slf dti segmentation 2005
TRANSCRIPT

8/3/2019 SLF DTI Segmentation 2005
http://slidepdf.com/reader/full/slf-dti-segmentation-2005 1/16
Segmentation of Subcomponents within theSuperior Longitudinal Fascicle in Humans:A Quantitative, In Vivo, DT-MRI Study
Nikos Makris1,2, David N. Kennedy 1, Sean McInerney 1, A. Gregory Sorensen3, Ruopeng Wang3, Verne S. Caviness, Jr1
and Deepak N. Pandya 2
1Harvard Medical School Departments of Neurology andRadiology Services, Center for Morphometric Analysis, HST A.Martinos Center, Massachusetts General Hospital, Boston, MA 02129, USA, 2Boston University School of Medicine,
Department of Anatomy and Neurobiology, Boston, MA 02215,USA and 3Harvard Medical School Departments of Neurology and Radiology Services, HST A. Martinos Center, MassachusettsGeneral Hospital, Boston, MA 02129, USA
Previous research in non-human primates has shown that the
superior longitudinal fascicle (SLF), a major intrahemispheric fiber
tract, is actually composed of four separate components. In humans,
only post-mortem investigations have been available to examine the
trajectory of this tract. This study evaluates the hypothesis that the
four subcomponents observed in non-human primates can also be
found in the human brain using in vivo diffusion tensor magnetic
resonance imaging (DT-MRI). The results of our study demonstrated
that the four subdivisions could indeed be identified and segmentedin humans. SLFI is located in thewhitematter of the superior parietal
and superior frontal lobes and extends to the dorsal premotor and
dorsolateral prefrontal regions. SLF II occupiesthe central coreof the
white matter above the insula. It extends from the angular gyrus to
the caudal--lateral prefrontal regions. SLF III is situated in the white
matter of the parietal and frontal opercula and extends from the
supramarginal gyrus to the ventral premotor and prefrontal regions.
The fourth subdivision of the SLF, the arcuate fascicle, stems from
the caudal part of the superior temporal gyrus arches around the
caudal end of the Sylvian fissure and extends to the lateral prefrontal
cortex along with the SLF II fibers. Since DT-MRI allows the precise
definition of only the stem portion of each fiber pathway, the origin
and termination of the subdivisions of SLF are extrapolated from theavailable data in experimental material from non-human primates.
Keywords: association fiber pathway, cerebral cortex, DT-MRI, superior
longitudinal fasciculus
Introduction
Traditionally the superior longitudinal fascicle (SLF) is consid-
ered to be a major association fiber pathway that connects the postrolandic regions (i.e. parieto-temporal association areas) with the frontal lobe and vice versa. Burdach (1822), followinga description by Reil and Autenrieth (1809), was the first todelineate this pathway in some detail. Subsequently, Dejerine
(1895), while describing different association fiber pathways,gave a detailed account of this fiber bundle that has remainedthe mainstay for interpreting many clinical disorders (e.g.Geschwind, 1965a). Moreover, Ludwig and Klingler (1956)and Heimer (1983), using gross dissection, further demon-
strated the trajectory of this fiber system in the human. Inrecent years, several studies have described the course of SLFusing the diffusion tensor magnetic resonance imaging (DT-MRI) technique (Makris et al ., 1997; Mori et al ., 1999; Pouponet al , 2000; Catani et al ., 2002; Mamata et al ., 2002). Although
the available descriptions of this fiber bundle provide basicanatomical information, the detailed understanding of SLFremains to be ascertained. Both Burdach and Dejerine sug-gested that there is a fiber tract stemming from the caudal part
of the superior temporal gyrus that arches around the Sylvian
fissure and advances forward to end within the frontal lobe.
They named this bundle the arcuate fascicle (AF) and consid-
ered it as part of SLF, using interchangeably the terms ‘superior
longitudinal’ or ‘arcuate fasciculus’ in their descriptions. Recent
observations from neuroimaging studies agree with this de-
scription. In contrast to these classical descriptions and the
neuroimaging studies, non-human primate studies showed the
SLF and AF to be two separate entities that have separatetrajectories (Petrides and Pandya, 1984,1988). How the AF and
SLF relate to each other remains to be clarified in humans.Experimental observations in non-human primates have
shown that SLF is composed of four components (Petrides
and Pandya, 1984). The major component of SLF, i.e. SLF II, is
derived from the caudal--inferior parietal region corresponding
to the angular gyrus in the human and terminates within the
dorsolateral frontal region. The dorsal component of SLF (SLF I)
appears to originate from the medial and dorsal parietal cortex
and ends in the dorsal and medial part of the frontal lobe. The
ventral component of SLF (SLF III) originates from the supra-
marginal gyrus and terminates predominantly in the ventral
premotor and prefrontal areas. It has been reported that the AF,the fourth division of SLF, originates from the caudal part of the
superior temporal gyrus and runs contiguous with the fibers of
SLF II above the Sylvian fissure and insula (Petrides and Pandya,
1988). Comparable organization of SLF in the human remains to
be delineated. Available anatomical information regarding the trajectories of
SLF in the human is derived from post-mortem material. More
refined neuroimaging techniques such as DT-MRI, as mentionedabove, have made possible the segmentation and volumetric
analysis of various fiber bundles (Makris, et al ., 2002a) as well as
the study of their trajectory (Mori, et al ., 1999; Lori, et al ., 2002)
in the human in vivo . In the present study we have identified
and segmented various subcomponents of SLF, including the AF,
and investigated their volumetry, anisotropy and topographic
anatomy, using the DT-MRI method combined with anatomicalknowledge in four human normal subjects in vivo . We have also
extrapolated the non-human primate literature data for the SLF
to the human DT-MRI SLF data presented herein to provide
anatomical inferences for these fiber pathways in terms of their
origins and terminations (Makris, et al ., 2002b). Combination of
this detailed information regarding the quantification of this
fiber system (Makris et al ., 2002b) with tractographic ap-
proaches for trajectory information (Mori et al ., 1999; Lori
et al ., 2002) would allow better anatomical--functional correla-
tional studies as well as evaluation of white matter structures in
clinical conditions.
Cerebral Cortex V 15 N 6 Ó Oxford University Press 2004; all rights reserved
Cerebral Cortex June 2005;15:854--869doi:10.1093/cercor/bhh186
Advance Access publication December 8, 2004

8/3/2019 SLF DTI Segmentation 2005
http://slidepdf.com/reader/full/slf-dti-segmentation-2005 2/16
Materials and Methods
Four normal right-handed male human subjects (mean age 35 years) were scanned using the DT-MRI technique to outline the SLF includingtheAF. Forthe acquisition of these data thefollowing protocol was used.Magnetic resonance imaging was performed using a General Electric1.5 T Signa imaging system (modified for instant imaging by AdvancedNMR, Inc., Wilmington, MA). Subjects were positioned in the magnetalong their anterior commissure (AC)--posterior commissure (PC) line.
This alignmentwas adjusted andverified using quick scout imaging priorto the remainder of the study MR acquisitions. Scans included conven-tional MRI (multi-slice T 1-weighted sagittal scan for prescription of subsequent images) and echo-planar DT-MRI. The echo-planar-based
protocol included automatic magnetic field shimming and diffusiontensorimaging in axial planes. Theaxial imaging covered theentire brain(45 axial sections). We assumed Gaussian behavior of the water self-diffusion, and sampled the diffusion tensor, D , using an echo-planarimaging (EPI) technique to sample the diffusion tensor on a voxel-
by-voxel basis. For each tensor acquisition six directions of high b valueimages and one low b value image (b ~ 0) were acquired. The followingtechnical parameters were used: T R = 8000 ms, T E = 96 ms, the ‘catch andhold’ readout option was employed (Sorensen, et al ., 1999), averages =12,numberof slices=45, slicethickness=3 mm, no interslice gap, field of
view = 403 20 cm,matrix = 2563 128, in-plane voxel resolution=1.5631.56 mm2, high b value images used b = 568 s/mm2. We acquired each
dataset of45 axial slices in three slabs of15 slices eachfor 11.2min/slab. The signal-to-noise ratio (SNR) was 30.
All of the axially acquired images were reformatted into the coronalorientation prior to the tensor calculations to facilitate anatomicinterpretation. The locations of AC, PC and interhemispheric fissureas well as the cerebral exteriors (anterior, posterior, superior andinferior) were recorded on the coronal images so that all voxel locations
within each image could be represented in the Talairach coordinatesystem. Note that the Talairach transformation was only applied to theresults of pathway segmentation for group trajectory analysis (see
below); all image analysis was performed in the native acquisition. Analysis of SLF was performed on contiguous coronal sections thatsampled the SLF in the anterior--posterior orientation in its completetrajectory. We then identified and delineated within SLF four separatefiber tracts following the criteria described below. These tracts wereSLF I, SLF II (including the horizontal part of the AF or AFh), SLF III, and
the vertical part of the AF, i.e. the AFv. It should be pointed out thatthese four components of the SLF constitute the stem portion of thisfiber bundle. The remaining mass of the fiber tract in the frontal, parietaland temporal lobes where the fibers fan out into sprays and theirextreme peripheries, i.e. where the fibers originate or terminate, couldnot be investigated in the present study (Makris et al ., 1997, 2002a).
This is due to the fact that DT-MRI does not provide the capability todistinguish among multiple fiber orientations within a single voxel(Makris et al ., 2002b).
Tensor solutions, generation of principal eigenvector maps (PEMS)and color coding were performed as follows. The orientation of thediffusion tensor, i.e. the orientation of diffusion, was assessed by aneigen-decomposition of the diffusion tensor. The largest eigenvaluecorresponds to the major axis of the diffusion ellipsoid, and hencerepresents the major orientation of diffusion at that location (Basser
et al . 1994). Subsequently, color was assigned for each voxel locationusing the primary eigenvector (corresponding to the largest eigenvalue)of the diffusion tensor. At each voxel the absolute value of the x , y and z
components of the principal eigenvector were used as the red, greenand blue color values (RGB) respectively, such that a red voxel in theimage represents right--left (or left--right), green indicates anterior--
posterior (or posterior--anterior), and blue represents superior--inferior(or inferior--superior). In Figure 1 this color coding scheme is shown
with the appropriate color painted onto a sphere, and the principaleigenvector map (PEM) is the result of color coding a tensor image(Makris et al ., 1997; Pajevic and Pierpaoli, 1999). To further distinguish
white matter fiber pathways from other regions, the color wasmodulated by a measure of anisotropy of the voxel. This emphasizesthe stem portions of the fiber tracts by diminishing the brightness of everything else. Here, ‘anisotropy’ is either fractional (Basser andPierpaoli, 1996) or lattice (Pierpaoli and Basser, 1996a,b) and is
measured, respectively, by the fractional anisotropy index (FA) andlattice anisotropy index (LA). Both fractional and lattice anisotropy arescalar metrics that quantify the degree of anisotropy of a medium suchas the white matter of the brain. Fractional anisotropy is an estimate of the anisotropy of a voxel that is rotationally invariant and latticeanisotropy is basically an averaging of the fractional anisotropy overa neighborhood of voxels. By computing this average over a neighbor-hood of voxels that are contiguous and surround the reference voxeland weighting their contributions according to their distance from the
reference voxel, lattice anisotropy offers a ‘smoother’ representation of the perimeter of the compact stem of a fiber tract. FA and LA range fromzero to one, with zero representing maximally isotropic diffusion andone representing maximally anisotropic diffusion.
Segmentation
Based upon a priori knowledge of the SLF subcomponents from theexperimental animal literature, we were able to define features of relative location and diffusion orientation in order to infer the existenceand location of the stem portions of the SLF I, SLF II, SLF III and AFv fiber
pathways in the human brain. Specifically, progressing through thecoronal sections, the voxels pertaining to the stem portions of fiber
pathways were determined by two factors: (i) relative location and (ii)orientation of diffusion properties of the tissue. It should be pointed outfor clarity that the AF has been considered to be composed of a more
rostral, horizontal portion or AFh and a more caudal, vertical portion or AFv. The regions were evaluated as follows. For SLF I, voxels with tensorinformation that was oriented anterior--posteriorly contained within thedorsomedial part of the hemisphere in the frontal and parietal lobes
were selected. SLF II consisted of voxels with tensor information that was oriented anterior--posteriorly in the region above the insula, theextreme capsule, the claustrum, the external capsule, the lenticularnucleus and the internal capsule. Note that this included the horizontal
part of the arcuate fascicle, AFh, and that the AFh runs contiguous to the ventral surfaceof SLF II and both fiber tracts are oriented in the anterior-- posterior dimension. Because of their proximity and their identicalorientation, the stem portions of these two fiber bundles are in-discernible from each other using images of fiber orientation in DT-MRI. Voxels with tensor information that was oriented vertically at thecaudal end of SLF II were labeled as AFv, corresponding to the vertical
part of the arcuate fascicle. Finally, voxels with tensor information that
was oriented anterior--posteriorly within the frontoparietal opercularregion were assigned to SLF III. Anterior--posteriorly oriented tensors
were constrained to be within a 33° solid angle of the pure anterior-- posterior orientation. A similar constraint was imposed upon thesuperior--inferiorly oriented tensors. This procedure generated a regionof interest (ROI) for the right and left SLF I, SLF II(including AFh), SLF IIIand AFv in each subject.
Characterization of ROI
For each ROI, the size (as number of voxels) and the mean and SD of anisotropy, in terms of FA and LA, were calculated (Basser and Pierpaoli,1996; Pierpaoli and Basser, 1996a). Calculations of left--right volumetricsymmetry were based upon a symmetry coefficient (L -- R)/[0.5(L + R)](Galaburda et al ., 1987). The course of SLF I, SLF II (including AFh),SLF III and AFv was defined in each individual subject using the seg-
mentation ‘color map approach’ (Makris et al ., 1997, 2002a; Mori, 2002).Moreover,the trajectories of SLFI, SLF II (including AFh), SLFIII and AFv were determined in the Talairach coordinate system in each individualsubject. From anatomical landmarks identified (see above), the averagecourse for the group as a whole was derived by calculating the Talairachcoordinate of the center of mass for the SLF I, SLF II (including AFh),SLF III and AFv respectively in each coronal section in which they wereobserved (Talairach and Tournoux, 1988). The tool used for two-dimensional visualization and quantitative processing of DT-MRI data has been described previously (Makris et al ., 2002b). Using in-housesoftware (named ‘surface and volume visualizer’ or SV 2) we createdthree-dimensional reconstructions from the series of two-dimensionaloutlines, which included the voxels manually selected on each coronalsection. Three-dimensional surfaces were displayed and visualized usingtransparency and color to elucidate the context and relationships
between structures in the same coordinate space.
Cerebral Cortex June 2005, V 15 N 6 855
8/3/2019 SLF DTI Segmentation 2005
http://slidepdf.com/reader/full/slf-dti-segmentation-2005 3/16
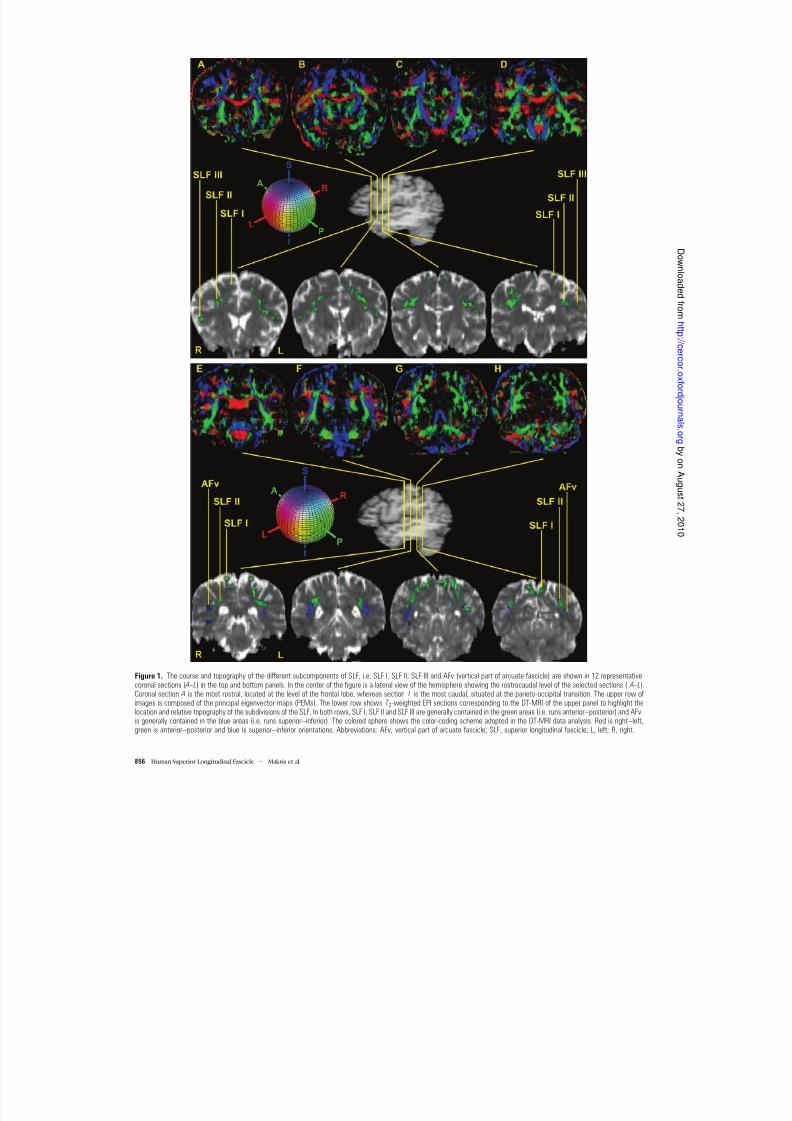
Figure 1. The course and topography of the different subcomponents of SLF, i.e. SLF I, SLF II, SLF III and AFv (vertical part of arcuate fascicle) are shown in 12 representativecoronal sections ( A-- L) in the top and bottom panels. In the center of the figure is a lateral view of the hemisphere showing the rostrocaudal level of the selected sections ( A-- L).Coronal section A is the most rostral, located at the level of the frontal lobe, whereas section L is the most caudal, situated at the parieto-occipital transition. The upper row ofimages is composed of the principal eigenvector maps (PEMs). The lower row shows T 2-weighted EPI sections corresponding to the DT-MRI of the upper panel to highlight thelocation and relative topography of the subdivisions of the SLF. In both rows, SLF I, SLF II and SLF III are generally contained in the green areas (i.e. runs anterior--posterior) and AFvis generally contained in the blue areas (i.e. runs superior--inferior). The colored sphere shows the color-coding scheme adopted in the DT-MRI data analysis. Red is right--left,green is anterior--posterior and blue is superior--inferior orientations. Abbreviations: AFv, vertical part of arcuate fascicle; SLF, superior longitudinal fascicle; L, left; R, right.
856 Human Superior Longitudinal Fascicle d Makris et al.
8/3/2019 SLF DTI Segmentation 2005
http://slidepdf.com/reader/full/slf-dti-segmentation-2005 4/16
Results
In this study five main objectives were accomplished: (i) the SLFfiber system was delineated bilaterally in four normal subjects by segmentation of its four subcomponents. Figures 1 and 2 illus-
trate this in one of the four subjects; (ii) biophysical measure-ments such as FA and LA as well as the volumes of SLF I, SLF II(including the horizontal part of the arcuate fascicle, AFh), SLFIII and the vertical part of the arcuate fascicle, AFv, for eachindividual and group were obtained as shown in Table 1; (iii) the
coarse trajectories of SLF I, SLF II (including AFh), SLF III and AFv for each individual were portrayed in Talairach coordinatespace as shown in Figures 4 and 5; (iv) the symmetry coefficientfor left/right volume was calculated for each individual fiber pathway and group as shown in Table 1; (v) three-dimensional
reconstructions of SLF I, SLF II (including AFh), SLF III and AFv were created as shown in Figure 3.
Delineation of the Subcomponents of the SLF in the
Human Brain
In this study four distinct subcomponents of the human SLF were delineated using DT-MRI, based on a priori knowledge
from non-human primates. The entire extent of the stem portions of the four subcomponents of SLF were identified in39 consecutive rostrocaudal coronal sections in four humansubjects. Of these 39 sections, 12 representative sections are
shown in Figure 1A --L in one of the four subjects. The qualitativemorphological analysis was performed in terms of relativelocation, and orientation of diffusion properties of the tissuefor each fiber pathway by direct inspection of the images. InFigure 1, DT-MRI views are shown in the upper part (sections
A --L , in the rostrocaudal dimension in the coronal plane). In thecenter of the figure is a lateral view of the hemisphere showingthe rostrocaudal level of the selected sections (sections A --L ).
The lower panel in Figure 1 shows T 2-weighted EPI sectionscorresponding to the DT-MRI of the upper panel to highlightthe location and relative topography of the subdivisions of the
SLF. In the lower panel sections the locations of SLF I, SLF II,SLF III and AFh are shown in green. In this figure, AFv is shownin blue (sections E --H ).
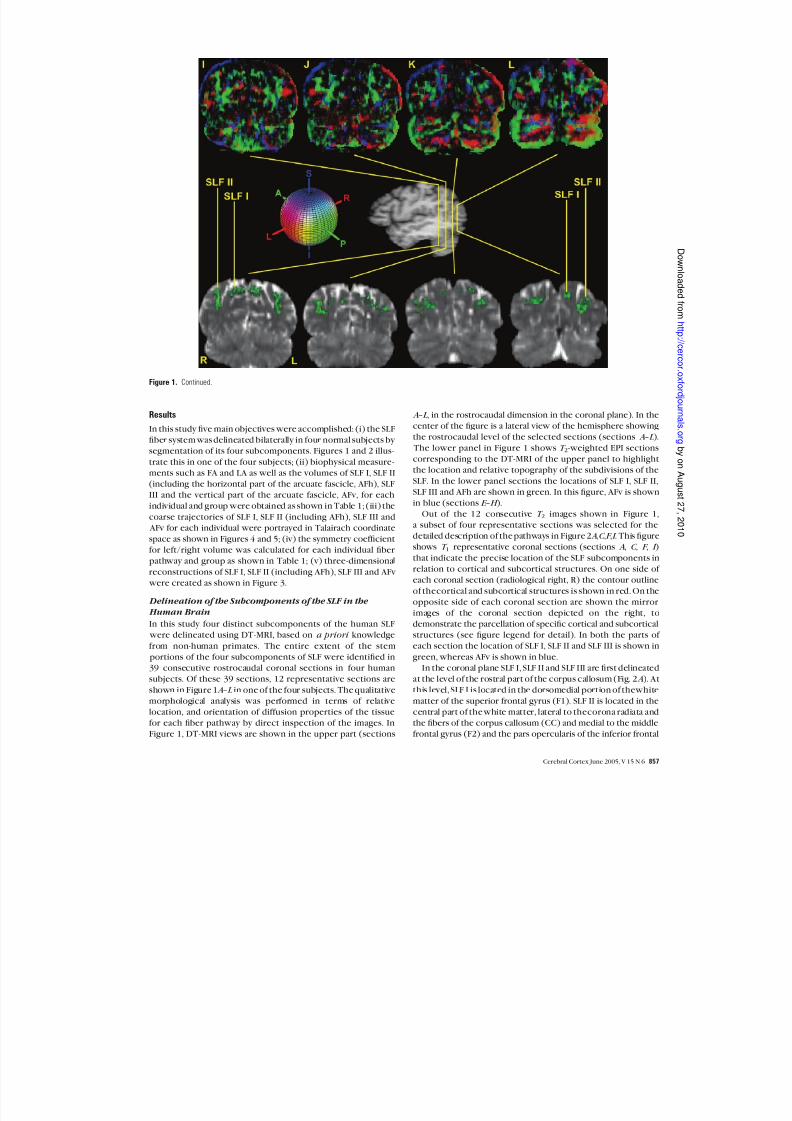
Out of the 12 consecutive T 2 images shown in Figure 1,a subset of four representative sections was selected for thedetailed description of the pathways in Figure 2A ,C ,F ,I . This figureshows T 1 representative coronal sections (sections A , C , F , I )
that indicate the precise location of the SLF subcomponents inrelation to cortical and subcortical structures. On one side of each coronal section (radiological right, R) the contour outline
of thecortical and subcortical structures is shown in red. On theopposite side of each coronal section are shown the mirrorimages of the coronal section depicted on the right, todemonstrate the parcellation of specific cortical and subcorticalstructures (see figure legend for detail). In both the parts of each section the location of SLF I, SLF II and SLF III is shown in
green, whereas AFv is shown in blue.In the coronal plane SLF I, SLF II and SLF III are first delineated
at the level of the rostral part of the corpus callosum (Fig. 2A ). Atthis level, SLF I is located in the dorsomedial portion of thewhite
matter of the superior frontal gyrus (F1). SLF II is located in thecentral part of the white matter, lateral to thecorona radiata andthe fibers of the corpus callosum (CC) and medial to the middlefrontal gyrus (F2) and the pars opercularis of the inferior frontal
Figure 1. Continued.
Cerebral Cortex June 2005, V 15 N 6 857
8/3/2019 SLF DTI Segmentation 2005
http://slidepdf.com/reader/full/slf-dti-segmentation-2005 5/16
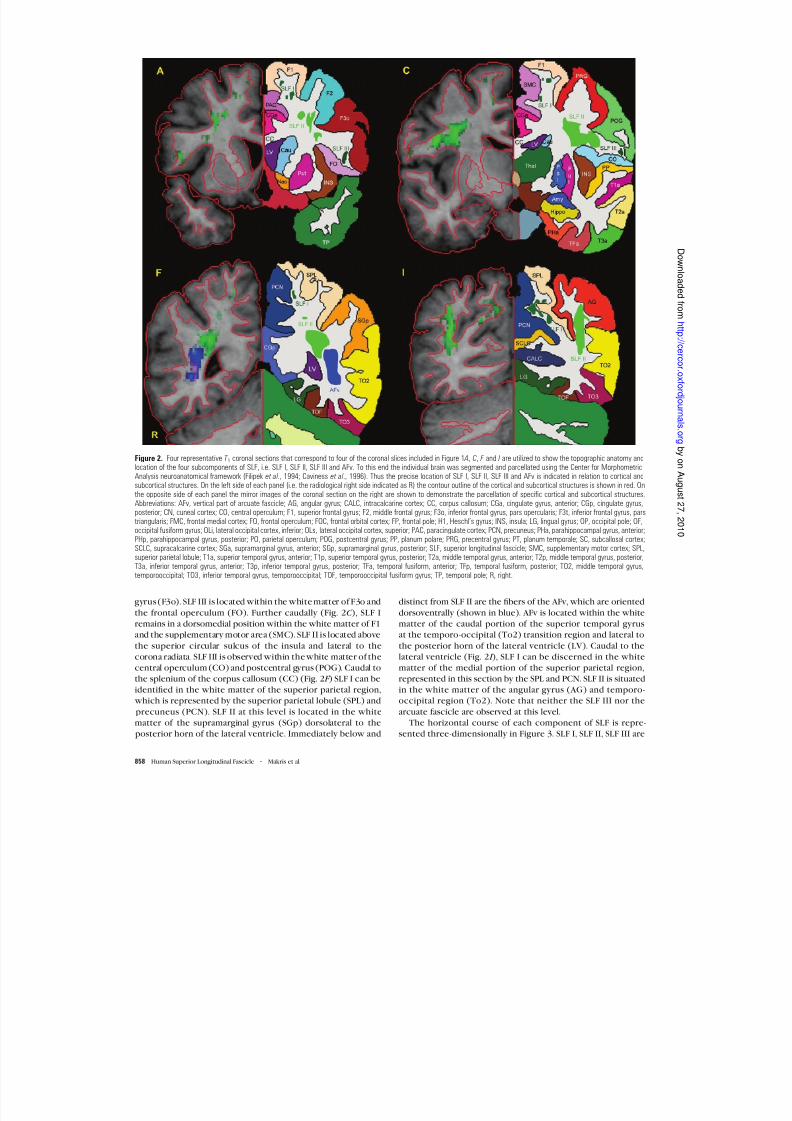
gyrus (F3o). SLF III is located within the white matter of F3o and
the frontal operculum (FO). Further caudally (Fig. 2C ), SLF Iremains in a dorsomedial position within the white matter of F1and the supplementary motor area (SMC). SLF II is located above
the superior circular sulcus of the insula and lateral to thecorona radiata. SLF III is observed within the white matter of thecentral operculum (CO) and postcentral gyrus (POG). Caudal tothe splenium of the corpus callosum (CC) (Fig. 2F ) SLF I can beidentified in the white matter of the superior parietal region, which is represented by the superior parietal lobule (SPL) and
precuneus (PCN). SLF II at this level is located in the whitematter of the supramarginal gyrus (SGp) dorsolateral to the posterior horn of the lateral ventricle. Immediately below and
distinct from SLF II are the fibers of the AFv, which are oriented
dorsoventrally (shown in blue). AFv is located within the whitematter of the caudal portion of the superior temporal gyrusat the temporo-occipital (To2) transition region and lateral to
the posterior horn of the lateral ventricle (LV). Caudal to thelateral ventricle (Fig. 2I ), SLF I can be discerned in the whitematter of the medial portion of the superior parietal region,represented in this section by the SPL and PCN. SLF II is situatedin the white matter of the angular gyrus (AG) and temporo-occipital region (To2). Note that neither the SLF III nor the
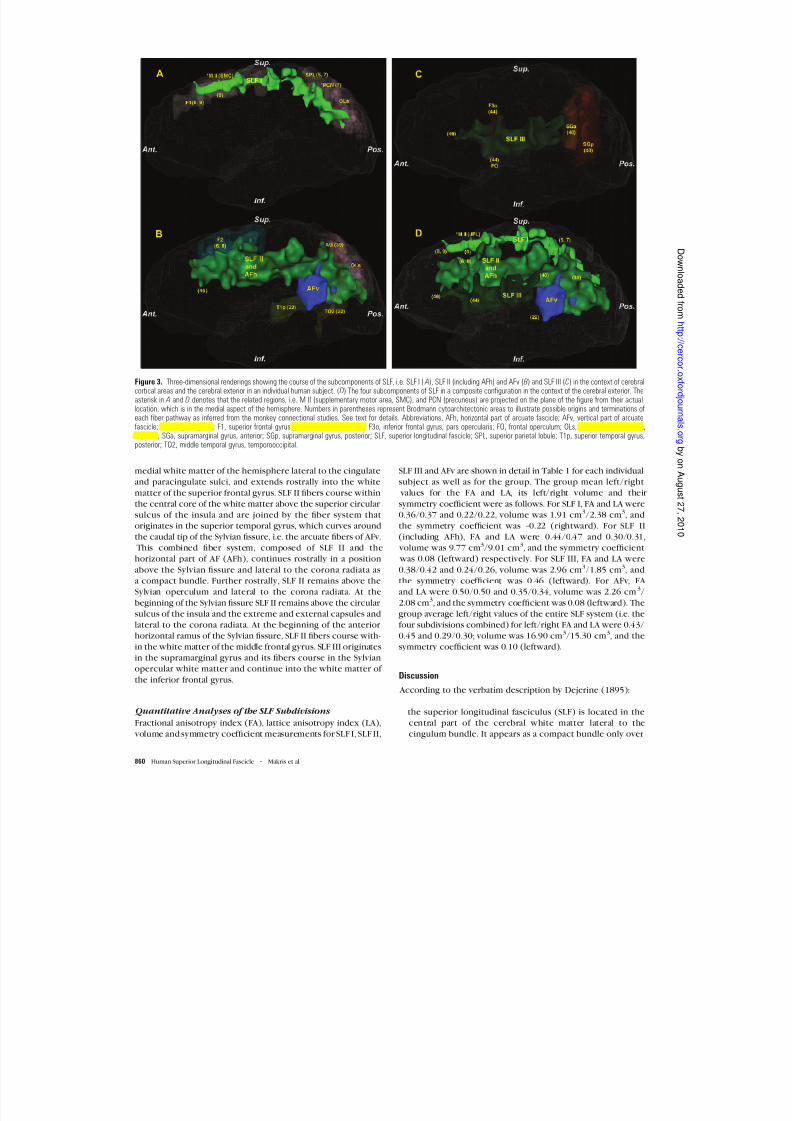
arcuate fascicle are observed at this level. The horizontal course of each component of SLF is repre-
sented three-dimensionally in Figure 3. SLF I, SLF II, SLF III are
Figure 2. Four representative T 1 coronal sections that correspond to four of the coronal slices included in Figure 1 A, C, F and I are utilized to show the topographic anatomy andlocation of the four subcomponents of SLF, i.e. SLF I, SLF II, SLF III and AFv. To this end the individual brain was segmented and parcellated using the Center for MorphometricAnalysis neuroanatomical framework (Filipek et al., 1994; Caviness et al ., 1996). Thus the precise location of SLF I, SLF II, SLF III and AFv is indicated in relation to cortical andsubcortical structures. On the left side of each panel (i.e. the radiological right side indicated as R) the contour outline of the cortical and subcortical structures is shown in red. Onthe opposite side of each panel the mirror images of the coronal section on the right are shown to demonstrate the parcellation of specific cortical and subcortical structures.Abbreviations: AFv, vertical part of arcuate fascicle; AG, angular gyrus; CALC, intracalcarine cortex; CC, corpus callosum; CGa, cingulate gyrus, anterior; CGp, cingulate gyrus,posterior; CN, cuneal cortex; CO, central operculum; F1, superior frontal gyrus; F2, middle frontal gyrus; F3o, inferior frontal gyrus, pars opercularis; F3t, inferior frontal gyrus, parstriangularis; FMC, frontal medial cortex; FO, frontal operculum; FOC, frontal orbital cortex; FP, frontal pole; H1, Heschl’s gyrus; INS, insula; LG, lingual gyrus; OP, occipital pole; OF,occipital fusiform gyrus; OLi, lateral occipital cortex, inferior; OLs, lateral occipital cortex, superior; PAC, paracingulate cortex; PCN, precuneus; PHa, parahippocampal gyrus, anterior;PHp, parahippocampal gyrus, posterior; PO, parietal operculum; POG, postcentral gyrus; PP, planum polare; PRG, precentral gyrus; PT, planum temporale; SC, subcallosal cortex;SCLC, supracalcarine cortex; SGa, supramarginal gyrus, anterior; SGp, supramarginal gyrus, posterior; SLF, superior longitudinal fascicle; SMC, supplementary motor cortex; SPL,superior parietal lobule; T1a, superior temporal gyrus, anterior; T1p, superior temporal gyrus, posterior; T2a, middle temporal gyrus, anterior; T2p, middle temporal gyrus, posterior;T3a, inferior temporal gyrus, anterior; T3p, inferior temporal gyrus, posterior; TFa, temporal fusiform, anterior; TFp, temporal fusiform, posterior; TO2, middle temporal gyrus,temporooccipital; TO3, inferior temporal gyrus, temporooccipital; TOF, temporooccipital fusiform gyrus; TP, temporal pole; R, right.
858 Human Superior Longitudinal Fascicle d Makris et al.

8/3/2019 SLF DTI Segmentation 2005
http://slidepdf.com/reader/full/slf-dti-segmentation-2005 6/16
shown in different shades of green, whereas the vertical part of AF (AFv) component is shown in blue. The overall orientation
of the SLF fiber system is horizontal, except for the vertical partof the arcuate fascicle. SLF I is more medial and dorsal (Figs 1, 2,3A ,D ), whereas SLF III is more ventral and lateral (Figs 1, 2,
3C ,D ). SLF II (including the horizontal part of AF, i.e. AFh) islocated between these two components (Figs 1, 2, 3B ,D ). Thehorizontal part of the arcuate fascicle (AFh) cannot be separatedfrom the SLF II, because the two fiber tracts run horizontally and
adjacent to one another. However, SLF II and the vertical portionof AF (AFv) can be distinguished caudally, where the fibers of the arcuate fascicle arch around the caudal portion of the
Sylvian fissure and their trajectory is vertical in nature (Figs 1, 2,3B ,D ). SLF III is restricted mainly to the anterior part of the cere- bral hemisphere, whereas SLF I and SLF II extend more caudally (Figs 1--3).
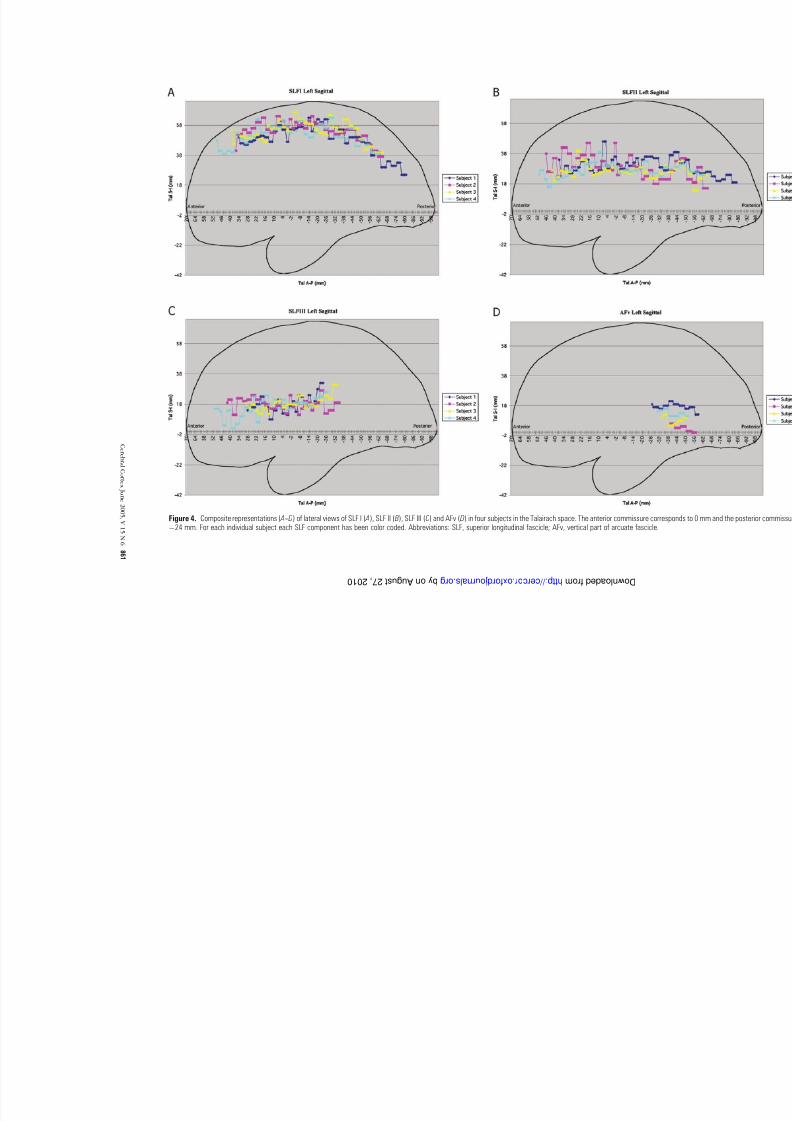
Whereas Figures 1--3 show the coarse trajectory of the SLF inone brain from among the four subjects, Figure 4 showscomposite views of the ROI centers of mass in four subjects
in Talairach coordinate space in lateral cardinal views for theleft side. Each individual case is represented with different colorin these diagrams. Specifically, Figure 4A portrays the sagittal
view of SLF I; Figure 4B represents the course of SLF II (includ-ing the horizontal component of the arcuate fascicle, AFh) in
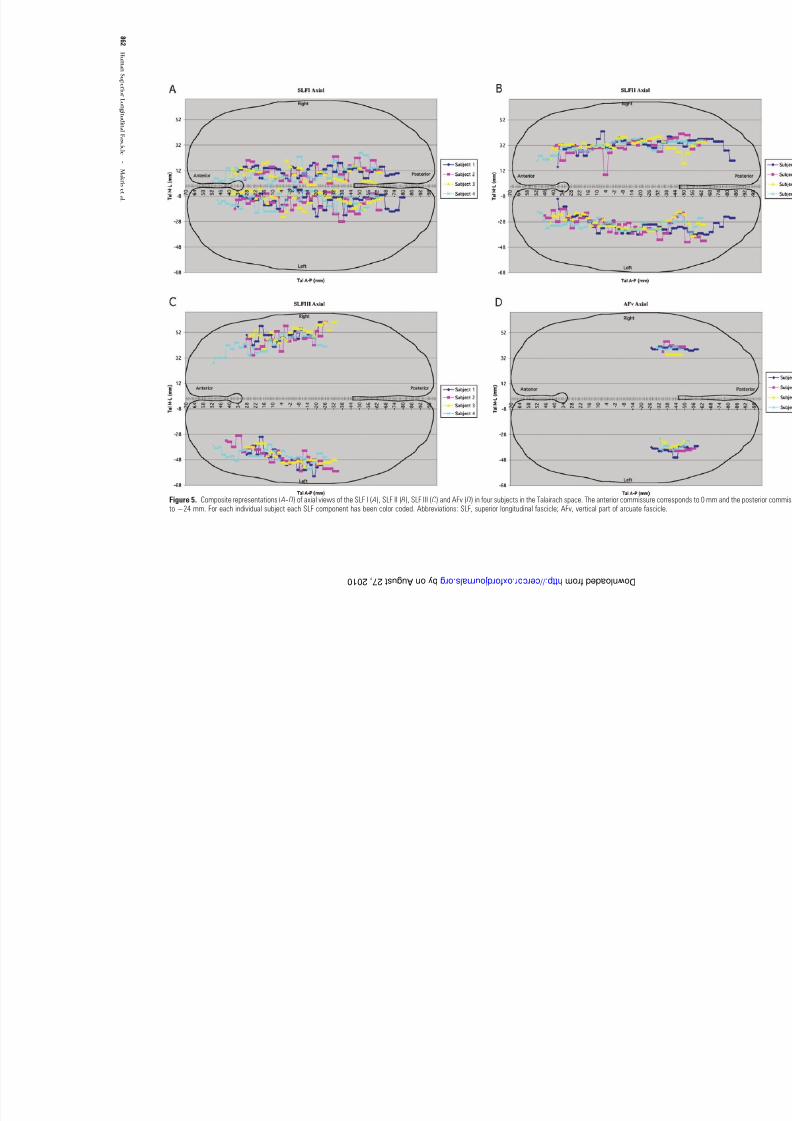
sagittal plane; and Figures 4C ,D show the sagittal representationof SLF III and AFv respectively in four subjects. Figure 5A --D
portrays the axial view of the extent of SLF I, SLF II (including AFh), SLF III and AFv respectively of both hemispheres, in four
subjects in Talairach coordinate space. In each of the casesstudied in Talairach coordinate space, the observed relationship between the pathway coarse trajectories and adjacent cortical
structures is in agreement with the individual analysis results asdemonstrated above (Figs 1 and 2).
From the above description it can be inferred that the SLF, with its four subdivisions, runs through the core of the whitematter of the cerebral hemisphere. SLF I and SLF II originate inthe occipitoparietal region. SLF I is situated around the cingulatesulcus and continues further forward rostrally within the dorsal
Table 1
Statistical results of anisotropy indices (i.e., FA and LA) and volumetry of total SLF and of the SLFI, SLFII, SLFIII and Afv subcomponents of SLF
Region Measure Side Subject 1 Subject 2 Subject 3 Subject 4 Group Mean Group SD
SLF I FA R 0.38 (0.21) 0.36 (0.21) 0.35 (0.16) 0.39 (0.18) 0.37 (0.02)L 0.38 (0.19) 0.36 (0.26) 0.34 (0.19) 0.36 (0.15) 0.36 (0.01)
LA R 0.25 (0.19) 0.22 (0.16) 0.19 (0.11) 0.23 (0.13) 0.22 (0.03)L 0.25 (0.17) 0.23 (0.23) 0.19 (0.14) 0.21 (0.10) 0.22 (0.03)
No. of voxels R 587 266 215 341 352 (164.63)
L 475 210 202 243 282 (129.32)Volume (cm3) R 3.96 1.80 1.45 2.30 2.38 (1.11)L 3.20 1.42 1.36 1.64 1.91 (0.87)
SC ÿ0.21 ÿ0.24 ÿ0.06 ÿ0.34 ÿ0.21 (0.12)SLF II FA R 0.46 (0.17) 0.43 (0.16) 0.48 (0.15) 0.50 (0.16) 0.47 (0.03)
L 0.43 (0.17) 0.41 (0.17) 0.48 (0.15) 0.46 (0.16) 0.44 (0.03)LA R 0.32 (0.15) 0.27 (0.13) 0.32 (0.13) 0.34 (0.15) 0.31 (0.03)
L 0.30 (0.15) 0.26 (0.14) 0.33 (0.14) 0.30 (0.15) 0.30 (0.03)No. of voxels R 1783 922 1529 1105 1335 (392.22)
L 1918 980 1805 1084 1447 (483.11)Volume (cm3) R 12.03 6.22 10.32 7.46 9.01 (2.65)
L 12.95 6.62 12.18 7.32 9.77 (3.26)SC 0.07 0.06 0.17 ÿ0.02 0.07 (0.08)
SLF III FA R 0.41 (0.18) 0.40 (0.15) 0.43 (0.14) 0.44 (0.15) 0.42 (0.02)L 0.41 (0.19) 0.35 (0.16) 0.39 (0.15) 0.38 (0.14) 0.38 (0.02)
LA R 0.27 (0.16) 0.24 (0.12) 0.27 (0.12) 0.28 (0.13) 0.26 (0.02)L 0.28 (0.16) 0.22 (0.13) 0.24 (0.13) 0.24 (0.11) 0.24 (0.03)
No. of voxels R 446 251 162 238 274 (120.80)L 545 485 385 342 439 (92.41)
Volume (cm3
) R 3.01 1.69 1.09 1.61 1.85 (0.82)L 3.68 3.27 2.60 2.31 2.96 (0.62)
SC 0.20 0.64 0.82 0.36 0.51 (0.28)Afv FA R 0.51 (0.17) 0.50 (0.18) 0.44 (0.14) 0.53 (0.15) 0.50 (0.04)
L 0.47 (0.15) 0.52 (0.16) 0.52 (0.14) 0.50 (0.15) 0.50 (0.02)LA R 0.37 (0.16) 0.34 (0.15) 0.28 (0.11) 0.37 (0.14) 0.34 (0.05)
L 0.34 (0.14) 0.36 (0.15) 0.36 (0.12) 0.35 (0.13) 0.35 (0.01)No. of voxels R 493 242 121 375 308 (161.19)
L 333 295 255 457 335 (87.34)Volume (cm3) R 3.33 1.63 0.82 2.53 2.08 (1.09)
L 2.25 1.99 1.72 3.08 2.26 (0.59)SC ÿ0.39 0.20 0.71 0.20 0.18 (0.45)
Total SLF FA R 0.44 (0.18) 0.42 (0.17) 0.46 (0.15) 0.48 (0.17) 0.45 (0.02)L 0.42 (0.18) 0.41 (0.18) 0.46 (0.16) 0.44 (0.16) 0.43 (0.02)Total 0.43 (0.18) 0.42 (0.18) 0.46 (0.16) 0.46 (0.17) 0.44 (0.02)
LA R 0.31 (0.17) 0.27 (0.14) 0.30 (0.14) 0.32 (0.15) 0.30 (0.02)L 0.29 (0.16) 0.26 (0.16) 0.31 (0.14) 0.29 (0.14) 0.29 (0.02)Total 0.30 (0.16) 0.26 (0.15) 0.30 (0.14) 0.31 (0.15) 0.29 (0.02)
No. of voxels R 3308 1681 2027 2059 2269 (713.42)L 3271 1970 2647 2126 2504 (587.89)Total 6579 3651 4674 4185 4772 (1274.68)
Volume (cm3) R 22.33 11.35 13.68 13.90 15.31 (4.82)L 22.08 13.30 17.87 14.35 16.90 (3.97)Total 44.41 24.64 31.55 28.25 32.21 (8.60)
SC ÿ0.01 0.16 0.27 0.03 0.11 (0.13)
Abbreviations: L, left; R, right; SC, symmetry coefficient (L-R)/[0.5(L;plR)]; FA, fractional anisotropy index; LA, lattice anisotropy index.
Cerebral Cortex June 2005, V 15 N 6 859
8/3/2019 SLF DTI Segmentation 2005
http://slidepdf.com/reader/full/slf-dti-segmentation-2005 7/16
medial white matter of the hemisphere lateral to the cingulateand paracingulate sulci, and extends rostrally into the white
matter of the superior frontal gyrus. SLF II fibers course withinthe central core of the white matter above the superior circularsulcus of the insula and are joined by the fiber system thatoriginates in the superior temporal gyrus, which curves aroundthe caudal tip of the Sylvian fissure, i.e. the arcuate fibers of AFv. This combined fiber system, composed of SLF II and thehorizontal part of AF (AFh), continues rostrally in a position
above the Sylvian fissure and lateral to the corona radiata asa compact bundle. Further rostrally, SLF II remains above the
Sylvian operculum and lateral to the corona radiata. At the beginning of the Sylvian fissure SLF II remains above the circularsulcus of the insula and the extreme and external capsules andlateral to the corona radiata. At the beginning of the anteriorhorizontal ramus of the Sylvian fissure, SLF II fibers course with-in the white matter of the middle frontal gyrus. SLF III originatesin the supramarginal gyrus and its fibers course in the Sylvianopercular white matter and continue into the white matter of
the inferior frontal gyrus.
Quantitative Analyses of the SLF Subdivisions
Fractional anisotropy index (FA), lattice anisotropy index (LA), volume and symmetry coefficient measurements for SLF I, SLF II,
SLF III and AFv are shown in detail in Table 1 for each individualsubject as well as for the group. The group mean left/right values for the FA and LA, its left/right volume and their
symmetry coefficient were as follows. For SLF I, FA and LA were0.36/0.37 and 0.22/0.22, volume was 1.91 cm3/2.38 cm3, andthe symmetry coefficient was – 0.22 (rightward). For SLF II(including AFh), FA and LA were 0.44/0.47 and 0.30/0.31, volume was 9.77 cm3/9.01 cm3, and the symmetry coefficient was 0.08 (leftward) respectively. For SLF III, FA and LA were
0.38/0.42 and 0.24/0.26, volume was 2.96 cm3/1.85 cm3, andthe symmetry coefficient was 0.46 (leftward). For AFv, FA
and LA were 0.50/0.50 and 0.35/0.34, volume was 2.26 cm
3
/2.08 cm3, and the symmetry coefficient was 0.08 (leftward). Thegroup average left/right values of the entire SLF system (i.e. thefour subdivisions combined) for left/right FA and LA were 0.43/0.45 and 0.29/0.30; volume was 16.90 cm3/15.30 cm3, and thesymmetry coefficient was 0.10 (leftward).
Discussion
According to the verbatim description by Dejerine (1895):
the superior longitudinal fasciculus (SLF) is located in thecentral part of the cerebral white matter lateral to thecingulum bundle. It appears as a compact bundle only over
Figure 3. Three-dimensional renderings showing the course of the subcomponents of SLF, i.e. SLF I ( A), SLF II (including AFh) and AFv ( B) and SLF III (C) in the context of cerebralcortical areas and the cerebral exterior in an individual human subject. ( D) The four subcomponents of SLF in a composite configuration in the context of the cerebral exterior. Theasterisk in A and D denotes that the related regions, i.e. M II (supplementary motor area, SMC), and PCN (precuneus) are projected on the plane of the figure from their actuallocation, which is in the medial aspect of the hemisphere. Numbers in parentheses represent Brodmann cytoarchitectonic areas to illustrate possible origins and terminations ofeach fiber pathway as inferred from the monkey connectional studies. See text for details. Abbreviations, AFh, horizontal part of arcuate fascicle; AFv, vertical part of arcuatefascicle; AG, angular gyrus; F1, superior frontal gyrus; F2, middle frontal gyrus; F3o, inferior frontal gyrus, pars opercularis; FO, frontal operculum; OLs, lateral occipital cortex,superior; SGa, supramarginal gyrus, anterior; SGp, supramarginal gyrus, posterior; SLF, superior longitudinal fascicle; SPL, superior parietal lobule; T1p, superior temporal gyrus,
posterior; TO2, middle temporal gyrus, temporooccipital.
860 Human Superior Longitudinal Fascicle d Makris et al.
8/3/2019 SLF DTI Segmentation 2005
http://slidepdf.com/reader/full/slf-dti-segmentation-2005 8/16
Figure 4. Composite representations ( A-- D) of lateral views of SLF I ( A), SLF II ( B), SLF III (C) and AFv ( D) in four subjects in the Talairach space. The anterior commissure corresÿ24 mm. For each individual subject each SLF component has been color coded. Abbreviations: SLF, superior longitudinal fascicle; AFv, vertical part of arcuate fascicle.
Cer eb r a l Cor t exJ une2 0 0 5 V1 5 N6
8 6 1
b y o n A u g u s t 2 7 , 2 0 1 0 h t t p : / / c e r c o r . o x f o r d j o u r n a l s . o r g D o w n l o a d e d f r o m
8/3/2019 SLF DTI Segmentation 2005
http://slidepdf.com/reader/full/slf-dti-segmentation-2005 9/16
Figure 5. Composite representations ( A-- D) of axial views of the SLF I ( A), SLF II ( B), SLF III (C) and AFv ( D) in four subjects in the Talairach space. The anterior commissure corrto ÿ24 mm. For each individual subject each SLF component has been color coded. Abbreviations: SLF, superior longitudinal fascicle; AFv, vertical part of arcuate fascicle.
8 6 2
Huma nS uper i or L ongi t ud i na l F a s ci cl e
d
Ma k r i s et a l
b y o n A u g u s t 2 7 , 2 0 1 0 h t t p : / / c e r c o r . o x f o r d j o u r n a l s . o r g D o w n l o a d e d f r o m

8/3/2019 SLF DTI Segmentation 2005
http://slidepdf.com/reader/full/slf-dti-segmentation-2005 10/16
the parietal operculum. It remains above the dorsal Sylvian
operculum and the circular sulcus, and lateral to the corona radiata. In coronal sections its shape is triangular: its infero-lateral angle corresponds to the base of the Sylvian oper-culum, its infero-medial angle corresponds to the externalcapsule entering in the interspace between the externalcapsule and the claustrum, and its superior angle is merged
in the neighboring layer of fibers. Caudally, at the level of the
posterior tip of the Sylvian fissure, SLF curves downwardcrossing the fibers of the corona radiata and the corpus
callosum and descending into the parieto-occipito-temporalgyri. The most superficial fibers radiate to the anterior part of the superior temporal gyrus; the middle layer of fibers endsin the posterior part of the superior temporal and middletemporal gyri. The deepest fibers radiate into the supramar-ginal gyrus, the angular gyrus, and the gyri of the externaloccipital surface.
Dejerine also reported that rostrally, according to Meynert(1865), SLF ends in the Rolandic operculum and in the
operculum of the inferior frontal gyrus and that, according to
Schnopfhagen (1890), it crosses the midline at the genu of thecorpus callosum and ends in the contralateral frontal corticalregions. In summary, SLF, as described above, would be anassociation fiber pathway connecting the lateral part of thetemporo-parieto-occipital regions with the convexity of thefrontal lobe. This bundle was designated the fasciculus arcuatus
by Burdach (Dejerine, 1895). Dejerine considered that this fiber pathway is composed of short association fibers connecting twoneighboring gyri and that only its deep layers, particularly the
ones that are in relationship with the external capsule, containsome longer fibers that can connect more distant gyri. In hislesion material Dejerine was not able to follow degeneratedfibers of SLF beyond the gyri adjacent to the lesion.
Most of Dejerine’s conclusions regarding the SLF were based
on human material prepared with myelin stain techniques. Incontrast to descriptive studies of fiber bundles in humans, in themonkey the fiber pathways have been traced using experi-mental techniques that allow detection of the exact origin andtermination of the association pathways. Thus, it has been
shown in the monkey that, from the parietal region, fibers travellongitudinally in the central core of the white matter, remainingabove the circular sulcus of the insula and lateral to the corona radiata (Petrides and Pandya, 1988). The main contingent of fibers of SLF has been designated SLF II. These fibers terminate indorsal frontal Brodmann (1905) areas 6 and 8 as well as area 46.
The contingent of fibers that originates from the caudal superiortemporal gyrus and the cortex of the superior temporal sulcus
curves around the depth of the caudal portion of the Sylvianfissure and runs along with the fibers of SLF II (Petrides and
Pandya, 1984). This group of fibers has been designated thearcuate fasciculus (AF). AF fibers continue into the frontal lobeand terminate predominantly in the dorsal part of area 8 and inarea 46. AF becomes distinct mainly at the parietal opercularlevel. It can be distinguished in most of its course from sur-
rounding fiber systems, such as the corona radiata, the sub-callosal fasciculus of Muratoff, the occipitofrontal fascicle, theexternal and extreme capsules, and the U-fibers. The experi-mental observations in monkeys also have revealed two addi-
tional components of SLF, i.e. SLF I and SLF III. SLF I originates inthe superior and medial parietal cortex, runs around thecingulate sulcus as well as in the white matter of the superior
parietal and frontal regions and terminates in the dorsal part of frontal areas 6, 8, area 9 and in the supplementary motor region
(M II). SLF III originates from the rostral part of the inferior parietal lobule (supramarginal gyrus) and terminates in the ventral part of area 6 and in areas 44 and 46 in the frontal lobe. Thus, in non-human primates the SLF is quite broad and itsterminations in the frontal lobe involve a number of regions. It
should be pointed out that the SLF I, SLF II, SLF III and AF fiber
pathways are bidirectional (Petrides and Pandya, 2002). According to our in vivo DT-MRI analysis, SLF also can be
divided into four components, in humans, i.e. SLF I, SLF II, SLF III
and AF. The analyses of the individual data sets (Figs 1, 2 and 3)as well as of the individual and grouped data mapped onto the Talairach coordinate space (Figs 4 and 5) have enabled us toelucidate the topography of the stem portion of the foursubdivisions of SLF in the human brain. To interpret the locationof the various pathways from the T 1-weighted and DT-MRI
data, we have followed the criteria of the semi-automated MRI- based morphometric method described in previous reports(Rademacher et al ., 1992; Filipek et al ., 1994; Caviness et al .,
1996). Figure 2 shows an example of the application of this
method to segment and parcellate the cortical and subcorticalstructures of T 1-weighted coronal sections and to compare theDT-MRI observations of SLF I, SLF II, SLF III and AF with thecortical and subcortical parcellation units.
Our observations suggest that SLF I is situated within the white matter of the superior parietal lobule (Brodmann [1909]
area 7), the superior parts of the precuneus (medial area 7), postcentral gyrus (area 5), precentral gyrus (area 4), and the posterior part of superior frontal gyrus (areas 8 and 9) as well as
the supplementary motor area (area 6) (Figs 2, 3A , 4A and 5A ).SLF II, including the horizontal portion of AF (AFh), is located within the white matter of the occipito-temporo-parietal transi-tion region (area 19), angular gyrus (area 39), supramarginalgyrus (area 40), postcentral gyrus (areas 3, 1 and 2), precentral
gyrus (area 4), and middle frontal gyrus (areas 6 and 46) (Figs 2,3B , 4B and 5B ). SLF III maintains a lateral position in the hemi-spheric white matter, coursing in the anterior--posterior dimen-sion within the dorsal Sylvian operculum (i.e. parietal area 40 —supramarginal gyrus, ventral part of pericentral areas 43, 2, 4 and
6, and frontal premotor area 44 — pars opercularis) (Figs 2, 3 C ,4C and 5C ). The vertical portion of the arcuate fascicle (AFv) ismapped within the white matter of the supramarginal gyrus(area 40), the posterior part of the superior temporal gyrus(area 22), and the temporo-occipital region (area 37) (Figs 2, 3B ,4D and 5D ).
The observations shown in Figures 1 and 2 also correspond tothe Talairach coordinate space analysis as shown in Figures 4
and 5 for the SLF I, SLF II, SLF III and AFv. The trajectories andthe topographic anatomy of the stem portion of these fiber path-
ways, in the four human subjects studied using DT-MRI, showedsimilarities to the observed trajectories and topography of thestem portion of SLF I, SLF II, SLF III and AF in the monkey usingexperimental techniques. In addition, compared to the monkey data, the present results showed that in the human the SLF I,
SLF II and SLF III extend more posteriorly within the dorsal parietal regions and parieto-occipital border zones. These fiberfascicles also extend more anteriorly within the frontal associ-ation areas. This may be due to the fact that the parietal and fron-
tal association regions are considerably expanded in the human.Using DT-MRI we were able to define fiber pathways at the
level of their stem portion (Makris et al ., 1997) in cases where
Cerebral Cortex June 2005, V 15 N 6 863

8/3/2019 SLF DTI Segmentation 2005
http://slidepdf.com/reader/full/slf-dti-segmentation-2005 11/16
the stem portion of pathways have characteristic orientations
that are contrasted from the orientations of adjacent compon-ents. However, to delineate the entire trajectory of a fiber tract,its origins and terminations must be traced as well. Althoughthis is possible with experimental methods in non-human primates, it is not feasible for long fiber tracts in the human(Makris et al ., 2002b). Therefore, regarding the specific origins
and terminations of the SLF, we could extrapolate from the
monkey experimental material. Taking into account the evolu-tionary expansion of associational cortical regions in the human,
the topological similarities observed for the stem portions of theSLF I, SLF II, SLF III and AFv in the human subjects as comparedto the monkey lend credence to the possibility that there may be similarities with respect to the origins and terminations of these pathways. It can be inferred that the SLF I in humans may originate from the superior and medial parietal cortex (superior parietal lobule areas 5 and 7 and precuneus area 7), run through
the white matter of the superior parietal and frontal regions andterminate in the dorsal part of areas 6, 8 and 9 and thesupplementary motor region (Fig. 3A ). Likewise, SLF II may
originate from the posterior--inferior parietal region (angular
gurus — area 39), run through the central core of the whitematter above the Sylvian fissure and terminate in the prefrontalregion (areas 46, 6 and 8) (Fig. 3B ). AF may connect the caudalsuperior temporal area 22 with the frontal lobe areas 8 and 46(Fig. 3B ). In contrast, SLF III may originate from the supra-marginal gyrus of the inferior parietal lobule (area 40), occupy
the white matter of the pericentral and frontal opercularregions and connect preferentially with area 44 (pars opercu-laris) and ventral area 6 of the frontal lobe and to a lesser degree
with area 46 (Fig. 3C). It should be pointed out that theseconnections may be bidirectional in nature in humans as has been shown in monkeys (Petrides and Pandya, 2002).
Functional Role of Subdivisions of SLF
Given that DT-MRI offers a method to delineate and measurethe four subcomponents of SLF, it is worth reviewing thefunctional role of this fiber system to aid in reasoning based onquantitative observations of these structures. Inference aboutthe role of SLF in humans comes from multiple sources such asthe course of its fiber trajectories, non-human primate studies,lesion-deficit correlations in humans as well as the known
functional role of the cortical areas that are interconnected by these fiber pathways derived from functional neuroimaging.
SLF I
Several studies have shown that the superior parietal lobulecodes the location of body parts in a body-centered coordinate
system (e.g. Duffy and Birchfiel, 1971; Sakata et al ., 1973;Mountcastle et al ., 1975; Lacquaniti et al ., 1995). Therefore,SLF I, by virtue of its interconnection with medial and superior parietal regions, with M II and with the dorsal premotor region,can contribute to the regulation of higher aspects of motor
behavior. Lesions of the dorsal premotor regions in humans andmonkeys result in severe deficits in conditional associative tasksin which different competing motor acts must be selected on
the basis of appropriate conditional rules (Petrides, 1982, 1987;Halsband and Passingham, 1982; Petrides and Pandya, 2002).
SLF II
It has been shown that the caudal part of the inferior parietallobule plays a major role in visual and oculomotor aspects of
spatial function in monkeys (Mountcastle etal ., 1975; Hyvarinenand Shelepin, 1979; Mesulam, 1981; Posner et al ., 1984;
Andersen and Gnadt, 1989; Goldberg and Segraves, 1989; Bisley and Goldberg, 2003). Moreover, humans with damage in thecaudal inferior parietal area exhibit severe impairment in spatialattention (e.g. Mesulam, 1981; Posner et al ., 1984). Therefore,SLF II can be viewed as a major link providing the prefrontal
cortex with information from the parietal lobe concerning the
perception of the visual space. Since the SLF II pathway asmentioned above in humans is bidirectional, the fibers originat-ing from the prefrontal cortex and directed back to the
posterior parietal region could provide a means by which the prefrontal cortex can regulate the focusing of attention indifferent parts of space. In addition, damageto SLF II could resultin disorders of spatial working memory by virtue of interruptingrelationships with prefrontal area 46 (Preuss and Goldman-Rakic, 1989; Petrides and Pandya, 2002).
SLF III
In monkeys, SLF III connects the rostral part of the inferior parietal lobule with the lateral inferior frontal lobe in a bi-
directional way (Preuss and Goldman-Rakic, 1989; Petrides andPandya, 2002). The rostral portion of the inferior parietal lobulehas been shown to receive input from the ventral precentralgyrus (Jones and Powell, 1970; Pandya and Seltzer, 1982) and itsneurons exhibit complex somatosensory responses related to
the face and arm in monkeys (Hyvarinen and Shelepin, 1979;Leinonen et al ., 1979a,b; Robinson and Burton, 1980; Taira et al .,1990). Therefore, SLF III probably provides the ventral premotor
region and the adjacent area 44 with higher order somatosens-ory input. In humans, the gestural impairments observed inclinical syndromes of ideomotor apraxia are often the result of lesions of the rostral inferior parietal cortex or of the underlying white matter (De Renzi, 1989). The possible connections bet- ween frontal area 44 (pars opercularis) and the supramarginal
gyrus (area 40) via SLF III suggest that SLF III may have a role inthe articulatory component of language. The bidirectional con-nections between the inferior parietal lobule and the ventral prefrontal area 46 via this fiber pathway may be critical for mon-
itoring orofacial and hand actions since this region has beenshown to play a role in working memory (Preuss and Goldman-Rakic, 1989; Petrides, 1994, 1996; Petrides and Pandya, 2002).
AF
The fibers of the arcuate fascicle connect the caudal part of the superior temporal gyrus (area Tpt) with the dorsal part of
prefrontal area 8 in the monkey. These fibers run contiguously with the fibers of SLF II. Observations using neuroimaging along
with the monkey data indicate that in humans the AF likely hasa similar trajectory. Through this pathway the caudal dorsolat-
eral prefrontal cortex may receive auditory spatial information. There is evidence that neurons in area Tpt respond selectively to the sound source (Leinonen et al ., 1980). Numerous neuro-imaging as well as anatomical and physiological observations in both humans and monkeys are consistent with these findings(Petrides and Pandya, 1988; Hackett et al ., 1999; Kaas and
Hackett, 1999; Romanski et al ., 1999a,b; Kaas and Hackett, 2000;Rauschecker and Tian, 2000; Hackett etal ., 2001; Morosan etal .,2001). Like SLF II, which may provide a means for visuospatial
interaction between the prefrontal cortex and the posterior parietal region, AF by linking the frontal and temporal corticesmay be viewed as providing a means by which the prefrontal
864 Human Superior Longitudinal Fascicle d Makris et al.

8/3/2019 SLF DTI Segmentation 2005
http://slidepdf.com/reader/full/slf-dti-segmentation-2005 12/16
cortex can receive and modulate audiospatial information
(Petrides and Pandya, 2002).
Quantitative Analysis
In the present study we performed measurements of bio- physical parameters for anisotropy such as FA and LA, and of volume and of symmetry for SLF I, SLF II, SLF III and AFv. FA
ranged from 0.36 to 0.50 on the left and from 0.37 to 0.50 on theright, whereas LA ranged from 0.22 to 0.35 on the left and from0.22 to 0.34 on the right. The overall FA and LA for SLF (all foursubcomponents combined) were 0.44 and 0.29 respectively. The FA and LA values for the stems of the four subcomponentsof SLF were similar to the FA and LA reported in other studies of
different fiber tracts in the normal human brain. Pierpaoli andBasser reported FA and LA values of 0.46 and 0.28 respectively insubcortical white matter (Pierpaoli and Basser, 1996a), and
Pierpaoli etal . (1996) showed an LA of 0.31 in the centrum semi-ovale. Klingberg et al . (2000) showed mean unscaled fractionalanisotropy values in temporoparietal white matter that rangedfrom 0.38 to 0.59. In a study by Virta et al . (1999), the LA rangedfrom 0.30 to 0.53 across the different locations of the cortico-
spinal tract. In a previous study of the cingulum bundle we haveshown that FA and LA values of this corticocortical association
pathway ranged from 0.45 to 0.54 and from 0.29 to 0.43 res- pectively (Makris et al ., 2002a). Similarly Rose et al . (2000)reported LA values for the corticospinal tract of 0.39, for the
cingulum bundle of 0.36, and for the superior longitudinal fas-ciculus of 0.46. Thus, it seems that our results of FA and LA for the subcomponents of SLF are in agreement with the data reported for other fiber systems.
The overall volume of SLF (all four subcomponents com- bined) was 32.21 cm3. These values should be contrasted with
measurements derived from purely topographic, constrainedmethods such as ‘white matter parcellation’ (Makris et al .,
1999). Specifically, the white matter of the superior sagittalstratum sector or ‘Ss total’ as measured in 20 healthy adult
subjects using T 1-weighted MRI was estimated to be 28.42 cm3. This corresponds to ~7% of total cerebral white matter. Thesuperior sagittal stratum sector corresponds approximately tothe SLF and these results are comparable in magnitude, althoughthey reflect different underlying methodologies and anatomic
considerations. The coefficient of variation (CV) is large for volumes (27% for total SLF) and more stable for anisotropy interpretations (5% for total fractional anisotropy). This is in part due to the low number of subjectsused in the present study (i.e., N=4), combined with the large volumetric variation in the
general population in specific white matter regions [superiorstrata regions, which include the SLF, CV range 12--70% (Makris
et al ., 1999)]. Due to the small number of subjects in this report,however, care should be taken in interpretation of the pop-ulation variance of these volumetric observations.
Of the four subcomponents of SLF, SLF III showed leftwardasymmetry. Our overall measurements on the linearity of thefour subcomponents of SLF showed a slight total leftwardasymmetry of this fiber system. These results, although repre-sentative of a small group of subjects, are consistent with the
possibility that SLF III may be more closely related to language.
DTI Tractography of SLF
We used the DTI Task Card software (Massachusetts GeneralHospital) to obtain SLF I, SLF II, SLF III and AFv. The algorithm
used by the software was a streamline method that develops
fiber tracts by following the direction of the principle eigen- vector at each step starting from a seed point (Mori et al ., 1999;Lori et al ., 2002). Figure 6 illustrates the results of thistractographic approach in one of the brains analyzed in thisstudy. Of the four subdivisions of SLF we were able to delineateonly SLF II (including AFh) and AFv. In Figure 6A , the color
fractional anisotropy map or PEM is shown in a coronal section
passing through the mid-thalamus. The grayscale fractionalanisotropy map of the identical coronal section is used as background in Figure 6B ,C . In Figure 6B , one ROI for each
hemisphere was selected as seed locations to initiate the fibertracking process. These two ROIs were strategically placed onthe stem of SLF II and AFh based on the fractional anisotropy color map, following the identical criteria that we used tosegment SLF II and AFh. It should be pointed out that anatom-ically driven selection of seed locations and a priori anatomical
knowledge of the tracts are necessary for the successfuldelineation and interpretation of a fiber pathway using tractog-raphy. In Figure 6C , the rostral portion of SLF II in each
hemisphere is shown as resulted from the fiber tracking
process of neighboring voxels that had a primary diffusionorientation close (as determined by an angular threshold) tothe orientation of the seed voxel. The color gives theorientation of the eigenvector as shown in the colored sphere.In Figure 6D , three-dimensional representations of SLF II and AFv are shown. In Figure 6D , SLF II (including AFh) and AFv are
contrasted against a background consisting of a fractionalanisotropy map of a mid-sagittal plane. The fractional anisot-ropy map of a coronal plane was added for reference. The stem
of SLF II and AFh course as a compact structure rostrally, whereas caudally there is a bifurcation, giving rise to the stemof the vertical part of the arcuate fasciculus (AFv) ventrally, anddorsally to the stem of the parietal projection of SLF II. Thecortical regions related to SLF II and AFv were added in Figure
6E . These are represented by the middle frontal gyrus (F2) andthe inferior frontal gyrus, pars opercularis (F3o) in the frontallobe, by the angular gyrus (AG) in the parietal lobe, by the posterior part of the superior temporal gyrus (T1p) andthe superior temporo-occipital transition region (TO2) in the
temporal lobe, and by the dorsolateral occipital region (OLs) inthe occipital lobe. While these findings seem to be generally inagreement with what we presented in this study as a result of extrapolation from the monkey autoradiographic material,there are some considerations that should be raised. First,tractography is a versatile tool for elucidating trajectory of fiber
pathways but less so for their volumetric analysis (Lori et al .,2002). One important component for this is the dependence of
the details of the tracts on the specific parameters such asanisotropy and angular thresholds as well as ROI selection.
Second, the size of a fiber bundle plays an important role in itsidentification. For instance, the streamline algorithms tend tofollow the way of the major bundle. This interrupts fibertractographic identification in regions where large bundlessuch as the corticospinal tract are predominating over smalltracts such as SLF I. This does affect the fiber pathway
volumetric analysis as well; however, discrete portions of thesmaller bundle may be still identifiable and contribute to the volumetry of the tract’s stem. Finally, tractography as well as
segmentation for volumetry of fiber pathways are heavily relying on a priori knowledge of the anatomical and topo-graphical details of the bundles.
Cerebral Cortex June 2005, V 15 N 6 865

8/3/2019 SLF DTI Segmentation 2005
http://slidepdf.com/reader/full/slf-dti-segmentation-2005 13/16
The aim of the present study was the identification and volumetric analysis of the four subdivisions of SLF. Therefore, we concentrated on the segmentation of the orientation
information rather than trying to identify the presence of the pathways from a purely tractographic perspective.
In conclusion, in this study we have identified the location of SLF and its four subdivisions in the human brain. According toour observations there are four major subcomponents of the
SLF. These four components have been discerned using knowl-edge of similar pathways in non-human primates observed withexperimental techniques. In classical behavioral neurology, as well as in current studies in cognitive neuroscience and inhuman neuroanatomy, the SLF fiber system has been considered
a single entity and not as composed of distinct subdivisions. Inaddition, the term arcuate fasciculus has been used interchange-ably with superior longitudinal fasciculus. Historically, SLF orthe AF has been central to the neurobiological interpretation of higher brain function generally and for language and languagedisorders in particular (e.g. Wernicke, 1874; Geschwind,
1965a,b; Lichtheim, 1884; Heilman et al ., 1970; Goodglass,1993). Similarly, in recent years SLF has been investigated in
a number of anatomical (Curnes et al ., 1988; Shen et al ., 1991; Aboitiz and Garcia, 1997; Mori et al ., 1999; Axer et al ., 2000;Catani et al ., 2002; Mamata et al ., 2002; Hagmann et al ., 2003),functional (Chertkow and Murtha, 1997, Duffau et al ., 2002),developmental (Okuda, 1994; Schmithorst et al ., 2002), cogni-tive (Damasio and Damasio, 1980; Caplan, 1987, 1992; Kosslynand Koenig, 1992), and clinical (Naeser and Hayward, 1979;
Tanabe et al ., 1987, 1989; Whittle and Fraser, 1991; Demeurisseand Capon, 1991; Kosslyn et al ., 1993; Shuren et al ., 1995;Pramstaller and Marsden, 1996; Pujol et al ., 1997; Saito et al .,1997; Nass et al ., 1998; Anderson et al ., 1999; Pujol et al ., 2000;
Ikeda et al ., 2001; Rose et al ., 2000; Mori, 2002; Catani et al .,2003) studies using available neuroimaging techniques. In termsof its volume, SLF may be the major cortical association fiber
pathway in the human brain. It interconnects frontal, temporaland parietal association areas, which emphasizes its centrality tomany associative or higher brain functions. Thus, clarification of
its anatomy is necessary in order to correlate structural andfunctional parameters more precisely in cognitive investiga-tions. The segregation of this fiber system into four componentsin the human in vivo with different topography and potentially different functional affiliations may provide a finer level of
analysis for human clinical studies. Such novel understanding of the anatomy of the SLF fiber system in humans will increasespecificity in investigations of structural--functional andanatomical--clinical relationships and will extend the functionalcharacterization of SLF beyond the language modality, namely to
the domain of other higher cortical functions such as memory,attention, and executive as well as visuospatial and audiospatial processing.
Notes
The authors gratefully acknowledge Dr Edward H. Yeterian, Mr GeorgeM. Papadimitriou, Mr Joseph J. Normandin and Mr James D. Howard fortheir valuable contributions to the preparation of this manuscript.Preparation of this article was supported in part by grants from: theNational Association for Research in Schizophrenia and Depression(NARSAD), and the Amyotrophic Lateral Sclerosis Association (ALSA) toN.M.; the Fairway Trust to D.K.; the National Center for ResearchResources (P41RR14075), and the National Institutes of Health (R01-NS38477) to A.G.S., and the Mental Illness and Neuroscience Discovery (MIND) Institute.
Address correspondence to Nikos Makris, Massachusetts GeneralHospital, Center for Morphometric Analysis, Building 149, 13th Street,Charlestown, MA 02129, USA.Email: [email protected].
Human Research Statement
The experiments undertaken in this paper were performed with the understanding and written informed consent of eachsubject.
Figure 6. Illustration of tractographic results for SLF II (including AFh) and AFv. ( A) The color fractional anisotropy (fractional anisotropy) map or principal eigenvector map (PEM) ina coronal section. The grayscale FA map of the identical coronal section is used as background in ( B) and (C). In ( B), one region of interest (ROI) (shown in green), which is the SLF II(including the AFh) for each hemisphere was selected as seed location to initiate the fiber tracking process. In (C), the rostral portion of SLF II in each hemisphere is shown asresulted from the fiber-tracking process. The color gives the orientation of the eigenvector as shown in the colored sphere. Red is right--left, green is anterior--posterior and blue is
superior--inferior orientations. ( D
) Three-dimensional representations of SLF II and AFv. SLF II (including AFh) and AFv are contrasted against a background consisting of a fractionalanisotropy map of a mid-sagittal plane. The fractional anisotropy map of a coronal plane was added for reference. The cortical regions related to SLF II and AFv were added in ( E ).Abbreviations, AFh, horizontal part of arcuate fascicle; AFv, vertical part of arcuate fascicle; AG, angular gyrus; F2, middle frontal gyrus; F3o, inferior frontal gyrus, pars opercularis;OLs, lateral occipital cortex, superior; SLF, superior longitudinal fascicle; T1p, superior temporal gyrus, posterior; TO2, middle temporal gyrus, temporooccipital.
866 Human Superior Longitudinal Fascicle d Makris et al.

8/3/2019 SLF DTI Segmentation 2005
http://slidepdf.com/reader/full/slf-dti-segmentation-2005 14/16
References
Aboitiz F, Garcia R (1997) The anatomy of language revisited. Biol Res30:171--183.
Andersen RA, Gnadt JW (1989) Posterior parietal cortex. Rev OculomotRes 3:315--335.
Anderson JM, Gilmore R, Roper S, Crosson B, Bauer RM, Nadeau S,Beversdorf DQ, Cibula J, Rogish M, 3rd, Kortencamp S, Hughes JD,Gonzalez Rothi LJ, Heilman KM (1999) Conduction aphasia and thearcuate fasciculus: a reexamination of the Wernicke--Geschwindmodel. Brain Lang 70:1--12.
Axer H, Berks G, Keyserlingk DG (2000) Visualization of nerve fiberorientation in gross histologicalsections of thehuman brain. MicroscRes Tech 51:481--492.
Basser PJ, Pierpaoli C (1996) Microstructural and physiological featuresof tissues elucidated by quantitative-diffusion-tensor MRI. J MagnReson B 111:209--219.
Basser PJ, Mattiello J, LeBihan D (1994) Estimation of the effective self-diffusion tensor from the NMR spin echo. J Magn Reson B 103:247--254.
Bisley JW, Goldberg ME (2003) Neuronal activity in the lateral intra- parietal area and spatial attention. Science 299:81--86.
Brodmann K (1905) Beitrage zur histologischen Lokalisation derGrosshirnrinde. III. Mitteilung. Die Rindenfelder der niederen Affen.
J Psychol Neurol 4:177--226.
Brodmann K (1909) Vergleichende Lokalisationslehre der Grosshirn-rinde in ihren Prinzipien dargestellt auf Grund des Zellenbaues.Leipzig: Verlag von Johann Ambrosius Barth.
Burdach CF (1822) Baue und Leben des Gehirns. Leipzig: in deerDyk’schen Buchhandlung.
Caplan D (1987) Discrimination of normal and aphasic subjects on a testof syntactic comprehension. Neuropsychologia 25:173--184.
Caplan D (1992) Language: structure, processing and disorders. Cam- bridge, MA: MIT Press.
Catani M, Howard RJ, Pajevic S, Jones DK (2002) Virtual in vivo
interactive dissection of white matter fasciculi in the human brain.Neuroimage 17:77--94.
Catani M, Piccirilli M, Cherubini A, Tarducci R, Sciarma T, Gobbi G,Pelliccioli G, Petrillo SM, Senin U, Mecocci P (2003) Axonal injury
within language network in primary progressive aphasia. Ann Neurol
53:242--247.Caviness VS, Jr, Makris N, Meyer JW, Kennedy DN (1996) MRI-based
parcellation of human neocortex: an anatomically specified method with estimate of reliability. J Cogn Neurosci 8:566--588.
Chertkow H, Murtha S (1997) PET activation and language. ClinNeurosci 4:78--86.
Curnes JT, Burger PC, Djang WT, Boyko OB (1988) MR imaging of com- pact white matter pathways. AJNR Am J Neuroradiol 9:1061--1068.
Damasio H, Damasio AR (1980) The anatomical basis of conductionaphasia. Brain 103:337--350.
De Renzi E (1989) [Agnosia]. Recenti Prog Med 80:633--637.Dejerine J (1895) Anatomie des Centres Nerveux. Paris: Rueff et Cie
(repr. 1980, Masson).Demeurisse G, Capon A (1991) Brain activation during a linguistic task
in conduction aphasia. Cortex 27:285--294.Duffau H, Capelle L, Sichez N, Denvil D, Lopes M, Sichez JP, Bitar A,
Fohanno D (2002) Intraoperative mapping of the subcorticallanguage pathways using direct stimulations. An anatomo-functionalstudy. Brain 125:199--214.
Duffy FH, Burchfiel JL (1971) Somatosensory system: organizationalhierarchy from single units in monkey area 5. Science 172:273--275.
Filipek PA, Richelme C, Kennedy DN, Caviness VS, Jr (1994) The youngadult human brain: an MRI-based morphometric analysis. Cereb Cort4:344--360.
Galaburda AM, Corsiglia J, Rosen GD, Sherman GF (1987) Planumtemporale asymmetry: Reappraisal since Geschwind and Levitsky.Neuropsychologia 25:853--868.
Geschwind N (1965a) Disconnexion syndromes in animals and man. I.Brain 88:237--294.
Geschwind N (1965b) Disconnexion syndromes in animals and man. II.Brain 88:585--644.
Goldberg ME, Segraves MA (1989) The visual and frontal cortices. Rev Oculomot Res 3:283--313.
Goodglass H (1993) Understanding aphasia. San Diego: Academic Press.Hackett TA,Stepniewska I, Kaas JH (1999) Prefrontal connections of the
parabelt auditory cortex in macaque monkeys. Brain Res 817:45--58.Hackett TA, Preuss TM, Kaas JH (2001) Architectonic identification of
the core region in auditory cortex of macaques, chimpanzees, andhumans. J Comp Neurol 441:197--222.
Hagmann P, Thiran JP, Jonasson L, Vandergheynst P, Clarke S, Maeder P,
Meuli R (2003) DTI mapping of human brain connectivity: statisticalfibre tracking and virtual dissection. Neuroimage 19:545--554.
Halsband U, Passingham R (1982) The role of premotor and parietalcortex in the direction of action. Brain Res 240:368--372.
Heilman KM, Pandya DN, Geschwind N (1970) Trimodal inattentionfollowing parietal lobe ablations. Trans Am Neurol Assoc 95:259--261.
Heimer L (1983) The human brain and spinal cord: functionalneuroanatomy and dissection guide. New York: Springer Verlag.
Hyvarinen J, Shelepin Y (1979) Distribution of visual and somaticfunctions in the parietal associative area 7 of the monkey. BrainRes 169:561--564.
Ikeda A, Matsumoto R, Ohara S, Kunieda T, Shirakashi Y, Kaji R,Fukuyama H, Shibasaki H (2001) Asymmetric tonic seizures with
bilateral parietal lesions resembling frontal lobe epilepsy. EpilepticDisord 3:17--22.
Jones EG, Powell TPS (1970) An anatomical study of converging sensory pathways within the cerebral cortex of the monkey. Brain93:793--820.
Kaas JH, Hackett TA (1999) ‘What’ and ‘where’ processing in auditory cortex. Nat Neurosci 2:1045--1047.
Kaas JH, Hackett TA (2000) Subdivisions of auditory cortex and processing streams in primates. Proc Natl Acad Sci USA 97:11793--11799.
Klingberg T, Hedehus M, Temple E, Salz T, Gabrieli JD, Moseley ME,Poldrack RA (2000) Microstructure of temporo-parietal whitematter as a basis for reading ability: evidence from diffusion tensormagnetic resonance imaging. Neuron 25:493--500.
Kosslyn SM, Koenig O (1992) Wet mind: the new cognitive neurosci-ence. New York: The Free Press.
Kosslyn SM, Daly PF, McPeek RM, Alpert NM, Kennedy DN, Caviness VS, Jr (1993) Using locations to store shape:an indirect effect of a lesion.
Cereb Cortex 3:567--582.Lacquaniti F, Guigon E, Bianchi L, Ferraina S, Caminiti R (1995)
Representing spatial information for limb movement: role of area 5in the monkey. Cereb Cortex 5:391--409.
Leinonen L, Nyman G (1979b) II. Functional properties of cells inanterolateral part of area 7 associative face area of awake monkeys.Exp Brain Res 34:321--333.
Leinonen L, Hyvarinen J, Sovijarvi AR (1980) Functional properties of neurons in the temporo-parietal association cortex of awakemonkey. Exp Brain Res 39:203--215.
Leinonen L, Hyvarinen J, Nyman G, Linnankoski I (1979a) I. Functional properties of neurons in lateral part of associative area 7 in awakemonkeys. Exp Brain Res 34:299--320.
Lichtheim O (1884) On aphasia. Brain 7:443--484.Lori NF, Akbudak E, Shimony JS, Cull TS, Snyder AZ, Guillory RK,
Conturo TE (2002) Diffusion tensor fiber tracking of human brainconnectivity: aquisition methods, reliability analysis and biologicalresults. NMR Biomed 15:494--515.
Ludwig E, Klingler J (1956) Atlas cerebri humani. Boston, MA: Little,Brown and Company.
Makris N, Pandya DN, Normandin JJ (2002a) Quantitative DT-MRIinvestigations of the human cingulum bundle. Cent Nerv Sys Spectr7:522--528.
Makris N, Meyer JW, Bates JF, Yeterian EH, Kennedy DN, Caviness VS(1999) MRI-Based topographic parcellation of human cerebral whitematter and nuclei II. Rationale and applications with systematics of cerebral connectivity. Neuroimage 9:18--45.
Makris N, Papadimitriou GM, Worth AJ, Jenkins BG, Garrido L, Sorensen AG, Wedeen V, Tuch DS, Wu O, Cudkowicz ME, Caviness VS, Jr,Rosen B, Kennedy DN (2002b) Diffusion tensor imaging. In: Neuro-
psychopharmacology: the fifth generation of progress (Davis KL,
Cerebral Cortex June 2005, V 15 N 6 867

8/3/2019 SLF DTI Segmentation 2005
http://slidepdf.com/reader/full/slf-dti-segmentation-2005 15/16
Charney D, Coyle J, Nemeroff C, eds), pp. 357--371. New York:Lippincott, Williams, and Wilkins.
Makris N, Worth AJ, Sorensen AG, Papadimitriou GM, Wu O, Reese TG,
Wedeen VJ, Davis TL, Stakes JW, Caviness VS, Kaplan E, Rosen BR,Pandya DN, Kennedy DN (1997) Morphometry of in vivo human
white matter association pathways with diffusion-weighted mag-netic resonance imaging. Ann Neurol 42:951--962.
Mamata H, Mamata Y, Westin CF, Shenton ME, Kikinis R, Jolesz FA,
Maier SE (2002) High-resolution line scan diffusion tensor MR
imaging of white matter fiber tract anatomy. AJNR Am J Neuroradiol23:67--75.
Mesulam MM (1981) A cortical network for directed attention and
unilateral neglect. Ann Neurol 10:309--325.Meynert T (1865) Anatomie der Hirnrinde und ihre Verbindungsbahnen
mit den empfindenden Oberflachen und den bewegenden Massen.In: Leidesdorf’s Lehrbuch der phychischen Krankheiten. Erlangen.
Mori S (2002) Twoand three-dimensional analyses of brain white matter
architecture using diffusion imaging. Cent Nerv Sys Spectr 7:529--534.
Mori S, Crain BJ, Chacko VP, van Zijl PC (1999) Three-dimensionaltracking of axonal projections in the brain by magnetic resonance
imaging. Ann Neurol 45:265--269.Morosan P, Rademacher J, Schleicher A, Amunts K, Schormann T, Zilles
K (2001) Human primary auditory cortex: cytoarchitectonic sub-
divisions and mapping into a spatial reference system. Neuroimage13:684--701.
Mountcastle VB, Lynch JC, Georgopoulos A, Sakata H, Acuna C (1975)
Posterior parietal association cortex of the monkey: commandfunctions for operations within extrapersonal space. J Neurophysiol38:871--908.
Naeser MA, Hayward RW (1979) The resolving stroke and aphasia. A
case study with computerized tomography. Arch Neurol 36:233--235.
Nass R, Leventhal F, Levine B, Lebron D, Maxfield C, McCaul P, George A,
Allen J (1998) Conduction aphasia in a 3-year-old with a left posterior cortical/subcortical abscess. Brain Lang 62:70--88.
Okuda Y (1994) [Brain development during the first year of life:quantitative assessment with ADC imaging]. Nippon Igaku Hoshasen
Gakkai Zasshi 54:1245--1251.
Pajevic S, Pierpaoli C (1999) Color schemes to represent the orientationof anisotropic tissues from diffusion tensor data: application to white
matter fiber tract mapping in the human brain. Magn Reson Med42:526--540.
Pandya DN, Seltzer B (1982) Intrinsic connections and architectonics of
posterior parietal cortex in the rhesus monkey. J Comp Neurol204:196--210.
Petrides M (1982) Motor conditional associative-learning after selective prefrontal lesions in the monkey. Behav Brain Res 5:407--413.
Petrides M (1987) Conditional learning and the primate frontal
cortex. In: The frontal lobes revisited (Perecman E, ed.), pp. 91--08.New York: Lea.
Petrides M (1994) Frontal lobes and behaviour. Curr Opin Neurobiol
4:207--211.Petrides M (1996) Specialized systems for the processing of mnemonic
information within the primate frontal cortex. Philos Trans R Soc
Lond B Biol Sci 351:1455--1461; discussion 1461--1452.Petrides M, Pandya DN (1984) Projections to the frontal cortex from the
posterior parietal region in the rhesus monkey. J Comp Neurol228:105--116.
Petrides M, Pandya DN (1988) Association fiber pathways to the frontal
cortex from the superior temporal region in the rhesus monkey. J Comp Neurol 273:52--66.
Petrides M, Pandya DN (2002) Association pathways of the prefrontalcortex and functional observations. In: Principles of frontal lobe
function (Struss DT, Knight RT, eds), pp. 31--50. Oxford: OxfordUniversity Press.
Pierpaoli C, Basser PJ (1996a) Toward a quantitative assessment of
diffusion anisotropy. Magn Reson Med 36:893--906.Pierpaoli C, Basser PJ (1996b) Toward a quantitative assessment of
diffusion anisotropy (erratum). Magn Reson Med 37:972.
Pierpaoli CJ, Basser PJ, Barnett A, Di Chiro G (1996) Diffusion tensor MR imaging of the human brain. Radiology 201:637--648.
Posner MI, Walker JA, Friedrich FJ, Rafal RD (1984) Effects of parietalinjury on covert orienting of attention. J Neurosci 4:1863--1874.
Poupon C, Clark CA, Frouin V, Regis J, Bloch I, Le Bihan D, Mangin J
(2000) Regularization of diffusion-based direction maps for thetracking of brain white matter fascicles. Neuroimage 12:184--195.
Pramstaller PP, Marsden CD (1996) The basal ganglia and apraxia. Brain
119 (Pt 1):319--340.
Preuss TM, Goldman-Rakic PS (1989) Connections of the ventralgranular frontal cortex of macaques with perisylvian premotor andsomatosensory areas: anatomical evidence for somatic representa-tion in primate frontal association cortex. J Comp Neurol 282:
293--316.Pujol J, Bello J, Deus J, Marti-Vilalta JL, Capdevila A (1997) Lesions in the
left arcuate fasciculus region and depressive symptoms in multiple
sclerosis. Neurology 49:1105--1110.Pujol J, Bello J, Deus J, Cardoner N, Marti-Vilalta JL, Capdevila A (2000)
Beck Depression Inventory factors related to demyelinating
lesions of the left arcuate fasciculus region. Psychiatry Res 99:151--159.
Rademacher J, Galaburda AM, Kennedy DN, Filipek PA, Caviness VS, Jr(1992) Human cerebral cortex: localization, parcellation andmorphometry with magnetic resonance imaging. J Cogn Neurosci 4:
352--374.Rauschecker JP, Tian B (2000) Mechanisms and streams for processing
of ‘what’ and ‘where’ in auditory cortex. Proc Natl Acad Sci USA
97:11800--11806.Reil DJC, Autenrieth DJHF (1809) Archiv fur die Physiologie. Halle: In
Der Curtschen Buchhandlung.Robinson CJ, Burton H (1980) Organization of somatosensory receptive
fields in cortical areas 7b, retroinsula, postauditory and granular
insula of M. fascicularis . J Comp Neurol 192:69--92.Romanski LM, Bates JF, Goldman-Rakic PS (1999a) Auditory belt and
parabelt projections to the prefrontal cortex in the rhesus monkey. J Comp Neurol 403:141--157.
Romanski LM, Tian B, Fritz J, Mishkin M, Goldman-Rakic PS, Rau-
schecker JP (1999b) Dual streams of auditory afferents targetmultiple domains in the primate prefrontal cortex. Nat Neurosci
2:1131--1136.Rose SE, Chen F, Chalk JB, Zelaya FO, Strugnell WE, Benson M, Semple J,
Doddrell DM (2000) Loss of connectivity in Alzheimer’s disease: anevaluation of white matter tract integrity with colour codedMR diffusion tensor imaging. J Neurol Neurosurg Psychiatry 69:
528--530.Saito Y, Kita Y, Bando M, Nagura H, Yamanouchi H, Ishii K (1997)
[Neuropsychological analysis in 2 cases of infarction in the left
precentral gyrus — with special reference to apraxia of speech andagraphia]. Rinsho Shinkeigaku 37:487--491.
Sakata H, Takaoka Y, Kawarasaki A, Shibutani H (1973) Somatosensory
properties of neurons in the superior parietal cortex (area 5) of therhesus monkey. Brain Res 64:85--102.
Schmithorst VJ, Wilke M, Dardzinski BJ, Holland SK (2002) Correlationof white matter diffusivity and anisotropy with age during childhoodand adolescence: a cross-sectional diffusion-tensor MR imaging
study. Radiology 222:212--218.Schnopfhagen F (1890) Die Entstehung der Windungen des Grosshirns.
Jahrbucher fur Psychiatrie 9:197--318.Shen WC, Yang CF, Chang CY, Ho WL (1991) [MRI of white matter
pathways]. Zhonghua Yi Xue Za Zhi (Taipei) 47:91--100.Shuren JE, Schefft BK, Yeh HS, Privitera MD, Cahill WT, Houston W
(1995) Repetition and the arcuate fasciculus. J Neurol 242: x596--598.
Sorensen AG, Wu O, Copen WA, Davis TL, Gonzalez RG, Koroshetz WJ,Reese TG, Rosen BR, Wedeen VJ, Weisskoff RM (1999) Human acute
cerebral ischemia: detection of changes in water diffusion aniso-tropy by using MR imaging. Radiology 212:785--792.
Taira M, Mine S, Georgopoulos AP, Murata A, Sakata H (1990) Parietal
cortex neurons of the monkey related to the visual guidance of handmovement. Exp Brain Res 83:29--36.
868 Human Superior Longitudinal Fascicle d Makris et al.

8/3/2019 SLF DTI Segmentation 2005
http://slidepdf.com/reader/full/slf-dti-segmentation-2005 16/16
Talairach J, Tournoux P (1988) Co-planar stereotaxic atlas of the human brain. New York: Thieme.
Tanabe H, Sawada T, Inoue N, Ogawa M, Kuriyama Y, Shiraishi J (1987)Conduction aphasia and arcuate fasciculus. Acta Neurol Scand76:422--427.
Tanabe H, Ikeda M, Murasawa A, Yamada K, Yamamoto H, Nakagawa Y,Nishimura T, Shiraishi J (1989) A case of acquired conductionaphasia in a child. Acta Neurol Scand 80:314--318.
VirtaA, Barnett A, PierpaoliC (1999) Visualizingand characterizingwhitematter fiber structure and architecture in the human pyramidal tractusing diffusion tensor MRI. Magn Reson Imaging 17:1121--1133.
Wernicke C (1874) Der aphasiche Symptomenkomplex. Breslau: Cohnund Weigert.
Whittle IR, Fraser DE (1991) Resolution of fluent dysphasia followingexcision of metastatic carcinoma from the arcuate fasciculus. Br JNeurosurg 5:647--649.