universidade federal do amazonas - início loebens final.pdf · universidade federal do amazonas...
TRANSCRIPT

UNIVERSIDADE FEDERAL DO AMAZONAS
FACULDADE DE CIÊNCIAS AGRÁRIAS
DEPARTAMENTO DE CIÊNCIAS PESQUEIRAS
ENGENHARIA DE PESCA
COMPOSIÇÃO E ESTRUTURA TRÓFICA DAS ASSEMBLEIAS DE PEIXES DA
FLORESTA DE IGAPÓ NO PARQUE NACIONAL DE ANAVILHANAS, AMAZONAS,
BRASIL.
Sara de Castro Loebens
MANAUS
2016

UNIVERSIDADE FEDERAL DO AMAZONAS
FACULDADE DE CIÊNCIAS AGRÁRIAS
DEPARTAMENTO DE CIÊNCIAS PESQUEIRAS
ENGENHARIA DE PESCA
COMPOSIÇÃO E ESTRUTURA TRÓFICA DAS ASSEMBLEIAS DE PEIXES DA
FLORESTA DE IGAPÓ NO PARQUE NACIONAL DE ANAVILHANAS, AMAZONAS,
BRASIL.
DISCENTE: Sara de Castro Loebens
ORIENTADOR:Dra. Kedma Cristine Yamamoto
MANAUS
2016
Monografia apresentada ao
Departamento de Ciências
Pesqueiras da Faculdade de
Ciências Agrárias como quesito
parcial para obtenção do grau de
Bacharel em Engenharia de Pesca
pela Universidade Federal do
Amazonas.

Membros da banca:
1. Dra. Cláudia Pereira de Deus
2. Dr. Cleber Duarte
3. Dra. Kedma Cristine Yamamoto
Prof. Dr. Bruno Adam SagratzkiCavero
Chefe do Departamento
Prof. Dr. Pedro Roberto de Oliveira
Coordenador de monografia
Profa. Dra. Sanny Maria de Andrade Porto
Coordenador do Curso de Engenharia de Pesca

Ao planeta Terra

“Eu dou um recado para vocês brancos, para que não poluam os rios. Por que isso
vai afetar o futuro não só dos índios, mas dos filhos de vocês também.”
AriatanaYawalapiti, líder do povo Yawalapiti.
“Não digo: eu descobri essa terra porque meus olhos caíram sobre ela, portanto a
possuo. Ela existe desde sempre, antes de mim.”
Davi Yanomami, pajé e líder do povo Yanomami

Agradecimentos
Agradeço primeiramente a Deus, pela força, proteção, por estar sempre comigo e
por me dar a vida;
Agradeço a minha família por ser meu apoio e alicerce, sem o qual não seria quem
sou e nem chegaria onde estou;
Agradeço ao meu amor Daniel Brandt Galvão por ser companheiro, carinhoso,
amigo e confidente, por estar ao meu lado em tudo e sempre me motivar, por fazer
parte da minha vida;
Agradeço a Pachamama (Mãe Terra) por ser incrível e trazer paz aos olhos e
também ao coração, eu luto por você;
A Amazônia por ser o paraíso, minha terra, meu quintal, meu mestre, e por possuir o
Rio Negro (o rio mais lindo do mundo) e Anavilhanas, meu local de estudo, minha
paixão, e todos os lugares incríveis que já conheci e que espero conhecer um dia;
Ao CNPq (Conselho Nacional de Desenvolvimento Cientifico e Tecnológico) e
FAPEAM (Fundação de Amparo à Pesquisa do Estado do Amazonas) pelo apoio
financeiro;
Ao ICMBio (Instituto Chico Mendes de Conservação da Biodiversidade) por apoiar
esse projeto e sempre ser solícito as nossas coletas;
A Universidade Federal do Amazonas, Faculdade de Ciências Agrárias,
Departamento de Ciências Pesqueiras, pela oportunidade de estudo;
A professora Kedma Yamamoto pelos ensinamentos, paciência, conselhos,
orientação, confiança e principalmente por ser para a figura do que busco ser um
dia, sou sua fã, por ter me proporcionado momentos indescritíveis na universidade e
principalmente em Anavilhanas, local de estudo e de paz. Obrigada mesmo
professora!;
Ao professor Carlos Edwar pela ajuda essencial e ensinamentos sobre estatística;
A todos os professores, meus mestres, que me ensinaram e me formaram;
Ao professor Jansen Zuanon pela ajuda na identificação dos peixes, ensinamentos e
por sempre ser solicito mesmo muito ocupado;
Aos técnicos que também foram meus mestres nessa jornada;
Ao Manoel do Laboratório de Botânica Agro florestal pela identificação dos frutos;
Aos professores Ronis da Silveira e Rafael de Fraga pela identificação e
ensinamentos sobre ofídios;

Aos meus parceiros do Laboratório de Ictiologia que me apoiaram e ajudaram muito
para que esse trabalho fosse concluído;
A todos os meus amigos de dentro e fora da UFAM que sempre estiveram ao meu
lado, me apoiaram, me ajudaram nos momentos difíceis e de alegria, me fizeram rir
muito e ter momentos inesquecíveis;
A todos que ajudaram no trabalho em campo, Jairo, Ilgner, professor Álvaro, Ray.
Ao técnico Ivanildo e o piloteiro Valdeci pela ajuda em campo e sem a qual esse
trabalho não seria possível, pela amizade, pelas histórias e lendas compartilhadas e
que levaram a muitas gargalhadas e pela paciência com a Sarinha;
Aos seguranças da Base I – Lago Prato, que eram parceria certa e apoiavam em
tudo durante as coletas;
As tias da ADAP por limpar o laboratório que ficava daquele jeito depois das
análises;
Agradeço aos doutores Cleber Duarte e Cláudia de Deus pela disponibilidade e por
aceitarem fazer parte da banca examinadora;
A todos que direta e indiretamente me ajudaram e apoiaram, podem ter certeza que
estarão sempre em meu coração.

RESUMO
A floresta de igapó é um dos biótopos formados na bacia Amazônica de grande
importância para os peixes uma vez que são utilizadas para abrigo e alimentação
por esses organismos. Com o intuito de caracterizar a as assembleias de peixes da
floresta de igapó quanto a sua composição e estrutura trófica foram realizadas
amostragens em quatro lagos do Parque Nacional de Anavilhanas, Amazonas,nos
meses de abril e setembro de 2014, e fevereiro de 2015. Foram coletados ao todo
931 indivíduos, distribuídos em 4 ordens, 20 famílias, 48 gêneros e 65 espécies. A
diversidade de Shannon-Wiener e a riqueza de Margalef foram maiores no ambiente
de floresta alagada. Para as amostras analisadas os índices de Berger-Parker e
Equitabilidade indicaram baixa dominância e alta homogeneidade no número de
indivíduos por espécie. Os resultados da perMANOVA demonstraram diferenças
significantes entre a composição da floresta de igapó e da água aberta.De acordo
com o grau de repleção, os exemplares apresentaram a maior parte dos estômagos
pouco cheios (0%-25%) e totalmente cheios (75%-100%). Os itens alimentares mais
abundantes foram: peixes, insetos, crustáceos e microcrustáceos, material vegetal,
frutos e sementes, material de origem animal e vegetal. O igapó apresentou alta
diversidade, e maior quantidade de espécies que a água aberta. A classificação em
guildas tróficas evidenciou a importância da floresta alagada na composição das
espécies ali presentes.
Palavras-chave: Rio Negro, diversidade, floresta alagada, alimentação, guildas
tróficas.

ABSTRACT
The igapó forest is one of the biotope formed in the Amazon basin of great
importance for the fish since they are used for shelter and food for these organisms.
In order to characterize the fish assemblages of igapó forest as their composition and
trophic structure samples were taken at four lakes in the Anavilhanas National Park,
Amazon, in April and September 2014 and February 2015. Were collected in all 931
individuals, distributed in 4 orders, 20 families, 48 genera and 65 species. The
diversity of Shannon-Wiener and richness of Margalef were higher in the flooded
forest environment. For samples analyzed the Berger-Parker indices and Equitability
indicated low dominance and high homogeneity in the number of individuals per
species. The results of PERMANOVA showed significant differences between the
composition of igapó forest and open water. According to the degree of fullness, the
specimens showed most bit full stomachs (0% -25%) and fully filled (75% -100%).
The most abundant food items were: fish, insects, crustaceans and
microcrustaceans, plant material, fruit and seeds, animal and plant material. The
igapó showed high diversity, and more species than open water. The classification of
trophic guilds highlighted the importance of flooded forest in the composition of the
species present there.
Keywords: Rio Negro, diversity, flooded forest, feeding, trophic guilds.

SUMÁRIO
1. INTRODUÇÃO .................................................................................................... 13
2. REFERENCIAL TEÓRICO .................................................................................. 15
2.1 O Rio Negro ....................................................................................................... 15
2.2 Diversidade das assembleias de peixes ............................................................ 17
2.3 Diversidade trófica ............................................................................................. 19
3. OBJETIVOS ........................................................................................................ 20
4. HIPÓTESES ........................................................................................................ 20
5. MATERIAL E MÉTODOS .................................................................................... 20
5.1 Área de estudo e amostragens ..................................................................... 20
5.2 Análise da diversidade de espécies .............................................................. 22
5.3 Análise da composição alimentar ................................................................. 23
5.4 Análise estatística ......................................................................................... 25
6. RESULTADOS E DISCUSSÃO ........................................................................... 26
6.1 Diversidade das assembleias ....................................................................... 26
6.2 Estrutura trófica das assembleias de peixes da floresta de igapó ................. 34
7. CONCLUSÕES ................................................................................................... 43
8. REFERÊNCIAS BIBLIOGRÁFICAS .................................................................... 44
APÊNDICE ................................................................................................................. 55

Lista de Figuras
Figura 1. Localização dos lagos estudados (Prato, Arraia, Canauiri Grande e
Canauiri Pequeno) e do Parque Nacional de Anavilhanas. ....................................... 21
Figura 2. Porcentagem do número de indivíduos dentro de cada ordem para floresta
de igapó e água aberta dos lagos. ............................................................................ 26
Figura 3. Porcentagem do número de espécies dentro de cada ordem para floresta
de igapó e água aberta dos lagos. ............................................................................ 27
Figura 4. Porcentagem do número de indivíduos por famílias capturadas na floresta
de igapó e água aberta dos lagos estudados. ........................................................... 28
Figura 5. Porcentagem do número de espécies presentes em cada família para a
floresta de igapó e água aberta dos lagos estudados. .............................................. 29
Figura 6. Abundância relativa do número de indivíduos das dez espécies mais
abundantes na floresta de igapó e água aberta dos lagos. ....................................... 30
Figura 7. Curvas de rarefação e o seu respectivo desvio padrão para os ambientes
de floresta de igapó (A) e água aberta (B) no PARNA Anavilhanas. ........................ 31
Figura 8. Ordenação da composição taxonômica utilizando os dois primeiros eixos
gerados pela nMDS para os dados de abundância das amostras de peixes da
floresta de igapó e água aberta. Legenda: Floresta de Igapó (F.I.); Água Aberta
(A.A.); coletas (1, 2, 3). FCG1=F. I., C. Grande, col. 1; FCG2=F. I., C. Grande, col. 2;
FCG3=F. I., C. Grande, col. 3; FCP1=F. I., C. Pequeno, col. 1 FCP2=F.I. C.
Pequeno, col. 2; FCP3=F.I. C. Pequeno, col. 3; FA1=F.I., Arraia, col. 1; FA2=F.I.,
Arraia, col. 2; FA3=F.I., Arraia, col. 3; FP1=F.I., Prato, col. 1; FP2=F.I., Prato, col. 2;
FP3=F. I., Prato, col. 3; ACG1=A.A.; C. Grande, col. 1; ACG2=A.A.; C. Grande, col.
2; ACG3=A.A.; C. Grande, col. 3; ACP1=A.A.; C. Pequeno, col. 1; ACP2=A.A.; C.
Pequeno, col. 12; ACP3=A.A.; C. Pequeno, col. 3; AA1=A.A., Arraia, col.1; AA2
=A.A., Arraia, col.2; AA3=A.A, Arraia, col.3; AP1=A.A., Prato, col. 1; AP2=A.A.,
Prato, col. 3; AP3=A.A., Arraia, col. 3........................................................................ 33

Figura 9. Frequência relativa (%) das categorias tróficas encontradas para as
espécies analisadas pertencentes a floresta de igapó do PARNA Anavilhanas. ...... 36
Figura 10. Índice de importância alimentar (IAi%) para a espécie carnívora mais
abundante P. flavipinnis. ........................................................................................... 37
Figura 11. Índice de importância alimentar (IAi%) para as espécies insetívoras mais
abundantes T. intermedia, T. elongatuse A. longimanus. ........................................ 37
Figura 12. Índice de importância alimentar (IAi%) para a espécie onívora mais
abundante H. immaculatus. ....................................................................................... 38
Figura 13. Índice de importância alimentar (IAi%) para as espécies piscívoras mais
abundantes A. polystictus, S. gouldingi e S. rhombeus. ............................................ 39
Figura 14, Índice de importância alimentar (IAi%), para a espécie zooplanctívora
mais abundante H. edentatus. ................................................................................... 40
Figura 15. Índice de importância alimentar (IAi%), para as espécies herbívoras mais
abundantes M. hypsauchen e M. asterias. ................................................................ 40
Figura 16. Dendrograma da similaridade da dieta dos peixes analisados com valores
de IAi (%) presentes na floresta de igapó do PARNA Anavilhanas. .......................... 41

Lista de tabelas
Tabela 1. Descrição dos aspectos externos para estimativa do grau de repleção dos
estômagos com os respectivos pontos atribuídos (BENNEMANN 1985; YABE;
BENNEMANN 1994). ................................................................................................ 24
Tabela 2. Parâmetro de diversidade calculados para diferentes biótopos do rio
Negro. S = riqueza de espécies; N = número de indivíduos; H’ = diversidade de
Shannon-Wiener; J’ = equitabilidade. ........................................................................ 32

13
1. INTRODUÇÃO
A Região Amazônica possui uma bacia hidrográfica de aproximadamente 7,9
milhões de km2 e no pico da cheia pode inundar algo em torno 0,3 milhão de km2 por
ano sendo a sazonalidade pluvial responsável pela flutuação no nível dos rios
(SAINT-PAUL et al., 2000). Assim, a bacia amazônica é considerada um ambiente
complexo sendo formada por inúmeros biótopos como igarapés, planícies
inundáveis, lagos e florestas alagadas (SIOLI, 1984; LOWE-McCONNELL, 1999).
Esse número de habitats disponível é responsável pela grande quantidade de
espécies de peixes (JUNK, 1984), aproximadamente 2400 (LÉVÊQUE et al., 2008).
Carvalho et al. (2007) sugerem que a imensa área de drenagem da bacia podem
abrigar ainda mais, algo em torno de 3000 espécies.
Durante os períodos de enchente e cheia, surgem biótopos importantes para
os peixes, como a floresta alagada (LOW-McCONNELL,1999; JUNK, 1984;
LOEBENS et al., 2016) e bancos de macrófitas aquáticas (ARAÚJO-LIMA et al.,
1986; PRADO et al., 2010; SOARES et al., 2014). As florestas alagadas podem ser
classificadas em igapó ou várzea, de acordo com as características da água onde
são encontradas (PRANCE, 1980; PIEDADE et al., 2015). Os rios de águas pretas
originam as florestas de igapó, que são oriundas dos escudos guianenses,
altamente lixiviadas, com pH ácido devido as concentrações de substâncias
orgânicas dissolvidas e baixa carga de nutrientes (SIOLI, 1984). Os rios de águas
brancas, que originam as florestas de várzea, são oriundas da região Andina ainda
sofrendo processos erosivos e carreando sedimentos, apresentam pH relativamente
neutro e elevadas cargas de nutrientes (SIOLI, 1984).
Afloresta de igapó possui solo arenoso, sustentando assim uma vegetação
muito mais pobre do que a encontrada no ambiente de várzea (PRANCE,
1980).Entretanto, esse ambiente apresenta grande importância para os peixes uma
vez que servem de local de refúgio contra predadores e para a alimentação, através
de frutos, sementes e insetos (GOULDING, 1980; LOWE-McCONNELL, 1999;
RODRIGUEZ; LEWIS, 1997; ADIS, 1999; CLARO-Jr., 2004; CORREA et al., 2008).
Poucos são os estudos realizados acerca da estruturação de assembleias de
peixes em ambientes alagáveis de águas pretas. Noveras et al. (2012) estudaram o
uso do igapó pelas assembleias de peixes durante o dia e a noite no Parque
Nacional de Anavilhanas, Saint-Paul et al. (2000),na mesma área, realizaram um

14
comparativo entre as florestas de várzea e igapó, encontrando maior diversidade e
menor CPUE existente no ambiente de água preta. Loebens et al. (2016) também
evidenciaram a diversidade das assembleias de peixes presentes no ambiente de
igapó.
A baixa concentração de íons orgânicos no rio Negro faz com que esse não
seja um ambiente propício ao desenvolvimento de altas biomassas de fitoplâncton e
macrófitas aquáticas, como na água branca, demonstrando assim a importância da
floresta na cadeia alimentar (KENSLEY; WALKER, 1982; GOULDING et al., 1988;
WALKER, 1990). Goulding et al. (1988) observaram que a energia que sustenta as
comunidades de peixes no sistema de águas pretas são oriundas principalmente das
florestas alagadas uma vez que esses organismos utilizam a matéria disponibilizada
nesse ambiente para alimentação. Diante disso, a presença de alimentos de origem
alóctone é de grande importância como já relatado em estudos realizados no
sistema amazônico (GOULDING, 1980; GOULDING et al., 1988; CLARO-Jr et al.,
2004; NOVERAS et al., 2012). Entretanto, a organização trófica do ecossistema e
dos mecanismos de interação entre as espécies de peixes ainda representam um
dos aspectos ecológicos menos estudados (PAZIN, 2004).
Dentre os ambientes que compõem a bacia do rio Negro, destaca-se o
Parque Nacional de Anavilhanas (PARNA Anavilhanas), mencionado em alguns
estudos como sendo um ambiente importante para a manutenção da diversidade
íctica (ARAUJO-LIMA et al., 1986; GARCIA, 1995; SAINT-PAUL et al., 2000;
NOVERAS et al., 2012; YAMAMOTO et al., 2014; LOEBENS et al., 2016). O parque
é uma Unidade de Conservação (UC) de uso indireto, não sendo permitida assim a
utilização de seus recursos a não ser para fins didáticos e científicos (SNUC, 2000).
As UC`s tem mostrado um papel importante na conservação de recursos e
manutenção da biodiversidade (AGOSTINHO et al., 2005). Além disso, por se
localizar em uma região intermediária do rio Negro, o arquipélago pode ser utilizado
como passagem por diversas espécies de peixes, fornecendo abrigo e alimento
(GOULDING et al., 1988).
Com o intuito de gerar informações para conhecer a diversidade
ictiofaunística da floresta de igapó do PARNA Anavilhanas e sua dinâmica de
interação com este biótopo, este estudo propôs caracterizar a diversidade,
composição e estrutura trófica das assembleias de peixes presentes na floresta de
igapó em lagos do PARNA Anavilhanas. Nossos resultados poderão subsidiar

15
informações que possam auxiliar no plano de manejo da UC,além de enriquecer os
conhecimentos sobre as assembleias de peixes formadas em ambientes alagados
de águas pretas.
2. REFERENCIAL TEÓRICO
2.1 O Rio Negro
O rio Negro é o maior tributário do rio Amazonas, possuindo cerca de
1.700Km (GOULDING et al., 1988). Apresenta águas escuras, pobres em nutriente,
pH baixo e rico em ácidos húmicos e taninos, solubilizados durante o processo de
decomposição da biomassa que é produzida pela floresta de igapó o que caracteriza
sua coloração (GOULDING et al., 1988; SIOLI, 1965). É formado a partir dos
escudos guianenses, mais antigos e altamente lixiviados (SIOLI, 1965), carreando
poucos sedimentos, resultado do desgaste das rochas (PIEDADE et al., 2015).
O sistema hidrológico do rio Negro é caracterizado pela variabilidade de
biótopos, praias arenosas, corredeiras, remansos, ilhas, paranás, lagos, igarapés e
florestas alagadas (SIOLI, 1984; LOWE-McCONNELL, 1999). Com isso, durante os
períodos de águas altas possibilitam uma maior disponibilidade de área, alimento e
abrigo para a ictiofauna, influenciando diretamente na ecologia alimentar das
espécies (SANTOS, 1981; ALMEIDA,1984; SOARES et al., 1986; CLARO-Jr, 2004;
YAMAMOTO, 2004).
Caracterizado como um rio pouco produtivo, pela baixa concentração de
nutrientes dissolvidos, o rio Negro apresenta ictiofauna rica e diversificada,
abrangendo cerca de 450 espécies, das quais provavelmente 30% ainda não foram
registradas (GOULDING et. al., 1988).Soares e Yamamoto (2005) não consideram
as características químicas da água preta fatores limitantes para a diversidade, e
apontam a necessidade do incremento de amostragens nos biótopos desse
ambiente. Da mesma maneira que nas várzeas, a variabilidade de biótopos
existentes nas águas pretas também sofre influência da variação sazonal,
decorrente da flutuação do nível do rio (GOULDING et al., 1988; JUNK etal., 1989;
SAINT-PAUL etal., 2000).

16
Estudos realizados com assembleias de peixes em ambientes de água preta
abordam aspectos de diversidade e relações tróficas (GOULDING etal., 1988;
NOVERAS etal., 2012), distribuição de peixes (GARCIA, 1995; SOARES;
YAMAMMOTO, 2005), assembleias presentes em floresta de igapó (SAINT-PAUL
etal., 2000, NOVERAS etal., 2012; LOEBENS etal., 2016),interações com bancos de
macrófitas aquáticas (ARAÚJO-LIMA, 1986), atividades migratórias (RIBEIRO, 1983;
RIBEIRO; PETRERE, 1990), e taxonomia e genética (MAIA; ALVES-GOMES, 2012;
FERNANDES et al., 2015).
No sistema do rio Negro se encontra o Parque Nacional de Anavilhanas
(PARNA Anavilhanas), uma unidade de conservação federal no município de Novo
Airão, a cerca de 40 km da cidade de Manaus, que possui um complexo de ilhas que
têm sua formação a partir da deposição de sedimentos oriundos do rio Branco
(LEENHEER; SANTOS, 1980). O PARNA apresenta uma área de 335.018 ha com
aproximadamente 400 ilhas (OLIVEIRA, 2003), sendo considerado o segundo maior
arquipélago fluvial do mundo. Delimitada por rios de águas pretas e ácidas, as ilhas
apresentam formato alongado distintas das encontradas nas áreas de várzea,
entretanto também sofrendo forte influência do ciclo de enchentes e vazantes do rio
Negro estando assim em constante processo de erosão e sedimentação
(LEENHEER; SANTOS, 1980; FITTKAU, 1964). A vegetação circundante dos lagos
é composta por florestas inundáveis denominadas de floresta de igapó estacional
(PRANCE, 1980).
O PARNA foi criado pela Lei nº. 11.799, de 29 de outubro de 2008, a partir da
Estação Ecológica de Anavilhanas, criada pelo Decreto nº 86.061, de 2 de junho de
1981. Na época em que o parque foi criado ali residiam cinquenta e três
comunidades que tiveram a área desapropriada e se instalaram no município de
Novo Airão, ao redor do parque (BARRETO FILHO, 2001).
Devido a sua localização e potencial, o complexo de ilhas de Anavilhanas
tornou-se foco de estudos voltados à ecologia, diversidade e taxonomia de peixes
(FERREIRA, 1981; GOULDING; CARVALHO, 1982; ARAÚJO-LIMA et al., 1986;
GOULDING et al., 1988; ZUANON, 1993; GARCIA 1995; SAINT-PAUL et al., 2000;
NOVERAS et al., 2012; MAIA; ALVES-GOMES, 2012; FERNANDES et al., 2015,
LOEBENS etal., 2016), distribuição e abundância ictioplanctônica (OLIVEIRA, 2003)
e manutenção da ictiofauna através de atratores artificiais (YAMAMOTO et al.,
2014).

17
2.2 Diversidade das assembleias de peixes
A organização e compreensão dos mecanismos e processos de
caracterização das assembleias de peixes são pressupostos para estudos sobre a
ecologia de comunidades (ANGERMEIER; KARR, 1984). Estudos sobre as relações
entre as variações ambientais e estratégias de alimentação, reprodução, dispersão e
interação das espécies com o ambiente têm tido sucesso em compreender padrões
ecológicos (GOLDSTEIN; MEADOR, 2004).
O entendimento dos ecossistemas aquáticos parte do pressuposto da
existência de uma interação aquática e terrestre, onde os organismos coexistem a
partir do ambiente físico, químico e biológico, determinado em biomassa, energia e
características biológicas (CROPP; GABRIC, 2002). As assembleias ícticas
funcionam como indicadoras da qualidade ambiental, refletindo o estado biótico e
abiótico de seu ambiente (KARR, 1981; ARAÚJO, 1998; VIEIRA; SHIBATTA, 2007).
Sendo assim, o estudo de comunidades envolve uma síntese dos fatores ambientais
e das interações bióticas, além disso, a estrutura de uma comunidade pode ser
analisada sobre vários aspectos, utilizando-se parâmetros como diversidade, riqueza
e equitabilidade (WOOTTON, 1995).
A grande variedade de espécies, formas e estratégias da ictiofauna da região
amazônica é oriunda de fatores bióticos e abióticos (JUNK et al., 1989) aliados a
grande diversidade de habitats (SIOLI, 1984; LOWE-McCONNELL, 1999). Podemos
citar fatores como predação e competição (RODRIGUEZ; LEWIS, 1997; JAKSON et
al, 2001; SIQUEIRA-SOUZA; FREITAS, 2004), características físico-quimicas, como
oxigênio dissolvido (JUNK et al., 1983), transparência e profundidade da água
(RODRIGUEZ; LEWIS, 1997; TEJERINA-GARRO et al., 1998), complexidade
estrutural (ARRINGTON et al., 2005; ARRINGTON; WINEMILLER, 2006; JUNK et
al., 1989), dificuldade exploratória e pulsos de inundação (JUNK, 1989).
As variações no nível da água na Amazônia Central fazem com que a
ictiofauna necessite se adaptar às profundas modificações que ocorrem no ambiente
(JUNK et al., 1989). Os biótopos criados pela inundação e os movimentos dos
peixes dentro das florestas alagadas promovem um rearranjo na dinâmica das
comunidades (CORREA et al., 2008).
Segundo Junk et al. (1989), planícies inundáveis são áreas periodicamente
alagadas, resultando em condições físicas e químicas que levam a biota a responder

18
através de adaptações, produzindo estruturas de comunidades características
desses sistemas. Esses ambientes são classificados assim como ecótonos ou zona
de transição entre o ambiente aquático e terrestre (ATTZ) (JUNK et al., 1989). Em
áreas alagáveis, o ecótono água-terra representa um importante componente para a
conservação das comunidades de peixes de água doce. Na interface destes
ambientes ocorre grande quantidade de habitats servindo assim de abrigo, local
para desova e alimentação para os peixes, sendo responsáveis pela manutenção da
abundância e diversidade desses organismos (HENRY, 2003).
Noveras et al. (2012), em estudo sobre assembleias de peixes em igapó de
Anavilhanas, encontraram maior diversidade nesse ambiente em relação ao
ambiente de água aberta dos lagos. Saint-Paul et al. (2000) também reportam
valores de riqueza superiores em florestas alagadas de água branca e preta em
relação ao ambiente de água aberta. Essa diferença entre os ambientes pode ser
explicada pela ATTZ que dispõe de refúgio e alimentação para os peixes, formando
assim vários nichos ecológicos (JUNK et al., 1989).
Diante de tais informações, faz-se necessária a análise detalhada das
características da estrutura das assembleias de peixes (SAINT-PAUL, 2000;
NOVERAS et al., 2012; SOARES; YAMAMOTO, 2005) através da utilização de
índices de diversidade (MARTINS; SANTOS, 1999). Os índices são usados como
descritores da diversidade (HURLBERT, 1971; MAGURRAN, 1988) e demonstram a
relação entre as espécies existentes e o ambiente (MARTINS; SANTOS, 1999).
Segundo Magurran (1988), a diversidade pode ser medida pelo número de
espécies e distribuição da abundância relativa. Ricklefs (1990), afirmou que a
diversidade demonstra o número de espécies em uma área, a variedade de
organismos, proveniente da substituição de espécies entre hábitats, e representa em
uma comunidade a abundância relativa de cada espécie. O número de espécies
encontrado seria a mais simples forma de medida de diversidade, entretanto, essa
classificação depende totalmente do tamanho da amostra (MARTINS; SANTOS,
1999), ou seja, a diversidade deve relacionar a variação de espécies quanto ao
número de indivíduos da amostra dentro de um intervalo determinado (MAY, 1975).

19
2.3 Diversidade trófica
Na Amazônia Central, com a flutuação média anual do nível d’água de 10
metros, e alternância das fases terrestre e aquática, ocorrem modificações drásticas
nas condições ambientais fazendo com que a ictiofauna tenha adaptações
específicas (JUNKet al., 1989). O aumento da área alagada durante o período da
cheia favorece a exploração de vários habitats incrementando o espectro alimentar
dos peixes (SANTOS, 1981; ALMEIDA, 1984; SOARES et al, 1986; YAMAMOTO,
2004; CLARO-Jr et al., 2004). Florestas inundadas são fonte de uma grande
variedade de recursos alimentares alóctones, como plantas terrestres, folhas, flores,
sementes, frutos, perifíton e invertebrados (GOULDING,1980).
Segundo Silva et al. (2008), estudos sobre a dieta de peixes e a relação desta
com o meio fornecem informações ecológicas importantes. Essas informações
trazem subsídios para a compreensão dos mecanismos que permitem a
coexistência e a exploração dos recursos por várias espécies em um mesmo
sistema (GOULDING, 1980). Claro-Jr et al. (2004), em estudo realizado na floresta
alagada em lagos de várzea da Amazônia Central demonstraram a relação direta de
três espécies onívoras com esse ambiente. Noveras et al. (2012), em estudo
realizado em floresta de igapó em Anavilhanas, demonstraram a utilização desse
ambiente para alimentação e distribuição de assembleias de peixes ali presentes.
O processo de frutificação das árvores, que ocorre durante o período da
cheia, faz com que os frutos sejam dispersos através da água para outros locais,
participando de forma importante na alimentação dos peixes (GOULDING, 1980). Os
invertebrados também têm uma importante participação na dieta do peixe e estão
presentes o ano inteiro, embora estejam mais disponíveis na época das cheias
(ZAVALA-CAMIN, 1996; ADIS, 1997; JUNK et al., 1997).
Segundo Abelha et al. (2001), a grande variedade de alimentos presentes na
dieta de peixes tropicais demonstra as características oportunistas desses
organismos principalmente em rios sazonais (GOULDING, 1980; LOWE-
McCONNELL, 1999; SANTOS; FERREIRA, 1999),demonstrando que a dieta dos
peixes é regulada principalmente pela dinâmica do pulso de inundação (JUNK et al.,
1989).

20
3. OBJETIVOS
Geral:
Caracterizar as assembleias de peixes na floresta de igapó do Parque
Nacional de Anavilhanas quanto a sua diversidade e estrutura trófica.
Específicos:
Identificar a composição taxonômica das assembleias de peixes
presentes na floresta de igapó no PARNA Anavilhanas;
Estimar a diversidade e abundância numérica das assembleias de
peixes da floresta de igapó e compará-la com a área aberta dos lagos;
Determinar a dieta das assembleias de peixes da floresta de igapó;
Identificar as guildas tróficas de peixes presentes na floresta de igapó.
4. HIPÓTESES
H0,1: A composição das assembleias de peixes da floresta de igapó é semelhante à
da água aberta dos lagos.
H1,1: A composição das assembleias de peixes da floresta de igapó difere da água
aberta dos lagos.
5. MATERIAL E MÉTODOS
5.1 Área de estudo e amostragens
As amostragens foram realizadas nos anos de 2014 e 2015 no Parque
Nacional de Anavilhanas,nos lagos Prato, Arraia, Canauiri Grande e Canauiri
Pequeno (Figura 1). Esses quatro lagos são historicamente estudados e possuem
localização chave, uma vez que os lagos Canauiri Pequeno e Arraia estão mais

21
próximos do canal principal, e os lagos Canauiri Grande e Prato se encontram mais
a dentro do parque.
O presente estudo trabalhou com a base de dados do projeto “A influência de
atratores artificiais nas assembleias de peixes do Médio Rio Negro,
Amazonas/BR”Edital PPP/FAPEAM para os dados de floresta de igapó.
Figura 1. Localização dos lagos estudados (Prato, Arraia, Canauiri Grande e Canauiri
Pequeno) e do Parque Nacional de Anavilhanas.
As coletas foram realizadas nos meses de abril (enchente) e setembro
(vazante) de 2014, e fevereiro (enchente) de 2015, períodos onde havia a floresta
alagada, sob a autorização n0 24518-2 do Instituto Chico Mendes de Conservação
da Biodiversidade – ICMBio. As pescarias experimentais foram realizadas através de
baterias de malhadeiras (rede-de-espera) com tamanhos de malha variando de 30 a
110 mm entre nós opostos (2,5 metros de profundidade x 25 metros de
comprimento), dispostas aleatoriamente no igapó e água aberta. As malhadeiras
foram armadas ao amanhecer, sendo expostas por um período de duas horas (06h
às 08h), seguido da despesca e, outra captura ao anoitecer (19h às 21h). Após a
despesca os peixes foram etiquetados, fixados em formol (10%) e transportados ao
laboratório de Ictiologia na UFAM. No laboratório foram pesados, medidos e lavados

22
em água corrente para serem preservados em álcool (70%) e identificados através
de chaves de identificação (FERREIRA et al., 1998, GÈRY, 1977) e com a ajuda de
especialistas.
5.2 Análise da diversidade de espécies
Para estimar a riqueza específica da comunidade capturada foi calculado o
índice de Margalef (LUDWIG; REYNOLDS, 1988), que analisa a relação entre o
número total de espécies e o número total de indivíduos observados, na fórmula a
seguir:
Onde:
S = Número total de espécies na amostra;
n = Número total de indivíduos na amostra.
Entre os índices de diversidade propostos com base na dominância de uma
ou mais espécies, escolhemos o índice de Berger-Parker (BERGER; PARKER,
1970) que representa a dominância de uma espécie na estrutura da comunidade, de
acordo com a fórmula.
Onde:
nmax= número de indivíduos da espécie mais abundante; e
N= número de indivíduos presentes na amostra.
Para estimar a diversidade foi calculado o índice de Shannon-Wiener
(SHANNON; WIENER, 1949):
Onde:
e
ni= número de indivíduos da espécie i,
N= número de indivíduos presentes na amostra.
n
SR
ln
11
Nn
d max
)(ln' ii PPH
Nn
p ii

23
Foi calculada também a Equitabilidade (MAGURRAN, 1988), que expressa à
maneira pela qual o número de indivíduos está distribuído entre as espécies,
representando uma situação onde todas as espécies são igualmente abundantes:
Onde:
H’ é o índice de diversidade de Shannon;
S é o número de espécie presentes na amostra.
A riqueza potencial de espécies foi estimada pela curva de Rarefação
(KREBS, 1989).
5.3 Análise da composição alimentar
No laboratório de Ictiologia da Universidade Federal do Amazonas foi
realizada a análise da dieta dos peixes. Através de uma incisão ventral nos peixes,
os estômagos foram retirados e pesados. Os estômagos retirados dos tratos
digestivos foram examinados sob microscópio estereoscópio e os itens alimentares
identificados até o nível taxonômico mais inferior possível.
A determinação da dieta foi efetuada através da análise do conteúdo
estomacal, utilizando os métodos da frequência de ocorrência (F.O) e do volume
relativo proposto por Hynes (1950). A F. O. foi calculada conforme a fórmula:
F.O = i.100/ N
Onde:
F.O.= Frequência de ocorrência (%),
i = Número de estômagos em que cada item alimentar foi identificado,
N = Número de estômagos analisados com alimento.
SHE ln'

24
O volume relativo é a estimativa visual do volume de cada item alimentar em
relação ao volume total do alimento em cada estômago, em percentagem (HYSLOP,
1980).
Os resultados individuais de ambos métodos foram combinados no Índice
Alimentar (IAi) (KAWAKAMI; VAZZOLER, 1980) e expressos em percentagem.Este
índice avalia o grau de importância que cada alimento possuiu na dieta dos peixes
(KAWAKAMI; VAZZOLER, 1980). Foi calculado o IAi(%) para cada item, adotando a
expressão:
Onde:
i = item alimentar;
F = frequência de ocorrência (%) do determinado item;
V= volume (%) de determinado item.
As guildas tróficas foram determinadas com base no IAi (%), considerando o
item alimentar predominante na dieta (IAi> 50%) (GASPAR-DA-LUZ et al., 2001).
Foram escolhidas para análise as espécies com número de exemplares acima de
seis. Posteriormente, foi determinada a estrutura trófica das assembleias de peixes.
A atividade alimentar foi avaliada através da identificação do Grau de
Repleção dos estômagos conforme a escala utilizada por Bennemann (1985) e
modificada por Yabe e Bennemann (1994) (Tabela 1).
Tabela 1. Descrição dos aspectos externos para estimativa do grau de repleção dos estômagos com os respectivos pontos atribuídos (BENNEMANN 1985; YABE; BENNEMANN 1994).
Grau de
repleção Características externas
0% - vazio Estômago sem alimento presente. Parede das porções cárdica e
pilórica muito espessa. Diâmetro da porção pilórica
correspondente à espessura da parede que ao ser palpada, sente-
se muito consistente.
25% - pouco cheio (1/4 cheio) Alimento na porção cárdica em maior quantidade que o grau

25
anterior.
50% - frequente (1/2 cheio) Alimento ocupando as duas porções, estando a maior Quantidade
na região cárdica, cuja parede torna-se mais delgada em metade
de sua extensão. O diâmetro da porção pilórica aumenta na região
anterior.
75% - cheio (3/4 cheio) Alimento ocupando toda a porção cárdica e metade da pilórica. A
parede da porção cárdica torna-se delgada em toda a sua
extensão. O diâmetro da porção pilórica aumenta muito na metade
anterior e em menos graus na restante.
100% - distendido (4/4 cheio) Alimento ocupando totalmente todo o estômago. A parede das
duas porções torna-se tão delgada, que ao ser manuseada
facilmente se rompe. O diâmetro da porção pilórica aumenta em
toda a extensão.
5.4 Análise estatística
Para verificar possível diferença estatística (p<0,05) entre os ambientes
amostrados, foi aplicada a Análise Multivariada de Variância Permutacional usando
Distância de Matrizes (perMANOVA) proposta por Anderson (2001; 2005) utilizando
como descritores as espécies coletadas e como objetos os ambientes amostrados e
calculando as matrizes de distâncias a partir do coeficiente de Bray-curtis. Essa
análise teve como objetivo testar possíveis diferenças entre as composições
ictiofaunísticas presentes nos ambientes amostrados. A posteriori foi utilizado o
Escalonamento Multi-dimensional não-paramétrico (nMDS), que utiliza a mesma
matriz de dados, para visualizar graficamente os resultados encontrados.
A similaridade entre as espécies e a composição alimentar das mesmas foi
verificada utilizando a análise de cluster tendo como medida de distância o
coeficiente de Bray-curtis (LEGENDRE; LEGENDRE,1983). Em seguida foi realizado
o cálculo da hierarquia utilizando o método da Distância Mínima ou Vizinho Mais
Próximo (Single Linkage) onde é calculada a matriz de distâncias entre os ‘n’
indivíduos da população e em seguida os indivíduos mais próximos são agrupados.
O coeficiente de correlação cofenética foi utilizado para se demonstrar a fidelidade
do dendrograma em relação a matriz inicial, sendo que o valor de 0,75 foi definido

26
como mínimo para que o dendrograma fosse considerado fiel à matriz de
similaridade (JONGMAN et al. 1995).
As análises foram realizadas utilizando-se o software estatístico R 3.3 (R
Development Core Team 2016).
6. RESULTADOS E DISCUSSÃO
6.1 Diversidade das assembleias
Foram coletados um total de 931 indivíduos, distribuídos em 4 ordens, 20
famílias, 48 gêneros e 65 espécies (Apêndice 1). Na floresta de igapó, foram
coletados 650 indivíduos e 62 espécies (Apêndice 1). Já na água aberta, foram
coletados 281 indivíduos e 37 espécies (Apêndice 1).
No igapó, a ordem predominante em relação à abundância numérica foi
Characiformes (63,33%), seguido por Siluriformes (28,35%), Perciformes (5,24%) e
Clupeiformes (3,08%) (Figura 2). Já na água aberta, a ordem predominante foi
Characiformes (57,04%), seguida de Siluriformes (34,75%), Clupeiformes (7,04%) e
Perciformes (1,06%) (Figura 2).
Figura 2. Porcentagem do número de indivíduos dentro de cada ordem para floresta de igapó e
água aberta dos lagos.
0 10 20 30 40 50 60 70
Clupeiformes
Perciformes
Siluriformes
Characiformes
Número de Indivíduos (%)
Igapó
Água Aberta

27
A floresta de igapó e o ambiente de água aberta apresentaram distribuição
semelhante em relação ao número de espécies dentro das ordens. A ordem
Characiformes foi a mais representativa (56,45% e 54,05%, respectivamente),
seguida de Siluriformes (25,81%e 35,14%, respectivamente), Perciformes (12,90% e
5,41%, respectivamente) e Clupeiformes (4,84% e 5,41%, respectivamente) (Figura
3). Perciformes e Clupeiformes foram as duas ordens com mesma
representatividade na água aberta. Diante disso, Characiformes apresentou o maior
número de espécies, gêneros e famílias nos dois ambientes.
Figura 3. Porcentagem do número de espécies dentro de cada ordem para floresta de igapó e
água aberta dos lagos.
Estudos realizados em ambientes alagados da Amazônia demonstram a
predominância das ordens Characiformes e Siluriformes (GOULDING, 1980; SAINT-
PAUL et al., 2000; CLARO-Jr., 2003; SIQUEIRA-SOUZA; FREITAS, 2004;
CORREDOR, 2004; SOARES; YAMAMOTO, 2005; NOVERAS et al., 2012,
LOEBENS et al., 2016).
As famílias mais representativas em número de indivíduos no igapó foram
Serrasalmidae (25,12%), Auchenipteridae (19,88%) e Hemiodontidae (17,87%)
(Figura 4). Já para água aberta foram Hemiodontidae (21,99%), Serrasalmidae
(19,15%) e Pimelodidae (18,79%) (Figura 4). A família Serrasalmidae também
apresentou a maior abundância em número de espécies (14,52%), seguida de
Cichlidae (11,29%) e Auchenipteridae (11,29%) no igapó (Figura 5). Já na água
aberta a família Auchenipteridae (18,92%) foi a mais representativa (Figura 5). A
família Pimelodidae (13,51%) foi a segunda e Hemiodontidae e Serrasalmidae,
0 10 20 30 40 50 60
Clupeiformes
Perciformes
Siluriformes
Characiformes
Número de espécies (%)
Igapó
Água Aberta

28
ambas com 4 espécies, se encontram em terceiro lugar como mais abundantes
(10,81%) (Figura 5).
As famílias Acestrorhynchidae, Doradidae e Characidae tiveram ocorrência
apenas na floresta de igapó. As famílias Engraulidae e Loricariidae apresentaram
um indivíduo e uma espécie cada para ambiente.
Figura 4. Porcentagem do número de indivíduos por famílias capturadas na floresta de igapó e
água aberta dos lagos estudados.
0 5 10 15 20 25 30
Engraulidae
Ctenoluciidae
Loricariidae
Doradidae
Prochilodontidae
Curimatidae
Bryconidae
Acestrorhynchidae
Characidae
Iguanodectidae
Cynodontidae
Sciaenidae
Pristigasteridae
Cichlidae
Anostomidae
Triportheidae
Pimelodidae
Hemiodontidae
Auchenipteridae
Serrasalmidae
Número de Indivíduos (%)
Igapó
Água Aberta

29
Figura 5. Porcentagem do número de espécies presentes em cada família para a floresta de
igapó e água aberta dos lagos estudados.
A presença dos Serrasalmidae foi constante em ambos os ambientes, sendo
os indivíduos do gênero Serrasalmus os mais abundantes nas coletas. O
gêneroapresenta ampla distribuição no ambiente aquático e habitam
preferencialmente ambientes onde a correnteza é menor (GOULDING, 1980), porém
não necessariamente ficam confinados em um único ambiente uma vez que durante
a formação de áreas alagadas ocorre a interconectividade dos lagos e a formação
dos mais variados habitats (LOWE-Mc-CONNELL, 1999).
As três espécies mais abundantes em número de indivíduos no igapó foram
Hemiodus immaculatus (16,33%), Auchenipterichthys longimanus (12,33%) e
Serrasalmus rhombeus (9,24%) (Figura 6) e as mais abundantes na água aberta
foram Hemiodus immaculatus (15,60%), Serrasalmus rhombeus (15,25%) e
Hypophthalmus edentatus (7,45%) (Figura 6).
O número de indivíduos por espécie demonstrou que poucas dominaram
numericamente. As espécies L. batesii, A. longiceps e A. vittatus apresentaram
apenas um único indivíduos em cada ambiente. H. immaculatus foi a espécie
dominante nos dois ambientes e também em três, dos quatro lagos amostrados.
0 5 10 15 20
Engraulidae
Acestrorhynchidae
Ctenoluciidae
Iguanodectidae
Curimatidae
Loricariidae
Sciaenidae
Pristigasteridae
Cynodontidae
Bryconidae
Prochilodontidae
Doradidae
Triportheidae
Characidae
Anostomidae
Hemiodontidae
Pimelodidae
Auchenipteridae
Cichlidae
Serrasalmidae
Número de espécies (%)
Igapó
Água Aberta

30
Das espécies amostradas 37 espécies (56,92%) foram comuns aos dois
ambientes, sendo 3 espécies (4,65%) coletadas apenas a água aberta e 25 espécies
(38,46%) encontradas apenas na floresta de igapó.
Figura 6. Abundância relativa do número de indivíduos das dez espécies mais abundantes na
floresta de igapó e água aberta dos lagos.
A curva de rarefação para a floresta de igapó apresentou um padrão de
inclinação menor do que para água aberta(Figura 7). Entretanto, existe ainda um
padrão crescente no número de espécies coletadas em função do número de
indivíduos para os dois ambientes. Esse resultado demonstra a possibilidade da
inclusão de novas espécies com um provável aumento do esforço de pesca.
Normalmente, estudos realizados na região amazônica não apresentam grande
esforço de pesca apesar de apresentar grandes listas de espécies (CLARO-Jr.,
2007). Segundo Goulding et al. (1988), o rio Negro necessita ainda de mais estudos
uma vez que, provavelmente, ainda possui um acréscimo de 30% do número de
espécies de peixes.
0 2 4 6 8 10 12 14 16 18 20
Bryconops alburnoides
Anodus elongatus
Agoniates halecinus
Hypophthalmus marginatus
Pellona flavipinnis
Metynnis hypsauchen
Ageneiosus polystictus
Tatia intermedia
Hypophthalmus edentatus
Pristobrycon sp.
Serrasalmus gouldingi
Triportheus elongatus
Serrasalmus rhombeus
Auchenipterichthys longimanus
Hemiodus immaculatus
Número de Indivíduos (%)
Igapó
Água Aberta

31
Figura 7. Curvas de rarefação e o seu respectivo desvio padrão para os ambientes de floresta
de igapó (A) e água aberta (B) no PARNA Anavilhanas.
O arquipélago de Anavilhanas possui inúmeros biótopos que vão desde
ambientes de terra firme a formação de florestas alagadas, aliando isso as
mudanças causadas anualmente pelo pulso de inundação e as poucas amostragens
realizadas no parque e em geral na floresta de igapó (SAINT-PAUL et al., 2000;
NOVERAS et al., 2012; LOEBENS et al., 2015) são grandes as probabilidades do
número de espécies presentes nesse habitat ser muito maior do que o encontrado
no presente trabalho.
A riqueza absoluta, a diversidade de Shannon-Wiener e a riqueza de Margalef
também foram maiores no ambiente de floresta alagada (Tabela 2). As amostras
analisadas os índices de Berger-Parker e Equitabilidade indicaram baixa dominância
e alta homogeneidade no número de indivíduos por espécie (Tabela 2), uma vez que
poucas espécies apresentaram abundância elevada no número de indivíduos nos
dois ambientes.
Os valores encontrados para os índices de diversidade corroboram com
resultados encontrados para florestas alagadas de águas pretas na Amazônia
Central (Tabela 2). A grande diferença entre o número de espécies encontrados por
Saint-Paul et al. (2000) deve-se ao fato do esforço de pesca ser muito maior do que
o do presente estudo. Segundo Goulding (1988), valores de índice de diversidade
superiores a 3,0 são indicativos de alta diversidade.

32
Tabela 2. Parâmetro de diversidade calculados para florestas de igapó do rio Negro. S =
riqueza de espécies; N = número de indivíduos; H’ = diversidade de Shannon-Wiener; J’ =
equitabilidade.
A perMANOVA, demonstrou efeito significativo do tipo de ambiente (p=0,001)
e da época de realização das coletas (p=0,027) na composição das assembleias de
peixes (Figura 8). O eixo 1 evidenciou as diferenças entre ambientes, agrupando do
lado direito as amostras realizadas na água aberta (AA), enquanto, no lado esquerdo
do gráfico, predominam as amostras efetuadas na floresta alagada (FA). O eixo 2,
por outro lado, agrupou as amostras da coleta 2 e as amostras da coleta 1 (Figura
1). Com exceção da amostra realizada na água aberta do lago Prato, as amostras
da coleta 3 estão situadas na porção central do gráfico.
Local de Coleta Parâmetros de diversidade
S N Biomassa (g) H' Margalef J' Berger-Parker Fonte
Igapó 62 650 66936,2 3,318 9,418 0,8038 0,1631 Presente estudo
Loebens et al., 2016 Água Aberta 37 281 25572,1 2,967 6,385 0,8216 0,1566
Igapó 150 5648 - 3,8 - - - Saint-Paul et al., 2000
Água Aberta 110
Igapó 41 337 - 2,912 - 0,784 - Novares et al., 2012
Água Aberta 30 877 - 2,435 - 0,717 -

33
Figura 8. Ordenação da composição taxonômica utilizando os dois primeiros eixos gerados
pela nMDS para os dados de abundância das amostras de peixes da floresta de igapó e água
aberta. Legenda: Floresta de Igapó (F.I.); Água Aberta (A.A.); coletas (1, 2, 3). FCG1=F. I., C.
Grande, col. 1; FCG2=F. I., C. Grande, col. 2; FCG3=F. I., C. Grande, col. 3; FCP1=F. I., C.
Pequeno, col. 1 FCP2=F.I. C. Pequeno, col. 2; FCP3=F.I. C. Pequeno, col. 3; FA1=F.I., Arraia,
col. 1; FA2=F.I., Arraia, col. 2; FA3=F.I., Arraia, col. 3; FP1=F.I., Prato, col. 1; FP2=F.I., Prato, col.
2; FP3=F. I., Prato, col. 3; ACG1=A.A.; C. Grande, col. 1; ACG2=A.A.; C. Grande, col. 2;
ACG3=A.A.; C. Grande, col. 3; ACP1=A.A.; C. Pequeno, col. 1; ACP2=A.A.; C. Pequeno, col. 12;
ACP3=A.A.; C. Pequeno, col. 3; AA1=A.A., Arraia, col.1; AA2 =A.A., Arraia, col.2; AA3=A.A,
Arraia, col.3; AP1=A.A., Prato, col. 1; AP2=A.A., Prato, col. 3; AP3=A.A., Arraia, col. 3.
Os resultados encontrados na análise evidenciam a importância da floresta
alagada na estruturação das assembleias de peixes. As diferenças encontradas
entre as coletas realizadas durante os períodos de enchente e vazante podem ser
explicadas pelo aumento espacial da área alagada e profundidade dos lagos. A
dinâmica da planície de inundação amazônica é controlada pela variação anual do
nível da água (JUNK et al.,1989). Com a ampliação das áreas marginais além do
aumento no número de hábitats disponíveis para os peixes, ocorre a dispersão e
redistribuição desses organismos nos lagos e no canal principal do rio
(RODRIGUEZ; LEWIS, 1994) sendo a conectividade em planícies de inundação
(LOWE-Mc-CONNELL, 1999) um processo de grande importância para a estrutura
das comunidades de peixes tropicais.

34
6.2 Estrutura trófica das assembleias de peixes da floresta de igapó
Foi analisado o conteúdo estomacal de 533 exemplares pertencentes a 24
espécies do igapó, sendo que desse total, 110 foram classificados como estômagos
vazios (20,64%), 162 como pouco cheios (30,39%), 52 como frequente (9,76%), 70
como cheios (13,13%) e 139 como distendidos (26,08%). Ressalta-se a presença de
material digerido que apesar de não ser considerado como item para o presente
estudo participa como porção alimentar encontrada nos estômagos durante as
análise do grau de repleção.
Os resultados das análises dos conteúdos estomacais revelaram uma dieta
diversificada onde os peixes consumiram como itens alimentares principais: insetos
(IN), representados pelos insetos imaturos, larvas, ninfas, e insetos adultos,
fragmentos de insetos não identificados; crustáceos (CR), constituídos por camarões
e caranguejos; microcrustáceos (ZOO), constituídos por cladóceras e copépodos;
peixes (P), constituídos por restos de peixes e escamas; frutos e sementes (FS),
constidos por frutos e sementes, inteiros ou não; e material vegetal (MV),
constituídos por pedaços de folhas, flores e raízes (Apêndice 2).
No estudo, é perceptível a grande quantidade de espécies consumidoras de
frutos e sementes pertencentes às famílias Arecaceae, Myrtaceae, Poaceae,
Burseraceae e Lecythidaceae e também, por insetos pertencentes às ordens
Hymenoptera, Coleoptera, Hemiptera, Diptera, Lepidoptera. Isso se deve ao fato da
frutificação das árvores ocorrer durante o período das cheias (GOULDING, 1980) e
da floresta alagada apresentar grande oferta de invertebrados, que são fonte rica em
proteínas (ADIS, 1997; JUNK et al., 1997; GOULDING, 1980; ZAVALA-CAMIN,
1996), sendo a presença de arbustos e árvores o substrato para esses organismos
(ADIS, 1997).
As florestas alagadas de água branca e preta se apresentam visualmente
parecidas (PRANCE, 1980), entretanto as várzeas são abundantes em frutos
carnosos e em quantidade produzida (SAINT-PAUL et al., 2000) e os igapós muito
mais ricos na produção de invertebrados. Isso pode ser observado no presente
estudo, uma vez que a grande maioria das espécies analisadas apresentaram o item
insetos em sua alimentação (Apêndice 2).
Ao contrário do período de águas baixas, onde o alimento se torna mais
escasso e com isso os organismos tendem a se alimentar de itens não comumente

35
encontrados em sua dieta durante o período de águas altas e formação de florestas
alagadas apresentam uma maior disponibilidade de itens no meio, demonstrando
suas preferências alimentares (GOULDING, 1980). Essa condição, aliada a
otimização dos ganhos energéticos, demonstra a grande interação dos organismos
com o meio.
Apesar de consumirem seus itens preferenciais, a diversidade de itens
encontrados na alimentação dos peixes provavelmente deve-se ao fato de sua
grande adaptação trófica (LOWE-McCONNELL., 1999). Isso demonstra um alto grau
de generalismo, de versatilidade alimentar e elevada plasticidade trófica nas
espécies tropicais (ABELHA et al., 2001; DEUS; PETRERE-JUNIOR, 2003),
conforme evidenciado em estudos realizados em ambientes tropicais com a
presença de rios sazonais (GOULDING, 1980; LOWE-McCONNELL, 1999;
SANTOS; FERREIRA, 1999; CLARO-Jr., 2003; ANJOS, 2007; PEREIRA, 2010).
A partir do cálculo do Índice de Importância Alimentar (%) (IAi), as 24
espécies foram agrupadas em categorias, segundo Zavala-Camin (1996), de acordo
com o regime alimentar sendo definidas em carnívoras(ingerem principalmente itens
de origem animal), piscívoras (ingerem principalmente peixes), insetívoras (ingerem
principalmente insetos), frugívoras (ingerem principalmente frutos e sementes),
onívoras (ingerem alimentos de origem animal e vegetal), herbívoras (ingerem
alimentos de origem vegetal) e zooplanctívoras (ingerem microrganismos de origem
animal ou zooplâncton) (Apêndice 2).
A categoria dos piscívoros apresentou o maior número de espécies (29,17%),
seguidas dos insetívoros (20,00%) e herbívoros (16,67%) (Figura 9). Os insetívoros
apresentaram a maior abundância (28,89%), seguida dos piscívoros (26,49%) e dos
onívoros (22,33%) (Figura 9). Apesar disso, a dieta de 38 espécies com 191
indivíduos não pode ser determinada, uma vez que apresentaram poucos indivíduos
coletados e estômagos analisados, inviabilizando assim uma conclusão confiável.

36
Figura 9. Frequência relativa (%) das categorias tróficas encontradas para as espécies
analisadas pertencentes a floresta de igapó do PARNA Anavilhanas.
A variação de espécies presentes nas guildas tróficas pode ser explicada pela
alta disponibilidade de determinados recursos no ambiente e pelo fato de alguns
recursos serem mais explorados pelos organismos ali presentes (ABELHA et al.,
2001).
A espécie mais abundante representante da guilda dos carnívoros foi o apapá
P. flavipinnis, alimentando-se de insetos, camarão e peixes e apresentando uma
forte tendência a insetivoria (69,52%) (Figura 10). Moreira-Hara et al. (2009) em
estudo sobre a alimentação da espécie em áreas alagadas da Amazônia Central,
demonstrou o elevado consumo de peixes e insetos e atividade alimentar maior
durante o período da cheia.
0,00
5,00
10,00
15,00
20,00
25,00
30,00
35,00 F
req
uên
cia
Rela
tiv
a(%
)
Abundância
Riqueza

37
Figura 10. Índice de importância alimentar (IAi%) para a espécie carnívora mais abundante P.
flavipinnis.
O cangati T. intermedia apresentou o IAi de 98,37% para insetos das ordens
Hymenoptera e Coleoptera bem como restos de insetos e ovos não identificados, o
também cangati A. longimanus com de 94,53% para insetos das ordens
Hymenoptera, Hemiptera e Lepidoptera, e a sardinha-comprida T. elongatus
apresentando insetos das ordens Lepidoptera e Hymenoptera, bem como restos de
insetos não identificados (71,92%) caracterizam-se como as espécies mais
abundantes pertencentes a guilda dos insetívoros, respectivamente. (Figura 11).
Figura 11. Índice de importância alimentar (IAi%) para as espécies insetívoras mais
abundantes T. intermedia, T. elongatuse A. longimanus.
As sardinhas geralmente são espécies onívoras alimentando-se de itens
abundantes disponíveis, como já foi observado para mesma espécie por Almeida
0,00
10,00
20,00
30,00
40,00
50,00
60,00
70,00
80,00
Insetos Camarão Peixe Material vegetal
0,00
10,00
20,00
30,00
40,00
50,00
60,00
70,00
80,00
90,00
100,00
Insetos Camarão Peixe Material vegetal
Frutos/ sementes
T. intermedia
T. elongatus
A. longimanus

38
(1984) e para T. albus por Ferreira et al. (1998). As diferenças encontradas para a
mesma espécie ocorrem pela grande plasticidade tróficas dos peixes tropicais
(ABELHA et al., 2001)
A principal representante da categoria trófica de onívoros foi o cubiu H.
immaculatus onde constatou-se a presença de vários itens tanto de origem vegetal
como animal (Figura 12). Dentre o item insetos, sendo o mais representativo com
50,46%, destacam-se os insetos das ordens Hymenoptera, Coleoptera, Hemiptera e
Diptera. Isso demonstra uma forte tendência à insetivoria, entretanto a espécie não
foi caracterizada como tal uma vez que a presença de frutos e sementes e material
de origem vegetal foi evidenciada nas análises. Freitas (2008), em estudo sobre a
composição da dieta de peixes do lago Prato no arquipélago de Anavilhanas durante
a seca, classificou a espécie como insetívora. A presença de mais itens na dieta da
espécie pode ser determinada pelo aumento na quantidade de recursos alimentares
no meio.
Figura 12. Índice de importância alimentar (IAi%) para a espécie onívora mais abundante H.
immaculatus.
A espécie mais representativa na guilda trófica dos piscívoros foi a piranha
preta S. rhombeus que apresentou valores elevados para o item alimentar peixe
(76,77%) com a presença de escamas ctenóides e ciclóides e restos de peixe,
alguns identificados como pertencentes à ordem Siluriformes (Figura 13). A
presença do ofídio Hrydops cf.martii (Colubridae) provavelmente é ocasional uma
vez que cobras não são itens normalmente encontrados na alimentação de peixes.
As espécies piranha-branca S. gouldingi e mandubé A. polystictus foram a segunda
0,00
10,00
20,00
30,00
40,00
50,00
60,00
Insetos Crustáceos Material vegetal Frutos/ sementes

39
e terceira apresentando 73,34% e 81,82% pra o item peixes, respectivamente
(Figura 14).
Figura 13. Índice de importância alimentar (IAi%) para as espécies piscívoras mais abundantes
A. polystictus, S. gouldingi e S. rhombeus.
A espécie Pristobrycon sp., apresentou uma grande quantidade de frutos e
sementes com o IAi (%) de 94,07%. Foram encontrados em menor quantidade os
itens peixe, material digerido, material vegetal, insetos e camarão. Com isso, ela foi
caracterizada como a única espécie representante pertencente à guilda dos
frugívoros.
Entre os serrasalmídeos ocorre a preferência pelas nadadeiras das presas
(GOULDING, 1980), entretanto, existe também consumo de outros itens como
peixes inteiros, frutos, vegetais e invertebrados (GOULDING, 1980; LEÃO et al.,
1991).
A espécie mapará H. edentatus foi a principal representante da guilda dos
zooplanctívoros. Em sua alimentação foram encontrados cladóceros das famílias
Bosminidae e Daphiniidae, que constituem o zooplânctone em pequenas
quantidades, material vegetal, frutos e sementes e insetos (Figura 14). Previattelli e
Santos-Silva (2011), em estudo realizado no lago Tupé, apontam a grande presença
de cladóceros na alimentação desses organismos. A presença de itens alimentares
como insetos e material de origem vegetal também foi evidenciado por Carvalho
(1980), em estudo realizado no lago do Castanho, sendo associado a predação dos
insetos nas raízes das macrófitas. Uma vez que a espécie apresenta rastros
0,00
10,00
20,00
30,00
40,00
50,00
60,00
70,00
80,00
90,00
A. polystictus
S. gouldingi
S. rhombeus
40
branquiais longos e numerosos e ausência de dentes é classificada como uma
espécie planctófaga segundo Nikolsky (1963) e Carvalho (1980).
Figura 14, Índice de importância alimentar (IAi%), para a espécie zooplanctívora mais
abundante H. edentatus.
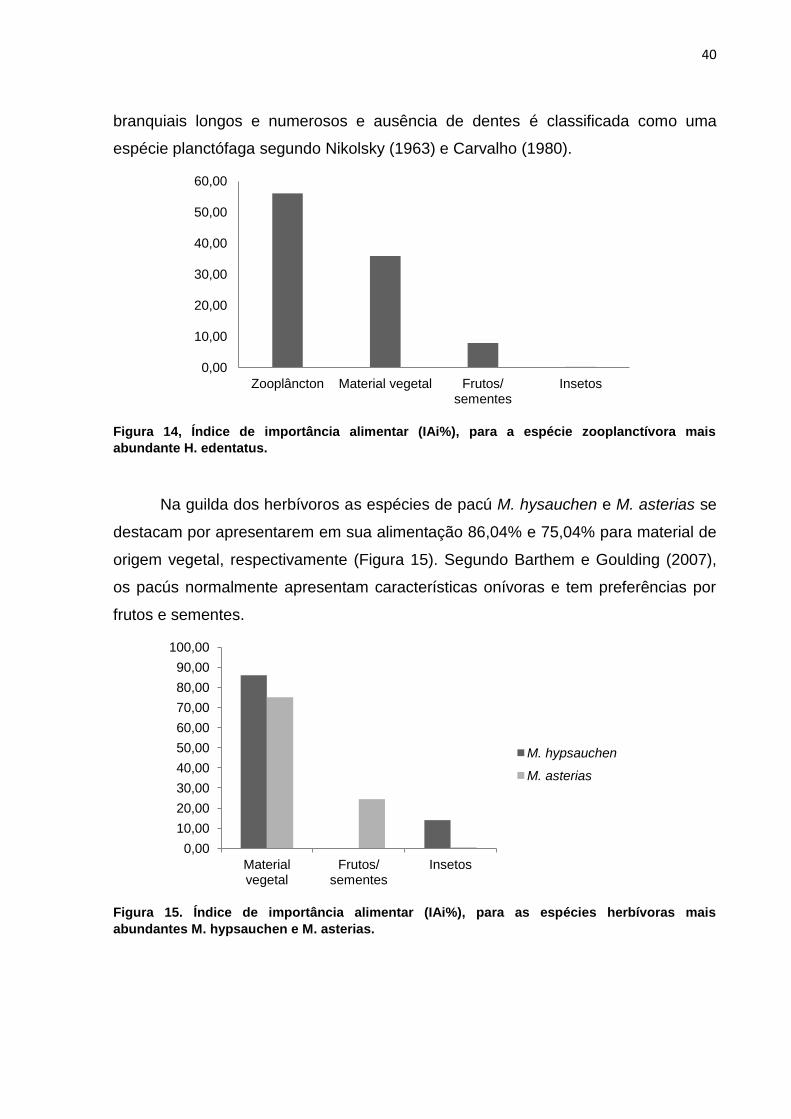
Na guilda dos herbívoros as espécies de pacú M. hysauchen e M. asterias se
destacam por apresentarem em sua alimentação 86,04% e 75,04% para material de
origem vegetal, respectivamente (Figura 15). Segundo Barthem e Goulding (2007),
os pacús normalmente apresentam características onívoras e tem preferências por
frutos e sementes.
Figura 15. Índice de importância alimentar (IAi%), para as espécies herbívoras mais
abundantes M. hypsauchen e M. asterias.
0,00
10,00
20,00
30,00
40,00
50,00
60,00
Zooplâncton Material vegetal Frutos/ sementes
Insetos
0,00
10,00
20,00
30,00
40,00
50,00
60,00
70,00
80,00
90,00
100,00
Material vegetal
Frutos/ sementes
Insetos
M. hypsauchen
M. asterias

41
Através da analise de agrupamento, constatamos que a composição da dieta
dividiu as espécies em seis guildas tróficas. O primeiro grupo formado por
Pristobryconsp., o segundo formado por H. edentatuse H. fimbriatus, o terceiro
formado por L. fasciatus, H. immaculatus, H. marginatus, P. flavipinnis, T. elongatus,
A. longimanus, T. angulatus, B. alburmoidese T. intermédia, o quarto formado por L.
taeniata, M. hypsauchen, M. asterias, P. trimaculatus, o quinto formado por S.
eigenmannie o sexto formado por P. squamosissimus, R. vulpinus, A. microlepis, A.
ucayalensis, S. gouldingi, A. polytictus e S. rhombeus (Figura 16).
Figura 16. Dendrograma da similaridade da dieta dos peixes analisados com valores de IAi (%)
presentes na floresta de igapó do PARNA Anavilhanas.

42
As espécies evidenciadas no terceiro grupo provavelmente foram agrupadas
de tal forma na análise, pela proximidade na alimentação uma vez que fazem parte
da grande categoria dos carnívoros. Entretanto, durante a categorização foram
agrupadas em carnívoros, insetívoros e onívoros pela distribuição dos valores
encontrados de IAi (%).
A espécie S. eigenmanni foi classificada como onívora pelo presente estudo,
porém diferenciando-se das outras alocadas na mesma categoria (onívora) por sua
tendência a piscivoria sendo o provável motivo de ter sido alocada individualmente
na análise, porém próxima às espécies consideradas plenamente piscívoras.
O mapará H. marginatus, foi classificado como insetívoro, apesar de sua
semelhança com as outras espécies do mesmo gênero e que foram classificadas em
zooplanctívoras, pela presença elevada do item insetos em sua alimentação.
Ressalta-se que os estômagos analisados apresentavam grande quantidade de
material digerido e que sendo retirado da análise podem ter influenciado no
resultado encontrado.
A oferta de alimentos nas regiões tropicais é regulada pelo pulso de
inundação (ZAVALA-CAMIN, 1996). As espécies onívoras aproveitam grande
variedade de alimentos disponíveis em diversos locais e com isso são favorecidas
em épocas de cheia. A presença de insetívoros, carnívoros e onívoros com
tendências a piscivoria e insetivoria deve-se ao fato da utilização dos ambientes
alagados por predadores, provavelmente para acompanharem a migração lateral
das presas. Outro fato que pode ser levado em consideração é que as malhadeiras
são altamente seletivas a organismos que se movimentam com frequência em busca
das presas. Espécies predadoras controlam a abundância de muitas espécies de
pequeno e médio porte, mantendo-as em níveis que não possam causar um colapso
no ambiente (JANZEN, 1970; PAYNE, 1966) e estão presentes em grandes
quantidades em estudos de estrutura de comunidades (SAINT-PAUL et al., 2000;
CLARO-Jr., 2007; FERREIRA, 1998). Com isso, demonstra-se a importância da
piscivoria em sistemas tropicais (RODRIGUEZ; LEWIS 1997; SÚAREZ et al. 2001).
Segundo proposto por Welcomme (1985), a coexistência de espécies,
pertencentes a diferentes guildas tróficas, demonstra adaptações das mesmas para
utilização do mesmo ambiente. Além disso, como encontrado por Claro-jr et al.
(2004), em estudo sobre o efeito da floresta alagada em espécies onívoras no

43
sistema Solimões/Amazonas, a determinação da estrutura trófica pode fornecer
informações sobre a ecologia das mesmas.
7. CONCLUSÕES
A estrutura das assembleias de peixes encontradas no ambiente de floresta
de igapó do PARNA Anavilhanas é formada por ordens de peixes esperadas para a
bacia Amazônica e, ambientes de água preta, com dominância de Characiformes,
seguida de Siluriformes.
A elevada presença de espécies, apesar do baixo número de indivíduos
distribuídos, demonstra a grande riqueza presente no igapó, sendo necessários mais
estudos voltados para esse ambiente.
A riqueza de espécies provavelmente é maior do que o encontrado no
presente trabalho principalmente se um esforço de pesca maior for empregado.
Os resultados encontrados demonstram que na zona de transição aquático-
terrestre (ATTZ) existe uma maior diversidade de organismos que provavelmente
estão explorando este ambiente para abrigo e alimentação.
A composição alimentar das assembleias de peixes presente na floresta de
igapó é diversificada e reflete a importância desse habitat para a manutenção da
biota aquática.
As principais guildas encontradas foram insetívoras, carnívoras e onívoras.
Isso demonstra a relação dos organismos com a floresta uma vez que esta possui
grande quantidade de itens disponíveis principalmente de insetos e frutos, além da
elevada presença de presas que adentram o igapó em busca de alimento.

44
8. REFERÊNCIAS BIBLIOGRÁFICAS
ABELHA, M. C. F.; AGOSTINHO, A. A.; GOULART, E. Plasticidade trófica em peixes
de água doce. Acta Scientiarum, Maringá, v. 23, n. 2, p. 425-434, 2001.
ADIS, J. Terrestrial invertebrates: survival strategies, group spectrum, dominance
and activity patterns, vol. 126, p. 318-330. In: JUNK, W. J. (Ed.) The central
Amazon floodplain. Berlin: Ecological Studies, 1997. p. 525.
AGOSTINHO, A. A.; THOMAZ, S. M.; GOMES, L. C. 2005. Conservação da
biodiversidade em águas continentais do Brasil. Megadiversidade,v. 1, n. 1, p.70-
78.
ALMEIDA, R. G. Biologia alimentar de três espécies de Triportheus (Pisces:
Characoidei: Characidae) do lago Castanho, Amazonas. Acta Amazônica,
Amazonas, v. 14, p. 48-76. 1984.
ANDERSON, M. J. A new method for non-parametric multivariate analysis of
variance. Aust. Ecol, n. 26, p. 32-46, 2001.
ANDERSON, M. J. Permanova: a FORTRAN computer program for permutational
multivariate analysis of variance. Auckland: Department of Statistics, University
of Auckland, New Zealand, 24 p, 2005.
ANGERMEIER, P. L.; KARR, J. R. Fish communities along environmental gradients
in a system of tropical streams. In: ZARET, T. M. Evolutionary ecology of
neotropical freshwater fishes. Holanda: The Hague, 1984. p.39-57.
ARAÚJO, F. G. Adaptação do índice de integridade biótica usando a comunidade de
peixes para o Rio Paraíba do Sul. Revista Brasileira de Biologia, São Carlos, v.
58, n. 4, p. 547-558, 1998.
ARAUJO-LIMA, C. A.; PORTUGAL, L. P. S.; FERREIRA, E. G. 1986.
Fishmacrophytesrelationship in the Anavilhanas Archipelago, a blackwater system in
the central Amazon. Journal of Fish Biology, 29:1-11.

45
ARAÚJO-LIMA, C.; FORSBERG, B. R.; VICTORIA, R.; MARTINELLI, L. Energy
sources for detritivores fishes in the Amazon. Science, EUA, v. 234, p.1256-1258,
1986.
ARRINGTON, D. A., WINEMILLER, K. O.; LAYMAN, C. A. Community assembly at
the patch scale in a species-rich tropical river. Oecologia, v. 144, p. 157–167, 2005.
ARRINGTON, D. A.; WINEMILLER, K. O. Cyclical flood pulses, littoral habitats and
species associations in a Neotropical floodplain river. Journal of the North
American Benthological Society, EUA, v. 25, n. 1, p. 126-141, 2006.
BARRETO FILHO, H. T. Da Nação ao Planeta através da Natureza: uma
abordagem antropológica das unidades de conservação de proteção integral
na Amazônia brasileira. São Paulo: Tese de Doutorado,Universidade de São
Paulo. 2001.
BARTHEM, R.B.; GOULDING, M. Um ecossistema inesperado: a Amazônia
revelada pela pesca.Amazon Conservation Association (ACA), Sociedade Civil
Mamirauá, Belém, 2007, 241 pp.
BENNEMANN, S. T. Aspectos da sistemática, alimentação e reprodução de
SchizodonIlasutus e Schizodonplatae do Rio Ibicuí-Mirim, RS
(PiscesAnostomidae). Santa Maria, Dissertação de Mestrado, não publicada,
Universidade Federal de Santa Maria, 99p., 1985.
BERGER, W. H.; PARKER, F. L. Diversity of Planktonic Foraminifera in Deep-Sea
sediments. Science, EUA, v. 168, p. 1345-1347, 1970.
CALLISTO, M.; MORENO, P.; BARBOSA, F. A. R. Habitat diversity and benthic
functional trophic groups at Serra do Cipó, Southeast Brazil. Revista Brasileira de
Biologia, São Paulo, v. 61, n.2, p. 259-266, 2001.
CARVALHO, L. N., ZUANON, J.; SAZIMA, I. Natural history of Amazon fishes. In: K.
Del Claro, P. S. Oliveira, V. Rico-Gray, A. Ramirez, A. A. A. Barbosa, A. Bonet, F. R.
Scarano, F. L. Consoli, F. J. M. Garzon, J. N. Nakajima, J. A. Costello, and M.

46
Vinicius. (ed.). Encyclopedia of Life Support System. Oxford: Eolss Publishers,
UNESCO, 2007, p. 1-24.
CLARO Jr., L.; FERREIRA, E.; ZUANON, J. A.; ARAUJO-LIMA, C. O efeito da
floresta alagada na alimentação de três espécies de peixes onívoros em lagos de
várzea da Amazônia central, Brasil. Acta Amazônica, Amazonas, v. 34, n. 1, p. 133-
137, 2004.
CORREA, S. B., CRAMPTON W. G. R., CHAPMAN L. J.; ALBERT J. S. A
comparison of flooded forest and floating meadow fish assemblages in an upper
Amazon floodplain. Journal of Fish Biology, EUA, v. 72, p. 629–644, 2008.
CROPP, R.; GABRIC, A. Ecosystem adaptation: Do ecosystems maximize
resilience? Ecology, EUA, v. 83, n. 7, p. 2019-2026, 2002.
DEUS, C. P.; M. PETRERE-JUNIOR. Seasonal diet shifts on seven fish species in an
Atlantic Rainforest stream in southeastern Brazil. Brazilian Journal of Biology, São
Carlos, v. 63, n.4, p. 579-588, 2003.
FERNANDES, C. C.; NOGUEIRA. A.; WILLISTON, A.; ALVES-GOMES, J. A. A new
species of electric knifefish from the rio Negro, Amazon basin
(Gymnotiformes: Hypopomidae, Microsternarchini). Proceedings of the Academy
of Natural Sciences of Philadelphia, EUA, v.164, p. 213-227, 2015.
FERREIRA, E. J. G. Alimentação dos adultos de doze espécies de ciclídeos
(Perciformes, Cichlidae) rio Negro, Brasil. Manaus, Amazonas: Dissertação de
Mestrado, INPA/FUA, 254 p., 1981.
FERREIRA, E. J. G.; ZUANON, J. A. S.; SANTOS, G. M. Peixes comerciais do
médio Amazonas: região de Santarém, Pará. Brasília: Ibama, 1998. 211pp.
FITTKAU, E. J. Remarks on limnology of central-Amazon rainforest streams. Verh.
Internat. Verein. Limnol, v. 15, p. 1092-1096, 1964
GARCIA, M. Aspectos ecológicos dos peixes das águas abertas de um lago no
arquipélago de Anavilhanas, rio Negro, AM. Manaus, Amazonas: Dissertação de
Mestrado, INPA/UFAM, pp. 95, 1995.

47
GASPAR DA LUZ, K. D.; ABUJANRA, F.; AGOSTINHO, A. A.; GOMES, L. C.
Caracterização trófica da ictiofauna de três lagoas da planície aluvial do alto rio
Paraná, Brasil. ActaScientiarum, Biological Sciences, v. 23, n. 2, p.401-407, 2001.
GÉRY, J. Characoids of the world. Neptune City, T.F.H. Publications, 1977, 672 p.
GOLDSTEIN, R. M.; MEADOR, M. R. Comparisons of fish species traits from small
streams to large rivers. Transactions of the American Fisheries Society, USA, v.
133, p. 971–983, 2004.
GORMAM, O. T.; KARR, J. R. Habitat structure and stream fishes communities.
Ecology, USA, v. 59, p. 507-515, 1978.
GOULDING, M.; CARVALHO, M. L. Life history and management of tambaqui
(Colossomamacropomum, Characidae), an important Amazonian food fish.
RevistaBrasileira de Zoologia, v. 1, p. 107-133, 1982.
GOULDING, M.; CARVALHO, M. L.; FERREIRA, E. G. Rio Negro Rich life in poor
water: Amazonian diversity and food chain ecology as seem through fish
communities. The Hague: SPB AcademicPublishing, 1988. 200p.
HENRY, R. Ecótonos nas interfaces dos ecossistemas aquáticos. RIMA,
SãoCarlos: Rima Editora, 2003. p. 1-28.
HYNES, H. B. N. The food of freshwater sticklebacks (Gasterosteusaculeatus and
Pygosteuspungitius) with a review of methods used in the studies of the food of
fishes. Journ. Anim. Ecol., v. 19, p. 36-58, 1950.
HYSLOP, E. J. Stomach content analysis: a review of methods and their
applications. J. Fish Biol., Southampton, v. 17, n.4, p.411-429, 1980.
JACKSON R. B.; CARPENTER S. R.; DAHM C. N. Water in a changing world.
EcolAppl, v. 11, p. 1027–45, 2001.
JONGMAN, R. H. G.; TER BRAAK, C. J. F.; VAN TONGEREN, O. F. R. Data
analysis in communityandlandscapeecology. Cambridge Univ. Press, Cambridge.
1995.

48
JUNK, W. J. Ecology, fisheries and fish culture in Amazônia. In: SIOLI, H. (Ed.). The
Amazon. Liminology and landscape ecology of mighty tropical river and its
basin. Dordrecht: Dr. W.Junk Publishers, 1984. p. 443-476.
JUNK, W. J. General aspects of floodplain ecology with special reference to
Amazonian floodplains, pp. 3-20. In: JUNK, W. J. (Ed.). The Central Amazon
Floodplain. Berlin Heidelberg: Springer-Verlag, 1997.
JUNK, W. J.; BAYLEY, P. B.; SPARKS, R. E. The flood pulse concept in river-
floodplains systems. In: DODGE, D. P. (Ed.). Proceedings of the International
Large River Symposium. Canadian SpecialPublication of Fisheries and Aquatic
Science, 1989. v. 106, p. 110-127.
KARR, J. R. Assessment of Biotic Integrity using fish communities. Fisheries,
Maryland, v. 6 n. 6, p. 21-27, 1981.
KAWAKAMI, E.; VAZZOLER, G. Método gráfico e estimativa de índice alimentar
aplicado no estudo da alimentação de peixes. Bol. Inst. Oceanogr., v. 29, p. 205-
207, 1980.
KENSLEY, B.; WALKER, I. Paleomonidae shrimps from the Amazon basin, Brazil,
(Crustacea: Decapoda: Natantia). Smithsonian Contributions to Zoology, p. 362,
1982.
KRAMER, D. L., LINDSEY, C. C., MOODIE, G. E. E.; STEVENS, E. D. The fishes
and the aquatic environment of the central Amazon basin, with particular reference to
respiratory patterns. Can. J. Zool., v. 56, n. 4, p. 717-729, 1978.
KREBS, C. J. Ecological Methodology . New York: Harper-Collins Publ., 1989,
370p.
LEENHEER, J. A.; SANTOS, V. Consideração sobre os processos de sedimentação
na água preta ácida do rio Negro (Amazônia Central). Acta Amazônica, Amazonas,
v. 10, n. 2, p. 343-355, 1980.
LEGENDRE, L.; LEGENDRE, P. Numericalecology. Developments in
environmentalmodeling, 3. ElsevierScientific, Amsterdan, The Netherlands. 1983.

49
LÉVÊQUE, C., OBERDORFF, T., PAUGY, D., STIASSNY, M. L. J. & TEDESCO, P.
A. Global diversity of fish (Pisces) in freshwater. Hydrobiologia, n. 595, p. 545-567,
2008.
LOEBENS, S. C.; FARIAS, E. U.; YAMAMOTO, K. C.; FREITAS, C. E. C.
Diversidade de assembleias de peixes em floresta alagada de águas pretas da
Amazônia Central. Scientia Amazonia, Amazonas, v. 5, n.1, 37-44, 2016.
LOWE-McCONNELL, R. H. Estudos Ecológicos de Comunidades de Peixes
Tropicais. São Paulo: Edusp, 1999. 366 p.
LUDWIG, J. A.; REYNOLDS, J. F. Statistical ecology. Nova York: J. Wiley, 1988.
MAGURRAN, A. E. Ecological diversity and its measurement. Universidade de
Princeton, Princeton, Nova Jersey, EUA, 1988.
MAIA, C. R.; ALVES-GOMES, J. A. Utilização do código de barras de DNA na
estimativa da diversidade de peixes elétricos do gênero Microsternarchus
(Ostariophysi: Gymnotiformes) na bacia do Rio Negro, Amazonas. In: Eloy Guillermo
Castellon; Luiz Augusto Gomes de Souza. (Org.). Desvendando as Fronteiras do
conhecimento na região amazônica do Alto Rio Negro. Manaus: INPA,2012.v.
01, p. 185-202.
MARTINS, F. R.; SANTOS, F. A. M. Técnicas usuais de estimativa da
biodiversidade. HolosEnvironment, Rio Claro, v. 1, n. 1, p. 236-267, 1999.
MAY, R. M. Patterns of species abundance and diversity. In: CODY, M. L.;
DIAMOND, J. M. (Ed.) Ecology and evolution of communities. Cambridge:
Belknap Press of the Havard University Press, 1975. p. 81-120.
MAYLY, B. F. J. Multivariate Statistical Methods: A primer. 3rdEditon. Chapman &
Hall/CRC, London, 2005. 214pp.
MOREIRA-HARA, S. S.; ZUANON, J. A. S.; AMADIO, S. A. Feeding of
Pellonaflavipinnis (Clupeiformes, Pristegasteridae) in a Central Amazonia floodplain.
Iheringia Série Zoologia, v. 99, n. 2, p. 153-157, 2009.

50
NOVERAS, J.; YAMAMOTO, K. C.; FREITAS, C. E. C. Uso do igapó por
assembléias de peixes nos lagos no Parque Nacional das Anavilhanas (Amazonas,
Brasil). ActaAmazonica, Amazonas, v. 42, n. 4, p. 567 – 572, 2012.
OLIVEIRA, A. K.; CARAVELLO, J. C. Fish assemblage composition in a tributary of
the Mogi Guaçu river basin, south-eastern Brazil. Iheringia, Série Zoológica, v. 93, n.
2, p. 369-376, 2003.
PAZIN, V. F. V. Assembleias de peixes em poças temporárias marginais a
riachos de terra Firme Amazônia Central. Manaus, Amazonas: Dissertação de
mestrado, INPA/UFAM, 40 p., 2004.
PIEDADE, M. T. F.; SCHONGART, J.; WITTMANN, F.; PAROLIN, P.; BLEICH, M.
E.; LOPES, A. Iniciando a viagem pelas áreas úmidas amazônicas. In: LOPES, A.;
PIEDADE, M. T. F. Conhecendo as áreas úmidas amazônicas: uma viagem
pelas várzeas e igapós.Manaus: INPA, 2015. p. 16-22.
PRADO, K. L. L.; FREITAS, C. E. C.; SOARES, M. G. M. Assembleias de peixes
associadas às macrófitas aquáticas em lagos de várzea do baixo rio Solimões.
Revista Biotemas, Santa Catarina, v. 23, p. 131-142, 2010.
PRANCE, G. T. A terminologia dos tipos de florestas amazônicas sujeitas à
inundação. Acta Amazonica, Amazonas, v. 10, n. 3, p. 495-504, 1980.
PREVIATELLI, D.; SANTOS-SILVA, E. N. Predação de zooplâncton por peixes no
lago Tupé, Manaus-AM. In: SANTOS-SILVA, E. N; SCUDELLER, V. V.;
CAVALCANTI, M. J. (Eds.). BioTupé: Meio Físico, Diversidade Biológica e
Sociocultural do Baixo Rio Negro, Amazônia Central. Manaus, Rizoma Editorial,
2011, p.242-252.
R Development Core Team. R: A language and environment for statistical
computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-
900051-07-0, 2016. URL: http://www.R-project.org.
RAHEL, F. J. Factors structuring fish assemblages along a bog lake successional
gradient. Ecology, USA, v. 65, p. 1276-1289, 1984.

51
RIBEIRO, M. C. L. B. As migrações dos jaraquis (Pisces, Prochilodontidae) no
Rio Negro, Amazonas, Brasil. Manaus, Amazonas: Dissertação de Mestrado,
Instituto Nacional de Pesquisas da Amazônia, 192pp., 1983.
RIBEIRO, M. C. L. B.; PETRERE JR., M. Fisheries ecology and management of the
jaraqui (Semaprochilodus taeniurus, S. insignis) in the central Amazonia. Regulated
river. Res. &Manag. v. 5, p. 195-215, 1990.
RICKLEFS, R. E. Ecology. 3 ed. New York: W. H. Freeman and Company. 1990.
RODRÍGUES, M. A.; LEWIS-JR. Structure of fish assemblages along environmental
gradients in floodplain lakes of the Orinoco River. Ecological Monographs, v. 67, n.
1, p. 109-128, 1997.
SAINT-PAUL, U.; ZUANON, J.; VILLACORTA CORREA, M. A.; GARCÍA, M.;
FABRÉ, N. N.; BERGER, U.; JUNK, W. J. Fish communities in central Amazonian
white-and blackwater floodplains. Environmental BiologyofFishes, v. 57, p. 235-
250, 2000.
SANTOS, G. M. Estudos de alimentação e hábitos alimentares de Schizodon
fasciatus Agassiz, 1829, Rhytiodus microlepis Kner, 1859 e Rhytiodus
argenteofuscus Kner 1859, do lago Janauacá – AM (Osteichthyes, Characoidei,
Anostomidae). Acta Amazonica, Amazonas, v. 11, n. 2, p. 267-283, 1981.
SANTOS, G. M.; FERREIRA, E. J. G. Peixes da bacia Amazônica. In: LOWE-
McCONNELL, R. H. Estudos ecológicos de comunidades de peixes tropicais.
Tradução de Anna Emília A. M. Vazzoler; Angelo Antonio Agostinho; Patrícia T. M.
Cunningham. São Paulo: EDUSP, 1999. p. 345-354.
SANTOS, G. M.; FERREIRA, E. J. G.; ZUANON, J. A. S. Peixes Comerciais de
Manaus. Manaus: Ibama/AM, ProVárzea, 2006. 144pp.
SANTOS, G. M.; JURAS, A. A.; MÉRONA, B. JÉGU, M. Peixes do baixo rio
Tocantins: 20 anos depois da usina hidrelétrica de Tucuruí. Brasília, Eletronorte,
2004. 216p.

52
SHANNON, C. E.; WEAVER, W. The Mathematical Theory of Communication.
UniversityofIllions Press, Urbana, USA, 1949.
SILVA, C. C.; FERREIRA, E. F. J.; DEUS, C. P. Dieta de cinco espécies de
Hemiodontidae (Teleostei, Characiformes) na área de influência do reservatório de
Balbina, rio Uatumã, Amazonas, Brasil. Iheringia, Sér. Zool., Porto Alegre, v. 98, n.
4, p. 464-468, 2008.
SIOLI, H. A limnologia e a sua importância em pesquisas da Amazônia.
Amazoniana, p. 11-35, 1965.
SIOLI, H. Hydrochemistry and geology in the Brazilian Amazon region. Amazoniana,
v.1, p.74-83, 1984.
SIQUEIRA-SOUZA, F. K. & FREITAS, C. E. C. Fish diversity of floodplain lakes on
the lower stretch of the Solimões river. Brazilian Journal of Biology, v. 64, n. 3, p.
501-510, 2004.
SNUC, SISTEMA NACIONAL DE UNIDADES DE CONSERVAÇÃO DA NATUREZA.
Lei Federal n. 9.985/2000, 2000.
SOARES, M. G. M.; ALMEIDA, R. G.; JUNK, W. J. The trophic status of the fish
fauna in LagoCamaleão, a macrophyte dominated floodplain lake in the middle
Amazon. Amazoniana, v. 9, n. 4, p. 511-526, 1986.
SOARES, M. G. M.; FREITAS, C. E. C.; OLIVEIRA, A. C. B. Assembleias de peixes
associadas aos bancos de macrófitas aquáticas em lagos manejados da Amazônia
Central, Amazonas, Brasil. Acta Amazonica, v. 44, n. 1, p. 143 – 152, 2014.
SOARES, M. G. M.; YAMAMOTO, K. C. Diversidade e composição da ictiofauana do
lago tupé. In: SILVA, E. N. S.; APRILE, F. M.; SCUDELLER, V. V.; MELO, S. (Eds.).
Biotupé Meio Físico, diversidade biológica e sociocultural do baixo rio Negro,
Amazônia Central. Manaus: INPA, 2005. p. 181-197.
SOARES, M. G. M; COSTA, E. L.; SIQUEIRA-SOUZA, F. K.; ANJOS, H. D. B.;
YAMAMOTO, K. C. Peixes de Lagos do Médio Rio Solimões. Manaus. Reggo
Edições, 2011. 160p.

53
TEJERINA-GARRO, F. L.; FORTIN, R.; RODRÍGUEZ, M. A. Fish community
structure in relation to environmental variation in floodplain lakes of the Araguaia
River, Amazon Basin. Environmental Biology of Fishes, v. 51, p. 399-410, 1998.
TER BRAAK, C. J. F. Ordination. In: JONGMAN, R, H. G.; TER BRAAK, C. J. F.;
VAN TONGEREN, O. F. R. (Eds.). Data analysis in community and landscape
ecology. Cambrigde University Press, Cambrigde, 1995. p.91-173.
VIEIRA, D. B.; SHIBATTA, O. A. Peixes como indicadores da qualidade ambiental do
ribeirão Esperança, município de Londrina, Paraná, Brasil. Biota Neotropica,
Campinas, v. 7, n. 1, p. 57-66, 2007.
WALKER, I. Ecologia e biologia dos igapós e igarapés. Ciência Hoje, v. 11, n. 64, p.
44-53, 1990.
WELCOMME, R. L. River fisheries. FAO Fish. Tech. Pap., 1985, n. 262, p. 330.
WOOTTON, R. J. Ecology of teleost fishes. Chapman & Hall Press, 1995. 404 p.
YABE, R. S.; BENNEMANN, T. Regime alimentar de Schizodon intermedius
Garavello & Britski do rio Tibagi, Paraná, e sua relação com algumas características
morfológicas do trato digestivo (Osteichthyes, Anostomidae). Rev. bras. Zool, v. 11,
n. 4, p. 777-788, 1994.
YAMAMOTO, K. C. A estrutura de comunidades de peixes em lagos manejados
da Amazônia Central. Manaus, Amazonas. Dissertação de Mestrado: INPA/UFAM,
2004. 71 pp.
YAMAMOTO, K. C; FREITAS, C. E. C.; ZUANON, J.; HURD, L. E. Fish diversity and
species composition in small-scale artificial reefs in Amazonian floodplain lakes:
Refugia for rare species?. Ecological Engineering. v. 67, p. 165–170, 2014.
ZAR, J. H. Biostatistical analysis. Prentice-Hall, Inc., Englewood cliffs, N. J., 1999.
4a edição. 663pp.
ZAVALA-CAMIM, L. A. Introdução aos estudos sobre alimentação em peixes.
Maringá: EDUEM, 1996. 129 pp.

54
ZUANON, J. A. Taxonomical aspects of the fish communities from a white water and
a black water in the Central Amazon. Summaties of the 1st SHIFT Workshop,
Belém, p. 5, 1993.

55
APÊNDICE
Apêndice 1. Espécies capturadas nos períodos de enchente e vazante de 2014 e enchente de 2015 em lagos do PARNAAnavilhanas.
Tabela taxonômica
Canauiri Pequeno Canauiri Grande Arraia Prato
TOTAL Igapó
Água aberta
Ordem, Família, Nome científico, Autor N Peso (g) N Peso (g) N Peso (g) N Peso (g) N Peso (g) N N
CLUPEIFORMES
Pristigasteridae
Pellona flavipinnis (Valenciennes, 1837) 7 736,9 4 720,8 8 646,9 15 1351,3 34 3455,9 15 19
Ilisha amazonica(Miranda Ribeiro, 1920) 1 42,6 0 0,0 2 104,3 1 37,6 4 184,5 4 0
Engraulidae
Lycengraulis batesii(Günther, 1868) 1 7,7 1 9,1 0 0,0 0 0,0 2 16,8 1 1
CHARACIFORMES
Acestrorhynchidae
Acestrorhynchus microlepis(Jardine, 1841) 4 314,4 2 333,9 1 22,9 1 33,3 8 704,5 8 0
Characidae
Chalceus erythrurus (Cope, 1870) 1 68,7 1 168,3 0 0,0 0 0,0 2 237,0 2 0
Ctenobrycon hauxwellianus (Cope, 1870) 1 10,1 0 0,0 0 0,0 0 0,0 1 10,1 1 0
Poptella compressa (Günther, 1864) 0 0,0 2 11,6 0 0,0 1 6,9 3 18,5 3 0
Tetragonopterus chalceus Spix&Agassiz, 1829 1 21,0 2 55,1 0 0,0 0 0,0 3 76,1 3 0

56
Ctenoluciidae
Boulengerella lucius (Cuvier, 1816) 1 106,6 0 0,0 1 155,1 1 179,7 3 441,4 1 2
Cynodontidae
Cynodon gibbus (Agassiz, 1829) 1 16,9 1 71,8 0 0,0 2 177,7 4 266,4 4 0
Rhaphiodon vulpinus Spix & Agassiz, 1829 0 0,0 5 1351,8 1 421,7 6 1600,4 12 3373,9 9 3
Iguanodectidae
Bryconops alburnoides Kner, 1858 4 157,8 5 145,9 3 90,3 7 266,0 19 660,0 10 9
Bryconidae
Brycon melanopterus (Cope, 1872) 0 0,0 2 555,1 0 0,0 1 200,9 3 756,0 3 0
Brycon pesu Müller &Troschel, 1845 0 0,0 3 110,1 0 0,0 0 0,0 3 110,1 2 1
Serrasalmidae
Serrasalmus gouldingi (Fink& Machado-Allison, 1992) 7 704,7 9 666,0 10 1106,9 7 702,4 33 3180,0 27 6
Serrasalmus elongatus Kner, 1858 1 61,8 0 0,0 0 0,0 0 0,0 1 61,8 0 1
Serrasalmus hastatus Fink & Machado-Allison, 2001 0 0,0 0 0,0 0 0,0 1 119,0 1 119,0 1 0
Serrasalmus rhombeus (Linnaeus, 1766) 41 1895,6 33 4105,4 16 2269,3 13 1755,9 103 10026,2 60 43
Metynnis hypsauchen (Müller &Troschel, 1844) 6 407,1 4 454,5 5 312,2 2 162,6 17 1336,4 17 0
Serrasalmus eigenmanni Norman, 1929 0 0,0 0 0,0 0 0,0 9 407,6 9 407,6 9 0
Metynnis argenteus Ahl, 1923 0 0,0 0 0,0 0 0,0 1 82,4 1 82,4 1 0
Metynnis sp 2 159,3 1 77,1 0 0,0 2 151,8 5 388,2 5 0
Myleus asterias (Müller &Troschel, 1844) 1 265,0 5 1091,5 0 0,0 10 1947,3 16 3303,8 16 0
Pristobrycon sp. 4 446,1 9 831,3 8 842,4 10 1189,4 31 3309,2 27 4
Hemiodontidae Argonectes longiceps (Eigenmann, 1912) 0 0,0 2 171,8 0 0,0 0 0,0 2 171,8 1 1

57
Anodus elongatus Agassiz, 1829 10 1014,4 4 249,0 0 0,0 0 0,0 14 1263,4 2 12
Hemiodus atranalis (Fowler, 1940) 0 0,0 0 0,0 1 15,5 0 0,0 1 15,5 1 0
Hemiodus immaculatus Kner, 1858 55 2922,1 12 817,8 10 718,6 73 3668,6 150 8127,1 106 44
Hemiodus unimaculatus (Bloch, 1794) 3 111,9 3 271,9 2 108,9 3 137,0 11 629,7 6 5
Triportheidae
Triportheus angulatus (Spix & Agassiz, 1829) 2 175,3 1 71,2 1 77,4 9 631,7 13 955,6 12 1
Triportheus elongatus (Günther, 1864) 10 447,4 17 1467,2 5 588,0 1 93,1 33 2595,7 28 5
Agoniates halecinus Müller &Troschel, 1845 2 83,2 0 0,0 5 148,2 8 294,6 15 526,0 3 12
Anostomidae
Anostomoides laticeps(Eigenmann, 1912) 5 946,6 0 0,0 0 0,0 0 0,0 5 946,6 5 0
Laemolyta taeniata (Kner, 1858) 4 241,2 8 881,0 3 127,3 4 472,1 19 1721,6 14 5
Leporinus fasciatus (Bloch, 1794) 2 175,1 5 967,1 2 555,9 1 203,7 10 1901,8 9 1
Pseudanos trimaculatus (Kner, 1858) 1 74,9 3 80,2 4 122,8 0 0,0 8 277,9 7 1
Prochilodontidae
Semaprochilodus insignis (Jardine, 1841) 0 0,0 0 0,0 1 355,1 0 0,0 1 355,1 1 0
Semaprochilodus taeniurus (Valenciennes, 1821) 0 0,0 5 1083,4 2 479,8 0 0,0 7 1563,2 3 4
Curimatidae Cyphocharax abramoides (Kner, 1858) 1 96,7 0 0,0 3 244,4 1 130,3 5 471,4 4 1
SILURIFORMES
Loricariidae
Ancistrus dolichopterus Kner, 1854 0 0,0 1 17,8 0 0,0 0 0,0 1 17,8 1 0
Dekeyseria scaphirhyncha (Kner, 1854) 1 56,4 0 0,0 0 0,0 0 0,0 1 56,4 0 1

58
Pimelodidae 0 0,0
Pimelodina flavipinnis (Steindachner, 1877) 0 0,0 1 468,0 3 815,8 1 600,2 5 1884,0 5 0
Hypophthalmus marginatus (Valenciennes, 1840) 8 893,8 2 339,7 9 1515,1 8 1477,5 27 4226,1 11 16
Hypophthalmus fimbriatus (Kner, 1858) 1 121,1 6 723,5 2 276,4 7 850,9 16 1971,9 10 6
Hypophthalmus edentatus (Spix & Agassiz, 1829)
8 906,1 1 87,9 23 2921,7 9 1176,0 41 5091,7 20 21
Brachyplatystoma platynemum Boulenger, 1898 0 0,0 1 148,3 0 0,0 0 0,0 1 148,3 1 0
Pinirampus pirinampu (Spix &A gassiz, 1829) 2 264,6 9 4026,1 2 671,1 1 55,1 14 5016,9 5 9
Sorubim lima (Bloch & Schneider, 1801) 0 0,0 0 0,0 0 0,0 1 50,9 1 50,9 0 1
Auchenipteridae Auchenipterus nuchalis (Spix & Agassiz, 1829) 0 0,0 2 50,7 0 0,0 3 106,4 5 157,1 2 3
Auchenipterichthys longimanus (Günther, 1864) 20 666,5 31 950,5 15 498,5 33 1136,0 99 3251,5 80 19
Centromochlus macracanthus (Soares-Porto de 2000) 0 0,0 2 73,5 0 0,0 2 31,4 4 104,9 2 2
Ageneiosus polystictus Steindachner, 1915 0 0,0 11 3342,9 10 2277,1 7 1517,4 28 7137,4 18 10
Ageneiosus ucayalensis Castelnau, 1855 3 89,2 4 445,4 5 187,2 4 201,3 16 923,1 8 8
Ageneiosus vittatus Steindachner, 1908 1 9,0 1 9,8 0 0,0 0 0,0 2 18,8 1 1
Tatia intermedia (Steindachner, 1877) 6 89,3 7 110,4 3 51,0 3 59,9 19 310,6 18 1
Doradidae
Astrodoras asterifrons (Kner, 1853) 1 1,9 0 0,0 0 0,0 0 0,0 1 1,9 1 0
Trachydoras nattereri (Steindachner, 1881) 0 0,0 0 0,0 0 0,0 1 8,4 1 8,4 1 0
PERCIFORMES
Sciaenidae Plagioscion squamosissimus (Heckel, 1840) 5 633,1 6 2263,1 5 1258,0 1 166,0 17 4320,2 15 2
Cichlidae

59
Geophagus proximus (Castelnau, 1855) 2 385,7 1 167,8 1 186,8 0 0,0 4 740,3 4 0
Cichla temensis Humboldt, 1821 1 412,0 1 51,6 2 195,3 1 66,7 5 725,6 4 1
Cichla monoculus Agassiz, 1831 0 0,0 0 0,0 1 86,1 2 850,8 3 936,9 3 0
Heros efasciatus Heckel, 1840 0 0,0 0 0,0 0 0,0 1 178,4 1 178,4 1 0
Heros severus Heckel, 1840 1 161,8 0 0,0 0 0,0 1 126,3 2 288,1 2 0
Mesonauta festivus (Heckel, 1840) 0 0,0 0 0,0 0 0,0 1 17,8 1 17,8 1 0
Uaru amphiacanthoides Heckel, 1840 1 237,6 0 0,0 1 185,1 2 452,5 4 875,2 4 0
TOTAL 240 16639,2 236 30096,9 176 20639,1 279 25133,2 931 92508,4 649 282

60
Apêndice 2. Número de estômagos analisados por indivíduo, Índice de importância alimentar (IAi) e guilda trófica encontrada para as 24 espécies
analisadas pertencentes a floresta de igapó dos quatro lagos do PARNA Anavilhanas. IN = insetos; P = peixes; FS = frutos/sementes; MV =
material vegetal; MD = material digerido; CR = crustáceos; RE = reptéis; ZOO = zooplâncton.
Espécie N Índice de Importância alimentar (%)
Guilda trófica IN P FS MV CR RE ZOO
Pellona flavipinnis 15 69,52 8,81 0,00 4,87 16,80 0,00 0,00 carnívora com
tendência a insetivoria
Pristobrycon sp. 27 2,23 9,02 84,07 4,55 0,09 0,00 0,00 frugívora
Laemolyta taeniata 14 0,00 0,00 1,62 98,38 0,00 0,00 0,00 herbívora
Metynnis hypsauchen 16 13,96 0,00 0,00 86,04 0,00 0,00 0,00 herbívora
Myleus asterias 16 0,52 0,00 24,44 75,04 0,00 0,00 0,00 herbívora
Pseudanos trimaculatus 7 0,00 18,18 9,09 72,73 0,00 0,00 0,00 herbívora
Auchenipterichthys longimanus 80 94,53 0,04 5,40 0,03 0,00 0,00 0,00 insetívora
Bryconops alburnoides 10 97,30 2,70 0,00 0,00 0,00 0,00 0,00 insetívora
Hypophthalmus marginatus 8 72,41 0,00 0,00 0,00 0,00 0,00 27,59 insetívora
Tatia intermedia 17 98,37 0,00 0,00 0,81 0,81 0,00 0,00 insetívora
Triportheus angulatus 11 96,20 0,00 1,19 2,61 0,00 0,00 0,00 insetívora
Triportheus elongatus 28 71,92 0,00 21,06 7,02 0,00 0,00 0,00 insetívora
Leporinus fasciatus 8 53,73 7,46 0,00 34,83 3,98 0,00 0,00 onívora
Hemiodus immaculatus 102 50,46 0,00 9,06 16,28 24,19 0,00 0,00 onívora com tendência
a insetivoria
Serrasalmus eigenmanni 9 0,00 60,98 0,00 39,02 0,00 0,00 0,00 onívora com tendência
a piscivoria
Acestrorhynchus microlepis 8 0,00 100,00 0,00 0,00 0,00 0,00 0,00 piscívora
Ageneiosus polystictus 17 9,09 81,82 0,00 0,00 9,09 0,00 0,00 piscívora
Ageneiosus ucayalensis 8 0,00 100,00 0,00 0,00 0,00 0,00 0,00 piscívora
Plagioscion squamosissimus 15 0,35 97,89 0,00 0,00 1,75 0,00 0,00 piscívora
Rhaphiodon vulpinus 8 1,27 98,73 0,00 0,00 0,00 0,00 0,00 piscívora

61
Serrasalmus gouldingi 26 3,42 73,34 22,30 0,00 0,94 0,00 0,00 piscívora
Serrasalmus rhombeus 59 12,56 76,77 6,74 3,24 0,63 0,06 0,00 piscívora
Hypophthalmus edentatus 16 0,20 0,00 7,89 35,90 0,00 0,00 56,02 zooplanctívora
Hypophthalmus fimbriatus 8 0,77 0,00 7,89 3,85 9,23 0,00 86,15 zooplanctívora