hydrogenase of alcaligenes - journal of bacteriology - american
TRANSCRIPT

JOURNAL OF BACTERIOLOGY, Oct. 1992, p. 6277-62890021-9193/92/196277-13$02.00/0Copyright C 1992, American Society for Microbiology
Vol. 174, No. 19
A Gene Complex Coding for the Membrane-BoundHydrogenase of Alcaligenes eutrophus H16
CHRISTIANE KORTLUKE,lt KARIN HORSTMANN,1 EDWARD SCHWARTZ,' MANFRED ROHDE,2tRALF BINSACK,1 AND BARBEL FRIEDRICH'*
Institut fur Pflanzenphysiologie und Mikrobiologie der Freien Universitat Berlin, D-1000 Berlin 33,1 andInstitut fir Mikrobiologie der Universitat Gottingen, D-3400 Gottingen,2 Germany
Received 13 May 1992/Accepted 22 July 1992
One of the key enzymes in the chemolithoautotrophic metabolism ofAlcaligenes eutrophus H16 is a dimeric,membrane-associated hydrogenase. The genetic determinants of this enzyme are located on the endogenousmegaplasmid pHG1 (G. Eberz, C. Hogrefe, C. Kortluke, A. Kamienski, and B. Friedrich, J. Bacteriol.168:636-641, 1986). Complementation studies showed that the information required for the formation of activemembrane-bound hydrogenase occupies more than 7.5 kb of megaplasmid DNA. We cloned and sequenced thisregion and identified the genes encoding the two hydrogenase subunits (hoxK and hoxG). The nucleotidesequence contains nine additional closely spaced open reading frames. Immunoelectron microscopy showedthat the gene product of one of these open reading frames (hoxM) is involved in the process leading to theattachment of hydrogenase to the membrane. Other open reading frames may encode additional processingfunctions and components of a hydrogenase-linked electron transport chain. Analysis of TnS-B21-mediatedtranscriptional fusions provided evidence that the structural genes and accessory functions belong to at leastthree coordinately regulated transcriptional units.
The aerobic, facultatively chemolithoautotrophic bacte-rium Alcaligenes eutrophus H16 can utilize hydrogen as itssole energy source. Hydrogen oxidation is catalyzed by twodistinct [NiFe] hydrogenases, which differ in catalytic prop-erties, cellular location, electron acceptor, cofactor content,and subunit composition (3, 18). The cytoplasmic hydroge-nase (SH) is a tetramer composed of four heterologouspolypeptides. This NAD-reducing enzyme provides reduc-ing equivalents primarily for autotrophic carbon dioxidefixation (3). The membrane-bound hydrogenase (MBH) ofA.eutrophus is a dimer consisting of two heterologous poly-peptides, the 31-kDa small subunit (SSU) and the 62-kDalarge subunit (LSU) (51). The oxidation of hydrogen cata-lyzed by the MBH is coupled to the respiratory chain andthus contributes to the generation of ATP. TheA. eutrophusMBH is a member of the more widespread family of mem-brane-associated hydrogenases, whereas the soluble NAD-reducing type has been found only in Alcaligenes andNocardia strains (3).
Previous studies on deletion and insertion mutants of A.eutrophus H16 led to the mapping of the determinants of theMBH on the 450-kb conjugative megaplasmid pHG1 (23, 28).This locus, designated hoxP, lies in a region containingseveral hydrogenase-related genes arranged in functionalgroups. The genes encoding the four subunits of the SH(hoxFUYH) (61) and adjacent accessory functions form onegroup. Another group comprises genes that encode functionsessential to the formation of both enzymes (10, 14). Adjacentto these pleiotropic functions is a gene (hoxA) encoding aresponse-regulator-type transcriptional activator (14). Ahigh-affinity nickel transporter is encoded at another site(hoxN) (13, 16).
* Corresponding author.t Present address: Institut fur Biologie II (Mikrobiologie), Uni-
versitat Freiburg, D-7800 Freiburg i. Br., Germany.t Present address: Gesellschaft fur Biotechnologische Forschung
mbH, Abteilung Mikrobiologie, D-3300 Braunschweig, Germany.
In this communication we report the cloning, nucleotidesequence, and organization of the hoxP locus, which en-codes the MBH. We present evidence that this regioncontains two structural genes and at least eight accessorygenes organized in at least three transcriptional units. Wealso describe results indicating that one of the accessorygenes is involved in the process leading to the attachment ofthe MBH enzyme to the membrane.
MATERIALS AND METHODS
Strains and plasmids. The bacterial strains, phages, andplasmids used in this study are listed in Table 1. The MBH-deletion strain HF328 was constructed via gene replacementafter manipulation of a subcloned fragment. A 6.7-kb frag-ment of the hoxP region contained in plasmid pCH312 wasexcised by cutting with BglII and replaced with a Kmrcassette prepared as a BamHI fragment, yielding plasmidCH313. A 2.3-kb fragment of pCH313 encompassing theinserted Kmr cassette was transferred to the Tcr vectorpSUP202. The resulting plasmid, pCH314, was introducedvia conjugative transfer from Escherichia coli S17-1 into A.eutrophus H16, and Kmr Tcr transconjugants were selected.Since pCH314 does not replicate inA. eutrophus, the doublyresistant transconjugants represent cointegrates arising viahomologous recombination. Homogenotes were selected asKmr Tcs segregants. The physical structure of the site of thedeletion in one of these derivatives was verified by restric-tion analysis of pHG1 DNA, and the isolate was designatedHF328.A series of plasmids carrying various segments of the
hoxP region was constructed for complementation studies.One group of plasmids was derived from the hybrid phageAAEP1, taken from a lambda library (25) of A. eutrophusH16. A partial EcoRI digest of XAEP1 was cloned into theEcoRI site of the broad-host-range plasmid pVK101 (26). Ahybrid plasmid containing a 15.2-kb insert consisting of fourEcoRI fragments (11.0, 3.5, 0.5, and 0.2 kb) was selected and
6277
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
17
Dec
embe
r 20
21 b
y 31
.170
.60.
86.

6278 KORTLUKE ET AL.
TABLE 1. Bacterial strains and plasmids used in this studya
Strain, plasmid, or phage Characteristics Source or reference
StrainsAlcaligenes eutrophusH16H20HF221HF328
MBH+ SH+MBH- SH+MBH- SH+MBH- SH+ Kmr, Kmr cassette inserted in place of a 6.7-kbsegment of the hoxP locus via gene replacement
DSM 428, ATCC 17699ATCC 1770028This study
Escherichia coliS17-1HB101WM1280
NM522K38
Tra+, recA pro thi hsdR, chr::RP4-2recA13 supE44 hsdS20 ara-14proA2 lacYl galK2 rpsL20xyl-5 mtl-lrecA his ilvA rpsL relA A(chlD-pgt)8 A c1857 A(int-tLl)BAM
A(cro)H1hsdR A (lac-proAB) (F' proAB lacIq lacZAM15)HfrC (X)
PlasmidspVK101pVK102pSUP202pTZ18RpTZ19RpJRD215pBluescript SK+
pBluescript KS+pGP1-2pRME1pCH311pCH312
pCH313
pCH314
pCH319pCH321pCH323pCH327pGE1pGE52pGE53pGE69pGE111pGE125pGE151
pGE194pGE195pGE215pGE219pGE220pGE221pGE223pGE231pGE244pGE245
Kmr Tcr Mob', RP4 onKmr Tcr Mob', RP4 onApr Cmr Tcr Mob+Apr, lacZ', fl onApr, lacZ', fl onKMr Smr Mob', RSF1010 onApr, lacZ', T7 410 promoter, fl on
Apr, lacZ', T7 +10 promoter, fl onKmr, T7 gene 1, plSA onApr, Kmr cassette1.1-kb PstI-SmaI fragment of pGE1 in pBluescript SK+6.4-kb SmaI-NheI (Klenow treated) fragment in pCH311 betweenSmaI and SpeI (Klenow treated)
6.7-kb BglII fragment of pCH312 replaced by a BamHI-excisedKmr cassette
2.3-kb PstI-XbaI (Klenow treated) fragment of pCH313 inpSUP202 between PstI and EcoRI (Klenow treated)
4.8-kb PstI-SalI fragment of pGE195 in pBluescript KS+1.5-kb BamHI-PvuII fragment of pGE195 in pBluescript KS+1.5-kb PvuII-EcoRI fragment of pGE195 in pBluescript SK+4.8-kb EcoRI fragment of pGE195 in pBluescript SK+11-kb EcoRI fragment of pCH122 in pVK10115.2-kb fragment of XAEP1 in pVK1017.1-kb Sall fragment of pGE52 in pVK1014.4-kb BglII-SalI fragment of pGE52 in pVK1012.5-kb XhoI fragment of pGE52 in pVK101Derivative of pGE52 with deletion of a 8.0-kb PstI fragmentDerivative of pRK404 (11), EcoRI site in the onV region removedby treatment with Klenow polymerase, HindIII-EcoRI fragmentof the pUC18 polylinker inserted between HindIII and EcoRI
7.5-kb PstI-XbaI fragment of pCH312 in pGE1519.2-kb PstI fragment of pHG1 in pVK1019.2-kb PstI fragment of pGE195 in pJRD215hoxG::TnS-B21 derivative of pGE215hoxM::TnS-B21 derivative of pGE215hoxL::TnS-B21 derivative of pGE215hoxV::Tn5-B21 derivative of pGE215Derivative of pGE219 with deletion of a 2.1-kb KpnI fragment4.0-kb NsiI-BglII fragment of pGE221 inserted in pJRD215Derivative of pGE223 with deletion of a 8.1-kb BglIl fragment
26265636369Stratagene Cloning
Systems, Inc.Stratagene59W. MesserThis studyThis study
This study
This study
This studyThis studyThis studyThis study15This studyThis studyThis studyThis studyThis studyA. Tran-Betke
This studyThis studyThis studyThis studyThis studyThis studyThis studyThis studyThis studyThis study
BacteriophagesXAEP1M13K07X::Tn5-B21
18.5 kb of pHG1 inserted in phage XAL47
b221 cI857 Pam8O, transposon delivery system carrying a modifiedTn5 with a pomoterless lacZ gene
a Mob, mobilizability; Tra, transfer of mobilizable plasmids; on (oriV), origin of replication.
564W. Messer
2147
25Pharmacia57
J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
17
Dec
embe
r 20
21 b
y 31
.170
.60.
86.

A. EUTROPHUS MBH GENES 6279
designated pGE52. Plasmids pGE53, pGE69, pGE111, andpGE125 were obtained from pGE52 via deletion or subclon-ing (Table 1). A 7.5-kb PstI-XbaI fragment of pCH312 wasligated into pGE151 to give plasmid pGE194. A 9.2-kb PstIfragment containing the MBH locus was isolated directlyfrom a digest of megaplasmid DNA and introduced into theunique PstI site ofvector plasmid pGE151 (Table 1), yieldingplasmid pGE195.A 9.2-kb PstI fragment of plasmid pGE195 carrying the
hoxP locus was inserted into the NsiI site of the broad-host-range plasmid pJRD215. One of the resulting pair of plas-mids, designated pGE215 (9), was used for mutagenesis withTnS-B21. Four other plasmids, pCH319, pCH321, pCH323and pCH327, were constructed for use in the T7 expressionsystem.Media and growth conditions. Strains ofA. eutrophus were
grown in Luria broth with 0.5% (wt/vol) sodium chloride (LBmedium) (42) or in mineral salts medium as describedpreviously (14, 52). Synthetic media for heterotrophic cul-tures contained 0.4% (wt/vol) fructose (FN medium), 0.2%fructose and 0.2% (wt/vol) glycerol (FGN medium), or 0.4%(wt/vol) succinate (SN medium). Lithoautotrophic cultureswere grown in mineral salts medium under an atmosphere ofhydrogen, carbon dioxide, and oxygen (8:1:1, vol/vol/vol).Strains of E. coli were grown in LB medium. Solid mediacontained 1.5% (wt/vol) agar. Antibiotics were added at thefollowing concentrations: 350 ,g of kanamycin per ml and 15,ug of tetracycline per ml for A. eutrophus and 25 ,ug ofkanamycin per ml, 15 ,g of tetracycline per ml, and 100 ,gof ampicillin per ml for E. coli.
Cloning and sequencing. Standard DNA techniques wereemployed (34). Large-scale isolation of plasmid DNA wasdone by the alkaline lysis procedure followed by ethidiumbromide-cesium chloride gradient centrifugation. A. eutro-phus megaplasmid DNA was isolated as described previ-ously (15). DNA fragments used in plasmid constructionswere isolated from agarose gels by electroelution onto NA45DEAE membranes (Schleicher and Schuell, Dassel, Germa-ny). For sequence analysis, restriction fragments were in-serted into the vector plasmids pTZ18R and pTZ19R (36).Serial deletions were generated by treatment with exonucle-ase III-S1 nuclease (22) with an Erase-a-Base kit (PromegaCorp., Madison, Wis.) as recommended by the manufac-turer. Single-stranded templates were isolated from transfor-mants of E. coli NM522 after infection with the helper phageM13KO7 and sequenced by the dideoxy-chain terminationmethod (49) with a T7 sequencing kit (Pharmacia LKBBiotechnology, Freiburg, Germany) and [5S]dATP (Amer-sham-Buchler GmbH, Braunschweig, Germany). Nucleotidesequences were compiled and analyzed with MacMollysequence analysis software (Softgene GmbH, Berlin, Ger-many) and the PC/GENE software package (IntelliGenetics,Inc., Mountain View, Calif.).Transposon mutagenesis. Plasmid pGE215 was introduced
into E. coli WM1280. Fresh cultures of the transformants(approximately 5 x 107 cells) were infected with X::TnS-B21at a multiplicity of infection of 5. After 1 h of incubation at30°C, sodium citrate was added to a final concentration of 5mM. Cells were then harvested, washed with buffer (35 mMpotassium phosphate [pH 7.01), resuspended in LB mediumcontaining kanamycin and tetracycline, and grown over-night. This culture was used for batch preparation of plas-mids. The resulting plasmid DNA was used to transform E.coli S17-1, and insert-carrying plasmids were selected byplating on LB plates containing kanamycin and tetracycline.Plasmid DNA was isolated from the doubly resistant trans-
formants, and the insertions were mapped by restrictionanalysis.Enzyme assays and immunological screening. The activities
of SH (hydrogen:NAD+ oxidoreductase, EC 1.12.1.2) andMBH (no EC number) were determined with cells grownautotrophically on hydrogen or heterotrophically on FN,FGN, or SN medium. SH activity was measured by spec-trophotometric determination of H2-dependent NAD reduc-tion after treatment of cells with detergent (19). MBHactivity was assayed in isolated membranes by spectropho-tometric measurement of H2-dependent methylene blue re-duction (51). Determination of P-galactosidase activity wasessentially as described previously (14).
Protein expression studies. The T7 RNA poymerase-pro-moter system of Tabor and Richardson (59) was used todetect expression of hax genes in E. coli. Derivatives ofplasmids pBluescript SK+ and pBluescript KS+ were intro-duced into E. coli K38, and the transformants were inducedat 42°C in the presence of rifampin (200 ,ug/ml) andL-[35Slmethionine (Amersham-Buchler). Proteins were sep-arated by sodium dodecyl sulfate-polyacrylamide gel elec-trophoresis in 6% (wt/vol) polyacrylamide gels (30).Immunoelectron microscopy. A rabbit polyclonal antise-
rum was raised against the purified MBH holoenzyme.Purification of the antibody was done by affinity chromatog-raphy on protein A-Sepharose CL-4B (Pharmacia). Theimmunoglobulin G antibodies were eluted with 0.1 M gly-cine-HCl (pH 3.0). The stock solution of the antibodiescontained 2.6 mg of immunoglobulin G protein per ml.
Exponential- and stationary-phase cells were fixed in asolution containing 0.5% (wt/vol) formaldehyde and 0.3%(vol/vol) glutaraldehyde in 50 mM potassium phosphatebuffer-0.9% sodium chloride (pH 6.9) for 1 h on ice. Thesamples were embedded by a method involving progressivelowering of the temperature with Lowicryl K4M resin andmethanol as the dehydrating agent (46). Immunolabellingwas done with protein A-gold complexes (particle size, 15nm) after preparation of ultrathin sections (postembeddingprocedure) (45). The protein A-gold complexes were pre-pared by standard techniques (58). The optimal concentra-tions of antibody and protein A-gold markers were deter-mined by serial dilutions. Electron micrographs were takenwith a Philips EM 301 electron microscope at calibratedmagnifications and at an acceleration voltage of 80 kV.Sections were evaluated according to the following criteria.Colloidal gold particles lying within 25 nm (approximate totallength of the antibody and the gold particle) of the cytoplas-mic membrane were scored as MBH in the attached state.Gold particles more than 25 nm away from the membranewere counted as cytoplasmic MBH or as background label-ling if they were outside the cells. The results reported belowrepresent averages of 50 and 25 sections of comparable sizesfor the exponential- and stationary-phase cells, respectively.The values are corrected for background labelling.
Nucleotide sequence accession number. The nucleotidesequence reported here has been deposited in the EMBL,GenBank, and DDBJ data bases under accession no.M96433.
RESULTS
Cloning of the genes for the MBH of A. eutrophus H16.Previous mapping studies showed that the MBH is encodedat a locus designated hoxP contained in part in an 11-kbsegment of megaplasmid pHG1 bounded by EcoRI sites (28).In an attempt to define the minimal region required for the
VOL. 174, 1992
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
17
Dec
embe
r 20
21 b
y 31
.170
.60.
86.

6280 KORTLUKE ET AL.
AI
E SP P PS P P PS NP NPN7Xrl 11n n
lIII IIII I I III III
E EE X
11S XSSE BgNSE XP
nliii1 1iii II
lkb
MBH activity(UImg)
HF221 HF328
E 11.0 E
E 15.2 E EE E
E P P E-16.3l
$ SS 7.1
LU 4.5
xX 2.3
P 7.5p 9.2
0 0
pGEI 0.16 nt
pGE52 2.15 nt
pGE125 1.40 0
pGE53 0.39 n t
pGE69 0 n t
pGE1I1 0 nt
N pGE194 1.18 0.68
pp pGE1S5 1.38 2.03
FIG. 1. Complementation analysis of the MBH- A. eutrophus mutants HF328 and HF221 with subcloned fragments from the hoxP region.A restriction map of the hoxP region of megaplasmid pHG1 is given in the upper part of the figure. The insertion in strain HF221 is representedas a hairpin. The deletion in strain HF328 is indicated by an open box above the restriction map. The subcloned fragments that were testedfor complementation are represented as bars below the restriction map (fragment sizes are given in kilobases). The MBH activities assayedfor the corresponding transconjugants appear at the right next to the plasmid designations. All values are for autotrophically grown cells andrepresent means of triplicate assays. The wild-type strain H16 produced 2.4 U/mg under these conditions. Restriction sites: Bg, BglII; E,EcoRI; N, NheI; P, PstI; S, Sall, X, XhoI.
synthesis of active MBH enzyme, we subcloned fragmentscontaining different segments of the hoxP locus in a broad-host-range vector and tested them for complementation ofMBH- mutants. The resulting recombinant plasmids wereintroduced by triparental crosses into the independentlyisolated MBH- mutants HF221 and HF328. Strain HF221carries a 1.3-kbA. eutrophus insertion element integrated atthe haxP locus (53). In strain HF328, 6.7 kb of the hoxPregion were deleted by gene replacement. Both mutantslacked MBH activity and were devoid of immunologicallyreactive MBH protein. The results of the complementationtests are given in Fig. 1.A plasmid carrying the 11-kb EcoRI fragment spanning the
site of the insertion in HF221 (pGE1) failed to restore MBHactivity, confirming an earlier observation (15). Plasmidscontaining an additional 4 kb or mnore of adjacent sequence(pGE52, pGE125, pGE194, and pGE195) restored MBHactivity in the insertion mutant. This suggested that theinserted sequences exert a polar effect on MBH functionsextending beyond the EcoRI site. A plasmid containing a6.3-kb PstI-EcoRI fragment that complemented the insertion(pGE125) failed to complement the deletion. Another plas-mid carrying an additional 1.2 kb of adjacent sequence(pGE194) gave a low level of MBH activity. Only a plasmidharboring a 9.2-kb PstI fragment fully restored MBH activ-ity. This indicated that functions essential for the synthesisof active MBH occupy between 7.5 and 9.2 kb of themegaplasmid DNA. Interestingly, the transconjugant HF328(pGE125), which failed to produce detectable MBH activity,contained a normal level of cross-reacting MBH protein(data not shown). One explanation for this could be that thecomplementing fragment carries the structural genes for thehydrogenase itself but lacks accessory functions necessaryfor the formation of active enzyme.
Sequence analysis of the MBH gene complex. The 9.2-kb
PstI fragment of pGE195 (Fig. 1) shown to contain thegenetic determinants of the MBH was sequenced. Thenucleotide and derived amino acid sequences are shown inFig. 2, and a summary of the analysis is given in Fig. 3. Thenucleotide sequence revealed 11 closely spaced open readingframes (ORFs) (Fig. 2). Statistical analysis predicted that allof the ORFs, with the exception of ORF4, have codingcharacteristics. The codon usage of the ORFs, again with theexception of ORF4, is typical ofA. eutrophus genes (16, 61).We therefore assigned 10 of the 11 ORFs genetic designa-tions: hoxK, hoxG, hoxZ, hoxM, hoxL, hoxO, hoxQ, hoxR,hoxT, and hoxV.Comparison of the deduced amino acid sequences with the
previously published NH2-terminal amino acid sequences(32) showed that the first two genes of the cluster, hoxK andhoxG, code for the SSU (apparent molecular mass, 31 kDa)and the LSU (apparent molecular mass, 62 kDa) of theMBH. The 16 NH2-terminal amino acids of the mature smallsubunit match amino acids 44 through 59 of the derivedamino acid sequence. This indicates that the 5' region ofhoxK codes for a 43-amino-acid signal peptide. The first 30residues of the large subunit are colinear with the NH2terminus of the nucleotide-derived sequence ofhoxG, exceptthat the initial methionine is missing. Apparently the initialmethionine residue is removed posttranslationally.Two possible RpoN-specific promoter sequences with
typical -24/-12 motifs (60) upstream of the hoxK initiationcodon were identified (Fig. 2). Sequences resembling the E.coli consensus ur70 promoter were absent in this region.Gene hoxZ could encode a protein of 27.6 kDa. The
deduced amino acid sequence of the hoxZ gene product ishomologous to the hyaC (38) gene product of E. coli as wellas to the translation products of genes located downstreamof hydrogenase genes in Bradyrhizobium japonicum (50),Rhodobacter capsulatus (31), Azotobacter vinelandii (40),
r
J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
17
Dec
embe
r 20
21 b
y 31
.170
.60.
86.

CTGCAGTAGCTGCGTGGCATTGCTCGGACGACAGACCCGCTCCTGCCGCGCATGCAGCAGGGTACGAAGGCTGCGAGCAGCGCCGGCTCATYGCCTr 100
TCCGTTGGGCGGGGCGAGACGCCGGGGGCGGGGGCTCATCCGATCAACGCCGATCACTGAACTTCCTTCTGATGCATTCAAGCGAAAACCCAGTGAGC 200
ATCTG-GCGTCGGCTAGCGCGAGGCG;ACGTCATATCGlATGACTATTTAATAGATCCGGT GTCCTt; GTlzGTAGGTTCGCCGAATAGi 300
GCGCTGTCGGGGACGCACGAACCTACG7CACAGATGCTCATACATCCTTCTCGGTATCAATCTTr AAACAAGCCATCCAACTCAGGATGGTAG 400
CGGGGGTTTC CCAGGTCTTCCAGCATTCGTGCGCGAGGGCGGTATCGCTCCCC= G 500
M V E TF Y E V M RR QG I S R R 17GCGCATC=4ACGAATGCCAATACCAATrACASAATAOClGAGAGTTAl;GlTCaAMACATTTAIr;AAGTG^ATGYCAGGCAGGGOCAi77 GCGACGA 600
boxKS F L Y SLTA TSLGL P S F L P Q I A H A M E T K P R T P 51AGT77CCGAAGTA CTAcAGCAcCCTGAC ACC GCGCAGATCGCGCACGCGATGGAAI CAAGCCGCGTACAC 700
V L W L H GLEL TC )S E S F I R S A H P L A K D V V L S M I S 84CAGTCTIGTT :CAiT:T:GCTGCCGGCGC GATCITMTCGCTCGGCCCATCCCGCTGGCAAlGGACGTCTGCTATCGATGATCTC 800
L D Y D D T L M A A A G H QA E A I L E E I M T K Y K G N Y I L A 117ACTGGACTACGACGACACTGM~GCGCTGCCGGCCACCAGGCCGAGGCCATCCTCGAGGAGATCATGACGAAGTACAAGGGCAACTATATTCTGGCG 900
V E G N P P L N Q D G M S Q L K Y V A XDAXA 15VEGNPPLNQDGMS~~~:ACI IIG G R PF IEL YV K DAK A 151GTGGAGGGGAACCGCCACTCA CAGGATGGCATGT GCATCATCGGGGGCGGCCATTCATTGAGCAGCTCAAATACGTGGCCAAGGATGCCAAGG 1000
I I S W G IS C) A S W G C) V Q A 4 K P N P T Q A T P V H K V I T D K 184CCATTATCTCCTGGGGTCCTGCGCATCCTGGGGATGCGTGCAGGCAGCCAACCTAATCCCACTCAGGCCACACCGGTTCACAAGGTCACCGACAA 1100
P I I K V P G C P P I A E V M T G V I T Y M L T F D R I P E L D R 217GCCGATTATCAAGGTCCCGC GCCTCCGATTGCCGAGTGATGAGGTGTCATTACCTACATGCTCACCTCGATCGTATTCCCGAACTGGATCGA 1200
Q G R PK M F Y S Q R I H D K YRRPHFDAGQFVEEWDDE 251CAGGGTCGGCCGAAGATGTICTATAGCCAGCGCATCCACGACAAATG CTACCGGCGTCCACACTTCGATGCCGGCCAG1TCGTCGAGGAATGGGACGACG 1300
S A R K G F Q L Y X M GG QK G P T T Y N A S T T R W N E G T S F 284AATCAGCCCGCAAAGGC TGClATATCAAGATG AAAGGCCCGACCAGTACACGCCGCACCACGCGCTGAACGAGGGGACGAGTTT 1400
P IQ S G H GOI G®S E D G F N D K G S F Y D R L T G I S 0 F G 317CCCCATTCAGTCGGGCCACGGl C GCrCCGAGGATGGCT?GGACAAAGGCTCATTCTACGATCGTCTGACCGGCATCAGCCAGTYCGGC 1500
V E A N A D K I d G T A S V V V G A A V T A H A A A S A I K R A S K 351GTTGAGGCCAACGCCGACAAGATGGCGGACGGCCTCCGTCGTGTGGGGGCGGCCGTGAGGCGCATGCCGCAGCGTCTGCGATCAAGCGTGCGTCGA 1600
boxGK N E T S G S E H* (M) S A Y A T Q G F N 9
AGAAGAACGAAACCAGCGGCAGTGACACTAAGCCGCCGGGGAAACGACTGAATCAGGaAMATCGAAATAATGTCAGCTTACGCAACCCAMGGCTTCAAT 1700
L D D R G R R I V V D P V T R I E G H M R @ E V N V D A N N V I R N 43CTTGACGACCGCGGCCGTCGCATTGTCGTCGATCCCGTCACCCGCATCGAGGGTCATATGCGCTGCGAGGTGAATGTCGATGCCAACAATGTCATTCGCA 1800
A V S T G T M W R G L E V I L K G R D P R D A W A F V E R I G V 76ACGCTGTTTCCACST=ACCATGTGGCGCGGACTGAAGTGATTCTCAAkGGCCGCGATCCGCGCGACGCCTGGGCGTTCGTAGAACGCATCCCGCGGTGT 1900
T G ALAS V R A V E N A L D I R I P K N A H L I R E I M A 109TGTACCGG Gc cGGTGcGTGccGTGGAAAACGCGCTCGACACAGAATTCCAAAGAACGCCCATCTGATCCGAGAGATcATGGCC 2000
K T L Q V H D H A V H F Y H L H A L D W V D V M S A L K A D P X R T 143AACURC CCCAAAGCCCCGA 2100
S E L Q Q L V S P A H P L S S A G Y F R D I Q N R L K R F V E S G 176C'ITCCGAGTTGCAGCAGTTAGTIGCCTGCGCATCCGCTGCTCGGCAGGCTATrCC GCGATATTCAAAATCGAC7CAA.CGCTTTGTCGAGAGTGG 2200
Q L G P F M N G Y W G S K A Y V L P P E A N L M A V T H Y L E A L 209TAGCTTGGCCTATGAGGTACTGGGGATCCAAGGCTTATTGCTGCCGCCGGAGGCCAATCTGATGGCGGTCACGCATTATTTGGAAGCGCTG 2300
D L Q K E W V K I H T I F G G K N P H P N Y L V G G V P O A I N L D 243GACCTACAGAAGGMA1GGTGAAAACCACACCATCTTCGGCGGCAAGAATCCGCACCCGAACTACTTGGTCGGTGGCGTGCCGTGCGCGATCAATCTCG 2400
G I G A A S A P V N M E R L S P V K A R I D E I I E F N K N V Y V 276ATGGTATCG C TA C A CA GCATCGACGAGATCATCGAATTCAATAAGAMTGTATACGT 2500
P D V L A I G T L Y KQ A G W L Y G G G L A A T N V L D Y G E Y P 309GCcAGACGTGCTCGCCATCGGCACACTGTATAAACAGGCCGGGTGGCTGTACGGCGGCGGGCTGGCAGCCACCAACGTGCTTGACTACGGCGAGTACCCG 2600
N V A Y N K S T D Q L P G G A I L N G N W D E V F P V D P R D S Q 0 343AACGTTGCCTACAACAAGAGCACTGACCAACTGCCCGGCGGCGCGAI'CCTCAACGGCAACTGGGACGAAGTATTTCCAGTGGATCCGCGCGACTCCCAAC 2700
V Q E F V S H S W Y K Y A D E S V G L H P W D G V T E P N Y V L G 376AGGTGCAGGAMTTCGTGTCGCACAGCTGGTACAAGTATGCCGACGAGAGCGTAGGTCTGCATCCCTGGGACGGCGTGACTGAGCCCAATTACGTGCTCGG 2800
A N T K G T R T R I E Q I D E S A K Y S W I K S P R W R G H A M E 409TGCAAACACTAAGGGTACACGCACGCGCATCGAGCAAATCGACGAGAGCGCGAAGTACT'CGTOGATTAAATCGCCGCGCTGGCGCGGCCACGCGATGGAG 2900
V 6 P L S R Y I L A Y A H A R S G N K Y A E R P K E Q L E Y S A Q M 443GTAGGGCCGCTGTCGCGCTACATCCoGCCTATGCCCATGCGCGGAGCGGCAACAAGTACGCTGAGCGTCCCeAGGAGCAGCTTGAGTACTCCGCGCAGA 3000
I N S A I P K A L G L P E T Q Y T L K Q L L P S T I G R T L A R A 476TGMTCAACAGTGCGATACCAAAGGCATTGGGATTGCCAG AACACAATACACGCTCAAGCAGTTGTTGCCCAGCACGATCGGTCGTACGCTGGCGCGCGC 3100
L E S Q Y C G E M M H S D W H D L V A N I R A G D T A T A N V D K 509ACTCGAGAGCCAATAIG)CGGAGAAATGATGCATAGCGACTGGCATGATCTGGTCGCCAACATCCGGGCGGGCGATACGGCAACCGCCAACGTTGACAAG 3200
W D P A T W P L Q A K G V G T V A A P R G A L G H W I R I K D G R I 543TiGGGATCCTGCCACCTGGCCGCTGCAAGCCAAGGGCGTTGGGACCGTCGCTGCGCCGCGCGGCGCTCTCGGACACTGGATTCGTATCAAGGACGCCGG;A 3300
FIG. 2. Nucleotide sequence of theA. eutrophus H16 hoxP region. The amino acid sequence deduced from the major ORFs is given in thesingle-letter code above the nucleotide sequence. Cysteine residues are circled. The leader peptide of the SSU is italicized. Stop codons areindicated by asterisks. The conserved dinucleotides of two possible or promoters and Shine-Dalgarno sequences are underlined. The nucleotidesequence is numbered beginning with the first C of the PstI site. Coordinants of the nucleotide and amino acid sequences appear at the right.
6281
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
17
Dec
embe
r 20
21 b
y 31
.170
.60.
86.

6282 KORTLUKE ET AL. J. BACTERIOL.
E N Y Q C V V P T T W N G S P R D Y K G Q I G A F E A S L M N T P 576TCGAGANCTATCAGTGTAGTGCCTACCACGTGGAATGCAGTCCGCGTGATTACAAGGGGCAGATCGGCGCATTTGAGGCTTCGCTGATGAACACCCC 3400
V V N P E Q P V E I L R T L H S F D P @) L A (@ S T H V M S A E G Q 609AGTGGTCAACCCGGAGCAGCCGGTGGAAATCTTGCGCACGCTGCATCGTTCGATCCCTGTCTGGCGTGTTCGACTCACGTCATGAGCGCGGAAGGCCAG 3500
hoxgE L T T V K V R * M S T 3GAACTCACTACAGTCAAGGTGCGATAAAAGGTGCCGGACCGGCGTCTGGCCAGCGAGCGGATATCGGTTCGGCAACGGCACAAGGATAGCATGAGCAC 3600
K M Q A D R I A D A T G T D E G A V A S G K S I K A T Y V Y E A P 36AAAAATGCAGGCGGATCGCATGCAGATGCGACCGGGACCGACGAAGGAGCGGTAGCCAGCGGGAAGTCAATCAAGGCCACTTATGTTATAAGGCGCCA 3700
V R L W H W V N A L A I V V L A V T G F F I G S P P A T R P G E A S 70GTGAGCTGTGGCACTGGGTCAATGCGCTGGCGATCGTAGTGCTGCAGTGACCGGAT TTTATCGGCTCGCCGCCCGCGACCAGGCCGGGGGAGGCCA 3800
A N F L M G Y I R F A H F V A A Y I F A I G M L G R I Y W A T A G 103GCGCAAAC2TTCTGATGGGCTATATTCGCTTTGCCCACTTTGTCGCAGCTTACATATTCGCGATCGGCATGCTGGGCCGCATCTACTGGGCGACGGCAGG 3900
N H H S R E L F S V P V F T R A Y W Q E V I S M L R W Y A F L S A 136GAATCATCATrCCCGCGAACTCTTCCGTGCCGGTGTCACTCGGGCGTACTGGCAGGAGGTGATTTCGATGCTGCGTTGGTACGCCTTCCTATCTGCG 4000
R P S R Y V G H N P L A R F A M F F I F F L S S V F M I L T G F A M 170CGTCCAGCCGGTATGTCGGTCACAATCCGCTGGCCCGTTTCGCGATGCTTCA TTCCTGGTCGGTGTTCATGATCCTCACGGGCTTCGCGA 4100
Y G E G A Q M G S W Q E R M F G W V I P L L G Q S Q D V H T W H H 203TGTACGGCGAAGGCGCCAGATGGGCTCGTGGCAGGAGCGCATGTCGGCTGGGTCATTCCTTTGCTCGGTCAATCTCAGGATGTGCATACCTGGCATCA 4200
L G M W F I V V F V I V H V Y A A I R E D I M G R Q S V V S T M V 236TTGGGTATGTGGrGATTTTGGATCGTCCATGTCTATGCAGCGATTCGCGAGGACATCATGGGCCGCCAGAGCGTAGTGAGCACGATGGTC 4300
S G Y R T F K D* M Q P K R S Q A F V D T M H P L P E V L P S V 23TCGGGCThTCGGACCTmUCGATGCGCAATGCAACCGAAGCGATCGCAGGCTTTCGTCGATACCATGCACCCGCTACCGGAGGTATTGCCCTCGG 4400
W W H P G V S R R V S V H S G ) F L S S H Y L K P L G L S Q P H A 56TATGGTGGCATCCCGGCGTGTCACGGCGCGTGTCGGTTCATTCGGGCTGTCTGAGTAGTCAnACCTCAAGCCGCTCGGCCTGTCGCAGCCCCACGC 4500
R C L L R I A K R R L H D I I I W N S R Q A W K G A ®) S S F R V S 89CCGC GTCTGCTCAGGATTGCGAAACGGCGCTTGCATGATATCATCAATTGGAACAGCCGTCAGGCTTGGAAAGGCGCCTGTTCCTCCTTTCGTGTCTCG 4600
V A R R T V H G S L F R * 101GTCGCTCGCCGCACGGTACACGGTTCCCTTTTTCGCTGACCACCGCCATGAGACCAACTCAAACGTCCGATATCGCACCTGGAACCAGCTGTAACCAATC 4700
M V V A M G I G N V L W A D E G F G V RC) I E T L Q Q R 28CCGTGACAGAkCC CGGTGGTAGTTGCAATGGGCATTGGCAATGTGCTCTGGGCGGATGAGGGCTTIGGCGTGCGCTGCATTGAAACCCTCCAGCAGCGC 4800
Y Q F A P Q V L V D G G T Q G L Y L I H H V Q A A S R L L I F D A 62TATCAGTTCGCACCGCoAGGTA9eCCTGGTCGACGGGGGCACGCAGGGGCTCTATCTGA7TCCACCACGTGCAGGCTGCGTCGCGCCTGCTGATCTTCGATG 4900
I D Y G L P P G T L R I I E D E A V P K F L G A K K M S L H Q T G 95CGATCGACTATGGCCTGCCTCCAGGAACGCTGAGAATTATCGAAGACGAGGCGGTGCCGAAATTCCTTGGGGCCAAGAAGATGAGCCTGCACCAGACCGG 5000
F Q E V L L L A Q L T G Q Y P Q Q V V L I G ® Q P E E L E D Y G G 128CTTCCAGGAAGTACTGTTGCTCGCCCAGTTGACGGGACAGTACCCGCAGCAGGTGGTGCTGATCGGCTGTCAGCCGGAGGAGTTGGAGGATTATGGCGGC 5100
S L R R V M K A A V E D A V E K G A D L L R R W G G M P V P R T A E 162AGTCTAAGGCGGGTGATGAAGGCTGCGGTGGAGGATGCGGTGGAGAAAGGGGCGGACTTGCTGCGGAGATGGGGCGGGATGCCGGTGCCTCGCACTGCGG 5200
L A P A E A V T V P H L A L D R Y E A E R P S P R A A ' R I G D E 195AGCTrGCGCCTGCCGAGGCCGTGACCGTGCCGCATCTGGCGCTCGATCGCTACGAAGCCGAACGTCCTTCGCCTCGGGCCGCCTGCCGGATTGGCGATGA 5300
R F M P Q D L * MC I G I P M Q V M A I E P G Y A V C A G R 22AC TTTATGCCACAGGATTG2WGGC CATTCCCATGCAGGTCATGGCGATCGAGCCTGGATATGCGGTC1R TGCGGGACGC 54 00
G E R R R V S S A L V G D ®) Q Q G D W L L V F L D S A R E R I D A L 56GGCGAACGACGGAGAGTCTCGAGCGCGCTGGTTGGTGACTGCCAGCAGGGGACTGGCTGCTGGTCTrCCTGGAITCGGCGCGTGAGCGGATCGATGCGC 5500
R A H E I D A T L D M L Q A A C W V L T H P K R N H V L N C H R R 89TCAGGGCGCATGAAATCGATGCCACGCTGGATATGCTGCAAGCGGCCTGCTGGGTCTTGAcGCATCCGAAGCGCAACCACGTT AA CCATCGGCG 5600
hoxOW P E D L A V L S N S R D S S S Y* M S T E S A M F L A P I 12ATGGCCCGAGGACCICGCCGTCTATCCAATCAGAGATTCTrCCTC1'TACTM( IAkCTCATG TCGACCGAATCAGCGATGTTTCTTGCCCCCAT 5700
P T T D A V P P V I E R L V E Q HQ A A W V D Q D S I D D W L A G 45CCCGACGACGGACGCAGTACCGCCGGTGATCGAACGTCTGGTCGC5AGCATCAGGCTGCCTGGGTTGACCAGGATTCCATGACGACTGGCTGGCTGGC5800G G C @ V L F I A G D P V R F P E (® V D V A V V L P E L Q R V F F N 79GGCGG CGTGCTGTATCGCCGGCGACCCGGTGCGCTTCCCGAGTGCGTGGATGTCGCTTCGTGTGCCGGAGTTGCAAAGGGTGTTCTrCA 5900
G F R I G V A K R E R E H E D V L A N R F G T Q R R P S L V F L R 1120GCATTGGCGTGGCGAAACGTGAACGTGAGCACGAAGATGTGCTGGCGAATCGCmGGCACGCAGCGTCGACCGTCGCTGGTGTGCG 6000
D G A Y V S V I A G M R D W D E Y V R E V R R A L A M P T S R P P 145AGACGGTGCCTATGTGAGCGTAATCGCCGGCATGCGCGACTGGGACGAATACGTGCGCGAGGTCCGGCGGGCGCTICWGATGCCGACT7CACGGCCGCCA 61 00
hmQS I G I P V I S A A A G G S ( H * M N S F P I 6TCCATCGGCATTCCAGTGATTCGGCGGCAGCCGGCGGCAGTTGCCACTGATCCGTCGAACAATGCCTCACA,CAACGCGATGAATTCATTCCCA7T 6200
P V V L L G P G S Q A E D E T L D Y I II M P Q G M S V Y T P P V L P 40CCGGTCGTGCTGCTCGGTCCCGGTCGC0AGCCGAGGAT0ACCCTCGATTACATACACATGCCGCAGGWATGAGTGTCTATACACCGCCTGCTGC6300
E P E Q I AC) L T Q A Q S V L R A I L D A L D R A D P K Q P N P R 73CGGAACCCGAGCAGATTGCCTGCCTGACCCAAGCGCAAAGTGTGCTGCGGGCGATTCTCGACGCGCTGGACCGCGCGGACCCGAAACAGCCCAATCCAAG 6400
I L W A L D E E N R Q L L K Q V L G E G E V S A R I E G E Y Q V 106AATCGAA6500GCATTGGACGAAGAAAACCGTCAACTGCTCAAGCAATCCGGTGAGGGGGAGGIGAGTGCAAGGATrGAGGGCGAGTACCAGGTG6500
FIG. 2-Continued.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
17
Dec
embe
r 20
21 b
y 31
.170
.60.
86.

A. EUTROPHUS MBH GENES 6283
Y I Q E A V F A G V W R V V S T P D A T T D Y I E V G A A P E V L 140TATATCCAGGAAGCTGTATTTGCAGGGGTGTGGCGCGTGGTG ACTTGCCCCGATGCGACGACGGACTACATCGAGGTTGGCGCAGCACCGGAAGTGC 6600
R G A A R Y G A N Q W S L D V G R V P D G V M N A P S I L A E I D 173
TGCGCGGCGCGGCGCGTTATGGCGCCAATCAGTGGAGTCTGGATGTCGGGCGGGTGCCCGACGGTGTCATGAACGCGCCCTCGATCCTGGCGGAGATCGA 6700
D Q L L S W H P G K P V H V V N L T L L P M S P Q D I T F L D E TTGACCAATTGCTGAGTTGGCATCCCGGCAAGCCGGTGCATGTCGTCAACCTGACATTACTGCCGATGTCGCCGCAAGATATTACGTTTCTCGACGAAACTL G A A S V T I L S R G Y G N C R I S S T A V P Y W R V V Y F N SCTCGGTGCTGCAAGCGTCACCATCCTTTCGCGCGGCTATGGCAACCQTCGCATTAGCAGACGGCAGTGCC TGGCGGGTGGTGTATTTCAATT
2066800
2406900
Q D T T I L N T V E I T D L P E V A A A P E D L R D S H E R L T 273CGCAGGACACCACCATTCTCAACACCGTCGAGATTACCGATCTGCCGGAAGTGCC700CGCGGCCCCGGAAGACCTGCGGGATTCTCACGAACGTCTAC7000
E V L K W V G S IAr:Arr.rArqr.AAATGGGTGGGGAMCW
hoxRM N D A G M G R F E G S Y L G D R G R L A A D A R 25* 282
.TATGAACGACGCAGGTATGGGGCGCTTCGAAGCAGTTACCTGGGAGATCGGGGGCGCCTGGCGGCCGACGCGC 7100
L ES X I W W E Y D P E V G D P V WQ I A P G T S F S A L P A HGGCT'GGAATGCAAGATCRFCWTGGTGGGAGTACGATCCTGAAGTCGGCGATC CAGTTTGGCAGATTGCGCCCGGCACTTCATTTTCGGCGCTCCCGGCGCAW R P N CnD G E A E 0 F M V L G P 0 A *
h LATM L L A N Y G A
587200
787300
C E V D R V R H L E E A F ®®(DI A A T R M A D I P V V N R A L S V E 42'gGGAAGTGGACCGGGTGCGCCATCTGGAAGAGGCTTTCTGCTGCATTGCGGCAASGCATGGCCGATATCCCCGTGGTGAACCGCGCGTTGTCGGTGG 7400
A L G F E Q®C A E S A G G S D G E M G I L I T P W F M N L I W L AAAGCATTGGGGTTCG fCAGTGCCGAATCAGCGGGCGGCTCGGATGGCGAAATGGGGAT CTCCCTG-TCAGACCTGATGGCTAGC
757500
P Y G P C L G E R D A S T L P V G K T ® M R R F G S H D F E F I G 108GCCATACGGCCCA CCTCGGAGAGCGGGATGCGTCGACGCTGCCGGTGGGAAAGACC FGTATGCGACGATTTGGAAGCCACGACTTCGAATTCATCGGT 7600
A S E P Q F G P Y Q F C S L F S P M F E F A N Q A S A R A T G T E V 142GCCTCGGAGCCACAGTTTGGTCCCTACCAATTTQCTCATrATTTTCGCCGATGTTTGAATTCGCCAACCAGGCTAGTGCGCGCGCAACCGGCACGGAAG 7700
L R L L R A S P D A P L P P T ® A P H R D R R G F L F G R R R I E 175TATTGCGTCTGTTACGGGCGTCGCCAGATGCGCCT 7ACCGCCA8CGTGCGCCCCGACCCGAGACAGGCGGGGGTTTCTGTTCGGTAGGCGTCGAATTGA7800
hozVM T R A V A L T G R L T F R P G Q V P G I A G E R P Q L A E
G E S L *GGGGjaC,TCGTTATGACTCGGGCCGTCGCGTTACCGGTCGGCTGACATTCGCCCAGGGCAGGTGCCAGGCATTGCGGGTGAGCGCCCCCAACCGCCGR L L R G R P G E A A P H L L P R L F A L QDG E A H G V T A A L A
AACGATTGCTGCGAGGCCGGCCCGGCGAGGCTGCACCGCACCTCCTGCCGAGGCTCTTTGCTCTGTGCGGCGAGGCGCACGGCGTCACCGCGGCGCTCGCV N A A L G R V A A P E P A L F R R L A H E T A @DE H I R R I W LCGTGAACGCTACGCTTGGTCGR GCCGCGCCCGAACCGGCAGCCCATGAGACGGCATGTGAGCATATCCGCCGCAACRR GCTG
D W P L H L A S G P V P S T F N S R V P Q R E L I D @(P M L K A S HGACTGGCCGCTGCATCTGGCCAGCGGGCCGGTGCCGTCCACGTTTAATAGTCGAGTGCCGCAGCGCGACTGATTGCCCGCGTTGAA CGC
S E S A A M L A W L E R A V L G T A P R R W L A H W H E D P A GATTCGGAGTCGGCCGCCATGCCTGGTG AGGGCGGTGCTGGGAACCGCACCCCGCCGCTGGCTGGCGCATIGGCATGAAGACCCGGCCGGC
L S T W A T K I H T W P A M A M R Q Q M Q V A Q A M A S M P A P LC rCACGCGACCAAGATCCACACCTGGCCTGCAATGGCAATGCGTCAGPCATGCA,GGTAGCGCAGGCCATGGCGAGCATGCCTGCCCCCCTG
30179
7900
638000
968100
1308200
1638300
1968400
L P H A S T D A L R E L A Q S L A G E A D F P R P S W Q G R V F E 230CTGCCGCACGCGTCAACGGACGCCTTGCGCGAACTGGCGCAGTCCTTGGCCGGGGAGGCGGAC CCTTGCCGCCCAAGCTGGCAAGGCCGGGTGTTCG 8500
T G S W T R L G L A D (DS A F G N M W L R L G A R I A E L V L L S 263AGACTGGCAG'GGACACGACTGGGGCTGGCGGATTGCAGCGCCTTCGGCAACATGTGGT00GCGGCTGGGCGCGCGCATCGCGGAGTTGGTGTTGCTGTC8600
L R D G M Q A S G E A H D S P PCL Q M G A L A L A P G Q A L A W S
GCTGCGGGACGGTATGCAGGCATCCGGGGAGGCACATGATTCACC TGAAATGGGCGCATGGCGCTGGCTCCCGGCCAGGCGTGGCATGGAGC2968700
E M A R G L L M H W V R L V D T P Q G P V I G G Y R I I A P T E W N 330GAAATGGCCCGTGGCTTGCTGATGCACTGGTACGTCTGGTGGATACGCCGCAAGGACCGGTGATCGGCGGCTATCGGATCATTGCGCCGACTGAALTGGA 8800
F H P D G A V A H M L A H L A P F D A A D V R R S I G I L V A A Y 363
ACTTCCATCCGGACGGGGCGGTGGCGCACATGCTCGCCCACCTCGCGCCGTTCGACGCGGCCGATIGTGCGGCGCAGCATCGGCATTCTGGTCGCGGCGTA 8900
D P O) V P Y T V E F A E S L G D T V H A *TGATCCTTGCGTCCCCTATACGGTGGAGTTCGCGGAGTCFTTGGGAGATACTGTCCATGCATGAGCTCAGCCTGGCGGGTGGCATCT.GCAGTTGCTCGAAGACGCTGCGCGGCGCGAACGATTTGCTCGGGTCACGCTCTTGCGCCTCGAGGCAGGCAAGTTGTGT'GGCGTGGAGGTACGCGCGCTGCGTTTGCGCTCGAGGCCATTGCTTCGGGAACCTGCCTGGAGGGCGCTTGTATCGAGATCGAGGAACCTGrAGGG%ACAGGC'CTGTGCCTGCAG
FIG. 2-Continued.
3839000
9100
9181
Azotobacter chroococcum (17), and Wolinella succinogenes(12). A hydropathy plot (29) of the deduced amino acidsequence of protein HoxZ revealed four hydrophobic do-mains with the characteristics of transmembrane helices(data not shown). This pattern suggested that HoxZ is an
integral membrane protein. A data base search indicated a
similarity between HoxZ and cytochrome b of Neurosporacrassa (8).
The 22.2-kDa translation product of gene hoxM predicts a
soluble protein. HoxM is homologous to the hyaD gene
product of E. coli (38). A possible role of protein HoxM inthe formation of active hydrogenase is discussed below.The next gene in the series, hoxL, could encode an
11.9-kDa acidic polypeptide homologous to the translationproduct of gene hypC of the E. coli hyp operon (33).Moreover, the hoxL gene product is also homologous to the
VOL. 174, 1992
n 8% %W r L'i LO %N " 916 " %I . 1. . - - . log ..
CTGGCGGTC;TCCG;A'T%cGAcGGTGAAGccGAGcAATTTATGGT,GcTcGGTccGcAr.GcGTamacTGCAACGATGTrWTGGCCAACTATGGAGCC
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
17
Dec
embe
r 20
21 b
y 31
.170
.60.
86.

6284 KORTLUKE ET AL.
bpo 1 000 2000 3000 4000 5000 6000 7000 8000 9000
I i
PNsNB Bg X K E B E BEPv XNsB PvS X S S E Bg NSEE NsXXP
ho,G11hx R hoxM11 hoIL IIh I ho" hoxVhoxK hoxG hoxZ ORF4 hoxM hoxL hoxO hoxQ hoxR hoxT hoxV
Size of genes/predicted gone products:
bp 1080
a 360
kDa 39.5
1854
617
68.7
732 303 606 318 483
244 101 202 106 161
27.6 11.6 22.2 11.9 17.7
846 234 541
282 78 179
30.9 8.6 19.7
Length of intergenic regions:
68 9 80 12 13 34
Homologous E. coilgenes:
hyaA
% identity 70
hyafB
40
hyaC hyaD
50
hyaE hyaF
43 26 33
FIG. 3. Physical and genetic map of the hoxP region. ORFs are indicated as boxes below the line. A synopsis of the data for the ORFsis given in the lower part of the figure. The scale is in base pairs. Restriction sites: B, BamHI; Bg, BglII; E, EcoRI; K, KpnI; N, NheI; Ns,NsiI; P, PstI; Pv, PvuII, S, Sall, X, XhoI.
product of hoxC, an A. eutrophus hox gene located in theadjacent pleiotropic region (14). The function of hoxL and itshomologs is unknown. A notable feature of the deducedamino acid sequence is the motif M-C-I-X-X-X-X-X-V. Thismotif, which is also found at the NH2 terminus of the E. colihomolog, is reminiscent of the metal binding domains offerredoxins (7) and suggests that HoxL is a metalloprotein.The deduced amino acid sequences of genes hoxO and
hoxQ are homologous to the translation products of geneshyaE and hyaF, respectively, of the E. coli hydrogenase-1operon (38). The function of these genes is unknown. Thestart codon of the next gene, hoxR, overlaps the stop codonof hoxQ. This motif of overlapping stop and start codons issuggestive of translational coupling (43). The nucleotidesequence of hoxR predicts an 8.6-kDa gene product. Thedouble cysteine motifs (C-X-X-C) are reminiscent of the
A.e. MNDB.m. MQ-C.1. MQ-D.g. MD-D.v.H MK-D.v.M MK-C.pa. MK-C.pe. MK-C.t. ME-M.e. MD-D.d. MQ-P.a. MQ-
DAGMGRFEGSYLGDRGRLAADARLECKICWWEYDPEVGD----------------------KYVCDICGYVYDPAVGD----------------------KYVCSVCGYVYDPADGE----------------------IYVCTVCGYEYDPAKGD----------------------KYVCTVCGYEYDPAEGD-_____---------------KY'VCTVCGYEYDPAEGD]
TCTVCGYIYDPEDGD]----------------------KFICDVCGYIYDPAVGD;----------------------KWQCTVCGYIYDPEVGD:----------------------KYECSICGYIYDEAEGD----------------------KYVCNVCGYEYDPAEHD
metal binding sites of bacterial redox proteins. Indeed, database searches showed clear-cut homologies between hoxRand several bacterial rubredoxins, including those of Butyr-ibacterium methylotrophicum (48), Chlorobium limicola(62), Desulfovibrio gigas (6), Desulfovibrio vulgaris Hilden-borough (5), D. vulgaris Miyazaki (55), Clostridium pasteur-ianum (35), Clostridium perfringens (54), Clostridium ther-mosaccharolyticum (41), Megasphaera elsdenii (2),Desulfovibrio desulfuricans (24), and Peptostreptococcusasaccharolyticus (1) (Fig. 4).The last two genes in the sequenced region, hoxT and
hoxV, are arranged with overlapping stop and start codons.hoxT and hoxVcould encode products of 19.6 and 41.5 kDa,respectively. Data base searches failed to reveal significanthomologies to any known proteins. An interesting feature ofthe hoxV translation product is the COOH-terminal motif
PVWQIAPGTSFSALPAHWRCPNCDGEAEQFMVLGPQA 78PDNGVAPGTAFADLPEDWVCPECGVSKDEFSP---EA 53:PDDPIDPGTGFEDLPEDWVCPVCGVDKDLFEP---ES 53IPDSGIKPGTKFEDLPDDWACPVCGASKDAFEK----Q 52IPDNGVKPGTSFDDLPADWVCPVCGAPKSEFEA----A 52IPDNGVKPGTAFEDVPADWVCPICGAPKSEFEP----A 52PDDGVNPGTDFKDIPDDWVCPLCGVGKDEFEEV--EE 54IPDNGVEPGTEFKDIPDDWVCPLCGVDKSQFSET--EE 54tPTQNIPPGTKFEDLPDDWVCPDCGVGKDQFEKI---- 52I-DGNVAAGTKFADLPADWVCPTCGADKDAFVKM--D- 521-------NVPFDQLPDDWCCPVCGVSKDQFSP----A 45
----------------------KFECTLCGYIYDPALVGPDTPDODGA-FEDVSENWVCPLCGAGKEDFEVY--ED 53* * * ** * * ** * *
FIG. 4. Multiple alignment of the derived amino acid sequence of the A. eutrophus H16 hoxR gene product and the sequences of 11bacterial rubredoxins. The alignment was generated by using the CLUSTAL program of the PC/GENE package. Asterisks in the bottom lineindicate consensus amino acids. A.e., A. eutrophus; B.m., B. methylotrophicum (48); C.I., C limicola (62); D.g., D. gigas (6); D.v.H., D.vulgans Hildenborough (5); D.v.M., D. vulgaris Miyazaki (55); C.pa., C. pasteunianum (35); C.pe., C. perfringens (54); C.t., C
thermosaccharolyticum (41); M.e., M. elsdenii (2); D.d., D. desulfuricans (24); P.a., P. asaccharolyticus (1).
bp 41
1149
383
41.5
0 15
J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
17
Dec
embe
r 20
21 b
y 31
.170
.60.
86.

A. EUTROPHUS MBH GENES 6285
PNsSg K Ns es NaP
K G Z ML O OR T V
ppGE219 I I P
K P
pGE231 I I
p p
pGE220 I t I
p p
pGE221 I tI
pGE244 I t
pGE223 I I
pGE245 IAII
B-Galactosidase activity(U/mg protein)
Growth conditions
H2/CO2/02 FGN Succinate
5.4
0
1.9
2.0
1.5
3.0
1.6
5.3
0
2.0
1.9
1.6
2.3
1.6
0.2
0
0.1
0.2
0.1
0.3
0.1
FIG. 5. Synthesis of ,-galactosidase by TnS-B21-mediated transcription fusions in cells grown under hydrogenase-repressing conditions(succinate medium) and hydrogenase-derepressing conditions (H2-CO2-02 or FGN medium). The subcloned segments of the hoxP regioncarrying TnS-B21 insertions (pinheads) in different genes are shown under a simplified restriction map. MBH genes are indicated as shadedboxes. Restriction sites: Bg, BglII; K, KpnI; Ns, NsiI; P, PstI. P-Galactosidase activities are given at the right. The values shown are meanenzyme activities determined for two independent transconjugants, each assayed in triplicate.
A-T-N-P-C, which resembles the COOH-terminal domainsof the [NiFe] hydrogenases believed to be involved in Niligation (44).
Analysis of TnS-B21-mediated transcriptional fusions. Toinvestigate the organization and regulation of the genes ofthe hoaxP locus, we subjected a plasmid-borne copy tomutagenesis with TnS-B21, a TnS derivative carrying apromoterless lacZ gene (57). The broad-host-range plasmidpGE215, carrying the entire sequenced region, served as thetarget. Insertion mutations were identified by tranformationof plasmid batch preparations and screening for doublyresistant transformants. Insertions were mapped by restric-tion analysis. Several plasmid derivatives carrying insertionsin hox genes were identified (Fig. 5). These plasmids wereintroduced into A. eutrophus H16 via conjugative transferfrom E. coli S17-1, and the resulting transconjugants wereassayed for ,B-galactosidase activity. The TnS-B21 deriva-tives pGE219 (hoxG::TnS-B21), pGE220 (hoxM::TnS-B21),pGE221 (hoxL::TnS-B21), and pGE223 (hoxV::TnS-B21) alldirected the synthesis of moderate amounts of ,3-galactosi-dase (Fig. 5). Several other TnS-B21 derivatives carrying thetransposon in the opposite orientation did not producep-galactosidase under the conditions tested (data notshown).
Plasmid pGE219 carries an insertion in the hoxG gene,which encodes the LSU of the MBH. This plasmid synthe-sized significant levels of ,-galactosidase activity when cellswere grown under hydrogenase-inducing conditions (H2-C02-02 or FGN medium) but only basal levels when cellswere grown in medium containing a preferentially utilizedcarbon source such as succinate. This pattern of P-galactosi-dase activity reflects the pattern of MBH activity synthe-sized under the same conditions (20), showing that the lacZgene in this transcriptional fusion is transcribed from apromoter (or promoters) subject to a hydrogenaselike regu-latory regime. The insertions in the downstream ORFshoxM, hoxL, and hoxV all gave the same basic pattern of,B-galactosidase activity, although the levels of activity werelower than those directed by the hoxG insertion.
The simplest explanation for these results would be theexistence of a single promoter-regulatory region directingthe transcription of the entire complex of ORFs, includinghoxV. To investigate this point, we constructed derivativesof the original insertion-harboring plasmids in which se-quences upstream of the transcriptional fusions were de-leted, and we measured the p-galactosidase activity aftertransfer to A. eutrophus H16 (Fig. 5). A deletion removing1.8 kb of the region 5' of the insertion site in the allelehoxG::TnS-B21 completely abolished P-galactosidase activ-ity. Shortening of the fragment harboring the insertion site inhoxL::TnS-B21 (pGE244) led to a reduction in the levels ofP-galactosidase activity, but significant amounts of enzymewere still produced. Truncation of the region 5' of theinsertion in the allele hoxV::TnS-B21 (pGE245) reduced butdid not eliminate synthesis of 3-galactosidase. Together,these results indicate that the genes of the MBH region areorganized in at least three coordinately regulated transcrip-tional units.
Heterologous expression studies. DNA fragments from theMBH region (Fig. 6) were inserted in both orientationsdownstream of the T7 promoter in the vectors pBluescriptSK+ and pBluescript KS+. The resulting plasmids wereintroduced into E. coli K38, and the transformants were usedin expression studies. A subcloned 4.8-kb PstI-SalI fragment(pCH319) directed the synthesis of three proteins withapparent molecular masses of 72, 42, and 24 kDa (Fig. 6).The apparent molecular masses of the first two polypeptidescorrespond to the theoretical values for the primary transla-tion products of genes hoxK and hoxG (39.5 and 68.7 kDa,respectively). This assignment is corroborated by the obser-vation that plasmids lacking the intact hoxK and hoxG ORFsproduced neither the 42-kDa protein nor the 72-kDa protein.A third polypeptide was in the size range expected for thehoxZ gene product (27.6 kDa). A plasmid (pCH321) carryingan insert with only hoxZ and the small downstream ORFproduced a protein of the same size (Fig. 6), confirming itsassignment as the hoxZ gene product. Plasmids (pCH323 andpCH327) harboring hoxM, hoxL, and hoxO produced poly-
VOL. 174, 1992
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
17
Dec
embe
r 20
21 b
y 31
.170
.60.
86.
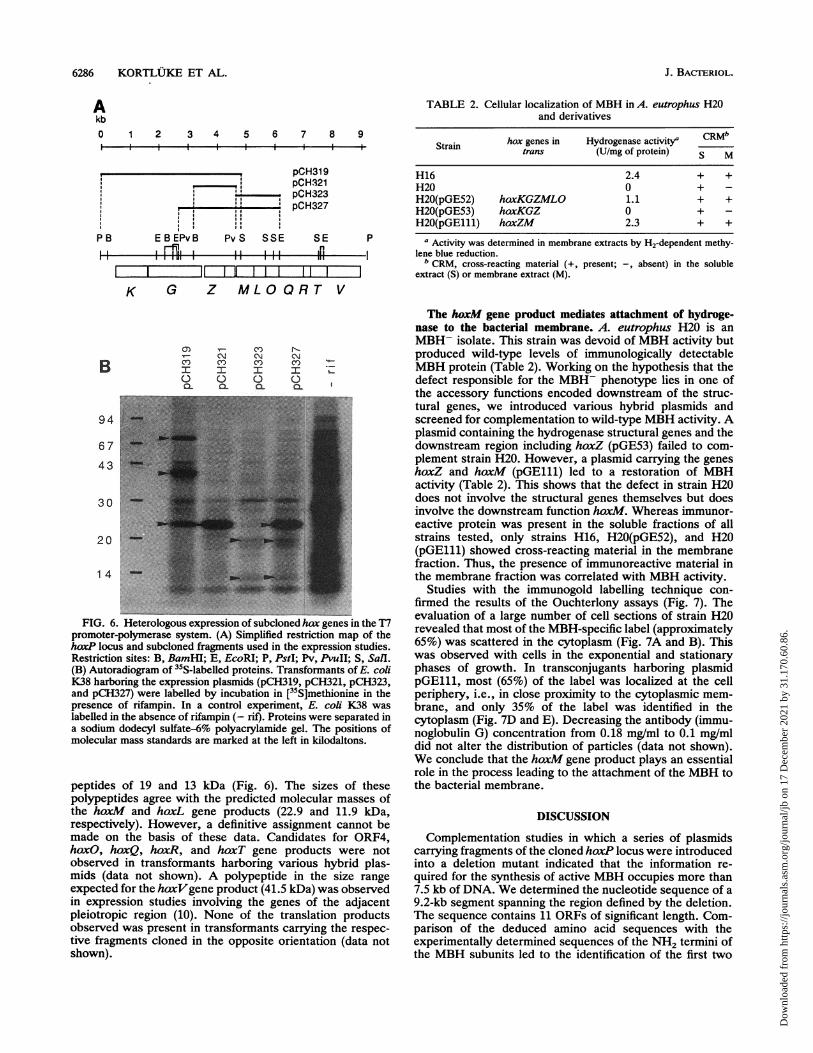
6286 KORTLUKE ET AL.
Akb0 1 2 3 4 5 6 7 8 9
pCH319pCH321
! ! pCH323pCH327
PB E B EPvB Pv S SSE SE P
L 1 I II 1
K G Z MLOQRT V
B0) X C)
_- NC\ CMj C\jCID CO CY) c
I: I Io o C0Q
94
67
43
30
20 I 1
144
FIG. 6. Heterologous expression of subcloned hax genes in the T7promoter-polymerase system. (A) Simplified restriction map of thehoxP locus and subcloned fragments used in the expression studies.Restriction sites: B, BamHI; E, EcoRI; P, PstI; Pv, PvuII; S, SalI.(B) Autoradiogram of 35S-labelled proteins. Transformants of E. coliK38 harboring the expression plasmids (pCH319, pCH321, pCH323,and pCH327) were labelled by incubation in [35S]methionine in thepresence of rifampin. In a control experiment, E. coli K38 waslabelled in the absence of rifampin (- rif). Proteins were separated ina sodium dodecyl sulfate-6% polyacrylamide gel. The positions ofmolecular mass standards are marked at the left in kilodaltons.
peptides of 19 and 13 kDa (Fig. 6). The sizes of thesepolypeptides agree with the predicted molecular masses ofthe hoxM and hoxL gene products (22.9 and 11.9 kDa,respectively). However, a definitive assignment cannot bemade on the basis of these data. Candidates for ORF4,hoxO, hoxQ, hoxR, and hoxT gene products were notobserved in transformants harboring various hybrid plas-mids (data not shown). A polypeptide in the size rangeexpected for the htxVgene product (41.5 kDa) was observedin expression studies involving the genes of the adjacentpleiotropic region (10). None of the translation productsobserved was present in transformants carrying the respec-tive fragments cloned in the opposite orientation (data notshown).
TABLE 2. Cellular localization of MBH in A. eutrophus H20and derivatives
Strain hox genes in Hydrogenase activitya CRMbtrans (U/mg of protein) S M
H16 2.4 + +H20 0 + -H20(pGE52) haxKGZMLO 1.1 + +H20(pGE53) hoaxKGZ 0 + -H20(pGE111) hoxZM 2.3 + +
a Activity was determined in membrane extracts by H2-dependent methy-lene blue reduction.
b CRM, cross-reacting material (+, present; -, absent) in the solubleextract (S) or membrane extract (M).
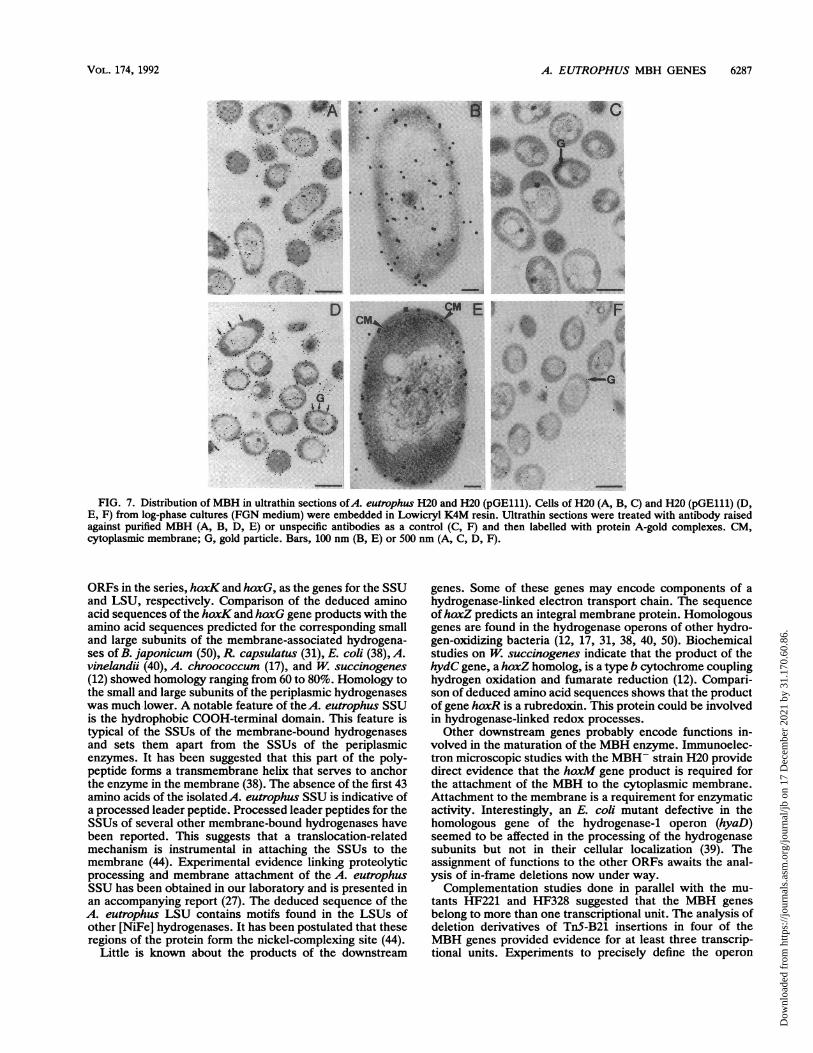
The hoxM gene product mediates attachment of hydroge-nase to the bacterial membrane. A. eutrophus H20 is anMBH- isolate. This strain was devoid of MBH activity butproduced wild-type levels of immunologically detectableMBH protein (Table 2). Working on the hypothesis that thedefect responsible for the MBH- phenotype lies in one ofthe accessory functions encoded downstream of the struc-tural genes, we introduced various hybrid plasmids andscreened for complementation to wild-type MBH activity. Aplasmid containing the hydrogenase structural genes and thedownstream region including hoxZ (pGE53) failed to com-plement strain H20. However, a plasmid carrying the geneshoxZ and hoxM (pGE111) led to a restoration of MBHactivity (Table 2). This shows that the defect in strain H20does not involve the structural genes themselves but doesinvolve the downstream function hoxM. Whereas immunor-eactive protein was present in the soluble fractions of allstrains tested, only strains H16, H20(pGE52), and H20(pGE111) showed cross-reacting material in the membranefraction. Thus, the presence of immunoreactive material inthe membrane fraction was correlated with MBH activity.
Studies with the immunogold labelling technique con-firmed the results of the Ouchterlony assays (Fig. 7). Theevaluation of a large number of cell sections of strain H20revealed that most of the MBH-specific label (approximately65%) was scattered in the cytoplasm (Fig. 7A and B). Thiswas observed with cells in the exponential and stationaryphases of growth. In transconjugants harboring plasmidpGE111, most (65%) of the label was localized at the cellperiphery, i.e., in close proximity to the cytoplasmic mem-brane, and only 35% of the label was identified in thecytoplasm (Fig. 7D and E). Decreasing the antibody (immu-noglobulin G) concentration from 0.18 mg/ml to 0.1 mg/mldid not alter the distribution of particles (data not shown).We conclude that the hoxM gene product plays an essentialrole in the process leading to the attachment of the MBH tothe bacterial membrane.
DISCUSSION
Complementation studies in which a series of plasmidscarrying fragments of the cloned hoxP locus were introducedinto a deletion mutant indicated that the information re-quired for the synthesis of active MBH occupies more than7.5 kb of DNA. We determined the nucleotide sequence of a9.2-kb segment spanning the region defined by the deletion.The sequence contains 11 ORFs of significant length. Com-parison of the deduced amino acid sequences with theexperimentally determined sequences of the NH2 termini ofthe MBH subunits led to the identification of the first two
J. BAC1TERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
17
Dec
embe
r 20
21 b
y 31
.170
.60.
86.
A. EUTROPHUS MBH GENES 6287
E
? >/. ,.*g * '' - t?. W C
.. o. La5.>.,.
s.. ..CDi_ F > s; _('
., ;? 19.
W- Z jg ' ': r % z? 1* 8; _
*S BS t_
' S- ...' ...
h ve->-vag:!d :. 1 1|?:, ." l:f
k ?,? .wX ir
R r s F
^ w 41£: F ; 't 2; X S. .: -.. , e>"*^sw .... w . '#'\-
?#sai7; .s >; %'Sui.. ?'
.. .. '? ^ _.
'wf > r X .::e *: .Z. su + ':.
,
K *,.,. "'s_ jF}F. o:
*'ty .,ls x.' , ' ,,^S: ,.w -
FIG. 7. Distribution of MBH in ultrathin sections ofA. eutrophus H20 and H20 (pGE111). Cells of H20 (A, B, C) and H20 (pGE111) (D,E, F) from log-phase cultures (FGN medium) were embedded in Lowicryl K4M resin. Ultrathin sections were treated with antibody raisedagainst purified MBH (A, B, D, E) or unspecific antibodies as a control (C, F) and then labelled with protein A-gold complexes. CM,cytoplasmic membrane; G, gold particle. Bars, 100 nm (B, E) or 500 nm (A, C, D, F).
ORFs in the series, haxK and hoxG, as the genes for the SSUand LSU, respectively. Comparison of the deduced aminoacid sequences of the hoxK and hoxG gene products with theamino acid sequences predicted for the corresponding smalland large subunits of the membrane-associated hydrogena-ses of B. japonicum (50), R. capsulatus (31), E. coli (38), A.vinelandii (40), A. chroococcum (17), and W. succinogenes(12) showed homology ranging from 60 to 80%. Homology tothe small and large subunits of the periplasmic hydrogenaseswas much lower. A notable feature of the A. eutrophus SSUis the hydrophobic COOH-terminal domain. This feature istypical of the SSUs of the membrane-bound hydrogenasesand sets them apart from the SSUs of the periplasmicenzymes. It has been suggested that this part of the poly-peptide forms a transmembrane helix that serves to anchorthe enzyme in the membrane (38). The absence of the first 43amino acids of the isolatedA. eutrophus SSU is indicative ofa processed leader peptide. Processed leader peptides for theSSUs of several other membrane-bound hydrogenases havebeen reported. This suggests that a translocation-relatedmechanism is instrumental in attaching the SSUs to themembrane (44). Experimental evidence linking proteolyticprocessing and membrane attachment of the A. eutrophusSSU has been obtained in our laboratory and is presented inan accompanying report (27). The deduced sequence of theA. eutrophus LSU contains motifs found in the LSUs ofother [NiFe] hydrogenases. It has been postulated that theseregions of the protein form the nickel-complexing site (44).
Little is known about the products of the downstream
genes. Some of these genes may encode components of ahydrogenase-linked electron transport chain. The sequenceof haxZ predicts an integral membrane protein. Homologousgenes are found in the hydrogenase operons of other hydro-gen-oxidizing bacteria (12, 17, 31, 38, 40, 50). Biochemicalstudies on W. succinogenes indicate that the product of thehydC gene, a hoxZ homolog, is a type b cytochrome couplinghydrogen oxidation and fumarate reduction (12). Compari-son of deduced amino acid sequences shows that the productof gene hoxR is a rubredoxin. This protein could be involvedin hydrogenase-linked redox processes.
Other downstream genes probably encode functions in-volved in the maturation of the MBH enzyme. Immunoelec-tron microscopic studies with the MBH- strain H20 providedirect evidence that the hoxM gene product is required forthe attachment of the MBH to the cytoplasmic membrane.Attachment to the membrane is a requirement for enzymaticactivity. Interestingly, an E. coli mutant defective in thehomologous gene of the hydrogenase-1 operon (hyaD)seemed to be affected in the processing of the hydrogenasesubunits but not in their cellular localization (39). Theassignment of functions to the other ORFs awaits the anal-ysis of in-frame deletions now under way.
Complementation studies done in parallel with the mu-tants HF221 and HF328 suggested that the MBH genesbelong to more than one transcriptional unit. The analysis ofdeletion derivatives of TnS-B21 insertions in four of theMBH genes provided evidence for at least three transcrip-tional units. Experiments to precisely define the operon
VOL. 174, 1992
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
17
Dec
embe
r 20
21 b
y 31
.170
.60.
86.

6288 KORTLUKE ET AL.
structure of the region are presently under way. When theactivity of the transcriptional fusions was monitored underhydrogenase-repressing conditions (SN medium) and hy-drogenase-derepressing conditions (H2-C02-02 or FGN me-dium), the insertions in the downstream ORFs gave the samebasic pattern of ,-galactosidase activity as that of the hoxGinsertion. This indicates that these genes are coordinatelytranscribed, as would be expected for gene products that arecomponents of a complex system or that participate in theassembly of such a system. The synthesis of active MBH isdependent on a positive regulator encoded by the gene hoxA(14). The coordinate expression of the downstream genessuggests that they are also under the control of hoxA.Taken together, the results discussed above indicate that
several if not all of the genetic functions affected by thedeletion in mutant HF328 are essential for the synthesis ofactive MBH. However, these functions alone are not suffi-cient to support the formation of the active enzyme. Asidefrom the positive regulator encoded by hoxA4, at least twoother loci mapped in the hoxBCDE region of the megaplas-mid pHG1 are required for the synthesis of wild-type levelsof enzyme atctivity (14). These loci seem to be involved in theincorporation of nickel into the nascent LSU (10). Westernimmunoblot studies reported in an accompanying publica-tion (27) indicate that both MBH subunits undergo posttrans-lational modification and that the genes of the hoxBCDEregion play an essential role in these processes. The deletionmutant HF328 produces wild-type levels of SH activity,showing that the functions affected by this mutation are notessential for SH synthesis. On the other hand, hoxB andhoxD mutations are pleiotropic, affecting the synthesis ofboth the MBH and SH enzymes (14). Thus, it appears thatthe formation of active, membrane-attached enzyme is amultistep process requiring the concerted action of bothspecific and nonspecific functions.Comparison of the organizations of theA. eutrophus MBH
genes and the E. coli hydrogenase-1 genes reveals a con-served arrangement of homologous functions. Recent stud-ies on the A. vinelandii hox genes indicate an arrangementremarkably similar to that of the other two bacteria (37). Themembrane-associated hydrogenases of these unrelated bac-teria belong to very different physiological contexts and aresubject to different regulatory regimes. The linkage of aseries of homologous genes implies that the blueprint for amulticomponent system and the apparatus needed for itsassembly originated before the divergence of these threebacterial species and has not been essentially modified.Current research efforts in our laboratory are directed atunderstanding the processes involved in the formation ofcatalytically active, membrane-bound hydrogenase from theinactive, cytoplasmic precursor.
ACKNOWLEDGMENTS
We are grateful to A. Strack for expert technical assistance and A.Kroger and R. L. Robson for communicating results before publi-cation.
This work was supported by grants from the Deutsche Forschun-gsgemeinschaft, Bundesministerium fur Forschung und Technolo-gie, and Fonds der Chemischen Industrie.
REFERENCES1. Bachmayer, H., A. M. Benson, K. T. Yasunobu, W. T. Garrard,
and H. R. Whiteley. 1968. Non-heme iron proteins. IV. Struc-tural studies of Micrcoccus aerogenes (Peptococcus aerogenes)rubredoxin. Biochemistry 7:986-996.
2. Bachmayer, H., K. T. Yasunobu, J. L. Peel, and S. G. Mayhew.
1968. Non-heme iron proteins. V. The amino acide sequence ofrubredoxin from Peptostreptococcus elsdenii. J. Biol. Chem.243:1022-1030.
3. Bowien, B., and H. G. Schlegel. 1981. Physiology and biochem-istry of aerobic hydrogen-oxidizing bacteria. Annu. Rev. Micro-biol. 35:405-452.
4. Boyer, H. W., and D. Roulland-Dussoix. 1969. A complementa-tion analysis of the restriction and modification of DNA inEscherichia coli. J. Mol. Biol. 41:459-472.
5. Brumlik, M. J., and G. Voordouw. 1989. Analysis of thetranscriptional unit encoding the genes for rubredoxin (rub) anda putative rubredoxin oxidoreductase (rbo) in Desulfovibriovulgaris Hildenborough. J. Bacteriol. 171:4996-5004.
6. Bruschi, M. 1976. The amino acid sequence of rubredoxin fromthe sulfate-reducing bacterium Desulfovibrio gigas. Biochem.Biophys. Res. Commun. 70:615-621.
7. Bruschi, M., and F. Guerlesquin. 1988. Structure, function andevolution of bacterial ferredoxins. FEMS Microbiol. Rev. 54:155-176.
8. Burke, J. M., C. Breitenberger, J. E. Heckman, B. Dujon, andU. L. RajBhandary. 1984. Cytochrome b gene from Neurosporacrassa mitochondria. J. Biol. Chem. 259:504-511.
9. Davison, J., M. Heusterspreute, N. Chevalier, V. Ha-Thi, and F.Brunel. 1987. Vectors with restriction site banks. V. pJRD215,a wide-host-range cosmid vector with multiple cloning sites.Gene 51:275-280.
10. Dernedde, J., M. Eitinger, and B. Friedrich. Unpublished data.11. Ditta, G., T. Schmidhauser, E. Yakobson, P. Lu, X.-W. Liang,
D. R. Finlay, D. Guiney, and D. R. Helinski. 1985. Plasmidsrelated to the broad host range vector pRK290, useful for genecloning and monitoring gene expression. Plasmid 13:149-153.
12. Dross, F., V. Geisler, R. Lenger, F. Theis, T. Krafft, F. Fahren-holz, E. Kojro, A. Duchene, D. Tripier, K. Juvenal, and A.Kr6ger. 1992. The quinone-reactive Ni/Fe-hydrogenase of Wo-linella succinogenes. Eur. J. Biochem. 206:93-102.
13. Eberz, G., T. Eitinger, and B. Friedrich. 1989. Genetic determi-nants of a nickel-specific transport system are part of theplasmid-encoded hydrogenase gene cluster in Alcaligenes eu-trophus. J. Bacteriol. 171:1340-1345.
14. Eberz, G., and B. Friedrich. 1991. Three trans-acting regulatoryfunctions control hydrogenase synthesis in Alcaligenes eutro-phus. J. Bacteriol. 173:1845-1854.
15. Eberzo G., C. Hogrefe, C. Kortlufke, A. Kamienski, and B.Friedrich. 1986. Molecular cloning of structural and regulatoryhydrogenase (hox) genes of Alcaligenes eutrophus H16. J.Bacteriol. 168:636-641.
16. Eitinger, T., and B. Friedrich. 1991. Cloning, nucleotide se-quence and heterologous expression of a high-affinity nickeltransport gene from Alcaligenes eutrophus. J. Biol. Chem.266:3222-3227.
17. Ford, C. M., N. Garg, R. P. Garg, K. H. Tibelius, M. G. Yates,D. J. Arp, and L. C. Seefeldt. 1990. The identification, charac-terization, sequencing and mutagenesis of the genes (hupSL)encoding the small and large subunits of the H2-uptake hydrog-enase of Azotobacter chroococcum. Mol. Microbiol. 4:999-1008.
18. Friedrich, B. 1990. The plasmid-encoded hydrogenase genecluster in Alcaligenes eutrophus. FEMS Microbiol. Rev. 87:425-430.
19. Friedrich, B., E. Heine, A. Finck, and C. G. Friedrich. 1981.Nickel requirement for active hydrogenase formation in Alcali-genes eutrophus. J. Bacteriol. 145:1144-1149.
20. Friedrich, C. G. 1982. Derepression of hydrogenase duringlimitation of electron donors and derepression of ribulosebi-sphosphate carboxylase during carbon limitation ofAlcaligeneseutrophus. J. Bacteriol. 149:203-210.
21. Gough, G. A., and N. E. Murray. 1983. Sequence diversityamong related genes for recognition of specific targets in DNAmolecules. J. Mol. Biol. 166:1-19.
22. Henikoff, S. 1984. Unidirectional digestion with exonuclease IIIcreates targeted break points for DNA sequencing. Gene 28:351-359.
23. Hogrefe, C., D. Romermann, and B. Friedrich. 1984. Alcali-
J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
17
Dec
embe
r 20
21 b
y 31
.170
.60.
86.

A. EUTROPHUS MBH GENES 6289
genes eutrophus hydrogenase genes (hox). J. Bacteriol. 158:43-48.
24. Hormel, S., K. A. Walsh, B. C. Pickril, K. Titani, J. LeGall, andL. Sieker. 1986. Amino acid sequence of rubredoxin fromDesulfovibrio desulfuricans strain 27774. FEBS Lett. 201:147-150.
25. Husemann, M., R. Klintworth, V. Bfittcher, J. Salnikow, C.Weissenborn, and B. Bowien. 1988. Chromosomally and plas-mid-encoded gene clusters for CO2 fixation (cfx genes) inAlcaligenes eutrophus. Mol. Gen. Genet. 214:112-120.
26. Knauf, V. C., and E. W. Nester. 1982. Wide host range cloningvectors: a cosmid clone bank of an Agrobacterium Ti plasmid.Plasmid 8:45-54.
27. Kortlufke, C., and B. Friedrich. 1992. Maturation of membrane-bound hydrogenase of Alcaligenes eutrophus. J. Bacteriol.174:6290-6293.
28. Kortifike, C., C. Hogrefe, G. Eberz, A. Puhler, and B. Friedrich.1987. Genes of lithoautotrophic metabolism are clustered on themagaplasmid pHG1 inAlcaligenes eutrophus. Mol. Gen. Genet.210:122-128.
29. Kyte, J., and R. F. Doolitle. 1982. A simple method for display-ing the hydropathic character of a protein. J. Mol. Biol. 157:105-132.
30. Laemmli, U. K. 1970. Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature (London)227:280-285.
31. Leclerc, M., A. Colbeau, B. Cauvin, and P. Vignais. 1988.Cloning and sequencing of the genes encoding the large andsmall subunits of the H2 uptake hydrogenase (hup) ofRhodobacter capsulatus. Mol. Gen. Genet. 214:97-107.
32. Lorenz, B., K. Schneider, H. Kratzin, and H. G. Schlegel. 1989.Immunological comparison of subunits isolated from varioushydrogenases of aerobic hydrogen bacteria. Biochim. Biophys.Acta 995:1-9.
33. Lutz, S., A. Jacobi, V. Schiensog, R. Bohm, G. Sawers, and A.Bock. 1991. Molecular characterization of an operon (hyp)necessary for the activity of the three hydrogenase isoenzymesin Escherichia coli. Mol. Microbiol. 5:123-135.
34. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecularcloning: a laboratory manual. Cold Spring Harbor Laboratory,Cold Spring Harbor, N.Y.
35. McCarthy, K. F. 1972. Ph.D. thesis. George Washington Uni-versity, St. Louis, Mo.
36. Mead, D. A., E. Szczesna-Skorupa, and B. Kemper. 1986. Singlestranded DNA 'blue' T7 promoter plasmids: a verstile tandempromoter system for cloning and engineering. Protein Eng.1:67-74.
37. Menon, A. L., L. E. Mortensen, and R. L. Robson. Personalcommunication.
38. Menon, N. K., J. Robbins, H. D. Peck, Jr., C. Y. Chatelus, E.-S.Choi, and A. E. Przybyla. 1990. Cloning and sequencing of aputative Escherichia coli [NiFe] hydrogenase-1 operon contain-ing six open reading frames genes. J. Bacteriol. 172:1969-1977.
39. Menon, N. K., J. Robbins, J. C. Wendt, K. T. Shanmugam, andA. E. Przybyla. 1991. Mutational analysis and characterizationof the Escherichia coli hya operon, which encodes [NiFe]hydrogenase 1. J. Bacteriol. 173:4851-4861.
40. Menon, A. L., L. W. Stultz, R. L. Robson, and L. E. Mortensen.1990. Cloning, sequencing and characterization of the [NiFe]hydrogenase encoding structural genes (hoxK and hoxG) fromAzotobacter vinelandii. Gene 96:67-74.
41. Meyer, J., J. Gagnon, L. C. Sieker, A. van Dorsselner, and J.-M.Moulis. 1990. Rubredoxin from Clostridium thermosaccharo-lyticum. Amino acid sequence, mass spectrometric and prelim-inary crystallographic data. Biochem. J. 271:839-841.
42. Miller, J. H. 1972. Experiments in molecular genetics. ColdSpring Harbor Laboratory, Cold Spring Harbor, N.Y.
43. Normark, S., S. Bergstdm, T. Edlund, G. Grundstr6m, B.Jaurin, F. P. Lindberg, and 0. Olsson. 1983. Overlapping genes.
Annu. Rev. Genet. 17:499-525.44. Przybyla, A. E., J. Robbins, N. Menon, and H. D. Peck, Jr. 1992.
Structure-function relationships among the nickel-containinghydrogenases. FEMS Microbiol. Rev. 88:109-136.
45. Rohde, M., H. Gerberding, T. Mund, G.-W. Kohring. 1988.Immunoelectron microscopic localization of bacterial enzymes:pre- and post-embedding labelling techniques on resin-embed-ded samples. Methods Microbiol. 20:175-210.
46. Roth, J., M. Bendayan, E. Carlemalm, M. Villiger, and M.Garavito. 1981. Enhancement of structural preservation andimmunocytochemical staining in low temperature embeddedpancreatic tissue. J. Histochem. Cytochem. 29:663-669.
47. Russel, M., and P. Model. 1984. Replacement of the fip gene ofEscherichia coli by an inactive gene cloned on a plasmid. J.Bacteriol. 159:1034-1039.
48. Saeki, K., Y. Yao, S. Wakabayashi, G.-J. Shen, J. G. Zeikus,and H. Matsubara. 1989. Ferredoxin and rubredoxin fromButyribacterium methylotrophicum: complete primary struc-tures and construction of phylogenetic trees. J. Biochem. 106:656-662.
49. Sanger, F., S. Nicklen, and A. R. Coulsen. 1977. DNA sequenc-ing with chain-terminating inhibitors. Proc. Natl. Acad. Sci.USA 74:5463-5467.
50. Sayavedra-Soto, L. A., G. K. Powell, H. J. Evans, and R. 0.Morris. 1988. Nucleotide sequence of the genetic loci encodingsubunits of Bradyrhizobium japonicum uptake hydrogenase.Proc. Natl. Acad. Sci. USA 85:8395-8399.
51. Schink, B., and H. G. Schlegel. 1979. The membrane-boundhydrogenase ofAkaligenes eutrophus. I. Solubilization, purifi-cation, and biochemical properties. Biochim. Biophys. Acta567:315-324.
52. Schlegel, H. G., H. Kaltwasser, and G. Gottschalk. 1961. EinSubmersverfahren zur Kultur wasserstoffoxidierender Bakte-rien: Wachstumsphysiologische Untersuchungen. Arch. Mikro-biol. 38:209-222.
53. Schwartz, E., C. Kortluke, and B. Friedrich. Unpublished data.54. Seki, Y., S. Seki, M. Sato, A. Ikeda, and M. Ishimoto. 1989.
Rubredoxin from Clostridium perfringens complete amino acidsequence and participation in nitrate reduction. J. Biochem.106:336-341.
55. Shimizu, F., M. Ogata, T. Yagi, S. Wakabayashi, and H.Matsubara. 1989. Amino acid sequence and function of rubre-doxin from Desulfovibrio vulgaris Miyazaki. Biochimie 71:1171-1177.
56. Simon, R., U. Priefer, and A. Piihler. 1983. A broad host rangemobilization system for in vitro genetic engineering: transposonmutagenesis in Gram-negative bacteria. Bio/Technology 1:784-791.
57. Simon, R., J. Quandt, and W. Klipp. 1989. New derivatives oftransposon TnS suitable for mobilization of replicons, genera-tion of operon fusions and induction of genes in Gram-negativebacteria.' Gene 80:161-169.
58. Slot, J. W., and H. J. Geuze. 1981. Sizing of protein A colloidalgold probes for immunoelectron microscopy. J. Ultrastruct.Res. 26:31-43.
59. Tabor, S., and C. C. Richardson. 1985. A bacteriophage 17RNA polymerase promoter system for controlled exclusiveexpression of specific genes. Proc. Natl. Acad. Sci. USA82:1074-1078.
60. Thony, B., and H. Hennecke. 1989. The -121-24 promotercomes of age. FEMS Microbiol. Rev. 63:341-357.
61. Trgn-Betcke, A., U. Warnecke, C. B6cker, C. Zaborosch, and B.Friedrich. 1990. Cloning and nucleotide sequences of the genesfor the subunits of NAD-reducing hydrogenase of Alcaligeneseutrophus H16. J. Bacteriol. 172:2920-2929.
62. Wooley, K. J., and T. E. Meyer. 1987. The complete amino acidsequence of rubredoxin from the green phototrophic bacteriumChlorobium thiosulphatophilum strain PM. Eur. J. Biochem.163:161-166.
VOL. 174, 1992
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
17
Dec
embe
r 20
21 b
y 31
.170
.60.
86.