the preimplantation conceptus of the mouseat
TRANSCRIPT

J. Anat. (1967), 102, 1, pp. 1-11With 11 figuresPrinted in Great Britain
The preimplantation conceptus of the mouse at90 hours post coitum
D. M. POTTS
Department of Anatomy, University of Cambridge
AND I. B. WILSON
Department of Zoology, University College of North Wales, Bangor
INTRODUCTION
The morphology of the conceptus in normal pregnancy has been studied in about100 of the several thousand species of eutherian mammals. Descriptions of develop-ment in the mouse are amongst the earliest available (Salenka, 1883; Duval, 1891;Burckhard, 1901; Sobotta, 1903). Recent reviews include Amoroso (1952), Boyd &Hamilton (1952) and Blandau (1961).
In mice from a randomly bred hybrid colony, fertilized eggs reach the uterusabout 72 h after copulation, in the morula or early blastocyst stage. Implantationstarts approximately 24-30 h later. In this paper a description is given of the con-ceptus and surrounding uterine tissue at 90 h after copulation, at the stage when theblastocysts have become spaced out along the uterus and located at their anti-mesometrial implantation sites, immediately preceding loss of the zona pellucida.
METHODS
Hybrid mice, from an original cross between A2G and C57BL, were bred atrandom under artificial conditions of 16 h light and 8 h dark. Mature females wereplaced with males for 2 h towards the end of the dark period and those with copula-tion plugs were killed at selected intervals after mating. The middle of the 2 h periodtaken as zero.The excised uteri were fixed in the extended state in 1% osmic acid at 4°C for
2 h. Preimplantation blastocysts were located in the uterus by a double sectioningtechnique (Potts, 1966a). Araldite sections were also taken at 1 ,ctm and stainedwith methylene blue and toluidine blue for light microscopy.For comparative purposes some uteri were fixed in Clarke's fluid (3/1 absolute
alcohol/glacial acetic) and some by the process of freeze-substitution (Feder &Sidman, 1958) using 1 % mercuric chloride in absolute alcohol. These were embeddedin ester wax, sectioned at 3-5 ,tm and stained with either toluidine blue or Heiden-hain's haematoxylin, for conventional light microscopy. Some sections stained withtoluidine blue were photographed then the cover-slip and mountant were strippedoff. These sections were incubated, after destaining, in a solution of RNAase(1 0 mg/cc at 37 °C for 1 h) and then restained with toluidine blue.
Material from a series of twenty-one uteri obtained between 88-92 h p.c. (postcoitum) was fixed by freeze-substitution. Six animals from this series killed at 90 h p.c.
I Anat. 102
D. M. POTTS AND I. B. WILSON
,t . W :FT,,,.:i, s w. :-
Ak
*:: s l ~~~~~~~4 ::::: .*.-~ ~~~~ik
."A*I'll ;,,,:;,.,"Cl-i-i
:-..;t
I
5
2

Preimplantation conceptus of the mouseprovided one uterine horn for fixation in Clarke's fluid. The uteri from five animalskilled 90 h p.c. were fixed for electron microscopy.
OBSERVATIONS
At 90 h p.c. the mouse blastocyst has reached its implantation site at the anti-mesometrial boundary of the uterine lumen. It is a sphere about 90 ,tm in dia-meter with an intact zona pellucida, the cells of the trophoblast and inner cellmass are of a relatively uniform size and there is no sign as yet of a trophoblastic giantcell transformation (not for another 6 h or more). The uterine lumen is occludedand the blastocyst is tightly enclosed by epithelium (though the lumen may openduring fixation and preparation of material for microscopy).
Light microscopyIn routine cytological fixatives, all of which (except formalin) have a fairly acid
pH, the zona pellucida is broken down and appears as diffuse material sticking tothe trophoblast and the uterine epithelium (Fig. 1). Fixation by freeze-substitutionpreserves the zona as a highly vacuolated, tenuous, but complete covering aroundthe blastocyst, with an irregular thickness of about 6 ysm (Fig. 2). In the series oftwenty-one animals killed from 88-92 h p.c. the vacuolation and dissolution of thezona was progressive so that it appeared to disintegrate in situ rather than ruptureand slip off the blastocyst.
Orientation, with mesometrial positioning of the inner cell mass, is not determineduntil after the zona disintegrates (Kirby, Potts & Wilson, 1967).
In the inner cell mass as many as six, but usually fewer, basiphilic bodies can bedistinguished (Fig. 1), and they stain deeply with toluidine blue (Fig. 3). If thesections are stripped, treated with RNAase and restained, the staining of theseobjects is almost completely abolished (Fig. 4).The uterine epithelium, at this stage, appears intact and cellular, though a little
flattened immediately around the blastocyst. There is slight oedema in the stromaaround the implantation chamber.
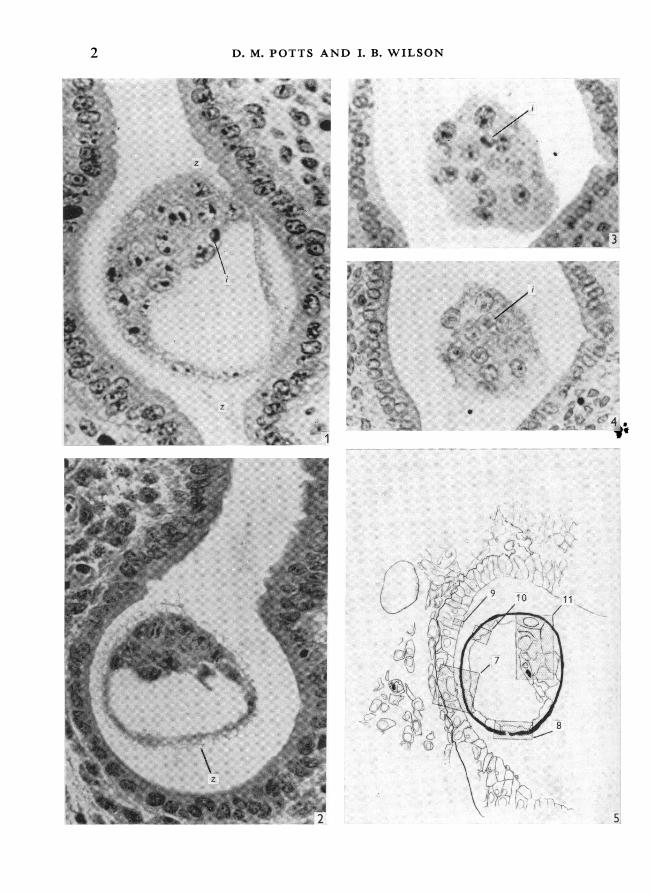
Figures 1-4 are transverse sections of uterus and blastocyst; magnification approx. x 450.The mesometrium is towards the top of the page. The space around the blastocyst is an artefact.Fig. 1. Fixation in Clarke; stained in Heidenhain's iron haematoxylin. The zona pellucida (z) isdiffuse. There is a darkly stained inclusion (i) within the inner cell mass.
Fig. 2. Fixation by freeze-substitution; stained in toluidine blue. The zona is vacuolate andfilamentous but forms a complete covering.Fig. 3. Fixation in Clarke; stained in toluidine blue. The inclusion stains intensely. The sectionpasses obliquely through the inner cell mass of a blastocyst lying 'horizontally' in the uterus.Fig. 4. Same section as Fig. 3 after stripping, treatment with RNAase and restaining withtoluidine blue. The inclusion stains only lightly.Fig. 5. Longitudinal section of uterus and blastocyst. A composite diagram of blastocystand surrounding uterine epithelium prepared from serial sections. Cell and nuclear outlines takenfrom electron micrographs, tissue outlines from light micrographs. Areas shown in subsequentfigures shaded.
I-2
3
D. M. POTTS AND I. B. WILSON
!:,
it A"
4
:: ...~4
*:
*:S I..... :.
a. ..
::
..
..
::sisi? .. ::<-v ::.:
:. q': . .,:: ...... :.,: .^
.::..:.. ...::.::: ::. .
.:: ' :: : .,
.>' ,: Je '. ...: ...... ::: ::. :::: :*:___._l. |
.iiE'.: ?_ ,,' :__ .:.|r :sllir. Biili=w:,..
:a_:
_ti ..,_.i
r !6
4
j,. .
INI. Pplk.
.i.,i:
lllv::.;4::---...-.--
...6

Preimplantation conceptus of the mouse
Electron microscopyThe trophoblast forms a single layer of cells lining the zona pellucida. About ten
trophoblast cells are cut in cross-section at the maximum diameter of a blastocyst.The plasma membrane facing the zona is irregular and in many cells a line of smallvesicles can be traced running parallel to the plasma membrane and about 0-25 ,uminside it (Fig. 10). The trophoblastic plasma membranes facing the primitive yolk-saccavity and adjacent to the inner cell mass are similar and show no special features.The trophoblast cells are united at their lateral borders by junctional complexes.Junctional complexes are uncommon between the trophoblast and inner mass cells.The endoplasmic reticulum is poorly developed but the cytoplasm of the trophoblast isrich in free ribosome particles. Mitochondria are numerous, and in many the cristaeare distorted so as to lie concentrically. A small Golgi can sometimes be identified.Approximately six to eight inner mass cells can be seen in a cross-section of the
blastocyst at its maximum diameter. Their nuclei are similar to those of the tropho-blast as is the content and the form of the mitochondria. The cells can be distinguishedfrom the trophoblast by the relative sparsity of free ribosomes and the latter cells canbe made out running peripherally to the inner cell mass, separating it from thezona pellucida (Fig. 11). In certain of the inner mass cells which are adjacent to thetrophoblast a zoning of ribosomes is apparent, with a higher concentration ofribosomes in those areas of cytoplasm nearest the trophoblast cells.A variety of cytoplasmic inclusions is found both in the trophoblast and in the
inner cell mass. One or two small lipid droplets 0-5-0{75 ,tm in diameter, some ofwhich have a finely laminated structure, are present in sections of most cells. Slightlylarger membrane bound vesicles up to 2 /um in diameter, with a dark crescenticdeposit along a portion of their circumference, are also found in many cells andmay be a second type of lipid inclusion (Fig. 11). Another species of inclusion, lesscommon than the preceding two, consists of a membrane-bound vesicle containinga dark angular body (Fig. 10).The largest inclusions are limited to the inner cell mass. They are up to 5 ,m in
diameter and consist of membranous and particulate components (Fig. 11). Insidethe bounding membrane are twisted and fragmented lamellae. In some areas theparticles are gathered as an aggregate, structurally parallel to RNA particles in thenucleolus, and elsewhere they are more loosely arranged. Amongst the looselypacked, finely granular material are regular particles approximately 750 A in diameter.A careful check of the 1 ,tm Araldite sections must be made in order to obtainultrathin sections of a blastocyst containing these large inclusions.The zona pellucida is between 2 and 2-5 ,um thick. It is finely granular with an
irregular outer edge and it has no bounding membranes. It is not pierced by anycanals or cavities but at two points in one specimen the continuity of the zona wasbroken and at both sites the trophoblast extends into the gap (Fig. 8). Two regionscan be distinguished in the zona, an inner homogeneous layer and an outer, slightlyless dense, layer of more irregular distribution (Fig. 11).
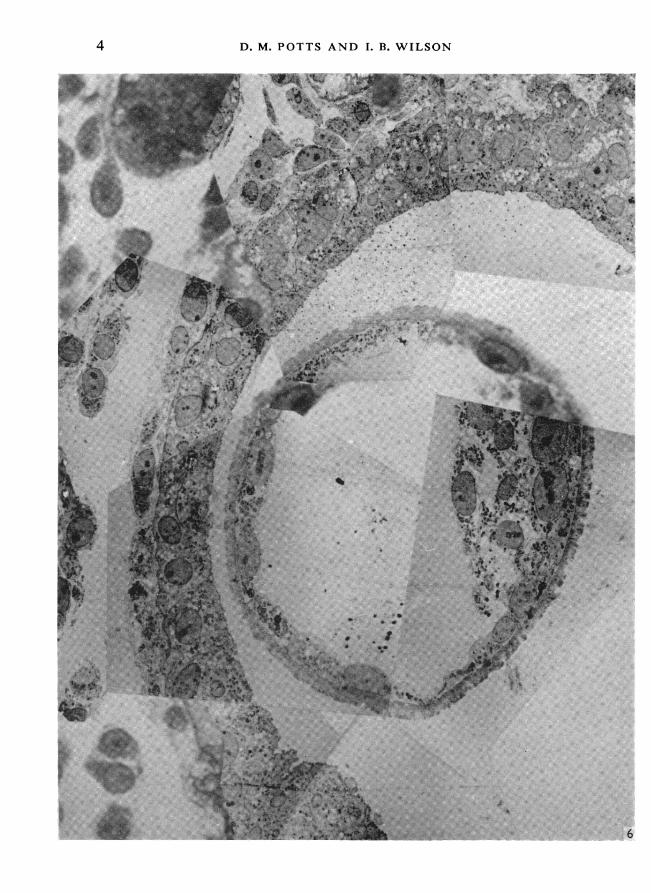
Fig. 6. Montage of electron micrographs prepared from consecutive ultrathin sections.Obscuring grid bars have been cut out and the micrographs superimposed on a light micrographtaken from a corresponding 1 ,tm section.
5

6 ~~~~~~~~~~~~D.M. POTTS AND I. B. WILSON
W *.
ML ~ ~ ~ ~ ~ ~ ~ * ~ { '
.A ~~:.C
,W ~ ~ I
V~~~~~~~~~V
'7

Preimplantation conceptus of the mouseThe cells of the uterine epithelium lining the cup-shaped depression containing
the conceptus are slightly flattened immediately below the blastocyst (Fig. 6). Theepithelial cell nuclei are regular with small nucleoli and are situated in the basalhalf of the cells. The cells carry stunted microvilli on their free border and they reston a well-defined basement membrane (Fig. 7). Mitoses have been observed in theepithelial cells. The lateral walls of adjacent cells run closely parallel, with occasionalinterdigitations of the plasma membranes, and for distances of up to 1 pm or morethe plasma membranes are absent. At some sites of symplasma formation theconjoined plasma membranes form a small hemispherical dilatation (Fig. 9). Similarsyncytial changes are observed throughout the length of the uterus and not only atthe site of implantation.
There is an extensive extracellular space between the subepithelial stromal cells.The subepithelial tissues often pull apart in the neighbourhood of the blastocystalong a line parallel to the epithelial basement membrane. This is almost certainlyan artefact but it is a consistent finding among several specimens and probablyreflects a genuine loosening of the tissues (Fig. 6).
DISCUSSION
Previous electron-microscope studies have been concerned with preimplantationblastocysts flushed from the uterus of the rat (Schlafke & Enders, 1963; Enders &Schlafke, 1965), the armadillo (Enders, 1962) and the rabbit (Hadek & Swift,1962). Rat cleavage stages have been studied by Izquierdo & Vial (1962). There isno detailed published account of the blastocyst in situ in the uterus.
It has been mentioned previously (Wilson, 1962) that the uterine lumen closesup at the time the blastocyst is moved to its antimesometrial implantation site(about 86 h p.c.). We suggest that this is a causal relationship, that the blastocystis in fact moved by closure of the lumen. In carefully fixed and prepared material,microvilli on apposed epithelial cells actually interdigitate. However, even if thewalls do become separated (as in the specimens illustrated in this paper) minorirregularities on the one side are mirrored on the opposite side, indicating theirclose apposition in vivo.
In material prepared for electron microscopy, the zona pellucida appears as aregular, dense layer about 2,um thick. Using freeze-substitution it is irregular,vacuolated and about 6,um thick; this is thought to more accurately represent thein vivo condition. The break in the zona shown in Fig. 8 probably indicates thatthere are weak spots in it at this stage of development, which may account forobservations of 'hatching' that have been made in vitro. We believe that the zonasimply disintegrates at the presumptive implantation site, partly because of ourobservations using freeze-substitution and also because the hatching movementswould be precluded by the close contact between the blastocyst and the epithelium.
In an electron-microscope study on the rabbit, Larsen (1962) has described thefusion of neighbouring uterine epithelial cells. Symplasmic changes in the maternal
Fig. 7. Electron micrograph of uterine epithelium with overlying blastocyst and zona pellucida.At several sites (arrows) the plasma membranes between adjacent epithelial cells have brokendown. x 7 800.
7

8 D. M. POTTS AND I. B. WILSON
. l.b ,* N.
8
i.,
.
Preimplantation conceptus of the mouisetissues at an implantation site have been described for a number of mammals butnot for rodents (Amoroso, 1952). The breakdown of plasma membranes and fusionof maternal and embryonic cytoplasm occurs in the mouse at about 100 p.c. (Potts,1966b). The lateral fusion of epithelial cells described in this paper appears to bea transitory phenomenon and is not found 10 h later. This change has been observedin one specimen only, it may be normal at this stage or it may be an atypicalmanifestation of the symplasmic maternal-embryonic changes which occur at thelater stage.
a^. t',..S w ! R............................ , .. ... . .
# x ; 1 1
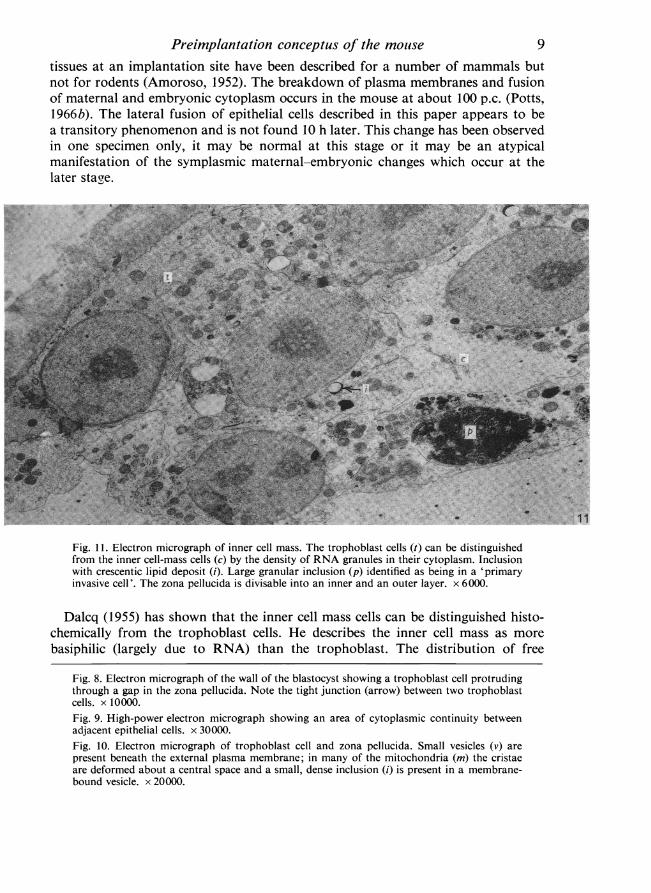
Fig. 11. Electron micrograph of inner cell mass. The trophoblast cells (t) can be distinguishedfrom the inner cell-mass cells (c) by the density of RNA granules in their cytoplasm. Inclusionwith crescentic lipid deposit (i). Large granular inclusion (p) identified as being in a 'primaryinvasive cell'. The zona pellucida is divisable into an inner and an outer layer. x 6000.
Dalcq (1955) has shown that the inner cell mass cells can be distinguished histo-chemically from the trophoblast cells. He describes the inner cell mass as morebasiphilic (largely due to RNA) than the trophoblast. The distribution of free
Fig. 8. Electron micrograph of the wall of the blastocyst showing a trophoblast cell protrudingthrough a gap in the zona pellucida. Note the tight junction (arrow) between two trophoblastcells. x 10000.Fig. 9. High-power electron micrograph showing an area of cytoplasmic continuity betweenadjacent epithelial cells. x 30000.Fig. 10. Electron micrograph of trophoblast cell and zona pellucida. Small vesicles (v) arepresent beneath the external plasma membrane; in many of the mitochondria (m) the cristaeare deformed about a central space and a small, dense inclusion (i) is present in a membrane-bound vesicle. x 20000.
9

D. M. POTTS AND I. B. WILSON
ribosomes demonstrated in this study appears to be the opposite, withmore ribosomesin the trophoblast than in the inner cell mass. Enders & Schlafke (1965) have alsodrawn attention to the disparity between histochemical and electron-microscopicstudies of the blastocyst.
In 1963, one of us (Wilson, 1963) described 'primary invasive cells' as dark,basiphilic bodies present in the inner cell mass prior to loss of the zona. Theirdevelopment and migration from the inner cell mass to the trophoblast and intothe uterine epithelium, as implantation started, was described. More detailed workand the present electron-microscopic observations make it possible to distinguishlarge complex inclusions within cells with cytoplasm and nucleus typical of theinner mass. Whilst the cytoplasm contains very little ribosomal RNA, the testscarried out using RNAase show quite clearly that the inclusion contains a very highconcentration of RNA. It is the inclusions which had previously been identified as'-primary invasive cells' as they appeared under the light microscope to be discrete, ifunusual, cells: the term still seems appropriate for the inclusion and its host cell.The description of their migration from the inner cell mass was based upon circum-stantial evidence; at one stage the inclusions can be seen in the inner cell mass, whilstat a later stage they disappear from this site and, simultaneously, similar onesappear in the trophoblast. They have been identified in the trophoblast and betweenthe uterine epithelial cells at approximately 100 h p.c., as will be described in a laterpaper.
SUMMARY
The morphology of the mouse blastocyst and surrounding maternal tissues isdescribed at 90 h post coitum. The embryos have reached their presumptive im-plantation sites and the uterine lumen is occluded. The zone pellucida is intact butits appearance varies with different methods of preparation. It is suggested that thezona disintegrates in situ. The trophoblast and inner cell-mass cells differ in thestructure of their plasma membranes, in their cell-to-cell junctions and in theirRNA content. Some inner cell-mass cells are also distinguished by large cytoplasmicinclusions. These inclusions are shown to be rich in RNA and it is suggested thatthey can be identified with the 'primary invasive cells' which are a factor that hasbeen described in normal implantation in the mouse.
This work has been partly supported by a generous grant from the Lalor Founda-tion, Delaware, U.S.A. We are grateful to Mrs Sheila Barton and Messrs K. W.Thurley and G. A. M. Oakes for technical assistance.
REFERENCES
AMOROSO, E. C. (1952). Placentation. In Marshall's Physiology of Reproduction, 3rd ed. Ed. A. S. Parkes.London: Longmans, Green.
BLANDAU, R. J. (1961). Biology of eggs and implantation. In Sex and Internal Secretions, 3rd ed. Ed.W. C. Young. Baltimore: Williams and Wilkins.
BoYD, J. D. & HAMILTON, W. J. (1952). Cleavage, early development and implantation of the egg. InMarshall's Physiology of Reproduction, 3rd ed. Ed. A. S. Parkes. London: Longmans, Green.
BURCKHARD, G. (1901). Die Implantation des Eies der Maus in die Uterinschleimhaut. Arch. mikrosk.Anat. EntwMech. 57, 528-569.
10

Preimplantation conceptus of the mouse 11DALCQ, A. M (1955). Processes of synthesis during early development of rodents' eggs and embryos.
Proc. Soc. Study Fert. 7, 113-122.DUVAL, M. (1891). Le placenta des rongeurs. J. Anat. PhYsiol., Paris 27, 24-73.ENDERS, A. C. (1962). The structure of the armadillo blasto:yst. J. Anat. 96, 39-48.ENDERS, A. C. & SCHLAFKE, S. J. (1965). The fine structure of the blastocyst: some comparative studies.
In Ciba Fdn Symp. on Preimplantation Stages of Pregnancy. Eds. G. E. W. Wolstenholme and M.O'Connor. London: Churchill.
FEDER, N. & SIDMAN, R. L. (1958). Methods and principles of fixation by freeze-substitution. J. biophys.biochem. Cytol. 4, 593-602.
HADEK, R. & SWIFT, H. (1962). Nuclear extrusion and intracisternal inclusions in the rabbit blastocyst.J. Cell Biol. 13, 445-451.
IZQUIERDO, L. & VIAL, J. D. (1962). Electron microscope observations on the early development of therat. Z. Zellforsch. mikrosk. Anat. 56, 157-179.
KIRBY, D. R. S., Porrs, D. M. & WILSON, I. B. (1967). On the orientation of the implanting blastocyst.J. Embryol. exp. Morph. (in the Press).
LARSEN, J. F. (1962). Electron microscopy of the uterine epithelium in the rabbit. J. Cell Biol. 14, 49-64.PoTTs, D. M. (1966a). A method for locating specific histological features for electron microscopy.
Jl R. microsc. Soc. 85, 97-102.PoTTs, D. M. (1966b). The ultrastructure of ovoimplantation in the mouse. Am. J. Obstet. Gynec. 96,
1122-1128.SCHLAFKE, S. & ENDERS, A. C. (1963). Observations on the fine structure of the rat blastocyst. J. Anat.
97, 353-360.SELENKA, E. (1883). Keimblatter und Primitivorgane der Maus. Stud. EntwGesch. Tiere 1, 1-27.SOBOTTA, J. (1903). Die Entwicklung des Eies der Maus vom Sclusse der Furchungsperiode bis zum
Auftreten des Amnionsfalten. Arch. mikrosk. Anat. EntwMech. 61, 274-330.WILSON, I. B. (1962). Mechanisms of implantation of the ovum. Yb. Carnegie Instn Wash. 61, 421.WILSON, I. B. (1963). A new factor associated with the implantation of the mouse egg. J. Reprod. Fert.
5, 281-282.