switch in 39 splice site recognition between exon
TRANSCRIPT

MOLECULAR AND CELLULAR BIOLOGY,0270-7306/01/$04.0010 DOI: 10.1128/MCB.21.6.1986–1996.2001
Mar. 2001, p. 1986–1996 Vol. 21, No. 6
Copyright © 2001, American Society for Microbiology. All Rights Reserved.
Switch in 39 Splice Site Recognition between Exon Definition andSplicing Catalysis Is Important for Sex-lethal Autoregulation
LUIZ O. F. PENALVA,† MARIA JOSE LALLENA, AND JUAN VALCARCEL*
Gene Expression Programme, European Molecular Biology Laboratory, D-69117 Heidelberg, Germany
Received 7 September 2000/Returned for modification 31 October 2000/Accepted 14 December 2000
Maintenance of female sexual identity in Drosophila melanogaster involves an autoregulatory loop in whichthe protein Sex-lethal (SXL) promotes skipping of exon 3 from its own pre-mRNA. We have used transienttransfection of Drosophila Schneider cells to analyze the role of exon 3 splice sites in regulation. Our resultsindicate that exon 3 repression requires competition between the 5* splice sites of exons 2 and 3 but isindependent of their relative strength. Two 3* splice site AG’s precede exon 3. We report here that, while thedistal site plays a critical role in defining the exon, the proximal site is preferentially used for the actualsplicing reaction, arguing for a switch in 3* splice site recognition between exon definition and splicingcatalysis. Remarkably, the presence of the two 3* splice sites is important for the efficient regulation by SXL,suggesting that SXL interferes with molecular events occurring between initial splice site communicationacross the exon and the splice site pairing that leads to intron removal.
Alternative splicing is a versatile mechanism of gene expres-sion regulation that can turn on or off genes or generate dif-ferent protein isoforms with distinct biological properties (forrecent reviews, see references 12 and 32). Despite the preva-lence of alternative splicing in higher eukaryotes, relativelylittle is known about the underlying molecular mechanisms ofregulation. Genes that participate in the Drosophila melano-gaster sex determination cascade have become good modelsystems for understanding splicing control because geneticdata have defined both regulatory factors and their target pre-mRNAs (reviewed in reference 46).
Sex-lethal (SXL), an RNA- binding protein with preferencefor U-rich sequences, promotes female-specific patterns ofsplicing on at least three transcripts: (i) its own pre-mRNA,where SXL promotes exon 3 skipping (6); (ii) transformer pre-mRNA, where SXL promotes a switch between alternative 39splice sites (9); and (iii) male-specific-lethal 2, where SXL pro-motes retention of an intron at the 59 untranslated region (4,28, 62). The biological consequences of these alternative splic-ing events are dramatic and control multiple aspects of sexualdetermination in the fruitfly. Sxl and tra transcripts spliced inthe female-specific mode encode SXL and TRA proteins,while transcripts that follow the alternative splicing pathwaycan only encode truncated polypeptides. Expression of full-length TRA controls somatic sexual differentiation and sexualbehavior, while expression of SXL maintains the female dif-ferentiation state throughout the life of the fly. Retention ofthe msl-2 intron allows SXL to act as a translational repressorand to inhibit MSL-2 protein expression, thereby turning offdosage compensation in female flies (5, 20, 29).
The mechanism by which SXL controls tra splicing has beeninvestigated in vivo and in vitro. Two 39 splice sites are present
in intron 1. The proximal site, used in both males and females(hence the name non-sex-specific) contains a high-affinity bind-ing site for SXL at the polypyrimidine (Py) tract. The distal siteis used in a female-specific fashion. Evidence from experimentsin transgenic flies and transient transfections of cells in culture,as well as in vitro biochemical analysis, indicate that SXLrepresses the use of the non-sex-specific site (27, 51, 54). Invivo and in vitro results are consistent with a model in whichSXL prevents the binding of the splicing factor U2AF to the Pytract of the non-sex-specific site, thereby diverting U2AF andsplicing to the female-specific site (21, 54). Blockage of U2AFbinding is also important for regulation of msl-2 splicing invitro (34).
Several lines of evidence suggest a different mechanism forSxl autoregulation. First, although the Py tract associated withone of the 39 splice sites preceding exon 3 in D. melanogastercontains a relatively long stretch of uridines and is a potentialbinding site for SXL (25), its mutation does not abolish regu-lation (26, 41), in contrast to tra (27, 51). Second, multipleU-rich sequences, relatively distant from the 59 and 39 splicesites, contribute to exon 3 skipping (26, 41), and cooperativebinding of SXL to these sequences, mediated through an ami-no-terminal glycine and an asparagine-rich domain, is impor-tant for regulation (55). Third, ectopic expression in maletransgenic flies of a chimeric protein in which the splicingactivation domain of U2AF was fused to SXL RNA-bindingdomains results in disruption of tra regulation but not of Sxlregulation (21). Because this chimeric protein promotes thesplicing of pre-mRNAs containing SXL binding sites at the Pytract (54), as U2AF does, these data argue that antagonizingU2AF activity is insufficient to explain SXL-mediated exonskipping.
Results using transgenic flies suggest that a key regulatorystep in Sxl autoregulation is the inhibition of exon 3 59 splicesite (26). Inhibition of 59 splice site recognition can result inexon skipping because of defects in exon definition. Exon def-inition is the process by which early recognition of splice sitesin relatively short exons embedded in long introns occurs
* Corresponding author. Mailing address: Gene Expression Pro-gramme, European Molecular Biology Laboratory, Meyerhofstrasse 1,D-69117 Heidelberg, Germany. Phone: 49-6221-387-156. Fax: 49-6221-387-442. E-mail: [email protected].
† Present address: Duke University Medical Center, Durham, NC27710.
1986
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 10
Feb
ruar
y 20
22 b
y 22
3.16
.72.
134.

through molecular interactions involving factors bound to the39 splice site preceding the exon and the downstream 59 splicesite (reviewed in reference 7). It is believed that the factorsinvolved in this process are the same as those involved in earlycommunication between splice sites across introns: U1 snRNPbound to the 59 splice site, U2AF65 bound to the Py tract, andbridging activities, for which members of the SR family offactors are strong candidates. How the communication be-tween splice sites across exons is switched to communicationacross introns for selection of the actual splicing partners iscurrently unknown.
Here we report a role for the 39 splice site AG in exondefinition that can be physically separated from its role insplicing catalysis, arguing for a switch in splice site recognitionin the transition between these two processes. Since the pres-ence of two 39 splice sites appears to be important for regula-tion of exon skipping by SXL, this observation opens the pos-sibility that the presence of SXL interferes with molecularevents leading to such a switch.
MATERIALS AND METHODS
Plasmids and mutagenesis. Plasmids copia-SxlTE234, pBShsp-cat, and pB-Shsp-SxlF1cDNA were described previously (41).
Deletion or substitution mutants in the Sxl minigene including exons 2 to 4were obtained from plasmid copia-SxlTE234 using the procedure described byL. O. F. Penalva and J. Valcarcel (Techical tips online [http://tto.trends.com]).All mutations were confirmed by sequencing. The sequence changes introducedwere as follows: 59 splice site mutants, as indicated in Fig. 2B; 59ss ex2Mut, thesequence of exon 2 59 splice site (in DNA form AG/GTAAA, the slash indicatingthe exon-intron boundary) was changed into CAATGAA; 39 splice site mutants,as indicated in Fig. 3B and 5A; DInt2, deletion of intron 2 sequences betweenpositions 12 and 2919 (the sequence between exon 2 59ss and the distal 39 splicesite of exon 3 is indicated in Fig. 3D); and DInt2MutAG, the sequence AGCCCAGAAAGAAGCAG, corresponding to the distal 39 splice site AG and the59 end of exon 3 was replaced by (CA)8.
Transfection and RNA isolation. Transfections were performed using Lipo-fectin (Gibco) according to the manufacturer’s recommendations. Typically, 3 mgof each plasmid (pBSHS-TE234 or mutant derivatives and either pBShsp-Sxl-CF1 or pBShsp-cat) were used to transfect Schneider cell cultures at 80% densityin 60-cm2 plates. At 30 h after transfection, cells were harvested, washed oncewith cold 1 3 phosphate-buffered saline, and then lysed in 0.5% NP40, 10 mMTris-HCl (pH 8.5), 1.5 mM MgCl2, and 150 mM NaCl. Nuclei were spun downfor 5 min at 800 3 g, and the supernatant (cytoplasmic fraction) was transferredto a new tube, digested for 20 min at 65°C with proteinase K (0.2 mg/ml) in 1%Sodium dodecyl sulfate – 100 mM Tris (pH 7.5)–12.5 mM EDTA–150 mM NaCl,extracted twice with phenol-chloroform and once with chloroform, and precip-itated with isopropanol; the pellet was then washed with 75% ethanol andresuspended in H2O. The purified RNA was quantified by determining theabsorbance at 265 nm.
Preparation of total RNA from Drosophila adult flies. Total RNA from adultflies was prepared as described previously (11).
RT-PCR. A total of 15 mg of total RNA purified from transfected cells wastreated twice with 10 U of RNase-free DNase (Roche) for 1 h at 37°C. RNA waspurified after each DNase digestion by extraction with phenol-chloroform andethanol precipitation. Reverse transcription (RT) was carried out using ExpandReverse Transcriptase (Roche) according to the manufacturer’s instructions.PCR was performed as described by Sakamoto et al. (41): 2 min at 94°C, followedby 25 cycles of 1 min at 94°C, 1 min at 60°C, and 1 min at 72°C, and then a final5 min at 72°C, using primers PT1 and PT2, designed by Sakamoto et al. (41) toprovide sequence tags for the transfected minigenes. PCR products were ana-lyzed in 1.8% agarose gels. For the analysis of intron retention in clone 59ssex2Mut (Fig. 2D), a primer corresponding to exon 3 sequences (CACTGACTCTTAAGATAGTATGTAG) replaced PT1 in the PCR. For the analyses shownin Fig. 5, primer PT1 was replaced by primer ex2-14 (TCAAGTCAACTGCAACTCACC).
The RT-PCRs were quantitative under these conditions (see Fig. 1 and datanot shown). All of the data presented were obtained using these conditions, and
results that required further amplification (for example, because of low trans-fection efficiency or low RNA yields) were disregarded.
Analysis of exon 3 3* splice site usage. A total of 15 mg of total RNA fromadult males (see Fig. 4) or from transfected cells (see Fig. 6) was analyzed byRT-PCR using the following primers: GGTTGCTTTGCGTTACAAAAAC (an-tisense, exon 3) and CCCCCATATGGCTACAACAA (sense, exon 2). PCRreactions were amplified for 25 cycles of 1 min at 94°C, 1 min at 55°C, and 30 sat 72°C, followed by a final extension of 10 min at 72°C. The products ofamplification were analyzed by electrophoresis on 6% polyacrylamide non de-naturing gels.
RESULTS
To address the role of competing splice sites in Sxl autoreg-ulation, Drosophila Schneider cells were transiently transfectedwith a minigene containing the Sxl genomic region betweenexons 2 and 4 (Fig. 1A) under the copia transposable elementpromoter or with mutants of this construct. RNA was isolatedfrom the transfected cultures and analyzed by RT-PCR asdescribed previously (41) using oligodeoxynucleotides corre-sponding to transcribed vector sequences (PT1 and PT2 in Fig.1A), thus allowing specific detection of transcripts derivedfrom the Sxl minigene and not endogenous Sxl transcripts.
To verify the sensitivity of our detection procedure tochanges in the relative accumulation of alternatively splicedtranscripts, we performed the experiment shown in Fig. 1B.Cotransfection of the plasmid containing the Sxl minigene (co-pia-SxlTE234) with the vector pBShsp-cat resulted in accumu-lation of transcripts that included exon 3 (lane 1), as is the casein male flies. Cotransfection of copia-SxlTE234 with an expres-sion plasmid containing SXL cDNA under a heat shock pro-moter (pBShsp-SxlF1cDNA) resulted in the accumulation oftranscripts that skipped exon 3 (lane 2), as in female flies.When these two types of RNAs were mixed in equal amounts,the products of RT and PCR amplification accumulated insimilar proportion (lane 3). When the RNAs were mixed at 1:3or 3:1 ratios, the relative abundance of the amplification prod-ucts changed proportionally (lanes 4 and 5). Figure 1C showsa comparison between the ratios of input RNAs and the ratiosbetween the signals of the amplified DNA products quantifiedby SYBR Green (Molecular Probes) staining and phosphorim-ager analysis. The data indicate that our assay system allowsthe accurate detection of threefold (and probably smaller)changes in the relative accumulation of alternatively splicedproducts. They also indicate that there are no significant dif-ferences in the relative amplification efficiencies of the twoproducts.
Regulation of 5* splice site usage. Competing sites of differ-ent strengths often establish default patterns of alternativesplicing that are then subject to modulation by regulatory fac-tors (for a review, see reference 12). It has been proposed thatblockage of the exon 3 59 splice site is the key regulatory stepin SXL-induced skipping of exon 3 (26). Both exon 2 and exon3 59 splice sites are conserved among three Drosophila species(melanogaster, suboscura, and virilis) which have diverged for;60 million years (10, 38). The 59 splice site of exon 3 ispredictably stronger in the three species, particularly becauseof the presence of a guanidine at intronic position 5 which isconserved in 85% of higher eukaryotic 59 splice sites (22) butabsent from the 59 splice site of exon 2. This configuration ofsplice site strengths could reflect that proper default and/orregulated Sxl splicing requires a strong 59 splice site in exon 3,
VOL. 21, 2001 SWITCH IN 39 SPLICE SITE RECOGNITION 1987
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 10
Feb
ruar
y 20
22 b
y 22
3.16
.72.
134.

a weak 59 splice site in exon 2, or both. To test these hypoth-eses, a series of mutant minigenes were prepared (Fig. 2B) inwhich (i) both exons contained identical 59 splice sites, eitherthat of exon 2 [mutant 59ss(2-2)] or that of exon 3 [mutant
59ss(3-3)]; (ii) the relative position of the two 59 splice sites wasswapped [mutant 59ss(3-2)]; and (iii) a strong consensus 59splice site was placed in exon 2, in competition with either thenatural 59 splice site of exon 3 [mutant 59ss(CS-3)] or with theexon 2 59 splice site placed in exon 3 [mutant 59ss(CS-2)].Figure 2C shows that the accumulation of alternatively splicedproducts was very similar in all cases, both in the absence or inthe presence of SXL. These data argue against a role for theprecise identity and/or strength of the competing 59 splice sitesin either establishing the default splicing pathway or in achiev-ing regulation by SXL.
We noticed that, in this particular series of experiments, anadditional product of amplification accumulated in the absenceof SXL. This product was purified and cloned. Sequence anal-ysis revealed that it corresponds to the use of a cryptic 39 splicesite 24 nucleotides upstream from the natural 39 splice siteassociated with exon 4. This cryptic site is spliced to exon 3 59splice site in the absence of SXL and apparently is not splicedto exon 2 in the presence of SXL.
To test whether inhibition of exon 3 59 splice site could occurin the absence of a competing 59 splice site, exon 2 59 splice sitewas inactivated by mutation (59ss ex2Mut). The results shownin Fig. 2D indicate that in the absence of a functional 59 splicesite in exon 2, the blockage of the exon 3 59 splice site wassignificantly less dramatic than for wild-type RNA, despite thefact that all the sequences required for SXL function arepresent in the transcript. Although we cannot rule out thatSXL still affects the kinetics of splicing but shows little effect onthe accumulation of final products, the result argues that theeffects of SXL are reduced in the absence of a functionalcompeting 59 splice site. This conclusion is also consistent withprevious results analyzing minigenes in which both exon 2 andintron 2 were deleted (41).
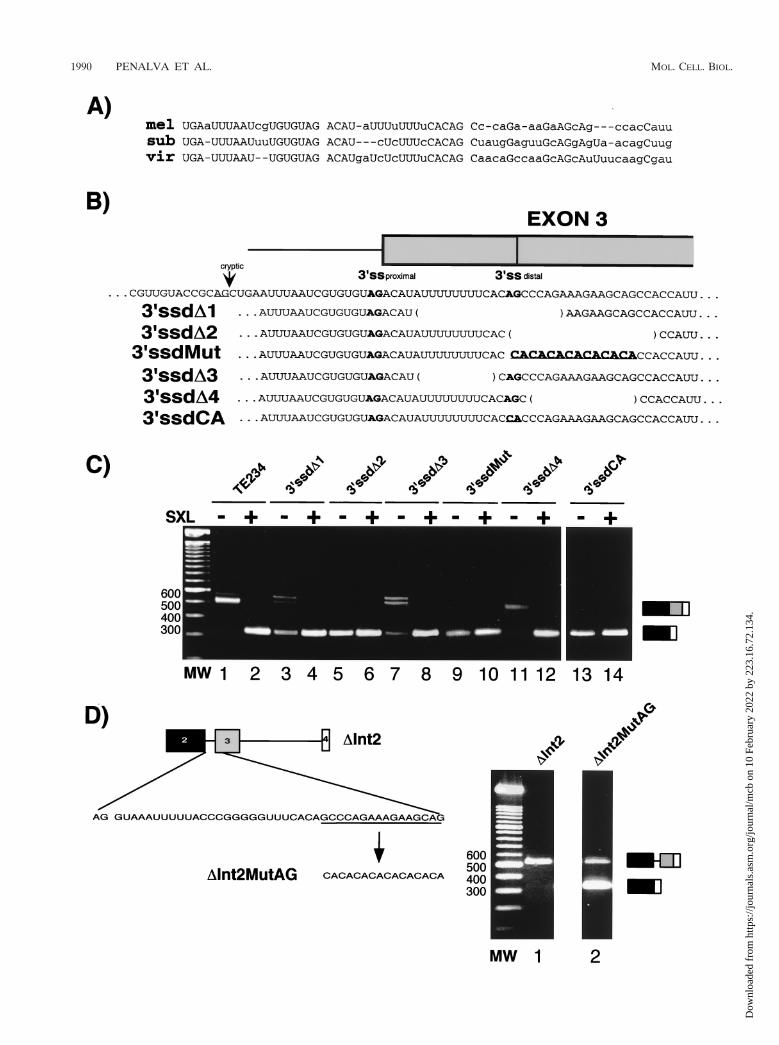
Regulation of 3* splice site usage. A feature conservedamong Sxl genes is the presence of two 39 splice sites precedingexon 3, of which the distal site is associated with a moreextensive Py tract (Fig. 3A). Sequencing of cDNA clones fromD. melanogaster male flies indicated that both sites are used(6). To test the possible roles of the alternative sites, a series ofmutant minigenes were generated (Fig. 3B). Deletion of thedistal site, including both the Py tract and the 39 splice site AG(39ssdD1), resulted in significant accumulation of transcripts inwhich exon 3 was skipped in the absence of SXL (Fig. 3C, lane3). This result suggests that the distal 39 splice site, with its longand U-rich Py tract, plays an important role in exon 3 defini-tion.
Because several AGs follow the distal 39 splice site andbecause it has been reported that downstream AGs can beactivated upon mutation of natural 39 splice sites (50), wereasoned that in order to determine the effect of mutations atthe distal AG and to prevent activation of the downstreamAGs, these needed to be either deleted (39ssdD2) or mutated(39ssdMut), together with the natural AG. Remarkably, dele-tion (39ssdD2) or substitution (39ssdMut) of the 39 splice siteAG, without introducing changes at the Py tract, resulted ineven stronger defects in exon definition (lanes 5 and 9, respec-tively). The downstream AG-rich stretch had, by itself, noeffect in either default or regulated splicing (39ssdD4, lanes 11and 12), indicating that the loss of exon definition in mutants39ssdD2 and 39ssdMut could be attributed exclusively to muta-
FIG. 1. Minigene structure, conserved sequences, and validation ofthe assay. (A) Schematic representation of the Sxl minigene present inplasmid copia-SxlTE234. Boxes represent exons 2 to 4 of Sxl mRNAs,thick lines represent introns, and patterns of splicing in the absence(2SXL) or in the presence (1SXL) of the female-specific protein SXLare indicated. The position of primers PT1 and PT2 used in RT-PCRsare indicated by arrows. (B) Analysis of the splicing pattern of copia-SxlTE234 transcripts by RT-PCR. Lane 1, cotransfection of copia-SxlTE234 and control plasmid pBShsp-cat; lane 2, cotransfection ofcopia-SxlTE234 and SXL expression vector pBShsp-SxlF1cDNA (5 mgof RNA was used in each experiment); lane 3, RT-PCR reaction using5 mg of RNA from cells cotransfected with copia-SxlTE234 plus pB-Shsp-cat and 5 mg of RNA from cells cotransfected with copia-Sx-lTE234 and pBShsp-SxlF1cDNA; lane 4, same as in lane 3 but using2.5 and 7.5 mg of the RNAs from the same cotransfections of lane 3,respectively; lane 5, same as in lane 3 but using 7.5 and 2.5 mg of theRNAs from the same cotransfections of lane 3, respectively. The struc-ture of the alternative PCR products is indicated on the right hand sideof the figure. The gray code for the exons is as in panel A. MW,molecular weight marker (100-bp ladder; Gibco). Some of the sizes areindicated. The expected sizes for the amplification products are 515 bp(splicing 2-3-4) and 325 bp (splicing 2–4) (41). The products of ampli-fication were cloned, and sequence analyses confirmed that they cor-respond to the predicted alternatively spliced products. (C) Values ofinput and measured RNA ratios. A, B, C, D, E, and F refer to thebands indicated in panel B. For band quantitation, the gel was stainedwith SYBR Green (Molecular Probes) and analyzed in a Fuji FLA-2000 phosphorimager using ImageQuant software (Molecular Dynam-ics).
1988 PENALVA ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 10
Feb
ruar
y 20
22 b
y 22
3.16
.72.
134.
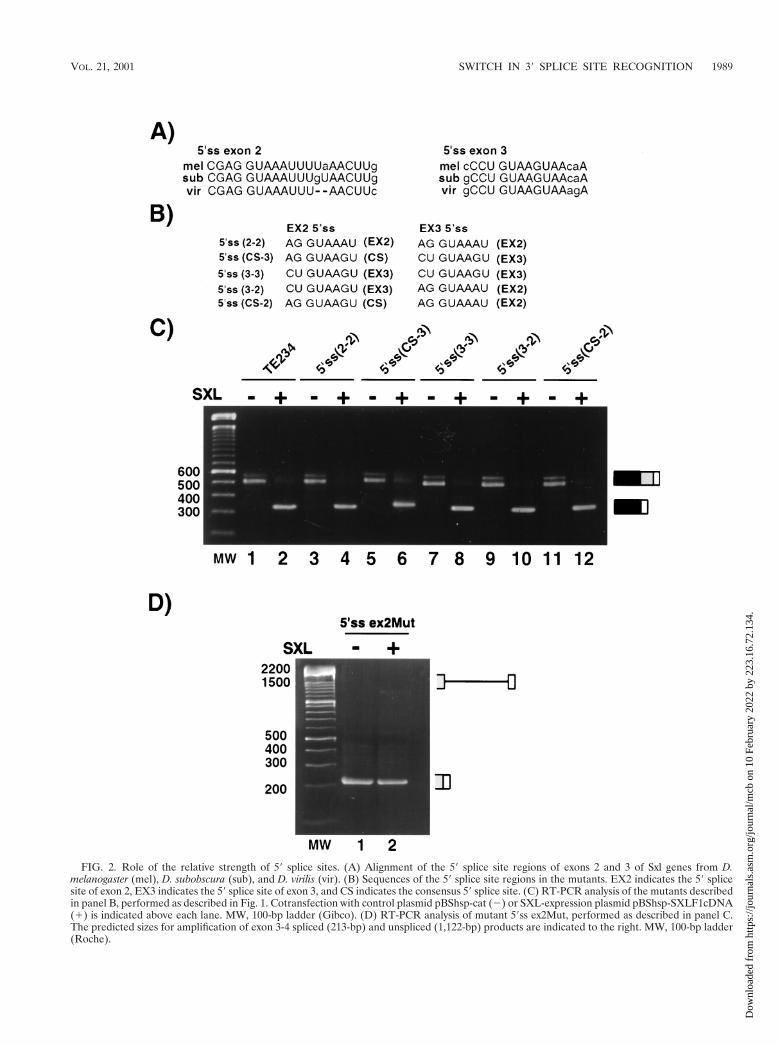
FIG. 2. Role of the relative strength of 59 splice sites. (A) Alignment of the 59 splice site regions of exons 2 and 3 of Sxl genes from D.melanogaster (mel), D. subobscura (sub), and D. virilis (vir). (B) Sequences of the 59 splice site regions in the mutants. EX2 indicates the 59 splicesite of exon 2, EX3 indicates the 59 splice site of exon 3, and CS indicates the consensus 59 splice site. (C) RT-PCR analysis of the mutants describedin panel B, performed as described in Fig. 1. Cotransfection with control plasmid pBShsp-cat (2) or SXL-expression plasmid pBShsp-SXLF1cDNA(1) is indicated above each lane. MW, 100-bp ladder (Gibco). (D) RT-PCR analysis of mutant 59ss ex2Mut, performed as described in panel C.The predicted sizes for amplification of exon 3-4 spliced (213-bp) and unspliced (1,122-bp) products are indicated to the right. MW, 100-bp ladder(Roche).
VOL. 21, 2001 SWITCH IN 39 SPLICE SITE RECOGNITION 1989
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 10
Feb
ruar
y 20
22 b
y 22
3.16
.72.
134.
1990 PENALVA ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 10
Feb
ruar
y 20
22 b
y 22
3.16
.72.
134.

tion of the distal 39 splice site AG. Accordingly, mutation ofthe distal 39 splice site AG alone to CA also resulted in efficientexon skipping even in the absence of SXL (39ssdCA, lanes 13and 14), indicating that downstream AGs could not be used forexon 3 definition.
The effects of deletion of the Py tract alone (39ssdD3, lane 7)or of deletion of both signals (39ssdD1, lane 3) appear lessdramatic than the effects of deletion or mutation of the 39splice site alone. This difference may be related to the fact thatcryptic splice sites become activated in these mutants: theamplification products corresponding to inclusion of exon 3 inthese mutants appear as a doublet (lanes 3 and 7); cloning andsequencing of the products revealed that both the proximal 39splice site and, unexpectedly, a cryptic 39 splice site located 20nucleotides upstream from the proximal 39 splice site in intron2 (indicated by an arrow in Fig. 3B) were utilized. Sequencingalso revealed an additional minor source of heterogeneity inthe use of a cryptic 39 splice site 24 nucleotides upstream ofexon 4, as observed in Fig. 2C.
We conclude that deletion of the Py tract associated with thedownstream 39 splice site of exon 3 causes, in the majority oftranscripts, either skipping of the exon or activation of a cryptic39 splice site upstream. Deletion of the AG dinucleotide asso-ciated with the distal 39 splice site causes efficient exon skip-ping. Taken together, the results argue that sequences associ-ated with the distal 39 splice site play a critical role in theproper definition of exon 3. Efficient exon 3 inclusion in theabsence of SXL is essential for the viability of male flies (46)(see also Discussion).
Previous work identified transcripts that used either theproximal or the distal 39 splice sites associated with Sxl exon 3(6). The strong effects of mutation of the distal 39 splice site inexon definition, however, suggest that the majority of the splic-ing events between exons 2 and 3 implicate recognition of thedistal site. This would also be consistent with the predictedrelative strength of their Py tracts.
To analyze the relative use of the distal and proximal sites inRNAs isolated from the transfected cells, RT-PCR analysiswas performed using oligonucleotide primers that allowed theamplification of the sequences at the junction between exons 2and 3. The products of amplification were cloned, and 20independent clones were sequenced. Surprisingly, 70% of theclones sequenced corresponded to cDNAs in which the prox-imal (and predictably weaker) 39 splice site was used. Similarestimates were obtained by electrophoretic analysis of the am-plification products (see below).
To further validate the analysis, the same RT-PCR approach
was used to analyze the use of exon 3 alternative 39 splice sitesin RNAs purified from male flies. The products of amplifica-tion were fractionated on a polyacrylamide gel that allowsseparation of the alternative products, that differ by 18 bp. Theresults (Fig. 4) show that the majority of the transcripts werespliced to the proximal site (compare lanes 1 and 3). This resultwas also confirmed by cloning the amplification products andsequencing 25 independent clones. As was the case for theminigene used in transient-transfection assays, more than 70%of the transcripts were spliced to the proximal 39 splice site. Weconclude that while the distal 39 splice site of exon 3 has an
FIG. 4. Relative use of exon 3 39 splice sites in Drosophila maleflies. RT-PCR was performed using primers corresponding to exons 2and 3 that should generate products of amplification differing by 18 bp,depending on whether the proximal or the distal 39 splice site areutilized. Lane 1, analysis of RNA from adult males; lane 2, controlPCR amplification of a cDNA in which exon 2 is spliced to the distal39 splice site of exon 3; lane 3, control PCR amplification of a cDNAin which exon 2 is spliced to the proximal 39 splice site of exon 3; MW,pBR322 MspI molecular weight marker. Some of the sizes (in basepairs) are indicated at the left.
FIG. 3. Role of exon 3 alternative 39 splice sites. (A) Alignment of exon 3 39 splice site region of Sxl genes from D. melanogaster (mel), D.subobscura (sub), and D. virilis (vir). The proximal and distal AG’s are separated by a space from the subsequent sequence. (B) Sequences of exon3 distal 39 splice site mutants. Exon 3 is represented as a gray box, the positions of the proximal, distal, and a cryptic 39 splice site, as well as thenucleotide sequence of the region, are indicated below the diagram. The sequences deleted in the different mutants are indicated between brackets.Nucleotide substitutions are underlined and in bold face. (C) RT-PCR analysis of the mutants indicated in panel B, performed as described in Fig.1. Cotransfection with control plasmid (2) or SXL expression plasmid (1) is indicated above each lane. MW, 100-bp ladder (Gibco). (D) On theleft is a schematic representation of 59 splice site switch constructs DInt2 and DInt2MutAG. The boxes show the exons, and the lines show theintrons. The sequence of the region between exon 2 59 splice site and the 59 end of exon 3 is indicated. The region mutated in clone DInt2MutAGis underlined, and the mutant sequence is indicated after the arrow. On the right is an RT-PCR analysis, performed as described in Fig. 1. Theproducts of amplification corresponding to the use of exon 2 or exon 3 59 splice sites are indicated. The expected sizes for the amplification productsare 325 and 537 bp, respectively. The products were cloned, and sequencing confirmed that they correspond to the expected alternatively splicedproducts. MW, 100-bp ladder (Roche).
VOL. 21, 2001 SWITCH IN 39 SPLICE SITE RECOGNITION 1991
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 10
Feb
ruar
y 20
22 b
y 22
3.16
.72.
134.

important role in exon definition, the majority of the actualsplicing events take place using the proximal 39 splice site.
To further substantiate a role for the 39 AG in exon defini-tion, uncoupled from its function in splicing catalysis, we pre-pared a minigene in which most of intron 2 was deleted suchthat splicing between exons 2 and 3 could not take place. Thedeletion also removed the proximal 39 splice site as well as partof the Py tract associated with the distal site (Fig. 3D). In thisconstruct (DInt2) the 59 splice sites of exons 2 and 3 competefor the only available 39 splice site of exon 4. In the absence ofSXL, exon 3 was spliced to exon 4 (Fig. 3D, lane 1). The AGcorresponding to exon 3 distal 39 splice site, as well as otherAGs immediately downstream, were mutated in constructDInt2MutAG (Fig. 3D). A significant fraction of splicing tookplace between exons 2 and 4 in these transcripts (Fig. 3D, lane2). This result indicates that the distal AG plays a critical rolein the recognition and activation of exon 3 59 splice site despitethe fact that this AG cannot participate in splicing reactions,and the associated Py tract has been significantly shortened(from eight to three U’s).
Role of the proximal 3* splice site. The results presentedabove indicate that the distal 39 splice site is essential for exondefinition, while the proximal site is preferentially used incatalysis. The analysis of mutants at the proximal 39 splice sitewas therefore particularly intriguing. Different concentrationsof SXL-encoding plasmid were cotransfected together with thereporter to quantitatively assess differences in response to theSXL protein, which accumulated in proportion to the amountof DNA transfected (data not shown). Mutation of the AGassociated with the proximal 39 splice site to CA (Fig. 5A,39sspCA) did not have any effect in exon 3 definition (Fig. 5B,lane 5). Cloning and sequencing of the amplification product,as well as RT-PCR analyses (Fig. 6, lanes 3 and 4), indicatedthat the distal 39 splice site AG was utilized, indicating that useof the proximal site is not strictly required for exon 3 inclusion.Strikingly, however, exon skipping in the presence of SXL wasstrongly reduced (Fig. 5B, compare lanes 2 to 4 with lanes 6 to8): 100-fold-higher concentrations of SXL-encoding plasmidwere required to achieve 50% exon skipping. These differencesare likely to have physiological relevance because even twofoldvariations in SXL expression can have important developmen-tal consequences (46). The dramatic effects of these minimalmutations are in sharp contrast with the modest effects thatindividual (or even multiple) deletion of other regulatory se-quences (e.g., SXL-binding sites) have in SXL function (26,41). Taken together, the data suggest that dual recognition ofthe 39 splice sites of exon 3 is critical to allow efficient regula-tion by SXL.
One possible caveat of this result was that the mutation notonly inactivated the proximal 39 splice site but also affected aputative branch point associated with the distal site. Althoughthe precise location of the branch point is not known, it isconceivable that the sequence changes introduced in mutant39sspCA could have improved the strength of the distal 39splice site branch point, somehow antagonizing the repressiveeffect of SXL. To address this issue, two new mutants wereprepared in which (i) the proximal AG was mutated to GG,which should be neutral regarding the creation of crypticbranch points, and (ii) the proximal AG and adjacent se-quences were deleted (Fig. 5A, 39sspD), thus disrupting a
branch point associated with the distal 39 splice site in thatregion. Analysis of these mutants indicated that neither theAG-to-GG substitution (data not shown) nor deletion of theAG affected exon 3 inclusion (Fig. 5B, lane 9), but they signif-icantly reduced the effect of SXL in exon skipping (Fig. 5B,compare lanes 2 to 4 and lanes 10 to 12), as was the case withthe AG-to-CA mutation. RT-PCR analysis showed that thedistal AG was used in the 39sspD1 mutant as well (Fig. 6, lane5), confirming that the distal AG can functionally undergocatalysis in the absence of a proximal AG. Taken together, theresults of Fig. 5B suggest that mutation or deletion of theproximal 39 splice site compromises SXL regulation and arguethat the occurrence of a switch in 39 splice site recognitionbetween exon definition and splicing catalysis allows SXL toefficiently interfere with exon 3 inclusion.
The switch in 39 splice site recognition could take place byvery different mechanisms, depending on whether or not the Pytract associated with the proximal site is functionally recog-nized by early splicing factors such as U2AF65/35 (see Discus-sion). To address this issue, a mutant was generated in whichall the uridines in the Py tract associated with the proximal 39splice site were mutated to cytidines (Fig. 5A). Such mutationsare predicted to result in significant decreases in the affinity ofU2AF65 binding (47, 48, 61). The results of Fig. 5C indicatethat the mutation was rather neutral regarding both exon def-inition and regulation by SXL, suggesting that the strength ofthe Py tract associated with the proximal site is not importantfor 39 splice site activation or regulation and arguing that theswitch in 39 splice site recognition involves a single recognitionevent at the Py tract and possibly a single branch point recog-nition event.
DISCUSSION
We report in this manuscript that two 39 splice site AGsassociated with Sxl exon 3 have very different roles in alterna-tive splicing of this exon. The distal AG is critical for properexon inclusion, presumably through interactions with factorsbound to the 59 splice site (e.g., U1 snRNP) across the exon(7). The proximal AG, in contrast, is preferentially used forcatalysis, and its mutation, rather than affecting exon defini-tion, compromises efficient regulation by SXL.
Two separable steps in 3* splice site recognition. Analyses ofprotection to RNase H-mediated degradation indicated thatthe 39 splice site AG is recognized twice during the splicingreaction in higher eukaryotes, first at early steps of spliceosomeassembly and then again at the time of the second transterifi-cation reaction leading to exon ligation (45). Several lines ofevidence argue that the two steps are, at least to some degree,independent. First, pre-mRNAs containing 39 splice sites withstrong Py tracts can undergo the first transterification reactionin vitro in the absence of a 39 splice site AG (39), as is thegeneral case for budding yeast pre-mRNAs (36, 40). Second, 39exon ligation can be accomplished in trans by exposing a spli-ceosome that has carried out the first step to an exogenouslyadded 39 exon (2), persuasively demonstrating that catalysisdoes not necessarily require early AG recognition within thesame molecule. Our results show that the particular configu-ration of 39 splice sites in Sxl exon 3 allows the physical sepa-ration of the two steps on two different AGs, and they also
1992 PENALVA ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 10
Feb
ruar
y 20
22 b
y 22
3.16
.72.
134.
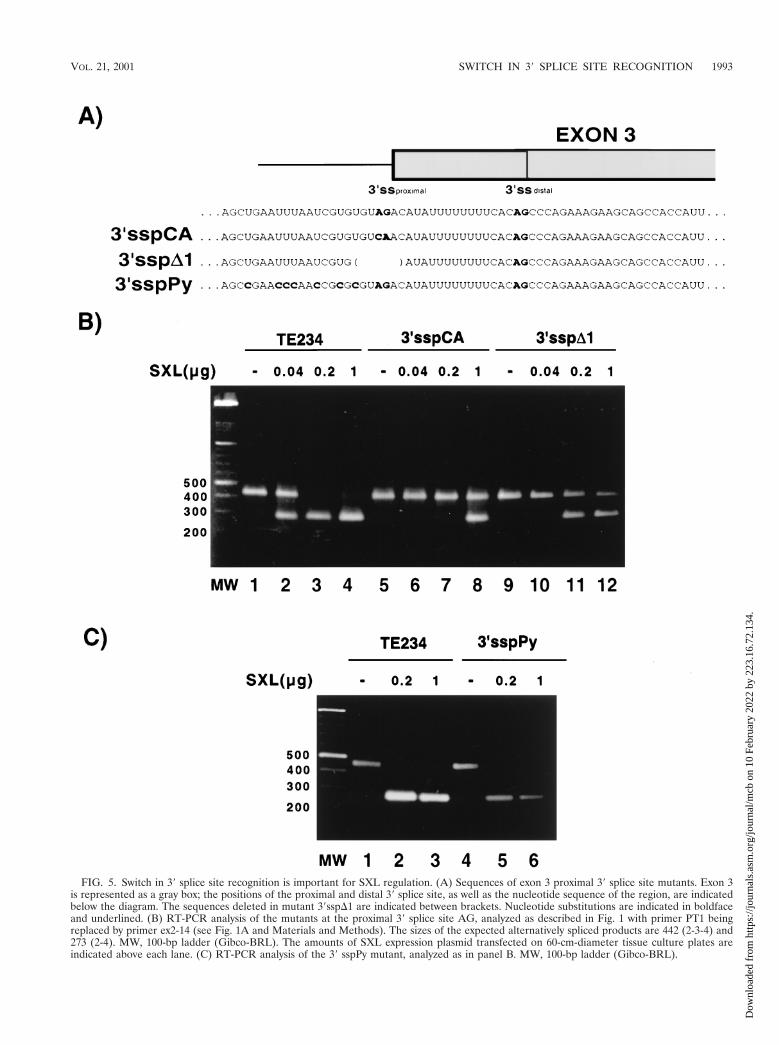
FIG. 5. Switch in 39 splice site recognition is important for SXL regulation. (A) Sequences of exon 3 proximal 39 splice site mutants. Exon 3is represented as a gray box; the positions of the proximal and distal 39 splice site, as well as the nucleotide sequence of the region, are indicatedbelow the diagram. The sequences deleted in mutant 39sspD1 are indicated between brackets. Nucleotide substitutions are indicated in boldfaceand underlined. (B) RT-PCR analysis of the mutants at the proximal 39 splice site AG, analyzed as described in Fig. 1 with primer PT1 beingreplaced by primer ex2-14 (see Fig. 1A and Materials and Methods). The sizes of the expected alternatively spliced products are 442 (2-3-4) and273 (2-4). MW, 100-bp ladder (Gibco-BRL). The amounts of SXL expression plasmid transfected on 60-cm-diameter tissue culture plates areindicated above each lane. (C) RT-PCR analysis of the 39 sspPy mutant, analyzed as in panel B. MW, 100-bp ladder (Gibco-BRL).
VOL. 21, 2001 SWITCH IN 39 SPLICE SITE RECOGNITION 1993
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 10
Feb
ruar
y 20
22 b
y 22
3.16
.72.
134.

argue for a critical function for the 39 splice site AG sequencein exon definition.
The first AG recognition step is important for definition ofthe 39 splice site region in cooperation with the Py tract andinvolves the two subunits of U2AF. U2AF65 binds to the Pytract (48, 59, 60), and U2AF35 binds to the AG region (34, 58,63). Recognition of the AG by U2AF35 is particularly criticalfor the splicing of pre-mRNAs containing a weak 39 splice site,which are therefore classified as AG-dependent substrates (39,58; reviewed in reference 35). Interestingly, although the Pytract associated with the distal site of Sxl exon 3 is relativelystrong (containing eight uridines in a row), the adjacent AGplays a critical role in exon 3 definition. The distal AG isimportant for the use of exon 3 59 splice site even when thesequence cannot undergo splicing reactions and the associatedPy tract has been significantly shortened (Fig. 3D). It is possi-ble that the stabilization of U2AF65 binding afforded byU2AF35 is critical for exon definition or that interaction ofU2AF35 with the distal AG promotes other steps in spliceo-some assembly (23, 57). Remarkably, the effects of deletion ofthe distal 39 splice site AG are more dramatic than the effectsof deleting only the Py tract or even of deleting both signals.One possible explanation for these observations is that recog-nition of the Py tract by U2AF65 initiates a cascade of eventsleading to exon definition and that this cascade relies on rec-ognition of the 39 splice site AG by U2AF35. If this interactiondoes not occur, abortive exon definition ensues. In the absenceof the Py tract, however, U2AF is directed to 39 splice sitesupstream and exon definition occurs through those signals,albeit less efficiently. Efficient exon 3 inclusion in the absenceof SXL is a critical feature of the Sxl genetic switch: evenmodest levels of exon skipping in males could produce enoughSXL protein to initiate the autoregulatory loop, promoting theaccumulation of more transcripts lacking exon 3 and therefore
the accumulation of more SXL protein, ultimately leading tolethality (reviewed in reference 46).
Despite the important role of the distal AG in exon defini-tion, catalysis occurs primarily using the proximal AG (Fig. 4and 6). One possible explanation for these results could be thatthe distal AG is not a functional splice site but rather acts as anexonic splicing enhancer sequence (ESE) of the proximal site.Indeed, the sequence encompassing the distal AG and se-quences immediately downstream is purine-rich, and ESEsmade of purine repeats have been shown to facilitate the use ofupstream weak 39 splice sites (56; reviewed in reference 8).Several results make this scenario very unlikely and argue thatthe distal AG is being recognized as a functional 39 splice sitefor purposes of exon definition. First, all known ESEs arecomposed of more than two functionally relevant nucleotidesand, as in the case of purine-rich enhancers, their activity isproportional to the number of short sequence repeats (52). Incontrast, only deletion or mutation of the distal AG (and not ofall the AGs downstream) has an effect in exon 3 definition.Second, the distal AG is preceded by a functional Py tract (Fig.3C), whereas the proximal site is not (Fig. 5C). Third, the distalPy tract and AG can function as a 39 splice site region when theproximal AG is mutated (Fig. 5 and 6). Fourth, the distal AGhas functions in splice site activation even when the proximalsite has been deleted and the distal site itself cannot undergosplicing (Fig. 3D).
Mechanisms of 3* splice site switch. Two molecular mech-anisms could account for the switch in 39 splice site recogni-tion. First, early factors such as U2AF65/35 could reassemble onthe upstream 39 Py tract and AG or be simultaneously boundto both sites and be differentially efficient for establishing in-teractions leading to exon definition (distal . proximal) orleading to intron removal (proximal . distal). The absence ofeffects of mutations at the Py tract upstream from the proximalAG (Fig. 5C), however, suggest that the proximal AG is notactivated from an upstream Py tract following the conventionalmechanism of 39 splice site activation.
An alternative scenario is that recognition of the distal siteresults in branch point definition, from which the AG used forcatalysis will be specified. How the 39 AG is determined at thetime of exon ligation is not completely understood, but workon substrates containing long Py tracts distant from the 39splice sites, as well as trans-splicing assays in vitro, suggestedthe existence of a 59-to-39 linear scanning mechanism thatwould identify the first AG from the Py tract-branch pointregion (13, 49, 50). Competition and local context effectswithin a window of sequence from the branch point also con-tribute to defining the AG undergoing catalysis (13, 14, 33, 37,50, 53).
In Sxl exon 3, early recognition of the distal site allows theuse of the proximal one for catalysis. Other examples of suchdual use have been reported previously. First, a mutation in thehuman b-globin gene, associated with a form of b-thalassemia,generates a 39 splice site AG between the branchpoint and thenormal 39 splice site, leading to aberrant splicing and the syn-thesis of a nonfunctional globin mRNA. Surprisingly, experi-mental inactivation of the wild-type AG in the mutant RNAaffected the efficiency with which the upstream cryptic splicesite was used in vitro (30, 61). A second example was found inthe early transcriptional unit of polyomavirus, which produces
FIG. 6. Use of AGs in proximal 39 splice site mutants. RNA iso-lated from transfected cells was analyzed by RT-PCR as in Fig. 4. Thetransfected reporter plasmid is indicated at the top. Cotransfectionwith control plasmid (2) or the SXL expression plasmid (1) is alsoindicated. The positions of the amplification products correspondingto use of the proximal and distal sites are indicated. Amplificationproducts were cloned, and sequence analyses confirmed that they cor-respond to splicing to the assigned AGs.
1994 PENALVA ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 10
Feb
ruar
y 20
22 b
y 22
3.16
.72.
134.

three tumor antigens (StAg, MTAg, and LTAg) encoded byalternatively spliced transcripts. It was found that the use of aStAg 39 splice site required the presence of a MTAg 39 splicesite located 14 nucleotides downstream (19). These studies ledto the conclusion that selection of the proximal site could occurafter branch formation, possibly through a scanning mecha-nism, and therefore that the 39 splice site requirements for 59cleavage and lariat formation can be uncoupled from thoserequired for 39 cleavage and ligation. Furthermore, observa-tions made using plant introns in which 39 splice sites seques-tered within stems of hairpin structures could be activated by“helper” downstream 39 splice sites were consistent with thepossibility that 39 splice sites can be recognized at least twiceduring spliceosome assembly prior to catalysis (31).
Switch in 3* splice site recognition and SXL function. Ourmost intriguing result was the apparent requirement of a switchin 39 splice AG recognition for efficient SXL function. Analternative explanation would be that the proximal AG is im-portant for SXL binding. The binding specificity of SXL, how-ever, is mainly dictated by U-rich sequences (24, 42, 47, 48),and although Sakashita and Sakamoto reported a preferencefor adjacent AG’s, these were located 39, not 59, from U-richstretches (42). In addition, while mutation of the proximal AGstrongly inhibits SXL function (Fig. 5B), drastic changes in asingle SXL binding site (particularly in the U8 Py-tract asso-ciated with the distal site) do not have substantial effects inSXL regulation (26, 41).
How can the switch in AG recognition facilitate SXL func-tion? The second AG recognition step has been associatedwith the second step of the splicing reaction (15, 53). To ac-complish its role in promoting exon skipping, however, SXLneeds to act even before the first step of catalysis, either byforcing branch point formation in the vicinity of exon 4 ratherthan exon 3 or by affecting splice site pairing across the introns.Our observations therefore suggest that SXL discriminateswhether the proximal or the distal site will be used even beforethe 29-59 branch is formed. The first implication of this hypoth-esis is that recognition of different AGs results in exon defini-tion complexes of different composition and/or different prop-erties for establishing partnerships with neighboring 59 splicesites. The second implication is that the different complexesare differentially sensitive to SXL function. It is conceivablethat splitting the two steps of 39 splice site recognition betweentwo physically different AGs opens a window of opportunity forSXL repression. Binding of U2 snRNP to the branch point, forexample, could be less stable when the proximal site is recog-nized. This lower stability could have little effect in splice siteactivation in the absence of SXL but be very detrimental in itspresence.
SXL could interfere with the assembly of splicing factors orsomehow inactivate the factors bound. Genetic results haveshown that mutant alleles of the gene sans-fille (snf), whichencodes a protein component of both U1 and U2 snRNPs,show deficient Sxl autoregulation when combined with partic-ular Sxl mutants (1, 18, 43, 44). It is conceivable that interac-tions between SXL and SNF (16, 17) compromise splice siteactivation.
If the precise structure of the 39 splice sites and the apparentswitch in 39 splice site recognition is important for SXL func-tion, why are the cryptic 39 splice sites that are activated by
mutation of the natural sites properly regulated, both in tran-sient tranfections (Fig. 5) and in transgenic flies (26)? Onepossibility is that the cryptic sites also have a similar configu-ration of nearby competing 39 splice sites. Indeed, analysis of 39splice site utilization indicated that both the cryptic site and theproximal site (now in a distal configuration relative to thecryptic site) are used (Fig. 3C and data not shown).
A plausible reason for maintaining the duality in 39 splicesite recognition is that other 39 splice sites regulatable by SXLwould not be strong enough to prevent even small levels ofexon skipping in the absence of SXL. As mentioned above,efficient inclusion of exon 3 is critical for male survival (46).The strong signals required for full exon definition would thendemand a mechanism like the switch in 39 splice site recogni-tion mentioned above to allow efficient regulation by SXL.Thus, the configuration of 39 splice sites preceding exon 3could have evolved to accomplish both stringent exon inclusionin the absence of regulator and an opportunity for the regula-tor to interfere.
ACKNOWLEDGMENTS
We thank Hiroshi Sakamoto, Kunio Inoue, and Yoshiro Shimura forthe kind gift of plasmids and protocols; Douglas Black, Peter Nielsen,and Chris Smith for discussions; Witold Filipowicz for pointing outrelated results in plants; an anonymous reviewer for pointing out asimilar example of 39 splice site recognition in polyoma early tran-scripts; and Iain Mattaj and Bertrand Seraphin for suggestions on themanuscript.
L.O.F.P. was supported by an EMBO postdoctoral fellowship.M.J.L. was supported by Fundacion Ramon Areces (Spain) and MarieCurie-European Union postdoctoral fellowships.
L.O.F.P. and M.J.L. contributed equally to this study.
REFERENCES
1. Albrecht, E. B., and H. K. Salz. 1993. The Drosophila sex determination genesnf is utilized for the establishment of the female-specific splicing pattern ofSex-lethal. Genetics 134:801–807.
2. Anderson, K., and M. J. Moore. 1997. Bimolecular exon ligation by thehuman spliceosome. Science 276:1712–1716.
3. Anderson, K., and M. J. Moore. 2000. Biomolecular exon ligation by thehuman spliceosome bypasses early 39 splice site AG recognition and requiresNTP hydrolysis. RNA 6:16–25.
4. Bashaw, G. J., and B. S. Baker. 1995. The msl-2 dosage compensation geneof Drosophila encodes a putative DNA-binding protein whose expression issex specifically regulated by Sex-lethal. Development 121:3245–3258.
5. Bashaw, G. J., and B. S. Baker. 1997. The regulation of the Drosophila msl-2gene reveals a function for Sex-lethal in translational control. Cell 89:789–798.
6. Bell, L. R., J. I. Horabin, P. Schedl, and T. W. Cline. 1991. Positive auto-regulation of sex-lethal by alternative splicing maintains the female deter-mined state in Drosophila. Cell 65:229–239.
7. Berget, S. M. 1995. Exon recognition in vertebrate splicing J. Biol. Chem.270:2411–2414.
8. Blencowe, B. J. 2000. Exonic splicing enhancers: mechanism of action, di-versity and role in human genetic diseases. Trends Biochem. Sci. 25:106–110.
9. Boggs, R. T., P. Gregor, S. Idriss, J. M. Belote, and M. McKeown. 1987.Regulation of sexual differentiation in D. melanogaster via alternative splic-ing of RNA from the transformer gene. Cell 50:739–747.
10. Bopp, D., G. Calhoun, J. I. Horabin, M. Samuels, and P. Schedl. 1996.Sex-specific control of Sex-lethal is a conserved mechanism for sex determi-nation in the genus Drosophila. Development 122:971–982.
11. Campuzano, S., L. Carramolino, C. V. Cabrera, M. Ruiz-Gomez, R. Villares,A. Boronat, and J. Modolell. 1985. Molecular genetics of the achaete-scutegene complex of D. melanogaster. Cell 40:327–338.
12. Chabot, B. 1996. Directing alternative splicing: cast and scenarios. TrendsGenet. 12:472–478.
13. Chen, S., K. Anderson, and M. J. Moore. 2000. Evidence for a linear searchin bimolecular 39 splice site AG selection. Proc. Natl. Acad. Sci. USA 97:593–598.
14. Chiara, M. D., L. Palandjian, R. F. Kramer, and R. Reed. 1997. Evidencethat U5 snRNP recognizes the 39 splice site for catalytic step II in mammals.EMBO J. 16:4746–4759.
VOL. 21, 2001 SWITCH IN 39 SPLICE SITE RECOGNITION 1995
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 10
Feb
ruar
y 20
22 b
y 22
3.16
.72.
134.

15. Chua, K., and R. Reed. 1999. The RNA splicing factor hSlu7 is required forcorrect 39 splice site choice. Nature 402:207–210.
16. Cline, T. W., D. Z. Rudner, D. A. Barbash, M. Bell, and R. Vutien. 1999.Functioning of the Drosophila integral U1/U2 protein Snf independent of U1and U2 small nuclear ribonucleoprotein particles is revealed by snf(1) genedose effects. Proc. Natl. Acad. Sci. USA 96:14451–14458.
17. Deshpande, G., M. E. Samuels, and P. D. Schedl. 1996. Sex-lethal interactswith splicing factors in vitro and in vivo. Mol. Cell. Biol. 16:5036–5047.
18. Flickinger, T. W., and H. K. Salz. 1994. The Drosophila sex determinationgene snf encodes a nuclear protein with sequence and functional similarity tothe mammalian U1A snRNP protein. Genes Dev. 8:914–925.
19. Ge, H., J. Noble, J. Colgan, and J. L. Manley. 1990. Polyoma virus smalltumor antigen pre-mRNA splicing requires cooperation between two 39splice sites. Proc. Natl. Acad. Sci. USA 87:3338–3342.
20. Gebauer, F., L. Merendino, M. W. Hentze, and J. Valcarcel. 1998. TheDrosophila splicing regulator Sex-lethal directly inhibits translation of male-specific-lethal 2 mRNA. RNA 4:142–150.
21. Granadino, B., L. O. F. Penalva, M. R. Green, J. Valcarcel, and L. Sanchez.1997. Distinct mechanisms of splicing regulation in vivo by the Drosophilaprotein Sex-lethal. Proc. Natl. Acad. Sci. USA 94:7343–7348.
22. Green, M. R. 1986. Pre-mRNA splicing. Annu. Rev. Genet. 20:671–708.23. Guth, S., C. Martinez, R. K. Gaur, and J. Valcarcel. 1999. Evidence for
substrate-specific requirement of the splicing factor U2AF(35) and for itsfunction after polypyrimidine tract recognition by U2AF(65). Mol. Cell. Biol.19:8263–8271.
24. Handa, N., O. Nureki, K. Kurimoto, I. Kim, H. Sakamoto, Y. Shimura, Y.Muto, and S. Yokoyama. 1999. Structural basis for recognition of the tramRNA precursor by the Sex-lethal protein. Nature 398:579–585.
25. Horabin, J. I., and P. Schedl. 1993. Regulated splicing of the Drosophilasex-lethal male exon involves a blockage mechanism. Mol. Cell. Biol. 13:1408–1414.
26. Horabin, J. I., and P. Schedl. 1993. Sex-lethal autoregulation requires mul-tiple cis-acting elements upstream and downstream of the male exon andappears to depend largely on controlling the use of the male exon 59 splicesite. Mol. Cell. Biol. 13:7734–7746.
27. Inoue, K., K. Hoshijima, H. Sakamoto, and Y. Shimura. 1990. Binding of theDrosophila Sex-lethal gene product to the alternative splice site of trans-former primary transcript. Nature 344:461–463.
28. Kelley, R. L., I. Solovyeva, L. M. Lyman, R. Richman, V. Solovyev, and M. I.Kuroda. 1995. Expression of msl-2 causes assembly of dosage compensationregulators on the X chromosomes and female lethality in Drosophila. Cell.81:867–877.
29. Kelley, R. L., J. Wang, L. Bell, and M. I. Kuroda. 1997. Sex lethal controlsdosage compensation in Drosophila by a non-splicing mechanism. Nature387: 195–199.
30. Krainer, A. R., R. Reed, and T. Maniatis. 1985. Mechanisms of humanb-globin pre-mRNA splicing, p. 353–382. In P. Berg (ed.), The Robert A.Welch Foundation conferences on chemical research. XXIX. Genetic chem-istry: the molecular basis of heredity. Welch Foundation, Houston, Tex.
31. Liu, H.-X., G. J. Goodall, R. Kole, and W. Filipowicz. 1995. Effects ofsecondary structure on pre-mRNA splicing: hairpins sequestering the 59 butnot the 39 splice site inhibit intron processing in Nicotiana plumbaginifolia.EMBO J. 14:377–388.
32. Lopez, A. J. 1998. Alternative splicing of pre-mRNA: developmental conse-quences and mechanisms of regulation. Annu. Rev. Genet. 32:279–305.
33. Luukkonen, B. G. M., and B. Seraphin. 1997. The role of branchpoint-39splice site spacing and interaction between intron terminal nucleotides insplice site selection in Saccharomyces cerevisiae. EMBO J. 16:779–792.
34. Merendino, L., S. Guth, D. Bilbao, C. Martinez, and J. Valcarcel. 1999.Inhibition of msl-2 splicing by Sex-lethal reveals interaction betweenU2AF35 and the 39 splice site AG. Nature 402:838–841.
35. Moore, M. J. 2000. Intron recognition comes of AGe Nat. Struct. Biol.7:14–16.
36. Parker, R., and B. Patterson. 1987. Architecture of fungal introns: implica-tions for spliceosome assembly, pp. 133–150. In M. Inouye and B. Dudock,(ed.), Molecular biology of RNA: new perspectives. Academic Press, NewYork, N.Y.
37. Patterson, B., and C. Guthrie. 1991. A U-rich tract enhances usage of analternative 39 splice site in yeast. Cell 64:181–187.
38. Penalva, L. O. F., H. Sakamoto, A. Navarro-Sabate, E. Sakashita, B. Gra-nadino, C. Segarra, and L. Sanchez. 1996. Regulation of the gene Sex-lethal:a comparative analysis of Drosophila melanogaster and Drosophila subob-scura. Genetics 144:1653–1664.
39. Reed, R. 1989. The organization of 39 splice sequences in mammalian in-trons. Genes Dev. 3:2113–2123.
40. Rymond, B. C., and M. Rosbash. 1985. Cleavage of 59 splice site and lariatformation are independent of 39 splice site in yeast mRNA splicing. Nature317: 735–737.
41. Sakamoto, H., K. Inoue, I. Higuchi, Y. Ono, and Y. Shimura. 1992. Controlof Drosophila Sex-lethal pre-mRNA splicing by its own female-specific prod-uct. Nucleic Acids Res. 20:5533–5540.
42. Sakashita, E., and H. Sakamoto. 1996. Protein-RNA and protein-proteininteractions of the Drosophila Sex-lethal mediated by its RNA-binding do-mains. J. Biochem. 120:1028–1033.
43. Salz, H. K. 1992. The genetic analysis of snf. a Drosophila sex determinationgene required for activation of Sex-lethal in both the germline and the soma.Genetics 130:547–554.
44. Salz, H. K., and T. W. Flickinger. 1996. Both loss-of-function and gain-of-function mutations in snf define a role for snRNP proteins in regulatingSex-lethal pre-mRNA splicing in Drosophila development. Genetics 144:95–108.
45. Sawa, H., and Y. Shimura. 1991. Alterations of RNase H sensitivity of the 39splice site region during the in vitro splicing reaction. Nucleic Acids Res.19:3953–3958.
46. Schutt, C., and R. Nothiger. 2000. Structure, function and evolution ofsex-determining systems in dipteran insects. Development 127:667–677.
47. Singh, R., H. Banerjee, and M. R. Green. 2000. Differential recognition ofthe polypyrimidine-tract by the general splicing factor U2AF65 and thesplicing repressor Sex-lethal. RNA 6:901–911.
48. Singh, R., J. Valcarcel, and M. R. Green. 1995. Distinct binding specificitiesand functions of higher eukaryotic polypyrimidine tract-binding proteins.Science 268:1173–1176.
49. Smith, C. W. J., E. B. Porro, J. G. Patton, and B. Nadal-Ginard. 1989.Scanning from an independently specified branch point defines the 39 splicesite of mammalian introns. Nature 342:243–247.
50. Smith, C. W. J., T. T. Chu, and B. Nadal-Ginard. 1993. Scanning andcompetition between AGs are involved in 39 splice site selection in mam-malian introns. Mol. Cell. Biol. 13:4939–4952.
51. Sosnowski, B. A., J. M. Belote, and M. McKeown. 1989. Sex-specific alter-native splicing of RNA from the transformer gene results from sequence-dependent splice site blockage. Cell 58:449–459.
52. Tanaka, K., A. Watakabe, and Y. Shimura. 1994. Polypurine sequenceswithin a downstream exon function as a splicing enhancer. Mol. Cell. Biol.14:1347–1354.
53. Umen, J. G., and C. Guthrie. 1995. Prp16p, Slu7p, and Prp8p interact withthe 39 splice site in two distinct stages during the second catalytic step ofpre-mRNA splicing. RNA 1:584–597.
54. Valcarcel, J., R. Singh, P. D. Zamore, and M. R. Green. 1993. The proteinSex-lethal antagonizes the splicing factor U2AF to regulate alternative splic-ing of transformer pre-mRNA. Nature 362:171–175.
55. Wang, J., and L. R. Bell. 1994. The Sex-lethal amino terminus mediatescooperative interactions in RNA binding and is essential for splicing regu-lation. Genes Dev. 8:2072–2085.
56. Watakabe, A., K. Tanaka, and Y. Shimura. 1993. The role of exon sequencesin splice site selection. Genes Dev. 7:407–418.
57. Wu, J. Y., and T. Maniatis. 1993. Specific interactions between proteinsimplicated in splice site selection and regulated alternative splicing. Cell 17:1061–1070.
58. Wu, S., C. M. Romfo, T. W. Nilsen, and M. R. Green. 1999. Functionalrecognition of the 39 splice site AG by the splicing factor U2AF35. Nature402:832–835.
59. Zamore, P. D., and M. R. Green. 1991. Biochemical characterization of U2snRNP auxiliary factor: an essential pre-mRNA splicing factor with a novelintranuclear distribution. EMBO J. 10:207–214.
60. Zamore, P. D., J. G. Patton, and M. R. Green. 1992. Cloning and domainstructure of the mammalian splicing factor U2AF. Nature 13:609–614.
61. Zhuang, Y., and A. M. Weiner. 1990. The conserved dinucleotide AG of the39 splice site may be recognized twice during in vitro splicing of mammalianmRNA precursors. Gene 90:263–269.
62. Zhou, S., Y. Yang, M. J. Scott, A. Pannuti, K. C. Fehr, A. Eisen, E. V. Koonin,D. L. Fouts, R. Wrightsman, J. E. Manning, and J. Lucchesi. 1995. Male-specific lethal 2, a dosage compensation gene of Drosophila, undergoes sex-specific regulation and encodes a protein with a RING finger and a metal-lothionein-like cysteine cluster. EMBO J. 14:2884–2895.
63. Zorio, D. A., and T. Blumenthal. 1999. Both subunits of U2AF recognize the39 splice site in Caenorhabditis elegans. Nature 402:835–838.
1996 PENALVA ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 10
Feb
ruar
y 20
22 b
y 22
3.16
.72.
134.