mapping intersubunit interactions of the regulatory … intersubunit interactions of the regulatory...
TRANSCRIPT

Mapping Intersubunit Interactions of the RegulatorySubunit (RIa) in the Type I Holoenzyme of ProteinKinase A by Amide Hydrogen/Deuterium ExchangeMass Spectrometry (DXMS)
Yoshimoto Hamuro1†, Ganesh S. Anand2,3†, Jack S. Kim1
Celina Juliano2, David D. Stranz4, Susan S. Taylor2,3 andVirgil L. Woods Jr1*
1Department of MedicineUniversity of California, SanDiego, La Jolla, CA 92093-0656USA
2Departments of Chemistry andBiochemistry, University ofCalifornia, San Diego, La JollaCA 92093-0654, USA
3Howard Hughes MedicalInstitute, University ofCalifornia, San Diego, La JollaCA 92093-0654, USA
4Sierra Analytics, LLC 2105Lancey Drive, Suite 1AModesto, CA 95355, USA
Protein kinase A (PKA), a central locus for cAMP signaling in the cell, iscomposed of regulatory (R) and catalytic (C) subunits. The C-subunitsare maintained in an inactive state by binding to the R-subunit dimer ina tetrameric holoenzyme complex (R2C2). PKA is activated by cAMP bind-ing to the R-subunits which induces a conformational change leading torelease of the active C-subunit. Enzymatic activity of the C-subunit isthus regulated by cAMP via the R-subunit, which toggles between cAMPand C-subunit bound states. The R-subunit is composed of a dimeriza-tion/docking (D/D) domain connected to two cAMP-binding domains(cAMP:A and cAMP:B). While crystal structures of the free C-subunitand cAMP-bound states of a deletion mutant of the R-subunit areknown, there is no structure of the holoenzyme complex or of the cAMP-free state of the R-subunit. An important step in understanding thecAMP-dependent activation of PKA is to map the R–C interface andcharacterize the mutually exclusive interactions of the R-subunit withcAMP and C-subunit. Amide hydrogen/deuterium exchange mass spec-trometry is a suitable method that has provided insights into the differentstates of the R-subunit in solution, thereby allowing mapping of the effectsof cAMP and C-subunit on different regions of the R-subunit. Our studyhas localized interactions with the C-subunit to a small contiguous surfaceon the cAMP:A domain and the linker region. In addition, C-subunitbinding causes increased amide hydrogen exchange within both cAMP-domains, suggesting that these regions become more flexible in theholoenzyme and are primed to bind cAMP. Furthermore, the differencein the protection patterns between RIa and the previously studied RIIbupon cAMP-binding suggests isoform-specific differences in cAMP-dependent regulation of PKA activity.
q 2004 Elsevier Ltd. All rights reserved.
Keywords: cAPK; PKA; DXMS; amide hydrogen/deuterium exchangemass spectometry*Corresponding author
0022-2836/$ - see front matter q 2004 Elsevier Ltd. All rights reserved.
†Y.H. and G.S.A. contributed equally to this work.Present address: Y. Hamuro, ExSAR Corporation, 11 Deer Park Drive, Suite 103, Monmouth Junction, NJ 08852,
USA.
E-mail address of the corresponding author: [email protected]
Abbreviations used: aX:Y, a-helix X in cAMP-binding domain Y; bX:Y, b-sheet X in cAMP-binding domain Y;C-subunit, catalytic subunit; cAMP, adenosine 30,50-cyclic monophosphate; cAMP:A, cAMP-binding domain A;cAMP:B, cAMP-binding domain B; cGMP, guanosine 30,50-cyclic monophosphate; D/D domain, dimerization/dockingdomain; DXMS, amide hydrogen/deuterium exchange mass spectrometry; GuHCl, guanidine hydrochloride; H/Dexchange, hydrogen/deuterium exchange; Mops, 3-(N-morpholino)propane sulfonic acid; PBC, phosphate-bindingcassette; PKA, protein kinase A; R-subunit, regulatory-subunit; TFA, trifluoroacetic acid.
doi:10.1016/j.jmb.2004.05.042 J. Mol. Biol. (2004) 340, 1185–1196
Introduction
Protein kinase A (PKA) is a key enzyme ineukaryotic cell signaling. Activation of PKA byadenosine 30,50-cyclic monophosphate (cAMP)leads to a variety of cellular responses throughregulation of downstream effector enzymes andion channels as well as transcription of specificgenes mediating cell growth and differentiation.1
In the absence of cAMP, PKA exists in an inactivestate as a tetrameric holoenzyme composed of ahomodimeric regulatory (R) subunit and twocatalytic (C) subunits. cAMP binding leads to dis-sociation of the holoenzyme to unleash the activeC-subunit.2 The R-subunit is thus a primary locusfor cAMP-binding in the cell. By toggling betweena C-subunit-bound state and a cAMP-bound state,the R-subunit functions as a sensor for cAMP andas a molecular switch for PKA catalysis.
There are two principal classes of R-subunits(type I and II), each further divided into a and bsubtypes. While the two isoforms of the R-subunitare similar in their domain organization, theydiffer in their subcellular localization and physio-logical function. The R-subunits are highly
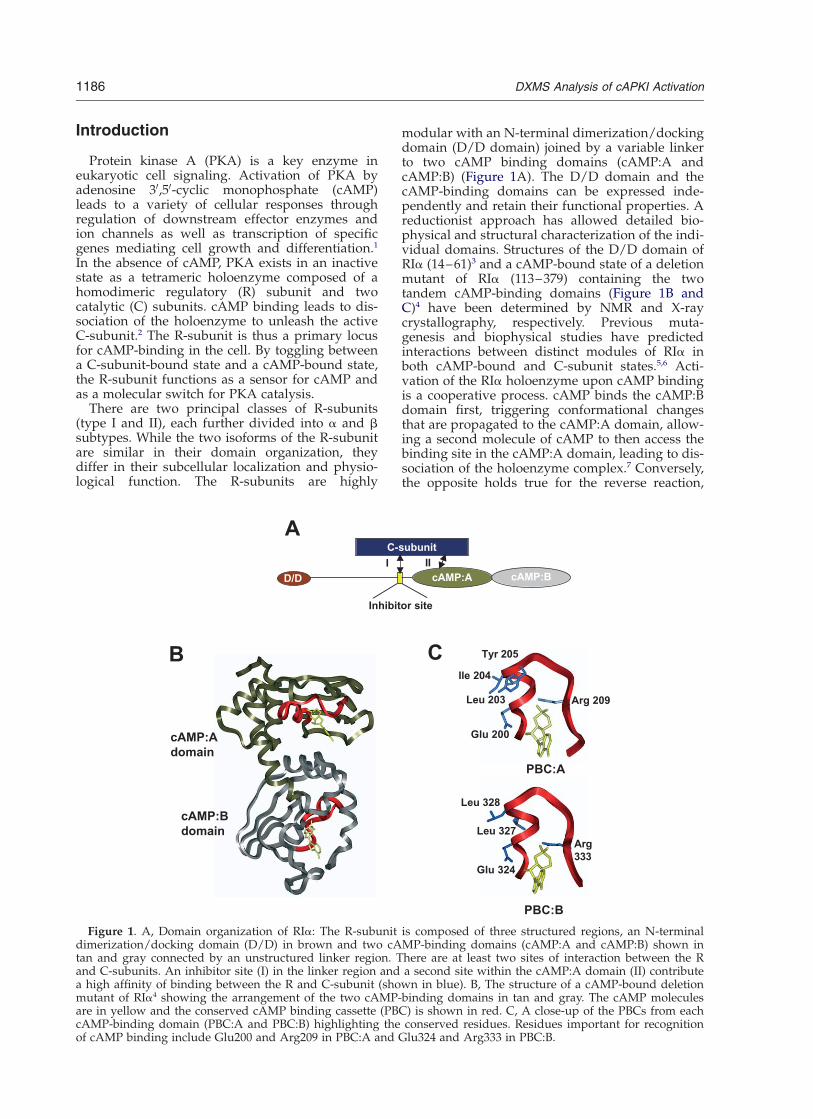
modular with an N-terminal dimerization/dockingdomain (D/D domain) joined by a variable linkerto two cAMP binding domains (cAMP:A andcAMP:B) (Figure 1A). The D/D domain and thecAMP-binding domains can be expressed inde-pendently and retain their functional properties. Areductionist approach has allowed detailed bio-physical and structural characterization of the indi-vidual domains. Structures of the D/D domain ofRIa (14–61)3 and a cAMP-bound state of a deletionmutant of RIa (113–379) containing the twotandem cAMP-binding domains (Figure 1B andC)4 have been determined by NMR and X-raycrystallography, respectively. Previous muta-genesis and biophysical studies have predictedinteractions between distinct modules of RIa inboth cAMP-bound and C-subunit states.5,6 Acti-vation of the RIa holoenzyme upon cAMP bindingis a cooperative process. cAMP binds the cAMP:Bdomain first, triggering conformational changesthat are propagated to the cAMP:A domain, allow-ing a second molecule of cAMP to then access thebinding site in the cAMP:A domain, leading to dis-sociation of the holoenzyme complex.7 Conversely,the opposite holds true for the reverse reaction,
Figure 1. A, Domain organization of RIa: The R-subunit is composed of three structured regions, an N-terminaldimerization/docking domain (D/D) in brown and two cAMP-binding domains (cAMP:A and cAMP:B) shown intan and gray connected by an unstructured linker region. There are at least two sites of interaction between the Rand C-subunits. An inhibitor site (I) in the linker region and a second site within the cAMP:A domain (II) contributea high affinity of binding between the R and C-subunit (shown in blue). B, The structure of a cAMP-bound deletionmutant of RIa4 showing the arrangement of the two cAMP-binding domains in tan and gray. The cAMP moleculesare in yellow and the conserved cAMP binding cassette (PBC) is shown in red. C, A close-up of the PBCs from eachcAMP-binding domain (PBC:A and PBC:B) highlighting the conserved residues. Residues important for recognitionof cAMP binding include Glu200 and Arg209 in PBC:A and Glu324 and Arg333 in PBC:B.
1186 DXMS Analysis of cAPKI Activation

with C-subunit binding to the cAMP-bound R-sub-unit promoting cAMP release from the cAMP:Adomain first, followed by release of cAMP fromthe cAMP:B domain.8
Experimental data on the PKA holoenzyme haveindicated that binding of R-subunits to the C-sub-unit requires at least two sites to achieve bindingwith high affinity (KD ¼ 0.2 nM).9 An inhibitor sitewithin the linker region mimics the substrate anddocks to the active-site cleft of the C-subunit.10 Asecond non-contiguous surface C-terminal to theinhibitor site is responsible for conferring high-affinity binding for the C-subunit (Figure 1A). InRIa, this peripheral site is contributed primarilyby the cAMP:A domain.11,12
Amide hydrogen/deuterium exchange (H/Dexchange) measured by mass spectrometry is amethod that has been used with increasingsophistication in the analysis of protein structure/dynamics,13 – 15 protein–protein interactions,16,17
and protein–ligand interactions.18 – 20 H/Dexchange mass spectrometry has been used exten-sively to study the dynamics of PKA; C-subunitbound to ATP or an inhibitor peptide,21 C-subunitbound to ADP,22 a deletion mutant of RIa boundto C-subunit or cAMP23 and RIIb bound to C-sub-unit or cAMP.24 In this study, we used H/Dexchange coupled to electrospray mass spec-trometry to monitor amide hydrogen exchangewithin full-length RIa when it interacts withcAMP or the C-subunit. Our results confirm theresult of previous studies that, in addition tothe inhibitor site, the helical subdomain within thecAMP:A domain provides a surface of interactionwith the C-subunit.23 Furthermore, our studieshighlight propagated conformational changes inboth cAMP-binding domains upon binding theC-subunit. Comparison of H/D exchange studieson cAMP and C-subunit-bound states of RIa in
this study with previous studies on RIIb is furtherdiscussed and our analysis suggests a basis forunderstanding R-subunit isoform-specificregulation of PKA.
Results
RIa proteolytic fragmentation
Prior to H/D exchange experiments, digestionconditions that produced RIa fragments of optimalsize and distribution for exchange analysis wereestablished (see Materials and Methods). Pepsindigestion of RIa was carried out by diluting thedeuterated sample (1 : 1.5, v/v) with cold quenchsolution (1.6 M GuHCl (pH 2.0) in 0.8% (v/v) for-mic acid). The quenched sample was flowed overimmobilized pepsin (66 ml bed volume) at a flow-rate of 300 ml/minute at 0 8C, allowing a digesttime of 13 s. These conditions generated 92 pep-tides covering the entire amino acid sequence ofRIa (Figure 2). Forty-one of the identified peptidesthat spanned 100% of the entire sequence were ofhigh quality (high signal/noise) for amide hydro-gen/deuterium exchange mass spectrometry(DXMS) analysis of the three states of RIa (boldlines in Figure 2). Since both the amino group ofthe first amino acid residue and the amide hydro-gen atom of the second amino acid residue tend toexchange too rapidly to retain deuterons duringthe experiment,25 the total number of amide hydro-gen atoms followed by DXMS was 314 out of 363non-proline residues (86%).
Characterization of three states of RIa by DXMS
To map the effects of activation of PKA, cAMP-bound and C-subunit bound states of RIa were
Figure 2. Pepsin digest peptide coverage map of RIa. Bold lines indicate the peptides identified, analyzed and usedin this study. Dotted lines indicate the peptides identified and analyzed but not used in this study.
DXMS Analysis of cAPKI Activation 1187

characterized by DXMS in this study. Additionally,to discriminate between the effects of cAMPremoval and C-subunit binding, a third cAMP-free state of RIa was analyzed. Larger than 10%changes in average deuterium incorporationduring the exchange time employed (10–3000 s)were considered significant in our analysis.
Deuterium on-exchange of cAMP-bound RIa
cAMP-bound RIa was incubated in deuteratedMops buffer (pH 7.0) for a time-course (10–3000 s)at 23(^1) 8C and quenched with low-pH buffer at0 8C. The average deuterium back-exchange duringthe analysis was 13% (see Materials and Methods).The deuteration level of cAMP-bound RIa as afunction of time is illustrated in Figure 3. RIa canbe divided into three regions; an N-terminal dimer-ization/docking domain (D/D domain), twotandem cAMP-binding domains, and a long linkerregion connecting the cAMP:A domain and D/Ddomain. Regions with different rates of exchangewere observed in full-length RIa. The fast-exchanging region spanned the N-terminal region(residues 1–120) that consists of a highly solvent-exposed linker region and D/D domain, while amore ordered C-terminal region of the protein con-taining tandem cAMP-binding domains showedslower exchange overall.
While a structure of full-length RIa is not avail-able, the structures of two cAMP-binding domains4
and the D/D domain3 have been used to map theresults of the deuterium on-exchange. In the D/Ddomain (residues 1–61), residues 3–18 exchangedvery rapidly, while segment 31–36, which mapsonto the center of helix I, exchanges relativelyslowly. Segment 39–47, spanning the turn connect-ing helices I and II, exchanged faster than thecenter of helix I.
Two segments (residues 80–101 and 105–110)from the linker region (residues 62–120) werefully deuterated even at the shortest exchangetime in our experiments (ten seconds), suggestingthis linker region is highly disordered. Themajority of amide hydrogen atoms in segment50–77 also exchanged very rapidly, as over 60% ofthem were already exchanged after ten seconds ofon-exchange. Residues 113–125 and 128–135 fromthe aXn helix showed slower exchange than thepeptides from the linker region, reflecting a moreordered structure for the aXn helix. None of thesegments analyzed in the cAMP-binding domains,except segment 377–379, show more than 90%deuteration even after the longest exchange timeemployed (3000 seconds). The most slowlyexchanging regions are segments 151–156 (b1:A),174–180 (b3:A and b4:A), 204–221 (b7:A andb8:A), 224–229 (b8:A and aB:A), 257–270 (aA:B),
Figure 3. Amide H/D exchange results of cAMP-bound RIa. Each block indicates peptide analyzed. Each block con-tains six time-points; from the top, ten seconds, 30 seconds, 100 seconds, 300 seconds, 1000 seconds and 3000 seconds.The deuteration levels of each segment at each time-point are shown by different colors from blue (,10% deuteration)through red (.90% deuteration), as indicated on the right of the Figure. The secondary structural elements are shownon top of the primary sequence. aX:Y in the Figure indicates a-helix X in cAMP-binding domain Y and bX:Y indicatesb-sheet X in cAMP-binding domain Y.
1188 DXMS Analysis of cAPKI Activation
294–298 (b3:B), 324–327 (b6:B and aB0:B), and330–345 (b7:B and b8:B). Interestingly, all thesesegments spanned b-strands, reflecting that, ingeneral, b-sheet elements show slower exchangewith solvent than a-helical regions (see Figure 3).26
Effects of C-subunit binding to RIa
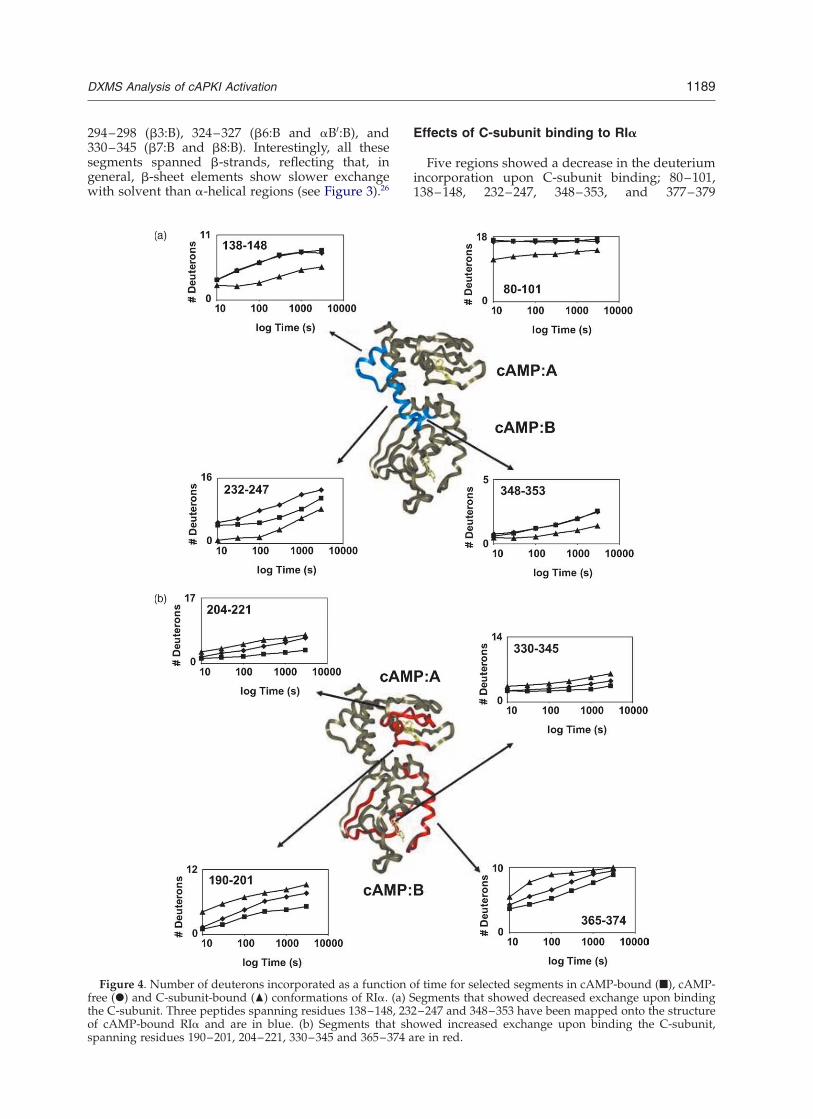
Five regions showed a decrease in the deuteriumincorporation upon C-subunit binding; 80–101,138–148, 232–247, 348–353, and 377–379
Figure 4. Number of deuterons incorporated as a function of time for selected segments in cAMP-bound (B), cAMP-free (X) and C-subunit-bound (O) conformations of RIa. (a) Segments that showed decreased exchange upon bindingthe C-subunit. Three peptides spanning residues 138–148, 232–247 and 348–353 have been mapped onto the structureof cAMP-bound RIa and are in blue. (b) Segments that showed increased exchange upon binding the C-subunit,spanning residues 190–201, 204–221, 330–345 and 365–374 are in red.
DXMS Analysis of cAPKI Activation 1189
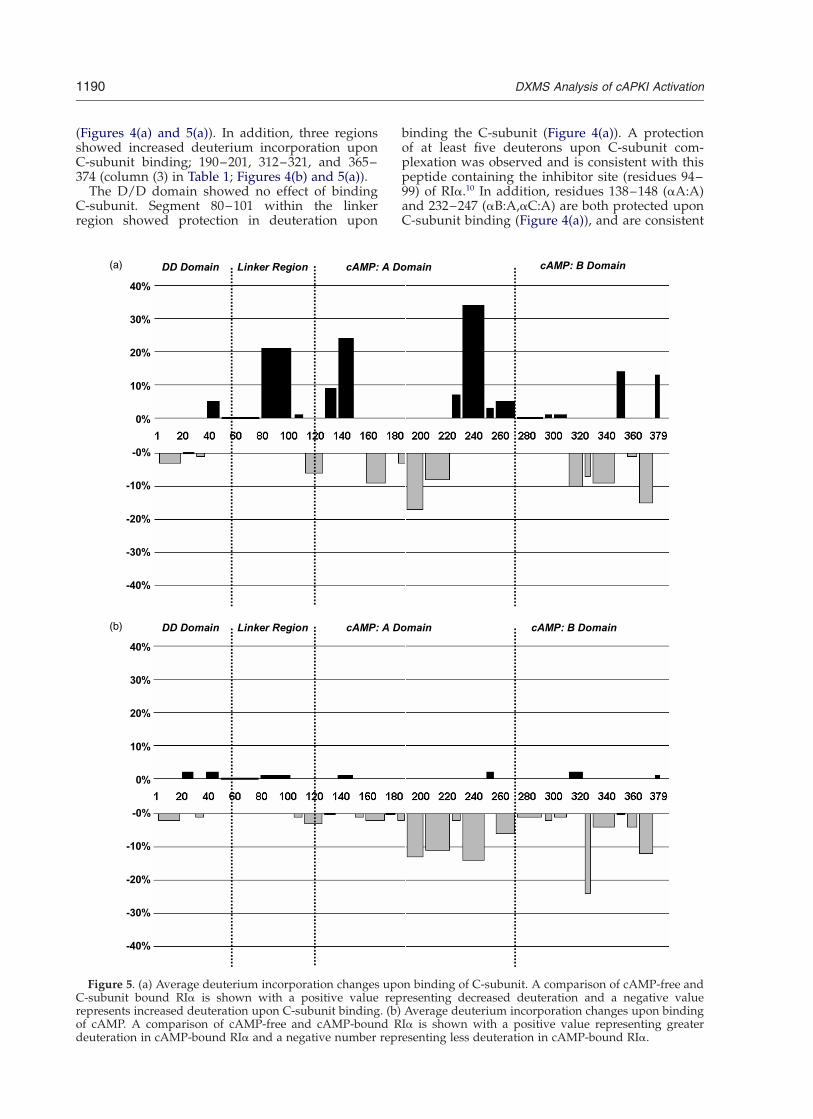
(Figures 4(a) and 5(a)). In addition, three regionsshowed increased deuterium incorporation uponC-subunit binding; 190–201, 312–321, and 365–374 (column (3) in Table 1; Figures 4(b) and 5(a)).
The D/D domain showed no effect of bindingC-subunit. Segment 80–101 within the linkerregion showed protection in deuteration upon
binding the C-subunit (Figure 4(a)). A protectionof at least five deuterons upon C-subunit com-plexation was observed and is consistent with thispeptide containing the inhibitor site (residues 94–99) of RIa.10 In addition, residues 138–148 (aA:A)and 232–247 (aB:A,aC:A) are both protected uponC-subunit binding (Figure 4(a)), and are consistent
Figure 5. (a) Average deuterium incorporation changes upon binding of C-subunit. A comparison of cAMP-free andC-subunit bound RIa is shown with a positive value representing decreased deuteration and a negative valuerepresents increased deuteration upon C-subunit binding. (b) Average deuterium incorporation changes upon bindingof cAMP. A comparison of cAMP-free and cAMP-bound RIa is shown with a positive value representing greaterdeuteration in cAMP-bound RIa and a negative number representing less deuteration in cAMP-bound RIa.
1190 DXMS Analysis of cAPKI Activation
with previous studies that identified the sameregions as part of a second contiguous surface thatinteracts with the C-subunit.23
Some segments showed increased exchangerates in RIa upon the binding of C-subunit andthe removal of cAMP (column (1) in Table 1;Figure 4(b)). These include residues 190–201, and204–221 in the cAMP:A domain, and residues324–327, 330–345 and 365–374 in the cAMP:Bdomain (Figure 4(b)). Residues 190–201 and204–221 are part of the b-barrel subdomain of
RIa. Residues 204–221 span the conserved cAMP-binding motif and phosphate-binding cassette(PBC). Residues 324–327 and 330–345 span thePBC of cAMP:B domain, while the segment 365–374 is close to the C terminus. While these regionsexchange faster upon removal of cAMP (goingfrom the cAMP-bound form to the cAMP-freeform, Figures 4(b) and 5(b)), the same regionsexchange even faster upon C-subunit binding, pre-sumably reflecting C-subunit-induced allostericchanges in RIa (comparing cAMP-free RIa andholoenzyme; Figures 4(b) and 5(a)).
Change in deuteration upon cAMP binding
A comparison of deuterium incorporation incAMP-bound and cAMP-free states of RIa pro-vides insights into the effects of cAMP binding(column (2) in Table 1; Figure 5(b)). No significantchange in deuterium incorporation was observedupon cAMP binding in the D/D domain and linkerregion. Four regions in the cAMP-binding domainswere protected from H/D exchange upon cAMPbinding; residues 190–221 (covered by two seg-ments, 190–201 and 204–221), 232–247, 324–327,and 365–374.
Discussion
C-subunit-binding surface of RIa
A primary interaction between the C-subunitand RIa is mediated by the inhibitor site(Figure 1A, residues 94–98), which mimics sub-strate binding. RIa contains Ala in place of thephosphorylatable Ser in the consensus site forphosphorylation by PKA, and MgATP is essentialfor formation of this complex.10 In our H/Dexchange experiments, segment 80–101, whichspans this region, showed protection upon C-sub-unit binding. The maximum difference in theextent of deuteration between holoenzyme andcAMP-free (or cAMP-bound) RIa was approxi-mately five deuterons at the shortest exchangetime employed (ten seconds), indicating that aminimum of five amide hydrogen atoms showedslower exchange with solvent upon binding theC-subunit (Figure 4(a)). Because the minimumconsensus phosphorylation site for PKA is fiveresidues long, this five amide protection can mostlikely be attributed to direct interface contactswith the C-subunit binding at the inhibitor site.Although previous studies have indicated theimportance of additional residues N-terminal tothe inhibitor site in interactions with theC-subunit,27,28 our results do not show any additionalprotection in those regions upon C-subunit bindingin the time-course of the H/D exchange experi-ments. While these results do not exclude the possi-bility of additional C-subunit binding sites in thelinker region, these are presumably unobservable,
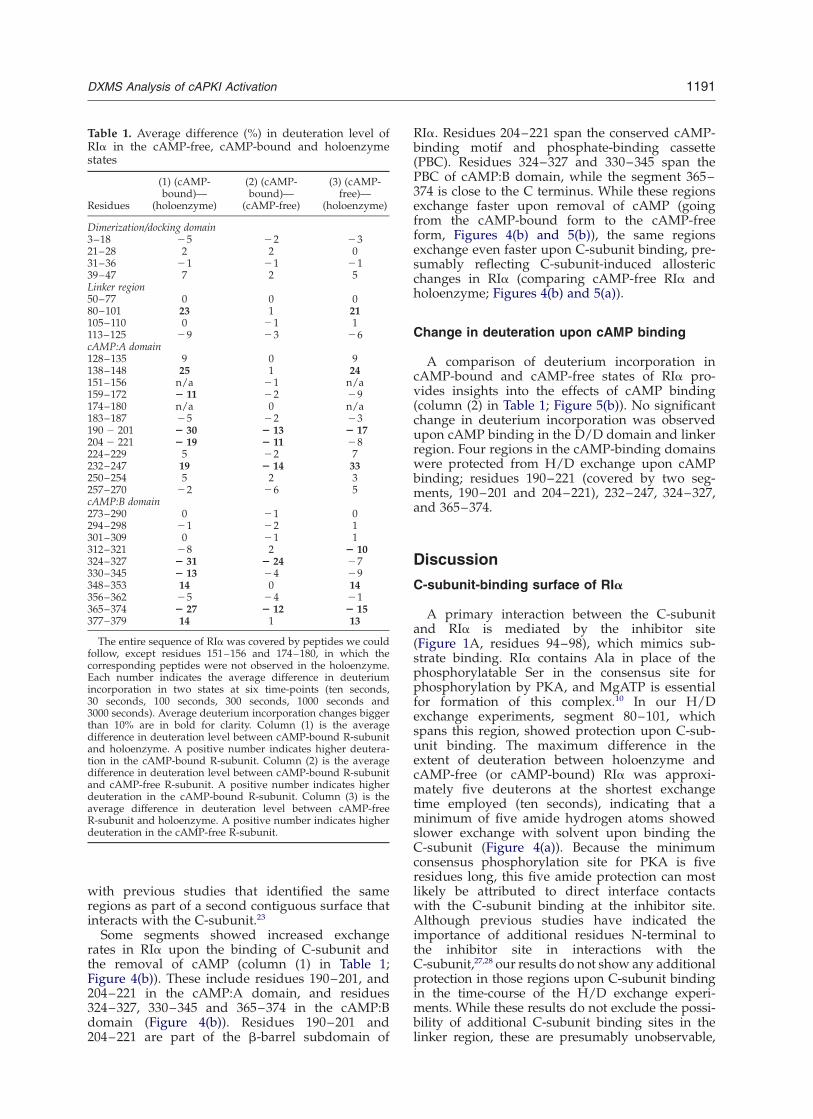
Table 1. Average difference (%) in deuteration level ofRIa in the cAMP-free, cAMP-bound and holoenzymestates
Residues
(1) (cAMP-bound)—
(holoenzyme)
(2) (cAMP-bound)—
(cAMP-free)
(3) (cAMP-free)—
(holoenzyme)
Dimerization/docking domain3–18 25 22 2321–28 2 2 031–36 21 21 2139–47 7 2 5Linker region50–77 0 0 080–101 23 1 21105–110 0 21 1113–125 29 23 26cAMP:A domain128–135 9 0 9138–148 25 1 24151–156 n/a 21 n/a159–172 2 11 22 29174–180 n/a 0 n/a183–187 25 22 23190 2 201 2 30 2 13 2 17204 2 221 2 19 2 11 28224–229 5 22 7232–247 19 2 14 33250–254 5 2 3257–270 22 26 5cAMP:B domain273–290 0 21 0294–298 21 22 1301–309 0 21 1312–321 28 2 2 10324–327 2 31 2 24 27330–345 2 13 24 29348–353 14 0 14356–362 25 24 21365–374 2 27 2 12 2 15377–379 14 1 13
The entire sequence of RIa was covered by peptides we couldfollow, except residues 151–156 and 174–180, in which thecorresponding peptides were not observed in the holoenzyme.Each number indicates the average difference in deuteriumincorporation in two states at six time-points (ten seconds,30 seconds, 100 seconds, 300 seconds, 1000 seconds and3000 seconds). Average deuterium incorporation changes biggerthan 10% are in bold for clarity. Column (1) is the averagedifference in deuteration level between cAMP-bound R-subunitand holoenzyme. A positive number indicates higher deutera-tion in the cAMP-bound R-subunit. Column (2) is the averagedifference in deuteration level between cAMP-bound R-subunitand cAMP-free R-subunit. A positive number indicates higherdeuteration in the cAMP-bound R-subunit. Column (3) is theaverage difference in deuteration level between cAMP-freeR-subunit and holoenzyme. A positive number indicates higherdeuteration in the cAMP-free R-subunit.
DXMS Analysis of cAPKI Activation 1191

due to the rapid exchange seen for the linker regionin all three states of RIa.
Secondary site of C-subunit binding iscontributed primarily by cAMP:A
Previous deletion mapping studies had shownthat a construct containing the inhibitor site andcAMP:A domain retained relatively high-affinitybinding to the C-subunit.11,12 This suggested that,in addition to the inhibitor site, the cAMP:Adomain was the smallest determinant for highaffinity binding to the C-subunit. Recent H/Dexchange studies on the cAMP-bound, free andC-subunit-bound states of a deletion mutant ofRIa (RIa (94–244) localized C-subunit binding to asmall surface on the helical subdomain ofcAMP:A domain formed by residues 137–148(aA:A) and 223–238 (aB:A and aC:A).23 Further-more, on the basis of these studies, computationaldocking programs enabled modeling of the inter-subunit interface of type I holoenzyme using thestructures of RIa and C-subunit that is mostconsistent with earlier experimental data.29
Our results confirm that the cAMP:A domain, inaddition to the inhibitor site, is the main locus forC-subunit binding. The two segments (138–148and 232–247) outside the inhibitor site that showstrong protection upon C-subunit binding liewithin helices aA:A and aC:A, forming a con-tiguous surface within cAMP:A domain, as seenpreviously (Figures 4(a) and 5(a)).23
Effects of C-subunit binding to cAMP-bindingdomains A and B: global conformationalchange upon binding the C-subunit
Of 28 segments in Table 1 that were analyzed inthe holoenzyme, 11 showed significant change indeuteration upon binding the C-subunit andremoval of cAMP. Six of these segments (159–172,190–201, 204–221, 324–327, 330–345 and 365–374) showed an increase in deuterium exchangeupon C-subunit binding (column (1) in Table 1).All of these segments span the cAMP-bindingpockets of cAMP:A and cAMP:B domains directlyor are proximal to the cAMP-binding sites.Although the changes seen in the holoenzymestate might well be a consequence of not havingcAMP occupying the two sites in the R-subunit,characterization of cAMP-free R-subunit allowedus to separate the effects of C-subunit bindingfrom the removal of cAMP. These segments showgreater exchange in the C-subunit-bound staterelative to both the cAMP-bound state and thecAMP-free state (column (3) in Table 1; Figure 5).The greatest increases in exchange comparingcAMP and C-subunit-bound states were seen inthree segments, 190–201, 324–327 and 365–374(column (1) in Table 1). These results point tolong-distance effects of C-subunit binding that arepropagated to the PBCs of both cAMP:A andcAMP:B domains. This suggests that the cAMP-
binding pockets in both domains are moredynamic in the holoenzyme, and thus are primedto bind cAMP, even though the PBCs remain well-shielded overall from solvent in all three states ofthe R-subunit. While these effects are consistentwith the results of previous studies on thecAMP:A domain,23 our results on full-length RIashow that these effects of C-subunit binding areclearly seen in the cAMP:B domain as well.
The overall effects of C-subunit and cAMP bind-ing to full-length RIa are identical with previousstudies on a deletion mutant of RIa (94–244),23
considering the different time-course of analysis inthe two studies. One major difference was seen inthe two studies. In the cAMP:A domain, segment113–125 was protected upon C-subunit binding inthe RIa (94–244) study,23 while the same segmentin the present study was found to show greaterdeuteration upon C-subunit binding. We believethe basis of this difference is that the deletionmutant RIa (94–224) exists as a monomer, whilefull-length RIa exists as a dimer.
Previous studies have shown that cAMP:Bdomain plays a role of “gatekeeper” to regulateaccess of a second molecule of cAMP to thecAMP:A domain in the holoenzyme.5,11 Further-more, deletion of the cAMP:B domain does notabolish binding to the C-subunit.10 Our results areconsistent with these studies, and only a small sur-face within the cAMP:B domain showed decreasedexchange upon binding the C-subunit. However,C-subunit binding results in increased exchange inboth cAMP:A and cAMP:B domains, indicatingthat C-subunit binding induces changes that arepropagated to distal regions of RIa. This suggeststhat, in addition to playing the role of gatekeeperfor cAMP binding, it influences indirectly C-sub-unit association and dissociation. These resultsprovide insight into the cooperativity of cAMP-binding to the R-subunit and dissociation of theC-subunit in the holoenzyme state. Residues 348–353 in the B-helix of cAMP:B domain (aB:B) andresidues 377–379 at the C terminus showdecreased deuteration upon C-subunit binding(Table 1; Figure 5(a)).
Effects of cAMP binding on RIa
Five segments are protected upon cAMP binding(column (2) in Table 1 and Figure 5(b)), includingresidues 190–201 and 204–221 spanning the PBCof cAMP:A domain, residues 324–327 spanningthe PBC of the cAMP:B domain, and residues365–374 spanning the C-terminal end of themolecule. The decreased deuterium incorporationin these residues upon cAMP binding reflects thedirect effect of cAMP binding. Residues 365–374spanning the C-terminal end of the protein showeddecreased deuterium exchange upon cAMP bind-ing. This segment includes Tyr371, whose aromaticring stacks against the adenosine ring of cAMP.Presumably, the interaction with cAMP decreasesthe flexibility of this region and resulted in the
1192 DXMS Analysis of cAPKI Activation

decreased deuterium incorporation in this seg-ment. The increased flexibility around Tyr371 isconsistent with the structure of the cAMP-free Bdomain of RIa, where there is a change observedin the C-terminal region upon removal of cAMP.30
Segment 232–247 is the only region that showedsignificant decrease in deuteration upon cAMPbinding and is distal to the cAMP-binding sites.This segment covers most of the C-helix ofcAMP:A domain, and the changes in deuteriumexchange suggest an allosteric change upon bind-ing cAMP.
The lack of deuteration change in the D/Ddomain upon binding cAMP and C-subunit mayimply the lack of communication between cAMP-binding sites and D/D domain or linker region(Table 1; Figure 5). This is consistent with the factthat the D/D domain behaves as an independentmodule with little difference in the extent of deu-terium exchange in the D/D domain relative tothe full-length protein. The binding affinity of theisolated D/D domain with effector AKAP proteinsis identical with that of the full-length protein,suggesting that the rest of the molecule does notalter the properties of the D/D domain.31
Comparative analysis of RIa and RIIb by DXMS
Characterization of cAMP and C-subunit bind-ing to RIIb by DXMS have been reported.24 In ourstudy on RIa, we have used the same experimentalconditions, and this allows us to compare the tworegulatory subunit isoforms. While the two iso-forms of the R-subunit show a high level ofsequence similarity, they are functionally verydifferent both in terms of their distinct localizationwithin the cell and potentially through differencesin their association and dissociation with theC-subunit. Both isoforms show bipartite bindingto the C-subunit with a primary site of interactionlocated in the linker region and a secondary site ofinteraction within the cAMP-binding domains. Inboth isoforms, the D/D domain shows little or nodifference upon C-subunit and cAMP-binding andappears to be independent of the rest of themolecule. However, there are isoform-specificdifferences in the cAMP-binding domains bothupon cAMP and C-subunit binding.
Regions in RIIb that show protection uponC-subunit binding included the segment spanningthe entire C-helix of cAMP:A domain (aC:A,residues 253–268). This segment is highly homo-logous between RIa and RIIb32 and, interestingly,an equivalent pepsin digest fragment in RIa wasanalyzed in the present study (residues 232–247).While this segment shows protection upon C-sub-unit binding in both isoforms, the extent of protec-tion in RIIb was much more than in RIa for thissegment in the timescale of our experiments.There are differences also in the effects of cAMPbinding. While the segment spanning the C-helixof the cAMP domain (residues 253–268) did notshow any change upon cAMP binding in the RIIb
isoform,24 the equivalent region in RIa (residues232–247) showed decreased deuteration uponcAMP binding. The different effects of cAMP bind-ing on RIa compared to RIIb are interesting andsuggest isoform-specific differences in the mechan-ism of cAMP-dependent activation. Indeed, earlystudies characterizing the type I and II isoforms ofthe PKA holoenzymes had observed that the typeI holoenzyme is activated in vitro at lower concen-trations of cAMP than the type II holoenzyme.33
However, it was pointed out that this difference insensitivity to cAMP is lost when MgATP is boundto the C-subunit and upon autophosphorylation ofthe RII subunit.2 This is especially relevant, sinceMgATP binding to RIa is obligatory for holo-enzyme formation.34 It must be pointed out thatthe above observations on the RII subunit weremade from studies on RIIa, and it appears thatproperties of the RIIa and RIIb homodimers andholoenzymes in solution are different.35 It is inter-esting to speculate that cAMP binding to RIainduces conformational changes in the C-helix,which is on the binding surface of the C-subunit,to facilitate the release of the C-subunit and acti-vation of PKA.
Conclusion
Amide H/D exchange coupled with proteolysisand mass spectrometry applied to full-length PKARIa in its cAMP-bound, cAMP-free and C-sub-unit-bound states provides an understanding ofthe dynamics of activation of PKA. While theeffects of cAMP binding to R-subunit are localizedprimarily within two cAMP-binding cassettes,except for the C-helix of cAMP-binding domain A(aC:A), binding of the C-subunit causes changesin all regions of the R-subunit outside the D/Ddomain. The binding of C-subunit to RIa slowsthe amide H/D exchange rates at a bindingsurface that spans the linker region and partof cAMP binding domain A, and accelerates theH/D exchange rates around both cAMP-bindingcassettes. The wide applicability and relative highresolution of this H/D exchange coupled withmass spectrometry method can provide novelinsight into protein dynamics by complementinghigh-resolution X-ray crystallographic and NMRstudies.
Materials and Methods
Materials
ATP, cAMP, Mops and cAMP immobilized on 6%(w/v) agarose were obtained from Sigma. Deuteriumoxide, 2H2O (99.9% deuterium) was obtained fromCambridge Isotopes.
Expression and purification
Proteins were expressed in Escherichia coli BL21 (DE3)
DXMS Analysis of cAPKI Activation 1193

cells (Novagen) and purified as described, using cAMP-agarose resin.36 Following cell lysis, protein was precipi-tated from the soluble fraction by 45% saturatedammonium sulfate (AS) at 4 8C. The AS pellets wereresuspended, incubated overnight with the cAMP-agarose resin, and eluted at room temperature. ForcAMP-bound RIa, the protein was eluted with buffercontaining 25 mM cAMP. For cAMP-free RIa, the proteinwas eluted with buffer containing 25 mM cGMP. Theprotein eluates were then dialyzed overnight at 4 8Cagainst buffer A (20 mM Mops (pH 7.0), 200 mM NaCl,2 mM EDTA, 2 mM EGTA, 10 mM DTT).
Holoenzyme formation
The cAMP-free RIa was added to wild-type C-subunitin a 1 : 1.2 molar ratio and dialyzed overnight at 4 8Cagainst buffer B (10 mM Mops (pH 6.5), 2 mM MgCl2,50 mM NaCl, 0.2 mM ATP, 1 mM DTT). To removeexcess C-subunit, the complex was purified by elutionthrough a S200 gel-filtration column and then concen-trated with 30 kDa cutoff Millipore concentrators, andstored in buffer B at 4 8C.
DXMS analysis
General operation procedure
A 20 ml H/D-exchanged protein solution (5 mM) wasquenched by shifting to pH 2.2–2.5, 0 8C with 30 ml of3.2 M GuHCl in 0.8% formic acid (the final pH wasmeasured on a non-deuterated mock solution at roomtemperature using a model 250 pH meter (DenverInstrument Co., Arvada, Colorado)). At 0 8C, thequenched solution was immediately passed over acolumn (66 ml bed volume; Upchurch Scientific, cat #C.130B) filled with porcine pepsin (Sigma, cat # p6887)immobilized on Poros 20 AL medium at 30 mg/mlfollowing the manufacturer’s instructions, with 0.05%(v/v) trifluoracetic acid (TFA) (300 ml/minute) for oneminute with contemporaneous collection of proteolyticproducts by a C18 column (Vydac cat # 218MS5105).The duration of digestion was 13 s. Subsequently, theC18 column was eluted with a linear gradient of10–40% B over ten minutes (solvent A was 0.05% TFAin water, and solvent B was 80% (v/v) acetonitrile, 20%(v/v) water, 0.01% TFA). Mass spectrometric analyseswere carried out with a Finnigan LCQ mass spec-trometer with capillary temperature at 200 8C.
Optimization of fragmentation conditions
Two major parameters optimized were the concen-tration of guanidine hydrochloride (GuHCl) in thequenching buffer and the pump C flow-rate over thepepsin column. A 2 ml stock solution of RIa (4 mg/ml)was diluted with 18 ml of water and then quenchedwith 30 ml of 0.8% formic acid containing various con-centrations of GuHCl (0–6.4 M). The quenched samplewas immediately injected into the DXMS apparatus andthe system started. The flow-rate passing over thepepsin column was varied between 100 ml/minute and300 ml/minute.
Sequence identification of pepsin-generated peptides
To quickly identify pepsin-generated peptides for eachdigestion condition employed, spectral data were
acquired in “triple play” mode. The triple play data setwas then analyzed employing the Sequest softwareprogram (Finnigan Inc.) to identify the sequence of thedynamically selected parent peptide ions. This tentativepeptide identification was verified by visual con-firmation of the parent ion charge state presumed bythe Sequest program for each peptide. This set of pep-tides was then further examined to determine if the“quality” of the measured isotopic envelope of peptideswas sufficient to allow accurate measurement of the geo-metric centroid of isotopic envelopes on deuteratedsamples.
Deuterium exchange experiments
Deuterated samples were prepared at 23(^1) 8C bydiluting 2 ml of RIa stock solution with 18 ml of deuter-ated buffer (20 mM Mops (pHread 7.0), 50 mM NaCl,1 mM DTT), followed by “on-exchange” incubation forvarying times (10–3000 s) prior to quenching in 30 ml of0.8% formic acid, 1.6 M GuHCl at 0 8C. These function-ally deuterated samples were then subjected to DXMSprocessing as above, along with control samples of non-deuterated and fully deuterated RIa (incubated in 0.5%formic acid in 90% 2H2O for two days at room tempera-ture).
The centroids of probe peptide isotopic envelopeswere measured using the in-house-developed programin collaboration with Sierra Analytics. The correctionsfor back-exchange were made employing methodsdescribed:37
Deuteration level ð%Þ ¼mðPÞ2mðNÞ
mðFÞ2mðNÞ£ 100
Deuterium incorporation ðnumberÞ
¼mðFÞ2mðNÞ
mðFÞ2mðNÞ£ MaxD
where m(P), m(N), and m(F) are the centroid value ofpartially deuterated peptide, non-deuterated peptide,and fully deuterated peptide, respectively. MaxD is themaximum deuterium incorporation calculated by sub-tracting the number of proline residues in the third orlater amino acid residue and two from the number ofamino acid residues in the peptide of interest (assumingthe first two amino acid residues cannot retaindeuterons.25 The deuteron recovery of fully deuteratedsample ((m(F) 2 m(N))/MaxD)/0.9 was, on average, 87%.
Acknowledgements
We thank Simon Brown for assistance in proteinpurification. These studies were funded byUniversity of California BioStar Technology trans-fer grants S97-90 and S99-44 (to V.L.W.) and Uni-versity of California Life Sciences Informatics (LSI)Technology transfer grant L98-30 (to V.L.W.).ExSAR Corporation is the matching partner forthese grants. V.L.W. has financial interests inExSAR Corporation. This work was supportedalso by an NIH grant to V.L.W. (CA-099835) S.S.T.(DK-54441).
1194 DXMS Analysis of cAPKI Activation

References
1. Shabb, J. B. (2001). Physiological substrates of cAMP-dependent protein kinase. Chem. Rev. 101, 2381–2411.
2. Beebe, S. J. & Corbin, J. D. (1986). Cyclic nucleotide-dependent protein kinases. Enzymes, 17, 43–111.
3. Banky, P., Roy, M., Newlon, M. G., Morikis, D., Haste,N. M., Taylor, S. S. & Jennings, P. A. (2003). Relatedprotein–protein interaction modules presentdrastically different surface topographies despite aconserved helical platform. J. Mol. Biol. 330,1117–1129.
4. Su, Y., Dostmann, W. R., Herberg, F. W., Durick, K.,Xuong, N. H., Ten Eyck, L. et al. (1995). Regulatorysubunit of protein kinase A: structure of deletionmutant with cAMP binding domains. Science, 269,807–813.
5. Herberg, F. W., Dostmann, W. R. & Taylor, S. S.(1996). Active site mutations define the pathway forthe cooperative activation of cAMP-dependentprotein kinase. Biochemistry, 35, 2934–2942.
6. Herberg, F. W., Dostmann, W. R., Zorn, M., Davis, S. J.& Taylor, S. S. (1994). Crosstalk between domains inthe regulatory subunit of cAMP-dependent proteinkinase: influence of amino terminus on cAMP bind-ing and holoenzyme formation. Biochemistry, 33,7485–7494.
7. Ogreid, D. & Doskeland, S. O. (1981). The kinetics ofassociation of cyclic AMP to the two types of bindingsites associated with protein kinase II from bovinemyocardium. FEBS Letters, 129, 287–292.
8. Ogreid, D. & Doskeland, S. O. (1983). Cyclic nucleo-tides modulate the release of [3H] adenosine cyclic30,50-phosphate bound to the regulatory moiety ofprotein kinase I by the catalytic subunit of the kinase.Biochemistry, 22, 1686–1696.
9. Gibson, R. M., Ji-Buechler, Y. & Taylor, S. S. (1997).Interaction of the regulatory and catalytic subunitsof cAMP-dependent protein kinase. Electrostaticsites on the type Ialpha regulatory subunit. J. Biol.Chem. 272, 16343–16350.
10. Poteet-Smith, C. E., Shabb, J. B., Francis, S. H. &Corbin, J. D. (1997). Identification of critical determi-nants for autoinhibition in the pseudosubstrateregion of type I alpha cAMP-dependent proteinkinase. J. Biol. Chem. 272, 379–388.
11. Ringheim, G. E. & Taylor, S. S. (1990). Dissecting thedomain structure of the regulatory subunit ofcAMP-dependent protein kinase I and elucidatingthe role of MgATP. J. Biol. Chem. 265, 4800–4808.
12. Huang, L. J. & Taylor, S. S. (1998). Dissecting cAMPbinding domain A in the RIalpha subunit of cAMP-dependent protein kinase. Distinct subsites forrecognition of cAMP and the catalytic subunit.J. Biol. Chem. 273, 26739–26746.
13. Hoofnagle, A. N., Resing, K. A. & Ahn, N. G. (2003).Protein analysis by hydrogen exchange mass spec-trometry. Annu. Rev. Biophys. Biomol. Struct. 32, 1–25.
14. Engen, J. R. & Smith, D. L. (2001). Investigatingprotein structure and dynamics by hydrogenexchange MS. Anal. Chem. 73, 256A–265A.
15. Hamuro, Y., Coales, S. J., Southern, M. R., Nemeth-Cawley, J. F., Stranz, D. D. & Griffin, P. (2003). Rapidstructural analysis of protein therapeutics byhydrogen/deuterium exchange (H/D-Ex) massspectrometry. J. Biomol. Tech. 14, 171–182.
16. Baerga-Ortiz, A., Hughes, C. A., Mandell, J. G. &Komives, E. A. (2002). Epitope mapping of amonoclonal antibody against human thrombin by
H/D-exchange mass spectrometry reveals selectionof a diverse sequence in a highly conserved protein.Protein Sci. 11, 1300–1308.
17. Ehring, H. (1999). Hydrogen exchange/electrosprayionization mass spectrometry studies of structuralfeatures of proteins and protein/protein interactions.Anal. Biochem. 267, 252–259.
18. Engen, J. R., Gmeiner, W. H., Smithgall, T. E. &Smith, D. L. (1999). Hydrogen exchange showspeptide binding stabilizes motions in Hck SH2.Biochemistry, 38, 8926–8935.
19. Hamuro, Y., Wong, L., Shaffer, J., Kim, J. S., Stranz,D. D., Jennings, P. A. et al. (2002). Phosphorylationdriven motions in the COOH-terminal Src kinase,CSK, revealed through enhanced hydrogen–deuterium exchange and mass spectrometry(DXMS). J. Mol. Biol. 323, 871–881.
20. Englander, J. J., Del Mar, C., Li, W., Englander, S. W.,Kim, J. S., Stranz, D. D. et al. (2003). Protein structurechange studied by hydrogen–deuterium exchange,functional labeling, and mass spectrometry. Proc.Natl Acad. Sci. USA, 100, 7057–7062.
21. Mandell, J. G., Falick, A. M. & Komives, E. A. (1998).Identification of protein–protein interfaces bydecreased amide proton solvent accessibility. Proc.Natl Acad. Sci. USA, 95, 14705–14710.
22. Andersen, M. D., Shaffer, J., Jennings, P. A. & Adams,J. A. (2001). Structural characterization of proteinkinase A as a function of nucleotide binding. Hydro-gen–deuterium exchange studies using matrix-assisted laser desorption ionization-time of flightmass spectrometry detection. J. Biol. Chem. 276,14204–14211.
23. Anand, G. S., Hughes, C. A., Jones, J. M., Taylor, S. S.& Komives, E. A. (2002). Amide H/2H exchangereveals communication between the cAMP and cata-lytic subunit-binding sites in the R(I)alpha subunitof protein kinase A. J. Mol. Biol. 323, 377–386.
24. Hamuro, Y., Zawadzki, K. M., Kim, J. S., Stranz, D. D.,Taylor, S. S. & Woods, V. L. J. (2003). Dynamics ofcAPK type IIbeta activation revealed by enhancedamide H/2H exchange mass spectrometry (DXMS).J. Mol. Biol. 327, 1065–1076.
25. Bai, Y., Milne, S., Mayne, L. & Englander, S. W.(1993). Primary structure effects on peptide grouphydrogen exchange. Proteins: Struct. Funct. Genet. 17,75–86.
26. Kossiakoff, A. A. (1982). Protein dynamics investi-gated by the neutron diffraction-hydrogen exchangetechnique. Nature, 296, 713–721.
27. Potter, R. L. & Taylor, S. S. (1979). Relationshipsbetween structural domains and function in theregulatory subunit of cAMP-dependent proteinkinases I and II from porcine skeletal muscle. J. Biol.Chem. 254, 2413–2418.
28. Cheng, X., Phelps, C. & Taylor, S. S. (2001). Differen-tial binding of cAMP-dependent protein kinaseregulatory subunit isoforms Ialpha and IIbeta to thecatalytic subunit. J. Biol. Chem. 276, 4102–4108.
29. Anand, G. S., Law, D., Mandell, J. G., Snead, A. N.,Tsigelny, I., Taylor, S. S. et al. (2003). Identification ofthe protein kinase A regulatory RIapha-catalytic sub-unit interface by amide H/2H exchange and proteindocking. Proc. Natl Acad. Sci. USA, 100, 13264–13269.
30. Wu, J., Brown, S., Xuong, N. H. & Taylor, S. S. (2004).RIa Subunit of PKA: A cAMP-free Structure Revealsa Hydrophobic Capping Mechanism for DockingcAMP into Site B. Structure, 12, 1057–1065.
31. Burns, L. L., Canaves, J. M., Pennypacker, J. K.,
DXMS Analysis of cAPKI Activation 1195

Blumenthal, D. K. & Taylor, S. S. (2003). Isoformspecific differences in binding of a dual-specificityA-kinase anchoring protein to type I and type IIregulatory subunits of PKA. Biochemistry, 42,5754–5763.
32. Canaves, J. M. & Taylor, S. S. (2001). Classificationand phylogenetic analysis of the cAMP-dependentprotein kinase regulatory subunit family. J. Mol.Evol. 54, 17–29.
33. Hofmann, F., Beavo, J. A., Bechtel, P. J. & Krebs, E. G.(1975). Comparison of adenosine 3-monophosphate-dependent protein kinases from rabbit skeletal andbovine heart muscle. J. Biol. Chem. 250, 7795–7801.
34. Herberg, F. W. & Taylor, S. S. (1993). Physiologicalinhibitors of the catalytic subunit of cAMP-depen-dent protein kinase: effect of MgATP on protein–protein interactions. Biochemistry, 32, 14015–14022.
35. Vigil, D., Blumenthal, D. K., Heller, W. T., Brown, S.,Canaves, J. M., Taylor, S. S. & Trewhella, J. (2004).Conformational Differences Among Solution Struc-tures of the Type Ia, IIa and IIb Protein Kinase ARegulatory Subunit Homodimers: Role of the LinkerRegions. J. Mol. Biol., 337, 1183–1194.
36. Diller, T. C., Madhusudan, T. C., Xuong, T. C. &Taylor, T. C. (2001). Molecular basis for regulatorysubunit diversity in cAMP-dependent proteinkinase: crystal structure of the type II beta regulatorysubunit. Structure, 9, 73–82.
37. Zhang, Z. & Smith, D. L. (1993). Determination ofamide hydrogen exchange by mass spectrometry: anew tool for protein structure elucidation. ProteinSci. 2, 522–531.
Edited by P. Wright
(Received 30 March 2004; accepted 13 May 2004)
1196 DXMS Analysis of cAPKI Activation