motor grammar of swallowing - words-and-pictures.de grammar of... · 2 eine motorische grammatik...
TRANSCRIPT

1
A Motor Grammar of Swallowing12
© Norbert Rüffer, 2012
Korrespondenzadresse: Dr. Norbert Rüffer, Fuchsweg 10, 65719 Hofheim,
Abstract
Swallowing is characterised by a sequence of motor events in the oral, pharyngeal and
oesophageal space. This sequence is controlled by a complex motor system that includes
the Central Pattern Generator for Swallowing (CPGS), located in the ponto-medullary region
of the brain stem. In this paper we will discuss some facts that challenge the assumption that
neural control systems for iterative movements, such as swimming, walking or breathing
(iterative CPGs), can serve as a model for the CPGS (iterative CPGS hypothesis).
In contrast to the iterative CPGS hypothesis, we propose a processing model of motor
planning and control for swallowing that is based on two fundamental principles that are not
specific to swallowing: motor coordination and motor learning. Motor coordination is a
combinatory planning process generating sequentially and hierarchically organised motor-
temporal structures or motor plans. Being free within biomechanical and functional
constraints, motor coordination enables a range of more or less fixed motor-temporal
relations within the pharyngeal swallowing sequence and between bolus transit and
swallowing motor events, depending in part on bolus features.
Automaticity of swallowing is based on motor learning, that is, improvement of motor
coordination with respect to bolus transit and airway protection, long-term storage of the
resulting motor-temporal patterns and activation of these structures. The CPGS is not an
iterative CPG but plays the role of the motor memory for the swallowing system.
The current work does not present new data but rather provides a new interpretation of data
recently published by Kendall, Mendell, Logemann, Martin-Harris and others on the motor-
temporal structure of the pharyngeal motor sequence.
1 This paper is based partly on presentations at the 6th World Congress for NeuroRehabilitation (WCNR), March 21–25, 2010, in Vienna, Austria, and at an expert meeting on ‘Motor Grammar and Presbyphagia’ at the University of Bielefeld, April 16, 2011. 2 I want to thank Andrea Hofmayer and Sönke Stanschus for important comments.

2
Eine motorische Grammatik des Schluckens
Schlucken ist durch eine Sequenz motorischer Aktionen im oralen, pharyngealen und
ösophagealen Raum charakterisiert, die durch ein komplexes motorisches System
kontrolliert wird, das den in der ponto-medullären Region des Hirnstamms lokalisierten
Central Pattern Generator des Schluckens (CPGS) einschließt. Wir diskutieren Fakten, die
die Annahme in Frage stellen, dass neuronale Kontrollsysteme für iterative Bewegungen wie
schwimmen, laufen oder atmen (iterative CPGs) ein geeignetes Modell für den CPGS
darstellen (iterative CPGs-Hypothese).
Als Alternative zur iterativen CPGs-Hypothese wird ein Verarbeitungsmodell der motorischen
Planung und Kontrolle des Schluckens vorgeschlagen, das auf zwei fundamentalen
Prinzipien basiert, die nicht schluckspezifisch sind: motorische Koordination und motorisches
Lernen. Motorische Koordination ist ein kombinatorischer Planungsprozess, der sequentiell
und hierarchisch organisierte motorisch-zeitliche Strukturen oder Motorpläne generiert. Die
auf motorischer Koordination basierende motorische Planung ist im Rahmen
biomechanischer und funktionaler Outputbedingungen variabel und ermöglichen ein
Spektrum von mehr oder weniger fixierten, zum Teil von Boluseigenschaften abhängigen
motorisch-zeitlichen Relationen innerhalb der pharyngealen Schlucksequenz und zwischen
Bolustransit und Schluckmotorik.
Die Automatizität des Schluckens basiert auf motorischem Lernen, d.h. einer Optimierung
der schluckmotorischen Koordination im Hinblick auf den Bolustransport und die
Atemwegsprotektion, der Speicherung dieser optimierten motorisch-zeitlicher Strukturen
sowie deren Aktivierung. Der CPGS ist kein iterativer Mustergenerator, sondern fungiert als
motorischer Speicher des Schlucksystems.
Die vorliegende Arbeit präsentiert keine neuen Daten, sondern re-interpretiert aktuelle Daten
von Kendall, Mendell, Logemann, Martin-Harris und anderen zur motorisch-zeitlichen
Struktur der pharyngealen Schlucksequenz.
Keywords: Central Pattern Generator for Swallowing (CPGS), processing model of
swallowing, motor planning of swallowing, motor coordination, motor learning, biomechanical
and functional constraints of swallowing
Schlüsselwörter: Central Pattern Generator des Schluckens (CPGS), Verarbeitungsmodell
des Schluckens, motorische Planung des Schluckens, motorische Koordination, motorisches
Lernen, biomechanische und funktionale Beschränkungen des Schluckens

3
The Central Pattern Generator for Swallowing
Swallowing is characterised by a sequence of motor events in the oral, pharyngeal and
oesophageal space that is controlled by a complex motor system, which includes cortical,
subcortical und brain stem areas. Kendall and colleagues proposed the following pharyngeal
swallowing sequence based on videofluoroscopy data (Kendall et al., 2000):
Spstart>Aestart>H1>SPmax>Aeclose>PESop>H2>PESmax>HL>H3>Pamax>PEScl
SPstart=soft palate begins to elevate; Aestart=aryepiglottic folds begin to elevate; H1=hyoid bone begins to elevate;
Spmax=soft palate has reached maximal elevation; Aeclose=aryepiglottic folds contact the epiglottis;
PESop=pharyngoesophageal sphincter (PES) begins to open; H2=hyoid bone has reached maximal elevation; PESmax=PES
reaches maximum distension; HL=larynx approximates the hyoid bone; H3=hyoid begins to descend; Pamax=maximum
pharyngeal constriction; PEScl=PES closes
According to this sequence, pharyngeal swallowing starts with the motor events soft palate
and aryepiglottic folds elevation (SPstart, Aestart), moves through hyoid and laryngeal
elevation (H1-3, HL) in association with laryngeal closure at the epiglottis level and
pharyngeal contraction (Pamax), and terminates in pharyngoesophageal sphincter
relaxation, opening and finally closure (PESop, PESmax, PEScl) after the bolus has reached
the oesophagus.
The Central Pattern Generator for Swallowing (CPGS), which is located in the ponto-
medullary region of the brain stem, plays an important role in neural control of swallowing.
While neural control of swallowing is not restricted to the CPGS, it is nevertheless based to a
large extent on the CPGS network. It is assumed that the CPGS generates a basic motor
plan for pharyngeal swallowing which can be modulated by superior levels of the swallowing
motor system (see Daniels&Huckabee, 2008, p. 19ff for an overview).
Although there has been no attempt to clarify the concept of the CPGS ‘basic motor plan’ to
date, there is a prominent few in dysphagia research, suggesting that the CPGS output can
be identified with a rhythmic pattern that serves as a frame for the motor-temporal structure
of swallowing. The following statement by Lang is typical in this respect (Lang, 2009, p. 340,
underlines added):
‘Electrophysiologic studies have found that neurons of the brain stem contain the timing pattern-
generating circuitry that governs the oral, pharyngeal, and esophageal phases of swallowing.
One can find neurons that respond at a time delay and duration that corresponds to the expected
timing of rhythmical movements of the jaw or peristalsis of the pharynx or esophagus. These neural
events are not caused by feedback from periphery, e.g., propagating peristalsis, because this pattern

4
persists even in paralyzed animals. Therefore, these neural events found in premotor neurons must
control the timing of these phases of swallowing...
Swallowing is initiated by a voluntary act but much of this process is composed of stereotyped motor
activity that is controlled by brain stem central pattern generators.’
According to Lang, the CPGS is a ‘timing pattern-generating circuitry’ that generates a
‘rhythmical’ and ‘stereotyped motor activity’ without being ‘caused by feedback from
periphery’. The CPGS function is viewed as being similar to Central Pattern Generators in
the sense of neural control systems for rhythmic and repetitive movements such as
swimming, walking or breathing. We will call Central Pattern Generators in this sense
iterative CPGs.
What are iterative CPGs? Iterative CPGs are neural networks that function as a biological
oscillator generating a rhythmical pattern. They may differ in neural architecture and rhythmic
pattern output as shown by the examples of the leech heartbeat CPG and the lobster pyloric
CPG in Hooper (2001):
Fig. 1. Neural network and rhythmic pattern output of central pattern generators; Hooper, 2001, p. 3, Fig. 1 Permission for reprint by John Wiley & Sons Ltd.
Iterative CPGs are related to motor systems and the CPG-generated rhythmic pattern serves
as a frame for the temporal relations of the motor activity output of the respective motor
system. Sensory integration may modify the CPG rhythmic pattern, which in turn would lead
to temporal modification of the correlated motor activity. An example is the locust flying CPG,
in which sensory feedback of wing elevation affects cycle period and burst duration of the
generated rhythmic pattern, which in turn affects the flying behaviour of the locust (Hooper,
2001).

5
Although the role of iterative CPGs within motor systems has only been partly explored, the
available data seem to support the following modular model. Motor systems generate motor
activities by selecting a set of muscles and activating this set in a particular temporal order:
Fig. 2. Motor-temporal control of iterative CPGs embedded in motor systems
Within the resulting motor-temporal sequence, the rhythmic pattern of an associated iterative
CPG translates into a frequency pattern related to shorter or longer intervals between
designated time points of the motor-temporal structure. Iterative CPGs control the intervals
between onset times of distinct muscles of the sequence, intervals between onset and offset
times of identical muscles (activation lengths) and define the onset and offset time of an
iteration cycle (motor-temporal loop). Iterative CPGs temporally modify motor activities but do
not change them in a qualitative way, since the effects of iterative CPGs are limited to
particular selection of a set of muscles and their activation sequence. Qualitative change of
motor activity is based on a different selection of muscles and their activation at the motor
system level.
Lateral turns in lampreys provide an example. In lampreys the swimming CPG is located in
the spinal cord and is neuroanatomically arranged as a sequence of spinal segments, which
each control a set of muscles called a myotome. Swimming results from laterally coordinated

6
waves of myotomal contractions along both sides of the fish’s body. Fagerstedt & Ullén
(2001) have shown that if the lamprey executes a lateral turn in response to a stimulus on
one side of the fish, this motor activity is the result of a lateral change in the swimming motor
sequence caused by longer cycle duration together with increasing burst duration, burst
proportion (within the movement cycle) and burst intensity. Apart from the change in burst
intensity, lateral turns in lampreys can be described as connected to a change in frequency:
cycle duration, burst duration and burst proportion are related to the intervals between onset
times and to activation lengths. That there is also an effect on burst intensity, that is, motor
force, does not contradict the assumption that iterative CPGs primarily modify the temporal
structure of motor activities, since temporal motor features and motor force are closely
related for biomechanical reasons. An increase in the frequency of iterative movements
normally requires an increase in the motor force.
Although the assumption that swallowing is controlled by an iterative CPG – which we will
call the iterative CPGS hypothesis – seems at first sight to have certain plausibility because
both iterative motor sequences and the pharyngeal swallowing sequence show a high
degree of automaticity, the facts contradict this assumption. If the CPGS neural control of
swallowing were similar to iterative CPG motor control, we would expect a similar structure
for neural control in both cases. For swallowing this would imply that swallowing should have
an iterative motor-temporal structure and that motor-temporal changes due to sensory
integration of bolus properties should be related to a change in frequency, that is, change in
the intervals within a certain selection and activation order of motor events.
Five problems with the iterative CPGS hypothesis
In the following, some facts will be discussed that challenge the assumption that iterative
CPGs are an appropriate model for the swallowing CPG. There are five main problems with
the iterative CPGS hypothesis:
Table 1. Iterative CPGs vs. CPGS
Iterative CPGs Five problems with
the iterative CPGS hypothesis
Generate iterative cycles (motor-temporal
loops)
1. The swallowing sequence has – with the
exception of mastication – no iterative
structure

7
Relate sensory integration to frequency in
the sense of shorter or longer intervals
between designated time points in a motor
sequence including onset times, activation
length and iterative cycle
2. Sensory integration of bolus properties is
only in part linked to change in frequency
Control the temporal structure of motor
activities within a certain activation sequence
3. The activation order of the swallowing
sequence is only partly fixed
4. It varies with bolus properties
5. It varies with bolus position
Problem no. 1: The pharyngeal swallowing sequence does not have an iterative motor-
temporal structure
As the pharyngeal swallowing sequence of Kendall et al. (2000) shows, swallowing is
composed of distinct motor events (swallowing gestures) that are activated in a particular
sequence with no repetition and therefore has no iterative structure:
Spstart>Aestart>H1>SPmax>Aeclose>PESop>H2>PESmax>HL>H3>Pamax>PEScl
SPstart=soft palate begins to elevate; Aestart=aryepiglottic folds begin to elevate; H1=hyoid bone begins to elevate;
Spmax=soft palate has reached maximal elevation; Aeclose=aryepiglottic folds contact the epiglottis;
PESop=pharyngoesophageal sphincter (PES) begins to open; H2=hyoid bone has reached maximal elevation; PESmax=PES
reaches maximum distension; HL=larynx approximates the hyoid bone; H3=hyoid begins to descend; Pamax=maximum
pharyngeal constriction; PEScl=PES closes
Of the swallowing motor events only mastication is iterative but since mastication is subject
to a greater degree of voluntary control, it seems to be a questionable example of
neurological control by the CPGS.
Problem no. 2: Sensory integration of bolus properties is only in part linked to change in
frequency
With the assumption that sensory integration of bolus properties (volume,
viscosity/consistency) is linked to a modification of frequency, we would expect bolus

8
properties to temporally affect onset times and activation lengths of the pharyngeal motor
events within a certain activation order.
The data in Kendall et al. (2000) partly support this expectation:
Fig. 3. Swallowing gesture timing related to increasing bolus volume: 1-ml liquid vs. 20-ml liquid; Kendall et al. 2000, adapted from Fig. 5, p. 81 B1=the first movement of the bolus from a stable or “hold” position that passes the posterior nasal spine, Spstart=soft palate begins to elevate, Aestart=aryepiglottic folds begin to elevate, H1=hyoid bone begins to elevate, Spmax=soft palate has reached maximal elevation, Aeclose=aryepiglottic folds contact the epiglottis, PESop=PES begins to open, H2=hyoid bone has reached maximal elevation, PESmax=PES reaches maximum distension, H3=hyoid begins to descend, PAmax=maximum pharyngeal constriction, PEScl=PES closes
Most of the expected motor events show simultaneously earlier onset, target and reset times
with increasing bolus volume related to faster oropharyngeal bolus transit (relative to the
bolus head).
However, a closer look at the data reveals that
some motor events differ with respect to the degree of temporal change caused by
increasing bolus volume (e.g., SPstart vs. PESop), and
some show no temporal change at all (H1)
The data show that subsequences of the pharyngeal swallowing sequence may behave
differently with respect to sensory integration of bolus properties. This is an unexpected fact
if it is assumed that sensory integration is linked to frequency change since frequency
change should affect the pharyngeal swallowing sequence in a continuous way.
9
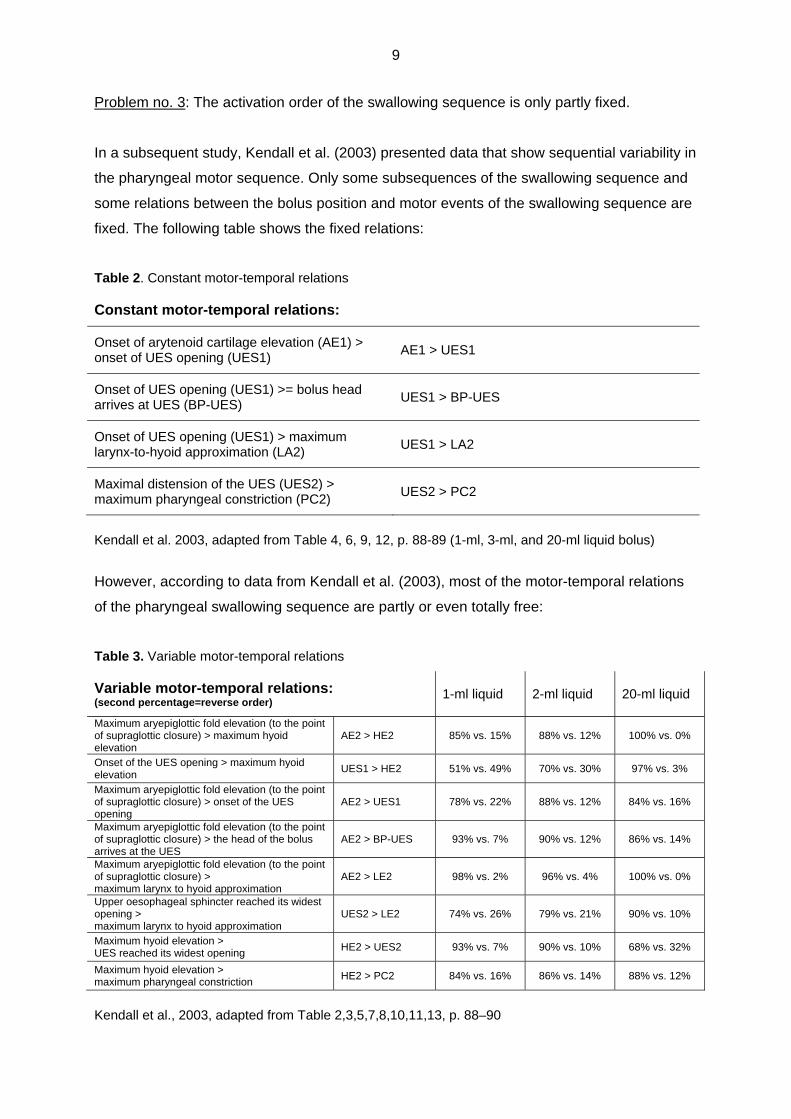
Problem no. 3: The activation order of the swallowing sequence is only partly fixed.
In a subsequent study, Kendall et al. (2003) presented data that show sequential variability in
the pharyngeal motor sequence. Only some subsequences of the swallowing sequence and
some relations between the bolus position and motor events of the swallowing sequence are
fixed. The following table shows the fixed relations:
Table 2. Constant motor-temporal relations
Constant motor-temporal relations:
Onset of arytenoid cartilage elevation (AE1) > onset of UES opening (UES1)
AE1 > UES1
Onset of UES opening (UES1) >= bolus head arrives at UES (BP-UES)
UES1 > BP-UES
Onset of UES opening (UES1) > maximum larynx-to-hyoid approximation (LA2)
UES1 > LA2
Maximal distension of the UES (UES2) > maximum pharyngeal constriction (PC2)
UES2 > PC2
Kendall et al. 2003, adapted from Table 4, 6, 9, 12, p. 88-89 (1-ml, 3-ml, and 20-ml liquid bolus)
However, according to data from Kendall et al. (2003), most of the motor-temporal relations
of the pharyngeal swallowing sequence are partly or even totally free:
Table 3. Variable motor-temporal relations
Variable motor-temporal relations: (second percentage=reverse order)
1-ml liquid 2-ml liquid 20-ml liquid
Maximum aryepiglottic fold elevation (to the point of supraglottic closure) > maximum hyoid elevation
AE2 > HE2 85% vs. 15% 88% vs. 12% 100% vs. 0%
Onset of the UES opening > maximum hyoid elevation
UES1 > HE2 51% vs. 49% 70% vs. 30% 97% vs. 3%
Maximum aryepiglottic fold elevation (to the point of supraglottic closure) > onset of the UES opening
AE2 > UES1 78% vs. 22% 88% vs. 12% 84% vs. 16%
Maximum aryepiglottic fold elevation (to the point of supraglottic closure) > the head of the bolus arrives at the UES
AE2 > BP-UES 93% vs. 7% 90% vs. 12% 86% vs. 14%
Maximum aryepiglottic fold elevation (to the point of supraglottic closure) > maximum larynx to hyoid approximation
AE2 > LE2 98% vs. 2% 96% vs. 4% 100% vs. 0%
Upper oesophageal sphincter reached its widest opening > maximum larynx to hyoid approximation
UES2 > LE2 74% vs. 26% 79% vs. 21% 90% vs. 10%
Maximum hyoid elevation > UES reached its widest opening
HE2 > UES2 93% vs. 7% 90% vs. 10% 68% vs. 32%
Maximum hyoid elevation > maximum pharyngeal constriction
HE2 > PC2 84% vs. 16% 86% vs. 14% 88% vs. 12%
Kendall et al., 2003, adapted from Table 2,3,5,7,8,10,11,13, p. 88–90

10
In the table the second percentage shows the reverse order of the respective subsequence.
The data demonstrate a range of different degrees of sequential variability, from relatively
fixed to relatively or even completely free. In most cases variability was greater with small
bolus volumes.
There was no constant complete pharyngeal motor sequence with all swallows and all bolus
volumes. The most common sequence of a certain length
AE2>UES1>HE2>UES2>LE2>PC2
was associated with only 25% of the observed swallows.
Variability in the pharyngeal motor sequence cannot be expected under the iterative CPGS
hypothesis, because iterative CPGs control the temporal structure of motor events within a
certain activation sequence generated by the connected motor system.
Problem no. 4: The activation order of the swallowing sequence varies with bolus properties.
Comparable to the data of Kendall et al. (2003), Mendell & Logemann (2007) found a partly
variable and, with respect to order, bolus-dependent pharyngeal swallowing sequence:
Table 4. Frequency of temporal sequences observed for each bolus type
Sequence 3-ml liquid 10-ml liquid 1-3-ml paste
LE-HE-BOT-LC 24 8 20
HE-LE-BOT-LC 23 12 42
LE-HE-LC-BOT 17 38 2
HE-LE-LC-BOT 14 25 7
(HE=LE)-BOT-LC 13 2 23
Each cell represents the number of participants who followed this pattern (n=100). LE=onset of laryngeal elevation, defined as the first displacement in movement at the area of the posterior vocal folds and arytenoid area, HE=onset of hyoid elevation, BOT=onset of tongue base movement toward the posterior pharyngeal wall, LC=first frame showing complete laryngeal closure; Mendell & Logemann, 2007, adapted from Table 4, p. 1264

11
The table shows variable subsequences between the onsets of hyoid elevation, tongue base
retraction and laryngeal elevation, which was identified by arytenoid movement, as well as
complete laryngeal closure. The sequential order was rather free with low bolus volume and
viscosity and shows a more fixed order with increasing bolus volume and viscosity.
Of particular interest was the activation order of tongue base retraction, laryngeal closure
and UES opening:
Fig. 4. Fronting of laryngeal closure and UES opening with increasing bolus volume. HE=onset of hyoid elevation, BOT=onset of tongue base movement toward the posterior pharyngeal wall, LC=first frame showing complete laryngeal closure, UES=onset of upper esophageal sphincter (UES) opening; Mendell & Logemann, 2007, adapted from Table 4+5, p. 1264 (3-ml liquid, 10-ml liquid, 1-3-ml paste)
With low bolus volume the activation order of these motor events corresponds to the order of
swallow phases and the sequence was mostly initiated by oral tongue base retraction and
terminated by pharyngeal UES opening. If the bolus volume increases, however, the Mendel
& Logemann data show that this may speed up the motor response by means of motor-
temporal fronting of UES opening (UES) and laryngeal closure (LC) before the onset of
tongue base retraction (BOT). Such structures do respond to high bolus volumes causing
accelerated bolus transit with a combination of earlier airway protection and earlier UES-
opening. Motor-temporal transformations cannot be expected with the iterative CPGS
hypothesis, since they change the activation order of a motor sequence beyond modification
of internal time intervals. Fronting of UES opening and laryngeal closure generates a
qualitatively different pharyngeal sequence, which is specialised with respect to high bolus
volume and fast bolus transit.
Problem no. 5: The activation order of the swallowing sequence varies with the bolus
position.
Unlike the widespread concept of ‘delayed pharyngeal swallow’, the swallow onset,
identifiable by the onset of hyoid movement, is not invariably synchronised with trigger areas

12
in the posterior oral space. Up to 80% of normal subjects swallow late, that is, with
pharyngeal bolus positions at swallow onset but without subsequent decrease in airway
protection (Martin-Harris et al., 2007). Pharyngeal bolus positions at swallow onset are a
normal feature of non-instructed spontaneous swallows (Dua et al., 1997), of swallows with
bolus accumulation in the valleculae during mastication (Hiiemae & Palmer, 1999), of
sequential swallows (Daniels & Foundas, 2001) and of swallows of healthy elderly (Leonard
& MCKenzie, 2006). Therefore, it is necessary to distinguish between late swallows with
pharyngeal bolus positions at swallow onset and sufficient airway protection and delayed
swallows with pharyngeal bolus positions at swallow onset and reduced airway protection.
Late swallows without reduced airway protection are possible because there is an early level
of airway protection that is independent of epiglottic closure (Rüffer, 2012):
Fig. 5. Early airway protection in case of late swallows; Rüffer, 2012, adapted from Fig. 9, p. 52

13
This early level of airway protection is based on several factors:
Anatomy: Spaces within the pharynx (valleculae, lateral pharyngeal channels, pyriform sinuses) are able to contain bolus material that has reached the pharynx before swallow onset thereby reducing the probability of penetration.
Dwell time: There is a time slot between the time point where bolus material enters the pharynx before swallow onset and decrease of airway protection (Dua et al., 1997).
Arytenoid movement: Early onset of arytenoid adduction and elevation reconfigures the laryngeal vestibule and its surroundings and increases airway protection at the biomechanical level (Dua et al., 1997).
Swallow-related apnoea: Early onset of swallow-related apnoea prevents aspiration of bolus material (Martin-Harris et al., 2007).
Of particular interest with respect to the connection between the bolus position and the
activation order of the swallowing sequence is arytenoid movement. While full arytenoid
adduction and elevation is localised after the onset of hyoid elevation and results in glottic
closure as well as arytenoid to epiglottic closure, the onset of arytenoid adduction and
elevation is shifted before the onset of hyoid elevation in case of late swallows.3 This early
movement leads to modification of the laryngeal vestibule and its surroundings by widening
the lateral pharyngeal channels, closing the interarytenoid space and heightening the barrier
of the aryepiglottic folds. It contributes to airway protection at the biomechanical level
because it reduces the probability that liquid boluses entering the hypopharynx before
swallow onset will overflow into the laryngeal vestibule. Together with the other factors in
early airway protection, fronting of arytenoid movement enables late swallows with sufficient
airway protection.
As late swallows demonstrate, the activation order of the pharyngeal swallowing sequence
may vary with bolus position. Both bolus properties and bolus position may induce motor-
temporal change of the pharyngeal swallowing sequence which goes beyond modification of
temporal distances within a certain activation order, a fact that is not expected under the
iterative CPGS hypothesis.
The data from Kendall, Mendell & Logemann and others challenge the assumption that the
CPGS is similar to iterative CPGs. The motor-temporal structure of swallowing has features
that are not expected with the assumption of iterative CPG control. The pharyngeal 3 Data from Mendell & Logemann (2007) provide evidence of a variable activation order for the onsets of arytenoid elevation and hyoid elevation in case of small bolus volume (see Table 4). We hypothesize that – apart from the influence of bolus volume – this variable order of the onsets of arytenoid movement and hyoid elevation is a feature of early swallows. It can be assumed that Mendell & Logemann saw early swallows mainly because they observed instructed swallows in a videofluoroscopy setting.

14
swallowing sequence does not include iterative loops, is in part variable, and bolus properties
as well as bolus position may induce motor-temporal transformations of the sequence. The
data show that the motor-temporal structure of swallowing is much more variable and
context-sensitive than one would expect with the assumption of the iterative CPGS
hypothesis. Since variability and context sensitivity are features of voluntary movement,
pharyngeal swallowing seems to be closer to voluntary movement than usually expected. It is
voluntary movement and not reflexes or iterative movements that provide an adequate
blueprint for the pharyngeal swallow.
Elements of a new model for motor processing of swallowing
Let me focus on the question of how to explain the facts challenging the iterative CPGS
hypothesis. Unlike the iterative CPG model, we want to propose a model of motor planning of
swallowing that is based on two fundamental principles of motor processing which are not
specific to swallowing, namely motor coordination and motor learning:
Principles of motor processing
1. motor coordination
2. motor learning
We assume that there is no difference between swallowing and other motor activities with
respect to the basic principles of motor processing, which we identify with motor coordination
and motor learning. As the following assumptions show, the combined effects of motor
coordination und motor learning can explain the data that are problematic for the iterative
CPGS hypothesis:
1. Motor coordination
The neurological control of swallowing is based on a combinatory motor planning
process that coordinates muscle activities in a sequential and hierarchical way and
thus generates more complex muscle activities.
This can explain why sensory factors (bolus properties) may be correlated to
subsequences of the pharyngeal sequence (Kendall et al., 2000). Pharyngeal
subsequences correspond to intermediate levels of motor coordination.
2. Selective conditions for motor coordination
Swallowing motor coordination is free within selective conditions, leaving a range of
possible variations (voluntary movement as blueprint).
This can explain why the pharyngeal swallowing sequence is only fixed in part and

15
may be subject to motor-temporal transformations (Kendall et al., 2003;
Mendell&Logemann, 2007).
3. Motor learning
Automaticity of swallowing is the result of motor learning.
This can explain why the pharyngeal swallow is reflexive without assuming that it is
controlled by an iterative CPG.
A closer look at 1–3 may clarify the concepts behind the proposed model for motor planning
of swallowing:
1. Motor coordination
Motor activities of a certain complexity are based on motor planning. Motor planning means
that a motor activity is not simply a response to a particular stimulus but that it is mediated by
some kind of computational process which integrates perception but does not map it directly
to motor activity. Elementary motor systems like reflexes are characterised by direct
stimulus-response mapping without motor planning. Possible evidence for motor planning
derives from the computational complexity and flexibility of a motor activity, although even
unspectacular motor activities such as pointing, for example, are presumably rather complex
(Shadmehr&Wise, 2005). We do not believe that motor planning presupposes voluntary
control. Speaking is an example of a motor activity that is certainly planned but nevertheless
not voluntarily controlled, at least under normal circumstances. Apart from the complexity
and flexibility of a motor activity, motor planning is also indicated by the ability of a motor
system to control and correct its output, which presupposes something like a comparison of a
motor plan with an actual motor output. Examples are preservation of function despite
developmental or age-related change in the biomechanical, neuromotor or sensory features
of a motor system (for example, change in biomechanical context due to growth). Further
possible evidence in support of motor planning is found in motor planning errors, that is,
apraxia. In the case of speech apraxia, speech movements are affected at the motor-
temporal planning level whereas the muscle level of speech is, in contrast to dysarthria,
intact (Ziegler & Cramon, 1986). Although we do not believe that motor planning is restricted
to cortical processing, if it can be demonstrated using an imaging technique – for example,
fMRI – that a motor activity relies on cortical processing, this could also be interpreted as
evidence of motor planning. This is particularly true if cortical processing precedes a
connected motor activity, which can be demonstrated with EEG data showing a pre-motor
potential (readiness potential (RP), Kornhuber & Deecke, 1965). Pre-activation is not a
requisite feature of motor planning, however, since motor planning can be nearly

16
simultaneous with motor output as is true in the case of language planning and execution of
speaking (‘incremental production’, Levelt, 1989, pp 24–27). The swallowing motor system
has all the hallmarks of motor planning. It is able to flexibly synchronise its motor output with
bolus properties; it can control and correct its motor output, preserving its functionality even if
the biomechanical and neuromotor environment undergoes change, which is true both in the
development (Weckmueller et al., 2010) and ageing of the swallowing system
(Wilmskötter&Stanschus, 2012); it can be affected by motor planning errors (Huckabee &
Kelly, 2006); and it is at least in part controlled by cortical processing (Hamdy et al., 1999).
There is also some evidence that pre-activation is relevant for swallowing, at least for
volitional swallowing (Huckabee et al., 2003).
We assume that motor planning of swallowing (as well as motor planning in general) is
localised within a processing system that includes a motor grammar, which generates motor-
temporal structures or motor plans and that the swallowing motor response is the result of
mapping a motor plan into muscle activity:
Fig. 6. Motor processing
The planning level of a complex motor activity like swallowing must at least represent a
selection of a set of muscles along with temporal specification of the activation of each
muscle within the set. Motor planning is primarily a temporal process since what makes
muscles interact is their being simultaneously active. We therefore assume a syntactic rule
coordinate, generating motor plans by linking motor events with the same time feature:
17
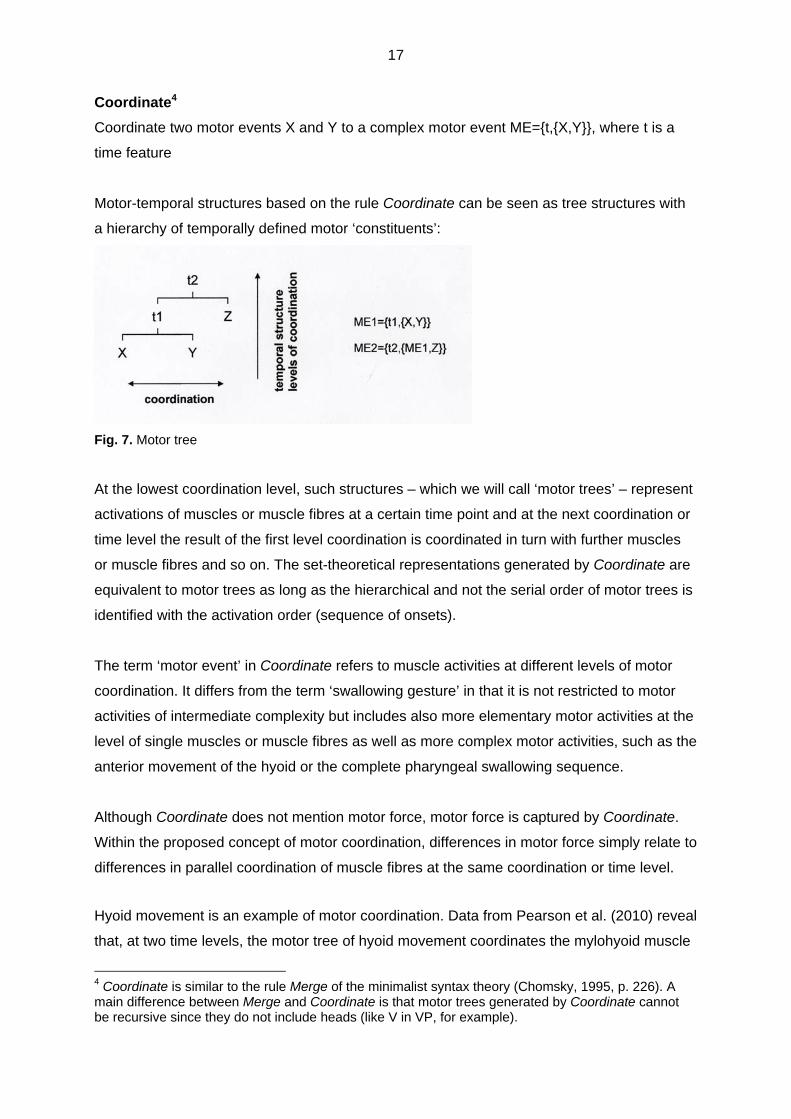
Coordinate4
Coordinate two motor events X and Y to a complex motor event ME={t,{X,Y}}, where t is a
time feature
Motor-temporal structures based on the rule Coordinate can be seen as tree structures with
a hierarchy of temporally defined motor ‘constituents’:
Fig. 7. Motor tree
At the lowest coordination level, such structures – which we will call ‘motor trees’ – represent
activations of muscles or muscle fibres at a certain time point and at the next coordination or
time level the result of the first level coordination is coordinated in turn with further muscles
or muscle fibres and so on. The set-theoretical representations generated by Coordinate are
equivalent to motor trees as long as the hierarchical and not the serial order of motor trees is
identified with the activation order (sequence of onsets).
The term ‘motor event’ in Coordinate refers to muscle activities at different levels of motor
coordination. It differs from the term ‘swallowing gesture’ in that it is not restricted to motor
activities of intermediate complexity but includes also more elementary motor activities at the
level of single muscles or muscle fibres as well as more complex motor activities, such as the
anterior movement of the hyoid or the complete pharyngeal swallowing sequence.
Although Coordinate does not mention motor force, motor force is captured by Coordinate.
Within the proposed concept of motor coordination, differences in motor force simply relate to
differences in parallel coordination of muscle fibres at the same coordination or time level.
Hyoid movement is an example of motor coordination. Data from Pearson et al. (2010) reveal
that, at two time levels, the motor tree of hyoid movement coordinates the mylohyoid muscle
4 Coordinate is similar to the rule Merge of the minimalist syntax theory (Chomsky, 1995, p. 226). A main difference between Merge and Coordinate is that motor trees generated by Coordinate cannot be recursive since they do not include heads (like V in VP, for example).

18
of the suprahyoid muscle group, which seems to be primarily responsible for the superior
movement of the hyoid, with the geniohyoid muscle of the suprahyoid muscle group, on
which the anterior movement of the hyoid seems to be primarily based:
Fig. 8. Motor tree of hyoid movement
2. Conditions for motor coordination
We assume that motor coordination of swallowing at the planning level is free within certain
constraints. Constraints on swallowing motor coordination serve as output conditions that
filter out motor-temporal structures if they are lacking obligatory motor features. There are
two types of conditions for motor coordination: biomechanical and functional.
Biomechanical conditions for swallowing motor coordination
Motor systems are embedded into biomechanical systems or, to use Rodney Brooks’ term
from robotics, motor systems are ‘embodied’ (Brooks, 2002, p. 51). This embodiment has far-
reaching consequences for motor systems, because it makes both the output of motor
systems and motor coordination at the planning level depend on biomechanical features:
Fig. 9. Biomechanical effects on motor output and motor coordination
Laryngeal closure at the epiglottic level may illustrate the dependence of the output of the
swallowing system on biomechanics. Epiglottic closure is a complex biomechanical process

19
depending on the biomechanical features of the hyoid, larynx and epiglottis but at the muscle
level it is initiated simply by contraction of the suprahyoid musculature.
The effects of embodiment are not restricted to the output of the swallowing system however
because they also affect motor coordination at the planning level. The reason is that
biomechanical hardware and neuromotor control of motor systems must be adjusted. If there
is no adjustment between the biomechanical hardware and neuromotor control of a motor
system, there will be a decrease in motor optimality at least, if not a decrease in function.
Motor optimality is an important concept in the field of sports science (training theory) and
robotics that is also important for swallowing but has not yet been systematically applied to
swallowing (see Todorov, 2006 for an overview). Optimal movements – or ‘physiological
movements’ – are characterised by maximising temporal synchronisation and minimising
motor force. An example of the effects of embodiment on swallowing motor coordination is
that the onset of hyoid movement must take into account the biomechanically caused
temporal costs of the laryngeal movement. The activation order of the pharyngeal motor
sequence is also in part dependent on biomechanics:
Biomechanical conditions for swallowing coordination are evident with motor events
that are biomechanically dependent in a direct way. Hyoid elevation (HE) should
therefore precede laryngeal elevation (LE), epiglottic closure (EC) and UES opening
(UES): HE > LE > EC/UES
Other biomechanical conditions for swallowing coordination are related to the space
within which swallowing muscles must act. Epiglottic closure (EC) should therefore
precede pharyngeal constriction (PC), since early pharyngeal constriction would
biomechanically obstruct epiglottic closure: EC > PC
There are also biomechanical conditions related to pressure. Soft palate closure
(SPC) should therefore precede tongue base retraction (BOT) to enable tongue base
retraction generating enough pressure for bolus transit: SPC > BOT
Functional conditions for swallowing motor coordination
Although it is difficult to draw a sharp line between biomechanical and functional conditions
on swallowing motor coordination because biomechanical conditions also contribute to
swallowing functionality, it makes sense to differentiate between conditions for swallowing
that depend on the embodiment of the swallowing motor system and conditions that do not.
Biomechanically independent functional conditions for swallowing motor coordination are
related to synchronisation of bolus transit with the sequence of swallowing motor events.

20
This synchronisation should at least link bolus arrival at the laryngeal vestibule with laryngeal
closure at some level (airway protection) and bolus arrival at the UES with UES opening
(bolus transport). We assume hypothetically that this minimal requirement fulfils swallowing
functionality:
Fig. 10. Selective synchronisation Selective synchronisation implies a clustering of temporally fixed motor temporal relations
around the synchronisation points of laryngeal closure and UES opening and enables more
free motor coordination elsewhere. Martin-Harris et al. (2003) were the first to discover
motor-temporal clustering of the pharyngeal swallowing sequence with respect to related
respiratory events. Selective synchronisation should lead to further clustering with respect to
bolus transit.
Selective synchronisation enables sensory integration of bolus properties by transforming the
pharyngeal swallowing sequence. Fronting of UES opening and laryngeal closure in case of
high volume in the data of Mendell & Logemann (2007) is an example of this (see Fig. 4).
The swallowing system can respond to a change in bolus transit by varying bolus properties
with transformations of the pharyngeal swallowing sequence to preserve selective
synchronisation of laryngeal closure and UES opening with the bolus transit.
Selective synchronisation allows variable bolus positions at swallow onset as long as airway
protection is preserved (‘larynx closure at some level’). Although in conflict with the common
assumption in dysphagia research that swallows with pharyngeal bolus positions are always
delayed, this implication of selective synchronisation is supported by data showing that late
swallows with pharyngeal bolus positions and sufficient airway protection are a common
feature of normal swallowing (Martin-Harris et al., 2007; Dua et al., 1997; Hiiemae&Palmer,
1999; Daniels&Foundas, 2001; Leonard&McKenzie, 2006). As we have seen, late swallows

21
with sufficient airway protection are possible because there is an early level of airway
protection independent of epiglottic closure which is based in part on fronting of arytenoid
movement (see Fig. 5).
The fixed relations of the Kendall et al. (2003) data can be explained by the proposed
functionality of selective synchronisation of bolus transit with laryngeal closure and UES
opening together with biomechanical conditions:
Constant motor-temporal relations (Kendall et al., 2003)
Explanation in terms of conditions for motor coordination
Onset of arytenoid cartilage elevation (AE1) > onset of UES opening (UES1)
AE1 > UES1
Functional condition: Since the bolus reaches the laryngeal vestibule before UES for anatomical reasons, laryngeal closure – particularly at the arytenoid level – should precede UES opening to fulfil swallowing functionality with respect to airway protection
Onset of UES opening (UES1) >= bolus head arrives at UES (BP-UES)
UES1 >= BP-UES
Functional condition: the arrival of the bolus at the UES should be synchronised with UES opening to fulfil swallowing functionality with respect to bolus transport
Onset of UES opening (UES1) > maximum larynx-to-hyoid approximation (LA2)
UES1 > LA2
Biomechanical condition: UES should relax before maximal laryngeal elevation to enable laryngeal elevation to dilate the UES in time
Maximal distension of the UES (UES2) > maximum pharyngeal constriction (PC2)
UES2 > PC2
Biomechanical condition: Maximum pharyngeal constriction should be triggered late in the swallowing sequence to prevent biomechanical obstruction of laryngeal elevation which would obstruct UES opening
Tab. 6. Constant motor-temporal relations; left column contains data from Kendall et al., 2003
3. Motor learning
Since the facts do not support the assumption that swallowing is controlled by an iterative
CPG, the automaticity of the pharyngeal swallowing sequence cannot be explained by
iterative CPG control. We believe that motor learning provides an alternative explanation.
Automaticity of swallowing is based on improvement of swallowing motor coordination with
respect to bolus transport and airway protection, long-term storage of the resulting motor-
temporal patterns and activation of these stored patterns during swallowing:

22
Fig. 11. Motor learning
The assumption that automaticity of swallowing is based on motor learning does of course
not imply that the swallowing motor sequence is acquired by conscious learning. As pointed
out by Shadmehr&Wise (2005), ‘motor learning’ must be extended to biological learning of
unconscious sensory-motor functions, such as instinctive behaviour for example. Biologically
determined motor systems are acquired by genetic learning – evolutionary learning that
accrues over generations and becomes encoded genetically – and biological maturation.
With such an extended concept of motor learning, it is possible to trace the automaticity of
the pharyngeal swallow back to motor learning. How the swallowing motor system is
acquired based on genetic encoded principles and how this acquisition integrates experience
is unclear.
The integration of memorised motor-temporal structures makes the pharyngeal swallow
automatic and reflex-like (without being a reflex). Activating a pattern is always faster than
generating it. The drawback of increasing speed using pattern activation is a loss of flexibility,
however. The invariance of activated patterns in opposition to generated patterns does not
contradict the facts supporting variability of the pharyngeal motor sequence, as long as it is
assumed that activation of stored motor-temporal structures can be restricted to
subsequences of the pharyngeal motor sequence (selective synchronisation).
We assume that memorised motor-temporal structures are linked to bolus properties in
particular. Activation of motor plans that are specialised with respect to bolus volume or
bolus viscosity enables rapid synchronisation of the swallowing motor response with bolus-
induced variations in bolus transit. The data of Kendall et al. (2003) and Mendell &
Logemann (2007) support the assumption that if volume is high, a stored pattern with fronted
onset times is activated. The greater variability of the pharyngeal swallowing sequence in the
case of low bolus volume would follow if, in this case, no stored pattern is activated. A
preliminary hypothesis for stored motor-temporal structures that are specialised with respect

23
to bolus properties could be:
high volume + low viscosity: early onsets
low volume + high viscosity: late onsets
CPGS as motor memory
The discussion so far has underlined the need for rethinking the role of the ponto-medullary
region of the brain stem where the CPGS is assumed to be localised. If swallowing is not
controlled by an iterative CPG, what else might the role of the ponto-medullary region of the
brain stem be? It seems plausible, at least initially, that the motor memory of the swallowing
system is localised here. We therefore hypothesize that the CPGS is a motor memory which
emerged from ontogenetic development of swallowing motor coordination leading to long-
term storage of motor-temporal patterns. Following the extension of motor learning to
biological learning by Shadmehr&Wise (2005), it is evident that motor memories are not
restricted to cortical motor processing. The CPGS might be an example of an implicit or non-
declarative subcortical motor memory that is specific for swallowing. Although dysphagia
research has provided some insight into motor processing at the CPGS level, such as
identification of the ‘programming interneurons responsible for the spatio temporal
organization of the swallowing motor sequence’ (Amri et al., 1990, p. 384), a theory of the
CPGS as motor memory is still outstanding.
Cortical swallowing processing integrates the output of the CPGS motor memory into a
superior planning process that captures bolus properties that are not yet integrated at the
brain stem level (olfactory and optic bolus properties). Although the specific role of cortical
motor processing of swallowing apart from higher sensory integration is unclear, it can be
hypothetically assumed that cortical embedding enables a shift from activation of stored
motor-temporal structures to generation of motor-temporal structures by motor coordination.
Given an appropriate feedback, pharyngeal motor coordination becomes voluntarily
accessible and changeable to a certain extent, despite its automaticity, as swallowing
manoeuvres such as the ‘Mendelsohn manoeuvre’ show. This opens up opportunities for
swallowing rehabilitation.

24
Summary and Discussion
The following model summarises the assumptions made for motor processing of swallowing
as proposed in this paper:
Fig. 12. Motor processing of swallowing
Complexity and flexibility in the swallowing motor system with respect to sensory integration
of bolus features (volume, viscosity, position) as well as other data support the assumption
that swallowing is not simply a response to a stimulus related to the bolus but is based on a
process of motor planning that integrates perception related to the bolus but does not map it
directly to motor activity as basic reflexes do. However, data on the motor-temporal structure
of the pharyngeal swallowing sequence also contradict the assumption that motor planning of
swallowing is mediated by an iterative CPG. The pharyngeal swallowing sequence differs
from iterative movements: it does not include motor-temporal loops, sensory integration of
bolus properties is not restricted to a change in motor-temporal relations within a certain

25
activation order of the sequence (frequency change), the activation order of the sequence is
in part variable, and it is subject to motor-temporal transformations related to bolus properties
and bolus position. To explain these facts we have proposed a model for motor processing of
swallowing that is based on motor coordination and motor learning, two basic principles of
motor processing, which are not specific for the swallowing system. We assume that every
motor activity with a certain complexity – including swallowing – is based on a free
combinatory process of motor planning, constrained by biomechanical and functional output
conditions. Motor learning leads to storage of coordinated motor-temporal structures in motor
memories and enables rapid motor responses by activation of those structures.
In the case of swallowing, free motor coordination limited by biomechanical and functional
output conditions leads to a pharyngeal swallowing sequence that is not completely
temporally ordered. Apart from biomechanical conditions, the temporal order of the
pharyngeal swallowing sequence is based on selective synchronisation of laryngeal closure
and UES-opening with pharyngeal bolus transit, leading to motor-temporal clusters around
these synchronisation points and less motor-temporal order elsewhere. The response to
bolus properties affecting bolus transit (high volume or high viscosity) is activation of
specialised motor temporal structures, stored in the CPGS, which cause motor-temporal
transformations of the pharyngeal sequence to preserve selective synchronisation.
Localisation of the motor memory of the proposed processing model within the medullary
region of the brain stem, where the CPGS is assumed to be situated, is a first step towards
clarifying localisation of the whole processing model. We hypothetically assumed that a shift
from activation to generation of motor-temporal structures by the rule ‘coordinate’ is
associated with a shift to cortical processing. Although this is prima facie a plausible
assumption, it is unclear if motor coordination is restricted to cortical processing. The
characteristic feature of motor temporal structures generated by ‘coordinate’ is variability, in
contrast to the invariance of memorised motor temporal structures. If we look at the sensory-
motor systems of animals, however, the data contradict the assumption that absence of
cortical processing excludes variability. The sensory-motor systems of animals can be
variable and flexible with respect to integration of perception and generation of motor
response even if there is no cortex at all. Returning to swallowing, the question is whether
subcortical motor planning of swallowing can play a role beyond storing motor-temporal
structures. The identification of ‘programming’ interneurons within the medullary region of the
brainstem (Amri et al., 1990) indicates that this may be the case.

26
This paper is intended to be a step towards explaining the concept of motor planning of
swallowing. If the goal is to explore the motor planning of swallowing, localising brain regions
which have motor functions associated with swallowing is not sufficient. What is needed
beyond localisation is an attempt towards developing a theory that makes explicit the
computational structure – the algorithm – of swallowing motor planning. With the motor
grammar of swallowing outlined in this paper, we intended to take a first step towards this.
The attempt to clarify motor planning at the computational level might be brought into doubt
because it still has a weak data pool, however. In fact, techniques to visualise neuromotor
function such as Functional Magnetic Resonance Imaging (fMRI) are not yet specific enough
to allow identification of aspects of motor planning in sufficient detail. On the other hand, we
are certainly not left empty-handed. Videofluoroscopy (VFS), possibly combined with
complementary techniques such as Electromyography (EMG), gives us some access to
motor planning. VFS data provide an insight into motor planning because they provide an
insight into the motor-temporal structure of swallowing. If it is true that motor coordination is
primarily a temporal process, as we assumed with the motor grammar rule ‘coordinate’, then
it can be assumed that motor-temporal data mirror motor planning. Temporal VFS data
provide a possible but not optimal data pool for a computational theory of motor planning of
swallowing, however, since VFS does not provide direct access to muscle activity but only to
muscle activity within a certain biomechanical context (embodiment). Therefore, if future
imaging techniques will some day enable visualisation of muscle activity and neuromotor
control of muscles, then this will provide a better data pool than the current temporal VFS
data. Until then, temporal VFS data are not optimal but are nevertheless sufficient to
empirically validate attempts to develop a computational theory of motor planning of
swallowing.
If this paper is to be an appropriate step towards developing a computational theory of motor
planning of swallowing, the next steps should include the following:
1. Coordinate
We proposed a motor plan of hyoid elevation in terms of Coordinate based on data
from Pearson et al. (2010). A description of other oral and pharyngeal motor events in
terms of Coordinate is outstanding.
2. Motor constituents
Swallowing gestures such as hyoid elevation or tongue base retraction describe
swallowing at an intermediate level of motor coordination. Given the proposed
concept for motor coordination, both more elementary and more complex (temporally
defined) motor ‘constituents’ should be also relevant for motor planning of swallowing,

27
however. It should be a goal of future research to explore this ‘constituent structure’
of swallowing motor plans.
3. Biomechanical constraints
Although dysphagia research has discovered many of the biomechanical constraints
on swallowing, a specific theory of biomechanical conditions for swallowing motor
coordination has not yet been developed.
4. Selective synchronisation
Is it true that synchronisation of bolus arrival at the laryngeal vestibule with laryngeal
closure at some level and bolus arrival at the UES with UES opening satisfies
swallowing functionality? Are there other synchronisation points?
5. Specialised motor-temporal structures
Given that sensory integration of bolus properties is linked to activation of specialised
motor-temporal structures, what structures can be identified?

28
References
Amri M, Car A & Roman C (1990): Axonal branching of medullary swallowing neurons projecting
on the trigeminal and hypoglossal motor nuclei: demonstration by electrophysiological and
fluorescent double labeling techniques. Exp Brain Res (1990) 81:384-390
Brooks RA (2002): Robot: The Future of Flesh and Machines. The Penguin Press.
Chomsky N (1995): The Minimalist Program. Cambridge/London: MIT Press.
Daniels SK & Foundas AL (2001): Swallowing Physiology of Sequential Straw Drinking.
Dysphagia 16:176–182.
Daniels SK & Huckabee M-L (2008): Dysphagia following stroke. San Diego/Oxford/Brisbane:
Plural Publishing.
Dua KS, Ren J, Bardan E, Xie P, Shaker R (1997): Coordination of Deglutitive Glottal Function
and Pharyngeal Bolus Transit During Normal Eating. Gastroenterology 112:73–83.
Fagerstedt P & Ullén F (2001): Lateral Turns in the Lamprey. I. Patterns of Motoneuron Activity. J
Neurophysiol 86:2246–2256.
Hamdy S, Mikulis DJ, Crawley A, Xue S, Lau H, Henry S & Diamant NE (1999): Cortical activation
during human volitional swallowing: an event-related fMRI study. Am J Physiol Gastrointest Liver
Physiol 277:219-225, 1999.
Hiiemae KM & Palmer JB (1999): Food Transport and Bolus Formation during Complete Feeding
Sequences on Foods of Different Initial Consistency. Dysphagia 14:31–42.
Hooper SL (2001): Central pattern generators. ENCYCLOPEDIA OF LIFE SCIENCES, John
Wiley & Sons, Ltd., www.els.net.
Huckabee M-L & Kelly B (2006): Rehabilitationsmodelle im Management von Dysphagien:
Kasuistiken zur Schluckapraxie und zur Spastischen Dysphagie. In: Stanschus S (ed.):
Rehabilitation von Dysphagien. Idstein: Schulz-Kirchner Verlag, 2006, pp 137–171
Huckabee M-L, Deecke L, Cannito MP, Gould HJ, Mayr W (2003): Cortical Control Mechanisms in
Volutional Swallowing : the Bereitschaftspotenzial. Brain Topography 16:3–17.
Kendall KA, Leonard RJ, McKenzie SW (2003): Sequence Variability During Hypopharyngeal
Bolus Transit. Dysphagia 18(2):85–91.
Kendall KA, McKenzie S, Leonard RJ, Goncalves MI, Walker A (2000): Timing of Events in
Normal Swallowing: A Videoflouroscopic Study. Dysphagia 15:74–83.
Kornhuber H.H. & Deecke L. (1965): Hirnpotentialänderungen bei Willkürbewegungen und

29
passiven Bewegungen des Menschen: Bereitschaftspotential und reafferente Potentiale. Pflügers
Arch 284: 1–17
Lang IM (2009): Brain Stem Control of the Phases of Swallowing. Dysphagia 24:333–348.
Leonard R & McKenzie S (2006): Hyoid-Bolus Transit Latencies in Normal Swallow.
Dysphagia:183–190.
Levelt WJM (1989): Speaking. From Intention to articulation. MIT Press, Cambridge
(Massachusetts)/London (England)
Martin-Harris B, Brodsky MB, Michel Y, Lee F-S, Walters B (2007): Delayed Initiation of the
Pharngeal Swallow : Normal Variability in Adult Swallows. Journal of Speech, Language, and
Hearing Research 50:585–594.
Martin-Harris B, Brodsky MB, Price CC, Michel Y, Walters B (2003): Temporal coordination of
pharyngeal and laryngeal dynamics with breathing during swallowing: single liquid swallows. J
Appl Physiol. 94:1735–1743.
Mendell DA & Logemann JA (2007): Temporal Sequence of Swallow Events During the
Oropharyngeal Swallow. Journal of Speech, Language, and Hearing Research 50:1256–1271.
Pearson WG Jr, Langmore SE, Zumwalt AC (2010): Evaluating the Structural Properties of
Suprahyoid Muscles and their Potential for Moving the Hyoid. Dysphagia online: 11 November
2010
Rüffer, N (2012): Late Swallows. DysphagiEforum 1/2012, p. 40-56
Shadmehr R & Wise SP (2005): The Computational Neurobiology of Reaching and Pointing. MIT
Press, Cambridge (Massachusetts)/London (England)
Todorov E (2004): Optimality principles in sensorimotor control (review). Nat Neurosci. 2004
September; 7(9): 907–915.
Weckmueller J, Easterling C & Arvedson J (2010): Preliminary Temporal Measurement Analysis
of Normal Oropharyngeal Swallowing in Infants and Young Children. Dysphagia online: 9 June
2010
Wilmskötter J & Stanschus S (2012): Presbyphagie - im Alter wird alles schlechter?.
Sprachheilarbeit 57 (2012), 2-11
Ziegler W., von Cramon DY (1986): Timing deficits in apraxia of speech. European Archives of
Psychiatry and Neurologic Sciences. 1986;236:44-49