the relationships between behavioural categories and social
TRANSCRIPT

RESEARCH PAPER
The Relationships between Behavioural Categories and SocialInfluences in the Gregarious Big Brown Bat (Eptesicus fuscus)R. Julia Kilgour & R. Mark Brigham
Department of Biology, University of Regina, Regina, SK, Canada
Correspondence
R. Julia Kilgour, Conservation and Science,
Lincoln Park Zoo, 2001 North Clark Street,
Chicago, IL 60614, USA.
E-mail: [email protected]
Received: October 22, 2012
Initial acceptance: November 14, 2012
Final acceptance: December 4, 2012
(L. Ebensperger)
doi: 10.1111/eth.12052
Abstract
Behavioural plasticity is a critical component of natural selection leading
to evolution. However, a surge of studies in the last two decades has dis-
covered a distinct limit to behavioural plasticity, commonly referred to as
behaviour types and behavioural syndromes. We set out to understand
the relationships across behavioural categories in wild-caught adult,
female big brown bats and how they compare between social and solitary
behaviours. Using bats sampled from four different maternity colonies, we
ran a series of behavioural assays to create a behavioural profile for each
individual. The behavioural profile encompassed exploratory, learning,
competitive and aggressive categories. We found that Big brown bats exhi-
bit a mean profile relatively unique to other well-documented species,
where aggression was linked to increased competitive ability but not to
boldness. Our results indicate that the solitary and socially directed behav-
iours of individuals are not necessarily related and that behaviours per-
taining to social interactions are linked most closely to learning abilities.
Furthermore, we found evidence that poor body condition may be a pre-
dictor of increased social interactions and that behaviours exhibited in the
presence of conspecifics are unrelated to those exhibited in solitude. These
findings indicate importance of social affiliations on individual behaviours
in this species and their uniqueness relative to other well-studied taxa.
Introduction
Plasticity and phenotypic variation in inherited traits
are the driving forces behind evolutionary change in
ecological traits. Behavioural plasticity allows organ-
isms to adapt to the surrounding environment, and
variation in behavioural traits is thus considered
adaptive given the heterogeneity of habitats (Via et al.
1995; Dingemanse et al. 2004). However, our current
understanding of individual differences implies that
animals actually exhibit a limited range of behaviour-
al variation across changing environmental contexts
(Wilson 1998; Sih et al. 2004a,b). For example,
numerous studies have reported consistency in indi-
vidual aggressive behaviour (a behavioural type)
between contexts (Huntingford 1976; Riechert &
Hedrick 1993; McGhee & Travis 2010); but see
(Coleman & Wilson 1998). Concepts such as the
aggressive spillover hypothesis suggest that juvenile
voracity and adult aggression are contextually adap-
tive but are also linked to pre-copulatory sexual can-
nibalism in female fishing spiders (Dolomedes triton)
(Johnson & Sih 2005). The intensity of the behaviour
displayed may differ among individuals, where some
individuals consistently exhibit lesser or higher
degrees of that behaviour (e.g. high explorers and low
explorers in Great Tits, Parus major (Verbeek et al.
1996)). Furthermore, the expression of consistent
individual differences has been demonstrated across a
wide range of taxa (mammals: Vervaecke et al. 1999;
Dochtermann & Jenkins 2007; birds: Verbeek et al.
1996; Dingemanse et al. 2002; fish: Harcourt et al.
2009; Bell & Sih 2007; invertebrates: Wilson et al.
2010; Sih & Watters 2005).
Despite the wealth of knowledge that has accumu-
lated on the topics of consistent individual differences,
Ethology 119 (2013) 189–198 © 2013 Blackwell Verlag GmbH 189
Ethology

the range of behaviours examined across studies
remains limited. Generally, authors have focused on
expressions of boldness and/or activity between indi-
viduals (Johnson & Sih 2005; Dochtermann & Jenkins
2007; Duckworth & Badyaev 2007; Reaney &
Backwell 2007). However, alternate selection pres-
sures imposed by differences in ecology and environ-
ments among species suggest that behaviour types
and how they correlate can be unpredictable within
and across species (Sih & Bell 2008). For example,
optimal behaviour types likely differ between solitary
and social organisms where behavioural types such as
aggression can have very different long-term impacts
on individual fitness, and therefore cannot be pre-
dicted using the same paradigm. Among cooperatively
breeding cichlids, the most effective helpers in intru-
der defence are the most aggressive individuals (Le
Vin et al. 2011). Despite this, studies examining the
social effects of individual differences have generally
focused on single aspects of social relationships such
as aggressive or agonistic interactions (Budaev 1997;
McGhee & Travis 2010; David et al. 2011; Pruitt et al.
2011) or, alternatively, on the general interaction pat-
terns among all group members (Moretz et al. 2007;
Pike et al. 2008; Croft et al. 2009; Harcourt et al.
2009). These measures provide information about just
two dimensions of a long list of complex interaction
types between conspecifics. Many animal species are
highly social, meaning there has been emphasis
placed on benefits received from iterative interactions
with group members through predator defence,
assisted breeding or information transfer (Wilson
2000). Therefore, behavioural categories such as
learning proficiency represent an important skill
among social animals, where key behaviours are
derived from exposure to experienced individuals
(Kavaliers et al. 2005). Learning ability may be pre-
dictable based on other behaviours such as explora-
tion of the environment and could also affect an
individual’s competitive nature with its groupmates.
Examination of a broad range of behavioural catego-
ries is necessary to gain a broader understanding of
how an individual’s behaviour is impacted by its
groupmates and context.
In this study, we sought to examine the relation-
ships between behaviours across a wide dimension of
behavioural categories and across different social
environments in long-lived, gregarious female big
brown bats (Eptesicus fuscus). We addressed the follow-
ing questions: (1) What, if any, is the relationship
between traits encompassing activity, learning, com-
petitiveness and aggressiveness? Activity and aggres-
siveness are behavioural categories commonly
examined in studies of behavioural correlates,
although they commonly show differential degrees of
relatedness depending on species and context (Boog-
ert et al. 2006; Biro et al. 2010; Kralj-Fiser et al.
2010; Wilson et al. 2010). Learning behaviour and
competitiveness can heavily impact an individual’s
relationship among its social partners, and yet, is
rarely examined in studies of behavioural traits and
syndromes (Wilson et al. 2010; Pruitt et al. 2011); (2)
How do behaviours exhibited in solitary treatments
compare with those exhibited in the presence of a
conspecific? While other studies have examined
behavioural syndromes and group dynamics, little is
known about the relationships between behaviours
exhibited in solitary and social contexts (Bergmuller
& Taborsky 2010).
Methods
Study Animals
We selected the big brown bat (Eptesicus fuscus) as
our study species. Big brown bats are a common
vespertilionid bat, which occurs in a wide range of
habitats across North America. During the summer,
females form maternity colonies, in which adult
males usually do not occur and exhibit strong
philopatry to specific roosting areas (Willis &
Brigham 2004 for forests; Brigham 1991 for build-
ings). Within maternity colonies of forest-dwelling
populations, females conform to a fission–fusionsocial system (Willis & Brigham 2004). The prefer-
ential associations between females in subgroups are
not based on genetic relatedness (Metheny et al.
2008), indicating that female roost-mate preferences
are mediated by other factors. Females typically
have one offspring per year, and juveniles have low
overwinter survival (O’Shea et al. 2010). Like many
small mammals, thermoregulation represents a high
energetic cost. Bats rely extensively on torpor to
save energy during daytime roosting periods
throughout the summer (Racey & Swift 1981).
However, many species adjust torpor use during
certain periods of the reproductive cycle (Audet
& Fenton 1988; Grinevitch et al. 1995; Dzal &
Brigham 2012), placing greater importance on social
thermoregulation (Willis et al. 2006; Pretzlaff et al.
2010) between members of maternity colonies dur-
ing those periods. Therefore, social associations
unique between females are likely critical to big
brown bats given the potential benefits.
We collected individuals from four different
building-roosting maternity colonies in southern
Ethology 119 (2013) 189–198 © 2013 Blackwell Verlag GmbH190
The Relationships between Behavioural Categories and Social Influences R. J. Kilgour & R. M. Brigham

Saskatchewan and south-western Alberta (hereafter
referred to colonies 1 through 4, listed in chronologi-
cal order). Individuals were either caught using mist
nets (Colonies 3 and 4) or manually removed from
building roosts by hand where feasible (Colonies 1
and 2). Trapping sessions only took place during the
early or late summer to minimize the risk of disturb-
ing parturition and lactation. During the early sum-
mer, bats were expected to be in early to mid-stages of
pregnancy. Individuals from colonies 1 (n = 8) and 2
(n = 4) were caught during the typical period of preg-
nancy (May–June). No individuals caught were detec-
tably pregnant. Individuals from colonies 3 (n = 8)
and 4 (n = 8) were trapped during the post-lactation
period and all females but two exhibited signs of lacta-
tion from that year.
Experiments were conducted at the University of
Regina Cypress Hills Field Station (49°34′N and 109°53′W) during the summer of 2009, with only one
group of bats (from a single colony) held in captivity
at a time. Bats were transported to the field station
in individual cotton bags. Each bat was injected with
a passive integrative transponder (PIT) tag for identi-
fication and given a unique combination of reflec-
tive forearm bands to allow for individual
identification during video recording. Bats were
hand fed Tenebrio molitor larvae, to monitor individ-
ual food intake and had access to water ad libitum.
Bats were group-housed indoors in a 145-l Exo
Terra flexarium (Rolf C. Hagen Inc., Montreal, Can-
ada) when not participating in trials. The seasonal
light cycle was maintained, except when conducting
experiments and lights were turned on for assays,
and room temperature was maintained at 20–24°C.All behavioural assays were conducted between
20:00 and 04:00, during the natural period of activ-
ity. Bats were held in captivity for no longer than
11 d and following the experimental sessions, they
were returned to the colony where they were
trapped. All protocols were approved by the Univer-
sity of Regina President’s Committee on Animal
Care (protocol 08-01).
To examine relationships between behavioural cat-
egories, we compared the expression of seven
behavioural variables by all individuals, three solitary
behaviours (learning ability, solitary exploration and
latency to feed in captivity) and four social behaviours
(competitive ability, social exploration, latency to
explore in social contexts and biting frequency).
We also examined the relationship between behavio-
ural variables and body condition. The methodologies
used to assess behaviours and traits measured are
described below.
Body Condition Index (BCI)
To compensate for differences in overall body form,
the body condition of each bat was calculated as the
ratio of body mass (g) divided by forearm length
(mm). Forearm length was measured as the length
of the right forearm (� 0.1 mm), and body mass
(� 0.1 g) was the initial mass measured when the
individual was caught, prior to feeding. Therefore, a
high value represents an individual with a better body
condition.
Learning Ability (Learning)
The learning ability of each bat was measured during
training sessions prior to the competitive ability
experiment. Individuals were trained to walk to a
food piece located on a plastic tray. Individuals were
placed in an unfamiliar triangular arena and trained
to feed in a series of six steps. At each step, the bat
was placed farther away from the dish with the food.
After a bat successfully acquired the food at a set dis-
tance five times in succession, the bat proceeded to
the next step, where the food tray was placed farther
away. Therefore, a bat with the highest score for
learning ability completed the training in 25 trials,
although most individuals required more trials to
effectively ‘pass’ a step. Prior to trials, food was with-
held for 20 h to increase motivation. Observers
remained in the room during trials, but out of sight
for each session. Noise was kept to a minimum. Indi-
viduals were scored between 1 and 6 for their learning
ability based on the number of training sessions nec-
essary for them to complete the prescribed training
procedure. A score of 6 represents a fast-learning indi-
vidual, and a score of 1 represents a slow-learning
individual. For further information, please see
(Table S1).
Competitive Ability (Comp)
Females were placed in dyadic food competitions with
group members to assess competitive ability. Bats
were trained to acquire a piece of Tenebrio larvae from
a food dish (see ‘learning ability’). Individuals were
then tested in dyadic combinations with all other bats
from the same colony and each trial was repeated five
times. In other words, each dyad underwent five com-
petition trials. If neither competitor consumed the
food piece after 60 s, it was considered a mistrial. The
individuals used in trials were randomized, although
no female competed twice consecutively. Therefore,
each female had a minimum rest period of 1 min
Ethology 119 (2013) 189–198 © 2013 Blackwell Verlag GmbH 191
R. J. Kilgour & R. M. Brigham The Relationships between Behavioural Categories and Social Influences

between trials. When not being tested, bats were kept
in cloth holding bags. Prior to trials, food was with-
held for 20 h to increase motivation. Trials took place
over 2–3 nights to reduce the effects of satiation. After
trials were complete for a night, bats were re-tested
for their ability to consume food pieces and all suc-
cessfully acquired and consumed them, indicating
that performance was not due to a lack of training or
satiation.
Competition trials consisted of two bats placed in a
triangular arena (35 9 47 9 47 cm; walls 19 cm
high) at the same time, equidistant from food. Trials
lasted until one bat successfully ate the food (typi-
cally, <30 s). Because unequal numbers of individuals
were caught at each colony, there were unequal
numbers of trials (and thus the potential number of
wins) between individuals from different colonies.
To compensate, competitive ability was standardized
by calculating the number of wins relative to the total
number of trials per individual. Competitive ability of
individuals was not measured between groups due to
limitations in acquiring bats from maternity colonies
and the logistics of holding multiple colonies simulta-
neously within the captive environment.
Exploration of a Novel Environment in a Solitary
Context (SoloEX)
To measure exploration as an indicator of an individ-
ual’s activity levels, we placed bats in a triangular
arena (different from that used in the competitive
experiment; 40 9 50 9 48 cm; walls 42 cm high)
and measured the amount of time bats exhibited
exploratory behaviour. Two novel objects were placed
in the arena during each trial, and these were chan-
ged after 8–10 trials to prevent habituation. The novel
objects were the following: a bean bag sack, a ceramic
mug, a plastic toy gun, a large plastic cup, a rock, a set
of plastic mini-speakers, a roll of duct tape and a plas-
tic lid from a small animal cage. The bats had no expo-
sure to the arena prior to trials. To encourage
exploration, bats were not allowed any habituation
period with the objects.
Exploratory behaviour was defined as any active
movement within the arena and most often took the
form of escape behaviours (e.g. attempting to climb
over the walls of the arena) and object exploration
(e.g. where bats walked under or over objects). There-
fore, an individual’s exploration score was quantified
as the number of seconds it exhibited exploratory
behaviours over the duration of the trial. We recorded
trials with a Sony Handycam (HDR-XR200V, Sony,
Tokyo, Japan) positioned directly above the arena.
Once bats were placed in the arena, observers left the
room. Trials lasted three min, and each individual was
run in three trials daily for three nights.
Exploration of a Novel Environment in a Social Setting
(SocialEX)
Exploratory behaviour was also measured in the pres-
ence of other bats. Individuals were run in explor-
atory trials in dyadic combinations with all other
group members. Each pairwise combination was
repeated three times. The rotation of novel objects,
the duration and execution of trials and the measure-
ment of exploratory behaviour were identical to the
solitary exploration trials. Pairwise combinations were
randomized and no individual participated in two
consecutive trials. Bats were kept in cloth holding
bags between trials. This treatment was also used to
measure activity levels of individuals.
Latency of Exploration in Social Settings (SocialLat)
The latency to explore was measured as the number
of seconds from the beginning of the trial to the time
when an individual began its first exploration bout of
Social Exploration trials. If an individual did not
explore during the trial, they were given a latency
score of 300 based on the 300 s duration of the trial.
Frequency of Biting Behaviour (BiteFrq)
Biting, or attempted biting, is an aggressive behaviour
used among groupmates of many different species.
Biting behaviour was assessed as the frequency of suc-
cessful or unsuccessful attempts made by each bat to
bite the other bat during the social exploration trials.
Any biting that occurred during the competitive
experiment trials was not included, as the motivation
for aggression was unequal between the two experi-
ments. Furthermore, there was a low frequency of
observed biting attempts during competitive trials.
Bats received a score for biting behaviour based on
the frequency of biting (or attempting to bite) com-
panion animals during social exploration trials.
Latency to Feed in Captivity (LatFeed)
Bats were given a score based on their ability to feed
consistently, as another measure of a bat’s learning
ability. For the 10 d that bats were maintained in cap-
tivity, they were fed mealworms (Tenebrio larvae) by
hand which allowed us to monitor food intake by
each individual. We detected variation among bats in
Ethology 119 (2013) 189–198 © 2013 Blackwell Verlag GmbH192
The Relationships between Behavioural Categories and Social Influences R. J. Kilgour & R. M. Brigham

how quickly they began to feed, and each bat was
given a score for their latency to feed. Bats were given
scores ranging from 1 to 5 based on the number of
Tenebio larvae consumed during the first feeding bout
and consecutive bouts. A score of 1 was given to bats
who successfully consumed six larvae on the first
feeding attempt and a minimum of six larvae at all
future feeding times. Successive increases in score
number were based on number of feeding attempts
and days passing until successfully consuming a mini-
mum of six larvae at one feeding. Bats removed from
colonies by hand were caught in the afternoon and
brought to the field station that evening, and bats
caught in mist nets in the evening as they exited from
day roosts. Thus, all bats were presumed to be hungry
upon arrival at the field station. Bats were fed 6–8mealworms in two feeding sessions each night. They
were considered to be feeding consistently once they
ate six full larvae consecutively and continued to
consume at least six mealworms per feeding bout. For
more detailed information, see (Tables S2 and S3). All
bats learned to eat consistently in captivity within
4 d.
Statistical Analysis
We used Exploratory Factor Analysis (EFA) to exam-
ine the relationships among the behavioural catego-
ries measured. EFA is useful in estimating the
relationships between variables without a priori
hypotheses. Unlike Principal Component Analysis,
EFA does not expect communality to be 1, allowing
for error variance in contributing variables. The num-
ber of factors was determined based on the Scree plots
and the Chi-square statistic, which tests the null
hypothesis that the model does not fit the data. We
used promax rotation, a type of oblique rotation,
which allows for correlations between factors. All
analyses were conducted in R, Version 2.13.0 (2009).
Results
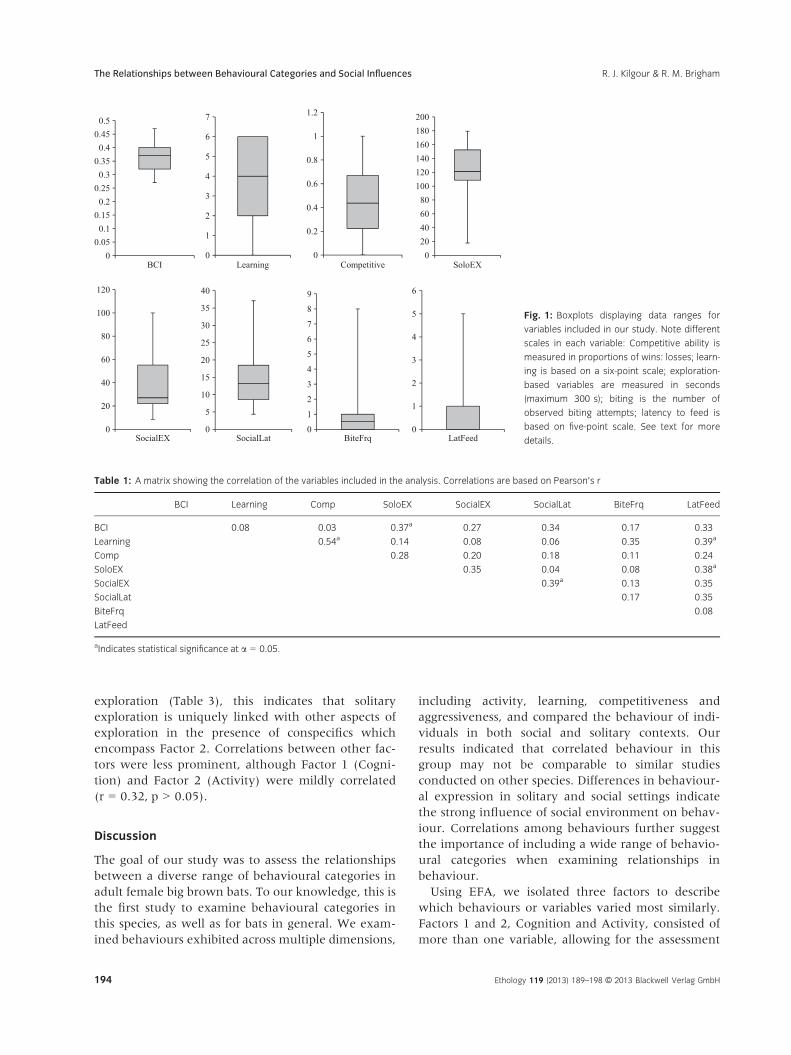
Bats exhibited considerable variation across the seven
categories of behaviours we measured (Fig. 1). With
the data pooled across all colonies, we found that the
majority of bats learned the feeding task quickly
(mean score of 3.7 � 0.42 of 6, where a score of six
indicates that an individual passed through each level
without repetition). Bats also generally had low laten-
cies to feed in captivity, as exhibited by a low mean
score (2.03 � 0.29 of 5), demonstrating that few lev-
els had to be repeated before moving on to the next
level. Bats exhibited higher exploratory behaviour in
solitary vs. social trials (overall mean seconds explor-
ing, 124.75 � 7.16 and 39.94 � 5.0, respectively).
Further analysis of these variables in a correlation
matrix allows for greater understanding of the relat-
edness between the variables (Table 1). Statistical sig-
nificance at the a = 0.05 level was observed between
several behavioural pairings (BCI and SoloEx, Learn
and Comp, Learn and LatFeed, SoloEx and LatFeed,
SocEx and SocLat); however, significance was lost
between all pairings when Bonferoni corrections were
applied. The correlations between the variables were
generally weak, with the exception of Learning and
Competitive behaviours (r = 0.54, p < 0.05).
Multivariate results from the EFA showed three fac-
tors resulting from the eight variables included in the
analysis (Fig. 2). A Chi-square analysis testing the
hypothesis that the model does not fit significantly
worse than a model where the variables correlate
freely was rejected (v2 = 1.99, df = 7, p = 0.961)
(Table 2). Barlett’s test of sphericity supported our
previous findings of sufficient correlations between
the behavioural variables (v2 = 43.74, df = 28,
p = 0.03). Kaiser–Meyer–Olkin (KMO) tests show
that, despite our small sample size, our sampling was
adequate to support the EFA (KMO = 0.58) (Budaev
2010). KMO and Barlett’s test were run in the paf
function from Rela library in R (Chajewski 2009). The
first factor relates to the measures of learning, compe-
tition, biting frequence and latency to feed, where
bats who learned quickly to obtain a food piece in the
training arena (Learning) were also individuals who
won the highest proportion of their competitive trials
(Comp), as well as had the shortest latency to feed
from the hand when first brought into captivity
(FeedLat) and also most likely to bite a conspecific
(BiteFrq). As these four categories refer to behaviours
requiring assessment and potentially cognitive skill,
the first factor is labelled ‘Cognition’. The second fac-
tor links a high BCI to a shorter latency to explore in a
social environment (SocialLat), as well as less time
spent exploring overall in the presence of a social
companion (SocialEX). This second factor also relates
to a high biting frequency (BiteFrq) and a longer
latency to feed from the hand (FeedLat). This factor
describes the overall activity level of the individual
and is therefore labelled ‘Activity’. The final factor
contains only the variable SoloEX, describing the
average exploration exhibited by a bat without any
conspecific present.
Correlations between the factors (Table 3) indicate
that Factor 2 (Activity) is highly correlated with Fac-
tor 3 (SoloEx) (r = �0.52, p < 0.05). Considering Fac-
tor 3 consists entirely of the measure of solitary
Ethology 119 (2013) 189–198 © 2013 Blackwell Verlag GmbH 193
R. J. Kilgour & R. M. Brigham The Relationships between Behavioural Categories and Social Influences
exploration (Table 3), this indicates that solitary
exploration is uniquely linked with other aspects of
exploration in the presence of conspecifics which
encompass Factor 2. Correlations between other fac-
tors were less prominent, although Factor 1 (Cogni-
tion) and Factor 2 (Activity) were mildly correlated
(r = 0.32, p > 0.05).
Discussion
The goal of our study was to assess the relationships
between a diverse range of behavioural categories in
adult female big brown bats. To our knowledge, this is
the first study to examine behavioural categories in
this species, as well as for bats in general. We exam-
ined behaviours exhibited across multiple dimensions,
including activity, learning, competitiveness and
aggressiveness, and compared the behaviour of indi-
viduals in both social and solitary contexts. Our
results indicated that correlated behaviour in this
group may not be comparable to similar studies
conducted on other species. Differences in behaviour-
al expression in solitary and social settings indicate
the strong influence of social environment on behav-
iour. Correlations among behaviours further suggest
the importance of including a wide range of behavio-
ural categories when examining relationships in
behaviour.
Using EFA, we isolated three factors to describe
which behaviours or variables varied most similarly.
Factors 1 and 2, Cognition and Activity, consisted of
more than one variable, allowing for the assessment
Table 1: A matrix showing the correlation of the variables included in the analysis. Correlations are based on Pearson’s r
BCI Learning Comp SoloEX SocialEX SocialLat BiteFrq LatFeed
BCI 0.08 �0.03 �0.37a �0.27 �0.34 0.17 0.33
Learning 0.54a 0.14 0.08 0.06 0.35 �0.39a
Comp 0.28 0.20 0.18 0.11 �0.24
SoloEX 0.35 0.04 0.08 �0.38a
SocialEX 0.39a �0.13 �0.35
SocialLat �0.17 �0.35
BiteFrq 0.08
LatFeed
aIndicates statistical significance at a = 0.05.
00.050.10.150.20.250.30.350.40.450.5
BCI0
1
2
3
4
5
6
7
Learning0
0.2
0.4
0.6
0.8
1
1.2
Competitive020406080100120140160180200
SoloEX
0
20
40
60
80
100
120
SocialEX0
5
10
15
20
25
30
35
40
SocialLat0
1
2
3
4
5
6
7
8
9
BiteFrq0
1
2
3
4
5
6
LatFeed
Fig. 1: Boxplots displaying data ranges for
variables included in our study. Note different
scales in each variable: Competitive ability is
measured in proportions of wins: losses; learn-
ing is based on a six-point scale; exploration-
based variables are measured in seconds
(maximum 300 s); biting is the number of
observed biting attempts; latency to feed is
based on five-point scale. See text for more
details.
Ethology 119 (2013) 189–198 © 2013 Blackwell Verlag GmbH194
The Relationships between Behavioural Categories and Social Influences R. J. Kilgour & R. M. Brigham

of relationships between these variables. Factor 1,
labelled ‘Cognition’, describes characteristics of cogni-
tion and specific aspects of social interaction. This
encompasses variables of ease of learning a new task
(Learning), ease of learning a new feeding technique
in a new environment (LatFeed), competitive ability
over conspecifics (Comp) and frequency of biting
aggression to conspecifics (BiteFrq) (Table 2). Com-
bined, these variables describe an organism that can
rapidly learn and adapt to a new environment and
out-compete others for limited resources. An individ-
ual who exhibits all of these characteristics may be
more likely to acquire access to limited resources.
Although we did not examine dominance relation-
ships in this study, dominance is often defined as an
individual’s resource holding power (Parker 1974).
Further, many aspects of a successful competitor lie in
the ability to assess the social environment prior to
competition, making behavioural plasticity crucial in
acquiring socially derived benefits (Galef & Wigmore
1983; Moscovice & Snowdon 2006). Based on our
data, body size is not a predictor of competitive ability
in female bats, which may reflect the uniqueness of
this study system.
Factor 2 considers behavioural categories of social
activity, as well as body size. We found that bats with
a high BCI also exhibited the shortest latency to
explore in the presence of a conspecific (SocialLat),
were overall less exploratory in the presence of a con-
specific (SocialEX), most frequently bit a companion
(BiteFrq) and also took the longest to learn to feed by
hand in captivity (LatFeed) (Table 2). When placed in
the novel environment with a conspecific, bats who
were not exploring spent most of the time engaged in
social activity with their companion. Therefore, we
see that bats who initiate exploration quickly in a
social environment also are mostly likely to spend the
trial period interacting with their companion. Fur-
thermore, the high BCI reflects individuals who were
of improved body condition were those who were
more often engaged in social interactions. The
increased biting frequency may be a result of
increased social interaction and increased opportunity
for aggressive encounters. Bats that are of better body
condition may be this way because they glean energy
savings through increased social interactions. Individ-
uals who are more active in the presence of compan-
ions may lose energy savings or the ability to roost
socially based on decreased interactions with group-
mates. Among shoaling Trinidadian guppies (Poecilia
reticulata), individuals who engage more in bold
behaviours (predatory inspection) were also had
fewer social connections with groupmates (Croft et al.
2009).
Comparing bats to other species, our examination
of the relationships between behaviour categories is
atypical. For example, correlations between behav-
iours are often observed in aggression and boldness,
although not always (Wilson et al. 2010). Exploratory
behaviour has often been considered a measure of
boldness and neophobia (Wilson et al. 1994). When
exploratory behaviour is measured in a completely
novel environment, the motivation underlying the
exploratory behaviour may be similar to boldness
(Hughes 1997). Our results indicate that aggression
level was not prominently related to boldness, where
Table 2: Inclusive variables listed with factor loadings. Factors were
determined based on Scree plots and non-significance of chi-square
statistics
Variable Factor 1 Factor 2 Factor 3
Learning 1.02
Competitive 0.5
Biting 0.36 0.39
Latency to Feed �0.31 0.48
Social Exploration �0.48
Body Composition Index 0.42
Latency for Social Exploration �0.83
Solitary Exploration 1.11
Cumulative Proportion of
Variance Explained
0.20 0.39 0.57
Table 3: Correlations between the factors isolated using exploratory
factor analysis. Values represent correlations as calculated using Pear-
son’s r
Factor 2 Factor 3
Factor 1 0.32 �0.12
Factor 2 �0.52a
aIndicates statistical significance at a = 0.05.
BCI
LearningComp
SoloEX
SocialEX
SocialLat
BiteFrqLatFeed
–1
–0.8
–0.6
–0.4
–0.2
0
0.2
0.4
0.6
–0.4 –0.2 0 0.2 0.4 0.6 0.8 1 1.2
Fact
or 1
Factor 2
Fig. 2: Relationships between variables across Factors 1 and 2 as
derived from exploratory factor analyses.
Ethology 119 (2013) 189–198 © 2013 Blackwell Verlag GmbH 195
R. J. Kilgour & R. M. Brigham The Relationships between Behavioural Categories and Social Influences

biting was negatively related to exploration in the
presence of a conspecific (Factor 2, Table 2) and was
positively related to learning ability and competive-
ness among bats (Factor 1, Table 2). Examination of
the correlations between variables shows a slight posi-
tive relationship between individuals who most fre-
quently bit a conspecific and those who trained to
acquire a food piece from a designated area most
quickly (Table 1). These results may suggest, how-
ever, that aggression might play a role in competitive
interactions between groupmates. In social animals,
aggression towards conspecifics is typically used as a
means of acquiring resources through dominance
relationships (Chase et al. 1994; Draud et al. 2004).
Our results imply that this may be consistent in big
brown bat maternity groups. Although bats do not
compete for food in the wild, they might compete for
roosting locations, where the individual who occupies
the warmest location in the roost is less likely to go
into deep torpor during the roosting period (McGo-
wan et al. 2006). Among females, this could lead to
greater energy savings to a developing foetus. Big
brown bats might use aggressive behaviours such as
biting to gain access to high valued roosting locations.
Therefore, unlike in other species, aggression towards
group mates was not a predictor of activity levels or
boldness. Considering other behavioural measures,
such as learning ability, bats exhibited similar rela-
tionships to starlings (Sturnus vulgaris). Measures of
learning ability in wild-caught starlings suggested that
learning ability did correlate with competitive ranks
but not to boldness (Boogert et al. 2006). Other stud-
ies that have examined the relationship between
exploratory behaviour and dominance interactions in
black-capped chickadees (Fox et al. 2009) show that
low-exploring individuals out-compete subordinate
individuals for roosting locations. Our results suggest
that neither solitary nor social exploratory behaviour
in bats is an indicator of competitive or aggressive
pairwise interactions (Tables 1 and 2). This study sup-
ports the growing body of evidence suggesting that
behavioural categories, and the relationships between
them, differ across species.
In addition to our examination of multiple behavio-
ural categories included in this analysis, we also con-
sidered behaviours exhibited in solitary and social
settings. Solitary behaviours included learning ability,
latency to feed in captivity and solitary exploration.
Variables measured when bats were in social settings
included biting frequency, competitive ability, social
exploration and social latency. Exploratory behaviour
was measured in both solitary and social settings, and
we found only a slight positive correlation between
those two variables (Table 1). However, solitary
exploration was best described as a behaviour in and
of itself, as it was the sole contributor to Factor 3. Fur-
thermore, Factor 2, which included the variables
describing exploration in the presence of a conspe-
cific, was negatively correlated with Factor 3 suggest-
ing that solitary exploration in bats is negatively
related to activity in the presence of a conspecific.
However, bats were less exploratory of their sur-
roundings when in the presence of a social compan-
ion (Table 1), implying that bats alter their behaviour
in the presence of social companions, a finding consis-
tent with other studies (Metcalfe et al. 1987; Pintor
et al. 2008; Mainwaring et al. 2011).
Our data demonstrate that correlated behavioural
categories vary across different taxa, perhaps imposed
by unique selection pressure imposed on animals
with unique ecologies. Furthermore, our findings
emphasize the importance of social context on
behavioural phenotypes. It is clear from our data that
the presence of a conspecific has the potential to alter
the behaviours exhibited by bats and that the rela-
tionships between behaviour of individuals found in
bat maternity colonies may be unique among social
animals. More study is needed to understand how
the behavioural categories we included relate to indi-
vidual behaviour, fitness consequences and social
relationships. A larger sample size as well as measur-
ing social behaviours such as competitive behaviour
across colonies would provide greater insight into
behavioural profiles compare across individuals. Our
measure of competitive ability was limited to com-
parisons only between dyads of group members,
potentially confounding the independence of the
result. To our knowledge, this is the first study to
examine correlations in behaviour in this species.
Given the differences in correlated behaviours that
we discovered when compared with other taxa, it is
clear that understanding the ecological and evolu-
tionary context of each species and population is cru-
cial when examining behavioural categories and
their relationships.
Acknowledgements
We thank M Dunbar, E Gillam, GG McNickle and J
Ratcliffe for helpful comments on the manuscript.
We also thank D Braun, A Matheson and J Poissant
for their assistance with field work and care of captive
bats. Funding was provided by the Natural Sciences
and Engineering Research Council of Canada (Discov-
ery Grant to RMB) and the American Society of
Mammalogists (Grants-in-Aid of Research, RJK).
Ethology 119 (2013) 189–198 © 2013 Blackwell Verlag GmbH196
The Relationships between Behavioural Categories and Social Influences R. J. Kilgour & R. M. Brigham

Literature Cited
Audet, D. & Fenton,M. B. 1988: Heterothermy and the use
of torpor by the bat eptesicus-fuscus (Chiroptera, Vesper-
tilionidae) – a field-study. Physiol. Zool. 61, 197—204.
Bell, A. M. & Sih, A. 2007: Exposure to predation gener-
ates personality in threespined sticklebacks (Gasterosteus
aculeatus). Ecol. Lett. 10, 828—834.
Bergmuller, R. & Taborsky, M. 2010: Animal personality
due to social niche specialisation. Trends Ecol. Evol. 25,
504—511.
Biro, P. A., Beckmann, C.& Stamps, J. A. 2010: Smallwithin-
day increases in temperature affects boldness and alters per-
sonality in coral reef fish. Proc. Biol. Sci. 277, 71—77.
Boogert, N. J., Reader, S. M. & Laland, K. N. 2006: The
relation between social rank, neophobia and individual
learning in starlings. Anim. Behav. 72, 1229—1239.
Brigham, R. M. 1991: Flexibility in foraging and roosting
behavior by the big brown bat (Eptesicus fuscus). Can. J.
Zool./Rev. Can. Zool. 69, 117—121.
Budaev, S. V. 1997: ‘Personality’ in the guppy (Poecilia re-
ticulata): a correlational study of exploratory behavior
and social tendency. J. Comp. Psychol. 111, 399—411.
Budaev, S. V. 2010: Using principal components and factor
analysis in animal behaviour research: caveats and
guidelines. Ethology 116, 472—480.
Chajewski, M. 2009: rela: Scale item analysis. R package
version 4.1.
Chase, I. D., Bartolomeo, C. &Dugatkin, L. A. 1994: Aggres-
sive interactions and inter-contest interval – how long do
winners keepwinning. Anim. Behav. 48, 393—400.
Coleman, K. & Wilson, D. S. 1998: Shyness and boldness
in pumpkinseed sunfish: individual differences are con-
text-specific. Anim. Behav. 56, 927—936.
Croft, D. P., Krause, J., Darden, S. K., Ramnarine, I. W.,
Faria, J. J. & James, R. 2009: Behavioural trait assort-
ment in a social network: patterns and implications.
Behav. Ecol. Sociobiol. 63, 1495—1503.
David,M., Auclair, Y. &Cezilly, F. 2011: Personality predicts
social dominance in female zebra finches, Taeniopygia
guttata, in a feeding context. Anim. Behav. 81, 219—224.
Dingemanse, N. J., Both, C., Drent, P. J., Van Oers, K. &
Van Noordwijk, A. J. 2002: Repeatability and heritabil-
ity of exploratory behaviour in great tits from the wild.
Anim. Behav. 64, 929—938.
Dingemanse, N. J., Both, C., Drent, P. J. & Tinbergen, J. M.
2004: Fitness consequences of avian personalities in a
fluctuating environment. Proc. Biol. Sci. 271, 847—852.
Dochtermann, N. A. & Jenkins, S. H. 2007: Behavioural
syndromes in Merriam’s kangaroo rats (Dipodomys merri-
ami): a test of competing hypotheses. Proc. Biol. Sci.
274, 2343—2349.
Draud, M., Macias-Ordonez, R., Verga, J. & Itzkowitz, M.
2004: Female and male Texas cichlids (Herichthys
cyanoguttatum) do not fight by the same rules. Behav.
Ecol. 15, 102—108.
Duckworth, R. A. & Badyaev, A. V. 2007: Coupling of dis-
persal and aggression facilitates the rapid range expan-
sion of a passerine bird. Proc. Natl Acad. Sci. USA 104,
15017—15022.
Dzal, Y. A. & Brigham, R. M. 2012: The tradeoff between
torpor use and reproduction in little brown bats (Myotis
lucifugus). J. Comp. Physiol. B. doi 10.1007/s00360-012-
0705-4.
Fox, R. A., Ladage, L. D., Roth, T. C. & Pravosudov, V. V.
2009: Behavioural profile predicts dominance status in
mountain chickadees, Poecile gambeli. Anim. Behav. 77,
1441—1448.
Galef, B. G. & Wigmore, S. W. 1983: Transfer of informa-
tion concerning distant foods – a laboratory investiga-
tion of the information-center hypothesis. Anim.
Behav. 31, 748—758.
Grinevitch, L., Holroyd, S. L. & Barclay, R. M. R. 1995:
Sex-differences in the use of daily torpor and foraging
time by big brown bats (Eptesicus fuscus) during the
reproductive season. J. Zool. 235, 301—309.
Harcourt, J. L., Sweetman, G., Johnstone, R. A. & Manica,
A. 2009: Personality counts: the effect of boldness on
shoal choice in three-spined sticklebacks. Anim. Behav.
77, 1501—1505.
Hughes, R. N. 1997: Intrinsic exploration in animals:motives
andmeasurement. Behav. Process. 41, 213—226.
Huntingford, F. A. 1976: Relationship between anti-preda-
tor behavior and aggression among conspecifics in 3-
spined stickleback, Gasterosteus aculeatus. Anim. Behav.
24, 245—260.
Johnson, J. C. & Sih, A. 2005: Precopulatory sexual cannibal-
ism in fishing spiders (Dolomedes triton): a role for behav-
ioral syndromes. Behav. Ecol. Sociobiol. 58, 390—396.
Kavaliers, M., Colwell, D. D. & Choleris, E. 2005: Kinship,
familiarity and social status modulate social learning
about ‘‘micropredators’’ (biting flies) in deer mice.
Behav. Ecol. Sociobiol. 58, 60—71.
Kralj-Fiser, S., Weiss, B. M. & Kotrschal, K. 2010: Behavio-
ural and physiological correlates of personality in grey-
lag geese (Anser anser). J. Ethol. 28, 363—370.
Le Vin, A. L., Mable, B. K., Taborsky, M., Heg, D. &
Arnold, K. E. 2011: Individual variation in helping in a
cooperative breeder: relatedness versus behavioural
type. Anim. Behav. 82, 467—477.
Mainwaring, M. C., Beal, J. L. & Hartley, I. R. 2011: Zebra
finches are bolder in an asocial, rather than social, con-
text. Behav. Process. 87, 171—175.
McGhee, K. E. & Travis, J. 2010: Repeatable behavioural
type and stable dominance rank in the bluefin killifish.
Anim. Behav. 79, 497—507.
McGowan, A., Sharp, S. P., Simeoni, M. & Hatchwell, B. J.
2006: Competing for position in the communal roosts of
long-tailed tits. Anim. Behav. 72, 1035—1043.
Ethology 119 (2013) 189–198 © 2013 Blackwell Verlag GmbH 197
R. J. Kilgour & R. M. Brigham The Relationships between Behavioural Categories and Social Influences

Metcalfe, N. B., Huntingford, F. A. & Thorpe, J. E. 1987:
The influence of predation risk on the feeding motiva-
tion and foraging strategy of juvenile atlantic salmon.
Anim. Behav. 35, 901—911.
Metheny, J. D., Kalcounis-Rueppell, M. C., Willis, C. K.
R., Kolar, K. A. & Brigham, R. M. 2008: Genetic rela-
tionships between roost-mates in a fission-fusion society
of tree-roosting big brown bats (Eptesicus fuscus). Behav.
Ecol. Sociobiol. 62, 1043—1051.
Moretz, J. A., Martins, E. P. & Robison, B. D. 2007: Behav-
ioral syndromes and the evolution of correlated behav-
ior in zebrafish. Behav. Ecol. 18, 556—562.
Moscovice, L. R. & Snowdon, C. T. 2006: The role of social
context and individual experience in novel task acquisi-
tion in cottontop tamarins, Saguinus oedipus. Anim.
Behav. 71, 933—943.
O’Shea, T. J., Ellison, L. E., Neubaum, D. J., Neubaum, M.
A., Reynolds, C. A. & Bowen, R. A. 2010: Recruitment
in a Colorado population of big brown bats: breeding
probabilities, litter size, and first-year survival. J. Mam-
mal. 91, 418—428.
Parker, G. A. 1974: Assessment strategy and evolution of
fighting behavior. J. Theor. Biol. 47, 223—243.
Pike, T. W., Samanta, M., Lindstrom, J. & Royle, N. J.
2008: Behavioural phenotype affects social inter-
actions in an animal network. Proc. Biol. Sci. 275,
2515—2520.
Pintor, L. M., Sih, A. & Bauer, M. L. 2008: Differences in
aggression, activity and boldness between native and
introduced populations of an invasive crayfish. Oikos
117, 1629—1636.
Pretzlaff, I., Kerth, G. & Dausmann, K. H. 2010: Commu-
nally breeding bats use physiological and behavioural
adjustments to optimise daily energy expenditure. Na-
turwissenschaften 97, 353—363.
Pruitt, J. N., Demes, K. W. & Dittrich-Reed, D. R. 2011:
Temperature mediates shifts in individual aggressive-
ness, activity level, and social behavior in a spider.
Ethology 117, 318—325.
R Development Core Team. 2009: R: A Language and
Environment for Statistical Computing. R Foundation
for Statistical Computing, Vienna, Austria.
Racey, P. A. & Swift, S. M. 1981: Variations in gestation
length in a colony of pipistrelle bats (Pipistrellus pipistrel-
lus) from year to year. J. Reprod. Fertil. 61, 123—129.
Reaney, L. T. & Backwell, P. R. Y. 2007: Risk-taking
behavior predicts aggression and mating success in a
fiddler crab. Behav. Ecol. 18, 521—525.
Riechert, S. E. & Hedrick, A. V. 1993: A test for correlations
among fitness-linked behavioral traits in the spider age-
lenopsis-aperta (Araneae, Agelenidae). Anim. Behav.
46, 669—675.
Sih, A. & Bell, A. M. 2008: Insights for behavioral ecology
from behavioral syndromes. Adv. Study Behav. 38,
227—281.
Sih, A. & Watters, J. V. 2005: The mix matters: behaviour-
al types and group dynamics in water striders. Behav-
iour 142, 1417—1431.
Sih, A., Bell, A. & Johnson, J. C. 2004a: Behavioral syn-
dromes: an ecological and evolutionary overview.
Trends Ecol. Evol. 19, 372—378.
Sih, A., Bell, A. M., Johnson, J. C. & Ziemba, R. E. 2004b:
Behavioral syndromes: an integrative overview. Quart.
Rev. Biol. 79, 241—277.
Verbeek, M. E. M., Boon, A. & Drent, P. J. 1996: Explora-
tion, Aggressive Behavior and Dominance in Pair-wise
Confrontations of Juvenile Male Great Tits. Behaviour
133, 945—963.
Vervaecke, H., De Vries, H. & Van Elsacker, L. 1999: An
experimental evaluation of the consistency of competi-
tive ability and agonistic dominance in different social
contexts in captive bonobos. Behaviour 136, 423—442.
Via, S., Gomulkiewicz, R., Dejong, G., Scheiner, S. M.,
Schlichting, C. D. & Vantienderen, P. H. 1995: Adaptive
phenotypic plasticity – consensus and controversy.
Trends Ecol. Evol. 10, 212—217.
Willis, C. K. R. & Brigham, R. M. 2004: Roost switching,
roost sharing and social cohesion: forest-dwelling big
brown bats, Eptesicus fuscus, conform to the fission-
fusion model. Anim. Behav. 68, 495—505.
Willis, C. K. R., Brigham, R. M. & Geiser, F. 2006: Deep,
prolonged torpor by pregnant, free-ranging bats. Natur-
wissenschaften 93, 80—83.
Wilson, D. S. 1998: Adaptive individual differences within
single populations. Philos. Trans. R. Soc. Lond., B, Biol.
Sci. 353, 199—205.
Wilson, E. O. 2000: Sociobiology: The New Synthesis. Har-
vard University Press, Cambridge, MA.
Wilson, D. S., Clark, A. B., Coleman, K. & Dearstyne, T.
1994: Shyness and boldness in humans and other ani-
mals. Trends Ecol. Evol. 9, 442—446.
Wilson, A. D.M.,Whattam, E.M., Bennett, R., Visanuvimol,
L., Lauzon, C. & Bertram, S.M. 2010: Behavioral correla-
tions across activity, mating, exploration, aggression, and
antipredator contexts in the European house cricket,Ach-
eta domesticus. Behav. Ecol. Sociobiol. 64, 703—715.
Supporting Information
Additional supporting information may be found in
the online version of this article:
Table S1: The protocol used to assess learning abil-
ity in bats. If a bat took longer than 3 min at any stage
in the training procedure, it was considered a “mis-
trial” and was returned to the previous step.
Table S2: Description of how bats were assigned
learning scores.
Table S3: Description of how bats were scored for
latency to feed in captivity.
Ethology 119 (2013) 189–198 © 2013 Blackwell Verlag GmbH198
The Relationships between Behavioural Categories and Social Influences R. J. Kilgour & R. M. Brigham