
●
●
●●
●●
●
●
●
●●●
●
●●
●
●●
●●●
●
●●
●
●
●●●
●
●●
●●
●
●●
●●●
●●●
●●
●●
●●
●
●●●●●●●●
●●●●●
●
●
●●
●●●●●
●
●●●
●
●
●●●●●●
●
●●
●●
●
●
●
●●●●
●
●●
●
●●●
●●
●
●●●
●●●
●●●●
●
●●
●
●
●●●●
●
●
●
●●●●●
●
●
●
●●
●
●●●
●●●●
●●
●●
●●●●
●
●●
●
●●●
●
●
●●●
●●
●●
●
●●●
●●●
●●
●●●
●●●●●
●●
●
●●●
●
●●●
●●
●
●
●●●●●●●●●
●●●●
●
●
●●●●
●●●●●●●
●●
●●●●●
●●
●●●●●
●●●●●
●●
●●●
●●
●●
●
●●●
●
●●●●●●
●
●●●
●
●
●
●
●●●●●
●
●
●
●
●●
●
●●●●●●●●
●
●●
●
●●
●
●●●
●
●
●
●
●
●
●
●●
●●
●
●
●●●●
●●●●
●
●
●
●●●
●●●●
●●●●●●●●
●●●
●●
●●●●
●
●
●●
●
●●●
●
●●●
●●
●●●
●●●
●●●
●
●
●●●
●
●
●●
●●
●●●●●●●●●
●
●
●
●
●●●
●●●●●
●
●●●●●
●●
●
●
●
●●
●●
●
●
●
●
●●
●●
●●●
●●
●
●●●●●
●
●●●●●
●
●●●●
●●●●
●
●●●
●
●●
●
●
●●
●
●●●
●●●
●
●
●●
●
●
●●●
●
●
●●
●
●
●
●●
●
●
●●●
●●●●●●
●●
●●
●●
●●
●
●●●●●●●●
●●
●●●●●●
●
●
●●
●●
●
●
●●●●●●●
●
●
●
●
●●
●●●
●
●
●●●
●
●●
●
●
●
●●●●
●
●●●●●●
●●●
●●●●●●●●●●●
●●●●●
●
●
●●
●●●
●●
●●●
●
●●
●
●●●
●●●●
●●
●
●●●●
●
●●●●
●●
●●
●
●●
●
●
●
●●●●●
●●
●
●●
●●●●●
●
●●●
●
●
●
●
●●●●
●●●●
●●●
●●●●
●●●●
●
●
●
●●
●
●
●●●
●
●
●
●
●●
●●●●●●
●●
●●●
●●●●
●●●●
●●
●
●●●●●●●●
●●●●●●●●●
●●
●
●
●
●
●●
●●●●●●●●●●●
●●
●
●●
●
●
●●
●●●
●
●●●
●●●
●●
●●●●
●●●●●
●
●●
●
●●
●●●●●
●●
●●●
●●●●
●
●●●●●
●●
●
●●
●●
●
●●
●●
●●
●●
●
●●
●●
●●
●●
●●●●
●●
●●●
●
●●
●●
●●
●
●●
●●
●●●●●●●
●
●●●●●●
●●
●
●
●
●
●
●●●●
●
●
●
●
●
●
●
●●
●●●
●
●
●
●
●
●●●
●
●
●
●
●●●
●
●●●
●●●
●●
●
●●
●●●●●●
●●●●●●
●●
●●
●
●
●
●
●
●●
●
●●●●
●
●
●
●
●
●●
●●
●
●●●●●
●
●●●●
●●●●●
●●●●●●
●●
●
●
●
●●●●●●
●
●●●●●●
●●
●●●●●●
●●●●
●●●●●●●
●●
●
●●●●
●
●
●●●
●●
●
●●
●
●●●●
●●●
●
●●●●●●●●●●●●●●●●
●●
●
●
●●
●
●●●
●
●●
●●●
●●
●
●
●
●
●
●
●
●
●●●●
●
●●●
●●●●●●●
●●
●●●●●●●●●
●●●
●●
●●
●
●●
●
●●
●
●
●
●●●●●
●●●●●●●●●
●
●
●●
●●●
●
●
●
●●●
●
●●
●●
●●
●●●
●●
●
●●●
●●●●●
●
●●●
●
●●
●●
●●
●
●
●●
●
●●●●
●
●
●●●●●
●
●
●
●●
●
●●●
●●●●
●
●●●
●●●
●
●●
●
●
●●
●●●
●●●
●●●●●●●
●
●
●
●●●●●
●●●●
●
●
●●●
●●
●
●
●
●●
●●
●
●●●
●●●●●●
●
●●●●●●●●●●
●●●●●●●●
●●●●
●
●
●●●●
●
●
●
●
●●●●●
●●●
●
●●●
●
●
●
●
●
●
●●●●●
●●●●
●●●
●
●●●
●
●
●●
●●
●●●
●
●●●
●●●●●●●●
●
●●●
●
●●
●
●●●
●●●
●
●●●
●●
●●●
●
●
●●
●●
●
●
●
●
●●●
●
●●●
●
●
●
●
●
●
●●●●●
●●●●
●●●●
●●●
●●
●●●●●
●●●
●
●
●●
●
●●
●●●●●
●●
●●
●●
●●
●
●●●
●●●
●●
●
●●
●
●
●●
●●
●
●●●
●●
●●●●●
●
●
●●
●
●●●
●●●●●
●
●●●
●●●●●
●●●●●●●●●●
●●●●
●
●●
●●
●●●●●
●
●
●●
●
●
●●
●
●
●●●●●●●
●
●●
●
●
●
●●●●
●
●●
●
●
●
●
●
●●●
●
●●
●
●●●●
●
●●
●
●●
●●
●●●●●●●●
●●●●
●●●●●●●●●
●
●
●
●●●●
●●
●
●●
●
●●●●
●
●●
●
●●
●
●
●
●●
●●
●●●
●●●
●
●●●●●
●
●
●●
●
●
●
●
●
●
●●
●●
●
●●●●●●●
●●●●
●●
●
●
●●●
●●●●
●
●●●
●●
●
●
●
●
●
●●●●●●●
●●
●
●●
●●●
●●●●●
●
●●●●●
●●
●
●●●●
●●
●●●●
●
●
●
●●
●●●●
●●
●
●●●●
●
●
●●
●
●
●
●
●●●
●●●
●
●●●●●●●
●
●●●●●
●●●
●●
●
●
●●●●
●●●
●
●●●
●●
●
●
●●
●●●
●●
●●●●●●
●●
●
●
●
●●●
●●●●●
●
●●●●
●●●
●
●
●
●
●
●●●●●●
●●
●
●●
●●
●●●
●●●●●
●●
●●●●●
●●●
●
●●
●●●●
●
●●
●
●●●
●●●●
●●
●●●●●●●●●
●
●
●
●
●
●●
●●●●●
●●
●●●
●●●●●●
●
●●●
●●
●●
●●
●●●●
●
●
●
●●●●●●
●●●●
●●●
●●●●●
●●
●●●
●
●●●
●●
●●●●●
●
●●●●●●●●
●
●
●
●
●●●
●●●●●●●●
●●●●●●●●
●
●●
●●
●●
●●
●●●●
●
●
●●●●
●●
●●●●
●●
●●
●
●
●●
●
●●●●●
●
●
●●
●●●●●●●●●●
●●●
●
●●●
●
●●
●●●
●
●●●
●●●●
●●●
●
●
●●
●
●
●●●●
●●
●
●●●●●●●●●●●
●●●●
●●
●●
●●●●●●●
●●●
●●●
●
●
●
●●
●●
●
●●
●●●●●
●●
●●
●●
●
●●
●●●●●●●
●
●
●
●●●●
●●
●●
●
●●●●●●●●
●●
●●
●●
●
●
●●●●
●
●●
●
●
●●
●
●●
●
●
●
●●
●●●●●●
●●●●●●
●●●
●
●
●●
●
●●●
●●●●●●●
●●●
●●
●
●
●●●●●●
●
●
●●●
●●●●●●●●
●
●
●●
●
●●●●●
●
●●●
●●
●●●
●●
●
●●
●●
●
●
●●
●
●●●●●
●●●●
●
●
●●●
●
●
●●●
●●
●●
●
●●●
●
●
●
●●
●●
●●
●
●●●
●
●
●●●●
●
●●●●
●●●
●
●
●●●●●●
●●
●●
●
●●
●●
●
●
●
●
●
●
●
●
●●●
●●●●
●
●●●
●
●●●
●
●●
●
●●●●●
●●●
●●●●●
●●●●
●●●●●●
●●●●●●●●●●●●●●
●●●●●
●●
●●●
●
●
●●●●●●
●
●●
●●
●
●
●●●●●●●
●
●
●●●●
●
●●●●
●
●●
●●●●
●
●
●●
●●
●●
●●●
●●●●
●
●
●
●
●
●●●●
●●
●
●
●
●
●●
●●●
●●
●●
●●
●●
●
●
●
●●●●
●●●
●●●
●
●●●
●●●
●
●
●●●●
●
●
●●●●●
●●●●
●
●●
●●
●
●
●●
●●
●
●●●
●●●
●
●●●
●
●●●
●
●●
●
●
●●●●
●●
●●
●●
●
●●●
●●●●●●●
●●
●
●●●●
●
●
●
●●●●●
●●
●●
●●
●
●●
●●●
●●●●
●●
●●●
●
●
●
●●●
●
●●●●●●●
●●
●●
●●
●
●●●●
●
●●
●
●●●
●
●●●●●●●
●
●
●●●
●
●
●
●●●●●●
●
●●
●●●●
●●●●●●
●●●●●●●●
●
●●●●●
●
●●
●
●
●●
●
●
●●●●●●
●
●
●
●●
●●●●●
●●●●●●●●
●●●
●
●
●
●
●●
●
●
●
●●
●●
●
●●●●●
●
●●
●●●
●
●
●●●
●●
●●●●●●●●●●●
●●●●
●
●●
●
●
●●
●●
●
●
●●
●
●●●●●
●
●●
●
●
●
●
●●●
●●
●●●●●●
●●
●●
●●●●●
●
●●
●
●
●●
●
●●
●●●
●
●●
●
●
●●
●●
●
●●●
●
●
●●●
●●
●
●
●●
●●
●
●
●●
●●
●
●●
●●●
●
●●●●●●●
●
●●●
●
●
●●●
●●
●
●●
●
●●●●●●●●
●●
●●●●
●
●
●
●●
●●
●●●●●
●●●●●
●●●●
●●●●●●●●
●●
●
●
●
●
●●
●●●●●
●
●●●
●
●
●
●●
●●●●
●●●●●●
●●●
●●
●●
●●
●●
●
●●●
●
●●
●
●●
●
●
●●●●●●●
●
●
●
●
●
●
●●●●
●●●
●
●●●
●
●
●
●●
●
●●
●●●●●●
●●●●●●●●●●
●●
●●●
●●
●
●
●●●●●
●●●●●●
●●
●●
●
●●
●
●
●●
●
●●●●●
●●
●
●●●
●●
●●
●
●●
●●●
●●●
●
●●
●●
●
●●
●●●●●
●●●●●●●
●
●●●
●
●●●●
●●●
●●●
●●
●
●
●●●
●
●
●●
●
●●
●●●
●●
●●
●
●
●
●
●
●
●●●●●●
●
●●●●
●●●
●●
●●●
●
●●●●
●●
●
●●
●
●●●
●
●●●
●●
●
●●●●
●●●●●
●
●
●●●
●
●
●●
●●●●●
●●●
●
●
●
●
●●
●●
λ10 λ90 µ10 µ900.0
0.2
0.4
0.6
0.8
1.0
Size
cor
rect
ed ra
tes
North American Perissodactylaλ0 = 0.42µ0 = 0.75µ = mm
b10 = 81b90 = 1159µ = mm
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●● ●
●●●●●●
●
●●●●
●
●●●●
●●
●
●●●
●
●●●●●●●
●●●●
●
●
●
●●
●●
●●●●
●
●
●●
●●●
●
●
●
●●
●
●
●
●
●
●
●●
●●●
●●
●●
●●●●●●●●
●●
●●●●●
●●
●●●●●
●●●
●
●
●●
●
●
●
●●
●●
●
●●●
●
●●
●●●
●●●●●●●
●●
●
●
●●
●●
●●
●●
●
●●●●●
●●●●
●
●
●●●●
●●●●●
●●●
●
●●●●●●●
●
●
●●●●●●
●
●●●
●
●
●●●●●
●●
●●
●
●
●
●●●
●
●●●●●●
●●●●●●●●●●
●●●●●●●●●●●●
●●●●●●
●
●●●●●
●
●
●
●●
●
●●●
●
●●●
●
●●
●
●●●●
●●●●●
●●
●●●●
●●●●●
●●●
●●●
●
●●●●●●
●●●
●
●
●
●
●●●●●
●
●
●●●●
●
●
●
●●●●●●●●●●●
●●●●●
●●
●●
●●●
●●●●
●●●
●
●
●●
●
●
●
●
●
●
●●
●
●
●●●●
●●●●●
●
●●●
●
●●
●●●●
●
●●●●●
●
●●●●●●●●
●
●
●
●
●
●●
●●●
●
●●
●●●
●
●●
●●●
●●●
●●●●●●●●●
●●
●●
●●●
●
●●●●●●●
●
●
●
●●
●●●
●
●
●●
●●
●
●
●●●
●●
●●
●●
●●●●●●
●●●●●●
●
●●●●●
●
●●
●
●
●
●
●
●
●●●
●●●
●●●●●
●●
●
●●
●
●
●
●●●●●
●
●
●●●●
●
●●●
●●●●
●
●
●
●●●●●
●
●
●●
●●●
●
●●
●
●●●
●●●
●
●●
●
●●
●●
●●●●●
●●
●
●
●●
●●●
●●
●
●●●
●
●
●●
●●
●
●
●
●●
●●
●●●●●●●
●
●
●
●●●●●●●●●●
●●
●
●●●●●
●
●●
●●
●
●●
●
●
●●●●●●
●●●
●
●●●●●
●
●
●●●
●●●
●●
●●
●●●
●
●
●●●●●
●●●●●●●
●
●
●
●
●●
●●
●●
●●●●●
●●
●
●●●
●●●●●●
●
●●
●●●●●●
●
●●●●●●●●
●
●●●●
●●●
●
●
●●●●
●●●
●
●●
●●●●●
●
●●
●●●●●
●
●●
●●●●●●
●
●
●●
●●
●
●●●
●●●●●
●
●
●
●
●
●
●●●
●
●
●
●●
●
●
●
●●●
●●●●●●
●
●
●●
●●●
●●●●●
●●
●●●●
●
●
●
●
●
●●●
●●
●●●
●
●●
●●
●●
●
●
●
●
●
●●
●
●●●●●●
●●
●●●●●
●●
●
●
●●
●
●
●
●●
●●
●●●●●
●
●●
●
●●●
●
●
●●
●●●●●●●●
●
●●
●●●●●●●
●
●●●
●●●●●●●
●
●
●
●
●
●
●●●●●
●●
●
●
●
●
●●●
●
●
●●
●●●●●●
●●●
●
●●
●
●
●●●●
●●●
●
●
●
●
●
●
●
●
●●●●●●
●
●●
●●
●●
●
●
●
●
●●●●●●
●
●●●●●●●●●
●●
●●
●
●●●●●●
●
●●●
●●
●●
●●
●●●
●
●●
●
●
●●
●●
●
●●●
●●●●●●●
●
●●●●
●
●●
●●●●●●●●●
●
●
●
●
●
●●●●●
●●●●
●
●●
●●
●
●●
●●●
●●●●●●●●●●
●
●●
●
●
●
●●
●●●●
●●
●●●
●●●
●●
●
●●●
●●
●●●●
●●●
●●
●●●●●
●
●●
●●
●●
●●●
●●●●●●●●
●●●
●●
●●●
●●
●●
●●●
●
●●
●
●●●
●●
●
●
●
●●●
●●
●
●●
●
●●●●●●
●
●●
●●●●
●●
●●●●●
●●●
●
●●●●
●●●●●●●
●●●●●●●●●●●●●●●●
●●●●
●●●
●●●
●●
●●●●●●●●
●
●●●
●●●
●
●
●●
●●
●
●●●●●●
●
●●●●●
●
●
●
●●●●
●●●
●●
●●
●
●
●
●
●
●●●●●
●
●●
●●●
●●●●●
●
●●●
●
●●●●
●
●●
●●
●
●
●
●●
●
●●
●●●●●●●
●●●
●●
●●
●●●
●●
●
●●●●
●●●●
●●●●●●●
●
●
●
●●●
●●
●●●
●
●
●●
●●●
●
●
●
●●●●●
●
●●●●
●●
●●●
●●●●
●
●●●●
●
●
●
●●●●●
●
●
●
●●●
●
●
●
●
●
●●
●
●●●
●
●●●●
●
●●
●
●
●●
●●
●
●●●●●●●●●
●●
●
●
●●
●●●●
●
●●●
●●●●●●
●
●
●
●●
●
●
●
●
●
●
●
●●●●
●
●
●●
●
●●
●●●
●●●
●●
●
●
●●
●●●●
●
●●●●●●●
●●
●
●●
●●
●●●
●
●
●●●●
●
●●●●●●●●
●●●●●●●●●●●●●
●●
●●●
●
●
●●
●●
●●●●●
●●●●●●●●●●
●
●●
●●●
●
●●●●
●●
●
●●
●
●●●
●●●●
●●
●●●
●●●
●●●
●
●●
●●●●●
●
●
●
●
●
●
●
●●
●●
●●
●
●●●
●●
●●●●
●●●●●●●
●●●
●●●●●●
●
●●●●●●
●
●●●●
●
●
●
●
●●
●●●●●
●●●
●●●●●●●
●●●
●●
●
●
●
●●●
●●●
●●●●●●
●●●
●●
●
●●
●●●●●
●
●
●●●
●●●●
●●
●
●●●●●
●
●●
●●●
●●●
●
●
●
●●
●
●
●●●
●●●
●●●●
●
●●
●
●
●●
●
●
●
●
●●●●●
●
●
●
●●●●
●●●
●●●
●
●
●●
●
●
●●●
●
●
●●
●
●●
●
●●●●●
●
●●●
●
●
●
●●●
●
●●
●●●●●●●●●●
●
●●
●
●●
●●
●
●
●●
●●
●
●●
●●
●●●●●●●●●●
●●●●●
●●●●
●
●
●
●
●●
●
●
●●●●
●
●
●●
●●●●●●●●
●●●
●
●●
●
●
●●●●●●●●●●●●●●●
●
●
●●
●●●
●●●●
●●
●●●●●●
●●●●
●●●●
●
●
●●●
●●●●●
●
●
●●●●
●●
●
●
●
●●●●
●●
●
●●
●
●
●●●●
●
●
●
●
●
●
●●●
●●●●●
●
●●●
●●●●
●●
●
●●
●
●●●
●
●●●●
●
●●●●●
●
●
●
●●
●●●●●
●●●●●●
●●●
●
●●
●●
●●
●●●●
●●●
●●●●●
●●●●●
●●●●
●
●
●
●●●●●
●
●●●●
●
●●●●
●●
●●
●
●●
●
●●●●
●
●
●●
●●
●
●
●●
●
●
●●●●●
●●
●
●●
●●●●●●
●
●
●
●
●
●
●●●●●●●●
●●●
●●●
●
●●●
●
●
●
●
●
●●●●●
●
●●
●
●●●
●●●●●●●●
●●●●
●●●●●●●●●
●
●
●●●●●
●
●
●●●
●
●●
●●●●●●●
●●●●
●●●●●
●
●●●●●●●
●
●●
●
●
●
●●
●●●●
●●
●
●
●●●●●●●●
●
●●●●●
●●
●
●●●
●
●
●●●●
●
●●●
●
●●
●●
●●●
●
●
●●
●●●●●
●●
●
●●●
●●●●●●●●
●●●●●
●●●●●●
●
●
●●●●
●
●●
●
●●●●
●
●●●
●●
●●●●
●●●
●●
●
●
●●
●
●
●●●●
●●
●
●
●
●●●
●
●●●●
●●●●●
●●
●
●●
●●●
●●●●●
●●●●
●●
●●
●
●●●
●●
●●●●●
●●●●
●
●
●●●
●●●●●●●●●
●●
●●●
●●●●●●●●
●
●●●
●
●
●
●
●
●
●●●
●●●●
●
●●
●
●●●
●●●●
●
●
●
●●
●●●●●●●
●●
●●●●●●
●●●●
●
●●
●
●●●●●
●
●●●●
●●●
●
●
●●
●●●●●
●
●
●●●●●●●●
●●●
●
●●
●
●
●
●
●
●●●
●●●
●
●
●
●●
●●●●●
●●
●
●●●●
●●
●
●●
●
●●
●
●●●
●
●
●●
●●
●
●●●●●●●
●●●●●●
●●●●●●●
●
●●●
●
●●●
●●●
●●●●●●●●●
●
●
●
●●●
●
●●
●●●●
●
●●
●
●
●
●
●
●●●●
●●●●●
●●
●
●●●●●
●●
●●●
●●
●●●●●●●●●●●
●●●
●●
●●
●
●●●●
●
●●
●
●●
●●●
●●●●●●
●●
●●
●●
●●
●
●
●●
●
●
●●
●●●
●
●
●●
●●●●
●
●
●
●
●●
●
●
●
●●●●●●●●
●●●●●
●●
●●
●●
●●
●●
●●●●●●●●
●●●
●
●
●
●●●●●●
●●●●
●
●
●●
●
●●●
●
●
●
●
●
●●
●●●
●●
●●
●
●●●
●●●●
●●
●
●●●
●●
●●●
●●●●●
●●
●●●
●
●●
●●●●●
●●
●
●
●
●
●●●
●●
●
●
●●●
●●
●●●●
●
●
●●
●●●●●●
●
●●●
●●●●●
●
●●●
●●●●●●●●
●●
●●●
●●
●●●
●
●●●●●●
●●●●
●
●●●
●
●
●●●
●
●●
●●
●●
●●●●
●
●
●
●
●
●
●●●●●
●●●●●●
●●●●●●
●
●●●●●●●●●●●
●
●●
●●●
●
●●
●●●●
●●●●
●
●
●●●
●●●
●●
●
●
●
●●
●
●●
●●●●●●
●
●●
●●●●
●
●
●
●●
●●●
●
●●●●
●●●●●●●●
●●
●●
●●●
●
●
●
●●
●●
●
●
●●
●●
●●
●
●●
●●
●●●●●
●
●●●●
●●●●●●
●●●●
●
●●●●●●●●
●●
●
●●
●●●●
●
●
●●
●
●
●
●
●●●
●
●
●
●●●
●
●
●
●●
●●
●
●
●●
●
●●●
●●
●
●
●●
●●●●
●●●●
●
●●●●●●
●
●●●
●●
●●
●
●
●●
●
●●●●
●●●
●
●●
●●●●●
●●●
●●●
●●●●
●
●●
●●●●
●●
●
●●●●●
●●
●
●
●●●●●●
●
●
●●●●●●
●●
●
●
●
●●
●●
●
●●
●
●●●●
●●●●●●
●
●
●
●●●●
●
●●●●
●
●●●●
●
●●●
●
●●●●
●
●
●
●●●
●●●●●●●
●●
●
●
●●
●●
●●●●●●●
●
●
●●●●●●●●●●●
●●
●
●
●●
●●
●●●●
●●
●●
●●●●●
●
●
●
●
●●●●
●●
●
●●
●
●●●●●●●
●
●●
●●
●●●
●●
●
●
●
●●
●●
●●●●
●●●●
●●
●●●●●
●●
●●
●●●
●
●●●
●
●
●
●
●
●●
●●
●
●●
●●
●●●●●●●
●
●●
●●
●
●●●●●
●
●●●
●●
●●●
●●●●
●
●●●
●
●
●●●
●
●
●
●●●●●●●●●●
●
●●
●
●●
●●●
●
●●
●●●●●●●●●
●
●●
●●●
●
●●●●
●
●●●●●
●
●
●●●●
●●●●●●
●●
●
●●●
●
●●●●●●
●●●
●●●●●
●
●
●
●
●
●●●
●●
●
●●●●
●
●●●●
●
●●
●●●
●
●
●●
●●●●●
●
●
●
●
●●
●●●●●
●●
●●●●●
●●●
●
●●
●
●
●●
●
●●
●
●
●
●
●●●●●●
●
●
●●●
●
●●
●
●
●●●
●●●
●
●
●●●●
●
●
●
●●●
●●
●●
●●
●
●
●●●
●
●
●●
●●
●●
●
●●●●●●●●
●
●
●
●●
●
●
●●●●
●
●●●●
●●
●
●●●●●●●●●●
●●●●●●●●●
●●
●●●
●●
●
●●●●
●●
●●●
●●
●●●
●●
●
●
●●●●●
●●●
●●●●●●●●
●
●
●●
●
●
●
●●
●●
●
●●
●
●
●●●
●
●
●●
●●
●●●●●●●●●
●●●●
●
●●
●
●●●●
●●
●●
●●●
●
●●●
●
●
●●●
●●
●●●●
●●●●
●
●
●
●
●
●●●
●
●●●
●
●
●●●●●●●●●
●
●
●●●●
●●●●
●●
●
●
●●
●
●●
●●●●●●
●
●
●
●
●●
●
●
●●●●●●●
●
●●●
●●
●●
●●●●
●●
●●
●●●
●
●●●●●●
●
●
●
●
●
●
●
●
●
●
●●●
●●
●●●
●
●
●●●●●●●
●
●
●●●
●
●
●●●●●
●●●●●●●
●
●●
●
●
●
●
●●●●●●
●●●
●
●●
●●
●●●●●
●
●●●
●●●
●
●●
●
●●
●
●
●
●●●●●●●●
●
●
●
●●●
●●●●●
●●●
●●●●●
●●●●●●●
●
●
●
●
●
●●
●●●●●●●
●●●●●●●●●●
●
●●●●●●
●
●
●●●
●
●
●
●●
●●●●
●●
●
●●
●●
●●●
●
●●●●
●●●●
●
●
●
●
●●●●●●●
●
●●
●●●
●
●
●
●●
●●
●
●●
●
●●●●
●●●●●●●●●●
●
●
●
●
●
●●
●●
●●
●●●
●
●●●
●●●●●●●
●
●●
●
●●●
●●
●
●
●●●●
●●
●
●
●●●●●
●●●
●●●●●●●●
●
●●●●
●●●●●●●●●
●
●
●●
●●●
●
●
●●●●●●●●●●●●●●●●●●●●
●●
●●●●●●
●
●●●
●
●●
●
●
●
●●●●
●●●
●
●●●●●●●●
●●●
●●●●
●●
●
●●●
●●
●●
●●●
●●
●●●●
●
●●●
●●●●●●●●●●●
●
●
●●
●●●
●
●
●●●●
●●●
●●●●
●●●●●●●●
●●●
●
●●●●
●
●●●●●●●●
●●
●
●●●●●
●
●
●
●●●
●
●●
●
●●
●●●●
●
●●●
●●
●
●●●●
●●●●●●
●●●●●
●
●●●
●●●●
●●●
●
●
●
●●●●●
●
●
●●
●●
●●●●
●●●●●●●●
●
●
●●●●●●●●●
●●
●●
●●
●
●●●
●●●●●
●●●●
●
●●●
●
●●
●
●●●●●
●●●
●
●
●
●
●●
●
●●●
●●
●●●●●●●●●
●
●●●●
●
●
●●
●●●●●●●
●
●
●●
●
●
●●
●●●●●●●●●●
●●
●
●
●●●●
●●●●●●
●●●●
●
●
●
●
●
●
●
●
●
●●●
●
●●●●●
●●
●●
●●●●
●●
●●
●
●●●
●●
●●
●
●
●●●
●●●●●●●●●●
●
●
●●●●
●
●
●●
●●
●●●
●●
●
●●
●●●●●●●●●●
●●
●●
●
●●
●●●
●●●
●●
●●
●
●
●
●
●●●
●●●●●
●●
●●
●
●
●●●●●
●
●●
●●●●●●
●●●●
●●
●●
●●●●
●●●●●●●
●
●
●●
●
●
●●●
●●
●●
●●●●●●●
●
●
●
●●●●●
●
●
●
●●
●
●●●●●●●●
●
●●●●●
●●
●●
●
●●
●
●●
●●●
●
●●●
●●
●●●
●
●●●●
●●●●
●●●
●●
●●●●
●●●●●
●
●
●
●
●
●
●
●
●●●
●●●●
●●●●
●●●
●●●●●●
●●●●
●●●●●●●
●
●
●
●●●●
●
●
●●
●●
●●●
●●●●
●●●
●●●
●
●●
●
●●●
●●
●
●
●
●●●
●●●●●
●●●
●●
●
●
●●●●●
●●
●●●
●
●●●
●●
●●●
●
●●
●●●●●●●
●
●●●
●●●●●●
●
●
●●
●
●●●●●●●
●●●
●
●●●
●●
●●●
●●
●
●
●
●●
●●
●
●●●●
●
●●●●●
●
●●●●●●●●●●
●
●●
●
●
●●
●●●●●
●●●●●●●
●●
●
●
●●●
●●
●●
●●●●●●●●
●●
●●
●
●
●●
●
●
●●
●●●
●
●●●
●
●
●●
●●●●●
●●●
●
●
●●●●
●●●●
●●
●●
●●
●
●●●
●
●
●
●●●
●
●●
●
●
●
●●
●
●●
●●●
●
●●
●
●
●
●
●
●
●●●●●●●
●●●
●●
●●
●●●●
●
●
●
●●●●
●
●
●
●
●
●
●
●●●
●●
●
●
●
●●
●●
●
●●●
●●
●
●
●●
●●
●●
●●●●
●
●●●●
●●●●●
●
●
●
●
●●●
●
●●
●●
●●●●
●●
●●
●
●●
●
●
●●
●●
●
●●●●●●
●●●●
●●●●●
●●
●●●●●●●●●
●
●
●
●
●●
●●
●
●●
●●
●
●
●
●
●●
●●
●●
●●●●
●
●
●●●●
●
●
●●●
●
●●
●●●
●
●●
●
●●
●●
●●●
●
●●
●●
●●●
●●
●
●●
●●
●
●
●●●●●●●●●●
●
●●●●●●
●●●●
●●●●
●●
●●
●
●●
●
●●●●●●●
●
●●●●●
●
●
●
●
●
●●
●●●
●●
●
●
●
●●●●
●
●
●●●●●
●●
●●
●●●●●
●●
●●
●●●●●●●
●●
●
●●●
●●●
●
●●
●
●
●
●
●
●
●
●●
●●
●
●
●
●●●
●
●●
●
●
●
●●●●
●
●
●●
●●●
●
●●●
●
●
●
●●●
●●●
●●
●●●●●●●
●●●
●●●
●
●●●
●
●●●
●●
●
●
●●●●●
●●●●●●
●
●
●●●●
●●
●
●●
●
●
●
●●
●
●
●
●●
●
●●●●●●●●
●
●
●●●●
●●●
●●●
●●●●●●●●●●
●●●
●●●
●
●●
●
●●●
●
●
●●●
●
●
●
●●
●●
●●●●
●
●●
●
●
●
●●●●●●●●
●
●●
●
●
●●●●
●
●
●●●●●●
●●●
●
●●●
●
●
●
●●●
●
●●
●●●●●
●●
●
●
●●
●●●
●●●●●●●●
●●●●●
●
●
●
●
●●
●
●●●
●
●●●●●●
●
●●●
●
●●
●●
●●
●●
●●
●
●
●●
●●
●●●●●●
●
●●
●●●
●●●●●●●
●
●●
●●●
●●
●●●●
●●●●●●●
●
●
●
●●●●
●●●
●
●
●●●●●●
●
●●
●●●
●●●●
●
●
●●
●
●
●●●
●
●
●●●●●
●●
●●
●●●●
●
●●
●●●
●●
●●●●●
●●
●●●●●●●●
●●
●
●●
●●●
●●
●●●●●●●●●
●●
●●
●
●
●
●
●
●
●●●●
●
●●●●●●●
●
●
●●●
●
●
●
●●●
●●●
●
●
●●
●
●●
●●
●●●●●●
●●
●●
●●
●●
●
●
●
●
●
●●●
●
●
●
●
●●●
●
●●
●
●●
●●
●
●●●
●
●
●
●●
●
●
●●●●●●
●
●
●●
●
●●
●
●●●●●●
●●●●●●●
●●
●
●
●●
●
●●
●●●●
●
●●●
●
●●
●
●●●●●●●●●
●
●●●●
●●
●
●
●●
●
●●
●
●●
●●●●●
●
●●●
●
●
●
●●
●●●●●●●●●●●
●
●●●●●●●●●
●
●●●●
●
●●●
●
●
●
●●
●●
●
●
●
●
●●●●
●
●
●
●
●
●●●
●
●
●
●
●●
●
●
●
●●●●●
●●●
●
●●●
●●●●●●●
●
●
●
●●
●●●●
●
●
●
●
●
●●
●●●
●
●●●●●●●
●●●
●
●
●●●●
●
●
●●●
●
●●●●●●
●●●●
●
●
●●
●
●●●
●
●●
●●●●●●
●●●●
●
●
●●
●●●
●●●
●
●●●●●●
●
●●●●
●●
●●
●●●
●●
●
●●●●●●
●
●●●
●
●●●●●●
●
●
●
●
●●●●●●
●
●●●●●
●
●
●
●●
●●●●●
●
●
●
●●
●
●●●
●●●
●
●
●
●●●
●●●●●●
●●●●●●
●
●
●
●●
●
●
●●
●
●
●
●
●
●●
●
●●
●
●●
●
●●
●
●●●●●●
●●
●●
●
●●●
●●
●●
●●
●●
●
●
●
●
●●
●
●
●
●●●●
●
●●
●
●●●●●●
●●
●●●●
●
●
●●
●
●●
●●
●●●
●
●
●●
●
●●
●
●
●●
●
●●
●●●
●●
●●●●
●
●
●●●
●●●
●●
●●●●●●●
●●
●
●
●●●
●●
●
●●●●●●
●
●●●●
●●
●●●
●
●●
●
●●●
●
●●
●●●
●
●●
●●●●
●●●●●●●
●●●●●
●●
●
●●●
●●
●
●
●
●●●
●●●●
●
●●●●●
●●
●
●●●
●
●
●●
●
●●●
●●
●
●●●
●
●●
●
●
●
●
●●●●
●
●●●●●●●
●
●●●
●
●●●●●●●
●●
●
●
●
●
●
●
●●
●●●
●
●●
●●●●●●●
●
●●
●
●
●
●●
●●●●
●●
●
●●●●
●●●●●●●●
●
●●●
●●
●
●●●●●●●●●●
●●
●●●●
●●●
●
●●●●
●
●●●
●
●
●●
●●●
●
●
●
●●
●●
●●●●
●
●
●●
●●●
●●
●
●●
●●●●●
●●●
●●
●
●●●●
●
●●●●
●●
●
●
●●
●●
●●●
●
●●
●
●●●●
●
●
●●
●
●
●●●
●●●
●
●●●●●●
●●
●
●
●
●●
●●●
●
●
●●
●
●●
●●●
●
●●
●●
●
●
●
●●
●●
●●
●
●
●●
●●●●●
●
●
●
●●
●
●
●
●●
●●
●
●●●
●●●
●●
●
●●
●
●●
●
●●●
●●
●●
●●●●●●
●
●
●
●●●
●
●
●●●●
●●
●●●
●●
●●
●●●●●
●
●
●●●
●●●●●●
●●●●●
●●●
●●
●
●●●
●●
●●●●●●●
●●
●●
●
●●
●
●●●●
●●
●
●
●●●●
●●
●
●
●●
●●
●
●●●●●●●●●
●
●●
●●●
●
●●●●
●
●●●●
●
●
●
●
●●●●●●
●
●●●●
●●
●●
●
●●
●●●
●
●●
●
●
●
●●
●●●●●●
●
●
●●
●●●●
●
●
●●
●●
●
●●●●●●●●●●
●●
●
●●
●
●●●
●
●●●●●●●●●
●●●
●●●
●●●
●
●
●●●●●
●●●●●●
●●
●●●●
●
●
●●●●
●
●
●●
●●
●
●●
●●
●●●
●●●●●●
●
●
●
●●●
●●●●●
●
●●●●●
●
●●●●●●
●
●
●
●●
●
●●●
●●●●●●●●●
●
●
●●
●
●●●●●
●
●
●
●●
●●●●●●●●
●●●
●
●
●
●●
●●●●●●
●
●
●
●●●●
●
●
●●●●●
●
●●●●
●●
●
●●●
●
●
●
●
●
●
●●●●
●●
●●
●
●●●
●●
●
●
●
●●●●●
●
●●●●
●
●
●●●●
●
●●●●
●●●
●●●●●●●
●
●
●●
●●●
●
●
●
●
●●●●
●
●
●●●
●
●●●●●●●●●●
●
●●●
●●●●
●●
●●●●
●
●●●●●
●●
●
●●●●●●
●
●●●●
●
●●●●●
●●●
●●●●
●
●●●●
●●
●●
●
●●
●●●●
●●●●
●●
●●
●
●
●
●
●
●
●●●●●●●●
●
●
●●●●
●
●
●●●●●
●
●●
●
●
●
●●
●●●
●●
●●●
●
●●●●●●●
●●●●●●●●●
●●
●
●●●
●●●
●●
●
●●●●●●●●
●
●●●●●
●
●
●●●
●
●●
●
●●
●●
●
●
●
●
●●●●●●●
●
●●●
●●
●●●●
●
●
●●
●●
●●
●●●●
●●
●●
●●
●●
●
●
●
●●●
●
●●●
●
●●●
●
●
●●●●●●●●●
●
●
●●●
●
●
●●●
●●
●●●
●●●●●●
●●●●●
●
●●
●●●
●
●
●●●●●●
●●●
●●
●●
●●●
●●
●●●●●●●
●
●●
●●
●
●
●
●●●●
●
●
●
●●●
●●
●
●●●●
●
●
●●●
●
●
●
●●●
●
●
●
●●
●●●●
●●●●●●
●
●
●
●
●●●●
●
●●●
●●
●
●●
●
●●
●●
●
●
●●
●
●●
●●●
●●
●
●●
●●
●●
●
●●●●●
●●●●●●●●●●●
●●●
●●●●●
●
●
●
●
●●
●
●
●
●●●●●
●
●●
●●
●
●●
●
●●●●●
●
●●
●●
●●●●
●
●●
●
●●●●●
●
●●
●●
●●●
●
●●
●
●●●●●
●●●●●
●●
●
●
●●●
●
●
●●●●
●●●●
●●●
●
●
●
●●●
●
●●
●●
●
●●
●
●●●
●
●●
●●
●
●●●
●●
●●●●●
●●
●
●●●●
●
●●●●●●
●●
●●
●
●●
●●●
●
●●●
●
●
●●●
●●●
●●●●
●
●●●●
●●●●●
●
●
●●
●
●
●●●
●
●
●
●
●
●●●●●●●●●
●●
●
●
●
●
●
●●●●
●
●
●●
●●●●●
●
●
●
●
●●
●●●●●●●
●
●●●●●
●
●
●
●
●
●●●
●
●●●●●
●●
●●●●
●
●
●
●●●
●
●●
●
●
●●
●●
●
●●●●
●
●
●
●●●●
●●
●
●●●●●
●●●●●●●●●
●●
●
●●●
●
●
●●
●
●
●●
●●●●●●●
●
●●
●●●
●
●●
●●●●
●
●
●●●
●●
●●●
●●●●
●●
●●
●●
●
●
●●●
●●●●
●●
●●●●●●
●
●
●
●●●●●●
●
●●
●
●●
●●●●●
●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●●
●
●
●●●
●●●●
●
●
●
●
●
●
●
●
●
●
●●●●●●
●
●
●●
●
●
●
●●
●●
●●●
●●●●
●●
●●●
●
●
●●
●
●
●●
●
●●
●
●
●
●
●●
●
●
●
●
●●●
●
●●
●●●●●
●●
●
●
●
●
●
●●
●
●●
●
●●●
●
●●
●
●●
●
●●
●
●
●
●●
●●
●
●
●●
●●
●●
●
●
●
●●
●
●
●
●
●●●
●●
●●
●●
●●
●
●●
●●
●
●
●
●
●
●●
●●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●●●●
●●●●
●
●
●●●
●
●
●
●
●●
●
●●●
●
●●●
●
●●
●●●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●●
●●
●
●●●●
●
●
●●
●
●●●
●●
●●●
●
●●
●●
●●
●
●●●●
●●
●
●
●●
●●
●
●●●
●●
●
●
●
●●●●
●●
●
●
●
●
●
●
●
●
●●
●●
●●
●
●●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●●
●●
●
●
●●●
●
●
●
●●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●●●●●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●●
●●●●
●
●●●●●
●
●
●
●
●
●
●
●
●
●●
●●
●●●●
●
●●●
●●
●
●
●
●
●●●
●
●
●
●
●●
●
●●
●
●●
●●
●
●
●
●
●
●
●
●
●
●●●●●
●●
●
●
●
●●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●●●●
●
●
●
●
●
●
●
●●●●
●
●
●
●
●
●
●●
●
●
●
●●●
●●
●●
●
●●
●
●
●
●●
●
●
●●
●●
●
●●●
●
●
●
●
●●
●
●
●
●●●
●
●
●
●
●●●●
●
●
●
●
●
●●
●
●●
●●
●
●
●
●
●●
●●●
●
●
●
●
●●
●●●
●●
●
●●●
●
●
●
●
●
●●●
●●
●
●
●
●
●
●
●●
●
●
●
●
●●
●●
●
●●
●
●
●
●
●
●
●
●●
●
●●
●
●●
●
●●
●
●●
●
●
●
●
●
●●
●●●●●
●
●●●
●
●●
●
●●
●●●
●
●●●
●
●●
●●
●
●
●
●
●
●
●●
●
●●●
●
●
●●
●
●●●●
●
●●●●●
●●
●
●●
●
●
●
●
●
●
●
●
●
●●●●
●●
●
●●
●
●●●
●●
●
●●
●●
●●
●
●
●●
●
●
●
●
●●
●●
●
●
●●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●●
●●●
●
●
●
●●
●
●●●
●
●
●
●
●
●
●
●●●
●●
●
●
●
●●●●
●
●
●●
●●
●
●●●
●
●
●●
●●●●
●●
●
●●
●●
●●
●●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●●●
●
●
●
●
●●●●●
●
●
●●
●
●
●●
●
●●●●
●
●●
●●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●●●
●●●
●
●●
●
●
●
●
●
●
●●●●
●
●
●
●●
●
●●●
●●●●●
●
●●
●
●
●
●●
●●
●●
●
●
●●
●●
●●
●
●●
●
●●
●
●●
●●
●
●
●
●
●
●●●●●
●●
●
●
●
●
●
●●
●●
●
●●
●●●●
●
●
●
●
●
●●
●●
●
●
●
●●
●
●●●
●
●●
●●●
●●
●●●
●
●●●
●●
●
●
●
●
●●
●●●
●●●●●●●
●
●
●
●●
●
●
●●●●
●
●●●●
●
●
●
●
●
●
●
●●
●
●
●●●
●
●
●●
●
●
●
●
●●
●
●●
●
●●●
●●
●
●
●●●
●
●
●
●●●●
●
●●
●
●
●
●
●●●●
●
●
●●
●●
●
●●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●●●
●●
●●
●
●●
●
●
●●●●
●●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●●
●
●
●
●
●
●●●●
●
●
●
●●●
●●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●●
●
●
●
●
●●
●
●
●●
●●
●●
●●
●
●●●
●
●
●
●●
●●●
●
●
●●●●
●
●
●
●●
●
●●
●●
●●●
●●
●
●●
●●
●
●●
●
●
●●
●●●●
●●
●
●●●●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●●
●
●●
●
●
●●●
●●●●●
●
●●
●●
●
●●
●
●
●
●
●●●●
●
●
●●
●●●
●
●●
●
●
●
●
●
●
●●
●
●
●
●●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●●●
●
●
●
●
●●●
●
●
●
●
●
●●
●
●●
●
●●●●
●
●
●
●●
●
●●●
●
●●
●
●●
●●
●
●●
●
●
●
●
●●
●●
●
●●
●
●
●●●
●
●
●
●
●
●
●
●●●●
●
●
●
●●
●
●
●
●●●
●
●
●●●
●
●
●●●●●
●
●
●
●●●●
●●
●
●
●
●
●●●●
●
●
●
●
●
●
●●
●
●
●
●●
●
●●●
●●
●
●●
●●
●●
●
●●
●
●
●
●
●
●●
●●
●●●
●●
●
●
●●
●
●●
●●
●●
●●●
●
●
●
●●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●●
●
●
●●●
●
●
●●
●
●
●●●
●●●
●
●
●
●
●●
●●●
●
●
●●●
●
●●●
●
●
●
●●
●●●
●
●●
●
●
●
●
●
●
●
●
●●●●●●●
●
●
●
●
●
●●
●
●
●
●
●●
●
●●●
●
●
●
●●
●●
●
●
●
●●
●
●
●
●●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●
●●
●
●
●
●●
●●
●
●●
●
●
●●●
●
●
●
●
●●●●
●
●●
●●
●●
●
●●●
●
●
●
●●
●●●●
●
●●
●●
●
●
●●●●●●
●
●
●●
●
●●
●
●
●
●
●●
●
●
●●
●
●●●
●
●
●●●●
●●●
●
●●●●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●●●
●
●●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●●
●●●●
●
●
●
●
●●●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●●●
●●●
●
●
●
●●
●●
●
●●●
●●
●
●
●
●●
●
●
●
●●●
●
●
●●●●
●
●
●●
●
●
●
●
●
●●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●●●●●
●
●
●●
●
●●
●
●
●
●
●
●●
●●●●
●●
●
●
●●
●●
●
●●
●
●●●
●●
●●
●
●
●
●
●●
●
●
●
●
●●
●
●●
●●●
●●
●
●●
●
●●
●
●
●●●●
●●
●
●
●●
●
●
●
●●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●●●●
●
●●●
●
●●
●
●
●
●
●
●●
●●
●●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●●
●
●●
●
●
●
●●
●●
●
●●
●
●●●●●
●●●
●
●●
●
●
●●
●●
●
●●●
●
●
●
●●
●
●
●
●
●●
●●●●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●●●
●●
●
●
●●●
●
●●
●
●●
●
●●●●●●●●
●
●●●
●
●
●
●●
●
●
●●
●●●●
●
●
●
●
●
●
●
●
●●
●
●●●
●
●●
●●
●
●●●
●
●
●●●
●
●●
●●
●
●
●●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●●
●●
●
●●●
●●●
●●●
●
●●
●
●●
●●
●
●
●
●●●●
●●●
●●●
●
●
●●
●
●
●●●
●
●●
●●
●
●
●
●●
●
●●
●●
●
●
●●
●
●●
●●
●
●●●●●
●
●
●
●●
●
●●
●
●
●●
●●●
●
●
●●●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●●
●
●●
●●
●●
●●
●
●
●
●●
●●
●●●
●●●●
●●●●
●
●
●
●
●●●
●
●
●●
●
●
●●
●
●
●
●
●
●●●●
●
●●
●●
●●●
●
●
●
●
●
●
●
●
●●
●
●●
●●
●
●
●
●
●
●
●
●
●●
●
●●
●
●●
●
●●
●
●
●●●
●
●
●
●
●
●
●
●
●
●●●
●●●●●
●●●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●●●
●
●●●●●
●●
●●●
●●●●
●●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●●
●
●
●
●
●
●
●●●
●
●●
●
●
●
●
●
●
●
●
●●●
●●
●●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●●
●
●●●●●
●●
●
●●
●●●
●●●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●●
●
●●
●
●●
●●●●
●
●
●
●
●
●●●
●●●
●●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●●●
●
●●
●●
●
●
●●
●
●●●
●
●●
●
●●
●●
●
●
●
●
●
●●
●●
●
●●
●
●
●
●
●●●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●●
●
●
●●●
●●●●
●●
●
●●
●●●
●
●
●
●
●
●
●
●
●
●●●
●
●
●●
●●●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●●
●
●●
●
●●●●
●
●
●
●●●
●●●●●●●
●●●●
●●●●●●
●
●●●●
●
●
●
●●●
●●
●●
●
●
●
●●
●●●
●
●●
●
●
●
●
●●
●
●
●
●
●
●●
●●
●●●
●●
●
●●
●
●●
●
●
●
●●●●
●●
●●
●
●
●●
●●●●
●
●●●
●
●
●●
●●
●
●
●
●
●
●●
●
●
●
●
●●
●
●●●●
●●
●●
●●
●●
●●
●
●
●●
●●●
●
●
●
●
●
●
●
●●●
●
●●
●●
●
●
●●●
●
●●
●
●
●
●
●
●
●
●
●●●
●
●
●●
●
●
●
●●
●●
●●●
●●●
●
●●●●●
●●
●
●
●
●●
●
●
●●●●●●●
●
●●
●●
●
●
●
●
●
●
●●
●
●
●●
●●
●●
●●●
●●●
●
●●●
●
●●
●●●●●●●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●●
●
●
●
●
●●●
●
●
●
●
●
●
●●
●●
●
●
●
●
●●
●●
●●
●
●●●
●●
●●●
●
●
●
●
●●●
●
●
●
●●●●●●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●●●
●●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●●
●
●●
●
●
●●●
●●●
●
●
●●●
●
●
●●●●●●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●●●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●
●●
●●●●
●●
●●●
●●●
●
●
●
●
●
●
●
●●●
●
●●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●●●
●
●●●
●●
●
●●
●●
●
●●
●
●
●●●
●
●●●
●●
●●
●●●
●
●●
●●●●
●
●
●●
●
●
●●●
●
●
●
●
●
●
●
●
●
●●●
●●
●
●
●●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●●
●●●●●
●
●
●
●
●
●
●●
●
●
●●
●
●●●●●
●●●
●●●●
●
●
●●●
●
●●
●
●●●
●●
●●
●
●●●
●
●
●
●●
●
●
●●
●●●
●●
●●●
●●
●
●
●●
●
●
●
●
●●●
●●
●
●
●●
●
●
●●
●●●
●
●
●
●
●●●●
●●
●
●
●●
●
●
●
●
●
●●●
●
●●●
●
●
●●
●●●
●●
●
●
●
●
●
●
●●
●●
●
●
●
●●●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●●●●●
●
●●
●●●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●●
●
●
●
●●●
●
●
●
●
●●
●
●
●
●●●
●
●
●
●●
●●
●●
●●
●
●
●
●
●●
●
●
●
●●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●●
●
●●●●
●
●●●
●
●
●
●
●
●●●●●
●
●●
●●
●
●●
●●
●
●
●
●
●●
●●
●
●
●
●
●
●
●●
●
●●●
●●●●
●●
●
●
●
●●
●●●●●
●
●
●●
●
●
●●
●●●
●
●
●
●
●●
●
●
●
●●●
●●
●
●
●
●●
●
●●●
●
●
●
●
●●
●
●
●●
●
●●●●
●●●●●
●
●
●
●
●●
●
●●
●
●●●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●●●●
●
●
●
●
●
●●
●●
●
●
●
●●●
●
●
●
●
●
●
●
●
●●
●
●●●●
●●
●
●●
●
●
●●
●●
●●●●
●
●
●●
●●●
●
●
●
●
●
●
●●
●●
●●
●●
●
●
●
●
●
●●
●
●●
●
●●●
●●
●
●●
●
●
●
●
●●
●●
●
●
●
●●●
●●
●
●●
●●
●●
●
●
●
●●
●
●
●●
●
●
●
●●
●●
●
●
●
●
●●●
●●●
●
●
●
●
●
●
●●
●
●●
●●
●
●
●●
●●
●
●
●
●
●
●
●
●●
●
●●
●●
●●
●
●●
●●
●●
●●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●●●
●
●
●
●
●
●●
●●●●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●●●
●
●
●●
●
●
●●
●●
●
●●
●
●●
●
●●●
●
●
●
●
●
●
●●●
●●●
●
●
●●●
●
●
●
●●
●
●●●
●●
●●
●
●
●●●
●●
●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●●
●●
●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●●
●
●
●
●
●
●●
●●
●
●
●
●●
●●
●
●
●
●
●
●
●●●
●
●
●
●
●
●●●●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●●
●
●●
●
●●
●●
●
●
●
●●
●
●
●●
●
●
●
●
●●
●
●●
●●
●●
●
●
●
●
●●
●
●●
●
●●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●●●
●●
●●●
●●
●●
●
●
●
●
●●
●
●●
●
●●
●
●
●
●●
●
●
●
●●
●●
●●
●●●●
●
●
●
●
●●
●●
●
●
●●
●
●
●●●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●●
●
●●●●
●
●
●
●
●
●
●
●
●●●●●
●●●
●●●●●●●
●●
●●●
●●
●
●
●●
●
●
●
●●●●
●
●
●
●
●
●●●
●
●
●●●
●
●●●●
●
●
●●
●●
●
●
●●
●
●
●●
●●
●
●
●
●
●
●
●●
●
●
●
●●●
●●
●
●●
●
●●●
●
●
●
●
●●●●●
●
●
●
●●
●
●●●
●
●
●
●●
●●
●●●●●
●
●●
●
●
●●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●●●●●
●
●
●
●
●
●
●●●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●●●
●
●
●
●
●
●●
●
●
●
●●
●
●●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●●●●
●
●●
●
●●
●
●
●
●●●
●
●
●
●
●●
●
●
●
●
●
●
●
●●●
●●
●●
●
●●
●
●●
●●
●●●●●●●
●●
●●
●
●●●●
●
●●●●
●
●
●
●
●
●●
●
●●●●
●
●●●
●●●●
●●●
●
●
●
●
●●
●
●
●
●
●
●●●
●
●
●
●
●
●
●
●●●
●●
●
●
●●
●
●●
●
●
●
●
●
●
●●
●●●●
●●
●
●●
●
●
●
●
●
●●
●●
●
●
●●
●
●
●
●●
●
●●
●●
●
●
●
●
●●
●
●
●●
●
●
●●
●●
●
●
●●
●
●●●
●
●●●●
●
●●●
●●
●●
●
●
●
●●
●
●
●●
●
●
●●
●●●●●
●●
●
●
●●
●●●●●
●●
●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●●●
●
●
●
●●
●
●
●
●
●
●●●
●
●
●
●●
●●
●
●●
●
●
●●
●
●
●●
●
●
●
●
●
●●●
●
●
●
●●●
●
●
●
●
●●
●
●●
●
●
●●
●
●
●
●
●
●
●●
●
●
●●●●
●
●
●●
●
●●●
●
●●●●●
●●
●
●
●
●
●
●
●●
●●
●
●
●
●
●●
●
●
●●●●
●
●
●●●●
●
●
●
●●
●●●
●
●
●
●●●●
●
●
●
●
●
●
●●
●
●●
●●
●
●
●
●
●●
●
●
●
●●●●
●●
●
●●●
●●●
●
●
●
●
●
●●
●
●●
●
●●●
●●●●●
●
●
●
●
●
●
●
●
●●
●
●●●
●
●
●
●
●●●
●
●
●
●●●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●●●●●
●
●
●
●
●●●
●●
●●
●
●
●
●
●
●
●●
●
●
●●●
●
●●
●●
●
●●
●
●
●
●
●
●
●●
●
●
●●●
●
●●
●
●
●
●
●
●
●●
●
●●
●
●
●●
●
●●
●●
●●●
●●●
●●
●
●
●●●●
●
●●●
●
●●●●●
●
●
●
●
●●
●
●
●●●●●
●●
●●●
●
●
●
●
●
●
●
●
●
●
●●●
●
●●
●●
●
●
●●●
●
●
●●
●
●
●
●
●●
●●●
●
●
●
●●●
●●●
●●
●●●
●
●
●●
●
●●●
●
●●●
●●●●●●
●
●
●●
●●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●●●
●
●
●●
●●
●●●●●
●
●●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●●●●●
●
●
●
●
●●●
●●
●●
●●●●
●
●
●
●
●
●
●
●
●●
●●
●●
●
●
●
●
●
●●
●
●
●
●
●●●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●
●●●
●●●
●●
●
●
●
●
●
●
●
●●
●
●●
●
●
●●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●
●●
●●●●
●●
●
●
●
●
●
●
●●●
●●
●
●
●
●
●●
●
●
●
●●●●
●●●
●●
●
●
●
●●●
●
●●
●
●
●
●
●
●
●
●
●●●
●
●
●●
●●
●●
●
●
●●
●
●●●
●
●
●
●
●
●●
●
●
●
●
●●
●●
●●
●●
●●●
●
●
●
●
●
●
●
●
●
●
●
●●●
●●
●
●
●●
●
●●●●●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●●
●
●
●
●
●●●
●
●
●
●
●
●●
●
●●
●
●
●●●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●●
●
●●
●●●
●
●
●●●
●●
●
●●
●
●
●
●●
●
●
●
●
●●●●
●
●
●
●
●●●
●
●●●
●
●
●
●
●
●●
●
●
●
●
●●
●
●●
●
●●●●
●●
●●
●
●
●●
●●●
●
●
●
●●●
●
●●
●
●
●
●
●
●
●●●●
●●●
●
●
●
●
●
●
●
●●●
●
●
●
●
●
●●●
●
●●●●●●
●
●
●
●
●
●
●
●●
●●
●●●
●
●●
●
●
●
●●
●●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●●
●●
●●●●
●●
●●
●●
●
●
●
●●
●●
●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●●
●
●
●●
●
●●●
●
●●
●
●●●
●
●●
●
●
●
●
●●
●
●
●
●
●●
●
●●●
●●
●●
●
●
●
●
●
●●
●●
●
●●●
●
●
●
●
●●
●
●
●
●●
●
●●
●
●●
●
●
●
●
●
●●
●
●
●
●●●●
●●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●●●●
●
●●●
●●●
●
●●●
●
●
●
●
●●
●
●●●
●
●
●●
●
●●●●●
●
●●
●●●
●●●
●●
●
●●
●
●
●●●
●
●
●●
●●●
●●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●●●
●
●
●
●●
●●
●
●
●
●●
●
●
●
●
●
●
●●
●
●●
●●●●
●●
●
●●●
●●●
●
●●
●
●
●
●●
●
●●
●●●
●
●
●
●●
●●
●
●
●
●
●●
●●
●
●●
●
●
●
●
●●●●●●●
●
●●●
●
●
●
●
●
●●
●●
●
●
●
●●●
●
●
●
●●●●
●●●
●●
●
●●
●●●
●●●●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●●
●●
●
●
●●
●
●
●
●●
●●
●
●●
●
●●
●●
●●●
●
●●
●●
●
●
●●
●●
●
●●
●
●
●
●●●●
●
●
●
●
●
●
●●
●●
●
●●●
●
●●
●
●
●●●●
●
●
●
●
●●●
●
●
●
●●●
●●
●
●
●
●●
●
●
●
●
●
●●
●●●
●●
●●●
●
●
●
●
●
●●
●
●
●●
●
●●
●
●●
●
●
●
●
●
●
●●
●
●
●
●●
●●●
●
●●
●
●●
●
●
●
●●●
●●
●
●●●
●
●●●●
●●●●●
●
●●
●
●
●●
●
●●
●
●
●●
●
●●
●●
●
●
●●●●●
●
●●
●●
●
●
●●●
●●
●
●
●●●●●●
●●●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●●
●●
●
●
●
●●
●
●●●
●
●●●
●●●
●
●●
●
●
●
●●●●
●
●●
●
●
●
●●
●
●
●●
●
●●●
●
●
●●
●
●
●
●
●●●●●
●●
●
●●
●
●
●●●
●
●
●●
●
●
●
●●●
●
●●
●
●
●
●●●
●●●
●
●●●●
●
●
●
●
●●●
●
●
●
●
●
●
●
●
●●●
●
●
●
●●●
●
●
●
●●
●
●
●●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●●●●●●
●
●●
●
●
●●
●
●●
●
●
●
●
●●●
●
●●
●●●
●
●
●●
●
●●
●●
●
●
●
●●
●
●●●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●
●●●●
●●
●●●
●
●
●
●
●
●
●
●
●
●
●
●●●
●●
●●
●●
●●●
●
●●●●
●●
●
●
●
●
●
●
●
●●
●
●
●●
●●
●
●●
●●●
●
●
●
●●
●●●●●●●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●●
●
●
●●
●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●
●
●
●
●
●
●●
●●
●●●
●●●
●
●
●●●
●●
●
●
●
●●●●
●●
●
●
●●
●
●
●
●●
●●●
●
●●●
●
●●
●●●
●
●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●
●
●●
●●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●●●●●
●
●
●●
●
●
●
●●
●●
●●●●●●
●
●
●
●
●
●●
●
●●
●
●
●
●●
●
●
●●
●
●●●
●
●
●●
●
●
●●
●
●●
●
●
●●●
●
●●
●
●
●●●
●
●
●
●
●
●●●
●
●●
●
●●
●
●
●●
●
●●
●●
●
●
●
●●●●
●
●
●●●
●
●
●●●
●
●
●
●●●●
●●●
●
●
●●●●
●
●
●
●●
●
●
●
●
●
●
●
●●
●
●●
●●●
●
●
●
●
●●
●
●●
●●
●●●●
●
●
●●
●●
●
●
●
●
●●●
●●
●
●
●
●
●●
●
●●
●
●
●
●
●●
●●
●
●●
●●
●
●
●
●
●●
●
●●●
●
●
●
●
●
●
●●
●
●
●
●
●●●
●●●
●●●
●
●●
●●
●
●
●●
●
●
●●
●
●●
●●
●
●
●
●●
●
●●
●
●●●
●
●
●
●
●
●
●●●●●
●●
●
●●●●
●●
●
●
●
●●●●●
●
●
●●
●
●●
●
●●●
●
●
●●
●
●
●
●
●●
●●●
●
●●●
●
●●
●●
●
●●
●
●
●
●●●
●●●●
λ10 λ90 µ10 µ900.0
0.2
0.4
0.6
0.8
1.0
Size
cor
rect
ed ra
tes
European Perissodactylaλ0 = 0.36µ0 = 0.73b10 = 138b90 = 1688µ = mm

Electronic Supplementary Materials
Mammal body mass evolution in North America and Europe over 20 million years:
similar trends generated by different processes
Shan Huang1,*, Jussi T. Eronen2, 3, Christine M. Janis4, Juha J. Saarinen2, 5, Daniele
Silvestro6, Susanne A. Fritz1,7
1Senckenberg Biodiversity & Climate Research Centre (BiK-F), Frankfurt am Main,
Germany;
2Department of Geosciences and Geography, University of Helsinki, Helsinki, Finland;
3BIOS Research Unit, Helsinki, Finland;
4Department of Ecology and Evolutionary Biology, Brown University, Providence, USA;
5Natural History Museum, London, UK;
6Department of Biological and Environmental Sciences, University of Gothenburg,
Gothenburg, Sweden;
7Institute of Ecology, Evolution and Diversity, Goethe University, Frankfurt am Main,
Germany.
*e-mail address: [email protected]

Supplementary information on the PyRate analysis
This study presents a cross-continent investigation of the temporal dynamics of species body
size in relation to the means of generation and maintenance of biodiversity over 20 million
years (myr). We analysed the fossil data in a Bayesian framework using the Python program
PyRate [1, 2]. For each order in each continental fauna, we generated 99 replicate datasets to
sample random occurrences within the geological time bins [following the procedure in 1, 3],
and with each of these 99×4 datasets, we ran 10 million iterations (excluding the additional
50,000 burn-in iterations), using Birth-Death Markov Chain Monte Carlo to sample the
parameter distributions. We modeled preservation as a stochastic Poisson process, in which
the preservation rate is heterogeneous among species [2] and may change as a function of
body size. Two parameters were needed to define the preservation process: a baseline
preservation rate ! and a correlation parameter "! determining how preservation rate
correlates with body size across lineages. To assess the temporal trend of body size evolution
for each group, we summarized the overall body size distribution in each geologic time bin
using the absolute origination and extinction times estimated by PyRate and averaged over the
99 replicates. For each group, we tested for significant trend through time based on the
Spearman’s rank-order correlations of the minimum, median, and maximum body size with
the time to Recent (at the mid-point of each time bin).
To further assess the relationship between body size and the diversification history of
each clade on each continent respectively, we used the Covar birth-death model with
temporally variable rates described in [1] (also implemented in PyRate [1]). The Covar
analysis jointly estimates two forms of heterogeneity in the origination rate # and extinction
rate $: changes through time by implementing clade-wide rate shifts through time and
variation among lineages as a function of body size. Thus, the birth-death model implemented
in our PyRate analyses inferred 1) the temporal dynamics of origination and extinction rates
(baseline rates # and $) and 2) correlation parameters, "#and "$, that describe lineage-

specific deviations of origination and extinction rates respectively from the baseline rates due
to variation in body size. Correlation parameters for preservation ("!), origination ("#), and
extinction rates ("$) were assigned a normal prior distribution with mean m = 0 (i.e. no
correlation) and precision (1/variance) = τ. We treated τ as a free parameter with a gamma
hyperprior [τ ~ Γ(2,2)] and estimated it from the data, a very important procedure allowing
definition of informative priors based on the data, rather than on an arbitrary choice. We
evaluated the correlations between body size and the various rates based on the posterior
distribution from all iterations.

Supplementary tables & figures
Table S1 The relationship between species body size and origination order within each genus,
assessed in linear mixed-effect models. In addition to the fixed effect of origination order
(significance indicated by the p-value), we included genus assignment as a random effect on
the intercept (model 1), the slope (model 2) and both (assumed uncorrelated, model 3). Only
genera with more than 1 species are included, so that the sample sizes are different from the
number of species reported in Table 1 for each assemblage. Using AIC as the model
comparison criterion, model 1 is the best model (in bold) for all cases (AIC weights shown in
parentheses) and shows no evidence for Cope’s rule as the dominant process driving body
size evolution in any of the assemblages. The only signal of Cope’s rule is seen in model 2
(DAIC = 2 log units higher than in the best model) for Artiodactyla when its two continental
assemblages were combined, but inference of the whole-clade evolutionary scenario must be
made with caution considering the biogeographic barrier between the two continents.
Models
Artiodactyla Perissodactyla
Europe (N = 161)
North America (N = 41)
Combined (N = 202)
Europe (N = 59)
North America (N = 57)
Combined (N = 119)
Model 1 AIC = 379
(0.59) p = 0.20
AIC = 113 (0.57)
p = 0.71
AIC = 487 (0.54)
p = 0.17
AIC = 78 (0.62)
p = 0.25
AIC = 113 (0.62)
p = 0.75
AIC = 196 (0.45)
p = 0.14
Model 2 AIC = 381
(0.22) p = 0.07
AIC = 115 (0.21)
p = 0.47
AIC = 489 (0.20)
p = 0.03
AIC = 80 (0.23)
p = 0.33
AIC = 114 (0.28)
p = 0.93
AIC = 196 (0.39)
p = 0.23
Model 3 AIC = 381
(0.20) p = 0.20
AIC = 115 (0.23)
p = 0.71
AIC = 488 (0.26)
p = 0.17
AIC = 81 (0.15)
p = 0.62
AIC = 116 (0.10)
p = 0.89
AIC = 198 (0.16)
p = 0.31

Figure S1 The distribution of our fossil samples in North America (left) and Europe (right),
with a color gradient indicating species body mass (occurrences of species without body mass
data are in black). Slight noises were added to the georeferenced locations of all occurrences
to illustrate multiple samples from the same or nearby sites. Detailed location information can
be found in [4], originally compiled from [5] and [6] for North American occurrences and the
NOW database [7] for European occurrences.
−120 −100 −80 −70
10
20
30
40
50
60North America
Longitude (°)
Latit
ude
(°)
●
●
●
●
●●
●●
●●
● ●●
●
●
●●
●
●●● ●●
●
●
●●
● ●●
●
●
● ●
●
●
●●●●
●●●●
●
●
●
●●●
●
●
●●
●●●●
●
●●●
●●
●
●●
●
● ●●
●●
●
●
●
●●
●
●
●
●
●●●
●
●
●●●
●
●
●
●
●●
●
●
●
●
●●●
●●●
●
●●
●
●●
●●●
●● ●●
●
●
●
●
●
●
●●●
●●●
●
● ●
●●
●
●●●
●
●
●
●
●
●●●
●
●●●
●●
●●
●
●
●
●
●
●
●●
●
●
●
●
● ●
●
●
●●●
●
●
●●
● ●
●
●
●●●
●
●
●●
●
●●
●
●
●
●●
● ●
●
●
●●●●●
●
●●
● ●
●
●
●●
●
●●
●
●●●
●
●
●●●●
●●
●
●
●●
●
●●
●
●
●●
●
●
●●
●
●●●●
●●
●
●
●●●
●
●●
●
●
●●
●●
●●
●
●●
●
●●
●●
●●
●●●
●●
●
●
●●
●●●●
● ●●
●●
● ●●●
●
●
●
●
●
●
●
●
●
●
●●
●●●●
●●
●
●
●
●●● ●
●
●
●●
●●
●●
●●
●
●
●●
●●
●●
●
●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●
●
●●
●
●
●
●●
●●
●
●
●
●●
●
●
●
●
●●
●●●
●
●
●
●●
●●
●
●●
●
●
●
● ●
●
●
● ●
●●●●
●●●
●●
●● ●
●
●
●
●
●●
●●●
●
● ●●●
● ●●●
●
●
●● ●
●
●●
● ●
●●●
●
●
●
●
●●●● ●
●
●
●
●●●
●
●
●
●●●
●
●
●●●● ●●
●
●●
● ●●●
●● ●
●
●
●●●
●●
●
●
●
●
● ●
●
●
●●●●●
●●●
●
●
●●●
●
●
●
●
●
●●
●
●●
●
●
●●●
●●●
●
●
●
●
●●●
●●
●●
●●
●
●
●
●
●
●
●●●●●
●
●
●●
●
●
●
●
●
●
●●
●
●● ●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●
●●●
●●
●
●●●
●
●●●
●●●●●●
● ●●●
●●●
●
●●
●
●
●
●
●●
●
●
● ●
●
●
●
●
●
●●
●
●
●
●
●
● ●●
●
●●
●●
●●●●●
●
●●●
●
●
●
● ●●●
●●●
●●●
●● ●●
●
●
●●
●
●●●
●●●●
●
●
●●
●● ●
●
●●●
●
●
●
●
●
●
●
●
●●
●●●●●
●●
●
●●
●
● ●
●
●●● ●●
●●
●
●
● ●●
●
●
●
●●
●
●●
●
●
●
●●
●●●●●●
●●●
●
● ●
●
●
●●
●●
●●●●
●
●
●
●●
●●●●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●●
●●●
●
●
●●
●
●
●
●●
●
●
●
●
●
●
●●●● ●
●
●
●●
●
●●●
●●
●●●
●●
●●
●●
●
●
●● ●●
●●●
●
●
●
●
● ●●●●
●
●●
●●
● ●
●
●●●● ●
●●
●
●●
●
●
●
●
●
●
●● ●
●
●
●●
●●
●
●
●
●
●
●
● ●
●
●
●
●●
●
●
●
● ●
●
●● ●
●
●●
●●●●●
●
●
●●
●●●
●
●●
●
●
●●
●●●
●●
●●●
●
●
●
●
●
●
●●●●●●
●●●● ●●●
● ●●
●●
● ●
●
●
●
●
●●
●●
●
●
●
●
●●
●●
●●
●
●●●
●● ●●●
●●●
●●
●
●
●●
●
●●●
●●
●
●
●
●●●
●●
●
● ●
●●●
●
●
●●
●●●
●●
●
●
●
●
●
●●
●
●●●
●
●
●●
●
●
●●
●
●
●
●
●
●
● ●
●●
●●
●
●
●
●
●●
●●●
●
●
●
●●
●
●●
●
●●●● ●● ●●●
●
●
●●
● ●
●●
●
●●
●●
●●
●●
●
●
●
●
● ●●●
●
●●●
●●
●●
●●
●●●●
●●
●
●●
●
●●
●●
●
●●
●
●
●
●●
●●
●
●
●
●
●●●
●
●●
●●●●
●
●
●
●
●●
●
●
●●●●●● ●
●●●●
●
●●●
●●
●
●
●●●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●●
●
●
●●
●
●
●●●
●
●
●●
●●●
●
●●●●
●
●
●
●●●●
●●●●
●●
●●●
●●
●●
●
●
●
●
●
●
● ●●
●
● ●
●
●●
●●
● ●●
●
●
●
●●●
●●●
●●
●
●●●●
● ●●
●
●●●
●
●●
●
●●
●●
●●●
●
●
●
●
●●
●
● ●
● ●
●
●●
●
●
●
●
●●
●
●
● ●
●
●● ●
●●
●
●
●
●
●
●●
●
●●●
●
●●●● ●
●
●● ●
●
●●
●●
●
●
●●
●
●
●●●
●
●●
●
●
● ●
●●● ●● ●
●
●●●
●
●●●
●●●
●
●
●●
●
●●
●
●●●●
●
●
●●
●
●
●
●
●
●●●
●●●
●●●
●●●
●●
●●
●
●
●●●●●
●● ●●
●●
●
●●
●●
●
●●●
●
● ●●
●
●● ●
●●●
●●●
●
●
●
●
●
●●●
●●●●●
●
●
●
●
●
●● ●●
●●
● ●
●●
●
●
●
● ●
●●●
●●
●●●●●●
●●● ●
●
●
●
●●
●●
●
●●●
●●
●●
●
●
●
●●●●
● ●● ●●
●
●
●
● ●
●
●●●●
●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●●
●●
●
●
●●●
●●
●●
●
●●
● ●●
●
●●
●
●
●
●
● ●●
●
●●●
●●●
●●
●
●
●
●
●
● ●
●●●●●●●●●
●
●●
●●●
●●
●●
●
●
●
●
●
●●●
●
●●
●●● ●
●
● ● ●
●● ●
●
●●●
●● ●
●
●●●
●●●●
●
●●●
●
●
●
●
●
●●
●●
●
●
●
●
●
●●
●
●●
●
●
●●●
●
●
●
●● ●
●●
●●●●●
●●
●
●● ●
●
●●●
●● ●
●●
●● ●
●
●●
●●
●
●
●
●●
●●●
●
●
●
●● ●
●
●
●●●
●
●
●●
●
● ●
●
●●
●
●
●●
●
●
●
●
●
●●
●
●
●●●
●●
● ●
●
●●
●
●●
●
●
●
●●
●
●
●
●
●
●●
●
●
●●
●
●
●
●●
●
●
●
● ●
●●
●●●
●
●●
●●
●
●
●
●● ●
●●
●
●●●●
●●●●
●
●
●
●●
●●
●
●
●●●●
●
●
●
●●
●
●●
●
●
●
●
●
●
●●
●●●
●●
●●●●●
●● ●
●
●●●●
●
●●
●●●
●●
●
●
●●
●
●
●●
●
●
●
●●
●
●●
●
●
●
● ●
●
●
●●
●
●●●
●●
●●●●●●
●●
●
●●
●●●
●
●
●●●
●●●
●
●
●
●●●
● ●
●
●
●●
●●
●
●
●●
●
●
●
●
●● ●
●●●
●●●
●
●
●
● ●
●●
●
●●
●●
●●
●
●
●●
●●●
●●● ●●
●
●
●●
●●
●●
●●●
●●
●●
●●●●
●●
●
●
● ●
●
●●
●
●●
●
●
●●
●
●
●
●
●
●
●
●
●●
●
●●●●●
● ● ●●●
●
●
● ●● ● ●
●
●
●●
●●●● ●
●
●
●
●
●●
●
●
●
●
●
−10 0 10 20 30 40 50
30
35
40
45
50
55
Europe
Longitude (°)
Latit
ude
(°)
●
●
●
●
●
●
●●●●●●
●
●●
●
●
●
●
●
●
●
● ●● ●
●●
●
●●
●●
●●●●●●●●
●
●
●
● ●●●●● ●
●●
●
●
●
●●
●
●
●●
●
●
●●
●●● ●
●
●
●●
●●
●●
●●
●
●●
●
●●
●●●
●
●
●
●
●
●
●●
●
● ●
●
●●
●●
●
●
●
●
●
● ●
●●
●
●●● ●●●
●
●●●
●●
●
●
●● ●●●●●
●● ●
● ●●●
●
●
●●●
●●
●●●●
●
●
●●
●
●
●
●●
●●
●
●●● ●●
●●●●
●●●●●●●●●●
●●●●●
●
●
●
●●
●
●●●
●●●
●
●
●
●●●●
●●●●
●
●
●
● ●●
● ●●
●
●
●●
●●
●
●●●
● ●●
●
●
●●●
●
●●●
●●
●●●●
●
●
●●
●
●●
●●●
●●●●●●●●●●●●
●●●●●●
●●
●● ●● ●●
● ●
●●
●●●
●●
●●
●●●●●●
●●●●●●● ●
●● ●●●●●●
●
●●
●●●●
●
●
●●●●●●
●
●● ●
●
●
●●
●
●●
●●●●●●
●● ● ●● ●●
● ●●
●●●
●
●●
●●
●●
●●● ●●●
●
●●●●●●
●
●●
●
●
●
●
●
●
●●●●●●●●
●●
●
●
●●
●●
●●
●
●
●● ●
●
●●●
●
●
●●●●●●●
●●
●●
● ●
●
●●
●
●
●
●
●●
●
●
●●●
●●●●●●
●
●●●●●●●●●
●
●●
●●
●
●
●●●●●●●●●●●●●●●●
●●
●●●
●●●
● ●●
●●●
●●●
●
●●
●
●
●●
●●
●●
●●
●
●
●
●●● ●●●
●
●
● ● ●
●
●
●●
●
● ●●●●
●●
●
●
●
●
●
●●●
● ●●
●
●
●●●●●
●
● ●●
●●
●●●●●●
● ● ●
●
●●
●
●
●●
●
●●
●●●●●●
●●●●●●
●
●
●
●
●
●●
●●
●●
●
●●
●●
●●
●
●●●
●●●●
●●●
●●
●● ●●●●
●●●●●●●
●●
●●
●●●●●●●
●●●
●●●●●● ● ●● ●
●●
●
●●●
●●●●● ●
●●
●
●●
●
●
●●●●●
●●
● ●
●
●
●
●●●●●●
●
●
●●
●●
●●
●
●●●
●●
●●●●●●
●
●●●●●●●
●●
●●●
●
●
●●
● ●●●●
●●●●●
●●●
●
●
●
●
●●
●
●
●
●
●
●●
●●
●●●●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●●●●
●●●●●
●
●●●
●●●● ●
●●●●●
●●
●●●●●
● ●●●
●
●●●
●
●●●
●●●●
●
●●
●
●
●
●
●
●
●
●●
●
●
●●●●
●●
●●●●●●
●●
●
●●
●
●
●
●
●
●
●●
●
●
●
●●● ●
●
●●
●
●●
● ● ●●
●●
●
●●
●
●
●
●●
●
●●●
●●●
●●
●
●●
●
●
●
●●
●●
●
● ●
●●●
●●
●●●
●●●
●
●
●
●● ●
●
●●
●●
●●
●
●
●
●●
●
●
● ●●●
●
●
●●
●
● ●●
●
●●
●
●●●●
●●
●
●
●●●
●
●
●●●●●●●● ●
●●●●
●
●●●●
●●
●●
●
●
●
●
●●
●
●●●●●●●●●●
●●
●●
●●
●
●●
●
●
●
●
●●● ●
●●●
●●
●●
●
●●
●
●●
●
●
●●●
●
●
●●
●●●
●●
●
●●
●●
●
●
●●●●●●●
●●●●
●
●
●●●
●
●
●●
●
●●
●
● ●●●
●●●●●●
●
●
●
●●●
●
●
●
●
●●
●
●●
●
●●
●
●●
●●
●●●
●
●●
●
●
●
●●●
●
●●
●●
●●●●
●●●●●●●●●●●●
●
●●
●
●●
●●●
●
●
●
●
●
●
●
●●
●
●●
●●●
●
●●●●●
●●●●●
●●
●●●●●●●●●
●●●●●●●
●●
●
●
●●
●
●●
●
●
●
●●
●
●●
●
●
●
●●
●●●●●
●
●
● ●●
●●
●●●
●
●● ●●
●
● ●● ●
●●●
●●
●
●
●●
●●
●
●
●
●
●●●●●
●
●
●
●●●
●
● ● ●
●●
●
●
●●●
●●●
●
●●
●
●●●
●
●
●●
●
●●●●●
●
●
●
●
●
●●
●
●
●
●
●
●
●
●●●●
●
●●●●●
●●●●●●
●
●●
●●●
●●●●
●●●●●●●●
●●●
●●
●●●●
●●●● ●
●●●●
●●
●
●
●●●
●
●●
●
●●
●
●
●
●
● ●●
●
●●
●●● ●●●●
●
●
● ●●
●●
●
●●
●
●●
●● ●
●●
●
●
●
●● ●●
●
●
●●●
●●
●
●●●
●
●●●●
●
●
●● ●
●
●
● ●●●●
●
●●
●●
●
●
●
●
●
●
● ●
●●
●
●
●
●
● ●
●
●
●
●
●●
●●
●●
● ●●
● ●
●●
●
●
●
●
●
●
●●●●
●●●●●
●
●
●●
●●●
●●
●
●
●
●
●
●
●●●●
●●
●●
●
●●
●
●
●
●
●
●
●
●● ●
●
●
●●
●●
●●
●
●
●●
●
●
●
●●
●●
●
●
●
●
●
●●
●●●
●
●
●
●
●
●
● ●●
●
●
●
●
●●
●●●●
●
●
●●
●
●●
●●
●●
●
●
●
●
●
●
●
●
●
●
●●
●●●
●
●●
●●●●
●
●●
●
●
● ●
●●
●●
●●
●
●
●●
●● ●●●●●
●●●●
●
● ●●●●
●
●
●●
●●
●
●●●●
●●
●
●
●●●●
● ●
●
●●
●
●
●
●
●●
●●
●●
●
●●●●●
●
●
●
●●
●●
●
●
●
●
●●
●
● ●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●
●●
●●●●●
●
●●
●
●
●●●
●●
●●●●●
●
●
●
●●●●
●●●●
●●
●
●
●
●
●
●
●
●●
●
●
●●
●●
●
●●●
●
●
●
●●●
●●●
●
●●●●
● ●
●●●
●
●
●●●
●●●
●
●●
●● ●
●
●
●
●
●
●●
●●
●
●●
●●●● ●
●●
●
●
●
●
●
●
●●
●
●
●
●●
●
●
●
●
●●
●
●
●
●
●●
●
●
●●
●●
●
●
●
●●●●
●●●●●●●●
●●
●●●●
●●
●
●
●
●●●
●
●
●
●●
●
●●
●
●●
●●●
●
●
●
●●
●●●●
●
●●
● ●●●●●
●
●●
●
●●
●●
●
●●●●●
●
●
●●
●
●
●●●
●
●
●●●●●●●●●●
●
●
●
●
●
●
●
●
●●
●
●
●
●●●●
●●
●
●●●
●
●
●
●
●
●●●
●
●●
●●
●
●
●●
●●●
●● ●
●
●
●
●
●●
●
●
●
●
● ●
●●●●●●
●●● ●●
●
● ●
●●●
●
●
●●●
●
●● ●
●
●●
●
●
●●●
●●
●●●
●
●
●
●
●●●
●
●●
●
●●
●●
●
●
●●
●● ●●
●
●●
●●●●●●●●
●●●●
●
●
●●
●
●●●
●● ●
●
●
●●
●
●●
●●●●●●●
●●
●
●●
●
●
●
●● ●
●
●●●
●●
●●
●
●
●●
●●●
●●●●
●● ●
●●
●●
●
●
●
●
●●●
●
●
●
●
●
●
●●●●●●●●
●●
●●●●●
●
●
●●
●
●●●
●
●
●●
●
●●●●
●●●●●●
●●
●
●●●
●●
●
●
●●
●
●
●● ●
● ●●
●
●
●●
●● ●●
●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●●
●
●
●
●
●●
●
●●
●
●
●●●
●
●●●
●●●
●●
●
●
●●●
●●
●●●●●
●●●
●
●●●●●
●●●● ●
●● ● ●
●●
●●●●
●●●●
●●
●
● ●
●●●
●●
●
●
●
●
●
●●●
●●
●
●●
●
●
●
●●●●●
●●
●
●
●●
●●
●●
●●
●
●●●
●●
●
●
●
●●●●●●●●● ●
●● ● ●
●●
●
●● ●
●
● ●●
●●●●●●●
●
●●●●●●
●●
●
●
●
●●●
●
●
●●●
●
●
● ●
●
●
●● ●●●
● ●
●●
●●●●●
●●●
●
●● ●
●
●●
● ●●
●
●●●●●●●
●
●
●●●
●
●●
●●
●●
●●
●
●
●●●
●●
●●●
●●
●
●●●
●●●●
●●●●●
●
●
●●
●
●●● ●
● ●●
●
●●
●
●●●
●●
●●●●●●
●
●●●
●●
●●●
●●●●●●●●●
●●
●●●●
●
●
●●
●
●
●
●
●●
●
●
●●●●●●
●●●●
● ●
●●
●●●
●●●
● ●
●●● ●
●●
●●●●
●●●●●
●●●
●●●
●● ●●●
●
●
●
●
●
●● ●●
●●
●
●●
● ●●
●●
●
●
●
●
●●
●
●
●
●
● ●●●●
●
●
●
●
●●
●●
●
●●
●● ●●
●●
●
●
●
●●
●●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●●●●
●
●
●●
●●●●●●●
●
●●●
●● ●
●
●●●●●
●
●●●
●
●●●
●
●
●
●●
●●
●●●●●●●●
●●●●●●
●●
●
●●
●
●●
●●●●
●
●
●
●
●
●●● ●
● ●
●●
●●●
●
●
●●●
●
●
●●●
●
●●
●●
● ●
●
●●●●
●●●● ●●●
●●●●
●
●
●●
●●
●
●●●●●
●
●●
●●
●●●
●
●
●●
●●
●●
●
●●
● ●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●●
●
●●
●●●
●
●
●●
●●●●
●●
● ●
●
●●●●
●
●
●
●
●
●●
●
●
●●
●●
●●
● ●●
●●
●●●
●●●●
●
●●
●
●
●
●●
●
●
●●
●
●●
●●
●●
●●●
●
●●
●
●
●●
●
●
●
●●
●●
●
●●●●
●
●●
●
●●●●●●●
●●●
●
●
●
●
●
●
●●●●
●●●
●●
● ●
●
●
●●
●
●
●
●
●
●
●
●
●
●
●●●● ●
●
●●
●●
●
●●●
●●
● ●
●
● ●●
● ●
●
●
●●●●
●
●
●● ●●●●
●●●●●●●
●●● ● ●
●
●
●●
●●●
●●
●
● ●
●●●
●●
●
●
●
●●
●●●
●
●●●●
●
●
●●
●
●
●
●●●
●●●
●●
●
●●●●
●
●
●
●
●●
●
●
●
●●
●
●
●●
●● ●●●●●
●●
●
●●●●●● ●
●
●●●
●●
●
● ●
●●
●
●
●
●
●
●
●
●
●
●
●
●●
●●●
●
●
●●●
●
●●
●
●
●●
● ●
●●
●●
●
●●●
●
●●
●●
●●●●●●●●
●●● ●
●●
●●●
●●
●
●
●
●●●
●
●●●
●●●●
●
●●●●
●
●
●
●
●
●
●
●
●
●●
●
●●
●
●
●
●●
●
●
●●
●
●●●●●●●●
●●
●●●●
●
●●
● ●●
●
●
●
●●●
●●●
●●●
●● ●●●
●
●
●
●●
●
●
●●
●●●●●●
●
●●
●●●●●●
● ●●
●●●●
●●
●
●
●
●
●
●●
●
●●
●
●
●
●●●●●●●●
●●
●
●●
●●
●●●
●
●
●
●●
●
●●●●●
●●●
●
●
●●
●●
●●●
●●●●●
●●●
●
●
●●
●
● ●
●●
●●
●
●
●
●●●
●● ●●
●
●
●●
●
●
●
●
●
●
●
●
●
●●●●●
●
●●●●
●●●●
●●
●
●●
●
●
●●●●●●●●●●●
●●
●●●●
●●●
●●●
●●●●●●●
●●●●
●
●●●
●
●
●●● ●●● ●●●
●●● ●
●● ●●
● ●●
●
●●
●●
●
●
●
●
●
●
●●
●
●●
●●
●
●
●●●
●
●●
●
●●
●
●
●●
●●
●
●
●
●
●
●
●
●●●
●
● ●
●
● ●
●
●
●
●
●●
●
●●
●
●●
●
●
●●
●●●
●●
●
●
●
●
●
●
●
●●
●●
●
●
●●
●
●
●●●
●●
●●●●
●
●●●●●●●●●●●●
●●●●● ●● ●●
●●
●
●●
●●● ●
●●●●●●
●
●●●
●
●●●●
●●●
●
●●●
●
●
●
●
●
●●
●●
●●
●●●
●●●●
●
●
●
●●
●
●●
●
●
●
●
●
●
●●
●
●●●●
●
●●●●●
●●
●
●
●
●●
●●● ●●
●●●
●
●
●●
●
●
●●
●
●
●●
●●●●●●●●●●●
●●●●
●
●
●●
●●
●●
●
●
●●●
●● ●
●
●●
●●●
●
●●
●●
●
● ●●●
●
●
●
●
●
●
●
●
●
●●● ●●
●● ●
●●●●
●●
●●●
●●
●
●
●
●●
●
●●
●
●●
●
●
●
●●
●●●
●●●●● ●
●●●●●●
●●
●●
●●
●●●
●
●● ●
●●
●
●
●●
●
●
●
●
●●
●●●
●
●●●●
●●●●
●●
●
● ●●
●●
●●
●
●
●
●
●●
●
●
●
●●●●
●●●●
●
●●●
●
●
●●●●●
● ● ●
●●●●
●●●●
●●●
●
●
●●
●●●
●
●
●●●
●
●
●
●
●
●
●
●● ●●
●● ●●
●●
●
●● ●●
●●
●
●
●● ●
●
●
●
●
●
●
●
●●
●
●
●
●
●●
●
●●
●
●●●●
●●●
●
●●
●
●●●●●●●●
●●
●
●
●
●
●
●
●●●
●●●
●●
●●
●●
●●
●●●●
● ●●●
●● ●
●●
●
●●●
●●●
●●●
● ●
●●●
●●
●●
●
●
● ●●
●●
●●
●
●
●
●
●
●
●●
●
●●
●
●●
●
●●
●
●
●●
●●
●
●
●
●
●
●●
●
●
●
●●●
●
●
●
●●
●
●●
●●● ●
●
●
●
●●●●●
●●
●
●●●
●
●
●
●
●
●
●
●
●●
●
●●●●
●●
● ●●
●●●
●●
●●●
● ●
●
●
●●
●
●●●
●●●●
●
●
●●●
●
●●●
●●
●
●●
●●
●
●● ●
●
●
●●
●
●●
● ●
●●
●●
●
●
●
●
●
●
●●
●
●●●
●
●●
●●●
●●
●●●
●●●● ●●
● ●
●●●
●●●
●● ●
●
●
●●
●
●●●●
●
●
●
●●
●● ●●●●
●●●
●
●
●●
●●
●
●
●●
●
●
●
●● ●●●●●
●
●●●
●●●
●
●
●
●●
●
●
●●
●●
●●●●
●
●●
●●●●●
●
●●●●
●
●●
●
●
●
2.8
3.7
4.7
5.7
6.7
●NA
log 1
0 Bod
y M
ass
(g)

Figure S2 The number of species based on recorded occurrences in each time bin (sampled
data, circles with solid lines) and based on species durations between first and last
appearances in the fossil record (inferred data, crosses with dashed lines). No correlation was
found between time to Recent and the difference between the two lines (p > 0.29 in
Spearman’s rank order correlation tests for all four cases). In addition, we report the number
of singletons (triangles with dotted lines) which indicates no temporal bias of sampling effort.
●
●
●
●
●●●
●
●
●●
●●
●
●
20 15 10 50
10203040506070
Geologic time (Ma)
Num
ber o
f spe
cies Artiodactyla
●
●
●
●
●
●
●
●
●
●
●●
●
●●
20 15 10 50
20
40
60
80
100
Geologic time (Ma)
Num
ber o
f spe
cies Artiodactyla
●●
●●
●
●
●●
●
●●
●
●
●
●
20 15 10 50
10
20
30
40
50
Geologic time (Ma)
Num
ber o
f spe
cies Perissodactyla
●
●
●
●
●
●●
●
●
●●
●
●
●●
20 15 10 50
10
20
30
40
Geologic time (Ma)
Num
ber o
f spe
cies Perissodactyla
North America Europe
● Sampled Inferred Singleton
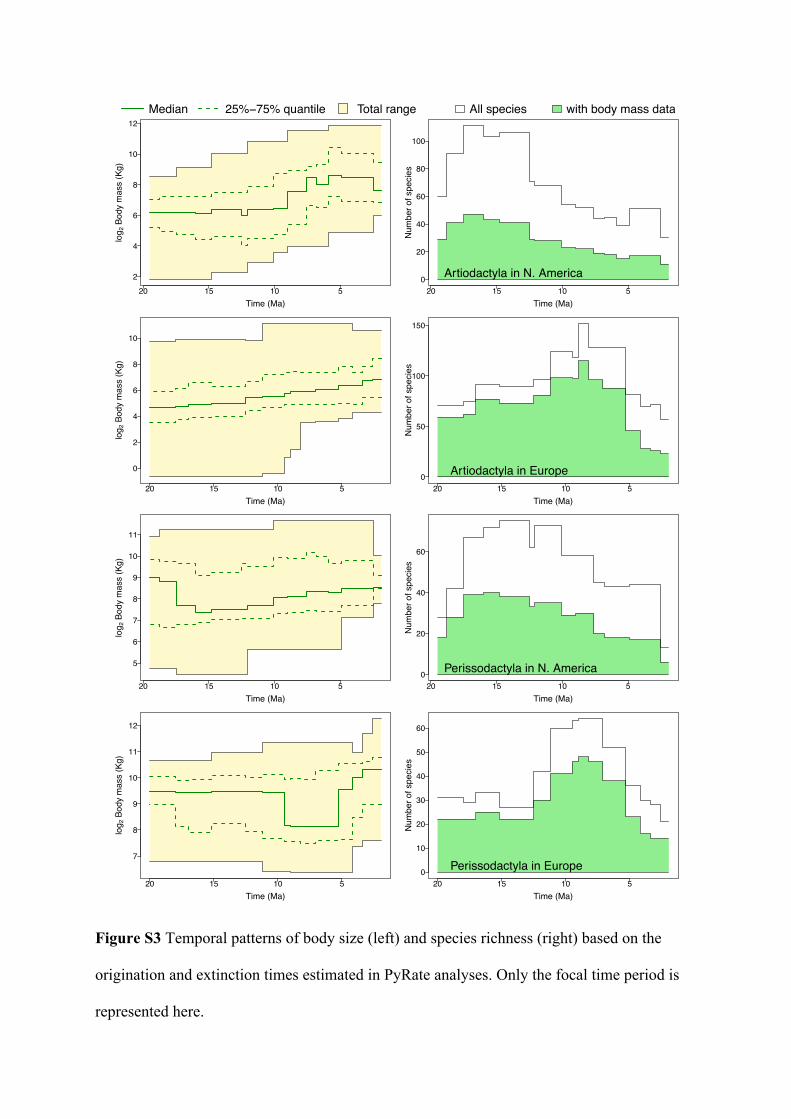
Figure S3 Temporal patterns of body size (left) and species richness (right) based on the
origination and extinction times estimated in PyRate analyses. Only the focal time period is
represented here.
20 15 10 52
4
6
8
10
12
Time (Ma)
log 2
Bod
y m
ass
(Kg)
20 15 10 50
20
40
60
80
100
Time (Ma)
Num
ber o
f spe
cies
Artiodactyla in N. America
20 15 10 5
0
2
4
6
8
10
Time (Ma)
log 2
Bod
y m
ass
(Kg)
20 15 10 50
50
100
150
Time (Ma)
Num
ber o
f spe
cies
Artiodactyla in Europe
20 15 10 5
5
6
7
8
9
10
11
Time (Ma)
log 2
Bod
y m
ass
(Kg)
20 15 10 50
20
40
60
Time (Ma)
Num
ber o
f spe
cies
Perissodactyla in N. America
20 15 10 5
7
8
9
10
11
12
Time (Ma)
log 2
Bod
y m
ass
(Kg)
20 15 10 50
10
20
30
40
50
60
Time (Ma)
Num
ber o
f spe
cies
Perissodactyla in Europe
Median 25%−75% quantile Total range All species with body mass data
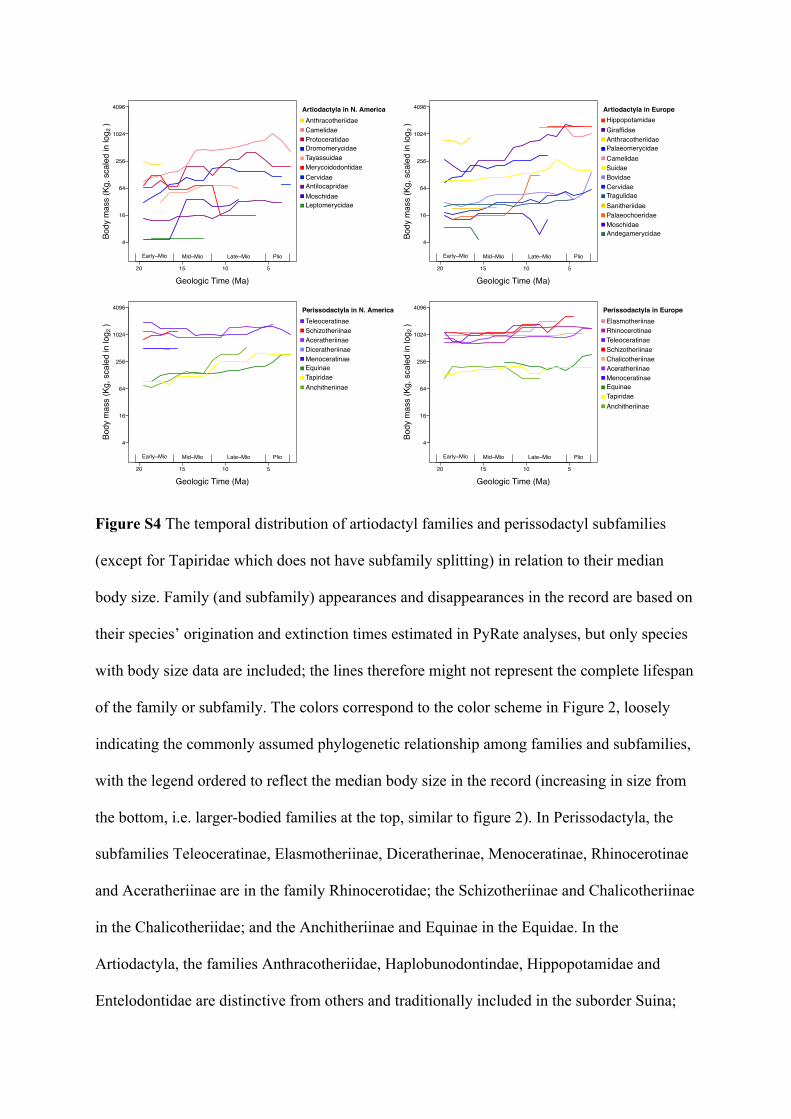
Figure S4 The temporal distribution of artiodactyl families and perissodactyl subfamilies
(except for Tapiridae which does not have subfamily splitting) in relation to their median
body size. Family (and subfamily) appearances and disappearances in the record are based on
their species’ origination and extinction times estimated in PyRate analyses, but only species
with body size data are included; the lines therefore might not represent the complete lifespan
of the family or subfamily. The colors correspond to the color scheme in Figure 2, loosely
indicating the commonly assumed phylogenetic relationship among families and subfamilies,
with the legend ordered to reflect the median body size in the record (increasing in size from
the bottom, i.e. larger-bodied families at the top, similar to figure 2). In Perissodactyla, the
subfamilies Teleoceratinae, Elasmotheriinae, Diceratherinae, Menoceratinae, Rhinocerotinae
and Aceratheriinae are in the family Rhinocerotidae; the Schizotheriinae and Chalicotheriinae
in the Chalicotheriidae; and the Anchitheriinae and Equinae in the Equidae. In the
Artiodactyla, the families Anthracotheriidae, Haplobunodontindae, Hippopotamidae and
Entelodontidae are distinctive from others and traditionally included in the suborder Suina;
20 15 10 5
Geologic Time (Ma)
Body
mas
s (K
g, s
cale
d in
log 2
)
4
16
64
256
1024
4096
PlioLate−MioMid−MioEarly−Mio
Artiodactyla in N. America
LeptomerycidaeMoschidaeAntilocapridaeCervidaeMerycoidodontidaeTayassuidaeDromomerycidaeProtoceratidaeCamelidaeAnthracotheriidae
20 15 10 5
Geologic Time (Ma)
Body
mas
s (K
g, s
cale
d in
log 2
)
4
16
64
256
1024
4096
PlioLate−MioMid−MioEarly−Mio
Artiodactyla in Europe
AndegamerycidaeMoschidaePalaeochoeridaeSanitheriidaeTragulidaeCervidaeBovidaeSuidaeCamelidaePalaeomerycidaeAnthracotheriidaeGiraffidaeHippopotamidae
20 15 10 5
Geologic Time (Ma)
Body
mas
s (K
g, s
cale
d in
log 2
)
4
16
64
256
1024
4096
PlioLate−MioMid−MioEarly−Mio
Perissodactyla in N. America
AnchitheriinaeTapiridaeEquinaeMenoceratinaeDiceratheriinaeAceratheriinaeSchizotheriinaeTeleoceratinae
20 15 10 5
Geologic Time (Ma)
Body
mas
s (K
g, s
cale
d in
log 2
)
4
16
64
256
1024
4096
PlioLate−MioMid−MioEarly−Mio
Perissodactyla in Europe
AnchitheriinaeTapiridaeEquinaeMenoceratinaeAceratheriinaeChalicotheriinaeSchizotheriinaeTeleoceratinaeRhinocerotinaeElasmotheriinae

the Tayassuidae, Suidae, Sanitheriidae, and Palaeochoeridae in the Suoidea within the Suina;
the Camelidae, Protoceratidae, Merycoidodontidae and Cainotheriidae in the Tylopoda; the
Tragulidae, Hypertragulidae, Leptomerycidae and Andegamerycidae are in the Tragulina
within the Ruminantia; and the Dromomerycidae, Gelocidae, Bovidae, Cervidae,
Antilocapridae, Moschidae, Giraffidae and Palaeomerycidae are in the Pecora within the
Ruminantia.

Figure S5 Temporal patterns of clade-wide baseline origination (#) and extinction ($) rates of
Artiodactyla (upper) and Perissodactyla (lower) in Neogene Europe (left) and North America
(right), until 3 million years ago (Ma). Solid lines represent the mean in each million-year
interval, and shaded area indicate the 95% credible interval. The PyRate analysis also allows
variation among lineages (see Supplementary information for methods), which was modeled
in relation to body size and is illustrated in Figure 4.
20 15 10 50.0
0.5
1.0
1.5
2.0
Geologic Time (Ma)
Rate
s
20 15 10 50.0
0.2
0.4
0.6
0.8
Geologic Time (Ma)
Rate
s
20 15 10 5
0.2
0.4
0.6
0.8
1.0
Geologic Time (Ma)
Rate
s
20 15 10 5
0.5
1.0
1.5
Geologic Time (Ma)
Rate
s
OriginationExtinction
North America EuropePerissodactyla
Artiodactyla

Additional references
1. Silvestro D., Salamin N., Schnitzler J. 2014 PyRate: a new program to estimate
speciation and extinction rates from incomplete fossil data. Methods in Ecology and Evolution
5(10), 1126-1131. (doi:10.1111/2041-210X.12263).
2. Silvestro D., Schnitzler J., Liow L.H., Antonelli A., Salamin N. 2014 Bayesian
estimation of speciation and extinction from incomplete fossil occurrence data. Syst Biol.
(doi:10.1093/sysbio/syu006).
3. Silvestro D., Antonelli A., Salamin N., Quental T.B. 2015 The role of clade
competition in the diversification of North American canids. Proc Natl Acad Sci USA
112(28), 8684-8689. (doi:10.1073/pnas.1502803112).
4. Fritz S.A., Eronen J.T., Schnitzler J., Hof C., Janis C.M., Mulch A., Böhning-Gaese
K., Graham C.H. 2016 Twenty-million-year relationship between mammalian diversity and
primary productivity. Proc Natl Acad Sci USA. (doi:10.1073/pnas.1602145113).
5. Janis C.M., Scott K.M., Jacos L.L. 1998 Evolution of Tertiary Mammals of North
America: Volume 1, Terrestrial Carnivores, Ungulates, and Ungulate Like Mammals.
(Cambridge, UK, Cambridge University Press.
6. Janis C.M., Gunnell G.F., Uhen M.D. 2008 Evolution of Tertiary Mammals of North
America: Volume 2, Small Mammals, Xenarthrans, and Marine Mammals. (Cambridge, UK,
Cambridge University Press.
7. Fortelius M. (coordinator) 2015 New and Old Worlds Database of Fossil Mammals
(NOW). University of Helsinki. Date of access: November 1st, 2015.