universidade federal do parÁ instituto de … rubens alex de... · ipa incidência parasitária...
TRANSCRIPT

UNIVERSIDADE FEDERAL DO PARÁ
INSTITUTO DE CIÊNCIAS BIOLÓGICAS
PROGRAMA DE PÓS-GRADUAÇÃO
BIOLOGIA DE AGENTES INFECCIOSOS E PARASITÁRIOS
COINFECÇÃO DO PLASMODIUM VIVAX E ENTEROPARASITOS NO MUNICÍPIO
DO OIAPOQUE, ESTADO DO AMAPÁ, FRONTEIRA BRASIL-GUIANA
FRANCESA
RUBENS ALEX DE OLIVEIRA MENEZES
Belém-Pará
2017

Dados Internacionais de Catalogação- na-Publicação (CIP) Biblioteca do Instituto de Ciências Biológicas - UFPA
Menezes, Rubens Alex de Oliveira
Coinfecção do Plasmodium vivax e enteroparasitos no município do
Oiapoque, estado do Amapá, fronteira Brasil-Guiana Francesa / Rubens
Alex de Oliveira Menezes ; Orientador, Ricardo Luiz Dantas Machado. -
2017.
139 f.: il.
Inclui bibliografias
Tese (Doutorado) - Universidade Federal do Pará, Instituto de
Ciências Biológicas, Programa de Pós-graduação em Biologia de Agentes
Infecciosos e Parasitários, Belém, 2017.
1. Doenças parasitárias – Oiapoque (AP). 2. Plasmodium vivax –
Oiapoque (AP). 3. Parasitos intestinais. 4. Malária – Oiapoque (AP) I.
Machado, Ricardo Luiz Dantas, orientador. II. Titulo.
CDD – 22 ed. 571.999098116

RUBENS ALEX DE OLIVEIRA MENEZES
COINFECÇÃO DO PLASMODIUM VIVAX E ENTEROPARASITOS NO MUNICÍPIO
DO OIAPOQUE, ESTADO DO AMAPÁ, FRONTEIRA BRASIL-GUIANA
FRANCESA
Tese apresentada ao Programa de Pós-Graduação
em Biologia de Agentes Infecciosos e Parasitários
do Instituto de Ciências Biológicas da
Universidade Federal do Pará como requisito
parcial para a obtenção do grau de Doutor em
Biologia de Agentes Infecciosos e Parasitários.
Orientador: Prof. Dr. Ricardo Luiz Dantas
Machado
Belém-Pará
2017

1
RUBENS ALEX DE OLIVEIRA MENEZES
COINFECÇÃO DO PLASMODIUM VIVAX E ENTEROPARASITOS NO MUNICÍPIO
DO OIAPOQUE, ESTADO DO AMAPÁ, FRONTEIRA BRASIL-GUIANA
FRANCESA
Tese apresentada ao Programa de Pós-Graduação em Biologia de Agentes Infecciosos e
Parasitários do Instituto de Ciências Biológicas da Universidade Federal do Pará como
requisito parcial para o grau de Doutor em Biologia de Agentes Infecciosos e Parasitários.
Orientador: Prof. Dr. Ricardo Luiz Dantas Machado
Departamento de Microbiologia e Parasitologia
Universidade Federal Fluminense
Banca examinadora: Prof. Dr. Antonio Carlos Rosário Vallinoto
Laboratório de Virologia ICB, UFPA
Prof. Dra. Ana Maria Revorêdo da Silva Ventura
Instituto Evandro Chagas, SVS – MS e UEPA
Prof. Dr. Adriano Penha Furtado
Laboratório de Biologia Celular e Helmintologia, ICB-UFPA
Prof. Dra. Edilene Oliveira da Silva
Instituto de Ciências Biológicas ICB-UFPA
Prof. Dra. Jeannie Nascimento dos Santos (Suplente)
Laboratório de Biologia Celular e Helmintologia, ICB/UFPA
Belém, 20 de Setembro de 2017

2
Dedico este trabalho à minha família pelo apoio,
compreensão e por compartilhar todos os momentos de
minha vida, além de todos que me apoiaram direta ou
indiretamente durante minha jornada acadêmica e
profissional.

3
AGRADECIMENTOS
A Deus força iluminada e infinita por ter guiado nesta longa trajetória profissional,
colocando sempre em meu caminho pessoas que me ajudaram e ensinaram muito.
Ao meu orientador Dr. Ricardo Luiz Dantas Machado, minha imensa gratidão pela
confiança, acolhimento e pela oportunidade de aprendizado nas orientações referentes ao
andamento do projeto, sempre muito valioso para o desenvolvimento dessa tese, contribuindo
para o meu crescimento acadêmico e profissional.
À Dra. Margarete do Socorro Mendonça Gomes por sua amizade incondicional, pela
dedicada orientação e sabedoria nas abordagens dos problemas científicos e por me receber de
portas abertas todas as vezes que precisei, compartilhando sua sabedoria e me incentivando a
ver a grandiosidade do mundo científico.
A toda minha família por todo amor carinho, dedicação, zelo, apoio, consolo e atitudes
certas em horas incertas além de paciência e ajuda tão necessários em todo o decorrer de
minha jornada acadêmica e profissional.
A minha esposa e companheira, Petille Santos Souza pelo seu carinho, amor, ajuda,
incentivo, compreensão e tolerância nas horas e momentos difíceis de ausência.
Ao Laboratório Central de Saúde Pública do Amapá (LACEN-AP), pelo apoio
logístico, autorizando minhas atividades laborais e de pesquisa no Laboratório de Fronteira
(LAFRON) em Oiapoque, durante a fase de acompanhamentos clínico-laboratorial dos
pacientes inseridos na pesquisa.
Aos técnicos, microscopistas e notificantes de Endemias do Município de Oiapoque
pelo auxilio na coleta das amostras e apoio técnico.
Ao setor de controle de qualidade de endemias do Município de Oiapoque e do
Laboratório Central de Saúde Pública do Amapá, pela avaliação semi quantitativa das lâminas
de malária.
Aos biomédicos: Romulo Lima de Sousa do LAFRON e Silvestre Rodrigues do
Nascimento do Hospital Estadual de Oiapoque, que realizaram as análises hematológicas,
processaram e armazenaram todas as amostras sanguíneas.
As pessoas que me ofereceram moradia e me acolheram em Oiapoque, Belém e em
Fortaleza, nas muitas vezes em que precisei me ausentar de minha cidade para desenvolver as
atividades da tese: Anapaula Martins Mendes, Leila Margareth Carrera Menezes e Heli da
Silva Araújo Salles (in memoriam).

4
À Coordenação do Curso de Pós-Graduação em Biologia de Agentes Infecciosos e
Parasitários-UFPA, pelo acolhimento humanizado e conhecimentos adquiridos. Aos
professores do Curso pela competência, generosidade intelectual e conhecimentos repassados
ao longo de minha jornada acadêmica.
À Dra. Maristela Gomes da Cunha, pela oportunidade de realizar os testes de ELISA
para a MSP-119 no laboratório de Microbiologia e imunologia do Instituto de Ciências
Biológicas da Universidade Federal do Pará (ICB-UFPA).
Aos profissionais do Instituto Evandro Chagas - IEC/SVS/MS pela forma cordial e
profissional que sempre fui tratado.
Aos profissionais da Seção de Parasitologia do Instituto Evandro Chagas, em especial
a Tamirys Pina Simão do Laboratório de Imunogenética em Malária, por ter me recebido para
as análises de citocinas, por sua prestimosa atenção durante a fase de execução das análises
laboratoriais e sua inestimável contribuição na análise estatística dos resultados.
À Maria Izabel de Jesus responsável pelo Laboratório de Biologia Humana da Seção
de Meio Ambiente/IEC, pela disponibilidade dos kits de citocinas e do laboratório para as
análises e Aline Collares Pinheiro de Sousa por toda hospitalidade, gentileza e prontidão em
colaborar com este trabalho, me acompanhando no citômetro e nas dosagens sérica do teor de
citocinas.
À Coordenação e professores do curso de enfermagem da universidade Federal do
Amapá (UNIFAP), na qual estou vinculado, pela liberação das minhas atividades laborais,
durante a fase final de redação da tese.
Ao grupo de pesquisadores que contribuíram na análise, interpretação e discussão dos
dados, para submissão e publicação de artigos em periódicos científicos nacionais e
internacionais.
Antecipadamente, aos membros da banca por aceitarem o convite para avaliar esta
tese.
Aos pacientes que colaboraram com o projeto e a todos que contribuíram direta ou
indiretamente para a realização desse trabalho, e que me é impossível relatar individualmente.
A cada um o meu muito obrigado!

5
SÚMARIO
LISTA DE FIGURAS.................................................................................. 7
LISTA DE ABREVIATURAS.................................................................... 8
RESUMO...................................................................................................... 10
ABSTRACT.................................................................................................. 11
1 INTRODUÇÃO........................................................................................... 12
1.1 CONSIDERAÇÕES GERAIS DA MALÁRIA............................................ 12
1.2 ASPECTOS EPIDEMIOLÓGICOS DA MALÁRIA................................... 13
1.2.1 Epidemiologia da Malária no Município de Oiapoque............................ 20
1.3 PARASITOS GASTROINTESTINAIS NO BRASIL.................................. 26
1.4 COINFECÇÃO MALÁRIA E PARASITOS GASTROINTESTINAIS...... 29
1.5 IMUNOPATOGÊNESE DA COINFECÇÃO MALÁRIA E
ENTEROPARASITOSES.............................................................................
32
1.5.1 Resposta imune de anticorpos e citocinas.................................................. 38
1.6 JUSTIFICATIVA.......................................................................................... 42
1.7 HIPÓTESES.................................................................................................. 43
1.7.1 Hipótese Nula............................................................................................... 43
1.7.2 Hipótese Alternativa................................................................................... 43
1.8 OBJETIVOS................................................................................................. 44
1.8.1 Geral............................................................................................................. 44
1.8.2 Específicos.................................................................................................... 44
2 MATERIAL E MÉTODOS........................................................................ 45
2.1 ÁREA DE ESTUDO E POPULAÇÃO........................................................ 45
2.1 CRITÉRIOS DE SELEÇÃO......................................................................... 46
2.1.1 Critérios de inclusão para paciente maláricos.......................................... 47
2.1.2 Critérios de exclusão................................................................................... 47
2.1.3 Critérios de inclusão para indivíduos não maláricos............................... 47
2.1.4 Critérios de exclusão................................................................................... 47
2.3 TAMANHO DA AMOSTRA........................................................................ 48
2.4 COLETA E EXAME DE SANGUE............................................................. 48
2.4.1 Coleta Sanguínea......................................................................................... 48
2.4.2 Diagnóstico de Malária................................................................................ 48
2.4.3 Detecção de Gametócitos............................................................................. 49

6
2.4.4 Concentração de Hemoglobina................................................................... 49
2.4.5 Controle de Qualidade do diagnóstico parasitológico.............................. 49
2.5 COLETA E EXAME DE AMOSTRAS FECAIS......................................... 49
2.6 DIAGNÓSTICO MOLECULAR DE PLASMÓDIO................................... 50
2.7 AVALIAÇÃO DE ANTICORPOS IgG CONTRA A PVMSP-119.............. 50
2.8 NÍVEIS SÉRICOS DE CITOCINAS............................................................ 51
2.9 ANÁLISE ESTATÍSTICA............................................................................ 52
2.10 CONSIDERAÇÃO ÉTICA........................................................................... 53
3 RESULTADOS............................................................................................ 54
3.1 MANUSCRITO 1.......................................................................................... 54
3.2 MANUSCRITO 2.......................................................................................... 67
4 CONCLUSÃO.............................................................................................. 92
REFERÊNCIA BIBLIOGRÁFICA........................................................... 93
APÊNDICE A - Termo de Consentimento Livre e Esclarecido.................. 110
APÊNDICE B – Formulário epidemiológico............................................... 112
APÊNDICE C - Resumos Aprovados em Congresso.................................. 113
APÊNDICE D - Manuscrito Extra............................................................... 126
ANEXO A - Parecer Consubstanciado do Comitê de Ética de Pesquisa...... 138
ANEXO B - Financiamento pelo Edital/CNPq/PPSUS – Amapá 2013....... 139

7
LISTA DE FIGURAS
Figura 1 - Série histórica do número de casos na região Amazônica no período de
1970 a 2014......................................................................................................................
15
Figura 2 - Áreas de transmissão de malária no Brasil de acordo com Incidência
Parasitária Anual (IPA) no ano de 2015..........................................................................
16
Figura 3 - Casos de malária e Incidência Parasitária Anual (IPA) no ano de 2014....... 17
Figura 4 - Número de casos de malária notificados no Estado do Amapá e diferença
percentual entre os anos de 2003 a 2017..........................................................................
18
Figura 5 - numero de caso de malária no estado do Amapá, por categoria de local
provável de infecção e diferença percentual entre os anos de 2012 a 2016.....................
19
Figura 6 - Série histórica de casos totais e Incidência Parasitária Anual durante o
período de 2003 a 2014 no município de Oiapoque-Amapá...........................................
22
Figura 7 - Casos de malária por P. vivax segundo local provável de infecção e
importados dos garimpos clandestinos da Guiana Francesa para Oiapoque...................
22
Figura 8 - Serie histórica do número de casos de malária notificados no município de
Oiapoque e diferença percentual entre os anos de 2003 a 2016......................................
24
Figura 9 - Serie histórica do número de casos de malária por categoria de local
provável de infecção notificados no município de Oiapoque e diferença percentual
entre os anos de 2003 a 2016...........................................................................................
25
Figura 10 - Frequência da distribuição de geohelmintos no Brasil de 1995 a 2013. a)
A. lumbricoides; b) T. trichiura e c) Ancilostomídeos....................................................
28
Figura 11 - Mapa da América do Sul, mostrando a localização da Guiana Francesa e
Oiapoque no Estado do Amapá........................................................................................
45

8
LISTA DE ABREVIATURAS
AC Estado do Acre
ADCI Inibição celular dependente de anticorpos
AM Estado do Amazonas
AMA-1 Antígeno apical de Membrana-1
ANOVA Análise de variância
AP Estado do Amapá
BAIP Biologia de Agentes Infecciosos e Parasitários
CONSEP Conselho Superior de Ensino, pesquisa e extensão
DATASUS Departamento de Informática do Sistema Único de Saúde
DSEI Distrito Sanitário Especial Indígena
ELISA Ensaio de imunoabsorção enzimática
FC€RII Receptor para IgE de baixa afinidade
Grupo CE Controle endêmico (sem infecção para malária e parasitos intestinais)
Grupo CI Coinfectados (Infectados com malária vivax e parasitos intestinais)
Grupo E Enteroparasitas (Infectados com parasitos intestinais)
Grupo M Maláricos (infectados com malária vivax)
GM-CSF Fator Estimulador de Colônias de Granulócitos e Macrófagos
HEMOPA Fundação Centro de Hemoterapia e Hematologia do Pará
IBGE Instituto Brasileiro de Geografia e Estatística
IFN-γ Interferon gama
IgG Imunoglobulina G
IgG1 Imunoglobulina variantes isotípicas ou subclasses 1
IgG2 Imunoglobulina variantes isotípicas ou subclasses 2
IgG3 Imunoglobulina variantes isotípicas ou subclasses 3
IgG4 Imunoglobulina variantes isotípicas ou subclasses 4
IgA Imunoglobulina A
IgE Imunoglobulina E
IgM Imunoglobulina M
IL-2 Interleucina-2
IL-3 Interleucina-3
IL-4 Interleucina-4
IL-5 Interleucina-5

9
IL-6 Interleucina-6
IL-10 Interleucina-10
IPA Incidência Parasitária Anual
IR Índice de reatividade
LACEN/AP Laboratório Central de Saúde Pública Amapá
LAFRON Laboratório de Fronteira
MA Estado do Maranhão
MFI Media de Intensidade de Fluorescência
MS Ministério da Saúde
MSP-1 Proteína de superfície de merozoíto-1
MT Estado do Mato Grosso
NEM Número de episódios de malária anterior
NK Exterminadoras naturais
OD Densidade ótica
OPD Cromógeno Orto-fenilenodiamina
PA Estado do Pará
PBS Salina tamponada com fosfato
PCR Reação em cadeia da polimerase
PNM Programa Nacional de controle de malária
PUM Período da última malária (meses)
RO Estado do Rondônia
RO Estado do Roraima
SINAN Sistema de Informação de Agravos de Notificação
SIVEP Sistema Informatizado de Vigilância Epidemiológica
SVS Secretaria de Vigilância em Saúde
TCLE Termo de consentimento livre e esclarecido
TGF-β Fator de transformação do crescimento beta
TH1 Subdivisões fenotípicas de linfócitos auxiliares 1
TH2 Subdivisões fenotípicas de linfócitos auxiliares 2
TNF-α Fator de necrose tumoral alpha
TO Estado do Tocantins
TR Tempo de residência no Oiapoque (ano)
UFPA Universidade Federal do Pará

10
RESUMO
A coinfecção malária-parasitoses intestinais são comuns nas regiões tropicais do planeta,
embora prevalentes, pouco se sabe sobre essa interação na epidemiologia e no impacto da
resposta imune. Este estudo avaliou a influência da coinfecção das enteroparasitoses na
malária causada pelo Plasmodium vivax em uma área fronteiriça entre Brasil e Guiana
Francesa. Levantamento transversal realizado em Oiapoque, município do Amapá, norte do
Brasil. O diagnóstico da malária foi realizado por meio da gota espessa, a dosagem de
hemoglobina por método automatizado, o exame coproparasitológico pelos métodos de
Hoffman-Pons-Janer e Faust. Avaliação de anticorpos IgG contra a PvMSP-119 foi realizada
por ELISA e a quantificação das citocinas Th1 (IFN-γ, TNF-α e IL-2) e Th2 (IL-4, IL-5 e IL-
10) por citometria de fluxo. Análise estatística foi utilizada para avaliar os parâmetros
investigados, sendo comparados os resultados a partir de quatro grupos distintos agrupados
em M os monoinfectados com malária vivax; CI os coinfectados malária vivax-
enteroparasitos; E os monoinfectados com enteroparasitos e CE controle endêmico (negativo
para ambas enfermidades). Foram incluídos 441 indivíduos agrupados de acordo com o seu
estado de infecção: [M 6,9% (30/441)], [CI 26,5% (117/441)], [E 32,4% (143/441)] e [CE
34,2% (151/441)]. O gênero masculino prevaleceu entre os grupos M (77% - 23/30) e CI
(60% - 70/117). Nos indivíduos investigados, 59% (261/441) foram infectados por algum tipo
de parasito intestinal. Entre estes, 45,2% (118/161) estava infectado somente por helmintos,
40,9% (107/261) somente protozoários, e 13,8% (36/261) tinham infecções associadas.
Dentre os helmintos detectados o Ascaris lumbricoides 19,9% (52/261) foi o mais prevalente,
seguido de Entamoeba histolytica como protozoários mais frequentes 5% (13/261). A
pesquisa evidenciou uma correlação entre o nível de hemoglobina e a presença das parasitoses
detectadas (coeficiente C = 0.2195 e p < 0.0001). Adicionalmente pelo teste qui-quadrado de
Tendência houve correlação entre a modalidade de enteroparasitos e o nível de Hemoglobina,
sendo apenas helminto (p < 0.0001), monoparasitado (p < 0.0001) e poliparasitado (p <
0.0031). Também foram calculadas utilizando um Tukey a partir e uma análise de variância,
ANOVA um critério, havendo diferença entre os níveis de hemoglobina nos diferentes grupos
estudados: CE/E, CE/CI, E/M e CI/M com (p < 0.01). A anemia foi expressa em percentuais,
considerado anêmico indivíduos com níveis de hemoglobina de ≤ 13 g/dL para homens e ≤ 12
g /dL para mulheres, utilizado o teste Kruskal-Wallis-Dunn entre os grupos, sendo [CI-CE (p
< 0.05)]. Foram observadas diferenças entre a parasitemia e de gametócitos entre os grupos M
e CI grupos (Wilcoxon-Mann-Whitney, p < 0.05). Com relação aos anticorpos contra PvMSP-
119 51,2% (226/441) da população foram respondedores, sobretudo para o grupo CI 81,1%
(95/117) apresentando perfil diferente quanto à reatividade. As análises dos níveis séricos de
citocinas da população revelou uma grande variação nas concentrações em todas as citocinas
de perfil inflamatório e anti-inflamatório, sendo as citocinas TNF-α e IL-10 aumentados para
os indivíduos do grupo coinfectado. A coinfecção não esteve associada com a redução de
malária (incidência, prevalência ou redução da parasitemia). Adicionalmente os resultados
apontam a coinfecção malária-enteroparasitoses um aspecto real no município do Oiapoque, e
avaliar a possibilidade de que populações infectadas por parasitoses intestinais possam
influenciar na dinâmica de transmissão, no cenário e desfecho da resposta imune na malária
de Plasmodium vivax, são fundamentais para estratégias diagnósticas e preventivas.
Palavras chaves: Malária, enteroparasitoses, coinfecção, citocinas, Amazônia brasileira.

11
SUMMARY
Malaria coinfection-intestinal parasites are common in tropical regions of the planet, although
prevalent, little is known about this interaction in epidemiology and the impact of immune
response. This study evaluated the influence of coinfection of enteroparasitoses on malaria
caused by Plasmodium vivax in a border area between Brazil and French Guiana. Transverse
survey carried out in Oiapoque, Amapá municipality, northern Brazil. The diagnosis of
malaria was made by means of a thick drop, hemoglobin concentration by automated method,
parasitological analyses of faeces by Hoffman-Pons-Janer and Faust methods. Moreover,
evaluation of IgG antibodies against PvMSP-119 was performed by ELISA and the
quantification of Th1 cytokines (IFN-γ, TNF-α and IL-2) and Th2 (IL-4, IL-5 and IL-10) by
flow cytometor. Statistical analysis was used to evaluate the parameters investigated,
comparing the results from four different groups grouped in M monoinfected with vivax
malaria; CI co-infected malaria vivax-enteroparasites; plus the enteroparasites monoinfected
infection and endemic control EC (negative for both diseases). A total of 441 individuals were
grouped according to their infection status: [M 6.9% (30/441)], [CI 26.5% (117/441)], [E
32.4% (143/441 )] and [EC 34.2% (151/441)]. The male gender prevailed among the M (77%
- 23/30) and CI (60% - 70/117) groups. In the investigated individuals, 59% (261/441) were
infected by some type of intestinal parasite. Among these, 45.2% (118/161) was infected only
by helminths, 40.9% (107/261) only protozoa, and 13.8% (36/261) had associated infections.
Among the helminths detected, Ascaris lumbricoides 19.9% (52/261) was the most prevalent,
followed by Entamoeba histolytica as most frequent protozoa 5% (13/261). The research
showed a correlation between the hemoglobin level and the presence of parasites detected
(coefficient C = 0.2195 and p <0.0001). In addition, the chi-square test showed a correlation
between enteroparasites and hemoglobin levels, being only helminth (p <0.0001),
monoparasite (p <0.0001), and polyparasite (p <0.0031). They were also calculated using a
Tukey from and an analysis of variance, ANOVA a criterion, with difference between
hemoglobin levels in the different groups studied: CE/E, CE/CI, E/M and CI/M with (p <0.01
). Anemia was expressed in percentages, considered an anemic individuals with hemoglobin
levels of ≤ 13 g/dL for men and ≤ 12 g/dL for women, using the Kruskal-Wallis-Dunn test
between the groups, being [CI-CE (p <0.05)]. Differences of parasitemia and gametocytes
between groups M and CI were observed (Wilcoxon-Mann-Whitney test, p <0.05). Regarding
the antibodies response against PvMSP-119, 51.2% (226/441) of the population were
responders, especially for the CI group 81.1% (95/117) presenting a different reactivity
profile. Serum cytokine evaluation in the population revealed a wide variation in
concentrations of all inflammatory and anti-inflammatory cytokines, where TNF-α and IL-10
being increased for individuals in the coinfected group. Coinfection was not associated with
the reduction of malaria (incidence, prevalence or reduction of parasitemia). In addition, the
results indicate that malaria-intestinal parasitosis coinfection is a real issue in the municipality
of Oiapoque, and evaluate the possibility that populations infected by intestinal parasites may
influence the transmission dynamics, scenario and outcome of the immune response in
Plasmodium vivax malaria, are fundamental for diagnostic and preventive strategies.
Key words: Malaria, enteroparasitoses, coinfection, cytokines, Brazilian Amazon.

12
1 INTRODUÇÃO
1.1 CONSIDERAÇÕES GERAIS DA MALÁRIA
A malária é uma infecção de manifestação principalmente aguda, caracterizada pela
tríade de calafrios, febre e sudorese excessiva, sendo uma das maiores causas de morbidade e
mortalidade em muitos países tropicais e subtropicais do mundo (Jain et al., 2010). Os
parasitos dessa nosologia são protozoários pertencentes ao gênero Plasmodium e atualmente,
cinco espécies reconhecidas infectam o homem, Plasmodium falciparum, Plasmodium vivax,
Plasmodium malariae, Plasmodium ovale e o Plasmodium knowlesi. Dentre esses, os dois
primeiros estão associados aos maiores índices de mortalidade/virulência e disseminação no
mundo (Andrade et al., 2010; Gonçalves et al., 2010; WHO, 2011), e a forma crônica em
humanos é mais observada pelos P. vivax, P. ovale e P. malariae.
Dos conhecidos parasitos da malária humana, somente o P. falciparum, P. vivax e o P.
malariae são detectados no Brasil (Oliveira-Ferreira et al., 2010). Nos últimos anos, o P.
vivax tem sido a espécie mais prevalente no país, responsável por aproximadamente 80% dos
casos, enquanto o P. falciparum é responsável por aproximadamente 20%, e o P. malariae
raramente detectado. Historicamente casos clínicos graves por P. vivax são raros (Andrade et
al., 2010), no entanto, estudos recentes reforçam a associação dessa espécie com
complicações clínicas e casos fatais, sendo este um motivo de preocupação para a saúde
pública (Alecrim et al.; 2006; Lacerda et al., 2008; Andrade et al., 2010).
A malária é uma doença de transmissão vetorial. O ciclo do Plasmodium é bastante
complexo, incluindo o homem como hospedeiro intermediário e as fêmeas do mosquito do
gênero Anopheles como hospedeiros definitivos. Observa-se que o mosquito anofelino
exercendo a hematofagia no hospedeiro intermediário infectado, ingere as formas sanguíneas
do parasito, porém, somente os gametócitos são capazes de evoluir no inseto, as demais
formas degeneram-se no estômago do inseto, o gametócito feminino amadurece e transforma-
se no macrogameta, o gametócito masculino, por um processo de exoflagelação, origina os
microgametas (quatro a oito) (Mueller et al., 2009).
O P. vivax possui algumas particularidades diferentes do P. falciparum sendo uma
delas persistir como hipnozoíto nos hepatócitos por meses ou anos antes de iniciar uma
infecção sanguínea, o que explica as recaídas tardias após a cura da infecção original. Até o
momento, os fatores inerentes ao hospedeiro e ao parasito que determinam o número e a
duração destas recaídas ainda não estão plenamente esclarecidos (Mueller et al., 2009).
Alguns autores acreditam que a picada de mosquitos não infectados leva a uma sinalização

13
para ativação dos hipnozoítos latentes, resultando em episódios de recaída (Huldén et al.,
2008) propondo que este seja um mecanismo de sobrevivência do parasito e conservação da
espécie (Wells et al., 2010).
1.2 ASPECTOS EPIDEMIOLÓGICOS DA MALÁRIA
A malária é uma doença da antiguidade, foi reconhecida por Hipócrates e descrita
possivelmente em textos chineses antigos. Acompanhou a saga migratória do ser humano
pelas regiões do Mediterrâneo, Mesopotâmia Índia e Sudeste Asiático. Constituiu-se, assim,
como um dos principais obstáculos ao desenvolvimento de várias comunidades em diversos
países (Cesário & Cesário, 2006). Contudo, o homem passou a ser hospedeiro ocasional ao
ocupar os espaços onde antes existiam apenas os animais, alvos preferidos dos mosquitos
transmissores da malária. Adicionalmente o início da atividade agrícola e os processos
migracionais potencializaram a relação tênue entre o homem e a malária (Ferreira et al.,
2010).
No ano de 2012, 3,4 bilhões de pessoas no mundo estavam em áreas de risco de
malária, com uma estimativa de 207 milhões de casos e 627.000 mortes. Das mortes
estimadas, 90% ocorreram na África Subsaariana em crianças menores de 5 anos de idade
(77% dos casos). As taxas de mortalidade por malária caíram em todo o mundo cerca de 45%,
devido às medidas de controle empregadas contra a doença. Isso tem sido observado desde
2000, onde mais da metade dos países com transmissão contínua da malária registram
diminuições na incidência, internação e morte por malária (WHO, 2013).
Essa contínua diminuição na incidência da malária no mundo de 227 milhões, no ano
2000, para 198 milhões em 2013, reflete na redução de 100% na região Europeia, 76% na
região das Américas, e 34% na região Africana, ao longo dos anos. Na última década, houve
um progresso de controle e prevenção da malária sem precedentes na história da região
africana, as taxas de mortalidade, nesse período, ocasionada pela malária, foram reduzidas em
54%. Essa redução se deu pelo fato do significativo aumento de intervenções de controle do
vetor, na oferta de testes rápidos de diagnóstico e tratamento com terapias combinadas à base
de artemisinina (WHO, 2014).
Cerca de 60% dos casos de malária registrados na América do Sul são provenientes da
Amazônia Legal (Areválo-Herrera et al., 2012). No Brasil, a área endêmica compreende a
região da Amazônia Legal, que inclui os Estados do Acre, Amapá, Pará, Rondônia,
Amazonas, Tocantins, Mato Grosso, Maranhão e Roraima. Os casos registrados nessa região

14
respondem por 99% dos casos autóctones de malária, no país, e 80% dos casos registrados
fora da região Amazônica são importados dos estados pertencentes à esta área (Brasil, 2014;
Parise, 2014).
Em 2003, foi implantado o sistema Informatizado de Vigilância Epidemiológica em
malária (SIVEP-Malária) pelo Programa Nacional de controle de malária (PNCM) do
Ministério da Saúde (MS), permitindo monitorar e dar cobertura a toda a região malárica,
além de orientar os programas estaduais e municipais e gerar informações de forma a
estratificar as áreas de alto risco com maior facilidade. Mas, apesar de a ferramenta ter
melhorado os serviços de vigilância epidemiológica, não se tornou prática rotineira o uso de
mecanismos automatizados para detecção de epidemias, conforme sugestões internacionais
(Braz et al., 2013).
Em 2010, foram notificados 333.460 mil casos no Brasil, sendo a maioria na Região
Amazônica. Apesar do número de óbitos e internações ter sido reduzido, a questão da
transmissibilidade ainda preocupa. Apesar dos esforços do programa nacional e dos
programas estaduais, em quase uma década, não se obteve uma redução efetiva desse
indicador (Braz et al., 2014). A incidência da malária nas últimas quatro décadas sofreu
significativa variação (Figura 1). Até a década de 1990, o aumento foi devido à construção de
grandes rodovias na Região Norte, expansão de áreas de garimpos e projetos de colonização
(Hermes et al., 2013).
Após esse período, houve um período de relativa tendência à estabilização, com
bruscas quedas e elevações no número de casos. O último pico significativo se deu no ano de
2005, onde o aumento dos criadouros do mosquito vetor da malária foi o principal
responsável, em função da atividade de piscicultura desordenada com a construção de tanques
artificiais nas periferias de diversas cidades da Região (Brasil, 2013). No ano de 2014, os
números voltaram a se comparar com os registrados no final da década de 1970 (Brasil,
2015).
Na Região Amazônica, a incidência se alia aos fatores já largamente conhecidos como
sendo causa da disseminação da doença, tais como: ocupação do solo de forma inadequada,
fatores ambientais e biológicos e transformações oriundas do comportamento humano (Braz
et al., 2013). Segundo Braz et al (2014), para que haja um controle realmente eficiente, é
necessário que se conheçam previamente as variações sazonais, cíclicas e históricas da
doença, por localidade, descrevendo onde e quando ocorre, identificando precocemente as
mudanças não esperadas, ou seja, as epidemias e, de posse das informações, montar
estratégias viáveis a cada localidade específica.

15
Figura 1 - Série histórica do número de casos na região Amazônica no período de 1970 a
2014. (Fonte: Brasil, 2015).
Nos estudos de Braz et al (2013), foi observado que em 2010 foram registrados mais
de 300.000 mil casos de malária por ano no Brasil, sendo 99,6% na Região Amazônica.
Durante esse período, foram introduzidos diversos programas com o objetivo de controlar a
endemia e, mesmo com todos os esforços nas três esferas do governo, os casos continuam em
patamares elevados. Para Braz et al (2012), este alto índice de parasitas indicou a necessidade
de melhorar a vigilância da doença para apoiar medidas de controle e prevenir uma situação
epidêmica, com mais casos relatados do que o esperado. Além disso, uma das estratégias da
Organização Mundial da Saúde (OMS) para controle de doenças é fortalecer a capacidade
local para permitir e promover a avaliação regular da malária, visando intervenções adequadas
e oportunas.
Também é necessário avaliar rotineiramente se essas ações estão contribuindo para
reduzir a incidência da doença. Segundo o ministério da saúde, desde 2000 houve uma
redução de mais de 50% no número de casos de malária no país. Em novembro de 2015, o
governo lançou o Plano de Eliminação da Malária no Brasil para ajudar o país a cumprir a
meta dos Objetivos de Desenvolvimento Sustentável, da Organização das Nações Unidas, de
reduzir em 90% o número de casos da doença até 2030. Em 2014, o Brasil registrou 143.250
casos de malária, o menor número desde 1989 (Penido, 2016). Apesar dos poucos casos
autóctones na região Extra-Amazônica, a doença não pode ser negligenciada diante do risco
de reintrodução, agravado pelo fluxo migratório em áreas suscetíveis, bem como pela

16
possibilidade de aumento da letalidade devido ao diagnóstico tardio e manejo clínico
inadequado.
Embora o principal vetor (Anopheles darlingi) esteja presente em cerca de 80% do
país, atualmente, a incidência de malária no Brasil é quase exclusivamente (99,5% dos casos)
restrita à Região da Amazônia Legal. Nesta área, uma combinação de condições
socioeconômicas e ambientais favorece a proliferação do mosquito e, consequentemente, a
exposição de grandes contingentes populacionais de residentes ou visitantes ao risco
(Oliveira- Ferreira et al., 2010).
Nesse contexto, o risco da ocorrência anual de casos em determinado espaço
geográfico é estimado pela Incidência Parasitária Anual (IPA), que é expressa pelo número de
exames positivos de malária a cada mil habitantes (Figura 2). A IPA pode ser classificada
como de baixo risco (IPA<10); médio risco (IPA: 10-49) e alto risco (IPA>50). O Estado do
Amapá apresentou IPA variável entre os municípios distribuídos em baixo, médio e alto risco
de transmissão.
Figura 2 - Áreas de transmissão de malária no Brasil de acordo com Incidência Parasitária
Anual (IPA) no ano de 2015. (Fonte: SIVEP-MALÁRIA/SVS 2015. Acesso em 18/07/2017).
Esta heterogeneidade torna-se ainda mais evidente nos inquéritos realizados em
unidades espaciais como vilarejos e pequenas comunidades rurais. Pode ser explicados por
diversos fatores como a presença de criadouros do anofelino e a distância entre eles e os
domicílios humanos, assim como as características dos domicílios, a densidade demográfica e

17
o uso de medidas de proteção contra o mosquito e contra o parasito (Silva et al., 2008; Silva-
Nunes et al., 2010).
Na Amazônia brasileira, bem como em diversas áreas endêmicas do mundo, a
distribuição da malária apresenta grande heterogeneidade. Adicionalmente a esse contexto, no
início da década de 1990, a Amazônia Legal compreendida pelos Estados do Amapá (AP),
Acre (AC), Amazonas (AM), Maranhão (MA), Mato Grosso (MT), Pará (PA), Rondônia
(RO), Roraima (RO) e Tocantins (TO) registra elevada incidência de malária (Oliveira- Filho
& Martinelli, 2009). De acordo com o Instituto Brasileiro de Geografia e Estatística (IBGE), a
Amazônia Legal possui cerca de 26.9 milhões de habitantes (13,4% da população brasileira) e
59,75% do território nacional (IBGE, 2014). O Estado do Amapá apresentou como numero de
casos notificados de malária de 12.830 e IPA de 17,1 no ano de 2014 (Figura 3).
Figura 3 – Casos de malária e Incidência Parasitária Anual (IPA) no ano de 2014. (Fonte:
SIVEP-MALÁRIA/SVS/MS 2017. Acesso em 18/07/2017).
Na Amazônia Brasileira, a incidência da malária está associada às interações entre a
forma de uso da terra, o comportamento humano, os elementos físicos e biológicos do meio
natural e as transformações causadas pela intervenção humana (Confalonieri, 2005), como
exemplo, colonização agrícola, pecuária, extrativismo vegetal e mineral, além da construção
de estradas e de usinas hidrelétricas (Barbieri, 2005). No contexto brasileiro, deve ser
fortemente destacada os fatores socioeconômicos que modulam o autocuidado, a exposição
aos fatores de risco, a identificação da doença como problema de saúde pública, as condições
para a adoção de meios de prevenção e a adesão ao tratamento (Braz et al., 2013).
Adicionalmente, os determinantes políticos interagem nessa complexa rede de causalidade,

18
com especial destaque para os diferenciais de gestão em saúde pública nos três níveis de
governo ao longo do tempo.
No Estado do Amapá, a malária tem acometido grande parte da população ao longo
dos anos. Dentre os principais motivos está a sua condição de área endêmica, devido às
características ambientais propícias ao hábitat e proliferação do vetor, bem como a circulação
dos protozoários do gênero Plasmodium. Fazendo a análise de uma série histórica do número
de casos de malária notificados no Estado, é possível observar uma redução ao longo dos anos
(Figura 4).
Figura 4 - Número de casos de malária notificados no Estado do Amapá e diferença
percentual entre os anos de 2003 a 2017 (Fonte: SIVEP-malária/DATASUS/MS, 2017). Os
dados de 2017 são dados preliminares até Junho, sujeito a alteração.
Na malária, a dinâmica do número de casos se dá em razão do comportamento
humano, como ocupação do solo através dos assentamentos e construção de roças, de estradas
de rodagem ou de hidroelétricas, além de frentes de trabalho na mineração, que fazem com
que aumentem esses valores (Gomes et al., 2015). Da mesma forma, há mobilidade desses
números com relação aos serviços oferecidos pelos programas de controle, ou seja, se há
investimento com equipe de saúde, educação e intervenção com diagnóstico e tratamento em
tempo hábil, esses valores são reduzidos (Cardoso, 2014; Gomes et al., 2016). A redução ou
aumento no número de casos de malária está diretamente relacionado à rede de notificação e
diagnóstico deste agravo nos municípios, bem como à realização de ações de controle
vetorial.
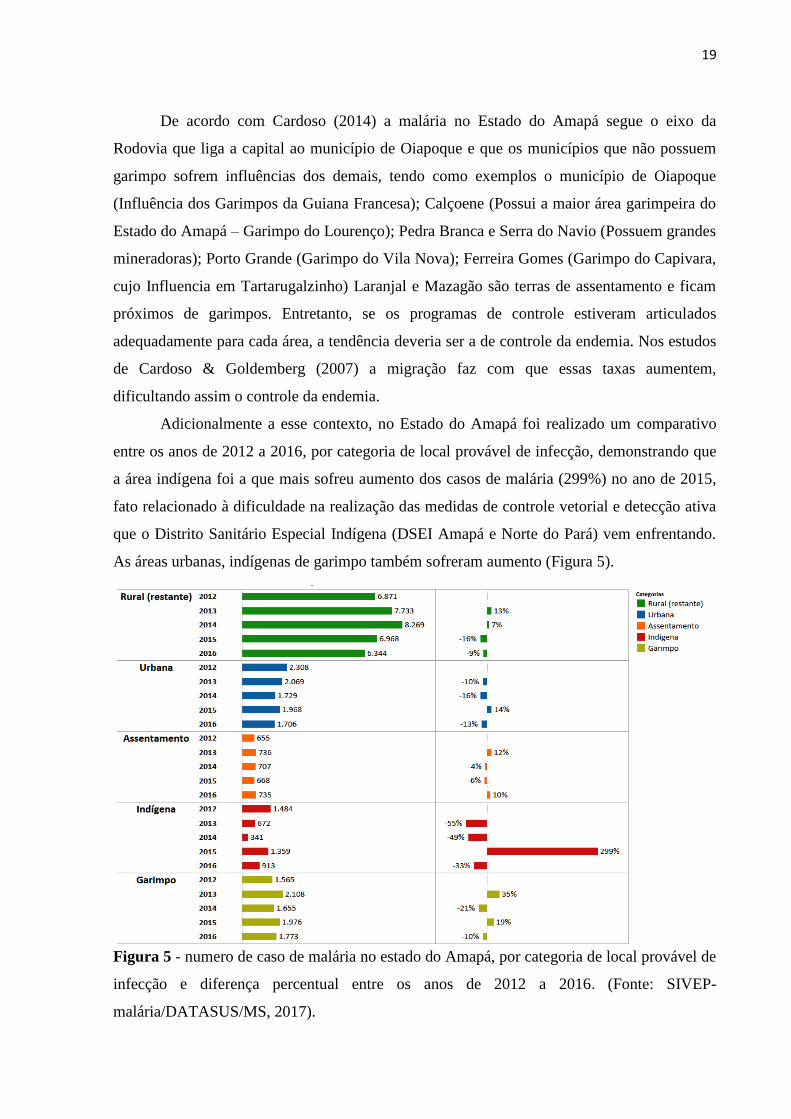
19
De acordo com Cardoso (2014) a malária no Estado do Amapá segue o eixo da
Rodovia que liga a capital ao município de Oiapoque e que os municípios que não possuem
garimpo sofrem influências dos demais, tendo como exemplos o município de Oiapoque
(Influência dos Garimpos da Guiana Francesa); Calçoene (Possui a maior área garimpeira do
Estado do Amapá – Garimpo do Lourenço); Pedra Branca e Serra do Navio (Possuem grandes
mineradoras); Porto Grande (Garimpo do Vila Nova); Ferreira Gomes (Garimpo do Capivara,
cujo Influencia em Tartarugalzinho) Laranjal e Mazagão são terras de assentamento e ficam
próximos de garimpos. Entretanto, se os programas de controle estiveram articulados
adequadamente para cada área, a tendência deveria ser a de controle da endemia. Nos estudos
de Cardoso & Goldemberg (2007) a migração faz com que essas taxas aumentem,
dificultando assim o controle da endemia.
Adicionalmente a esse contexto, no Estado do Amapá foi realizado um comparativo
entre os anos de 2012 a 2016, por categoria de local provável de infecção, demonstrando que
a área indígena foi a que mais sofreu aumento dos casos de malária (299%) no ano de 2015,
fato relacionado à dificuldade na realização das medidas de controle vetorial e detecção ativa
que o Distrito Sanitário Especial Indígena (DSEI Amapá e Norte do Pará) vem enfrentando.
As áreas urbanas, indígenas de garimpo também sofreram aumento (Figura 5).
Figura 5 - numero de caso de malária no estado do Amapá, por categoria de local provável de
infecção e diferença percentual entre os anos de 2012 a 2016. (Fonte: SIVEP-
malária/DATASUS/MS, 2017).

20
Com relação à malária em áreas urbanas, especificamente na capital, Macapá, a
malária é periurbana, onde se encontram áreas de ressaca, fragmentos de florestas e
assentamentos desordenados (invasões). Num estudo realizado nos anos de 2007 e 2008, na
comunidade de Lagoa dos Índios, a abundância dos vetores se associou com o padrão sazonal
das chuvas. Deste modo, considerou-se que a transmissão de malária se mantém durante todo
o ano na cidade de Macapá (Galardo et al., 2013). Embora a malária se mantenha como
padrão de permanência entre os municípios do estado do Amapá é necessário um
monitoramento continuo dessa enfermidade, visando atingir cada vez mais o que é
preconizado pelo Ministério da Saúde: Diagnóstico precoce e Tratamento imediato e
adequado na quebra da cadeia de transmissão da doença.
Com relação às áreas indígenas, pouco se sabe sobre a ecologia dos vetores da malária
e suas relações com as populações indígenas. Os estudos demostram o vetor Anopheles
darlingi exibir comportamento predominantemente exofílico (Coimbra Jr et al., 2005). A
presença diária e nos mais variados horários de indígenas às margens de cursos d’água no
entorno das aldeias, inclusive nos momentos de pico da atividade anofélica, expõe os
indivíduos de ambos os sexos e de todas as idades ao risco de infecção, possivelmente
elevando esses percentuais a cada ano.
Para as áreas de garimpo, o desenvolvimento do ciclo malarígeno depende não apenas
das características naturais do meio ambiente, como também da presença do homem no
manuseio e forma de uso da terra influenciando os níveis de ocorrência de malária (Silva-
Nunes et al., 2008). Para compreender como a heterogeneidade da ocupação se relaciona com
a incidência da malária, é preciso abordar não apenas a dimensão biológica da doença, mas
também como esse processo ocorre no espaço regional, influenciando as características
populacionais e ambientais (Barbieri & Sawyer, 2007). Dentre as características importantes,
para áreas garimpeiras, na transmissão da malária é a quantidade de locais propícios à
reprodução dos vetores, como: cavas abandonadas que acumulam águas de chuva, grande
aglomerado de pessoas em um espaço pequeno e uma grande migração interna facilita a
entrada e saída de pessoas infectadas ou não, e a constante renovação de população de
suscetíveis.
1.2.1 Epidemiologia da Malária no Município de Oiapoque
O município de Oiapoque, pertencente ao Estado do Amapá, região Amazônica,
extremo norte do Brasil, fronteira ocidental da região amazônica. Faz fronteira ao norte com a
Guiana Francesa, ao sul com os municípios de Calçoene, Serra do Navio e Pedra Branca do

21
Amapari, ao leste é banhado pelo Oceano Atlântico. A área geográfica de Oiapoque é de
22.625 km2, possui população de 20.426 habitantes segundo IBGE (2010), além de uma
população flutuante (fluxo migratório de curto prazo) que não entra nas estatísticas oficiais
em virtude da intensa mobilidade de pessoas entre as cidades gêmeas de Oiapoque no Amapá
e Saint-Georges na Guiana Francesa com objetivo de trabalhar em cidades da Guiana
Francesa ou nos garimpos clandestinos, fator de aumento da vulnerabilidade à malária no
município de Oiapoque.
O clima é equatorial, com uma umidade relativa média de 82%. A precipitação anual
varia entre 2.700 e 3.300 mm. A temperatura média é de 27ºC, variando entre 26 e 33°C.
Temperaturas mais altas coincidem com os meses mais secos do ano de setembro a novembro.
O município também possui características que contribuem para as ocorrências dos casos de
malária, condições ambientais favoráveis ao Anopheles vetor da malária inclusive com áreas
indígenas isoladas e com presença de transmissão da doença (Galardo et al., 2013; Gomes et
al., 2016).
As áreas fronteiriças entre os países são muitas vezes caracterizadas por intensos
fluxos populacionais transfronteiriços (Peite et al., 2008). A cidade de Oiapoque encontra-se
na fronteira entre a Guiana Francesa e o Estado brasileiro do Amapá, onde há um fluxo
populacional intenso e continuo com o município francês de Saint Georges. Na Guiana
Francesa, a malária é endêmica e distribuída ao longo dos rios Maroni e Oiapoque, enquanto a
área costeira que faz fronteira com o Oceano Atlântico quase não possui transmissão da
malária (Nacher et al., 2013).
O contínuo fluxo migratório na fronteira Brasil - Guiana Francesa tem como ponto de
convergência o município de Oiapoque, que apresenta um índice de parasitemia anual
considerado como área de médio risco para transmissão de malária (SIVEP-
malária/DATASUS/MS, 2017). De fato, o Ministério da Saúde observou uma redução no
número de casos de malária ao longo dos anos na maioria dos municípios amazônicos em que
ocorre a transmissão, mas o mesmo não ocorreu nas áreas que se localizam na faixa de
fronteira (Gomes et al., 2016).
Esta falha no controle é uma preocupação para as autoridades de saúde e vários fatores
têm sido atribuídos, sendo necessária pesquisas mais aprofundadas para entender os
determinantes dessa malária a fim de estabelecer as melhores medidas de controle possíveis.
Segundo Andrade (2005), ao estudar a expansão da malária e sua vinculação com o processo
migratório no período de 1990 - 2003 observou que a situação da malária importada pela
fronteira, no período analisado, influenciou na manutenção e propagação da doença, visto

22
que, não somente manteve o IPA de alto risco de transmissão, como também contribuiu com
o aumento nos casos de infecção. De 2003 a 2014 foram notificados 53.958 casos de malária,
com IPA durante o período de 250,2/1000 habitantes (Figura 6).
Figura 6 - Série histórica de casos totais e Incidência Parasitária Anual durante o período de
2003 a 2014 no município de Oiapoque-Amapá (SIVEP-malária/DATASUS/MS, 2017).
Acompanhando a redução que ocorreu no país, o IPA em 2014 chegou a 39,5/1000
habitantes. Entre os casos notificados 27.301 (50,6%) foram autóctones e 26.657 (50,1%),
foram importados. Em relação aos casos autóctones, 8.470 (31%) foram na área urbana e
21.212 (69%) na área rural. Plasmodium vivax (61,4%) foi a espécie parasitaria predominante
seguido do P. falciparum (35,4%). A IPA média nas áreas indígenas foi de 254,6/1000
habitantes (SIVEP-malária, 2017). A Guiana Francesa contribuiu com 98,1% de casos
importados, sendo vários garimpos clandestinos relatados como local provável de infecção,
com o maior percentual oriundo do garimpo do Sikini com 21,9% dos casos (Figura 7).
Figura 7 - Casos de malária por P. vivax segundo local provável de infecção e importados dos
garimpos clandestinos da Guiana Francesa para Oiapoque (Fonte: Adaptada de Gomes et al.,
2015).

23
Em se tratando do fluxo migratório, essa fronteira internacional vem implicando
trânsito intenso, principalmente feito por brasileiros em direção aos garimpos clandestinos ou
aos trabalhos em cidades da Guiana Francesa, onde, muitos dos residentes em Oiapoque
possuem familiares envolvidos na extração de ouro ou outro tipo de atividade nos garimpos.
Entretanto, como essa população não é registrada por ocasião dos recenseamentos feitos no
município, não aparece nos censos populacionais (Cardoso, 2014; Gomes et al., 2015; Gomes
et al., 2016).
Essa relação entre homem e meio ambiente perdura ao longo dos anos e estabelece
uma articulação necessária de subsistência que envolve diversos fatores sociais e econômicos
que registram a presença de garimpeiros clandestinos nesta região e que vivem em condições
de vida e de trabalho de extrema precariedade. Esse grupo se desloca constantemente nos dois
sentidos e, especialmente, vem à procura de tratamento para repetidas infecções por malária.
Esse é o fenômeno conhecido como “malária importada”. No entanto, um hábito comum entre
os garimpeiros de ingerir medicamentos sem o devido acompanhamento e orientação médica,
acaba se refletindo nas resistências aos antimaláricos indicados pelo Ministério da Saúde do
Brasil (Gomes et al., 2015; Gomes et al., 2016).
As áreas de fronteira internacional são regiões que merecem atenção especial em
relação à saúde. Os limites internacionais criam empecilhos técnicos, políticos e operacionais
no controle das endemias, com explicita dificuldades em fornecer atenção integral à saúde aos
cidadãos fronteiriços que vivem e circulam entre a cidades gêmeas (Silva-Nunes et al., 2008;
Silva et al., 2010; Gomes et al., 2016). A vigilância e o controle da malária na fronteira
compreendem um grande desafio, pois a mobilidade transfronteiriça faz parte do cotidiano da
população e dificulta o controle dessa enfermidade.
O processo saúde-doença, geralmente, transcende as fronteiras e pode afetar ambos os
lados, podendo propagar-se, até mesmo, para regiões remotas dos limites dos países, no
entanto, a informação e direitos não atravessam o limite internacional, mas os agentes e
vetores produtores de doenças circulam livremente por essas áreas (Levino & Carvalho,
2011). Nesse contexto, as precárias condições de vida de grande parte das populações
fronteiriças, a ocupação desordenada do espaço, as intervenções politicas mal planejadas e os
diversos fluxos migratórios contribuem para o aumento da receptividade aos vetores e da
vulnerabilidade da população nessas áreas.
No Município de Oiapoque, a malária tem acometido grande parte da população ao
longo dos anos. Fazendo a análise de uma série histórica do número de casos de malária
notificados no município, é possível observar uma redução ao longo dos anos (Figura 8). No

24
ano de 2003, Oiapoque apresentou um cenário de 3.877 casos, nos anos posteriores, houve
aumento e diminuição, fechando o ano de 2016 com 897 notificações positivas. Essa redução
no número de casos de malária ao longo dos anos está diretamente relacionado à rede de
notificação e diagnóstico deste agravo no município, bem como à realização de ações de
controle vetorial.
Figura 8 - Serie histórica do número de casos de malária notificados no município de
Oiapoque e diferença percentual entre os anos de 2003 a 2016. (Fonte: SIVEP-
malária/DATASUS/MS, 2017).
Adicionalmente, no Município de Oiapoque foi realizado um comparativo entre os
anos de 2003 a 2016, por categoria de local provável de infecção, demonstrando que a área
indígena foi a que mais sofreu aumento dos casos de malária (59%) no ano de 2016 (Figura
9), fato relacionado à dificuldade na realização das medidas de controle vetorial e detecção
ativa dos casos, por serem locais de difícil acesso. Estatísticas vitais e essenciais para
monitorar o perfil de saúde/doença e planejar ações de saúde e educação, não são disponíveis
para a ampla maioria dos grupos. Neste sentido, Coimbra jr. & Santos (2000) chamam
atenção para as implicações do que denominam uma danosa invisibilidade demográfica e
epidemiológica.

25
Figura 9 - Serie histórica do número de casos de malária por categoria de local provável de
infecção notificados no município de Oiapoque e diferença percentual entre os anos de 2003 a
2016 (Fonte: SIVEP-malária/DATASUS/MS, 2017).
O perfil epidemiológico dos povos indígenas no Brasil é muito pouco conhecido, o
que decorre da exiguidade de investigações, da ausência de censos e de outros inquéritos
regulares e da precariedade dos sistemas de registro de informações sobre morbidade,
mortalidade, cobertura vacinal, dentre outros fatores (Coimbra Jr. & Santos, 2000). A
relevância da malária no perfil epidemiológico da população indígena é inquestionável
(Ianelli, 2000). Grupos vivendo em certas áreas da Amazônia em especial aquelas sob a
influência de fluxos migratórios, atividades de mineração e garimpos ou de implantação de
projetos de desenvolvimento, são particularmente vulneráveis (Gomes et al., 2016).
Adicionalmente a esse contexto, nas áreas urbanas e rurais tem sido observada uma elevada
taxa de morbidade da malária geralmente proveniente de outras localidades (Figura 9).
A maioria dos casos de malária importados do município de Oiapoque é proveniente
da Guiana Francesa. Por conta da fronteira, muitos brasileiros se deslocam para lá para
desempenhar atividades garimpeiras e voltam contaminados com malária, além dos que lá
residem e mantém intenso fluxo de entrada e saída no referido município, contribuindo para a
transmissão de novos casos (Gomes et al., 2016). Áreas fronteiriças e de garimpo representam

26
um desafio para o controle da malária, por apresentarem intenso fluxo migratório, e um
ambiente favorável para a manutenção e disseminação da doença na região.
1.3 PARASITOS GASTROINTESTINAIS NO BRASIL
Os parasitos gastrointestinais são cosmopolitas e comuns em muitas partes do mundo,
sendo responsáveis pela morbidade e mortalidade de grande parte da população,
especialmente em países de baixo status socioeconômico. É considerada um sério problema
de saúde pública nos países subdesenvolvidos e em desenvolvimento (Mahmud et al., 2013).
São mais prevalentes em ambientes quentes e úmidos, sobretudo de populações que ocupam
ambientes em condições sanitárias inapropriadas e que, ocasionalmente, concentram-se em
grupos de grande mobilidade e pluralidade com complexo e dinâmico quadro de saúde
relacionado a processos sociais, econômicos e ambientais (Oliveira et al., 2015).
Embora a prevalência de infecções gastrointestinais seja maior nos países em
desenvolvimento, essas doenças são frequentes nos países industrializados, provavelmente em
associação com a globalização através abastecimento alimentar e com a imigração e/ou
adoção de regiões endêmicas para viagens (Calderaro et al., 2014). Estima-se que infecções
intestinais causadas por helmintos e protozoários afetem mais de 2 bilhões de pessoas no
mundo, e estão entre os muitos problemas de saúde nos países em desenvolvimento,
principalmente, em locais com crescimento rápido e desordenado da população, sem as
adequadas estruturas de abastecimento de água potável e saneamento básico (Lodo et al.,
2010; Siqueira et al., 2011; Vieira & Benetton, 2013).
Embora populações inteiras estejam em risco geograficamente, as crianças são
observadas com a maior carga de infecção. Esta desproporção tem bases comportamentais,
biológicas e ambientais, na qual as crianças tendem a ser mais ativas no ambiente infectado e
raramente empregam bons comportamentos sanitários (Harhay et al., 2010). Adicionalmente a
prevalência das doenças parasitárias pode variar de acordo com a região, havendo influência
na infraestrutura física, no clima e cultura da localidade, estando esses fatores associados à
quantidade e ao tipo das parasitoses que infectam os indivíduos (Semedo et al., 2014). O
Brasil possui índices variáveis de parasitoses intestinais em diferentes municípios e regiões,
resultante das condições climáticas, sociais, econômicas e sanitárias das localidades, assim
como, condições do solo, água e alimentos dessas regiões (Hailegebriel, 2017).
Os parasitos encontrados no trato gastrointestinal humano podem ser categorizados em
dois grupos, helmintos e protozoários. Os helmintos transmitidos pelo solo (Ascaris

27
lumbricoides, Ancilostomíase e Trichuris trichiura) são os mais prevalentes, no Brasil entre
os protozoários, Giardia intestinalis e o complexo Entamoeba histolytica/díspar protozoários
patogênicos, sendo os comensais, Endolimax nana e Entamoeba coli não menos importantes
nos estudos de prevalência (Kubiak et al., 2017). Esses comensais indicam a ingestão de água
ou alimentos contaminados com resíduos fecais e que, portanto, estão sob risco de
contaminação de amebas patogênicas reforçando a importância do diagnóstico e descrição
destes comensais, a fim de se programar medidas preventivas para evitar infecção pela
Giardia intestinalis e E. histolytica (Belloto et al., 2011).
A frequência de giardíase sofre variações quanto à distribuição mundial, ficando a taxa
de incidência entre 11% e 30% (Mbuh et al., 2010). Globalmente, estima-se que existem 280
milhões de casos de infecção por G. intestinalis por ano em todo o mundo, mas
principalmente na Ásia, África e América Latina, a giardíase sintomática envolve cerca de
200 milhões de pessoas e, a cada ano, cerca de 500 mil novos casos sintomáticos são
detectados. Na União Europeia em 2012, foram documentados 16 368 (5,43 por 100 000
habitantes) de casos confirmados de giardíase (Kubiak et al., 2017). Nos Estados Unidos da
América, G. lamblia é considerado o parasito intestinal mais comumente encontrado no
homem representando a principal causa de diarreia causada por protozoário intestinal. Sabe-se
que a frequência de infecção por giardíase e do complexo Entamoeba histolytica/díspar varia
de acordo com a população e a região estudada e que é maior em países em desenvolvimento
(Huang & White, 2006).
Considerando os esforços para uma melhor compreensão da epidemiologia de
parasitos intestinais, relativamente poucos estudos equivalentes são feitos em protozoários
intestinais. Isso é surpreendente, porque a amebíase intestinal causada pelo protozoário
Entamoeba histolytica é a terceira maior doença parasitária responsável pela morte no mundo
após a malária e esquistossomose (Ouattara et al., 2010). No Brasil, a ampla diversidade das
características socioeconômicas, climáticas e geográficas tem sido apontada como fator crítico
para o perfil dos agentes etiológicos na diarreia, modelando assim a frequência destes
diferentes enteropatógenos (Belloto et al., 2011). Adicionalmente para Sánchez-Arcila et al
(2014), a falta de conhecimento e cuidados com a saúde potencializa a manutenção desses
patógenos, acarretando numa grande frequência de enteroparasitas patogênicos.
Ademais, o Brasil, principalmente a região Amazônica, as geohelmintíases apresentam
uma prevalência elevada e os helmintos intestinais mais comuns que infectam os seres
humanos são o A. lumbricoides, T. trichiura e Ancilostomídeos (Araújo & Fernandez, 2005).
Estes são amplamente distribuídos em países tropicais, infectando mais de um bilhão de

28
pessoas (Figura 10). Essa distribuição helmíntica no hospedeiro humano pode abrigar poucos
ou muitos parasitos, e tem consequências clínicas para os hospedeiros, pois é principalmente a
intensidade da infecção que determina a gravidade da morbidade (Wammes et al., 2014). A
maioria dos nemátodos intestinais como A. lumbricoides e T. trichuris são adquiridos através
da ingestão de ovos infectantes do solo contaminado com fezes humanas infectadas, enquanto
as infecções por Ancilostomídeos ocorrem quando as larvas infectantes do solo penetram na
pele do individuo (Morenikeji et al., 2009).
As geohelmintíases humanas, geralmente negligenciadas, têm grande importância no
contexto universal, o A. lumbricoides é o nematódeo mais frequente a infectar o trato
intestinal humano, a maioria das infecções envolve pequeno número de parasitos adultos e é
assintomática. A manifestação dos sintomas da ascaridíase depende do número de parasitos
adultos albergados pelo indivíduo. Infecções maciças podem resultar em bloqueio mecânico
do intestino delgado, principalmente em crianças (Melo et al., 2004). Os T. trichiura
colonizam o ceco (intestino grosso), sua penetração nos tecidos provoca inflamações e
pequenas hemorragias de modo que, quando presente em um grande número, podem causar
disenteria e até mesmo um prolapso retal (Hall et al., 2008). A ancilostomose raramente é
sintomática, até que seja estabelecida uma carga significativa de parasitas intestinais. A maior
preocupação da infecção é a espoliação de sangue, os ancilostomídeos consomem cerca de
0,25 mL de sangue por dia, podendo causar anemia e que pode estar associada a um retardo
físico e mental dos indivíduos afetados (Kucik et al., 2004).
Figura 10 - Frequência da distribuição de geohelmintos no Brasil de 1995 a 2013. a) A.
lumbricoides; b) T. trichiura e c) Ancilostomídeos (Adaptada de Charmmartin et al., 2014).
A infecção por helmintos influencia o estado nutricional, mas seu impacto sobre a
anemia não é clara. Na região amazônica, como em muitas outras partes do mundo, as áreas
endêmicas de malária coincidem com locais de alta prevalência de helmintíase intestinal

29
(Nacher, 2012). Essas características sugerem uma longa coadaptação evolutiva entre esses
parasitos e o homem. A chave para esta parceria é a interação imunológica entre helmintos e
seus hospedeiros mamíferos, polarizando as respostas imunes e modulando os processos
regulatórios, o que pode explicar a sua sobrevivência dentro de um hospedeiro a longo prazo
(Allen & Maizels, 2011). Além de representar uma tendência temporal de permanecia com
variabilidade e complexidade epidemiológica de cada parasito. Esta diferença destaca a
importância de estudos na busca de estimativa do risco de infecção por helmintos e outras
doenças tropicais negligenciadas.
Nesse contexto, a principal consequência da associação entre malária e das
geohelmintoses é a anemia. Por meio dos mecanismos de hemólise a malária se torna um
importante contribuinte para a ocorrência de anemia, enquanto que infecções com moderada ou
elevada intensidade causada pelo Ascaris lumbricoides, Trichuris trichiura e Ancilostomídeos,
são perniciosas, produzindo frequentemente anemia, e tendo como consequência à espoliação
sanguínea que quando não tratada e associada a desnutrição, se torna muitas vezes fatal
(Stephenson et al., 2000; Nacher, 2011; David et al., 2013). Estudos epidemiológicos que
avaliam concomitantemente as parasitoses intestinais, malária e a anemia de uma população
são importantes para o conhecimento da realidade local, contribuindo para que novas
estratégias de prevenção e controle sejam aplicadas com maior eficiência.
1.4 COINFECÇÃO MALÁRIA E PARASITOS GASTROINTESTINAIS
O múltiplo parasitismo é comum e as taxas de coinfecção dependem da distribuição
espacial das condições ambientais que permitem a transmissão dos parasitos, além das
interações imunológicas e de outros fatores, pelos parasitos, que possam alterar a
susceptibilidade ou o comportamento do hospedeiro (Knowles, 2011). Nesse contexto, o
maior obstáculo ao controle efetivo de parasitas intestinais em populações em risco é o
conhecimento inadequado da distribuição geográfica da infecção e as variáveis demográficas
que influenciam a sua prevalência (Brooker et al., 2006a). Apesar da elevada frequência e da
importância global para a saúde essas doenças são negligenciadas e poucos estudos retratam
sua prevalência a nível global, contribuindo para a compreensão geral da epidemiologia
desses patógenos na coinfecções.
Dessa forma, protozoários do gênero Plasmodium agente etiológico da malária, e
muitas espécies de parasitos intestinais (protozoários e helmintos) compartilham da mesma
área de distribuição geográfica, podendo assim ambos os tipos de parasitos coinfectar uma

30
mesma população de hospedeiros, principalmente nas áreas que apresentam características
geográficas, climáticas e socioeconômicas peculiares que favoreçam a sua existência e
manutenção (Brasil, 2005). Essa coinfecção de um mesmo hospedeiro pode influenciar de
maneira positiva (facilitação) ou negativa (competição) através de diferentes mecanismos
como competição por recursos e interações mediadas por via imune, potencializando ou
inibindo a malária no hospedeiro (Knowles, 2011). Desta forma, tem-se centrado sobre as
interações da coinfecção entre a malária e as helmintíases intestinais, e diversos estudos com
dados preliminares sugerem um decréscimo na gravidade da malária, entre os coinfectados
com helmintos intestinais (Degarege et al., 2012). Esse contexto fortalece as pesquisas de
coinfecção malária-parasitoses intestinais focadas na coinfecção malária-helmintos, não se
tendo amplo conhecimento a respeito das coinfecções de malária-protozoários intestinais.
Quando se trata dessas coinfecções o Ascaris lumbricoides é o helminto mais
investigado e resultados contraditórios tem sido detectado. Estudos revelaram que a
coinfecção com o A. lumbricoides foi associada com uma diminuição da malária (incidência,
prevalência ou redução da parasitemia) (Brutus et al., 2006; Boel et al., 2010). Para a malária
cerebral ou insuficiência renal, dois estudos identificaram o A. lumbricoides como o único
helminto associado à proteção contra a malária grave em adultos (Bejon et al., 2008).
Ademais, um trabalho observou um aumento da malária grave em crianças infectadas por este
nematelminto no continente africano (Le Hesran et al., 2004).
As infecções por Ancilostomídeos parecem também estar associado a casos de malária
por P. falciparum na África, essa alta prevalência de ambas as infecções entre os indivíduos é
extremamente comum nesta localidade (Brooker et al., 2006b). Os ancilóstomos são o
segundo nematoide gastrointestinal relatado em interações com malária (Nacher et al., 2000).
De fato, diversos estudos demonstram aumento da frequência de malária na coinfecção com
estes enteroparasitos (Boel et al., 2010; Pullan et al., 2011; Degarege et al., 2012), e aumento
da gravidade da anemia devido aos distintos mecanismos através dos quais cada parasita
causa anemia (Brooker et al., 2006b), enquanto que, apenas um estudo não encontrou
nenhuma associação entre a presença de Ancilostomídeos e a infecção malárica (Shapiro et
al., 2005).
A maioria dos estudos envolve o Plasmodium falciparum, no entanto, o efeito protetor
da malária pelo Plasmodium vivax foi também observado em três estudos - dois fora do Brasil
(Murray et al., 1978; Boel et al., 2010) e um no Brasil, onde Melo et al (2010) evidenciaram
que as helmintíases intestinais estão associadas com a proteção para a redução dos níveis de
hemoglobina, durante os episódios de malária por P. vivax, em uma população infantil da

31
cidade de Manaus no Estado do Amazonas. Adicionalmente, há evidências crescentes de que
a coinfecção com múltiplos parasitas pode prejudicar a resposta imune do hospedeiro a
parasitas isolados e pode aumentar a suscetibilidade à doença clínica de maneira que
atualmente não são claramente compreendidas (Druilhe et al., 2005; Mwangi et al., 2006).
Outro ponto interessante sobre as coinfecções entre o plasmódio e helmintos é o
aumento de gametócitos circulantes, aumento de duração da infecção e o aumento da
atratividade do hospedeiro para o vetor (Le Hesran et al., 2004; Nacher, 2012). Tem sido
descrito na literatura que o processo de gametogênese pode ser influenciado pelo estresse
parasitário e múltiplos fatores ambientais (Ono et al., 1993; Dyer & Day, 2000), dentre eles as
baixas concentrações de hemoglobina estão associadas com a circulação de gametócitos na
corrente sanguínea (Price et al., 1999). De fato, em 2001, estudo de coorte realizado na
Tailândia mostra um aumento de duas vezes a quantidade de gametócitos do Plasmodium
falciparum circulantes vinculados ao decréscimo dos níveis de hemoglobina em pacientes
infectados por helmintos (Nacher et al., 2001).
Embora a coinfecção entre o plasmódio e os helmintos seja prevalente em países
tropicais, o efeito de suas interações permanece obscuro (Roussilhom et al., 2010). Alguns
estudos relataram que indivíduos infectados por helmintos são suscetíveis à infecção pelo
plasmódio (Midzi et al., 2011; Ojurongbe et al., 2011), ocasionando o aumento de
gametócitos circulantes (Nacher et al., 2011), e redução dos níveis de hemoglobina (Boel et
al., 2010), bem como suprimindo manifestações clínicas agudas (Murray et al., 1978) e ainda,
aumentando o risco de malária (Spiegel et al., 2003; Nacher, 2011; Nacher 2012). Sabe-se
ainda que o aumento da incidência e da prevalência da malária pode interferir no
desenvolvimento de infecções mistas pelo P.vivax e o P. falciparum e que, a diversidade de
parasitos pode ser maior em pacientes infectados por helmintos (Nacher et al., 2001;
Chaorattanakawee et al., 2003).
Segundo Sánchez-Arcila et al (2014), as diferenças nos resultados obtidos nos
diferentes estudos, podem depender da espécie de parasitos intestinais e da idade de uma
população. Enquanto que a maioria dos estudos de coinfecção retratam a prevalência de
helmintos em crianças, na população do Estado de Rondônia foram encontrados uma
prevalência de protozoários e os participantes do estudo eram adultos (Sánchez-Arcila et al.,
2015). Além disso, o aumento da diversidade genética dos oocistos nos mosquitos
alimentados com sangue de pacientes coinfectados, teoricamente, poderia levar a uma maior
diversidade genética dos parasitos da malária que circulam na população.

32
As implicações evolutivas da coinfecção também podem estender-se à reprodução de
helmintos, que tem interesse de proteger seu hospedeiro para sobreviver e se reproduzir
(Nacher et al., 2000). Estas associações entre helmintíases e malária são bem documentadas
em regiões endêmicas da África e Ásia, principalmente com a malária causada por P.
falciparum. No Brasil, a infecção pelo P. vivax é predominante e, a má-nutrição e a forma
grave são pouco frequentes quando comparado com o continente africano. No entanto, em
áreas endêmicas brasileiras de malária vivax apenas um estudo de associação foi realizado em
uma população infantil da Amazônia oriental brasileira (Melo et al., 2010).
A erradicação da malária vem sendo discutida e a disseminação da resistência aos
antimaláricos é um grande problema para o seu efetivo controle. Portanto, avaliar a
possibilidade de que populações infectadas por parasitas intestinais possam influenciar na
dinâmica de transmissão dos parasitos da malária, bem como na compreensão da doença e sua
progressão, assim como fatores que influenciam o resultado clínico da infecção, são
fundamentais para estratégias diagnósticas e preventivas. A investigação proposta poderá
confirmar os achados já existentes no Brasil e avaliar o efeito da intensidade das
enteroparasitoses no curso da infecção pelo P. vivax.
1.5 IMUNOPATOGÊNESE DA COINFECÇÃO MALÁRIA E ENTEROPARASITOSES
O processo de aquisição de imunidade na malária é, ainda hoje, pouco compreendido,
sendo que inúmeros fatores o influenciam, como: a complexidade do ciclo biológico dos
plasmódios, sua extensa diversidade antigênica, o perfil de transmissão da área endêmica, a
maturidade do sistema imunológico relacionado à idade, status imunológico do indivíduo e/ou
os mecanismos inatos de resistência que o diferem (Langhorne et al., 2008). A resposta imune
contra malária envolve mecanismos inatos e adaptativos, com a participação de vários tipos
de células e anticorpos que podem levar a eliminação do agente ou a imunopatologia (Medina
et al., 2011).
Assim como o P falciparum, o P vivax também induz uma resposta imune específica
no hospedeiro por dois mecanismos: 1 - humorais (TH2) por meio de anticorpos que
fornecem proteção pelo bloqueio da invasão de merozoítos em novas hemácias, reduzindo a
citoaderência e melhoram a atividade fagocítica de macrófagos e monócitos. 2- celulares
(TH1) pelos linfócitos T que ativam os linfócitos B para produção de anticorpos e pela
liberação de citocinas que ativam macrófagos, natural killer (NK), células dendríticas e
neutrófilos para combater a infecção (Wipasa et al., 2011; Medina et al., 2011).

33
A invasão dos eritrócitos pelos plasmódios é um processo fundamental para o
estabelecimento da infecção malárica, constituindo assim um dos principais alvos para a
resposta imune protetora. Diversos trabalhos têm demonstrado que a imunidade direcionada
ao estágio eritrocítico pode contribuir para a redução/eliminação dos parasitos assim como
para o desenvolvimento das manifestações clínicas da doença (Polley et al., 2006; Osier et al.,
2008). As citocinas são moléculas protéicas secretadas por inúmeras células e pelo sistema
imune, com função sinalizadora para o sistema imune. Na malária vivax as citocinas são
liberadas depois da ruptura do eritrócito, com a consequente liberação de merozoitos no ciclo
eritrocítico (Medina et al., 2011).
O equilíbrio entre as citocinas pró e antiinflamatórias se faz necessário para garantir o
controle das alterações imunopatológicas, além de determinar os níveis de anemia, carga
parasitária, grau de pirexia, gravidade clínica, evolução da doença e até mesmo a morte
(Gonçalves et al., 2010; Jain et al., 2010). No entanto, pequenos níveis destas citocinas não
são suficientes para inibir o crescimento do parasito (Sohail et al., 2008). Apesar das
descrições clínicas da doença causada por P. vivax padrões referentes à resposta imunológica
e padrão de citocinas são escassos, e não completamente esclarecidos (Andrade et al., 2010;
Gonçalves et al., 2010; Medina et al., 2011). Vários estudos descrevem o papel do TNF-α,
IFN-γ, IL-10 e IL-4 para a infecção por P. falciparum e sua relação na patogênese da malária
(Vafa et al., 2007; Jain et al., 2010).
Adicionalmente a essa discussão, a superprodução e persistência desses mediadores
podem levar a imunopatologia, com gravidade da doença e até a morte (Sohail et al., 2008).
Os helmintos são conhecidos como reguladores de linfócitos T, levando a produção de
citocinas regulatórias da resposta Th1 (Maizels & Yazdanbakhsh, 2003). Estes nematoides
também modulam as funções das células dendríticas, consequentemente alterando a resposta
imune para os antígenos maláricos (Hartgers & Yazdanbakhs, 2006). Ademais, os ligantes
expressos pelos helmintos estimulam os receptores Toll-like, regulando negativamente a
capacidade de resposta imune durante a infecção concomitante (Boutlis et al., 2006; Wammes
et al., 2010). Sabe-se também que os helmintos provocam anemias e isso aumenta a
atratividade dos hospedeiros ao vetor, podendo tornar a pele mais refratária à picada dos
anofelinos (Hagel et al., 1993), deixando os hospedeiros menos atentos e incapazes de
responder ao ataque, facilitando a passagem do esporozoíto e aumentando a chance da
infecção sanguínea.
Uma das hipóteses específicas para a associação de helmintos com a proteção da
malária grave é que estes parasitos intestinais levam ao aumento do complexo IgE que ativa o

34
receptor para IgE de baixa afinidade (FC€RII) e assim libera IL10 (citocina antinflamatória),
que ativa a óxido nítrico sintetase, que por sua vez leva a liberação de óxido nítrico e reduz o
sequestramento de hemácias parasitadas (Nacher, 2002). Outra hipótese é que a proteção pode
ser induzida por transferência passiva de IgG de indivíduos imunes para malária para
pacientes infectados com o plasmódio (Roussilhon et al., 2010). Altas cargas parasitárias de
helmintos estão associadas com o não balanceamento Th2/Th1 (Pearce et al., 1991) e,
portanto, pode ser responsável por um direcionamento de resposta imune Th2 mediado por
helmintos contra antígenos maláricos.
Os helmintos reduzem os níveis de anticorpos citofílicos (IgG1 e IgG3), enquanto que
aumentam os não citofílicos (IgG2, IgG4 e IgM), mecanismo conhecido como inibição celular
dependente de anticorpos (ADCI) (Druilhe et al., 2005). A proteína de superfície do
merozoíto (MSP) é identificada como o principal alvo de anticorpos mediado por ADCI
(Oeuvray et al., 1994). A proteína de superfície de merozoíto-1 (MSP-1) é uma glicoproteína
de 195 kDa expressa em todas as espécies de malária estudadas, considerada de grande
potencial vacinal (Holder et al., 1992). Tem sido demonstrado que em P. falciparum a MSP-1
é sintetizada como um grande polipeptídio precursor, que é então processada
proteoliticamente em fragmentos menores durante o final do desenvolvimento do esquizonte
(Holder, 1988).
Diversos estudos demonstraram a elevada antigenicidade de diferentes regiões do
MSP-1 do P. vivax no Brasil (Soares et al., 1997; Soares et al., 1999; Nogueira et al., 2006;
Barbedo et al., 2007) e em outros países (Dent et al., 2009; Pitabut et al., 2007; Zeyrek et al.,
2008). A especificidade de anticorpos, ocorre em particular com a região C-terminal (42 kD e
19 subfragmentos-kD), bloqueando a invasão do parasito in vitro (Chappel & Holder, 1993;
Uthaipibull et al., 2001), e indução de imunidade protetora em modelos de animais (Arévalo-
Herrera et al., 1992; Egan et al., 2000).
Estudos de coinfecção malária-helminto estão direcionados para os mecanismos de
interação entre os parasitos por via resposta imunológica mediada, isto em virtude dos
conhecidos efeitos imunomoduladores causados pelos helmintos (Knowles, 2011). Isso se
caracteriza por altos níveis de citocinas como IL-4, IL-5 e IL-13, eosinofilia, altas
concentrações no soro de IgE (Maizels & Yazdanbakhsh, 2003; Smits et al., 2010) e de
anticorpos não-citofílicos IgG2, IgG4 e IgM (Druilhe et al., 2005). Apesar desta forte resposta
Th2, helmintos adultos frequentemente sobrevivem por anos em hospedeiros
imunocompetentes, possivelmente através da indução de mecanismos imunorregulatórios
(Hartgers & Yazdanbakhsh et al., 2006; Knowles, 2011).

35
Estes mecanismos incluem a indução de células T regulatórias e modulação de células
do sistema imune inato, como macrófagos e células dendríticas, o que resulta em ambiente
anti-inflamatório caracterizado por níveis aumentados de IL-10 e TGF-β (Hartgers &
Yazdanbakhsh, 2006). Esta rede regulatória no hospedeiro pode ser caracterizada como
hiporresponsividade imunológica, a qual protege a eliminação dos helmintos, mas ao mesmo
tempo protege o hospedeiro contra a patologia em virtude de uma possível inflamação
excessiva (Maizels & Yazdanbakhsh, 2003; Taylor et al., 2005). Esta hiporresponsividade não
é direcionada somente contra antígenos dos helmintos, mas parece se estender para antígenos
de terceiros.
Dessa forma, a resposta imune fortemente regulatória desencadeada pela infecção
helmíntica poderia prejudicar a resposta imune Th1 eficaz contra a infecção por plasmódio, o
que tornaria o hospedeiro mais suscetível a desenvolver altos níveis de parasitemia e sintomas
de malária. Por outro lado, esta resposta regulatória também poderia ter um efeito benéfico ao
hospedeiro por diminuir os efeitos nocivos de uma possível reação inflamatória exagerada
causada por citocinas do perfil Th1, assim ajudando a controlar a imunopatologia resultante
da infecção por plasmódio, neste caso poderia diminuir a probabilidade de desenvolver
quadros de malária grave (Knowles, 2011).
Além da influência do perfil regulatório das infecções helmínticas na malária, o forte
perfil Th2 pode se contrapor ao Th1, pela clássica dicotomia Th1xTh2, podendo também
induzir maior produção de anticorpos não citofílicos como IgG4 no lugar dos anticorpos
citofílicos, prejudicando a resposta eficiente na diminuição da carga parasitária. Entretanto o
real papel desempenhado por helmintos no curso da malária ainda não está esclarecido.
Estudos tem investigado as interações existentes entre o plasmódio e helmintos durante uma
coinfecção e demonstrado resultados conflitantes. É sugerido que ocorra tanto a presença de
sinergismo quanto de antagonismo nas infecções simultâneas por nematódeos e protozoários
(Cox, 2001; Knowles, 2011).
Assim, algumas pesquisas realizadas com modelos animais indicaram a existência de
efeito facilitador de Schistosoma sp. sobre a infecção por Plasmodium sp., enquanto outras
sugeriram um efeito supressor. Alguns estudos epidemiológicos reportaram um efeito protetor
da infecção helmíntica sobre a densidade parasitária na infecção por P. falciparum ou sobre as
manifestações clínicas da doença, enquanto outros demonstraram um risco aumentado de
infecção por Plasmodium sp. ou malária naqueles indivíduos coinfectados com helmintos.
Estes resultados contraditórios podem ser atribuídos as diferentes espécies de helmintos e
fenótipos estudados (clínico ou parasitológico) e as diferentes populações expostas a níveis

36
distintos de transmissão das infecções (Le Hesran et al., 2004; Lyke et al., 2005; Knowles,
2011).
Diversos estudos demonstram mudanças na resposta de anticorpos aos antígenos
plasmodiais em indivíduos coinfectados com helmintos. A resposta imune contra protozoários
intestinais não é tão destacadamente imunorregulatória como a resposta induzida por
helmintos, entretanto infecções crônicas por protozoários foram sugeridas como
possivelmente associadas ao perfil de resposta imune reguladora do tipo 1 (Díaz & Allen,
2007; Courtin et al., 2011). Por exemplo, a resposta imune contra Giardia duodenale, um dos
protozoários intestinais mais prevalentes, envolve tanto a resposta celular quanto a humoral.
Estudos relatam que os anticorpos contra giárdia atuam ativando a via clássica do sistema
complemento que leva à lise do parasito, limitando a invasão dos tecidos pelos trofozoítas de
giárdia (Faubert, 2000; Boef et al., 2012).
Entretanto a defesa mais substancial está na imunidade de mucosa e ação de
anticorpos IgA. A produção de IL-6 por linfócitos T estava aumentada nos indivíduos com G.
duodenale comparado aos que não abrigavam esse protozoário. Também ocorreu aumento nos
níveis de IL-10 produzido por células da imunidade inata nos indivíduos infectados com altas
cargas de G. duodenale, tornando possível a ideia de que altas cargas deste protozoário
intestinal induzam resposta reguladora inata, ambas as estimulações para secreção das duas
citocinas foram realizadas com antígenos não pertencentes à giárdia. Dessa forma é razoável
pensar que existem influencias em outras infecções no mesmo hospedeiro (Boef et al., 2012).
Os enteroparasitos podem causar aos seus portadores danos como obstrução intestinal
(A. lumbricoides), desnutrição (A. lumbricoides e T. trichiura), quadros de diarreia (G.
duodenale e E. histolytica) e anemia (Ancilostomídeos). Embora a anemia seja mais evidente
por infecções com Ancilostomídeos, esta alteração hematológica também foi associada com
infecção de moderada a alta intensidade de T. trichiura (Knowles, 2011). O impacto das
infecções por A. lumbricoides sobre a anemia ainda não está esclarecido. Como a anemia é
uma característica hematológica também fortemente associada à malária, diversos trabalhos
avaliaram o impacto da coinfecção malária-helminto sobre a anemia nos indivíduos
portadores dos dois tipos de parasitos, e alguns destes trabalhos encontraram resultados
contrastantes (Abanyie et al., 2013).
Em um trabalho realizado em crianças nigerianas, a frequência de anemia nas crianças
coinfectadas com P. falciparum e A. lumbricoides não foi diferente da frequência de
indivíduos anêmicos portadores somente de P. falciparum, indicando que neste caso a malária
é a principal causa de anemia. No Zimbábue, a prevalência de anemia no grupo de crianças

37
coinfectadas com P. falciparum e helmintos foram maiores que no grupo de crianças com
somente uma infecção (Midzi et al., 2010; Boef et al., 2012). Estudo realizado no estado do
Amazonas demonstrou que a concentração de hemoglobina durante a infecção malárica
encontrava-se aumentada nas crianças, que além da malária por P. vivax também estavam
infectadas com helmintos intestinais quando comparado às crianças que estavam somente com
P. vivax, sugerindo que a infecção helmíntica concomitante estava protegendo as crianças de
desenvolver anemia (Melo el al., 2010).
As respostas imunorreguladora e Th2 induzidas pelas infecções helmínticas podem
atenuar o perfil Th1 da malária, levando a diminuição da gravidade da anemia malárica,
através da secreção de citocinas como IL-10 e TGF-β. A eosinofilia, número de eosinófilos
maior que 450 células/mm3, é uma alteração hematológica bastante comum nas infecções
helmínticas. Geralmente as infecções por protozoários como Plasmodium spp. G. lambia e E.
histolytica não causam eosinofilia. Os eosinófilos são estimulados por citocinas específicas
como fator estimulador de colônias de granulócitos e macrófagos (GM-CSF), IL-3 e IL-5.
Linfócitos T ativados por infecções parasitárias produzem IL-5, estímulo predominante para
eosinofilia que assim como IL-4 levam ao aumento de produção de IgE para a resolução da
infecção helmíntica (Leder & Weller, 2000; Mueller et al., 2013).
Mesmo que a eosinofilia não seja uma característica marcante na malária, um estudo
demonstrou que indivíduos com infecções assintomáticas por P. falciparum apresentavam
aumento na frequência de eosinófilos e que durante a fase aguda da malária o número de
eosinófilos diminuía, mas ocorria elevação nos níveis plasmáticos de proteína eosinofílica
catiônica e proteína eosinofílica, marcadores de função de eosinófilos. Dessa forma, o menor
número de eosinófilos durante a fase aguda da malária seria devido ao sequestro destas
células nos tecidos e não em virtude de menor produção de eosinófilos na medula óssea
(Roussilhon et al., 2010).
Assim, é possível que os eosinófilos ou seus grânulos possam participar na resposta
contra a malária. Assim, nas coinfecções de malária com parasitoses intestinais as alterações
nos números de eosinófilos poderiam ter alguma implicação na resposta imune direcionada
contra ambos os parasitos (Lacerda et al., 2012). Estes resultados podem ser atribuídos as
diferentes espécies de helmintos e fenótipos estudados e as diferentes populações expostas a
transmissão das infecções.

38
1.5.1 Resposta imune de anticorpos e citocinas
O estudo das respostas de anticorpos aos antígenos de Plasmodium é um processo
chave para a descoberta e desenvolvimento de vacinas contra a malária. Entretanto, a
dificuldade prática de uma vacinação contra a malária é, devido à necessidade de cultura dos
parasitos, em larga escala e livres de contaminação com componentes celulares do hospedeiro
e pela complexidade do ciclo de vida do Plasmodium, que resulta em uma resposta
imunológica específica para cada forma evolutiva, e pelo fato do parasito ter desenvolvido
diversas estratégias de escape à resposta imune do hospedeiro (Good et al., 2005). Além de
apresentar um custo relativamente alto, as preparações antigênicas são de difícil
reprodutibilidade e consequentemente, o desenvolvimento de uma vacina antimalárica
necessita de modernas técnicas para a produção de antígenos (Hill, 2011).
O desenvolvimento de vacinas contra a malária, sobretudo na busca de imunógenos
efetivos e de baixo custo, representa um desafio científico significante no controle dessa
infecção (Good et al., 2005). Nos últimos anos um progresso tem sido obtido na identificação
e caracterização molecular e imunológica de proteínas de vários estágios do P. falciparum
candidatas a uma vacina. No caso do P. vivax, apesar de sua alta prevalência em várias partes
do mundo, inclusive no Brasil, a busca de vacinas é pouco explorada e o número de genes
clonados que codificam essas proteínas antigênicas ainda é pequeno quando comparado ao P.
falciparum (Galinski; Barnwell, 1996).
Uma das razões principais é a falta de um sistema in vitro para o cultivo contínuo do
P. vivax, já estabelecido para o P. falciparum e as dificuldades de manutenção do parasito em
cultura se devem ao fato de que o P. vivax invade preferencialmente ou quase que
exclusivamente reticulócitos os quais normalmente correspondem a apenas 1% da população
total de eritrócitos no homem (Mons et al., 1988; Barnwell et al., 1989). Consequentemente, a
obtenção de reticulócitos em quantidades suficientes para manter um cultivo contínuo no
laboratório não é uma tarefa fácil.
Por essa razão, inexiste ainda uma vacina eficaz utilizada no combate à malária,
principalmente no que tange a espécie de P. vivax. É sabido que uma vacina contra esta
espécie é de suma importância, levando em consideração que as infecções por P. vivax são as
mais prevalentes em áreas endêmicas. Além disso, estima-se que as formas latentes
responsáveis pelas reinfecções (hipnozoíticas) contribuem com aproximadamente 80% de
todas as infecções do estádio sanguíneo por P. vivax. Como não há diagnóstico para as formas
latentes, os hipnozoítas são importantes fontes de reintrodução do P. vivax (White, 2011).

39
As vacinas pensadas para combater a malária são classificadas de acordo com a fase
em que elas atuam para suprimir o parasito. Uma vacina pré-eritrócitica reduziria a incidência
de infecções primárias, além de evitar o estabelecimento das formas latentes, reduzindo as
reinfecções que contribuem para a transmissão continuada. A vacina do estádio sanguíneo
bloquearia o crescimento do parasito e preveniria a doença e a morte do hospedeiro. Uma
vacina de bloqueio da transmissão impediria a infecção do mosquito vetor, dessa forma, iria
acelerar a erradicação da doença (Nilsson et al., 2015). Essas estratégias focam,
principalmente, a determinação de antígenos-alvo, essenciais para o desenvolvimento e
sobrevivência do parasito.
Diversos antígenos de estágios assexuados sanguíneos têm despertado interesse como
alvos potenciais de uma possível vacina protetora. Dentre esses antígenos, temos a proteína de
superfície de merozoítas (MSP-119) é uma glicoproteína polimórfica expressa sobre a
superfície do merozoíto de P. vivax (Good et al., 2005; Holder, 2009) e a proteína de antígeno
de membrana apical (AMA-1) proteína imunogênica integrante da membrana que é expressa
na superfície apical dos merozoítos e esporozoítos, e possuem importância durante a invasão
nos eritrócitos e hepatócitos (Hodder et al., 2001, Silvie et al., 2004). Ambas as proteínas tem
se destacado como um dos principais objetos de diferentes estudos em malária vivax (Herrera
et al., 2007). A resposta imune contra os estágios assexuados sanguíneos está associada com a
resposta mediada por anticorpos.
Estudos de transferência de soro de indivíduos imunes, quando transferidos para
indivíduos não imunes, protegem contra as formas eritrocíticas do parasito, demonstrando que
aqueles indivíduos que receberam o soro apresentaram proteção contra a malária clínica ou
diminuição na gravidade e carga parasitária da doença. Estas evidências sugerem que o
estágio eritrocítico é o principal alvo para a imunidade naturalmente adquirida (Doolan et al.,
2009). Estudos como esses demonstraram que os anticorpos desempenham um papel
importante na aquisição de imunidade contra a malária clínica.
Nesse contexto, Sánchez-Arcila et al (2015) avaliando a resposta imune de anticorpos
de P. vivax por AMA-1 e MSP-119 numa população de Rondônia no Brasil, evidenciam que
embora a malária seja mais frequente em indivíduos infectados com parasitos intestinais,
parâmetros hematológicos e de parasitemia não diferiram entre os indivíduos infectados com
parasitos intestinais, maláricos e coinfectados. Parece que a alteração hematológica observada
tanto na malária quanto nos grupos coinfectados deve-se ao efeito da malária, em vez de
parasitas intestinais ou coinfecção.

40
Apesar de haver mudanças em IgG total dirigidas a PvAMA-1 e PvMSP-119 no grupo
de parasitos intestinais, uma diminuição da IgG e nas respostas citofïlicos associados à
coinfecções não foi observado. Estas respostas talvez relacionem com outros fatores, como
propriedades de antígenos, número e tempo de exposição, idade e determinantes genéticos do
hospedeiro. Novos estudos devem ser realizados para determinar o efeito de protozoários
intestinais na resposta imune à diferentes antígenos da malária (Sánchez-Arcila et al., 2015).
Adicionalmente, é importante destacar que a presença da resposta de anticorpos para P. vivax
AMA-1 e MSP-119 proteínas de todos os grupos indicaram que os participantes haviam sido
expostos à infecção da malária e as respostas da subclasse de IgG foram em grande parte de
acordo com pré-resultados posteriormente publicados (Supali et al., 2010; Turner et al.,
2008).
Essas interações imunológicas entre helmintos e parasitos da malária não são claras e
há pouco consenso entre essa coinfecção. Entretanto, durante a infecção da malária, as
citocinas são referidas como sendo importantes marcadores moleculares mediadas por células
de resposta imune e conhecido por serem críticos na regulação de doenças (Sánchez-Arcila et
al., 2014). Essas infecções helmínticas tem um profundo efeito sobre o sistema imunológico,
e que podem inibir a capacidade do hospedeiro para montar uma resposta imune do tipo Th1.
Com efeito, a infecção com Ascaris sp. induz respostas antiinflamatórias Th2 associada pelos
níveis elevados de interleucina IL-10. Portanto, espera-se que a influência da infecção
helmíntica no sistema imunitário poderia estender-se a resposta imunitária contra o paludismo
devido ao efeito antiinflamatório de citocinas induzidas por helmintos e, assim,
possivelmente, afetar o curso da infecção de malária (Geiger et al., 2002; Figueiredo et al.,
2010).
Adicionalmente a maioria dos estudos enfatiza que o P. vivax induz uma maior
produção de citocinas em relação a P. falciparum mesmo com biomassa parasitária
semelhante (Anstey et al., 2009). Os estudos reportam a importância do padrão de citocinas e
de suas interações na resposta imune do hospedeiro durante infecção por P. vivax (Sortica et
al, 2012). O aumento da parasitemia, bem como a resposta imunológica durante o percurso da
doença são fatores que determinam sua gravidade (Sortida et al., 2012). Os níveis elevados de
citocinas do tipo Th1 e Th2 como IFN-γ e IL-4, respectivamente, têm sido associados à
gravidade da doença, no entanto existem muitas dúvidas sobre o papel imunológico destas
citocinas na patogênese da doença (Tangteerawatana et al., 2007).
Na infecção por P. vivax, estudos vêm evidenciando o papel protetor da citocina IL-4
como regulador negativo dos efeitos pró-inflamatórios ocasionada por P. vivax. Apesar de

41
possuir baixos níveis sistêmicos, IL-4 parece ser um mediador central com várias interações
significativas em indivíduos assintomáticos (Cabantous et al., 2009; Mendonça et al., 2013).
Por outro lado, citocinas antiinflamatórias como TGF-β e IL-10 nas fases iniciais da infecção
correlacionam-se a elevadas parasitemias, porém em fases tardias da infecção parecem
prevenir sintomas mais graves (Walther et al., 2009). Durante a infecção, as citocinas
desempenham um papel divergente, controlando o crescimento do parasito e exacerbando a
patologia (Kabyemela et al., 2013). Os efeitos antagônicos são atribuídos ao tempo de
expessão e ao equilíbrio das citocinas anti e pró-inflamatórias (Dodoo et al., 2002). No
entanto, esses mecanismos imunopatológicos da malária ainda não foram totalmente
elucidados, constituindo-se um campo amplo para novos estudos.
Com isso, à capacidade de alguns parasitos intestinais, principalmente helmintos
induzir respostas antiinflamatórias do hospedeiro para seu próprio benefício, fortalece a
hipótese de que pode haver um contrapeso entre citocinas pró-inflamatórias, associadas à
malária, e citocinas anti-inflamatórias, associadas a helmintos ou protozoários parasitos
intestinais. Embora as respostas de citocina tenham sido extensivamente descritas na infecção
pelo P. falciparum, poucos estudos analisaram a concentração de citocinas sistêmica em
coinfecção de parasitos intestinais e malária por P. vivax (Maizels et al., 2004).
No Brasil, a transmissão da malária concentra-se na Região Amazônica, sendo que o
P. vivax tem sido a espécie mais prevalente, responsável por aproximadamente 84% dos casos
nesta região (Gomes et al., 2016; Dotrário et al., 2016). A manutenção do número de casos,
juntamente com o rápido desenvolvimento do P. vivax nos mosquitos vetores (Olliaro et al.,
2016) e o controle dificultado por métodos tradicionais em regiões endêmicas, tem mostrado a
importância da identificação de técnicas efetivas e economicamente viáveis, que possam
interromper a transmissão da malária em regiões endêmicas (Mueller et al., 2015).
Indivíduos que estão frequentemente expostos ao parasito, tem a capacidade de
desenvolver respostas imunes específicas, protegendo-os de evoluir a uma infecção malárica
grave. Além disso, o soro desses indivíduos pode impossibilitar a fertilização dos gametas,
impedindo o desenvolvimento parasitário e, consequentemente, bloqueando a transmissão
desse parasito de pacientes infectados para os mosquitos (Greenwod et al., 2008) viabilizando
uma vacina de bloqueio de transmissão (Arévalo-Herrera et al., 2010, 2011, 2015). Em
síntese, estas vacinas são negligenciadas por não previrem a infecção em indivíduos
vacinados ou moderarem o curso da doença, sendo um empreendimento financeiro pouco
atraente para as empresas de vacinas nos países desenvolvidos.

42
1.6 JUSTIFICATIVA
As regiões tropicais são conhecidas pela endemicidade da coinfecção da malária-
parasitos intestinais. Embora prevalentes, pouco se sabe sobre esta interação e como as
infecções simultâneas afeta a epidemiologia e/ou patogénese de cada morbidade. Em
Oiapoque, município do Estado do Amapá, norte do Brasil, fronteira ocidental da Amazônia
com a Guiana Francesa, apresenta ao longo dos anos índice parasitário anual elevado, e um
conjunto de atributos naturais que reflete a influência imposta pela conjugação dos domínios
guianense e amazônico brasileiro, classificando-o como município ou área de risco
permanente e constante para transmissão da malária e parasitos intestinais.
Essa coinfecção humana por múltiplas espécies de parasitos são reais e relativamente
frequentes na natureza e diversos estudos epidemiológicos indicam que essa interação
representa um desafio para compreender a ecologia dos parasitos e criar estratégias
antiparasitárias fundamentais e essenciais na compreensão dos mecanismos que regem as
respostas imunes em benefícios a saúde humana. Adicionalmente, informações contraditórias
sobre a consequência benéfica ou não, alertam sobre os possíveis efeitos das coinfecções no
desenvolvimento de uma vacina contra a malária. Entretanto, os dados são controversos e
conflitantes, alguns mostrando uma maior susceptibilidade, outros mostram menor
susceptibilidade ou ainda, nenhuma influência na evolução clinica da malária.
Tendo em vista que os trabalhos que abordam as coinfecções entre plasmódio e
parasitoses intestinais são raros, e que estes são principalmente voltados para P. falciparum, é
importante o desenvolvimento de estudos direcionados para P. vivax, já que esta espécie é a
principal responsável pelos casos de malária no Brasil. Considerando que nas áreas endêmicas
de malária a população também esteja exposta é que a coinfecção de malária com parasitos
intestinais (helmintos e protozoários) possa causar alterações no balanço entre a resposta
imune e a evolução clínica do hospedeiro infectado, torna os estudos de coinfecção entre os
dois parasitos de grande importância no planejamento de ações antimaláricas em áreas
endêmicas de P. vivax.
Adicionalmente ao contexto das interações fronteiriças do Brasil com a Guiana
Francesa, a migração humana é uma realidade entre as diferentes regiões, onde as
intensidades de transmissão e os antecedentes genéticos humanos são bem distintos,
contribuindo para a transmissão de novos casos de malária, que teoricamente em pacientes
coinfectados poderia levar a uma maior diversidade genética dos parasitos que circulam na
população, constituindo um sério agravante para a saúde pública. Associado a estes aspectos

43
sociais, está o alastramento de casos, surgimento da resistência às drogas pelo P. vivax nos
últimos anos, complicações clínicas associadas com casos fatais, bem como, resultados não
promissores no desenvolvimento de vacinas, gerando um grande problema para o seu efetivo
controle, incapaz de erradicar a doença sem o auxílio de medidas inovadoras.
Além disso, as pesquisas precisam levar em consideração a influência do
poliparasitismo na imunogenicidade de antígenos candidatos a vacinas. A descrição de
determinantes de proteção ou gravidade são fundamentais para guiar futuros estudos no
manejo e controle epidemiológico da malária. A elucidação do padrão geográfico do
complexo P. vivax é importante para as avaliações de diversidade, caracterizando e
identificando ferramentas de predição do diagnóstico, além de possíveis alvos terapêuticos
que codificam antígenos candidatos a vacinas, assim como, na formulação e estruturação de
medidas de vigilância que visem o controle da malária.
Por essas razões, julga-se pertinente estudar o perfil epidemiológico, imunológico e
molecular dos povos de uma área amazônica e fronteiriça entre Brasil e Guiana Francesa, o
que decorre da exiguidade de investigações, da ausência de inquéritos e censos, bem como da
precariedade de informações sobre morbidade e mortalidade. Portanto, avaliar a possibilidade
de que populações infectadas por enteroparasitos possam influenciar na dinâmica de
transmissão dos parasitos do P. vivax, bem como na compreensão da doença e sua progressão,
e nos fatores que influenciam o resultado clínico da infecção, são fundamentais para
estratégias diagnósticas e preventivas aumentando a capacidade de controle da malária e
contribuindo para a eliminação da doença.
1.7 HIPÓTESES
1.7.1 Hipótese Nula
A coinfecção de enteroparasitos não influencia nos níveis de hemoglobina, no número de
gametócitos circulantes na corrente sanguínea e na resposta imune na malária causada pelo
Plasmodium vivax em pacientes de uma área endêmica da Amazônia.
1.7.2 Hipótese Alternativa
A coinfecção de enteroparasitos influencia na infecção pelo Plasmodium vivax em
pacientes de uma área endêmica da Amazônia.

44
Variável
independente
Coinfecção
pelos
Enteroparasitas
Variáveis
dependentes
Infecção pelo Plasmodium vivax
Quantidade de gametócitos circulantes
Concentração de hemoglobina
Níveis de anticorpos
Perfil de citocinas
1.8 OBJETIVOS
1.8.1 Geral
- Avaliar a influência da coinfecção de parasitos intestinais na malária causada pelo
Plasmodium vivax em pacientes do município de Oiapoque no Estado do Amapá, área
endêmica da Amazônia brasileira.
1.8.2 Específicos
- Caracterizar por gênero e idade da população os enteroparasitos (helmintos e protozoários)
detectados;
- Identificar a prevalência dos enteroparasitos da população estudada;
- Correlacionar a frequência dos enteroparasitos com os níveis de hemoglobina da população
de estudo;
- Determinar a distribuição e prevalência de indivíduos com diagnóstico de malária vivax e
parasitos intestinais entre os grupos e subgrupos de estudo;
- Correlacionar dados epidemiológicos e hematológicos na coinfecção malária-parasitos
intestinais;
- Avaliar os níveis de anticorpos IgG MSP-119 e as possíveis alterações associadas as
coinfecções malária-parasitos intestinais;
- Verificar o perfil de citocinas Th1 (IFN-γ, TNF-α e IL-2) e Th2 (IL-4, IL-5 e IL-10) e as
possíveis alterações associadas as coinfecções malária-parasitos intestinais;

45
2. MATERIAL E MÉTODOS
2.1 ÁREA DE ESTUDO E POPULAÇÃO
Estudo transversal foi realizado, no município de Oiapoque, pertencente ao Estado do
Amapá, norte do Brasil, fronteira ocidental da região amazônica. Possui IPA de alto risco para
transmissão de malária, fica localizado na parte mais setentrional do Estado do Amapá. Faz
fronteira ao norte com a Guiana Francesa, ao sul com os municípios de Calçoene, Serra do
Navio e Pedra Branca do Amapari, ao leste é banhado pelo Oceano Atlântico e a oeste limita-
se com o município de Laranjal do Jari. Está situado a 590 km da capital, Macapá, ligando-se
por via terrestre através da BR 156, onde a presença de trechos estreitos e perigosos,
principalmente no período das chuvas, quando diversos locais tornam-se intrafegáveis em
razão dos atoleiros (Gea, 2013). Possui uma população de 20.509 habitantes localizado a uma
latitude norte de 03° 50’ 35” e longitude oeste 51° 50’ 06”segundo IBGE (2010) (Figura 11).
Figura 11 - Mapa da América do Sul, mostrando a localização da Guiana Francesa e
Oiapoque no Estado do Amapá (Fonte: Adaptada de Gomes et al., 2016).
De acordo com Palhares & Guerra (2016), o município de Oiapoque apresenta um
conjunto de atributos naturais que reflete a influência imposta pela conjugação dos domínios
guianense e amazônico. A cobertura vegetal corresponde fisionomicamente a dois padrões de

46
vegetação distintos: o domínio das formações florestadas e o domínio das formações
campestres. É cortado por vários cursos d’água e com uma base geológica e geomorfológica
que apresenta características peculiares, como morros graníticos, quedas d’água, pedreiras,
entre outras. A área geográfica de Oiapoque é de 22.625 km2. O clima é equatorial, com
umidade relativa média de 82%. A precipitação anual varia entre 2.700 e 3.300 mm. A
temperatura média é de 27 ºC, variando entre 26 e 33 °C. Temperaturas mais altas coincidem
com os meses mais secos do ano de setembro a novembro (ANA, 2015).
É composto por uma sede municipal, Oiapoque, e quatro distritos: Clevelândia do
Norte (área de destacamento militar do exército); Vila Velha (área de propriedades agro-
extrativas); Vila Brasil (serve de apoio aos garimpos infiltrados nas Guiana Francesa);
Taperebá (área de apoio aos pescadores da costa marítima). Outras localidades na área
geográfica municipal: Ponte do Cassiporé (intercessão da BR-156); Rio Cassiporé, ponto de
apoio para o tráfego rodoviário da BR-156 e fluvial, tanto para agricultores da região como
para os povoados indígenas da região (Gea, 2013).
Na cidade de Oiapoque circulam moradores, visitantes, comerciantes, garimpeiros,
militares e índios. Em decorrência da fronteira com o município de Saint George (Guiana
francesa), apresenta um fluxo populacional intenso, com movimento frequente de catraias que
transportam passageiros de um lado para outro, franceses e brasileiros, numa relação que
envolve comércio, turismo, vida social e compras, mas, que também favorece a disseminação
de doenças transmissíveis (Andrade, 2005).
A economia local gira em torno do comércio de produtos brasileiros vendidos aos
Guianenses, do cultivo da mandioca, extrativismo, venda de produtos hortifrutigranjeiros
comercializados no mercado local. Não se pode deixar de mencionar a atividade do garimpo
que serve para dinamizar a economia local (comércio, hotéis, bares etc.). Também existe a
pesca e a caça de subsistência de diversas espécies nativas encontradas nos rios e florestas da
região (Silva, 2013). A exploração sexual também é fortemente encontrada no local.
2.2 CRITÉRIOS DE SELEÇÃO
Os indivíduos incluídos neste estudo foram triados em quatro postos de diagnóstico de
malária, localizado nos bairros Paraíso, Planalto, Nova Esperança e Infraero. Foram incluídos
neste estudo os pacientes maláricos (Plasmodium vivax) e não maláricos que preencheram os
critérios de seleção da pesquisa. Todos os casos positivos (malária e enteroparasitas) foram
devidamente tratados e acompanhados pela equipe técnica composta de dois agentes
notificantes, dois microscopistas, um agente de endemia, um enfermeiro, um biomédico e um

47
médico, na unidade mista de saúde do Oiapoque e no Laboratório de Fronteira, priorizando
sempre o bem estar do paciente. Para tal, foi utilizada a ficha de inclusão e acompanhamento
do projeto.
2.2.1 Critérios de inclusão para paciente maláricos
- Termo de consentimento assinado para maiores de 18 anos
- Termo de Assentimento assinado para menores de 18 anos
- Consentimento pós-informado dos pais/responsáveis pelas crianças e adolescentes (se
menores de 18 anos)
- Impressão digital para aqueles não alfabetizados
- Gota espessa positiva para P. vivax, independente da quantificação da parasitemia
- Idade: crianças maiores de 7 anos, adolescentes e adultos
- Primoinfecção ou história anterior de malária há mais de trinta dias
- Ser nativo da área de estudo
2.2.2 Critérios de exclusão
- Falta de consentimento/autorização para participação na pesquisa
- Gravidez referida
- Uso de tratamento antimalárico nos 30 dias anteriores
- Pacientes com doenças degenerativas, tuberculose e leishmanioses
- Crianças menores de 7 anos.
2.2.3 Critérios de inclusão para indivíduos não maláricos
- Termo de consentimento assinado para maiores de 18 anos
- Consentimento pós-informado dos pais/responsáveis pelas crianças e adolescentes (se
menores de 18 anos)
- Impressão digital para aqueles não alfabetizados
- Gota espessa negativa e sem sinais clínicos de malária
- Idade: crianças maiores de 7 anos, adolescentes e adultos
- Ser nativo da área de estudo.
2.2.4 Critérios de exclusão
- Falta de consentimento/autorização para participação na pesquisa;

48
- Gravidez referida;
- Crianças (maiores de 7 anos), adolescentes e adultos com malária por P. vivax, P.
falciparum, P. malariae ou malária mista (P. falciparum e P. vivax);
2.3 TAMANHO DA AMOSTRA
Os dados epidemiológicos, tais como idade, sexo e número de episódios de malária
anteriores foram obtidos durante uma entrevista realizada e também a partir dos prontuários
médicos. Considerando um tamanho finito de 20.509 indivíduos, com o erro tolerável de
amostragem de 5% (ε) = 0,05, associado ao intervalo de confiança de 95%, onde, tendo-se um
α = 0,05 e utilizando-se o valor maximizado para a Incidência de 0,25 seria necessário incluir
no mínimo 377 indivíduos na pesquisa, sendo incluídos 441 indivíduos que foram
comparados os resultados a partir de quatro grupos: 1) pacientes com coinfecções por
parasitas intestinais e malária; 2) pacientes com malária e resultado negativo para parasitas
intestinais, 3) pacientes sem malária e positivo para parasitas intestinais e 4) pacientes
negativos para as duas parasitoses.
2.4 COLETA E EXAME DE SANGUE
2.4.1 Coleta Sanguínea
Amostras de sangue foram colhidas por punção venosa para identificar os níveis de
hemoglobina (com anticoagulante) e para o preparo da gota espessa em lâmina (sem
anticoagulante). Após a coleta, a lâmina foi mantida em temperatura ambiente para secagem
da gota de sangue, sendo posteriormente corada e analisada por microscópia ótica de aumento
de 100X (Nikon, Japão), possibilitando quantificar a intensidade do parasitismo, mediante a
determinação da parasitemia por volume (µL) de sangue.
2.4.2 Diagnóstico de Malária
As lâminas foram preparadas como recomendado pela técnica de Walker (azul de
metileno e Giemsa) (Herrera et al., 2014) e avaliada pelo microscopista local do posto de
diagnóstico de malária do município. A contagem direta de parasitos, foi realizada pela
estimativa da parasitemia a partir da avaliação semiquantitativa, que registra o intervalo de
parasitemia por µl de sangue, com 200 campos e o número de parasitos encontrados foram
multiplicados por 5 (Bueno et al., 2012).

49
2.4.3 Detecção de Gametócitos
Foi realizada por contagem direta na lâmina, pelo microscopista local do posto de
diagnóstico de malária do município. A contagem das formas sexuais (gametocitemia) das
lâminas de Gota espessa foi realizada por meio de uma revisão das lâminas positivas para
quantificação dos gametócitos em 100 leucócitos. A conversão para gametócitos/µL foi
realizada por cálculo utilizando como valor de referência 8000 leucócitos/mm3, sendo
realizado a contagem direta de gametócitos na lâmina (Nº de gametócitos X Nº de leucócitos
(8000)/100 = gametócitos por µL). Este procedimento é semelhante aos métodos diagnósticos
preconizados pelas diretrizes do Clinical and Laboratory Standards Institute (CLSI) do
Centers for Disease Control (CDC) e também similar às diretrizes da Organização Mundial de
Saúde (OMS).
2.4.4 Concentração de Hemoglobina
A concentração de hemoglobina foi medida no sangue venoso, utilizando equipamento
automatizado (Mindray-BC-3000plus) do Hospital de Oiapoque. A anemia média foi definida
através dos valores de referencias da hemoglobina, sendo os indivíduos considerados
anêmicos quando seus níveis de hemoglobina eram ≤ 13 g/dL de sangue em homens e ≤ 12 g
/dL de sangue em mulheres.
2.4.5 Controle de Qualidade do diagnóstico parasitológico de malária
Todas as lâminas positivas foram revisadas e encaminhadas ao setor de controle de
qualidade de endemias do Laboratório Central do Amapá (LACEN/AP). Esse controle da
qualidade foi realizado através da avaliação de três fatores: coloração da lâmina, confecção da
lâmina e concordância ou divergência dos resultados. Posteriormente são expedidos laudos
com os resultados das análises de volta aos laboratórios e aos coordenadores de endemias do
município.
2.5 COLETA E EXAME DE AMOSTRAS FECAIS
A todos os indivíduos foi solicitado a fornecer amostras fecais pela manhã, sendo
proporcionados dois recipientes de plástico. Um com solução conservante (formol a 10%) e
outro sem solução conservante, para os casos negativos três amostras fecais foram coletadas e
examinadas utilizando os métodos: a fresco/direto sem e com lugol, Hoffman-Pons-Janer e
flutuação por centrifugação em solução de sulfato de zinco. Para cada amostra de fezes, duas

50
lâminas foram examinadas para a detecção dos parasitos por dois investigadores com
experiência na identificação, utilizando microscopia óptica (Nikon, Japão) com ampliações de
100X e 400X. As análises foram realizadas em Laboratório privado do município do
Oiapoque/AP.
2.6 DIAGNÓSTICO MOLECULAR DE PLASMÓDIO
Estas análises foram realizadas no Laboratório de Imunogenética de Malária no
Instituto Evandro Chagas/SVS/MS no Pará. O DNA foi extraído do sangue total utilizando o
kit Easy-DNATM (Invitrogen, Carlsbad, California, USA) and the QIAamp® DNA Blood Kit
(Qiagen, Inc., Chatsworth, CA). A confirmação do diagnóstico parasitológico foi realizada
pelo protocolo de PCR aninhado de acordo com o descrito previamente por Snounou et al.
(1993).
2.7 AVALIAÇÃO DE ANTICORPOS IgG CONTRA A PVMSP-119
A avaliação de anticorpos IgG total contra a MSP-119 foi realizada no Laboratório de
Microbiologia e Imunologia na Universidade Federal do Pará, seguindo o protocolo de Cunha
et al (2001). A proteína recombinante (His6-MSP-119) da MSP-1 do P. vivax (cepa Belém)
expressa em Escherichia coli. Os 96 poços das placas de ELISA (Costar, Corning Inc., NY,
EUA) foram sensibilizados com 50 μL da proteína His-MSP-119 (4 μg/mL) diluída em
tampão carbonato 0,05 M pH 9,0 (Na2CO3 15 mM, NaHCO3 34,9 mM), por 16 horas a
temperatura ambiente.
Posteriormente, as placas foram lavadas com PBS (phosphate buffered saline) Tween
0,05% e bloqueadas com 200 μL de PBS leite 5% (Molico, Nestlé®). Após 2 horas a 37°C, as
placas foram lavadas com PBS Tween 0,05% e 50 μL do plasma, diluído 1:100 em PBS leite
5%, foram adicionados em cada poço, em duplicata, e as placas foram incubadas por 16 horas
a temperatura ambiente. Após nova lavagem das placas com PBS Tween 0,05%, foram
adicionados, em cada poço, 50 μL de conjugado anti-IgG humano ligado a peroxidase
(DAKO polyclonal rabbit, Glostrup, Dinamarca), diluído 1:10.000 em PBS leite 5%, e as
placas foram incubadas por 2 horas em temperatura ambiente.
Em seguida, após nova lavagem com PBS Tween 0,05%, as placas foram reveladas
com a adição de 100 μL de OPD (orto-fenilenodiamina) (1 mg/mL) diluído em tampão
fosfato-citrato (NaH2PO4 0,2 M, C6H8O7 0,2 M, pH 5,0) contendo 0,03% de peróxido de

51
hidrogênio. Essa reação foi mantida no escuro e interrompida após 10 minutos pela adição de
25 uL de H2SO4 4 N em cada poço. A densidade ótica (OD) foi quantificada em leitora de
ELISA (EL800 Bio Tek, Winooski, EUA) em um comprimento de onda de 490 nm.
Para definição das amostras negativas ou positivas, foi calculado o índice de
reatividade (IR), determinado pela divisão do valor da OD da amostra pelo ponto de corte
(Corte: PvMSP-119 = 0.165). Amostras com IR ≥ 1 foram consideradas positivas, enquanto
que amostras com IR < 1 foram consideradas negativas. O ponto de corte foi estabelecido pela
média das ODs de 56 amostras de plasma de indivíduos sem histórico de malária, da
Fundação Centro de Hemoterapia e Hematologia do Pará (HEMOPA) residentes em Belém -
Pará, acrescida de três desvios padrões.
2.8 NÍVEIS SÉRICOS DE CITOCINAS
O Kit CBA (BD) (cytometric beads array) foi usado nesse estudo para a quantificação
das citocinas IL-2, INFγ e TNF (Th1) e IL-4, IL-5, IL-10 (Th2) em uma mesma amostra. Seis
populações de beads com distintas intensidades de fluorescência são conjugadas com um
anticorpo de captura específico para cada citocina, misturadas para formar o CBA e lidas no
citômetro de Fluxo do tipo FACSCanto II (Becton Dickinson, San José, CA, EUA),
previamente calibrado com “setup beads,” incubadas com Isotiocianato de fluoresceína
(FITC) ou Ficoeritrina (PE), de acordo com as recomendações do fabricante.
Uma curva padrão foi realizada para cada citocina e analisadas pelo FACSDiva
Software (Becton Dickinson, San Jose,CA, EUA). As populações de beads foram visualizadas
de acordo com as suas respectivas intensidades de fluorescência: da menos brilhante para a
mais brilhante. No CBA, as beads de captura das citocinas foram misturadas com o anticorpo
de detecção conjugado com o fluorocromo PE, e depois incubadas com as amostras para
formar o ensaio "em sanduíche". Os tubos para aquisição foram preparados com: 50 µL de
amostra, 50 µL da mistura de beads e 50 µL do reagente de detecção Th1/Th2 PE (Human
Th1/Th2 PE Detection Reagent/1 vial, 4mL). O mesmo procedimento foi realizado para a
obtenção da curva-padrão. Os tubos foram homogenizados e incubados por três horas, em
temperatura ambiente, no escuro.
Os resultados foram gerados em gráficos e tabelas utilizando-se o software FCAP
Array 3. (Becton Dickinson, San Jose,CA, EUA). Para cada citocina foram quantificados os
valores brutos de Media de Intensidade de Fluorescência (MFI). Os valores foram expressos
em pg/mL para cada citocina em comparação com a curva padrão, foram considerados 300

52
eventos para cada citocinas. Estas análises foram realizadas no Laboratório de Imunogenética
de Malária no Instituto Evandro Chagas/SVS/MS no Pará.
2.9 ANÁLISE ESTATÍSTICA
Os resultados dos dados epidemiológicos obtidos a partir do questionário, de cada
voluntário foram armazenados no banco de dados do programa Epi-Info 3.5.1 (CDC, Atlanta,
GA, EUA). Os valores de cada grupo (Malária, coinfectados, enteroparasitas e controle
endêmico) assim como os subgrupos (Helmintos, protozoários e associação de helmintos-
protozoários) foram expressos em percentuais. A idade, tempo de residência no Oiapoque
(ano) (TR), número de episódios de malária anterior (NEM), período da última malária
(meses) (PUM), níveis de hemoglobina (g/dL), parasitemia (parasites/𝜇L) e gametócitos
foram expressos em Mediana (1º e 3º Quartil) utilizando o software estatístico BioEstat 5.3.
As diferenças entre os grupos quanto a idade, TR, NEM, PUM e hemoglobina foram
calculadas utilizando um Tukey a partir e uma análise de variância, ANOVA um critério. Para
calcular o grau de anemia foi utilizado o teste Kruskal-Wallis-Dunn.
Os testes estatísticos utilizados foram escolhidos considerando-se o tipo e tamanho de
cada variável e o número de grupos avaliados, sendo as diferenças de parasitemia e de
gametocitos entre os grupos: malária e coinfectado calculadas usando o teste Wilcoxon-
Mann-Whitney. Para o índice de reatividade (IR) PvMSP119 entre os grupos estudados, foram
realizadas múltiplas correlações através do teste não paramétrico de Kruskal-Wallis seguido
de pós-teste de Dunn. Os dados foram expressos no formato Boxplot (valor mínimo ao valor
máximo, P25%-P75% e mediana), As diferenças significativas foram estimadas pela mediana
dos valores em cada grupo, sendo consideradas significativas aquelas onde p ≤ 0,05.
Para analise da expressão das citocinas, o nível de significância foi obtido pela
comparação entre as concentrações de citocinas dos grupos estudados, utilizando o programa
Graph-Pad Prism versão 6.0 (GraphPad Software, San Diego, CA, USA). Para à analise da
existência de correlação entre os grupos estudados, foi realizado pelo teste não paramétrico de
Kruskal-Wallis seguido de pós-teste de Dunn´s. Os dados foram expressos no formato
Boxplot (valor mínimo ao valor máximo, P25%-P75% e mediana). As diferenças
significativas foram estimadas pela mediana dos valores em cada grupo, sendo considerados
significativos àqueles onde P≤ 0,05.

53
2.10 CONSIDERAÇÃO ÉTICA
O estudo foi realizado com autorização do Comitê de Ética em Pesquisa da
Universidade Federal do Amapá (CEP /UNIFAP, protocolo 187404137/2013). Os candidatos
ao estudo foram incluídos, após receberem esclarecimento e manifestarem por escrito a sua
concordância em participar da pesquisa. Indivíduos positivos para malária vivax e parasitos
intestinais foram subsequentemente tratados por quimioterápicos recomendados pelo
Ministério da Saúde. Os participantes positivos para parasitos intestinais foram encaminhados
a equipe médica do hospital de Oiapoque para orientações e tratamento.

54
3 RESULTADOS
3.1 MANUSCRITO 1
Título: High Frequency of Enteroparasitoses in the Municipality of Oiapoque, Amapá
State, Brazil, on the Border With French Guiana
Autores: Rubens Alex de Oliveira Menezes, Margarete do Socorro Mendonça Gomes,
Anapaula Martins Mendes, Silvestre Rodrigues do Nascimento, Alvaro Augusto Ribeiro
DÀlmeida Couto, Mathieu Nacher, Martin Johannes Enk, Ricardo Luiz Dantas
Machado
Proposta do artigo: Respondendo aos objetivos específicos 1 a 3, este trabalho avaliou a
frequência de enteroparasitos numa população do município de Oiapoque no Estado do
Amapá e correlacionou com o nível de hemoglobina de cada indivíduo, tendo como propósito
produzir informações que possam subsidiar o planejamento, avaliação e intervenções voltadas
para a prevenção e controle das referidas infecções.
Resumo dos resultados: Nos indivíduos investigados, 58.6% (261/446) foram infectados por
algum tipo de parasito intestinal. Entre estes, 45,2% (118/161) estava infectado somente por
helmintos, 40,9% (107/261) com protozoários, e 13,8% (36/261) tinham infecções associadas.
Dentre os helmintos detectados o Ascaris lumbricoides 19,9% (52/261) foi o mais prevalente,
seguido de Entamoeba histolytica como protozoários mais frequentes 5% (13/261). A
pesquisa evidenciou uma correlação inversamente proporcional significativa entre o nível de
hemoglobina e a presença das parasitoses detectadas (coeficiente de contingência C=0.2195 e
p < 0.0001). O índice de enteroparasitoses é um reflexo claro da falta de saneamento da região
estudada, indicando um estado epidemiológico preocupante.
Status: Submetido – Revista Pan-Amazônica de Saúde (ISSN 2176-6223)

55
E-Mail de Submissão

56
High Frequency of Enteroparasitoses in the Municipality of Oiapoque, Amapá State,
Brazil, on the Border With French Guiana
Rubens Alex de Oliveira Menezes
Programa de Pós-Graduação Stricto Sensu em Biologia de Agentes Infecciosos e Parasitários,
Universidade Federal do Pará, Belém, Pará, Brasil, Departamento de Ciências biológicas e da
Saúde da Universidade Federal do Amapá, Macapá, Amapá, Brasil - email:
Margarete do Socorro Mendonça Gomes
Laboratório Central do Amapá, Macapá, Amapá, Brasil - email:
Anapaula Martins Mendes
Universidade Federal do Amapá/Campus Binacional-Oiapoque, Oiapoque, Amapá, Brasil -
email: [email protected]
Silvestre Rodrigues do Nascimento
Hospital Estadual do Oiapoque, Oiapoque, Amapá, Brasil - email: silvestre-
Alvaro Augusto Ribeiro DÀlmeida Couto
Laboratório Central do Amapá, Macapá, Amapá, Brasil - email: [email protected]
Mathieu Nacher
Université des Antilles et de la Guyane, Caiena, Guiana Francesa - email:
Martin Johannes Enk
Instituto Evandro Chagas, Secretaria de Vigilância Sanitária, Ministério da Saúde,
Ananindeua, Pará, Brasil - email: [email protected]
Ricardo Luiz Dantas Machado
Instituto Evandro Chagas, Secretaria de Vigilância Sanitária, Ministério da Saúde,
Ananindeua, Pará, Brasil, Universidade Federal Fluminense, Niterói, Rio de Janeiro, Brasil -
email: [email protected]
Corresponding author
Rubens Alex de Oliveira Menezes
Deptº Ciências Biológicas e da Saúde/Universidade Federal do Amapá. Rodovia Juscelino
Kubitscheck, Jardim Equatorial, 68903-419, Macapá, Amapá, Brasil.
Phone: 55 96 4009-2932; Mobile: 55 96 99151-5567
E-mail: [email protected]

57
ABSTRACT
Background: Enteroparasites represent a considerable proportion of infectious parasitic
diseases worldwide. This study evaluated the frequency of enteroparasites and the correlation
of enteroparasites with hemoglobin levels. Methods: A cross-sectional study was performed
in the municipality of Oiapoque in the state of Amapá in northern Brazil, which is located at
the western border of the Amazon region. Fecal samples collected over a one-year period
(2014/2015) were analyzed using direct methods and spontaneous sedimentation. Results:
The study included 446 individuals ranging in age from 7 to 61 years. Among the investigated
individuals, 58.6% (261/446) were infected with some type of enteroparasitas. Of these
infected individuals, 45.2% (118/161) were infected only by helminths, 40.9% (107/261)
were infected with protozoa, and 13.8% (36/261) had combined infections. Ascaris
lumbricoides (19.9%, 52/261) was the most commonly detected helminth, followed by
Entamoeba coli and Endolimax nana were the most frequent protozoans (17.2%, 45/261). The
study showed an inversely proportional correlation between the hemoglobin level and the
presence of detected parasitoses. Conclusions: In Oiapoque, enteroparasitic diseases are the
cause of anemia in the population. The high frequency of enteroparasites is a clear reflection
of the lack of sanitation in the studied region, indicating an epidemiological state of concern.
Keywords: Amazon Region, Enteroparasitoses, Hemoglobin, Oiapoque, Western Frontier
INTRODUCTION
Intestinal parasites represent a considerable segment of infectious parasitic diseases
worldwide, although the prevalence may vary depending on the characteristics of each
region1. Most enteroparasitoses are transmitted by the oral route via the ingestion of water or
food infected with parasitic structures. A high number of these parasitoses is associated with
places with poor sanitary hygiene and a lack of treated water and sewage, which facilitate the
dissemination of eggs, cysts and larvae2. Additionally, the lack of public policies aimed at
promoting changes in cultural habits and improving the socioeconomic conditions of the
population favor the establishment of this disease class3. Intestinal infections caused by
protozoa and helminths are estimated to afflict 3.5 billion people worldwide and cause
illnesses in approximately 450 million people, with risks of serious public health problems in
several countries, especially underdeveloped countries4.
In Latin American countries such as Brazil, 55.3% of the population is estimated to be
infected with enteroparasites5. Additionally, 10.9 million people in the country are infected
with soil-transmitted helminths (geohelminths), with the highest risk of transmission
occurring in the northern region of Brazil6. However, because no public policy requires
notification of enteroparasitoses, data are restricted to some scientific publications and cannot
truly address the prevalence and incidence of these infections in the different regions of this
country7. This lack of epidemiological data is more evident in the northern region of Brazil.
Because epidemiological studies addressing the occurrence of intestinal parasites in
populations in the State of Amapá are scarce, the responsible authorities cannot design and
implement effective control measures. Notably, the border areas between countries contain
international political boundaries and are often characterized by an intense population flow,
which creates a unique environment with effects on the incidence of diseases and the
availability of healthcare services8. This scenario is closely related to the local social
determinants of health.

58
In addition to the unique symptomatology of these parasites, infections also affect
nutritional status, growth and cognitive function. Moreover, environmental and
socioeconomic factors and hygiene habits considerably affect morbidity and mortality9.
Anemia is a relevant problem for individuals living in areas with limited resources and a
significant burden of enteroparasitoses10
. Although helminth infections affect nutritional
status, their impact on anemia is unclear11
. Some studies have indicated that moderate or high
intensity infections with the hookworm Ancylostoma cause anemia in men. Cases of Trichuris
trichiura enteroparasitoses are also associated with this clinical condition12
. This study
evaluated the frequency of enteroparasites in a population in the municipality of Oiapoque in
the state of Amapá, Brazil, and correlated infection with the hemoglobin level of each
individual to produce information in support of the planning and evaluation of interventions
aimed at the prevention and control of these infections.
MATERIALS AND METHODS
A cross-sectional study was performed from November 2014 to November 2015 in the
municipality of Oiapoque in the State of Amapá, which is located in the western border of the
Brazilian Amazon region. This municipality is located in the northernmost part of the state
and borders French Guiana to the north and the Atlantic Ocean to the east. In 2010, this region
had a population of 20,509 inhabitants according to the Brazilian Institute of Geography and
Statistics13
. After receiving a detailed explanation of the project and signing the informed consent
form, a questionnaire with socioepidemiological data was filled out, and two fecal samples
were collected in labeled containers without and with preservatives. The collected samples
were analyzed in the Laboratory of Clinical Analysis of the Municipal Hospital of Oiapoque.
The methods used for enteroparasite detection were direct examination and spontaneous
sedimentation. The results were presented as descriptive and inferential statistics in tables to indicate
the respective frequencies corresponding to the detected enteroparasites. The contingency
coefficient and Chi-square (χ2) tests were used to assess the combined proportions and to
evaluate the significance of the greatest contribution among age groups, genders and parasitic
species to the hemoglobin level. P-values equal to or lower than 5% (p < 0.05) were
considered significant. The study was approved by the Research Ethics Committee of the
Federal University of Amapá (protocol 187404137/2013). RESULTS
A total of 446 subjects were included in the study, as summarized in Table 1. The
participants were classified into the following age groups: 7 to 14 years old (5.9%, 26/446);
15 to 18 years old (27.8%, 124/446); 19 to 25 years (19.7%, 88/446) and ≥ 26 years old
(46.6%, 208/446). The highest parasitosis positivity rate was observed among individuals ≥
26 years of age (31.31%, 140/446), followed by individuals aged 15 to 18 years (13%,
58/446), 19-25 years (9.41%, 42/446) and 7 to 14 years (4.71%, 21/446). The majority of the
individuals included in the study were male (54.48%, 243/446). The Chi-square test (χ2)
revealed a significant (p = 0.0082) association between gender and the presence of intestinal
parasites.

59
Table 1. Characterization of helminths and protozoa detected in the study population in
Oiapoque (n = 446), Amapá state, Brazil, according to gender and age (years).
Oiapoque
Protozoa
(n = 107)
Helminths
(n = 118)
Both
(n = 36)
Negative
(n = 185) p-value
Gender (%)
Male 56 (23.0) 46 (19.0) 26 (11.0) 115 (47.0) 0.0082*
Female 51 (25.1) 72 (35.5) 10 (5.0) 70 (34.4)
Age group (%)
7-14** 8 (30.8) 10 (38.5) 3 (11.5) 5 (19.2) 0.0302*
15-18 20 (16.1) 34 (27.4) 4 (3.2) 66 (53.2) 0.0026*
19-25 20 (22.7) 16 (18.2) 6 (6.8) 46 (52.3) 0.0298*
≥26** 59 (28.4) 58 (27.9) 23 (11.0) 68 (32.7) 0.0006*
Source: Study data collection instrument.
*Chi-square test with significant results (p-value < 0.05).
**Meta-analysis test: Several combined proportions with statistical significance of a higher
contribution between the 7 to 14 year old (p = <0.0001) and ≥ 26 year old (p = <0.0001) age
groups. All age groups contributed to the presence of intestinal parasites. The non-parametric
Chi-square test (χ2) revealed a significant association between the 7 to 14 year old (p =
0.0302), 15 to 18 year old (p = 0.0026), 19 to 25 year old (p = 0.0298) and ≥ 26 year old age
groups (p = 0.0006). A meta-analysis test (several combined proportions) was performed to
assess the significance of the factor that had the greatest contribution among the age groups.
The results were significant between the 7 to 14 year old (p = <0.0001) and ≥ 26 year old (p =
<0.0001) age groups.
Among the individuals investigated, 58.6% (261/446) were infected with some type of
intestinal parasite. Of these, 45.2% (118/161) were infected only by helminths, 40.9%
(107/261) were infected with protozoa, and only 13.8% (36/261) of these individuals had
combined infections with these intestinal parasites. Ascaris lumbricoides (19.9%, 52/261) was
the most detected helminth, followed by Ancilostomídeos (7.7%, 20/261), Strongyloides
stercoralis (3.8%, 10/261) and Trichuris trichiura (2.3%, 6/261). Entamoeba coli and
Endolimax nana were the most frequent protozoans (17.2%, 45/261). Giardia intestinalis and
Entamoeba histolytica/Entamoeba dispar were detected in 3.1% (8/261) and 5% (13/261) of
individuals, respectively (Table 2). Table 2. Frequency of intestinal parasites in the studied population of Oiapoque (n = 446),
Amapá state, Brazil, from 2014 to 2015.
Oiapoque - Intestinal parasites (n) (%)
Protozoa 107 41
Entamoeba histolytica/Entamoeba dispar 13 5.0
Entamoeba coli + Endolimax nana 45 17.2
Entamoeba histolytica/Entamoeba dispar + Entamoeba coli 11 4.2
Entamoeba histolytica/Entamoeba dispar + Endolimax nana 1 0.4
Entamoeba histolytica/Entamoeba dispar + Endolimax nana +
Entamoeba coli
12
4.6
Entamoeba coli 8 3.1
Entamoeba coli + Giardia intestinalis 3 1.1
Giardia intestinalis + Endolimax nana + Entamoeba coli 5 1.9
Giardia intestinalis 8 3.1
Entamoeba histolytica/Entamoeba dispar + Endolimax nana +
Entamoeba coli + Giardia intestinalis
1 0.4
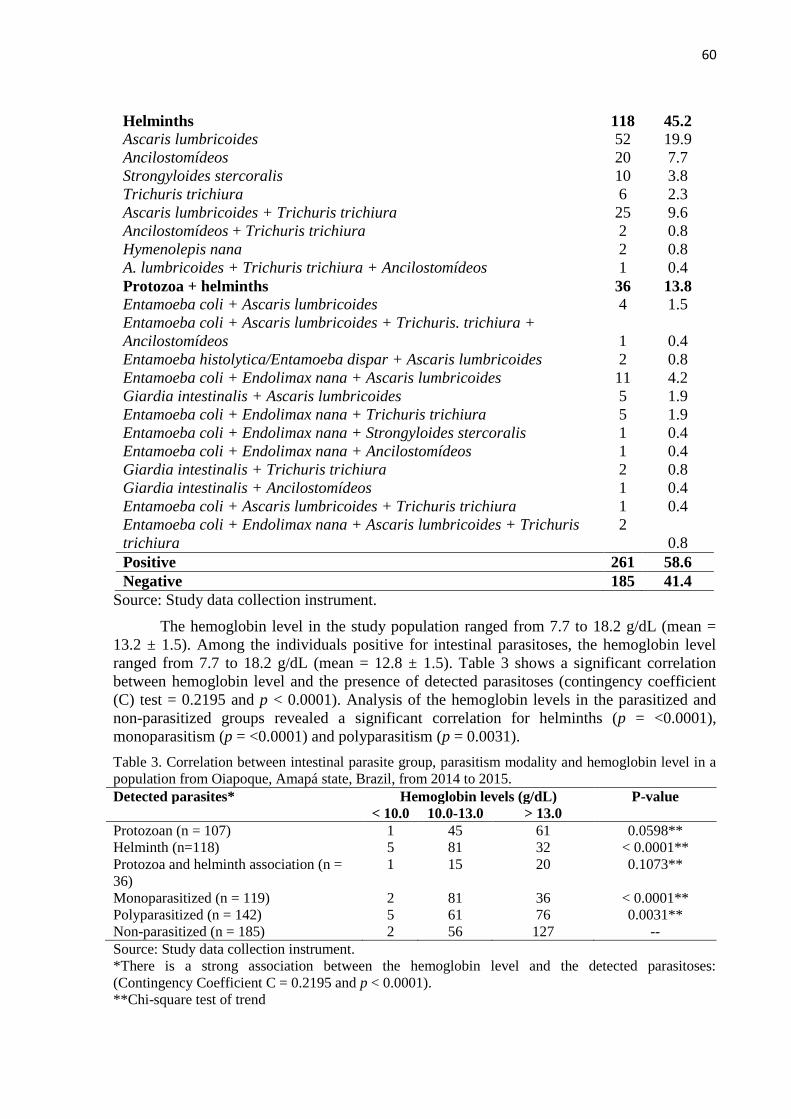
60
Helminths 118 45.2
Ascaris lumbricoides 52 19.9
Ancilostomídeos 20 7.7
Strongyloides stercoralis 10 3.8
Trichuris trichiura 6 2.3
Ascaris lumbricoides + Trichuris trichiura 25 9.6
Ancilostomídeos + Trichuris trichiura 2 0.8
Hymenolepis nana 2 0.8
A. lumbricoides + Trichuris trichiura + Ancilostomídeos 1 0.4
Protozoa + helminths 36 13.8
Entamoeba coli + Ascaris lumbricoides 4 1.5
Entamoeba coli + Ascaris lumbricoides + Trichuris. trichiura +
Ancilostomídeos
1 0.4
Entamoeba histolytica/Entamoeba dispar + Ascaris lumbricoides 2 0.8
Entamoeba coli + Endolimax nana + Ascaris lumbricoides 11 4.2
Giardia intestinalis + Ascaris lumbricoides 5 1.9
Entamoeba coli + Endolimax nana + Trichuris trichiura 5 1.9
Entamoeba coli + Endolimax nana + Strongyloides stercoralis 1 0.4
Entamoeba coli + Endolimax nana + Ancilostomídeos 1 0.4
Giardia intestinalis + Trichuris trichiura 2 0.8
Giardia intestinalis + Ancilostomídeos 1 0.4
Entamoeba coli + Ascaris lumbricoides + Trichuris trichiura 1 0.4
Entamoeba coli + Endolimax nana + Ascaris lumbricoides + Trichuris
trichiura
2
0.8
Positive 261 58.6
Negative 185 41.4
Source: Study data collection instrument. The hemoglobin level in the study population ranged from 7.7 to 18.2 g/dL (mean =
13.2 ± 1.5). Among the individuals positive for intestinal parasitoses, the hemoglobin level
ranged from 7.7 to 18.2 g/dL (mean = 12.8 ± 1.5). Table 3 shows a significant correlation
between hemoglobin level and the presence of detected parasitoses (contingency coefficient
(C) test = 0.2195 and p < 0.0001). Analysis of the hemoglobin levels in the parasitized and
non-parasitized groups revealed a significant correlation for helminths (p = <0.0001),
monoparasitism (p = <0.0001) and polyparasitism (p = 0.0031). Table 3. Correlation between intestinal parasite group, parasitism modality and hemoglobin level in a
population from Oiapoque, Amapá state, Brazil, from 2014 to 2015.
Detected parasites* Hemoglobin levels (g/dL) P-value
< 10.0 10.0-13.0 > 13.0
Protozoan (n = 107) 1 45 61 0.0598**
Helminth (n=118) 5 81 32 < 0.0001**
Protozoa and helminth association (n =
36)
1 15 20 0.1073**
Monoparasitized (n = 119) 2 81 36 < 0.0001**
Polyparasitized (n = 142) 5 61 76 0.0031**
Non-parasitized (n = 185) 2 56 127 --
Source: Study data collection instrument.
*There is a strong association between the hemoglobin level and the detected parasitoses:
(Contingency Coefficient C = 0.2195 and p < 0.0001).
**Chi-square test of trend

61
DISCUSSION
Due to the great diversity in socioeconomic and geographical characteristics among
Brazilian municipalities, enteroparasitic infections are endemic in various areas of the country
and thus constitute a basic and relevant public health problem14
. Currently, the prevalence of
intestinal parasites in the municipality of Oiapoque, which is an area bordering French
Guiana, is underestimated due to the lack of records in the municipality, which prevents the
development of specific and well-targeted control measures for this population. These issues
indicate the relevance and importance of discussing the behavior of these diseases in the
municipality and the health care of border populations and highlight their particularities and
geographical aspects related to the main constraints and determinants of health in border
areas. Variations in the frequencies of diseases between men and women may result from
physiological, intrinsic or behavioral differences and distributions based on influences of the
population structure15
. This study found an infestation of enteroparasites with a predominance
among males (Table 1), which might be related to work activities that have contact with the
soil, such as illegal gold mining and agriculture (the predominant activities in the region),
given the large percentage of geohelminths detected. The data presented here indicate that individuals from all age groups are affected by
intestinal parasitoses. Additionally, a greater contribution was observed for the 7 to 14 year
old and ≥ 26 year old age groups (meta-analysis: several combined proportions p = 0.0001)
(Table 1). Factors such as differences in hygiene habits and the resistance of individuals to
seeking health centers due to cultural and social issues may influence this process16
. In this study, a predominance of polyparasitized individuals was observed (54.4%,
142/261) (Table 3). This finding may be associated with the high frequency with which the
host comes into contact with the medium contaminated with different species or may be
related to the degree of host immunocompetence2. The frequent finding of polyparasitism is
due to similarities in life cycles given the elimination of large numbers of eggs and/or cysts
and their resistance in the environment, which act as an important focus for the maintenance
and transmission of these pathogens14
. Therefore, in this locality, intervention and control
measures for these parasites in soil and water are necessary to break this chain of
transmission. The prevalence of enteroparasites was 58.6% (261/446) in the study population (Table
2). Studies in other regions of Brazil with geographic and socioeconomic characteristics
different from the municipality of Oiapoque indicate variation in positivity between 7.4% in
the state of Santa Catarina in the southern region of Brazil17
. to 42.9% in the interior of the
state of Bahia in northeast Brazil18
. Although the prevalence is usually lower in urban
populations in the country, where there is usually a better quality of sanitation and hygienic
sanitary conditions, the results are still worrying. In the municipality of Oiapoque, the
precarious socioeconomic and sanitary conditions favor the transmission of enteroparasitoses.
The location of this area on the border with French Guiana, where the migratory flow related
to illegal mining is intense, is also a factor that may influence the results. Our study showed that Ascaris lumbricoides (19.9%, 52/261) was the most detected
helminth, but other geohelminths were detected at low frequencies, including Ancilostomídeos
(7.7%, 20/261), Strongyloides stercoralis (3.8, 10/261) and Trichuris trichiura (2.3%, 6/261).
Regarding the protozoa, Entamoeba histolytica/Entamoeba dispar (5%, 13/261) was observed
in the investigated individuals, but combined infections with non-pathogenic amoebas, such
as Entamoeba coli and Endolimax nana, were the most frequent (17.2% 45/261).
Additionally, Giardia intestinalis (3.1%, 8/261) was detected at a reduced frequency (Table

62
2). Several factors may explain these findings. First, the helminths detected can be transmitted
by waterborne routes, such as A. lumbricoides and Trichuris trichiura. Thus, consumption of
raw food and contaminated water may be a common practice among this population. Additionally, geohelminths known to actively penetrate skin (Strongyloides stercoralis
and Ancilostomídeos) were detected, which suggests that habits such as walking barefoot or
handling soil without hand protection (i.e., agriculture and illegal mining, the predominant
activities in the region) may be important factors in the transmission of these parasites in the
population. Second, the absence of a population treatment policy and the lack of treatment
plans for parasitized individuals (after therapeutic treatment) may stimulate the maintenance
of the transmission chain. Finally, the lack of soil and water research and decontamination for
these parasitoses complements the characteristics of maintenance of these helminths in the
municipality. Among the different ameba species, E. histolytica/Entamoeba dispar is the only
species considered invasive, especially in tropical regions and communities living in
inadequate sanitary conditions19
. Although many individuals are contaminated by commensal
amoebae, most infections are asymptomatic20,21
. The results show low numbers of this
parasite, which indicates that the parasite may not be endemic in the region. However, the
detection of commensal amoebae as evidenced by helminth infections indicates that the
population ingests water or food contaminated with fecal waste and therefore is at high risk of
contamination by this pathogenic amoeba. We must emphasize the importance of the diagnosis and description of these
commensals for the planning of preventive measures and avoidance of infection due to oro-
fecal contamination with pathogenic amoebas22,23
. Another important protozoan pathogen
infection found was giardiasis. The decrease in the rate of giardiasis usually increases with the
age group because successive contact with the parasite increases host immunity19,24
. This
parasite is often found in collective environments because transmission occurs where direct
person-person contact is habitual. However, in the municipality of Oiapoque, the analysis of
the samples revealed a low frequency of giardiasis, which may be related to host factors, such
as immune defense mechanisms, and the biological characteristics of the parasite, whose
elimination is intermittent. At least three alternate examinations are required for each patient
to obtain an accurate evaluation of this parasite. Another important factor in the drastic reduction of helminthoses and protozooses is
adequate drug treatments in conjunction with the establishment of health education programs,
personal hygiene, early treatment of symptomatic and asymptomatic infected individuals,
food storage care, water treatment and footwear use; these measures are key for health
promotion and disease prevention25
. When some form of intestinal parasite is identified, the
awareness of the infected individual/population is of great importance in addition to specific
drug treatment for the avoidance of reinfection26
. The infected individual is treated with
polyvalent antiparasitic agents. At least two therapeutic schemes should be performed, and the
soil should be treated to break the chain of transmission. Another important finding was the association between anemia and intestinal
parasitoses, which represented a serious public health problem. The presence of some
parasites usually determines the onset of anemia, and clinical manifestations are usually
proportional to the parasitic load of the individual. The study showed an inversely
proportional association between hemoglobin level and detected parasitoses (Contingency
Coefficient C = 0.2195 and p < 0.0001) (Table 3). However, the development of anemia in
parasitized individuals is multifactorial, and factors such as nutritional status, parasite species
and load, duration of infection, body iron store, iron intake and bioavailability and

63
physiological iron requirements are complicating factors of this clinical picture27,28
. No
significant differences in these factors were found between groups in this study. Among the
geohelminthiases detected in this study, ancylostomiasis had a marked relationship with
anemia. Despite this close relationship, our results did not support this finding, probably due
to the low circulation of this parasite or its non-endemic profile in the region. Thus, new
studies are necessary to determine the prevalence of this helminthiasis in the municipality. There is a gap in the literature regarding the risk of anemia development in individuals
infected with multiple helminth species but at low numbers. This low-intensity polyparasitism
may be related to the increased possibility of anemia. A recent study demonstrated that
polyparasitic infections were associated with a 5 to 8-fold increased risk of developing
anemia27
. Polyparasitism is a common condition in humans; however, little is known about
the interaction between parasites and their impact on the host's immune system and other
diseases28
. Investigations of the Th1/Th2 response patterns are of great relevance for
understanding host defense against infectious/parasitic diseases. The implications of
concomitant infections in humans have been evaluated mainly in relation to the effects of
intestinal helminth infections on Plasmodium falciparum malaria in non-Brazilian endemic
areas with conflicting results. In Brazil, studies with patients from the state of Rondônia
coinfected with P. vivax and intestinal parasites did not find a direct relationship with
anemia29,30
. Although coinfection with Plasmodium and helminths is prevalent in tropical
countries, the effect of their interactions remains unclear. Some studies have reported that
helminth-infected individuals are susceptible to infection by Plasmodium, causing an increase
in circulating gametocytes, reduced hemoglobin levels and suppression of acute clinical
manifestations and thus an increased risk of malaria11, 12
. An increased incidence and
prevalence of malaria can affect the development of mixed P. vivax and P. falciparum
infections. Additionally, parasite diversity may be higher in helminth-infected patients
31. Malaria
is an endemic disease in the municipality of Oiapoque, and almost half of the cases are from
French Guiana. This association may be relevant for the establishment of the anemic status in
this population. The municipality of Oiapoque is in an area bordering French Guiana, and
several diseases in addition to malaria, such as AIDS, tuberculosis and leishmaniasis, are
important public health problems in the region. New studies need to be carried out to clarify
the influence of intestinal parasitoses on coinfected individuals to understand the transmission
dynamics of these diseases. These studies will produce information that can support the
planning and evaluation of interventions aimed at the prevention and control of these
infections.
CONCLUSIONS
The frequency of intestinal parasitoses is high in the municipality of Oiapoque, and an
effective model of primary healthcare adapted to the region has been suggested to combat
these adverse conditions and to promote the implementation and success of public policies
supporting the universalization of access to education and sanitation and health services.
Many challenges will be faced given the peculiarities of the region. Improvement of the care
model emphasizing preventive health education and community participation conducted by
local health authorities and municipal managers aims at better health care for the population
living in the state of Amapá. Acknowledgments: We acknowledge and thank all those who accepted to participate in the
present study, as well as to the professionals of the Hospital of the municipality of Oiapoque,

64
in which the activities were developed in particular to the employees: Flaviano Feitosa and
Leudilene Marques for assistance in sample collection and technical support. Funding: This work was supported by the Research Program for the Unified Health System
(PPSUS): Shared Health Management with the Foundation for Research Support of the State
of Amapá [001/2013 EPF 00007631]. Competing interests: None declared. Ethical approval: Approved by the Research Ethics Committee of the Federal University of
Amapá (protocol 187404137/2013).
REFERENCES
1. Lemus-Espinoza D, Maniscalchi MT, Kiriakos D, Pacheco F, Aponte C, Villarroel O et al.
Enteroparasitosis en ninos menores de 12 anos del estado anzoategui, venezuela. Rev. Soc.
Ven. Microbiol. 2012 Abr; 32: 139-47.
2. Bellin M, Grazziotin NA. Prevalência de parasitos intestinais no município de
Sanandauva/RS. Newslab 2011; 18(104): 116-22.
3. Lima Junior OA, Kaiser J, Catisti, R. highoccurrenceof giardiasis in children living on a
’landless farm workers’ settlement in araras, são paulo, brazil. Rev. Inst. Med. Trop 2013
May-June; 55(3): 185-88.
4. Nxasana N, Baba K, Bhat VG, Vasaikar SD. Prevalence of intestinal parasites in primary
school children of mthatha, eastern cape province, south africa. Annals of Medical and Health
sciences Research 2013 Oct-Dec; 3(4): 511-16.
5. Pedraza DF, Queiroz D, Sales MC. Doenças infecciosas em crianças pré-escolares
brasileiras assistidas em creches. Cienc. Saúde Coletiva 2014 Jun; 19(2): 511-28.
6. Chammartin F, Guimaraes LH, Scholte RGC et al. Spatio-temporal distribution of soil-
transmitted helminth infection in Brazil. Parasite & Vectors 2014 Sep; 7(440): 2-16. doi:
10.1186/1756-3305-7-440.
7. Costa ACN, Borges BC, Costa AV, Ramos MF, Gomes JM, Gomes JM et al.
Levantamento de acometidos por enteroparasitoses de acordo com a idade e sexo e sua
relação com o meio onde está inserido o PSF Prado na cidade de Paracatu – MG. Rev Patol
Trop. 2012 Abr.-Jun; 41(2): 203-24.
8. Stefani A, Hanf M, Nacher M, Girod R, Carme B. Environmental, entomological,
socioeconomic and behavioural risk factors for malaria attacks in Amerindian children of
Camopi, French Guiana. Malar J. 2011 Aug; 10 (246). doi: 10.1186/1475-2875-10-246.
9. Furini AAC, Lima TAM, Rodrigues LV, Faschina F, Galao EG, Satin MS et al. Prevalence
of intestinal parasitosis in a population of children of a daycare in Brazil. Parasitaria 2015;
21(1): 1-5.

65
10. Cojulun AC, Bustinduy AL, Sutherland LJ, Mungai PL, Mutuku F, Muchiri E, el al.
Anemia among Children Exposed to Polyparasitism in Coastal Kenya. The American Journal
of Tropical Medicine and Hygiene 2015; 93 (5): 1099-05.
11. Nacher M. Helminth-infected patients with malaria: a low profile transmission hub? Malar
J. 2012; 11: 376. doi: 10.1186/1475-2875-11-376.
12. Nacher M. Interactions between worms and malaria: good worms or bad worms?. Malar J.
2011; 10: 259. doi: 10.1186/1475-2875-10-259.
13. Instituto Brasileiro de Geografia e Estatística. Censo Demográfico Brasileiro.
Características da população e dos domicílios: Resultados do Universo. Amapá: IBGE,
Brasília, 2010 [Citado 2017 Feb 1]. Disponivel em: http://cod.ibge.gov.br/4AJ
14. Belo VS, Oliveira RB, Fernandes PC, Nascimento BWL, Fernandes FV, Castro CLF, et
al. Factors associated with intestinal parasitosis in a population of children and adolescents.
Rev Paul Pediatr. 2012; 30 (2): 195-01.
15. Menezes RAO, Gomes MSM, Barbosa FHF, Brito GCM, Junior AAP, Couto AARD.
Parasitas intestinais na população residente em áreas úmidas em Macapá, Amapá, Brasil.
Bioterra 2013; 13(2): 10-8.
16. Espelage W, Heiden M, Stark K, Alpers K. Characteristics and risk factors for
symptomatic Giardia lamblia infections in Germany. BMC Public Health 2010; 10: 01-09.
17. Seger J, Souza WM, Marangoni JCF, Maschio VJ, Chielli EO. Prevalência de parasitas
intestinais na população do Bairro Salete, município de São Miguel do Oeste, SC. Unoesc &
Ciência – ACBS. 2010; 1(1): 53-6.
18. Matos MA, Cruz ZV. Prevalência de parasitoses intestinais no município de Ibiassucê -
Bahia. Rev Educ Meio Amb Saú. 2012; 5(1): 64-1.
19. Castro TG, Silva-Nunes M, Conde WL, Muniz PT, Cardoso MA. Anemia e deficiência de
ferro em pré-escolares da Amazônia Ocidental brasileira: prevalência e fatores associados.
Cad. Saúde Pública 2011; 27(1): 131-42.
20. Gil FF, Busatti HGNO, Cruz VL, Santos JFG, Gomes MA. High prevalence of
enteroparasitosis in urban slums of Belo Horizonte-Brazil. Presence of enteroparasites as a
risk factor in the family group. Pathogens and Global Health 2013; 107(6): 320-24.
21. Yihenew G, Adamu H, Petros B. The Impact of Cooperative Social Organization on
Reducing the Prevalence of Malaria and Intestinal Parasite Infections in Awramba, a Rural
Community in South Gondar, Ethiopia. Interdisciplinary Perspectives on Infectious Diseases
2014; ID 378780:1-6.
22. Inoue AP, Nigro S, Castilho VLP. Frequência de parasitas intestinais em um hospital
terciário com atendimento SUS. Arq Med Hosp Fac Cienc Med Santa Casa São Paulo 2015;
60(1): 7-11.

66
23. Cavasini CE, Cimerman S, Barbosa DRL, Silva MCME, Furini AAC, Machado RL.
Agreement and disagreement on intestinal commensal microorganisms. Rev Panam Infec.
2015; 17(1): 26-9.
24. Rezende HHA, Avelar JB, Figueiredo Junior J, Castro AM. Associação de
enteroparasitoses com quadros anêmicos e eosinofilia em moradores do município de caldas
novas - Goiás nos anos de 2007 a 2011. Newslab 2014; 124(1): 126-31.
25. Mendes AMN, Silva ACC, Koppe EC, Filgueiras LA. Incidência de ascaridíase em
comunidade quilombola de Cachoeiro de Itapemirim, Espirito Santo, Brasil. Boletim
Informativo Geum 2016; 7(1): 28-3.
26. Silva JC, Furtado LFV, Ferro TC, Bezerra KC, Borges EP, Melo ACFL. Parasitismo por
Ascaris lumbricoides e seus aspectos epidemiológicos em crianças do Estado do Maranhão.
Rev Soc Bras Med Trop. 2011; 44(1): 100-02.
27. Miotto JE, Caro DAS, Barros MF, Rego BEF, Santos FC, Macagnan R. Diagnóstico
laboratorial de enteroparasitoses e anemia e sua possível associação com eosinofilia em
crianças em idade escolar em Ubiratã –PR. Biosaúde 2014; 16(2): 52-2.
28. Cabral EA, Morais AMB, Gonçalves JS. Correlação entre a prevalência de anemias
associadas à enteroparasitoses: uma revisão de literatura. Temas em saúde. 2016; 16 (3):
2447-2131.
29. Sánchez-Arcila JC, Perce-da-Silva DS, Vasconcelos MPA, Rodrigues-da-Silva RN,
Pereira VA, Aprígio CJL. et al. Intestinal Parasites Coinfection does not Alter Plasma
Cytokines Profile Elicited in Acute Malaria in Subjects from Endemic Area of Brazil.
Mediators of Inflammation 2014; ID 857245: 1-12.
30. Sánchez-Arcila JC, França MM, Pereira VA, Vasconcelos MPA, Têva A, Perce-da-Silva
DS et al. The influence of intestinal parasites on Plasmodium vivax-specific antibody
responses to MSP-119 and AMA-1 in rural populations of the Brazilian Amazon. Malar J.
2015; 14: 442. doi: 10.1186/s12936-015-0978-7.
31. Chaorattanakawee S, Nataleng O, Hananantachai H, Nacher M, Brockman A, Nosten F, et
al. Trichuris trichiura infection is associated with the multiplicity of Plasmodium falciparum
infections, in Thailand. Ann Trop Med Parasitol 2003; 97(2): 199-02. doi:
10.1179/000349803125002968.

67
3.2 MANUSCRITO 2
Enteroparasite and vivax malaria co-infection on the Brazil-French Guiana border:
Epidemiological, haematological and immunological aspects
Autores: Rubens Alex de Oliveira Menezes, Margarete do Socorro Mendonça Gomes,
Anapaula Martins Mendes, Álvaro Augusto Ribeiro D’Almeida Couto, Mathieu Nacher,
Tamirys Pina Simão, Aline Collares Pinheiro de Sousa, Andrea Regina de Souza
Baptista, Maria Izabel de Jesus, Martin Johannes Enk, Maristela Gomes da Cunha,
Ricardo Luiz Dantas Machado
Proposta do artigo: Respondendo aos objetivos específicos 4 a 7, este artigo avaliou a
influência da coinfecção das enteroparasitoses na malária causada pelo Plasmodium vivax em
uma área fronteiriça entre Brasil e Guiana Francesa da Amazônia brasileira. Este é o primeiro
estudo que oferece um perfil imune da interação parasita-hospedeiro na malária vivax em
Oiapoque-Amapá. Tais resultados poderão contribuir para o conhecimento sobre a patogênese
dessas coinfecções e no delineamento de estratégias antiparasitárias.
Resumo dos resultados: Foram incluídos no estudo 441 indivíduos agrupados de acordo com
o seu estado de infecção: [Maláricos (M) 6,9% (30/441)], [Coinfectados (CI) 26,5%
(117/441)], [Enteroparasitas (E) 32,4% (143/441)] e [Controle endêmico (CE) 34,2%
(151/441)]. O gênero masculino prevaleceu entre os grupos (M) 77% (23/30) e (CI) 60%
(70/117). Houve diferença entre os níveis de hemoglobina nos diferentes grupos estudados:
[CE e E (p < 0.01)], [CE e CI (p < 0.01)], [E e M (p < 0.01)] e [CI e M (p < 0.01)]. A anemia
foi expressa em percentuais entre indivíduos [CI e CE (p < 0.05)]. Foi observado diferenças
na parasitemia entre os grupos [CI e M (p < 0.05)]. Com relação aos anticorpos contra
PvMSP-119 foram detectados em 51,2% (226/441) da população. As analíses dos níveis de
citocinas revelaram uma grande variação nas concentrações, com pistas imunológicas na
alteração da resposta imune expressa pelas citocinas TNF-α e IL-10 aumentados para os
indivíduos do grupo coinfectado. Os resultados apontam a coinfecção Malária e
Enteroparasitoses um aspecto real no município do Oiapoque. Adicionalmente, a coinfecção
não esteve associada com a redução de malária (incidência, prevalência ou redução da
parasitemia).
Status: Submetido – PLOS Neglected Tropical Diseases (ISSN: 1935-2735)

68
E-Mail de Submissão

69
Enteroparasite and vivax malaria co-infection on the Brazil-French Guiana border:
Epidemiological, haematological and immunological aspects
Short title: Enteroparasite and vivax malaria co-infection on the Amazon border
Rubens Alex de Oliveira Menezes1,2*
, Margarete do Socorro Mendonça Gomes3, Anapaula
Martins Mendes4, Álvaro Augusto Ribeiro D’ Almeida Couto
3, Mathieu Nacher
5, Tamirys
Pina Simão6,7
, Aline Collares Pinheiro de Sousa7, Andrea Regina de Souza Baptista
8, Maria
Izabel de Jesus7, Martin Johannes Enk
7, Maristela Gomes da Cunha
1, Ricardo Luiz Dantas
Machado1,7,8
1Postgraduate Program in the Biology of Infectious and Parasitic Agents, Federal University
of Pará (UFP), Belém, Pará State, Brazil
2 Federal University of Amapá (UNIFAP), Macapa, Amapá State, Brazil
3 Amapá State Secretary of Health (SESA)/Amapá Central Laboratory, Amapá state, Brazil
4 UNIFAP/Oiapoque Binational Campus, Oiapoque, Amapá State, Brazil
5 Université des Antilles et de la Guyane Caiena, French Guiana
6 Postgraduate Program in Neuroscience and Cell Biology, UFP, Belém, Pará State, Brazil
7 Evandro Chagas Institute/Brazilian Secretariat of Health Surveillance (SVS)/Brazilian
Ministry of Health (MS), Ananindeua, Pará State, Brazil
8 Fluminense Federal University, Niterói, Rio de Janeiro State, Brazil
* Corresponding author
E-mail: [email protected] (RAOM)

70
Abstract
Malaria-enteroparasitic co-infections are known for their endemicity. Although they are
prevalent, little is known about their epidemiology and effect on the immune response. This
study evaluated the effect of enteroparasite co-infections with malaria caused by Plasmodium
vivax in a border area between Brazil and French Guiana.
Methods: The cross sectional study took place in Oiapoque, a municipality of Amapá, on the
Amazon border. Malaria was diagnosed using thick blood smears, haemoglobin dosage by an
automated method and coproparasitology by the Lutz e Faust methods. The anti-PvMSP-119
IgG antibodies in the plasma were evaluated using ELISA and Th1 (IFN-γ, TNF-α and IL-2),
and Th2 (IL-4, IL-5 and IL-10) cytokine counts were performed by flow cytometry. The
participants were grouped into those that were monoinfected with vivax malaria (M), vivax
malaria-enteroparasite co-infected (CI), monoinfected with enteroparasite (E) and endemic
controls (EC), who were negative for both diseases.
Main findings: 441 individuals were included and grouped according to their infection status:
[M 6.9% (30/441)], [Cl 26.5% (117/441)], [E 32.4% (143/441)] and [EC 34.2% (151/441)].
Males prevailed among the (M) 77% (23/30) and (CI) 60% (70/117) groups. There was a
difference in haemoglobin levels among the different groups under study for [EC-E], [EC-Cl],
[E-M] and [Cl-M], with (p < 0.01). Anaemia was expressed as a percentage between
individuals [CI-EC (p < 0.05)]. In terms of parasitaemia, there were differences for the groups
[CI-M (p < 0.05)]. Anti-PvMSP-119 antibodies were detected in 51.2% (226/441) of the
population. The cytokine level analysis revealed a large variation in the TNF-α and IL-10
concentrations, being high for individuals in the co-infected group.
Conclusion: Co-infection was not associated with malaria reduction (the incidence,
prevalence or reduction of parasitaemia). Studies of this nature that offer immunological clues
about immune response changes are essential for diagnostic and preventive strategies.
Keywords: malaria, enteroparasites, co-infection, cytokines, Brazilian Amazon.

71
Author summary
Tropical regions are known for the endemicity of malaria-enteroparasite co-infections.
Although they are prevalent, little is known about these interactions and how simultaneous
infection affects the epidemiology and/or pathogenesis of each morbidity. Oiapoque, a
municipality in the state of Amapá, northern Brazil, on the western border of the Amazon,
was chosen to determine the epidemiology and effect of these co-infections. To understand
the development of anti-parasitic strategies, it is fundamental to uncover the mechanisms that
govern these immune responses. The authors demonstrate the differences between the
antibody profiles reported in the study and draw attention to the importance of performing
immunoepidemiological studies in different malaria-endemic areas, where the transmission
intensities and human genetic backgrounds are very distinct. Establishing P. vivax's complex
geographical pattern is important, both for diversity evaluations that help select candidate
antigens for vaccines and for formulating and structuring surveillance measures to control
malaria. The results show that malaria-enteroparasite co-infection is a reality in the
municipality of Oiapoque. This study evaluates the possibility that populations infected with
intestinal parasites can affect malarial transmission dynamics, which is essential knowledge
for diagnostic and preventive strategies.

72
Introduction
In Brazil, over 99% of reported malaria cases occur in the Amazon region [1,2]. The
cases recorded outside the Amazon region are imported from the Amazon states or from other
countries. There are rare indigenous accounts that have been restricted to the Atlantic Forest
in the south-eastern part of the country [3]. Socioeconomic and environmental conditions
favour disease transmission and vector survival [4]. In recent years, Plasmodium vivax has
been the most prevalent species in the country, accounting for approximately 80% of
episodes, while Plasmodium falciparum is responsible for approximately 20%, and
Plasmodium malariae is rarely detected. Historically, severe clinical P. vivax cases have been
rarely reported [5]. However, recent studies emphasize the association of this species with
clinical complications and fatal cases, which is a cause for public health concern [5-7].
Currently, it is estimated that more than a third of the world's tropical and sub-tropical
population is infected with one or more enteroparasites [8]. Human infections with these
organisms remain prevalent in countries where the malaria parasite is also endemic [9,10],
and they are one of the most important public health issues in the world [11,12]. Malaria and
intestinal parasite co-infections are widespread, and both have similar geographical
distributions and overlap in developing countries [9,13].
Although Plasmodium and helminth co-infections are prevalent in tropical countries,
the effect of their interactions remains unclear [14]. Some studies report that individuals who
are infected with helminths are susceptible to Plasmodium infection [15,16], resulting in an
increase in circulating gametocytes [17], a reduction in haemoglobin levels [18], the
suppression of acute clinical manifestations [19] and an increased risk of malaria transmission
[17,20,21]. It is known that increased incidence and prevalence of malaria can affect the
development of P. vivax and P. falciparum-mixed infections and that parasite diversity may
be greater in patients who are infected with helminths [22,23]. The evolutionary implications
of co-infection may also extend to the reproduction of helminths, which are interested in
protecting their hosts to survive and reproduce [24].
These associations between helminthiases and malaria are well documented in
malaria-endemic regions of Africa and Asia, especially where malaria is caused by P.
falciparum. However, the protective effect against malaria from P. vivax has also been
observed in two studies outside Brazil [18,19]. In the Brazilian Amazon region, intestinal
helminthiases were associated with protection against reduced haemoglobin levels during P.
vivax malaria episodes in a population of children in the city of Manaus, Amazonas State [25].
In addition, in the municipality of Porto Velho in the State of Rondônia, co-infection with
enteroparasites did not affect the immune response pattern to vivax malaria [26,27], but
differences were observed between the haemoglobin levels of malaria patients and individuals
who were not infected by enteroparasites [26].
The municipality of Oiapoque is in the French Guiana border region, and it has a close
relationship with local social health determinants. It also has an intense population flow. The
objective of this study was to evaluate the effect of enteroparasite co-infection on malaria
caused by Plasmodium vivax in this region of the Brazilian Amazon.
Methods
Study area
A cross-sectional study was conducted in the municipality of Oiapoque, Amapá State,
in northern Brazil, on the western border of the Amazon region. It has an Annual Parasite
Index that designates it as at high risk of malaria transmission, and it is located in the northern
part of the state of Amapá. Due to the border with the municipality of Saint Georges de
l’Oyapock in French Guiana, there is an intense population flow related to trade, mining,
tourism and social life, which also facilitates the spread of communicable diseases.
Inclusion criteria

73
Participants were required to meet the following inclusion criteria: (1) signed consent
form, (2) being native to the study area, (3) being over seven years of age, (4) consenting to
blood collection and (5) providing a faecal sample.
Sample
A total of 441 participants formed the study population, with the sample collection
taking place over one year between the months of November 2014 and November 2015. Data
collection was performed by passive detection in the municipality's Basic Health Units,
following phenotypic diagnoses using thick blood smears and prior to treatment. For the
uninfected group, samples were also collected in the same municipality so that the
participating individuals were from a similar background exposure conditions. After a
parasitological evaluation, the subjects were divided into the following four groups: 1)
patients with an intestinal parasite and malaria co-infection (n = 117); 2) patients with malaria
who were negative for intestinal parasites (n = 30); 3) patients without malaria who were
positive for enteroparasites (n = 143); and 4) patients who were negative for both (n = 151).
Epidemiological data such as the age, gender and number of previous episodes of malaria
were obtained during interviews and from medical records.
Ethical considerations
This study was authorised by the Federal University of Amapá (UNIFAP) Research
Ethics Committee (CEP, protocol 187404137/2013). The study candidates were included after
they received a briefing and expressed their consent to participate in writing. Donors who
were positive for malaria and enteroparasites were subsequently treated with
chemotherapeutic agents as recommended by the Ministry of Health. Participants who were
positive for enteroparasites were referred to the hospital's medical team for advice and
treatment.
Collection and laboratory analysis
Blood collection
Blood samples were collected by venipuncture. Ten millilitres of venous blood was
collected from each patient. Four millilitres was dispensed into a tube containing EDTA
(ethylenediamine tetraacetic acid) (Beckton & Dickson, USA) to perform haematological
analysis and to prepare thick blood smear slides, and 6 mL was dispensed into a tube (Becton
& Dickson, USA) with no anticoagulant for immunological analysis. After this collection,
each slide was maintained at room temperature to dry the blood drop. The slides were
subsequently stained and analyzed using light microscopy (Nikon, Japan) according to the
protocol described by the World Health Organization.
Malaria diagnosis
The slides were prepared by following the Walker technique (methylene blue and
Giemsa) [28] and evaluated by a local microscopist from the municipality’s malaria
diagnostic station. Direct parasite counting was performed by estimating the parasitaemia
using a semi-quantitative evaluation, which recorded the parasitaemia interval per µL of
blood from 200 fields. The number of counted parasites was multiplied by 5 [29]. These
results were subsequently confirmed in the Amapá Border Laboratory (LAFRON/AP) and by
the endemic quality control section of the Amapa Central Laboratory (LACEN/AP).
For the sexual form count (gametocytaemia), the thick blood slides were reviewed for
gametocyte quantification in 100 leukocytes. The conversion to gametocytes/μL was
performed by calculation using a reference value of 8000 leukocytes/mm3, and the
gametocytes were counted directly on the slide (Nº gametocytes X Nº of leukocytes

74
(8000)/100 = gametocytes per µL). This procedure is similar to the diagnostic methods
recommended by the Clinical and Laboratory Standards Institute (CLSI) guidelines of the
United States Centers for Disease Control (CDC) and the World Health Organization (WHO)
guidelines. The count was subsequently confirmed in LAFRON/AP and sent to LACEN/AP's
endemic quality control section.
Haemoglobin dosage
The haemoglobin concentration was measured in venous blood using the Oiapoque
Hospital's automated equipment (Mindray-BC-3000plus). Anemia was defined with
haemoglobin reference values. The evaluated haematological parameters were the total
number of erythrocytes (RBC; reference range: males 4.5 - 6.5 x 106/µL, females 3.9 - 5.6 x
106/µL) and the haemoglobin (Hb; males ≥ 13 g/dL, females ≥ 12 g/dL). Individuals were
considered anaemic when their haemoglobin levels were ≤ 13 g/dL of blood for males and ≤
12 g/dL of blood for females.
Evaluation of anti-PvMSP-119 IgG antibodies
The evaluation of the total anti-MSP-119 IgG antibodies was performed at the Federal
University of Pará (UFP) according to Wedge et al.'s (2001) protocol. The recombinant
protein (His6-MSP-119) of the MSP-1 from P. vivax (Belém strain) was expressed in
Escherichia coli. The 96 wells of the Enzyme-Linked Immunosorbent Assay (ELISA) plates
(Costar, Corning Inc. NY, USA) were sensitized with 50 μL of His-MSP-119 protein (4
μg/mL) diluted in 0.05 M of pH 9.0 carbonate buffer (15 mM Na2CO3, 34.9 mM NaHCO3)
for 16 hours at room temperature. The plates were subsequently washed with phosphate-
buffered saline (PBS) containing 0.05% Tween and blocked with 200 μL of 5% PBS-milk
(Molico, NESTLÉ®). After 2 hours at 37°C, the plates were washed with 0.05% Tween PBS,
and 50 μL of plasma diluted 1:100 in 5% PBS-milk was added to each well in duplicate. The
plates were then incubated for 16 hours at room temperature. After the further washing of the
plates with 0.05% Tween PBS, 50 μL of human peroxidase-linked anti-IgG (DAKO
polyclonal rabbit, Glostrup, Denmark) diluted 1:10,000 in 5% PBS-milk was added to each
well, and the plates were incubated for 2 hours at room temperature. Then, after further
washing with 0.05% Tween PBS, the plates were developed by adding 100 μL of OPD (ortho-
phenylenediamine) (1 mg/mL) diluted in phosphate-citrate buffer (0.2 M NaH2PO4, 0.2 M
C6H8O7, pH 5.0) containing 0.03% hydrogen peroxide. This reaction was maintained in the
dark and stopped after 10 minutes by adding 25 μL of 4 N H2SO4 to each well. The optical
density (OD) was measured using an ELISA reader (EL800 Bio Tek, Winooski, USA) at a
wavelength of 490 nm. The reactivity index (RI) was calculated to define the negative or
positive samples, determined by dividing each sample's OD value by the cut-off point (Cut-
off: PvMSP-119 = 0.165). Samples with an RI ≥ 1 were considered positive, while samples
with an RI < 1 were considered negative. The cut-off point was established by taking the
mean ODs of 56 plasma samples obtained from the Pará Haemotherapy and Haematology
Foundation Centre (HEMOPA) from individuals residing in Belém, Pará State, with no
history of malaria plus three standard deviations.
Cytokine dosage
A CBA (cytometric bead array) Kit (BD) was used in this study to quantify cytokines
IL-2, INFγ and TNF (Th1) and IL-4, IL-5, and IL-10 (Th2) in the same sample. Six bead
populations with distinct fluorescence intensities were conjugated with a capture antibody
specific for each cytokine. They were mixed to form the CBA and read on a FACSCanto II-
type flow cytometer (Becton Dickinson, San Jose, CA) that was previously calibrated with
"setup beads", incubated with fluorescein isothiocyanate (FITC) or phycoerythrin (PE)

75
according to the manufacturer's recommendations. A standard curve was performed for each
cytokine and analyzed using FACSDiva software (Becton Dickinson, San Jose, CA, USA).
The bead populations were displayed according to their respective fluorescence intensities,
from dimmer to brighter. In the CBA, the cytokine capture beads were mixed with detection
antibody conjugated with PE fluorochrome and then incubated with the test samples to form a
"sandwich" test. The acquisition tubes were prepared with 50 μL of sample, 50 μL of bead
mix and 50 μL of Th1/Th2 PE detection reagent (Human Th1/Th2 PE Detection Reagent/1
vial, 4 mL). The same procedure was performed to obtain the standard curve. The tubes were
homogenized and incubated for three hours at room temperature in the dark. The results were
presented on graphs and in tables using FCAP Array 3 software (Becton Dickinson, San Jose,
CA, USA). Raw MFI (media fluorescence intensity) values were quantified for each cytokine.
The values were expressed in pg/mL for each cytokine in comparison to the standard curve.
Three hundred events were considered for each cytokine.
Collection and examination of faecal samples
Two faecal samples were collected from each individual, each in a separate container.
One contained a preservative solution (10% formaldehyde), and one was kept without
preservative solution. For the negative cases, three faecal samples were collected and
examined using the fresh/direct approach with and without Lugol solution and the Hoffman-
Pons-Janer and Faust methods. For each faecal sample, two slides were examined using light
microscopy (Nikon, Japan) at magnifications of 100X and 400X for parasite detection by two
researchers who were experienced in these identifications. The analyses were performed in a
private laboratory in the municipality of Oiapoque.
Statistical analysis
Each volunteer's epidemiological data results, which were obtained from the
questionnaire, was stored in an Epi-Info 3.5.1 database (CDC, Atlanta, GA, USA). The values
for each group (malaria, co-infected, enteroparasite and endemic control), as well as the
subgroups (helminths, protozoa and helminth-protozoa association), were expressed as a
percentage. The age, residence time in Oiapoque (years) (RT), number of previous malaria
episodes (NPE), period since last malaria (months) (PLM), haemoglobin levels (g/dL),
parasitaemia (parasites/L) and gametocytes were expressed as medians (1st and 3rd Quartile)
using BioEstat 5.3 statistical software. The differences between groups with regard to age,
RT, NPE, PLM and haemoglobin were calculated using Tukey's test based on a one-way
analysis of variance (ANOVA). The Kruskal-Wallis-Dunn test was used to calculate the
degree of anaemia.
The statistical tests used here were chosen by considering the size and type of each
variable and number of evaluated groups. The differences in the parasitaemia and the
gametocytes between the malaria and co-infected groups were calculated using the Wilcoxon-
Mann-Whitney test. For the PvMSP119 reactivity index (RI) among the studied groups,
multiple correlations were performed using the non-parametric Kruskal-Wallis test, followed
by Dunn's post-test. The data were expressed in box plot format (minimum to maximum
values, P25%-P75% and median). Significant differences were estimated using the median
values for each group; those with p ≤ 0.05 were considered significant.
For the cytokine expression analysis, the significance level was obtained by
comparing the cytokine concentrations of the studied groups using the GraphPad Prism
program, version 6.0 (GraphPad Software, San Diego, CA, USA). An analysis to uncover the
correlation between the groups was performed using the non-parametric Kruskal-Wallis test
followed by Dunn's post-test. The data were expressed in box plot format (minimum to
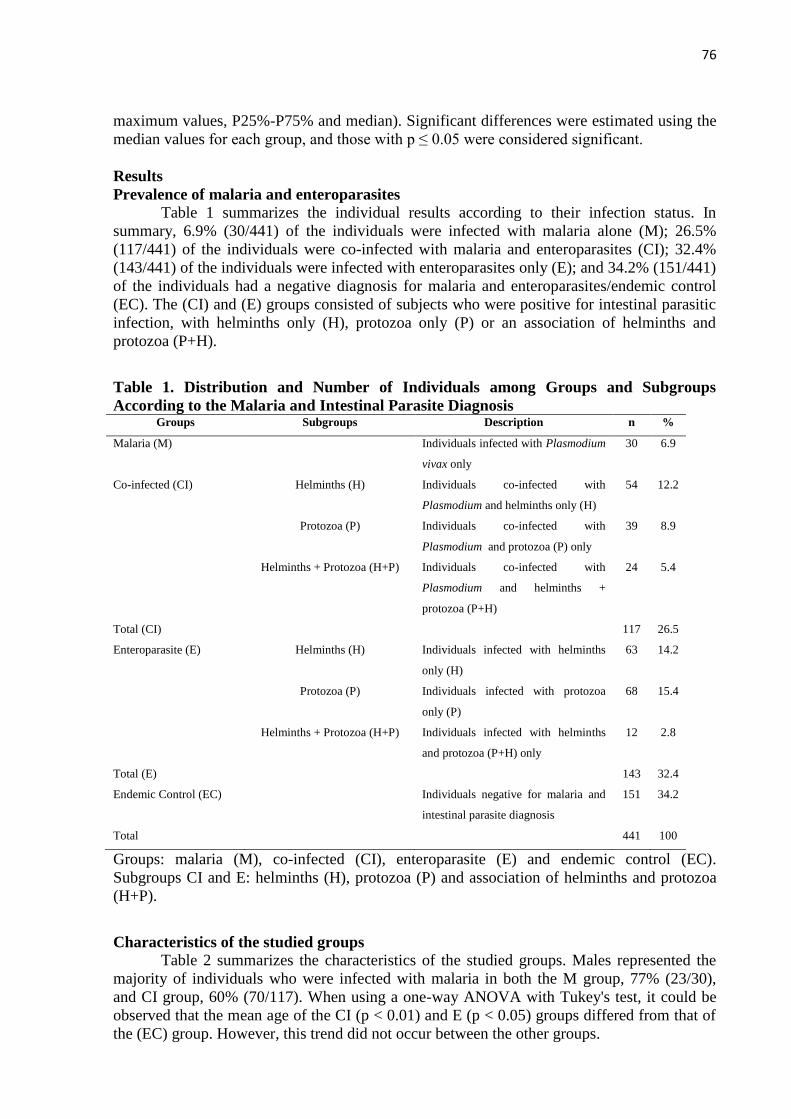
76
maximum values, P25%-P75% and median). Significant differences were estimated using the
median values for each group, and those with p ≤ 0.05 were considered significant.
Results
Prevalence of malaria and enteroparasites
Table 1 summarizes the individual results according to their infection status. In
summary, 6.9% (30/441) of the individuals were infected with malaria alone (M); 26.5%
(117/441) of the individuals were co-infected with malaria and enteroparasites (CI); 32.4%
(143/441) of the individuals were infected with enteroparasites only (E); and 34.2% (151/441)
of the individuals had a negative diagnosis for malaria and enteroparasites/endemic control
(EC). The (CI) and (E) groups consisted of subjects who were positive for intestinal parasitic
infection, with helminths only (H), protozoa only (P) or an association of helminths and
protozoa (P+H).
Table 1. Distribution and Number of Individuals among Groups and Subgroups
According to the Malaria and Intestinal Parasite Diagnosis Groups Subgroups Description n %
Malaria (M) Individuals infected with Plasmodium
vivax only
30 6.9
Co-infected (CI) Helminths (H) Individuals co-infected with
Plasmodium and helminths only (H)
54 12.2
Protozoa (P) Individuals co-infected with
Plasmodium and protozoa (P) only
39 8.9
Helminths + Protozoa (H+P) Individuals co-infected with
Plasmodium and helminths +
protozoa (P+H)
24 5.4
Total (CI) 117 26.5
Enteroparasite (E) Helminths (H) Individuals infected with helminths
only (H)
63 14.2
Protozoa (P) Individuals infected with protozoa
only (P)
68 15.4
Helminths + Protozoa (H+P) Individuals infected with helminths
and protozoa (P+H) only
12 2.8
Total (E) 143 32.4
Endemic Control (EC) Individuals negative for malaria and
intestinal parasite diagnosis
151 34.2
Total 441 100
Groups: malaria (M), co-infected (CI), enteroparasite (E) and endemic control (EC).
Subgroups CI and E: helminths (H), protozoa (P) and association of helminths and protozoa
(H+P).
Characteristics of the studied groups
Table 2 summarizes the characteristics of the studied groups. Males represented the
majority of individuals who were infected with malaria in both the M group, 77% (23/30),
and CI group, 60% (70/117). When using a one-way ANOVA with Tukey's test, it could be
observed that the mean age of the CI (p < 0.01) and E (p < 0.05) groups differed from that of
the (EC) group. However, this trend did not occur between the other groups.
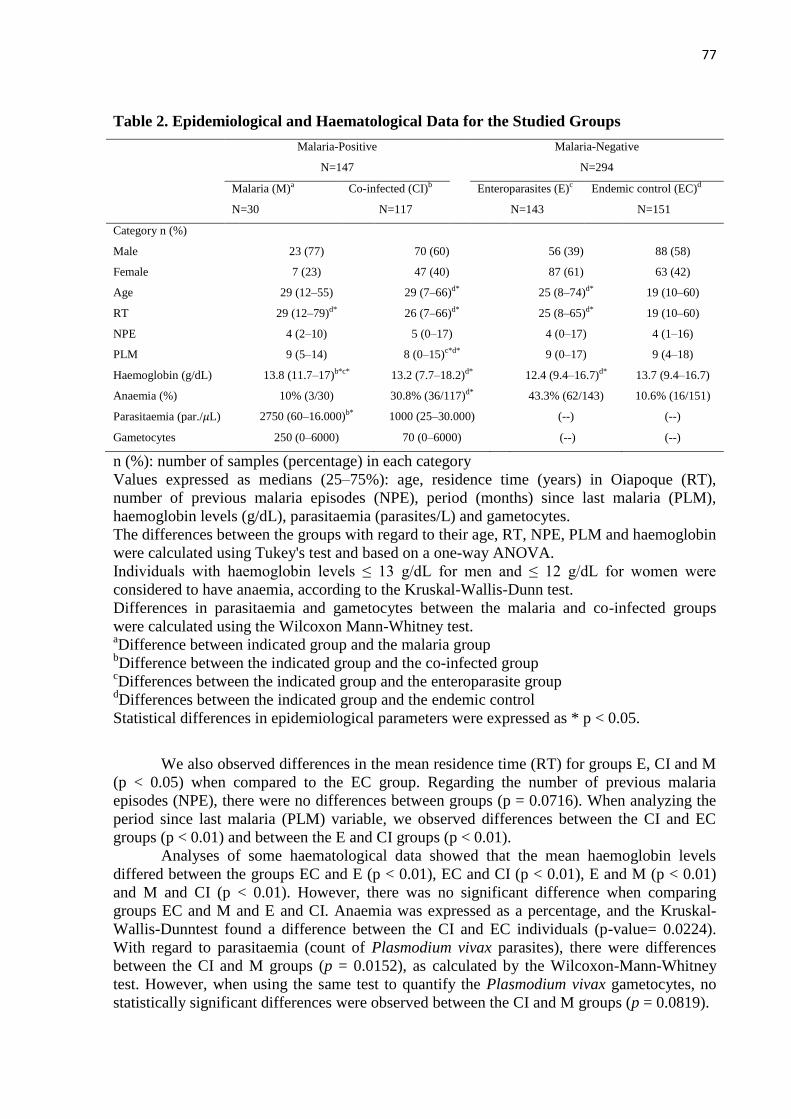
77
Table 2. Epidemiological and Haematological Data for the Studied Groups
Malaria-Positive
N=147
Malaria-Negative
N=294
Malaria (M)a
N=30
Co-infected (CI)b
N=117
Enteroparasites (E)c
N=143
Endemic control (EC)d
N=151
Category n (%)
Male 23 (77) 70 (60) 56 (39) 88 (58)
Female 7 (23) 47 (40) 87 (61) 63 (42)
Age 29 (12–55) 29 (7–66)d* 25 (8–74)d* 19 (10–60)
RT 29 (12–79)d* 26 (7–66)d* 25 (8–65)d* 19 (10–60)
NPE 4 (2–10) 5 (0–17) 4 (0–17) 4 (1–16)
PLM 9 (5–14) 8 (0–15)c*d* 9 (0–17) 9 (4–18)
Haemoglobin (g/dL) 13.8 (11.7–17)b*c* 13.2 (7.7–18.2)d* 12.4 (9.4–16.7)d* 13.7 (9.4–16.7)
Anaemia (%) 10% (3/30) 30.8% (36/117)d* 43.3% (62/143) 10.6% (16/151)
Parasitaemia (par./𝜇L) 2750 (60–16.000)b* 1000 (25–30.000) (--) (--)
Gametocytes 250 (0–6000) 70 (0–6000) (--) (--)
n (%): number of samples (percentage) in each category
Values expressed as medians (25–75%): age, residence time (years) in Oiapoque (RT),
number of previous malaria episodes (NPE), period (months) since last malaria (PLM),
haemoglobin levels (g/dL), parasitaemia (parasites/L) and gametocytes.
The differences between the groups with regard to their age, RT, NPE, PLM and haemoglobin
were calculated using Tukey's test and based on a one-way ANOVA.
Individuals with haemoglobin levels ≤ 13 g/dL for men and ≤ 12 g/dL for women were
considered to have anaemia, according to the Kruskal-Wallis-Dunn test.
Differences in parasitaemia and gametocytes between the malaria and co-infected groups
were calculated using the Wilcoxon Mann-Whitney test. aDifference between indicated group and the malaria group
bDifference between the indicated group and the co-infected group
cDifferences between the indicated group and the enteroparasite group
dDifferences between the indicated group and the endemic control
Statistical differences in epidemiological parameters were expressed as * p < 0.05.
We also observed differences in the mean residence time (RT) for groups E, CI and M
(p < 0.05) when compared to the EC group. Regarding the number of previous malaria
episodes (NPE), there were no differences between groups (p = 0.0716). When analyzing the
period since last malaria (PLM) variable, we observed differences between the CI and EC
groups (p < 0.01) and between the E and CI groups (p < 0.01).
Analyses of some haematological data showed that the mean haemoglobin levels
differed between the groups EC and E (p < 0.01), EC and CI (p < 0.01), E and M (p < 0.01)
and M and CI (p < 0.01). However, there was no significant difference when comparing
groups EC and M and E and CI. Anaemia was expressed as a percentage, and the Kruskal-
Wallis-Dunntest found a difference between the CI and EC individuals (p-value= 0.0224).
With regard to parasitaemia (count of Plasmodium vivax parasites), there were differences
between the CI and M groups (p = 0.0152), as calculated by the Wilcoxon-Mann-Whitney
test. However, when using the same test to quantify the Plasmodium vivax gametocytes, no
statistically significant differences were observed between the CI and M groups (p = 0.0819).

78
Anti-PvMSP119 IgG responses to Plasmodium vivax
The percentage of individuals in the investigated population that possessed naturally
acquired anti-PvMSP-119 IgG-1 antibodies is shown in Fig 1. Responders represented 51.2%
(226/441) of individuals (Fig 1a). The presence of anti-PvMSP-119 IgG antibodies among the
groups showed a higher prevalence of 81.1% (95/117) in the CI group. The lowest prevalence
was in the EC group, with 33.1% (50/151) (Fig 1b). To support the data (Fig 1c), each
individual's data were shown as a point and expressed in box plot format. Significant
differences were observed, as estimated by the median values for each group (a nonparametric
Kruskal-Wallis test followed by Dunn’s post-test). These values were considered significant,
where p < 0.05.
Fig 1. (a) Frequency-specific antibody response to PvMSP-119, as determined by ELISA. The
subjects were grouped into responders and non-responders to the recombinant protein. (b)
Prevalence of anti-PvMSP-19 IgG antibodies in the studied groups. (c) PvMSP-119 reactivity
index (RI) between the studied groups as expressed in box plot format, with individual data
shown as points. Multiple correlations were made using the nonparametric Kruskal-Wallis
test followed by Dunn’s post hoc test (minimum to maximum values, P25%–P75% and
median); significant differences were estimated using the median values for each group, and
those with p < 0.05 were considered significant. ** p < 0.05, *** p = 0.001 and **** p <
0.001.

79
Fig. 2 shows the seroprevalence in terms of the frequency of anti-MSP119-specific IgG
antibodies as distributed across the four study groups as well as the reactivity index, which
specified and subdivided the CI and E groups. Fig 2a shows that the helminths were most
prevalent in the CI group, with 85.1% (46/54). The highest values in group E were found in
the protozoa and helminth association subgroup, with 58.3% (7/12). Fig 2b expresses the
PvMSP-119 reactivity index (RI) among the studied groups structured as in Fig 2a. The data
are expressed in box plot format (minimum to maximum values, P25%-P75% and median),
with each individual's data shown as a point. Significant differences were observed in each
group’s median values, and those with p < 0.05 were considered significant. ** p < 0.05, ***
p = 0.001 and **** p < 0.001.
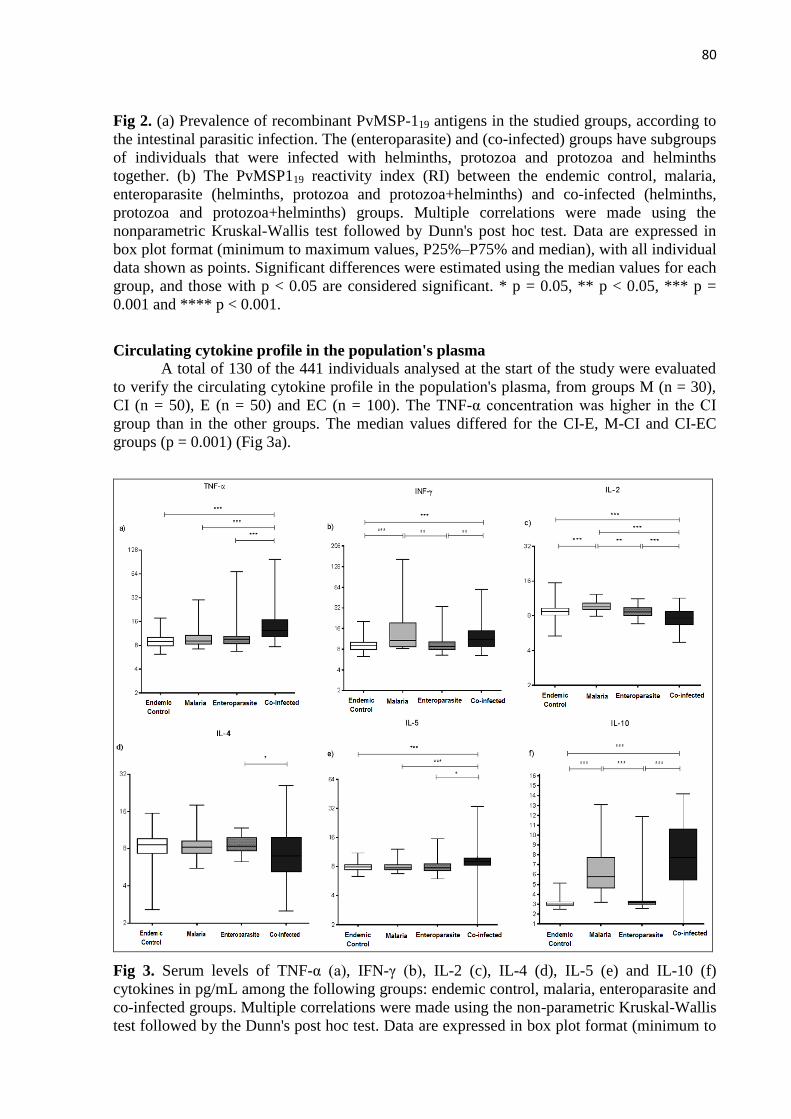
80
Fig 2. (a) Prevalence of recombinant PvMSP-119 antigens in the studied groups, according to
the intestinal parasitic infection. The (enteroparasite) and (co-infected) groups have subgroups
of individuals that were infected with helminths, protozoa and protozoa and helminths
together. (b) The PvMSP119 reactivity index (RI) between the endemic control, malaria,
enteroparasite (helminths, protozoa and protozoa+helminths) and co-infected (helminths,
protozoa and protozoa+helminths) groups. Multiple correlations were made using the
nonparametric Kruskal-Wallis test followed by Dunn's post hoc test. Data are expressed in
box plot format (minimum to maximum values, P25%–P75% and median), with all individual
data shown as points. Significant differences were estimated using the median values for each
group, and those with p < 0.05 are considered significant. * p = 0.05, ** p < 0.05, *** p =
0.001 and **** p < 0.001.
Circulating cytokine profile in the population's plasma
A total of 130 of the 441 individuals analysed at the start of the study were evaluated
to verify the circulating cytokine profile in the population's plasma, from groups M (n = 30),
CI (n = 50), E (n = 50) and EC (n = 100). The TNF-α concentration was higher in the CI
group than in the other groups. The median values differed for the CI-E, M-CI and CI-EC
groups (p = 0.001) (Fig 3a).
Fig 3. Serum levels of TNF-α (a), IFN-γ (b), IL-2 (c), IL-4 (d), IL-5 (e) and IL-10 (f)
cytokines in pg/mL among the following groups: endemic control, malaria, enteroparasite and
co-infected groups. Multiple correlations were made using the non-parametric Kruskal-Wallis
test followed by the Dunn's post hoc test. Data are expressed in box plot format (minimum to

81
maximum values, P25%–P75% and median). Significant differences were estimated using the
median values for each group, with p < 0.05 being considered significant. * p = 0.05, ** p <
0.05 and *** p = 0.001.
Additionally, the IFN-γ levels were higher in group M than in the other groups.
Comparisons of the M-EC (p = 0.001), M-E (p < 0.05), E-CI (p < 0.05) and EC-CI (p =
0.001) groups were statistically significant (Fig 3b). Increased IL-2 concentration values were
observed in the EC group. Statistically significant differences were observed for the EC-CI (p
= 0.001), EC-M (p = 0.001), M-CI (p = 0.001), M-E (p < 0.05) and CI-E (p = 0.001) groups
(Fig 3c).
The IL-4 concentration was higher in the CI group, and there was a statistically
significant difference for CI-E (p = 0.05). Comparisons between the remaining groups were
not statistically significant (Fig 3d). IL-5 showed high concentrations in the CI group, and
comparisons of the CI-E (p = 0.05), CI-M (p = 0.001) and EC-CI (p = 0.001) groups were
significant (Fig 3e). With respect to the IL-10 cytokine profile, we can observe increased
concentrations for groups CI and M. There were statistically significant differences for groups
CI-E, CI-EC, M-E and M-EC (p = 0.001) (Fig 3f).
Discussion
Microorganism co-infections are largely unexplored, and their effects can be either
favourable or harmful to the human host. Some studies have explored the effects of helminth
co-infections on the dynamics of P. falciparum malaria transmission and its correlation with
anaemia [26,27]. In Brazil, recent studies have described the haematological and
immunological profiles of vivax malaria and intestinal parasite co-infections [25-27] in two
areas of Brazil's western Amazon. We are the first to study the effect of intestinal parasites on
biological aspects of malaria in individuals who were naturally infected with P. vivax in a
malaria-endemic area located on the Brazilian border with French Guiana.
The parasitological findings of this study show a higher prevalence of individuals who
were infected only with E (32.4%), followed by the CI group (26.5%). Group M had the
lowest prevalence (6.9%). The CI and M groups were predominant in the male population,
emphasizing their geographical peculiarities and aspects related to the primary constraints and
health determinants in the border region related to work activities, more specifically, the
illegal mining (gold mining) and agricultural activities prevalent in the region where contact
with the vector and soil is frequent.
The results show that individual haemoglobin levels differ between the CI and M
groups. Anaemia was more prevalent in the CI and E individuals, with significant differences
in the plasmodial parasite load between the CI and M groups. This haematological change
observed between the groups emphasizes that anaemia is multifactorial and is a frequent early
manifestation of malaria that contributes to disease severity, especially in the context of
concurrent infections [30-32]. This disease leads to the destruction or sequestration of
erythrocytes, abnormal erythropoiesis and blood loss arising from eventual coagulopathy [33-
35]. Additionally, cytokine polymorphisms have been associated with susceptibility to severe
malarial anaemia, disturbing erythropoiesis [36,37]. Studies have shown that a number of co-
infections increase susceptibility to anaemia because they exacerbate the inflammation caused
by malaria [38,39]. Helminth infections are associated with the increased likelihood of
development and the greater severity of anaemia in Plasmodium infection [39] as well as
genetic changes - haemoglobinopathies - and nutritional deficiencies [40].
The molecular mechanisms underlying malarial anaemia are largely unknown,
although it is accepted that the disease is complex and multifactorial and has characteristics in

82
common with acute malaria [40,41]. In this context, the results of this study indicate that the
effect of enteroparasitic infection on the prevalence and severity of anaemia caused by vivax
malaria still requires further investigation. These differences may be explained by the fact that
in a co-infection, there may be a negative interaction between parasites, characterized by a
protective effect on the individual and a suppressive effect on one of the pathogens or a
positive interaction in which an infection is facilitated by the presence of the other parasite
[41-43]. Thus, it is possible that P. vivax and intestinal parasite co-infection could modify the
immune response profile against certain specific Plasmodium antigens.
Seroepidemiological studies to investigate the type and magnitude of the vivax malaria
immune response in naturally exposed populations, in both Brazil [44-46] and around the
world [47-50], have provided important information about the potential candidacy of
immunogenic P. vivax antigens for use in creating an anti-malarial vaccine. The differences
between antibody profiles reported in the present study indicate the importance of performing
immunoepidemiological studies in different malaria-endemic areas, where transmission
intensities and human genetic backgrounds are very distinct [51].
Studies on antibody responses to Plasmodium antigens offer a key process in the
discovery and development of anti-malarial immunotherapy. Several studies have reported
high antibody responses to P. vivax antigens in individuals who have been exposed to malaria
infections [27,52]. In this study, anti-PvMSP-119 IgG antibodies were detected in 51.2%
(226/441) of the studied population, indicating the presence of an immunogenic protein, with
results similar to those of other studies in the Amazon region [27,52-55]. Higher prevalence
and specific antibody RIs were observed in the CI and M groups, reaching 81.1% (95/117)
and 76.7% (27/30), respectively. As demonstrated in other endemic areas in the Brazilian
Amazon, PvMSP-119 appears as an immunogenic molecule during naturally acquired malaria
infections [56-58].
Corroborating the findings in the Western Brazilian Amazon, the presence of
enteroparasites also did not affect antibody responses to this antigen [27]. The reduced
prevalence of IgG and RI in the group of individuals who were infected with intestinal
parasites alone appears to be a general characteristic of this association, as in Rondônia, the
prevalence of protozoa in vivax malaria-co-infected individuals was higher than that detected
in the Oiapoque municipality, where geohelminths were more prevalent.
These results support those of other studies [59,60] that have suggested, theoretically,
the possibility that malaria-helminth co-infection potentiates parasitic genetic diversity in
individuals who are exposed to both infections [21,59,60]. Understanding this transmission
dynamic and the evolutionary implications of populational P. vivax co-infection is very
important for predicting the emergence and distribution of different strains of the parasite,
which may differ in their virulence and drug resistance [61]. This finding is a major concern
in these neglected border areas, where the usual practice of inadequate self-treatment
contributes to an increased risk of developing resistance to antimalarial drugs.
Due to Oiapoque's local genetic diversity and the combined effects of the geographical
structure of the border between Brazil and French Guiana in a regional, national and
international context, there is a dispersion of Plasmodium vivax populations between the two
countries [62]. Establishing P. vivax's complex geographical pattern is important, both for
evaluating diversity when encoding candidate antigens for vaccines and when formulating and
structuring surveillance measures to control malaria.
In fact, helminths were the most prevalent enteroparasites in the CI (85.1%, 46/54)
and E (58.3%, 7/12) groups. The reduction in the antibody prevalence and reactivity index in
group E may be explained by the association (protozoa-helminths) and/or presence of
intestinal protozoa. However, in the CI group, the IgG prevalence and reactivity index were
not reduced and were similar to those in infected group M. These data confirm the non-

83
importance of protozoa in the immune process during co-infection with P. vivax, as
previously shown in another Brazilian Amazon region [27]. However, the diversity of
enteroparasites meant that groups could not be formed with sample sizes large enough for
researchers to be able to infer some sort of effect on co-infection.
The general trend in co-infection studies (helminths, malaria) is convergent and points
to the possibility of increasing incidence and prevalence of malaria [63,64] and a tendency to
reduce malaria symptoms [24,65]. This finding suggests that these patients might be less
likely to seek treatment, making them a potential source of transmission [21]. In poor tropical
areas, health determinants and constraints affect the population's health care and services
sought when the disease is at an advanced stage.
In malaria, protective immunity is gradually acquired through the natural exposure of
people living in areas where the disease is endemic, but this immunity is rapidly lost when
exposure ceases, indicating that malaria's immunological memory is short term and requires
constant exposure to the parasite [66,67]. Although infection by many enteroparasites was
observed, the research revealed a prevalence of helminths, and it is generally believed that
helminth infection may alter the host's natural immune response to Plasmodium due to the
anti-inflammatory effect of helminth-induced cytokines [26].
Understanding the Th1/Th2 response pattern is of great importance for understanding
the host's defense, and these antagonistic responses allow for the homeostasis of the immune
system, which is important for parasite containment [68]. In cases in which an inflammatory
pattern is prevalent, the disease tends to be more severe [5]. In this regard, the ability to
measure numerous molecules and visualise inflammatory changes in individuals who are co-
infected with malaria and enteroparasites is extremely important for advancing the
understanding of the immune response to pathogens [26,27]. In this context, despite the
importance of the PvMSP-119 immunogenic molecule being present in response to malaria,
other mediators, such as pro- and anti-inflammatory cytokines, also act in the elimination of
plasmodial infections.
The individual analysis of mean serum cytokine levels in the studied population
showed a broad variation in the serum concentrations of all the Th1 inflammatory profile
cytokines (IFN-γ, TNF-α and IL-2) in the studied groups. Initially, we found that the
expression of these cytokines exhibited a very similar profile between groups, with high
levels for M and CI individuals. This observation suggests that only malarial infection, and
not intestinal parasites, affected the increase in these levels. Elevated serum cytokine levels
during infection have been reported previously, both in uncomplicated and severe malaria
[69,70]. However, their participation in reducing the parasitic load remains controversial.
Some studies have shown the importance of the cytokine pattern and its interactions in
the vivax malaria immune response [68].Increased parasitaemia and immune responses during
the course of the disease are factors that determine its severity [71]. Inflammatory patterns are
rare, and few studies have evaluated the various cytokines present using the same method and
the same group of malaria-infected patients [72]. This observation can also be corroborated
when we observe the Th2 profile of anti-inflammatory cytokines (IL-4, IL-5 and IL-10), with
increased serum concentrations in co-infected individuals. In fact, the high levels of these
cytokines and their increased frequency among the groups infected with malaria (Cl and M)
are consistent with the hypothesis that these cytokines play an anti-inflammatory role by
inhibiting pro-inflammatory mechanisms after the initial stages of infection [73].
In the literature, IL-4 and IL-5 concentrations have been shown to confer protection
against extracellular pathogens, including helminths [74]. Some studies have shown
conflicting results, in which the concentrations of these cytokines depend on the host's
parasitaemia levels [75]. Additionally, several studies focusing on IL-10 have shown that this
cytokine plays an important role in the control of the antiparasitic response and tissue damage

84
caused by this response, demonstrating its importance in regulating the immune response
[76]. Furthermore, studies have shown that interactions between IL-6 and IL-10 cytokines are
correlated with the parasitic load [77]. This study presents several variations in the Th1
cytokine expression (IFN-γ, TNF-α and IL-2), with similar medians among the studied
groups, but with high and varying levels of each cytokine. The Th2 cytokine (IL-4, IL-5 and
IL-10) medians differed from each other in the studied groups, showing high levels for
individuals in the co-infected group.
Variations in the cytokine production and high Th1 (TNF-α) and Th2 (IL-4, IL-5 and
IL-10) profile levels in the co-infected group therefore suggest that cytokines exhibit marked
increases in their plasma concentrations when individuals are infected with P. vivax, with a
shifting pattern inchanges associated with malaria and intestinal parasite co-infection. They
seem to act in inflammatory processes in a nonspecific way, not exerting any effect on co-
infection. However, only the IL-10 cytokine levels were affected in cases of malaria and
intestinal parasite co-infection. Further wide-ranging studies are needed to evaluate the
intestinal parasite species present in malaria-endemic areas and their relationship to the
immune response of infected individuals.
These results may reflect the fact that the number of parasites in the bloodstream at a
particular time may not always show the maximum immunogenic potential capable of
inducing a host's immune response because some parasite antigens may be in immune system
tissue and not just in the blood [78]. The overall magnitude of production of pro- and anti-
inflammatory cytokines and the imbalance between them during co-infection have been
proposed as important determinants in controlling or exacerbating malaria [79]. However,
new immunological and molecular epidemiological studies are required to better understand
the protective immune response scenario and outcome in Plasmodium vivax malaria. These
studies will be essential in the development of vaccines and new immunotherapeutic targets.
The present study has some limitations, we acknowledge. First the cross sectional
design is prone to bias and confounding and does not allow to infer causality. Secondly, the
small sample size, notably in the malaria alone group, may not have been sufficient to detect
some differences with other groups. Morevover, this evaluation did not allow to look at the
respective effects of different nematode species which may have contrasting effects.
Likewise, the current results highlight the possibility that the enteroparasite and vivax malaria
co-infection on the Brazil-French Guiana border mark biological differences, until
unexplained, towards the differential immune response and haematological profile of the host-
parasite interaction in vivax malaria in Oiapoque, Amapá state. Aditionaly, provide additional
information about the influence that enteroparasite could have in a malaria vivax
development; essential for diagnostic and preventive strategies.
Conclusion
The malaria by P. vivax is endemic in the region studied and its coinfection with intestinal
parasitosis is present in the municipality. Fecal findings demonstrate a diverse enteroparasitic
prevalence among the groups, highlighting the particularities and geographic aspects related
to the determinants of health in the border area. Hemoglobin levels in individuals appear to be
strongly attributed to malaria infection, especially in the context of concurrent infections, with
patients co-infected with lower parasitemia but were more likely to be anemic than patients
with isolated malaria. The high prevalence of sexual stage antibodies between groups
increases the premise that most individuals are exposed to malaria, making it an important
factor for the maintenance of transmission in the locality. The profile of inflammatory and
anti-inflammatory cytokines, coinfection may influence the two parasitoses investigated. We
must emphasize the need for longitudinal studies of statistical power enough to better
characterize the nuances of this co-infection.

85
Acknowledgments
We would like to extend our appreciation and gratitude to all who agreed to participate in this
study as well as to the local malaria control team in the municipality of Oiapoque for their
assistance in sample collection and technical support. We also thank the Central Laboratory
(LACEN) of Amapá, the Microbiology and Immunology Laboratory of the Federal University
of Pará and the Malaria Immunogenetics Laboratory/Parasitology Section of the Evandro
Chagas Institute for their sample analysis, logistical support and general support.
References
1. Arevalo-Herrera M, Chitnis C, Herrera S. Current status of Plasmodium vivax vaccine.
Hum Vaccin. 2010;6: 124-132.
2. Arevalo-Herrera M, Quinones ML, Guerra C, Cespedes N, Giron S, Ahumada M, et al.
Malaria in selected non-Amazonian countries of Latin America. Acta Trop. 2012;121: 303-
314.
3. Parise ÉV. Vigilância, prevenção e controle da malária em Palmas, Tocantins, Brasil, no
período 2000 a 2013. Hygeia. 2014;10: 190-206.
4. Lapouble OMM, Santelli ACF, Muniz-Junqueira MI. Situação epidemiológica da malária
na região Amazônica Brasileira, 2003 a 2012. Rev Panam Salud Publica. 2015;38: 300-306.
5. Andrade BB, Reis-Filho A, Souza-Neto SM, Clarencio J, Camargo LM, Barral A, et al.
Severe Plasmodium vivax malaria exhibits marked inflammatory imbalance. Malar J. 2010;9:
13.
6. Alecrim MG, Lacerda MV, Mourao MP, Alecrim WD, Padilha A, Cardoso BS, et al.
Successful treatment of Plasmodium falciparum malaria with a six-dose regimen of
artemether-lumefantrine versus quinine-doxycycline in the Western Amazon region of Brazil.
Am J Trop Med Hyg. 2006;74: 20-25.
7. Lacerda MV, Hipolito JR, Passos LN. Chronic Plasmodium vivax infection in a patient
with splenomegaly and severe thrombocytopenia. Rev Soc Bras Med Trop. 2008;41: 522-523.
8. Hotez PJ, Brindley PJ, Bethony JM, King CH, Pearce EJ, Jacobson J. Helminth infections:
the great neglected tropical diseases. J Clin Invest. 2008;118: 1311-1321.
9. Brooker S, Akhwale W, Pullan R, Estambale B, Clarke SE, Snow RW, et al. Epidemiology
of plasmodium-helminth co-infection in Africa: populations at risk, potential impact on
anemia, and prospects for combining control. Am J Trop Med Hyg. 2007;77: 88-98.
10. Salim N, Knopp S, Lweno O, Abdul U, Mohamed A, Schindler T, et al. Distribution and
risk factors for Plasmodium and helminth co-infections: a cross-sectional survey among
children in Bagamoyo district, coastal region of Tanzania. PLoS Negl Trop Dis. 2015;9:
e0003660.
11. Mboera LE, Senkoro KP, Rumisha SF, Mayala BK, Shayo EH, Mlozi MR. Plasmodium
falciparum and helminth coinfections among schoolchildren in relation to agro-ecosystems in
Mvomero district, Tanzania. Acta Trop. 2011;120: 95-102.

86
12. Adegnika AA, Kremsner PG. Epidemiology of malaria and helminth interaction: a review
from 2001 to 2011. Curr Opin HIV AIDS. 2012;7: 221-224.
13. Noland GS, Graves PM, Sallau A, Eigege A, Emukah E, Patterson AE, et al. Malaria
prevalence, anemia and baseline intervention coverage prior to mass net distributions in Abia
and Plateau states, Nigeria. BMC Infect Dis. 2014;14: 168.
14. Roussilhon C, Brasseur P, Agnamey P, Perignon JL, Druilhe P. Understanding human-
Plasmodium falciparum immune interactions uncovers the immunological role of worms.
PLoS One. 2010;5: e9309.
15. Midzi N, Mtapuri-Zinyowera S, Sangweme D, Paul NH, Makware G, Mapingure MP, et
al. Efficacy of integrated school based de-worming and prompt malaria treatment on
helminths-Plasmodium falciparum co-infections: a 33 months follow up study. BMC Int
Health Hum Rights. 2011;11: 9.
16. Ojurongbe O, Adegbayi AM, Bolaji OS, Akindele AA, Adefioye OA, Adeyeba OA.
Asymptomatic falciparum malaria and intestinal helminths co-infection among school
children in Osogbo, Nigeria. J Res Med Sci. 2011;16: 680-686.
17. Nacher M. Interactions between worms and malaria: good worms or bad worms? Malar J.
2011;10: 259.
18. Boel M, Carrara VI, Rijken M, Proux S, Nacher M, Pimanpanarak M, et al. Complex
Interactions between soil-transmitted helminths and malaria in pregnant women on the Thai-
Burmese border. PLoS Negl Trop Dis. 2010;4: e887.
19. Murray J, Murray A, Murray M, Murray C. The biological suppression of malaria: an
ecological and nutritional interrelationship of a host and two parasites. Am J Clin Nutr.
1978;31: 1363-1366.
20. Spiegel A, Tall A, Raphenon G, Trape JF, Druilhe P. Increased frequency of malaria
attacks in subjects co-infected by intestinal worms and Plasmodium falciparum malaria. Trans
R Soc Trop Med Hyg. 2003;97: 198-199.
21. Nacher M. Helminth-infected patients with malaria: a low profile transmission hub?
Malar J. 2012;11: 376.
22. Nacher M, Singhasivanon P, Silachamroon U, Treeprasertsuk S, Vannaphan S, Traore B,
et al. Helminth infections are associated with protection from malaria-related acute renal
failure and jaundice in Thailand. Am J Trop Med Hyg. 2001;65: 834-836.
23. Chaorattanakawee S, Natalang O, Hananantachai H, Nacher M, Brockman A, Nosten F,
et al. Trichuris trichiura infection is associated with the multiplicity of Plasmodium
falciparum infections, in Thailand. Ann Trop Med Parasitol. 2003;97: 199-202.
24. Nacher M, Gay F, Singhasivanon P, Krudsood S, Treeprasertsuk S, Mazier D, et al.
Ascaris lumbricoides infection is associated with protection from cerebral malaria. Parasite
Immunol. 2000;22: 107-113.

87
25. Melo GC, Reyes-Lecca RC, Vitor-Silva S, Monteiro WM, Martins M, Benzecry SG, et
al. Concurrent helminthic infection protects schoolchildren with Plasmodium vivax from
anemia. PLoS One. 2010;5: e11206.
26. Sánchez-Arcila JC, Perce-da-Silva DS, Vasconcelos MPA, Rodrigues-da-Silva RN,
Pereira VA, Aprígio CJL, et al. Intestinal parasites coinfection does not alter plasma cytokines
profile elicited in acute malaria in subjects from endemic area of Brazil. Mediators Inflamm.
2014;2014: 1-12.
27. Sánchez-Arcila JC, França MM, Pereira VA, Vasconcelos MPA, Têva A, Perce-da-Silva
DS, et al. The influence of intestinal parasites on Plasmodium vivax-specific antibody
responses to MSP-119 and AMA-1 in rural populations of the Brazilian Amazon. Malar J.
2015;14: 442.
28. Herrera S, Vallejo AF, Quintero JP, Arevalo-Herrera M, Cancino M, Ferro S. Field
evaluation of an automated RDT reader and data management device for Plasmodium
falciparum/Plasmodium vivax malaria in endemic areas of Colombia. Malar J. 2014;13: 87.
29. Bueno LL, Morais CG, Lacerda MV, Fujiwara RT, Braga EM. Interleukin-17 producing
T helper cells are increased during natural Plasmodium vivax infection. Acta Trop. 2012;123:
53-57.
30. Weatherall DJ, Miller LH, Baruch DI, Marsh K, Doumbo OK, Casals-Pascual C, et al.
Malaria and the red cell. Hematology Am Soc Hematol Educ Program. 2002;1: 35-57.
31. Lamikanra AA, Brown D, Potocnik A, Casals-Pascual C, Langhorne J, Roberts DJ.
Malarial anemia: of mice and men. Blood. 2007;110: 18-28.
32. Ekvall H. Malaria and anemia. Curr Opin Hematol. 2003;10: 108-114.
33. Gilson PR, Crabb BS. Morphology and kinetics of the three distinct phases of red blood
cell invasion by Plasmodium falciparum merozoites. Int J Parasitol. 2009;39: 91-96.
34. Holder AA, Blackman MJ, Borre M, Burghaus PA, Chappel JA, Keen JK, et al. Malaria
parasites and erythrocyte invasion. Biochem Soc Trans. 1994;22: 291-295.
35. Stoute JA, Odindo AO, Owuor BO, Mibei EK, Opollo MO, Waitumbi JN. Loss of red
blood cell-complement regulatory proteins and increased levels of circulating immune
complexes are associated with severe malarial anemia. J Infect Dis. 2003;187: 522-525.
36. Chang KH, Stevenson MM. Malarial anaemia: mechanisms and implications of
insufficient erythropoiesis during blood-stage malaria. Int J Parasitol. 2004;34: 1501-1516.
37. Awandare GA, Martinson JJ, Were T, Ouma C, Davenport GC, Ong'echa JM, et al. MIF
(macrophage migration inhibitory factor) promoter polymorphisms and susceptibility to
severe malarial anemia. J Infect Dis. 2009;200: 629-637.
38. McQueen PG, McKenzie FE. Host control of malaria infections: constraints on immune
and erythropoeitic response kinetics. PLoS Comput Biol. 2008;4: e1000149.

88
39. Haldar K, Mohandas N. Malaria, erythrocytic infection, and anemia. Hematology Am
Soc Hematol Educ Program. 2009;1: 87-93.
40. Calis JC, Phiri KS, Faragher EB, Brabin BJ, Bates I, Cuevas LE, et al. Severe anemia in
Malawian children. N Engl J Med. 2008;358: 888-899.
41. Midzi N, Mtapuri-Zinyowera S, Mapingure MP, Sangweme D, Chirehwa MT, Brouwer
KC, et al. Consequences of polyparasitism on anaemia among primary school children in
Zimbabwe. Acta Trop. 2010;115: 103-111.
42. Boef AG, May L, van Bodegom D, van Lieshout L, Verweij JJ, Maier AB, et al. Parasitic
infections and immune function: effect of helminth infections in a malaria endemic area.
Immunobiology. 2013;218: 706-711.
43. Abanyie FA, McCracken C, Kirwan P, Molloy SF, Asaolu SO, Holland CV, et al.
Ascaris co-infection does not alter malaria-induced anaemia in a cohort of Nigerian preschool
children. Malar J. 2013;12: 1.
44. Soares IS, Barnwell JW, Ferreira MU, Cunha MG, Laurino JP, Castilho BA, et al. A
Plasmodium vivax vaccine candidate displays limited allele polymorphism, which does not
restrict recognition by antibodies. Mol Med. 1999;5: 459-470.
45. Mourão LC, Morais CG, Bueno LL, Jimenez MC, Soares IS, Fontes CJ, et al. Naturally
acquired antibodies to Plasmodium vivax blood-stage vaccine candidates (PvMSP-119 and
PvMSP-3α359−798) and their relationship with hematological features in malaria patients
from the Brazilian Amazon. Microbes Infect. 2012;14: 730-739.
46. Rocha MV, Francoso KS, Lima LC, Camargo TM, Machado RL, Costa FT, et al.
Generation, characterization and immunogenicity of a novel chimeric recombinant protein
based on Plasmodium vivax AMA-1 and MSP119. Vaccine. 2017;35: 2463-2472.
47. Valencia SH, Rodriguez DC, Acero DL, Ocampo V, Arevalo-Herrera M. Platform for
Plasmodium vivax vaccine discovery and development. Mem Inst Oswaldo Cruz. 2011;106:
179-192.
48. malERA Consultative Group on Vaccines. A research agenda for malaria eradication:
vaccines. PLoS Med. 2011;8: e1000398.
49. Hoffman SL, Vekemans J, Richie TL, Duffy PE. The march toward malaria vaccines.
Am J Prev Med. 2015;49: S319-S333.
50. Mueller I, Shakri AR, Chitnis CE. Development of vaccines for Plasmodium vivax
malaria. Vaccine. 2015;33: 7489-7495.
51. Riccio EK, Totino PR, Pratt-Riccio LR, Ennes-Vidal V, Soares IS, Rodrigues MM, et al.
Cellular and humoral immune responses against the Plasmodium vivax MSP-119 malaria
vaccine candidate in individuals living in an endemic area in north-eastern Amazon region of
Brazil. Malar J. 2013;12: 326.

89
52. Vicentin EC, Francoso KS, Rocha MV, Iourtov D, Dos Santos FL, Kubrusly FS, et al.
Invasion-inhibitory antibodies elicited by immunization with Plasmodium vivax apical
membrane antigen-1 expressed in Pichia pastoris yeast. Infect Immun. 2014;82: 1296-1307.
53. Arruda ME, Zimmerman RH, Souza RMC, Oliveira-Ferreira J. Prevalence and level of
antibodies to the circumsporozoite protein of human malaria parasites in five states of the
Amazon region of Brazil. Memórias do Instituto Oswaldo Cruz. 2007;102: 367-372.
54. Barbedo MB, Ricci R, Jimenez MC, Cunha MG, Yazdani SS, Chitnis CE, et al.
Comparative recognition by human IgG antibodies of recombinant proteins representing three
asexual erythrocytic stage vaccine candidates of Plasmodium vivax. Mem Inst Oswaldo Cruz.
2007;102: 335-339.
55. Lima-Junior JC, Rodrigues-da-Silva RN, Banic DM, Jiang J, Singh B, Fabricio-Silva
GM, et al. Influence of HLA-DRB1 and HLA-DQB1 alleles on IgG antibody response to the
P. vivax MSP-1, MSP-3alpha and MSP-9 in individuals from Brazilian endemic area. PLoS
One. 2012;7: e36419.
56. Soares IS, Levitus G, Souza JM, del Portillo HA, Rodrigues MM. Acquired immune
responses to the N-and C-terminal regions of Plasmodium vivax merozoite surface protein 1
in individuals exposed to malaria. Infect Immun. 1997;65: 1606-1614.
57. Storti-Melo LM, da Costa DR, Souza-Neiras WC, Cassiano GC, Couto VS, Povoa MM,
et al. Influence of HLA-DRB-1 alleles on the production of antibody against CSP, MSP-1,
AMA-1, and DBP in Brazilian individuals naturally infected with Plasmodium vivax. Acta
Trop. 2012;121: 152-155.
58. Cassiano GC, Furini AA, Capobianco MP, Storti-Melo LM, Cunha MG, Kano FS, et al.
Polymorphisms in B cell co-stimulatory genes are associated with IgG antibody responses
against blood-stage proteins of Plasmodium vivax. PLoS One. 2016;11: e0149581.
59. Nacher M, Singhasivanon P, Gay F, Silachomroon U, Phumratanaprapin W,
Looareesuwan S. As infecções por plasmodium falciparum e Plasmodium vivax são
associadas a ascaris lumbricoides: efeito imunomodulador? J Parasitol. 2001;87: 912-915.
60. Nacher M, Singhasivanon P, Krudsood S, Phumratanaprapin W, Vanaphan S,
Tangpukdee N, et al. Inverse relationship between the number of fertilized Ascaris eggs
excreted and fever, in patients co-infected with Plasmodium vivax and Ascaris lumbricoides.
Ann Trop Med Parasitol. 2005;99: 623-625.
61. Gazzinelli RT, Kalantari P, Fitzgerald KA, Golenbock DT. Innate sensing of malaria
parasites. Nat Rev Immunol. 2014;14: 744-757.
62. Gomes MD, Vieira JL, Cassiano GC, Musset L, Legrand E, Nacher M, et al. Evaluation
of circumsporozoite protein of Plasmodium vivax to estimate its prevalence in Oiapoque,
Amapá state, Brazil, bordering French Guiana. Rev Inst Med Trop Sao Paulo. 2016;58: 72.
63. Nacher M, Singhasivanon P, Yimsamran S, Manibunyong W, Thanyavanich N, Wuthisen
R, et al. Intestinal helminth infections are associated with increased incidence of Plasmodium
falciparum malaria in Thailand. J Parasitol. 2002;88: 55-58.

90
64. Yatich NJ, Yi J, Agbenyega T, Turpin A, Rayner JC, Stiles JK, et al. Malaria and
intestinal helminth co-infection among pregnant women in Ghana: prevalence and risk
factors. Am J Trop Med Hyg. 2009;80: 896-901.
65. Degarege A, Animut A, Legesse M, Erko B. Malaria severity status in patients with soil-
transmitted helminth infections. Acta Trop. 2009;112: 8-11.
66. Struik SS, Riley EM. Does malaria suffer from lack of memory? Immunol Rev.
2004;201: 268-290.
67. Mueller I, Galinski MR, Baird JK, Carlton JM, Kochar DK, Alonso PL, et al. Key gaps in
the knowledge of Plasmodium vivax, a neglected human malaria parasite. Lancet Infect Dis.
2009;9: 555-566.
68. Ribeiro BP, Cassiano GC, de Souza RM, Cysne DN, Grisotto MA, Santos APA, et al.
Polymorphisms in Plasmodium vivax circumsporozoite protein (CSP) influence parasite
burden and cytokine balance in a pre-Amazon endemic area from Brazil. PLoS Negl Trop
Dis. 2016;10: e0004479.
69. Lyke KE, Burges R, Cissoko Y, Sangare L, Dao M, Diarra I, et al. Serum levels of the
proinflammatory cytokines interleukin-1 beta (IL-1beta), IL-6, IL-8, IL-10, tumor necrosis
factor alpha, and IL-12(p70) in Malian children with severe Plasmodium falciparum malaria
and matched uncomplicated malaria or healthy controls. Infect Immun. 2004;72: 5630-5637.
70. Baptista JL, Vanham G, Wery M, van Marck E. Cytokine levels during mild and cerebral
falciparum malaria in children living in a mesoendemic area. Trop Med Int Health. 1997;2:
673-679.
71. Sortica VA, Cunha MG, Ohnishi MD, Souza JM, Ribeiro-Dos-Santos AK, Santos NP, et
al. IL1B, IL4R, IL12RB1 and TNF gene polymorphisms are associated with Plasmodium
vivax malaria in Brazil. Malar J. 2012;11: 409.
72. Rodrigues-da-Silva RN, Lima-Junior JC, Fonseca BP, Antas PR, Baldez A, Storer FL, et
al. Alterations in cytokines and haematological parameters during the acute and convalescent
phases of Plasmodium falciparum and Plasmodium vivax infections. Mem Inst Oswaldo Cruz.
2014;109: 154-162.
73. Akanmori BD, Kurtzhals JA, Goka BQ, Adabayeri V, Ofori MF, Nkrumah FK, et al.
Distinct patterns of cytokine regulation in discrete clinical forms of Plasmodium falciparum
malaria. Eur Cytokine Netw. 2000;11: 113-118.
74. O'Shea JJ, Paul WE. Mechanisms underlying lineage commitment and plasticity of helper
CD4+ T cells. Science. 2010;327: 1098-1102.
75. Sarangi A, Mohapatra PC, Dalai RK, Sarangi AK. Serum IL-4, IL-12 and TNF-alpha in
malaria: a comparative study associating cytokine responses with severity of disease from the
Coastal districts of Odisha. J Parasit Dis. 2014;38: 143-147.
76. Freitas do Rosario AP, Langhorne J. T cell-derived IL-10 and its impact on the regulation
of host responses during malaria. Int J Parasitol. 2012;42: 549-555.

91
77. Da Costa AG, Antonelli LR, Costa PA, Pimentel JP, Garcia NP, Tarrago AM, et al. The
robust and modulated biomarker network elicited by the Plasmodium vivax infection is
mainly mediated by the IL-6/IL-10 axis and is associated with the parasite load. J Immunol
Res. 2014;2014: 318250.
78. Couper KN, Blount DG, Riley EM. IL-10: the master regulator of immunity to infection.
J Immunol. 2008;180: 5771-5777.
79. Medina TS, Costa SP, Oliveira MD, Ventura AM, Souza JM, Gomes TF, et al. Increased
interleukin-10 and interferon-gamma levels in Plasmodium vivax malaria suggest a reciprocal
regulation which is not altered by IL-10 gene promoter polymorphism. Malar J. 2011;10: 264.

92
4 CONCLUSÃO
- Na população estudada, o parasitismo intestinal acomete indivíduos de ambos os sexos e
tem relação com a faixa etária, nível de escolaridade, aspectos financeiros e as atividades de
garimpagem;
- O Ascaris lumbricoides foi o helminto mais frequente e a Entamoeba histolytica/díspar foi o
protozoário patogênico mais predominante;
- Observa-se uma relação inversamente proporcional entre o nível de hemoglobina e as
parasitoses detectadas, bem como correlação significante na modalidade de parasitismo;
- Os achados coproparasitológicos demonstram uma prevalência diversa entre os grupos,
salientando as particularidades e aspectos geográficos relacionados aos condicionantes e
determinantes de saúde na área de fronteira;
- A malária vivax é endêmica na região estudada e a coinfecção com enteroparasitoses está
presente no município;
- Há predomínio da população masculina entre os indivíduos maláricos, coinfectados e dos
não infectados com malária e enteroparasitoses;
- Os níveis de hemoglobina dos indivíduos parece estar fortemente atribuída a infecção
malárica, principalmente no contexto de infecções simultâneas;
- Prevalência elevada de anticorpos contra PvMSP-119 potencializa a premissa de que a
maioria dos indivíduos são expostos a malária, configurando um importante fator de
manutenção da transmissão na localidade;
- Observa-se que o perfil de citocinas inflamatórias e anti-inflamatórias pode influenciar no
perfil da coinfecção entre as duas parasitoses investigadas.

93
REFERÊNCIA
ABANYIE, F.A., MCCRACKEN, C., KIRWAN, P., MOLLOY, S.F., ASAOLU, S.O.,
HOLLAND, C.V., GUTMAN, J., LAMB, T.J. Ascaris co-infection does not alter
malaria-induced anaemia in a cohort of Nigerian preschool children. Malaria Journal
12 (1): 2-8, 2013.
ADEGNIKA, A.A., KREMSNER, P.G. Epidemiology of malaria and helminth interaction: a
review from 2001 to 2011. Curr Opin HIV AIDS 7(3): 221-4, 2012.
ALECRIM, M.G., LACERDA, M.V., MOURÃO, M.P., ALECRIM, W.D., PADILHA, A.,
CARDOSO, B.S., BOULOS, M. Successful treatment of Plasmodium falciparum
malaria with a six- dose regimen of artemether-lumefantrine versus quinine-doxycycline
in the Western Amazon region of Brazil. American journal of tropical medicine and
hygiene 74 (1): 20-25, 2006.
ALLEN, J.E, MAIZELS, R.M. Diversity and dialogue in immunity to helminths. Nat Rev
Immunol. 11 (6): 375-88, 2011.
ANA. Agência Nacional de Águas. Totais pluviométricos de longo período na Sub - bacia
30 - do rio Oiapoque ao rio Araguari. Disponível em: <http://hidroweb.ana.gov.br/.>
Acesso em Agosto de 2015.
ANDRADE, B.B., REIS-FILHO, A., SOUZA-NETO, S.M., CLARÊNCIO, J., CAMARGO,
L.M.A., BARRAL, A., BARRAL-NETTO, M. Severe Plasmodium vivax malaria
exhibits marked inflammatory imbalance. Malaria Journal 9: 9-13, 2010.
ANDRADE, R.F. Malária e migração no Amapá: projeção espacial num contexto de
crescimento populacional. Tese (Doutorado em Desenvolvimento Sócio-Ambiental,
Núcleo de Altos Estudos Amazônicos) - Belém, Universidade Federal do Pará, 2005.
ANSTEY, N.M., RUSSELL, B., YEO, T.W., PRICE, R.N. The pathophysiology of vivax
malaria. Trends Parasitol. 25(5): 220-27, 2009.
ARAUJO, C.F., FERNÁNDEZ, C.L. Prevalence of intestinal parasitosis in the city of
Eirunepé, Amazon. Revista Sociedade Brasileira Medicina Tropical 38 (1): 69, 2005.
ARÉVALO-HERRERA, M., ROSERO, F., HERRERA, S., CASPERS, P. ROTMANN, D.,
SINIGAGLIA, F., CERTA, U. Protection against malaria in Aotus monkeys immunized
with a recombinant blood-stage antigen fused to a universal T-cell epitope: correlation
of serum gamma interferon levels with protection. Infect Immun 60 (1): 154-158,
1992.
ARÉVALO-HERRERA, M., CHITNIS. C., HERRERA, S. Current status of Plasmodium
vivax vaccine. Hum Vaccin 6 (1): 124-32, 2010.
ARÉVALO-HERRERA, M., SOLARTE, Y., MARIN, C., SANTOS, M., CASTELLANOS,
J., BEIER, J.C., HERRERA, S.V. Malaria transmission blocking immunity and sexual

94
stage vaccines for interrupting malaria transmission in Latin America. Memórias do
Instituto Oswaldo Cruz 106 (1): 202-11, 2011.
ARÉVALO-HERRERA, M., QUINONES, M.L., GUERRA, C. Malaria in selected non-
Amazonian countries of Latin America. Acta Tropica 121 (3): 303-314. 2012.
ARÉVALO-HERRERA, M., VALLEJO, A.F., RUBIANO, K., SOLARTE, Y., MARIN, C.,
CASTELLANOS, A., CÉSPEDES, N., HERRERA, S. Recombinant Pvs48/45 Antigen
Expressed in E. coli Generates Antibodies that Block Malaria Transmission in
Anopheles albimanus Mosquitoes. Plos One 3 (10): 1-16, 2015.
BARNWELL, J.W., NICHOLS, M.E., RUBINSTEIN, P. In vitro evaluation of the role of the
Duffy blood group in erythrocyte invasion by Plasmodium vivax. J. Exp. Med.
169(5):1795-802, 1989.
BARBEDO, M.B., RICCI, R., JIMENEZ, M.C.S., CUNHA, M.G., YAZDANI, S.S.,
CHITNIS, C.E., RODRIGUES, M.M., SOARES, I.S. Comparative recognition by
human IgG antibodies of recombinant proteins representing three asexual erythrocytic
stage vaccine candidates of Plasmodium vivax. Memorias Instituto Oswaldo Cruz
102: 335-9, 2007.
BARBIERI, AF. Uso do solo e prevalência de malária em uma região da Amazônia
Brasileira. Cadernos de Geografia 15 (24): 9-30, 2005.
BARBIERI, A.F., SAWYER, D.O. Heterogeneity of malaria prevalence in alluvial gold
mining areas in Northern Mato Grosso State, Brazil. Cad. Saúde Pública 23 (12):
2878-2886, 2007.
BEJON, P., MWANGI, T.W., LOWE, B., PESHU, N., ADRIAN, V.S.H., MARSH, K.
Helminth infection and eosinophilia and the risk of Plasmodium falciparum malaria in
1- to 6- year-old children in a malaria endemic area. PLoS Negl Trop Dis 6 (2): 1-6,
2008.
BELLOTO, M.V.T., JUNIOR, J.E.S., MACEDO, E.A., PONCE, A., GALISTEU, K.J.,
CASTRO, E., TAUYR, L.V., ROSSIT, A.R.B., MACHADO, R.L.D. Enteroparasitoses
numa população de escolares da rede pública de ensino do Município de Mirassol, São
Paulo, Brasil. Revista Pan-Amazônica de Saúde, 2 (1): 37-44, 2011.
BOEL, M., CARRARA, V.I., RIJKEN, M., PROUX, S., NACHER, M., PIMANPANARAK,
M., PAW, M.K., MOO, O., GAY, H., BAILEY, W., SINGHASIVANON, P., WHITE,
N.J., NOSTEN, F., MCGREADY, R. Complex Interactions between soil-transmitted
helminths and malaria in pregnant women on the Thai-Burmese border. PLoS Negl
Trop Dis 16 (11): 887, 2010.
BOEF, A.G., MAY, L., VAN BODEGOM D., VAN LIESHOUT, L., VERWEIJ, J.J.,
MAIER, A.B., WESTENDORP, R.G., ERIKSSON, U.K. Parasitic infections and
immune function: Effect of helminth infections in a malaria endemic area.
Immunobiology 218 (5) 706-11, 2012.

95
BOUTLIS, C.S., YEO, T.W., ANSTEY, N.M. Malaria tolerance-forwhom the cell tolls?
Trends in Parasitology 22: 371-377, 2006.
BUENO LL, MORAIS CG, LACERDA MV, FUJIWARA RT, BRAGA EM. Interleukin-17
producing T helper cells are increased during natural Plasmodium vivax infection. Acta
Trop. 123 (1): 53-57, 2012.
BRASIL. Ministério da Saúde. Secretaria de Vigilância em Saúde. Plano Nacional de
Vigilância e Controle das Enteroparasitoses. Brasília: MS, 2005.
BRASIL. Ministério da Saúde. Secretaria de Vigilância em Saúde. Boletim Epidemiológico.
Situação epidemiológica da malária no Brasil, 2000 a 2011. Brasília: MS, v. 44, n°1,
2013.
BRASIL. Ministério da Saúde. Guia de Vigilância em Saúde. Brasília: MS, 2014.
BRASIL. Ministério da Saúde, Sistema de Informação de Vigilância Epidemiológica,
SIVEP-Malária. Brasília: MS, 2015.
BRAZ, R.M., DUARTE, E.C., TAUIL, P.L. Epidemiology of malaria in the municipality of
Cruzeiro do Sul State of Acre Brazil in 2010: uses of a control chart at the local level.
Rev Soc Bras Med Trop 45 (4): 526-9, 2012.
BRAZ, R.M., DUARTE, E.C., TAUIL, P.L. Caracterização das epidemias de malária nos
municípios da Amazônia Brasileira em 2010. Caderno Saúde Pública, 29 (5): 935-44,
2013.
BRAZ, R.M., DUARTE, E.C., TAUIL, P.L. Algorritmo para monitoramento da incidência da
malária na Amazônia Brasileira, 2003 a 2010. Revista Panamericana Saúde Pública 3
(3): 186-92, 2014.
BROOKER, S., CLEMENTS, A.C., BUNDY, D.A. Global epidemiology, ecology and
control of soil-helminth infections. Adv Parasitol 62: 221-61. 2006a.
BROOKER, S., CLEMENTS, A.C.A., HOTEZ, P.J., HAY, S.I., TATEM, A.J., BUNDY,
D.A.P., SNOW, R.W. The co-distribution of Plasmodium falciparum and hookworm
among African school children. Malaria Journal 3 (5): 99, 2006b.
BROOKER, S., AKHWALE, W., PULLAN, R., ESTAMBALE, B., CLARKE, S.E., SNOW,
R.W., HOTEZ, P.J. Epidemiology of Plasmodium-helminth co-infection in Africa:
populations at risk, potential impact on anemia, and prospects for combining control.
American Journal of Tropical Medicine and Hygiene 77 (6): 88-98, 2007.
BRUTUS, L., WATIER, L., BRIAND, V., HANITRASOAMAMPIONONA, V.,
RAZANATSOARILALA, H., COT, M. Parasitic co-infections: does Ascaris
lumbricoides protect against Plasmodium falciparum infection? American journal of
tropical medicine and hygiene 75 (2): 194-8, 2006.
CABANTOUS, S., POUDIOUGOU, B., OUMAR, A.A., TRAORE, A., BARRY, A., VITTE,
J., BONGRAND, P., MARQUET, S., DOUMBO, O., DESSEIN, A.J. Genetic evidence

96
for the aggravation of Plasmodium vivax malaria by interleukin 4. Journal of
Infectious Diseases 200 (10): 1530-1539, 2009.
CARDOSO, R.F., GOLDENBERG, P. Malária no Estado do Amapá, Brasil, de 1970 a 2003:
trajetória e controle. Caderno Saúde Pública 23 (6): 1339-48, 2007.
CARDOSO, R.F. Efeito da Sazonalidade na Curva Endêmica da Malária por
Plasmodium falciparum e vivax no Garimpo do Lourenço: Uma Série Temporal
Histórica na Zona da Amazônia Brasileira. Tese (Doutorado em Ciências Médicas) -
Porto Alegre, Universidade Federal do Rio Grande do Sul, Faculdade de Medicina,
2014.
CASTRO, G.T., MOURÃO, L.C., MELO, G.C., MONTEIRO, W.M., LACERDA, M.V.,
BRAGA, É.M. Potential Immune Mechanisms Associated with Anemia in Plasmodium
vivax Malaria: a Puzzling Question. Infection and Immunity 82: 3990-4000, 2014.
CALDERARO, A., MONTECCHINI, S., ROSSI, S., GORRINI, C., CONTO, F., MEDICI,
M.C., CHEZZI, C., ARCANGELETTI, M.C. Intestinal parasitoses in a tertiary-care
hospital located in a non-endemic setting during 2006-2010. BMC Infectious Diseases
14 (1): 264, 2014.
CESÁRIO, M., CESÁRIO, R.R. Malária, Amazônia e Desenvolvimento: migração,
degradação ambiental e doenças infecciosas caminham juntas na Amazônia. Scientific
American Brasil 4 (46), 54-55, 2006.
COIMBRA JR, C.E.A., SANTOS, R.V. Saúde, minorias e desigualdade: Algumas teias de
inter-relações, com ênfase nos povos indígenas. Ciência & Saúde Coletiva 5 (1):125-
132, 2000.
COIMBRA JR, C.E.A., SANTOS, R.V., ESCOBAR, A.L., orgs. Epidemiologia e saúde dos
povos indígenas no Brasil. Rio de Janeiro: Editora FIOCRUZ; ABRASCO, 2005.
260p. Disponível em: <http://static.scielo.org/scielobooks/bsmtd/pdf/coimbra-
9788575412619.pdf> Acesso em 01 de Junho de 2017.
CONFALONIERI, U.E.C. Saúde na Amazônia: um modelo conceitual para a análise de
paisagens e doenças. Estudos avançados 19 (53): 221-36, 2005.
COURTIN, D., DJILALI-SAÏAH, A., MILET, J., SOULARD, V., GAYE, O., MIGOT-
NABIAS, F., SAUERWEIN, R., GARCIA, A., LUTY, A.J. Schistosoma haematobium
infection affects Plasmodium falciparum-specific IgG responses associated with
protection against malaria. Parasite Immunology 33 (2): 124-31, 2011.
COX, F.E. Concomitant infections, parasites and immune responses. Parasitology. 122: 23-
38, 2001.
CUNHA, M.G., RODRIGUES, M.M., SOARES, I.S. Comparison of the immunogenic
properties of recombinant proteins representing the Plasmodium vivax vaccine
candidate MSP119 expressed in distinc bacterial vectors. Vaccine 20: 385-396, 2001.

97
CHAORATTANAKAWEE, S., NATALENG, O., HANANANTACHAI, H., NACHER, M.,
BROCKMAN, A., NOSTEN, F., LOOAREESUWAN, S., PATARAPOTIKUL, J.
Trichuris trichiura infection is associated with the multiplicity of Plasmodium
falciparum infections, in Thailand. Ann Trop Med Parasitol. 97 (2): 199-202, 2003.
CHAMMARTIN, F., GUIMARÃES, L.H., SCHOLTE, R.G., BAVIA, M.E., UTZINGER, J.,
VOUNATSOU, P. Spatio-temporal distribution of soil-transmitted helminth infections
in Brazil. Parasit Vectors. 18;7:440. 2014.
CHAPPEL, J.A., HOLDER, A.A. Monoclonal antibodies that inhibit Plasmodium falciparum
invasion in vitro recognise the first growth factor-like domain of merozoite surface
protein-1. Mol Biochem Parasitol. 60: 303-311, 1993.
DAVID, T.C., MACEDO, L.C., SÁ, F.M.P., JÚNIOR, M.P.S. Prevalência de enteroparasitos
no município de Ariquemes, Rondônia, BRASIL. Revista Cientifica da Faculdade de
Educação e Meio Ambiente 4 (2): 39-48, 2013.
DEGAREGE, A., LEGESSE, M., MEDHIN, G., ANIMUT, A., ERKO, B. Malaria and
related outcomes in patients with intestinal helminths: a cross-sectional study. BMC
Infect Dis. 9 (12): 291, 2012.
DENT, A.E., CHELIMO, K.S., PETER, O., SPRING, M.D. CRABB, S. BRENDAN, M.,
ANN. M., TISCH, D.J., KAZURA, J.W. Temporal stability of naturally acquired
immunity to Merozoite Surface Protein-1 in Kenyan Adults. Malaria Journal 8
(162): 1-10, 2009.
DÍAZ, A., ALLEN, J.E. Mapping immune response profiles: The emerging scenario from
helminth immunology. European Journal of Immunology 37 (12): 3319-26, 2007.
DODOO, D., OMER, F.M., TODD, J., AKANMORI, B.D., KORAM, K.A., RILEY, E.M.
Absolute levels and ratios of proinflammatory and anti-inflammatory cytokine
production in vitro predict clinical immunity to Plasmodium falciparum malaria. J
Infect Dis. 185(7): 971-79, 2002.
DOOLAN, D.L., DOBAÑO, C., BAIRD, J.K. Acquired immunity to malaria. Clinical
Microbiology Reviews 22 (1): 13-36, 2009.
DOTRÁRIO, A.B., MENON, L.J.B., BOLLELA, V.R., MARTINEZ,R., ARAÚJO, D.C.A.,
FONSECA, B.A.L., SANTANA, R.C. Malaria and other febrile diseases among
travellers: the experience of a reference centre located outside the Brazilian Amazon
Region. Malaria Journal 15 (1): 294, 2016.
DRUILHE, P., TALL, A. Worms can worsen malaria: towards a new means to roll back
malaria?. Trends in Parasitology 21: 359-362, 2005.
DYER, M., DAY, K.P. Commitment to gametocytogenesis in Plasmodium falciparum.
Parasitol Today 16 (3): 102-7, 2000.

98
EGAN, A.F., BLACKMAN, M.J., KASLOW, D.C. Vaccine efficacy of recombinant
Plasmodium falciparum merozoite surface protein 1 in malaria-naive-exposed, and/or -
rechallenged Aotus vociferans monkeys. Infect Immun. 68: 1418-27, 2000.
FAUBERT, G. Immune Response to Giardia duodenalis. Clin Microbiol Rev. 13 (1): 35-54,
2000.
FERREIRA, M.U., DA SILVA, N.S., DA SILVA-NUNES, M., MALAFRONTE, R.S.,
MENEZES, M.J., D'ARCADIA, R.R., KOMATSU, N.T., SCOPEL, K.K.G., BRAGA,
E.M., CAVASINI, C.E., CORDEIRO, J.A. Epidemiology and control of frontier
malaria in Brazil: lessons from community-based studies in rural Amazonia.
Transactions of the Royal Society of Tropical Medicine and Hygiene, 104, (5): 343-
50, 2010.
FIGUEIREDO, C.A. BARRETO, M.L. RODRIGUES, L.C. COOPER, P.J., SILVA, N.B.,
AMORIM, L.D., ALCANTARA-NEVES, N.M. Chronic intestinal helminth infections
are associated with immune hyporesponsiveness and induction of a regulatory network.
Infection and Immunity. 78 (7): 3160-3167, 2010.
GALARDO, A.K., ZIMMERMAN, R., GALARDO, C.D. Larval control of Anopheles
(Nyssorhinchus) darlingi using granular formulation of Bacillus sphaericus in
abandoned gold-miners excavation pools in the Brazilian Amazon rainforest. Revista
Sociedade Brasileira Medicina Tropical 46 (2): 172-7, 2013.
GALINSKI, M.R., BARNWELL, J.W. Plasmodium vivax: Merozoites, invasion of
reticulocytes and considerations for malaria vaccine development. Parasitol. Today
12(1):20-9, 1996.
GEA. Governo do Estado do Amapá. Município de Oiapoque. Disponível em
<http://www4.ap.gov.br/Portal_Gea/municipios/municipio-oiapoque.htm>. Acesso em
15 de maio de 2016.
GEIGER, S.M. MASSARA, C.L. BETHONY, J. SOBOSLAY, P.T. CARVALHO, O. S.,
CORRÊA-OLIVEIRA, R. Cellular responses and cytokine profiles in Ascaris
lumbricoides and Trichuris trichiura infected patients. Parasite Immunology 24: 499-
509, 2002.
GOMES, M.S.M., VIEIRA, J.L.F., MACHADO, R.L.D., NACHER, M., STEFANI, A.,
MUSSET, L., LEGRAND, E., MENEZES, R.A.O., JÚNIOR, A.A.P., SOUSA, A.P.M.,
COUTO, V.S.C.A., COUTO, A.A.R.A. Efficacy in the treatment of malaria by
Plasmodium vivax in Oiapoque, Brazil, on the border with French Guiana: the
importance of control over external factors. Malaria Journal 12: 402, 2015.
GOMES, M.S.M., VIEIRA, J.L.F., CASSIANO, G.C., MUSSET, L., LEGRAND, E.,
NACHER, M., COUTO, V.S.C.A., MACHADO, R.L.D., COUTO, A.A.R.A.
Evaluation of circumsporozoite protein of Plasmodium vivax to estimate its prevalence
in Oiapoque, Amapá state, Brazil, bordering French Guiana. Rev. Inst. Med. Trop. Sao
Paulo 28 (72): 2-4, 2016.

99
GONCALVES, R.M., SALMAZI, K.C., SANTOS, B.A.N., BASTOS, M.S., ROCHA, S.C.,
BOSCARDIN, S.B., SILBER, A.M., KALLÁS, E.G., FERREIRA, M.U., SCOPEL,
K.K.G. CD4_ CD25_ Foxp3_ Regulatory T Cells, Dendritic Cells, and Circulating
Cytokines in Uncomplicated Malaria: Do Different Parasite Species Elicit Similar Host
Responses?. Infect Immun 78 (11): 4763-4772, 2010.
GOOD, M.F. Vaccine-induced immunity to malaria parasites and the need for novel
strategies. Trends in Parasitology 21 (1): 29-34, 2005.
GOOD, M.F., XU, H., WYKES, M., ENGWERDA, C.R. Development and regulation of
cellmediated immune responses to the blood stages of malaria: implications for vaccine
research. Annu. Rev. Immunol. 23 (1):69-99, 2005.
GREENWOOD, B.M., FIDOCK, D.A., KYLE, D.E., KAPPE, S.H., ALONSO, P.L.,
COLLINS, F.H., DUFFY, P.E. Malaria: progress, perils, and prospects for eradication.
The Journal of Clinical Investigation 118 (4)1266-1276, 2008.
HAGEL, I., LYNCH, N.R., PEREZ, M., DI PRISCO, M.C., LOPEZ, R., ROJAS, E.
Modulation of the allergic reactivity of slum children by helminthic infection. Parasite
Immunology 15 (6): 311–315, 1993.
HAILEGEBRIEL, T. Prevalence of intestinal parasitic infections and associated risk factors
among students at Dona Berber primary school, Bahir Dar, Ethiopia. BMC Infectious
Diseases 17 (1): 362, 2017. HALL, A., HEWITT, G., TUFFREY, V., SILVA, N. A review and meta-analysis of the
impact of intestinal worms on child growth and nutrition. Maternal and Child
Nutrition, 4 (1): 118-236, 2008.
HARHAY, M.O., HORTON, J., OLLIARO, P.L. Epidemiology and control of human
gastrointestinal parasites in children. Expert Review of Anti-infective Therapy 8 (2):
219-34, 2010.
HARTGERS, F.C., YAZDANBAKHSH, M. Co-infection of hel minths and malaria:
modulation of the immune responses to malaria. Parasite Immunology 28 (10): 497-
506, 2006.
HERMES, S.C.M.N., NUNES, V.L.B., DORVAL, B.A.F. Aspectos Epidemiológicos da
malária humana no município de Aripuanã, Estado do Mato Grosso, Brasil, 2005 a
2010. Revista Brasileira de Geografia Médica e da Saúde - HYGEIA 9 (17): 42 - 51,
2013.
HERRERA, S., CORRADIN, G., ARÉVALO-HERRERA, M. An update on the search for a
Plasmodium vivax vaccine. Trends in Parasitology 23 (3): 122-28, 2007.
HILL, A.V.S. Vaccines against malaria. Philos Trans R Soc Lond B Biol Sci.
366(1579):2806-14, 2011.
HERRERA, S., VALLEJO, A.F., QUINTERO, J.P., ARÉVALO-HERRERA, M.,
CANCINO, M., FERRO, S. Field evaluation of an automated RDT reader and data

100
management device for Plasmodium falciparum/Plasmodium vivax malaria in endemic
areas of Colombia. Malaria Journal 13 (87): 1-10, 2014.
HODDER, A.N., CREWTHER, P.E., ANDERS, R.F. Specificity of the protective antibody
response to apical membrane antigen 1. Infection and Immunity 69 (5): 3286-3294.
2001.
HOLDER, A.A. The precursor to major merozoite surface antigens: structure and role in
immunity. Prog Allergy 41: 72-97, 1988.
HOLDER, A.A. BLACKMAN, M.J., BURGHAUS, P.A., CHAPPEL, J.A., LING, I.T.,
MCCALLUM-DEIGHTON, N., SHAI, S. A malaria merozoite surface protein (MSP1)-
structure, processing and function. Mem Inst Oswaldo Cruz. 87 (3): 37-42, 1992.
HOTEZ, P.J., BRINDLEY, P.J., BETHONY, J.M., KING, C.H., PEARCE, E.J.,
JACOBSON, J. Helminth infections: the great neglected tropical diseases. The Journal
of Clinical Investigation, 118 (4): 1311-21, 2008.
HUANG, D.B., WHITE, A.C. An Updated Review on Criptosporidium and Giardia.
Gastroenterol Clin North Am 35 (2): 291-314, 2006.
HULDÉN, L., HULDÉN, L., HELIÖVAARA, K. Natural relapses in vivax malaria induced
by Anopheles mosquitoes. Malaria Journal 7 (64): 1-11, 2008.
INTITUTO BRASILEIRO DE GEOGRAFIA E ESTATISTICA. Censo Demográfico
Brasileiro. Características da população e dos domicílios: Resultados do Universo.
Amapá: IBGE, Brasília, 2010.
IANELLI, R.V. Epidemiologia da malária em populações indígenas da Amazônia. In:
Doenças Endêmicas: Abordagens Sociais, Culturais e Comportamentais (R.B.
Barata & R.Briceño-León, orgs.), pp. 355-374. Rio de Janeiro: Editora Fiocruz. 2000.
JAIN, V., SINGH, P.P. SILAWAT, N., PATEL, R., SAXENA, A., BHARTI, P.K.,
SHUKLA, M., BISWAS, S., SINGH, N. A preliminary study on pro- and anti-
inflammatory cytokine profiles in Plasmodium vivax malaria patients from central zone
of India. Acta Trop 113 (3): 263-268, 2010.
KABYEMELA, E., GONÇALVES, B.P., PREVOTS, D.R., MORRISON, R.,
HARRINGTON, W., GWAMAKA, M., KURTIS, J.D., FRIED, M., DUFFY, P.E.
Cytokine profiles at birth predict malaria severity during infancy. PLoS One. 8(10): 1-
8, 2013.
KNOWLES, S.C.L. The effect of helminth co-infection on malaria in mice: a meta-analysis.
International Journal for Parasitology 15 (41): 1041-51. 2011.
KUBIAK, K., DZIKA, E., DOBRZAŃSKA, M., LEPCZYŃSKA, M., KORYCIŃSKA, J.,
THOENE, M., DZISKO, J. Giardiasis in the Warmia and Mazury province (north-
eastern Poland) - an epidemiological analysis. Polish Annals of Medicine 24 (1): 5-8,
2017.

101
KUCIK, C., MARTIN, G., SORTOR, B. Common intestinal parasites. American Family
Physician, 69 (5): 1161-8, 2004.
LACERDA, M.V.G., HIPÓLITO, J.R., LENY, N.M. Chronic Plasmodium vivax infection in
a patient with splenomegaly and severe thrombocytopenia. Rev Soc Bras Med Trop 41
(5): 522-523, 2008.
LACERDA, M.V.G., FRAGOSO, S.C.P., ALECRIM, M.G.C., ALEXANDRE M.A.A.,
MAGALHÃES, B.M.L., SIQUEIRA, A.M., FERREIRA, L.C., ARAÚJO, J.R.,
MOURÃO, M.P., FERRER, M., CASTILLO, P., MARTIN-JAULAR, L.,
FERNANDEZ-BECERRA, C., DEL PORTILLO, H., ORDI, J., ALONSO, P.L.,
BASSAT, Q. Postmortem characterization of patients with clinical diagnosis of
Plasmodium vivax malaria: to what extent does this parasite kill?. Clin. Infect. Dis. 55
(8): 67-74, 2012.
LANGHORNE, J., NDUNGU, F.M., SPONAAS, A.M., MARSH, K. Immunity to malaria:
more questions than answers. Nat Immunol. 9(7):725-32, 2008.
LEDER, K., WELLER, P.F. Eosinophilia and helminthic infections. Best Practice &
Research Clinical Haematology. 13 (2): 301-17, 2000.
LE HESRAN, J.Y., AKIANA, J., NDIAYE, E.H.M., DIA, M., SENGHOR, P., KONATE, L.
Severe malaria attack is associated with high prevalence of Ascaris lumbricoides
infection among children in rural Senegal. Transactions of the Royal Society of
Tropical Medicine and Hygiene 98 (7): 397-9, 2004.
LEVINO, A., CARVALHO, E.F. Análise comparativa dos sistemas de saúde da tríplice
fronteira: Brasil/Colômbia/Peru. Rev Panam Salud Publica 30(5): 490-500. 2011.
LODO, M., OLIVEIRA, C.G.B., FONSECA, A.L.A., CAPUTTO, L.Z., PACKER, M. L.T.,
VALENTI, V.E., FONSECA, F.L.A. Prevalência de enteroparasitos em município do
interior paulista. Revista Brasileira de Crescimento e Desenvolvimento Humano 20
(3): 769-777, 2010.
LYKE, K.E., DICKO, A., DABO, A., SANGARE, L., KONE, A., COULIBALY, D.,
GUINDO, A., TRAORE, K., DAOU, M., DIARRA, I., SZTEIN, M.B., PLOWE, C.V.,
DOUMBO, O.K. Association of Schistosoma haematobium infection with protection a
gainst acute Plasmodium falciparum malaria in Malian children. The American
Journal of Tropical Medicine and Hygiene 73 (6): 1124-30, 2005.
MAIZELS, R.M., YAZDANBAKHSH, M. Immune regulation by helminth parasites: cellular
and molecular mechanisms. Nature Reviews Immunology 3 (9): 733-44. 2003.
MAIZELS, R.M. BALIC, A. GOMEZ-ESCOBAR, N. NAIR, M. TAYLOR, M.D. ALLEN,
J.E. Helminth parasites-masters of regulation. Immunological Reviews 201 (1): 89-
116, 2004.
MAHMUD, M. A., SPIGT, M., BEZABIH, A.M., PAVON, I.L., DINANT, G.J., VELASCO,
R.B. Risk factors for intestinal parasitosis, anaemia, and malnutrition among school
children in Ethiopia. Pathogens and Global Health 107 (2): 58-65, 2013.

102
MEDINA, T. S., COSTA, S.P.T., OLIVEIRA, M.D., VENTURA, A.M., SOUZA, J.M.,
GOMES, T. F., VALLINOTO, A.C.R., PÓVOA, M.M., SILVA, J.S., CUNHA, M.G.
Increased interleukin-10 and interferon- levels in Plasmodium vivax malaria suggest a
reciprocal regulation which is not altered by IL-10 gene promoter polymorphism.
Malaria Journal 14 (10): 2-10, 2011.
MEEUSEN, E.N., BALIC, A. Do eosinophils have a role in the killing of helminth parasites?
Parasitol Today. 16 (3):95-101, 2000.
MELO, G.C., REYES-LECCA, R.C., VITOR-SILVA, S., MONTEIRO, W.M., MARTINS,
M., BENZECRY, S.G., ALECRIM, M.D., LACERDA, M.V. Concurrent helminthic
infection protects schoolchildren with Plasmodium vivax from anemia. PLoS One 5
(6): 11206, 2010.
MELO MCB, KLEM VGQ, MOTA JAC, PENNA FJ. Parasitoses intestinais. Rev Med 14:3-
12, 2004.
MENDONÇA, V.R.R., QUEIROZ, A.T.L., LOPES, F.M., ANDRADE, B.B., BARRAL-
NETTO, M. Networking the host immune response in Plasmodium vivax malaria.
Malaria Journal. 12(69): 1-10, 2013.
MIDZI, N., MTAPURI-ZINYOWERA, S., MAPINGURE, M.P., SANGWEME, D.,
CHIREHWA, M.T., BROUWER, K.C., MUDZORI, J., HLEREMA, G., MUTAPI, F.,
KUMAR, N., MDULUZA, T. Consequences of polyparasitism on anaemia among
primary school children in Zimbabwe. Acta Tropica 115 (1-2): 103-11, 2010.
MIDZI, N., MTAPURI-ZINYOWERA, S., SANGWEME, D., PAUL, N.H., MAKWARE,
G., MAPINGURE, M.P., BROUWER, K.C., MUDZORI, J., HLEREMA, G.,
CHADUKURA, V., MUTAPI, F., KUMAR, N., MDULUZA, T. Efficacy of integrated
school based de-worming and prompt malaria treatment on helminths -Plasmodium
falciparum co-infections: A 33 months follow up study. BMC Int Health Hum Rights
11 (9): 2-14, 2011.
MONS, B., COLLINS, W.E., SKINNER, J.C., VAN DER STAR W., CROON, J.J., VAN
DER KAAY, H.J. Plasmodium vivax: in vitro growth and reinvasion in red blood cells
of Aotus nancymai. Exp. Parasitol. 66(2):183-8, 1988.
MORENIKEJI, O., AZUBIKE, N., IGE, A. Prevalence of intestinal and vector-borne urinary
parasites in communities in south-west Nigeria. J Vector Borne Dis, 46 (2): 164-67,
2009.
MUELLER, I., GALINSKI, M.R., BAIRD, J.K., CARLTON, J.M., KOCHAR, D.K.,
ALONSO, P.L., DEL PORTILLO, H.A. Key gaps in the knowledge of Plasmodium
vivax, a neglected human malaria parasite. The Lancet Infectious Diseases 9 (9): 555-
566, 2009.
MUELLER, I., GALINSKI, M.R., TSUBOI, T., AREVALO-HERRERA, M., COLLINS,
W.E., KING, C.L. Natural Acquisition of Immunity to Plasmodium vivax:
Epidemiological Observations and Potential Targets. Adv. Parasitol 81: 77-131, 2013.

103
MUELLER, I.; SHAKRI, A. R.; CHITNIS, C. E. Development of vaccines for Plasmodium
vivax malaria. Vaccine 33 (52):7489-95, 2015.
MURRAY, J., MURRAY, A., MURRAY, M., MERRAY, C. The biological suppression of
malaria: an ecological and nutritional interrelationship of a host and two parasites. Am
J Clin Nutr 31 (8): 1363-6, 1978.
MBOERA, L.E.G., SENKORO, K.P., RUMISHA, S.F., MAYALA, B.K., SHAYO, E.H.,
MLOZI, M.R.S. Plasmodium falciparum and helminth coinfections among
schoolchildren in relation to agro-ecosystems in Mvomero District, Tanzania. Acta
Tropica 120(1-2):95-102, 2011.
MBUH, J.V., NTONIFOR, H.N., OJONGO, J.T. The incidence,intensity nad host morbidity
of human parasitic protozoan infections in gastrointestinal disorder outpatients in Buea
Sub division, Cameroom. J Infect Dev Ctries 4(1): 38-43, 2010.
NACHER, M., GAY, F., SINGHASIVANON, P., KRUDSOOD, S., TREEPRASERTSUK,
S., MAZIER, D., VOULDOUKIS, I., LOOAREESUWAN, S. Infection is associated
with protection from cerebral malaria. Parasite Immunol 22 (3): 107-13, 2000.
NACHER, M., SINGHASIVANON, P., SILACHAMROON, U., TREEPRASERTSU, S.,
KRUDSOOD, S., GAY, F., MAZIER, D., LOOAREESUWAN, S. Association of
helminth infections with increased gametocyte carriage during mild falciparum malaria
in Thailand. Am J Trop Med Hyg 65 (5): 644-7, 2001.
NACHER, M., SINGHASIVANON, P., YIMSAMRAN, S., MANIBUNYONG, W.,
THANYAVANICH, N., WUTHISEN, R., LOOAREESUWAN, S. Intestinal helminth
infections are associated with increased incidence of Plasmodium falciparum malaria in
Thailand. J Parasitol. 88 (1): 55-8, 2002.
NACHER, M. Interactions between worms and malaria: good worms or bad worms?.
Malaria Journal 12 (10): 259, 2011.
NACHER, M. Helminth-infected patients with malaria: a low profile transmission hub?
Malaria Journal 11: 376, 2012.
NACHER, M., STEFANI, A., BASURKO, C., LEMONNIER, D., DJOSSOU, F., DEMAR,
M., ELENGA, N., BROUSSE, P., VILLE, M., CARME, B. The burden of Plasmodium
vivax relapses in an Amerindian village in French Guiana. Malaria Journal 24 (12):
367, 2013.
NILSSON, S.K., CHILDS, L.M., BUCKEE, C., MARTI, M. Targeting human transmission
biology for malaria elimination. PLOS Pathog.11(6):e1004871. 2015.
NOGUEIRA, P.A., ALVES, F.P., FERNANDEZ-BECERRA, C., PEIN, O., SANTOS, N.R.,
PEREIRA, S.L.H., CAMARGO, E.P., DEL PORTILLO, H.A. A reduced risk of
infection with Plasmodium vivax and clinical protection against malaria are associated
with antibodies against the N terminus but the C terminus of merozoite surface protein
1. Infect Immun 74: 2726-33, 2006.

104
OEUVRAY, C., BOUHAROUN-TAYOUN, H., GRASS-MASSE, H., LEPERS, JP.,
RALAMBORANTO, L., TARTAR, A., DRUILHE, P. A novel merozoite surface
antigen of Plasmodium falciparum (MSP-3) identified by cellular-antibody cooperative
mechanism antigenicity and biological activity of antibodies. Memórias do Instituto
Oswaldo Cruz, 89 (2):77-80, 1994.
OJURONGBE, O., ADEGBAYI, A.M., BOLAJI, O.S., AKINDELE, A.A., ADEFIOYE,
O.A., ADEYEBA, O.A. Asymptomatic falciparum malaria and intestinal helminths
coinfection among school children in Osogbo, Nigeria. J Res Med Sci. 16 (5): 680-6,
2011.
OLIVEIRA, D., FERREIRA, F.S., ATOUGUIA, J., FORTES, F., GUERRA, A., CENTENO-
LIMA, S. Infection by Intestinal Parasites, Stunting and Anemia in School-Aged
Children from Southern Angola. PLOS ONE 10 (9): 2015.
OLIVEIRA-FERREIRA, J., LACERDA, M.V., BRASIL, P., LADISLAU, J.L., TAUIL, P.L.,
DANIEL-RIBEIRO, C.T. Malaria in Brazil: an overview. Malaria Journal 30 (9):115-
30, 2010.
OLIVEIRA-FILHO, A.B., MARTINELLI, J.M. Casos notificados de malária no Estado do
Pará, Amazônia Brasileira, de 1998 a 2006. Epidemiologia e Serviços de Saúde,
18(03): 277-284, 2009.
OLLIARO, P.L., BARNWELL, J.W., BARRY, A., MENDIS, K., MELLER, I., REEDER,
J.C., SHANKS, G.D., SNOUNOU, G., WONGSRICHANALAI, C. Implications of
Plasmodium vivax Biology for Control, Elimination and Research. The American
Journal of Tropical Medicine and Hygiene 95 (6): 4-14, 2016.
ONO, T., OHNISHI, Y., NAGAMUNE, K., KANO, M. Gametocytogenesis induction by
Berenil in cultured Plasmodium falciparum. Exp Parasitol. 77 (1): 74-8, 1993.
OSIER, F.H., FEGAN, G., POLLEY, S.D., MURUNGI, L., VERRA, F., TETTEH, K.K.,
LOWE, B., MWANGI, T., BULL, P.C., THOMAS, A.W., CAVANAGH, D.R.,
MCBRIDE, J.S., LANAR, D.E., MACKINNON, M.J., CONWAY, D.J., MARSH, K.
Breadth and magnitude of antibody responses to multiple Plasmodium falciparum
merozoite antigens are associated with protection from clinical malaria. Infect Immun.
76(5):2240-8, 2008.
OUATTARA, M., N’GUÉSSAN, N.A., YAPI, A., N’GORAN, E.K. Prevalence and Spatial
Distribution of Entamoeba histolytica/dispar and Giardia lamblia among
Schoolchildren in Agboville Area (Côte d’Ivoire). Eichinger D, ed. PLoS Neglected
Tropical Diseases 4 (1): e574. 2010. doi:10.1371/journal.pntd.0000574.
PALHARES, J. M., GUERRA, A. J. T. Potentialities in Oiapoque Municipality, Amapá
State, for the Development of Geotourism. Espaço Aberto, PPGG - UFRJ, v. 6, n.2, p.
51-72, 2016.
PARISE, E.V. Vigilância, prevenção e controle da malária em Palmas, Tocantins, Brasil, no
período de 2000 a 2013. Revista Brasileira de Geografia Médica e da Saúde -
HYGEIA 10 (18): 190 - 206, 2014.

105
PEARCE, E.J., CASPAR, P., GRZYCH, J.M., LEWIS, F.A., SHER, A. Downregulation of
Th1 cytokine production accompanies induction of Th2 responses by a parasitic
helminth, Schistosoma mansoni. J. Exp. Med. 173 (1): 159-66, 1991.
PEITER, P.C, MACHADO, L.O., IÑIGUEZ ROJAS, L. MIRANDA, A.C., BARCELLOS,
C., MOREIRA, J.C., MONKEN, M. Doenças transmissíveis na faixa de fronteira
Amazônica: o caso da malária. Território, ambiente e saúde. Rio de Janeiro: Editora
Fiocruz; 2008. p. 257-272.
PENIDO, A. Brasil registra menor número de casos de malária nos últimos 35 anos. Portal da
Saúde – Ministério da Saúde, 2016. Disponível em: <www.saude.gov.br>. Acesso em:
15/05/2016.
PITABUT, N., PANICHAKRON, J., MAHAKUNKIJ-CHAROEN, Y.,
HIRUNPETCHARAT, C. IgG antibody profile to C-terminal region of Plasmodium
vivax merozoite surface protein-1 n Thai individuals exposed to malaria. Southeast
Asia J Trop Med Public Health 38 (1): 1-7, 2007.
POLLEY, S.D., CONWAY, D.J., CAVANAGH, D.R., MCBRIDE, J.S., LOWE, B.S.,
WILLIAMS, T.N., MWANGI, T.W., MARSH, K. High levels of serum antibodies to
merozoite surface protein 2 of Plasmodium falciparum are associated with reduced risk
of clinical malaria in coastal Kenya. Vaccine. 24(19):4233-46, 2006.
PULLAN, R.L., KABATEREINE, N., BUKIRWA, H., STARDKE, S.G., BROOKER, S.
Heterogeneities and consequences of Plasmodium species and hookworm coinfection: a
population based study in Uganda. J Infect Dis. 203 (3): 406-17, 2011.
PRICE, R., NOSTEN, F., SIMPSON, J.A., LUXEMBURGER, C., PHAIPUN, L., KUILE, F.
T., VUGT, M.V., CHONGSUPHAJAISIDDHI, T., WHITE, N.J. Risk factors for
gametocyte carriage in uncomplicated falciparum malaria. Am J Trop Med Hyg 60
(6): 1019-23, 1999.
ROUSSILHON, C., BRASSEUR, P., AGNAMEY, P., PÉRIGNON, J.L., DRUILHE, P.
Understanding human-Plasmodium falciparum immune interactions uncovers the
immunological role of worms. PLoS ONE 5 (2): 930, 2010.
SALIM, N., KNOPP, S., LWENO, O., ABDUL, U., MOHAMED, A., SCHINDLER, T.,
ROTHEN, J., MASIMBA, J., KWABA, D., MOHAMMED, A.S., ALTHAUS, F.,
ABDULLA,S., TANNER, M., DAUBENBERGER, C., GENTON, B. Distribution and
Risk Factors for Plasmodium and Helminth Co-infections: A Cross-Sectional Survey
among Children in Bagamoyo District, Coastal Region of Tanzania. PLoS Negl Trop
Dis. 9: 4, 2015.
SÁNCHEZ-ARCILA, J.C., PERCE-DA-SILVA, D.S., VASCONCELOS, M.P.A.,
RODRIGUES-DA-SILVA, R.N., PEREIRA, V.A., APRÍGIO, C.J.L., LIMA, C.A.M.,
FONSECA, B.P.F.E., BANIC, D.M., LIMA-JUNIOR, J.C., OLIVEIRA-FERREIRA, J.
. Intestinal Parasites Coinfection Does Not Alter Plasma Cytokines Profile Elicited in
Acute Malaria in Subjects from Endemic Area of Brazil. Mediators of Inflammation,
2014: 1-12, 2014.

106
SÁNCHEZ-ARCILA, J.C., DE FRANÇA, M.M., PEREIRA, V.A., VASCONCELOS,
M.P.A., TÊVA, A., PERCE-DA-SILVA, D.S., NETO, J.R., APRÍGIO, C.J.L., LIMA-
JUNIOR, J.C., RODRIGUES, M.M., SOARES, I.S., BANIC, D.M., OLIVEIRA-
FERREIRA, J. The influence of intestinal parasites on Plasmodium vivax-specific
antibody responses to MSP-119 and AMA-1 in rural populations of the Brazilian
Amazon. Malaria Journal 14: 442, 2015.
SEMEDO, M. R. L.; SANTOS, M. M. A. S.; BAIÃO, M. R.; LUIZ, R. R., DA VEIGA G.V.
anemia and associated factors among school-age children in cape verde, west África.
AJFAND 14 (7): 1-18, 2014.
SILVA-NUNES, M., CODEÇO, C.T., MALAFRONTE, R.S., DA SILVA, N.S.,
JUNCANSEN, C., MUNIZ, P.T., FERREIRA, M.U. Malaria on the Amazonian
frontier: transmission dynamics, risk factors, spatial distribution, and prospects for
control. Am J Trop Med Hyg.79(4):624-35, 2008.
SILVA, N.S., SILVA-NUNES, M., MALAFRONTE, R.S., MENEZES, M.J., D'ARCADIA,
R.R., KOMATSU, N.T., SCOPEL, K.K., BRAGA, E.M., CAVASINI, C.E.,
CORDEIRO, J.A., FERREIRA, U.M. Epidemiology and control of frontier malaria in
Brazil: lessons from community-based studies in rural Amazonia. Trans R Soc Trop
Med Hyg 104 (5): 343-350. 2010.
SILVA, G. V. Desenvolvimento econômico em cidades da fronteira amazônica: ações,
escalas e recursos para Oiapoque-AP. Confins, 17, 2013. Disponível em:
<http://confins.revues.org/8250> Acesso em 02 de Outubro de 2017.
SILVIE, O., FRANETICH, J.F., CHARRIN, S., MUELLER, M.S., SIAU, A., BODESCOT,
M., RUBINSTEIN, E., HANNOUN, L., CHAROENVIT, Y., KOCKEN, C.H.,
THOMAS, A.W., GEMERT, G.J.V., SAUERWEIN, R.W., BLACKMAN, M.J.,
ANDERS, R.F., PLUSCHKE, G., MAZIERA, D. Role for apical membrane antigen 1
during invasion of hepatocytes by Plasmodium falciparum sporozoites. Journal of
Biological Chemistry 279: 9490-9496. 2004.
SIQUEIRA, L.O., ALBARELLO, K., YOUNES, S., HAHN, S. Diagnóstico de anemia e
parasitoses em crianças em situação de vulnerabilidade social. Rev. Diálogos:
Contribuição da extensão para a consolidação dos direitos humanos. 6: 18 -25,
2011.
SIVEP – MALÁRIA – Sistema de Informação de Vigilância Epidemiológica – Ministério da
Saúde. Brasil. Disponível em: <www.saude.gov.br/sivep_malaria> Acesso em:
18/07/2017.
SOARES, I.S., CUNHA, M.G., SILVA, M.N., SOUZA, J.M., DEL PORTILLO, H.A.,
RODRIGUES, M.M. Longevity of naturally acquired antibody responses to the N and
C-terminal regions of Plasmodium vivax merozoite surface protein 1. Am J Trop Med
Hyg. 60: 357-63, 1999.
SOARES, I.S., LEVITUS, G., SOUZA, J.M., PORTILHO, H.A., RODRIGUES, M.M.
Acquired immune responses to the N-and C-terminal regions of Plasmodium vivax

107
merozoite surface protein 1 in individuals exposed to malaria. Infection and
Immunity, 65 (5): 1606–1614, 1997.
SOHAIL, M., KAUL, A., BALI, P., RAZIUDDIN, M., SINGH, M.P., SINGH, O.P., DASH,
A.P., ADAK, T. Alleles -308A and -1031C in the TNF-α gene promoter do not increase
the risk but associated with circulating levels of TNF-α and clinical features of vivax
malaria in Indian patients. Mol Immunol 45: 1682-1692, 2008.
SORTICA ,V.A., CUNHA, M.G., OHNISHI, M.D., SOUZA, J.M., RIBEIRO-DOS-
SANTOS, A.K.C., SANTOS, N.P.C., CALLEGARI-JACQUES, S.M., SANTOS,
S.E.B., HUTZ, M.H. 2012. IL1B, IL4R, IL12RB1 and TNF gene polymorphisms are
associated with Plasmodium vivax malaria in Brazil. Malaria Journal. 11: 409, 2012.
SUPALI, T. VERWEIJ, J.J. WIRIA, A.E., DJUARDI, Y., HAMID, F., KAISAR, M.M.,
WAMMES, L.J., VAN LIESHOUT, L., LUTY, A.J., SARTONO, E.,
YAZDANBAKHSH, M. Polyparasitism and its impact on the immune system.
International Journal for Parasitology 40 (10): 1171-1176, 2010.
SHAPIRO, A.E., TUKAHEBWA, E.M., KASTEN, J., CLARKE, S.E., MAGNUSSEN, P.,
OLSEN, A., KABATEREINE, N.B., NDYOMUGYENYI, R., BROOKER, S.
Epidemiology of helminth infections and their relationship to clinical malaria in
southwest Uganda. Trans R Soc Trop Med Hyg. 99 (1):18-24, 2005.
SMITS, H.H., EVERTS, B., HARTGERS, F.C., YAZDANBAKHSH, M. Chronic helminth
infections protect against allergic diseases by active regulatory processes. Current
Allergy Asthma Reports 10 (1): 3-12, 2010.
SNOUNOU, G., VIRIYAKOSOL, S., ZHU, X.P., JARRA, W., PINHEIRO, L., ROSARIO,
V.E., THAITHONG, S., BROWN, K.N. High sensitivity of detection of human malaria
parasites by the use of nested polymerase chain reaction. Mol Biochem Parasitol.
61(2):315-20, 1993.
SPIEGEL, A., TALL, A., RAPHENON, G., TRAPE, J.F., DRUILHE, P. Increased frequency
of malaria attacks in subjects co-infected by intestinal worms and Plasmodium
falciparum malaria. Trans R Soc Trop Med Hyg 97 (2): 198-9, 2003.
STEPHENSON, L.S., HOLLAND, C.V., COOPER, E.S. The public health significance of
Trichuris trichiura. Parasitology 121: 73-95, 2000.
TANGTEERAWATANA, P., PICHYANGKUL, S., HAYANO, M., KALAMBAHETI, T.,
LOOAREESUWAN, S., TROYEBLOMBERG, M., KHUSMITH, S. Relative levels of
IL4 and IFN-γ in complicated malaria: Association with IL4 polymorphism and
peripheral parasitemia. Acta Trop. 101(3): 258-65, 2007.
TAYLOR, M.D., LEGOFF L., HARRIS, A., MALONE, E., ALLEN, J.E., MAIZELS, R.M.
Removal of regulatory T cell activity reverses hyporesponsiveness and leads to filarial
parasite clearance in vivo. The Journal of Immunology 15 (8): 4924–33. 2005.
TURNER, J.D., JACKSON,J.A. FAULKNER, H. BEHNKE, J., ELSE, K.J., KAMGNO, J.,
BOUSSINES, Q.M., BRADLEY, J.E. Intensity of intestinal infection with multiple

108
worm species is related to regulatory cytokine output and immune hyporesponsiveness.
The Journal of Infectious Diseases 197 (8): 1204-1212, 2008.
UTHAIPIBULL, C., AUFIERO, B., SYED, S.E., HANSEN, B., GUEVARA PATINO, J.A.,
ANGOV, E., LING, I.T., FEGEDING, K., MORGAN, W.D., OCKENHOUSE, C.,
BIRDSALL, B., FEENEY, J., LYON, J.A., HOLDER, A.A. Inhibitory and blocking
monoclonal antibody epitopes on merozoite surface protein 1 of the malaria parasite
Plasmodium falciparum. J Mol Biol 307: 1381-94, 2001.
VAFA, M., MAIGA, B., BERZINS, K., HAYANO, M., BERECZKY, S., DOLO, A., DAOU,
M., ARAMA, C., KOURIBA, B., FARNERT, A., DOUMBO, O.K., TROYE-
BLOMBERG, M. Associations between the IL-4-590 T allele and Plasmodium
falciparum infection prevalence in asymptomatic Fulani of Mali. Microbes Infect. 9:
1043-1048, 2007.
VIEIRA, D.E.A., BENETTON, M.L.F.N. Fatores ambientais e sócio econômicos associados
à ocorrência de enteroparasitoses em usuários atendidos na rede pública de saúde em
Manaus, AM, BRASIL. Biosci. J. Uberlândia 29 (2): 487-498, 2013.
WALTHER, M., JEFFRIES, D., FINNEY, O.C., NJIE, M., EBONYI, A., DEININGER, S.,
LAWRENCE, E., NGWAAMAMBUA, A., JAYASOORIYA, S., CHEESEMAN, I.H.,
GOMEZ-ESCOBAR, N., OKEBE, J., CONWAY, D.J., RILEY, E.M. Distinct Roles for
FOXP3+ and FOXP3
- CD4
+ T Cells in Regulating Cellular Immunity to Uncomplicated
and Severe Plasmodium falciparum Malaria. PLoS Pathog. 4(5): 1-14, 2009.
WAMMES, L.J., HAMID, F., WIRIA, A.E., DE GIER, B., SARTONO, E., MAIZELS, R.M.,
LUTY, A.J., FILLIÉ, Y., BRICE, G.T., SUPALI, T., SMITS, H.H.,
YAZDANBAKHSH, M. Regulatory T cells in human geohelminth infection suppress
immune responses to BCG and Plasmodium falciparum. Eur. J. Immunol. 40: 437-
442, 2010.
WAMMES, L.J., MPAIRWE, H., ELLIOTT, A.M., YAZDANBAKHSH, M. Helminth
therapy or elimination: epidemiological, immunological, and clinical considerations.
Lancet Infect Dis. 14 (11): 1150-62, 2014.
WELLS, T.N.C., BURROWS, J.N., BAIRD, J.K. Targeting the hypnozoite reservoir of
Plasmodium vivax: the hidden obstacle to malaria elimination. Trends in Parasitology,
26: 145-151, 2010.
WHITE, N. J. Determinants of relapse periodicity in Plasmodium vivax malaria. Malaria
Journal 10 (1): 297, 2011.
WIPASA, J., OKELL, L, SAKKHACHORNPHOP S, SUPHAVILAI C,
CHAWANSUNTATI K., LIEWSAREE, W., HAFALLA, J.C., RILEY, E.M. Short-
Lived IFN-γ Effector Responses, but Long-Lived IL-10 Memory Responses, to Malaria
in an Area of Low Malaria Endemicity. PLoS Pathog 7 (2): 2011.
WORLD HEALTH ORGANIZATION (WHO). World Malaria Report 2011. WHO -
Scientific Group, Technical Report Series. 2011.

109
WORLD HEALTH ORGANIZATION (WHO). World Malaria Report 2013. WHO -
Scientific Group, Technical Report Series. 2013.
WORLD HEALTH ORGANIZATION (WHO). World Malaria Report 2014. WHO -
Scientific Group, Technical Report Series. 2014.
ZEYREK, F.Y., OGLU, A., ERDOGAN, D.D., AK, M., COBAN, C. Analysis of naturally
acquired antibody responses to the 19-kd C-terminal region of merozoite surface
protein-1 of 26 Plasmodium vivax from individuals in Sanliurfa, Turkey. Am J Trop
Med Hyg 78: 729-732, 2008.

110
APÊNDICE A - Termo de Consentimento Livre e Esclarecido
TERMO DE CONSENTIMENTO LIVRE E ESCLARECIDO
(Obrigatório para Pesquisa Clínica em Seres Humanos – Resolução nº 466/2012 – CNS)
I – DADOS DE IDENTIFICAÇÃO
NOME (PACIENTE):_______________________________________________________
DOCUMENTO DE INDENTIDADE: __________________________________________
ENDEREÇO COMPLETO: __________________________________________________
BAIRRO: _____________________________ CEP: ______________________________
CIDADE: ___________________ ESTADO: _______________ FONE: ______________
II – DADOS SOBRE O ESTUDO
1.Título: Coinfecção do Plasmodium Vivax e enteroparasitos no Município do Oiapoque,
Estado do Amapá, Fronteira Brasil-Guiana Francesa
2. Equipe responsável: Rubens Alex de Oliveira Menezes, Margarete do Socorro Mendonça
Gomes, Ricardo Luiz Dantas Machado.
III. EXPLICAÇÕES SOBRE O PROJETO DE PESQUISA AO PACIENTE
1. A malária é uma doença muito frequente na Amazônia e causada por parasitos chamados
plasmódios;
2. A infecção de parasitas intestinais com o agente de malária pode ter o quadro de anemia
acentuado;
3. Visando esclarecer alguns desses aspectos, convidamos você a participar desta pesquisa.
Para conduzir os estudos, precisaremos realizar uma coleta do sangue da ponta dos dedos para
preparação de lâminas de Gota Espessa e Distensão Sanguínea para diagnóstico da doença.
Serão também coletadas amostras de sangue (10 mL) da veia do braço para realizar os estudos
genéticos e sorológicos e fezes para realização do exame parasitológico. As coletas de sangue
serão realizadas com materiais descartáveis e estéreis. Geralmente, não há dor no local da
coleta, mas pode ocorrer de intensidade limitada e por pouco tempo, em casos de coletas de
sangue da veia. Serão entregues frascos limpos para o exame de fezes;
4. Algumas complicações decorrentes da coleta de sangue podem ocorrer como manchas
roxas no local da coleta devido à retirada da agulha pelo profissional antes do garrote ou por
perfuração da veia; alergia ao álcool à 70% usado para limpeza local ou contaminações
devido à má limpeza feita ou uso de material não-descartável (estéril) contaminado;
5. É assegurado o diagnóstico e o tratamento de todos os pacientes com malária, independente
da participação neste estudo;
7. A participação neste estudo é voluntária e você poderá retirar o seu consentimento a
qualquer momento sem prejuízo do seu tratamento regular;
8. As informações obtidas serão analisadas em conjunto com outros pacientes/indivíduos, não
sendo divulgado a identificação de nenhum paciente/indivíduo;
9. Não há despesas pessoais para o participante em qualquer fase do estudo, incluindo exame
e consultas. Também não há compensação financeira relacionada à participação;
10. O material biológico obtido poderá ser utilizado para esta pesquisa será armazenado uma
alíquota no Laboratório de Malária da Seção de Parasitologia do Instituto Evandro Chagas
para a realização em outras pesquisas decorrentes deste estudo. Nova solicitação de utilização
dessas amostras será realizada junto ao Comitê de Ética;

111
11. Para obtenção de quaisquer informações e esclarecimentos, o paciente poderá entrar em
contato com os pesquisadores responsáveis no Instituto Evandro Chagas, situado na Rodovia
Br 316, s/ nº - Bairro: Levilândia, CEP: 67030 – 000. Telefone para contacto: (0XX91) 3214
2089. Poderá também contactar o Comitê de Ética em Pesquisa do Instituto Evandro Chagas,
situado no mesmo endereço citado anteriormente. Telefone para contacto: (0XX91) 3214-
2237, sob Coordenação do Dr. Ricardo Luiz Dantas Machado;
12. Todas as informações coletadas serão mantidas em sigilo.
IV. AFIRMAÇÕES DO PACIENTE OU DE SEU RESPONSÁVEL LEGAL
1. Fui esclarecido sobre os objetivos da pesquisa, os procedimentos, riscos, benefícios e
outros assuntos? SIM NÃO
2. Fui esclarecido sobre a liberdade de retirar o meu consentimento a qualquer momento, sem
que isso traga prejuízo quanto à continuidade do meu tratamento ou do menor do qual sou
responsável? SIM NÃO
3. Fui esclarecido de que não haverá remuneração financeira além do previsto para as
despesas do estudo? SIM NÃO
4. Fui esclarecido de que não haverá indenização além das previstas pela lei, em reparação a
dano imediato ou tardio, causado pela pesquisa em questão? SIM NÃO
5. Fui esclarecido de que a segurança da minha identidade ou do menor do qual sou
responsável será preservada, mantendo-se as informações em caráter confidencial?
SIM NÃO
V. CONSENTIMENTO PÓS-INFORMADO
Declaro que, após ter sido convenientemente esclarecido dos riscos e benefícios deste estudo,
conforme acima definido, autorizo a minha participação ou do menor do qual sou responsável,
na qualidade de paciente no Projeto de Pesquisa referido no inciso II.
Macapá, ........... de...................... de................
......................................................................... ...........................................................
Assinatura Paciente ou do Responsável Legal Assinatura e carimbo do Pesquisador
Documento em duas vias
Rubrica em todas as páginas do TCLE do participante da pesquisa e o pesquisador
responsável

112
APÊNDICE B – Formulário epidemiológico
Coinfecção do Plasmodium Vivax e enteroparasitos no Município do Oiapoque, Estado do
Amapá, Fronteira Brasil-Guiana Francesa
A Informação do Espécime
1. Número de ID do espécime:_____________ (por exemplo, CB 01) a ser codificado
conforme abaixo:
XX = primeira inicial do país e do centro de coleta, por exemplo CB para
Colômbia/Buenaventura
## = identificador exclusivo do espécime, por exemplo 01 para primeiro paciente recrutado
para a avaliação
2. Data de coleta do espécime: [____/____/_______] (DIA/MÊS/ANO)
B. Informações sobre o paciente
1. Número de ID do paciente:______________ (igual ao número de ID do espécime)
2. Sexo: Masculino Feminino
3. Idade: _____ [anos]
4. Residência: cidade ou distrito ______________________________
Estado (Província) e País __________________________
5. Temperatura corporal: ___________________________________________________
6. Histórico de viagens: “Você viajou para fora deste Estado (província)/distrito nas últimas
quatro semanas?” Sim Não
Se sim, para onde? (começar com a viagem mais recente)__________________________
Viagem 1: Cidade ou Povoação/Bairro ________________________Distrito ou Estado
(Província)_______________
C Resultados de investigação microscópica:
1. Investigação microscópica (realizada no campo):
Espécie: ______________________
Parasitemia: ______________________ (contagem de parasitos no campo)
Classificado como 1+/2+/3+/4+ ou % de parasitemia ou parasitos por
microlitro. 2. Investigação microscópica (realizada no laboratório):
Espécie: ______________________
Parasitemia: ______________________ (contagem precisa de parasitos)
Classificada como parasitos por microlitro de sangue
D Passado malárico
Exposição prévia à doença Sim Não
Primoinfecção Sim Não
Episódios prévios de malária Sim Não
Se sim Quantas vezes________________________
Assinatura do investigador principal _____________________________
Data: [___/___/______] (DIA/MÊS/ANO)
OBS: Este documento foi elaborado em duas vias, uma ficará guardada nos arquivos das
instituições responsáveis pela entrevista e coleta do paciente e sob a responsabilidade do
pesquisador e a outra será entregue aos participantes do estudo, sendo que todas as duas vias
estarão assinadas por quem de direito.

113
APÊNDICE C – Resumos aprovados em Congresso

114
Resumo Aprovado no Congresso de Medicina Tropical 2015
Código: 1645
Título: Perfil epidemiológico da coinfecção Plasmodium Vivax e Parasitoses Intestinais no
Município do Oiapoque, Estado do Amapá: Resultados Preliminares
Autores: Rubens Alex Oliveira de Menezes, Margarete do Socorro Mendonça Gomes,
Anapaula Martins Mendes, Jairo Lourenço da Silva Filho, Francicléa Lúcia Oliveira
Ferreira, Ederson Tavares Nascimento e Ricardo Luiz Dantas Machado*.
Instituição: Universidade Federal do Pará
Palavras-chave: Malária, parasitoses intestinais, coinfecção, município do Oiapoque,
Amazônia brasileira
Resumo: Introdução: Embora a coinfecção entre o plasmódio e os parasitos intestinais seja
prevalente em países tropicais, o efeito de suas interações permanece obscuro. Este estudo
tem como objetivo avaliar o perfil da coinfecção com parasitos intestinais com pacientes com
infecção por Plasmodium vivax em uma área endêmica da Amazônia brasileira. Metodologia:
Estudo transversal foi realizado, no município de Oiapoque, pertencente ao Estado do Amapá,
norte do Brasil, fronteira ocidental da região amazônica. O diagnóstico de malária e a
contagem de gametócitos foram executados pela gosta espessa e esfregaço sanguíneo. A
dosagem de hemoglobina foi realizada por método automatizado. O exame parasitológico das
fezes foi realizado pelo método de Hoffman-Pons-Jones. O teste do Qui-quadrado e Exato de
Fisher foram usados para avaliar as diferenças entre as proporções e o teste de Student t para
avaliar as diferença entre as médias. Resultados: Nos três meses de coleta foram incluídos no
estudo 37 indivíduos que concordaram participar da pesquisa. O P. vivax foi detectado em
95,7% dos indivíduos. A parasitemia variou de 150 a 13.500 parasitos/mm3 (DP ± 3,57). Os
gametócitos foram detectados em 94,6% das lâminas investigadas. Os níveis de hemoglobina
variam de 10,4 a 15,7 g/dL (DP ± 2,65). Dos pacientes analisados 83,78% apresentaram-se
parasitados, com pelo menos um parasito intestinal patogênico. A Entamoeba coli foi o
protozoário mais frequente (32,14%), seguido da Entamoeba histolytica (14,28%) e a Giardia
lamblia (7,14%). Os helmintos detectados foram: Ascaris lumbricoides (46,42%), Thicuris
trichuris e Ancilostomídeos foram detectados em 3,57% das amostras avaliadas. Verificou-se
associação significativa entre enteroparasitoses e os níveis de hemoglobina. Não se observou
significância estatística na comparação entre presença de gametócitos e a presença de
parasitos. Conclusão: Os dados sugerem que existe uma associação entre estes dois grupos de
parasitos, ou poderia ser explicado pela presença de determinantes estruturais comuns para
ambas as doenças. Um programa de educação continuada envolvido com a prevenção e
tratamento das infecções parasitárias e malária é uma medida fundamental para sua
erradicação.

115
Certificado do Resumo no Evento Científico

116

117
Resumo 1 Aprovado no Congresso de Medicina Tropical 2016
Código: 7533
Titulo: Perfil dos níveis de hemoglobina e concentração de gametócitos na coinfecção
Plasmodium Vivax/Enteroparasitoses no Município do Oiapoque, Estado do Amapá
Autores: Rubens Alex de Oliveira Menezes1, Margarete do Socorro Mendonça Gomes
2,
Anapaula Martins Mendes3, Silvestre Rodrigues do Nascimento
4, Ricardo Luiz Dantas
Machado1,5
1Programa de Pós-Graduação em Biologia de Agentes Infecciosos e Parasitários da
Universidade Federal do Pará; 2SESA/Laboratório Central do Amapá;
3Universidade
Federal do Amapá/Campus Binacional-Oiapoque; 4HEO - Hospital Estadual do Oiapoque,
Pela Secretaria Municipal de Saúde do Oiapoque-AP; 5Instituto Evandro Chagas/SVS/MS.
RESUMO
Introdução: Este estudo tem como objetivo avaliar parâmetros que influenciam a infecção por
Plasmodium vivax coinfectatos com parasitos intestinais em uma área endêmica da Amazônia
brasileira. Metodologia: Estudo transversal realizado no município de Oiapoque, Estado do
Amapá. O diagnóstico de malária e a contagem de gametócitos foram executados pela gosta
espessa e esfregaço sanguíneo. A dosagem de hemoglobina foi realizada por método
automatizado. O exame parasitológico das fezes foi realizado pelo método de Hoffman-
PonsJones. Análise estatística foi utilizada para avaliar os parâmetros investigados.
Resultados: Foram incluídos no estudo 446 indivíduos, o P. vivax foi detectado em 99,1% dos
indivíduos. A amostra foi dividida em quatro grupos: grupo I [Malária e Parasitose positivo
(n=118)], grupo II [Malária positivo e parasitose negativo (n=34)], grupo III [Malária
negativo e parasitose positivo (n=143)] e grupo IV [Malária e parasitose negativo (n=151)]. A
parasitemia variou de 25 a 30.000 parasitos/mm3 (DP ± 2,75). Os gametócitos foram
detectados em 88,5% das lâminas investigadas. Os níveis de hemoglobina variam de 7,7 a
18,2 g/dL (DP ± 0,5). Dos indivíduos incluídos no estudo, 118 (26,5%) foram coinfectados
com malaria e parasitos intestinais, esses indivíduos coinfectados, a prevalência de malária
por P. vivax foi de 117 (99,1%). Amebas comensais foram os protozoários mais frequentes
(84,7%) e o Ascaris lumbricoides foi o helminto mais detectado (88,4%). Não houve
significância estatística entre a concentração de gametócitos entre os grupos com malária
(coinfectados ou não) (p=0,1432).Não foi observado relação entre os grupos com malária
(coinfectados ou não) e a parasitemia (p = 0,06). . Houve diferença entre os níveis de
hemoglobina nos diferentes grupos estudados; entre o grupo I e o grupo II (p < 0,05), entre I e
IV (p<0,0001), entre II e III (p<0,005) e entre III e IV (p<0,0001). Conclusão: Na população
estudada, a coinfecção pode interferir nos níveis de hemoglobina da malária vivax.
Adicionalmente, a coinfecção não esteve associada com a redução de malária (incidência,
prevalência ou redução da parasitemia). A melhoria dos indicadores está nas ações de
educação em saúde e de mobilização da comunidade, além de uma maior cobertura da atenção
básica à saúde com relação aos casos suspeitos destes agravos.
Palavras-Chave: Malária, Plasmodium vivax, anemia, Oiapoque, Enteroparasitoses

118
Certificado do Resumo no Evento Científico

119
Resumo 2 Aprovado no Congresso de Medicina Tropical 2016
Código: 7534
Titulo: Caracterização da coinfecção da Malária e Parasitoses Intestinais no Município
do Oiapoque, Estado do Amapá, Brasil
Autores: Rubens Alex de Oliveira Menezes1, Margarete do Socorro Mendonça Gomes
2,
Anapaula Martins Mendes3, Silvestre Rodrigues do Nascimento
4, Ricardo Luiz Dantas
Machado1,5
1Programa de Pós-Graduação em Biologia de Agentes Infecciosos e Parasitários da
Universidade Federal do Pará; 2SESA/Laboratório Central do Amapá;
3Universidade
Federal do Amapá/Campus Binacional-Oiapoque; 4HEO - Hospital Estadual do Oiapoque,
Pela Secretaria Municipal de Saúde do Oiapoque-AP; 5Instituto Evandro Chagas/SVS/MS
RESUMO
Introdução: A coinfecção malária-parasitoses intestinais são comum nas regiões tropicais do
planeta. Embora essa coinfecção seja prevalente, o efeito de suas interações permanece
obscuro. Este estudo tem como objetivo avaliar o perfil da coinfecção de parasitos intestinais
com infecção por Plasmodium spp em uma área endêmica da Amazônia brasileira.
Metodologia: Estudo transversal foi realizado, no município de Oiapoque, pertencente ao
Estado do Amapá. O diagnóstico de malária e a contagem de gametócitos foram executados
pela gosta espessa e esfregaço sanguíneo. O exame parasitológico das fezes foi realizado pelo
método de Hoffman-Pons-Jones. Resultados: Em um ano de coleta foram incluídos no estudo
446 indivíduos, sendo 34 (7,6%) estavam infectados apenas com malária, 118 (26,5%) foram
co-infectados com malaria e parasitas intestinais, 143 (32%) foram infectados com parasitas
intestinais apenas, e 151 (33,9%) não foram infectadas com malária e parasitas intestinais. Em
indivíduos coinfectados, a prevalência de malária por P. vivax foi de 117 (99,1%) e P.
falciparum de 01 (0,9%). Ao analizarmos os casos apenas de malária, observamos 34 (7,6%),
sendo P. vivax 30 (85,8%), P. falciparum 3 (11,4%) e P. vivax + P. falciparum 01 (2,8%).
Dos 118 (26,5%) coinfectados com malaria e parasitos intestinais, observa-se 39 casos
(33,3%) de malaria-protozoários, sendo prevalente a associação de Entamoeba coli +
Endolimax nana (43,6%). Para os casos de malária-helmintos (46,6%), sendo Ascaris
lumbricoides mais predominante, com 47,2% dos casos. Os casos de tripla associação
(Malária/Protozoários/Helmintos foi verificado em (20,6%) das amostras, sendo mais
prevalente associação de E. coli + E. nana + A. lumbricoides (33,4%). Conclusão: Os
resultados apontam a coinfecção Malária e Enteroparasitoses um aspecto real no município do
Oiapoque. Avaliar a possibilidade de que populações infectadas por parasitoses intestinais
possam influenciar na dinâmica de transmissão da malária, são fundamentais para estratégias
diagnósticas e preventivas.
Palavras-Chave: Epidemiologia, Malária, Oiapoque, Plasmodium vivax

120
Certificado do Resumo no Evento Científico

121

122
Resumo Aprovado na 6th International Conference on Plasmodium Vivax Research 2017
Coding: 13427
Título: Malaria-Intestinal Parasite Co-infections: Intensity of Plasmodium vivax
Transmission Associated with Specific IgG Responses (PvMSP119) in Individuals from
Brazilian Amazon Border Community
Autores: Rubens Alex de Oliveira Menezes1,2
, Tamirys Pina Simão3,7
, Margarete do
Socorro Mendonça Gomes4, Anapaula Martins Mendes
5, Alvaro Augusto Ribeiro D Àlmeida
Couto4, Mathieu Nacher
6, Martin Johannes Enk
7, Maristela Gomes da Cunha
1, Ricardo Luiz
Dantas Machado1,7
1Postgraduate Program in Biology of Infectious and Parasitic Agents of the Federal University
of Pará, Brazil; 2Federal University of Amapá, Brazil;
3Postgraduate Program in
Neurosciences and Cell Biology of the Federal University of Pará, Brazil; 4SESA/ Central
Laboratory of Amapá; 5Federal University of Amapá/Binacional Campus-Oiapoque;
6Université des Antilles et de la Guyane Caiena/Guiana Francesa e da University of the West
Indies; 7Evandro Chagas Institute/Health Ministry, Pará, Brazil.
Backgrounds: Polyparasitism is a common condition in humans, but its impact on the
immune system and clinical diseases is still poorly understood. There are few studies on the
prevalence and effect of malaria-intestinal parasite co-infections in the immune response of
this interaction. Materials and Methods: We characterized the malaria-intestinal parasite co-
infection associated with specific IgG responses (PvMSP119) by evaluating the reactivity and
intensity of Plasmodium vivax transmission in 446 individuals from a community of the
Brazilian Amazon, located in the State of Amapá, Oiapoque municipality, bordering French
Guiana, and the samples was collected from 2014 to 2015. The IgG antibodies specific for
(MSP-119) were detected in the plasma by means of the enzyme-linked immunosorbent assay
(ELISA) and the samples were divided into four groups: group I [Malaria and Parasitosis (n =
118)], group II [Malaria only (n = 34)], group III [Parasitosis only (n = 143)] and group IV
[Control (n = 151)]. Results: The reactivity index of the samples ranged from 0.0182 to
18.4364 ± 3.3584 with 53.8% (240/446) presenting His6-MSP119 specific IgG, being these
82.2% (97 / 118) belonging to group I, 76.4% (26/34) to group II, 40.5% (58/143) to group III
and 39% (59/151) to group IV. The frequency of antibodies observed was 57.5% (61/106) for
protozoa, 57.9% (69/119) for helminths, 69.4% (25/36) for protozoa and helminths and 45.9%
(85/185) without infection. Analyzing the frequency in the groups I and III we obtained for
group I: 76.9% (30/39) with protozoa infection, 87.5% (49/56) with helminths and 75%
(18/24) protozoa and helminths infections; for group III: 45.5% (31/68) with protozoa
infections, 31.7% (20/63) with helminths and 58.3% (7/12) with protozoa and helminths
infections. Conclusions: Our results confirm the high antigenicity of this protein, especially
for group I, presenting a different profile regarding reactivity and intensity of transmission.
These data add important information that may help in understanding the immunological
aspects involved in the pathogenesis of malaria vivax.

123
Certificado do Resumo no Evento Científico

124

125
Resumo Aprovado no Congresso de Medicina Tropical 2017
Código: 15356
Título: Frequência de Enteroparasitoses no Município de Oiapoque, Amapá, Brasil,
Fronteira com a Guiana Francesa
Autores: Rubens A. O. Menezes1; Dayse C. G. Dias
2; Camila C. D. Melo
2; Margarete S. M.
Gomes3, Anapaula M. Mendes
4, Silvestre R. Nascimento
5, Alvaro A. R. D. Couto
3, Mathieu
Nacher6, Martin J. Enk
7, Ricardo L. D. Machado
1,7
1Programa de Pós-Graduação em Biologia de Agentes Infecciosos e Parasitários da
Universidade Federal do Pará; 2Discente de Medicina da Faculdade da Amazônia (FAMAZ),
Belém, PA, Brasil.3SESA/Laboratório Central do Amapá;
4Universidade Federal do
Amapá/Campus Binacional-Oiapoque; 5HEO - Hospital Estadual do Oiapoque, Pela
Secretaria Municipal de Saúde do Oiapoque-AP; 6Université des Antilles et de la Guyane
Caiena/Guiana Francesa e da University of the West Indies; 7Instituto Evandro
Chagas/SVS/MS.
RESUMO
Introdução: As enteroparasitoses representam um segmento considerável de doenças
infeccioso-parasitárias ao redor do mundo. Este estudo avaliou a frequência de enteroparasitos
e correlacionou com o nível de hemoglobina. Métodos: Estudo transversal realizado, no
município de Oiapoque, pertencente ao Estado do Amapá, norte do Brasil, fronteira ocidental
da região amazônica, no período de um ano de coleta de amostras fecais 2014/2015 utilizando
os métodos a fresco, direto, Hoffmann e sedimentação por centrifugação. Resultados: Foram
incluídos no estudo 446 indivíduos, todas as faixas etárias contribuíram para a presença de
parasitos intestinais, sendo realizado o teste de Meta-analise: Varias proporções combinadas,
cujo resultado apresentou-se entre as faixas etárias de 7 a 14 anos (p= <0.0001) e de ≥ 26 anos
(p=<0.0001). Nos indivíduos investigados, 58.6% (261/446) foram infectados por algum tipo
de parasito intestinal. Entre estes, 45,2% (118/161) estava infectado somente por helmintos,
40,9% (107/261) com protozoários, e 13,8% (36/261) tinham infecções associadas. Dentre os
helmintos detectados o Ascaris lumbricoides 19,9% (52/261) foi o mais prevalente, seguido
da associação da Entamoeba coli e a Endolimax nana como protozoários mais frequentes
17,2% (45/261). A pesquisa evidenciou uma correlação inversamente proporcional
significativa entre o nível de hemoglobina e a presença das parasitoses detectadas (coeficiente
de contingência C=0.2195 e p < 0.0001). Conclusões: O índice de enteroparasitoses é um
reflexo claro da falta de saneamento da região estudada, indicando um estado epidemiológico
preocupante. Dessa forma, um modelo eficaz de atenção primária à saúde tem sido a sugestão
no combate a essas condições adversas.
Palavra Chave: Enteroparasitoses, Região Amazônica, Oiapoque, Hemoglobina.

126
APÊNDICE D – Manuscrito Extra
Evaluation of the Anti-Pvs48/45 Humoral Immune Response in Natural Plasmodium
vivax Infections in the Brazilian Amazon
Este artigo avaliou a resposta de anticorpos naturalmente adquirida em infecções por
Plasmodium vivax contra a proteína Pvs48/45 na Amazônia Brasileira. Tais resultados
poderão contribuir para o entendimento do potencial imunogênico desse antígeno e
desenvolvimento de uma vacina de bloqueio de transmissão eficiente. Este é o primeiro
estudo que oferece um perfil imune humoral à proteína Pvs48/45, em amostras de pacientes
maláricos da região Amazônica brasileira.
Resumo dos resultados: A pesquisa de anticorpos IgG anti-Pvs48/45 foi realizada por
ELISA no soro de 281 amostras de pacientes maláricos residentes na região amazônica
brasileira. O ponto de corte foi estabelecido utilizando soro de voluntários que nunca tiveram
contato com o plasmódio, estabelecendo a média da densidade óptica (DO) mais três desvios
padrão. Dos 281soros analisados, em 22,4% foram encontradas respostas de anticorpos
específicos anti-Pvs48/45 e os índices de parasitemia e a presença de gametócitos não
influenciaram o reconhecimento antigênico desta proteína. Dentre os cinco subfragmentos
analisados, a região recombinante C-Terminal + Central da Pvs48/45 foi a que apresentou
maior reconhecimento antigênico na amostra em estudo (32%), apesar de não ser
estatisticamente significativo. Essas características dão suporte ao potencial promissor desse
antígeno no desenvolvimento de uma nova estratégia de vacina.

127
Evaluation of the Anti-Pvs48/45 Humoral Immune Response in Natural Plasmodium
vivax Infections in the Brazilian Amazon
Tatiana M. S. Cisne Pessoa1; Myrela C. Santos de Jesus
1; Rubens Alex de Oliveira
Menezes2; Ricardo L. D. Machado
3; Myrian Arevalo-Herrera
4; Sócrates Herrera
4;
Luciane M. Storti-Melo1*
1Post-Graduate Program in Parasitic Biology, Federal University of Sergipe (Universidade
Federal de Sergipe - UFS), 49100-000, São Cristóvão, Sergipe, Brazil 2Post-Graduate Program in Biology of Infectious and Parasitic Agents, Federal University of
Pará (Universidade Federal do Pará – UFPA) 3Evandro Chagas Instituto/Health Surveillance Service/Ministry of Health (IEC), Belém,
Pará, Brazil 4Caucaseco Scientific Research Centre, Cali, Colombia
*Corresponding author: Universidade Federal do Sergipe, Departamento de Biologia,
Centro de Ciências Biológicas e da Saúde
Address: Av. Marechal Rondon, s/n, Rosa Elze, São Cristóvão, Sergipe, Brasil, Cep. 49100-
000, E-mail address:[email protected] (Storti-Melo, L. M.)
Abstract
Background Malaria continues to be responsible for major morbidity and mortality in many
tropical and subtropical countries. Individuals who are frequently exposed to the malaria
parasite in endemic areas develop specific immune responses, leading to a reduction in the
parasitic load and clinical manifestations. The sera of these individuals may make it
impossible for gametes to fertilize in mosquitoes, thereby blocking the transmission of this
parasite to the vector. The Pvs48/45 protein is found on the surface of the gametocytes and is
used as an antibody target, which represents a viable vaccination strategy to block
transmission. The aim of this study was to evaluate naturally acquired antibody responses
against the Pvs48/45 protein in Plasmodium vivax-infected patients.
Materials and methods Anti-Pvs48/45 IgG antibody screening was performed by ELISA using 281 serum
samples from malarial patients living in the Brazilian Amazon region. The cut-off point was
established using sera from volunteers who had never had contact with Plasmodium to
establish the mean optical density (OD) plus three standard deviations. Fischer's exact test
was applied to compare the parasitaemia indices and the presence of gametocytes with the
frequency of humoral immune responses and the antigenicity of the Pvs48/45 protein and its
subfragments. The level of significance was 95%.
Results
Anti-Pvs48/45-specific antibody responses were found in 22.4% of the 281 analysed
sera. The parasitaemia indices and the presence of gametocytes did not influence antigenic
recognition of this protein. Among the five analysed subfragments, the recombinant C-
terminal + central region of Pvs48/45 presented the highest antigenic recognition in the study
sample (32%), although the difference was not significant.
Conclusion Studies using an isolated region of the protein may facilitate understanding of the
immunogenic potential of this antigen. This study aims to contribute to the development of an
efficient transmission-blocking vaccine against the Pvs48/45 protein in future trials.
Keywords: Pvs48/45; vaccine; immune response; malaria.

128
Introduction
Plasmodium vivax and the vectors involved in the biological cycle of malaria have
several distinct characteristics that allow the transmission of this parasite in both temperate
climates and tropical areas [1, 2]. However, none of these characteristics are as specific as the
ability of this Plasmodium species to form hypnozoites, which reactivate periodically and can
cause multiple malaria episodes that account for up to 80% of clinical episodes [3]. In Brazil,
approximately 99% of malaria transmission is concentrated in the Amazon region. P. vivax is
the most prevalent species and is responsible for approximately 84% of the cases in this
region [4-6]. The maintenance of the number of malaria cases, the rapid development of P.
vivax in vector mosquitoes [7] and the difficulty in the control of transmission using
traditional methods in endemic regions have shown the importance of identifying effective
and economically viable methods to interrupt the transmission of malaria in endemic regions
[8].
Individuals who are frequently exposed to the parasite have the ability to develop
specific immune responses that protect them from developing a serious malarial infection [9,
10]. Moreover, the serum of these individuals may make fertilization of the gametes
impossible, thereby preventing parasite development and consequently blocking the
transmission of this parasite from the infected patients to mosquitoes [11]. This exposure to
endemicity facilitates the development of a transmission blocking vaccine [12]. Specific
antibodies against Plasmodium antigens expressed in gametocytes have been experimentally
demonstrated to block Plasmodium transmission from the vertebrate host to the vector
mosquito [13]. Although this blockade does not prevent the individual from acquiring the
disease, the vaccination strategy interrupts the transmission from infected individuals to
uninfected individuals [12, 14] and prevents the re-emergence of transmission in areas where
complete elimination has been achieved [8].
The Pvs48/45 protein is located on the surface of the P. vivax gametocyte and acts on
the fertilization of the parasite. Thus, the protein represents a potential target for transmission
blocking and anti-parasitic activity [15]. The synthesis of this protein was recently described,
and a preliminary evaluation of its immunological properties, including its antigenicity,
immunogenicity and transmission blocking activity, was performed using samples from
patients with malaria vivax from Colombia. However, this study suggested the need for a
broader evaluation in areas endemic for malaria to verify the effectiveness of this protein in
transmission blocking. The estimated responses of antibodies acquired against the Pvs48/45
protein in natural P. vivax infections in an endemic area in Brazil may provide results that add
to and clarify the role of these antibodies and the relevance of this antigen for vaccine
development.
This protein together with the post-fertilization antigens Pvs25 and Pvs28 has a highly
limited polymorphic sequence and low nucleotide diversity compared to the blood stage
antigens [16]. These characteristics support the promising potential of this antigen in the
development of a novel vaccine strategy. Here, we present for the first time the humoral
immune response to the Pvs48/45 protein in samples from malarial patients from the Brazilian
Amazon region.
Materials and methods
Study population
For this study, 281 samples collected from seven different cities in the Brazilian
Amazon region were included to extend the range of samples for the descriptive antigenicity
study. The samples were taken from Rio Branco/state of Acre (n=26, collected in 2003), Porto
Velho/state of Rondônia (n=26, collected in 2003), Belém (n=25, collected in 2003),
Goianésia do Pará (n=20, collected in 2012) and Itaituba (n=19, collected in 2016) in the state
of Pará and the municipalities of Macapá (n=16, collected in 2003) and Oiapoque (n=150,

129
collected in 2015) in the state of Amapá (Fig. 1). The samples used in this study were
collected from patients who were randomly selected from individuals who spontaneously
sought a public health centre with symptoms suggestive of malaria, had a positive thick-drop
smear test result and were confirmed to have vivax malaria by molecular diagnosis [17-19].
Figure 1: Cities in the Brazilian Amazon region where the collections were performed.
Source: Geomapas and IBGE.
Ethical aspects
This project was submitted to the Ethics Committee of the Federal University of
Sergipe (Universidade Federal de Sergipe) to request the re-use of the samples and was
approved under number opinion no. 1,774,182.
Laboratory analysis
The blood plasma samples were analysed at the Caucaseco Scientific Research Centre
in Cali, Colombia. The evaluation of naturally acquired antibodies against the Pvs48/45
protein and its five subfragments (N-terminal, C-terminal, central, N-terminal + central and
C-terminal + central) was performed using an enzyme-linked immunosorbent assay (ELISA)
as described in Arévalo-Herrera et al. [15]. All samples were analysed in duplicate. Because
the protein purification procedure used the thioredoxin system, all samples were also
evaluated by ELISA for reactivity to thioredoxin in parallel with the protein and subfragment
analyses.
Samples that showed a humoral immune response against Pvs48/45 were titrated. The
titration was performed in serial dilutions of 1:200, 1:400, 1:800 and 1:1600.

130
Data analysis
The result for each sample was expressed using the reactivity index (RI). This index
was calculated by dividing the mean absorbance values of each sample by the cut-off value of
the negative controls used as a reference on each plate. The cut-off point was calculated by
summing the mean of the negative controls with three times the obtained standard deviation.
Thus, the positive samples were those with an RI ≥ 1.0, whereas the negative samples had an
RI < 1.0. Samples that showed a positive result for thioredoxin were considered false positive
because this protein was present in the organisms where the protein was expressed, including
E. coli, as described in Arévalo-Herrera [15].
Fisher's exact test was applied in the Bioestat 5.3 program with a 95% significance
level for comparison of the parasitaemia indices and the presence of gametocytes with the
frequency of humoral immune responses and verification of the antigenicity of the Pvs48/45
protein and its subfragments.
Results
Antigenicity of Pvs48/45 protein in the Brazilian Amazon
Of the samples analysed to evaluate the humoral immune response against the
Pvs48/45 protein, 63 (22.42%) samples showed a positive humoral immune response (Fig. 2),
and 20.28% of the samples reacted with thioredoxin in addition to Pvs48/45.
The results show that the antibody response profile against the Pvs48/45 antigen is
variable from one region to another. Samples from the city of Rio Branco showed a higher
percentage of positive immune responses (38.5%), whereas no samples from individuals from
Macapá presented antigenicity for the Pvs48/45 protein. Patients with vivax malaria from
Itaituba, Belém and Goianésia do Pará presented similar percentages of samples with positive
immune responses (15.8%, 12.5% and 10.0%, respectively).
Among the 63 samples positive for Pvs48/45, the maximum reactivity index (RI)
value obtained was 5.5, and the minimum value was 1.0 (estimated reactivity threshold). The
dispersion of the RI values (Fig. 3) shows that the values are distributed close to the reactivity
threshold between 1 and 2, with a median of 1.3 (mean=1.5; SD=0.605).
Only one sample presented an RI > 5, and three samples presented an RI ≥ 2. Because
the RI is a value that expresses how many times the optical density (OD) of the sample is
greater than the cut-off point, the majority of the samples presented little reactivity.
Samples showing a positive immune response were titrated. The maximum dilution at
which a positive immune response was still obtained was 1:1600.
Figure 2: Distribution of the humoral immune response frequency against Pvs48/45 by city.
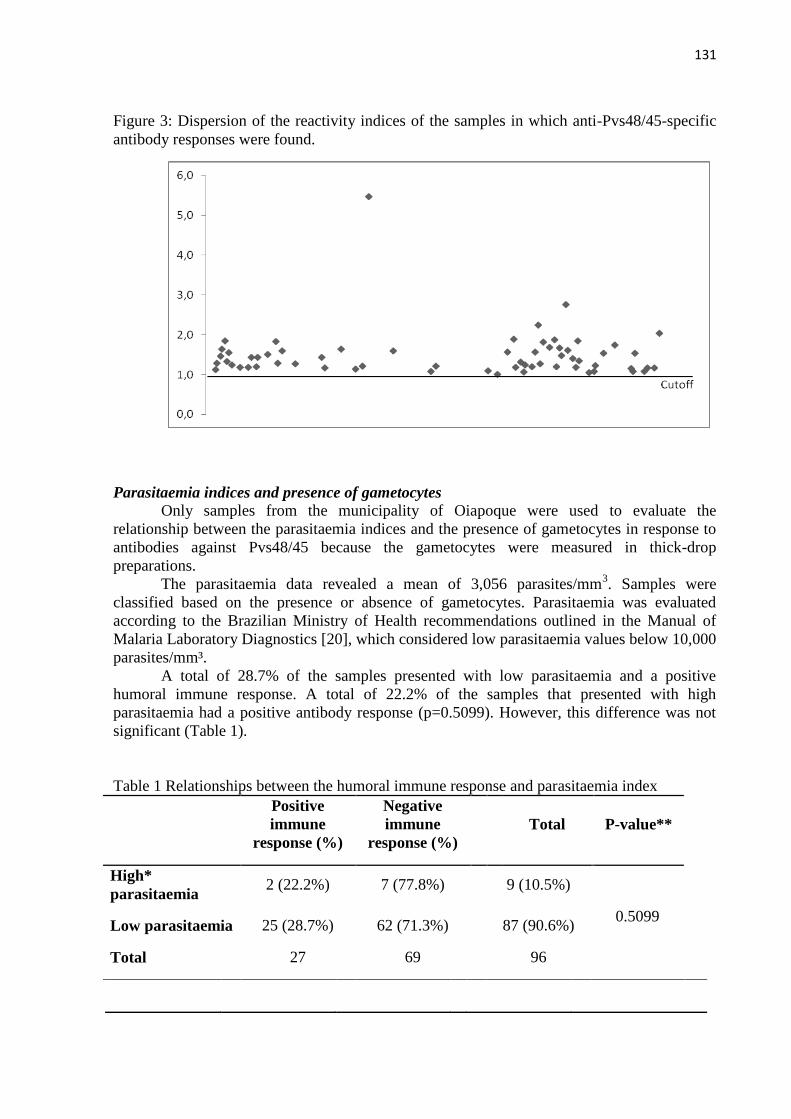
131
Figure 3: Dispersion of the reactivity indices of the samples in which anti-Pvs48/45-specific
antibody responses were found.
Parasitaemia indices and presence of gametocytes
Only samples from the municipality of Oiapoque were used to evaluate the
relationship between the parasitaemia indices and the presence of gametocytes in response to
antibodies against Pvs48/45 because the gametocytes were measured in thick-drop
preparations.
The parasitaemia data revealed a mean of 3,056 parasites/mm3. Samples were
classified based on the presence or absence of gametocytes. Parasitaemia was evaluated
according to the Brazilian Ministry of Health recommendations outlined in the Manual of
Malaria Laboratory Diagnostics [20], which considered low parasitaemia values below 10,000
parasites/mm³.
A total of 28.7% of the samples presented with low parasitaemia and a positive
humoral immune response. A total of 22.2% of the samples that presented with high
parasitaemia had a positive antibody response (p=0.5099). However, this difference was not
significant (Table 1).
Table 1 Relationships between the humoral immune response and parasitaemia index
Positive
immune
response (%)
Negative
immune
response (%)
Total P-value**
High*
parasitaemia 2 (22.2%) 7 (77.8%) 9 (10.5%)
0.5099 Low parasitaemia 25 (28.7%) 62 (71.3%) 87 (90.6%)
Total 27 69 96

132
Presence of
gametocytes
21 (25.3%) 62 (74.7%) 83 (86.5%)
0.1128 Absence of
gametocytes
6 (46.2%) 7 (53.8%) 13 (13.5%)
Total 27 69 96 (100%)
* High parasitaemia: ≥ 10,000 parasites/mm³; low parasitaemia <10,000 parasites/mm³.
** Fisher's exact test.
A humoral immune response positive for Pvs48/45 was detected in 25.3% of the
samples with the presence of gametocytes. Conversely, 46.2% of the samples in which no
gametocytes were found had a positive immune response. This difference was not significant
(p=0.1128) (Table 1).
Reactivity of the samples to the Pvs48/45 subfragments
In addition to the whole protein, 5 subfragments were expressed corresponding to the
N-terminal, C-terminal, central, N-terminal + central and C-terminal + central regions. A
subgroup of the investigated sample (n=169) distributed among the seven endemic areas was
used to assess the prevalence of antibodies against these specific regions of the protein and
thus estimate possible epitope locations.
Table 2 shows that the recombinant C-terminal + central sub-fragment showed the
highest percentage of reactivity (32%; p=0.0565), although the difference was not significant.
The central region of the peptide presented the lowest reactivity, with 17% (p=0.1645) of the
samples.
Table 2 Percentage of samples processed by peptide
Peptide n Positive RI
n (%) P-value*
Pvs48/45 281 63 (22.4%)
N-terminal 169 40 (23.7%) 0.4465
C-terminal 169 35 (20.7%) 0.4128
Central 169 29 (17.2%) 0.1645
N-terminal + Central 169 45 (26.6%) 0.2478
C-terminal + Central 169 54 (32.0%) 0.0565
* Fisher's exact test
Discussion
Different vaccine strategies that protect the individual from acquiring and developing
malarial infections using different parasite antigens have been evaluated [21, 22]. However,
effective results have not been achieved. Despite not preventing individuals from developing
the disease [23], the transmission blocking strategy blocks infected individuals from
transmitting the disease post-vaccination [15].
In this study, serum samples from naturally exposed malaria patients from the
Brazilian Amazon were analysed. Of these samples, 22.42% (63/281) presented an immune
response to the Pvs48/45 antigen. Although the seroprevalence was low, confirmation of this
antigenicity shows the production of anti-Pvs48/45 antibodies by individuals after exposure to
natural P. vivax infection. The recognition of the recombinant Pvs48/45 protein by these
natural antibodies shows that at least one epitope is present in the tested recombinant protein.

133
However, the low prevalence may be due to the expression system, which is based on the
thioredoxin model. The high percentage of samples that reacted to thioredoxin in addition to
Pvs48/45 (20.28%) caused a considerable portion of the samples to be classified as false
positive, leading to this low seroprevalence. In turn, the variable endemicity level of these
regions should be considered [24] because the endemicity differs from the malarial
transmission profile in the Colombian regions [15]. Moreover, the malarial history of these
patients investigated in the Brazilian Amazon showed that 34% of the samples represented
primary infections, which compromised the response to the investigated protein. Finally, the
Brazilian population is quite mixed due to the influence of colonization by the Portuguese and
contains crosses involving, mainly, Europeans, Africans and Native Americans [25].
Therefore, ethnicity may influence the frequency of polymorphisms of molecules involved in
the immune response and consequently may compromise their efficiency [26].
The low titre obtained is further evidence of the low reactivity of Pvs48/45 in samples
from infected individuals from the Brazilian Amazon. In a study conducted in different
endemic areas in the Brazilian Amazon region with the PV200L fragment of P. vivax MSP-1,
patients from the city of Macapá showed the lowest frequency and antibody levels. This result
was similar to the results obtained in the present study because no sample from this area
showed an immune response against Pvs48/45. One explanation for this finding was that
Macapá presented a low annual parasitaemia index (6.0 in 2009), which could contribute to
the absence of an immune response [19].
Individuals living in endemic areas are continually exposed to Plasmodium. These
individuals are more likely to develop clinical immunity by maintaining direct contact with
the parasite, which protects them against the development of severe malaria. These
individuals do not develop sterile immunity because this type of immunity is not achieved
under natural conditions. In a natural infection, individuals may not present high gametocyte
titres. Although P. vivax gametocytes are found in most infections prior to the onset of clinical
manifestations [8], the proportion in relation to the asexual stages does not reach 10%.
Additionally, these gametocytes remain in circulation in the blood stream for up to 3 days [7].
This phenomenon is perhaps another explanation for the low percentage of responses to the
Pvs48/45 antigen used in this study. In this sense, we cannot rule out the possibility that the
antigen tested is poorly recognized by individuals who are naturally exposed to Plasmodium
in the Brazilian Amazon region. Nevertheless, the importance of these findings can be
reinforced by evaluating the effectiveness of the antibodies produced by these individuals
through immunogenicity and transmission blocking tests to determine whether this antigen
has the ability to block the Plasmodium life cycle.
The results show that the presence or absence of gametocytes is not related to the
development of an antibody response against Pvs48/45 (Table 3). Although the gametocytes
might not have been found in the thick-drop test because they were present in low
concentrations [7], these concentrations were sufficient to ensure the contact of the
individuals with these parasitic forms. These results reinforce the need for further investments
and studies with the Pvs48/45 antigen as a candidate for a transmission blocking vaccine
because antibodies can develop independently of the high presence of gametocytes in the
bloodstream. Individuals who experience more malarial episodes present lower parasitaemias
than individuals who report fewer malarial episodes [27]. In the present study, both
individuals with high and low parasitaemias had similar anti-Pvs 48/45 antibody response
profiles. These results may indicate that the Pvs48/45 protein is antigenic in naturally exposed
populations regardless of the low parasitaemia indices commonly seen in endemic areas. This
finding may be supported by the presence of gametocytes in the early stages of P. vivax
infection even in cases with low parasitaemias [7].

134
When analysing the antigenicity of the C-terminal recombinant peptide plus the
central portion, we found that this region had a higher percentage of antigenicity compared to
the antigenicity obtained with the complete antigen (22.42%), although the difference was not
significant (32%). Studies with the homologous Pfs48/45 protein of P. falciparum revealed
that the amino-terminal (N-terminal) and carboxy-terminal (C-terminal) fragments induced
antibody responses capable of blocking transmission [28]. A study with the Pfs48/45 protein
from P. falciparum made progress in identifying conformational epitopes capable of inducing
blocking antibodies. Two epitopes (I and II) located in the C-terminal region of this protein
were identified as highly efficient targets because they contained domains crucial for the
development of a transmission blocking vaccine [29]. Pvs48/45 and its homologue Pfs48/45
are included in the group of proteins with cysteine-rich domains. As shown previously, the
cysteine residues located in the central and C-terminal regions of Pfs48/45 are crucial for the
presentation of the epitopes that induce blocking antibodies, which is an essential feature for
the effectiveness of a transmission blocking vaccine [29]. Although the expression of these
proteins, which are part of the cysteine class, in their proper conformation has been extremely
complex, some studies have achieved the expression of a fragment of the protein with a
correct conformational structure that has facilitated recognition of the protein. The C-terminal
fragment of the Pfs48/45 protein was fused to four catalysts (DsbA, DsbC, FkpA and SurA)
for expression to synthesize a correctly folded protein and generate high antibody titres [28].
This approach may represent a strategy for further expression studies with Pvs48/45 or its C-
terminal subfragment for future experimental immunogenicity assays.
Conclusion
Finally, further efficacy studies must be conducted to determine whether the results
obtained to date are on the right track or whether a course correction is required. The
successful development of a vaccine against P. vivax may take time, but investments are
needed because this neglected malaria parasite is widespread in the Americas, and reports of
major clinical complications have begun to emerge.
List of abbreviations
Declarations
Ethics approval and consent to participate
Ethical approval for this study was granted from the Research Ethics Committee of the
Universidade Federal do Sergipe (Protocol no. 1.088.855/2015), and all participants gave
written informed consent for enrolment in the study.
Consent for publication
Not applicable.
Availability of data and material
The authors confirm that all data underlying the findings are fully available without
restriction.
Competing interests
The authors declare that they have no competing interests.
Funding
Work reported in this manuscript was funded in part by Fundação de Amparo à Pesquisa do
Estado de Sergipe (FAPESP) and in part by the Conselho Nacional para o Desenvolvimento
Científico e Tecnológico (CNPq), Brasília, Brazil. Work in Colombia was jointly sponsored
by COLCIENCIAS, the Ministry of Social Protection and the National Institute of Allergy
and Infectious Diseases (NIAID grant #49486/TMRC).
Authors’ contributions
SH, RLDM and LMSM designed and initiated the study. TMSCP, MSCJ and RAOM
performed the laboratory analyses. TMSCP, RAOM, RLDM, SH and LMSM analysed and

135
interpreted the data. TMSCP, RAOM and LMSM wrote the manuscript. All authors read,
commented on and approved the final manuscript.
Acknowledgements
We thank all of the patients involved in this study.
Authors information
References
1. Alho RM, Machado KV, Val FF, Fraiji NA, Alexandre MA, Melo GC, et al. Alternative
transmission routes in the malaria elimination era: an overview of transfusion-transmitted
malaria in the Americas. Malar J. 2017;16:78.
2. Vallejo AF, Martinez NL, Tobon A, Alger J, Lacerda MV, Kajava AV, et al. Global
genetic diversity of the Plasmodium vivax transmission-blocking vaccine candidate
Pvs48/45. Malar J. 2016;15:202.
3. Howes RE, Battle KE, Mendis KN, Smith DL, Cibulskis RE, Baird JK, et al. Global
epidemiology of Plasmodium vivax. Am J Trop Med Hyg. 2016;95:15-34.
4. Brasil. Ministério da Saúde. Secretaria de Vigilância em Saúde. Manual de diagnóstico
laboratorial da malária. Brasília, p. 112, 2015.
5. Gomes MD, Vieira JL, Cassiano GC, Musset L, Legrand E, Nacher M, et al. Evaluation of
circumsporozoite protein of Plasmodium vivax to estimate its prevalence in Oiapoque,
Amapa state, Brazil, bordering French Guiana. Rev Inst Med Trop Sao Paulo. 2016;58:72.
6. Dotrario AB, Menon LJ, Bollela VR, Martinez R, de Almeida EADC, da Fonseca BA, et
al. Malaria and other febrile diseases among travellers: the experience of a reference centre
located outside the Brazilian Amazon region. Malar J. 2016;15:294.
7. Olliaro PL, Barnwell JW, Barry A, Mendis K, Mueller I, Reeder JC, et al. Implications of
Plasmodium vivax biology for control, elimination, and research. Am J Trop Med Hyg.
2016;95:4-14.
8. Mueller I, Shakri AR, Chitnis CE. Development of vaccines for Plasmodium vivax malaria.
Vaccine 2015;33:7489-95.
9. Geleta G, Ketema T. Severe malaria associated with Plasmodium falciparum and P. vivax
among children in Pawe Hospital, Northwest Ethiopia. Malar Res Treat. 2016.
10. Gupta P, Sharma R, Chandra J, Kumar V, Singh R, Pande V, et al. Clinical
manifestations and molecular mechanisms in the changing paradigm of vivax malaria in
India. Infect Genet Evol. 2016;39:317-24.
11. Greenwood BM, Fidock DA, Kyle DE, Kappe SH, Alonso PL, Collins FH, et al. Malaria:
progress, perils, and prospects for eradication. J Clin Invest. 2008;118:1266-76.

136
12. Arevalo-Herrera M, Solarte Y, Marin C, Santos M, Castellanos J, Beier JC, et al. Malaria
transmission blocking immunity and sexual stage vaccines for interrupting malaria
transmission in Latin America. Mem Inst Oswaldo Cruz. 2011;106:202-11.
13. Arévalo-Herrera M, Castellanos A, Yazdani SS, Shakri AR, Chitnis CE, Dominik R,
Herrera S. Immunogenicity and protective efficacy of recombinant vaccine based on the
receptor-binding domain of the Plasmodium vivax Duffy binding protein in Aotus
monkeys. Am J Trop Med Hyg. 2005;73(5 Suppl):25-31.
14. Tsuboi T, Tachibana M, Kaneko O, Torii M. Transmission-blocking vaccine of vivax
malaria. Parasitol Int. 2003;52:1-11.
15. Arevalo-Herrera M, Vallejo AF, Rubiano K, Solarte Y, Marin C, Castellanos A, et al.
Recombinant Pvs48/45 antigen expressed in E. coli generates antibodies that block malaria
transmission in Anopheles albimanus mosquitoes. PLoS One. 2015;10:E0119335.
16. Kang JM, Ju HL, Moon SU, Cho PY, Bahk YY, Sohn WM, et al. Limited sequence
polymorphisms of four transmission-blocking vaccine candidate antigens in Plasmodium
vivax Korean isolates. Malar J. 2013;12:144.
17. Cassiano GC, Furini AA, Capobianco MP, Storti-Melo LM, Almeida ME, Barbosa DR,
et al. Immunogenetic markers associated with a naturally acquired humoral immune
response against an N-terminal antigen of Plasmodium vivax merozoite surface protein 1
(PvMSP-1). Malar J. 2016;15:306.
18. Gomes MSM, Vieira JLF, Machado RLD, Nacher M, Stefani A, Musset L, et al. Efficacy
in the treatment of malaria by Plasmodium vivax in Oiapoque, Brazil, on the border with
French Guiana: the importance of control over external factors. Malar J. 2015;14:402.
19. Storti-Melo LM, Souza-Neiras WC, Cassiano GC, Taveira LC, Cordeiro AJ, Couto V, et
al. Evaluation of the naturally acquired antibody immune response to the Pv200l N-
terminal fragment of Plasmodium vivax merozoite surface protein-1 in four areas of the
Amazon region of Brazil. Am J Trop Med Hyg. 2011;84:58-63.
20. Ministério da Saúde. Manual de diagnóstico laboratorial da malária. Brasil: Secretaria de
Vigilância em Saúde; 2015.
21. Bennett JW, Yadava A, Tosh D, Sattabongkot J, Komisar J, Ware LA, et al. Phase 1/2a
trial of Plasmodium vivax malaria vaccine candidate VMP001/AS01B in malaria-naive
adults: safety, immunogenicity, and efficacy. PLoS Negl Trop Dis. 2016;10:E0004423.
22. Yadava A, Waters NC. Rationale for further development of a vaccine based on the
circumsporozoite protein of Plasmodium vivax. PLoS Negl Trop Dis. 2017;11.
23. Demissie Y, Ketema T. Complicated malaria symptoms associated with Plasmodium
vivax among patients visiting health facilities in Mendi town, Northwest Ethiopia. BMC
Infect Dis. 2016;16:436.
24. Mourao FR, da Cunha AC, Silva RA, Souza EB. A vigilância da malária na Amazônia
Brasileira. Open Journal System. 2014;4:161-8.

137
25. Furini AAC, Cassiano GC, Capobianco MP, dos Santos SEB, Machado RLD. Frequency
of TNFA, INFG, and IL10 gene polymorphisms and their association with malaria vivax
and genomic ancestry. Mediators Inflamm. 2016.
26. Cassiano GC, Santos EJ, Maia MH, Ada CF, Storti-Melo LM, Tomaz FM, et al. Impact
of population admixture on the distribution of immune response co-stimulatory genes
polymorphisms in a Brazilian population. Hum Immunol. 2015;76:836-42.
27. Vasquez-Jimenez JM, Arevalo-Herrera M, Henao-Giraldo J, Molina-Gomez K, Arce-
Plata M, Vallejo AF, et al. Consistent prevalence of asymptomatic infections in malaria
endemic populations in Colombia over time. Malar J. 2016;15:70.
28. Outchkourov NS, Roeffen W, Kaan A, Jansen J, Luty A, Schuiffel D, et al. Correctly
folded Pfs48/45 protein of Plasmodium falciparum elicits malaria transmission-blocking
immunity in Mice. Proc Natl Acad Sci U S A. 2008;105:4301-5.
29. Outchkourov N, Vermunt A, Jansen J, Kaan A, Roeffen W, Teelen K, et al. Epitope
analysis of the malaria surface antigen Pfs48/45 identifies a subdomain that elicits
transmission blocking antibodies. J Biol Chem. 2007;282:17148-56.
Figure captions
Fig. 1 Cities in the Brazilian Amazon region where the collections were performed
Source: Geomapas and IBGE.
Fig. 2 Distribution of the humoral immune response frequency against Pvs48/45 by city
Fig. 3 Dispersion of the reactivity indices of the samples in which anti-Pvs48/45-specific
antibody responses were found

138
ANEXO A – Parecer Consubstanciado do Comitê de Ética de Pesquisa

139
ANEXO B – Financiamento pelo Edital/CNPq/PPSUS – Amapá 2013