localization of acid phosphatase activity in the basidia
TRANSCRIPT

JOURNAL OF BACTERIOLOGY, JUlY 1973, p. 443-446Copyright i 1973 American Society for Microbiology
Vol. 115, No. 1Printed in U.SA.
NOTES
Localization of Acid Phosphatase Activity in theBasidia of Coprinus micaceus
JEROME L. NEHEMIAH
Department of Pathology, Albert Einstein College of Medicine, Bronx, New York 10461
Received for publication 27 March 1973
Gill lamellae from the young fruiting bodies of the basidiomycete Coprinusmicaceus possess cytoplasmic particles with cytochemically demonstrable acidphosphatase activity; they are presumed to be lysosomes.
In Coprinus micaceus, autolysis of gill tissueoccurs after sporulation. Although autolysis hasbeen described in detail by Buller (1), theintracellular mechanisms of the lytic processhave not been elucidated. From the work ofMatile (4) and Iten and Matile (3), this autol-ysis appears to be a lysosome-mediated phe-nomenon. This cytochemical study was used toleam if lysosomes are demonstrable in thepresporulated basidia of the Coprinus fruitingbody, before autolysis has begun.
Gill tissue of C. micaceus (collected nearAmherst, Mass.) was studied. Pilei of fruitingbodies were broken from the stipes and cut intosmall, wedge-shaped sections of approximately5 to 10 mm in width, and submerged intoice-chilled fixative in a shallow petri dish. Thefixative consisted of 2.5% glutaraldehyde(Fisher Scientific Co.), 1% paraformaldehyde,and 0.025% CaCl2 in 0.05 M cacodylate buffer,pH 6.8 (all expressed as final concentrations).The pileus was rapidly cut away from the gilllamellae. Lamellae were then separated and cutinto rectangular blocks of tissue approximately2 by 6 mm. The blocks were transferred to fresh,cold fixative for a total fixation time of 1.5 h.The blocks were rinsed three times with cold 0.1M cacodylate buffer, pH 6.8, and kept in coldbuffer until incubation.
For the demonstration of acid phosphatase,the blocks of lamellae were incubated at 37 C for15 min in the medium of Gomori modified byNovikoff (5). The ingredients were cytidine mon-ophosphate (25 mg; Sigma Chemical Corp.),distilled water (12 ml), 0.05 M acetate buffer (10ml, pH 5.0), and 1% lead nitrate (3 ml). After
incubation the blocks were rinsed in distilledwater, treated with ammonium sulfide, rinsedin distilled water again, and mounted in glyce-rogel. Controls were treated identically exceptthat the incubation medium contained a finalconcentration of 0.01 M NaF.The interstitial basidioles did not stain in the
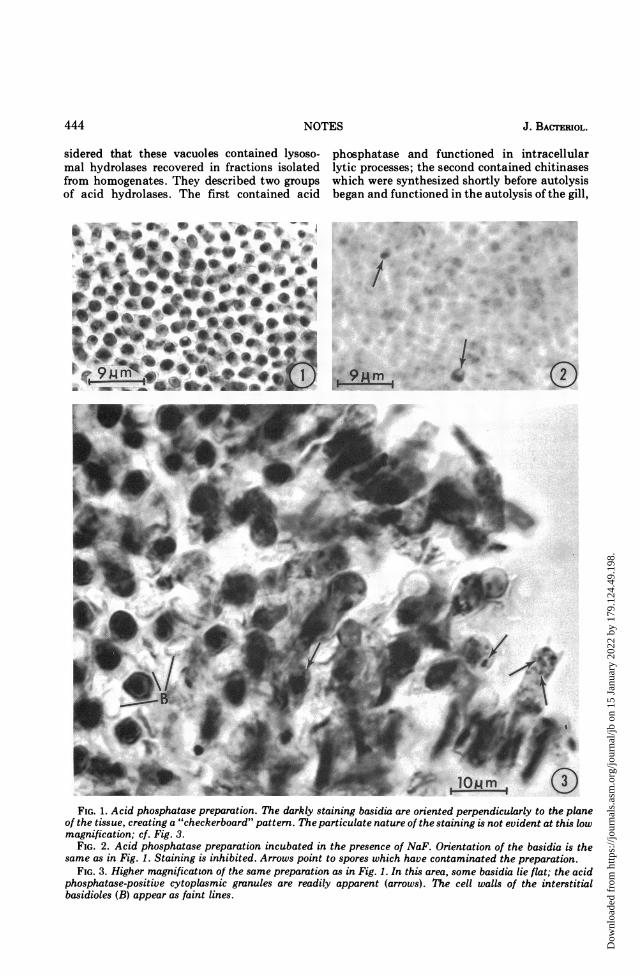
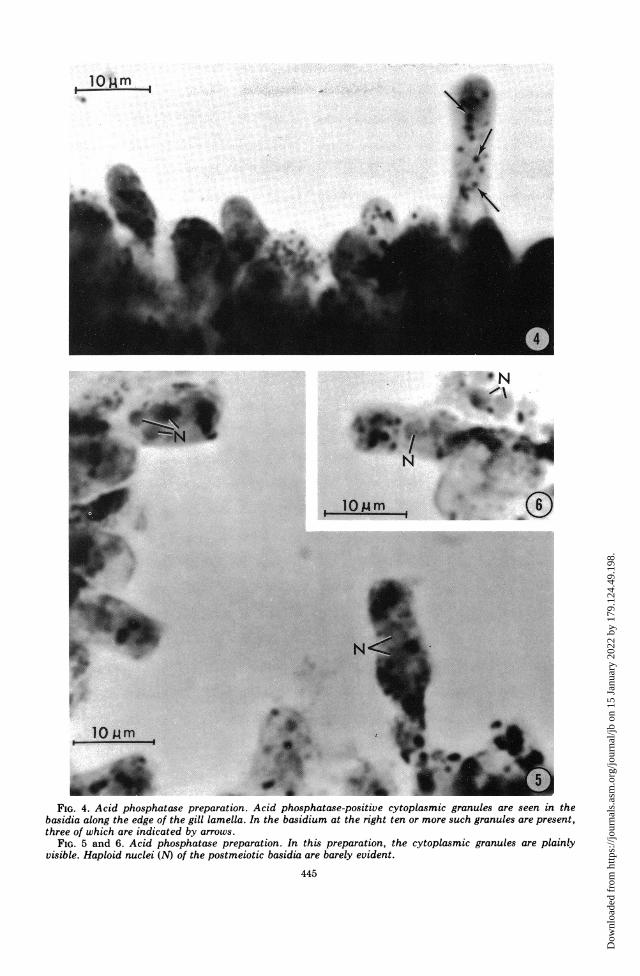
absence or presence of fluoride. In contrast, thebasidia stain darkly in the absence of fluoride(Fig. 1 and 3) but not in its presence (Fig. 2).When a piece of gill was placed flat on the slide,the staining of the basidia presented a "check-erboard" pattern. When oriented in this man-ner, the acid phosphatase granules were dif-ficult to visualize, because the cells were ap-proximately 25 i,m high. However, when aportion of the gill had been flattened, basidiawere often seen lying on edge. In this orienta-tion, acid phosphatase-positive granules wereeasily distinguished in the cytoplasm (Fig. 3-6).The granules ranged in size up to 0.7 gm indiameter. It was not unusual to see five or moregranules within a single basidium (Fig. 4). Theacid phosphatase activity was inhibited by 0.01M NaF.The finding that the postmeiotic, pre-
sporulated basidia of C. micaceus contain cyto-plasmic organelles which give a positive reac-tion for acid phosphatase makes it likely thatthese cells contain lysosomes (2, 5).To determine the types of lysosomes, elec-
tron microscopy of the cytoplasmic granules isrequired.
Iten and Matile (3) have described phase-contrast vacuoles in C. lagopus which measure0.5 to 4.0 jum in diameter. These authors con-
443
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
15
Janu
ary
2022
by
179.
124.
49.1
98.
J. BACTERIOL.
sidered that these vacuoles contained lysoso-mal hydrolases recovered in fractions isolatedfrom homogenates. They described two groupsof acid hydrolases. The first contained acid
phosphatase and functioned in intracellularlytic processes; the second contained chitinaseswhich were synthesized shortly before autolysisbegan and functioned in the autolysis of the gill,
,4,i,,~~~~~
4
I."Or
i 9;4m 1 (E
__ 1eJ 9W | WlomFIG. 1. Acid phosphatase preparation. The darkly staining basidia are oriented perpendicularly to the plane
of the tissue, creating a "checkerboard" pattern. The particulate nature of the staining is not evident at this lowmagnification; cf. Fig. 3.
FIG. 2. Acid phosphatase preparation incubated in the presence of NaF. Orientation of the basidia is thesame as in Fig. 1. Staining is inhibited. Arrows point to spores which have contaminated the preparation.
FIG. 3. Higher magnification of the same preparation as in Fig. 1. In this area, some basidia lie flat; the acidphosphatase-positive cytoplasmic granules are readily apparent (arrows). The cell walls of the interstitialbasidioles (B) appear as faint lines.
444 NOTES
_0-N
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
15
Janu
ary
2022
by
179.
124.
49.1
98.
10Mm
,
t
lOMm
FIG. 4. Acid phosphatase preparation. Acid phosphatase-positive cytoplasmic granules are seen in thebasidia along the edge of the gill lamella. In the basidium at the right ten or more such granules are present,three of which are indicated by arrows.
FIG. 5 and 6. Acid phosphatase preparation. In this preparation, the cytoplasmic granules are plainlyvisible. Haploid nuclei (N) of the postmeiotic basidia are barely evident.
445
.6& N
It
'T 194
d.
N
; I
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
15
Janu
ary
2022
by
179.
124.
49.1
98.

446 NOTES
rather than in intracellular digestion. Since thelysosomes in this study were visualized beforesporulation, it is likely that they belong to thefirst group described by Iten and Matile.Whether the chitinases are packaged in thesame population of lysosomes that contain acidphosphatase remains an unsettled question.
This investigation was supported by Public Health Servicegrant R01-CA-06576 from the National Cancer Institute, to A.B. Novikoff.
LITERATURE CITED1. Buller, R. T. 1922. Researches on fungi, vol. 2. Longmans,
J. BACTERIOL.
Green and Co., New York.2. de Duve, C. 1969. The lysosome in retrospect, p. 3-40. In J.
Dingle and H. B. Fell (ed), Lysosomes in biology andpathology. North Holland Publishing Co., London.
3. Iten, W., and P. Matile. 1970. Role of chitinase and otherlysosomal enzymes of Corprinus lagopus in the autolysisof fruiting bodies. J. Gen. Microbiol. 61:301-309.
4. Matile, H. 1969. Plant lysosomes, p. 406-430. In T. Dingleand H. B. Fell (ed), Lysosomes in biology and pathology.North Holland Publishing Co., London.
5. Novikoff, A. B. 1963. Lysosomes in the physiology andpathology of cells: contributions of staining methods, p.36-73. In A. V. S. de Reuck and M. P. Cameron (ed),Ciba Foundation Symposium on Lysosomes. Little,Brown and Co., Boston.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
15
Janu
ary
2022
by
179.
124.
49.1
98.