isolation, purification, characterization and mass spectroscopic analysis of carbonic anhydrase from...
TRANSCRIPT

Journal of Environmental Chemical Engineering 2 (2014) 31–39
Isolation, purification, characterization and mass spectroscopic analysis ofcarbonic anhydrase from Serratia sp. for sequestration of carbondioxide and formation of calcite
Randhir K. Bharti, Shaili Srivastava, Indu Shekhar Thakur *
School of Environmental Sciences, Jawaharlal Nehru University, New Delhi 110067, India
A R T I C L E I N F O
Article history:
Received 2 September 2013
Accepted 22 November 2013
Keywords:
16S rRNA sequencing
CO2 sequestration
Calcite
Carbonic anhydrase
Carboxysomes
Serratia sp.
A B S T R A C T
Bacterial community isolated from marble rock was enriched in the chemostat by continuous culture in
minimal salt medium with different concentrations of sodium bicarbonate. One of the isolate survived at
100 mM sodium bicarbonate exhibited capability to sequester carbon dioxide (CO2) was identified as
Serratia sp. by 16S rRNA sequencing. CO2 sequestering capacity was detected by carbonic anhydrase
enzyme activity and formation of calcite. The carbonic anhydrase enzyme from Serratia sp. was detected
after isolation, partial purification by ammonium sulfate precipitation, gel filtration chromatography,
affinity chromatography, sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and
MALDI-TOF/MS. The purified enzyme molecular weight was 29 kDa on SDS-PAGE and gel filtration
chromatography. The optimum pH and temperature of the enzyme were found to be 7.5 and 40 8Crespectively. The Km of CA was 2.25 mM and the Vmax 940 mmol/min/mg. The Kcat and Kcat/Km value of
purified carbonic anhydrase was 2.8 � 105 s�1 and 1.2 � 108 M�1 s�1 respectively. MALDI-TOF mass
spectrometry data showing highest matched sequence in database was similar to carbonic anhydrase.
Scanning electron microscopy (SEM) analysis revealed the formation of calcium carbonate. Major calcite
phase diffraction peaks occurred at 2u = 29.48, 36.18, 39.58 and 43.58 determined by X-ray diffraction
(XRD) analysis. Energy-dispersive X-ray (EDX) analysis showed the presence of CaO (72%) and carbon
(18%) indicated role of carbon concentrating mechanisms of Serratia sp., a member of Enterobacteriaceae
isolated from marble mines.
� 2013 Elsevier Ltd. All rights reserved.
Contents lists available at ScienceDirect
Journal of Environmental Chemical Engineering
jou r n al h o mep ag e: w ww .e lsev ier . co m / loc ate / jec e
Introduction
Global warming gases (GWGs) are one of the most seriousenvironmental issues which made a question mark on existence ofthe earth. Carbon dioxide (CO2), one of the major GWGs,concentration was stable at about 270 ppm which has increasedapproximately 38% at 380 ppm after industrial revolution. It ispredicted that by the middle of this century the concentration ofCO2 will be reached to 600 ppm and by the end of the century it islikely to reach 700 ppm [1]. Increase in CO2 concentration may bemitigated by autotrophic and heterotrophic carbon fixation byplants and microorganisms [1]. One of the most effective methodsto mitigate the challenge of the rising levels of CO2 is bymicroorganism as some of them are capable of fixing atmosphericcarbon dioxide in to valuable products.
* Corresponding author. Tel.: +91 11 26704321x10/26191370;
fax: +91 011 26717586.
E-mail addresses: [email protected], [email protected],
[email protected] (I.S. Thakur).
2213-3437/$ – see front matter � 2013 Elsevier Ltd. All rights reserved.
http://dx.doi.org/10.1016/j.jece.2013.11.018
Carbon capture and storage (CCS) is an approach to mitigateglobal warming gases based on capturing CO2 from large sourcepoints. The most important global carbon sink is forest, soil andocean where photosynthetic carbon dioxide is fixed by autotrophicbiota, however, some bacteria like chemoautotrophs, obligateautotrophs, facultative autotrophs and chemolithoautotrophs canfix atmospheric CO2 through ribulose-1,5-bisphosphate carboxyl-ase/oxygenase (Rubisco) and other carboxylating enzymes [2].Photosynthetic microorganisms and some chemolithotrophs canacclimate to a wide range of CO2 concentration from as low as 0.001%to 10% CO2 (v/v) in the air in equilibrium with their environment [2].Therefore, the search of bacterial strains for efficient sequestration ofcarbon dioxide titillates the scientific mind.
Physicochemical fixation of CO2 is in the form of carbonatemineral such as calcite, magnesite and dolomite. Carbon dioxideand dissolved inorganic carbon (bicarbonate) are essentialcomponents in microorganisms. It is well established that anumber of plants, alga, cyanobacteria and chemolithotrophicbacteria actively take up and accumulate inorganic carbon (CO2)intracellularly which allows cells to photosynthesize efficiently

R.K. Bharti et al. / Journal of Environmental Chemical Engineering 2 (2014) 31–3932
even under atmospheric CO2. This CO2 acquisition mechanism istermed as a carbon concentrating mechanism (CCM) and it hasbeen suggested that carbonic anhydrase (CA) plays a key role in theCCM [3]. CA (EC 4.2.1.1) is a zinc-containing enzyme that catalyzesthe reversible dehydration of HCO3� to CO2. This reaction is knownto play important roles in various biological processes such as ionexchange, respiration, pH homeostasis, CO2 acquisition, andphotosynthesis [3]. The CA functions to convert an accumulatedcytosolic pool of HCO3� into CO2 within the carboxysome. Thegeneration of CO2 coupled with a diffusive restriction to the effluxfrom the carboxysome, possibly imposed by the protein shell, leadsto the localized elevation of CO2 around the active site of Rubiscowithin the carboxysome. The substrate for the carboxysome,HCO3�, is accumulated in the cytosol by the operation of a numberof active CO2 and HCO3� transporters. These transporters arelocated on plasma membrane and exist in both low affinity andhigh affinity transporter forms [3,4]. The use of biotechnologicalapproach for the sequestration of carbon dioxide is the emergingtechnology and environmentally benign. Microorganisms can bestimulated by continuous enrichment in presence of CO2 or HCO3�
to acquire proteins and genes for sequestration processes andformation of calcite and other valuable products.
There are three evolutionary distinct groups of carbonicanhydrases, alpha, beta and gamma, which show no significantsequence identity or structural similarity. Some of the prokaryoteshave multiple carbonic anhydrase genes which suggest theimportance of carbonic anhydrase enzyme in prokaryotic physiol-ogy [5]. Some of bacterial strains have been shown to carbonicanhydrase activity including strains of Citrobacter freundii, Bacillus
subtilis and Cynobacteria like Synechocystis [6–8]. But there are noreports about carbonic anhydrase of Enterobacteriaceae, Serratia
sp., for CO2 sequestration processes. Many of the blue-green algaeand several chemolithotrophic bacteria contain polyhedral inclu-sion bodies [8]. Basically carboxysome is a polyhedral bacterialmicro-compartment that enhances carbon fixation by encapsulat-ing the Rubisco and CA enzymes. Carboxysomes carry out the finalstages of the carbon-concentrating mechanism (CCM) throughwhich autotrophic prokaryotes accumulate inorganic carbon toenhance CO2 fixation [9]. In the present study, we enriched anindigenous microbial population of marble rock to developchemolithotrophic bacteria by continuous enrichment in thechemostat in different concentrations of sodium bicarbonate ascarbon source, assayed enzymatic activity of carbonic anhydraseand mechanisms of bacterial CO2 concentrating mechanisms byisolation, purification, characterization of carbonic anhydrasewhich facilitate CO2 sequestration by enzyme Rubisco andformation of calcite.
Materials and methods
Sampling site and microorganism
Samples were collected from marble rock of the palaeoproter-ozoic metasediments of the Aravalli Supergroup in the Umra area,Rajasthan, India, for isolation of microbes. The upper portion of therock was scrapped and dissolved in autoclaved distilled water(1:10, w/v) which served as inoculums for enrichment of bacteriain the chemostat by continuous culture.
Enrichment and culture condition of bacteria
A chemostat culture was set up in a 2-L glass vessel, effectivevolume 1 L, with culture condition as stirring at 150 rpm,temperature at 30 8C, and pH 7.6 in the minimal salt medium(MSM). The composition of MSM (g/L) was: Na2HPO4, 7.8; KH2PO4,6.8; MgSO4, 0.2 g; NaNO3, 0.085; ZnSO4�7H2O, 0.05; ZnCl2, 0.02;
Ca(NO3)2�4H2O, 0.05 [10]. The supernatant containing microor-ganisms (50 mL) obtained from marble rock was served asinoculums in the chemostat. The chemostat culture was run inpresence of sodium bicarbonate (5 mM) initially, which increasedto 10, 20, 50, 100 and 150 mM and growth of microorganisms wasdetermined. After stabilization of growth of bacteria, theconcentration of sodium bicarbonate was increased in thechemostat. Bacterial community from chemostat enriched inpresence of sodium bicarbonate (5, 10, 20, 50, 100 and 150 mM)was inoculated in MSM in flasks for further enrichment. Theculture medium was removed after 12, 24, 36, 48, 60 and 72 h fordetermining the growth pattern and carbonic anhydrase activity.The growth patterns of bacterial strains were taken at O.D. at595 nm with a spectrophotometer (Cary, 100 Bio, Varian Co,Australia). One of the bacterial strains survived in the sodiumbicarbonate (100 mM) without losing its ability of production ofcarbonic anhydrase was cultured on LB-agar plate and selected forfurther study.
Identification of bacteria by 16S rRNA method
Genomic DNA from the bacterial strain was isolated with theGenome DNA Kit (Qiagen Inc., USA) as described by themanufacturer. The 16S rRNA gene was amplified from genomicDNA by using PCR with universal primers 50-GAGAGTTT-GATCCTGGCTCAG-30 (forward) and 50-CTACGGCTACCTTGT-TACGA-30 (reverse) [11]. PCR amplification was performed usingprimer with a DNA thermal cycler under the following conditions:10–20 ng template DNA, 5 mL 10� reaction buffer, 2.5 U Taq DNApolymerase, 1 mM forward primer, 1 mM reverse primer, 200 mMof each dNTP and H2O combined in a total volume of 50 mL. Thetubes were incubated at 94 8C for 5 min and then subjected to thefollowing thermal cycling program: denaturation at 94 8C for2 min, primer annealing at 55 8C for 2 min, and chain extension at72 8C for 2 min with an additional extension time of 10 min on thefinal cycle, for a total of 30 cycles. The amplified DNA was purifiedusing Qiaquick PCR Purification Kit (Qiagen Inc., USA), adjusted to200 ng/mL and cloned in the pDrive (Qiagen Inc., USA) and sent tothe M/S Bangalore Genei, Bangalore, India, for sequencing.Sequenced data was compared and analyzed with the existingdatabase of GenBank, National Center for Biotechnology Informa-tion. A phylogenetic tree was drawn on the basis of the sequences.A bootstrap consensus tree (1000 copies) was drawn by multiplesequence alignment with Neighbor-Joining method using softwareMega, version 3.1 with different species of bacteria [11].
Carbonic anhydrase enzyme assay
Culture medium was centrifuged at 7500 rpm for 5 min, cellpellet was washed, suspended in sonication buffer (10 mL)containing Tris–HCl (50 mM, pH 6.5) buffer and lysozyme (0.2%)and sonicated for 10 min. After centrifugation at 12,000 rpm, thepellet was discarded and supernatant containing enzyme wasstored. The protein concentration was determined by Bradfordmethod with bovine serum albumin as the standard. Carbonicanhydrase activity was assayed by the changing pH/dye indicatormethod of Wilbur–Anderson method [12]. The buffer/indicatorpairs used were 25 mM HEPES, 100 mM phenol red (pH 7.5,lmax = 558 nm) and 25 mM TAPS, 100 mM m-cresol purple (pH 8.4,lmax = 578 nm), and both solutions also contained 100 mM sodiumsulfate to maintain ionic strength of the reaction medium. Thereaction was initiated by addition of 0.5 ml CO2 (aq) to 0.5 ml of a2� buffer indicator solution containing enzyme samples. Theenzyme activity was determined by monitoring the time requiredfor the pH change of the reaction solution. The activity wascalculated from the formula given below. The time (tc) interval was

R.K. Bharti et al. / Journal of Environmental Chemical Engineering 2 (2014) 31–39 33
determined between the addition of CO2 solution the pH change ofthe reaction solution. In the same way, the time interval wasrecorded without an enzyme solution (to).
1 Wilber�Andersont unit ¼ to � tc
tc:
The carbonic anhydrase activity was also measured in aqueousphase by using nitrophenyl esters [13]. The enzymatic reactioncontained Tris–HCl buffer (50 mM, pH 7.5), 3 mM p-nitrophenylacetate (p-NPA), enzyme preparations (100 mL). One unit ofenzyme activity was expressed as 1 mmol of p-nitrophenyl acetatehydrolyzed per minute.
Production and partial purification of carbonic anhydrase
Enzyme present in cell extract was precipitated by ammoniumsulfate (40–75%) and the mixture was stirred for 2 h, andcentrifuged at 15,000 � g for 30 min. The ammonium sulphatefraction was dialyzed against Tris–HCl buffer (50 mM, pH 6.5) andapplied to Sephadex G-100 gel filtration column. Fractions werecollected and protein and carbonic anhydrase activity wasdetermined. Partially purified enzyme of gel filtration was appliedto the p-aminobenzenesulfonamide affinity column equilibratedwith 25 mM Tris–HCl/0.1 M Na2SO4 (pH 7.0) [14]. The affinity gelwas washed with 25 mM Tris–HCl/22 mM Na2SO4 (pH 7.0), and CAisozymes were eluted under different elution conditions [14].Fractions were collected and the absorbance of fractions wasmeasured at 280 nm. The carbonic anhydrase activity wasmeasured. Selected fractions which gave carbonic anhydraseenzyme activity were analyzed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (12%) [15]. Elution volumesof proteins of known molecular mass were measured and used asreference standards in comparison to carbonic anhydrase formolecular mass determination.
MALDI-TOF mass spectrometry and analysis of peptide sequences
Protein band visualized after Coomassie Brilliant Blue R250staining in SDS–PAGE gel was subjected to MALDI-TOF/MS analysis(Brucker Daltonik GmbH) [15]. Mass spectral data were analyzedby using data explorer software (Applied Biosystems, Foster City,CA). Acquired data were analyzed by comparison to in silicoinformation contained in the NCBI databases using peptide massfingerprinting (PMF). The 300 most intense peaks were searchedagainst the NCBI taxonomy subset ‘‘all bacteria’’ (>753,000sequences) at a mass tolerance of 50–100 ppm using mascot(http://www.matrixscience.com). Presence of the carbonic anhy-drase was confirmed by submission of detected fragment ions tothe NCBI database.
Characterization of carbonic anhydrase
Carbonic anhydrase enzyme activity was measured at 30 8C andat different pH of 50 mM acetate buffer (pH 3.0, 4.0 and 5.0),sodium phosphate (pH 6.0–7.0) and sodium carbonate (pH 8.0, 9.0and 10.0). The enzyme sample was incubated in different buffers(pH 3.0–10.0) at 30 8C for 2 h and the residual activity wasmeasured to check the pH stability. Effect of temperature wasstudied by measuring enzyme activity between 30 and 60 8C and atpH 7.5 for 30 min. It was then cooled in ice bath and the residualenzyme activity was measured. Activity of the carbonic anhydrasewithout pretreatment was considered as 100%.
The purified fractions having carbonic anhydrase activity wereused for enzyme kinetic studies. Enzyme activity at differentsubstrate concentrations (1.0–16.0 mM) was determined at 37 8C
by measuring absorbance at 348 nm. The Km, Vmax, Kcat and Kcat/Km
values were calculated at pH 7.6. The specific activity of enzymesCA was tested with increasing concentration of substrate. Thekinetic parameters Km and Vmax were estimated from MichaelisMenton equation. The data were fit in curve of the singlerectangular hyperbola (equation y = ax/(b + x)). From this the Vmax
and Km value has been derived. The rate of reaction was expressedas the number of mmoles of substrate reduced per min per mg oftotal protein in the enzyme sample. One unit specific activity isdefined as 1 mmol substrate conversion per milligram per minute.
Production and analysis of carbonate crystals
The precipitation of CO2 in the form of CaCO3 was performed ina 50 mL total reaction mixture containing 200 mM Tris buffer pH7.5, 25 mM CaCl2 solution, and 3 U/mL enzyme. A samplecontaining 1 mM of sulfonamide and 1 mM of EDTA was alsoadded to the reaction mixture to test the specific inhibition of CA.All the reaction mixture was performed in CO2 incubator at 30 8Cwith 5% CO2 and 120 rpm rotation. The precipitate formed afterdefinite time interval was filtered and dried in oven at 40 8C. Afterweighing, the precipitated sample was used to determine theamount of carbonate deposited due to enzymatic action andprecipitation of CaCO3 against blank. After 24 h reaction, pre-cipitates was filtered and dried in oven at 40 8C for further analysis.Scanning electron microscope (SEM) was performed to detectcalcite formation by bacterium [16]. In this case cells were fixed inglutaraldehyde (1% solution) and paraformaldehyde (2%) bufferedwith sodium phosphate buffer saline (0.1 M, pH 6.8). Fixation wasfor 12–18 h at 40 8C temperature, after cells were washed in freshbuffer, and post fixed for 2 h in osmium tetraoxide (1%) in the samebuffer at 40 8C. The bacterial cells fixed were smeared with poly-L-lysin for 30 min in wet condition. The specimen was washed withphosphate buffer, and then dehydrated in a series of ethanol–watersolution (30, 50, 70 and 90% ethanol, 5 min each), and dried under aCO2 atmosphere for 20 min. Mounting was done on aluminumstubs, and cells were coated with 90 A thick gold–palladiumcoating in polaron Sc 7640 sputter coater (Carl Zeiss, Germany) for30 min. Coated cells were viewed at 15 kV with scanning electronmicroscopy (Leo Electron Microscopy Ltd., Cambridge). Dx4 PrimeEnergy Dispersive X-ray spectrometer (Bruker, Germany, X-flashdetector) was performed at 20 kV for confirming the calciumcarbonate crystals in the bacterial cell. X-ray absorption spectros-copy provides information on the electronic and structural state ofan element [16]. In X-ray diffraction analysis, the dry cell powderform and extracts were prepared [16]. For diffraction analysis, X-pertsystem, PANalytical equipped with ‘X’ pert Pro software havingCu (K-a) radiations (45 kV, 40 mA) with scan rate of 18 2u/min wasused. Diffractogram was taken for each sample that gave a specificpick list in diffractogram. ‘X’ pert Pro software was used foranalysis of diffractogram which suggested the presence ofcompounds in samples. The analysis of calcium carbonate by X-ray diffraction method was performed to define its crystalstructure [16].
Results and discussion
Enrichment, isolation and characterization of bacterium
Microbial population of marble rock was enriched in MSM andsodium bicarbonate (5, 10, 20, and 50, 100 and 150 mM) in thechemostat by continuous enrichment. Result of the study indicatedmicrobial growth and enzyme production was higher in presenceof increasing concentration of sodium bicarbonate which reachedto maximum at 50 mM and after that it was decreased (Fig. 1). Eachbacterial community enriched in sodium bicarbonate was
Time (hour )
100806040200
S
peci
fic a
ctiv
ity o
f enz
yme
carb
onic
anh
ydra
se (µ
mol
e PN
PA /m
in/m
g pr
otei
n)
10
15
20
25
30
35
40
O.D
at 5
95 n
m
(gr
owth
of b
acte
ria)
0.1
0.2
0.3
0.4
0.5
0.6
0.7
spe cific activitygrowth of ba cteria
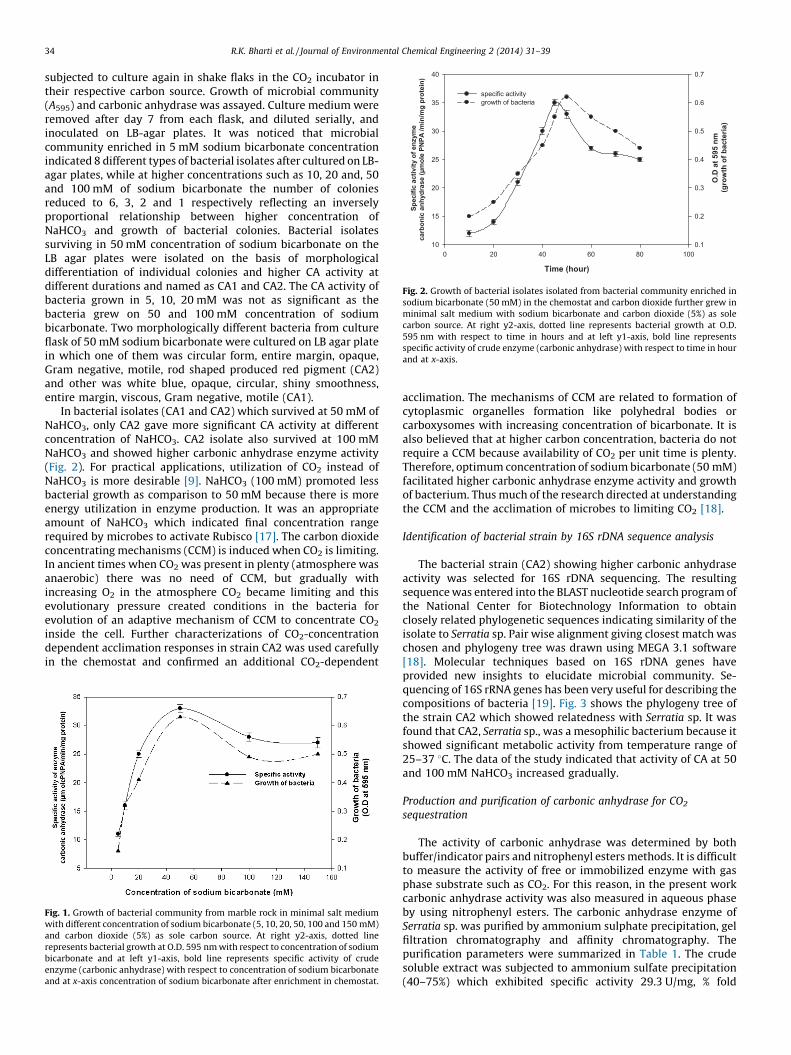
Fig. 2. Growth of bacterial isolates isolated from bacterial community enriched in
sodium bicarbonate (50 mM) in the chemostat and carbon dioxide further grew in
minimal salt medium with sodium bicarbonate and carbon dioxide (5%) as sole
carbon source. At right y2-axis, dotted line represents bacterial growth at O.D.
595 nm with respect to time in hours and at left y1-axis, bold line represents
specific activity of crude enzyme (carbonic anhydrase) with respect to time in hour
and at x-axis.
R.K. Bharti et al. / Journal of Environmental Chemical Engineering 2 (2014) 31–3934
subjected to culture again in shake flaks in the CO2 incubator intheir respective carbon source. Growth of microbial community(A595) and carbonic anhydrase was assayed. Culture medium wereremoved after day 7 from each flask, and diluted serially, andinoculated on LB-agar plates. It was noticed that microbialcommunity enriched in 5 mM sodium bicarbonate concentrationindicated 8 different types of bacterial isolates after cultured on LB-agar plates, while at higher concentrations such as 10, 20 and, 50and 100 mM of sodium bicarbonate the number of coloniesreduced to 6, 3, 2 and 1 respectively reflecting an inverselyproportional relationship between higher concentration ofNaHCO3 and growth of bacterial colonies. Bacterial isolatessurviving in 50 mM concentration of sodium bicarbonate on theLB agar plates were isolated on the basis of morphologicaldifferentiation of individual colonies and higher CA activity atdifferent durations and named as CA1 and CA2. The CA activity ofbacteria grown in 5, 10, 20 mM was not as significant as thebacteria grew on 50 and 100 mM concentration of sodiumbicarbonate. Two morphologically different bacteria from cultureflask of 50 mM sodium bicarbonate were cultured on LB agar platein which one of them was circular form, entire margin, opaque,Gram negative, motile, rod shaped produced red pigment (CA2)and other was white blue, opaque, circular, shiny smoothness,entire margin, viscous, Gram negative, motile (CA1).
In bacterial isolates (CA1 and CA2) which survived at 50 mM ofNaHCO3, only CA2 gave more significant CA activity at differentconcentration of NaHCO3. CA2 isolate also survived at 100 mMNaHCO3 and showed higher carbonic anhydrase enzyme activity(Fig. 2). For practical applications, utilization of CO2 instead ofNaHCO3 is more desirable [9]. NaHCO3 (100 mM) promoted lessbacterial growth as comparison to 50 mM because there is moreenergy utilization in enzyme production. It was an appropriateamount of NaHCO3 which indicated final concentration rangerequired by microbes to activate Rubisco [17]. The carbon dioxideconcentrating mechanisms (CCM) is induced when CO2 is limiting.In ancient times when CO2 was present in plenty (atmosphere wasanaerobic) there was no need of CCM, but gradually withincreasing O2 in the atmosphere CO2 became limiting and thisevolutionary pressure created conditions in the bacteria forevolution of an adaptive mechanism of CCM to concentrate CO2
inside the cell. Further characterizations of CO2-concentrationdependent acclimation responses in strain CA2 was used carefullyin the chemostat and confirmed an additional CO2-dependent
Fig. 1. Growth of bacterial community from marble rock in minimal salt medium
with different concentration of sodium bicarbonate (5, 10, 20, 50, 100 and 150 mM)
and carbon dioxide (5%) as sole carbon source. At right y2-axis, dotted line
represents bacterial growth at O.D. 595 nm with respect to concentration of sodium
bicarbonate and at left y1-axis, bold line represents specific activity of crude
enzyme (carbonic anhydrase) with respect to concentration of sodium bicarbonate
and at x-axis concentration of sodium bicarbonate after enrichment in chemostat.
acclimation. The mechanisms of CCM are related to formation ofcytoplasmic organelles formation like polyhedral bodies orcarboxysomes with increasing concentration of bicarbonate. It isalso believed that at higher carbon concentration, bacteria do notrequire a CCM because availability of CO2 per unit time is plenty.Therefore, optimum concentration of sodium bicarbonate (50 mM)facilitated higher carbonic anhydrase enzyme activity and growthof bacterium. Thus much of the research directed at understandingthe CCM and the acclimation of microbes to limiting CO2 [18].
Identification of bacterial strain by 16S rDNA sequence analysis
The bacterial strain (CA2) showing higher carbonic anhydraseactivity was selected for 16S rDNA sequencing. The resultingsequence was entered into the BLAST nucleotide search program ofthe National Center for Biotechnology Information to obtainclosely related phylogenetic sequences indicating similarity of theisolate to Serratia sp. Pair wise alignment giving closest match waschosen and phylogeny tree was drawn using MEGA 3.1 software[18]. Molecular techniques based on 16S rDNA genes haveprovided new insights to elucidate microbial community. Se-quencing of 16S rRNA genes has been very useful for describing thecompositions of bacteria [19]. Fig. 3 shows the phylogeny tree ofthe strain CA2 which showed relatedness with Serratia sp. It wasfound that CA2, Serratia sp., was a mesophilic bacterium because itshowed significant metabolic activity from temperature range of25–37 8C. The data of the study indicated that activity of CA at 50and 100 mM NaHCO3 increased gradually.
Production and purification of carbonic anhydrase for CO2
sequestration
The activity of carbonic anhydrase was determined by bothbuffer/indicator pairs and nitrophenyl esters methods. It is difficultto measure the activity of free or immobilized enzyme with gasphase substrate such as CO2. For this reason, in the present workcarbonic anhydrase activity was also measured in aqueous phaseby using nitrophenyl esters. The carbonic anhydrase enzyme ofSerratia sp. was purified by ammonium sulphate precipitation, gelfiltration chromatography and affinity chromatography. Thepurification parameters were summarized in Table 1. The crudesoluble extract was subjected to ammonium sulfate precipitation(40–75%) which exhibited specific activity 29.3 U/mg, % fold

Fig. 3. 16S rDNA analysis for identification and phylogenetic placement of CA2, Serratia sp. IST D104, isolated from marble rock and enriched in sodium bicarbonate in the
chemostat and carbon dioxide (5%).
R.K. Bharti et al. / Journal of Environmental Chemical Engineering 2 (2014) 31–39 35
purification 2.25 and yield 80.71. Partially purified protein wasfractionated by Sephadex G-100 gel filtration to remove otherproteins based on the basis of molecular size. Four fractions wereobtained in which one of the fraction, molecular weight 30 kDa,determined by known molecular weight proteins (phosphorylaseb, 98 kDa; bovine serum albumin, 67 kDa; ovalbumin, 44 kDa;glutathione S-transferase, 29 kDa; and Lysozyme, 16 kDa) hadactivity of carbonic anhydrase. Results indicated specific enzymeactivity 81.2 U/mg, % fold purification and yield was 6.24 and 61.19respectively. Partially purified enzyme of Sephadex G-100 wasfurther purified by affinity column chromatography. Resultsindicated specific enzyme activity 864.3 U/mg, % fold purificationand yield was 66.48 and 66.70 respectively. Fractions werecollected and protein was determined at 280 nm. Enzyme activitywas determined which was concentrated and subjected to SDS-PAGE. Result of the study indicated molecular mass of the purifiedenzyme from Serratia sp. 29.8 kDa on SDS-polyacrylamide gelelectrophoresis which was comparable to gel filtration chroma-tography (Fig. 4). Data of the study indicated single protein band ofenzyme may be monomeric indicated by SDS-PAGE. Carbonic
Table 1Purification and characterization of carbonic anhydrase in different stages of purificatio
from marble rock.
Purification stage Total protein (mg) Total ac
Crude 57.8 751.4
Ammonium sulphate precipitation (40–75%) 20.7 606.51
Sephadex G-100 6.4 519.68
Affinity chromatography 0.58 501.2
anhydrase has been isolated and purified from both prokaryoticand eukaryotic system which is comparable to our study [7].
MALDI-TOF analysis
The protein band of a 29 kDa excised from SDS–PAGE gelsurface subjected to MALDI-TOF mass spectrum of protein that wasidentified using the MASCOT search engine to query the NCBIprotein database. MALDI-TOF analysis indicated that enzymeobtained from Serratia sp. had highest matched sequence withcarbonic anhydrase which had been shown of mascot score. Thesequence coverage of the peptide against the Serratia sp. enzymewas 29.6%. There were seven significant spectra obtained thatcorrespond to peptides found to matched by peptide massfingerprinting (PMF). In this case, six peptides of Serratia sp. werefound to correspond exactly to internal sequence of carbonicanhydrase enzyme. The nominal mass of the homologous proteinof Serratia sp. was 31,324 Da. The molecular weights of the protein,determined by gel filtration chromatography, SDS-PAGE andMALDI-TOF/MS were almost similar. The molecular masses of
n processes from cell extract of chemolithotrophic bacterium, Serratia sp., isolated
tivity (U) Specific activity (U/mg) Yield (%) Purification fold
13 100 1
29.3 80.71 2.25
81.2 69.16 6.24
864.3 66.70 66.48

Fig. 4. SDS-polyacrylamide gel electrophoresis of carbonic anhydrase of Serratia sp.
In figure, lane M is marker, lane 1 is of crude protein of bacterium, lane 2 is
precipitated protein after ammonium sulphate precipitation, and lane 3 is protein
obtained after affinity chromatography.
R.K. Bharti et al. / Journal of Environmental Chemical Engineering 2 (2014) 31–3936
carbonic anhydrase isolated from various sources differ widely anda number of enzymes of similar molecular weight have beenreported from a variety of sources [20,21]. The isoelectric point andN-terminal sequences of carbonic anhydrase are also reported [6].Result of the MALDI-TOF/MS peptides data study indicatedsimilarity with bacterium isolated from marine sources. Therefore,present Serratia sp. isolated from marble rock and enriched inchemostat may be significant strain for evaluation of efficientsequestration of carbon dioxide concentrating mechanisms.
Characterization of carbonic anhydrase enzyme
The optimal temperature and pH were determined on the basisof p-nitrophenyl acetate as substrate in presence and absence ofthe enzyme. Carbonic anhydrase showed strong dependence onsubstrate concentration. The enzyme was subjected to treat atvarious temperatures and pH ranging from 30 to 60 8C and pHranges from 3 to 10. The optimum activity was detected at slightlybasic pH 7.5, and enzyme activity was too weak at lower pH 3 to 6and more alkaline range i.e. 8–10 range (Fig. 5a). The optimumtemperature for carbonic anhydrase enzyme was 40 8C andenzyme activity was decreased after 45 8C (Fig. 5b). The enzymewas not stable beyond 50 8C. While enzyme activity was increasingmoderately temperature ranges from 30 to 40 8C (Fig. 5b). Enzymekinetics measured Km and Vmax from Lineweaver–Burk plots of CAactivity at optimal pH and temperature using various concentra-tions of p-NPA substrate. The Km of CA was 2.3 mM and the940 mmol/min/mg (Fig. 5c). The line of regression graph has shownstrong positive linear correlation, r close to +1. The value ofcorrelation coefficient (r) is 0.88 (Fig. 5d). Carbonic anhydrase wasalso assayed in the presence of different metals. The increased CAactivity was detected in the presence of Zn2+ followed by Co2+, Fe3+,Mg2+ and Cu2+ (Fig. 5e). The enzyme that catalyzes the CO2 fixationreaction implicated as the limiting step in CO2 fixation because ofits poor turnover rate and the competition between O2 and CO2 atthe active site [22].
Higher activity of carbonic anhydrase in experiment revealedmore efficient bacteria for CO2 sequestration because carbonicanhydrase is supporting enzyme for the activity of Rubisco. The
enzyme is thought to dehydrate abundant cytosolic bicarbonateand provide ribulose 1,5-bisphosphate carboxylase/oxygenase(Rubisco) sequestered within the carboxysome with sufficientlyhigh concentrations of its substrate CO2 to permit its efficientfixation to ribulose 1,5-bisphosphate and hence availability of CO2
per minute toward the active site of the Rubisco may enhance andin this way fixation of CO2 would be enhanced [23]. Somechemolithotrophic bacteria and most of cyanobacteria have properchannels for transport of dissolved inorganic carbon (CO2, HCO3�,and CO3
2�). By enhancing the bicarbonate ion concentration orgaseous CO2 concentration in both the cases bacteria accumulateHCO3� in cytoplasm and its trigger higher CA activity andconversion of HCO3� to CO2. In this way bacteria acquainted tosequester more CO2. So activity of carbonic anhydrase is importantfactor for CO2 sequestration. Over expression of CA by strain CA2reported in various steps reveals more suitability for CO2
sequestration. Carbonic anhydrase (CA) has been associated infixation of dissolved inorganic carbon reported from hydrothermalvent habitat and other ecosystem [6,23]. Five different classes ofcarbonic anhydrase identified so far in which three are isolated andcharacterized from bacterial system [22–27]. Genes encodinghomologous of all three of these classes of carbonic anhydrases arereported from Thiomicrospira crunogena genome, Proteobacteria
and chemolithotrophic bacteria [28]. But we have isolated noblebacteria from marble rock and characterized carbonic anhydraseso that it can be applied for sequestration of carbon dioxide andevaluation of proteomic and genomics which is underway in thislaboratory.
Formation and determination of calcite
The mineralization process started using a concentration of25 mM CaCl2 after addition of the carbonic anhydrase (3 U/mL)which reached to maximum at 24 h, pH 7.5 and 25 8C. The additionof CA enzyme to the reaction mixture containing carbon dioxidesaturated calcium chloride solution resulted in the enhanceddeposition of calcium carbonate salts. The result of the studyindicated formation of 187 mg of calcium carbonate by purified CAenzyme, however, the carbonate deposition from crude enzymewas almost fifteen times lesser (Fig. 6). Addition of sulfonamide inone of the reaction mixture resulted in the inhibition of the purifiedenzyme and hence less deposition of carbonate salts. However,EDTA completely inhibited CA activity resulting in too littlecarbonate deposition. This study confirmed that carbon dioxidecould be sequestered using carbonic anhydrase enzyme asreported earlier [29–31].
To determine the presence of microbial calcite precipitation,the pure bacterial culture was examined under SEM analysis. TheSEM analysis has shown distinct calcite crystals as shown in Fig. 7aand b. This indicated that the bacteria served as the nucleationsites for the mineralization process. A hexagonal structure wasobtained with calcite phases. The calcite crystals displayed welldefined faceted rhombohedral characteristics in the study similarto earlier report [32]. EDX result verified that a precipitateproduced by the bacterium was composed of Ca, C, and O andother atomic percent ratios indicated the presence of CaCO3
(Fig. 7c). Major element present in EDX spectrum showed thepresence of CaO (72%) and carbon (18%). Calcite was the mainpolymorph precipitated by the bacterium (Fig. 7c). XRD analysis ofthe crystals precipitated by bacterium Serratia sp. ISTD04 duringthe 24 h of incubation identified in the form of crystals asrepresenting as calcite in the spectrum (Fig. 7d). The production ofEPS, cell aggregation, and Ca2+-aggregation were known to beimportant factors in the precipitation process [33]. The produc-tion of specific bacterial outer structures and their chemicalnature might also be crucial for the bacterial crystallization
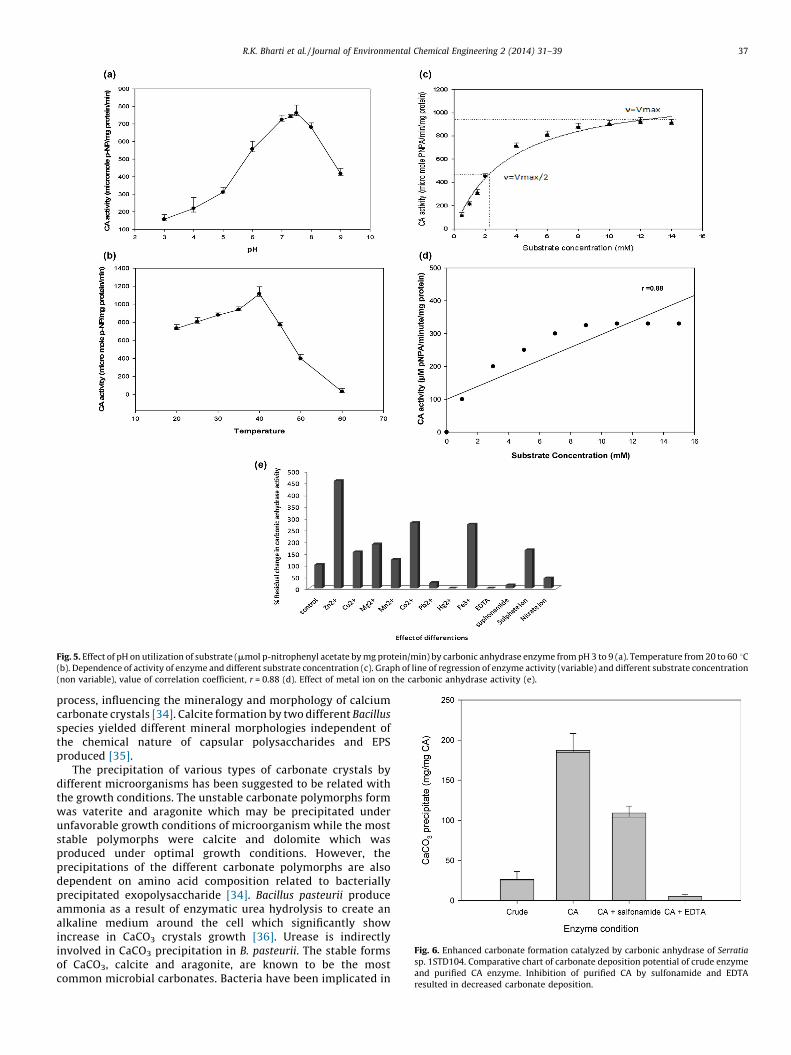
Fig. 5. Effect of pH on utilization of substrate (mmol p-nitrophenyl acetate by mg protein/min) by carbonic anhydrase enzyme from pH 3 to 9 (a). Temperature from 20 to 60 8C(b). Dependence of activity of enzyme and different substrate concentration (c). Graph of line of regression of enzyme activity (variable) and different substrate concentration
(non variable), value of correlation coefficient, r = 0.88 (d). Effect of metal ion on the carbonic anhydrase activity (e).
Fig. 6. Enhanced carbonate formation catalyzed by carbonic anhydrase of Serratia
sp. 1STD104. Comparative chart of carbonate deposition potential of crude enzyme
and purified CA enzyme. Inhibition of purified CA by sulfonamide and EDTA
resulted in decreased carbonate deposition.
R.K. Bharti et al. / Journal of Environmental Chemical Engineering 2 (2014) 31–39 37
process, influencing the mineralogy and morphology of calciumcarbonate crystals [34]. Calcite formation by two different Bacillus
species yielded different mineral morphologies independent ofthe chemical nature of capsular polysaccharides and EPSproduced [35].
The precipitation of various types of carbonate crystals bydifferent microorganisms has been suggested to be related withthe growth conditions. The unstable carbonate polymorphs formwas vaterite and aragonite which may be precipitated underunfavorable growth conditions of microorganism while the moststable polymorphs were calcite and dolomite which wasproduced under optimal growth conditions. However, theprecipitations of the different carbonate polymorphs are alsodependent on amino acid composition related to bacteriallyprecipitated exopolysaccharide [34]. Bacillus pasteurii produceammonia as a result of enzymatic urea hydrolysis to create analkaline medium around the cell which significantly showincrease in CaCO3 crystals growth [36]. Urease is indirectlyinvolved in CaCO3 precipitation in B. pasteurii. The stable formsof CaCO3, calcite and aragonite, are known to be the mostcommon microbial carbonates. Bacteria have been implicated in
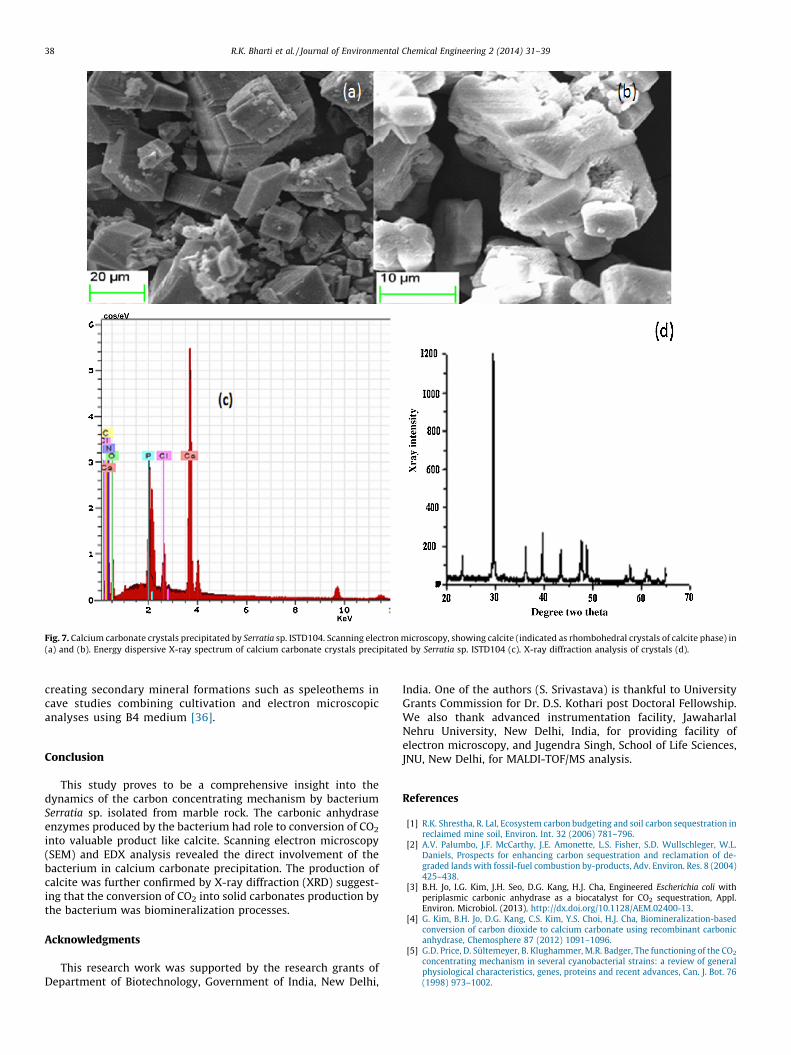
Fig. 7. Calcium carbonate crystals precipitated by Serratia sp. ISTD104. Scanning electron microscopy, showing calcite (indicated as rhombohedral crystals of calcite phase) in
(a) and (b). Energy dispersive X-ray spectrum of calcium carbonate crystals precipitated by Serratia sp. ISTD104 (c). X-ray diffraction analysis of crystals (d).
R.K. Bharti et al. / Journal of Environmental Chemical Engineering 2 (2014) 31–3938
creating secondary mineral formations such as speleothems incave studies combining cultivation and electron microscopicanalyses using B4 medium [36].
Conclusion
This study proves to be a comprehensive insight into thedynamics of the carbon concentrating mechanism by bacteriumSerratia sp. isolated from marble rock. The carbonic anhydraseenzymes produced by the bacterium had role to conversion of CO2
into valuable product like calcite. Scanning electron microscopy(SEM) and EDX analysis revealed the direct involvement of thebacterium in calcium carbonate precipitation. The production ofcalcite was further confirmed by X-ray diffraction (XRD) suggest-ing that the conversion of CO2 into solid carbonates production bythe bacterium was biomineralization processes.
Acknowledgments
This research work was supported by the research grants ofDepartment of Biotechnology, Government of India, New Delhi,
India. One of the authors (S. Srivastava) is thankful to UniversityGrants Commission for Dr. D.S. Kothari post Doctoral Fellowship.We also thank advanced instrumentation facility, JawaharlalNehru University, New Delhi, India, for providing facility ofelectron microscopy, and Jugendra Singh, School of Life Sciences,JNU, New Delhi, for MALDI-TOF/MS analysis.
References
[1] R.K. Shrestha, R. Lal, Ecosystem carbon budgeting and soil carbon sequestration inreclaimed mine soil, Environ. Int. 32 (2006) 781–796.
[2] A.V. Palumbo, J.F. McCarthy, J.E. Amonette, L.S. Fisher, S.D. Wullschleger, W.L.Daniels, Prospects for enhancing carbon sequestration and reclamation of de-graded lands with fossil-fuel combustion by-products, Adv. Environ. Res. 8 (2004)425–438.
[3] B.H. Jo, I.G. Kim, J.H. Seo, D.G. Kang, H.J. Cha, Engineered Escherichia coli withperiplasmic carbonic anhydrase as a biocatalyst for CO2 sequestration, Appl.Environ. Microbiol. (2013), http://dx.doi.org/10.1128/AEM.02400-13.
[4] G. Kim, B.H. Jo, D.G. Kang, C.S. Kim, Y.S. Choi, H.J. Cha, Biomineralization-basedconversion of carbon dioxide to calcium carbonate using recombinant carbonicanhydrase, Chemosphere 87 (2012) 1091–1096.
[5] G.D. Price, D. Sultemeyer, B. Klughammer, M.R. Badger, The functioning of the CO2
concentrating mechanism in several cyanobacterial strains: a review of generalphysiological characteristics, genes, proteins and recent advances, Can. J. Bot. 76(1998) 973–1002.

R.K. Bharti et al. / Journal of Environmental Chemical Engineering 2 (2014) 31–39 39
[6] R. Ramanan, K. Kannan, S.D. Sivanesan, S. Mudliar, S. Kaur, A.K. Tripathi, T.Chakrabarti, Bio-sequestration of carbon dioxide using carbonic anhydrase en-zyme purified from Citrobacter freundii, World J. Microbiol. Biotechnol. 25 (2009)981–987.
[7] R. Ramanan, K. Kannan, N. Vinayagamoorthy, K.M. Ramkumar, S.D. Sivanesan, T.Chakrabarti, Purification and characterization of a novel plant-type carbonicanhydrase from Bacillus subtilis, Biotechnol. Bioprocess Eng. 14 (2009) 32–37.
[8] J.M. Shively, F.L. Ball, B.W. Kline, Electron microscopy of the carboxysomes (poly-hedral bodies) of Thiobacillus neapolitanus, J. Bacteriol. 116 (1973) 1405–1411.
[9] T.O. Yeates, C.A. Kerfeld, S. Heinhorst, G.C. Cannon, J.M. Shivel, Protein-basedorganelles in bacteria: carboxysomes and related microcompartments, Nat. Rev.Microbiol. 6 (2008) 681–691.
[10] I.S. Thakur, Structural and functional characterization of a stable, 4 chlorosalicylicacid degrading, bacterial community in a chemostat, World J. Microbiol. Biotech-nol. 11 (1995) 643–645.
[11] P.K. Jaiswal, S. Kohli, M. Gopal, I.S. Thakur, Isolation and characterization ofalkalotolerant Pseudomonas sp. strain ISTD1 for degradation of dibenzofuran, J.Ind. Microbiol. Biotechnol. 38 (2010) 503–511.
[12] R.G. Khalifah, The carbon dioxide hydration activity of carbonic anhydrase. I.Stop-flow kinetic studies on the native human isoenzymes B and C, J. Biol. Chem.246 (1971) 2561–2573.
[13] A. Innocenti, A. Scozzafava, S. Parkkila, L. Puccetti, G.De. Simone, C.T. Supuran,Investigation of the esterase, phosphate, and sulfatase activities of the cytosolicmammalian carbonic anhydrase isoform I, II and XIII with 4-nitrophenyl esters assubstrates, Bioorg. Med. Chem. Lett. 18 (2008) 2267–2271.
[14] O. Ozensoy, O. Arslan, S.O. Sinan, A new method for purification of carbonicanhydrase isozymes by affinity chromatography, Biochemistry (Moscow) 69(2004) 216–219.
[15] M. Mishra, I.S. Thakur, Purification, characterization and mass spectroscopicanalysis of thermo-alkalotolerant b-1,4 endoxylanase from Bacillus sp. and itspotential for dye decolorization, Int. Biodeter. Biodegr. 65 (2011) 301–308.
[16] S. Srivastava, I.S. Thakur, Biosorption and biotransformation of chromium bySerratia sp. isolated from tannery effluent, Environ. Technol. 33 (2012) 113–122.
[17] Z. Dou, S. Heinhorst, E.B. Williams, C.D. Murin, J.M. Shively, G.C. Cannon, CO2
fixation kinetics of Halothiobacillus neapolitanus mutant carboxysomes lackingcarbonic anhydrase suggest the shell acts as a diffusional barrier for CO2, J. Biol.Chem. 283 (2008) 10377–10384.
[18] M.H. Spalding, Microalgal carbon-dioxide-concentrating mechanisms: Chlamy-domonas inorganic carbon transporter, J. Exp. Botany 59 (2008) 1463–1473.
[19] S. Kumar, K. Tamura, M. Nei, MEGA 3: integrated software for molecular evolu-tionary genetics analysis and sequence alignment brief, Bioinformatics 5 (2004)150–163.
[20] S.J. Giovannoni, T.B. Britschgi, C.L. Moyer, K.G. Field, Genetic diversity in SargassoSea bacterioplankton, Nature 345 (1990) 60–63.
[21] Z. Yu, L. Xie, S. Lee, R. Zhang, A novel carbonic anhydrase from the mantle of thepearl oyster (Pinctada fucata), Comp. Biochem. Physiol. B: Biochem. Mol. Biol. 143(2006) 190–194.
[22] S. Atsumi, W. Higashide, J.C. Liao, Direct photosynthetic recycling of carbondioxide to isobutyraldehyde, Nat. Biotechnol. 27 (2009) 1177–1180.
[23] P. Joseph, S.O. Bettache, J.L. Montero, I. Nishimori, T. Minakuchi, D. Vullo, A.Scozzafava, J.Y. Winumb, S. Kohler, C.T. Supuran, A new b-carbonic anhydrasefrom Brucella suis, its cloning, characterization, and inhibition with sulfonamidesand sulfamates, leading to impaired pathogen growth, Bioorg. Med. Chem. 19(2011) 1172–1178.
[24] K.P. Dobrinski, A.J. Boller, K.M. Scott, Expression and function of four carbonicanhydrase homologs in the deep-sea chemolithoautotroph Thiomicrospira cru-nogena, Appl. Environ. Microbiol. 76 (2010) 3561–3567.
[25] K.S. Smith, J.G. Ferry, Prokaryotic carbonic anhydrases, FEMS Microbiol. 24 (2000)335–366.
[26] L.G. Puskas, M. Inui, K. Zahn, H. Yukawa, A periplasmic alphatype carbonicanhydrase from Rhodopseudomonas palustris is essential for bicarbonate uptake,Microbiology 146 (2000) 2957–2966.
[27] E.A. Marcus, A.P. Moshfegh, D.R. Scott, The periplasmic alpha-carbonic anhydraseactivity of Helicobacter pylori is essential for acid acclimation, J. Bacteriol. 187(2005) 729–738.
[28] M.K. Fasseas, D. Tsikou, E. Flementakis, Molecular and biochemical analysis of theb class carbonic anhydrases in Caenorhabditis elegans, Mol. Biol. Report 37 (2010)2941–2950.
[29] G.M. Bond, J. Stringer, D.K. Brandvold, F.A. Simsek, M.G. Medina, G. Egeland,Development of integrated system for biomimetic CO2 sequestration using theenzyme carbonic anhydrase, Energy Fuels 15 (2001) 309–316.
[30] P. Mirjafari, K. Asghari, N. Mahinpey, Investigating the application of enzymecarbonic anhydrase for CO2 sequestration purposes, Ind. Eng. Chem. Res. 46(2007) 921–926.
[31] W.E. Muller, H.C. Schroder, U. Schlossmacher, M. Neufurth, W. Geurtsen, M.Korzhev, X. Wang, The enzyme carbonic anhydrase as an integral componentof biogenic Ca-carbonate formation in sponge spicules, FEBS Open Bio 3 (2013)357–362.
[32] V. Achal, A. Mukherjee, P.C. Basu, M.S. Reddy, Strain improvement of Sporosarcinapasteurii for enhanced urease and calcite production, J. Ind. Microbiol. Biotechnol.36 (2009) 981–988.
[33] M.C. Portillo, C. Saiz-Jimenez, J.M. Gonzalez, Molecular characterization of totaland metabolically active bacterial communities of white colonizations in theAltamira Cave, Spain Res. Microbiol. 160 (2009) 41–47.
[34] O. Braissant, G. Cailleau, C. Dupraz, P. Verrecchia, Bacterially induced mineraliza-tion of calcium carbonate in terrestrial environments: the role of exopolysac-charides and amino acids, J. Sediment. Res. 73 (2003) 485–490.
[35] C.P. Ercole, A.L. Cacchio, V. Botta, A. Lepidi, Bacterially induced mineralization ofcalcium carbonate: the role of exopolysaccharides and capsular polysaccharides,Microsc. Microanal. 13 (2007) 42–50.
[36] D. Tao, Z. Wenkun, Optimization of calcium carbonate: precipitation for Bacilluspausteurii, Appl. Mech. Mater. 178–181 (2012).