the actin regulator n-wasp is required for muscle-cell ... · the actin regulator n-wasp is...
TRANSCRIPT

The actin regulator N-WASp is required for muscle-cellfusion in miceYael Gruenbaum-Cohena, Itamar Harelb, Kfir-Baruch Umanskya, Eldad Tzahorb, Scott B. Snapperc, Ben-Zion Shiloa,1,and Eyal D. Schejtera,1
aDepartment of Molecular Genetics and bDepartment of Biological Regulation, Weizmann Institute of Science, Rehovot 76100, Israel; and cDepartment ofMedicine and Immunology, Massachusetts General Hospital, Harvard Medical School, Boston, MA 02115
Edited by Margaret Buckingham, Pasteur Institute, Paris, France, and approved May 21, 2012 (received for review September 30, 2011)
A fundamental aspect of skeletal myogenesis involves extensiverounds of cell fusion, in which individual myoblasts are incorporatedinto growing muscle fibers. Here we demonstrate that N-WASp, aubiquitous nucleation-promoting factor of branched microfilamentarrays, is an essential contributor to skeletal muscle-cell fusion indeveloping mouse embryos. Analysis both in vivo and in primarysatellite-cell cultures, shows that disruption of N-WASp functiondoes not interfere with the program of skeletal myogenic differ-entiation, and does not affect myoblast motility, morphogenesisand attachment capacity. N-WASp–deficient myoblasts, however,fail to fuse. Furthermore, our analysis suggests that myoblast fu-sion requires N-WASp activity in both partners of a fusing myo-blast pair. These findings reveal a specific role for N-WASp duringmammalian myogenesis. WASp-family elements appear thereforeto act as universal mediators of the myogenic cell-cell fusion mech-anism underlying formation of functional muscle fibers, in bothvertebrate and invertebrate species.
actin nucleation | myotube formation
Myoblast fusion provides a universal mechanism for forma-tion and growth of multinucleated muscle fibers (1, 2).
However, although the genetic regulatory networks governingskeletal mammalian myogenesis are well characterized (3, 4),many of the cellular mechanisms underlying execution of myo-genic differentiation, including muscle-cell fusion, are poorlyunderstood.Studies of myoblast fusion during Drosophila myogenesis
have put forward a molecular genetic framework, which ascribesmajor significance to the contribution of the actin-based cyto-skeleton, with a particularly prominent role assigned to thebranched actin polymerization machinery centered on theArp2/3 complex (5–7). Nucleation of branched actin poly-merization by Arp2/3 is commonly stimulated by nucleationpromoting factors (NPFs) belonging to the WASp-protein fam-ily. Therefore, the conserved nature of the Arp2/3 complex andits associated NPFs raises the possibility that elements of thismachinery are universal mediators of myoblast fusion. To ad-dress this issue, we examined the consequences of disrupting thefunction of N-WASp, the primary mammalian homolog ofWASp-family proteins, during embryonic myogenesis in mice.Here, we report that myogenesis in mouse embryos is severelyimpaired following disruption of N-WASp function in myogenictissue, throughout the skeletal muscle field. Although the sizeand distribution of the progenitor myoblast population is notaffected, these cells give rise to thin, mononucleated musclefibers. Using primary cell cultures, we show that N-WASp–deficient myoblasts are motile, differentiate properly, and as-sume the morphology of mature myogenic cells, yet fail to fuse.These observations identify a myogenic setting for N-WASpfunction, and suggest an essential, universal involvement forbranched actin nucleation, mediated by WASp-family elements,during the process of myoblast fusion.
ResultsConditional Disruption of Murine N-WASp Results in Abnormal SkeletalMyogenesis. Disruption of the N-WASp gene results in embry-onic lethality at embryonic day E11, characterized by smallbody size and prominent neural tube and cardiac defects (8).To circumvent these phenotypes, which bar proper study ofmyogenesis in the absence of N-WASp function, we made use ofa conditional, loxP-based allele (referred to as N-WASpfl), whichenables tissue-specific disruption of N-WASp (9). Two Cre driverlines, Myf5Cre (10) and MyoDCre (11), were used for this purpose.Both drivers mediate Cre-based recombination at early phases ofthe myogenic program, in all skeletal muscle progenitor cells, andare considered highly effective tools for studying the consequencesof single-gene disruption on skeletal myogenesis (12). Mice inwhich N-WASp was disrupted using N-WASpfl and either of thetwo myogenic Cre lines completed embryogenesis, but died im-mediately after birth. Although the general morphology of theseconditional knockout animals (referred to herein as N-WASpcKO) was normal, external examination, as well as tissuedissection, suggested that skeletal muscle mass was greatly re-duced (Fig. S1). Furthermore, these animals were incapable ofinflating their lungs, a common consequence of impairment tomuscle development in mouse embryos, and the likely cause ofpostnatal lethality. Indeed, immunostaining of histological sectionsrevealed that the musculature of Myf5Cre/N-WASpfl/- embryos atE16.5–E18, visualized with antibodies to the heavy chain of musclemyosin (MHC), is poorly developed (Fig. 1 A–D). The effect isextensive, as practically all skeletal muscle groups (body-wall, limb,respiratory, facial, etc.) are affected in this manner.
Myogenic Program Is Properly Initiated in N-WASpcKO Mice. To elu-cidate the basis for the dramatic effect of disrupting N-WASp onmuscle fiber formation, we examined several key features as-sociated with the onset of skeletal myogenesis in N-WASpcKO
embryos. The complex set of genetic programs that govern fatedetermination during skeletal myogenesis commonly lead toexpression of the key differentiation factor myogenin in allskeletal muscle progenitors (3, 4). Using RNA in situ hybridiza-tion, we determined that the normal expression pattern of myo-genin at E10.5 remains unaltered inN-WASpcKO embryos (Fig. 1 Eand F and Fig. S1). This observation implies both proper initiationof the program underlying skeletal muscle differentiation, as wellas proper myogenic patterning within the somites, which will giverise to body-wall skeletal muscles. Myf5Cre/N-WASpfl/- embryos
Author contributions: Y.G.-C., I.H., K.-B.U., E.T., B.-Z.S., and E.D.S. designed research;Y.G.-C., I.H., and K.-B.U. performed research; S.B.S. contributed new reagents/analytictools; Y.G.-C., B.-Z.S., and E.D.S. analyzed data; and Y.G.-C., B.-Z.S., and E.D.S. wrotethe paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.1To whom correspondence may be addressed. E-mail: [email protected] [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1116065109/-/DCSupplemental.
www.pnas.org/cgi/doi/10.1073/pnas.1116065109 PNAS | July 10, 2012 | vol. 109 | no. 28 | 11211–11216
DEV
ELOPM
ENTA
LBIOLO
GY

further display robust expression of MHC, a second marker ofadvanced myoblast differentiation, in E11.5 wholemount prepa-rations (Fig. 1 G and H). MHC expression at this stage is dis-played by a variety of myoblast populations, consistent with properspecification and differentiation of skeletal muscle progenitors indifferent anatomical locations.Given the well established contribution of Arp2/3-based
branched actin polymerization to cell motility (13), we nextascertained whether lack of N-WASp function interferes withthe motile capacity of skeletal muscle progenitors. To addressthis issue, we generated N-WASpcKO embryos using the N-WASpfl
allele, together with the Pax3Cre driver (14). In such embryos,N-WASp is disrupted in dermomyotome-derived muscle pre-cursors before their differentiation, allowing to assess theircapacity to migrate considerable distances away from theirsomitic origin (15). Pax3Cre/N-WASpfl/- embryos do not developpast E12, probably due to expression of the driver within theneural crest and heart progenitors, but survive long enough toallow for analysis of early myogenesis in the absence of N-WASpfunction. Importantly, MHC-expressing cells in the limb areproperly patterned within these embryos, demonstrating that theprogenitors migrate away from the somites and populate limbbuds and other sites of differentiation and muscle fiber formation(Fig. S1). Furthermore, the expression pattern of Pax7, an earlymyoblast fate marker, revealed that Pax3Cre/N-WASpfl/- embryospossess a normally sized field of limb bud myoblasts (Fig. S1).
Taken together, these observations imply that disruption ofmurine N-WASp does not interfere with the specification, migra-tion capacity, and differentiation program of skeletal myoblastsduring embryogenesis.
Skeletal Muscle Fibers of N-WASpcKO Mice Are Mononucleated. Thedemonstration that the initial phases of skeletal myogenesis donot require N-WASp led us to examine a “timeline” of histologicalsections of maturing N-WASpcKO embryos, stained for informativemarkers, to identify the stage at which phenotypic defects firstarise (Fig. 2). Mixed fields of Pax7-positive precursors and MHC-expressing cells at E10.5, demonstrate proper and timely onset ofthe early stages of myogenesis in the somites and limbs ofN-WASpcKO embryos (Fig. 2 A, B, E, F, I, and J). As muscledevelopment progresses, mild phenotypic abnormalities canbe detected by E14.5, when muscle fibers in N-WASpcKO em-bryos appear shorter and thinner than those in age-matched wild-type embryos (Fig. 2 C, G, and K). These phenotypes becomehighly pronounced as the embryos mature further (Fig. 2 D, H,and L). Significantly, N-WASpcKO embryonic muscle fibers remainmononucleated, suggesting that their underdeveloped natureresults from a failure to incorporate myoblasts via fusion intomaturing fibers.Fusion arrest in N-WASpcKO embryos is associated with en-
hanced activation of Caspase-3 (Fig. S2 A–C), implying increasedapoptosis within the myogenic field. The myogenic differentiationprogram is not perturbed, however, evidenced by the unalteredratio between myoblasts expressing Pax7 and those expressing themore advanced differentiation marker MyoD (Fig. S2 D–J).
N-WASp–Depleted Primary Cultures Differentiate Properly but Fail toFuse. We next turned to primary cell cultures, to analyze themyogenic requirement for N-WASp at cellular resolution. To-ward this end, satellite cell cultures were prepared from isolatedmuscle fibers of N-WASpfl/fl mice (16). Disruption of N-WASp inthese cultures was achieved following infection with the adeno-virus vector Ad-Cre-eGFP, containing both the Cre recombinaseand nuclear eGFP. Depletion of N-WASp protein was verified byWestern blot analysis (Fig. S3). Differentiation of the adenovirus-
Fig. 1. Skeletal muscle mass of N-WASpcKO embryos is severely reduced,whereas myogenic differentiation is unaffected. (A–D) Histological sectionsof wild-type (WT) embryos (A and C) and Myf5Cre/N-WASpfl/- embryos (B andD). Muscle fibers are visualized with anti-MHC (red) and nuclei with DAPI(blue). (A and B) Matching sections revealing body wall muscles (BM) andlimb muscles (LM) at E16.5. (C and D) Matching sections revealing facialelements at E16.5. EOM, extra ocular muscles. (E and F) RNA in situhybridization of a myogenin DNA probe to E10.5 WT (E) and Myf5Cre/N-WASpfl/- (F) embryos reveals an identical pattern of somitic expression.(G and H) Immuno-localization of MHC in E11.5 WT (G) and Myf5Cre/N-WASpfl/- (H) embryos reveals similar patterns of differentiated musclegroups. Somitic (S), forelimb (FL), hindlimb (HL), and temporalis (T) musclesare indicated. (Scale bars: 500 μm in A, 100 μm in C, and 2 mm in F and H.)
Fig. 2. Muscle fibers of N-WASpcKO embryos are properly initiated, butremain mononucleated. (A-D) Myogenesis in WT embryos. (A–D) Somitic(A) and limb (B–D) myogenic cell fields at E10.5 (A and B), E14.5 (C), andE16.5 (D). Pax7 (green) marks myoblast nuclei, MHC (red) marks fibers and allnuclei are visualized by DAPI (blue). (E–L) Corresponding panels showing theprogress of myogenesis inMyf5Cre/N-WASpfl/- embryos (E–H) and inMyoDCre/N-WASpfl/- embryos (I–L) of similar ages. Arrows point to nuclei within rep-resentative multinucleated wild-type fibers in C and D, and to single nucleiwithin mutant fibers in G, H, K, and L. Representative mononucleated fibersin E16.5 N-WASpcKO embryos are outlined in H and L. (Scale bar: 50 μm in A.)
11212 | www.pnas.org/cgi/doi/10.1073/pnas.1116065109 Gruenbaum-Cohen et al.

infected cultures was induced via serum starvation. N-WASpfl/fl–derived cultures, separately infected with a vector harboringeGFP alone (Ad-eGFP), harbor an intact N-WASp locus whileexpressing cytoplasmic eGFP, and therefore served as ourprimary control.Monitoring culture differentiation by visualization of cell
morphologies and expression of informative markers, revealeda striking difference between the Ad-eGFP– and Ad-Cre-eGFP–infected cultures of N-WASpfl/fl satellite cells (Fig. 3). Althoughthe control cells readily formed dense arrays of multinucleatedmyofibers by this protocol (Fig. 3 A–C and E–G), the N-WASp–depleted cultures were primarily composed of individual,mononucleated cells (Fig. 3 D and H). Quantification demon-strated that 67% of control myotubes harbored three or morenuclei, whereas 73% of N-WASp–depleted cells contained onlya single nucleus, with nearly the entire remainder composed ofbinucleated cells (Fig. 3I).Depletion of N-WASp from the satellite cell cultures was
associated with enhanced apoptosis, matching our reportedobservations in N-WASpcKO embryos. To ascertain that the blockin myoblast fusion was not a consequence of the reduced celldensity resulting from cell death, we reassessed the fusion ca-pacity of Ad-eGFP-Cre–infected cultures of N-WASpfl/fl satellitecells that were plated at a 2.5-fold higher confluency than thestandard assay. Myoblast fusion upon serum starvation is arrestedto a similar extent under these conditions (74% mononucleatedcells). In a corresponding experiment, control (Ad-eGFP–infected)cultures were plated at a fourfold lower confluency than thestandard assay. Myoblast fusion upon serum starvation declines
somewhat, but remains robust under these conditions (43% ofmyotubes harbor three or more nuclei).Although strongly deficient in their capacity to generate
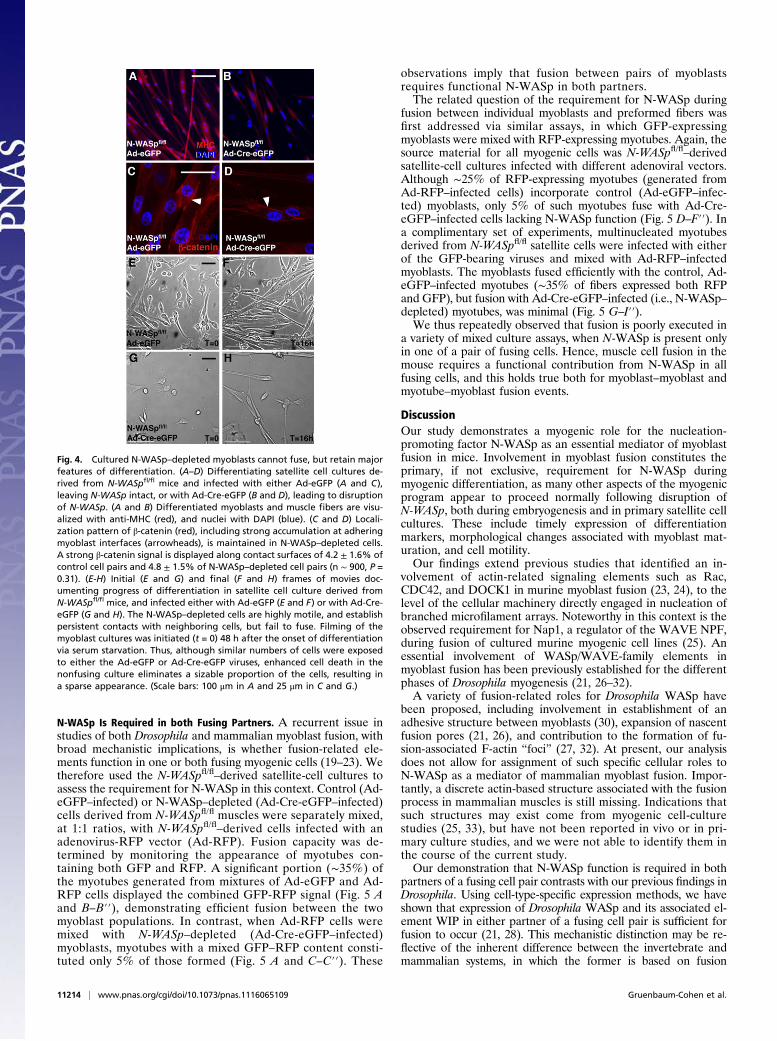
multinucleated myotubes, N-WASp–depleted cells displayed avariety of features associated with proper myogenic differentiationbefore onset of fusion. Thus, these cells adopted the elongated,spindle-like morphology characteristic of differentiated myocytes(Fig. 4 A and B), and expressed robust, normal levels of differ-entiation markers such as MyoD, Myogenin, and MHC (Fig. S3).Importantly, interfaces between neighboring pairs of N-WASp–depleted cells displayed pronounced recruitment of β-catenin,a marker and component of productive myogenic cell attach-ments (17, 18), suggesting that cell–cell contact was properlyinitiated (Fig. 4 C and D).Time-lapse imaging of live satellite cell cultures was used to
monitor their dynamic behavior following adenovirus infectionand serum starvation (Fig. 4 E–H and Movies S1 and S2). As canbe readily ascertained from this analysis, N-WASp–depletedcultures share many features with the Ad-eGFP–infected con-trols, but fail to generate myotubes. Thus, the N-WASp–de-pleted cells are highly motile, moving at an average speed of1.67 ± 0.23 μm/min, somewhat faster than the controls (1.14 ±0.35 μm/min, n = 14). Furthermore, the N-WASp–depleted cellsundergo a variety of cell-shape changes, and are clearly capableof prolonged cell–cell contact (up to 45 min). However, theseassociations consistently fail to result in fusion and myotubeformation. Taken together, the satellite-cell culture data stronglyimply that the failure of N-WASp–depleted cells to form mul-tinucleated myotubes results from a specific arrest in cell fusion,whereas all other aspects of myogenesis are unaffected.
Fig. 3. Disruption of N-WASp inhibits myoblast fusion insatellite cell cultures. (A–H) Primary cultures derived fromsatellite cells, isolated from EDL muscles of 4- to 6-wk-oldmice, following infection with various adenovirus vectorsand 48 h of serum-starvation-induced differentiation. Mus-cle fibers are visualized with anti-MHC (red) and nuclei withDAPI (blue). GFP (green) is an indicator of viral infection.(A and B) Cultures derived from WT (ICR) mice and infectedwith Ad-eGFP (A) or Ad-Cre-eGFP (B) viruses. (C and D)Cultures derived from N-WASpfl/fl mice and infected withAd-eGFP (C) or Ad-Cre-eGFP (D) viruses. (E–H) Magnifiedviews of cultures matching those shown in A–D. Controlcultures (A–C and E–G) formed multinucleated fibers,whereas the fibers in cultures in which N-WASp was dis-rupted (D and H) remained mononucleated. (I) Graphicpresentation of the fusion index assayed following 48 h indifferentiation medium (n = number of nuclei within in-dividual fibers; ∼800 nuclei were counted and classified foreach of the four infection protocols). (Scale bars: 100 μm inA and 50 μm in E.)
Gruenbaum-Cohen et al. PNAS | July 10, 2012 | vol. 109 | no. 28 | 11213
DEV
ELOPM
ENTA
LBIOLO
GY
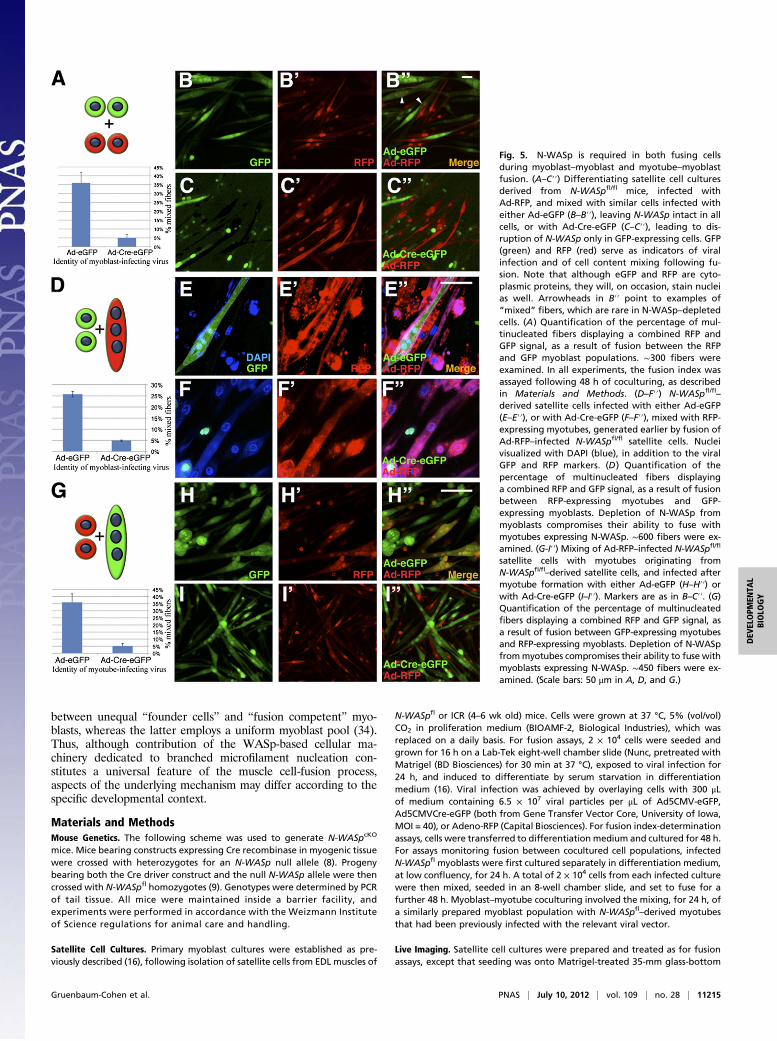
N-WASp Is Required in both Fusing Partners. A recurrent issue instudies of both Drosophila and mammalian myoblast fusion, withbroad mechanistic implications, is whether fusion-related ele-ments function in one or both fusing myogenic cells (19–23). Wetherefore used the N-WASpfl/fl–derived satellite-cell cultures toassess the requirement for N-WASp in this context. Control (Ad-eGFP–infected) or N-WASp–depleted (Ad-Cre-eGFP–infected)cells derived from N-WASpfl/fl muscles were separately mixed,at 1:1 ratios, with N-WASpfl/fl–derived cells infected with anadenovirus-RFP vector (Ad-RFP). Fusion capacity was de-termined by monitoring the appearance of myotubes con-taining both GFP and RFP. A significant portion (∼35%) ofthe myotubes generated from mixtures of Ad-eGFP and Ad-RFP cells displayed the combined GFP-RFP signal (Fig. 5 Aand B–B′′), demonstrating efficient fusion between the twomyoblast populations. In contrast, when Ad-RFP cells weremixed with N-WASp–depleted (Ad-Cre-eGFP–infected)myoblasts, myotubes with a mixed GFP–RFP content consti-tuted only 5% of those formed (Fig. 5 A and C–C′′). These
observations imply that fusion between pairs of myoblastsrequires functional N-WASp in both partners.The related question of the requirement for N-WASp during
fusion between individual myoblasts and preformed fibers wasfirst addressed via similar assays, in which GFP-expressingmyoblasts were mixed with RFP-expressing myotubes. Again, thesource material for all myogenic cells was N-WASpfl/fl–derivedsatellite-cell cultures infected with different adenoviral vectors.Although ∼25% of RFP-expressing myotubes (generated fromAd-RFP–infected cells) incorporate control (Ad-eGFP–infec-ted) myoblasts, only 5% of such myotubes fuse with Ad-Cre-eGFP–infected cells lacking N-WASp function (Fig. 5 D–F′′). Ina complimentary set of experiments, multinucleated myotubesderived from N-WASpfl/fl satellite cells were infected with eitherof the GFP-bearing viruses and mixed with Ad-RFP–infectedmyoblasts. The myoblasts fused efficiently with the control, Ad-eGFP–infected myotubes (∼35% of fibers expressed both RFPand GFP), but fusion with Ad-Cre-eGFP–infected (i.e., N-WASp–depleted) myotubes, was minimal (Fig. 5 G–I′′).We thus repeatedly observed that fusion is poorly executed in
a variety of mixed culture assays, when N-WASp is present onlyin one of a pair of fusing cells. Hence, muscle cell fusion in themouse requires a functional contribution from N-WASp in allfusing cells, and this holds true both for myoblast–myoblast andmyotube–myoblast fusion events.
DiscussionOur study demonstrates a myogenic role for the nucleation-promoting factor N-WASp as an essential mediator of myoblastfusion in mice. Involvement in myoblast fusion constitutes theprimary, if not exclusive, requirement for N-WASp duringmyogenic differentiation, as many other aspects of the myogenicprogram appear to proceed normally following disruption ofN-WASp, both during embryogenesis and in primary satellite cellcultures. These include timely expression of differentiationmarkers, morphological changes associated with myoblast mat-uration, and cell motility.Our findings extend previous studies that identified an in-
volvement of actin-related signaling elements such as Rac,CDC42, and DOCK1 in murine myoblast fusion (23, 24), to thelevel of the cellular machinery directly engaged in nucleation ofbranched microfilament arrays. Noteworthy in this context is theobserved requirement for Nap1, a regulator of the WAVE NPF,during fusion of cultured murine myogenic cell lines (25). Anessential involvement of WASp/WAVE-family elements inmyoblast fusion has been previously established for the differentphases of Drosophila myogenesis (21, 26–32).A variety of fusion-related roles for Drosophila WASp have
been proposed, including involvement in establishment of anadhesive structure between myoblasts (30), expansion of nascentfusion pores (21, 26), and contribution to the formation of fu-sion-associated F-actin “foci” (27, 32). At present, our analysisdoes not allow for assignment of such specific cellular roles toN-WASp as a mediator of mammalian myoblast fusion. Impor-tantly, a discrete actin-based structure associated with the fusionprocess in mammalian muscles is still missing. Indications thatsuch structures may exist come from myogenic cell-culturestudies (25, 33), but have not been reported in vivo or in pri-mary culture studies, and we were not able to identify them inthe course of the current study.Our demonstration that N-WASp function is required in both
partners of a fusing cell pair contrasts with our previous findings inDrosophila. Using cell-type-specific expression methods, we haveshown that expression of Drosophila WASp and its associated el-ement WIP in either partner of a fusing cell pair is sufficient forfusion to occur (21, 28). This mechanistic distinction may be re-flective of the inherent difference between the invertebrate andmammalian systems, in which the former is based on fusion
Fig. 4. Cultured N-WASp–depleted myoblasts cannot fuse, but retain majorfeatures of differentiation. (A–D) Differentiating satellite cell cultures de-rived from N-WASpfl/fl mice and infected with either Ad-eGFP (A and C),leaving N-WASp intact, or with Ad-Cre-eGFP (B and D), leading to disruptionof N-WASp. (A and B) Differentiated myoblasts and muscle fibers are visu-alized with anti-MHC (red), and nuclei with DAPI (blue). (C and D) Locali-zation pattern of β-catenin (red), including strong accumulation at adheringmyoblast interfaces (arrowheads), is maintained in N-WASp–depleted cells.A strong β-catenin signal is displayed along contact surfaces of 4.2 ± 1.6% ofcontrol cell pairs and 4.8 ± 1.5% of N-WASp–depleted cell pairs (n ∼ 900, P =0.31). (E-H) Initial (E and G) and final (F and H) frames of movies doc-umenting progress of differentiation in satellite cell culture derived fromN-WASpfl/fl mice, and infected either with Ad-eGFP (E and F) or with Ad-Cre-eGFP (G and H). The N-WASp–depleted cells are highly motile, and establishpersistent contacts with neighboring cells, but fail to fuse. Filming of themyoblast cultures was initiated (t = 0) 48 h after the onset of differentiationvia serum starvation. Thus, although similar numbers of cells were exposedto either the Ad-eGFP or Ad-Cre-eGFP viruses, enhanced cell death in thenonfusing culture eliminates a sizable proportion of the cells, resulting ina sparse appearance. (Scale bars: 100 μm in A and 25 μm in C and G.)
11214 | www.pnas.org/cgi/doi/10.1073/pnas.1116065109 Gruenbaum-Cohen et al.
between unequal “founder cells” and “fusion competent” myo-blasts, whereas the latter employs a uniform myoblast pool (34).Thus, although contribution of the WASp-based cellular ma-chinery dedicated to branched microfilament nucleation con-stitutes a universal feature of the muscle cell-fusion process,aspects of the underlying mechanism may differ according to thespecific developmental context.
Materials and MethodsMouse Genetics. The following scheme was used to generate N-WASpcKO
mice. Mice bearing constructs expressing Cre recombinase in myogenic tissuewere crossed with heterozygotes for an N-WASp null allele (8). Progenybearing both the Cre driver construct and the null N-WASp allele were thencrossed with N-WASpfl homozygotes (9). Genotypes were determined by PCRof tail tissue. All mice were maintained inside a barrier facility, andexperiments were performed in accordance with the Weizmann Instituteof Science regulations for animal care and handling.
Satellite Cell Cultures. Primary myoblast cultures were established as pre-viously described (16), following isolation of satellite cells from EDL muscles of
N-WASpfl or ICR (4–6 wk old) mice. Cells were grown at 37 °C, 5% (vol/vol)CO2 in proliferation medium (BIOAMF-2, Biological Industries), which wasreplaced on a daily basis. For fusion assays, 2 × 104 cells were seeded andgrown for 16 h on a Lab-Tek eight-well chamber slide (Nunc, pretreated withMatrigel (BD Biosciences) for 30 min at 37 °C), exposed to viral infection for24 h, and induced to differentiate by serum starvation in differentiationmedium (16). Viral infection was achieved by overlaying cells with 300 μLof medium containing 6.5 × 107 viral particles per μL of Ad5CMV-eGFP,Ad5CMVCre-eGFP (both from Gene Transfer Vector Core, University of Iowa,MOI = 40), or Adeno-RFP (Capital Biosciences). For fusion index-determinationassays, cells were transferred to differentiation medium and cultured for 48 h.For assays monitoring fusion between cocultured cell populations, infectedN-WASpfl myoblasts were first cultured separately in differentiation medium,at low confluency, for 24 h. A total of 2 × 104 cells from each infected culturewere then mixed, seeded in an 8-well chamber slide, and set to fuse for afurther 48 h. Myoblast–myotube coculturing involved the mixing, for 24 h, ofa similarly prepared myoblast population with N-WASpfl–derived myotubesthat had been previously infected with the relevant viral vector.
Live Imaging. Satellite cell cultures were prepared and treated as for fusionassays, except that seeding was onto Matrigel-treated 35-mm glass-bottom
Fig. 5. N-WASp is required in both fusing cellsduring myoblast–myoblast and myotube–myoblastfusion. (A–C′′) Differentiating satellite cell culturesderived from N-WASpfl/fl mice, infected withAd-RFP, and mixed with similar cells infected witheither Ad-eGFP (B–B′′), leaving N-WASp intact in allcells, or with Ad-Cre-eGFP (C–C′′), leading to dis-ruption of N-WASp only in GFP-expressing cells. GFP(green) and RFP (red) serve as indicators of viralinfection and of cell content mixing following fu-sion. Note that although eGFP and RFP are cyto-plasmic proteins, they will, on occasion, stain nucleias well. Arrowheads in B′′ point to examples of“mixed” fibers, which are rare in N-WASp–depletedcells. (A) Quantification of the percentage of mul-tinucleated fibers displaying a combined RFP andGFP signal, as a result of fusion between the RFPand GFP myoblast populations. ∼300 fibers wereexamined. In all experiments, the fusion index wasassayed following 48 h of coculturing, as describedin Materials and Methods. (D–F′′) N-WASpfl/fl
–
derived satellite cells infected with either Ad-eGFP(E–E′′), or with Ad-Cre-eGFP (F–F′′), mixed with RFP-expressing myotubes, generated earlier by fusion ofAd-RFP–infected N-WASpfl/fl satellite cells. Nucleivisualized with DAPI (blue), in addition to the viralGFP and RFP markers. (D) Quantification of thepercentage of multinucleated fibers displayinga combined RFP and GFP signal, as a result of fusionbetween RFP-expressing myotubes and GFP-expressing myoblasts. Depletion of N-WASp frommyoblasts compromises their ability to fuse withmyotubes expressing N-WASp. ∼600 fibers were ex-amined. (G-I′′) Mixing of Ad-RFP–infected N-WASpfl/fl
satellite cells with myotubes originating fromN-WASpfl/fl–derived satellite cells, and infected aftermyotube formation with either Ad-eGFP (H–H′′) orwith Ad-Cre-eGFP (I–I′′). Markers are as in B–C′′. (G)Quantification of the percentage of multinucleatedfibers displaying a combined RFP and GFP signal, asa result of fusion between GFP-expressing myotubesand RFP-expressing myoblasts. Depletion of N-WASpfrommyotubes compromises their ability to fuse withmyoblasts expressing N-WASp. ∼450 fibers were ex-amined. (Scale bars: 50 μm in A, D, and G.)
Gruenbaum-Cohen et al. PNAS | July 10, 2012 | vol. 109 | no. 28 | 11215
DEV
ELOPM
ENTA
LBIOLO
GY

dishes (MatTek). Time-lapse movies were obtained using an Applied PrecisionDeltaVision imaging system. Z stacks of five images were acquired at regularintervals and compiled to form maximum intensity point projection images.For velocity measurements, the temporal location of cells was trackedmanually using a Matlab GUI. Cell velocity was determined from the averagedistance made by the cell in 1 min.
Immunohistochemistry. Immunolabeling protocols of mouse tissue paraffinsections and of satellite cell cultures were as described (35, 36). Primaryantibodies used included mouse anti-MHC (MH-20, Developmental StudiesHybridoma Bank, 1:5), mouse anti-Pax7 (Developmental Studies HybridomaBank, 1:5–1:10), rabbit anti-MyoD (sc-304, Santa Cruz Biotechnology, 1:100),anti-cleaved Caspase 3 (Cell Signaling, 1:50), and mouse anti-β-catenin (BDBiosciences, 1:50). Cy2-, Cy3-, or Cy5-conjugated secondary antibodies (JacksonImmunoResearch) were used at a dilution of 1:200–1:500. Images were ac-quired using a Zeiss LSM710 confocal microscopy system. Immuno-detection ofMHC on embryo whole mounts was performed as described (37) using alkalinephosphatase-conjugated anti-MHC (clone MY32, Sigma-Aldrich, 1:400) andNBT/BCIP substrate (Roche Applied Science).
In Situ Hybridization.Whole-mount in situ hybridization was performed usingdigoxigenin-labeled antisense riboprobes synthesized from cDNA as de-scribed (38). Images were obtained using a Leica MZ16FA stereomicroscopeattached to a digital camera (DC300F, Leica Microsystems).
Western Blotting. Protein extracts were obtained following collection andsonication of cultured satellite cells in RIPA buffer (10mM Tris/150 mMNaCl/5mM EDTA/1% Triton X-100/0.1% SDS/1% Sodium deoxycholate, and freshlyadded protease inhibitors). Western blotting protocols were as described (39).Primary antibodies used included mouse anti-MyoD (sc-32758), mouse anti-Myogenin (sc-12732), rabbit anti N-WASp (sc-H-100), and rabbit anti-Emerin(sc-15378), all from Santa Cruz Biotechnology Monoclonal, as well as mouseanti-MHC n2.261 (Developmental Studies Hybridoma Bank). Secondary anti-bodies used were either anti-rabbit HRP or anti-mouse HRP (Jackson Immu-noResearch). Chemiluminescent detection was performed using EZ-ECL(Biological Industries), according to the manufacturer’s directions.
ACKNOWLEDGMENTS. We thank Paul Knopp and Peter Zammit (King’s Col-lege, London) for providing instruction and sharing their expertise on genera-tion and handling of satellite cell cultures. We thank our colleagues Ari Elson,David Goldhamer, Yoram Groner, Eran Hornstein, Ronen Schweitzer, and EliZelzer for advice, reagents, and use of laboratory facilities; Margaret Bucking-ham for insightful discussions; Sagi Levy for instruction and help in quantifyingmyoblast motility; R’ada Massarwa, Ariel Rinon, and Natti Weinblum for theircontributions to experiments performed in the course of this study; and allmembers of the B.-Z.S. laboratory for their help and support. This work wassupported by research grants from the Israel Science Foundation (ISF) and theMuscular Dystrophy Association (to B-Z.S. and E.D.S.) and a MYORES travelgrant (to Y.G-C.). K-B.U. was supported by ISF Legacy Grant 1875/08. B-Z.S. isan incumbent of the Hilda and Cecil Lewis Chair in Molecular Genetics.
1. Rochlin K, Yu S, Roy S, Baylies MK (2010) Myoblast fusion: When it takes more tomake one. Dev Biol 341:66–83.
2. Simionescu A, Pavlath GK (2011) Molecular mechanisms of myoblast fusion acrossspecies. Adv Exp Med Biol 713:113–135.
3. Bismuth K, Relaix F (2010) Genetic regulation of skeletal muscle development. ExpCell Res 316:3081–3086.
4. Buckingham M (2006) Myogenic progenitor cells and skeletal myogenesis in verte-brates. Curr Opin Genet Dev 16:525–532.
5. Haralalka S, Abmayr SM (2010) Myoblast fusion in Drosophila. Exp Cell Res 316:3007–3013.
6. Onel SF, Renkawitz-Pohl R (2009) FuRMAS: Triggering myoblast fusion in Drosophila.Dev Dyn 238:1513–1525.
7. Richardson BE, Nowak SJ, Baylies MK (2008) Myoblast fusion in fly and vertebrates:New genes, new processes and new perspectives. Traffic 9:1050–1059.
8. Snapper SB, et al. (2001) N-WASP deficiency reveals distinct pathways for cell surfaceprojections and microbial actin-based motility. Nat Cell Biol 3:897–904.
9. Cotta-de-Almeida V, et al. (2007) Wiskott Aldrich syndrome protein (WASP) andN-WASP are critical for T cell development. Proc Natl Acad Sci USA 104:15424–15429.
10. Tallquist MD, Weismann KE, Hellström M, Soriano P (2000) Early myotome specifi-cation regulates PDGFA expression and axial skeleton development. Development127:5059–5070.
11. Chen JC, Mortimer J, Marley J, Goldhamer DJ (2005) MyoD-cre transgenic mice:a model for conditional mutagenesis and lineage tracing of skeletal muscle. Genesis41:116–121.
12. Wamhoff BR, Sinha S, Owens GK (2007) Conditional mouse models to study de-velopmental and pathophysiological gene function in muscle. Handb Exp Pharmacol178:441–468.
13. Millard TH, Sharp SJ, Machesky LM (2004) Signalling to actin assembly via the WASP(Wiskott-Aldrich syndrome protein)-family proteins and the Arp2/3 complex. BiochemJ 380:1–17.
14. Engleka KA, et al. (2005) Insertion of Cre into the Pax3 locus creates a new allele ofSplotch and identifies unexpected Pax3 derivatives. Dev Biol 280:396–406.
15. Buckingham M, et al. (2003) The formation of skeletal muscle: From somite to limb.J Anat 202:59–68.
16. Beauchamp JR, et al. (2000) Expression of CD34 and Myf5 defines the majority ofquiescent adult skeletal muscle satellite cells. J Cell Biol 151:1221–1234.
17. Radice GL, et al. (1997) Developmental defects in mouse embryos lacking N-cadherin.Dev Biol 181:64–78.
18. Charrasse S, Meriane M, Comunale F, Blangy A, Gauthier-Rouvière C (2002) N-cadherin-dependent cell-cell contact regulates Rho GTPases and beta-catenin localization inmouse C2C12 myoblasts. J Cell Biol 158:953–965.
19. Haralalka S, et al. (2011) Asymmetric Mbc, active Rac1 and F-actin foci in the fusion-competent myoblasts during myoblast fusion in Drosophila. Development 138:1551–1562.
20. Jin P, et al. (2011) Competition between Blown fuse and WASP for WIP bindingregulates the dynamics of WASP-dependent actin polymerization in vivo. Dev Cell 20:623–638.
21. Massarwa R, Carmon S, Shilo BZ, Schejter ED (2007) WIP/WASp-based actin-poly-merization machinery is essential for myoblast fusion in Drosophila. Dev Cell 12:557–569.
22. Sohn RL, et al. (2009) A role for nephrin, a renal protein, in vertebrate skeletal musclecell fusion. Proc Natl Acad Sci USA 106:9274–9279.
23. Vasyutina E, Martarelli B, Brakebusch C, Wende H, Birchmeier C (2009) The smallG-proteins Rac1 and Cdc42 are essential for myoblast fusion in the mouse. Proc NatlAcad Sci USA 106:8935–8940.
24. Laurin M, et al. (2008) The atypical Rac activator Dock180 (Dock1) regulates myoblastfusion in vivo. Proc Natl Acad Sci USA 105:15446–15451.
25. Nowak SJ, Nahirney PC, Hadjantonakis AK, Baylies MK (2009) Nap1-mediatedactin remodeling is essential for mammalian myoblast fusion. J Cell Sci 122:3282–3293.
26. Gildor B, Massarwa R, Shilo BZ, Schejter ED (2009) The SCAR and WASp nucleation-promoting factors act sequentially to mediate Drosophila myoblast fusion. EMBO Rep10:1043–1050.
27. Kim S, et al. (2007) A critical function for the actin cytoskeleton in targeted exocytosisof prefusion vesicles during myoblast fusion. Dev Cell 12:571–586.
28. Mukherjee P, Gildor B, Shilo BZ, VijayRaghavan K, Schejter ED (2011) The actin nu-cleator WASp is required for myoblast fusion during adult Drosophila myogenesis.Development 138:2347–2357.
29. Richardson BE, Beckett K, Nowak SJ, Baylies MK (2007) SCAR/WAVE and Arp2/3 arecrucial for cytoskeletal remodeling at the site of myoblast fusion. Development 134:4357–4367.
30. Schäfer G, et al. (2007) The Wiskott-Aldrich syndrome protein (WASP) is essential formyoblast fusion in Drosophila. Dev Biol 304:664–674.
31. Schröter RH, et al. (2004) kette and blown fuse interact genetically during the secondfusion step of myogenesis in Drosophila. Development 131:4501–4509.
32. Sens KL, et al. (2010) An invasive podosome-like structure promotes fusion poreformation during myoblast fusion. J Cell Biol 191:1013–1027.
33. Duan R, Gallagher PJ (2009) Dependence of myoblast fusion on a cortical actin walland nonmuscle myosin IIA. Dev Biol 325:374–385.
34. Abmayr SM, Pavlath GK (2012) Myoblast fusion: Lessons from flies and mice. De-velopment 139:641–656.
35. Harel I, et al. (2009) Distinct origins and genetic programs of head muscle satellitecells. Dev Cell 16:822–832.
36. Ono Y, et al. (2011) BMP signalling permits population expansion by preventingpremature myogenic differentiation in muscle satellite cells. Cell Death Differ 18:222–234.
37. DeLaurier A, Schweitzer R, Logan M (2006) Pitx1 determines the morphology ofmuscle, tendon, and bones of the hindlimb. Dev Biol 299:22–34.
38. Tirosh-Finkel L, Elhanany H, Rinon A, Tzahor E (2006) Mesoderm progenitor cells ofcommon origin contribute to the head musculature and the cardiac outflow tract.Development 133:1943–1953.
39. Aziz-Aloya RB, et al. (1998) Expression of AML1-d, a short human AML1 isoform, inembryonic stem cells suppresses in vivo tumor growth and differentiation. CellDeath Differ 5:765–773.
11216 | www.pnas.org/cgi/doi/10.1073/pnas.1116065109 Gruenbaum-Cohen et al.