on the development of the male genitalia and the efferent genital
TRANSCRIPT


36 DBV RAJ MEHTA
ambulatory legs. In this connexion Kraepelin (1873), Dewitz(1875), and others described these appendages as arising fromimaginal discs, and Wheeler (1893) showed that there existsa direct continuity of the embryonic appendages with the
Zander (1900,1901,1903) actually described the developmentof the penis and the gonapophyses ('valvae') in the post-embryonic stages of Hymenoptera, Trichoptera, and Lepido-ptera from paired rudiments or buds on the ninth abdominalsegment. This formed a basis for later work on these lines, andhis conclusions were largely confirmed in certain other insectorders. In this connexion may be mentioned the work of Muir(1915, 1918) and Singh-Pruthi (1924) on Coleoptera, Christo-phers and his collaborators (1922, 1926) on Diptera, Kershawand Muir (1922), Singh-Pruthi (1924) and George (1929) onHomoptera and Zygoptera respectively.
Verson and Bisson (1896) also referred to the developmentof the male genitalia in Bombyx mori Linn, in their exten-sive memoir on the origin and nature of the efferent genitalducts in this species. Their conclusions, however, were criticizedby Zander (1903), and his work, since then, is the only compre-hensive account to which reference could be made for determin-ing the homologies of the elements composing the insectgenitalia.
It has also been realized that the mode of origin of the'valvae' as described by Zander (1903) is open to question(Singh-Pruthi, 1924), and his conclusions differ from those ofhis predecessors (Verson and Bisson, 1896, and others). Thedevelopment of the posterior abdominal segments and theirappendages in Lepidoptera need further reinvestigation, becauseZander's observations (1903) on these lines, though conclusive,were not confirmed by subsequent workers. It therefore isdesirable that the whole problem should be revised in the lightof existing knowledge of orders other than the Lepidoptera.With this in view I have studied the development of the malegenitalia in the following Lepidoptera, viz. H e p i a l u s lupu-l inus Linn., P ie r i s r a p a e Linn., E a r i a s fabia Stoll.,and Bombyx mori Linn., as representatives of the sub-

GENITALIA IN LEPIDOPTERA 87
order Homoneura and the super-families Papilionina, Noctuina,and Notodontina respectively.
In this account I have also included a general description ofthe development and nature of the efferent genital ducts in theabove forms. Much confusion prevails on this subject, and Ihave discussed at length the existing views on it throughoutthe whole class, supporting my arguments with the conditionswhich I find in the Lepidoptera mentioned above.
It is with great pleasure that I express my indebtedness toMr. L. E. S. Eastham, my supervisor of research studies, forsuggesting this problem and taking a keen interest in the pro-gress of this research. I am also thankful to Dr. A. D. Imms forvarious valuable suggestions in preparing the manuscript forpublication. To Professor J. Stanley Gardiner I am muchindebted for providing me with every facility for work in theZoological Laboratory, Cambridge.
The material for this work was obtained locally and waspartly sent to me from India. In this connexion, I wish to ex-press my sincere thanks to Mr. M. Afzal Husain, Entomologistto the Government of the Punjab, for supplying me withthe developmental stages of the silk-worm moth Bombyxmori Linn.
This investigation was conducted during the tenure of aUniversity Scholarship awarded by the Government of thePunjab, India. The grant made by the State is gratefullyacknowledged.
MATEBIAL AND TECHNIQUE.
The common British forms such as P i e r i s r a p a e Linn,and H e p i a l u s l u p u l i n u s Linn, were reared in the labora-tory. The developmental stages of E a r i a s fabia Stoll. andthe silk-worm moth Bombyx mori Linn, were receivedfrom the Entomological Section, Department of Agriculture,Punjab, by the courtesy of the authorities.
The specimens were fixed in Bouin's picro-formol and Carnoy'sfluids. Eeal difficulty was experienced in sectioning the lastlarval and pupal stages of large forms such as H e p i a l u s andBombyx , on account of the thick outer chitinous cuticle and





42 DEV RAJ MBHTA
of development are the same in these insects, therefore refer-ence to forms other than P ie r i s r a p a e Linn, has only beenmade where occasion demanded, as for example in B o m b y xmori Linn., which presents morphological features additionalto those of P i e r i s .
There are ten distinct abdominal segments in the larva. The
XT IX T
TEXT-FIG. 3.
Longitudinal vertical section through the posterior abdominal seg-ments of the larva of P ie r i s r a p a e .
segments 1-8 bear distinct spiracles and can therefore be easilyidentified. The ninth is comparatively small and the tenth bearsa pair of large pro-legs. The genitalia are formed only towardsthe end of the larval period, but the genital rudiments becomevisible soon after the caterpillar has passed through the thirdmoult. At this stage, in a longitudinal vertical section (Text-fig.3), the relation of different parts is fairly evident. In the earliestlarval instar there is an invagination of the body-wall on the


44 DBV RAJ MEHTA
and develop into a pair of lobes, more prominent in E a r i a sfabia Stoll. than in P ie r i s (Text-fig. 7, VI). They projectfreely within the genital cavity and represent the ' valvae lobes'.At this stage the surrounding mesodermal cells rich in tracheaemigrate into the lumen of these two pairs of cone-like lobes.
Before the larva pupates the penis lobes approximate andfuse in the median line, enclosing a central integumentarydepression which forms the 'ductus ejaculatorius simplex'(Text-figs. 5, 6, 8, E.jd"). The genital pouch has now almostdisappeared by a process of evagination, and the lateral pairof cones lies on the surface close to the ninth sternite anddevelops into the embryonic valvae (Text-figs. 5, 6, 8, Val).
During pupal development the finer modelling of the penisand the valvae takes place. The epidermal cells bordering thevalvae (Val) show intense activity, so that a layer of chitin issecreted around them. This is particularly evident in theNoctuid E a r i a s fabia Stoll (Text-fig. 8). The penis attainsits full size by now, and the 'ductus ejaculatorius simplex'becomes coated with concentric muscle-fibres which give ita peculiar appearance.
The above mode of the development of the penis and thevalvae is identical to that which has been described by Versonand Bisson (1896) in Bombyx mori Linn. But these authorswere not in a position to decide whether the fundaments of thepenis and the valvae are differentiated from a single broaderbud-growth, or that the 'valvae lobes' arise subsequentlybelow the 'penis lobes' and independently of them. Zander(1903) showed in the Pyralid P a r a p o n y x s t r a t i o t a r i athat the former view was more correct, but later workers(Singh-Pruthi, 1924, and others) dissented from it. As I haveshown above, the penis lobes are differentiated earlier as com-pared with rudiments of the valvae from a coniform thickeningof the epidermis. It is therefore quite clear that the origin ofthe ' penis' and the ' valvae' lobes takes place not only indepen-dently but also in succession. Besides the Lepidoptera, similarobservations have been recorded in Homoptera (Singh-Pruthi,1924), Orthoptera (Wheeler, 1893), and Hymenoptera (Michaelis,1900).


46 DEV RAJ MEHTA
segments 1-7 bear distinct spiracles, while on the eighth theyare rudimentary. The ninth and tenth segments closely ap-proximate, and Poulton (1889) showed that 'when a spine is
Pilm.m.
TEXT-FIG. 7.
Transverse section through the ninth abdominal segment in thefinal instar larva of Ea r i a s fabia .
Pup.Ct. Vallm.m.
TEXT-FIG. 8.
Transverse section through the ninth abdominal segment of theearly pupal stage of Ea r i a s fab ia .
absent and the terminal part of the pupa is rounded, the partabove the anus, nevertheless, corresponds to the larval analflap'. This terminal structure represents a portion of the tenthsegment and is known as the 'cremaster'.
In an advanced pupal stage, internally the relationship ofabdominal segments in the developing imago becomes more


48 DEV EAJ MEHTA
processes respectively of the tenth segment. Thus there canbe no scepticism regarding the existence of a distinct tenthsegment in addition to these two appendages. The limits ofthe former are well denned, and it is visibly differentiated intoa tergal and sternal part (X St), and the anal tube (A) passesbetween the two sclerites. In the imago the tenth segmentremains membranous and lies more or less in close associationwith the uncus. It is therefore seldom preserved in preparationsof genitalia treated with caustic potash. Probably it is on thisaccount that the uncus and the gnathos (' Scaphium' of Zanderand others) are regarded as the tergum and the sternum of thetenth somite (Peytoureau, 1894; Klinkhardt, 1900, and others).Zander (1903) was the first to define the limits of the tenthsegment in the pupal stages of the Pyralid P a r a p o n y xs t r a t i o t a r i a , and the above account of P i e r i s corroborateshis conclusions in all essential details.
In the absence of any further ontogenetic evidence confirm-ing Zander's views on these lines, considerable speculation hasbeen indulged in by several morphologists. Busck and Heinrich(1921) believed that the tegumen and the uncus both belongto the tenth somite, the ninth tergite being greatly reducedand continued dorsally as membrane only. In the same wayMacgillivray (1923) considered the tegumen as the tenth seg-ment, and regarded the uncus as the distal end of this struc-ture. Differing from these, Forbes (1923) regarded the tegumenas the ninth tergite and the uncus as the 'dorsal part' of thetenth segment. Finally, Philpott (1926), reviewing the uncus inthe Micropterygoidea, came to the conclusion' that the tegumenis formed from the ninth tergite and that the uncus is an out-growth of that tergite'. I have clearly shown by illustrationsfrom the longitudinal vertical sections through the pupal stageof B o m b y x mor i Linn. (Text-figs. 9, 10) that the limits ofthe tegumen or the ninth tergite (IX T) and the uncus (U) arewell denned by the presence of a distinct dorsal constriction.Also, the appendiculate nature of the uncus is evident from itsrelation with the tenth segment which is withdrawn below it.The tergal and sternal parts of the latter are clearly indicatedin my preparations, so that it is easily shown that the uncus

GENITALIA IN LEPIDOPTERA 49
and the gnathos are derived as dorsal and ventral processesof the tenth segment.
The ninth sternite (IX St) is large and develops a pair of
2m.m.
TEXT-FIG. 10.
Longitudinal vertical section through an advanced pupa of Bom-byx mori Linn, showing the development of the penis and theanellus lobes.
elongate lobiform processes called the ' anellus lobes' (Text-fig.10, An.l.) which lie on either side of the penis (P). The intro-mittent organ or the penis at this stage is fully formed anddevelops a distinct penis pouch (Pp), and its lumen is traversed
NO. 301 B

50 DEV RAJ MEHTA
by the 'ductus ejaculatorius simplex' (D.ej) which opens atits orifice.
Towards the end of pupal life the genitalia are presented intheir final arrangement. The relation of the different elements
rxst.
vi nstPup.Ct
TEST-FIG. 11.
Transverse section through the posterior abdominal region of thelate pupal stage of P ie r i s r a p a e .
which compose the genital armature in P i e r i s r a p a e Linn,and E a r i a s fabia Stoll. is fairly evident in a transversesection through an advanced pupa (Text-figs. 11 and 12).Chitinization now proceeds rapidly over the copulatory organs.The valvae (Val) assume a broad, leaf-like appearance with acovering of hair and spines. The tegumen (Tg) forms a dorsalhood over the anal tube (A) which passes below it. The uncus(17) has become more prominent and also develops a coveringof hairs and spines. The penis is heavily chitinized at this stageand supported on either side by the large anellus lobes (Text-fig.10, An.l.)

GENITALIA IN LEPIDOPTBBA 51
(b) E f fe ren t Gen i t a l Ducts.—This account refersprecisely to Pieris—the other species confirming it in allessentials.
In the caterpillar there persists from the earliest stages a pair
lm.m.
TEXT-PIG. 12.
Transverse section through the posterior region of the abdomenin the advanced pupal stage of Ea r i a s fab ia .
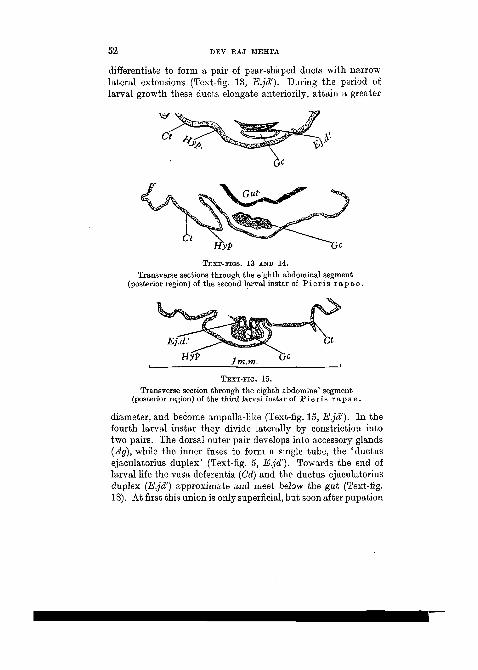
of very delicate ducts, the vasa deferentia (Vd), which arisefrom the testes (Text-fig. 4, Tes). During larval developmentthey possess a distinct lumen, grow in size, and become con-voluted. From the very beginning of larval life each vasdeferens terminates on either side of the eighth segment, moreor less in a latero-ventral position (Text-fig. 17, Vd). In theprepupal stage they meet the narrow extensions of the ' ductusejaculatorius duplex' on the posterior margin of the eighthsegment (Text-fig. 18, E.jd').
In the earliest larval instar of P i e r i s r a p a e Linn, thebody-wall invaginates to form the genital cavity (Gc). At thebase of this integumentary depression the epidermal cells
E2