original article digestive and liver disease...regulação epigenética no envelhecimento e no...
TRANSCRIPT

Regulação Epigenética no Envelhecimento e no Câncer Gástrico P á g i n a | 113
Artigos
4.2.2 Protein expression and promoter DNA methylation of SIRT1 and 3 in gastric
cancer
Submetido como Original Article à revista “Digestive and Liver Disease” em 08/03/11. Protein expression and promoter DNA methylation of SIRT1 and 3 in gastric cancer
Luara Carolina Frias Lisboa1; Carolina Oliveira Gigek1; Patrícia Natalia Oliveira Silva1;
Mariana Ferreira Leal1; Eleonidas Moura Lima2; Danielle Queiroz Calcagno3; Paulo
Pimentel Assumpção4; Rommel Rodriguez Burbano3; Marília de Arruda Cardoso
Smith1* 1 Disciplina de Genética, Departamento de Morfologia e Genética, Universidade
Federal de São Paulo, SP, Brasil. 2 Departamento de Biologia, Campus Ministro Reis Velloso/Parnaíba, Universidade
Federal do Piauí, PI, Brasil. 3 Laboratório de Citogenética Humana, Instituto de Ciências Biológicas, Universidade
Federal do Pará, PA, Brasil. 4 Serviço de Cirurgia, Hospital Universitário João de Barros Barreto, Universidade
Federal do Pará, PA, Brasil.
ABSTRACT
Background: Sirtuins are an important regulator of chromatin structure and gene
expression. This is the first study evaluating SIRT1 and SIRT3 expression and promoter
methylation in gastric carcinogenesis. Methods: Immunohistochemistry was analyzed
about 50 gastric tumor and in about 20 normal gastric mucosa samples. About 100
gastric cancer and 50 normal mucosa samples were investigated through methylation
specific PCR. Results: SIRT1 expression was associated with gastric cancer (OR=6;
p=0.0003). This is the first study to evaluate SIRT1 promoter methylation. We observed
no association between SIRT1 protein and methylation pattern in gastric tissue.
Concerning SIRT3, we originally report higher protein imunnostaining in 80% of non-
neoplastic, thus differing significantly from neoplastic samples (OR=0.313; p=0.05). The

Regulação Epigenética no Envelhecimento e no Câncer Gástrico P á g i n a | 114
Artigos
frequency of SIRT1 and SIRT3 promoter methylation did not differ between tumor and
normal gastric samples. Conclusion: SIRT1 expression was higher in tumor than in non-
neoplastic gastric tissue, and no association with DNA methylation pattern was
observed. In contrast, SIRT3 may reveal a protective role in gastric carcinogenesis.
Moreover, we originally demonstrated a correlation between SIRT3 methylation pattern
and protein expression in gastric tumor samples.
Keywords: SIRT1, SIRT3, gastric cancer, DNA methylation, epigenetic.
BACKGROUND
Sirtuin is a NAD dependent deacetylase family conserved in many
organisms [1], with seven known components (SIRT1-7) in humans. These proteins
participate in regulation of chromatin structure and gene expression and have a
determinant role on cellular identity [2].
SIRT1 deacetylates histones and non-histone targets, including genes
involved in cell cycle, survival, stress response and apoptosis, as MYC and p53 [3].
Normal SIRT1 expression has a protective effect against DNA damage, enhancing DNA
repair ability and prevention of tumorigenesis [4]. In contrast, in cancer, SIRT1
colocalizes on the promoter region of several aberrantly silenced tumor suppressor
genes whose DNA is hypermethylated, but its ultimate activity has not yet been proved
[5]. Hence, the role of SIRT1 as a tumor promoter or a tumor suppressor remains to be
determined; nevertheless there are strong evidences that SIRT1 may be a critical
regulator of cancer development, being an important therapeutic target [3].
Another sirtuin, SIRT3, has mitochondrial localization and its expression is
associated with reduction of mitochondrial damage [1], growth arrest, senescence and
apoptosis. The role of SIRT3 is still unknown, although high gene expression was
described in breast cancer node positive [6]. SIRT3 is located in 11p15.5, a known
imprinted chromosomal region, which comprises genes related to aging and tumor
suppressor [7].
Gastric cancer is the result of genetic, epigenetic and environmental
onmental factors [8]. Furthermore, gene promoter methylation is the most common

Regulação Epigenética no Envelhecimento e no Câncer Gástrico P á g i n a | 115
Artigos
epigenetic alteration and has been reported in gastric cancer [9-15]. SIRT1 has a CpG
island of about 1Kb and CG content up to 72% in promoter and exon 1 region [16].
SIRT3 promoter region was identified as a bidirectional promoter with a CG content of
67% and comprises two CpG islands [17]. Thus, the investigation of epigenetic
mechanisms involved in regulation of SIRT1 and SIRT3 expression is relevant to
elucidate these genes function in gastric carcinogenesis.
The aim of this study was to evaluate SIRT1 and SIRT3 promoter
methylation pattern in gastric carcinogenesis and correlate with their protein expression
as well as with clinicopathological characteristics. To our knowledge, this is the first
study to investigate the relationship between SIRT1 and SIRT3 promoter methylation
and protein expression in gastric cancer.
METHODS
1. Samples
Gastric samples were obtained surgically from João de Barros Barreto
University Hospital (HUJBB) in Pará State, Brazil. This population is composed of
interethnic crosses among three main origin groups: European (mainly represented by
Portuguese), Africans and Amerindians [18]. Informed consent with approval of the
ethics committee of HUJBB was obtained. All patients had negative histories of
exposure to either chemotherapy or radiotherapy before surgery and there was no other
co-occurrence of diagnosed cancers. All samples were classified according to Laurén
[19] and tumors were staged using standard criteria by TNM [20] staging.
2. Immunohistochemical staining
SIRT1 and SIRT3 protein expression were evaluated in formalin-fixed
paraffin embedded tissues of 70 and 74 samples of gastric mucosa, respectively (Table
1).
Antigen retrieval was performed by microwave treatment 20 min at 900 W
in a citrate buffer, pH 6.0. After cooling, sections were immersed in 0.3% hydrogen
peroxide in phosphate-buffered saline (PBS) for 10 minutes to block endogenous
peroxidase activity. Sections were then incubated in a humid chamber overnight with
SIRT1 primary antibody (C19 DB083, Delta biolabs, USA) or SIRT3 primary antibody

Regulação Epigenética no Envelhecimento e no Câncer Gástrico P á g i n a | 116
Artigos
(C73E3, Cell Signaling Technology, USA). After the PBS rinse, slides were incubated
with secondary antibody and then with streptavidin-biotin-peroxidase complex, both for
30 minutes at room temperature with a PBS wash between each step. Slides were
visualized with diaminobenzidine-hydrogen peroxide and counterstained with Harry’s
hematoxylin.
Positive protein expression was defined as clear nuclear staining in 50%
or more of the cells, whereas negative immunostaining was considered when no
positive cells were observed or in rare cases (less than 25% weakly stained tumor cells)
(Figures 1 and 2). Normal gastric mucosa was used as an internal control. Two
pathologists evaluated the immunostaining results independently.
3. Methylation specific PCR (MSP)
SIRT1 and SIRT3 methylation pattern were evaluated in 162 and 88
gastric tissue samples, respectively. Among these samples, around 70% are from
sporadic gastric adenocarcinomas and 30% of non-neoplastic gastric mucosa of
patients (distant location of primary tumor) (Table 2).
Genomic DNA (200 ng) of gastric tissue samples underwent bisulfite
modification using EpiTect Bisulfite kit (Qiagen, Germany) according to the
manufacturer’s instructions, converting unmethylated cytosines to uracils and leaving
methylated cytosines unchanged. MSP was performed on treated DNA as previously
described [21]. MSP for SIRT3 was performed as previously described [22]. Specific
primers for SIRT1 promoter, were as follows: 5’-GGGGTGAATTTGGTTGTATTATATG-
3’ (sense) and 5’-AAACAAAAACTATTACATCTACCACT-3’ (antisense) for the
unmethylated reactions; 5’-GGCGAATTTGGTTGTATTATACG-3’ (sense) and 5’-
GAACGAAAACTATTACGTCTACCG-3’ (antisense) for the methylated reactions, with
PCR products of 112 bp and 110 bp respectively. Briefly, PCR reaction was carried out
in a 25μL volume with 200μmol/L of MgCl2, 100ng of DNA, 200 pmol/L of primers and
1.25 units of Taq DNA polymerase. After initial denaturation for 5 min at 94ºC, 40 cycles
of 94ºC for 45 s, at 62ºC for 45 s, and 72ºC for 30 s were carried out, followed by a final
extension for 5 min at 72ºC.

Regulação Epigenética no Envelhecimento e no Câncer Gástrico P á g i n a | 117
Artigos
Results were scored when there was a clear and visible band on the
electrophoresis gel with the methylated or unmethylated primers [21]. Hypermethylation
was considered when the presence of a methylated band was observed (Figure 3).
SIRT1 and SIRT3 methylation pattern and respective protein expression
were both in all available gastric cancer samples with immunohistochemistry results
(Table 3).
4. Statistical analyses
Logistic regression was performed to evaluate imunnohistochemistry
results and clinicopathological characteristics χ2 test or Fisher’s exact was used to
assess associations between methylation status and clinicopathological characteristics.
p values less than 0.05 were considered significant.
RESULTS
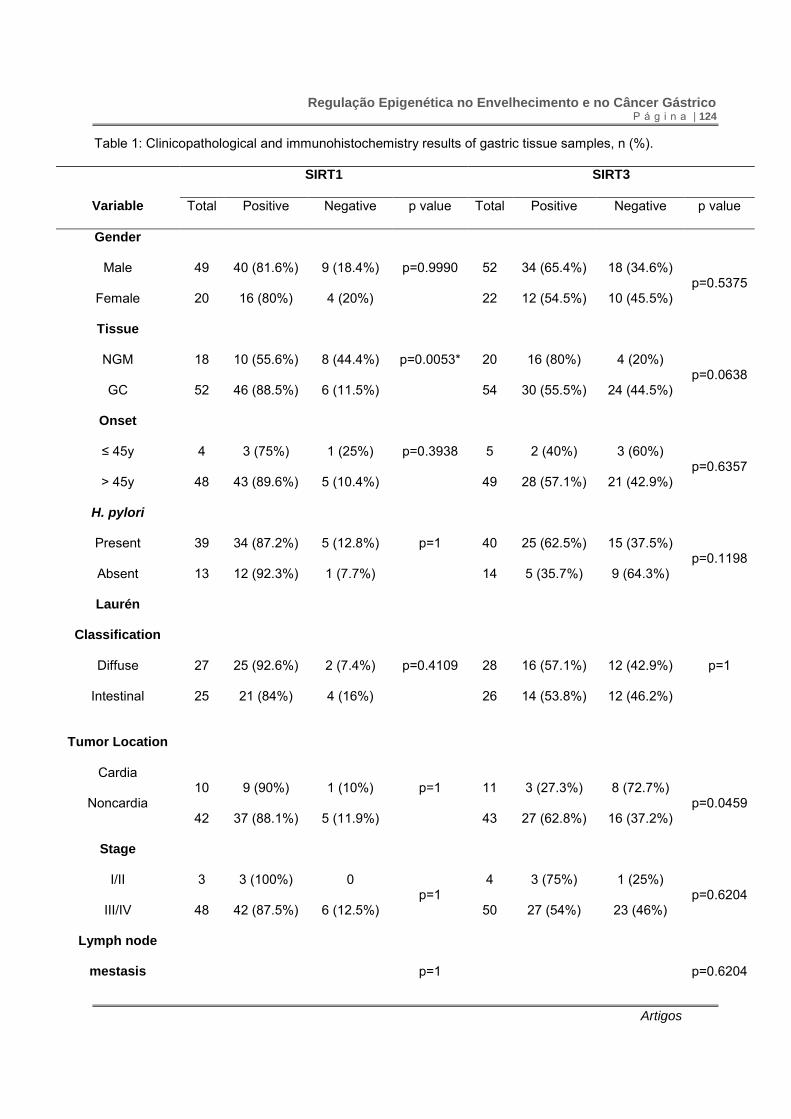
SIRT1 expression was more frequently observed in neoplastic than in non-
neoplastic samples (0.885 vs 0.556). The odds ratio for SIRT1 was 6 (95% CI 2.7-
21.17). (p=0.003; χ2 = 8.79, df=1). Compared to normal mucosa, frequency of SIRT1
staining was significantly higher in diffuse type gastric cancer (p=0.0079). However,
SIRT1 expression did not differ between diffuse and intestinal type gastric cancer
(p=0.4109) (Table 1).
Eighty per cent of normal gastric mucosa and 55.5% of gastric cancer
samples showed SIRT3 expression. Thus, SIRT3 we observed a difference between
neoplastic and non-neoplastic samples, with OR = 0.313 (0.092-1.059) (p=0.05; χ2 =
3.708, df=1). There was no difference of SIRT3 protein staining between diffuse and
intestinal type gastric cancer (p=1). Gastric cancer samples of the noncardia region
presented more frequent SIRT3 expression than the cardia region (p=0.0459). There
was no clear association between the frequency of SIRT1 and SIRT3 expression and
age, gender, H. pylori infection, tumor extension and presence of distant metastasis
(Table 1).
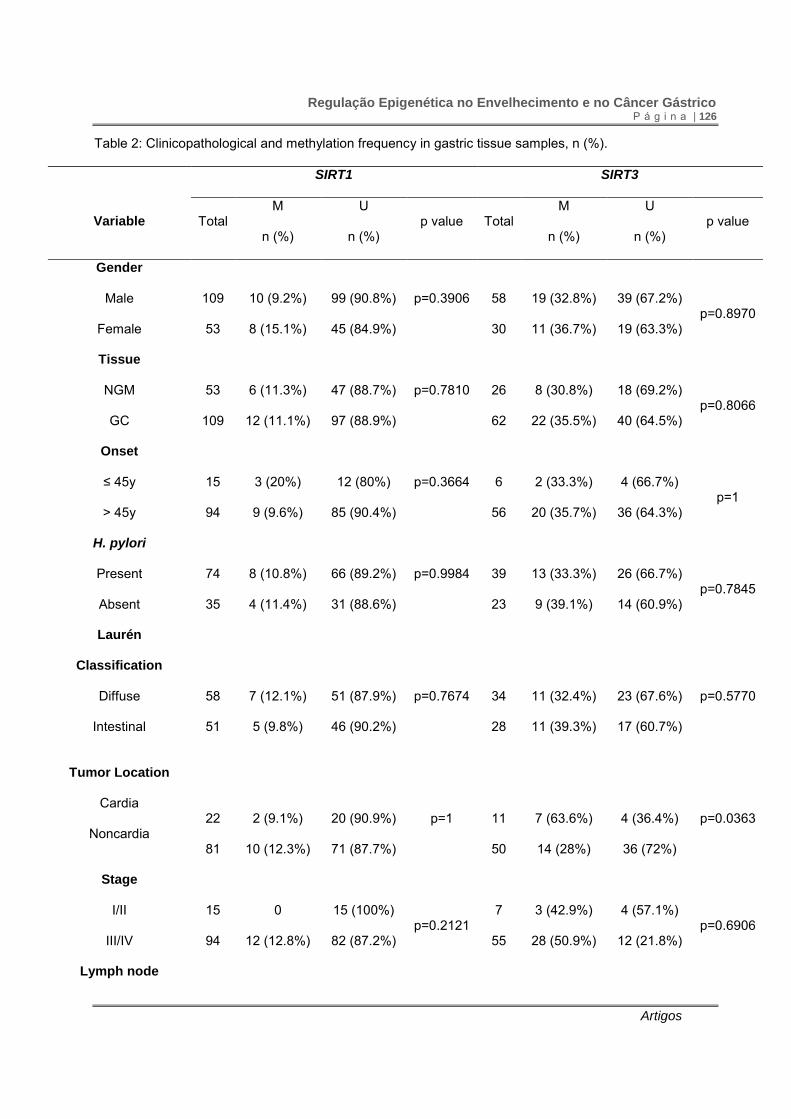
The frequency of SIRT1 and SIRT3 promoter methylation did not differ
between tumor and normal samples. SIRT3 promoter methylation was more frequently
observed in cardia than in noncardia tumors (p=0.0363), occurring mostly in cardia

Regulação Epigenética no Envelhecimento e no Câncer Gástrico P á g i n a | 118
Artigos
intestinal type gastric cancer (p=0.0152). Promoter methylation of SIRT1 and SIRT3 did
not show association with age, gender, H. pylori infection, tumor extension and
presence of distant metastasis (Table 2).
To clarify the relationship between promoter methylation and gene
expression, we compared the presence of methylation of SIRT1 and SIRT3 promoter
regions with their protein expression in gastric cancer and normal mucosa specimens.
There was no correlation between SIRT1 expression and its promoter methylation
pattern in our sample (p=0.8409, r=0.0308). In gastric specimens, SIRT3 methylation
pattern was associated with lack of protein expression (p=0.0033, r=0.4461) (Table 3).
DISCUSSION
In the present study, we observed higher immunostaining of SIRT1 protein
in 88.5% of neoplastic samples, thus indicating that presence of this protein increases in
6 times the risk of gastric cancer. A recent study reported that 73% of gastric cancer
samples presented positive SIRT1 immunoreactivity. Moreover, SIRT1 expression was
associated with poor prognosis, histological type and p53 expression [23].
Overexpression of SIRT1 allows rapid proliferation and loss of checkpoints, promoting
the progression of prostate [24] and colon cancer [25]. Taken together, these data
suggest that this protein participate in tumor promotion in carcinogenesis processes,
including gastric cancer.
Tumor growth depends on the loss of control of differentiation and
inhibition of apoptosis. Tumor suppressor p53 is a known substrate of SIRT1, and plays
a role in cell cycle regulation and apoptosis, regulating proliferation and stress response
[4]. We observed higher SIRT1 protein in diffuse type gastric cancer. This observation
might be correlated with our previous data in the same sample population: an increased
frequency of immunostaining of p53 in the intestinal type gastric cancer [26]. Cha et al
[23] demonstrated that 100% of p53-positive gastric cancer cases showed lower SIRT1
expression. Thereby, SIRT1 appears to promote gastric tumorigenesis via p53.
MYC is another protein involved in gastric tumorigenesis and binds to
SIRT1 promoter region, inducing SIRT1 expression [27]. We have previously described
MYC immunoreactivity in all gastric adenocarcinoma samples in individuals from Pará

Regulação Epigenética no Envelhecimento e no Câncer Gástrico P á g i n a | 119
Artigos
State [28-30], suggesting that MYC overexpression might contribute to higher SIRT1
levels found in our sample.
Furthermore, MYC induces SIRT1 expression in HeLa cells and SIRT1, in
turn, deacetylates and downregulates MYC, resulting in decreased MYC target gene
expression and cellular transformation. Consequently, this MYC-SIRT1 negative
feedback loop can restrain MYC’s transformational activities. However, MYC
overexpression and poor prognosis of advanced gastric neoplasia suggest that this
possible MYC-SIRT1 feedback is not sufficient to restrain the gastric malignization.
Therefore, SIRT1 might have both tumor suppression and promoting role depending on
the stage of cell transformation [27].
To our knowledge, this is the first study evaluating SIRT1 promoter
methylation, as well as its relation with protein expression. Here we observed that 11%
of all gastric tissue (non-neoplastic and neoplastic) presented SIRT1 promoter
methylation. The fact that both non-neoplastic and neoplastic samples presented SIRT1
promoter methylation might be due to the regular methylation levels previously
observed in health stomach tissue [31]. Moreover, SIRT1 overexpression was
previously associated with high-CpG island methylator phenotype in colorectal cancers
[32]. Hypermethylation of multiple tumor-related genes is frequently detected in gastric
carcinoma and adjacent normal tissues and is likely to be involved in the early gastric
carcinogenesis process [9-14, 33].
To clarify the relationship between promoter methylation and protein
expression, we compared the presence of methylation of SIRT1 promoter region with
protein immunostaining in gastric cancer and normal mucosa specimens. Therefore, no
association between protein expression and promoter methylation pattern was found in
our sample, indicating that this type of epigenetic control does not have any influence in
protein expression.
Concerning SIRT3, we found that 80% of non-neoplastic gastric mucosa
presented this protein immunostaining, diffenring significantly from neoplastic samples,
therefore having a protective role in this type of cancer. The mitochondrial localization of
SIRT3 is consistent with its role in active process to minimize mitochondrial damage
[34]. In colorectal carcinoma, SIRT3 mediates apoptosis by silencing of BCL2, a

Regulação Epigenética no Envelhecimento e no Câncer Gástrico P á g i n a | 120
Artigos
mitochondrial anti-apoptotic protein [1]. Likewise, high levels of BCL2 has been reported
in the early stage of gastric carcinogenesis process [35], which may explain the frequent
lack of SIRT3 protein in our neoplastic samples.
In addition, SIRT3 overexpression increases FOXO3A DNA-binding
activity, and thus, FOXO3A dependent gene expression [34]. In gastric cancer,
FOXO3A induces apoptosis cooperating with RUNX3 [36], a transcription factor
frequently inactivated in 40% of early stage and almost 90% of advanced gastric cancer
by hemizygous deletion and hypermethylation of its promoter [37]. In our population, we
also described RUNX3 methylation in about 50% of gastric cancer samples [10].
Our data corroborates with previous report that SIRT3 acts as tumor
suppressor, and its loss amplifies the phenotypic effects of oncogene expression [38].
The frequency of SIRT3 promoter methylation did not differ between tumor
and normal samples. In the present study, we originally reported that SIRT3 methylation
pattern association with lack of protein expression in our gastric specimens. We
observed SIRT3 methylation in more than 60% all of gastric samples and an inverse
relationship between promoter methylation and protein expression was also found. No
methylation in lymphocytes of healthy elderly individuals was observed in a previous
research and only 6.8% of young individuals presented no methylation in lymphocytes
[22]. In the present study, we identified an association between SIRT3 expression and
unmethylated status in gastric tumor samples. Taken together, our data suggest a
tissue-specific regulation of SIRT3.
SIRT1 expression was higher in tumor cells than in normal gastric tissue,
increasing the risk of this type of cancer. No association between protein and DNA
methylation pattern was observed. Moreover, we originally demonstrated the SIRT3
methylation pattern and protein expression was correlated with gastric tumor samples.
Further studies may reveal a possible therapy via sirtuin in gastric cancer.
REFERENCES
1. Allison, S.J. and J. Milner, SIRT3 is pro-apoptotic and participates in distinct
basal apoptotic pathways. Cell Cycle, 2007. 6(21): p. 2669-77.

Regulação Epigenética no Envelhecimento e no Câncer Gástrico P á g i n a | 121
Artigos
2. Metoyer, C.F. and K. Pruitt, The role of sirtuin proteins in obesity.
Pathophysiology, 2008. 15(2): p. 103-8.
3. Kim, E.J. and S.J. Um, SIRT1: roles in aging and cancer. BMB Rep, 2008.
41(11): p. 751-6.
4. Zeng, L., et al., Silent information regulator, Sirtuin 1, and age-related diseases.
Geriatr Gerontol Int, 2009. 9(1): p. 7-15.
5. Jones, P.A. and S.B. Baylin, The fundamental role of epigenetic events in cancer.
Nat Rev Genet, 2002. 3(6): p. 415-28.
6. Ashraf, N., et al., Altered sirtuin expression is associated with node-positive
breast cancer. Br J Cancer, 2006. 95(8): p. 1056-61.
7. Schwer, B., et al., The human silent information regulator (Sir)2 homologue
hSIRT3 is a mitochondrial nicotinamide adenine dinucleotide-dependent deacetylase. J
Cell Biol, 2002. 158(4): p. 647-57.
8. Shang, J. and A.S. Pena, Multidisciplinary approach to understand the
pathogenesis of gastric cancer. World J Gastroenterol, 2005. 11(27): p. 4131-9.
9. Gigek, C.O., et al., hTERT methylation and expression in gastric cancer.
Biomarkers, 2009. 14(8): p. 630-6.
10. Lima, E.M., et al., Study of Methylation Pattern of de Novo DNA
Methytransferase Genes and its Correlation with DNA Methylation Pattern of RUNX3 in
Individuals with Gastric Cancer from Northern Region of Brazil. Int J Morphol, 2007.
25(4): p. 817-824.
11. Leal, M.F., et al., Promoter hypermethylation of CDH1, FHIT, MTAP and PLAGL1
in gastric adenocarcinoma in individuals from Northern Brazil. World J Gastroenterol,
2007. 13(18): p. 2568-74.
12. Lima, E.M., et al., Methylation status of ANAPC1, CDKN2A and TP53 promoter
genes in individuals with gastric cancer. Braz J Med Biol Res, 2008. 41(6): p. 539-43.
13. Moura Lima, E., et al., DNA mismatch repair gene methylation in gastric cancer
in individuals from northern Brazil. Biocell, 2008. 32(3): p. 237-43.
14. Guimaraes, A.C., et al., Interrelationships among chromosome aneuploidy,
promoter hypermethylation, and protein expression of the CDKN2A gene in individuals

Regulação Epigenética no Envelhecimento e no Câncer Gástrico P á g i n a | 122
Artigos
from northern Brazil with gastric adenocarcinoma. Cancer Genet Cytogenet, 2007.
179(1): p. 45-51.
15. Gigek, C.O., et al., Insulin-like growth factor binding protein-3 gene methylation
and protein expression in gastric adenocarcinoma. Growth Horm IGF Res, 2010.
16. Li, L.C. and R. Dahiya, MethPrimer: designing primers for methylation PCRs.
Bioinformatics, 2002. 18(11): p. 1427-31.
17. Bellizzi, D., et al., Characterization of a bidirectional promoter shared between
two human genes related to aging: SIRT3 and PSMD13. Genomics, 2007. 89(1): p.
143-50.
18. Batista dos Santos, S.E., et al., Differential contribution of indigenous men and
women to the formation of an urban population in the Amazon region as revealed by
mtDNA and Y-DNA. Am J Phys Anthropol, 1999. 109(2): p. 175-80.
19. Lauren, P., The Two Histological Main Types of Gastric Carcinoma: Diffuse and
So-Called Intestinal-Type Carcinoma. an Attempt at a Histo-Clinical Classification. Acta
Pathol Microbiol Scand, 1965. 64: p. 31-49.
20. Sobin, L.H. and C.H. Wittekind, TNM: Classification of malignant tumours. 6th
ed2002, New York: Wiley-Liss
21. Herman, J.G., et al., Methylation-specific PCR: a novel PCR assay for
methylation status of CpG islands. Proc Natl Acad Sci U S A, 1996. 93(18): p. 9821-6.
22. Silva, P.N., et al., Promoter methylation analysis of SIRT3, SMARCA5, HTERT
and CDH1 genes in aging and Alzheimer's disease. J Alzheimers Dis, 2008. 13(2): p.
173-6.
23. Cha, E.J., et al., Expression of DBC1 and SIRT1 is associated with poor
prognosis of gastric carcinoma. Clin Cancer Res, 2009. 15(13): p. 4453-9.
24. Huffman, D.M., et al., SIRT1 is significantly elevated in mouse and human
prostate cancer. Cancer Res, 2007. 67(14): p. 6612-8.
25. Stunkel, W., et al., Function of the SIRT1 protein deacetylase in cancer.
Biotechnol J, 2007. 2(11): p. 1360-8.
26. Khayat, A.S., et al., Interrelationship between TP53 gene deletion, protein
expression and chromosome 17 aneusomy in gastric adenocarcinoma. BMC
Gastroenterol, 2009. 9(1): p. 55.

Regulação Epigenética no Envelhecimento e no Câncer Gástrico P á g i n a | 123
Artigos
27. Yuan, J., K. Minter-Dykhouse, and Z. Lou, A c-Myc-SIRT1 feedback loop
regulates cell growth and transformation. J Cell Biol, 2009. 185(2): p. 203-11.
28. Calcagno, D.Q., et al., Aneuploidy of chromosome 8 and C-MYC amplification in
individuals from northern Brazil with gastric adenocarcinoma. Anticancer Res, 2005.
25(6B): p. 4069-74.
29. Calcagno, D.Q., et al., Interrelationship between chromosome 8 aneuploidy, C-
MYC amplification and increased expression in individuals from northern Brazil with
gastric adenocarcinoma. World J Gastroenterol, 2006. 12(38): p. 6207-11.
30. Costa Raiol, L.C., et al., Interrelationship between MYC gene numerical
aberrations and protein expression in individuals from northern Brazil with early gastric
adenocarcinoma. Cancer Genet Cytogenet, 2008. 181(1): p. 31-5.
31. Esteller, M., CpG island hypermethylation and tumor suppressor genes: a
booming present, a brighter future. Oncogene, 2002. 21(35): p. 5427-40.
32. Nosho, K., et al., SIRT1 histone deacetylase expression is associated with
microsatellite instability and CpG island methylator phenotype in colorectal cancer. Mod
Pathol, 2009. 22(7): p. 922-32.
33. Leung, W.K., et al., Concurrent hypermethylation of multiple tumor-related genes
in gastric carcinoma and adjacent normal tissues. Cancer, 2001. 91(12): p. 2294-301.
34. Jacobs, K.M., et al., SIRT3 interacts with the daf-16 homolog FOXO3a in the
mitochondria, as well as increases FOXO3a dependent gene expression. Int J Biol Sci,
2008. 4(5): p. 291-9.
35. Tsamandas, A.C., et al., The potential role of Bcl-2 expression, apoptosis and
cell proliferation (Ki-67 expression) in cases of gastric carcinoma and correlation with
classic prognostic factors and patient outcome. Anticancer Res, 2009. 29(2): p. 703-9.
36. Ito, Y. and K. Miyazono, RUNX transcription factors as key targets of TGF-beta
superfamily signaling. Curr Opin Genet Dev, 2003. 13(1): p. 43-7.
37. Hanai, J., et al., Interaction and functional cooperation of PEBP2/CBF with
Smads. Synergistic induction of the immunoglobulin germline Calpha promoter. J Biol
Chem, 1999. 274(44): p. 31577-82.
38. Schumacker, P.T., A tumor suppressor SIRTainty. Cancer Cell, 2010. 17(1): p. 5-
6.
Regulação Epigenética no Envelhecimento e no Câncer Gástrico P á g i n a | 124
Artigos
Table 1: Clinicopathological and immunohistochemistry results of gastric tissue samples, n (%).
SIRT1 SIRT3
Variable Total Positive Negative p value Total Positive Negative p value
Gender
Male
Female
49
20
40 (81.6%)
16 (80%)
9 (18.4%)
4 (20%)
p=0.9990
52
22
34 (65.4%)
12 (54.5%)
18 (34.6%)
10 (45.5%)
p=0.5375
Tissue
NGM
GC
18
52
10 (55.6%)
46 (88.5%)
8 (44.4%)
6 (11.5%)
p=0.0053*
20
54
16 (80%)
30 (55.5%)
4 (20%)
24 (44.5%)
p=0.0638
Onset
≤ 45y
> 45y
4
48
3 (75%)
43 (89.6%)
1 (25%)
5 (10.4%)
p=0.3938
5
49
2 (40%)
28 (57.1%)
3 (60%)
21 (42.9%)
p=0.6357
H. pylori
Present
Absent
39
13
34 (87.2%)
12 (92.3%)
5 (12.8%)
1 (7.7%)
p=1
40
14
25 (62.5%)
5 (35.7%)
15 (37.5%)
9 (64.3%)
p=0.1198
Laurén
Classification
Diffuse
Intestinal
27
25
25 (92.6%)
21 (84%)
2 (7.4%)
4 (16%)
p=0.4109
28
26
16 (57.1%)
14 (53.8%)
12 (42.9%)
12 (46.2%)
p=1
Tumor Location
Cardia
Noncardia
10
42
9 (90%)
37 (88.1%)
1 (10%)
5 (11.9%)
p=1
11
43
3 (27.3%)
27 (62.8%)
8 (72.7%)
16 (37.2%)
p=0.0459
Stage
I/II
III/IV
3
48
3 (100%)
42 (87.5%)
0
6 (12.5%)
p=1
4
50
3 (75%)
27 (54%)
1 (25%)
23 (46%)
p=0.6204
Lymph node
mestasis
p=1
p=0.6204

Regulação Epigenética no Envelhecimento e no Câncer Gástrico P á g i n a | 125
Artigos
Present
Absent
49
3
43 (87.8%)
3 (100%)
6 (12.2%)
0
50
4
27 (54%)
3 (75%)
23 (46%)
1 (25%)
Distant
metastasis
Present
Absent
Unkown
17
33
2
13 (76.5%)
31 (93.9%)
4 (23.5%)
2 (6.1%)
p=0.1624
17
34
10 (58.8%)
18 (52.9%)
7 (41.2%)
16 (47.1%)
p=0.7707
IHC: immunohistochemistry assay; NGM: normal gastric mucosa; GC: gastric cancer; *significant p value
after multiple comparison corrections.
Regulação Epigenética no Envelhecimento e no Câncer Gástrico P á g i n a | 126
Artigos
Table 2: Clinicopathological and methylation frequency in gastric tissue samples, n (%).
SIRT1 SIRT3
Variable Total M
n (%)
U
n (%) p value Total
M
n (%)
U
n (%) p value
Gender
Male
Female
109
53
10 (9.2%)
8 (15.1%)
99 (90.8%)
45 (84.9%)
p=0.3906
58
30
19 (32.8%)
11 (36.7%)
39 (67.2%)
19 (63.3%)
p=0.8970
Tissue
NGM
GC
53
109
6 (11.3%)
12 (11.1%)
47 (88.7%)
97 (88.9%)
p=0.7810
26
62
8 (30.8%)
22 (35.5%)
18 (69.2%)
40 (64.5%)
p=0.8066
Onset
≤ 45y
> 45y
15
94
3 (20%)
9 (9.6%)
12 (80%)
85 (90.4%)
p=0.3664
6
56
2 (33.3%)
20 (35.7%)
4 (66.7%)
36 (64.3%)
p=1
H. pylori
Present
Absent
74
35
8 (10.8%)
4 (11.4%)
66 (89.2%)
31 (88.6%)
p=0.9984
39
23
13 (33.3%)
9 (39.1%)
26 (66.7%)
14 (60.9%)
p=0.7845
Laurén
Classification
Diffuse
Intestinal
58
51
7 (12.1%)
5 (9.8%)
51 (87.9%)
46 (90.2%)
p=0.7674
34
28
11 (32.4%)
11 (39.3%)
23 (67.6%)
17 (60.7%)
p=0.5770
Tumor Location
Cardia
Noncardia
22
81
2 (9.1%)
10 (12.3%)
20 (90.9%)
71 (87.7%)
p=1
11
50
7 (63.6%)
14 (28%)
4 (36.4%)
36 (72%)
p=0.0363
Stage
I/II
III/IV
15
94
0
12 (12.8%)
15 (100%)
82 (87.2%)
p=0.2121
7
55
3 (42.9%)
28 (50.9%)
4 (57.1%)
12 (21.8%)
p=0.6906
Lymph node

Regulação Epigenética no Envelhecimento e no Câncer Gástrico P á g i n a | 127
Artigos
mestasis
Present
Absent
95
14
11 (11.6%)
1 (7.1%)
84 (88.4%)
13 (92.9%)
p=1
57
5
21 (36.8%)
1 (20%)
36 (63.2%)
4 (80%)
p=0.6473
Distant metastasis
Present
Absent
Unkown
29
63
17
3 (10.3%)
7 (11.1%)
26 (89.7%)
56 (88.9%)
p=1
16
42
4
6 (37.5%)
15 (35.7%)
10 (62.5%)
27 (64.3%)
p=1
IHC: immunohistochemistry assay; NGM: normal gastric mucosa; GC: gastric cancer; *significant p value
after multiple comparison corrections.

Regulação Epigenética no Envelhecimento e no Câncer Gástrico P á g i n a | 128
Artigos
Table 3: SIRT1 and SIRT3 promoter methylation and protein expression in tumor and normal gastric samples.
SIRT1 SIRT3
IHC
MSP
Positive Negative p Positive Negative p
Tumor M
U
M
U
M
U
4 (66.7%) 2 (33.3%) p=0.1827 5 (33.3%) 10 (66.7%) p=0.0047*
42 (93.3%) 3 (6.7%) 20 (83.3%) 4 (16.7%)
Normal 3 (100%) 0 (0%) p=0.2452 2 (66.7%) 1 (33.3%) p=0.8206
6 (42.9%) 8 (57.1%) 9 (81.8%) 2 (18.2%)
Total 7 (77.8%) 2 (22.2%) p=0.8409 7 (38.9%) 11 (61.1%) p=0.0033*
48 (81.4%) 11 (18.6%) 29 (82.9%) 6 (17.1%)
IHC: immunohistochemistry assay; MSP: methylation specific PCR; M: methylated; U: unmethylated; *significant p value after multiple comparison
corrections.

Regulação Epigenética no Envelhecimento e no Câncer Gástrico P á g i n a | 129
Artigos
Figure 1: Immunohistochemical detection of SIRT1 antibody. a) and b) negative and
positive immunostaining of normal gastric tissue, respectively; c) and d) diffuse and
intestinal type gastric adenocarcinoma samples. Magnification: 40x.

Regulação Epigenética no Envelhecimento e no Câncer Gástrico P á g i n a | 130
Artigos
Figure 2: Immunohistochemical detection of SIRT3 antibody. a) and b) negative and
positive immunostaining of normal gastric tissue, respectively; c) and d) diffuse and
intestinal type gastric adenocarcinoma samples. Magnification: 100x (c) and 40x (a,b
and d).
Figure 3: Methylation analysis by MSP. a) and b) SIRT1 and SIRT3 promoter showing
unmethylated bands of 112bp and 167bp, respectively. L: size marker; M: methylated;
U: unmethylated.

Regulação Epigenética no Envelhecimento e no Câncer Gástrico P á g i n a | 131
Artigos
4.2.3 SIRT1, IGFBP-3 and CAV1 promoter DNA methylation in aging process
Submetido como Research Letter à “Age and Ageing” em 04/03/11.
SIRT1, IGFBP-3 and CAV1 promoter DNA methylation in aging process
Running head: DNA methylation in aging
Carolina Oliveira Gigek1; Luara Carolina Frias Lisboa1; Patricia Natalia Oliveira Silva1;
Mariana Ferreira Leal1, Roger W. de Labio2; Spencer Luiz Marques Payão2; Marilia de
Arruda Cardoso Smith1 1 Genetics Division, Departament of Morphology and Genetics Universidade
Federal de São Paulo, SP, Brazil. 2 Genetics and Molecular Biology, Hemocentro, Faculdade de Medicina de Mar´ılia
(FAMEMA), Marília, SP, Brazil
Keywords: aging, DNA methylation, epigenetic
SIR - Nowadays, epigenetics is defined as the heritable changes in gene
activity and expression, and, as the stable, long-term alterations in the transcriptional
potential of a cell that are not necessarily heritable [1]. The most known epigenetic
modifications are DNA methylation and histone post-transcriptional modifications. DNA
methylation consists in an addition of a methyl group on the aromatic ring of 5-carbon
from the cytosine ring of a CpG dinucleotide. This dinucleotide is concentrated in
particular gene promoters, called CpG islands, where it can regulate gene’s expression,
often blocking transcription when methyl group is present [1].
The human genome is a fragile and highly conserved structure which
accumulates a wide range of damaging alterations with age, despite continuous
surveillance and repair [2]. The relationship between epigenetics and aging was
proposed many years ago, showing a decrease in the genomic global DNA methyation
in ageing [3]. In addition to this global hypomethylation, a number of specific loci have

Regulação Epigenética no Envelhecimento e no Câncer Gástrico P á g i n a | 132
Artigos
been described as becoming hypermethylated during the ageing process [1].
Interesting, this events are known epigenetic alterations in cancer [4], which suggests
the accumulation of epigenetic alterations during ageing may directly contribute to
malignant transformation.
We selected three genes, SIRT1, IGFBP-3 and CAV-1, in order to
evaluate DNA promoter methylation in the aging process. To our knowledge, this is a
novel study of DNA promoter methylation of these genes in aging process.
Material and Methods
1. Samples
Methylation status of SIRT1, IGFBP-3 and CAV-1 were evaluated in
peripheral blood samples from about 50 healthy older adults, with exclusion criteria for
depression, stroke, dementia and low cognitive levels using Mini-Mental State Exam
(MMSE), and 60 young adults controls. The mean age was 70.06 ± 9.644 years and
21.46 ± 2.482 years, respectively. All older individuals were selected at Neurology
Department of Universidade Federal de São Paulo – UNIFESP. The younger group was
formed by healthy volunteers also of UNIFESP.
2. DNA Extraction and Methylation Specific PCR (MSP)
DNA extraction was performed using the QIAamp DNA Blood Midi Kit
(Qiagen). To evaluate methylation status, genomic DNA (200 ng) of all samples
underwent bisulfite modification using EpiTect Bisulfite Kit (Qiagen, Germany) according
to the manufacturer’s instructions. Specific primers for MSP, located within genes’
promoter, are described in Table 1. Briefly, PCR reaction was carried out in a 25μL
volume with 200μmol/L of MgCl2, 100ng of DNA, 200 pmol/L of primers and 1.25 units
of Taq (LGC, Brazil). After initial denaturating for 5 min at 94ºC, 40 cycles at 94ºC for 45
s, at respectively temperature (see Supplementary data) for 45 s, and at 72ºC for 30 s,
samples were carried out, followed by a final extension for 5 min at 72ºC.
Results were scored when emerged a clear visible band on the
electrophoresis gel with the methylated and unmethylated primers [5] (Figure 1).
3. Statistical analyses