the cyanobacterial uptake hydrogenase: regulation, maturation and
TRANSCRIPT



Yes, my friends, I believe that water will one day be employed as fuel, that hydrogen and oxygen which constitute it, used singly or together, will furnish an inexhaustible source of heat and light of an intensity of which coal is not capable Jules Verne


List of Papers
This thesis is based on the following papers, which are referred to in the text by their Roman numerals.
I Holmqvist, M., Stensjö, K., Oliveira, P., Lindberg, P., Lindblad,
P. (2009). Characterization of the hupSL promoter activity in Nostoc punctiforme ATCC 29133. BMC Microbiology, 9:54.
II Holmqvist, M., Lindberg, P., Agervald, A., Stensjö, K.,
Lindblad, P. (2010) Transcript analysis of the extended hyp-operon in the cyanobacteria Nostoc sp. strain PCC 7120 and Nostoc punctiforme ATCC 29133. Submitted
III Agervald, A., Stensjö, K., Holmqvist, M., Lindblad, P. (2008)
Transcription of the extended hyp-operon in Nostoc sp. strain PCC 7120. BMC Microbiology, 8:69.
IV Devine, E., Holmqvist, M., Stensjö, K., Lindblad, P. (2009)
Diversity and transcription of proteases involved in the maturation of hydrogenases in Nostoc punctiforme ATCC 29133 and Nostoc sp. strain PCC 7120. BMC Microbiology, 9:53.
V Camsund, D., Devine, E., Holmqvist, M., Lindblad, P., Stensjö,
K. (2010) A HupS-GFP fusion protein demonstrates a heterocyst specific localization of the uptake hydrogenase in the cyanobacterium Nostoc punctiforme. Manuscript.
VI Ekman, M., Ow, S.Y., Holmqvist, M., Zhang, X., Van
Wagenen, J., Wright, P.C, Stensjö, K. (2010) Metabolic adaptation in a H2 producing heterocystous cyanobacterium: potentials and implications for bio-engineering. Manuscript.
Reprints were made with permission from the respective publishers.

Author’s request This work, a result of the author’s research for a doctoral degree at Uppsala University, Sweden, contains unpublished material. The author reserves her right to publish it at appropriate time. If any of the unpublished material is taken for reproduction, further study or modification it must be duly acknowledged giving reference to this thesis.

Contents
Introduction...................................................................................................11 A future based on solar-H2 .......................................................................11
The current climate situation ...............................................................11 Solar energy.........................................................................................11 Hydrogen as an energy carrier .............................................................12 Cyanobacterial biohydrogen production..............................................12
Cyanobacteria...........................................................................................13 The phylum Cyanophyta......................................................................13 Cyanobacterial hydrogen metabolism .................................................14 Heterocysts ..........................................................................................14 Nitrogenases ........................................................................................15 Hydrogenases.......................................................................................16 Maturation of [NiFe] hydrogenases.....................................................17 Maturation of the hydrogenase large subunit ......................................18 Maturation of the hydrogenase small subunit......................................18
The model organisms ...............................................................................20 Nostoc punctiforme ATCC 29133 .......................................................20 Nostoc sp. strain PCC 7120 .................................................................20
The uptake hydrogenase...........................................................................21 Cellular localization.............................................................................21 Subcellular localization .......................................................................21
Transcriptional regulation of the uptake hydrogenase .............................22 Physical organization of the hupSL operon in N. punctiforme ............22 The transcription regulator NtcA.........................................................23 Transcription regulators of the Cal family...........................................24
Aim of this thesis ..........................................................................................25
RESULTS AND DISCUSSION ...................................................................27 Transcriptional regulation of the uptake hydrogenase .............................27
Which are the key players in the transcriptional regulation of the uptake hydrogenase? (Paper I).............................................................27
Maturation of the uptake hydrogenase .....................................................30 Which are the accessory proteins involved in the maturation of the hydrogenase small subunit? (Paper I, II and III) ................................30

Which are the regulatory proteins interacting with the extended hyp-operon promoter region? (Paper II) .....................................................35 How are the uptake hydrogenase specific proteases transcribed, and what confers the specificity? (Paper IV)..............................................36
Function of the uptake hydrogenase.........................................................40 What is the subcellular and cellular localization of the uptake hydrogenase? (Paper I and V)..............................................................40 How do the cells reprogram their metabolism when the uptake hydrogenase is no longer present? (Paper VI) .....................................41
SUMMARY AND FUTURE OUTLOOK ...................................................48
SUMMARY IN SWEDISH..........................................................................51
ACKNOWLEDGEMENT ............................................................................53
REFERENCES .............................................................................................57

Abbreviations
2DE 2-Dimensional Gel Electrophoresis 2-OG 2-Oxoglutarate 5’RACE 5’ Rapid Amplification of cDNA Ends AbrB Antibiotic resistance protein B ATCC American Type Culture Collection BLAST Basic Local Alignment Search Tool Bp Base pair CalA Cyanobacterial AbrB-like protein A CalB Cyanobacterial AbrB-like protein B cDNA Complementary DNA EMSA Electrophoretic Mobility Shift Assay FeS Iron Sulfur GC Gas Chromatography GFP Green Fluorescent Protein Hox Hydrogen oxidation protein Hup Hydrogen uptake protein Hyp Hydrogenase pleiotropic IHF Integration Host Factor iTRAQ Isobaric Tag for Relative and Absolute Quantification NHL NCL-1, HT2A and Lin-41 mRNA Messenger RNA MS Mass Spectrometry NAD(H) Nikotinamide Adenine Dinucleotide NHM5 Nostoc Hydrogenase Mutant nr 5 OD Optical Density OPP Oxidative Pentose Phosphate ORF Open Reading Frame PCC Pasteur Culture Collection PCR Polymerase Chain Reaction PS Photosystem RT-PCR Reverse Transcriptase PCR RTqPCR Reverse Transcriptase quantitative PCR TLC Thin Layer Chromatography TPR Tetratrico Peptide Repeat Tsp Transcriptional start point TW Terrawatt


11
Introduction
A future based on solar-H2 The current climate situation One of today’s greatest challenges is to alter the current energy regime and the way we use energy. The energy demand is increasing globally every year due to a combination of an energy intense way of living in the industrialized countries and rapid economic growth in the developing countries, especially China and India [1,2]. Even with more efficient energy usage the energy demand is predicted to double until 2050 [1]. This, together with prospects of oil supplies peaking in the future, problems with national security since oil and gas reserves are concentrated in politically unstable regions of the world, air pollution and global warming caused by emissions of CO2 and other greenhouse gases, has created an unsustainable situation [2,3]. Thus, the need for a clean, renewable, carbon-neutral, and abundant fuel is raising world wide.
Solar energy The sun is the most abundant renewable energy source available and has an immense potential [1,4]. Furthermore, solar energy is more evenly distributed around the world than oil and natural gas reserves. Currently, the annual global energy consumption rate is ~13 TW (terawatt) [1,4,5]. Even if taking into account a general decrease in energy intensity due to technical developments, this demand is estimated to increase to ~ 27 TW by year 2050 [1]. The additional energy needed must come from CO2 neutral energy sources, but renewable energy such as biomass, geothermal power, wind power or hydro power does not have the capacity to meet this demand [6]. However, the face of the Earth is hit by approximately 120 000 TW of solar energy every year. In fact the energy from the sun striking the Earth in 1h (~14 TW) is sufficient to cover the current energy demand for the whole world during one year [1,6]. If we could utilize just a few percent of the total solar energy dispersed over the globe daily we would have a secure, clean, abundant and infinite energy source more than capable of meeting the future global energy demands.

12
Hydrogen as an energy carrier Due to the intermittent and diurnal nature of solar irradiation there is a need to capture and convert solar energy for storage and transportation to places of demand. The ideal way of storing solar energy is to convert it into chemically bonds by production of solar fuels, such as molecular hydrogen [1], Molecular hydrogen (H2) is a non-toxic gas [7] with the highest gravimetric energy density of currently known fuels [8]. It has an non-polluting nature since, when burned or oxidized in a fuel cell, the only emission is water [4]. This makes hydrogen one of the most promising clean fuels for the future [4]. Many developed countries, such as Canada, US, Japan and the member states of EU have roadmaps for transition to hydrogen economy (an energy system based on electricity and hydrogen [3]) and have already started the implementation of these roadmaps [2]. H2 is a versatile energy carrier since it can be produced from a variety of sources, including fossil fuels and renewables, e.g. steam reformation of natural gas and coal, gasification, pyrolysis or fermentation of biomass and organic waste and water electrolysis using wind, hydro, thermal, nuclear or solar power [2,3]. Today, more than 90% of the H2 used is produced from fossil fuels through steam reforming of natural gas. Neither is this sustainable nor does it eliminate air pollution and release of greenhouse gases. Hence, the viability of the future hydrogen economy is entirely dependent upon development of cost-effective, sustainable, non-polluting, large scale H2 production system sufficient to meet the global energy needs [4]. One way to achieve this is to couple hydrogen production to a high potential renewable energy source, such as the sun.
Cyanobacterial biohydrogen production
In nature H2 can be produced by a number of microorganisms using solar energy either directly (photosynthesis) or indirectly (dark fermentation of organic substrates) [8]. In this context, cyanobacteria are a group of organisms which have gained a lot of attention lately. Cyanobacteria are, together with green algae, the only organisms capable of both oxygenic photosynthesis, using water as the electron donating substrate instead of organic acids, and subsequent hydrogen production [9-11]. The ability to use only solar energy and water for production of a biofuel have raised the hopes of using cyanobacteria as commercially viable H2 producers. One of the main drawbacks with renewable fuel is the current cost. Sustainable production of renewable fuel is simply too expensive for commercial use, compared to oil and natural gas, and this hampers transition from dependency on fossil fuels to renewables [2]. Compared to H2 production by fermentation or gasification, the combination of photosynthetic solar energy capture and hydrogen production within the same organism is far more

13
efficient than growing, harvesting and processing biomass and therefore offers greater potential [12]. Moreover, since cyanobacteria are photoautotrophic organisms they need only air, water, a few inorganic nutrients and light for their survival and are thus inexpensive to cultivate [13,14]. Many strains are also capable of fixing atmospheric dinitrogen (N2) which further eliminates the need of nitrogen supplements to their growth media. The main cost for cyanobacterial biohydrogen production would thus come from construction and operation of bioreactors. Biomass generation for H2 production is limited by the amount of land areas available for growing crops but bioreactors containing cyanobacteria could be placed on land unsuitable for habitation or cultivation and would thus not compete for valuable land areas. If marine strains are used, the bioreactors could be placed at sea thereby eliminating the need for freshwater, a limited resource in some areas of the world. Many strains of cyanobacteria grow at ambient temperatures (20-30°C) and hence bioreactors for cyanobacteria cultivation require modest energy input for heating. All this contribute to reducing the cost of commercial cyanobacterial biohydrogen production.
Cyanobacteria
The phylum Cyanophyta Cyanobacteria are gram negative prokaryotes within the domain Bacteria [15]. The name, cyanobacteria, is derived from the Greek word for blue (kyanós) and originates from the color of the organisms. Cyanobacteria are an ancient group of organisms dating back as early as 2 billion years ago [16,17]. They are widely distributed around the globe and inhabit a range of different environments, including marine, freshwater, terrestrial and even extreme habitats, e.g. hot springs, polar regions and deserts [9,16]. Cyanobacteria have played an important role early in the history of Earth and the evolution of life. They are the original source of atmospheric oxygen and are also suggested to be the predecessor of chloroplasts in plants [16,18]. Furthermore, as important primary producers and with the ability of many strains to fix N2, cyanobacteria play a significant role in the carbon and nitrogen cycles in the biosphere [16]. Some strains of cyanobacteria are free living while others can establish symbiosis with a diversity of hosts; fungi (forming lichen), non-vascular plants (such as hornworts and liverworts), spore-producing vascular plants (such as ferns), gymnosperms (such as cycads) and angiosperms (such as members of the genus Gunnera) [15]. Cyanobacteria is a large heterogeneous group and exhibits a variety of morphologies ranging from unicellular to colonial and filamentous forms [9,16]. The cyanobacteria are divided into five groups; group I and II, containing strains with symmetric respective asymmetric division patterns,

14
group III and IV, containing filamentous forms, and group V, containing branching filamentous forms [16]. Filamentous cyanobacteria from group III have only vegetative cells while those from group IV and V can differentiate into more advanced cell types. Those specialized cells are heterocysts (cells specialized for N2-fixation), akinetes (resting cells for surviving unfavorable growth conditions such as cold and desiccation) [16] and hormogonia (gliding filaments serving as infection units when establishing symbiosis) [19]. During symbiosis the cyanobacteria provides the host with combined nitrogen in exchange for carbohydrates [14].
Cyanobacterial hydrogen metabolism In cyanobacteria, two groups of enzymes are directly involved in the hydrogen metabolism; the nitrogenases and the hydrogenases [10,18]. In strains capable of N2-fixation, H2 is produced by nitrogenase as a by-product when fixing N2. The evolved hydrogen is subsequently recaptured and oxidized by an uptake hydrogenase to prevent energy losses from the cell [10,18,20]. In addition, some cyanobacterial strains harbor a bidirectional hydrogenase, capable of oxidizing or evolving H2 depending on the present redox potential in the cell [10,18,20]. The exact function of the bidirectional hydrogenase is unclear but suggested functions are acting as an electron valve for photosynthesis generated electrons and playing a role in the release of excess reducing power during fermentation [18]. Neither the uptake hydrogenase nor the bidirectional hydrogenase is universally present. There is however a strong connection between the uptake hydrogenase and N2-fixation since the enzyme has been found in all N2-fixing, filamentous strains examined so far [9,18,21]. Hydrogenases and nitrogenase are very oxygen sensitive enzymes and are quickly inactivated in the presence of oxygen [10,18]. Therefore, cyanobacteria have developed different ways of maintaining nitrogenase enzyme activity by separating nitrogenase from oxygenic photosynthesis either spatially, in heterocysts and vegetative cells [14,18,22], or temporally, with photosynthesis during the days and nitrogen fixation during the nights [22,23].
Heterocysts During periods of nitrogen starvation some of the vegetative cells in filamentous cyanobacteria undergo a number of metabolic and morphological changes and differentiate into heterocysts [24]. Heterocysts are terminally differentiated cells, where a low oxygen environment is provided by a combination of two additional layers of envelope surrounding the cells, higher respiration rate and loss of the O2 evolving activity of PSII [10,14,22]. Approximately 5-10% of all cells, evenly distributed in the filament, differentiate into heterocysts 24 hours after nitrogen depletion

15
[9,25]. Heterocysts can be distinguished from vegetative cells by their size, shape, thick cell envelope, reduced autofluorescence (due to loss of phycobiliproteins [26]) and storage of granules with cyanophycin in the polar regions adjacent to vegetative cells [24,25]. The storages of cyanophycin are believed to serve as nitrogen reserves [25]. The two additional layers of envelope consist of a inner glycolipid layer, serving as a permeability barrier for gases, and an outer polysaccharide layer, protecting the glycolipid layer from external damage [24,25]. Despite the thicker envelope surrounding the heterocysts small amounts of oxygen will still be able to enter the heterocysts, most likely through the poles, but will quickly be consumed by respiration [25]. In Nostoc PCC 7120 two terminal respiratory oxidases, Cox2 and Cox3 (cytochrome oxidase), were found to be heterocyst specific while the third terminal oxidase, Cox1, was expressed only in vegetative cells [27]. Heterocysts do not fix carbon and are therefore depending on being supplied with reductants and carbon skeletons from surrounding vegetative cells, probably in the form of sugars [25]. In the heterocysts the imported sugars are subsequently catabolised in the OPP pathway (oxidative pentose phosphate) to generate reducing power, NADPH [25]. In return for the sugars the heterocysts supply the vegetative cells with fixed nitrogen in the form of amino acids [24]. Ammonium is assimilated in the GS-GOGAT cycle in the heterocysts by incorporation into glutamate. Glutamate is produced from glutamine and 2-OG (2-oxoglutarate) by GOGAT (glutamine-2-oxoglutarate amidotransferase) while glutamine is produced from glutamate and ammonium by GS (glutamine synthetase) [25]. While GS is highly expressed in heterocysts the enzyme GOGAT is absent, suggesting an additional exchange of metabolites between heterocysts and vegetative cells in the form of glutamine and glutamate [25]. In filamentous cyanobacteria the periplasm appears to be continuous, not only between neighboring vegetative cells but also between vegetative cells and heterocysts, and might allow for transport of molecules between the cells [24,25].
Nitrogenases Nitrogenases are metal enzymes. They consists of a heterotetrameric nitrogenase part (NifD2K2) and a homodimeric dinitrogenase reductase part (NifH2) (Fig. 1). The dinitrogenase reductase part mediate electrons from the external electron donor, e.g. a ferredoxin or a flavodoxin, to the nitrogenase part [9]. The most common type of nitrogenase in diazotrophic cyanobacteria are Mo-nitrogenases, containing molybdenium in their active site. Additional nitrogenases can be found in e.g. Anabaena variabilis ATCC 29413 who, besides two Mo-nitrogenases, also contains a vanadium nitrogenase [28]. Mo-nitrogenases catalyze the reaction:

16
N2 + 8H+ +8e- + 16ATP 2NH3 + 16ADP + 16Pi + H2 Since 16 ATP are consumed per molecule of N2 fixed this process requires a lot of energy from the cell [9]. The amount of H2 produced per molecule of N2 fixed is not absolute and the electrons used for hydrogen production can be increased from a minimum of 25%, depending on external factors such as CO2 concentrations and light intensity [29]. Thus, the electron flow may be diverted towards higher H2 production rather than N2-fixation.
Hydrogenases Hydrogenases are multimeric enzymes, not uncommon in nature, that catalyze the simple reversible chemical reaction H2 ⇔ 2H+ + 2e- [30]. The cyanobacterial uptake hydrogenase consists of a small subunit (HupS) and a large subunit (HupL), encoded by hupSL [11,18,22] (Fig. 1). The bidirectional hydrogenase is a pentameric enzyme, encoded by hoxEFUYH, consisting of a diaphorase part (HoxEFU) and a hydrogenase part (HoxYH) [11,18,22] (Fig. 1). Hydrogenases can be classified into three groups depending on the metal content in their active center; [FeFe], [NiFe] and [Fe]-hydrogenases [20]. [NiFe] hydrogenases are further subdivided into four classes [20,31]: 1) Bacterial and Archaeal membrane bound uptake hydrogenases 2) a - (Cyano)bacterial uptake hydrogenases
b - Bacterial H2 sensors 3) a - Archaeal factor F420 reducing hydrogenases
b - Bacterial and Archaeal NADP reducing hydrogenases c - Archaeal methyl-viologen reducing hydrogenases d - Bacterial bidirectional NADP/NAD reducing hydrogenases
4) Bacterial and Archaeal energy converting, membrane associated, H2 evolving hydrogenases
All cyanobacterial hydrogenases belong to class 2a (uptake hydrogenases) or 3b (bidirectional hydrogenases) of [NiFe] hydrogenases [20,31], thereby containing a nickel (Ni) and an iron (Fe) atom in their active site [9,20]. The Fe atom in [NiFe] hydrogenases coordinates three, for biological systems unusual, ligands; one carbon monoxide molecule (CO) and two cyanide ions (CN-) [20,32]. The large subunit contains the active site where H2 is either split or formed. Hydrogen are proposed to enter or leave the hydrogenase large subunit through a hydrophobic channel connecting the active site with the surface of the molecule [11,20]. The small subunit mediates electron transport to and from the active site to redox partners and downstream applications through iron-sulphur (FeS) clusters [20,32].

17
Figure 1. Schematic picture of three enzymes directly involved in cyanobacterial hydrogen metabolism. At the expense of ATP, atmospheric dinitrogen (N2) is fixed by the nitrogenase into ammonia (NH3) and molecular hydrogen (H2) is produced as a by-product. The uptake hydrogenase recaptures the energy stored in the chemical bonds by oxidizing the H2 produced by the nitrogenase. If present, the bidirectional hydrogenase can either oxidize or evolve hydrogen using NADH and NAD+ as redox partners [10,18]. Subunits of the nitrogenase (NifHD2K2), the uptake hydrogenase (HupSL and the, in cyanobacteria so far unidentified, third subunit HupC) and the bidirectional hydrogenase subunits (HoxEFUYH) are shown. The dotted line separates the hydrogenase part (HoxHY) from the diaphorase part (HoxEFU) in the bidirectional hydrogenase.
Maturation of [NiFe] hydrogenases Hydrogenases are complex metalloenzymes and the assembly of the active site and the correct folding of the large subunit therefore requires assistance of seven accessory proteins; the Hyp-proteins (encoded by hypABCDEF) and a protease. The proteases are specific for one type of hydrogenases only and in cyanobacteria HupW and HoxW (encoded by hupW and hoxW) act on the uptake hydrogenase and the bidirectional hydrogenase respectively [20,32]. The hyp-genes (hydrogenase pleiotropic) and the Hyp-proteins have been studied mainly in Escherichia coli [32] but homologues have been found in cyanobacteria and are believed to have the same function. This has also been partially showed with mutational analyses in Synechocystis sp. PCC 6803 [9,33]. Since the hyp-genes have been found in only one copy in cyanobacterial genomes, irrespectively of if the cyanobacteria express the uptake hydrogenase, the bidirectional hydrogenase or both, it appears that the hyp-genes, in contrast to the proteases, act on both types of hydrogenases [18]. The hyp-genes in cyanobacteria are often clustered and located in proximity of the structural genes of either the uptake or the bidirectional hydrogenase [18].

18
Maturation of the hydrogenase large subunit The maturation process of the large subunit consists of formation of an apo-protein, supply of metals, assembly and integration of the active site and proteolytic cleavage of the C-terminal of the apo-protein (Fig. 2) [32]. While the CN-ligands are biosynthesized from carbamoyl phosphate by HypF, a carbamoyl transferase, at the HypF-HypE complex [32] the source of the CO-ligand is still not known [34]. The CN-ligand is transferred, via HypE, to the HypC-HypD complex where the Fe-part of the metallocenter (Fe(CN)2CO) is assembled before insertion into the apo-protein by HypC [34]. In addition, HypC acts as a chaperone by stabilizing the apo-protein in the open conformation until Ni is inserted [18]. Ni is bound by the zinc containing HypA, at the HypA-HypB complex, and inserted into the active site before proteolytic cleavage of the C-terminal by the hydrogenase specific protease [11,18,32]. Cleavage of the C-terminal tail occurs after the conserved motif DPCxxCxxH/R, leaving a histidine as the new C-terminal amino acid, and allows for structural reorganization and closure of the apo-protein [32,35] before association with the hydrogenase small subunit [36,37]. The exact mechanism for recognition of the different hydrogenases, and which part(s) of the protease determines specificity, are still unknown although Ni is known to be an important substrate recognition motif and the protease binds directly to the metal [38,39]. Hence, the insertion of Ni functions as a checkpoint and not until the insertion of is done the protease can act and cleave the C-terminal of the apo-protein [18,39]. In E. coli three amino acids (Glu16, Asp62 and His93) have been identified as important for Ni-binding of the hydrogenase specific protease HybD [38].
Maturation of the hydrogenase small subunit While the maturation process of the large subunit is to a large extent uncovered, the maturation process of the small subunit is still unclear. The small subunit of uptake hydrogenases contains three FeS cluster involved in electron transport [20]. For insertion of the FeS cluster into the small subunit mobilization of sulphur, by a cysteine desulfurase, and assembly of the FeS cluster on a scaffold are needed [20,40]. In cyanobacteria, the main FeS assembly scaffold protein is Nfu, a protein similar to the C-terminal domain of the FeS assembly scaffold protein NifU [41]. It is therefore likely that a protein involved in the assembly of FeS clusters to the cyanobacterial uptake hydrogenase small subunit is a NifU/Nfu like protein.

19
Figure 2. Schematic picture of the maturation process of the large subunit (HupL) of cyanobacterial uptake hydrogenases. The accessory proteins needed for insertion of metals and ligands into the active center and for correct folding of the protein are the Hyp-proteins (HypABCDEF) and a hydrogenase specific protease (HupW) [18]. The HypEF complex is involved in synthesizing the CN- ligands from carbamoyl phosphate and transferring them to the HypCD complex [32]. On the HypCD complex the iron atom (Fe) and the CO- and CN-ligands are assembled before insertion into the apo-protein [32]. HypC is involved in the insertion of the FeCN2CO complex while acting as a chaperone on the apo-protein by stabilizing the open formation [32]. The nickel atom (Ni) is delivered by the HypAB complex [32]. Once Ni is inserted into the active site HupW can cleave off the C-terminal tail before closure of the apoprotein and complex formation with the small subunit, HupS [37].

20
The model organisms Nostoc punctiforme ATCC 29133 Nostoc punctiforme ATCC 29133 (also known as Nostoc punctiforme PCC 73102) is a filamentous, heterocyst forming, N2-fixing cyanobacterium (Fig. 3). In addition to heterocysts, N. punctiforme can form akinetes and hormogonia. This strain harbors a nitrogenase and an uptake hydrogenase, but lacks the bidirectional hydrogenase (Table 1). N. punctiforme is an photoautotroph but can also grow heterotrophically [25]. The strain has an interesting origin as a symbiont to the cycad Macrozamia [42] where it during symbiosis exhibits decreased growth rate, high metabolism [43] and a higher heterocyst frequency - all desirable characteristics for biotechnological applications, such as hydrogen production. The genome is sequenced (http://genome.jgi-psf.org/nospu/nospu.home.html) with a size of 9.06 Mb, distributed on one chromosome and 5 plasmids [44], and both the transcriptome [45] and the proteome have been characterized [46,47]. The strain is relatively easy to cultivate and molecular methods are developed for the strain which makes it suitable as a model organism.
Table 1. Presence of uptake and bidirectional hydrogenases (H2ases) in the cyanobacterial strains Nostoc punctiforme ATCC 29133 and Nostoc sp. strain PCC 7120 are shown by listing the structural genes encoding the subunits of the enzymes. Precense of genes encoding accessory proteins (Hyp-proteins and hydrogenase specific proteases) involved in the maturation of the hydrogenase large subunits are also listed.
Strain Uptake H2ase
Bidirectional H2ase Hyp-proteins Protease
N. punctiforme hupSL - hypABCDEF hupW Nostoc PCC 7120 hupSL hoxEFUYH hypABCDEF hupW, hoxW
Nostoc sp. strain PCC 7120 Nostoc sp. strain PCC 7120 (also known as Anabaena sp. strain PCC 7120) is a filamentous, heterocyst forming, N2-fixing cyanobacterium (Fig. 3). Unlike N. punctiforme this strain can not form akinetes or hormogonia. Nostoc PCC 7120 harbors, in addition to a nitrogenase and an uptake hydrogenase, also a bidirectional hydrogenase (Table 1). Despite expressing two types of hydrogenases, only one set of hyp-genes is present in the genome. Nostoc PCC 7120 is an obligate photoautotroph [25]. The genome is sequenced (http://genome.kazusa.or.jp/cyanobase/Anabaena) with a genome size of 7.21 Mb, distributed on one chromosome and 6 plasmids [48], and the transcriptome [49] and the proteome have been characterized [50,51]. Nostoc PCC 7120 is, similarly to N. punctiforme, a suitable model organism since it is relatively easy to cultivate and methods for molecular and genetic studies are developed also for this strain.

21
Figure 3. Micrographs of two filamentous, heterocyst forming cyanobacterial strains. A) Nostoc punctiforme ATCC 29133 and B) Nostoc sp. strain PCC 7120. The scale bars on the images represents 5 μm. H: heterocyst, V: vegetative cell.
The uptake hydrogenase Cellular localization Even though the activity of the uptake hydrogenase is generally believed to be localized to the heterocysts, where the nitrogen fixation takes place, this belief is under discussion [18]. Is the enzyme expressed only in heterocysts or can it under some special growth conditions be expressed also in vegetative cells? If so, is it active? In Nostoc PCC 7120 the activity of the uptake hydrogenase were shown to be exclusively localized to heterocysts in aerobically grown filaments [52]. This localization of the uptake hydrogenase was recently confirmed [53]. However, in this strain a genomic reorganization occur upon transition from non N2-fixing conditions to N2-fixing conditions where an excision element, interrupting hupL, is removed enabling transcription of a functional transcript in the heterocysts [54]. Hence the strict confinement of the uptake hydrogenase to the heterocyst of Nostoc PCC 7120 is more or less expected. In N. punctiforme no such rearrangement of hupL occurs and immunolocalization studies have shown presence of the uptake hydrogenase in both vegetative cells and heterocysts [53,55,56].
Subcellular localization The subcellular localization of the uptake hydrogenase is also under debate. In other bacteria, the membrane bound [NiFe] uptake hydrogenase small subunit contain a Tat (twin-arginine translocation) signal peptide. This signal peptide serves as a signal for export of the fully matured and fully assembled heterodimer to the periplasm [20]. Cyanobacterial uptake hydrogenases lack this Tat-signal peptide and are therefore grouped with cytoplasmic H2-sensing hydrogenases instead of membrane bound uptake hydrogenases [20].

22
Moreover, the cyanobacterial uptake hydrogenases have an amino acid sequence more similar to the soluble sensor hydrogenases [18]. The cyanobacterial uptake hydrogenase does however possess H2 uptake activity, as shown by the H2 evolving, uptake hydrogenase-free mutant strain of N. punctiforme [57]. In other bacteria, such as Azotobacter vinelandii [58], the uptake hydrogenase is anchored to the periplasmic side of the cytoplasmic membrane by a third subunit, a b-type cytochrome believed to participate in electron transfer from the small subunit to the respiratory electron transport chain [20,58,59]. This subunit has not yet been identified in cyanobacteria, but as the cyanobacterial uptake hydrogenase is believed to be membrane associated the presence of such a subunit is likely [18,60]. The suggested membrane association was further strengthen by the recent finding of a membrane localization of the uptake hydrogenase in L. majuscula CCAP1446/4 [53]
Transcriptional regulation of the uptake hydrogenase Physical organization of the hupSL operon in N. punctiforme The small and the large subunit of the uptake hydrogenase are encoded by hupS (Npun_F0368) and hupL (Npun_F0369) respectively. The genes form a single transcript and hence share the same promoter [61]. In the promoter region, three consensus sequences, representing putative binding sites for regulatory proteins, have been identified together with a RNA polymerase binding site and a tsp (transcription start point) located 259 bp (base pairs) upstream of the start codon of hupS (Fig. 4) [61]. Downstream of hupL a putative transcription terminator has been identified [61]. One of the putative binding sites suggests binding of the transcriptional regulator NtcA to the hupSL promoter. This binding was later confirmed when NtcA were shown to bind to and retard a DNA fragment containing the N. punctiforme hupSL promoter in a gel shift assay [62]. The binding site identified is most similar to the first motif to be described for NtcA (TGT-N9 or 10–ACA) in Nostoc PCC 7120 [63]. The two other identified putative binding sites suggest binding of IHF (integration host factor). IHF is a DNA bending protein [64] known to be involved in transcriptional activation of nitrogenase genes in purple bacteria [65] and is also present upstream hydrogenase genes in Bradyrhizobium japonicum [66]. Proteins with high similarity to IHF in other organisms have so far not been identified in cyanobacteria. We can however not rule out the possibility that a protein with similar function as IHF and similar recognition sequence might be present in cyanobacteria.

23
The transcription regulator NtcA NtcA (nitrogen control) [67] is a transcriptional regulator belonging to the CRP (cyclic AMP receptor protein) family of regulatory proteins (also known as the CAP family; catabolite activator protein) [68,69] operating global nitrogen control in cyanobacteria [68,70]. NtcA controls the expression of a large number of genes involved in nitrogen assimilation by binding to their promoter regions and activate or repress their transcription [68]. In addition to atmospheric N2, cyanobacteria can use a number of different nitrogen sources, such as ammonium, nitrate, nitrite, urea and some amino acids [71,72]. Ammonium is the energetically most economical, and hence preferred, nitrogen source [71,72]. In the absence of ammonium NtcA promotes, in addition to its own expression, the expression of genes encoding proteins involved in assimilation of other nitrogen sources [68,73]. The metabolite 2-OG enhances the affinity of NtcA to DNA and is believed to be the link between C:N balance in the cell and NtcA mediated regulation [74,75]. Cyanobacteria lack the enzyme 2-OG dehydrogenase and hence the metabolic destiny for 2-OG is to serve as carbon skeleton for ammonium incorporation [71,72]. The synthesis of 2-OG depends on the carbon fixation and the consumption on the nitrogen assimilation, which makes the cellular concentration of 2-OG a direct indicator of the current C:N balance [71,72]. NtcA is transcribed early in heterocyst differentiation (4.5 to 6h after nitrogen depletion) and regulates the expression of the master regulator of cell differentiation, HetR [76]. Since HetR in turn upregulates expression of NtcA, the regulatory proteins appear to have a mutual dependency for expression [76]. In agreement, NtcA has been shown to be necessary for heterocyst differentiation in Nostoc PCC 7120 [77,78]. In the C-terminus NtcA contains a helix-turn-helix motif responsible for the interaction with DNA [70,72]. This DNA binding motif of NtcA, conserved in cyanobacteria, suggest that NtcA in different cyanobacterial strains bind to the same (or similar) motif in the promoter regions [71]. NtcA binds DNA as a dimer and hence the recognition sequence in the DNA is palindromic [71]. Different binding sites for NtcA have been reported [68]. The consensus sequence GTA–N8–TAC (where the GT-N10-AC subset appears to be most important) represents canonical NtcA activated promoters. These canonical NtcA activated promoters are similar to the bacterial Class II CRP activated promoters [79] and have an E. coli σ70-like -10 box in the form of TAN3T [68]. The NtcA binding site is centered at -41.5 bp upstream the tsp, overlapping or replacing the -35 box [25,70]. The sequence in one (or both) triplets can vary, along with increased or decreased spacer length, and are believed to result in a lower affinity of NtcA to the DNA [71]. Repressor NtcA binding sites have been found downstream of position -41.5 inhibiting promoter clearance or blocking access of the RNA polymerase by overlapping the tsp, the -10 box or the -35 box [68]. NtcA regulated

24
promoters can have complex regulation and harbor more than one NtcA binding site as well as binding sites for other regulatory proteins [71]. In addition, there are examples where NtcA acts as both activator and repressor of the same promoter [80].
Transcription regulators of the Cal family Cyanobacterial Cal proteins (cyanobacterial AbrB-like) belong to the AbrB family of transcription regulators (antibiotic resistance) and are subdivided into two clades, CalA and CalB [81]. In contrast to AbrB in other bacteria cyanobacterial Cal proteins contain the conserved DNA-binding region in their C-terminus instead of in their N-terminus [81]. The transcription factor AbrB has been extensively studied in Bacillus, where it is considered to be a global transition-state regulator due to its involvement (directly or indirectly) in the regulation of more than 60 different genes in a wide variety of metabolic and physiological processes [82,83]. Proteins belonging to the AbrB family of transcription regulators bind to promoters with different affinities and are assumed to recognize promoter regions based on their DNA 3D-conformation rather than through a consensus binding sequence [82,84]. Multimerization is necessary to form the DNA binding region of the protein [83,85] and is believed to play a role in the ability of AbrB to regulate promoters differently [83]. In addition, post-translational modifications of CalA in cyanobacteria have been suggested to influence the DNA binding affinity of the protein [86,87]. The functions of cyanobacterial Cal proteins are not clear but CalA has been associated with hydrogen metabolism [85,88], toxin production [86], cell division [89], oxidative stress responses [90] and repression of its own expression [85]. CalA appears to be essential since, despite attempts, no fully segregated knock-out mutant has been constructed [81,85,88]. CalB on the other hand has been successfully knocked-out [81] and has so far been connected to nitrogen uptake [81] and the carbon concentrating mechanism [91].

25
Aim of this thesis
The aim of this thesis was to, combining transcriptomics with proteomics, deepen the understanding about the transcriptional regulation, maturation and function of the uptake hydrogenase in filamentous, heterocystous cyanobacteria as well as to place the function of the uptake hydrogenase in a metabolic context. H2 can be produced by a number of hydrogenase or nitrogenase containing microorganisms, such as cyanobacteria, green algae, photosynthetic bacteria and fermentative bacteria. However, green algae and cyanobacteria are the only organisms capable of producing H2 from solar energy and water with no need for organic substrates as electron donors. Heterocystous cyanobacteria have an advantage compared to green algae since they can provide a natural microoxic environment for the O2 sensitive, H2 producing enzymes in their heterocysts. Heterocystous cyanobacteria are thus capable of producing H2 in a continuous culture, grown in the light and under aerobic conditions, while at the same time carrying on oxygenic photosynthesis. This eliminates the need to cycle the culture between conditions favorable for photosynthesis and conditions favorable for H2 production. The main drawback with cyanobacterial H2 production however, is that the cyanobacterial nitrogenases or hydrogenases are not very efficient enzymes. The cyanobacterial strain chosen for industrial H2 production must hence be a strain with more efficient enzymes, yet to be discovered, or more likely a genetically engineered strain. Some routes for creating a H2 producing cyanobacterial strain of commercial importance are; a) inactivation of the uptake hydrogenase, b) redirection of the electron flow in the cells to make more electrons available for H2 production, c) engineering of a more efficient bidirectional hydrogenase where the electron flow is directed towards hydrogen production, d) conversion of the uptake hydrogenases into a H2 evolving enzyme by turning the electron flow and e) introduction of a foreign more efficient hydrogenase. The uptake hydrogenase has been successfully inactivated in a number of cyanobacterial strains [54,57,92-94], therefore much work is currently focused on directing or redirecting the electron flow and introducing a fully active heterologous hydrogenases in a cyanobacterial host strain. For a foreign hydrogenase to function optimally in a new environment, and to genetically engineer an existing hydrogenase or redirect electron flow in the cells, we need detailed knowledge about the maturation,

26
regulation and function of the enzyme of interest. This knowledge include information about transcriptional control of the expression, the accessory proteins needed for correct assembly of the active enzyme as well as the function in the cells, i.e. the competing pathways and the redox partners - all of which have been studied in this thesis. In my work two cyanobacterial strains, Nostoc punctiforme ATCC 29133 and Nostoc sp. strain PCC 7120, have been used. Both strains are filamentous, heterocystous cyanobacteria. Although closely related, two of the main differences are that; a) N. punctiforme harbors only an uptake hydrogenase while Nostoc PCC 7120 harbors both an uptake hydrogenase and a bidirectional hydrogenase and b) N. punctiforme has a symbiotic origin while Nostoc PCC 7120 are a free-living strain. These similarities and differences make the hydrogen metabolism in these strains interesting to compare.

27
RESULTS AND DISCUSSION
Transcriptional regulation of the uptake hydrogenase Which are the key players in the transcriptional regulation of the uptake hydrogenase? (Paper I) In order to have full control over when, and in which amount, the hydrogenase in our genetically engineered strain should be expressed we need information about how the transcription is controlled. Even though it is known that the uptake hydrogenase is expressed during N2-fixing conditions [61] the exact regulation of the expression is unknown. Previously three putative binding sites for regulatory proteins have been identified in the promoter region of hupSL in N. punctiforme together with a tsp and a -10 and a -35 box [61]. These putative binding sites represent recognition sites for the transcriptional regulators NtcA and IHF [61]. NtcA binding sites have been found in the hupSL promoter in a number of other cyanobacteria, such as Anabaena variabilis ATCC 29413 [95], Nostoc sp. PCC 7422 [94], Lyngbya majuscula CCAP 1446/4 [96] and Gloeothece sp. ATCC 27152 [97], while IHF binding sites have so far only been found in the hupSL promoter in N. punctiforme [61] and L. majuscula [96]. The putative NtcA binding site appears to represent a true NtcA binding site since we could show that NtcA interacted specifically with a DNA fragment of the hupSL promoter from N. punctiforme containing the putative NtcA binding site and retarded it on a gel. This is in agreement with previous results [62]. In addition, NtcA has been shown to bind to the putative binding sites found in the hupSL promoter in A. variabilis [95], L. majuscula [96] and Gloeothece ATCC 27152 [97]. To investigate the importance of the putative NtcA and IHF binding sites and to identify other regulatory regions in the hupSL promoter, truncated versions of the N. punctiforme hupSL promoter (A-E) were fused with a reporter gene, gfp (encoding the fluorescent protein GFP; Green Fluorescence Protein) (Fig. 4). The constructs were cloned into a self-replicative vector and transferred into N. punctiforme by electroporation. The fluorescence intensities, and hence the promoter activities, of the resulting strains carrying the promoter-gfp fusion constructs were measured. The result showed that there were only minor variations in expression from the

28
four longer promoters constructs (A-D) and these differences were not always significant between experiments. However, the overall expression pattern was comparable between experiments. When looking at individual experiments, transcription from construct B seemed to be almost abolished in some experiments while showing low activity in others. This is interesting since the part of the hupSL promoter present in construct A, but deleted in construct B, harbors no known putative binding sites for regulatory proteins. Loss of the most upstream putative IHF binding site (construct C) and the NtcA binding site (construct D) seemed to have a weak positive respective weak negative effect on the transcription. This indicates that the importance of IHF and NtcA for the transcription of hupSL in N. punctiforme may be low. The result that NtcA did not have a key role in the transcriptional regulation of the uptake hydrogenase was a bit unexpected since NtcA controls the expression of a large number of genes involved in nitrogen fixation [68,70] and there is a strong connection between nitrogen fixation and uptake hydrogenase activity [9]. Furthermore, NtcA has been shown to regulate hupSL transcription in A. variabilis [95]. The results suggest additional regulation of the hupSL transcription in N. punctiforme by so far unidentified regulatory proteins and reflect the complexity of transcriptional regulation in cyanobacteria.
Surprisingly, the shortest promoter fragment, with the 5’ end located at -57 relative to the tsp, showed the highest transcription of all constructs in all experiments. This was additionally confirmed when measuring the bioluminescence from N. punctiforme cultures carrying the same promoter deletions fused to the reporter genes luxAB (encoding luciferase) instead of gfp (data not shown). The high expression level was not due to loss of heterocyst specificity, and hence high total expression from all cells, but represented localized expression in the heterocysts only. The short part of the promoter region present in construct E lacks the putative NtcA and IHF binding sites present in the longest promoter construct but is still sufficient to drive a strong expression of hupSL. The result suggests that an unknown transcription regulator binds this short promoter region or the untranslated leader sequence and promotes heterocyst specific transcription and that the region upstream this short promoter region harbors binding sites for regulatory proteins inhibiting transcription. It is not unlikely that this promoter region harbors an additional tsp undetected in the previous 5’ end mapping of the hupSL promoter region [61]. In cyanobacteria there are many examples where genes are transcribed from multiple promoters, e.g. ntcA [98] and the heterocyst differentiation genes hetC, devB [69] and hetR [99] in Nostoc PCC 7120. It have been shown, for both hetR and ntcA in Nostoc PCC 7120, that a combination of constitutive and inducible promoters contributes to a more fine tuned regulation and strong localized expression in differentiating heterocysts [69,100]. Moreover, it is suggested that the NtcA

29
dependent upstream promoters of hetC and devB are responsible for transcription early (3 hours) after nitrogen step down while the transcription from the HetR dependent downstream promoters are responsible for localized accumulation of transcripts in heterocysts [69]. A similar tandem promoter arrangement for the transcriptional regulation of hupSL in N. punctiforme is not unthinkable and additional putative NtcA binding sites can be found downstream of the tsp (data not shown). However, further experiments are needed to elucidate this possibility.
AGGCTTCGAGTCCTTTAGGCGGAATTCCCCTTTTGTCTGAAACTAGAGGCTAGGGATTAATGAT
GCTAGGGACTAGGAAAATTTTTCCCAATACCTAATACTTAATCCTCAATCCCCTAAATTGAGTGA
ATTAATTTTTTTACTATAAAACACTCTATTTCTAAAATTCTAGGGGGAAATTGCCAATATTAATGTC
AGCTTTAAACACCTTACATTTTGATAAGGTGTTCTGTATCATGACCTGACACAACGCAGTTCTTC
TATATTGAATTGATAACATCTTATATATTTAAGACCTAACTTTGTTATCAAAATATAACTATTTGCTTT
GAACTTAAAAATAAATTTATTTACTATTTACTTTTTTTGGCTAGTGCTGATATTTCTTAACGTACAA
GACTTCACCTTTAAAATCTTAGCCCATTTATATAGAAAAGTTGAGATTACAACTCTCTCAAAATGA
TTTAAAACAGCTATCTAAAATCAGTGTGCTTAGTTTGTTATTCAAGCTTTTGTGAAAAATTATGTT
CTTGATACAACCTTGTAAGCAATCTTAAGTAGATTTAAATTATTAGCAATGATTAATTTTTATTTAAC
AAAAGATAATTCACTAGAAAATCCACTAATATTAATGAATAAATATTGATAAATACGTGAGACTCTA
CAAGCATTATTTACTAATTCACGTATGGCAAACAATACAAAAACACCTAGCCCatg
A
B
C
D
E
WT
250bp
gfp/luxAB
hupS
Primers for Gel Shift Assay
putative NtcA bidning site putative IHF binding site
gfp/luxAB
gfp/luxAB
gfp/luxAB
gfp/luxAB695 bp
610 bp
559 bp
479 bp
316 bp
-436
-351
-300
-220
-57
751 bp
tsp -35 and -10 boxes
Figure 4. Detailed view of the nucleotides in the hupSL promoter region in Nostoc punctiforme ATCC 29133. Putative binding sites for regulatory proteins (IHF and NtcA) are shown together with the tsp and a -10 and a -35 box [61]. The primers used for gel shift assay are also indicated. Below the nucleotide sequence, the intergenic region between hupSL and its upstream gene is shown (WT) together with the five constructs where truncated versions of the hupSL promoter have been fused to the reporter genes gfp or luxAB (A to E). The total lengths of the truncated promoters (positive numbers) and the 5’ ends relative to the transcription start point (negative numbers) are given. The grey lines represent the hupSL promoter sequence and the white line the DNA sequence belonging to the self replicative vector carrying the truncated promoters, pSUN202 [101].

30
In conclusion: Although mobility shift assay showed that NtcA binds specifically to the hupSL promoter region in N. punctiforme, data obtained from the promoter deletion study suggest that transcription of the hupSL promoter in N. punctiforme is not strictly dependent on NtcA. The promoter deletion study therefore indicates that, besides NtcA, there must be other transcription factors involved in the regulation. Surprisingly, the shortest truncated promoter fragment, with the 5´end located at -57 bp in relation to the tsp, gave the highest GFP fluorescence and is apparently enough for a high expression of hupSL. The result, reflecting the complexity of cyanobacterial transcriptional regulation suggest interaction of positively acting regulatory proteins with this short promoter sequence or with the regions downstream of the tsp, as well as interaction of negatively acting regulatory proteins with the region upstream this short promoter.
Maturation of the uptake hydrogenase Which are the accessory proteins involved in the maturation of the hydrogenase small subunit? (Paper I, II and III) One of the routes to a genetically engineered cyanobacterial strain with high H2 production rate is introduction of a foreign, more efficient hydrogenase. In order to successful express a fully active heterologous hydrogenase in cyanobacteria we need to know which accessory proteins should be co-expressed with the foreign hydrogenase in the host strain. Information about the maturation process of hydrogenases and the additional proteins required for their maturation is thus essential. While the maturation processes of the hydrogenase large subunit in large parts have been uncovered, much of the maturation process of the small subunit, e.g. insertion of FeS clusters, remains unknown. Proteins involved in the maturation of the small subunit have been identified in aerobic bacteria, e.g. HupGHIJ in Rhizobium leguminosarum bv. viciae strain UPM791 [101,102]. The presence of hupGHIJ appears to be connected to oxygen since homologues can be found in other aerobic, [NiFe]-type uptake hydrogenase containing bacteria such as Azotobacter vinelandii, Rhodobacter capsulatus, Ralstonia eutropha H16, Azotobacter chrooccum and Bradyrhizobium japonicum [21,103,104] but are absent in strictly anaerobic bacteria [21,102]. The proposed functions of HupGHIJ in R. leguminosarum and of the HupGH homologues, HoxOQ, in R. eutropha are protecting the FeS clusters from oxygen, maintaining protein stability of the uptake hydrogenase small subunit precursor as well as preventing premature translocation of the precursor to the periplasm by binding to and masking the Tat-signal peptide [102,105]. The genes hupGHIJ, encoding HupGHIJ, caught our interest due to their genomic localization upstream, and in the same direction as, the hyp-genes and in

31
between the uptake hydrogenase structural genes and the hyp-genes. In the large cluster of genes involved in hydrogen metabolism in R. leguminosarum these genes form a subcluster consisting of five genes, hupGHIJK [101], preceded by a promoter (P3) upstream hupG [106]. Interestingly, a set of five ORFs, encoding unknown proteins, could be found in the same genomic localization in Nostoc PCC 7120, i.e. upstream, and in the same direction as, the hyp-genes and in between the uptake hydrogenase structural genes and the hyp-genes. The genomic localization of homologues to these ORFs appears to be conserved in other uptake hydrogenase containing cyanobacteria, such as the filamentous, heterocyst forming, N2-fixing strains N. punctiforme, A. variabilis (http://genome.jgi-psf.org/anava/anava.home.html) and Nodularia spumigena CCY9414 (AAVW01000003) as well as the filamentous, non-heterocyst forming, N2-fixing strains Trichodesmium erythraeum IMS101 (http://genome.jgi-psf.org/trier/trier.home.html) and L. majuscula (AF368526) (Fig. 5) but are absent in strains harboring only the bidirectional hydrogenase. Moreover, Npun_R0363, a homologue to one of the ORFs upstream the hyp-operon in Nostoc PCC 7120, has been shown to be a part of the hyp-operon in N. punctiforme, forming transcript with the hyp-genes [107]. Since gene clustering is often an indicator for physical or functional interaction of the encoded proteins [32], we suggested that these ORFs upstream the hyp-genes may be involved in the maturation of the uptake hydrogenase small subunit in cyanobacteria. FeS clusters in the hydrogenase small subunit fulfill an important function since they mediate electron transport between the active site and the hydrogenase redox partner [20,32]. However, how the FeS clusters are inserted into HupS is still unknown. In the FeS biosynthesis systems, identified so far in bacteria, two of the key players are a cysteine desulfurase, such as IscS, SufS and NifS, and a FeS cluster scaffold protein, such as IscU, IscA, SufA, and NifU [20,40]. The cyanobacterial protein Nfu is highly similar to the C-terminal domain of NifU and is considered to be the essential FeS cluster scaffold protein in cyanobacteria [41]. Therefore, a protein involved in the assembly of FeS clusters for the uptake hydrogenase small subunit could be expected to be similar to Nfu. Interestingly, in silico analyses showed that alr0692, one of the five ORFs upstream the hyp-genes in Nostoc PCC 7120, contains a domain similar to the C-terminal of NifU. Moreover, the domain is also present in the alr6092 homologue Npun_R0364 in N. punctiforme. The finding that these proteins, encoded by one of the ORFs upstream the hyp-genes in Nostoc PCC 7120 and the corresponding homologue in N. punctiforme, just like Nfu harbor a domain similar to the C-terminal of NifU supports the theory that these genes encodes proteins involved in the maturation of HupS. Other domains, known to be involved in protein-protein interactions (such as NHL [64] repeats and

32
TRP repeats [108,109]) found in two the ORFs upstream the hyp-genes (alr0691 and alr0693) in Nostoc PCC 7120 are also conserved in their corresponding homologues (Npun_R0367 and Npun_0363) in N. punctiforme. It is not surprising to find domains involved in protein-protein interactions in proteins believed to be involved in maturation. In the maturation process of HupL, direct contact between the Hyp-proteins and between the Hyp-proteins and the apo-protein occurs [32] and the situation is probably similar for the maturation process of HupS. In R. leguminosarum hupGHIJK are co-transcribed from a single promoter upstream hupG [106]. To investigate if the ORFs upstream the hyp-genes are co-transcribed and under the same regulation, or transcribed from different promoters and possibly regulated differently, the upstream regions of the ORFs upstream the hyp-genes were mapped for tsps using 5´RACE. When mapping 5' regions for the five ORFs upstream the hyp-genes a tsp was found upstream alr0689 in Nostoc PCC 7120 and upstream Npun_R0367 in N. punctiforme. Moreover, analysis of the transcripts from these promoters, using RT-PCR (Reverse Transcriptase PCR), showed that the five ORFs upstream the hyp-genes can be transcribed together with the hyp-genes forming an extended hyp-operon in both organisms. This supports the idea that the gene products from the hyp-genes and the ORFs upstream the hyp-genes fulfill similar functions e.g. are involved in maturation of the uptake hydrogenase. In addition, a tsp was found upstream hypC and hypF in Nostoc PCC 7120. However, no tsps have been found upstream these genes in N. punctiforme [107]. Since Nostoc PCC 7120, in addition to an uptake hydrogenase, also harbors a bidirectional hydrogenase one explanation for the presence of these extra tsps could be a need for additional regulation of the hyp-operon gene products when a bidirectional hydrogenase is present. Proteins involved in maturation of the uptake hydrogenase small subunit, must co-exist with HupS. It is therefore not unlikely that the expressions of the genes coincide. In order to elucidate if the ORFs upstream the hyp-genes are transcribed under the same conditions as the uptake hydrogenase, RNA was preparated from N. punctiforme and Nostoc PCC 7120 cultures 0, 24, 48 and 72 hours after nitrogen depletion, and the transcript level patterns studied. These transcript level patterns were compared to those for genes known to be involved in hydrogen metabolism, such as the uptake hydrogenase and nitrogenase structural genes, hupS and nifH, and the maturation genes, hypC and hypF. 23S, encoding the ribosomal large subunit, was used as a control for a non-induced transcript. As expected, transcript levels of hupS, nifH, hypC and hypF were readily upregulated 24 hours after nitrogen depletion in both N. punctiforme and Nostoc PCC 7120. Interestingly, all five ORFs upstream the hyp-genes in both organisms showed similar transcript level patterns as hupS, nifH, hypC and hypF with

33
upregulation of transcript levels 24 hours after a shift from non N2-fixing to N2-fixing conditions. Transcript levels of the ORFs upstream the hyp-genes peaked at 48 hours in N. punctiforme and at 72 hours in Nostoc PCC 7120. The control 23S, representing a non-induced transcript, showed as expected consistent high transcript levels for all time points tested, validating the transcript level patterns for the other genes. Analysis of the transcript level patterns revealed induction of transcripts of hupS and the ORFs upstream the hyp-genes 24 hours after nitrogen step down showing that the transcripts do indeed co-exist not only under the same growth conditions but also in time. These results support the suggestion that these ORFs may be involved in the maturation of the uptake hydrogenase small subunit. In conclusion: A set of genes, encoding unknown proteins, have been identified in the cyanobacterium Nostoc PCC 7120. These five ORFs are found in between hupSL and the hyp-genes and are located upstream of, and in the same direction as, the hyp-genes. Homologues can be found in the same genomic localization in other filamentous, N2-fixing cyanobacteria, such as N. punctiforme but are missing in strains unable to fix N2. Interestingly, genes involved in the maturation process of the uptake hydrogenase small subunit in aerobic bacteria are also located in between the uptake hydrogenase structural genes and the maturation genes in the same direction as the hyp-genes. Conserved regions in the ORFs upstream the hyp-genes in Nostoc PCC 7120 involved in protein-protein interactions and FeS assembly are retained in their corresponding homologues in N. punctiforme. The ORFs upstream the hyp-genes in both N. punctiforme and Nostoc PCC 7120 showed similar changes in transcript levels after nitrogen depletion as genes known to be involved in hydrogen metabolism and are transcribed together with the hyp-genes forming an extended hyp-operon in both strains. In agreement a tsp has been identified upstream the extended hyp-operon in both N. punctiforme and Nostoc PCC 7120. Together the results support the suggestion that these ORFs are involved in cyanobacterial hydrogen metabolism, putatively in the maturation of the small subunit of the uptake hydrogenase.
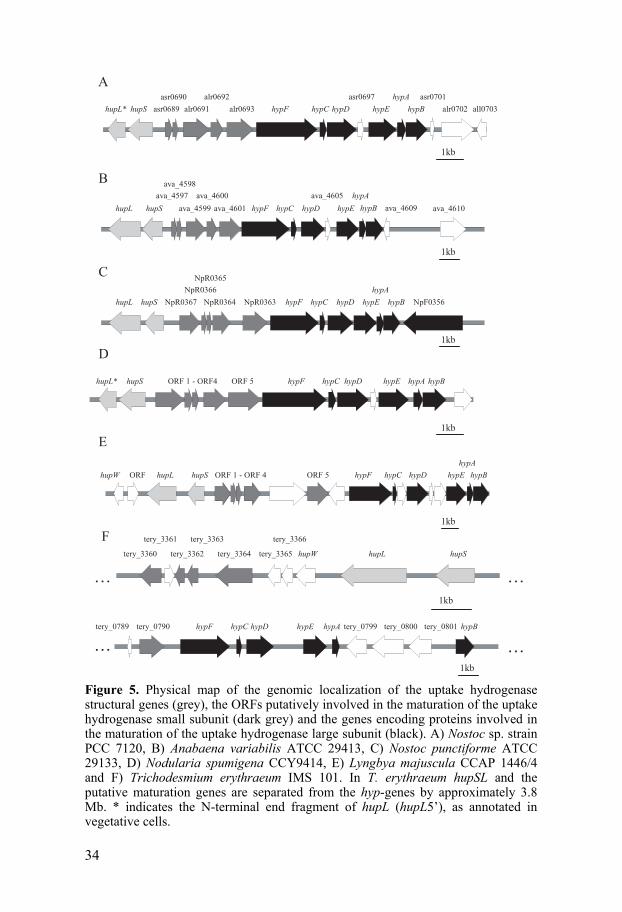
34
Figure 5. Physical map of the genomic localization of the uptake hydrogenase structural genes (grey), the ORFs putatively involved in the maturation of the uptake hydrogenase small subunit (dark grey) and the genes encoding proteins involved in the maturation of the uptake hydrogenase large subunit (black). A) Nostoc sp. strain PCC 7120, B) Anabaena variabilis ATCC 29413, C) Nostoc punctiforme ATCC 29133, D) Nodularia spumigena CCY9414, E) Lyngbya majuscula CCAP 1446/4 and F) Trichodesmium erythraeum IMS 101. In T. erythraeum hupSL and the putative maturation genes are separated from the hyp-genes by approximately 3.8 Mb. * indicates the N-terminal end fragment of hupL (hupL5’), as annotated in vegetative cells.

35
Which are the regulatory proteins interacting with the extended hyp-operon promoter region? (Paper II) In a cyanobacterial strain, genetically engineered for a high H2 production rate, it might be desirable to have a high expression of the H2 producing enzyme. When expressing a foreign hydrogenase at a high level, or overexpressing an enhanced endogenous hydrogenase, the maturation proteins must probably also be expressed at a higher level in order to get functionally active enzymes. Knowledge about the transcriptional regulation of the proteins involved in the maturation process of the chosen hydrogenase is therefore important. To search for transcription factors interacting with the promoter region of the extended hyp-operon DNA affinity assays were carried out. Interestingly, the DNA sequence forming the upstream region of the extended hyp-operon also forms the upstream region of the hupSL operon (Fig. 5). The promoter regions of hupSL and the extended hyp-operon are hence located in the same stretch of DNA. Investigation of this region with a DNA-protein binding assays will give information not only of regulatory proteins interacting with the promoter region of the extended hyp-operon but also regulatory proteins interacting with the promoter region of hupSL. This is highly interesting since the previous hupSL promoter deletion study indicated that the hupSL promoter is subjected to additional regulation by unknown regulatory proteins but also poses the problem that it is impossible to tell which of the promoters the proteins interacting with the hupSL/extended hyp-operon upstream region are regulating. DNA fragments covering the hupSL - Npun_R0367 and hupSL - asr0689 intergenic regions were amplified by PCR from genomic DNA of N. punctiforme and Nostoc PCC 7120 respectively. The resulting DNA fragments were covalently bound to magnetic beads and incubated with total protein extract from respective strain. The proteins interacting with the promoter regions were eluted and run on a SDS-PAGE gel before identification by MS (mass spectrometry). The identified peptides belonged to the transcription factors CalA and CalB. These transcription factors have recently been connected to regulation of the bidirectional hydrogenase in Synechocystis PCC 6803 [88], the maturation gene hypC in Nostoc PCC 7120 [85] and nitrogen metabolism in Synechocystis PCC 6803 [81]. It is therefore not unlikely that they are involved in the regulation of the uptake hydrogenase, the extended hyp-operon or both. Whether CalA and CalB bind the promoter region directly, or if one protein binds the DNA and the second protein is interacting with the DNA binding protein is however not possible to determine with this method. In addition, further experiments are needed to test if CalA and CalB act as activators or inhibitors of transcription and if they regulate transcription of the extended hyp-operon, hupSL or both. To test the specificity of the CalA interaction with the hupSL/extended hyp-operon upstream regions EMSAs (electrophoretic mobility shift assays) were

36
performed with the hupS/Npun_R0367 upstream region, amplified by PCR from N. punctiforme genomic DNA, and histidine-tagged CalA protein purified from Nostoc PCC 7120. The result showed that the interaction of CalA to the hupS/Npun_R0367 upstream region was specific and confirmed the result obtained by the DNA affinity assay, strengthening the indication that CalA is acting as a regulatory protein on the transcription of the extended hyp-operon, hupSL or both.
Table 2. The ORFs located upstream the hyp-genes in Nostoc sp. strain PCC 7120 and their homologues in Nostoc punctiforme ATCC 29133.
Nostoc PCC 7120 asr0689 asr0690 arl0691 arl0692 arl0693
N. punctiforme Npun_R0366 Npun_R0365 Npun_R0367 Npun_R0364 Npun_R0363
In conclusion: DNA binding assays showed interaction of two cyanobacterial transcription factors, CalA and CalB, with the hupSL/extended hyp-operon upstream region in N. punctiforme and Nostoc PCC 7120. In addition, gel shift assays showed that the interaction of CalA with the hupSL/extended hyp-operon upstream region in N. punctiforme was specific. Binding of CalA, newly associated with hydrogen metabolism, and CalB, associated with nitrogen metabolism, to the hupSL/extended hyp-operon usptream region indicates a role of these transcription factors in the transcriptional regulation of the extended hyp-operon and/or hupSL in filamentous, heterocystous cyanobacteria.
How are the uptake hydrogenase specific proteases transcribed, and what confers the specificity? (Paper IV) As discussed above, in order to get a fully active heterologous enzyme information about which accessory proteins should be co-expressed with the hydrogenase in the host strain is of great importance when introducing a foreign hydrogenase in cyanobacteria. The hydrogenase specific proteases are accessory proteins involved in the maturation of the large subunit [18,32]. As a last step in the maturation process they act on the apo-protein of HupL, cleaving off the amino acids forming the C-terminal tail [18,32]. Unlike the Hyp-proteins, these proteases are considered to be hydrogenase specific [37,110]. For example, while expressing both an uptake hydrogenase and a bidirectional hydrogenase Nostoc PCC 7120 harbors only one set of Hyp-proteins but two types of proteases, HupW and HoxW, acting on the uptake hydrogenase and the bidirectional hydrogenase respectively. The uptake hydrogenase specific proteases in N. punctiforme and Nostoc PCC 7120, identified by in silico analysis, are located independently of hupSL or the hyp-genes in the genomes and have been shown to be transcribed under both N2-fixing and non N2-fixing conditions indicating a

37
different regulation than for hupSL and the hyp-genes in both strains [111]. To investigate if hupW is transcribed alone or co-transcribed with other genes the 5´end of the transcript containing hupW were mapped and the length of the transcript analyzed by Northern blot in N. punctiforme and Nostoc PCC 7120. Northern blot analysis of the expression of hupW in N. punctiforme showed presence of a single transcript (approximately 1300 nt) only under N2-fixing conditions. The absence of transcript in N. punctiforme under non N2-fixing condition is in contradiction to a previous study where transcript of hupW in N. punctiforme were identified with RT-PCR during both non N2-fixing and N2-fixing conditions [111]. The size of the transcript corresponds to the theoretical size of hupW forming an operon with the upstream gene Npun_F0373 and the presence of such a transcript was also confirmed with RT-PCR. In agreement, a putative tsp was found, using 5’ RACE, upstream Npun_F0373 but not immediately upstream hupW. In the promoter region a putative -10 and -35 box were identified together with a putative NtcA binding site. In contrast to the result found for N. punctiforme, Northern blot revealed the presence of two transcripts in Nostoc PCC 7120; one shorter (approximately 500 nt) present under both N2-fixing and non N2-fixing conditions and a longer (approximately 1600 nt) present only under N2-fixing conditions. The length of the shorter transcript corresponds to the size of hupW while the length of the longer transcript corresponds to the theoretical size of hupW and the upstream gene alr1422 forming an operon. The presence of such an operon was also confirmed with RT-PCR showing that the two genes were co-transcribed. The search for 5’ ends revealed a tsp upstream hupW and in the promoter region a putative -10 and -35 box was identified together with two imperfect putative NtcA binding sites. In addition, two conserved regions, one of them resembling a putative IHF binding site, were identified in the promoter region. Two additional tsps were also identified upstream hupW inside arl1422. In silico analysis of alr1422 identified a novel putative translational start site 115 bp downstream the earlier described start site. This new translational start site fits better with the position of the two putative tsp which would then be located upstream of, instead of inside, alr1422. In the putative promoter region of alr1422 a -10 and a -35 box were identified together with two putative NtcA binding sites. From the results of the Northern blots and the 5’ mappings it appears that hupW in both N. punctiforme and Nostoc PCC 7120 is transcribed together with its upstream gene under N2-fixing conditions. These genes, Npun_F0373 and alr1422, are not homologous to each other and both encode unknown proteins. Thus, the functions of Npun_F0373 and alr1422, and the reason for them being co-transcribed together with the proteases are highly interesting and deserve further attention. In an extensive evolutionary study, including more than 80 different microorganisms, it was revealed that the large and the small subunit of

38
[NiFe] hydrogenases have evolved together and have been strongly connected for probably all their evolutionary history [21]. This raised the question whether a common evolutionary relationship also could be found between the hydrogenase large subunit and its hydrogenase specific protease. In order to investigate the relationship of hydrogenases and their specific proteases in different microorganisms a phylogenetic tree was constructed using both PAUP and MrBayes analysis. The resulting tree was a non-rooted tree since no suitable outgroup has been found for proteases. Interestingly, the resulting subgroups of the tree correlates with groups previously reported for [NiFe] hydrogenases [20,31]. These protease groups are; 1) Bacterial proteases (specific for group 1 hydrogenases) 2) Cyanobacterial proteases of HupW -type (specific for group 2
hydrogenases) 3) Bacterial and Archaean proteases
a) Archaean proteases (specific for group 3a hydrogenases) d) Bacterial proteases of HoxW-type (specific for group 3d hydrogenases)
4) Bacterial and Archaean proteases of Hyc-type (specific for group 4 hydrogenases)
HupW-type proteases and HoxW-type proteases act on different types of hydrogenases and are also clustered in different groups in the phylogenetic tree. To investigate what determines the specificity of the proteases amino acid alignments of HupW and HoxW from different cyanobacteria were performed and conserved regions, specific for each protease type, mapped. The study showed that one of the conserved regions [38,112] found in HoxW-type proteases differed when compared to HupW-type proteases. In HoxW-type proteases this region consists of the sequence H(Q/I)L (from now on referred to as the HOXBOX region) while among the HupW-type proteases the sequence is replaced by D(C/G/F)GT. In order to better understand the importance of these two different but conserved regions in silico analysis were performed of the 3D-model of the HupW-type protease HybD from E. coli and the putative 3D-models of HoxW and HupW from Nostoc PCC 7120, using the online program SWISS-MODEL (http://swissmodel.expasy.org). The study showed that the conserved residues appears to be clustered around the D(C/G/F)GT/HOXBOX region and two of the suggested Ni-binding residues (Glu16 and His93 in E. coli HybD [38]) for all three proteins. In addition, along α-helix 1, β-sheet 2 and α-helix 4 [38] conserved areas on the surface could be found in HupW and HybD. Protein-protein docking experiment with HoxW from Nostoc PCC 7120, HybD from E. coli and the HupW-type protease HynC from Desulfovibrio vulgaris str. Miyazaki F together with their corresponding hydrogenase large subunit, using the docking algorithm BiGGER, suggested

39
that the D(C/G/F)GT/HOXBOX region and the areas around α-helix 1, β-sheet 2 and α-helix 4 are in close contact with the large subunit. The HOXBOX region has previously been suggested to act as the catalytic site for proteolytic cleavage or to be involved in substrate binding [38,112]. However, the discovery of the two variants of the conserved region together with that the docking studies places the D(C/G/F)GT/HOXBOX region in a position unsuitable for C-terminal cleavage favors a role of the regions in substrate binding, recognition and possibly protease specificity. In conclusion: Transcript analysis of the uptake hydrogenase specific proteases in N. punctiforme and Nostoc PCC 7120 revealed that the uptake hydrogenase specific proteases are mainly transcribed under N2-fixing conditions. This transcription pattern follows the transcription pattern of the hydrogenase they cleave. In addition, during N2-fixing conditions the two hupW in N. punctiforme and Nostoc PCC 7120 are transcribed together with their non homologous upstream genes (Npun_F0373 and alr1422 respectively). In agreement, putative tsps were found upstream alr1422 in Nostoc PCC 7120 and upstream Npun_F0373 in N. punctiforme. The importance of this finding is unknown and the function of Npun_F0373 and alr1422 deserves further investigation. In silico analysis revealed the presence of putative binding sites for NtcA in the promoter regions, indicating that NtcA might be responsible for the co-ordination of hupW and hupSL transcription to N2-fixing conditions. When comparing the evolution of [NiFe] hydrogenases with the evolution of proteases a clear resemblance between the two phylogenetic trees is found indicating a co-evolution between the hydrogenase and its hydrogenase specific protease. Amino acid alignment of proteases revealed a conserved region existing in two different variants, the D(C/G/F)GT region and the HOXBOX region, when comparing HupW-type proteases to HoxW-type proteases. Protein-protein docking studies showed that the hydrogenase large subunit and its hydrogenase specific protease appear to have a close physical interaction during the cleavage event and suggest a role of the D(C/G/F)GT/ HOXBOX region in hydrogenase binding and recognition as well as hydrogenase specificity. The physical interaction between the hydrogenase and its protease could be responsible for the selectivity of the protease and would explain the similarity seen among the protease and the hydrogenase phylogenetic trees. In addition, the phylogenetic trees indicate that this interaction between hydrogenases and their hydrogenas specific proteases might be of an ancient origin.

40
Function of the uptake hydrogenase What is the subcellular and cellular localization of the uptake hydrogenase? (Paper I and V) To gain maximum efficiency when introducing foreign H2 producing enzymes in heterocyst forming cyanobacteria, the O2 sensitive enzymes should be expressed solely in the heterocysts. This can be done by using endogenous promoters believed to be active in heterocysts only. However, endogenous promoters are not always strong promoters and when expressing a foreign hydrogenase a high amount of the enzyme is probably desired. Furthermore, the regulation of the endogenous promoter might be unidentified. Knowing the motif conferring heterocyst specificity would enable us to achieve high, heterocyst specific expression of selected enzymes by integrating this motif into stronger promoters commonly used to overexpress proteins in the chosen strain. Although the uptake hydrogenase activity in heterocystous cyanobacteria, grown in aerobic and N2-fixing conditions, is believed to be mainly confined to heterocysts, the exact cellular and subcellular localization of the uptake hydrogenase in cyanobacteria is still unclear [18]. From the promoter deletion study, where truncated versions of the hupSL promoter region in N. punctiforme were fused to the reporter gene gfp, we saw that the localization of GFP fluorescence from all constructs were heterocyst specific (A-E, Fig. 4). Interestingly, the shortest promoter fragment, with the 5’ end of the promoter located at -57 bp upstream of the tsp, also retained heterocyst specific expression. The result indicates that, unless the hupSL transcript is subjected to post-transcriptional regulation, the motif conferring heterocyst specificity appears to be located in the short promoter region starting -57 bp upstream the tsp. If the region harboring the motif conferring heterocyst specificity could be narrowed down further to an even shorter DNA sequence, this would enhance the identification of the motif itself. Knowing the motif would be very useful when introducing foreign, more efficient but O2 sensitive hydrogenases into heterocystous cyanobacteria. From the study with truncated versions of the hupSL promoter fused to gfp it appears that the uptake hydrogenase is expressed specifically in the heterocysts of N. punctiforme. This is somewhat in contradiction to results obtained from immunolocalization studies where the uptake hydrogenase was detected in both vegetative cells and heterocysts [53,55,56]. However, it is important to keep in mind that transcription and localization of a transcript does not always translate to the presence and final localization of the protein. Hence, to further study the cellular localization of the uptake hydrogenase and to get clues of its subcellular localization we constructed a HupS-GFP fusion protein on a self replicative vector and introduced it into N.

41
punctiforme by electroporation. The GFP fluorescence from the transformed cultures was studied 0, 24, 48 and 96 hours as well as one week after induction of heterocysts by nitrogen depletion. In agreement with the previous result, when measuring GFP fluorescence from truncated versions of the hupSL promoter, the GFP fluorescence, and hence the cellular localization of HupS, was observed only in heterocyst for all time points studied. Unless GFP is physically hampering a putative transport of HupS, the result indicates a heterocyst specific localization of the uptake hydrogenase in N. punctiforme. The uptake hydrogenase is an oxygen sensitive enzyme and is quickly inactivated by O2 [18]. The restriction of the transcription and localization of this enzyme to heterocysts, in a photosynthesizing culture grown under aerobic conditions, therefore seems reasonable.
In conclusion: The promoter deletion study, using gfp as reporter gene, showed that the expression of all the truncated versions, of the hupSL promoter in N. punctiforme were heterocyst specific. The discovery that the shortest promoter fragment, with a 5’ end located at -57 bp in relation to tsp, retained heterocyst specific expression is highly interesting because it indicates that the information that determines heterocyst specific gene expression might be confined to this short DNA sequence. Additional studies, where the fluorescence of a HupS-GFP fusion protein was observed solely in the heterocysts of N. punctiforme, imply a heterocyst specific localization of the HupS protein. Together the two studies suggest that the transcripts of hupSL, as well as the uptake hydrogenase, are confined specifically to heterocysts in filamentous, heterocystous cyanobacteria.
How do the cells reprogram their metabolism when the uptake hydrogenase is no longer present? (Paper VI) In a heterocystous cyanobacterial strain with a H2 production rate of commercial practical use we want to direct electron flow away from competing pathways and towards the H2 producing enzyme. In addition, the uptake hydrogenase must be inactivated. Information about redox partners, to which cellular processes the uptake hydrogenase donates electrons and how the cells compensate for the loss of uptake hydrogenase generated electrons when the uptake hydrogenase is inactivated can give valuable clues how to redirect the electron flow for higher H2 production. The uptake hydrogenase has been successfully knocked-out by gene mutation in a number of cyanobacterial strains, such as N. punctiforme [57], Nostoc PCC 7120 [54,93], A. variabilis [92], and Nostoc sp. PCC 7422 [94]. All these mutant strains released significant amounts of H2 compared to their respective WT (wild type) strain while the rate of N2-fixation was

42
comparable (similar or higher) to WT N2-fixation, despite the loss of reducing power due to release of H2 to the environment [29,93,94,113]. Hence, to be able to keep up the N2-fixation rate compensatory mechanisms must be activated in the cells. By identifying the metabolic processes involved in adaptation to N2-fixation when the uptake hydrogenase is absent, we hoped to get information about basic functions of the uptake hydrogenase and clues how to metabolically re-engineer cyanobacteria for improved H2 production as well as how to adjust growth conditions to be more favorable for H2 production. The metabolism of N. punctiforme and NHM5 (the corresponding Nostoc punctiforme Uptake Hydrogenase Deficient Mutant) [57], were compared using a combination of two quantitative proteomic approaches, iTRAQ (isobaric Tag for Relative and Absolute Quantification) and 2DE (2- Dimensional Gel Electrophoresis). To be able to correlate proteomics data with phenotypic effects the rates of growth, nitrogenase activity and H2 evolution were established. The growth rate of N. punctiforme and NHM5, measured by OD (Optical Density) at 750 nm, were similar when grown in medium supplemented with combined nitrogen while NHM5 showed a decreased growth rate compared to the WT 6 days after nitrogen depletion. As shown earlier [29] the nitrogenase activity, measured by acetylene reduction, were similar in WT and NHM5 until 96 hours after nitrogen depletion where the activity was higher in NHM5. Analysis of the H2 concentration in the head space, using GC (Gas Chromatography), showed, in agreement with earlier reports [29], merely trace amounts of H2 evolution from WT while H2 evolution of NHM5 followed the nitrogenase activity, peaking at 40 hours after nitrogen depletion. The reduced growth rate of the uptake hydrogenase deficient mutant during N2-fixing conditions confirms that release of H2 to the environment represents a loss of energy for the cells and affects the organism negatively. To study how NHM5 manages to keep the N2-fixation rate high by altering its metabolism, iTRAQ and 2DE analysis were carried out where the proteome of whole filaments, as well as purified heterocysts, were analyzed comparing NHM5 to WT during N2-fixing conditions. In the iTRAQ and 2DE analysis 89 proteins were upregulated in heterocysts of NHM5 compared to WT while 65 proteins were downregulated. In whole filaments 58 proteins were upregulated in NHM5 compared to WT while 65 were downregulated. Only 10 of the proteins identified with 2DE overlapped with the result from the iTRAQ analysis, showing the complementarity of the two methods. This is in agreement with earlier reports [114-116]. From the proteins identified in this study 177 have previously been identified when the heterocyst specific metabolism was mapped in N. punctiforme [46]

43
Figure 6. Schematic picture of metabolic processes important or specific for the functions of vegetative cells and heterocysts in filamentous, heterocyst forming cyanobacteria. PSI/PSII: Photosystem I and II, OPP pathway: oxidative pentose phosphate pathway, SusA: sucrose synthase, TCA pathway: tricarboxylic acid pathway, 2-OG: 2-oxoglutarate, GS: glutamine synthetase, UH2ase: uptake hydrogenase, N2ase: nitrogenase, PM: plasma membrane, PG: peptidoglycan layer, OM: outer membrane, HEG; heterocyst envelope glycolipids, HEP; heterocyst envelope polysaccharides
In NHM5 the gene encoding the large subunit of the uptake hydrogenase is disrupted by insertional mutagenesis. The disruption caused not only low abundance of HupL in NHM5 but interestingly also lowered abundance of HupS. Despite the lower abundance of HupS, transcript levels of hupS, studied by RTqPCR (Reverse Transcriptase quantitative PCR), were higher in NHM5 than in WT suggesting a positive effect of higher intracellular H2 concentrations on hupSL transcription together with posttranscriptional regulation of hupS or instability of free HupS when the amount of HupL is insufficient.
Among the differently regulated proteins involved in nitrogen metabolism GS (encoded by Npun_R5387) was found in higher abundance in heterocysts of NHM5. Since GS is induced during high C:N ratios where nitrogen is in limited supply [78,117-119], this could indicate that the absence of a functional uptake hydrogenase causes reduced nitrogen availability in NHM5. The reduced nitrogen availability might be connected to reduced nitrogenase activity, caused by loss of H2 derived reducing equivalents, or the decreased abundance of the nitrogenase structural protein NifH (encoded by Npun_R0415) in heterocysts of NHM5. In contradiction, the abundance of another nitrogenase structural protein, NifK (encoded by Npun_R0390),

44
was unchanged. The downregulation of NifH might be oxygen related degradation since the oxygen levels in the heterocyst may be increased due to lowered respiratory activity when respiration is deprived of uptake hydrogenase generated electrons.
In heterocysts, the N2-fixation and respiration are depending on reducing equivalents (NADPH) supplied by the OPP pathway [120]. The OPP pathway proteins, encoded by the fbp-tal-zwf-opcA operon (Npun_F4023 to Npun_F4026), were upregulated in heterocysts of NHM5 suggesting an increased OPP pathway activity compensating for the loss of uptake hydrogenase generated reducing power. Abundance of proteins connected to carbon concentration and fixation, such as both subunits of Rubisco (encoded by rbcSL; Npun_F4195 and Npun_F4197) and sucrose synthase, SusA (encoded by susA; Npun_F1876), were in general lower in filaments of NHM5 compared to filaments of WT. Since NtcA acts as a repressor of both susA and rbcSL in Nostoc PCC 7120 during conditions of nitrogen limitation, the reduced overall carbon fixation rate in NMH5 could be due to an increased C:N ratio in the cells [121]. SusA, catalyzing the cleavage of sucrose [122], is located to vegetative cells [121] and has been suggested to be involved in the control of carbohydrate flow from vegetative cells to heterocyst [123,124]. Hence a down regulation of SusA may indicate an increased carbon transport from vegetative cells to heterocysts of NHM5, sustaining an upregulated OPP pathway activity. In addition, one alkaline/neutral invertase (encoded by Npun_F4643), irreversibly hydrolyzing sucrose to glucose and fructose [122], was found in higher abundance in heterocysts of NHM5 than in heterocyst of WT indicating an increased production of substrate for the OPP pathway.
The uptake hydrogenase has been suggested to act as an electron donor to both photosynthesis and respiration [125-127] and inactivation of the uptake hydrogenase should therefore affect both processes. This is also reflected in the different abundances of proteins related to photosynthetic and respiratoric electron transport in the heterocysts of NHM5 compared to heterocysts of WT. For example the Cox2 A subunit had higher abundance in heterocysts of NHM5 than in heterocyst of WT. Out of the three respiratory terminal oxidases found in N. punctiforme (Cox1, Cox2 and Cox3), only Cox 2 and Cox3 are expressed in the heterocysts [27,128]. One possible explanation for the increased abundance of the Cox2 A subunit (encoded by Npun_F0377) could be that, since Cox3 has been shown to be important for maintaining the microoxic environment in the heterocysts, the uptake hydrogenase generated electrons are primarily used by Cox3 and that in the absence of an uptake hydrogenase the amount of Cox2 is upregulated to maintain the high oxygen consumption rate in the heterocysts.

45
Since the water splitting, O2 evolving activity of PSII is lost in heterocyst, the main function of PSI is to generate ATP through cyclic electron flow [129]. This is supported by higher abundance of the PSI subunit PsaA (encoded by Npun_3818) in heterocysts of NHM5, compared to heterocysts of WT, and points to an important respiratory ATP production from uptake hydrogenase generated electrons. In addition, an inhibitor of the photosynthesis coupled F1F0 ATPase (encoded by Npun_R4416) were found in lower abundance while two proteins involved in respiratory and photosynthetic electron transport (encoded by Npun_F4970 and Npun_R5546) were found in higher abundance, indicating an altered electron flow in heterocysts of NHM5.
When looking at changes in the outer membrane and cell wall related proteins, six porins (encoded by Npun_F4220, Npun_F2347, Npun_F0722, Npun_F4555, Npun_F1139 and Npun_R0740) were found with lower abundance in heterocysts of NMH5 compared to heterocysts of WT. The exact function of these porins is unknown, but OprB in Pseudomonas aeruginosea has been suggested to be involved in glucose transport into the periplasmic space [130]. The reason for the downregulation of these porins could be to inhibit diffusion of external oxygen through the membranes, or more likely, due to the increased need for imported carbohydrates, to reduce an efflux of sugars through the outer membrane into the periplasmic space. Three cell wall/cell membrane associated proteins (encoded by Npun_R0039, Npun_R2885 and Npun_R5769) were found in higher abundance in either filaments or heterocysts of NHM5, probably correlating with a reduced capacity of respiration to remove oxygen since thickened glycolipid layer is a known response to elevated oxygen levels in the heterocyst [131]. When respiration is deprived of uptake hydrogenase generated electrons, the oxygen levels in the heterocyst are probably increased due to lowered respiratory activity. Such increase in intracellular oxygen concentrations would cause oxygen induced stress in the heterocysts. This is in agreement with the finding of increased abundances of proteins involved in protein turnover, i.e. chaperones and proteases, and ferritin-like iron binding proteins of the Dps (DNA-binding protein from starved cells) family in heterocysts of NMH5 indicating a heterocyst specific oxygen induced stress.
The NHM5 specific protein expression pattern (even though compared to an already nitrogen starved WT) is in many ways similar to the expression pattern of nitrogen starved cells, with the exception of the lowered abundance of NifH and the unchanged abundance of NtcA itself. Therefore, it seems like inactivation of the uptake hydrogenase causes an increase in the C:N ratio. Many of the genes encoding proteins found in different abundance

46
in NMH5 compared to WT are regulated by NtcA or have putative NtcA binding sites in their promoter regions. For example, the expression of GS [117], the Cox proteins [132], SusA [121] and Rubisco [121] have all been reported to be regulated by NtcA in cyanobacteria. This is however not surprising since NtcA, responding to the C:N balance in the cell, is the global regulator of nitrogen control as well as an important regulator of genes involved in heterocyst differentiation and there is a close connection between N2-fixation and hydrogen uptake metabolism. However, it does point to an important role of the C:N balance in initiation of the NHM5 specific protein expression pattern. Moreover, the C:N balance of the cell appears to have cross talks with other regulatory mechanisms, such as redox status and intracellular concentration of H2, since many genes regulated by NtcA have previously been reported to be regulated by redox status or electron transport chain related mechanisms, e.g. glnA (encoding GS) [119] and ntcA (encoding NtcA) [133,134]. In this context the roles of additional transcription factors, such as CalA, deserves some attention. In conclusion: Inactivation of the uptake hydrogenase in strain NHM5 caused only minor phenotypic effects, such as slowed growth rate during N2-fixing conditions, while the N2-fixation rate remained comparable to WT. However, when analyzing the effects on a global protein expression level several proteins were found in different abundance in NHM5 compared to WT. Among these, proteins involved in nitrogen and carbon fixation and metabolism, energy metabolism, stress responses and cell envelope function and structure were identified (Fig. 6). When the uptake hydrogenase is inactivated the heterocysts of NHM5 adapt to the loss of this energy recycling route. Those adaptations include higher respiratory activity, modifications of the heterocyst cell envelope and upregulation of stress response proteins indicating a significant supply of electrons from the uptake hydrogenase to respiration for reduction of O2 and that in the absence of an uptake hydrogenase alternative mechanisms are activated to keep the oxygen concentrations low. In order for heterocysts of NHM5 to maintain a high N2-fixation rate they need to compensate for the loss of uptake hydrogenase reducing power. These compensatory mechanisms to generate extra ATP and NADPH appears to involve upregulation of OPP pathway activity, increased carbohydrate transport from vegetative cells to heterocysts and increased abundance of PSI proteins and proteins involved in photosynthetic electron transport. At least 50% of the genes encoding the differently regulated proteins are NtcA regulated or have putative NtcA binding sites in their promoter regions suggesting a shift in the C:N balance when the uptake hydrogenase is absent. This is not strange considering that inactivation of the uptake hydrogenase most likely causes a reduction in the amount of ATP and NADPH available to drive N2-fixation, in turn leading to an increased C:N ratio. This C:N shift appears to induce an NtcA regulated activation of

47
compensatory pathways to maintain a high N2-fixation rate. Interestingly, the three metabolic processes mentioned above seem to be tightly connected since all of them affect the reductant pool in the cells. This study has given us a system-wide understanding of the NHM5 specific redirection of metabolic processes and shows the importance of the uptake hydrogenase in N2-fixation as well as the metabolic plasticity of N. punctiforme, viewed in the flexibility between different regulatory mechanisms, strengthening the suitability of this strain for bioengineering modifications.

48
SUMMARY AND FUTURE OUTLOOK
The main obstacles to overcome before the hydrogen economy is a reality concern storage and clean renewable production of H2 [8]. The idea of using cyanobacteria cultures as hydrogen factories is hence appealing but lies far away in the future, the main problem being the low H2 production rates [8]. Currently, large efforts are made to identify the largest barriers for high H2 production and to find ways to get around them. Some of the main reasons for the low production rates include efficient hydrogen oxidation by the cells, inefficient H2 evolving enzymes, oxygen sensitivity of the H2 producing enzymes and electron consuming pathways competing with H2 production, [11,18]. The H2 uptake activity of the cells has efficiently been addressed by knocking-out the uptake hydrogenase in several cyanobacterial strains [54,57,92-94]. Due to the low turnover of cyanobacterial [NiFe] hydrogenases [11] and the high ATP requirement of nitrogenases [10] there is an interest in introducing foreign hydrogenases in cyanobacteria. Algal [FeFe] hydrogenases have a higher turnover than cyanobacterial [NiFe] hydrogenases [4,10] and are thus promising candidates for heterologous expression in cyanobacteria. In addition, the H2 evolution could probably be improved further by overexpressing the H2 producing enzyme. Microorganisms have optimized their energy usage for survival and growth and not H2 production. Therefore, there are many competing electron consuming pathways in the cells, such as respiration and the Calvin cycle, limiting the electron flow to H2 evolving enzymes. In order to improve the H2 production rate those competing pathways must be identified and characterized and the electron flow redirected to H2 production. To be able to tackle all those obstacles mentioned above profound knowledge about the transcriptional regulation, maturation and function of the chosen H2 evolving enzyme and the metabolic and regulatory network in the target strain is needed.
The discovery of the D(C/G/F)GT/HOXBOX region in HupW-type respective HoxW-type proteases and the putative role of these regions in substrate recognition, binding and specificity may therefore be an important finding for understanding the selectivity of hydrogenase specific proteases. Heterologous gene products can’t be expected to work properly in an environment they are not optimized for. Hence, when introducing a foreign hydrogenase we need to know if the corresponding maturation genes and

49
hydrogenase specific protease need to be co-expressed together with the foreign hydrogenase or if the already existing endogenous maturation system is enough or can be modified to fit the heterologous enzyme. Mutational analysis of amino acids in the D(C/G/F)GT/HOXBOX region could thus provide useful information concerning substrate preferences. While the maturation process of the hydrogenase large subunit in cyanobacteria is rather well explored, the maturation processes of the small subunit remains clouded in darkness. This poses potential problems when aiming for successful expression of functionally active foreign hydrogenases in cyanobacteria. The identification, and partial characterization, of five genes, encoding proteins putatively involved in maturation of the uptake hydrogenase small subunit is therefore important for the basic understanding of the maturation process of the small subunit. Inactivation of these genes by gene interruption would provide further information about their physical role and importance for an active uptake hydrogenase in filamentous, N2-fixing cyanobacteria.
When expressing a foreign hydrogenase in a filamentous, heterocystous cyanobacteria the expression needs to be heterocyst specific and at a suitable high level. Therefore, understanding the regulatory mechanism for expression of hydrogenases is equally important as understanding the maturation process. Ideally, transcription should be inducible so that the transcription can be turned off when we quickly want to grow cells to high densities in newly started cultures and turned on in batch cultures ready to be used for H2 production. In order to achieve this goal we need information about key players regulating transcription of genes involved in H2 metabolism and what determines heterocyst specific expression. Both the transcript of hupSL, and the uptake hydrogenase itself, are located in the heterocysts in N. punctiforme as determined by observing GFP fluorescence in the hupSL promoter deletion study and from the HupS-GFP fusion protein. In both studies, GFP fluorescence was solely observed in the heterocysts. Moreover, the promoter deletion study showed that the promoter region of hupSL in N. punctiforme harbors a motif responsible for heterocyst specific expression. Identification of such a motif would be valuable and could be used when directing expression of foreign genes, such as hydrogenases, to the heterocysts. The finding that a very short hupSL promoter fragment (with the 5’ end located at -57 bp in relation to the tsp) is enough for a high and heterocyst specific expression narrows down the search for the motif conferring heterocyst specificity to a stretch of 316 bp. By fusing even shorter versions of the 316 bp DNA fragment to gfp and monitor the fluorescence the DNA region to investigate for motifs conferring heterocyst specificity could be shortened further before attempting to identify the motif, by exchanging or removing bases in the sequence, and regulatory protein(s) interacting with the motif. Three transcriptional

50
regulators were found interacting with the hupSL/extended hyp-operon promoter region in N. punctiforme. Those transcriptional regulators were, besides NtcA, two less well characterized transcription factors, CalA and CalB. Furthermore, CalA and CalB were found interacting with the hupSL/extended hyp-operon promoter region in Nostoc PCC 7120 as well. The importance and exact roles of these two cyanobacterial transcription factors remain to be elucidated, for example by overexpressing CalA and CalB separately in N. punctiforme and monitor the changes by transcriptomics and proteomics. CalA appears to be essential since no fully knock-out mutant has been created [81,85,88] but knocking-out CalB in N. punctiforme could give important clues of its function in filamentous, heterocystous cyanobacteria. In a commercially grown strain of cyanobacteria bioengineered for H2 production, the endogenous uptake hydrogenase activity must be inactivated to allow release of H2 to the surrounding. Inactivation of this energy recycling route represents a loss of an electron supplying pathway, as well as an energy loss for the cells, which the cells must adapt to. When analyzing the effects of inactivation of the uptake hydrogenase in an uptake hydrogenase deficient mutant strain of N. punctiforme several proteins were observed in different abundance compared to WT. Among these are proteins involved in energy metabolism, nitrogen and carbon fixation and metabolism as well as cellular stress responses and cell envelope function and structure. The knowledge that, in the absence of a functional uptake hydrogenase, alternative mechanisms are activated to keep the oxygen concentrations low has provided us with new targets when re-engineering cyanobacterial metabolism towards higher H2 production, such as oxygen scavenging components, cell envelope components and components involved in sugar transport from vegetative cells to heterocysts.
The work in this thesis has given valuable fundamental information about the transcriptional regulation, maturation and function of the uptake hydrogenase in filamentous, heterocystous cyanobacteria. Moreover, the function of the uptake hydrogenase has been inserted in a metabolic context illuminating the complexity and interplay of different metabolic pathways. This information could be used when introducing foreign hydrogenases into cyanobacteria or when attempting to enhance H2 production rates in cyanobacteria using endogenous H2 producing enzymes.

51
SUMMARY IN SWEDISH
Ett av våra största miljöproblem idag är den globala uppvärmningen. Ökande halter av växthusgaser i atmosfären har lett till högre medeltemperaturer på jorden och temperaturhöjningen har redan gett upphov till problem som smältande glaciärer, höjda havsnivåer och förändrade förutsättningar för jordbruket. Fossila bränslen som kol, olja och naturgas är stora utsläppskällor av växthusgaser och orsakar även skador på miljön i form av föroreningar. Att vi måste hitta nya koldioxidneutrala, förnyelsebara och framförallt rena energikällor har ytterligare belysts av oljekatastrofen i Mexikanska golfen 2010. Solenergi utgör just en sådan förnyelsebar energikälla med tillräcklig kapacitet inte bara för att förse västvärden med ren förnyelsebar energi, utan också för att tillgodose det snabbt växande energibehovet i utvecklingsländer som Kina och Indien. Ett samhälle baserat på solenergi har även den fördelen att länder inte längre blir beroende av andra länder för sin energiförsörjning, vilket kan bidra till att stabilisera det politiska klimatet. Eftersom mängden solljus är oregelbunden och varierar över årstiderna måste solljuset lagras i en energibärare för förvaring och transport. Vätgas (H2) är en energibärare vars enda utsläpp är vatten och är därför av många ansedd som det bästa valet för att lagra solenergi. Vätgasen kan användas som bränsle i bränsleceller och i många större städer rullar vätgasdrivna bussar och bilar redan på gatorna. Ett av de största problemen som måste lösas innan ett framtida vätgassamhälle kan bli verklighet är att det idag inte finns något miljövänligt kommersiellt gångbart sätt att producera vätgas från solenergi. Cyanobakterier, även kallade blågröna alger, är en grupp fotosyntetiska mikroorganismer som har fått mycket uppmärksamhet i det här sammanhanget eftersom de kan producera H2 från solljus och vatten. Tyvärr producerar cyanobakterier väldigt små mängder H2 och utnyttjar precis som vi vätgasen som bränsle vilket resulterar i att mängden H2 de släpper ut till oss är väldigt liten. En cyanobakterie odlad för kommersiell vätgasproduktion måste därför vara en genetiskt förbättrad sådan. Exempelvis måste det vätgaskonsumerande enzymet inaktiveras och de vätgasproducerande enzymerna göras mera effektiva eller bytas ut mot effektivare enzymer från andra organismer. För att kunna skapa denna förbättrade bakterie, anpassad för storskalig vätgasproduktion, krävs detaljerade kunskaper om bakteriens reglering av vätgasproducerande enzym och hur deras ämnesomsättning fungerar.

52
I cyanobakterier produceras H2 som en biprodukt när enzymet nitrogenas fixerar kväve från luften. Den producerade vätgasen tas upp av enzymet upptagshydrogenas som gör energiinnehållet i vätgasen återigen tillgänglig för organismen. Vissa cyanobakteriestammar innehåller även ett tredje enzym, det bidirektionella hydrogenaset, som kan både producera och konsumera vätgas. Nitrogenaser och hydrogenaser är syrekänsliga och inaktiveras snabbt av den syrgas som produceras av fotosyntesen. Cyanobakterier måste därför separera fotosyntes och vätgasproduktion antingen i tid, med fotosyntes på dagen och vätgasproduktion på natten, eller i rum, i celler specialiserade för kvävefixering. Dessa specialiserade celler kallas för heterocyster. I vilka celltyper hydrogenaserna ska uttryckas, och i vilken mängd, bestäms till stor del av så kallade transkriptionsfaktorer. Hydrogenaser är avancerade enzym som behöver hjälp av accessoriska proteiner för korrekt ihopsättning. Denna ihopsättning kallas för hydrogenasets mognadsprocess. Mitt arbete har gett fördjupad kunskap om regleringen, mognadsprocessen och funktionen av cyanobakteriella upptagshydrogenaser. För detta arbete valdes två cyanobakteriella stammar, Nostoc punctiforme ATCC 29133 och Nostoc sp. strain PCC 7120, som modellorganismer. Dessa stammar är närbesläktade och i många avseenden väldigt lika men skiljer sig åt eftersom N. punctiforme bara har ett upptagshydrogenas medan Nostoc PCC har både ett upptagshydrogenas och ett bidirektionellt hydrogenas.
Sammanfattningsvis så visar denna avhandling att: o Upptagshydrogenaset uttrycks specifikt i heterocyster o Inaktivering av upptagshydrogenaset ger upphov till en rad
strukturella och ämnesomsättningsrelaterade förändringar. Genom att studera dessa förändringar får vi större förståelse för upptagshydrogenaset funktion och var energin från den konsumerade vätgasen tar vägen i cellen.
o Fem gener, lokaliserade uppströms de gener som kodar för de assessoriska proteinerna, kodar troligtvis för andra, hittills okända mognadsprocessproteiner
o Två okända transkriptionsfaktorer, CalA och CalB, kan vara inblandade i regleringen av upptagshydrogenaset och/eller de accessoriska mognadsprocessproteinerna.
o En konserverad region i de hydrogenasspecifika proteaserna kan vara det som bestämmer specificiteten hos proteaser.
o Upptagshydrogenaser och deras specifika proteaser har följts åt genom evolutionen under en mycket lång tid.

53
ACKNOWLEDGEMENT
First I would like to thank my supervisors Peter and Karin for all the fruitful scientific discussions, for all the time you have spent reading my manuscripts and for teaching me how to write manuscripts and become an independent scientist. You have both taught me so much and for that I’m ever grateful. I would also like to thank Peter for giving me this great opportunity when accepting me as a PhD student in his group and for hosting so many Cyanogroup parties during the years and Karin for all the good advices, work related as well as non-work related, and for your never ending, highly contagious enthusiasm. You have been not only my mentor but also my close friend and older sister! Thanks to all the present and past members of the Cyanogroup: Ann (Wipawee), Daniel C, Ellenor, Fernando, Hsin-ho, Johannes, Josè, Olga, Martin, Namita, Paulo, Petra, Pia, Rikard, Tanai, Taras, Thorsten, Wanthanee, Xiaohui, Åsa A, Åsa S for your friendship and company and all the great scientific discussions during our group meetings. It has been a pleasure working with you. You have been like my second family for a long time and I will miss you all. Best of luck with your ongoing and future scientific projects and take care. I also would like to thank all the students, (Cissi, Daniel V, Helena, John W, Lei, Micke, Peter Y, Pond, Robert, Sean, Thiyagu and Thomas) who did shorter or longer projects in our group for bringing new ideas to the group and for nice company.
Ellenor and Åsa A – thanks for wonderful friendship. I never thought I would find two of my best friends at work, but I did and I count myself lucky. You both mean so much to me. I’m glad that you have been beside me during all this time, sharing the fun times as well as the hard times, being excellent travelling companions and for understanding me and my situation like no other of my friends who aren’t PhD students can. Love you both!
Pia – I have been trying to walk in your shoes for many years. You are one of my raw models and I enjoy the way I can toss scientific ideas back and forth with you while working in the lab. Thanks also for nice company in the lab and during breaks, for trusting me to continue your project when you left for your postdoc and for being such a down-to-earth, relaxed and nice person to be around.

54
Gunilla – besides being a very competent and efficient person to work with I must say that you are the coolest lady I know and I can only hope that I will turn into someone like you when I grow old(er). I miss all the friendly banter during fika!
Paulo - thank you for all the interesting scientific discussions and good advices during the years, your helpfulness and for taking the time to ask me how I am from time to time. I really, really appreciate it.
Fernando - thank you for your friendship, for all the crazy fun moments, for bringing me to Portugal and showing me your home and your country, for caring about me and if I feel ok and last, but not least, for taking care of Åsa S for me.
José - thank you for all fun moments and for your friendship and company. You, Fernando and Paulo made a very special trio together
Åsa S - thanks for being my friend and for being you. I also want to thank you and Fernando for bringing your wonderful boy Lucas into the world and for trusting me to hold him. It is wonderful when two friends fall in love and you happiness is my happiness.
Helena and Robert - thanks for being the positive persons you are. You were both great company when you were working here, and you both left an empty space behind. I miss having you around but I’m glad that I could keep you as friends when I lost you as colleagues.
John Larsson - thanks for being an excellent student and for putting up with me as your supervisor without complaining. It was fun working together with you and I learned a lot about supervising during that period. Good luck with your own PhD studies!
Thanks to all the present and past members at the Department of Photochemistry and Molecular Science for your friendliness, your company and for fun conversations and scientific discussions.
Leif – thanks for being such a good prefect, caring about the organization of the department, your employees and their working environment. There are not a coincidence that girls at Fotomol are dying their hairs and so many Fotomol babies are being born – people like working here.
Sven – thanks for being such a competent and reliable janitor and for nice company at fika. Is there something that you can not fix?
Åsa F, Susanne S and Maria L – thanks for being superb administrators, always helpful and constructive, and for fantastic and fun company. Fika breaks would not have been the same without you!
The cleaning staff – thanks for keeping tidy around the mess on my desk. Would have been great if you would have been paid to clean my desk too!
Erik – thanks for making sure that we never run out tea. A very important task!

55
Thanks to Stenbjörn for creating such a good environment for science in the Consortium of Artificial Photosynthesis.
Thanks to all the persons I spent so much time with teaching during the years: Hai-Peng, Christoph, Eva H, Paulo and Åsa A. It has been an honor and a pleasure teaching with you. Special thanks to Marianne for being so friendly and helpful and for all the technical service. You made teaching easy for us.
Thanks to the past and present members of Physiological Botany which I work with during my years at the department: Anna, Annelie, Annika, Anita, Agneta, Birgitta, Calle, Cecilia, Elin, Erika, Eva H, Eva S, Gunn-Britt, Henrik J (Jr), Henrik J. (Sr), Lars, Marie and Sandra. I very much enjoyed the time I spent with you all at Fysbot. Special thanks to Tage for all the apples!
Thanks to all the past members of the Mast cell group at the Rudbeck laboratory: Gunnar, Nicklas, Marie, Mikael, Christine, Erik, Jessica, Maria, Hanna. You were all a part of my first experience with the scientific community and inspired me to go for a PhD degree. Special thanks to Erik for being such a good supervisor and to Hanna and Maria and Jessica for all fun times and your friendship! Thanks for the financial support from Rektor’s travelling grant from Wallenberstiftelsen and Ångpanneföreningen that made it possible for me to attend so many great conferences. Thanks Mum and Dad and my dear sisters Sofia and Britta for never ending love, patience, support and shoulders to cry on when needed. Special thanks to Sofia invaluable babysitting when I needed to work late. Thanks to Daniel for everything! I lack words to describe my gratitude to you for being there for me, pushing me in the right direction when needed and for putting up with all my irregular working hours and my annoying attitude that “work comes first”. Especially, when we have missed trains because I had to feed my cultures before we left for vacation, or when I booked our flight tickets on the wrong day, or in the wrong direction, because my mind were still at work. Thank you for giving me the most wonderful gift on earth, our amazing son Edwind, and thank you Edwind for making my life complete. Love you both endlessly! Thanks to Ellenor, Karin, Peter L and Åsa A for reading and commenting my thesis.

56
Thanks to all my non-work related friends who are still my friends even though I have buried myself in work in periods and not stayed in touch. You are all very important to me even though I’m bad at showing it. I will try to make it up to you now when I have more time!
Thanks to Nostoc punctiforme ATCC 29133 and Nostoc sp. strain PCC 7120 for having, in contradiction to my plants at home, survived my loving care all these years. Finally a special thanks to all those people that I seem to have forgotten right now but that I never will forget in my heart! Thank you!!

57
REFERENCES
1. Lewis, N.S. and Nocera, D.G. (2006). Powering the planet: chemical challenges in solar energy utilization. Proc. Natl. Acad. Sci. U. S. A, 103(43):15729-15735.
2. Barbir, F. (2009). Transition to renewable energy systems with hydrogen as an energy carrier. Energy, 34(3):308-312.
3. Ogden, J., M. (2002). Hydrogen: The fuel of the future? Physics Today, 55(4): 69-75.
4. Rupprecht, J., Hankamer, B., Mussgnug, J.H., Ananyev, G., Dismukes, C. and Kruse, O. (2006). Perspectives and advances of biological H2 production in microorganisms. Appl. Microbiol. Biotechnol., 72(3):442-449.
5. Kamat, P.V. (2007). Meeting the clean energy demand: Nanostructure architectures for solar energy conversion. J. Phys. Chem. C, 111(7):2834-2860.
6. Eisenberg, R. and Nocera, D.G. (2005). Preface: overview of the forum on solar and renewable energy. Inorg. Chem., 44(20):6799-6801.
7. Suzuki, Y. (1982). On Hydrogen as Fuel Gas. Int. J. Hydrogen Energy, 7(3):227-230.
8. Levin, D.B., Pitt, L. and Love, M. (2004). Biohydrogen production: prospects and limitations to practical application. Int. J. Hydrogen Energy, 29(2):173-185.
9. Tamagnini, P., Axelsson, R., Lindberg, P., Oxelfelt, F., Wunschiers, R. and Lindblad, P. (2002). Hydrogenases and hydrogen metabolism of cyanobacteria. Microbiol. Mol. Biol. Rev., 66(1):1-20.
10. Ghirardi, M.L., Dubini, A., Yu, J. and Maness, P.C. (2009). Photobiological hydrogen-producing systems. Chem. Soc. Rev., 38(1): 52-61.
11. Ghirardi, M.L., Posewitz, M.C., Maness, P.C., Dubini, A., Yu, J. and Seibert, M. (2007). Hydrogenases and hydrogen photoproduction in oxygenic photosynthetic organisms. Annu. Rev. Plant. Biol., 58:71-91.
12. Prince, R.C. and Kheshgi, H.S. (2005). The photobiological production of hydrogen: potential efficiency and effectiveness as a renewable fuel. Crit. Rev. Microbiol., 31(1):19-31.
13. Sakurai, H. and Masukawa, H. (2007). Promoting R & D in photobiological hydrogen production utilizing mariculture-raised cyanobacteria. Mar. Biotechnol. (N.Y.), 9(2):128-145.

58
14. Meeks, J.C. and Elhai, J. (2002). Regulation of cellular differentiation in filamentous cyanobacteria in free-living and plant-associated symbiotic growth states. Microbiol. Mol. Biol. Rev., 66(1):94-121.
15. Meeks, J.C. (1998). Symbiosis between nitrogen-fixing cyanobacteria and plants. The establishment of symbiosis causes dramatic morphological and physiological changes in the cyanobacterium. Bioscience, 48(4):266-276.
16. Tomitani, A., Knoll, A.H., Cavanaugh, C.M. and Ohno, T. (2006). The evolutionary diversification of cyanobacteria: molecular-phylogenetic and paleontological perspectives. Proc. Natl. Acad. Sci. U. S. A., 103(14):5442-5447.
17. Cavalier-Smith, T. (2006). Cell evolution and Earth history: stasis and revolution. Philos. Trans. R. Soc. Lond. B. Biol. Sci., 361(1470):969-1006.
18. Tamagnini, P., Leitao, E., Oliveira, P., Ferreira, D., Pinto, F., Harris, D.J., Heidorn, T. and Lindblad, P. (2007). Cyanobacterial hydrogenases: diversity, regulation and applications. FEMS Microbiol. Rev., 31(6):692-720.
19. Campbell, E.L. and Meeks, J.C. (1989). Characteristics of hormogonia formation by symbiotic-Nostoc spp. in response to the presence of Anthoceros-Punctatus or its extracellular products. Appl. Environ. Microbiol., 55(1):125-131.
20. Vignais, P.M. and Billoud, B. (2007). Occurrence, classification, and biological function of hydrogenases: an overview. Chem. Rev., 107(10):4206-4272.
21. Vignais, P.M., Billoud, B. and Meyer, J. (2001). Classification and phylogeny of hydrogenases. FEMS Microbiol. Rev., 25(4):455-501.
22. Tsygankov, A.A. (2007). Nitrogen-fixing cyanobacteria: A review. Appl. Biochem. Microbiol., 43(3):250-259.
23. Bergman, B., Gallon, J.R., Rai, A.N. and Stal, L.J. (1997). N2- fixation by non-heterocystous cyanobacteria. FEMS Microbiol. Rev., 19(3):139-185.
24. Kumar, K., Mella-Herrera, R.A. and Golden, J.W. Cyanobacterial heterocysts. Cold Spring Harb. Perspect. Biol., 2(4):a000315.
25. Flores, E. and Herrero, A. Compartmentalized function through cell differentiation in filamentous cyanobacteria. Nat. Rev. Microbiol., 8(1):39-50.
26. Xu, X. and Wolk, C.P. (2001). Role for hetC in the transition to a nondividing state during heterocyst differentiation in Anabaena sp. J. Bacteriol., 183(1):393-396.
27. Valladares, A., Herrero, A., Pils, D., Schmetterer, G. and Flores, E. (2003). Cytochrome C oxidase genes required for nitrogenase activity and diazotrophic growth in Anabaena sp. PCC 7120. Mol. Microbiol., 47(5):1239-1249.
28. Thiel, T. (1993). Characterization of genes for an alternative nitrogenase in the cyanobacterium Anabaena variabilis. J. Bacteriol., 175(19):6276-6286.

59
29. Lindberg, P., Lindblad, P. and Cournac, L. (2004). Gas exchange in the filamentous cyanobacterium Nostoc punctiforme strain ATCC 29133 and its hydrogenase-deficient mutant strain NHM5. Appl. Environ. Microbiol., 70(4):2137-2145.
30. Vignais, P.M. and Colbeau, A. (2004). Molecular biology of microbial hydrogenases. Curr. Iss. Mol. Biol., 6:159-188.
31. Wu, L.F. and Mandrand, M.A. (1993). Microbial hydrogenases: primary structure, classification, signatures and phylogeny. FEMS Microbio. Rev., 10(3-4):243-269.
32. Bock, A., King, P.W., Blokesch, M. and Posewitz, M.C. (2006). Maturation of hydrogenases. Adv. Microb. Physiol., 51:1-71.
33. Hoffmann, D., Gutekunst, K., Klissenbauer, M., Schulz-Friedrich, R. and Appel, J. (2006). Mutagenesis of hydrogenase accessory genes of Synechocystis sp. PCC 6803. Additional homologues of hypA and hypB are not active in hydrogenase maturation. FEBS J., 273(19):4516-4527.
34. Forzi, L. and Sawers, R.G. (2007). Maturation of [NiFe]-hydrogenases in Escherichia coli. Biometals, 20(3-4):565-578.
35. Devine, E., Holmqvist, M., Stensjö, K. and Lindblad, P. (2009). Diversity and transcription of proteases involved in the maturation of hydrogenases in Nostoc punctiforme ATCC 29133 and Nostoc sp. strain PCC 7120. BMC Microbiol., 9:53.
36. Gollin, D.J., Mortenson, L.E. and Robson, R.L. (1992). Carboxyl-terminal processing may be essential for production of active NiFe hydrogenase in Azotobacter vinelandii. FEBS Lett., 309(3):371-375.
37. Menon, N.K., Robbins, J., Der Vartanian, M., Patil, D., Peck, H.D., Jr., Menon, A.L., Robson, R.L. and Przybyla, A.E. (1993). Carboxy-terminal processing of the large subunit of [NiFe] hydrogenases. FEBS Lett., 331(1-2):91-95.
38. Fritsche, E., Paschos, A., Beisel, H.G., Bock, A. and Huber, R. (1999). Crystal structure of the hydrogenase maturating endopeptidase HybD from Escherichia coli. J. Mol. Biol., 288(5):989-998.
39. Theodoratou, E., Paschos, A., Magalon, A., Fritsche, E., Huber, R. and Bock, A. (2000). Nickel serves as a substrate recognition motif for the endopeptidase involved in hydrogenase maturation. Eur. J. Biochem., 267(7):1995-1999.
40. Fontecave, M. and Ollagnier-De-Choudens, S. (2008). Iron-sulfur cluster biosynthesis in bacteria: Mechanisms of cluster assembly and transfer. Arch. Biochem. Biophys., 474(2):226-237.
41 Balasubramanian, R., Shen, G.Z., Bryant, D.A. and Golbeck, J.H. (2006). Regulatory roles for IscA and SufA in iron homeostasis and redox stress responses in the cyanobacterium Synechococcus sp strain PCC 7002. J. Bacteriol., 188(9):3182-3191.
42. Rippka, R. and Herdman, M. (1992) Pasteur Culture Collection of cyanobacterial strains in axenic culture. In Catalogue and Taxonomic Handbook, Institut Pasteur, Paris, p. 103.

60
43. Costa, J.-L. and Lindblad, P. (2002) Cyanobacteria in symbiosis with cycads. In Cyanobacteria in Symbiosis, Kluwer Academic Publishers, Dordrecht, The Netherlands, pp. 195-205.
43. Meeks, J.C., Elhai, J., Thiel, T., Potts, M., Larimer, F., Lamerdin, J., Predki, P. and Atlas, R. (2001). An overview of the genome of Nostoc punctiforme, a multicellular, symbiotic cyanobacterium. Photosynth. Res., 70(1):85-106.
45. Campbell, E.L., Summers, M.L., Christman, H., Martin, M.E. and Meeks, J.C. (2007). Global gene expression patterns of Nostoc punctiforme in steady-state dinitrogen-grown heterocyst-containing cultures and at single time points during the differentiation of akinetes and hormogonia. J. Bacteriol., 189(14):5247-5256.
46. Ow, S.Y., Noirel, J., Cardona, T., Taton, A., Lindblad, P., Stensjö, K. and Wright, P.C. (2009). Quantitative overview of N2-fixation in Nostoc punctiforme ATCC 29133 through cellular enrichments and iTRAQ shotgun proteomics. J. Proteome. Res., 8(1):187-198.
47. Anderson, D.C., Campbell, E.L. and Meeks, J.C. (2006). A soluble 3D LC/MS/MS proteome of the filamentous cyanobacterium Nostoc punctiforme. J. Proteome. Res., 5(11):3096-3104.
48. Kaneko, T. et al. (2001). Complete genomic sequence of the filamentous nitrogen-fixing cyanobacterium Anabaena sp. strain PCC 7120. DNA Res., 8(5):205-213, 227-253.
49. Ehira, S. and Ohmori, M. (2006). NrrA, a nitrogen-responsive response regulator facilitates heterocyst development in the cyanobacterium Anabaena sp. strain PCC 7120. Mol. Microbiol., 59(6):1692-1703.
50. Ow, S.Y., Cardona, T., Taton, A., Magnuson, A., Lindblad, P., Stensjö, K. and Wright, P.C. (2008). Quantitative shotgun proteomics of enriched heterocysts from Nostoc sp. PCC 7120 using 8-plex isobaric peptide tags. J. Proteome. Res., 7(4):1615-1628.
51. Stensjö, K., Ow, S.Y., Barrios-Llerena, M.E., Lindblad, P. and Wright, P.C. (2007). An iTRAQ-based quantitative analysis to elaborate the proteomic response of Nostoc sp. PCC 7120 under N2-fixing conditions. J. Proteome. Res., 6(2):621-635.
52. Peterson, R.B. and Wolk, C.P. (1978). Localization of an uptake hydrogenase in Anabaena. Plant Physiol., 61(4):688-691.
53 Seabra, R., Santos, A., Pereira, S., Moradas-Ferreira, P. and Tamagnini, P. (2009). Immunolocalization of the uptake hydrogenase in the marine cyanobacterium Lyngbya majuscula CCAP 1446/4 and two Nostoc strains. FEMS Microbiol. Lett., 292(1):57-62.
54. Carrasco, C.D., Holliday, S.D., Hansel, A., Lindblad, P. and Golden, J.W. (2005). Heterocyst-specific excision of the Anabaena sp. strain PCC 7120 hupL element requires xisC. J. Bacteriol., 187(17):6031-6038.
55. Lindblad, P. and Sellstedt, A. (1990). Occurrence and Localization of an uptake hydrogenase in the filamentous heterocystous cyanobacterium Nostoc PCC 73102. Protoplasma, 159(1):9-15.

61
56. Tamagnini, P., Oxelfelt, F., Salema, R. and Lindblad, P. (1995). Immunological characterization of hydrogenases in the nitrogen-fixing cyanobacterium Nostoc sp. strain PCC 73102. Curr. Microbiol., 31(2):102-107.
57. Lindberg, P., Schutz, K., Happe, T. and Lindblad, P. (2002). A hydrogen-producing, hydrogenase-free mutant strain of Nostoc punctiforme ATCC 29133. Int. J. Hydrogen Energy, 27(11-12):1291-1296.
58. Meek, L. and Arp, D.J. (2000). The hydrogenase cytochrome b heme ligands of Azotobacter vinelandii are required for full H2 oxidation capability. J. Bacteriol., 182(12):3429-3436.
59. Sayavedrasoto, L.A. and Arp, D.J. (1992). The hoxZ gene of the Azotobacter vinelandii hydrogenase operon is required for activation of hydrogenase. J. Bacteriol., 174(16):5295-5301.
60. Houchins, J.P. and Burris, R.H. (1981). Comparative characterization of two distinct hydrogenases from Anabaena sp. strain 7120. J. Bacteriol., 146(1):215-21.
61. Lindberg, P., Hansel, A. and Lindblad, P. (2000). hupS and hupL constitute a transcription unit in the cyanobacterium Nostoc sp. PCC 73102. Arch. Microbiol., 174(1-2):129-133.
62. Lindberg, P. (2003) Cyanobacterial hydrogen metabolism. Uptake hydrogenase and hydrogen production by nitrogenase in filamentous cyanobacteria. Ph. D. thesis. Uppsala University, Uppsala, Sweden
63. Ramasubramanian, T.S., Wei, T.F. and Golden, J.W. (1994). Two Anabaena sp. strain PCC 7120 DNA-binding factors interact with vegetative cell- and heterocyst-specific genes. J. Bacteriol., 176(5):1214-1223.
64. Wagner, R. (2000). The players or cellular components necessary for transcription. In Transcriptional Regulation in Procaryotes, Oxford University Press, New York, U.S.A., p. 17.
65. Hoover, T.R., Santero, E., Porter, S. and Kustu, S. (1990). The integration host factor stimulates interaction of RNA polymerase with NIFA, the transcriptional activator for nitrogen fixation operons. Cell, 63(1):11-22.
66. Black, L.K. and Maier, R.J. (1995). IHF- and RpoN-dependent regulation of hydrogenase expression in Bradyrhizobium japonicum. Mol. Microbiol., 16(3):405-413.
67. Vega-Palas, M.A., Madueno, F., Herrero, A. and Flores, E. (1990). Identification and cloning of a regulatory gene for nitrogen assimilation in the cyanobacterium Synechococcus sp. strain PCC 7942. J. Bacteriol., 172(2):643-647.
68. Herrero, A., Muro-Pastor, A.M., Valladares, A. and Flores, E. (2004). Cellular differentiation and the NtcA transcription factor in filamentous cyanobacteria. FEMS Microbiol. Rev., 28(4):469-487.
69. Muro-Pastor, A.M., Flores, E. and Herrero, A. (2009). NtcA-regulated heterocyst differentiation genes hetC and devB from Anabaena sp

62
Strain PCC 7120 exhibit a similar tandem promoter arrangement. J. Bacteriol., 191(18):5765-5774.
70. Herrero, A., Muro-Pastor, A.M. and Flores, E. (2001). Nitrogen control in cyanobacteria. J. Bacteriol., 183(2):411-425.
71. Luque, I., Forchhammer, K. (2008) Nitrogen assimilation and C/N balance sensing. In The Cyanobacteria, Caister Academic Press, Norfolk, U. K., pp. 335-382
72. Flores, E. and Herrero, A. (2005). Nitrogen assimilation and nitrogen control in cyanobacteria. Biochem. Soc. Trans. 33:164-167.
73. Muro-Pastor, M.I., Reyes, J.C. and Florencio, F.J. (2005). Ammonium assimilation in cyanobacteria. Photosynth. Res., 83(2):135-150.
74. Vazquez-Bermudez, M.F., Herrero, A. and Flores, E. (2002). 2-Oxoglutarate increases the binding affinity of the NtcA (nitrogen control) transcription factor for the Synechococcus glnA promoter. FEBS Lett., 512(1-3):71-4.
75. Muro-Pastor, M.I., Reyes, J.C. and Florencio, F.J. (2001). Cyanobacteria perceive nitrogen status by sensing intracellular 2-oxoglutarate levels. J. Biol. Chem., 276(41):38320-38328.
76 Muro-Pastor, A.M., Valladares, A., Flores, E. and Herrero, A. (2002). Mutual dependence of the expression of the cell differentiation regulatory protein HetR and the global nitrogen regulator NtcA during heterocyst development. Mol. Microbiol., 44(5): 1377-1385.
77. Wei, T.F., Ramasubramanian, T.S. and Golden, J.W. (1994). Anabaena sp. strain PCC 7120 ntcA gene required for growth on nitrate and heterocyst development. J. Bacteriol., 176(15):4473-4482.
78. Frias, J.E., Flores, E. and Herrero, A. (1994). Requirement of the regulatory protein NtcA for the expression of nitrogen assimilation and heterocyst development genes in the cyanobacterium Anabaena sp. PCC 7120. Mol. Microbiol, 14(4):823-832.
79. Barnard, A., Wolfe, A. and Busby, S. (2004). Regulation at complex bacterial promoters: how bacteria use different promoter organizations to produce different regulatory outcomes. Curr. Opin. Microbiol., 7(2):102-108.
80. Luque, I., Contreras, A., Zabulon, G., Herrero, A. and Houmard, J. (2002). Expression of the glutamyl-tRNA synthetase gene from the cyanobacterium Synechococcus sp PCC 7942 depends on nitrogen availability and the global regulator NtcA. Mol. Microbiol., 46(4):1157-1167.
81. Ishii, A. and Hihara, Y. (2008). An AbrB-like transcriptional regulator, sll0822, is essential for the activation of nitrogen-regulated genes in Synechocystis sp. PCC 6803. Plant Physiol., 148(1): 660-670.
82. Bobay, B.G., Benson, L., Naylor, S., Feeney, B., Clark, A.C., Goshe, M.B., Strauch, M.A., Thompson, R. and Cavanagh, J. (2004). Evaluation of the DNA binding tendencies of the transition state regulator AbrB. Biochemistry, 43(51):16106-16118.

63
83. Phillips, Z.E. and Strauch, M.A. (2002). Bacillus subtilis sporulation and stationary phase gene expression. Cell. Mol. Life Sci., 59(3):392-402.
84. Xu, K. and Strauch, M.A. (1996). In vitro selection of optimal AbrB-binding sites: comparison to known in vivo sites indicates flexibility in AbrB binding and recognition of three-dimensional DNA structures. Mol. Microbiol., 19(1):145-158.
85. Agervald, A., Zhang, X., Stensjö, K., Devine, E. and Lindblad, P. CalA, a cyanobacterial AbrB protein, interacts with the upstream region of hypC and acts as a repressor of its transcription in the cyanobacterium Nostoc sp. strain PCC 7120. Appl. Environ. Microbiol., 76(3):880-890.
86. Shalev-Malul, G., Lieman-Hurwitz, J., Viner-Mozzini, Y., Sukenik, A., Gaathon, A., Lebendiker, M. and Kaplan, A. (2008). An AbrB-like protein might be involved in the regulation of cylindrospermopsin production by Aphanizomenon ovalisporum. Environ. Microbiol., 10(4):988-999.
87. Oliveira, P. and Lindblad, P. (2009). Transcriptional regulation of the cyanobacterial bidirectional Hox-hydrogenase. Dalton Trans., 2009(45):9990-9996.
88. Oliveira, P. and Lindblad, P. (2008). An AbrB-Like protein regulates the expression of the bidirectional hydrogenase in Synechocystis sp. strain PCC 6803. J. Bacteriol., 190(3):1011-1019.
89. He, D. and Xu, X. CalA, a cyAbrB protein, binds to the upstream region of ftsZ and is down-regulated in heterocysts in Anabaena sp. PCC 7120. Arch. Microbiol., 192(6):461-469.
90. Agervald, A., Baebprasert, W., Zhang, X., Incharoensakdi, A., Lindblad, P. and Stensjö, K. The CyAbrB transcription factor CalA regulates the iron superoxide dismutase in Nostoc sp. strain PCC 7120. Environ. Microbiol., doi: 10.1128/AEM.02521-09
91. Lieman-Hurwitz, J., Haimovich, M., Shalev-Malul, G., Ishii, A., Hihara, Y., Gaathon, A., Lebendiker, M. and Kaplan, A. (2009). A cyanobacterial AbrB-like protein affects the apparent photosynthetic affinity for CO2 by modulating low-CO2-induced gene expression. Environ Microbiol., 11(4):927-36.
92. Happe, T., Schutz, K. and Bohme, H. (2000). Transcriptional and mutational analysis of the uptake hydrogenase of the filamentous cyanobacterium Anabaena variabilis ATCC 29413. J. Bacteriol., 182(6):1624-1631.
93. Masukawa, H., Mochimaru, M. and Sakurai, H. (2002). Disruption of the uptake hydrogenase gene, but not of the bidirectional hydrogenase gene, leads to enhanced photobiological hydrogen production by the nitrogen-fixing cyanobacterium Anabaena sp. PCC 7120. Appl. Microbiol. Biotechnol., 58(5):618-624.
94. Yoshino, F., Ikeda, H., Masukawa, H. and Sakurai, H. (2007). High photobiological hydrogen production activity of a Nostoc sp. PCC

64
7422 uptake hydrogenase-deficient mutant with high nitrogenase activity. Mar. Biotechnol. (N.Y.), 9(1):101-112.
95. Weyman, P.D., Pratte, B. and Thiel, T. (2008). Transcription of hupSL in Anabaena variabilis ATCC 29413 is regulated by NtcA and not by hydrogen. Appl. Environ. Microbiol., 74(7):2103-2110.
96. Leitao, E., Oxelfelt, F., Oliveira, P., Moradas-Ferreira, P. and Tamagnini, P. (2005). Analysis of the hupSL operon of the nonheterocystous cyanobacterium Lyngbya majuscula CCAP 1446/4: regulation of transcription and expression under a light-dark regimen. Appl. Environ. Microbiol., 71(8):4567-4576.
97. Oliveira, P., Leitao, E., Tamagnini, P., Moradas-Ferreira, P. and Oxelfelt, F. (2004). Characterization and transcriptional analysis of hupSLW in Gloeothece sp. ATCC 27152: an uptake hydrogenase from a unicellular cyanobacterium. Microbiology, 150:3647-3655.
98. Ramasubramanian, T.S., Wei, T.F., Oldham, A.K. and Golden, J.W. (1996). Transcription of the Anabaena sp strain PCC 7120 ntcA gene: Multiple transcripts and NtcA binding. J. Bacteriol., 178(3):922-926.
99. Buikema, W.J. and Haselkorn, R. (2001). Expression of the Anabaena hetR gene from a copper-regulated promoter leads to heterocyst differentiation under repressing conditions. Proc. Natl. Acad. Sci. U. S. A., 98(5):2729-2734.
100. Olmedo-Verd, E., Muro-Pastor, A.M., Flores, E. and Herrero, A. (2006). Localized induction of the ntcA regulatory gene in developing heterocysts of Anabaena sp strain PCC 7120. J. Bacteriol., 188(18):6694-6699.
101. Rey, L., Hidalgo, E., Palacios, J. and Ruiz-Argueso, T. (1992). Nucleotide sequence and organization of an H2-uptake gene cluster from Rhizobium leguminosarum bv. viciae containing a rubredoxin-like gene and four additional open reading frames. J. Mol. Biol., 228: 998-1002.
102 Manyani, H., Rey, L., Palacios, J.M., Imperial, J. and Ruiz-Argueso, T. (2005). Gene products of the hupGHIJ operon are involved in maturation of the iron-sulfur subunit of the [NiFe] hydrogenase from Rhizobium leguminosarum bv. viciae. J. Bacteriol., 187(20):7018-7026.
103. Burgdorf, T., Lenz, O., Buhrke, T., van der Linden, E., Jones, A.K., Albracht, S.P. and Friedrich, B. (2005). [NiFe]-hydrogenases of Ralstonia eutropha H16: modular enzymes for oxygen-tolerant biological hydrogen oxidation. J. Mol. Microbiol. Biotechnol., 10(2-4):181-196.
104. Menon, A.L., Mortenson, L.E. and Robson, R.L. (1992). Nucleotide sequences and genetic analysis of hydrogen oxidation (hox) genes in Azotobacter vinelandii. J. Bacteriol., 174(14):4549-4557.
105. Schubert, T., Lenz, O., Krause, E., Volkmer, R. and Friedrich, B. (2007). Chaperones specific for the membrane-bound [NiFe]-hydrogenase interact with the Tat signal peptide of the small subunit precursor in Ralstonia eutropha H16. Mol. Microbiol., 66(2):453-467.

65
106. Martinez, M., Brito, B., Imperial, J. and Ruiz-Argueso, T. (2004). Characterization of a new internal promoter (P3) for Rhizobium leguminosarum hydrogenase accessory genes hupGHIJ. Microbiology 150:665-75.
107. Hansel, A., Axelsson, R., Lindberg, P., Troshina, O.Y., Wunschiers, R. and Lindblad, P. (2001). Cloning and characterisation of a hyp-gene cluster in the filamentous cyanobacterium Nostoc sp. strain PCC 73102. FEMS Microbiol. Lett., 201(1):59-64.
108. Blatch, G.L. and Lassle, M. (1999). The tetratricopeptide repeat: a structural motif mediating protein-protein interactions. Bioessays, 21(11):932-939.
109. Das, A.K., Cohen, P.W. and Barford, D. (1998). The structure of the tetratricopeptide repeats of protein phosphatase 5: implications for TPR-mediated protein-protein interactions. EMBO J., 17(5):1192-1199.
110. Thiemermann, S., Dernedde, J., Bernhard, M., Schroeder, W., Massanz, C. and Friedrich, B. (1996). Carboxyl-terminal processing of the cytoplasmic NAD-reducing hydrogenase of Alcaligenes eutrophus requires the hoxW gene product. J. Bacteriol., 178(8):2368-2374.
111. Wunschiers, R., Batur, M. and Lindblad, P. (2003). Presence and expression of hydrogenase specific C-terminal endopeptidases in cyanobacteria. BMC Microbiol., 3:8.
112. Yang, F., Hu, W., Xu, H., Li, C., Xia, B. and Jin, C. (2007). Solution structure and backbone dynamics of an endopeptidase HycI from Escherichia coli: implications for mechanism of the [NiFe] hydrogenase maturation. J. Biol. Chem., 282(6):3856-3863.
113. Sveshnikov, D.A., Sveshnikova, N.V., Rao, K.K. and Hall, D.O. (1997). Hydrogen metabolism of mutant forms of Anabaena variabilis in continuous cultures and under nutritional stress. FEMS Microbiol. Lett., 147(2):297-301.
114. Wu, W.W., Wang, G., Baek, S.J. and Shen, R.F. (2006). Comparative study of three proteomic quantitative methods, DIGE, cICAT, and iTRAQ, using 2D gel- or LC-MALDI TOF/TOF. J. Proteome. Res., 5(3):651-658.
115. Seth, G., Philp, R.J., Lau, A., Jiun, K.Y., Yap, M. and Hu, W.S. (2007). Molecular portrait of high productivity in recombinant NS0 cells. Biotechnol. Bioeng., 97(4):933-951.
116. Choe, L.H., Aggarwal, K., Franck, Z. and Lee, K.H. (2005). A comparison of the consistency of proteome quantitation using two-dimensional electrophoresis and shotgun isobaric tagging in Escherichia coli cells. Electrophoresis, 26(12):2437-2449.
117. Valladares, A., Muro-Pastor, A.M., Herrero, A. and Flores, E. (2004). The NtcA-dependent P1 promoter is utilized for glnA expression in N2-fixing heterocysts of Anabaena sp. strain PCC 7120. J. Bacteriol., 186(21):7337-7343.

66
118. Tumer, N.E., Robinson, S.J. and Haselkorn, R. (1983). Different promoters for the Anabaena glutamine-synthetase gene during growth using molecular or fixed nitrogen. Nature, 306(5941):337-342.
119. Reyes, J.C. and Florencio, F.J. (1995). Electron-transport controls transcription of the glutamine-synthetase gene (glna) from the cyanobacterium Synechocystis sp. PCC 6803. Plant Mol. Biol., 27(4):789-799.
120. Summers, M.L., Wallis, J.G., Campbell, E.L. and Meeks, J.C. (1995). Genetic evidence of a major role for glucose-6-phosphate dehydrogenase in nitrogen fixation and dark growth of the cyanobacterium Nostoc sp. strain ATCC 29133. J. Bacteriol., 177(21): 6184-6194.
121. Curatti, L., Giarrocco, L. and Salerno, G.L. (2006). Sucrose synthase and RuBisCo expression is similarly regulated by the nitrogen source in the nitrogen-fixing cyanobacterium Anabaena sp. Planta, 223(5):891-900.
122. Cumino, A.C., Marcozzi, C., Barreiro, R. and Salerno, G.L. (2007). Carbon cycling in Anabaena sp. PCC 7120. Sucrose synthesis in the heterocysts and possible role in nitrogen fixation. Plant Physiol., 143(3):1385-1397.
123. Curatti, L., Flores, E. and Salerno, G. (2002). Sucrose is involved in the diazotrophic metabolism of the heterocyst-forming cyanobacterium Anabaena sp. FEBS Lett., 513(2-3):175-178.
124. Marcozzi, C., Cumino, A.C. and Salerno, G.L. (2009). Role of NtcA, a cyanobacterial global nitrogen regulator, in the regulation of sucrose metabolism gene expression in Anabaena sp. PCC 7120. Arch. Microbiol., 191: 255-63.
125. Eisbrenner, G. and Bothe, H. (1979). Modes of electron-transfer from molecular-hydrogen in Anabaena cylindrica. Arch. Microbiol., 123(1):37-45.
126. Houchins, J.P. and Burris, R.H. (1981). Light and dark reactions of the uptake hydrogenase in Anabaena 7120. Plant Physiol., 68(3):712-716.
127. Houchins, J.P. and Hind, G. (1982). Pyridine-nucleotides and H2 as electron-donors to the respiratory and photosynthetic electron-transfer chains and to nitrogenase in Anabaena heterocysts. Biochimica Et Biophysica Acta, 682(1):86-96.
128. Jones, K.M. and Haselkorn, R. (2002). Newly identified cytochrome c oxidase operon in the nitrogen-fixing cyanobacterium Anabaena sp. strain PCC 7120 specifically induced in heterocysts. J. Bacteriol., 184:2491-2499.
129. Telor, E. and Stewart, W.D.P. (1977). Photosynthetic components and activities of nitrogen-fixing isolated heterocysts of Anabaena cylindrica. Proc. R. Soc. Lond. Series B-Biol. Sci., 198(1130):61-86.
130. del Castillo, T., Ramos, J.L., Rodriguez-Herva, J.J., Fuhrer, T., Sauer, U. and Duque, E. (2007). Convergent peripheral pathways catalyze initial glucose catabolism in Pseudomonas putida: genomic and flux analysis. J. Bacteriol., 189(14):5142-5152.

67
131. Kangatharalingam, N., Priscu, J.C. and Paerl, H.W. (1992). Heterocyst envelope thickness, heterocyst frequency and nitrogenase activity in Anabaena flos-aquae - Influence of exogenous oxygen tension. J.Gen.Microbiol., 138:2673-2678.
132. Golden, J.W. and Yoon, H.S. (2003). Heterocyst development in Anabaena. Curr. Opin. Microbiol., 6(6):557-563.
133. Jiang, F., Mannervik, B. and Bergman, B. (1997). Evidence for redox regulation of the transcription factor NtcA, acting both as an activator and a repressor, in the cyanobacterium Anabaena PCC 7120. Biochem. J., 327:513-517.
134. Alfonso, M., Perewoska, I. and Kirilovsky, D. (2001). Redox control of ntcA gene expression in Synechocystis sp. PCC 6803. Nitrogen availability and electron transport regulate the levels of the NtcA protein. Plant Physiol., 125(2):969-981.
