on papain lipase. - journal of biological · pdf fileon papain lipase. by marta s9sdberg and...
TRANSCRIPT

ON PAPAIN LIPASE.
BY MARTA S9SDBERG AND ERWIN BRAND.
(From the Division of Luboratories, MonteJore IIospitaZ, New York.)
(Received for publication, March IS, 1925.)
It is well known that during germination of oil seeds an altera- tion of their lipase occurs. Contrary to older views, especially those of Connstein, IIoycr, and Wartenberg (l), the following explanation for this phenomenon is offered by Willstatter and Waldschmidt-Leitz (21, pp. 163, 207; 23, p. 145). In the resting seed the lipase exists as “spermatolipasc” (seed lipase) and is abso- lutely inactive in a neutral medium (pH 6.8). Germination sets in with the hydrolysis of the seminal proteins. As soon as this has attained a certain degree, the seminal enzymes of the peptic group begin to act on the lipasc. Through the action of these enzymes the spermatolipase is changed into another lipase, the “blastolipase” (germ lipase). This blastolipase possesses the ability, in contrast to the spermatolipasc, to split fats also in a neutral medium. Only now can mobilization of the fat reserves be started by the blastolipase. It is noteworthy that this char- acteristic proteolysis of the spermatolipase to blastolipase during germination is produced by the enzymes of the peptic group, but not by tryptic ferments (23, p. 146; 21, p. 209).
But the effect of papain, as well as of papain activated by HCN, on spermatolipase has not been studied from this point of view. This investigation would be of particular interest, because Willstiitter and Grassmann, in their noteworthy communication, were able to show that papain alone and papain activated by HCN behave like two specific enzymes from different proteolytic groups. Papain alone acts like pepsin, while “papain cyanhydrin” has a tryptic action; i.e., peptone-forming as well as peptone-splitting (13, pp. 186, 205).
While investigating the action of papain and of papain cyan- hydrinl on spermatolipase, it occurred to us that practically noth-
* We shall report on this question in the near future.
59
by guest on May 4, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Papain Lipase
ing is known of the presence of a lipase in crude papain. It is true that Gerber (.i, 6) has found lipases in the latex of several plants, but he did not investigate the latex of Calica papaya, the source of papain. Pratt (9, p. 31) refers to the possible existence of a lipase in papaya latex, but was not able to establish definitely its presence.
We found in crude papain a lipase of considerable activity. The properties of this papain lipase are briefly given.
Papain lipase splits both higher glycerides and lower esters. It is insoluble in water and has its optimum activity at 35-40°C. at a ~1% of 5.S to 6.2 in acetate buffer. As to the kinetics, the law of Schiitz is followed to a certain extent. HCN and bile salts have no effect at all on the activity of the lipase. Calcium chlor- ide, however, accelerates it in an alkaline medium (changing pH).
Papain lipase differs from castor lipase in its pH optimum and its susceptibility to activation by calcium chloride (21, pp. 163, 167, 171). Rut is this difference in itself sufficient reason to assumn two specific lipsses? To answer this question we have to consider the recent development of views on the specificity of animal lipases. I,oevcnhart (8) was the first to find that the most esscnt’ial difference betwen the lipases of the liver and the pan- creas consists in their greatly differing “quantitative specificity” (15, p. 23; 17, p. 231) for substrates of different constitution. These results of Locvcnhnrt were recently reinvestigated and their theoretical foundation (15) was broadened by Willst%tter and his coworkers. Willst~itter was able to improve thr, accuracy of Locvenhart’s estimations by the comparative use of crude and purifiod lipase prepnrst,ions and by eliminating the possibility of the influence of accompanying bodies by having the enzyme act under conditions that arc made comparable by the addition of activators or of inhibiting substances (19, p. 231; 22, p. 210; 23, p. 146). But even a con&lerable difference in the quantitative specificity of tlvo lipnses, while suggestive, does not in itself suffice to prove finally their dissimilarity. Stereochemical specificity, however, seems really to constitute a property of the enzyme and to be independent of accompanying bodies (2; 3; 14, p. 206; 18; 19, p. 221). As such exact and comparable estimations have not yet been carried out for castor and papain lipase, the difference in their pII optimum and in their susceptibility to activation does not
by guest on May 4, 2018
http://ww
w.jbc.org/
Dow
nloaded from

31. Sandberg and E. Brand 61
per se sufice to decide whether or not they are really two different enzymes, for Willstattcr, in his work with animal lipases, found that the dependence of lipolysis on hydrogen ion concentration, as it is experimentally found, is deceptive and is caused by other more complex phenomena (22, p. 97). Accompanying bodies influence the lipase in its relation to hydrogen ion concentration, to adsorption, activation, inhibition and poisoning, stability, optimum of temperature, and heat destruction (14, p. 205).
A lipase preparation obtained directly from papaya latex might be mom suitabIe for investigating the relative specificity of castor and papain lipnse.
EXPERIMEKTAL.
1. Estinaation of the Lipolytic Action.
In estimating the lipolysis, an olive oil of the saponification number 183 was usually employed as substrate.? The detcrmina- tions were done in accord with Willstatter in the following manner (22, pp. 103, 111; 21, p. 177).
The papain preparation which is to be determined is weighed into a srnall wide mouthed bottle of a capacity of about 30 cc. with well ground stopper, then the desired amount of water or buffer solution is added,3 and the mixture shaken for a short time, till the greater part of the papain has gone into solution. Now 2.5 grn. of olive oil arc added, the mixture is shaken by hand strongly and evenly for 3 minuDes (22 pp. 104, 112; 21, p. 166) and left in the thermostat at the desired temperature for the remaining time of the estimation. After the reaction has gone on for a given time, it is stopped by bringing the saponification mixture into an Erlenmeycr flask with the help of about ten times the amount of 96 per cent alcohol (distilled over ROH) and 20 cc. of ether. Then the fatty acid which has been formed is estimated, using I .5 drops of a 1 per cent alcoholic solution of thymolphthalein as indicator (22, p. 105), by titration with 0.5 N alcoholic IiOH (alcohol content DO per cent), till a distinctly blue color appears.
* ~&thy1 butyratc according to Willstiitter (17, p. 236): volume 20 cc.; 5 cc. of N acetate buffer (pII S.8), 0.2 gm. of methyl butyrate, 100 mg. of papain (LVallerstein), 60 min., 30°C. = 0.1 per cent cleavage.
3 The lipase being insoluble in water, it is necessary to add the water or buffer solution before adding the olive oil. Otherwise the papain would envelop the lipasc in the saponification mixture, thus preventing its action.
by guest on May 4, 2018
http://ww
w.jbc.org/
Dow
nloaded from

62 Papain Lipase
The buffer as well as the amphoteric substances of the papain preparation themselves use up some KOH when in alcoholic solu- tion (12; 20; 22, p. 105; 21, p. 177). Therefore, suitable controls are run with each experiment and this amount of KOH is deducted.
The percentage of cleavage is estimated from the saponifica- tion number of the olive oil in the following manner.
100 mg. of papain (Wallerstein) (Table III) reacted with 2.5 gm. of olive oil for 60 minutes at 30°C. in the presence of 4 cc. of 0.25 x acetate buffer (pH 6.0). The amount of alkali used to neutra- lize the fatty acid after that time was 2.5 cc. of 0.5 N KOH for the experiment and 0.75 cc. of 0.5 N KOH for the control. From the difference of 1.75 cc. of 0.5 s KOII, the saponification is estimated as follows:
1.75 x 56.11 x 100 ~ - = 10.7 2 x 2.5 x 183 per cent
9. Papain Lipase and Hydrogen Ion Concentration.
We find that the most favorable conditions for the action of the papain lipase are offered in a pH range from about 5.8 to 6.2, if acetate buffer of a normality of 0.1 to 0.5 is used. Phosphat’e buffers are less satisfactory, and the smallest amount of cleavage is found with citrate buffers, which coincides with Willstatter’s findings with castor lipase (21, p. 170).
3. Lipolytic Action of Dij’erent Papain Prepamtions.
We find that in different commercial preparations of papain the content of lipasc present varies very much. The highest content of lipase is shown by a preparation which we owe to the kindness of Mr. L. Wallerstein; a preparation from Parke, Davis and Company gave the next best results.
l+. Lipolytic Action and Temperature.
Papain lipase has its temperature optimum at about 35-4O”C. At higher temperatures, however, and when the experiments are continued for a longer time, in addition to the injurious influence of the temperature, destruction by the proteolytic enzyme must be taken into consideration.
by guest on May 4, 2018
http://ww
w.jbc.org/
Dow
nloaded from

IX. Sandberg and E. Brand 63
TABLE I.
Lipolytic Action and pH. ~~~
Acetate Buffer,*
(100 mg. papain (Iva~lcrstein), 2.5 gm. olive oil, 4 cc. 0.25 N acetate buffer, 60 min., 30°C.)
pH . . .._.._........ . .’ 4.0 4.8 1 5.0 ’ 5.2 -~
5.6 G.0 1 6.2 1 6.S Cleavage, per cent . ./ 2.2 5.6 1 6.6 j 7.7 8.8 110.7 i 9.S / 7.9 --
Phosphate Buffer. f
(100 mg. papain (Wallerstein), 2.5 gm. olive oil, 4 cc. M/IS phosphate buffer, Cfl min., 30°C.)
pH . . . . . . . . . . . . . . .I 5.3 6.6 6.8 7.3 8.0 Cleavage,percent ..__. ( 5.4 1 7.1 / 6.6 I 6.1 1 5.3
Citrate Uu.fer.f (100 mg. papain (Wallerstein), 2.5 gm. olive oil, 4 cc. 0.1 M citrate buffer,
60 min., 30°C.).
PH................................ . . . . . j5.0~5.3j5.616.016.3Ij:; Cleavagqpercent . . . . . . . . . . . . . . . . . . . . . . 1.7 ) 2.9 3.5 15.4 5.1
NII,-NH&l Buffer.1
(100 mg. papain (Parke, Davis and Company), 2.5 gm. olive oil, 6.0 cc. H&, 1 cc. 0.5 N NH&H&l buffer, pH 8.9, 60 min., 3OT!. = 3.5 per cent cleavage).
* Walpole (11). t According to Sorensen. $ According to -Michaelis, and Willstatter, Waldschmidt-Leitz, and
Memmen, (22), p. 104.
TABLE II.
Lipolytic Action and Buffer Concentration.
(100 mg. papain (Wallerstein), 2.5 gm. olive oil, 4 cc. acetate buffer, 60 min., 30°C.)
PH
5.6 5.8 6.0 6.2 6.8
Cleavage at various buffer concentrations.
ix. 0.5 N 0.2: N per cent per cent
pe;y;t
-; ;O&---
7.7 7.6 9.2 7.8 7.8 8.9 9.3 9.8 10.0 10.7 10.3 9.6 9.5 9.8 10.0 7.5 7.4 7.9 8.5
by guest on May 4, 2018
http://ww
w.jbc.org/
Dow
nloaded from

64 Papain Lipase
5. Influence of Activators.
When investigating animal lipases, WillstSitter, Waldschmidt- Leitz, and Memmen (22, p. 99) studied the influence of the com- bined action of several activators” (complex or coupled adsorbents) in acid and alkaline media. We found that addition of ovalbumin, HCN, or bile salts has no influence on the lipolytic action of papain lipase, either in an acid or an alkaline medium. Glycerol, how-
TABLE III.
Lipolytic Action of Different Papain Preparations. (100 mg. papain, 2.5 gm. olive oil, 4 cc. 0.25 N acetate buffer, pH 6.0, 60
min., 30°C.)
Wallerstein. l’ttrke, Davis and Company.
Merck I. Merck II. “i;Lra;d
per cent pa7 cell t per cent per cent per cmt
10.7 8.7 6.9 3.3 2.4
TABLE IV.
Lipolytic Action and Temperature.
(80 mg. papain (Parke, Davis and Company), 2.5 gm. olive oil, 4 cc. 0.25 N acetate buffer, pII 6.0, 60 min.)
“C. per cent per cent per cent 25 6.1 8.7 13.5 30 7.G 10.8 16.7 35 8.7 11.9 18.0 40 11.2 15.2 17.7 50 11.7 14.1 14.8 60 11.2 11.7 11.9
60 min.
Cleuvage.
120 min. 320 min.
ever, inhibits in acid and neutral media, but activates at an alka- line pH. CaClz activates distinctly at an alkaline reaction. Rut this activation of papain lipase is diminished by ovalbumin, quite in contrast to Willstlitter’s findings with pancreatic lipase (22, p. 109). This effect of CaC12 and ovalbumin can also be seen with a more purified preparation (Table XII).
WillstMer states in his work on castor lipasc that plant lipase
4 The effect of individual activators, used separately, was known before.
by guest on May 4, 2018
http://ww
w.jbc.org/
Dow
nloaded from

M. Sandberg and E. Brand 65
is not susceptible to activation (21, p. 163). He also finds, that an activation of castor lipase by manganese salts, which has been described by Hoyer (7), Tanaka (lo), and Falk and Hamlin (4), does not occur under optimal conditions (21, p. 171).
TABLE V.
Lipase Activation and pII.
(80 mg. papain (Parke, Davis and Company), 2.5 gm. olive oil, 60 min., 3O”C., pH 4.8 and 6.0; 2 cc. 0.5 N acetate buffer + 2 cc. H20, pH 8.9; 1 cc. NHJ-SH&I buffer + 5 cc. ILO. 2 per cent CaC12 solution, 3 per cent ovalbumin solution.)
I Cleavage I Without glycerol. With 50 per cent glycerol.
(Acetate buffer.) 6.0 7.6 7.4 7.8 7.9 4.1 4.3 4.1
(NHJ-NH&l buffer.) 8.9 3.2 6.1 4.0 5.1 4.3 5.6 4.4 5.7
Experiment I.-80 mg. of papain (Parke, Davis and Company) and 2.5 gm. of olive oil react for 60 minutes at 30°C. with: (a) 4 cc. of 0.25 N acetate bufrer (pH 6.0); (b) 10 mg. of HCN + 4 cc. of 0.25 N acetate buffer (pH ad- justed to 6.0); (c) 5 cc. of H20 + 1 cc. of 0.5 N NH,-NH&I, buffer (pH 8.9); and (d) 5 cc. of HQO + 10 mg. of HCN + 1 cc. of 0.5 N NHs-NH&l buffer (pE-I adjusted to 9.2). The amount of cleavage was: (a) 7.9 per cent, (b) 7.4 per cent, (c) 2.9 per cent, and (d) = 3.3 per cent. Previous treatment with these buffer solutions for 60 minutes at 30°C. has no influence on the amount of cleavage: (a) 7.5 per cent, (b) 7.6 per cent, (c) 3.2 per cent, and (d) 3.0 per cent.
Experiment 2.-80 mg. of papain (Parke, Davis and Company), 5 cc. of H,O, and 1 cc. of 0.5 N NHs-SH&l buffer (pH 8.9) were mixed with 2.5 gm.
by guest on May 4, 2018
http://ww
w.jbc.org/
Dow
nloaded from

66 Papain Lipase
of olive oil. Different amounts of the following activators were added and left 60 minutes at 30°C.
T No.
1 2 3 4 5 6 7 8 9
!-
Amount of cleavage
Without activator. + 1 mg. CaCll. + 2 “ “ + 5 “ “ + 10 “ “ + 20 ‘( “ + 40 “ “ + 10 (‘ sodium taurocholate. + 10 “ “ glycocholate.
per cent
3.0 4.6 4.5 6.0 5.8 6.3 6.1 2.9 3.4
6. Kinetics of the Lipolytic Action.
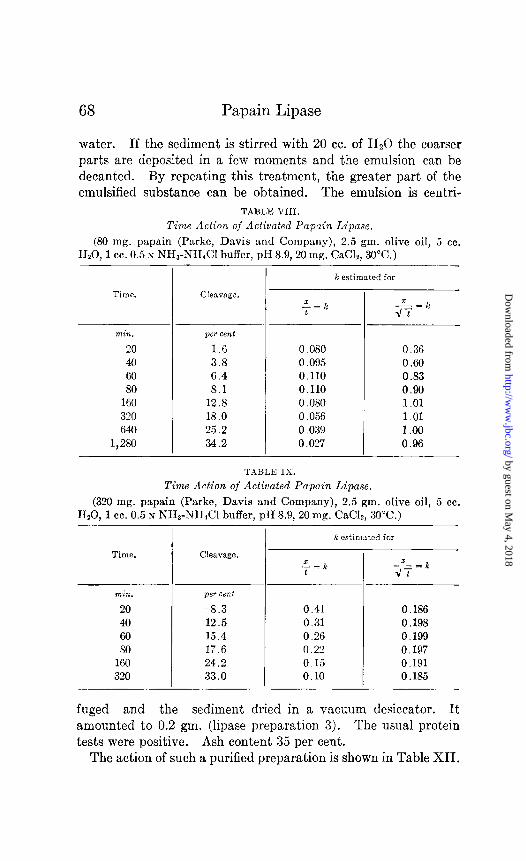
The lipolytic saponification follows to a certain extent the law of Schiitz (Tables VI to XI). The discrepancies that arise upon activation with CaClz in an alkaline medium (changing pH) warrant further investigation (Tables VIII, IX, and X).
We have postponed the introduction of a special papain lipase unit (as was done by Willst%ter for various other lipases (16, p. 23; 17, p. 237; 22, p. 115; 21, p. 178)) till a lipase has been prepared directly from the latex.
7. Stability of the Lipase.
The lipase is relatively stable against the action of the proteoly- tic enzyme. 100 mg. of papain (Wallerstein) were first treated with: (a) 2 cc. of Hz0 for 16 hours at 20°C.; (b) 2 cc. of Hz0 for 16 hours at 37°C.; (c) 2 cc. of Hz0 for 48 hours at 37’C.; (d) 10 mg. of HCN for 16 hours at 20°C.; (e) 10 mg. of HCN for 16 hours at 37°C.; and {fl 10 mg. of HCN for 48 hours at 37°C.
The amount of cleavage of 2.5 gm. of olive oil (acetate buffer, pH 6.0, 60 minutes, 30°C.) was: (a) 9.0 per cent, (b) 7.1 per cent, (c) 3.2 per cent, (d) 8.0 per cent, (e) 6.1 per cent, and (f) 1.0 per cent.
8. Pzrrification of the Lipase.
5 gm. of papain (Parke, Davis and Company) were extracted twice for 10 minute periods with 50 cc. of H20 at 25”C., centri- fuged, and the supernatant clear fluid was decanted. The amount
by guest on May 4, 2018
http://ww
w.jbc.org/
Dow
nloaded from

MI. Sandberg and E. Brand 67
of cleavage of 6.0 cc. of solution I with 2.5 gm. of olive oil and (a) 1 cc. of N acetate buffer (pII 6.0) and (b) 1 cc. of N SH,- NH&l buffer (pH 8.9) + 20 mg. of CaCL was (a) 0.0 per cent
TABLI? VI.
Time Action of Papain Lipase.
(80 mg. papain (Parke, Davis and Company), 2.5 gm. olive oil, 4 cc. 0.25 N acetate buffer, pH 6.0, 30°C.)
min. per cent
10 2.3 20 4.2 40 0.1 60 7.6 80 8.8
120 10.8 160 12.3 320 16.7
k estimated for
0.23 0.73 0.21 0.94 0.15 0.96 0.13 0.98 0.11 0.98 0.09 0.98 0.08 0.97 0.05 0.93
Action of Different Amounts of Papain Lipase.
(Papain (Parke, Davis and Company), 2.5 gm. olive oil, 4 cc. 0.25 s acetate buffer, pH 6.0, 60 min., 30°C.)
I’apain.
77W. per cent
20 2.0 40 3.9 80 7.6
100 8.7 160 10.9 200 12.2 320 14.8
Cleavage.
12 estimated for
r=k -f!-=k E de
0.10 0.45 0.097 0.62 0.095 0.85 0.087 0.87 0.068 0.86 0.061 0.86 0.046 0.83
and (b) 0.0 per cent. The sediment (0.95 gm.) has a dark brown color and consists of some sand, the remains of cell walls and of a substance which makes a whitish emulsion when stirred with
by guest on May 4, 2018
http://ww
w.jbc.org/
Dow
nloaded from
68 Papain Lipase
water. If the sediment is stirred with 20 cc. of I-I,0 the coarser parts are deposited in a few moments and the emulsion can be decanted. By repeating this treatment, the greater part of the emulsified substance can be obtained. The emulsion is centri-
TABLE VIII.
Time Action of Activated Pupuin Lipase.
(80 mg. papain (Parke, Davis and Company), 2.5 gm. olive oil, 5 cc. lI,O, 1 cc. 0.5 s NH3-NII&l buffer, pH 8.9, 20 mg. CaC12, 30°C.)
min. pet cent
20 1.6
40 3.8 60 6.4 80 8.1
160 12.8 320 18.0 640 25.2
1,280 34.2
Cleavage.
k estimated for
0.080 0.36 0.095 0.60 0.110 0.83 0.110 0.90 0.080 1 .Ol 0.056 1.01 0.039 1.00 0.027 0.96
TABLE IX.
Time Action of Activated Papain Lipase.
(320 mg. papain (Parke, Davis and Company), 2.5 gm. olive oil, 5 cc. H,O, 1 cc. 0.5 N NH,-NH&l buffer, pH 8.9, 20 mg. CM& 30°C.)
T k estimated for
Time. Cleavage.
min. pet cent
20 8.3 40 12.5 60 15.4 SO 17.6
160 24.2 320 33.0
.z_ = k t
0.41 0.186 0.31 0.198 0.26 0.199 0.22 0.197 0.15 0.191 0.10 0.185
-Z- =k dt
fuged and the sediment dried in a vacuum desiccator. It amounted to 0.2 gm. (lipase preparation 3). The usual protein tests were positive. Ash content 35 per cent.
The action of such a purified preparation is shown in Table XII.
by guest on May 4, 2018
http://ww
w.jbc.org/
Dow
nloaded from

M. Sandberg and E. Brand 69
TABLE X.
Action of Different Amounts of Activated Papain Lipase.
(Papain (Parke, Davis and Company), 2.5 gm. olive oil, 5 cc. I-IZO, 1 cc. 0.5 N NH,-NH&l buffer, pH 8.9, 20 mg. CaCI*, 60 min., 30°C.)
k estimated for
Papain. Cleavage. ?=k L=k e d\/e
m*. per cent
10 1.0 0.100 0.32 20 2.0 0.100 0.45 40 3.2 0.080 0.51 80 6.1 0.076 0.68
100 7.1 0.071 0.71 160 10.3 0.064 0.81 320 15.4 0.048 0.86
TABLE XI.
Amount of Enzyme and Percentage of Cleavage at Diferent Times. (Papain (Parke, Davis and Company), 2.5 gm. olive oil, 5 cc. HzO, I cc.
0.5 N N&NH4CI buffer, pH 8.9, 20 mg. CaC12, 30°C.)
Amount of cnsyme.
e = 320 mg.
1 (320 yd f (160 ) -:(80 1: )
6(4 1 A(20 “ )
- Amount of enzyme X time = e X 1
20 40 80 160 320
8.3 12.5 17.6 24.2 33.0 9.0 11.8 18.1 25.2 32.8 8.1 12.8 18.0 25.2 34.2 7.7 10.9 15.4 20.5 6.1 8.4 12.5
TABLE XII.
Action of a Purijied Papain Lipase Preparation.
(20 mg. lipase preparation 3, 2.5 gm. olive oil, 60 min., 30°C.)
Buffer.. . 4 cc. 0.25 N acetate buffer, pH 6.0. 3 cc. FLO, 1 cc. A WII,-SH,CI buffer. pII 8.2.
Activator.. None. 10 mp. 10 rrrE. 10 mg. CsClr CaC12. Sane. 30 mg.
ChCl?. ovaibumirl. + 30 mg. ovalburnin.
per cent per cent per cm2 per cent per cent per cent
8.6 8.9 1.6 10 .!I 2.6 61
by guest on May 4, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Papain I,ipase
SUMMARY.
Crude papain contains a lipasc of considerable activity. Papain lipase is insoluble in water and has its optimum effect at
3540°C. at a pH 5.8 to 6.2 in acetate buffer. Papain lipase follows the law of Schtitz to a certain extent. CaClz activates papain lipase in an alkaline medium (changing
PHI. Papain lipase differs from castor lipase in its pH optimum and its
possible activation by CaCL in an alkaline medium (changing PHI.
Purification is possible by removing the papain with water.
BIBLIOGRAPHY.
1. Connstein, W., Hoyer, E., and Wartenberg, H., Ber. them. GM., 1902, xxxv, 3983.
2. Dakin, H. D., J. Physiol., 1904, xxx, 253. 3. Dakin, II. D., J. Physiol., 19004-05, xxxii, 199. 4. Falk, K. G., and Hamlin, M. L., J. Am. Chem. SOL, 1913, xxxv, 210. 5. Gerber, C., Compt. rend. Sot. biol., 1913, lxxiv, 250,718,720,822. 6. Gerber, C., Compt rend. Sot. biol., 1913, lxxv, 147, 149. Gerber, C., and
Guiol, IT., Compt. rend. Sot. biol., 1913, lxxv, 151. 7. IIoyer, E., 2. physiol. Chem., 190607, 1, 414. 8. Loevenhart. 11. S., J. Biol. Chem., 1906-07, ii, 427. 9. Pratt, D. S., Philippine J. SC., Series A, 1915, x, 1.
10. Tanaka, Y., J. Coil. Eng. Imp. Univ. Tokyo, 1912, v, 142. 11. Walpolc, G. S., J. Chem. $oc., 1914, cv, 2501. 12. Willstattcr, R., Alkalimetrische. Bestimmung von .4minosburen und
Peptiden, in Abderhalden, E., Handbuch der biologischen Arbeits- methoden, 1. Abt., Berlin and Vienna, 1922, pt. 7, 289.
13. Willstatter, R., and Grassmann, W., 2. physiol. Chem., 1924, cxxxviii, 184.
14. Willstatter, R., Haurowitz, F., and Memmen, F., Z. physiol. Chem., 1924, cxl, 203.
15. Willstiitter, R., and Kuhn, R., Z. physiol. Chem., 1923, cxxv, 1. 16. Willstatter, R., and Xlemmen, F., Z. physiol. Chem., 1923, cxxix, 1. 17. Willstattcr, R., and Nemmen, F., Z. physiol. Chem., 1924, cxxxiii, 229. 18. Willstiitter, R., and ?vIemmen, F., Z. physiol. Chem., 1924, cxxxiii, 247. 19. Willstatter, R., and Memmcn, F., Z. physiol. Chem., 1924, cxxxviii, 216. 20. Willstiitter, R., and Waldschmidt-Leitz, E., Ber. them. Ges., 1921,
liv, 2988. 21. Willstatter, R., and Waldschmidt-Leitz, E., Z. physiol. Chem., 1924,
cxxsiv, 161. 22. Willstattcr, R., Waldschmidt-Leitz, E., and Nemmen, F., Z. physiol.
Chem., 1923, cxxv, 93. 23. Willstiitter, R., and Waldschmidt-Leitz, E., Z. physiol. Chem., 1923,
cxxv, 132.
by guest on May 4, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Marta Sandberg and Erwin BrandON PAPAIN LIPASE
1925, 64:59-70.J. Biol. Chem.
http://www.jbc.org/content/64/1/59.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
alerts to choose from all of JBC's e-mailClick here
#ref-list-1
http://www.jbc.org/content/64/1/59.citation.full.htmlaccessed free atThis article cites 0 references, 0 of which can be
by guest on May 4, 2018
http://ww
w.jbc.org/
Dow
nloaded from