integrated stratigraphy.pdf
TRANSCRIPT

New Zealand Journal of Geology & Geophysics, 1999, Vol. 42: 581-6140028-8306/99/4204-0581 $7.00/0 © The Royal Society of New Zealand 1999
581
Integrated stratigraphy of the Waitakian-Otaian Stage boundary stratotype,Early Miocene, New Zealand
H. E. G. MORGANS1
A. R. EDWARDS2
G. H. SCOTT1
I. J. GRAHAM3
P. J. J. KAMP4
T. C. MUMME1*
G. J. WILSON1
G. S. WILSON1
'institute of Geological & Nuclear Sciences LtdP.O. Box 30 368Lower Hutt, New Zealand
2Stratigraphic Solutions LtdP.O. Box 295Waikanae, New Zealand
institute of Geological & Nuclear Sciences LtdP.O. Box 31 312Lower Hutt, New Zealand
4Department of Earth SciencesUniversity of WaikatoPrivate Bag 3105Hamilton, New Zealand
*Present address: 47 Parkvale Road, Karori, Wellington,New Zealand.
Abstract The base of the type section of the Otaian Stageat Bluecliffs, South Canterbury, is recognised as thestratotype for the boundary between the Waitakian andOtaian Stages. Principal problems with the boundary are therestriction of existing bioevent proxies to shelf and upperslope environments and its uncertain age. These topics areaddressed by a multidisplinary study of a 125 m sectionabout the boundary, which examines its lithostratigraphy,depositional setting, biostratigraphy, correlation, andgeochronology.
The lower siltstone lithofacies (0-38.5 m) was depositedat upper bathyal depths (200-600 m) in a marginal basinwhich was partially sheltered from fully oceanic circulationby a submarine high and islands. The site was covered bycool-temperate water and was probably adjacent to theSubtropical Convergence. This unit is succeeded by thebanded lithofacies (38.5-106 m) and the upper siltstone
G 98007Received 26 February 1998; accepted 3 June 1999
lithofacies (basal 19 m studied). Paleodepth probablydeclined up-sequence, but deposition at shelf depths is notdefinitely indicated. A cyclic pattern of abundance spikes inbenthic and planktonic foraminifera commences 9 m abovebase and extends to 73 m in the banded lithofacies. Oxygenisotope excursions (up to 2.08%o) in Euuvigerina miozea andCibicides novozelandicus are greatest within the intervalcontaining the abundance spikes. The stage boundary occursin the banded lithofacies at the highest abundance spike(73 m). Although condensed intervals might affect thecompleteness of the section, they are not associated withsedimentary discontinuities, and we consider that the sectionis suitable as a biostratigraphic reference.
Spores, pollens, dinoflagellates, calcareous nannofossils,foraminifera, bryozoans, and ostracods are preserved nearthe boundary, but molluscs principally occur higher, in theshallower upper siltstone lithofacies. Siliceous microfossilsare rare. There is considerable scope for further biostrati-graphic research.
The primary event marking the boundary at 73 m is theappearance of the benthic foraminifer Ehrenberginamarwicki. This is a distinctive and widely distributed eventbut is restricted to shelf and upper bathyal environments.Supplementary events in planktonic foraminifera andcalcareous nannofossils were researched. Highest occur-rences of Globigerina brazieri and G. euapertura arerecorded at 47 and 58 m. There is a marked decline in relativeabundance of Paragloborotalia spp. at 62 m. Helicosphaeracarteri becomes more abundant than H. euphratis between56 and 87 m. These events are not exact proxies for theboundary but they may usefully indicate proximity to it. Theyoccur in the interval of prominent spikes in foraminiferalabundance.
The Waitakian-Otaian boundary is dated at 21.7 Ma bystrontium isotopes. Stable primary remanence could not bedetermined in a pilot paleomagnetic study of Bluecliffsspecimens. However, specimens trended towards reversedpolarity, and remagnetisation great circle analysis will allowdirections to be calculated in future collections.
Keywords Oligocene; Miocene; Waitakian; Otaian; stageboundary stratotype; lithostratigraphy; biostratigraphy;geochronology; integrated stratigraphy; Bluecliffs Silt;Otekaike Limestone; planktonic foraminifera; benthicforaminifera; calcareous nannofossils; Mollusca;palynoflora; dinoflagellates; Radiolaria; oxygen isotopes;carbon isotopes; strontium isotopes
INTRODUCTION
New Zealand Cenozoic stages were developed as biostrati-graphic units based on local bioevents to overcome problemsin recognising European stages in a distant biogeographicprovince. Pioneering work by Thomson (1916) and Allan

582 New Zealand Journal of Geology and Geophysics, 1999, Vol. 42
South Island, New Zealand
100 km
44 S-- Blue Cliffs
trigZ, Otiake46S-
170E 174'E
53
Craigmore Limestone section
3 4 -
& stream section"J39/f123-f146 f216 foi?
Fig. 1 Location of Bluecliffsexposures, Otaio River, showin j,sections sampled: "streansection", "cliff section',Craigmore Limestone section, andlocation of isolated samples.Access to Bluecliffs is fromMacAlwees Crossing Road.NZMS 260 co-ordinates for thebase of "stream section": J39/507333.
(1933) used brachiopod and mollusc distributions, but theresolution and biofacies coverage of the classification wereconsiderably enhanced by inclusion of foraminiferal events(Finlay & Marwick 1940, 1947). Following the latter work,the classification stabilised and became the standardstratigraphic reference in New Zealand Cenozoic geology.A large amount of earth science research has been reportedusing local stages (e.g., Suggateetal. 1978; Field etal. 1989).As well, the classification provides a framework for single-group biostratigraphies (e.g., Edwards 1971; Wilson 1984).Significantly, the local stages now act as a de factochronostratigraphic classification for Cenozoic time in NewZealand (Edwards et al. 1988; Morgans et al. 1996). Forstages to perform this role effectively it is necessary todefine, evaluate, and calibrate boundaries, and to identifybioevents that are suitable time proxies. The Waitakian-Otaian boundary has high priority for early review.Biostratigraphically, it is defined by benthic foraminiferalevents that occur primarily in shelf and upper bathyalsequences. More widely dispersed substitutes are needed.Currently, poor biostratigraphic resolution constrains studiesof the evolution of deep-water Early Miocene basins thatare prospective for hydrocarbons. Chronostratigraphically,the age of the boundary is poorly constrained because ofthe absence of geomagnetic polarity, isotopic or bioeventproxy data; this impacts on geohistory modelling. Edwardset al. (1988) rather arbitrarily located the boundary at 22 Ma,a moderately close assignment in the light of the presentresearch.
In this study we integrate data relevant to these problems.The boundary stratotype section is studied to identify itsdepositional setting, completeness, and suitability as achronostratigraphic and biostratigraphic reference. Theposition of the boundary within the section and itsenvironmental history are examined. Dating is attemptedusing strontium isotopes, oxygen isotopes, and magneto-stratigraphy. Major fossil groups present are reviewed withspecial attention to nannofossils and planktonic foraminifera.
The performance of the present boundary events is evaluated.We describe changes in planktonic taxa close to the presentboundary that might serve as proximity events and extendits utility. The boundary is correlated with other regionaland global biostratigraphic classifications.
Waitakian and Otaian StagesAllan (1933) showed that Park's (1918) original usage ofWaitakian was ambiguous, biostratigraphically. He resolvedthe problem by proposing that Waitakian should refer to"...the period of time represented by the deposition of theWaitaki Limestone and the Otiake beds of Waitaki Valley...'.He cited the Trig Z section (Fig. 1) as the most completereference, but noted that Waitakian, as redefined, might notbe contiguous with the overlying Hutchinsonian Stage. Gage(1957) revised the lithostratigraphy of the Waitaki valleyand proposed Otekaike Limestone for Waitaki Limestone.Four members were recognised, the uppermost (WaitouraMarl) having its type section at Trig Z.
The origin of the Otaian Stage dates from Allan's (1933)surmise that there might be a gap in the biostratigraphicclassification between Waitakian and Hutchinsonian Stages.Finlay (1939) cited supporting brachiopod evidence, andFinlay & Marwick (1940) gave criteria that distinguishedlower Hutchinsonian from Hutchinsonian s.s. Finlay &Marwick (1947) proposed the Otaian Stage for the informallower Hutchinsonian of Finlay & Marwick (1940) and citedBluecliffs as type locality (Otaio River, South Canterbury,Fig. 1). They stated that the "...lower limit is marked by thelowest occurrence of (primarily) Ehrenbergina and(secondarily) Spiroloculina novo-zealandica (Cush. andTodd)" and noted that the stage extends from "...somewhatbelow the middle of the banded unit at Blue Cliffs...upto the 'Brown Sandstone' which is Hutchinsonian."Importantly, Finlay & Marwick (1947) stated that "...thelower part of the banded beds is Waitakian." Gair (1959)proposed Bluecliffs Silt and Southburn Sand for thelithostratigraphic units referred to by Finlay & Marwick

•3 < — & (/Q T)
f 2. K o. 3 M— » 3 Sp K»
=3 ~ £ -"'
ofa
gft
,-a ~a-o3 C/) -^ gj
a. o -no|.3
oo
8J_I_L
8 Metresi i. i ' i '
1 •Hardbands starting at base of "cliff section' in I
CO 03 W 00M u ^ CJi
I XXXI I X03 COCO DO 03 0 3 CD
I I X X03 DO 03 03
<O O -* M ro ro ro
« S 2 £ &2 2223 2 222 2 2 22 2 22 2 2 =CO A 4 J £ £k Ui Cn (71 (Ji C7I (Ji Cft O> <3) O) CD O l ^
S - ^ N » C O u i ^i < o o ro •& oi ~>i t o e aSa 3 alsi Is I sis I I RS KS r3 rt Foraminifera ri?1 (this study) 2
i i2 2 2 222 2 2 2
SS5 £ 2 a2 2 2 2 2 2 22 5 i i t i I i F3 F3
' Calcareous5 nannoplanktono> (this study)
a"4 I"*
3J-1 2 -*
I? I32-h 5— 2-o * oi cn^ J CO CD
JC7]~ 5oT 30T Strontium isotope dating ^ '?S cnS S S of benthic foraminifera, coSiS"} J i * m"1 * = planktonics S^1
S co S rv)N Z M S 1S111 /
Jenkins (1971; table 32,column 18) NZMS1,(NZMS 260 = f7***)
3 3 j Pocknall (1982)
u>—i 1 i i i — r ~ i 1 r~r~i 1—i—n—i—i i i i 1 MM—m 1 1 i i i i—i—i—i—i—n—i 1 1 1—
2 2 2 2222 2 222 2 2 2222222 2 2222222 2 22222222222 2 2 2 203 CO CO frf A ^ ^ U1U1UI UI UI UI O> OS 0 ) 0 ) 0 1 O> O> C5 ~J~-I^J^I^J-J ^ l -J COCOCOCtS Q3 00 00 00 OOOO (O V (O CO
o> <j to fo^oi->i v -tfcco ui -vi o o - ' i o u l c n >>i co o-*wco*.ui ^ too-^rocoS cn cn joo<o -* £ os ©
""• ' ' ' Residue ' examined for '3 M M M siliceous 3 microfossils 3->co ft ui (J. Ashby, os pers. comm.) -Ji
2 2
5J S
—i 1 i 1 1—r~i—r~i 1 n \—i—n 1 r~i—
i i s si i sii^ I ss a is s is2 22 2 22 2 22 2 2 2 2 22 2 2 2
g sa s 23i 3 as & t a s ss 2 nF5 B BS ^ u
_. Stable isotope samples,S * = Cibicides only;^ ** = Euuvigerina only
(f116 = Craigmore Limestone; no dinoflagellates:f121 = Waitoura Marl)
Dinoflagellate samples
Paleomagneticorientated cores(Mumme1997)
m 3 » g 2^ ^ g i i ^•-1 S <»
GS3699 (f496) = lower thirdof banded beds, west end
of section
f593 = GS 9222 f592 = GS 9223 Macrofossil collections(IGNS files)

584 New Zealand Journal of Geology and Geophysics, 1999, Vol. 42
(1947). Hutchinsonian Stage was incorporated in AltonianStage by Scott (1972).
The stratotypes for Waitakian and Otaian Stages arecomponents of the same sedimentary sequence (Carter 1988;Field et al. 1989) but are 60 km apart in Canterbury Basin(Fig. 1). However, there is no ambiguity as to the locationof the boundary stratotype. Waitoura Marl, the highestWaitakian unit at Trig Z, lacks Otaian fossils and correlateswith strata near Bluecliffs that are significantly below theWaitakian-Otaian boundary. At Bluecliffs, the boundary,following Finlay & Marwick (1947), is well defined andthe surrounding section is well exposed. We recognise theboundary stratotype for the Waitakian and Otaian Stages asthe horizon at 73 m (Fig. 2) defined by the primary eventfor the base of the Otaian Stage, namely, the first appearancedatum (FAD) of the benthic foraminifer Ehrenberginamarwicki.
The Waitakian and Otaian Stages are regarded asbiostratigraphic units, defined by homotaxial biostratigraphicevents whose chronostratigraphic value is yet to beestablished. Therefore, we use "lower", "middle", and"upper" for their informal subdivisions.
MATERIAL AND METHODS
CollectionsSamples were taken for biostratigraphic, oxygen, carbon,and strontium isotope analysis, and for a pilot study ofpaleomagnetism (Fig. 2). We focused on the sectionspanning the Waitakian-Otaian boundary ("cliff section",Fig. 1), the preceding "stream section", and isolatedexposures down to Otekaike Limestone (Fig. 3). A further220 m of Bluecliff Silt overlies the cliff section, althoughmuch of it is poorly exposed. Previous collectors (e.g., Scott1965; Shelley 1974) commenced at the base of the cliffsection (Fig. 1; NZMS 260 grid ref. J39/5108-3351). Weextended our sampling further upstream and into a tributarystream ("stream section") and included a further 35 m ofstrata below the cliff section. Sample density varied between1 and 2 m; 35 magnetostratigraphic samples and 94biostratigraphic samples (J39/f 123—217) were taken.Material from the foraminiferal samples was used for isotopestudies.
Collections were also made for biostratigraphic controlthrough Craigmore Limestone (J39/fll4—120) and withinWaitoura Marl between Craigmore Limestone and the baseof the Bluecliffs section (J39/fl21-122).
Fossil groups examined and previous studiesForaminifer a
The section is highly fossiliferous and especially rich incalcareous groups. Planktonic taxa (Table 1) are consistentlymore abundant than benthic taxa (Table 2), typically rangingbetween c. 45 and 85% of total foraminiferal fauna. Benthicforaminifera become more abundant in the upper part of thesection. Previous foraminiferal studies by Finlay & Marwick(1947), Scott (1958, 1965, 1973), Jenkins (1971), andShelley (1974) underpin our present understanding of theWaitakian-Otaian boundary and its position at Bluecliffs.
NannofossilsCoccoliths, and associated calcareous nannofossils,constitute a small and variable part of the samples. Their
500-
400-
Fig. 3 Lithostratigraphic nomenclature for section at Bluecliffsand for underlying units.
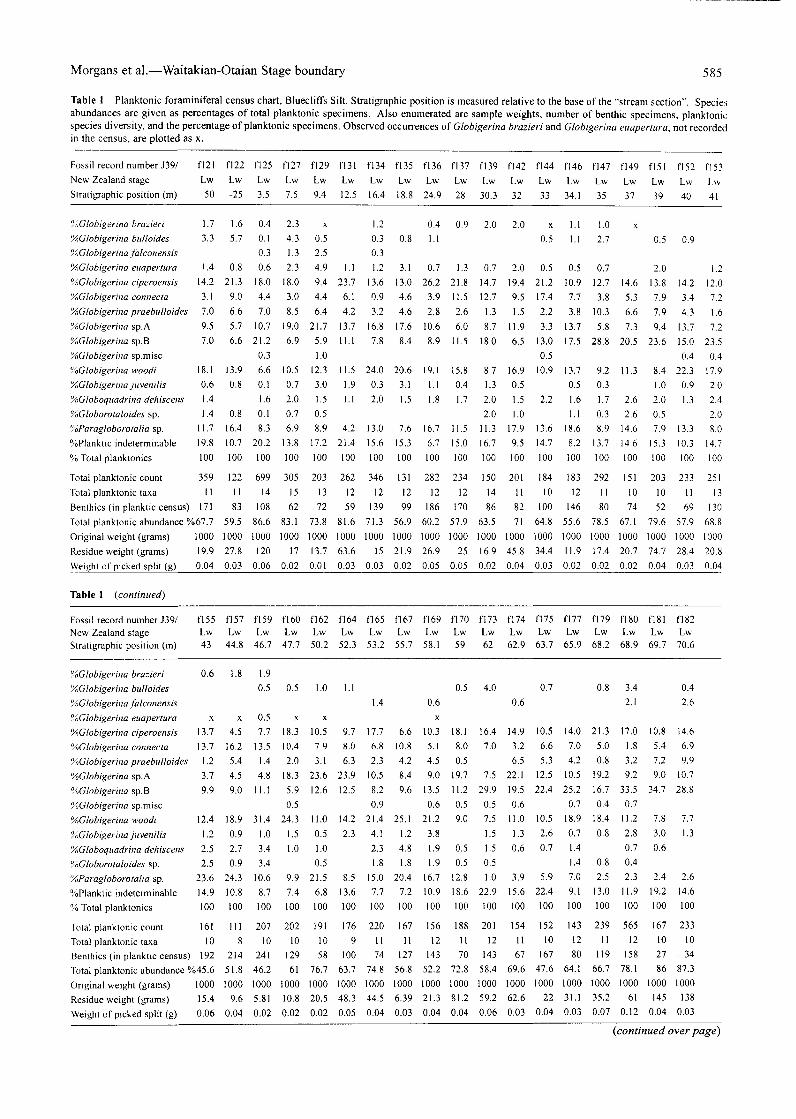
Morgans et al.—Waitakian-Otaian Stage boundary 585
Table 1 Planktonic foraminiferal census chart, Bluecliffs Silt. Stratigraphic position is measured relative to the base of the "stream section". Speciesabundances are given as percentages of total planktonic specimens. Also enumerated are sample weights, number of benthic specimens, planktonicspecies diversity, and the percentage of planktonic specimens. Observed occurrences of Globigerina brazieri and Globigerina euapertura, not recordedin the census, are plotted as x.
Fossil record number J39/New Zealand stageSlratigraphic position (m)
f!21 fl22 fl25 fl27 fl29 f 131 f 134 f 135 fl36 f 137 f 139 fl42 fl44 f!46 f!47 fl49 fl51 fl52 fl53Lw Lw Lw Lw Lw Lw Lw Lw Lw Lw Lw Lw Lw Lw Lw Lw Lw Lw Lw-50 -25 3.5 7.5 9.4 12.5 16.4 18.8 24.9 28 30.3 32 33 34.1 35 37 39 40 41
'/oGlobigerina brazieri'/oGlobigerina bulloides'/oGlobigerina falconensis'/oGlobigerina euapertura"/oGlobigerina ciperoensis"/oGlobigerina connecta"/oGlobigerina praebulloides"/oGlobigerina sp.A"/oGlobigerina sp.B"/oGlobigerina sp.misc"/oGlobigerina woodi"/oGlobigerina juvenilis'/oGloboquadrina dehiscens'/oGloborotaloides sp.'/oParagloborotalia sp.%Planktic indeterminable% Total planktonics
Total planktonic countTotal planktonic taxaBenthics (in planktic census)Total planktonic abundance %67.7Original weight (grams) 1000Residue weight (grams) 19.9
Weight of picked split (g) 0.04
1.7
3.3
1.4
14?
3.1
7 0
9.57.0
18.10.6
1.4
1.4
11 7198
100
35911
171
1.6
5.7
0.821.3
9.06.65.7
6.6
13.90.8
0.8
16.410.7100
122
11
83
0.4
0.1
0.3
0.6
18.04.4
7.010.721.2
0.3
6.6
0.1
1.6
0.1
8.3
20.2100
699
14
108
2.3
4.3
1.3
2.3
18.03.0
8.519.06.9
10.50.7
2.0
0.7
6.9
13.8100
30515
62
X
0.5
2.5
4.9
9.4
4.4
6.4
21 75.9
1.0
12.33.0
1.5
0.5
8.9
17.2100
2031372
1.1
23.76.1
4.2
13.711.1
11.51.9
1.1
4.2
21.4100
262
1259
1.2
0.3
0.3
1.2
13.60.9
3.216.87.8
24.00.3
2.0
13.015.6100
34612
139
0.8
3.1
130
4.6
4.617.68.4
20.63.1
1.5
7.6
15.3100
131
1299
0.4
1.1
0.7
26 23.9
2.8
10.68.9
19.11.1
1.8
16.76.7
100
28212
186
0.9
1.3
21 8
11.5
266.0
11.5
15.80.4
1.7
11 515 0100
234
12
170
2.0
0.7
14 712.7
1 3
8.7
18.0
8.7
1.3
2.0
2.0
11 3167
100
150
14
86
2.0
2.0
194
9.5
1 5
11.96.5
16.90.5
1.5
1.0
179
0 5
100
201
11
82
X
0.5
0.5
21 217.4
22
3.313.00.5
10.9
2.2
13.614.7100
184
10
100
1.1
1.1
0.5
109
7.7
3 8
13.717.5
13.70.5
1.6
1.1
18.68.2
100
18312
146
1.0
2.7
0.7
P 73.8
10 3
5.828.8
9.2
0.3
1.7
0.3
8.9
13.7100
29211
80
X
14.65.3
6 6
7320.5
11.3
2.6
2.6
14.614.6100
151
10
74
0.5
2.0
13.87.9
7.9
9 4
23.6
8.4
1.0
2.0
0.5
7.9
15.3100
2031052
0.9
14.23.4
4.3
13 715.00.4
22.30.9
1.3
13.310.3100
233
11
69
1.2
12.07.2
1.6
23.50.4
17.92.0
2.4
2.0
8.0
14.7100
251
13
130
59.5 86.6 83.1 73.8 81.6 71.3 56.9 60.2 57.9 63.51000 1000 1000 1000 1000 1000 1000 1000 1000 100027.8 120 17 13.7 63.6 15 21.9 26.9 25 16.90.03 0.06 0.02 0.01 0.03 0.03 0.02 0.05 0.05 0.02
71 64.8 55.6 78.5 67.1 79.6 57.9 68.8
1000 1000 1000 1000 1000 1000 1000 100045.8 34.4 11.9 17.4 20.7 74.7 28.4 20.80.04 0.03 0.02 0.02 0.02 0.04 0.03 0.04
Table 1 (continued)
Fossil record number J39/New Zealand stageStratigraphic position (m)
fl55 fl57 fl59 fl60 fl62 fl64 fl 65 fl67 fl69 fl70 fl73 f 174 fl75 fl77 fl79 f 180 fl81 f 182Lw Lw Lw Lw Lw Lw Lw Lw Lw Lw Lw Lw Lw Lw Lw Lw Lw Lw43 44.8 46.7 47.7 50.2 52.3 53.2 55.7 58.1 59 62 62.9 63.7 65.9 68.2 68.9 69.7 70.6
"/oGlobigerina brazieri"/oGlobigerina bulloides
'/oGlobigerina falconensis
"/oGlobigerina euapertura
"/oGlobigerina ciperoensis"/oGlobigerina connecta"/oGlobigerina praebulloides
"/oGlobigerina sp.A"/oGlobigerina sp.B"/oGlobigerina sp.misc"/oGlobigerina woodi
"/oGlobigerina juvenilis
"/oGloboquadrina dehiscens'/oGloborotaloides sp."/oParagloborotalia sp.%Planktic indeterminable% Total planktonics
Total planktonic countTotal planktonic taxaBenthics (in planktic census)
0.6
X
13.713.7
1.2
3.7
9.9
12.4
1.2
2.52.5
23.614.9100
161
10
192
Total planktonic abundance %45.6Original weight (grams)Residue weight (grams)Weight of picked split (g)
100015.40.06
1.8
X
4.5
16.25.4
4.5
9.0
18.9
0.92.7
0.9
24.310.8100
111
8
214
51.81000
9.6
0.04
1.90.5
0.5
7.7
13.51.4
4.8
11.1
31.4
1.0
3.43.4
10.68.7
100
207
10
241
46.210005.810.02
0.5
X
18.310.42.0
18.35.9
0.5
24.3
1.5
1.0
9.9
7.4
100
202
10
129
61
100010.80.02
1.0
X
10.57.9
3.1
23.612.6
11.00.5
1.0
0.5
21.56.8100
191
10
58
76.7100020.50.02
1.1
9.7
8.0
6.3
23.912.5
14.22.3
8.5
13.6100
176
9
100
63.7100048.30.05
1.4
17.76.8
2.3
10.58.20.9
21.4
4.1
2.3
1.8
15.07.7
100
220
11
74
74.8100044.50.04
6.6
10.84.2
8.4
9.6
25.11.2
4.8
1.8
20.47.2
100
167
11
127
56.810006.390.03
0.6X
10.35.1
4.5
9.0
13.50.6
21.23.8
1.9
1.9
16.710.9100
15612
143
52.2100021.30.04
0.5
18.18.0
0.5
19.711.20.5
9.0
0.5
0.5
12.818.6100
188
11
70
72.8100081.20.04
4.0
16.47.0
7.5
29.90.5
7.5
1.5
1.5
0.5
1.0
22.9100
201
12
143
58.4100059.20.06
0.6
14.93.2
6.5
22.119.50.6
11.01.3
0.6
3.9
15.6100
154
11
67
69.6100062.60.03
0.7
10.56.6
5.3
12.522.4
10.52.6
0.7
5.9
22.4100
152
10
167
47.61000
22
0.04
14.07.0
4.2
10.525.2
0.7
18.90.7
1.4
1.4
7.0
9.1
100
143
12
80
64.1100031.10.03
0.8
21.35.0
0.8
19.216.70.4
18.40.8
0.8
2.5
13.0100
239
11
119
66.7100035.20.07
3.4
2.1
17.01.8
3.2
9.233.5
0.7
11.22.80.7
0.4
2.311.9100
56512
158
78.11000
61
0.12
10.85.4
7.2
9.0
34.7
7.8
3.0
0.6
2.4
19.2100
167
10
27
86
1000145
0.04
0.4
2.6
14.66.9
9.9
10.728.8
7.7
1.3
2.614.6100
233
10
34
87.31000
138
0.03
{continued over page)
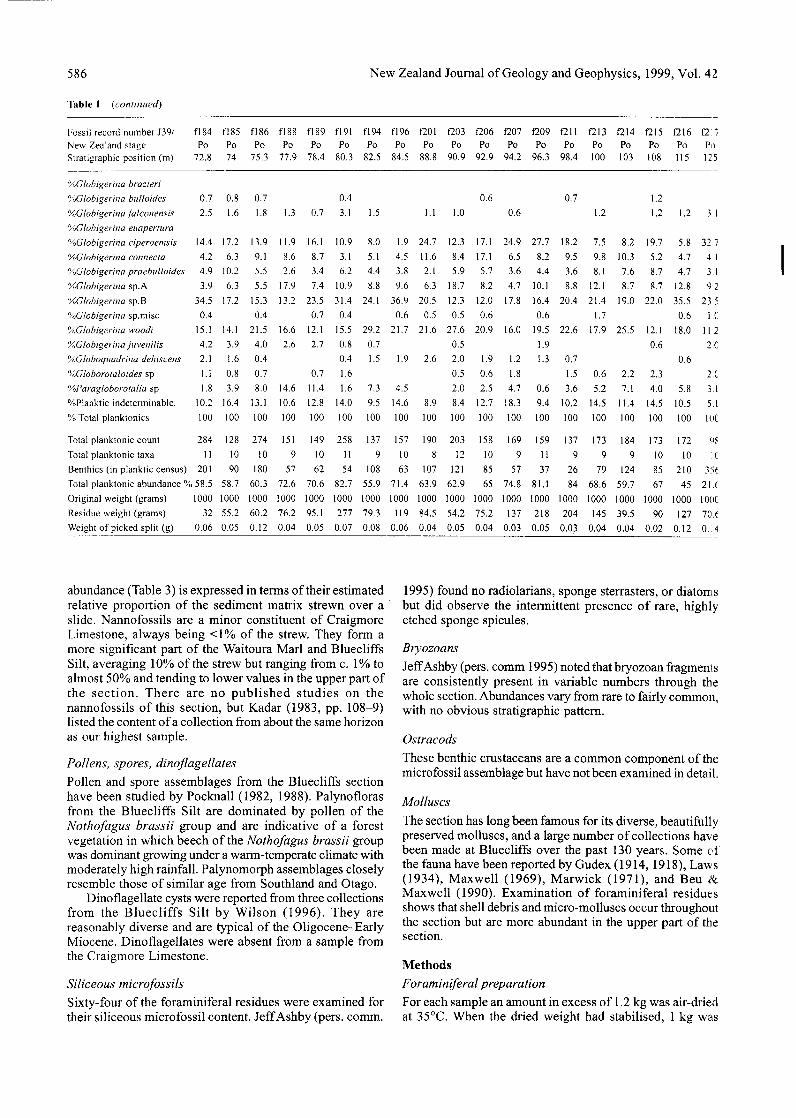
586 New Zealand Journal of Geology and Geophysics, 1999, Vol. 42
Table 1 (continued)
Fossil record number .139/New Zealand stageStratigraphic position (m)
'AiGlobigerina brazieri
"AiGlobigerina bidloides
'AiGlobigerina falconensis'AiGlobigerina euapet'tura
'AiGlobigerina ciperoemsis
'AGlobigerina connecta"AiGlobigerina praebidloides
"AiGlobigerina sp.A
"AiGlobigerina sp.B"AiGlobigerina sp.misc"AiGlobigerina woodi'AiGlobigerina juvenilis'AiGloboquadrina dehiscens
%Globorotaloides sp'AiParagloborolalia sp%Planktic indeterminable% Total planktonics
Total planktonic countTotal planktonic taxaBenthics (in planktic census)Total planktonic abundance %Original weight (grams)Residue weight (grams)Weight of picked split (g)
fl84Po
72.8
0.7
2.5
14.44 ~>
4.9
3.9
34.50.4
15.14.2
2.1
1.1
1.8
102
100
284
11
201
58.51000
32
0 06
t'185Po74
0.81.6
17.263
10.26.3
17.2
14.13.91.60.83.9
164
100
1281090
58.7100055.2OOS
fl86Po
75.3
0.71.8
13 9
9 15.5
5.5
15.30.4
21.54.0
0.4
0.7
8.0
13 1100
274
10
180
60.3100060.20 P
fl88Po
77 9
1.3
11 98 62.6
17.9
13.2
16.62.6
14.6106100
1519
5772.6100076.20 04
fl89Po
78 4
0.7
16 18 73.47.4
23.50.7
12.12.7
0.711.412 8
100
149
1062
70.6100095.10 05
fl91Po
80 3
0.43.1
1093 1
6.210.9
31 40.4
15.50.80.4
1.61.6
140100
25811
5482.71000277
0 07
fl94Po
87 5
1.5
805 1
4.4
8.824 1
29.20.71.5
7.395
100
1379
10855.9
100079.30.08
fl96Po
84 5
1 94 53.89.6
36 90.6
21.7
1.9
4.5146100
1571063
71.4
1000119
0.06
f201Po
88 8
1.1
24 7
11 62.1
6.320 50.5
21.6
2.6
8 9100
1908
10763.9
100084.50.04
f203Po
90.9
1.0
123845.9
18.712 30.5
27.60.52.00.52.084
100
20312
12162.9100054.20.05
f206Po
92.9
0.6
17 117 15.78.2
1200.6
20.9
1.9
0.62.5
127100
158108565
100075.20.04
f207Po
94.2
0.6
24 9653.64.7
178
16.0
1.2
1.84.7
183
100
1699
5774.81000137
0 03
f209Po
96 3
27 7824.4
10.1
1640.6
19.51.91.3
0.694
100
15911
3781.11000218
0 05
f211Po
98 4
0.7
18 29 5
3.68.8
20 4
22.6
0.71.53.6
102
100
1379
2684
1000204
0 03
f213Po100
1.2
7.59.88.1
12.1
21 4
1.717.9
0.65.2
14.5
100
1739
79
68.6
1000
1450 04
f214
Po103
8.210.3
7.68.7
190
25.5
2.2
7.111.4
100
1849
124
59.7
1000
39.5
0.04
f215Po108
1.21.2
19.7
5.28.78.7
22.0
12.1
0.6
2.34.0
14.5
100
173108567
1000
900.02
f216Po115
1.2
5.84.74.7
12.8
35.5
0.618.0
0.6
5.810.5
100
17210
21045
1000
1270 1?
PD
125
3,1
32.7
4,1
3.192
23.5
l.C
11.2
2.C
2.C3.15.1
IOC
OS
;c3,«
21.(
100C
70.<
0 4
abundance (Table 3) is expressed in terms of their estimatedrelative proportion of the sediment matrix strewn over aslide. Nannofossils are a minor constituent of CraigmoreLimestone, always being <1% of the strew. They form amore significant part of the Waitoura Marl and BluecliffsSilt, averaging 10% of the strew but ranging from c. 1% toalmost 50% and tending to lower values in the upper part ofthe section. There are no published studies on thenannofossils of this section, but Kadar (1983, pp. 108-9)listed the content of a collection from about the same horizonas our highest sample.
Pollens, spores, dinoflagellates
Pollen and spore assemblages from the Bluecliffs sectionhave been studied by Pocknall (1982, 1988). Palynoflorasfrom the Bluecliffs Silt are dominated by pollen of theNothofagus brassii group and are indicative of a forestvegetation in which beech of the Nothofagus brassii groupwas dominant growing under a warm-temperate climate withmoderately high rainfall. Palynomorph assemblages closelyresemble those of similar age from Southland and Otago.
Dinoflagellate cysts were reported from three collectionsfrom the Bluecliffs Silt by Wilson (1996). They arereasonably diverse and are typical of the Oligocene-EarlyMiocene. Dinoflagellates were absent from a sample fromthe Craigmore Limestone.
Siliceous microfossils
Sixty-four of the foraminiferal residues were examined fortheir siliceous microfossil content. Jeff Ashby (pers. comm.
1995) found no radiolarians, sponge sterrasters, or diatomsbut did observe the intermittent presence of rare, highlyetched sponge spicules.
Bryozoans
Jeff Ashby (pers. comm 1995) noted that bryozoan fragmentsare consistently present in variable numbers through thewhole section. Abundances vary from rare to fairly common,with no obvious stratigraphic pattern.
Ostracods
These benthic crustaceans are a common component of themicrofossil assemblage but have not been examined in detail.
Molluscs
The section has long been famous for its diverse, beautifullypreserved molluscs, and a large number of collections havebeen made at Bluecliffs over the past 130 years. Some ofthe fauna have been reported by Gudex (1914, 1918), Law s(1934), Maxwell (1969), Marwick (1971), and Beu &Maxwell (1990). Examination of foraminiferal residuesshows that shell debris and micro-molluscs occur throughoutthe section but are more abundant in the upper part of thesection.
MethodsForaminiferal preparation
For each sample an amount in excess of 1.2 kg was air-driedat 35°C. When the dried weight had stabilised, 1 kg was

Morgans et al.—Waitakian-Otaian Stage boundary 587
placed in water and left for 1-2 days. This was washed overa 75 |Xm screen and the residue again air-dried at 35°C andthen weighed. This treatment provided material suitable forstable isotope studies. Most of the material was in goodcondition. Only at the base of the section and from withinor near the hard bands in the cliff section were there problemswith preservation of the foraminifera. Where these problemswere severe, extra samples were prepared.
For the biostratigraphic study, a small statistical samplewas prepared from each bulk sample using a Carpcomicrosplitter, the sample being split repeatedly until a faunaof 100-200 specimens remained on a 150 jj,m screen. Thesplits were manipulated so that both planktonic and benthicsplits each contained 100-200 specimens, although for somebenthic splits, the final counts had fewer than 100 specimens.The number of benthic specimens in the final >150 .m split,and its weight, are given in Table 2. Planktonic foraminiferaare recorded as percentages in Table 1. All other fossilmaterial in the sample encountered during the census wasretained. This material consisted mainly of ostracods, whichwere counted in the census, as well as broken molluscs,which were not examined further.
One-half of all residues has been kept intact for furtherstudies. The remainder of the other splits, from which thestatistical samples were counted, were utilised for the isotopestudies. All calcareous benthic material in each sample above500 (j.m was taken for strontium isotope analysis, includingsome selected planktonic specimens. For oxygen and carbonisotope studies, undertaken at University of Waikato, we usedthe benthic foraminifera Euuvigerina spp. (Fig. 4J, K) andCibicides novozelandicus (Fig. 4M). Acquisition of thismaterial provided a more extensive conspectus ofassemblages than was gained during the census.
Nannofossil preparation
The suite of samples examined included 7 collected fromCraigmore Limestone and 50 from Waitoura Marl andBluecliff Silt (Fig. 2). The latter are evenly spacedstratigraphically but have a bias against the hard bands andtowards the lower and upper limits of the section. Despitethis, they are considered to be representative of the range ofenvironmental and stratigraphic changes occurring in thissection.
Standard methods were used to prepare and examinesamples selected for calcareous nannofossil analysis. A thinrepresentative slurry of the sample matrix was spread overa 22 x 32 mm coverglass, dried, and attached to a glass slideusing the permanent mounting medium Petropoxy 154.Examination was undertaken using a polarising microscopeequipped with a lambda plate to traverse the coverglass atmagnifications of 1500x (detail) and then, as appropriate,at either 900x or 750x (survey). As a minimum, each slideexamined was traversed twice at 1500x. The nature of eachassemblage was documented (Table 3) in terms of itsabundance (estimated relative cover of the smear), diversity(species number, both observed and adjusted), preservation(overall plus breakage, etching and plating), inferredenvironmental conditions (living and depositional), andbiostratigraphic position (datums and zones). Alsodocumented was the estimated abundance of each speciesor operational taxonomic unit and, as appropriate, otherrelevant information such as the nature, preservation, size,location, and inferred origin of individual specimens.
Stable isotopes
Analyses were carried out on the benthic foraminiferaCibicides novozealandicus and Euuvigerina spp. Specimenswere cleaned in an ultrasonic bath, vacuum roasted, andreacted with 100% orthophosphoric acid at 70°C for 20 minunder vacuum. The evolved CO2 was separated, purified,and isotopically analysed in a VG Micromass 602E massspectrometer at University of Waikato. The reference gaswas calibrated daily against the international standardNBS-19. The 13C and 18O isotope values are presented(Fig. 5) in conventional per mille deviations from the PDBstandard, and the analytical precision is better than ±0.05%o.
Paleomagnetism
Standard paleomagnetic samples (25 mm diameter x 22 mmheight) were collected from 5 m intervals throughout thesequence using a portable Tas drill (Mumme 1997). Aftercollection, samples were shielded until they were measuredon a two-axis ScT cryogenic magnetometer at VictoriaUniversity of Wellington (VUW). The NRM was routinelymeasured on all samples and found to be weak <2x\Q~A A/m. Six sample pairs were selected to evaluate behaviour andapplicability of thermal and alternating field (AF)demagnetisation methods. AF demagnetisation wasconducted in a Molspin AF demagnetiser, and samples weremeasured at peak fields of 5 mT to a maximum of 40 mT.Thermal demagnetisation was conducted at 50° incrementsfrom 100°C to 400°C in the VUW purpose-built zero-fieldoven. All other samples were subjected to thermaldemagnetisation only. After each thermal demagnetisationstep, the magnetic susceptibility was measured to monitorfor thermal alteration. In all measurements, susceptibilityand remanence increased markedly by 400°C of heating and,in several, increases occurred by the 350°C step of heating.Measurements after these heating steps were not used infurther analysis.
LITHOSTRATIGRAPHY
The lithostratigraphic classification adopted (Fig. 3) is basedon Gair (1959) and Field & Browne (1986). The followingunits are recognised in upward sequence.
Craigmore Limestone Member, Otekaike LimestoneCraigmore Limestone is exposed beside Gordons ValleyRoad (Fig. 1), where it consists of 16 m of well-bedded, palecream-grey, slightly glauconitic limestone. The lower 4 mis concretionary and nodular and is overlain by 12 m of asofter, well-bedded unit with a hard band near the top.Although the position of this sequence relative to the>80 ft (>25 m) of Craigmore Limestone recorded by Gair(1959) is unknown, the bedding suggests that it correlateswith the middle to upper parts. A basal greensand was notobserved.
Waitoura Marl Member, Otekaike LimestonePale brown-grey, soft, marly mudstone is poorly exposed inweathered hillside exposures (old cliffs) to the west of theBluecliffs section (Fig. 1, J39/fl21, 122). This unit liesbetween Craigmore Limestone and Bluecliffs Silt. Itsidentification as Waitoura Marl is tentative and cores areneeded to establish its content and relationships.

Table 2 Benthic foraminiferal census chart, Bluecliffs Silt. Stratigraphic position is measured relative to the base of the "stream section". Species abundances aregiven as raw numbers found in the split used for the census. Also enumerated for each sample are the diversity of benthic foraminiferal species, number of ostracods,weights of the census split, and the >75 ^m residue from which it was obtained. Some taxa are aggregated in the table and the listed species diversity may exceedthe number of taxa listed. Observed occurrences of Ehrenbergina marwicki and Spiroloculina novozealandica, not recorded in the census, are plotted as x.
oooo
Benthic foraminiferal distribution chart; Bluecliffs
"" sisFossil record number J39/
Stratigraphic position (m) _ j
New Zealand Stage
Alabamina tenuimarginataAmphicoryne hirsutaAmphicoryne_scalarisAmphistegina sp.Anomaiina aoteaAnomalinoides fasciotus
Anomalinoides macraglabraAnomalinoides orbicukjsAnomalinoides parvumbilioAnomalinoides sphericoArenodosaria antipoda _Astrononion parkiAstrononion pusillumAstrononion sp.AAstrononion spp.Baggina sp. _BoHvina lapsusBoAVina mahoenuicaBoiivina reticulatoBoBvina_ spp.Buiimina papulaBulp)inelia spicataCassiduSna cannata^ _CassiduSna c_urx}ata_CassiduSna laevigataCeratobuiimina sp.
n • n I o r j ' p ) j i o i w | ! j i j |
3 3 3 i 3 3 3 3 i 3 3 3 3 3 3 3 3 3 : 3 i l 3 ; 3 • 3 i 3~j 3 3 i 3 3 : 3
6 9 ; 10 ! 1 t 3 j 6 [ 1 • 3 j_2j_4_!_4 I 3 5_l_3_j_5 I 4 : 5_ i_4 _ 8 1 5 : 2 4 _5 | ' 2 2 2 | 4 [ 3 | 5 6 4 5 j 1 j 4
11 5 | 3 i 8 1 : 5 , 4 ] S | S I .4. ' 6 2 I 3 I 11 ! 10 | 11 , 7 j 9_ 9 I 13 | 5 | 10 | 15 13| I ? 1 7 j 131 7 | 22 19 , 6
3 4 I 9~1 2 I 4 4 1 2 1 3 I 6 : 4 " 4 ' 6 ^ 6 1 8 1 9 7 7 I 3 I R 4 I 5 T 2 T ~ 4 ' 1 0 : [ i l l T T T61 6 • 4 ! 3 I 4__4_M_ 5 _8 211 2 : 4 I 4 4 | I I
4 j s I J_2J J J | 3 | l2 |10! 1
Cibicidesi medJocris
Gbicides motestus _Cibicides novozelandicusCibicides perforatusCitHcides spp^Cibicides cf- pseudoungerianusCibicides vortexCribrorotalia dorreeni
1 j 20t ' 3 | 2_5
! 12[13113_ 7 ! 5_h? 11J. ! "l " ~T~3^1 j 2
; T'Z. -UUTT
j-5-!_4.r-nii.! 21 | 8 I 3 ! S_ 10J20
i 3": i !"i
7 19_^9
—u3
"7 1 s i 122'
: 13 I 19^171 24 j 21 | 6 I lT~6 j 23 25 15:6 I' 8"|"l6' 25J"i3 "iot"B I 12 '24: 18 | 19j"i2"f"i6j 12"t"l9'T 8 ' 171~9 ]~6 j 10[ 14j~9~t~6>"j '7 , i s t ^ p "i ^ . ^ L | H | t i I 1 I _ | 1 ' 3 2 '1 1 j 1 j 1 2 _^2 [ 1__"1 111 ! 1 " I 1 I " I j 1 [ _ | 2 j ~ _ i J_ [ J_ ~ j " . j
_ L L I T ' ? * " - 3 - - L . • " J - J L T 2 ^ 1 " ' '"'""I I T '"""psi i l i _ i _i j" [' ' I r2_T i-_X_j " ~ I J _ L T L L T - J ' 1 ' 1 " • " '
|"2TJl2|"a'"3T7 | 10i"l6"pl6J"6" 13 j"3~22"| laTTpp"" 13 j 11 j 21~j 7~2:1 2 | 2 j j 2 | 2 _ | !_^6_J__4_4 ^ |
23; 151 9 | 19 I 17J~9~! 7 '. 21 t 29 I 4 | 4 i 19"| 6 ; 12~| 16Tl4i 17 ' 121 8~T~9 ~i 10 [ 15 22 | 3 ] "l 2 j I B . 3T~
Et
~ 4 T 4 j T ! ~JX~V\ 7~t"2T4T '• "iT"i"t~TTT| "" ,
Eponides broeckhianusEuuvigerina mioschwageriEuuv^erinarniozeaEyolvocassickjlinasp.
Fissurina spp.
GlobocassiduSna subglobosaGuttuSna sppL
Gyroidina prominula
Gyroidinqidesze[andca_
Haeusierello hectori
Karreria maona _Lagena spp.Latjcarinha a/tocamerata
\2\-}- S|4?-TlJTdT? '~sT8 I 10~9
7]4^Tytirft4J f t '. iT__ 13 j s l 121 ; ; i_L I -_2
J i i I
'•A~<2\ r ] ""T1 I [ 2 1 ' p " j « ? I 3 j 3~l~iT3'i To"! 5 i "" 9 \ ' '"6 • i 6 [ ' 4 i 9 I 5 1 5 " r ^ T ' '*ioTT[T!~s"[Tl"i4 ' '"6 | '3Ti6| I [ , " p '"" I I ' ' "Ti | Z ± + l l I L f Jn';io/p]TP5jTj , p I i--4H
(20 22Tl«t l^7 ' 1 8 i 1 6 j 9 ' 6 fa.' 2\il Yj 23 : 10! 17~! u\ 17 j t: 7 l~6 J 1? ; 1 Tt~8;\ 12 I "7 PJ7 3~tT7T"27 Tl6 * l s i j b T i 6T5 6 1~9 j l O J T f i j 5~T 11 15 j 9 j 14~| 7 I 1
V lT" : i 4 I 4 I 1 1
P2 : 4 ! 2 2 \ T 6 l i l . T_\ I 2 i T t
NS3
o
IQ
B
a.O
i
o
(continued)

Table 2 (continued)
Laficannina pauperotoLenticuiina cultraiusLenticuiina dicampykjs _LenticuiinagibbaLenticuiina toculqsusLenticuiina mamHligeraLenticuiina spp^ __Morginulina subbullataMartinqttieUa communisMetonis cassidulinoidesMejqnis maoricaMeionisjpp.Nodqsaria iongiscata_NodosorigpyrutaNodosaha seipityoosq^Nodosariojp.Nqnione/ia^Notorotalio serrata
Notorotalia sp.Notorqtalia spinosa_Nqtorotaiia sp.C
2 j 1 3 i I ! r |[<> :""'i] 4 I 7 1 3 7 V 7 1121 7 t 6 5 9 j 6 j 4 |
7 5 j 4 : 2
j ' - - ' ' • • ' -i-ij i10 2 i l o j 5 : 4 i 1 i 6 i 4 T l l | 6 1 4 i 3 I 1
5 | 9 i 2 ^ 8 l r + 7 f *7" 7 + - 2 h t 2 |T
J- : -r f"- - [ ! - f - - " I — i I
* 2 i 4 i 2 ! l [ 5 ' 4 J
' 8 3 }_4_j 3 j 3 . 2 ! 7 7 ~3 ' [~6_i 2 '^2 ] 2 | 1 ]~2 j 6~T41 )~[ " I
i1" 1 " r ~* f"'T2i r r r i y f i T TV-t—
1 1 1
|J4 -..|_L'{- I - - i - : ,i'f:
_l 2 ] 1 ' 7 * 7 ' 1 J2 j 5 Iti j 10: 6 j 5 J ! 2 ; 2 j 4 ^ j 1 . ?_|.?.. 3
t '..I ' , I ?_ 2 ] J . 4
1 '. •> , ' _ * '_ [ 6 ! ! 1 2 r ! 1 5'. 2 3 --Li 3 i ' - 1 ' * LL' 1 2 ! ' ' j 3 i !-2-j I-3 I 'X? '. 71 -!!. 5_ I 3 _ 1 . 5 ' 6 I 24 1 0 J _ i 2 1 3 j - . "-
3 ' sT 4 p6 ] )5 ? T l ] l l ' 7
6 ! 1 } 1 : _ I ' „ i ' J '
1 ] ! 1
I J 4 h 2 JJ2!
i , ^ . . M -
^-a. -H
' 41
_L_2 1 7 _ _ j J_24 3 X j j 6 , _ j __5 j 5 9 _ 2_| 4 ; 6_^4 ; 4 j
* 2 :! 8 8_; 2 j Y i 10 _ 3 ^ 3 ] " l 3 ; 2 _3 [ 1 i J_ 2 j 5_ 2 j 6__ 1 j 1 _! 121 9 j_l 5 j 3 ; 5J 5 j 3_^ 9 f 5_ 8 [ 4_j 20
1 16 \l_ ' i 5 i 5 i_" ! ' 2 . 7 i 8 i _ J 4 j 5__3 j ._ i • ._!_3.22LlK 5 ; _6. J3. _
] 2 2 1 I 1 ; 1 ! 6 j 4 | I I ; 2 ! 2 | l i 1 1 [ 2 : 5 l 2 : 1
l _ 4 ! 7 |_5 : 5 j _ 6 6_j 4 • j _4 2j_^ I Li
Orthomorphina cf^cha/iengeriono^ t " 1
-_- . _ l .
Piectofrondicularia sp.Polymorphina sp.Pseudonodosaria aperto
1
I l l 1 3 j
! :
"
6 _7__ 5 J j / 5j »J 3 ;7 * 4 ^ 3 ' ~ ' ^ l '
: 4 J_5 2 _7 ] 3^_[ ' 2 j 5 i_ j 7 '_9_16 «_! 10| 5 j__4_J_14j_2 J 17 2
• ~ r! 1
Pseudonodosario symmetricoPuffenia bulloidesPuSenio quinqueloba
PuSenia sp. _L , ' ' I _ i _ ; _ 'Quinquefocu/ina spp. ! ~ L . 4 i__U 2 I — L 2 JJ-! •Romulina sp. __ ! _ |__^_ j • __L _.__,Rqsaltna s p ^ _ J _ _j | _| |_ _ j _ j _ _ j [_
Soracenorio sp, ^ | . ( _i j ' j__ jSigmoilina tenuis J j i J j _ ' t :
Siphoning australis j _ _ 1 1 J j v j ___ , [Siphotextularia gwomoana i _ 1 I ^ ] _^ 1 -Siphotextulgria emaciata _i ^ __• : i_ ' • j ' ^ _
Siphotextularia sp, |__^ l_j_"~: __| ' J[j | __ iSiphouvigerina probosciaea • 1 : !___! i ! _ : 2 _. j 1 'Siphouvigerina sp. _ _ , j _ , j i !__^ _ |_ ! _ ; _ j _, 1 \ __H__i ;Sphoeroidina bu/loides _ ^A_ " '__j 2 "u_J___4 ! 5_L10 _^_ j 7__ j 3 ^ 2X| 2 ' i 4 i _ , 3
Spiroloculino novozeolandico •, _ ^ •_ i i _ ' _^ i ! X_ '•• [
-,- f . . ._| . - . —|—
—*~ [ — - - • j —p • -
! 1 ~"~2 i 1 r
ix^Y't
i TLJTf" "~i
l " • " L i ^ i Z " 1 ^ i7P_LT__^
^ T 2 ' | 3 ~2"j 3~T
Stilostomello pqmuligeraStilostomella sp. _ _Textuloria miozea _Textulorio sp.Trifarino bradyi _Trifama esutiens_Mama spp.Vaginulinq elegansyaginulinq_sppL
Vaginulina vagina _Vaginulinopsis sp. _ _ _ _ j ; x _specimens indet. ' ' "
11
U 4_4 1
D-r I - J - M l i J , JI 1 i ;
3 [
I i_4 I ! 2 1 1 i l_l
"2r "* T '"1 rr i r1 IJ_
;_- iijx[..;xx']T; +• 1
1 1 J _ 2_
if 'i
^^y^^ti 1 i d u II i^f^ 1 • • I u I ' i J I t ^ 1 O 1 / l _ l i _ ' \ ~ Oi "* O ; I __l_ I __|_ ' ^ *J 1 O V ! *J ] ' 1 1 ' *. 1 *j ' ' 1 1 1 ! <
Bentnic dlveralty (species/somple ^43 j 41~29 4 8 * 4 5 ; 39 "l 42 j j i 3 I 36^35 j 40^ 47 |36 37 43^42 ' x \ 44 J36|47 j 30 : 57 * 42T36T38 j 30 j 26~34 ! 44 j3S '^u\ 36 28_32 j_44j_35 i j 5 j 45*47 ; 39 30*36 J 36 29T34J 3 4 j J 8 I 35T29I26J 3 7 [ 3 T
Bentnic specimens per sample
Ostrocoda per sample
Bulk weight all samples = lOOOg.Residue weight (grams) =
Weight of picked split (grams)
29124 26 5 4 , I 2 8 ' 2 8 | 1 4 [ l 7 1 6 J 2 9 ! 3 6 ; 3 6 ' 1 2 17 24141 20 I 40 | 56 2 2 ^ j 36 ' 13 j 34 26 ' 72 44 j 54 24 ' 461 641 601 96, 50 | 59j 88 | S ! 58 I 68 , 551 36 ' 58M8 I 48 j 46,' 4 o l l 8 , 22 | 36: 3 VsV^t, \ 10 ' 48 | 40 j 50
' § s l s \ 2 \ i s i s
I!" 5 S ^ f O DO <OP P ! o 1 p
; d 0 | d j d | d | o j o , o o ; d I o ' d I o 1 o d | d •!O p I O I p ]O i p p | O ! p ! o j p : p | p j P p'1— I •— I1— ! p | - \ •— O p ' •— I p •— i p Q I p I 6 |<N0 o j d d | d d d l c J J d i O d | o ' d 10 | d d d ! Q : d | o | o | o | o o ' d ' o | d | d | d ; d I d
oo3
a>
IP
6
or)
&
O"
o
a.

590 New Zealand Journal of Geology and Geophysics, 1999, Vol. 4
Table 3 Nannofossil distribution chart, Craigmore Limestone, Waitoura Marl, and Bluecliff Silt. Stratigraphic position is measure 1relative to the base of the "stream section". Data categories are shown in bold and codes used for each category are shown in tli.:footnotes. In category 6 abundance data are qualitative estimates of numbers observed during examination of smear slides. Numbe;;1 -4 are actual specimens observed.
Sample #(J39/f...)Stratigraphic unitStratigraphic position (metres)Overall abundance 'Overall preservation 2
Overall nannosize 3
Other microfossils 4
Lithofacies (on microslide) 5
Taxa: '•6 (see caption)Braarudosphaera bigelowiCalcidiscus leptoporusChiasmolithus altusChiasmolithus oamaruensisCoccolithus miopelagicusCoccolithus ovalisCyclicargolithus floridanusCystodytes sp. indeterminateDiscoaster deflandreiHelicosphaera carteriHelicosphaera euphratisHelicosphaera obliquaHolodiscolithus macropontsMicrascidites vulgarisPonlosphaera multipart*Pontosphaera sp. (punctate)Prinsiaceae indet. (small)Reticulofenestra bisectaReticulofenestra "clifdenensis " MSReticulofenestra gartneriiReticulofenestra placomorphaSphenolithus moriformisSyracosphaera clathrata
114
<fll5SMS
LM
2TS
T
115
0.5SPS
LM
3S
116 117 118Craigmore Limestone
5.2TM
LM
3T
8TM
LM
TT
1*
11TM
LM
TT
1
119
15.5TM
LM
2*3
120
15.9SMS
LM
TSS
1
3
>
121 122Waitourafl20>fl21CMS
M
TC
s
3S
T2
F1*
3
AMS
Z
scR
1*4S
41
A3*1
1
123
0RMS
LM
T
SS
S
1
R2
124
1.9RMS
LM
TRS
3
1
R
1
125 126 127 128Bluecliffs Silt (stream
3.5RMS
LM
TRS
1T
T3
R
21?*
5.5FMS
LM
1
TFR
3*3T
T21F2*
2
3
7.5CMS
z
1
scR
T*2T
T
C
2
8.6AMTbM
S
cs4*2S
c
3
T
129 130section)9.4CMSfM
SFT
1.5*11
T
C2
1
11.1FMS
M
TFS
4T
S1
R
1
2
132
14.4CMS
LM
S
cR
3T1
T
F
1
135
If.CMSf
LN/
R
cs123
T1
F2
3
1
Table 3 {continued)
Sample #(J39/f...)Stratigraphic unitStratigraphic position (metres)Overall abundance 'Overall preservation 2
Overall nannosize 3
Other microfossilsLithofacies (on microslide) 5
Taxa: '•* (see caption)Braarudosphaera bigelowiCalcidiscus leptoporusChiasmolithus altusChiasmolithus oamaruensisCoccolithus miopelagicusCoccolithus ovalisCyclicargolithus floridanusCystodytes sp. indeterminateDiscoaster deflandreiHelicosphaera carteriHelicosphaera euphratisHelicosphaera obliquaHolodiscolithus macropontsMicrascidites vulgarisPonlosphaera multiporaPontosphaera sp. (punctate)Prinsiaceae indet. (small)Reticulofenestra bisectaReticulofenestra "clifdenensis " MSReticulofenestra gartneriiReticulofenestra placomorphaSphenolithus moriformisSvracosphaera clathrata
136 137 140 145 146Bluecliffs Silt (stream section)24.9
FM
LM
TFS
2
1
R
1
28CM
LM
1SFS
14
31
C2*
30.7CM
fLM
SFS
T*31
S3
C
3
33.5CM
M
4
FCS
22
T
R1
34.1FM
bM
TFS
T
F
147
35CMS
M
1
SCR
TTT1
S
c
12
T
149
37AMM
LM
S
cT
1*1T
T
A2*
2
151
39CPS
LM
ScR
3T
31
F
1
1
153 157 160 162 164Bluecliffs Silt (cliff section
41FMS
M
TRS
24
T2
F1*
2
T
44.8FMS
LM
TR3
1T
32
F
1
1
47.7CMSfM
TFS
1
2
T
C
2
50.2APMfZ
TFT
2?
T2
A
2
52.2CMM
M
SCs
14T
S4
F3*
T
3
166 167 169Waitakian part)54.4AMSf
M
1
FT1T24
TT
A1?
T
T
55.7CMMf,sLM
TFR
T*2T
S
c
T
S
58.1FMMfM
RS
2*2
T
F
1
170
59CPs
LM
FS
1
1
C
2
174
62.9CPSf
LM
FS
23
S1
c1
1
1
i:
65(1\N
L
1
1
<:i
(continued)

Morgans et al.—Waitakian-Otaian Stage boundary 591
Table 3 (continued)
Sample # (J39/f...)Stratigraphic unit
178 180Bluecliffs
182Silt
(cliff section;Waitakian part)Stratigraphic position (metres)Overall abundance 'Overall preservation 2
Overall nannosize 3
Other microfossilsLithofacies (on microslide) 5
Taxa: ''6 (see caption)Braarudosphaera bigelowiCalcidiscus leptoporusChiasmolithus altusChiasmolithus oamaruensisCoccolilhus miopelagicusCoccolithus ovalisCyclicargolithus floridanusCystodytes sp. indeterminateDiscoaster deflandreiHelicosphaera carteriHelicosphaera euphratisHelicosphaera obliquaHolodiscolithus macroporusMicrascidites vulgarisPontosphaera multiporaPontosphaera sp. (punctate)Prinsiaceae indet. (small)Reticulofenestra bisectaReticulofenestra "clifdenensis " MSReticulofenestra gartneriiReticulofenestra placomorphaSphenolithus moriformisSyracosphaera clathrata
67TPS
LM
T2
1
T
68.9FMS
LM
RT
11T
TT
F2
121
70.6RPS
LMI
1RT*
121
T2
S
3
184
72.8CMS
LZ
11RR
TS2A1
1
T1
186
75.3FPS
z
RS
T42
1
F
1
2
188
77.9CMSf
z
1FS
23
T
C
4
1
190
79.4CPS
M
C
s141
C
194
82.5CP
s
LM
FT
1*1
13
F
T
1
198 200 202Bluecliffs Silt (main
85.8RPS
LM
1
1R1
2
T1
R
2
87.8FMS
LM
2F4
2T
T2
R
4
89.8FPS
LM
1F1132
2
31
F2?11
1
206 208 210 213section; Otaian part)
92.9FPS
LZ
4R3
34
TT
R1
1
95RPS
LM
4
ST
1*T1
TT
R1
211
97.3FPS
LZ
F1
3*31
3T1
F1
1
101RMS
LM
5S3
4T1
1T
R3
2*
1
214
103RMS
M
TS3
3T
2T
R
215
108CP
sf
LZ
TFT
1R
ST
C
216
115FPS
z
FT
3*1
3
TR1
2
1
217
125FPS
z
RT*
1*31
ST
F
3*
'A = abundant (>5000), C = common (2500-5000), F = few (1000-2500), R = rare (500-1000), S = sparse (100-500), T = trace(5-100). 2M = moderate, P = poor. 3T = tiny, S = small, M = medium. 4b = bryozoan, f = foraminifera, s = calcareous spongespicules. 5L = lime, M = mud, Z = silt. 6* = battered, bold = complete.
Bluecliffs Silt
Gair (1959) originally described Bluecliffs Silt as ".. .a bluesandy silt ('Blue clay' of Gudex) in which hard sandstonelayers 5 ft to 6 ft apart and about 1 ft thick give a veryregularly banded appearance. The silt is fossiliferousthroughout, but more so in the middle and upper portions,the macrofossils being chiefly Mollusca with a fewbrachiopods near the base. Pyritic concretions occurirregularly throughout, and gypsum in small films is widelydistributed." This description refers to the cliff section(Fig. 1) which contains the boundary stratotype. AlthoughField & Browne (1986) synonymised Bluecliffs Silt withTokama Siltstone, we consider the latter to be restricted toNorth Canterbury (Andrews et al. 1987). It was not depositedcontiguously with Bluecliffs Silt (Field et al. 1989, sheet 4,map 28).
From outcrop data we estimate that there is c. 100 m ofsection between Craigmore Limestone and the base of thecliff section. The upper 35 m (stream section, Fig. 1) ismassive blue-grey siltstone which is exposed in the riverbedand banks. It is in conformable contact with strata at thebase of the cliff section. For descriptive convenience wedistinguish three informal lithofacies in upward successionat Bluecliffs.
1. Lower siltstone lithofacies
The lower siltstone lithofacies (0-38.5 m) is a massive, softto moderately hard, medium blue-grey calcareous siltstonewith no obvious bedding. It is exposed in low banks and inchannels starting in a side stream 400 m to the west of theBluecliffs cliff section, and continues along the north bankof the Otaio River to the base of the latter.
2. Banded lithofaciesThe banded lithofacies (38.5-106 m) is a bedded, moderatelysoft, blue-grey, calcareous siltstone. Bedding varies fromindistinct to well defined and reflects variation in lithi-fication. Cemented beds, which appear to be more calcareousthan the surrounding siltstone, are termed hard bands. Thebase of the lithofacies is defined by the lowest hard band,and the top of the lithofacies is the topmost hard band. Werecognise 28 hard bands (Fig. 2). They vary considerably inprominence; some are most obvious at a distance from theexposure. The banded lithofacies is exposed in the cliffsection for 1 km along the north bank of the Otaio River.
3. Upper siltstone lithofaciesThe upper siltstone lithofacies (base at 106 m) is a massive,soft, blue-grey calcareous siltstone without obvious bedding.

592 New Zealand Journal of Geology and Geophysics, 1999, Vol. 42

Morgans et al.—Waitakian-Otaian Stage boundary 593
It overlies the highest hard band; only the lower part at theeastern end of the cliff exposure was examined in this study.Macrofossils become more abundant higher in the uppersiltstone lithofacies.
Southburn SandBluecliffs Silt grades up into the brown sands of SouthburnSand, which is characterised by fossiliferous sandstone andconcretions.
Sedimentary cyclesCarter (1988) included Bluecliffs Silt in Otakou Group,regarded by Carter et al. (1991) as a progradational highstandsystems tract that developed as sediment supply intoCanterbury Basin increased in the Early Miocene. In theirinterpretation, the condensed interval (not exposed in oursections) underlying Otekaike Limestone marks themaximum flooding surface between the highstand systemstract and the preceding transgressive systems tract in asecond order cycle of thermo-tectonic subsidence at a passivemargin. They did not resolve higher order cycles embracingBluecliffs Silt.
Maximum flooding surfaces separating transgressivesystems tract and highstand systems tract sequence units canbe characterised by microfossil abundance and diversitypeaks (Armentrout 1996; Mancini & Tew 1997) which, givenstable biotic productivity, reflect decreases in sediment fluxat highstands. At Bluecliffs there is a well-defined series offoraminiferal abundance peaks (Fig. 5), between 9 and 73 min the lower siltstone and banded siltstone lithofacies. Spikesin benthic abundance form a waxing series up to 47 m andthen wane to 73 m. The turning point at 47 m coincides withmaximum species diversity (57 taxa). The spikes occur in,
Fig. 4 Benthic foraminifera from Bluecliffs, SEM micrographs.J39/fxxx = locality (see Fig. 2). FPxxxxx = catalogue number ofspecimen in foraminiferal collections at Institute of Geological &Nuclear Sciences, Lower Hutt. md = maximum dimension. A,Ehrenbergina marwicki Finlay (J39/fl 84; FP4566;md = 0.41 mm). B, Spiroloculina novozealandica Cushman &Todd (J39/f215; FP4567; md = 1.02 mm). C, Trifarina bradyiCushman (J39/f134; FP4568; md = 0.34 mm). D, Gyroidinoideszelandica (Finlay) (J39/fl64; FP45469; md = 0.50 mm). E,Karreriella cf. chilostoma (Reuss) (J39/fl27; FP4570;md= 1.35 mm). F, Vulvulina pennatula (Batsch) (J39/H46;FP4571; md = 0.79 mm). G, Martinottiella communis(d'Orbigny) (J39/f7597; FP4572; md = 1.63 mm). H, Lenticulinasp. (J39/H35; FP4573; md = 0.75 mm). I, Cibicides cf.pseudoungerianus (Cushman) (J39/fl34; FP4574;md = 0.40 mm). J, Euuvigerina miozea (Finlay) (J39/f213;FP4575; md = 0.76 mm). K, Euuvigerina maynei (Chapman)(J39/H35; FP4576; md = 0.81 mm). L, Notorotalia spinosa(Chapman) (J39/T164; FP4577; md = 0.47 mm). M, Cibicidesnovozelandicus (Karrer) (J39/f 170; FP4578; md = 1.05 mm). N,Anomalinoides sphericus (Finlay) (J39/fl85; FP4579;md = 0.41 mm). O, Haeuslerella hectori Finlay (J39/f7603;FP4580; md = 1.56 mm). P, Q, Anomalinoides parvumbilia(Finlay). P (J39/f209; FP4581; md = 45 mm). Q (J39/O09;FP4582; md = 0.44 mm). R, Notorotalia serrata Finlay (J39/f206; FP4583; md = 0.53 mm). S, Cassidulina carinata Silvestri(J39/fl85; FP4584; md = 0.29 mm). T, Globocassidulinasubglobosa (Brady) (J39/H29; FP4585; md = 0.28 mm).
or near, minima in the coarse fraction signature. Spikes inthe planktonic abundance signature coincide with benthicspikes at 9 m and 30 m but are slightly offset at 34-36 mand 70-73 m. Overall, spikes in planktonic abundancedecline up-sequence from the major excursion at 9 m.
The generally inverse relation between the foraminiferalspecimen abundance and coarse fraction signatures isconsistent with a cyclic record of highstands which producedcondensed intervals due to decreases in sediment flux. Mostof the abundance peaks (e.g., 9,47,56,64 m) occur close tominima in the relative abundance of spinose planktonic taxa(Fig. 5). The latter signature is possibly a climate proxy inwhich maxima occur in cool episodes, while minimarepresent warm near-surface water (Li & McGowran 1994).These data support the interpretation of the benthicabundance spikes as marking highstands, possiblyrepresenting higher order maximum flooding surfaces. The18O signatures for two benthic foraminiferal species (Fig. 5)show inconsistent relationships with the sedimentary andfaunal signatures, except that excursions are largest withinthe interval containing the abundance spikes.
Significantly, the abundance spikes are not well definedin the exposure, and there is little evidence that the inferredcondensed intervals affect the completeness of the stratotypesection. They are not associated with prominent beddingplanes or concentrations of glauconite. The connectionbetween the spikes and cemented beds in the bandedlithofacies is not resolved by our data because sampling isrestricted to the less cemented interbeds. Marked changesin the composition of benthic foraminiferal assemblages atthe spikes is not observed. The principal macroscopicevidence suggesting slow sedimentation in the vicinity ofthe major benthic spike at 47 m is a significant increase inthe number of pyritised burrows. These considerationsindicate that, while the spikes probably represent condensedintervals, they may not significantly affect the completenessof the section. Particularly, the condensed interval at 73 m(boundary horizon) is minor as judged by the coarse fractionand benthic abundance signatures.
DEPOSITIONAL SETTING
PaleogeographyThe interval studied covers a period in which the earliestMiocene carbonate facies changed to predominantly clasticsedimentation, derived from uplift initiated by movementon the Alpine Fault (Kamp 1986). A lower Otaianreconstruction of Canterbury (Field et al. 1989, sheet 4, map28) showed Bluecliffs at shelf depths in a shallow basinbordered by land to the north and west, and by the submarineEndeavour High to the east. Fine clastic sediments in thevicinity of Bluecliffs lay seaward of a sand wedge progradingout from the western landmass. Peter King's (pers comm.1997) reconstruction (Fig. 6) suggests that the Bluecliffs sitewas at upper bathyal depths below the Waitakian-Otaianboundary. The relative abundance of planktonic fora-minifera, discussed later, indicates that, near the stageboundary, the basin was not closed to the extent shown byField etal. (1989).
PaleodepthKarreriella cf. chilostoma (Fig. 4E) and Vulvulina pennatula(Fig. 4F) occur in the lower siltstone lithofacies but

« - 10000 -
o £ 8000 -
m O 2000
1500 - -
Cibicides novozelandicus
Fig. 5 Sedimentary, foraminiferal, and isotope signatures for a section across the Waitakian-Otaian boundary at Bluecliffs. wt3/wtl: wt3 is the dry weight of >75 iim fractionweight of the bulk sample. Total benthic specimens/g, total planktic specimens/g: = the number of (benthic or planktonic) foraminifera in a weighed split of >150 |im fraction,
,r .*"
; wtl is the dryrecalculated toprc rpnnrtfri as
zNCD
faa.o
1
oera
Oo
a percentage of total planktonic specimens in the >150 |im fraction. Carbon and oxygen isotopes are reported as per mille deviations from the PDB standard.

Morgans et al.—Waitakian-Otaian Stage boundary 595
Fig. 6 Paleogeography andpaleobathymetry of the NewZealand region at 22 Ma (after P.R. King pers. comm. July 1997).Position of Subtropical Conver-gence after Edwards (1975).
Paleobathymetry
Emergent land mass
Shelf
Upper bathyal
Mid-lower bathyal
populations are very small and they do not appear in thecensus (Table 2). Both are restricted to slope and deep-sea environments (Hayward 1986). Crundwell et al.(1994) found that Vulvulina pennatula occurred below600 m in a Taranaki Basin Pliocene sequence. The rarityof these taxa, with the absence of other typical slope taxa(e.g.,Gyroidinoides neosoldanii, Planulina catilla,Sigmoilopsis schlumbergeri), suggests that the lowersiltstone lithofacies was deposited in the upper bathyalzone (200-600 m).
Apart from the absence of deeper water taxa, thetaxonomic composition of assemblages in the banded andupper siltstone lithofacies resembles that of the lowersiltstone lithofacies. Dominant or common taxa (Table 2)include Anomalinoides spp. (Fig. 4N, P, Q), Cassidulina spp.,and Globocassidulina subglobosa (Fig. 4S, T), Cibicidesspp. (Fig. 41, M), Gyroidinoides spp. (Fig. 4D), Haeuslerellahectori (Fig. 40), Lenticulina spp. (Fig. 4H), Martinottiellacommunis (Fig. 4G), Notorotalia spp. (Fig. 4L, R),Sphaemidina bulloides, and Trifarina spp. (Fig. 4C). Manyspecies persist throughout the interval sampled. Distinctiveshallow shelf and shoal species (e.g., Amphistegina sp.,Cribrorotalia ornatissima, Elphidium crispum, E. kanoum)are almost absent, although they are major constituents of aWaitakian assemblage (I38/f7607) from the prograding sandwedge to the west near Fairlie. Of these taxa, only two
specimens of Amphistegina are present in our collections.These data suggest that the banded lithofacies and lowerpart of the upper siltstone lithofacies were deposited at outershelf to uppermost bathyal depths (c. 150-300 m). Thesimilarity in taxonomic composition of assemblages isevidence that changes in depth during deposition of the threelithofacies were relatively small.
Planktonic foraminifera live at preferred positions in thewater column during growth (Be 1977) and, insofar as theoxygen and carbon isotope compositions of their shellsrecord environmental parameters (Hemleben et al. 1989),fossil taxa provide some information about relativepaleodepth. Of relevance here is the persistent presence inBluecliffs assemblages of small but typical populations ofGloboquadrina dehiscens (Fig. 7D). In stable isotopestudies of Miocene material (Corfield & Cartlidge 1991;Gasperi & Kennett 1992) this species ranks as one of thedeepest dwelling planktonic taxa. Although this relativeranking cannot be converted reliably to absolute depth inthe water column, the data show that Globoquadrinadehiscens calcified significantly below the 0-50 m photiczone and that the Bluecliffs sea probably exceeded shelfdepths.
Molluscs are rare in the lower siltstone and bandedlithofacies but are common and diverse in the upper siltstone.Assemblages include outer shelf and upper bathyal taxa

IocB9
ono
oCD
i
o
<

Morgans et al.—Waitakian-Otaian Stage boundary 597
(Alan Beu pers. comm. 1997). Nannofossil abundance anddiversity begins to decrease in the banded lithofacies(c. 57 m) and indicate the onset of gradual shoaling.
In overview, the lower siltstone lithofacies is identifiedas the deepest unit, estimated to have been deposited at upperbathyal depths (c. 200-600 m). Reduction in paleodepth isfirst detected within the banded lithofacies and continuedinto the upper siltstone. Whether the site had moved up toshelf depths at the top of the interval under study is uncertain.We consider that upper strata of the banded lithofacies andlower strata of the upper siltstone lithofacies were depositedat depths of 150-300 m in a regressive regime. These dataare uniformitarian estimates of paleodepth that provide poorresolution and are not well constrained. Cyclic patterns inspecimen abundance and sedimentation data discussedelsewhere suggest that eustatic oscillations in depth weresuperimposed on the long-term decline in depth.
PaleoclimateThe composition of nannofossil and planktonic foraminiferalassemblages indicate a cool-temperate water mass. Anindication is the almost total absence of Globigerinoid.es,whose Early Miocene record in southeastern Australia isrelated to southward incursions of warm water (Chaproniere1992). Oscillatory marine climate is indicated by the relativeabundance signature of spinose globigerinids (Fig. 5), whichChaproniere (1992) took as a proxy for cool near-surfacewater. McGowran & Li (1993, 1996) and Li & McGowran(1994) noted the association of spinose taxa related toGlobigerina bulloides with cool, nutrient-rich water (Weaveret al. 1997) and linked their abundance signature withnorthward fluctuations in the location of the SubtropicalConvergence over southeastern Australian neritic sites inthe Early Miocene. This signature, represented byGlobigerina bulloides (Fig. 80, P), Globigerinapraebulloides (Fig. 8I-N), and variants Globigerina sp. A(Fig. 8Q, R) and Globigerina sp. B (Fig. 8S, T), andGlobigerina ciperoensis (Fig. 8F-H), is cyclic at Bluecliffswith strong spikes between c. 35 and 70 m. The largest is at70 m and immediately precedes the Waitakian-Otaianboundary.
Constraints on the interpretation of oxygen isotope datafor two benthic foraminifera (Fig. 5) are the wide, variablyspaced sample scheme, which may not reliably resolve
Fig. 7 Planktonic foraminifera from Bluecliffs, SEM micro-graphs. J39/fxxx = locality (see Fig. 2). FPxxxxx = cataloguenumber of specimen in foraminiferal collections at Institute ofGeological & Nuclear Sciences, Lower Hutt. md = maximumdimension. A-C, Globigerina woodi woodi Jenkins; J39/fl85.A (FP4520; md = 0.40 mm). B (FP4521; md = 0.36 mm).C (FP4522; md = 0.34 mm). D, Globoquadrina dehiscens(Chapman, Parr & Collins) (J39/fl85; FP4523; md = 0.39 mm).E-G, Globigerina woodi connecta Jenkins; J39/f 185. E (FP4524;md = 27 mm). F (FP4525; md = 0.34 mm). G (FP4526;md = 0.34 mm). H, Globorotaloides suteri Bolli (J39/H34;FP4527; md = 0.32 mm). I-M, Globigerina brazieri Jenkins.! (J39/T134; FP4528; md = 0.35 mm). J (J39/f7602; FP4529;md = 0.33 mm). K (J39/f7602; FP4530; md = 0.35 mm).L (J39/f7602; FP453 f; md = 0.37 mm). M (J39/f7602; FP4532;md = 0.33 mm). N-R, Globigerina euapertura Jenkins;J39/f7601 N (FP4533; md = 0.26 mm). O (FP4534;md = 0.26 mm). P (FP4535; md = 0.28 mm). Q (FP4536;md = 0.28 mm). R (FP4537; md = 0.37 mm).
periodic signals, and the weak, and sometimes negative,covariance between the signatures. However, the data areparticularly variable between 25 and 63 m. The amplitudeof the heavy excursion in ' 80%o between 34 and 43 m inEuuvigerina spp. is large (2.08%o) and greatly exceedsamplitudes recorded by Zachos et al. (1997) for the basalMiocene Mi la glacial maximum at an equatorial site. Theyconsidered that the increase might equate with a cooling of6°C. Although the excursion between 34 and 43 m may notrepresent part of a single cycle, and may indicate change inice volume as well as temperature, the data flag thelikelihood of significant instability in climate before theWaitakian-Otaian Stage boundary at 73 m. While highamplitude excursions in 180%o all occur within the 9-73 msection containing spikes in specimen abundance, the dataoften do not covary closely with the spinose globigerinidclimatic proxy. If both contain climatic information, thissuggests that near-surface and bottom water regimes differedsignificantly.
Whereas the marine signatures likely identify significantclimatic change before the Waitakian-Otaian boundary,Pocknall (1982) did not observe comparable evidence in landfloras. His data suggest that terrestrial climate was warm-temperate, with moderately high rainfall evenly distributedthrough the year. Landscapes were fully forested, with brassibeech as the dominant group. Montane taxa are absent(Dallas Mildenhall pers. comm. 1997). We have not resolvedthe discordance between marine and terrestrial climaticreconstructions. Southward in Canterbury Basin, Bueninget al. (1998, fig. 7) analysed brachiopod carbonate anddetermined paleotemperatures of c. 11.2-14.5°C (correctedfor depth) from a Waitakian locality and c. 9.5-ll°C froman Otaian locality.
OceanicityNannofossil assemblages in Bluecliff Silt and underlyingWaitoura Marl have limited diversity (Table 3) and arenumerically dominated by small or minute taxa. They ofteninclude specimens of Coccolithus ovalis (Fig. 9E-H) andCyclicargolithus floridanus (Fig. 9T) that are smaller thannormal. Taxa considered to prefer warmish water coastal,marginal sea, or oceanic conditions are either rare (e.g.,Braarudosphaera bigelowi, Discoaster deflandrei (Fig. 90),Helicosphaera spp. (Fig. 9I-N), Pontosphaera multipora(Fig. 9Q-S), and Sphenolithus moriformis) or absent (e.g.,Discoaster druggi, high-spired Sphenolithus spp., Thora-cosphaera spp., and Triquetrorhabdulus carinatus).Coronocylus nitescens, a ubiquitous oceanic species, is alsoabsent. These rather featureless nannofloras imply depositionfrom a temperate, vertically well mixed, eutrophic watermass at a shallow offshore location, intermediate betweenneritic and fully oceanic.
Relative abundance of planktonic foraminifera (Fig. 5)varies between 45 and 87% of total specimens through thesection but drops to 22% in the highest sample. The patternis cyclical, possibly at several frequencies. Generally,planktonic species are open-ocean organisms and are scarcein low salinity, turbid coastal water (Hemleben et al. 1989),although the gross pattern is commonly perturbed by variousphysical and biotic factors. Gibson (1989) examined a largeset of data from off northeastern United States and reportedrelative abundances comparable with Bluecliffs in thevicinity of the shelf break at depths between c. 90 and 500 m

598New Zealand Journal of Geology and Geophysics, 1999, Vol. 42

Morgans et al.—Waitakian-Otaian Stage boundary 599
in a zone separating neritic from oceanic water. In this region,the shelf break is usually >100 km from the shoreline.Although it is unlikely that Bluecliffs paleogeography wassimilar to that sampled by Gibson (1989), the comparisonindicates that Bluecliffs was in an intermediate positionbetween inshore neritic and fully oceanic water. Theoscillatory pattern may have a climatic component.Generally, the planktonic representation is greatest whenspinose taxa, possibly proxies for cool nutrient-rich water,are most abundant. This may be related to the position ofthe Subtropical Convergence, an important water massboundary in the region in the Early Miocene (Edwards 1975;McGowran & Li 1996; Weaver et al. 1997).
Field et al. (1989, sheet 4, map 28) envisioned that theBluecliffs basin was closed to the north and west. However,only at the top of our section, c. 50 m above the boundarybetween Waitakian and Otaian Stages, is there evidence ofa neritic water mass over the site. Nannofossil andforaminiferal data from the lower siltstone, banded, andlower 15 m of the upper siltstone lithofacies suggestincomplete closure to the west. Figure 6 shows anarchipelago scenario with a submarine ridge (EndeavourHigh) and isolated westward landmasses. These wouldpartially deflect oceanic water yet provide insufficient shelterfor neritic water to establish.
Bottom environmentBenthic foraminiferal assemblages are diverse and areprimarily composed of shallow infaunal-epifaunal taxa.Species abundances tend to be equitably distributed.Particularly, abundance spikes of infaunal opportunisttaxa (e.g., Bolivina, Euuvigerina) typical of nutrient-richand/or oxygen-deficient environments (Sen Gupta &Machain-Castillo 1993) are absent. An overview of thedata suggests that the sediment interface was welloxygenated, with moderate nutrient flux. Bottom energywas low. Laminated sediments are absent whereasbioturbation is commonly observed. An active metazoaninfauna is suggested.
Fig. 8 Planktonic foraminifera from Bluecliffs, SEM micro-graphs. J39/fxxx = locality (see Fig. 2). FPxxxxx = cataloguenumber of specimen in foraminiferal collections at Institute ofGeological & Nuclear Sciences, Lower Hutt. md = maximumdimension. A, B, Paragloborotalia nana nana Bolls; J39/T134.A (FP4538; md = 0.27 mm). B (FP4539; md = 0.26 mm).C, D, Paragloborotalia nana semivera Hornibrook; J39/fl34.C (FP4540; md = 0.29 mm). D (FP4541; md = 0.30 mm).E, Paragloborotalia nana pseudocontinuosa Jenkins (J39/T134;FP4542; md = 0.34 mm). F-H, Globigerina ciperoensisciperoensis Bolli; J39/T185. F (FP4543; md = 0.27 mm).G (FP4544; md = 0.26 mm). H (FP4545; md = 0.23 mm).I-N, Globigerina praebulloides Blow. I (J39/H85; FP4546;md = 0.48 mm). J (J39/fl85; FP4547; md = 0.35 mm).K (J39/f 185; FP4548; md = 0.37 mm). L (J39/f 164; FP4549;md = 0.38 mm). M (J39/H64; FP4550; md = 0.39 mm).N (J39/H64; FP4551; md = 0.45 mm). O, P, Globigerinabulloide- d'Orbigny. O (J39/T185; FP4552; md = 0.49 mm).P (J39/fl64; FP4553; md = 0.47 mm). Q, R, Globigerina sp. A;J39/fl85. Q (FP4554; md = 0.26 mm). R (FP4555;md = 0.28 mm). S, T, Globigerina sp. B; J39/fl 85. S (FP4556;md = 0.29 mm). T (FP4557; md = 0.25 mm).
BIOSTRATIGRAPHY
We evaluate the nature, distribution, and utility of selectedbioevents in the vicinity of the Waitakian-Otaian boundary.Benthic foraminiferal taxa cited by Finlay & Marwick (1947)for the boundary are considered, as well as planktonicforaminiferal and nannofossil events that might potentiallyextend the range of environments in which the boundarycan be recognised. Data from several other groups arereviewed briefly.
Benthic foraminiferaEhrenbergina marwicki (Fig. 4A)
RECOGNITION: This species is highly distinctive (Scott1973) and its occurrence is unlikely to be misidentified.Ehrenbergina cf. bicornis (FAD in Waitakian Stage) is tightlycoiled whereas E. marwicki is largely uncoiled (Scott 1973).
DISTRIBUTION: The FAD occurs at 73 m (J39/fl 84, Fig. 5).The event is widely recorded in shelf and upper bathyalsequences in South Island and western North Island(Fig. 10). It is generally absent from deep-water sequencesin Northland, eastern North Island, and Taranaki. Hayward& Buzas (1979, fig. 15,21) recorded the greatest abundanceof Ehrenbergina marwicki in upper bathyal deposits fromnorthern New Zealand. Conversely, a study by Hayward &Brook (1994) of inner-outer shelf Otaian sequences fromWaiheke Island, Auckland, failed to recover any Ehren-bergina marwicki. Examination of collections higher in thesection (Scott 1965) show Ehrenbergina marwicki is absentfrom the upper siltstone lithofacies and Southburn Sand atBluecliffs. Early populations are small. At Bluecliffs, thespecies was not recorded in counts of <200 specimens until5.1m above its first occurrence in strews of several thousandspecimens. Li & McGowran (1994) suggested thatEhrenbergina marwicki was a "deep-cold" species in theneritic Miocene assemblages at Lakes Entrance, Victoria;Li & McGowran (1997) placed it in their "outer neritic"group which inhabited depths of c. 150 m or more. AtBluecliffs, its FAD follows a spike in abundance of spinoseglobigerinids at 70 m, which possibly signals a cool-climateepisode.
BIOSTRATIGRAPHIC UTILITY: No ancestral taxon has beenidentified in the New Zealand region, and the FAD isinterpreted as a migration event. As such, it is sharplydefined, although small population sizes may constrainprecision in locating its exact position. The principalconstraint on the use of the FAD is its restriction to shelfand upper slope sequences, although Hayward & Buzas(1979) documented its widespread but rare occurrence froma wide paleodepth range in Otaian strata of northern NewZealand. Whether the FAD maintains a consistent strati-graphic position from basin to basin is poorly tested. Thereare significantly different values of an index of evolutionarymorphology in Haeuslerella near the FAD of E. marwickiin southeastern South Island sections (Scott 1973, fig. 16).If the FAD is related to the inferred cool-climate event withwhich it is associated at Bluecliffs, it may be useful inregional chronology. Data from the Lakes Entrance sequencein Victoria are ambiguous. There the FAD is closely abovethe LAD of Globigerina euapertura (Li & McGowran 1997),as at Bluecliffs (Fig. 5). However, they also position the FADof Ehrenbergina marwicki below the FAD of Globigerina

600 New Zealand Journal of Geology and Geophysics, 1999, Vol. 42

Morgans et al.—Waitakian-Otaian Stage boundary 601
woodi and FAD of G. connecta, whereas at Bluecliffs it iswell above the latter events.
Spiroloculina novozealandica (Fig. 4B)
RECOGNITION: Spiroloculina canaliculata (Bortonian-Tongaporutuan) might be confused with S. novozealandica.In both taxa, chambers are rectangular in cross-section. Walljunctions are weakly rounded in Spiroloculina novo-zealandica but are usually angular in S. canaliculata. In somespecimens of Spiroloculina canaliculata, side walls areslightly concave, with raised sutures. Commonly, specimensof Spiroloculina novozealandica are larger and more robustthan S. canaliculata. If the material is well preserved andcarefully observed, the two taxa can be discriminatedreliably.
DISTRIBUTION: The FAD is at 89 m (J39/f201, Fig. 5) atBluecliffs. This species is mainly confined to shelf and upperbathyal environments. It is widely distributed in thesehabitats throughout New Zealand (Fig. 10) althoughHayward & Buzas (1979) and Hayward & Brook (1994)failed to record it from a variety of paleodepths from northernNew Zealand. It is generally absent from deeper waterassemblages in eastern North Island, Taranaki, andNorthland. Populations are often small. For example, itoccurs only twice in the benthic census (counts <200specimens) at Bluecliffs. Examination of material strati-graphically higher in the section than examined for this study(earlier collections re. Scott 1965) show it to be a persistentcomponent of the fauna. It is found throughout the uppersiltstone lithofacies (Fig. 3) but is absent from the SouthburnSand.
Fig. 9 Calcareous nannofossils from Bluecliffs, SEM micro-graphs. J39/fxxx = locality (see Fig. 2). Gxxxx/xx = negative incollections of Stratigraphic Solutions Ltd, Waikanae.A-D, Coccolithus miopelagicus Bukry. A (J39/f215; x2650;G1650/16; distal view). B (J39/f215; X2850; G1651/14; distalview). C (J39/fl47; x2850; G1652/19; proximal view—withcentral cross). D(J39/f215; x2800; G1651/34; proximal view—with cross). E-H, Coccolithus ovalis (Black); J39/f215.E (X4700; G1650/14; oblique distal view). F (x5500; G1655/22;distal view). G (x2800; Gl650/4; coccosphere). H (x2500;G1650/19; coccosphere). Note prominent diagenetic ring of tubecycle laths. I-L, Helicosphaera carteri (Wallich); J39/Q15.I (x3950; G1651/35; distal view, plated). J (x6100; G1651/31;distal view, plated). K (x3650; G1650/11; proximal view, flaredflange). L (X4100; G1651/8; proximal view). M, Heli-cosphaera obliqua (Bramlette & Wilcoxon) (J39/f215; X3550;G1655/24; distal view). N, Helicosphaera euphratis Haq (J39/fl47; X5000; G1655/16; proximal view). O, Discoasterdeflandrei Bramlette & Reidel (J39/O15; x3000; G1650/21;plan view—note corrosion pits). P, Micrascidites vulgarisDeflandre & Deflandre-Rigaud (J39/f215; x2250; G1651/10;ascidian sclerite, aragonitic). Q-S, Pontosphaera multipora(Kamptner); J39/f215. Q (x5800; G1651/13; distal view).R (X4800; G1652/8; distal view, corroded). S (X5000; G1656/14; proximal view). T, Cyclicargolithus floridanus (Roth &Hay) (J39/f215; x3200; G1650/15; damaged coccosphere).U, V, Calcidiscus leptoporus (Murray & Blackman); J39/fl47. U (X6100; G1658/19; proximal view—note oval distalshield). V (x5900; G1658/20; proximal view). W, X,Reticulofenestra sp.; included in Table 3 as Prinsiaceae indet.(small). W (J39/H47; X10 200; G1658/7; proximal view).X (J39/f215; x2950; G1656/16; coccosphere).
BIOSTRATIGRAPHIC UTILITY: Provided that confusion withSpiroloculina canaliculata is avoided, the FAD is welldefined morphologically. Small population sizes affectprecision in locating the FAD which, commonly, is in thenear vicinity of that for Ehrenbergina marwicki. Becausethe biofacies distribution of this species is similar toEhrenbergina marwicki, it does not significantly extend therange of environments in which the Waitakian-Otaianboundary can be identified.
Planktonic foraminiferaPotentially, the wide distribution of planktonic foraminiferafrom middle neritic through oceanic environments mightreduce the biogeographic limitations of benthic taxa usedto define the stage boundary. However, there are fewbioevents in the small cool-temperate assemblages, and nonecoincide with the stage boundary. We focus on thediscrimination of several that occur in the vicinity of theboundary and should be useful in regional biostratigraphy.
Globigerina brazieri (Fig. 7I-M)RECOGNITION Jenkins (1966), Topping (1978),Chaproniere (1988), and Hornibrook et al. (1989) all notedthe close morphological similarity of this species toGlobigerina woodi s.l., with which it usually occurs. Topping(1978) showed that variation in measured charactersoverlapped among Globigerina woodi woodi (Fig. 7A-C),G. woodi connecta (Fig. 7E-G), G. apertura, G. labia-crassata, and G. brazieri. However, Globigerina brazieriwas discriminated from G. woodi woodi with only minormisclassification when 20 variates were jointly considered.Such analyses are impractical for routine biostratigraphy,and it is necessary to use qualitatively determined characterstates to discriminate Globigerina brazieri from G. woodis.l. The most objective is a high primary aperture whichspans the penultimate and antepenultimate chambers(Fig. 71) (Jenkins 1966, fig. 6 #43, #49; Kennett &Srinivasan 1983, pi. 7 fig. 7; Chaproniere 1988, pi. 2 fig. 1,4; not Berggren 1992, pi. 1, fig. 4, 5). Of the two characters,aperture attachment is considered to be more significant thanaperture height. Usually, specimens with these features arecompact, with 3-3.5 chambers in the outer whorl. Specimensin which the aperture spans the penultimate (n-lth) and n-3rd chambers typically are less compact as they include mostof a fourth chamber in the outer whorl.
DISTRIBUTION: The LAD is placed at 47 m (J39/H59,Fig. 5) at Bluecliffs. Using the abundance of spinoseglobigerinids as a climate proxy, the LAD occurs at the closeof a warm-climate episode associated with reduced sedimentflux. The section may be condensed in the vicinity of theLAD. The event is recognised throughout New Zealand(Fig. 10) in Jenkins' (1966) Globigerina woodi connectaZone. It is similarly placed in southeastern Australia(Chaproniere 1988). Northward, Jenkins & Srinivasan(1986) recorded the LAD in Zone N8 from DSDP Site 588(26°S) and this position is also cited by Kennett & Srinivasan(1983). They regarded it as a warm-temperate to subtropicalspecies.
BIOSTRATIGRAPHIC UTILITY: The principal merit of theLAD is its wide dispersal in the New Zealand region(Fig. 10), complementing the landward, shallow-water

602 New Zealand Journal of Geology and Geophysics, 1999, Vol. 42
Waitakian localities
Globigerina euaperturaGlobigerina brazieri
EhrenberginaSpiroloculina
166'E 168'E 170 E 172'E 174'E 166 E 168 E 170 E 172 E 174 E 176 E 178 E 180
Fig. 10 (Left) Waitakian records of Globigerina euapertura and G. brazieri. (Right) Otaian records of Ehrenbergina and Spiroloculina.Both were extracted from New Zealand Fossil Record File database, July 1997. The data refer primarily to collections kept on open fileat the Institute of Geological & Nuclear Sciences, Lower Hutt. A significant number of identifications made before 1950 are not in thedatabase. Summary biostratigraphic data were inspected for each record but specimens were not examined. The records are not necessarilyadjacent to the boundary between Waitakian and Otaian Stages. Only localities with eight-digit New Zealand Map Grid references areplotted. Due to these constraints, some subsurface records of G. euapertura from Taranaki Basin are not shown. However, for thegenerally deep water eastern North Island basin, the plots show the greater dispersal of Globigerina euapertura relative to Ehrenberginaand Spiroloculina.
distribution of the benthic foraminiferal events that definethe Waitakian-Otaian boundary. While the LAD is less welldefined, morphologically, than the benthic events, the criteriapreviously given enable it to be recognised objectively.Significant demerits as a boundary proxy are its 26 mdisplacement below the boundary at Bluecliffs, imper-sistence of populations (Fig. 11), and low abundance, evenin offshore assemblages with abundant planktonicspecimens. Although the event is apparently diachronous inthe Southern Hemisphere, it may not be significantly so inNew Zealand and southeastern Australian temperate watermasses.
Globigerina euapertura (Fig. 7N-R)RECOGNITION: Although Globigerina euapertura is readilyidentifiable for much of its history, discrimination of its latestpopulations from G. woodi is difficult. This problem has
been largely ignored in the literature yet affects use of theevent in biostratigraphy. Hornibrook et al. (1989, p. 126)commented that it was "Not a particularly distinctive taxon."Topping (1978) and Kennett & Srinivasan (1983) did notdiscuss Globigerina euapertura, whereas Chaproniere(1988, 1992) referred it to Turborotalia. Although theGlobigerina woodi plexus may be phyletically distinct fromG. euapertura, as is implicit in Chaproniere (1988, 1992),the taxa have some major characters in common. Theseinclude cancellate ("honeycomb") wall topography, lowtrochospiral shells with c. 3.5-4 chambers in the outer whorl,moderate rates of chamber expansion in the outer whorl,and apertures that span the n—lth and «-3rd chambers andprincipally face into the umbilicus. Variation in othercharacters overlap and do not provide a basis for simplecategorical distinctions. In combination, several of thefollowing are useful. In Globigerina woodi the aperture

Morgans et al.—Waitakian-Otaian Stage boundary 603
OTAIAN
WAITAKIAN
20 15 10I I I I I I I I I I I5 4 3 2 1 0
Paragloborotalia spp. G. euapertura
% Planktonic specimens
G. brazieri
I I I I I I I I60 40 20 0
spinose taxa
Fig. 11 Census data on potentially useful biostratigraphic events below the Waitakian-Otaian boundary. There is an abrupt decline inParagloborotalia spp. above 55 m, which is interpreted as a population "crash". Populations of Globigerina euapertura decline up totheir disappearance from the census at 47 m. There is one higher record at 58 m (not present in the census data). The pattern is consistentwith niche contraction leading to extinction. Globigerina brazieri is rare and impersistent before its extinction. In the census data thiscoincides with an increase in spinose globigerinid specimens, possibly marking the inception of a cool-climate episode.
typically is high and the locus of maximum elevation facesdirectly into the umbilicus. It has a conspicuous porelessrim. The aperture forms a lower arch in most specimens ofGlobigerina euapertura. It faces directly into the umbilicusin some individuals, possibly including the holotype (Jenkins1960, pi. 1 fig. 8b), but in others (Fig. 7Q) it is weaklyoblique in axial orientation and very weakly extraumbilical.Often, the rim is narrow and sometimes concealed (Jenkins1971, pi. 16, fig. 462), but there are specimens in which itis raised and prominent. Sutures are less depressed inGlobigerina euapertura than in G. woodi and the shelloutline is consequently less lobulate. Although chamberoutlines are nearly hemispherical in Globigerina woodi, theyare slightly elongated in the direction of coiling in G.euapertura, creating a weakly depressed outline. In someindividuals the last chamber is set back farther from theumbilicus than in Globigerina woodi.
DISTRIBUTION: The number of specimens fluctuates(Fig. 11) but progressively declines through the lowersiltstone lithofacies at Bluecliffs. The highest record in thecensus is with Globigerina brazieri at 47 m (J39/T159), atthe end of a warm-climate episode as inferred by therelatively low abundance of spinose taxa (Fig. 5). Fromexamination of much larger strews, we locate the LAD 11 mhigher (J39/fl69). This horizon is at the termination of thenext warm-climate episode and is in the near vicinity of J39/
7602 (Slll/f602) from which Jenkins (1971, p. 147)recorded Globigerina euapertura. We have inspected hismaterial and confirm the identification. Globigerinaeuapertura is widely distributed throughout New Zealandin Waitakian strata (Fig. 10).
BIOSTRATIGRAPHIC UTILITY: Although Hornibrook et al.(1989) stated that the LAD was a lower Waitakian event,our data show that it occurs 15 m below the FAD ofEhrenbergina marwicki and might be regarded moreproperly as a stratigraphically inferior proxy for theWaitakian-Otaian boundary. The LAD is more widelyrecorded than that of Globigerina brazieri (Fig. 10) but maynot have been recognised consistently because of theproblem of discriminating specimens from G. woodi. Thisis its principal demerit. Another is the indication of small,fluctuating populations immediately preceding the LAD atBluecliffs (Fig. 11). The data suggest that specimen numbersmay be climate related. The rarity of specimens, and likelyimpersistency of populations, indicates that the LAD isdifficult to determine accurately.
Abundance of Paragloborotalia (Fig. 8A-E)RECOGNITION: The principal taxa are Paragloborotalianana nana (Fig. 8A, B), P. nana semivera (Fig. 8C, D), andP. nana pseudocontinuosa (Fig. 8E). Although these taxahave many similarities and are difficult to separate, the

604 New Zealand Journal of Geology and Geophysics, 1999, Vol. 4
Fig. 12 Planktonic foraminifera from Bluecliffs, SEM micrographs. J39/fxxx = locality (see Fig. 2). FPxxxxx = catalogue number i fspecimen in foraminiferal collections at Institute of Geological & Nuclear Sciences, Lower Hutt. md = maximum dimensiaA-D, Globorotalia cf. kugleri Bolli; J39/f7617. A (FP4558; md = 0.11 mm). B (FP4559; md = 0.14 mm). C (FP456 :md = 0.13 mm). D (FP4561; md = 0.14 mm). E-H, Globorotalia cf. incognita Walters; J39/fl34. E (FP4562; md = 0.25 miF(FP4563;md = 0.27mm). G (FP4564; md = 0.28 mm). H (FP4565; md = 0.26 mm).
abundance data (Fig. 11) refer to the generic representation,and discrimination is not a significant problem. Some formsare referred to Globorotalia cf. incognita (Fig. 12E-H) butare a minor part of the Paragloborotalia plexus. Althoughthere are excursions, the relative abundance of Para-globorotalia climbs from 8% near the base of the section to24% at 45 m. Larger excursions follow and abundance spikesdecrease between 45 and 55 m. The latter position marksthe inception of a steep decline in relative abundance to 1%at 62 m (J39/fl72). There is a recovery to 15% at 78 m, butfor most of the interval above the boundary values are < 10%.In detail, the signature appears to be cyclic at frequencies<10 m, stratigraphically, but more closely spaced samplesare needed to establish the pattern. Biostratigraphically,the most distinctive feature is the population crash at62 m.
DISTRIBUTION: Paragloborotalia is widely distributed inEarly Miocene planktonic foraminiferal assemblages but welack comparable abundance data. The crash at 62 m iscoincident with a small increase in spinose globigerinids(Fig. 11) and excursions in oxygen isotopes (Fig. 5). Aclimatic response is suggested.
BIOSTRATIGRAPHIC UTILITY: If confirmed elsewhere, thepopulation event between 56 and 62 m is potentially usefulbecause it is sharply defined, stratigraphically, and occursin an easily recognised taxon. It is approximately coincident
with the LAD of Globigerina euapertura. It usefully mayindicate inferior proximity to the boundary.
NannofossilsOverview
There are no published studies on the nannofossils i fBluecliffs other than Kadar's (1983, pp. 108-9) list of thetaxa in a sample collected from about the same horizon asour highest sample. His list is similar to ours oncenomenclatural changes and likely different taxonoimcconcepts are taken into account. The main differences areKadar's records of Coronocyclus nitescens and Triquetr< •-rhabdulus milowii. Coronocyclus nitescens is a distinctivetaxon that preferred fully oceanic conditions (A.R.Cunpubl.), and its apparent complete absence from oursamples at Bluecliffs was not anticipated. Converse] r,Triquetrorhabdulus milowii, a stenothermal warm-w;it rtaxon, was not expected to occur at Bluecliffs, and i. erecords may refer to specimens of Micrascidites vulgai is(Fig. 9P) in which some arms have been disarticulated.
Special mention should be made of the presence ofReticulofenestra sp. ("clifdenensis") (Table 3). This large,distinctive, unnamed taxon resembles the Late Eocene -Early Oligocene species R. oamaruensis. Always rare, itoccurs sporadically in New Zealand in strata of upperWaitakian - lower Lillburnian (Early-Middle Miocene) age,including the Altonian of Clifden. Also worth noting is the

Morgans et al.—Waitakian-Otaian Stage boundary 605
occasional presence of specimens of Calcidiscus leptoporus(Fig. 9U, V). They represent one of the oldest known recordsof this long-ranging Neogene species.
Recycled nannofossils sporadically present at BlueclifFs,some as well-preserved specimens, include Chiasmolithusaltus, C. oamaruensis, Reticulofenestra bisecta, R.placomorpha, and Zygrhablithus bijugatus. Their presenceindicates the occurrence, somewhere along orupslope, of afully marine sequence of Late Eocene and Oligocene age.
Craigmore Limestone
None of the Craigmore Limestone nannofloras aresufficiently diverse to allow them to be dated accurately orplaced with any certainty in any particular zone (Table 3).However, their stratigraphic position indicates that theymight be impoverished Helicosphaera euphratis Zoneassemblages. Furthermore, it may be significant that therichest two assemblages lack Reticulofenestra bisecta, arobust and ubiquitous taxon that became extinct in the lowerWaitakian. Thus, a middle or late Waitakian age is plausiblefor this part of the Craigmore Limestone.
Waitoura Marl, Bluecliffs Silt
The nannofloras of the Waitoura Marl and Bluecliffs Siltcontain Helicosphaera carteri, and Discoaster deflandrei isthe only discoaster present (Table 3). Thus, they can beplaced in the Helicosphaera euphratis Zone of Edwards(1982). Although this zone was originally thought to rangein age from earliest Otaian to middle Altonian (Edwards1982), the results of the present study indicate that the baseof the zone occurs within the upper Waitakian. Furthermore,Calcidiscus macintyrei s.l., Discoaster variabilis, andReticulofenestra pseudoumbilica, all of which first appear
in the younger (upper Otaian - mid Altonian) part of thiszone, are not present. Also not observed, possibly due tocooler water temperatures, are several other biostrati-graphically useful early Neogene nannofossils. They includeDiscoaster druggi, Helicosphaera ampliaperta, H. recta,Ilselithina fusa, Sphenolithus belemnos, S. conicus, andTriquetrorhabdulus carinatus.
Abundance of Helicosphaera
The only biostratigraphically significant nannofossil eventobserved in the sequence at Bluecliffs is a reversal in theabundance of Helicosphaera carteri and H. euphratis. Itoccurs very close to the Waitakian-Otaian Stage boundaryand probably is best characterised as the top of the acme ofH. euphratis (Table 3). An important bioevent in theMediterranean region (Fornaciari & Rio 1996), it may alsobe a useful correlation tool within the New Zealand region.
Dinoflagellates
Dinoflagellate cysts occur throughout the Bluecliffs Silt inreasonable diversity (Table 4), with assemblages typical ofthe Oligocene-Middle Miocene. It is difficult to assign aprecise age because of the apparent lack of stratigraphicallyimportant taxa, and no obvious bioevents near the Waitakian-Otaian Stage boundary were observed. The relative lack ofdata on known ranges of Oligocene and Miocene dino-flagellates in New Zealand hinders the usefulness of thisgroup in this part of the column.
Palynoflora
Pocknall (1982) recorded diverse floras of 30 spore, 8gymnosperm, and 88 angiosperm taxa in 21 collections(Fig. 2) from Bluecliffs Silt and assigned them all to the
Table 4 Dinoflagellate distribution chart for selected samples (Fig. 2) from BlueclifFs.
Fossil record number J39/Catalogue number
fl 16 fl21 fl25 f7594 f7600 f7606 fl82 f7612 f7613 17616 f7619 f7626L17626 L17627 L17628 L10142 L10143 L10144 L17629 L10145 L10146 L10147 L10148 L10149
Chiropteridium lobospinosumCassiculosphaeridia sp.Systematophora placacanthaOperculodinium centrocarpumDeflandrea cf. heterophlyctaLejeunecysta sp.Phtanoperidinium sp.Cordosphaeridium sp."Pyxidinopsis sp.
Saturnodinium sp.Chiropteridium spp.Hystrichokolpoma rigaudiaeImpagidinium cf. victorianumNematosphaeropsis balcombianaOperculodinium tiaraThalassiphora sp.Spiniferites ramosusHystrichokolpoma sp.ICorrudinium sp.Deflandrea phosphoriticaCleistosphaeridium sp.Plerodinium cingulatumMembranilarnacia sp.Deflandrea leptodermataPolysphaeridium cf. zoharyiOperculodinium sp.Thalassiphora pelagica
X
X
X X
X X
X
X
X
X
X
X
X
X
X
X
X
X
X
X
X X X
X X
X
X
X X X X X
X
X X X X
X X
X
X X X X X X
X X X
X X X
X
X
X
X
X

606 New Zealand Journal of Geology and Geophysics, 1999, Vol. 42
Proteacidites isopogiformis Zone (Pocknall & Mildenhall1984). The palynofloras were notable for the consistency ofdominant palynomorphs, and no significant events were seennear to the Waitakian-Otaian boundary.
MolluscsThe following data were supplied by Alan Beu (pers. comm.1997).
The lowest localised collection (J39/f579), from the baseof the banded lithofacies, includes Limopsis, Globisinummiocaenicum, Proximitra rutidoloma, Spinomelon, Comitasfusiformis, and Zeacuminia. It includes a specimen ofNotocorbula humerosa whose size resembles small Otaianpopulations rather than the large specimens of Waitakianpopulations. Otherwise the collection is not age diagnostic.
A collection (GS3699, J39/f7496) from the lower thirdof the banded lithofacies has a typical Otaian assemblage,including Spissatella trailli, Zeacolpus woodhouseae,Bathytoma haasti, and Austrotoma lawsi and was assignedto Otaian Stage by Marwick (1971). Other collections fromthe banded siltstone lithofacies below the Otaian boundarylack diagnostic Waitakian taxa and resemble Otaian faunasfrom higher in the section.
Bluecliffs Silt assemblages are uniform in compositionand compare with those from similar lithofacies in MountHarris Beds and Rifle Butts Formation to the south. Theycontrast with the shallower assemblage in the Waitakianstratotype at Trig Z. This overview indicates that somemolluscan FAD events cited for Otaian by Finlay & Marwick(1947) occur below the Waitakian-Otaian boundarystratotype as defined by the FAD of the foraminiferEhrenbergina marwicki.
CORRELATION
New Zealand zonesJenkins (1966) cited Bluecliffs section in his originalproposal of the planktonic foraminiferal Globigerina woodiconnecta Zone (Fig. 13). He noted that the LAD ofGlobigerina brazieri and LAD of G. euapertura occur inthe lower part of the zone. Globigerina woodi connecta ispresent throughout the stream section and cliff section atBluecliffs. The base of the zone probably lies in theunexposed interval above Waitoura Marl (Fig. 3). CraigmoreLimestone and Waitoura Marl are correlated with theGlobigerina woodi woodi Zone. Jenkins (1966) stated thatthe upper part of this zone is present in Otekaike Limestonein the Waitakian stratotype at Trig Z, Waitaki valley.
Exposures of Craigmore Limestone and Waitoura Marland all of the stream section and cliff section sampled inthis study lie in the lower part of the calcareous nannofossilHelicosphaera euphratis Zone (Edwards 1982).
Pocknall (1982) assigned Bluecliffs floras, includingthose from higher strata in the upper siltstone lithofacies(Fig. 2), to the Proteacidites isopogiformis Zone (Pocknall& Mildenhall 1984).
Tropical-subtropical zonesJenkins (1971) recorded Globorotalia cf. kugleri in S i l l /f617, 619, and 626 from strata above the Waitakian-Otaianboundary at Bluecliffs. The material is tiny and atypical ofGloborotalia kugleri (Fig. 12A-D). We consider that it is
inadequate to identify reliably planktonic foraminiferal ZoneMl (Berggren et al. 1995), which is defined by the totalrange of the nominate species. Zone M2 refers to the intervalbetween the LAD of Globorotalia kugleri and the FAD oiGlobigerinatella insueta. The only record of the latter irNew Zealand is from V22/f6530, southern Hawke's Bay.
The Bluecliffs nannofloras lack the primary indexspecies needed for exact correlation with either the standardlow-latitude zonal scheme of Bukry (1973,1975) and Okadji& Bukry (1980) or the similar constructs (Fig. 13) of Martini(1971) and Martini & Muller (1986). However, some broadcorrelations are possible, after making nomenclatural andtaxonomic adjustments, using some of the other bioeventf;mentioned by these authors. Thus, undifferentiatedcorrelation with Bukry's (1973, 1975) Early Miocene CN1.CN2, and lower CN3 Zones is indicated by the knownstratigraphic position of the Bluecliffs section above theLAD Reticulofenestra bisecta (a top CP19b bioevent) andbelow the FAD Calcidiscus macintyrei s.l. (upper CN3)Similarly, correlation with Martini's (1971) Early Mioceneupper NN1 and NN2 Zones is indicated by the knownstratigraphic position of the Bluecliffs section above tin:LAD Reticulofenestra bisecta (a basal NN1 bioevent), abovuthe FAD Helicosphaera carteri group (mid NN1), and belev,the FAD Calcidiscus macintyrei s.l. (top NN2).
Temperate-transitional zonesBerggren et al. (1995) integrated zonations developed fojseveral mid-latitude regions in hemispheres into a unifiesscheme (Fig. 13). Zone Mtl, like low-latitude Zone Ml, i-based on the range of Globorotalia kugleri and is ncrecognisable with certainty at Bluecliffs. Zone Mt2 is define:by the partial ranges of Globorotalia incognita an*:Globorotalia semivera between the LAD of Globorotaiikugleri and the FAD of G. praescitula. The latter event mark.'the Otaian-Altonian boundary and was recorded frorSouthburn Sand at Bluecliffs (Scott 1972). However, becausiof their dependency on the LAD of Globorotalia kuglerZones Mtl-Mt2 cannot be resolved satisfactorily gBluecliffs. This situation applies throughout New Zealand
Tasman Sea
DSDP Site 593 (Jenkins & Srinivasan 1986) records a lowerbathyal oceanic sequence at the western edge of ChallengerPlateau, c. 500 km northwest of Bluecliffs, and provides aninformative perspective on the zones developed by Jenkins(1966) and Edwards (1982) from onshore sequencesGlobigerina woodi woodi and Globigerina woodi connectaZones were recorded by Jenkins & Srinivasan (1986), whoidentified Globorotalia kugleri from both zones. PossiblZones Mtl-Mt2 can be identified at DSDP Site 593. This i -not possible in sequences around the New Zealand landmas, -and indicates the value of the globigerinid zones propose iby Jenkins (1966) in neritic settings.
The calcareous nannofossil correlation betweenBluecliffs and Site 593 is based on the interval between thetop of consistently present Reticulofenestra bisecta (andcoincident top of Helicosphaera recta and Zygrhablitb.itbijugatus) to the base of Calcidiscus macintyrei, an intervalthat Lohmann (1986) assigned to Martini's NN1, NN2, ar ilower NN3 Zones. This correlation is compatible with, butsomewhat broader than, that indicated by the plankton i;foraminifera. The most obvious difference between the tw >

5"re
reQ.pc?
5'the
b
oc3d
ary
sec
5'3
Oeoch
i
o3Olo
gi
oal da
P'—*
3
2!
'ent
pro
xi
12
era'
Oligo
oft
?-1
<<*
ioc©
n
3 "-on
olc
lN
Epoch
Age c?
Time Ma <"(D
•a 2
§ 12 <?5" 3
:iona
1 tim
OP
3T
3 -
—
p
calceireou
3
33O_
1£—Noa
tio
3
DUNTROONIAN
G. euaperturaZone
WAITAKIAN OTAIAN ALTONIAN
G. dehiscens f G. woodi woodi ••>>Zone ^J_ Zone -i>
P22Globigerina clperoensis IZ
AP16Globigerina euapertura IZ
Mt1 Globorotalia kugleri TRZ
primodrdius)lb <Gt- k u a l e r i -
AN1Globigerina brazieri PRZ
G. woodi connectaZone
Mt2Gt. incognita - Gt. semivera PRZ
AN2Globorotalia incognita PRZ
•s> G. trilobus trilobus• • p Zone
Mt3 Gt. praescitula -Gt. miozea IZ
AN3Gt. praescitula IZ
NP25 NN1 NN2 NN3
CP19
MNP25a MNP25b
CN1CN1a&b CN1b
MNN1MNN1a MNN1b|MNN1C MNN1d MNN2a MNN2b
CN2
MNN3a
NN4
CN3
MNN3b MNN4a
NEW ZEALAND STAGES
Jenkins (1966,1971)
TransitionalBerggren et al. (1995)
(Sub)AntarcticBerggren (1992)
Helicosphaera euphratis Zone
Martini (1971)
Bukry 1973,1975Okada&Bukry 1980
Fornaciari& Rio (1996)
i Edwards (1982)
Bluecliffs section as constrained by strontium isotope ages70 metres — * • • <— 50 metres —*•'.
i'i !iUpper Bluecliffs section not examinedin this study (see Fig. 3)
Globigerina euapertura -4
Globigerina brazieri -4
Paragloborotalia spp.
Helicosphaera euphratis(more abundant)
p | Strontium ages Graham et al. (in prep.)
— • Ehrenbergina marwicki
— • Spiroloculina novozealandicaGloborotalia praescitula (Scott, 1972: S119/f482 =
J39/f9482 = F6574)below (average above
f173=4.4%)
\ \ • Helicosphaera carteri(more abundant)
IO33O
II1o03
COrjjQ
Cm-n<•nmm-z
CO
3
o
CTQft)
croa
IL I li . i i . ii'llli: ii: it 11iii 11 i l : \l CO
522 Ma Strontium ages Graham et al. (in prep.) 21.7 Ma-
Globigerina brazieri
Globigerina euapertura
Paragloborotalia spp. (average below f 173 = 13.2%)
Waitakian -«t»- OtaianEhrenbergina marwicki |
H Spiroloculina novozealandica
20.5Ma
Helicosphaera euphratis (more abundant) Paragloborotalia spp. (average above f 173 = 4.4%)
EgrndODET3Jn>
Tl-rjcom
Helicosphaera carteri (more abundant)

608 New Zealand Journal of Geology and Geophysics, 1999, Vol. 42
nannofloras is that Coronocyclus nitescens, Discoasterdruggi, Sphenolithus belemnos, S. dissimilis, andTriquetrorhabdulus carinatus occur rarely at Site 593 buthave not been found at Bluecliffs, and that Helicosphaerahas not been recorded from this part of the Early Mioceneat Site 593. It is likely that these dissimilarities reflectpaleoenvironmental rather than biostratigraphic differ-ences.
Southern oceansAs with other interregional Early Miocene schemes, theplanktonic foraminiferal zonation (Fig. 13) proposed byBerggren (1992) for subantarctic sequences fits poorly withBluecliffs data. The LAD of Globigerina euapertura islocated at the base of Zone AN1 (= base of Miocene),whereas at Bluecliffs it occurs at the level of the LAD of G.brazieri, which defines the top of Subantarctic Zone AN1.However, Zone AN2 (for the interval between the LAD ofGlobigerina brazieri and the FAD of Globorotaliapraescitula) is recognisable at Bluecliffs. The Waitakian-Otaian boundary lies near the base of this zone.
The Bluecliffs nannofloras correlate with an unnamedEarly Miocene biostratigraphic interval recognised by Wei& Wise (1992) at ODP Sites 747, 748, and 751. Theirunnamed interval, which they tentatively correlated withBukry's Zones CN1-CN3, extends up from the top ofReticulofenestra bisecta to the base of "Calcidiscusleptoporus-C. macintyrei". In New Zealand, the LAD ofReticulofenestra bisecta is in middle Waitakian, whereas theFAD of "Calcidiscus leptoporus-C. macintyrei" is in upperOtaian (Edwards 1982).
MediterraneanThe Waitakian-Otaian boundary at Bluecliffs (Fig. 13) canbe correlated directly with the calcareous nannofossil ZoneMNN1-MNN2 boundary of Fornaciari & Rio (1996). Bothare marked by a reversal in the abundance of Helicosphaeracarteri and H. euphratis, specifically the top of the acme ofH. euphratis. Fornaciari & Rio (1996) also indicated thatthis event provides an easy means of distinguishing betweenthe Aquitanian and Burdigalian Stages in the Mediterraneanregion. The LAD of Chiropteridium spp. (Dinophyceae),which is present up to J39/O82 (Fig. 2), occurs in theLemme-Carrisio Section, Italy, 1 m below the proposedposition of the Oligocene-Miocene boundary (Steininger etal. 1997), although other evidence suggests the genus rangesinto the Middle Miocene (Williams et al. 1993).
DATING
PaleomagnetismPrevious studies have shown New Zealand Neogenemudstones to have complex and variable paleomagneticproperties and behaviour. They generally have low-intensityprimary magnetisation (generally <10~3 A/m) commonlyoverprinted by secondary magnetisations (e.g., Wright &Vella 1988; Roberts et al. 1994; Wilson & McGuire 1995).Rock magnetic and petrographic investigations havedemonstrated that common remanence carriers for thesesediments are titanium-rich ferrimagnetic iron-oxides(titano-magnetite and hemo-ilmenite) and iron sulfide(greigite and pyrrhotite) minerals (Roberts & Turner 1993;
Wilson & Roberts 1999). For these reasons, and also becausethe sediments from South Canterbury have not previouslybeen studied for paleomagnetism, we conducted a pilot studyof the Bluecliffs section to assess paleomagnetic behaviourand stability of remanence and to assess appropriatedemagnetisation techniques.
All specimens measured from Bluecliffs were over-printed by a low-coercivity normal overprint from the presentgeomagnetic field. This was generally cleaned by 150°C ofheating or peak alternating fields of 15 m l Vectorcomponent plots were noisy, and stable endpoints were netreached before thermal alteration of the clay component ofthe specimens. Thermal demagnetisation allowed identi-fication of trends in demagnetisation. Where primaryremanence was of reversed polarity, a further normalgeomagnetic field overprint overlapping in blockingtemperature spectra prevented elucidation of a stable primarycomponent (e.g., Fig. 14A, B). AF demagnetisation was noteffective in cleaning this component and, in general, vectorcomponent plots for specimens cleaned by AF methods werenoisy and showed random behaviour at peak fields above20 mT (e.g., Fig. 14F, H). The minerals holding the magneticremanence in the Bluecliffs section are of low coercivityand low unblocking temperature and do not always respondto AF demagnetisation methods. The signal is perhaps dueto a combination of titanomagnetite and ilmeno-hematitegrains, but the presence of partially reduced sulfides cannoibe ruled out. Petrographic analysis revealed ubiquitous pyri cein several thin sections prepared from unheated paleo-magnetic samples and minor titano-magnetite in one thirsection (D. Smalepers. comm. 1993). The ubiquitous pyritesuggests full reduction of the iron-sulfide component of theBluecliffs sediments.
Remagnetisation great circle analysis (for methodologysee McFadden & McElhinny 1988) on several samples fromwithin a small stratigraphic range (Fig. 15) demonstrate;that, although stable endpoints were not identifieddemagnetisation trends from a number of specimens can beused to identify a reliable endpoint for sites that are clearlytrending towards a primary remanence of reversed polarityMore detailed and comprehensive paleomagnetic samplingand analysis will identify a magnetostratigraphy for the;Bluecliffs section. However, several specimens must becollected from each site and they must be subject to arigorous and closely spaced thermal demagnetisationprocedure for site polarity means to be identified by greatcircle remagnetisation analysis.
Strontium isotopesWhole rock, benthic, and planktonic foraminiferal shell-were analysed for 87Sr/86Sr data. Sample locations are showrin Fig. 2. Analytical methods are discussed by Graham eial. (in press). Ages were obtained using the "look-up" table:of Howarth & McArthur (1997), relative to the time-scakof Cande & Kent (1995). For most of the section, benthuand planktonic foraminiferal shells give consistent buislightly offset ratios. Overall, they increase gradually up th<section, although there are several excursions to lower ratio •caused by interaction with pore fluids. Samples from tinlower siltstone lithofacies between R16440 and R1669/(Fig. 13) have similar ratios corresponding to a mean age o"c. 22 Ma. From 66 m (R16453), upward through tlruWaitakian-Otaian boundary at 73 m, to the highest sampk

Morgans et al.—Waitakian-Otaian Stage boundary 609
A) 2821.3 (Thermal, °C) B) 2821.2 (AF, mT)
N U N,Up
C) 2848.3 (Thermal, °C)
N,Up
D) 2848.1 (AF, mT)
NXIp
~>NRM
NRM=9.8x105 A/m0 Demag Level 3 5 ° 0 Demag Level 4 0 0 Demag Level 350 0 Demag Level 4 0
E) 2873.3 (Thermal, °C) F) 2873.1 (AF, mT) G) 2889.2 (Thermal, °C) H) 2889.1 (AF, mT)
N,Up N,UpNRM
NRM=1.1x10"a A/m
N,Up
NRM-1.3X10-1 A/m
Demag Level0 Demag Level 4 0
0 Demag Level 3 5 ° 0 Demag Level 40
Fig. 14 Vector component diagrams of demagnetisation behaviour of representative samples (Fig. 2) from the pilot study of theBluecliff section. A, B, Comparison of thermal and AF demagnetisation of reversed polarity trending samples 2821.3 and 2821.2,respectively. C, D, Comparison of thennal and AF demagnetisation of reversed polarity trending samples 2848.3 and 2848.l'respectively. E, F, Comparison of thennal and AF demagnetisation of reversed polarity trending samples 2873.3 and 2873.1,respectively. G, H, Comparison of thermal and AF demagnetisation of normal polarity samples 2889.2 and 2889.1, respectively.
(R16462) at 125 m, ratios increase smoothly. In theimmediate vicinity of the Waitakian-Otaian boundary, theycorrespond to an age of 21.7 Ma. At 125 m the age isc 20.5 Ma, equivalent to the Aquitanian-Burdigalianboundary. Confidence limits for the ratios at the 95% levelcorrespond with age ranges of ±0.25 Ma.
Stable isotopesWright & Miller (1992) located the base of oxygen isotopeZone Mi la at a prominent enrichment event in EarlyMiocene benthic foraminiferal isotope data at 282.8 m bsfin north Atlantic DSDP Site 563. It occurs in Chron C6Ar(21.32-21.77 Ma, Cande & Kent 1995). To determine thepresence of this event at Bluecliffs, carbonate from thebenthic foraminifera Cibicides novozelandicus andEuuvigerina spp. was analysed from 46 samples at c. 2 mintervals through the section (Fig. 2, 5). Major features ofthe data are the generally heavier values and largerexcursions for Euuvigerina spp. than for Cibicidesnovozelandicus, variable covariance of the signatures, andthe presence of several enrichment/depletion cycles between25 and 63 m. The largest excursion is the enrichment of2.08%o in Euuvigerina spp. between 34 and 43 m (Fig. 5).However, this is not mirrored by Cibicides novozelandicus,and we are uncertain of its climatic significance. The
signatures covary more closely near the enrichmentmaximum in Euuvigerina spp. (1.73%o) at 63 m, althoughthe amplitude of the excursion is much greater than inCibicides novozelandicus.
Widely spaced samples, variable and sometimes weakcovariance between the benthic signatures, use of benthictaxa not previously analysed in stable isotope studies, lackof data from planktonic carbonate, and the possibility ofcontamination or exchange with sedimentary carbonate insome samples inhibit robust interpretations of the data.Therefore, we rely on the gross pattern of the signatures,both of which record larger amplitude enrichment/depletion cycles between 25 and 63 m than elsewhere inthe sequence, and identify this interval as a possiblerecord of Mi la.
Bioevent proxiesGlobigerina brazieriLi et al. (1992) recorded the LAD of Globigerina brazieriat 124 m bsf at ODP Site 747A, Kerguelen Plateau, in ChronC6Bn (Heider et al. 1992, 22.59-23.07 Ma). Berggren(1992) recorded the LAD in Chron C5En (18.28-18.78 Ma)at ODP Sites 747A and 748B, and in C5Dn (17.28-17.61 Ma) at ODP Site 751A (all Kerguelen Plateau).However, the specimens figured by Berggren (1992, pi. 1,

610 New Zealand Journal of Geology and Geophysics, 1999, Vol. 42
Poles to remagnetlsed great circles
Circles2813.32819.22821.32826.12830.3
Df)053.0025.9298.5340.9091.8
If)40.041.722.735.309.1
Solution
D= 190.5' l=45.9i =2.9'
Fig. 15 Example of remagnetisation great circle analysis forobtaining the characteristic remanence direction for severalspecimens from a short stratigraphic interval.
fig. 4, 5) do not match our interpretation of Globigerinabrazieri, and his data may not be relevant.
Globigerina euaperturaFrom records in ODP Sites 747A and 747B, Berggren et al.(1995) placed the LAD of Globigerina euapertura in ChronC6Cn.2n (23.68-23.80 Ma), at the Oligocene-Mioceneboundary and before the FAD of Globoquadrina dehiscens(Chron C6Br, 23.07-23.35 Ma). This order conflicts withour record of the LAD of Globigerina euapertura well abovethe FAD of Globoquadrina dehiscens. The discrepancy maybe due to the earlier extinction of Globigerina euaperturain southern high latitudes than at Bluecliffs, or converselyan earlier appearance of Globoquadrina dehiscens in NewZealand, or to differences in taxonomic concept.
Abundance of HelicosphaeraThe abundance reversal of the nannofossil taxa H. euphratisand Helicosphaera carteri occurs close to the Waitakian-Otaian Stage boundary. Correlation of this event with theMediterranean Neogene zonal scheme of Fornaciari & Rio(1996), and with data in Steininger et al. (1997), indicatesan age of less than 21.3 Ma, or c. 21 Ma. Although Fornaciari& Rio (1996) used the latter date for the Aquitanian-Burdigalian boundary, Berggren et al. (1995) placed it at20.5 Ma. The age of the reversal event, less than 21.3 Ma,is too young compared to the strontium isotope date of21.7 Ma. This inconsistency may be a combination of erroron the strontium isotope date (±0.25 Ma) and a slightlyearlier abundance reversal in New Zealand.
Chiropteridium spp.The highest occurrence of this taxon is in sample J39/fl82,In the proposed Paleogene-Neogene boundary stratotype irthe Lemme-Carrosio section, Italy, its LAD is 1 m abovethe boundary (23.80 Ma, Steininger et al. 1997). However,other records suggest that the genus Chiropteridium extendsinto the Middle Miocene (Williams et al. 1993).
EVALUATION
We focus on features of the Bluecliffs section relevant to its;utility as a reference for the Waitakian-Otaian boundary.
Sedimentary recordAlthough the stage boundary is conceptually a surface, wehave examined a 125 m sequence about the boundary tcplace it in environmental context and to locate events in itsvicinity that might serve as "near boundary" markers. Thesection meets primary reference requirements of goodexposure and accessibility. The fine clastic lithofacies l-relatively uniform throughout, apart from the cementedhorizons in the banded lithofacies. Fossils are well preservedThe sequence has no structural complications and there an:no indications of breaks in deposition.
In closer focus, the depositional record at Bluecliffs i-much less uniform than field examination indicates. A-judged by patterns of foraminiferal specimen abundanceand supported by the coarse fraction signature, sedimeminflux was cyclic with a succession of well-defined minimathat identify condensed intervals. The cyclic pattern centreson prominent events at 64 and 27 m below the boundaryand wanes upward to be a minor feature at, and above, theboundary. The pattern suggests that the stratigraphic scaleis unlikely to be a linear measure of time, particularly belowthe boundary. The pattern might affect the separation ofclosely spaced events occurring within any of the condensedintervals. To the extent that the sedimentary pattern is aeustatic response, it should be recorded below the boundarythroughout the region.
Paleogeographically, the section represents a depositional site at upper bathyal depths, intermediate betweershoreline and open ocean. No significant changes in thi>setting occur in the vicinity of the boundary, although th:basin shallowed. New Zealand Cenozoic stages are locaunits used by stratigraphers studying landward marine an:terrestrial basins. Only at the outer limits of the offshorteconomic zone are fully oceanic sequences encountered. I:this perspective, the intermediate location of the Bluecliff i-site is valuable as it records both terrestrial and oceanievents, although neither record is optimal. More limitiii;,for correlation of local biostratigraphic units with regionaand international equivalents is the southern setting of tkisite in the vicinity of a major water-mass boundary, tb:Subtropical Convergence (Edwards 1975). The absence o:typical Globorotalia kugleri from the Early MiocemCanterbury Basin is an example of this paleogeographi:restriction.
BioeventsFor biostratigraphy, the diverse biota preserved in BlueclitfsSilt is valuable as it associates bioevents with distinc;biogeographies and potentially allows more rigorous testin L;

Morgans et al.—Waitakian-Otaian Stage boundary
of their stratigraphic order. While large molluscs and othermacro-invertebrates are poorly represented near theboundary, probably because of the paleodepth, smallmolluscs occur in foraminiferal residues, as do ostracods.Among the protists, the foraminiferal record is excellent,although several significant siliceous groups (radiolarians,diatoms) have not been found in our material. Algae are wellrepresented by coccoliths and dinoflagellate cysts.Importantly, for terrestrial correlations, there is a fair recordof pollens and spores. In overview, with several exceptions,there is a good fossil record near the boundary at Bluecliffs.To a degree, the small number of bioevents currently suitablefor biostratigraphy reflects the small amount of research onthe preserved biota.
Climate is a powerful engine of biogeographic andevolutionary change. Although the existence of an episodeof cool climate in the immediate vicinity of the boundary isnot confirmed by the land flora, and the isotope data areinconclusive, patterns of marine bioevents in this vicinityindicate environmental change and faunal turnover. Thereare extinctions (Globigerina brazieri and G. euapertura)below the boundary. The waning and then intermittentrepresentation of the latter (Fig. 11) before extinction suggestniche contraction and eventual loss (Maynard Smith 1989).Other taxa (e.g., Paragloborotalia spp.) survived throughthe boundary in small numbers, or responded with changesin dominance (Helicosphaera euphratis, H. carteri). Incontrast, the appearance of the benthic foraminifersEhrenbergina marwicki and Spiroloculina novozealandicamight be viewed as migrants taking advantage of newlyavailable territory. While these scenarios are speculative,they suggest that the present stratigraphic position of theWaitakian-Otaian boundary possibly is tuned to a significantenvironmental event and usefully located for biostratigraphy.Further events similar to those above might be found nearthe boundary and in the preceding c. 60 m.
CONCLUSIONS
The base of the Early Miocene Otaian Stage in its stratotypesection at Bluecliffs is recognised as the boundary stratotypefor the Waitakian and Otaian Stages. The FAD of the benthicforaminifer Ehrenbergina marwicki defines the boundaryand occurs at 73 ± 1 m above the base of the section. Theboundary is well exposed.
The fine clastic sequence adjacent to the boundary wasdeposited on a passive margin at upper bathyal depths in abasin defined by a submarine high and archipelago. The sitewas distant from shorelines. The cool-temperate water masswas in an intermediate position between neritic and oceanicwater and possibly adjacent to the Subtropical Convergence.There was good southward fetch. Landmasses had a warm-temperate climate, were forest covered, and wet.
Abundance signatures of foraminiferal specimens belowthe boundary are cyclic. The spikes covary with declines inrelative weights of coarse sediment (>75 |J,m fraction) andlikely identify condensed intervals produced by highstands.On this interpretation there is a minor condensed interval atthe boundary but neither here, nor through the precedingsection which contains larger amplitude cycles, is there fieldevidence that the sequence is significantly incomplete. Weconsider that the sedimentary record of the section does notcompromise its utility as a reference at the current level ofbiostratigraphic resolution.
611
Climatic instability through the c. 40 m sectionimmediately underlying the boundary is indicated by cyclesin the relative abundance of spinose globigerinids (a cool-water proxy) and by the oxygen isotope data for benthic taxa,although the two signals do not closely covary. Severalplanktonic foraminifera become extinct in this interval andthere are major changes in the relative abundances ofnannofossil and planktonic foraminiferal taxa. These events,and the cyclic sedimentary record in the same interval, maybe connected with global oxygen isotope Zone Mi la. Datingis insufficiently precise to confirm the linkage but it ispossible that the Waitakian-Otaian Stage boundary is locatedclose to a significant global climatic event and might be auseful local proxy.
The diverse biota in the vicinity of the boundary includesplankton and bottom dwellers, as well as a record ofterrestrial vegetation. This diversity assists biostratigraphicstudies by allowing events in several groups to beinterrelated. However, the paleogeographic position of thesection in a cool-temperate water mass at a southern sitelimits its utility as a reference, particularly in interregionalcorrelation.
The FAD of Ehrenbergina marwicki, the primary eventmarking the Waitakian-Otaian boundary, is well defined inthe section and widely distributed in outer shelf to upperbathyal marine biofacies around New Zealand. It should beretained. Molluscan events cited for the boundary by Finlay& Marwick (1947) occur in Waitakian strata significantlybelow the boundary and need to be reassessed. Supple-mentary, widely dispersed bioevents that identify theboundary are required.
We did not find high-resolution planktonic foraminiferalor calcareous nannofossil events in the immediate vicinityof the boundary that assist its recognition in offshoresequences. Below the boundary there are events (abundancechanges in Paragloborotalia spp. and Helicosphaera spp.,LAD of Globigerina euapertura) in these groups that appearto be useful "proximity" indicators.
Strontium isotopes date the boundary at 21.7 Ma.Although our work slightly modifies the date conjecturedby Edwards et al. (1988), the need for improved precisionremains. The conflict with the nannofossil date of c. 21 Maalso needs to be resolved. Paleomagnetic studies indicatethat it should be possible to resolve a magnetostratigraphyfor the section. A priority for future chronostratigraphicresearch should be the selection of a boundary para-stratotype with quality polarity and diverse bioeventsignatures.
ACKNOWLEDGMENTS
We are grateful to Jeff Ashby, Alan Beu, Dallas Mildenhall, andPeter King for data, and Malcolm Laird for assistance in the field.Wendy St George undertook the scanning electron microscopy andprepared some illustrations. Critical comment from Alan Beu,George Chaproniere, Brad Field, Bruce Hayward, and DavidWaghorn significantly improved the manuscript. The research wassupported by FRST contracts CO5617 and STR401.
REFERENCES
Allan, R. S. 1933: On the system and stage names applied tosubdivisions of the Tertiary strata in New Zealand.Transactions of the New Zealand Institute 63: 81-108.

612 New Zealand Journal of Geology and Geophysics, 1999, Vol. 42
Andrews, P. B.; Field, B. D.; Browne, G. H.; McLennan, J. M.1987: Lithostratigraphic nomenclature for the UpperCretaceous and Tertiary sequence of Central Canterbury,New Zealand. New Zealand Geological Survey Record 24.40 p.
Armentrout, J. M. 1996: High resolution sequence biostratigraphy:examples from the Gulf of Mexico Plio-Pleistocene. In:Howell, J. A.; Aitken, J. F. ed. High resolution sequencestratigraphy: innovations and applications. GeologicalSociety Special Publication 104: 65-86.
Be, A. W. H. 1977: An ecological, zoogeographic and taxonomicreview of Recent planktonic foraminifera. In: Ramsay,A. T. S. ed. Oceanic micropaleontology, Vol. 1. London,Academic Press. Pp. 1-100.
Berggren, W. A. 1992: Neogene planktonic foraminifermagnetobiostratigraphy of the southern Kerguelen Plateau(Sites 747, 748, and 751). In: Wise, S. W. Jr; Schlich, R.ed. Proceedings of the Ocean Drilling Program, ScientificResults 120: 631-647.
Berggren, W. A.; Kent, D. V.; Swisher, C. C ; Aubry, M-P. 1995: Arevised Cenozoic geochronology and chronostratigraphy.In: Berggren, W. A.; Kent, D. V.; Aubry, M-A.; Hardenbol,J. ed. Geochronology, time scales and global stratigraphiccorrelation. SEPM Special Publication 54: 129-212.
Beu, A. G.; Maxwell, P. A. 1990: Cenozoic Mollusca of NewZealand. New Zealand Geological Survey PaleontologicalBulletin 58. 518 p.
Buening, N.; Carlson, S. J.; Spero, H. J.; Lee, D. E. 1998: Evidencefor the Early Oligocene formation of a proto-SubtropicalConvergence from oxygen isotope records of New ZealandPaleogene brachiopods. Palaeogeography, Palaeo-climatology, Palaeoecology 138: 43-68.
Bukry, D. 1973: Low-latitude coccolith biostratigraphic zonation.Initial Reports of the Deep Sea Drilling Project 15:685-703.
Bukry, D. 1975: Coccolith and silicoflagellate stratigraphy,northwestern Pacific Ocean, Deep Sea Drilling Project Leg32. Initial Reports of the Deep Sea Drilling Project 32:677-701.
Cande, S. C ; Kent, D. V. 1995: Revised calibration of thegeomagnetic polarity timescale for the Late Cretaceous andCenozoic. Journal of Geophysical Research 100: 6093-6095.
Carter, R. M. 1988: Post-breakup stratigraphy of the KaikouraSynthem (Cretaceous-Cenozoic), continental margin,southeastern New Zealand. New Zealand Journal ofGeology and Geophysics 31: 405-429.
Carter, R. M.; Abbott, S. T; Fulthorpe, C. S.; Haywick, D. W.;Henderson, R. A. 1991: Application of global sea-level andsequence stratigraphic models in Southern HemisphereNeogene strata from New Zealand. Special Publicationsof the International Association of Sedimentologists 12:41-65.
Chaproniere, G. C. H. 1988: Globigerina woodi from the LateOligocene and Early Miocene of southeastern Australia.Journal of Foraminiferal Research 18 (2): 124-129.
Chaproniere, G. C. H. 1992: The distribution and development oflate Oligocene and early Miocene reticulate globigerinesin Australia. Marine Micropaleontology 18: 279-305.
Corfield, R. M.; Cartlidge, J. E. 1991: Isotopic evidence for thedepth stratification of the fossil and recent Globigerinina:a review. Historical Biology 5: 37-63.
Crundwell, M. P.; Scott, G. H.; Thrasher, G. P. 1994: Calibrationof paleobathymetry indicators by integrated seismic andpaleontological analysis of foreset sequences, TaranakiBasin, New Zealand. 1994 New Zealand PetroleumConference Proceedings. Wellington, Publicity Unit,Crown Minerals Operations Group, Energy and ResourcesDivision, Ministry of Commerce. Pp. 169-178.
Edwards,A. R. 1971: A calcareous nannoplankton zonation of theNew Zealand Paleogene. In: Farinacci, A. ed. Proceedingsof the Second Planktonic Conference, Rome 1970. RomeEdizioni Tecnoscienza. Pp. 381-419.
Edwards, A. R. 1975: Southwest Pacific Cenozoic paleogeograph)and an integrated Neogene paleocirculation model. InitialReports of the Deep Sea Drilling Project 30: 667-684.
Edwards, A. R. 1982: Calcareous nannofossils. In: Hoskins, R. Hed. Stages of the New Zealand marine Cenozoic: ;:.synopsis. New Zealand Geological Survey Report NZGS107. 75 p.
Edwards, A. R; Hornibrook, N. de B.; Raine, J. I; Scott, G. H.Stevens, G. R.; Strong, C. P.; Wilson, G. J. 1988: A NewZealand Cretaceous-Cenozoic time scale. New ZealandGeological Survey Record 35: 135-149.
Field, B. D.; Browne, G. H. 1986: Lithostratigraphy of Cretaceousand Tertiary rocks, southern Canterbury, New Zealand.New Zealand Geological Survey Record 14. 55 p.
Field, B. D.; Browne, G. H. and others 1989: Cretaceous andCenozoic sedimentary basins and geological evolution inthe Canterbury Region, South Island, New Zealand. NewZealand Geological Survey Basin Studies 2. 94 p.
Finlay, H. J. 1939: New Zealand Foraminifera: key species instratigraphy—1. Transactions of the Royal Society of NewZealand 68: 504-533.
Finlay, H. J.; Marwick, J. 1940: The divisions of the UpperCretaceous and Tertiary in New Zealand. Transactions ofthe Royal Society of New Zealand 70: 11-13 5.
Finlay, H. J.; Marwick, J. 1947: New divisions of the New ZealandUpper Cretaceous and Tertiary. New Zealand Journal ofScience and Technology B28: 228-236.
Fornaciari, E.; Rio, D. 1996: Latest Oligocene to early MiddleMiocene quantitative calcareous nannofossil bio-stratigraphy in the Mediterranean region. Micro-paleontology 42: 1-36.
Gage, M. 1957: The Geology of the Waitaki Subdivision. NewZealand Geological Survey Bulletin 55. 135 p.
Gair, H. S. 1959: The Tertiary geology of the Pareora District,South Canterbury. New Zealand Journal of Geology andGeophysics 2: 265-296.
Gasperi, J. T.; Kennett, J. P. 1992: Isotopic evidence for deptlstratification and paleoecology of Miocene planktonicforaminifera: western equatorial Pacific DSDP Site 289In: Tsuchi, R.; Ingle, J. C. Jr. ed. Pacific Neogene—Environment, evolution, and events. Tokyo, University ofTokyo Press. Pp. 117-147.
Gibson, T. G. 1989: Planktonic-benthonic foraminiferal ratiosmodern patterns and Tertiary applicability. MarineMicropaleontology 15: 29-52.
Graham, I. J.; Morgans, H. E. G.; Waghorn, D. B.; Trotter, J. A.:Whitford, D. J. in press: Sr isotope stratigraphy of theOligocene Otekaike Limestone (Trig Z section) in southernNew Zealand: age of the Duntroonian-Waitakian Stageboundary. New Zealand Journal of Geology and Geo-physics.
Gudex, M. C. 1914: List of fossil Mollusca from Bluecliffs, SouthCanterbury. Transactions of the New Zealand Institute 46:278.
Gudex, M. C. 1918: The succession of Tertiary beds in the Pareoradistrict, South Canterbury. Transactions of the NewZealand Institute 50: 244-262.
Hayward, B. W. 1986: A guide to paleoenvironmental assessmentusing New Zealand Cenozoic foraminiferal faunas. New-Zealand Geological Survey Report Pal 109. 13 p.
Hayward, B. W.; Brook, F. J. 1994: Forminiferal paleoecology andinitial subsidence of the early Miocene Waitemata Basin,Waiheke Island, Auckland. New Zealand Journal ofGeology and Geophysics 37: 11-24.

Morgans et al.—Waitakian-Otaian Stage boundary 613
Hayward, B. W.; Buzas, M. A. 1979: Taxonomy and paleoecologyof early Miocene benthic foraminifera of northern NewZealand and the north Tasman Sea. Smithsonian Contri-bution to Paleobiology 36: 154 p.
Heider, F.; Leitner, B.; Inokuchi, H. 1992: High southern latitudemagnetostratigraphy and rock magnetic properties ofsediments from sites 747, 749, and 751. Proceedings ofthe Ocean Drilling Program, Scientific Results 120:225-245.
Hemleben, C ; Spindler, M.; Anderson, O. R. 1989: Modernplanktonic foraminifera. New York, Spinger-Verlag. 363 p.
Hornibrook, N. de B.; Brazier, R. C ; Strong, C. P. 1989: Manualof New Zealand Permian to Pleistocene foraminiferalbiostratigraphy. New Zealand Geological Survey Paleon-tological Bulletin 56. 175 p.
Howarth, R. J.; McArthur, J. M. 1997: Statistics for strontiumisotope stratigraphy: a robust LOWESS fit to the marineSr-isotope curve for 0 to 206 Ma, with look-up table forderivation of numeric age. Journal of Geology 105:441^56.
Jenkins, D. G. 1960: Planktonic foraminifera from the LakesEntrance oil shaft, Victoria, Australia. Micropaleontology6: 345-371.
Jenkins, D. G. 1966: Planktonic foraminiferal zones and new taxafrom the Danian to Lower Miocene of New Zealand. NewZealand Journal of Geology and Geophysics 8: 1088-1126.
Jenkins, D. G. 1971: New Zealand Cenozoic planktonicforaminifera. New Zealand Geological Survey Paleon-tological Bulletin 42. 278 p.
Jenkins, D. G.; Srinivasan, M. S. 1986: Cenozoic planktonicforaminifers from the equator to the sub-Antarctic of theSouthwest Pacific. Initial Reports of the Deep Sea DrillingProject 90: 795-834.
Kadar, A. P. 1983: Late Eocene-Late Miocene calcareousnannofossils from the Kahuranaki-Elsthorpe-Kairakauarea, southern Hawke's Bay. Unpublished MSc thesis,lodged in the Library, University of Auckland, Auckland,New Zealand. 113 p.
Kamp, P. J. J. 1986: Late Cretaceous-Cenozoic tectonicdevelopment of the southwest Pacific region. Tectono-physics 121: 225-251.
Kennett, J. P.; Srinivasan, M. S. 1983: Neogene planktonicforaminifera. Stroudsburg, Hutchinson Ross PublishingCompany. 265 p.
Laws, C. R. 1934: New Tertiary Mollusca from Timaru District,South Canterbury, New Zealand. Transactions of the NewZealand Institute 63: 315-329.
Li, Q.; McGowran, B. 1994: Miocene upwelling events: neriticforaminiferal evidence from southern Australia. AustralianJournal of Earth Sciences 41: 593-603.
Li, Q.; McGowran, B. 1997: Miocene climatic oscillation recordedin the Lakes Entrance oil shaft, southern Australia: benthicforaminiferal response on a mid-latitude margin.Micropaleontology 43: 149-164.
Li, Q.; Radford, S. S.; Banner, F. T. 1992: Distribution ofmicroperforate tenuitellid planktonic foraminifers in holes747A and 749B, Kerguelen Plateau. Proceedings of theOcean Drilling Programme, Scientific Results 120:569-594.
Lohmann, W. H. 1986: Calcareous nannoplankton biostratigraphyof the southern Coral Sea, Tasman Sea, and southwesternPacific Ocean, Deep Sea Drilling Project Leg 90: Neogeneand Quaternary. Initial Reports of the Deep Sea DrillingProject 90: 763-793.
McFadden, P. L.; McElhinny, M. W. 1988: The combined analysisof remagnetisation circles and direct observations inpaleomagnetism. Earth and Planetary Science Letters 87:161-172.
McGowran, B.; Li, Q. 1993: Miocene planktonic foraminifera fromLakes Entrance in Gippsland: midlatitude neritic signalfrom a transforming ocean. Memoirs of the Association ofAustralasian Paleontologists 15: 395^05.
McGowran, B.; Li, Q. 1996: Ecostratigraphy and sequencebiostratigraphy, with a neritic foraminiferal example fromthe Miocene in southern Australia. Historical Biology 11:137-169.
Mancini, E. A.; Tew, B. H. 1997: Recognition of maximumflooding events in mixed siliciclastic-carbonate systems:key to global chronostratigraphic correlation. Geology 25:351-354.
Martini, E. 1971: Standard Tertiary and Quaternary calcareousnannoplankton zonation. In: Farinacci, A. ed. Proceedingsof the Second Planktonic Conference, Rome 1970. Rome,Edizioni Tecnoscienza. Pp. 739-785.
Martini, E.; Muller, C. 1986: Current Tertiary and Quaternarycalcareous nannoplankton stratigraphy and correlations.Newsletters on Stratigraphy 16: 99-112.
Marwick, J. 1971: New Zealand Turritellidae related to ZeacolpusFinlay (Gastropoda). New Zealand Geological SurveyPaleontological Bulletin 44. 87 p.
Maxwell, P. A. 1969: Middle Tertiary Mollusca from North Otagoand South Canterbury, New Zealand. Transactions of theRoyal Society of New Zealand, Geology 6: 155-185.
Maynard Smith, J. 1989: The causes of extinction. PhilosophicalTransactions of the Royal Socielv of London B325:241-252.
Morgans, Ff. E.G.; Scott, G. H.; Beu, A. G.; Graham, 1. J.; Mumme,T. C ; St George, W.; Strong, C. P. 1996: New ZealandCenozoic timescale (version 11/96). Institute of Geological& Nuclear Sciences Science Report 96/38. 12 p.
Mumme, T. C. 1997: Paleomagnetism of the Waitakian-Otaianboundary at Bluecliffs, Otaio River, South Canterbury, andthe Duntroonian-Waitakian boundary at Trig. Z, Otiake,North Otago. Institute of Geological & Nuclear SciencesScience Report 97/37. 21 p.
Okada, H.; Bukry, D. 1980: Supplementary modification andintroduction of code numbers to the low-latitude coccolithbiostratigraphic zonation (Bukry, 1973; 1975). MarineMicropaleontology 5: 321-325.
Park, J. 1918: The geology of the Oamaru District, North Otago(Eastern Otago Division). New Zealand Geological SurveyBulletin 20. 119 p.
Pocknall, D. T. 1982: Palynology of the Bluecliffs Siltstone (EarlyMiocene), Otaio River, South Canterbury, New Zealand.New Zealand Geological Survey Report PAL 55. 25 p.
Pocknall, D. T. 1988: Bluecliffs section. In: Pocknall, D. T.;Tremain, R. ed. Tour LB1, 7th International PalynologicalConference, Brisbane, Australia, August 1988. NewZealand Geological Survey Record 33: 88-90.
Pocknall, D. T; Mildenhall, D. C. 1984: Late Oligocene-EarlyMiocene spores and pollen from Southland, New Zealand.New Zealand Geological Survey Paleontological Bulletin51. 66 p.
Roberts, A. P.; Turner, G. M. 1993: Diagenetic formation offerrimagnetic iron sulphide minerals in rapidly depositedmarine sediments, South Island, New Zealand. Earth andPlanetary Sciences Letters 115: 257-273.
Roberts, A. P.; Turner, G. M.; Vella, P. P. 1994: Magneto-stratigraphic chronology of late Miocene to early Pliocenebiostratigraphic and oceanographic events in New Zealand.Geological Society of America Bulletin 106: 665-683.
Scott, G. H. 1958: Distribution of populations of fossilforaminifera. New Zealand Journal of Geology andGeophysics 1: 474-484.

614 New Zealand Journal of Geology and Geophysics, 1999, Vol. 42
Scott, G. H. 1965: Utility of Haeuslerella Parr (Foraminifera) inNew Zealand middle Tertiary stratigraphy. New ZealandGeological Survey Paleontological Bulletin 38. 48 p.
Scott, G. H. 1972: Revision of Hutchinsonian, Awamoan andAltonian Stages (Lower Miocene, New Zealand)—2. NewZealand Journal of Geology and Geophysics 15: 49-70.
Scott, G. H. 1973: Ehrenbergina (Foraminiferida): Variability andapplication to Lower Miocene biostratigraphy in NewZealand. New Zealand Journal of Geology and Geophysics16: 52-67.
Sen Gupta, B. K..; Machain-Castillo, M. L. 1993: Benthicforaminifera in oxygen-poor environments. MarineMicropaleontology 20: 183-201.
Shelley, J. M. 1974: A paleoenvironmental study of the definitionof the Otaian Stage at its type section, Bluecliffs, OtaioRiver, South Canterbury, New Zealand. Unpublished MScthesis, lodged in the Library, University of Canterbury,Christchurch, New Zealand. 89 p.
Steininger, F. F.; Aubry, M. P.; Berggren, W. A.; Biolzi, M.;Borsetti, A. M.; Cartlidge, J. E.; Cati, F.; Corfield, R.;Gelati, R.; Iaccarino, S.; Napoleone, C; Ottner, F.; Rogl,F.; Roetzel, S.; Spezzaferii, S.; Tateo, F.; Villa, G.;Zevenboom, D. 1997: The global stratotype section andpoint (GSSP) for the base of the Neogene. Episodes 20:23-28.
Suggate, R. P.; Stevens, G. R.; Te Punga, M. T. ed. 1978: Thegeology of New Zealand. Wellington, Government Printer.820 p.
Thomson, J. A. 1916: On stage names applicable to the divisionsof the Tertiary in New Zealand. Transactions of the NewZealand Institute 48: 28^0 .
Topping, R. M. 1978: Foraminifera from the Mahoenui GroupNorth Wanganui Basin. Unpublished PhD thesis, lodgedin the Library, University of Auckland, Auckland, NewZealand. 414 p.
Weaver, P. P. E.; Neil, H.; Carter, L. 1997: Sea surface temperatureestimates from the Southwest Pacific based on planktonicforaminifera and oxygen isotopes. Palaeogeography,Palaeoclimatology, Palaeoecology 131: 241-256.
Wei, W.; Wise, S. W. 1992: Oligocene-Pleistocene calcareousnannofossils from Southern Ocean sites 747, 748, and 75 :.Proceedings of the Ocean Drilling Programme, ScientificResults 120: 509-521.
Williams, G. L.; Stover, L. E.; Kidson, E. J. 1993: Morphologyand stratigraphic ranges of selected Mesozoic-Cenozoicdinoflagellate taxa in the Northern Hemisphere. GeologicalSurvey of Canada Paper 92-10. 137 p.
Wilson, G. J. 1984: New Zealand late Jurassic to Eocenedinoflagellate biostratigraphy—a summary. Newsletters onStratigraphy 13: 104-117.
Wilson, G. J. 1996: Preliminary dinoflagellate biostratigraphy ofthe Bluecliffs sequence (Bluecliffs Siltstone and CraigmoieLimestone), South Canterbury. Unpublished Institute ofGeological & Nuclear Sciences internal report GJW 29<V96. 2 p.
Wilson, G. S.; McGuire, D. M. 1995: Distributed deformation dueto coupling across a subduction thrust: mechanism ofyoung tectonic rotation within the south Wanganui Basin,New Zealand. Geology 23: 645-648.
Wilson, G. S.; Roberts, A. P. 1999: Impact of diagenesis onmagnetic mineral assemblages in multiply redepositedsiliciclastic marine sediments, Wanganui Basin, NewZealand. In: Tarling, D. H.; Turner, P. ed. Palaec-magnetism and diagenesis in sediments. GeologicalSociety of London Special Publication 151: 95-108.
Wright, I. C ; Vella, P. P. 1988: A New Zealand late Miocenemagnetostratigraphy: glacioeustatic and biostratigraphiccorrelations. Earth and Planetary Sciences Letters 80:348-352.
Wright, J. D.; Miller, K. G. 1992: Miocene stable isotopestratigraphy, Site 747, Kerguelen Plateau. Proceedings ofthe Ocean Drilling Program, Scientific Results 120:855-866.
Zachos, J. C ; Flower, B. P.; Paul, H. 1997: Orbitally paced climaticoscillations across the Oligocene/Miocene boundarv.Nature 388: 567-570.